



1 23
EcoHealthConservation Medicine: HumanHealth:Ecosystem Sustainability Officialjournal of International Association forEcology and Health ISSN 1612-9202 EcoHealthDOI 10.1007/s10393-011-0725-7
Rodent-Borne Hantaviruses in Cambodia,Lao PDR, and Thailand
Kim Blasdell, Jean François Cosson,Yannick Chaval, Vincent Herbreteau,Bounneuang Douangboupha, SathapornJittapalapong, Ake Lundqvist, et al.

1 23
Your article is protected by copyright and all
rights are held exclusively by International
Association for Ecology and Health. This e-
offprint is for personal use only and shall not
be self-archived in electronic repositories. If
you wish to self-archive your work, please
use the accepted author’s version for posting
to your own website or your institution’s
repository. You may further deposit the
accepted author’s version on a funder’s
repository at a funder’s request, provided it is
not made publicly available until 12 months
after publication.

Rodent-Borne Hantaviruses in Cambodia, Lao PDR,and Thailand
Kim Blasdell,1,2 Jean Francois Cosson,3 Yannick Chaval,4 Vincent Herbreteau,5
Bounneuang Douangboupha,6 Sathaporn Jittapalapong,7 Ake Lundqvist,8 Jean-Pierre Hugot,9
Serge Morand,2 and Philippe Buchy1
1Virology Unit, Institut Pasteur du Cambodge, 5 Monivong Boulevard, BP 983, Phnom Penh, Cambodia2Institut des Sciences de l’Evolution, CNRS, IRD, Universite Montpellier 2, CC065, 34095 Montpellier Cedex 05, France3INRA, UMR CBGP (INRA/IRD/Cirad/Montpellier SupAgro), Campus International de Baillarguet, CS 30016, 34988 Montferrier sur Lez, France4Centre de Biologie et de Gestion et des Populations (CBGP), International de Baillarguet, CS 30016, 34988 Montferrier sur lez, France5CIRAD, UR AGIRs (Animal et Gestion Integree des Risques), Campus International de Baillarguet, Montpellier, France6National Agricultural Research Centre, National Agricultural and Forestry Research Institute, Vientiane, Lao PDR7Department of Parasitology, Faculty of Veterinary Medicine, Kasetsart University, Bangkok 10900, Thailand8Swedish Institute for Infectious Disease Control, Stockholm, Sweden9Origine, Structure et Evolution de la Biodiversite, Museum National d’Histoire Naturelle, Paris, France
Abstract: In order to evaluate the circulation of hantaviruses present in southeast Asia, a large scale survey of
small mammal species was carried out at seven main sites in the region (Cambodia, Lao People’s Democratic
Republic, and Thailand). Small scale opportunistic trapping was also performed at an eighth site (Cambodia).
Using a standard IFA test, IgG antibodies reacting to Hantaan virus antigens were detected at six sites.
Antibody prevalence at each site varied from 0 to 5.6% with antibodies detected in several rodent species
(Bandicota indica, B. savilei, Maxomys surifer, Mus caroli, M. cookii, Rattus exulans, R. nitidius, R. norvegicus,
and R. tanezumi). When site seroprevalence was compared with site species richness, seropositive animals were
found more frequently at sites with lower species richness. In order to confirm which hantavirus species were
present, a subset of samples was also subjected to RT-PCR. Hantaviral RNA was detected at a single site from
each country. Sequencing confirmed the presence of two hantavirus species, Thailand and Seoul viruses,
including one sample (from Lao PDR) representing a highly divergent strain of Seoul virus. This is the first
molecular evidence of hantavirus in Lao PDR and the first reported L segment sequence data for Thailand
virus.
Keywords: hantavirus, southeast Asia, rodents, serology, phylogeny, seoul virus variant
INTRODUCTION AND PURPOSE
Hantaviruses are single-stranded, tri-segmented, negative
sense RNA viruses belonging to the family Bunyaviridae.
Electronic supplementary material: The online version of this article
(doi:10.1007/s10393-011-0725-7) contains supplementary material, which is available
to authorized users.
Correspondence to: Philippe Buchy, e-mail: [email protected]
EcoHealthDOI: 10.1007/s10393-011-0725-7
Original Contribution
� 2011 International Association for Ecology and Health
Author's personal copy

At present, 45 distinct hantavirus species are recognized, all
associated with either rodent or soricomorph hosts (Dearing
and Dizney 2010). The genus is further subdivided into
species hosted by Sigmodontinae, Neotominae, Murinae,
and Arvicolinae rodent families and by the Soricidae and
Talpidae insectivorous families (Henttonen et al. 2008).
Hantaviruses are found throughout Eurasia, the
Americas, and Africa, where pathogenic variants are jointly
responsible for up to 200,000 human clinical cases annually
(Dearing and Dizney 2010; Song et al. 1984). Two syn-
dromes are caused by hantaviruses in humans: hantavirus
cardiopulmonary syndrome (HCPS), which is restricted to
the Americas and hemorrhagic fever with renal syndrome
(HFRS), with a much wider distribution including Eurasia,
Africa, and a few isolated cases in the Americas. Hantavirus
species hosted by Sigmodontinae and Neotominae rodents
are responsible for HCPS, while HFRS-associated hantavi-
ruses are generally hosted by Murinae rodents. However,
infections with Puumala virus, an arvicoline-borne virus,
are also associated with HFRS (Terajima et al. 2004). Tula
virus, another arvicoline-borne virus, is also potentially
linked with human infections, although disease has not
been confirmed (Schultze et al. 2002; Klempa et al. 2003).
With the exception of China, hantaviruses have been
poorly studied in southeast (SE) Asia. In China, 40,000–
60,000 human cases are reported annually (Bi et al. 2008)
but only isolated cases have been reported from other
countries in the region (e.g., Thailand, Suputthamongkol
et al. 2005; Singapore, Wong et al. 1989). However, pre-
vious studies have identified at least six hantavirus species
in rodent hosts in the region. These include Hantaan virus,
hosted by Apodemus agrarius (Lee et al. 1978), restricted
primarily to China; Seoul virus hosted by Rattus norvegicus
(Lee et al. 1982), which probably has a worldwide distri-
bution; Thailand virus, hosted by Bandicota indica and
present in Thailand (Xiao et al. 1994; Hugot et al. 2006);
Serang virus, hosted by R. tanezumi and recently identified
in Indonesia (Plyusnina et al. 2009); Thottapalayam virus,
hosted by Suncus murinus and found in India, Thailand,
and Indonesia (Carey et al. 1971; Okumura et al. 2007);
Cao Bang virus, hosted by Anurosorex squamipes and found
in Vietnam (Song et al. 2007). Other hantavirus species
may also be present, including partially characterized spe-
cies hosted by Rattus species in Cambodia (Reynes et al.
2003) and R. tanezumi in Singapore (Johansson et al. 2010)
which appear to be variants of Serang virus. Distinct
genetic variants of Seoul virus have also been identified in
China (Zou et al. 2008).
Although hantaviruses were previously thought to
share co-phylogeny with their rodent hosts (Khaiboullina
et al. 2005), recent findings of multiple new hantaviruses
and in particular, the sorciomorph-borne viruses, pose
some problems for the co-phylogeny theory and have called
this idea into debate (Henttonen et al. 2008; Ramsden et al.
2009; Schmidt-Chanasit et al. 2010). Each hantavirus spe-
cies or lineage is thought to be hosted predominantly by a
single rodent or soricomorph species or group of closely
related species. However, spill-over infections into other
species also commonly occur [spill-over of Dobrava virus
A. agrarius lineage into A. flavicollis (Schlegel et al. 2009);
Hantaan virus, usually hosted by Apodemus species but also
detected in R. norvegicus and Niviventer spp. (Yao et al.
2002; Li et al. 2005; Zou et al. 2008); Seoul virus infecting
several Rattus species and Mus musculus (Wang et al. 2000;
Jiang et al. 2008)]. These spill-over events may even have
led to host-switching events in some cases, further com-
plicating co-phylogeny analyses. Evidence for host-switch-
ing events has been proposed for both rodent- and
soricomorph-borne hantaviruses (Vapalahti et al. 1999;
Kang et al. 2009).
As SE Asia is a biodiversity hotspot with numerous
rodent and soricomorph species, unrecognized virus spe-
cies and/or hantavirus host species and lineages may be
present in the region. Our study aimed to determine the
prevalence of hantaviruses in a variety of SE Asian habitats
(including both disturbed and relatively undisturbed areas)
and to assess the virus species present. The data were also
assessed to establish if hantavirus presence was related to
the level of species richness in the rodent communities.
METHODS
Rodent Samplings
This research is part of the CERoPath (Community Ecology
of Rodents and their Pathogens in a changing environ-
ment) project, which aims to investigate the dynamics of
murine rodents and their pathogens (macro and micro-
parasites) in SE Asia. This region is characterized by high
biodiversity and growing modification of habitats due to
rapid economic development and insertion into the global
economy. Seven main sampling locations were selected:
Nan (19�150N; 100�830E), Loei (17�390N; 101�770E) and
Buriram (14�890N; 103�010E) in Thailand; Luang Prabang
(19�620N; 102�050E) and Champasak (15�120N; 105�800E)
in Laos; and Preah Sihanouk (10�710N; 103�820E) and
Kim Blasdell et al.
Author's personal copy

Mondolkiri (12�120N; 106�890E) in Cambodia (see Fig. 1).
These locations represent a variety of habitats, in relation to
human pressures and land usage (see below). Opportunistic
trapping was also carried out on a much smaller scale in the
environs of Phnom Penh, Cambodia (11�330N; 104�550E).
For the seven main localities, two sampling sessions
were conducted, one during the wet season and one during
the dry season. At each of these locations, rodents were
sampled from four habitat types: forest (including natural
forest and plantations); non-flooded habitats (including
shrub, orchards and upland agriculture); lowland flooded
habitats (rice paddies); and peridomestic (houses and
immediate surrounding areas). Trapping took place within
an area of approximately 100 km2. For each session 30 lines
of ten traps were set over 4 days, amounting to 1,200 trap
nights. These trap nights were evenly distributed between
the first three habitat types (forest, non-flooded habitats,
and lowland flooded habitats) with 400 trap nights set in
each habitat. In addition, animals were also trapped by
local hunters in peridomestic habitats, ranging from 334
(Loei) to 822 (Buriram) trap nights per site with a mean of
546 trap nights. Greater use of local hunters was employed
at sites with low species richness in an attempt to reach
local rodent species richness. Local handmade wire live
traps, baited with cassava, banana, and sticky rice were set
for 4 days at each line and checked daily. Animals were
euthanized and a blood pellet, serum, and organs were
collected, stored temporarily in liquid nitrogen and then
transferred to long term storage at -80�C.
In Phnom Penh, animals were captured at several loca-
tions within and around the city from both domestic
dwellings and agricultural land on an opportunistic basis.
Traps, baits, and animal processing were as described above.
Rodent Species Determination
Rodent species were determined in the field when possible
using morphological criteria as described in the literature
(Marshall 1988; Aplin et al. 2003; Pages et al. 2010). Most
Bandicota were determined using the criteria used by Aplin
et al. (2003) but ambiguous specimens (mainly subadults)
were subjected to a PCR targeting the D-loop gene on the
mitochondrial DNA (Chaval et al. 2010). As morphological
criteria were not fully discriminant between some species
within the genera Rattus, Niviventer, and Mus, a molecular
approach was thus carried out by sequencing the COI gene.
Species determination was performed using the freely
available molecular tool developed for SE Asian rodent
species molecular identification (http://www.ceropath.org/
barcoding_tool/rodentsea).
Within the seven main sites, rodent species richness was
estimated both by the observed number of species (Sobs) and
the estimated number corrected for sample size (Sk). Sk was
calculated by the rarefaction procedure in BiodivR v. 1.1
(Hardy 2009) and corresponds to the expected number of
species in a sample of k rodents. In our case, k was fixed to 170,
i.e., the lowest sample size recorded among our seven sites.
Serology
Hantaan virus-infected Vero E6 cells fixed on slides were
used to perform indirect fluorescent antibody (IFA)
serology using previously described methods (Schilling
et al. 2007). As hantavirus antibodies are known to cross
react between hantavirus species (Hujakka et al. 2003),
species determination was not possible. Serum samples
were diluted 1:30 with PBS. Secondary antibody,
Figure 1. Geographic locations of trapping sites.
Hantaviruses in Southeast Asian Rodents
Author's personal copy

comprising a mixture of goat anti mouse IgG and goat anti-
rat IgG, both conjugated with fluorescein isothiocyanate
(FITC) (Sigma, Steinheim, Germany), was used at a dilu-
tion of 1:50 in PBS. Fluorescence was visualized using ultra
violet microscopy.
Hantavirus Molecular Identification
Lung samples from seropositive animals, from additional
individuals belonging to species with high seroprevalence
and from individuals where serum samples were not
available for screening by IFA, were selected for molecular
testing. Lung samples were homogenized (50 mg sample in
500 ll PBS) using the MagNA Lyzer Instrument (230 V,
Roche, cat. no. 03358976001) and bead system, then RNA
extracted using the Qiagen Viral RNA extraction kit (cat.
no. 52960). To determine hantavirus species, samples were
screened for hantavirus RNA using previously described
reverse-transcription nested RT-PCRs targeting the L
segment (Klempa et al. 2006) and S segment (Papa et al.
1998). The Superscript� III one-step RT-PCR system with
Platinum� Taq (Invitrogen, cat. no. 12574026) was used
for first round PCR and Promega GoTaq� Flexi DNA
polymerase (cat. no. M8295) was used for nested PCRs.
Amplicons of the expected size (324 and 945 bp for L and S
segments, respectively) were sequenced using a MJ
Research PTC-225 Peltier Thermal Cycler and ABI PRISM
BigDyeTM Terminator Cycle Sequencing Kits with Amp-
liTaq DNA polymerase (FS enzyme: Applied Biosystems).
For phylogenetic analysis, sequences for other hantavi-
ruses were obtained from GenBank. Specifically, published
data documented from the Asian region was collated (Sup-
plementary Table). Because the rodents collected in this
study almost entirely consisted of species belonging to the
Murinae subfamily, viruses isolated from murines (rats and
mice) and from human patients were focused on. Hantavi-
ruses from voles, sigmodontines, and insectivores were not
included in the analysis due to their distant phylogenetic
relationship to murine-borne hantaviruses. Data sets of S and
L segments were analyzed separately. Sequences were aligned
using the Muscle option in Seaview software v.4.2.12 (Gouy
et al. 2010). As both partial segments corresponded to coding
sequences, predicted amino acid sequences were also ana-
lyzed. Percentage differences between sequences (constitut-
ing the number of nucleotides/amino acids differing between
two sequences, by sequence length) were calculated in MEGA
4.0 (Tamura et al. 2007). The best model of evolution for each
data set was determined using jModelTest 0.1.1 (Posada
2008). Phylogenetic reconstructions were performed using
the Maximum Likelihood method of Phyml 3.0 (Guindon
and Gascuel 2003), implemented in Seaview using the model
of evolution selected by jModelTest (GTR model), and
visualized in FigTree v1.3.1 (http://tree.bio.ed.ac.uk/). Nodal
supports were determined using the approximate likelihood
ratio test approach.
RESULTS
Trapping Effort
During sessions where species richness was low, hunter
trapping pressure was increased. Hence there was a sig-
nificant negative correlation between collected rodent
species richness and local hunter trapping nights (in log;
slope = -0.37, df = 7, adjusted R2: 0.45, P 0.028), indi-
cating that increasing trapping by local hunters did not
significantly increase rodent species richness.
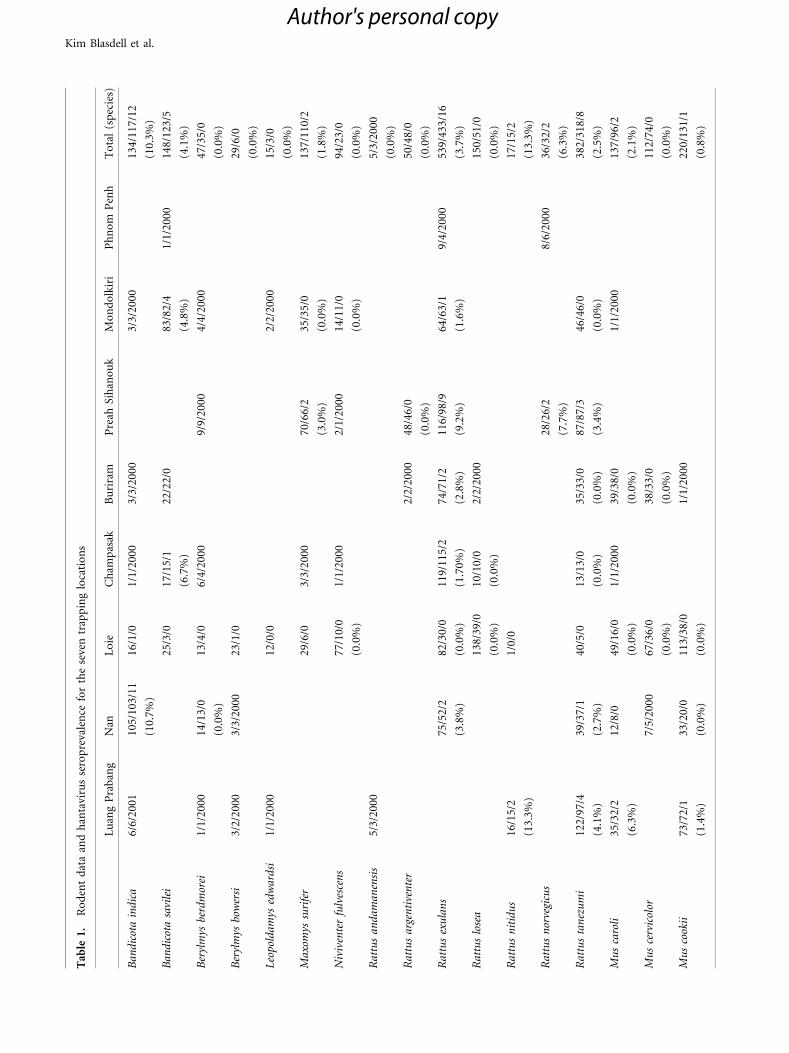
Hantavirus Seroprevalence
A total of 2,364 small mammals were collected during this
study, of which 1,640 rodents were tested by IFA for IgG
antibodies to Hantaan virus and 50 tested positive. Anti-
Hantaan virus antibodies were detected in nine species,
namely (in order of prevalence) R. nitidius, B. indica,
R. norvegicus, B. savilei, R. exulans, R. tanezumi, M. caroli,
Maxomys surifer, and M. cookii (Table 1). Four species did
not test positive despite a significant number of samples
analyzed (�35): Berylmys berdmorei, R. argentiventer,
R. losea, and M. cervicolor. Six species captured and tested
in low numbers (�23) also all tested negative: B. bowersi,
Leopoldamys edwardsi, Niviventer fulvescens, R. andaman-
ensis and the non-murine species Cannomys badius and
Rhizomys pruinosus. Given the low number of captures for
these species, we would be unlikely to be able to detect a
low prevalence of hantavirus infection in these populations.
Between zero and five species were found seropositive at
any given site. Seropositive R. exulans were identified at five
sites; R. tanezumi at three sites; B. indica and B. savilei at
two sites; and R. nitidus, R. norvegicus, M. surifer, M. cookii,
and M. caroli each at a single site (Table 1).
Anti-hantavirus antibodies were detected in every
country and at six of the seven sites studied. All main sites
were sampled twice, but serum was only available from the
second trapping session in Loei, Thailand. Overall hanta-
virus antibody prevalence at the trapping sites was 0% for
Kim Blasdell et al.
Author's personal copy

Phnom Penh, 3.9% for Mondulkiri in March 2009 but 0%
in November 2009; 0% in April 2009 and 3.2% in
November 2009 for Champasak; 4.4% for Luang Prabang
in July 2008 and 3.9% in February 2010; 5.1% for Nan in
November 2008 and 6.4% in March 2010; 1.2% for Buri-
ram in November 2008 and 0.8% in June 2009; 4.5% in
November 2008 and 5.1% in July 2009 for Preah Sihanouk.
Serological testing of samples did not reveal any seroposi-
tive individuals in Loei despite a large sampling size (196
rodents tested from session two only) (Table 1).
Overall, serological evidence of previous exposure to
hantavirus was found to be negatively correlated with
rodent richness (Kendall’s rank correlation, Sobs:
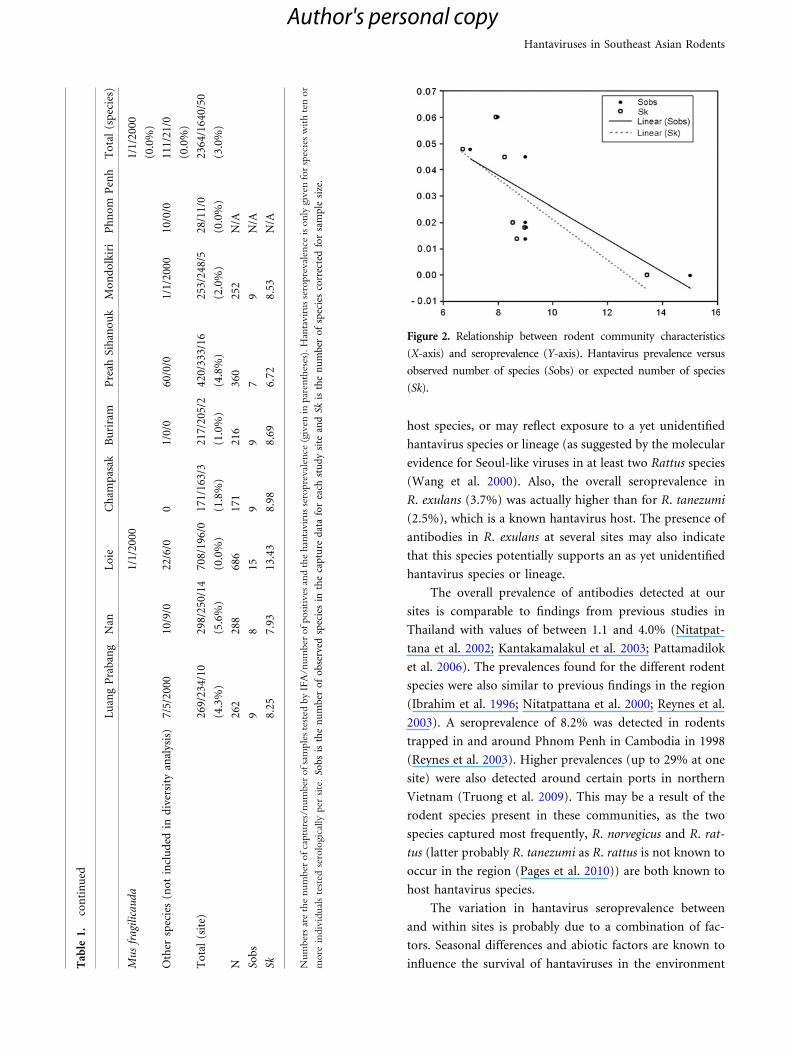
P = 0.020; Sk: P = 0.011) (Table 1; Fig. 2).
Molecular Analysis
As the serological data could not determine the infecting
hantavirus species, a subset of 520 samples were subjected
to the partial L segment nested RT-PCR and of these, 132
were also tested by the partial S segment nested RT-PCR
(data not shown) to allow species identification. All sero-
positive samples were subjected to this test, along with a
partially random subset of other samples. Due to the
unavailability of sera from the first trapping session in Loei,
a large number of the samples from this session were tested
by RT-PCR (159 in total) to compensate for the lack of
serological testing. Of the 520 samples tested, ten were
confirmed positive by both RT-PCRs (although sequences
for the partial S segment were only successfully obtained
from nine samples). The majority of these positive samples
(eight samples) were obtained from B. indica from the Nan
site in Thailand (sample IDs R5320, R5370, R6083, R6098,
R6107, R6108, R6109, and R6153). One sample was from a
R. tanezumi captured in Luang Prabang, Lao PDR (L0199),
and one was from a R. norvegicus captured in Phnom Penh,
Cambodia (PP21).
Sequencing and subsequent genetic analysis indicated
that these samples represented at least two species of han-
tavirus, namely Thailand virus (found at the Nan site,
Thailand) and Seoul virus (in Phnom Penh, Cambodia and
Luang Prabang, Lao PDR). Details of sequences included in
the p-distance and phylogenetic analysis can be found in
the Supplementary Table. Nucleotide sequence compari-
sons (P distances, data not shown) found PP21, from
Phnom Penh, Cambodia, to be closely related to several
strains of Seoul virus (up to 99 and 94% sequence
homology for S and L segments, respectively). The isolates
from Nan, Thailand, showed high sequence homology for
the S segment with previous isolates of Thailand virus. This
is the first known study to include L segment sequences of
Thailand virus. The virus L0199 from Luang Prabang, Lao
PDR, appeared to be a highly divergent variant of Seoul
virus. The S segment of this virus showed highest nucleo-
tide homology to a previously isolated Cambodian strain of
Seoul virus (AJ427506, GenBank) and to PP21, which was
also isolated in Cambodia (945 nucleotides: 85%). How-
ever, for the partial L segment (324 nucleotides) L0199 was
found most closely related (86% nucleotide identity) to two
viruses from China, CGRn8316 (GenBank accession
number EF990931) and CGRn9415 (GenBank accession
number EF990930), isolated from R. norvegicus and
believed to be natural recombinants between Seoul and
Hantaan viruses (Zou et al. 2008).
Phylogenetic analyses supported the genetic analysis
for all viruses. The divergent Laos virus grouped distantly
with other Seoul virus isolates at the nucleotide level for
both S and L segments and more closely for the partial
nucleoprotein sequence. S segment analysis indicated that
this Laotian isolate was the most basal of the Seoul samples
(Fig. 3), while L segment analyses showed L0199 forming a
sister taxa with the two suspected Seoul–Hantaan
recombinant viruses (Fig. 4). In contrast, when predicted
amino acid identities were compared, strain L0199 grouped
closely with other isolates of Seoul and was found to share
approximately 97 and 100% homology for L protein and
nucleoprotein, respectively.
DISCUSSION
The findings from this study increase the number of areas in
SE Asia where hantaviruses are known to circulate. Of the
sites studied, anti-hantavirus antibodies and hantaviral RNA
have previously only been detected at Buriram (Nitatpattana
et al. 2002; Pattamadilok et al. 2006) so this is the first evi-
dence of the presence of hantavirus at Luang Prabang,
Champasak, Mondulkiri, Preah Sihanouk, and Nan and the
first molecular detection of hantaviruses in Lao PDR.
This study supports previous findings that many SE
Asian rodent species can be infected with hantaviruses
(Ibrahim et al. 1996; Nitatpattana et al. 2000, 2002), with
nine species found seropositive. The antibodies detected in
some of the rodent species here may represent spillover
infections of known hantavirus species into non-reservoir
Hantaviruses in Southeast Asian Rodents
Author's personal copy
Tab
le1.
Ro
den
td
ata
and
han
tavi
rus
sero
pre
vale
nce
for
the
seve
ntr
app
ing
loca
tio
ns
Lu
ang
Pra
ban
gN
anL
oie
Ch
amp
asak
Bu
rira
mP
reah
Sih
ano
uk
Mo
nd
olk
iri
Ph
no
mP
enh
To
tal
(sp
ecie
s)
Ban
dic
ota
ind
ica
6/6/
2001
105/
103/
1116
/1/0
1/1/
2000
3/3/
2000
3/3/
2000
134/
117/
12
(10.
7%)
(10.
3%)
Ban
dic
ota
savi
lei
25/3
/017
/15/
122
/22/
083
/82/
41/
1/20
0014
8/12
3/5
(6.7
%)
(4.8
%)
(4.1
%)
Ber
ylm
ysbe
rdm
orei
1/1/
2000
14/1
3/0
13/4
/06/
4/20
009/
9/20
004/
4/20
0047
/35/
0
(0.0
%)
(0.0
%)
Ber
ylm
ysbo
wer
si3/
2/20
003/
3/20
0023
/1/0
29/6
/0
(0.0
%)
Leo
pold
amys
edw
ard
si1/
1/20
0012
/0/0
2/2/
2000
15/3
/0
(0.0
%)
Max
omys
suri
fer
29/6
/03/
3/20
0070
/66/
235
/35/
013
7/11
0/2
(3.0
%)
(0.0
%)
(1.8
%)
Niv
iven
ter
fulv
esce
ns
77/1
0/0
1/1/
2000
2/1/
2000
14/1
1/0
94/2
3/0
(0.0
%)
(0.0
%)
(0.0
%)
Rat
tus
and
aman
ensi
s5/
3/20
005/
3/20
00
(0.0
%)
Rat
tus
arge
nti
ven
ter
2/2/
2000
48/4
6/0
50/4
8/0
(0.0
%)
(0.0
%)
Rat
tus
exu
lan
s75
/52/
282
/30/
011
9/11
5/2
74/7
1/2
116/
98/9
64/6
3/1
9/4/
2000
539/
433/
16
(3.8
%)
(0.0
%)
(1.7
0%)
(2.8
%)
(9.2
%)
(1.6
%)
(3.7
%)
Rat
tus
lose
a13
8/39
/010
/10/
02/
2/20
0015
0/51
/0
(0.0
%)
(0.0
%)
(0.0
%)
Rat
tus
nit
idu
s16
/15/
21/
0/0
17/1
5/2
(13.
3%)
(13.
3%)
Rat
tus
nor
vegi
cus
28/2
6/2
8/6/
2000
36/3
2/2
(7.7
%)
(6.3
%)
Rat
tus
tan
ezu
mi
122/
97/4
39/3
7/1
40/5
/013
/13/
035
/33/
087
/87/
346
/46/
038
2/31
8/8
(4.1
%)
(2.7
%)
(0.0
%)
(0.0
%)
(3.4
%)
(0.0
%)
(2.5
%)
Mu
sca
roli
35/3
2/2
12/8
/049
/16/
01/
1/20
0039
/38/
01/
1/20
0013
7/96
/2
(6.3
%)
(0.0
%)
(0.0
%)
(2.1
%)
Mu
sce
rvic
olor
7/5/
2000
67/3
6/0
38/3
3/0
112/
74/0
(0.0
%)
(0.0
%)
(0.0
%)
Mu
sco
okii
73/7
2/1
33/2
0/0
113/
38/0
1/1/
2000
220/
131/
1
(1.4
%)
(0.0
%)
(0.0
%)
(0.8
%)
Kim Blasdell et al.
Author's personal copy
host species, or may reflect exposure to a yet unidentified
hantavirus species or lineage (as suggested by the molecular
evidence for Seoul-like viruses in at least two Rattus species
(Wang et al. 2000). Also, the overall seroprevalence in
R. exulans (3.7%) was actually higher than for R. tanezumi
(2.5%), which is a known hantavirus host. The presence of
antibodies in R. exulans at several sites may also indicate
that this species potentially supports an as yet unidentified
hantavirus species or lineage.
The overall prevalence of antibodies detected at our
sites is comparable to findings from previous studies in
Thailand with values of between 1.1 and 4.0% (Nitatpat-
tana et al. 2002; Kantakamalakul et al. 2003; Pattamadilok
et al. 2006). The prevalences found for the different rodent
species were also similar to previous findings in the region
(Ibrahim et al. 1996; Nitatpattana et al. 2000; Reynes et al.
2003). A seroprevalence of 8.2% was detected in rodents
trapped in and around Phnom Penh in Cambodia in 1998
(Reynes et al. 2003). Higher prevalences (up to 29% at one
site) were also detected around certain ports in northern
Vietnam (Truong et al. 2009). This may be a result of the
rodent species present in these communities, as the two
species captured most frequently, R. norvegicus and R. rat-
tus (latter probably R. tanezumi as R. rattus is not known to
occur in the region (Pages et al. 2010)) are both known to
host hantavirus species.
The variation in hantavirus seroprevalence between
and within sites is probably due to a combination of fac-
tors. Seasonal differences and abiotic factors are known to
influence the survival of hantaviruses in the environmentTab
le1.
con
tin
ued
Lu
ang
Pra
ban
gN
anL
oie
Ch
amp
asak
Bu
rira
mP
reah
Sih
ano
uk
Mo
nd
olk
iri
Ph
no
mP
enh
To
tal
(sp
ecie
s)
Mu
sfr
agil
icau
da
1/1/
2000
1/1/
2000
(0.0
%)
Oth
ersp
ecie
s(n
ot
incl
ud
edin
div
ersi
tyan
alys
is)
7/5/
2000
10/9
/022
/6/0
01/
0/0
60/0
/01/
1/20
0010
/0/0
111/
21/0
(0.0
%)
To
tal
(sit
e)26
9/23
4/10
298/
250/
1470
8/19
6/0
171/
163/
321
7/20
5/2
420/
333/
1625
3/24
8/5
28/1
1/0
2364
/164
0/50
(4.3
%)
(5.6
%)
(0.0
%)
(1.8
%)
(1.0
%)
(4.8
%)
(2.0
%)
(0.0
%)
(3.0
%)
N26
228
868
617
121
636
025
2N
/A
Sob
s9
815
99
79
N/A
Sk8.
257.
9313
.43
8.98
8.69
6.72
8.53
N/A
Num
ber
sar
eth
enum
ber
ofca
ptu
res/
num
ber
ofsa
mple
ste
sted
by
IFA
/num
ber
ofpositives
and
the
han
tavir
usse
ropre
val
ence
(giv
enin
par
enth
eses
).H
anta
vir
usse
ropre
val
ence
isonly
giv
enfo
rsp
ecie
sw
ith
ten
or
more
indiv
idual
ste
sted
sero
logic
ally
per
site
.So
bs
isth
en
um
ber
of
ob
serv
edsp
ecie
sin
the
cap
ture
dat
afo
rea
chst
ud
ysi
tean
dSk
isth
en
um
ber
of
spec
ies
corr
ecte
dfo
rsa
mp
lesi
ze.
Figure 2. Relationship between rodent community characteristics
(X-axis) and seroprevalence (Y-axis). Hantavirus prevalence versus
observed number of species (Sobs) or expected number of species
(Sk).
Hantaviruses in Southeast Asian Rodents
Author's personal copy

itself (Kallio et al. 2006; Linard et al. 2007; Dearing et al.
2009) and the difference in primary host density (and
population and community demographics) will also have
an effect (Calisher et al. 2007; Suzan et al. 2009). Previous
studies have also identified a trend for hantavirus preva-
lence to increase with decreasing species diversity (Mills
2006; Clay et al. 2009; Dizney and Reudas 2009; Suzan et al.
2009). It has been postulated that, because in many cases
the rodent species remaining in communities of low species
richness are those that are the reservoir hosts of hantavi-
ruses, this would allow for hantaviruses to circulate at a
higher prevalence in these communities (Mills 2006). It has
been suggested that this negative correlation may in part be
due to the dilution effect (Ostfeld and Keesing 2000), where
increasing species diversity is predicted to decrease patho-
gen prevalence through several mechanisms including
reduced encounters between hosts and decreased host
density (Keesing et al. 2006). Due to limitations in our data
set, we were unable to explore this finding further.
In this study, hantaviruses were found to infect animals
living in many different habitats, and particularly species
associated with human habitats (settlements and agricul-
ture). Similar findings have been observed for hantaviruses
circulating in South America (Mills 2006; Armien et al.
2009). As these areas also probably pose the greatest
potential for human transmission it is here that transmis-
sion prevention needs to be implemented in priority.
Of the three murine species found positive for hanta-
virus RNA in this study, two were the presumed natural
hosts of the viruses detected [Thailand virus in B. indica
(Xiao et al. 1994) and Seoul virus in R. norvegicus (Lee et al.
1982)]. However, Seoul virus was also identified in
R. tanezumi, a species previously associated with Serang
virus (Plyusnina et al. 2009). At least one isolate of Seoul
virus (Gou3 strain) has been collected from a rodent of the
R. rattus species complex previously (Wang et al. 2000) and
this isolate was also found to be genetically divergent from
other Seoul virus strains. It is conceivable that R. tanezumi
is the reservoir host for a distinct lineage of Seoul virus in
this area of Laos, as no R. norvegicus were encountered
during extensive trapping in this region. The divergent
nature of the nucleotide sequence of this sample may reflect
adaptation to R. tanezumi, as may also be the case for Gou3
virus to R. rattus.
Seoul virus regularly causes human infections, pri-
marily in east and SE Asia (Wong et al. 1989; Plyusnina
Figure 3. Maximum likelihood
tree for the partial S segment of
murine hantaviruses, showing the
position of sequences identified in
this study. Bootstrap values for the
major clades are indicated.
Kim Blasdell et al.
Author's personal copy

et al. 2004; Kariwa et al. 2007) and sporadically elsewhere
(Cueto et al. 2008; Heyman et al. 2009). There is also
serological data suggesting human infection by Thailand
virus (Pattamadilok et al. 2006) although this virus has
never been isolated from a patient. However, all three
rodent host species found positive for hantavirus RNA in
this study are associated with human habitats, indicating
the potential for zoonotic transmission. Rattus norvegicus is
primarily associated with human urban habitation and
both R. tanezumi and B. indica are found in a variety of
habitats including agricultural land and around (and in the
case of R. tanezumi, in) human habitation in rural areas
(Aplin et al. 2003).
This association with human environments has been
demonstrated in at least one case to increase the reproduc-
tive capacity of rodent species by increasing the duration of
the breeding season (Kuenzi et al. 2001). Most known out-
breaks of hantavirus disease have occurred in anthropo-
logically disturbed habitats, where natural biodiversity has
been reduced to low levels (Mills 2006). As with elsewhere,
most of the rodent species that host hantaviruses in SE Asia
are highly adaptable to many environments and they often
constitute one of only two or three rodent species present
within certain urban sites. These species can often reach high
densities due to the lack of competitive pressure. The low
rodent species richness (as a measure of biodiversity) asso-
ciated with high seroprevalence observed in our study sup-
ports previous findings from Vietnam and Cambodia
(Reynes et al. 2003; Truong et al. 2009). This finding may
have serious implications for SE Asian habitats, as many are
undergoing modification and no doubt, subsequent reduc-
tions in their natural species richness.
CONCLUSION
Nine murine rodent species were found to have been in-
fected with hantavirus in this study, with molecular analysis
confirming the presence of at least two species of hanta-
virus. This study presents the first molecular evidence for
hantaviruses in Lao PDR and indicates the presence of a
divergent strain of Seoul virus circulating in this country.
This study also presents the first L segment sequence data
for Thailand hantavirus. Our findings also suggest that
hantavirus seroprevalence is related to rodent species
richness, which is a potentially important finding in a
Figure 4. Maximum likelihood
tree for the partial L segment of
murine hantaviruses, showing the
position of sequences identified in
this study. Bootstrap values for the
major clades are indicated.
Hantaviruses in Southeast Asian Rodents
Author's personal copy

region where species richness is changing rapidly in many
areas.
ACKNOWLEDGMENTS
The authors wish to thank Mr. Hul Vibol and Mr. Y
Bunthin for their technical assistance in both the laboratory
and the field and to Mr. Kim Aun for his assistance in the
field. Also to Maria Walhstrom for kindly preparing the IFA
slides used in this study. The CERoPath project (Commu-
nity ecology of rodents and their pathogens in South East
Asia: effects of biodiversity changes and implications in
health ecology/ANR 07 BDIV 012) is funded by the Agence
Nationale de la Recherche (ANR, France).
REFERENCES
Aplin KP, Brown PR, Jacob J, Krebs CJ, Singleton GR (2003) FieldMethods for Rodent Studies in Asia and the Indo-Pacific, Mel-bourne: ACIAR
Armien AG, Armien B, Koster F, Pascale JM, Avila M, Gonzalez P,et al. (2009) Hantavirus infection and habitat associationsamong rodent populations in agroecosystems of Panama:implications for human disease risk. American Journal ofTropical Medicine and Hygiene 81:59–66
Bi Z, Formenty PBH, Roth CE (2008) Hantavirus infection: areview and global update. Journal of Infection in DevelopingCountries 2:3–21
Calisher CH, Wagoner KD, Amman BR, Root JJ, Douglass RJ,Kuenzi AJ, et al. (2007) Demographic factors associated withprevalence of antibody to Sin Nombre virus in deer mice in thewestern United States. Journal of Wildlife Disease 43:1–11
Carey DE, Reuben R, Panicker KN, Shope RE, Myers RM (1971)Thottapalayam virus: a presumptive arbovirus isolated from ashrew in India. Indian Journal of Medical Research 59:1758–1760
Chaval Y, Dobigny G, Michaux J, Pages M, Corbisier C, CossonJF, et al. (2010) A multi-approach survey as the most reliabletool to accurately assess biodiversity: an example of Thai murinerodents. Kasetsart Journal of Natural Science 44:590–603
Clay CA, Lehmer EM, Jeor SS, Dearing MD (2009) Sin nombrevirus and rodent species diversity: a test of the dilution andamplification hypotheses. Public Library of Science One 4:e6467.doi:10.1371/journal.pone.0006467
Cueto GR, Cavia R, Bellomo C, Padula PJ, Suarez OV (2008)Prevalence of hantavirus infection in wild Rattus norvegicus andR. rattus populations of Buenos Aires City, Argentina. TropicalMedicine and International Health 13:46–51. doi:10.1111/j.1365-3156.2007.01968.x
Dearing MD, Dizney L (2010) Ecology of hantaviruses in achanging world. Annals of the New York Academy of Sciences1195:99–112
Dearing MD, Previtali MA, Jones JD, Ely PW, Wood BA (2009)Seasonal variation in Sin Nombre virus infections in deer mice:preliminary results. Journal of Wildlife Diseases 45:430–436
Dizney LJ, Reudas LA (2009) Increased host species diversity anddecreased prevalence of Sin Nombre virus. Emerging InfectiousDiseases 15:1012–1018
Gouy M, Guindon S, Gascuel O (2010) SeaView version 4: amultiplatform graphical user interface for sequence alignmentand phylogenetic tree building. Molecular Biology and Evolution27:221–224
Guindon S, Gascuel O (2003) A simple, fast, and accurate algo-rithm to estimate large phylogenies by maximum likelihood.Systems Biology 52:696–704
Hardy OJ (2009) BiodivR 1.1. A program to compute indices ofspecies diversity within sample and species similarity betweensamples using rarefaction principles to reduce sampling bias.Available via http://ebe.ulb.ac.be/ebe/Software.html
Henttonen H, Buchy P, Suputtamongkol Y, Jittapalapong S,Herbreteau V, Laakkonen J, et al. (2008) Recent discoveries ofnew hantaviruses widen their range and question their origins.Animal Biodiversity and Emerging Diseases 1149:84–89
Heyman P, Baert K, Plyusnina A, Cochez C, Lundkvist AEsbroeckMV, et al. (2009) Serological and genetic evidence for thepresence of Seoul hantavirus in Rattus norvegicus in Flanders,Belgium. Scandinavian Journal of Infectious Diseases 41:51–56
Hugot JP, Plyusnina A, Herbreteau V, Nemirov K, Laakkonen J,Lundkvist A, et al. (2006) Genetic analysis of Thailand hanta-virus in Bandicota indica trapped in Thailand. Virology Journal3:72–81. doi:10.1186/1743-422X-3-72
Hujakka H, Koistinen V, Kuronen I, Eerikainen P, Parviainen M,Lundkvist A, et al. (2003) Diagnostic rapid tests for acutehantavirus infections: specific tests for Hantaan, Dobrava andPuumala viruses versus a hantavirus combination test. Journal ofVirological Methods 108:117–122
Ibrahim IN, Sudomo M, Morita C, Uemura S, Muramatsu Y,Ueno H, et al. (1996) Seroepidemiological survey of wild rats forSeoul virus in Indonesia. Japanese Journal of Medical Science andBiology 49:69–74
Jiang JF, Zuo SQ, Zhang WY, Wu XM, Tang F, De Vlas SJ, et al.(2008) Prevalence and genetic diversities of hantaviruses inrodents in Beijing, China. American Journal of Tropical Medicineand Hygiene 78:98–105
Johansson P, Yap G, Low HT, Siew CC, Kek R, Ng LC, et al.(2010) Molecular characterization of two hantavirus strainsfrom different Rattus species in Singapore. Virology Journal7:15–24. doi:10.1186/1743-422X-7-15
Kallio ER, Klingstrom J, Gustafsson E, Manni T, Vaheri A,Henttonen H, et al. (2006) Prolonged survival of Puumalahantavirus outside the host: evidence for indirect transmissionvia the environment. Journal of General Virology 87:2127–2134.doi:10.1099/vir.0.81643-0
Kang HJ, Bennett SN, Dizney L, Sumibcay L, Arai S, Ruedas LA,et al. (2009) Host switch during evolution of a genetically dis-tinct hantavirus in the American shrew mole (Neurotrichusgibbsii). Virology 388:8–14 . doi:10.1016/j.virol.2009.03.019
Kantakamalakul W, Siritantikorn S, Thongcharoen P, Singchai C,Puthavathana P (2003) Prevalence of rabies virus and Hantaanvirus infections in commensal rodents and shrews trapped inBangkok. Journal of the Medical Association of Thailand 86:1008–1014
Kariwa H, Yoshimatsu K, Arikawa J (2007) Hantavirus infectionin East Asia. Comparative Immunology, Microbiology and Infec-tious Diseases 30:341–356
Keesing F, Holt RD, Ostfeld RS (2006) Effects of species diversityon disease risk. Ecology Letters 9:485–498
Kim Blasdell et al.
Author's personal copy

Khaiboullina SF, Morzunov SP, St Jeor SC (2005) Hantaviruses:molecular biology, evolution and pathogenesis. Current Molec-ular Medicine 5:773–790
Klempa B, Fichet-Calvet E, Lecompte E, Auste B, Aniskin V,Meisel H, et al. (2006) Hantavirus in African wood mouse,Guinea. Emerging Infectious Diseases 12:838–840
Klempa B, Meisel H, Rath S, Bartel J, Ulrich R, Kruger DH (2003)Occurrence of renal and pulmonary syndrome in a region ofnortheast Germany where Tula hantavirus circulates. Journal ofClinical Microbiology 41:4894–4897
Kuenzi AJ, Douglass RJ, White D Jr, Bond CW, Mills JN (2001)Antibody to sin nombre virus in rodents associated with peri-domestic habitats in west central Montana. American Journal ofTropical Medicine and Hygiene 64:137–146
Lee HW, Baek LJ, Johnson KM (1982) Isolation of Hantaan virus,the etiological agent of Korean hemorrhagic fever from wildurban rats. Journal of Infectious Diseases 146:638–644
Lee HW, Lee PW, Johnson KM (1978) Isolation of the etiologicalagent of Korean hemorrhagic fever. Journal of Infectious Diseases137:298–308
Li L, Yang DJ, Chen JY, Ding JQ, Su X, Tian YQ, Wei JF (2005)Cloning and sequencing of S segment of hantavirus strain TJJ16isolated in Tianjin. Chinese Journal of Microbiology and Immu-nology 25:652–655 (In Chinese)
Linard C, Tersago K, Leirs H, Lambin EF (2007) Environmentalconditions and Puumala virus transmission in Belgium. Inter-national Journal of Health Geographics 6:55–66 . doi:10.1186/1476-072X-6-55
Marshall JT (1988) Family Muridae. In: Mammals of Thailand,Lekagul B, McNeely JA (editors), Bangkok: Darnsutha Press, pp397–487
Mills JN (2006) Biodiversity loss and emerging infectious disease:an example from the rodent-borne hemorrhagic fevers. Biodi-versity 7:9–17
Nitatpattana N, Henrich T, Palabodeewat S, Tangkanakul W,Poonsuksombat D, Chauvancy G, et al. (2002) Hantaan virusantibody prevalence in rodent populations of several provincesof northeastern Thailand. Tropical Medicine and InternationalHealth 7:840–845
Nitatpattana N, Chauvancy G, Dardaine J, Poblap T, JumronsawatK, Tangkanakul W, et al. (2000) Serological study of hantavirusin the rodent population of Nakhon Pathom and NakhonRatchasima Provinces Thailand. Southeast Asian Journal ofTropical Medicine and Public Health 31:277–282
Okumura M, Yoshimatsu K, Kumperasart S, Nakamura I, OginoM, Taruishi M, et al. (2007) Development of serological assaysfor Thottapalayam virus, an insectivore-borne Hantavirus.Clinical Vaccine Immunology 14:173–181 . doi:10.1128/CVI.00347-06
Ostfeld RS, Keesing F (2000) Biodiversity and disease risk: the caseof Lyme disease. Conservation Biology 14:722–728
Pages M, Chaval Y, Herbreteau V, Waengsothorn S, Cosson JF,Hugot JP, et al. (2010) Revisiting the taxonomy of the Rattinitribe: a phylogeny-based delimitation of species boundaries.BioMed Central Evolutionary Biology 10:184–210. doi:10.1186/1471-2148-10-184
Papa A, Johnson AM, Stockton PC, Bowen MD, Spiropoulou CF,Alexiou-Daniel S, et al. (1998) Retrospective serological andgenetic study of the distribution of hantaviruses in Greece.Journal of Medical Virology 55:321–327
Pattamadilok S, Lee BH, Kumperasart S, Yoshimatsu K, OkumuraM, Nakamura I, et al. (2006) Geographical distribution of
hantaviruses in Thailand and potential human health signifi-cance of Thailand virus. American Journal of Tropical Medicineand Hygiene 75:994–1002
Plyusnina A, Ibrahim IN, Plyusnin A (2009) A newly recognizedhantavirus in the Asian house rat (Rattus tanezumi) in Indo-nesia. Journal of General Virology 90:205–209. doi:10.1099/vir.0.006155-0
Plyusnina A, Ibrahim IN, Winoto I, Porter KR, Gotama IB,Lundkvist A, et al. (2004) Identification of Seoul hantavirus inRattus norvegicus in Indonesia. Scandinavian Journal of Infec-tious Diseases 36:356–359
Posada D (2008) jModelTest: phylogenetic model averaging.Molecular Biology and Evolution 25:1253–1256
Ramsden C, Holmes EC, Charleston MA (2009) Hantavirusevolution in relation to its rodent and insectivore hosts: noevidence for codivergence. Molecular Biology and Evolution26:143–153. doi:10.1093/molbev/msn234
Reynes JM, Soares JL, Hue T, Bouloy M, Sun S, Kruy SL, et al.(2003) Evidence of the presence of Seoul virus in Cambodia.Microbes and Infection 5:769–773
Schilling S, Emmerich P, Klempa B, Auste B, Schnaith E, SchmitzH, et al. (2007) Hantavirus disease outbreak in Germany: lim-itations of routine serological diagnostics and clustering of virussequences of human and rodent origin. Journal of ClinicalMicrobiology 45:3008–3014 . doi:10.1128/JCM.02573-06
Schlegel M, Klempa B, Auste B, Bemmann M, Schmidt-Chanasit J,Buchner T, et al. (2009) Dobrava-Belgrade virus spilloverinfections, Germany. Emerging Infectious Diseases 15:2017–2020
Schmidt-Chanasit J, Essbauer S, Petraityte R, Yoshimatsu K,Tackmann K, Conraths FJ, et al. (2010) Extensive host sharingof central European Tula virus. Journal of Virology 84:459–474.doi:10.1128/JVI.01226-09
Schultze D, Lundkvist A, Blauenstein U, Heyman P (2002) Tulavirus infection associated with fever and exanthema after a wildrodent bite. European Journal of Clinical Microbiology andInfectious Disease 21:304–306
Song JW, Kang HJ, Song KJ, Truong TT, Bennett SN, Arai S, et al.(2007) Newfound hantavirus in Chinese mole shrew, Vietnam.Emerging Infectious Diseases 13:1784–1787
Song G, Hang CS, Liao HX, Fu JL, Gao GZ, Qiu HL, et al. (1984)Antigenic difference between viral strains causing classical andmild types of epidemic hemorrhagic fever with renal syndromein China. Journal of Infectious Diseases 150:889–894
Suputthamongkol Y, Nitatpattana N, Chayakulkeeree M, Palab-odeewat S, Yoksan S, Gonzalez JP (2005) Hantavirus infectionin Thailand: first clinical case report. Southeast Asian Journal ofTropical Medicine and Public Health 36:700–703
Suzan G, Marce E, Giermakowski JT, Mills JN, Ceballos G, OstfeldRS, et al. (2009) Experimental evidence for reduced rodentdiversity causing increased hantavirus prevalence. Public Libraryof Science One 4:e5461. doi:10.1371/journal.pone.0005461
Tamura K, Dudley J, Nei M, Kumar S (2007) MEGA4: molecularevolutionary genetic analysis (MEGA) software version 4.0.Molecular Biology and Evolution 24:1596–1599
Terajima M, Vapalahti O, Van Epps HL, Vaheri A, Ennis FA(2004) Immune responses to Puumala virus infection and thepathogenesis of nephropathia epidemica. Microbes and Infection6:238–245
Truong TT, Yoshimatsu K, Araki K, Lee BH, Nakamura I, Endo R,et al. (2009) Molecular epidemiological and serological studiesof hantavirus infection in northern Vietnam. Journal of Veteri-nary Medical Science 71:1357–1363
Hantaviruses in Southeast Asian Rodents
Author's personal copy

Vapalahti O, Lundkvist A, Fedorov V, Conroy CJ, Hirvonen S,Plyusnina A, et al. (1999) Isolation and characterization of ahantavirus from Lemmus sibiricus: evidence for host switchduring hantavirus evolution. Journal of Virology 73:5586–5592
Wang H, Yoshimatsu K, Ebihara H, Ogino M, Araki K, Kariwa H,et al. (2000) Genetic diversities of hantaviruses isolated in Chinaand characterization of novel hantaviruses isolated from Nivi-venter fulvescens and Rattus rattus. Virology 278:332–345
Wong TW, Chan YC, Joo YG, Lee HW, Lee PW, Yanagihara R(1989) Hantavirus infections in humans and commensal ro-dents in Singapore. Transactions of the Royal Society of TropicalMedicine and Hygiene 83:248–251
Xiao SY, Leduc JW, Chu YK, Schmaljohn CS (1994) Phylogeneticanalyses of virus isolates in the genus Hantavirus, family Bun-yaviridae. Virology 198:205–217
Yao ZH, Dong GM, Yu YX, Zhang JK, Yan DY, Liu XC, Zhang LL(2002) A new subtype of hantavirus SN7 isolated from Nivi-venter confucianus in Sichuan province, China. Chinese Journalof Infectious Diseases 20:79–82 (in Chinese)
Zou Y, Hu J, Wang ZX, Wang DM, Yu C, Zhou JZ, et al. (2008)Genetic characterization of hantaviruses isolated from Guizhou,China: evidence for spillover and reassortment in nature. Jour-nal of Medical Virology 80:1033–1041. doi:10.1002/jmv.21149
Kim Blasdell et al.
Author's personal copy






























