Sex differences in the physiology of eating Lori Asarian 1 and Nori Geary 2 1 Institute of Veterinary Physiology and Center for Integrated Human Physiology, University of Zurich, Zurich, Switzerland; and 2 Schwerzenbach, Switzerland Submitted 27 September 2012; accepted in final form 5 July 2013 Asarian L, Geary N. Sex differences in the physiology of eating. Am J Physiol Regul Integr Comp Physiol 305: R1215–R1267, 2013. First published July 31, 2013; doi:10.1152/ajpregu.00446.2012.—Hypothalamic-pituitary-gonadal (HPG) axis function fundamentally affects the physiology of eating. We review sex differences in the physiological and pathophysiological controls of amounts eaten in rats, mice, monkeys, and humans. These controls result from interactions among genetic effects, organizational effects of reproductive hormones (i.e., permanent early developmental effects), and activational effects of these hormones (i.e., effects dependent on hormone levels). Male-female sex differences in the physi- ology of eating involve both organizational and activational effects of androgens and estrogens. An activational effect of estrogens decreases eating 1) during the periovulatory period of the ovarian cycle in rats, mice, monkeys, and women and 2) tonically between puberty and reproductive senescence or ovariectomy in rats and monkeys, sometimes in mice, and possibly in women. Estrogens acting on estrogen receptor- (ER) in the caudal medial nucleus of the solitary tract appear to mediate these effects in rats. Androgens, prolactin, and other reproductive hormones also affect eating in rats. Sex differences in eating are mediated by alterations in orosensory capacity and hedonics, gastric mechanoreception, ghrelin, CCK, glucagon-like peptide-1 (GLP-1), glucagon, insulin, amylin, apolipoprotein A-IV, fatty-acid oxidation, and leptin. The control of eating by central neurochem- ical signaling via serotonin, MSH, neuropeptide Y, Agouti-related peptide (AgRP), melanin-concentrating hormone, and dopamine is modulated by HPG function. Finally, sex differences in the physiology of eating may contribute to human obesity, anorexia nervosa, and binge eating. The variety and physiological impor- tance of what has been learned so far warrant intensifying basic, translational, and clinical research on sex differences in eating. neuroendocrinology; estrogens; testosterone; eating disorders; obesity SEX DIFFERENCES ARE PERVASIVE in physiology and medicine (51, 64, 73, 109, 110, 466, 797, 826). The controls of eating and energy homeostasis are no exceptions. It was observed approximately 100 years ago that removal of the ovaries leads to marked accretion of adipose tissue in rats (697), that daily food intake expressed as kilocalories per gram body weight differs between male and female rats (778), and that food intake varies regularly through the ovarian cycle in intact female rats (674, 779). Sex differences in eating have been the subject of physiological research ever since. The clinical relevance of this work is increas- ingly evident. In the United States, women are approximately threefold more vulnerable than men to psychiatric eating disorders (346, 351) and approximately twofold more vulnerable to severe and morbid obesity (BMI 35 and 40 kg/m 2 , respectively, mass/height 2 ) (226). Women also appear to suffer more from these disorders in terms of physical and psychological functioning and quality of life (24, 84, 273, 292, 465, 531, 762). The increased obesity burden suffered by women is reflected in the fact that 80% of bariatric surgery patients in the United States are women (568, 630). Obesity also decreases fertility and increases the risks of miscarriage and serious health problems for mother and child during pregnancy and after birth (357). In short, eating and weight management are special challenges for women’s health. In light of this, our goals are to critically review present understanding of sex differences in the physiology of eating, to identify important gaps in current knowledge, and to highlight opportunities for basic and translational research. We focus on eating, that is, the controls of the “consummatory” behavior of meal taking and related measures of the total amount consumed. Except for a few instructive examples, we restrict our review to laboratory rats and mice and to anthropoid primates, i.e., mon- keys, apes, and humans (infraorder Simiiformes or Anthropoidea). We consider both male-female sex differences and sex- specific effects, i.e., effects that occur only in one sex, such as effects related to ovarian cycles, pregnancy, and lactation, as well as effects controlled by gonadal steroid hormones. As reflected in our review, there is much more work on females, especially ovarian-cycle effects and estrogen-mediated effects, than on male-female sex differences or androgen-mediated effects. We focus on biological sex differences, but emphasize at the outset that it is impossible to draw sharp lines between purely bio- logical and nonbiological causes of sex differences in behavior (e.g., 48, 250, 797). We consider food choice only in the context of the total amount eaten. Although we review some Address for reprint requests and other correspondence: Nori Geary, Zielacker- strasse 10, 8603 Schwerzenbach, Switzerland (e-mail: [email protected]). Am J Physiol Regul Integr Comp Physiol 305: R1215–R1267, 2013. First published July 31, 2013; doi:10.1152/ajpregu.00446.2012. Review 0363-6119/13 Copyright © 2013 the American Physiological Society http://www.ajpregu.org R1215 by 10.220.32.247 on September 18, 2016 http://ajpregu.physiology.org/ Downloaded from

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Sex differences in the physiology of eating
Lori Asarian1 and Nori Geary2
1Institute of Veterinary Physiology and Center for Integrated Human Physiology, University of Zurich, Zurich, Switzerland;and 2Schwerzenbach, Switzerland
Submitted 27 September 2012; accepted in final form 5 July 2013
Asarian L, Geary N. Sex differences in the physiology of eating. Am J PhysiolRegul Integr Comp Physiol 305: R1215–R1267, 2013. First published July 31,2013; doi:10.1152/ajpregu.00446.2012.—Hypothalamic-pituitary-gonadal (HPG)axis function fundamentally affects the physiology of eating. We review sexdifferences in the physiological and pathophysiological controls of amounts eatenin rats, mice, monkeys, and humans. These controls result from interactions amonggenetic effects, organizational effects of reproductive hormones (i.e., permanentearly developmental effects), and activational effects of these hormones (i.e.,effects dependent on hormone levels). Male-female sex differences in the physi-ology of eating involve both organizational and activational effects of androgensand estrogens. An activational effect of estrogens decreases eating 1) during theperiovulatory period of the ovarian cycle in rats, mice, monkeys, and women and2) tonically between puberty and reproductive senescence or ovariectomy in ratsand monkeys, sometimes in mice, and possibly in women. Estrogens acting onestrogen receptor-� (ER�) in the caudal medial nucleus of the solitary tract appearto mediate these effects in rats. Androgens, prolactin, and other reproductivehormones also affect eating in rats. Sex differences in eating are mediated byalterations in orosensory capacity and hedonics, gastric mechanoreception, ghrelin,CCK, glucagon-like peptide-1 (GLP-1), glucagon, insulin, amylin, apolipoproteinA-IV, fatty-acid oxidation, and leptin. The control of eating by central neurochem-ical signaling via serotonin, MSH, neuropeptide Y, Agouti-related peptide (AgRP),melanin-concentrating hormone, and dopamine is modulated by HPG function.Finally, sex differences in the physiology of eating may contribute to humanobesity, anorexia nervosa, and binge eating. The variety and physiological impor-tance of what has been learned so far warrant intensifying basic, translational, andclinical research on sex differences in eating.
neuroendocrinology; estrogens; testosterone; eating disorders; obesity
SEX DIFFERENCES ARE PERVASIVE in physiology and medicine (51, 64,73, 109, 110, 466, 797, 826). The controls of eating and energyhomeostasis are no exceptions. It was observed approximately100 years ago that removal of the ovaries leads to markedaccretion of adipose tissue in rats (697), that daily food intakeexpressed as kilocalories per gram body weight differs betweenmale and female rats (778), and that food intake varies regularlythrough the ovarian cycle in intact female rats (674, 779). Sexdifferences in eating have been the subject of physiologicalresearch ever since. The clinical relevance of this work is increas-ingly evident. In the United States, women are approximatelythreefold more vulnerable than men to psychiatric eating disorders(346, 351) and approximately twofold more vulnerable to severeand morbid obesity (BMI � 35 and 40 kg/m2, respectively,mass/height2) (226). Women also appear to suffer more fromthese disorders in terms of physical and psychological functioningand quality of life (24, 84, 273, 292, 465, 531, 762). The increasedobesity burden suffered by women is reflected in the fact that�80% of bariatric surgery patients in the United States arewomen (568, 630). Obesity also decreases fertility and increases
the risks of miscarriage and serious health problems for motherand child during pregnancy and after birth (357). In short, eatingand weight management are special challenges for women’shealth. In light of this, our goals are to critically review presentunderstanding of sex differences in the physiology of eating, toidentify important gaps in current knowledge, and to highlightopportunities for basic and translational research. We focus oneating, that is, the controls of the “consummatory” behavior ofmeal taking and related measures of the total amount consumed.Except for a few instructive examples, we restrict our review tolaboratory rats and mice and to anthropoid primates, i.e., mon-keys, apes, and humans (infraorder Simiiformes or Anthropoidea).
We consider both male-female sex differences and sex-specific effects, i.e., effects that occur only in one sex, such as effectsrelated to ovarian cycles, pregnancy, and lactation, as well aseffects controlled by gonadal steroid hormones. As reflected inour review, there is much more work on females, especiallyovarian-cycle effects and estrogen-mediated effects, than onmale-female sex differences or androgen-mediated effects. Wefocus on biological sex differences, but emphasize at the outsetthat it is impossible to draw sharp lines between purely bio-logical and nonbiological causes of sex differences in behavior(e.g., 48, 250, 797). We consider food choice only in thecontext of the total amount eaten. Although we review some
Address for reprint requests and other correspondence: Nori Geary, Zielacker-strasse 10, 8603 Schwerzenbach, Switzerland (e-mail: [email protected]).
Am J Physiol Regul Integr Comp Physiol 305: R1215–R1267, 2013.First published July 31, 2013; doi:10.1152/ajpregu.00446.2012. Review
0363-6119/13 Copyright © 2013 the American Physiological Societyhttp://www.ajpregu.org R1215
by 10.220.32.247 on Septem
ber 18, 2016http://ajpregu.physiology.org/
Dow
nloaded from

subjective phenomena that are closely connected to eating perse, such as ratings of palatability, we do not review the widerange of subjective and behavioral phenomena integral to a fullunderstanding of eating, for example, foraging and other “ap-petitive” behaviors (41), cognitive and social controls of eat-ing, and stress- or immune-related controls.
Neuroendocrine Background
We begin with an overview of hypothalamic-pituitary-go-nadal (HPG) axis function for several reasons. 1) A commonsource of error is the failure to recognize differences in HPGaxis function among women, rats, mice, and other species, apotential problem that is compounded by the ever-increasingunderstanding of HPG axis physiology. 2) Most of the knownHPG mechanisms underlying sex differences in eating involvegonadal steroid hormones. But because the mechanisms ofmany sex differences in eating remain unclear, it would bepremature to assume that other HPG mechanisms are notinvolved. For example, changing levels of estrogens alone mayfully explain changes in eating during the ovarian cycle in miceand rats, but do not do so in women. 3) The many metabolicfeedbacks onto the hypothalamic controls of ovarian cyclingand ovulation (187, 330, 646, 787, 812) suggest it is likely thathypothalamic reproductive physiology also controls eating,although such controls have not yet been identified. 4) Neu-roendocrinology is a vibrant area, and many novel discoveriesand concepts are likely to be relevant to the physiology ofeating. We also discuss in this section some criteria that weused to select physiologically reasonable methods for hormonetreatment and to distinguish apparently aphysiological results.
Gonadal steroid hormones. Estrogens, androgens, and pro-gestins (or progestagens) are groups of gondadal steroid hor-mones, each defined by its biological activity (74). In rats andanthropoid primates, 17�-estradiol (or estradiol) is the mostpotent estrogen and usually circulates in the highest concen-trations (236, 556, 821). For example, rats’ ovaries secrete�5–8-fold more estradiol than estrone (655), and exogenousestradiol inhibited eating �10-fold more potently than exoge-nous estrone in ovariectomized rats (766). Testosterone is theprimary androgen, and progesterone is the primary progestin.Gonadal steroid hormones act on cognate receptors, i.e., estro-gen receptors (ER), progestin receptors (PR), and androgen re-ceptors (AR). Classical steroid receptors are nuclear receptors,although, as described below, the importance of membrane-mounted steroid receptors is increasingly apparent. For example,the principal ER, ER�, and ER�, are expressed in tissue-specificpatterns both in nuclei and on membranes (470, 479).
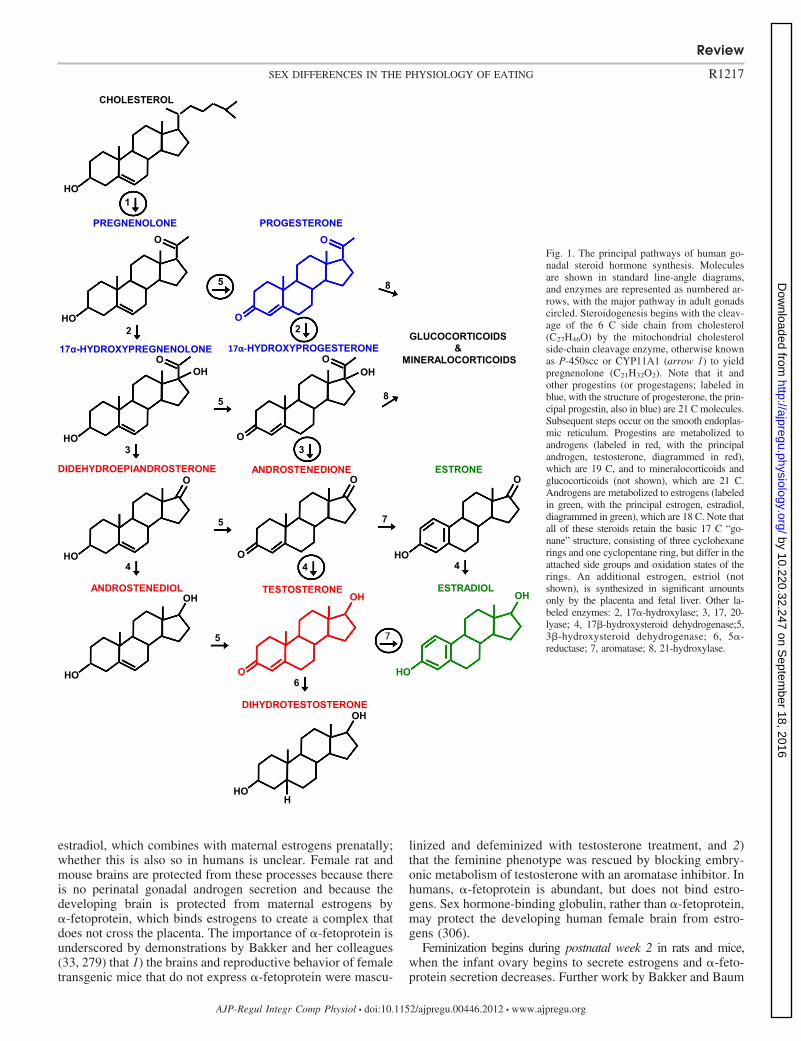
Figure 1 schematizes the principal pathways of humangonadal steroidogenesis (for a detailed review, see Ref. 484).Males and females produce both androgens and estrogens, andboth have biological effects in each sex. The gonads are thesource of almost all circulating androgens in men and ofcirculating estrogens and progestins in premenopausal women,and these molecules act as hormones. In contrast, most circu-lating estrogens and progestins in men, a significant amount ofcirculating androgens in premenopausal women, and all of thegonadal steroids in the plasma of postmenopausal womenderive mainly from other tissues, and these molecules appear tohave their main biological actions in those tissues beforereaching the circulation; i.e., they do not act as classical
hormones (411, 448, 669). This is sometimes referred to asintracrine function. Recognition of its importance has funda-mentally changed endocrinology in recent decades. In thebrain, locally produced steroids are called neurosteroids (240,477, 478, 611). In anthropoid primates, the adrenal glands area major source of precursor molecules for local steroid syn-thesis. In rats and mice, synthesis begins with cholesterol,which cannot pass the blood-brain barrier and is synthesized denovo in the brain, or with gonadal steroid hormones taken upfrom the circulation. For example, in rat hypothalamic neurons,circulating estrogens regulate the expression of 3�-hydroxys-teroid dehydrogenase, which controls the synthesis of proges-terone from cholesterol, and recent studies indicate that it isthis neuroprogesterone, not endocrine progesterone, that initi-ates the LH surge and ovulation (477, 478). There is a reportthat another neurosteroid, 17�-estradiol, may affect eating(104). Finally, 2-hydroxyestradiol and 2-hydroxyestrone arecatabolic products of estrogens that circulate in the blood andmay act in the brain to affect food reward (29) (please seePhysiological Sex Differences in Disordered Eating).
Origins of sex differences in brain and behavior. Biologicalsex differences derive from two evolutionary forces: naturalselection, due to the different biological roles of males andfemales, and sexual selection, due to competition for mates.The proximal causes of biological sex differences comprise acomplex interplay of genetic and endocrine mechanisms (9, 10,32, 271, 425, 466, 471, 671, 807). In most mammals, thedevelopment of phenotypic sex differences is initiated by geneson the X and Y sex chromosomes, with females typicallypossessing the XX karyotype and males, the XY karyotype.There are relatively few genes on the sex chromosomes(�0.15% of human genes are Y-linked and �4.5% are X-linked), and their functions are not yet fully understood. At theblastocyst stage of embryological development, correspondingto 70–100 cells in humans, most cells inactivate one or theother X chromosome, according to a random process. In thesecells, only one of the two alleles of each gene is active. Inhumans, however, 15–25% of genes escapes X inactivation. Akey sex-determination gene is Sry (Sex determining region onY), whose presence causes the undifferentiated fetal gonad todevelop into a testis; in the absence of Sry, autosomal or Xgenes induce ovarian differentiation. De Vries et al. (174)deleted Sry from the Y chromosome and inserted it in into anautosome, thus enabling dissociation of sex-chromosome ef-fects from Sry-determined effects, i.e., mainly effects mediatedby gonadal hormones. Analyses of this “four core-genotype”model indicate that while the majority sex differences inreproductive behaviors and the related brain structures in miceare controlled by Sry via gonadal hormones, a variety of othersex differences are controlled more by sex chromosomes thangonadal steroids (425). Two tests of eating in the four core-genotype model were recently reported (please see Sex differ-ences in eating in rats and mice).
Relatively permanent effects of gonadal steroid hormones inearly development or during puberty are called “organizationaleffects” (8, 551). Early masculinization and defeminization ofmale brains results from the surge in androgen secretion by thetestes that occurs during the end of the embryonic periodthrough the first postnatal day in rats and mice and betweenweek 10 and 20 of pregnancy in humans (652). In mice andrats, these effects of androgens require their aromatization to
Review
R1216 SEX DIFFERENCES IN THE PHYSIOLOGY OF EATING
AJP-Regul Integr Comp Physiol • doi:10.1152/ajpregu.00446.2012 • www.ajpregu.org
by 10.220.32.247 on Septem
ber 18, 2016http://ajpregu.physiology.org/
Dow
nloaded from
estradiol, which combines with maternal estrogens prenatally;whether this is also so in humans is unclear. Female rat andmouse brains are protected from these processes because thereis no perinatal gonadal androgen secretion and because thedeveloping brain is protected from maternal estrogens by�-fetoprotein, which binds estrogens to create a complex thatdoes not cross the placenta. The importance of �-fetoprotein isunderscored by demonstrations by Bakker and her colleagues(33, 279) that 1) the brains and reproductive behavior of femaletransgenic mice that do not express �-fetoprotein were mascu-
linized and defeminized with testosterone treatment, and 2)that the feminine phenotype was rescued by blocking embry-onic metabolism of testosterone with an aromatase inhibitor. Inhumans, �-fetoprotein is abundant, but does not bind estro-gens. Sex hormone-binding globulin, rather than �-fetoprotein,may protect the developing human female brain from estro-gens (306).
Feminization begins during postnatal week 2 in rats and mice,when the infant ovary begins to secrete estrogens and �-feto-protein secretion decreases. Further work by Bakker and Baum
HO
O
17α-HYDROXYPREGNENOLONE
DIDEHYDROEPIANDROSTERONE
HO
PREGNENOLONE
CHOLESTEROL
1
2
HO
OOH
3
HO
O
ANDROSTENEDIOL
4
OH
HO
O
OPROGESTERONE
5
O
O17α-HYDROXYPROGESTERONE
5
OH
O
ANDROSTENEDIONE
5
3
O
O
TESTOSTERONE
5
4
OH
ESTRONEO
4
7
HO
HO
DIHYDROTESTOSTERONE
6
OH
H
OH
7
HO
ESTRADIOL
8
8
GLUCOCORTICOIDS &
MINERALOCORTICOIDS
2
Fig. 1. The principal pathways of human go-nadal steroid hormone synthesis. Moleculesare shown in standard line-angle diagrams,and enzymes are represented as numbered ar-rows, with the major pathway in adult gonadscircled. Steroidogenesis begins with the cleav-age of the 6 C side chain from cholesterol(C27H46O) by the mitochondrial cholesterolside-chain cleavage enzyme, otherwise knownas P-450scc or CYP11A1 (arrow 1) to yieldpregnenolone (C21H32O2). Note that it andother progestins (or progestagens; labeled inblue, with the structure of progesterone, the prin-cipal progestin, also in blue) are 21 C molecules.Subsequent steps occur on the smooth endoplas-mic reticulum. Progestins are metabolized toandrogens (labeled in red, with the principalandrogen, testosterone, diagrammed in red),which are 19 C, and to mineralocorticoids andglucocorticoids (not shown), which are 21 C.Androgens are metabolized to estrogens (labeledin green, with the principal estrogen, estradiol,diagrammed in green), which are 18 C. Note thatall of these steroids retain the basic 17 C “go-nane” structure, consisting of three cyclohexanerings and one cyclopentane ring, but differ in theattached side groups and oxidation states of therings. An additional estrogen, estriol (notshown), is synthesized in significant amountsonly by the placenta and fetal liver. Other la-beled enzymes: 2, 17�-hydroxylase; 3, 17, 20-lyase; 4, 17�-hydroxysteroid dehydrogenase;5,3�-hydroxysteroid dehydrogenase; 6, 5�-reductase; 7, aromatase; 8, 21-hydroxylase.
Review
R1217SEX DIFFERENCES IN THE PHYSIOLOGY OF EATING
AJP-Regul Integr Comp Physiol • doi:10.1152/ajpregu.00446.2012 • www.ajpregu.org
by 10.220.32.247 on Septem
ber 18, 2016http://ajpregu.physiology.org/
Dow
nloaded from

(32) on transgenic mice lacking aromatase clearly demon-strated the active role of estrogens in this process. They found,for example, that female mice lacking aromatase failed todevelop normal female adult reproductive behavior (34) andthat estradiol treatment between postnatal days 15 and 25, butnot before day 15, was sufficient to reinstate normal adultbehaviors (88).
In contrast to the permanent or “organizational” sex-differ-entiating effects of gonadal hormones early in development,effects at other life stages are often reversible and are called“activational effects” (8, 551). These occur only in the pres-ence of the hormones involved and, therefore, wax and waneduring reproductive life. Often activational sex differencesrequire anatomic substrates generated in early development bysexually differentiated organizational processes.
HPG function in adults. The fundamentals of HPG axisfunction in rats (and presumably in mice, but this has not beenas extensively characterized) and humans (understood mainlyfrom studies in monkeys) are similar (236, 556, 821) (134, 289,290). Gonadotropin-releasing hormone (GnRH; formerly calledluteinizing hormone-releasing hormone) is secreted pulsatilelyfrom neurons located in the hypothalamic preoptic area in ratsand mice and in the arcuate nucleus in monkeys and humansinto the hypophyseal-portal circulation. This leads to secretionof follicle-stimulating hormone (or FSH) and luteinizing hor-mone (or LH) from the anterior pituitary into the generalcirculation. These stimulate the secretion of gonadal steroids.GnRH secretion is also regulated by a pulse generator, whichhas a constant period of about 2–3 h in men and 60–90 min inwomen. Both slower and faster frequencies fail to producenormal gonadal steroid levels. Gonadal steroids, LH, and otherHPG-axis hormones provide feedback signals to both thepituitary and hypothalamic levels. Feedback is mainly negativein males and contributes to a relatively constant hormonesecretion in adult males. The resulting plasma levels of testos-terone are �2–3 ng/ml in mice, �1–3 ng/ml in rats, �8–15ng/ml in cynomolgus monkeys, and �3–10 ng/ml in men(520). Both positive and negative feedbacks occur in females,leading to changing hormone secretion through the ovariancycle, as described in the next sections.
SPONTANEOUS OVARIAN CYCLES. HPG axis function and thecontrol of ovulation vary widely across mammalian species.Many are seasonal ovulators (454), some are mating-inducedovulators (364), and a few, including rats, mice and anthropoidprimates, display spontaneous cycles. In this latter group,ovulation occurs in spontaneous rhythms, or ovarian cycles,that occur regularly throughout the year between puberty andreproductive senescence, except during pregnancy and lacta-tion. These are known as estrous cycles in rats, mice andseveral other species and as menstrual cycles in women,monkeys, apes, and a few other species in which the cycle endswith discharge of endometrial tissue.
Ovulation and behavioral estrus occur in 4–5-day cycles inrats and mice. Rats’ sexual receptivity is maximal in the middleof the nocturnal phase of estrus and near zero during diestrus(estrous phases are defined in the next section) (72, 825). Incontrast, women are sexually receptive throughout their ovar-ian cycle, although the degree of receptivity apparently varies(247, 295). There is little or no seasonal variation in reproduc-tive function of mice and rats (421). In addition to reproductivebehaviors, eating, locomotor activity, nest building, fluid in-
take, food hoarding, and other behaviors vary rhythmicallyduring the estrous cycle (217, 224, 251, 727). The maxima ofthese cycles are not all in phase, and a variety of evidenceindicates that they are separately controlled. For example,although facilitation of the copulatory behavior lordosis anddecreased eating both occur during the night of estrus in rats,facilitation of lordosis requires progestins, as well as estrogens(549), but the estrous decrease in eating does not (15, 259,766). Ovarian cycle effects are the most researched sex differ-ences in eating, and we review them in detail.
OVARIAN CYCLE PHASE. Rat and mouse ovarian cycle phasesor days are most frequently categorized by “vaginal cytology”(48, 236) based on Long and Evans’s (440) classical descrip-tion of the associations among ovulation, reproductive-tracthistology, and reproductive behavior, and named as suggestedby Heape (321). Vaginal estrus, marked by the cornification ofvaginal epithelial cells, begins around the LH surge, whichoccurs near dark onset, and ends during the subsequent lightphase. Thus, vaginal cytology is best sampled in the nocturnalphase or early in the diurnal phase, as indicated by the hatchedbar at the bottom of Fig. 2. As mentioned above, behavioralestrus and the estrous decrease in eating are most prominentduring the nocturnal phase after the LH surge. In order to havethis nocturnal phase occur during the nominal day of estrus,cycle days should not begin at the midpoint of the dark phase,i.e., midnight, as ordinary clock-time days do. Beginning cycledays at the midpoint of the dark and using diurnal vaginalcytology to assign day names leads to the unfortunate conse-quence that estrous behaviors occur during the day labeledproestrus. This is a major a source of confusion in across-experiment comparisons. Therefore, here, we begin cycle daysat dark onset and assign names based on early light-periodvaginal cytology (15). The preovulatory phase of the cycleusually lasts 3 days, labeled diestrus 1 (or metestrus), diestrus2 (or diestrus), and proestrus.
Women’s cycle days are numbered either 1) forward fromthe first day of menses, which is the beginning of the follicularphase, and with day 14, the presumed day of ovulation,dividing the follicular and luteal phases, or 2) backward (fol-licular stage) and forward (luteal phase) from the LH peak.Detection of the LH peak by assaying plasma or urinary LH isconsidered the gold standard for ovarian-cycle research (55,342). If LH is measured, the periovulatory phase is the 4 daysaround the LH peak (ovulation usually occurs within 1 day ofthe LH peak).
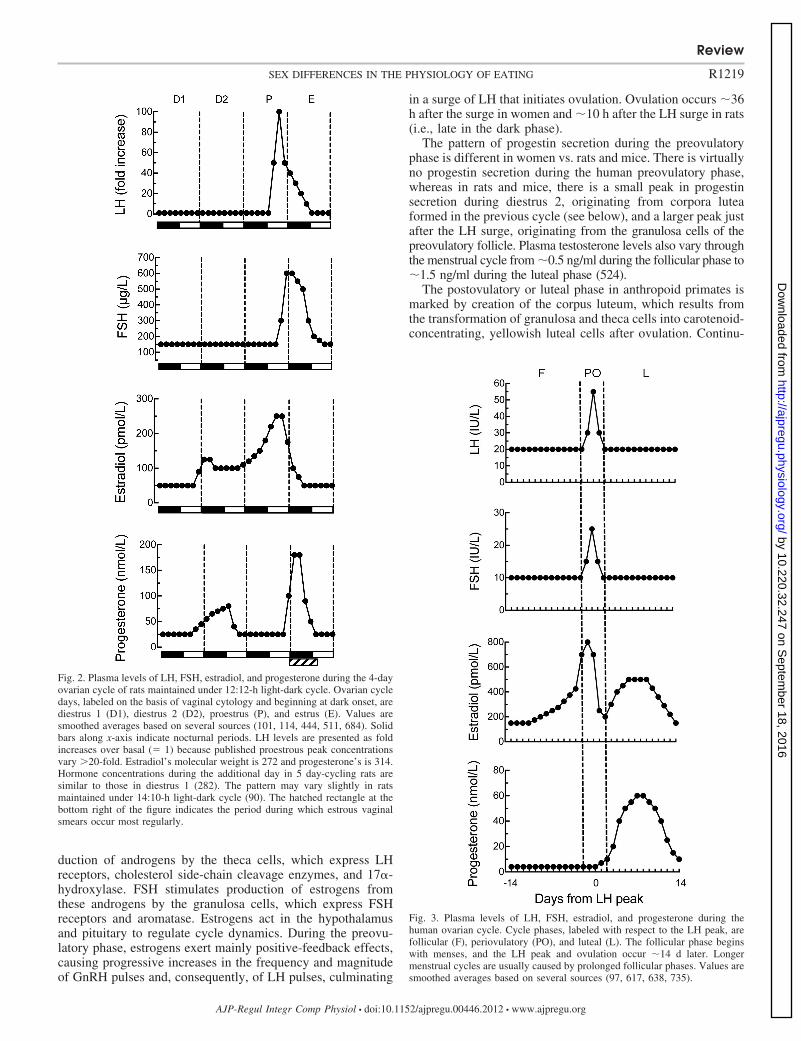
NEUROENDOCRINE CONTROL OF THE OVARIAN CYCLE. The cyclicchanges in LH, FSH, estradiol, and progesterone in 4 day-cycling rats and women are shown in Figs. 2 and 3, respec-tively. Some obvious differences are 1) cycle length is muchlonger in women (�28 days) than in rats and mice (usually 4or 5 days); 2) absolute levels of estradiol are much higher inwomen;3) absolute levels of progesterone are lower in women; and4) the pattern of hormone secretion after ovulation is verydissimilar in humans and rats (discussed below).
The patterns of LH, FSH, and estrogen secretion are similarin women and rats and mice during the preovulatory phase ofthe cycle, i.e., during the follicular phase in women anddiestrus 1 through early estrus in rats and mice. The preovu-latory levels of LH and FSH, although low, are required toincrease follicular production of estrogens. LH stimulates pro-
Review
R1218 SEX DIFFERENCES IN THE PHYSIOLOGY OF EATING
AJP-Regul Integr Comp Physiol • doi:10.1152/ajpregu.00446.2012 • www.ajpregu.org
by 10.220.32.247 on Septem
ber 18, 2016http://ajpregu.physiology.org/
Dow
nloaded from
duction of androgens by the theca cells, which express LHreceptors, cholesterol side-chain cleavage enzymes, and 17�-hydroxylase. FSH stimulates production of estrogens fromthese androgens by the granulosa cells, which express FSHreceptors and aromatase. Estrogens act in the hypothalamusand pituitary to regulate cycle dynamics. During the preovu-latory phase, estrogens exert mainly positive-feedback effects,causing progressive increases in the frequency and magnitudeof GnRH pulses and, consequently, of LH pulses, culminating
in a surge of LH that initiates ovulation. Ovulation occurs �36h after the surge in women and �10 h after the LH surge in rats(i.e., late in the dark phase).
The pattern of progestin secretion during the preovulatoryphase is different in women vs. rats and mice. There is virtuallyno progestin secretion during the human preovulatory phase,whereas in rats and mice, there is a small peak in progestinsecretion during diestrus 2, originating from corpora luteaformed in the previous cycle (see below), and a larger peak justafter the LH surge, originating from the granulosa cells of thepreovulatory follicle. Plasma testosterone levels also vary throughthe menstrual cycle from �0.5 ng/ml during the follicular phase to�1.5 ng/ml during the luteal phase (524).
The postovulatory or luteal phase in anthropoid primates ismarked by creation of the corpus luteum, which results fromthe transformation of granulosa and theca cells into carotenoid-concentrating, yellowish luteal cells after ovulation. Continu-
Fig. 3. Plasma levels of LH, FSH, estradiol, and progesterone during thehuman ovarian cycle. Cycle phases, labeled with respect to the LH peak, arefollicular (F), periovulatory (PO), and luteal (L). The follicular phase beginswith menses, and the LH peak and ovulation occur �14 d later. Longermenstrual cycles are usually caused by prolonged follicular phases. Values aresmoothed averages based on several sources (97, 617, 638, 735).
Fig. 2. Plasma levels of LH, FSH, estradiol, and progesterone during the 4-dayovarian cycle of rats maintained under 12:12-h light-dark cycle. Ovarian cycledays, labeled on the basis of vaginal cytology and beginning at dark onset, arediestrus 1 (D1), diestrus 2 (D2), proestrus (P), and estrus (E). Values aresmoothed averages based on several sources (101, 114, 444, 511, 684). Solidbars along x-axis indicate nocturnal periods. LH levels are presented as foldincreases over basal (� 1) because published proestrous peak concentrationsvary �20-fold. Estradiol’s molecular weight is 272 and progesterone’s is 314.Hormone concentrations during the additional day in 5 day-cycling rats aresimilar to those in diestrus 1 (282). The pattern may vary slightly in ratsmaintained under 14:10-h light-dark cycle (90). The hatched rectangle at thebottom right of the figure indicates the period during which estrous vaginalsmears occur most regularly.
Review
R1219SEX DIFFERENCES IN THE PHYSIOLOGY OF EATING
AJP-Regul Integr Comp Physiol • doi:10.1152/ajpregu.00446.2012 • www.ajpregu.org
by 10.220.32.247 on Septem
ber 18, 2016http://ajpregu.physiology.org/
Dow
nloaded from

ing LH secretion stimulates the corpus luteum to secreteestrogens and progestins; in women, this phase lasts �10–14days. Progestins maintain the corpus luteum and stimulateangiogenesis and hypertrophy of the uterine endometrium (thedecidual response). Estradiol levels are higher during most ofthe luteal phase than during the follicular phase, althoughlower than during the periovulatory phase. The human lutealphase ends with degeneration of the corpus luteum, sheddingof the uterine decidua, and menstruation. The drop in secretionof progestins, estrogens, and inhibin releases the hypothalamusand pituitary from inhibition and increases GnRH pulse fre-quency, thus increasing LH and FSH secretion and initiatingthe follicular phase of a new cycle. If pregnancy occurs,secretion of chorionic gonadotropin maintains the corpus lu-teum.
Rats and mice do not provide an adequate model of thehuman luteal phase. Unlike women, rats and mice lack aprolonged postovulatory phase during which functional cor-pora lutea maintain high plasma estrogen and progestin levels(236, 313, 314). First, estrogen levels are basal by the time ofovulation and do not increase again until the next cycle.Second, rat corpora lutea begin to develop not after ovulation,as in women, but one or two cycles earlier; thus, two or threegenerations of corporal lutea are present simultaneously. Thesereach their maximal size and secretory potential during theirfinal diestrus, which explains the small peak in progestinsecretion during diestrus 2, and thereafter begin to degenerate.Another factor that decreases plasma progesterone levels anddeciduation is that rat corpora lutea express 20�-dihydroxys-teroid dehydrogenase, which converts progesterone to 20�-hydroxyprogesterone, a less biologically active progestin. Asecond, larger peak in progesterone occurs simultaneously withthe LH surge and ends near ovulation. As already noted, thisprogesterone originates in the preovulatory follicles, not thecorpora lutea. By a few hours after ovulation, progesteronelevels are no longer sufficient to maintain the decidual re-sponse. Follicular estrogen secretion and the next cycle beginduring the light phase after ovulation, after only 6–8 h ofrecovery of HPG axis and reproductive tract.
Stimulation of the rat uterine cervix during mating causessecretion of prolactin from the anterior pituitary, which main-tains the corpora lutea for about the duration of the humanluteal phase after nonfertile mating (“pseudopregnancy”) andthroughout pregnancy (usually 21 days) after fertile mating.Pseudopregnancy also occurs spontaneously. Despite the main-tenance of the corpora lutea and the similar durations of ratpseudopregnancy and the luteal phase, the endocrine profilesare different. Plasma levels of progesterone increase duringpseudopregnancy as in the luteal phase, but plasma estradiollevels are minimal, as in normal pregnancy (249, 728). Inad-vertent induction of pseudopregnancy can hamper studies ofintact cycling rats.
Kisspeptin and gonadatropin-inhibitory peptide. The re-cently discovered hypothalamic peptides kisspeptin (268, 283,377, 552) and gonadatropin-inhibitory peptide (GnIH) or RF-amide-related peptide-3 (127, 407, 745) play important roles inHPG axis function. In mice and rats, kisspeptin is expressedmost densely in the preoptic area, anteroventral periventricularnucleus (AVPV), and arcuate nucleus (the latter is often calledthe infundibular nucleus in humans). GnRH neurons are amajor target of kisspeptinergic fibers. Kisspeptin is vital for
pubertal development. In mice or humans lacking the kisspep-tin receptor Kiss1R, GnRH secretion is insufficient to supportnormal pubertal development (hypogonadotropic hypogonad-ism). An elegant recent study by Lomniczi et al. (439) indicatesthat the timing of puberty in female rats depends upon epige-netic silencing of two transcription-repressor genes that sup-press kisspeptin expression.
After puberty, kisspeptin is involved in the feedback controlof gonadal steroids on GnRH secretion. In both sexes, gonad-ectomy leads to increased kisspeptin mRNA in the arcuatenucleus and decreased kisspeptin mRNA in the AVPV, sug-gesting negative and positive feedbacks, respectively (in fe-males, negative feedback predominates in the early and mid-follicular phases, and positive feedback predominates in thelate follicular and preovulatory phases; in males the twoinfluences apparently are in tonic balance). Rometo et al. (610)showed that the estrogenic feedback effect also occurs inwomen by demonstrating 1) that the increases in GnRH secre-tion that occurs in postmenopausal women and in ovariecto-mized cynomolgus macaques (Macaca fascicularis) were as-sociated with increased kisspeptin expression in the arcuatenucleus, and 2) that in ovariectomized monkeys estradioltreatment reversed both effects. In another interesting study(682), kisspeptin mRNA expression in the preoptic area andcaudal arcuate nucleus and GnIH mRNA expression in thedorsal medial and paraventricular nuclei were higher in thefollicular than the luteal phases of rhesus macaques (Macacamulatta).
Estrogen signaling via ER� mediates kisspeptin function ina complex fashion. For example, conditional knockout of ER�in kisspeptin neurons both advances the onset of puberty andretards subsequent pubertal development in female mice (464).A recent study by Frazao et al. suggests that these disparateeffects may be due in part to the different effects of estradiolacting via ER� on the electrophysiological activity of AVPVvs. arcuate kisspeptin neurons (233).
The synchrony of rat and mouse ovarian cycles with thecircadian pacemaker, which results in cycle periods that areeven multiples of days and which times the LH surge to occurat dusk, depends on kisspeptin neurons in the AVPV (236, 377,793). In rats, estrogens stimulate these kisspeptin neurons,which, in turn, drive GnRH neurons in the preoptic area toproduce the LH surge and ovulation; in contrast, in women,estrogenic positive feedback appears to be mediated by kiss-peptin neurons in the arcuate nucleus that project to themediobasal hypothalamus, with preoptic area and circadianinputs not required (236, 793).
GnIH is synthesized by neurons in the dorsomedial nucleusof the hypothalamus in rats and mice (127, 407, 408, 745, 747).In one study, GnIH was found in the arcuate nucleus in womenand female cynomolgus macaques (610); in another study, itwas found in the intermediate periventricular (which is adja-cent to the dorsomedial nucleus) and paraventricular nuclei infemale rhesus macaques (746); and in a third study, it was foundin the arcuate, dorsomedial and intermediate periventricular nuclei offemale rhesus macaques (682). Interestingly, in the latter study,kisspeptin mRNA expression in the preoptic area and arcuatenucleus and GnIH mRNA expression in the dorsomedial andparaventricular nuclei were higher in the follicular phase thanin the luteal phase (682). GnIH fibers project to similar areas asthe GnRH neurons, as well as several other brain areas (127,
Review
R1220 SEX DIFFERENCES IN THE PHYSIOLOGY OF EATING
AJP-Regul Integr Comp Physiol • doi:10.1152/ajpregu.00446.2012 • www.ajpregu.org
by 10.220.32.247 on Septem
ber 18, 2016http://ajpregu.physiology.org/
Dow
nloaded from
407, 745). GnIH is thought to be involved in the negativefeedback effects of estrogens early in the follicular phase andto sharpen the control of LH and FSH secretion by acting as afunctional antagonist to GnRH.
Kisspeptin and GnIH both appear to affect eating. Centraladministration of kisspeptin inhibited eating in male mice(692), and central administration of GnIH stimulated eating inmale mice, rats, and cynomolgus monkeys (127, 352, 498,682). In addition, in female rats, knockdown of Arc kisspeptinneurons with saporin conjugated to the neurokinin-3 receptor,which the Arc kisspeptin neurons also express, reduced theeffect of ovariectomy to increase body weight; unfortunatelyfood intake was not measured (487). Fu and van der Pol (243)described a potential mechanism for the effects of kisspeptinand GnIH on eating. They found that kisspeptin fibers makeexcitatory synapses on arcuate nucleus neurons that expressproopiomelanocortin, the precursor of the eating-inhibitoryneuropeptide �-melanocortin-stimulating hormone (�-MSH),and indirectly inhibit arcuate neurons that express neuropeptideY (NPY), which stimulates eating (sex differences in theeffects of �-MSH and NPY are described below). Furthermore,GnIH inhibited the proopiomelanocortin neurons and reducedthe effect of kisspeptin. Clearly, more work directed at ana-lyzing sex differences in the eating effects of kisspeptin andGnIH is called for.
Reproductive senescence. The loss of fertility in aging fe-males has different causes in rats vs. anthropoid primates. Inrats, the repeated exposure of the brain to estrogens throughreproductive life leads to progressive degeneration and unre-sponsiveness of the arcuate nucleus, such that cycling endswell before the ovaries are depleted of ova (347). Older femalerats retain potentially viable ovaries in a state of arrestedfollicular development for months, during which time theydisplay vaginal estrus. In contrast to rats, although agingaffects numerous facets of HPG function in women, cycling ismaintained until the ovaries are depleted (98, 189, 235, 303,420, 508, 638). Thus, from the perspective of hypothalamic-pituitary function, young ovariectomized rats provide a bettermodel of human menopause than do older, reproductivelysenescent rats.
Menopause occurs between 45 and 55 yr of age in �95% ofwomen (276, 327, 629, 730). The menopausal transition ismarked by increasing cycle-length variability, steady increasesin basal FSH, and steady decreases in inhibin and anti-Mülle-rian hormone. Indeed, plasma levels of anti-Müllerian hormonein women 20–50 yr of age predicted the onset of menopausewith an mean error of only 6 mo (730). In contrast, averageplasma estradiol levels do not change much before menopause.Rather, they remain similar to those in younger women untilmenopause and then decrease over about a year to a fraction ofthe premenopausal basal level (98, 99, 303). Postmenopausalplasma estrogens derive mainly from the adipose tissue, andtheir levels are associated with adiposity (361). Obesity is alsoassociated with slightly later age of menopause [i.e., a mediandelay of �1 yr in overweight and obese women in a recentcareful study (492)]. The mechanisms for this are not under-stood (28, 424, 492). Aging also affects HPG function in men,with the result that bioavailable testosterone levels decrease by�2%/yr after age 40 (220, 308).
Hormone treatment regimens. Gonadectomy and hormonereplacement are classic endocrine methods. Because endoge-
nous testosterone levels are relatively constant, mimickingthem is simple. Endogenous testosterone levels in adult ratsvary according to age, strain, etc., from �1–7 ng/ml, andnear-physiological replacement is usually achieved with con-stant-release silicone-capsule implants (117, 118, 659). Be-cause reproductive hormone levels cycle in female rats, how-ever, constant-release pellets cannot be considered physiolog-ical. Indeed, daily or continuous peripheral administration oflow estradiol doses or even single high doses of long-lastingestrogens such as estradiol valerate can disrupt ovarian cycling,induce pseudopregnancy, elicit progressive, aphysiological changesin several brain neurochemical receptor systems and in behav-ior, and greatly accelerate the degeneration of the arcuatenucleus (83, 179, 347, 467, 468, 528, 529, 647). In contrast, aweekly cyclic estradiol injection regimen in which 10 �gestradiol benzoate was injected on Tuesdays and Wednesdaysand progesterone priming and sexual-receptivity tests weredone on Fridays led to stable, normal levels of progestin andoxytocin receptors and in sexual receptivity (647). Plasmaestradiol levels in rats maintained on this regimen, however,increased to more than 4 times the proestrous maximum forseveral days and never decreased below the proestrous maxi-mum (804). Reducing the two estradiol benzoate doses to 2 �gled to a more normal magnitudes of plasma estradiol, but stillmaintained the proestrous level for an abnormal duration anddid not fully reproduce normal patterns of food intake (255). Incontrast to these weekly schedules, subcutaneous injection of 2�g estradiol benzoate once each 4th day produced near-phys-iological 4-day cycles of plasma estradiol concentration (15,476) (Fig. 4) and led to normal spontaneous meal patterns, foodintake, and body weight (15). Note that because of the rapidrates of esterification of estradiol benzoate in the plasma andclearance of estradiol from the plasma in rats (417, 724), thedurations of the estradiol increases in these studies are duemainly to the slow entry of subcutaneously injected estradiolinto the circulation.
Doses more than 20 �g estradiol consistently elicit signs ofaversion in female rats, such as abnormal latency and durationof the eating-inhibitory effect, abnormal orofacial expressions,and the formation of conditioned taste aversions (253, 254,333). We do not consider estradiol’s aversive effects to be
Fig. 4. Plasma estradiol concentration during chronic, cyclic estradiol treat-ment. Plasma samples were taken in the 9th cycle of subcutaneous injection of2 �g estradiol benzoate (EB) once each 4th day, at the middle of the light phaseof day 2 (D2, arrow), which models diestrus 2 in intact rats. D4 of the treatmentregimen modeled estrus based on maximally decreased eating behavior andincreased sexual receptivity in progesterone-primed rats. Values below thedetection threshold of our radioimmunoassay (30 pmol/l) are shown as 30pmol/l. Reprinted from Hormones and Behavior, Cyclic estradiol treatment nor-malizes body weight and restores physiological patterns of spontaneous feedingand sexual receptivity in ovariectomized rats, 42: 461–471, 2002; republished withpermission from Elsevier; from Asarian and Geary (15).
Review
R1221SEX DIFFERENCES IN THE PHYSIOLOGY OF EATING
AJP-Regul Integr Comp Physiol • doi:10.1152/ajpregu.00446.2012 • www.ajpregu.org
by 10.220.32.247 on Septem
ber 18, 2016http://ajpregu.physiology.org/
Dow
nloaded from

useful in the analysis of its physiological effects on eating anddo not consider high-dose studies here.
Subcutaneous injection of 0.5 mg/rat progesterone or 0.5–2mg/100 g body wt progesterone increased plasma progesteroneconcentration to about the estrous maximum, although the timecourse was prolonged compared with estrus (3). Less informa-tion is available concerning appropriate doses for mice or forother HPG hormones. Becker et al. (48) provide an excellentdiscussion of technical and interpretational issues surroundingperipheral gonadal steroid hormone treatment.
Dose is also crucial in interpreting central steroid hormonetreatments. Implants of �3 ng 3H-labeled estradiol, preparedby filling the distal 1 mm of 28-gauge cannulas with 1:300estradiol:cholesterol mixtures, into the ventromedial hypotha-lamic area (VMH) produced measurable label within only�500 �m of the implant site and, in combination with periph-eral progesterone injections, were sufficient to increase sexualreceptivity in ovariectomized rats (168, 169). A formal map-ping study for the inhibition of eating has not been done. Testsperformed by Butera and colleagues (103, 108) of intra-hypothalamic implants of �100 ng estradiol in the distal 1 mmof 28-gauge cannulas indicated that estradiol spread �1 mm inamounts sufficient to inhibit eating and did not produce pe-ripheral estrogenic effects, such as increased uterine weight orcornification of vaginal epithelial cells. Intrahypothalamic im-plants of more concentrated estradiol mixtures, both in thestudy by Butera and Beikirch (103) and many earlier studies,were sufficient to produce peripheral effects. Because thethreshold peripheral estradiol dose for the inhibition of eatingseems to be less than that for cornification of vaginal epithelialcells (190), such large doses clearly cannot be used to identifylocal effects. We (732) demonstrated that doses of �200 ng3H-labeled estradiol applied to the surface of the dorsal hind-brain just posterior to the area postrema in 1-mm2 pieces ofabsorbable surgical fabric produced measurable label only�200 �m caudally, �600 �m rostrally, and �500 �m ven-trally and did not lead to detectable amounts of estradiol in theplasma.
Latency of estrogen’s effect on eating. A frequent source ofconfusion is that in rats and mice, most estrogen-dependentresponses occur during the nocturnal phase of estrus, whenplasma estrogen levels are low, not high (please see Fig. 2).This timing probably reflects the dependence of the behaviorson transcriptional effects of estrogens, whose downstreamconsequences require hours or days to complete. This reason-ing suggests that events during the ovarian cycle that dependon gene expression are likely to be due to the increases inplasma estrogens during diestrus, i.e., 1–2 days prior to estrus,and not to the peak of estrogen concentration during proestrus.This has been shown to be the case both for the LH surge (236)and for lordosis, a reflexive proceptive behavior characteristicof estrus that depends on increased expression of progestinreceptors (549). For example, acute antagonism of estrogenicfunction during diestrus blocked the proestrus surge of LH andovulation, whereas the same treatment early in proestrus hadno effect (221, 510).
The estrogenic inhibition of eating in ovariectomized miceand rats has a latency that suggests a similar interpretation.Physiological or modest pharmacological peripheral doses ofestradiol in a lipid vehicle, such as sesame oil, decrease eating�24–48 h later in mice and rats, depending on the circadian
time of administration (255, 287, 637, 731). Central adminis-tration of estradiol inhibited eating with a similar latency in rats(732). These data suggest that endogenous estrogens normallyact in diestrus to initiate effects that result in reduced eatingduring estrus. We consider the typical �24–48 h latency of theestrogenic inhibition of eating to be a useful criterion for thephysiological relevance of estrogenic treatments in rats andmice. That is, if an estrogen or estrogen agonist decreaseseating in �24 h in mice or rats, it is unlikely to mimic thephysiological action of endogenous estrogens (please see Refs.203, 286, 333, 731 and Site of ER Controlling Eating forfurther discussion). Unfortunately, we know of no data on thetime course of any estrogenic effect on eating or on HPG axisfunction in monkeys, apes, or women.
Sex Differences in Eating in Rats and Mice
Male-female differences. Total daily energy intake in malerats exceeds that in females to an extent greater than predictedby their larger lean body mass and metabolic rate (790, 803).Normal “homeostatic” eating also contributes to the mainte-nance of significantly less body fat content in male than femalerats (129). As described below, both organizational and acti-vational effects of estrogens and androgens appear to contrib-ute to these differences. There may be a species difference inhow males’ greater intake is expressed in spontaneous mealpatterns: the greater total food intake of male than femaleLong-Evans rats maintained on a palatable liquid diet resultedmainly from larger meals (457), whereas the greater foodintake of similarly maintained male than female C57BL/6Jmice resulted entirely from more frequent meals (701).
Activational effects of estrogens and androgens contribute tothe maintenance of normal levels of food intake in rats, but doso in opposite ways. With few exceptions, ovariectomy in-creases rats’ daily food intake and body weight by increasingmeal size, and estradiol treatment normalizes all three mea-sures; in contrast, orchiectomy decreases daily food intake andbody weight by decreasing meal frequency, and testosteronetreatment normalizes them (15, 18, 77, 102, 115, 191, 202, 203,267, 726, 764, 776). As we review below, the estrogeniccontrol of eating in rats is the best understood of these phe-nomena. There are many species differences in the effects ofgonadectomy on eating and weight. For example, as discussedbelow, ovariectomy often fails to elicit overeating in mice. Inaddition, in many species orchiectomy increases food intakeand adiposity (341). This may be the case for monkeys andhumans, as we also discuss below.
There is an interesting male-female sex difference in regu-latory or homeostatic eating. Male mice that were acutelyfood-deprived for 24 h (513), chronically food-restricted untilthey lost about 15% body weight (660), or underwent partiallipectomy (660) all compensated by overeating, whereas sim-ilarly challenged female mice compensated by decreasing en-ergy expenditure without overeating. A similar sex differencein postdeprivation eating occurred in both rats (751) andhumans (820). The developmental origins of this sex differenceare reviewed in the next section; whether activational effectsalso contribute is unknown.
There is also a sex difference in conditioned taste aversionlearning in rats. In several tests, males and females acquiredtaste aversions to unconditioned stimuli such as LiCl similarly,
Review
R1222 SEX DIFFERENCES IN THE PHYSIOLOGY OF EATING
AJP-Regul Integr Comp Physiol • doi:10.1152/ajpregu.00446.2012 • www.ajpregu.org
by 10.220.32.247 on Septem
ber 18, 2016http://ajpregu.physiology.org/
Dow
nloaded from

but females’ taste aversions extinguished faster after acquisi-tion, i.e., began to ingest the conditioned stimulus, typically, asweet solution, in normal amounts sooner when it was pre-sented repeatedly in the absence of the unconditioned stimulus(160). Activational effects of both estrogens and androgensappear to contribute to this sex difference (116, 819). Thesefindings merit further research because conditioned taste aver-sions are probably important in the control of eating in humans,especially in certain clinical populations, for example patientsundergoing radiation or chemotherapy and patients with buli-mia nervosa (65, 86, 641).
Development. Work begun in the 1970s by Wade andcolleagues (266, 764) and others (56, 507, 452) demonstratedthat neonatal masculinization of female rat pups increased theirfood intake and decreased their sensitivity to the eating-inhib-itory effects of estrogens as adults. The latter effect suggeststhat, as is the case for numerous sexually differentiated brainfunctions, activational effects of estrogens in adults requireorganizational programming of the developing neural sub-strate. Nohara et al. (513) recently discovered some of thissubstrate. They found that female mice that were masculinizedwith neonatal testosterone treatment ate like intact males in that1) they ate more than intact females at 6 wk of age, aspreviously described, and 2), unlike intact females, they in-creased eating following a 24-h fast when tested as adults.Nohara et al. (513) also identified two changes in the physiol-ogy of hypothalamic proopiomelanocortin (POMC) circuitsthat may underlie the sex differences in eating (POMC is theprecursor of the neurotransmitters �- and �-melanocyte-stim-ulating hormone, which are involved in energy homeostasis;please see Sex Differences in Central Controls of Eating). Thatis, both hypothalamic expression of the Pomc gene and thearborization of hypothalamic POMC neurons were reduced inneonatally masculinized females from the intact-female to theintact-male level. Comparison of neonatal treatment with es-tradiol and 5�-dihydrotestosterone, which cannot be convertedto estradiol, verified that these effects were AR-mediated.Masculinization also led to hyperleptinemia and reduced thesensitivity of exogenous leptin to upregulate POMC, decreaseeating and prevent adipose-tissue mass accumulation. Theseeffects were estrogen dependent. The changes in plasma leptinconcentrations and leptin sensitivity, however, lay outside thenormal range, suggesting that the neonatal manipulations werenot entirely physiological.
Chen et al. (122, 123) reported the first measurements ofeating in the four core-genotype model of HPG axis develop-ment (described in The origins of sex differences in brain andbehavior). They found that in C57BL/6J mice, both gonadalsex (testes or ovaries) and chromosomal sex (XX or XY)affected eating: 1) gonadal females (i.e., without Sry) ate morethan gonadal males (with Sry) during the dark regardless oftheir chromosomal sex (XX or XY) (Fig. 5), and 2) gonadaland chromosomal females (XX with Sry) ate more than allother groups during the light and had an approximately twofoldmore fat mass (not shown). Estrogen and androgen treatmentswere not tested. Chen et al.’s (122, 123) and Nohara et al.’s (513)elegant studies, together with recent translational work on orga-nizational influences on eating disorders (please see PhysiologicalSex Differences in Disordered Eating), should rekindle interest inthe development of sex differences in eating (290).
Puberty involves changes in the secretion of HPG and otherhormones, tissue sensitivity, and neuronal architecture (670,671). Pubertal brain maturation appears to be necessary for theestrogenic inhibition of eating: 1) exogenous estradiol inhibitedeating only in postpubertal rats (649, 765, 771), and 2) theamplification of endogenous cholecystokinin satiation by ex-ogenous estradiol began at puberty (please see Cholecysto-kinin). In contrast, testosterone treatment did increase eating inprepubertal male rats (518), indicating that the maturation ofthe neural substrate for HPG-control of eating is sex-specific.
An organizational effect of estrogens also appears to benecessary for the stimulation of eating by prolactin treatmentthat occurs in adult female, but not male, rats (323). That is,prolactin stimulated eating in adult males whose brains werefeminized by castration on postnatal day 1, but not in adultfemales masculinized by neonatal testosterone treatment. Thisorganizational effect may be required for the increased eatingthat occurs in pregnancy and lactation described below.
Ovarian cycle. The initial reports that rats eat least duringthe periovulatory (estrous) phase of the ovarian cycle and mostduring diestrus have been replicated countless times in rats (forreviews, see Refs. 18 and 764) and extended to mice (527, 547,731), humans, and many other species. The estrous minimumin daily food intake is typically �20% less than the diestrousmaximum. Rarely, food intake did not vary across the cycle.For example, Varma et al. (758) reported that Fischer 344 rats,a small, lean strain, did not eat less during estrus, and Petersen(547) reported that mice fed a honey-laced wheat-cereal dietate more during estrus, although chow-fed fed mice ate less.
Importantly, the estrous decrease in eating in rats is duesolely to a decrease in the size of spontaneous meals, with nocontribution from a decrease in meal frequency (15, 77, 190,207, 527, 547). Indeed, meal frequency usually increases duringestrus [meal size was also reduced in the Fischer 344 ratsmentioned above, but this was fully compensated for by theincrease in meal frequency (758)]. These and data reviewed inthe next section suggest that these two parameters of spontaneousfeeding are controlled separately. The proximal physiologicalmechanisms for the estrous decrease in eating are discussed
Fig. 5. Developmental effects of both gonadal sex (testes or ovaries) andchromosomal sex (XX or XY) affect food intake. Mice were gonadectomized4 wk before testing to eliminate the activational effects of sex hormones. Notethat 1) during the dark phase (left), gonadal females ate more than gonadalmales, regardless of their chromosomal sex, and 2) during the light phase(right; note altered scale), gonadal and chromosomal females (XX) ate morethan all other groups; these mice also had an approximately twofold more fatmass (not shown). Tests (not shown) of mice with XO and XXY chromosomesindicated that these effects were due to X-gene dosage, not the presence of theY chromosome. *P � 0.05; **P � 0.01. Republished with permission fromChen et al. (122).
Review
R1223SEX DIFFERENCES IN THE PHYSIOLOGY OF EATING
AJP-Regul Integr Comp Physiol • doi:10.1152/ajpregu.00446.2012 • www.ajpregu.org
by 10.220.32.247 on Septem
ber 18, 2016http://ajpregu.physiology.org/
Dow
nloaded from

below. Fessler (222) has advanced an interesting hypothesis con-cerning its ultimate adaptive meaning.
There are several reports of altered macronutrient selectionduring the estrous cycle (42, 261, 358, 423, 808). The forms ofmacronutrients used in these studies and the specific changes inmacronutrient selection observed varied widely, however, sug-gesting that food properties unrelated to macronutrient typecaused the results. For example, as reviewed below, cyclicchanges in the rewarding effect of sweet taste may contributeto cyclic changes in eating.
The estrous inhibition of eating in rats follows the diestrusincrease in plasma levels of estrogens with the time lagdiscussed above (please see Latency of estrogen’s effect oneating). In contrast, neither the smaller peak in plasma proges-tin levels during diestrus 2 nor the larger periovulatory peak isrelated to changed eating. The estrous inhibition of eating inrats is not secondary: 1) to stimulation of locomotor activitybecause the former is expressed as a decrease in meal size andthe latter causes a decrease in meal frequency (207); 2) toincreases in appetitive or consummatory reproductive behaviorbecause both estradiol and progesterone are necessary to nor-malize most or all reproductive behaviors in ovariectomizedrats (242, 549, 690); or 3) to estrogen-dependent changes inwater intake (151, 224, 237, 355, 384, 406, 726, 727) becausethese are not synchronous in intact, cycling rats (maximumwater intake occurs on diestrus 1 and decreases on diestrus 2)and because food intake increased �3 days before water intakeincreased after ovariectomy (727) [the estrogenic controls ofeating and drinking also were dissociated in several tests inguinea pigs (155)]. Nevertheless, it would be useful to deter-mine spontaneous meal patterns in a more naturalistic environ-ment permitting social interactions, reproductive behavior,foraging for food, etc. (for example, Refs. 386 and 473).
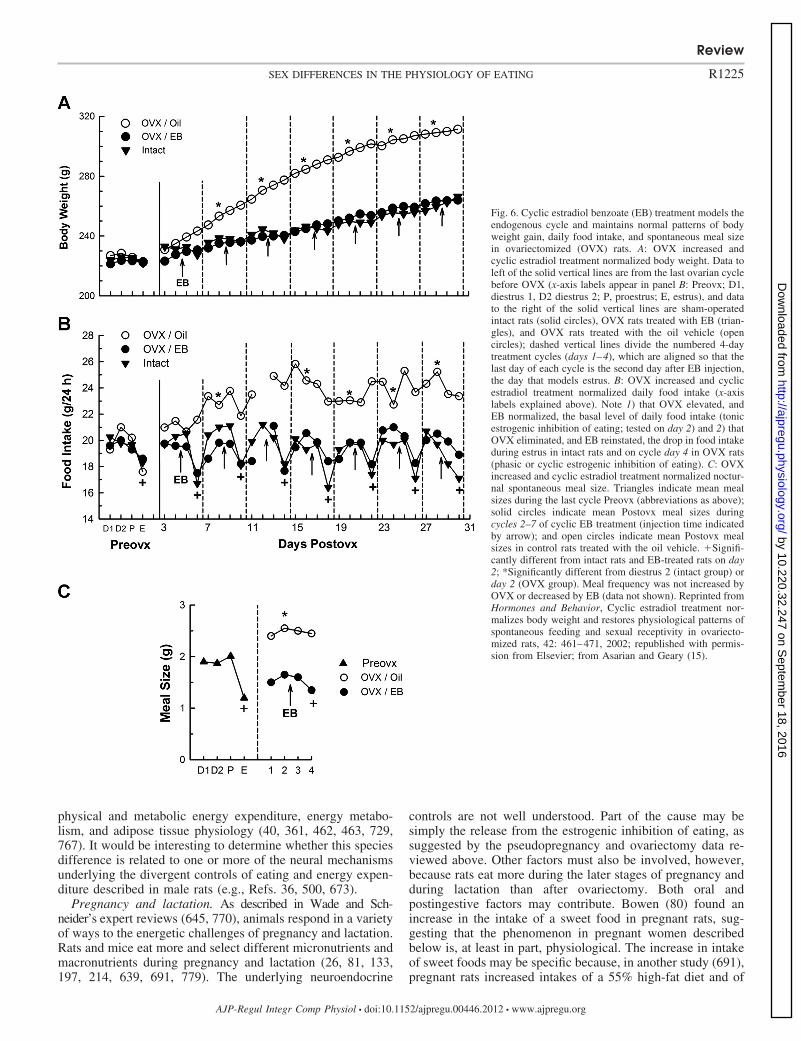
Ovariectomy and hormone treatment. Ovariectomy both abol-ishes the cyclicity of eating, as would be expected by disrup-tion of the ovarian cycle, and increases daily food intake abovethe diestrous maximum for several weeks, leading to increasedbody weight and adiposity (15, 77, 130, 589, 662, 726, 764)(Fig. 6A). In contrast to the effect of menopause in women,however, the gain in adipose tissue in rats is mainly in thesubcutaneous depots, not intra-abdominal depots, and leanbody mass is increased, not decreased (274).
As Drewett (191) originally pointed out, the acyclicity andthe increased overall level of eating after ovariectomy suggestthat HPG-axis function normally exerts two influences oneating: 1) a tonic inhibition, whose loss after ovariectomyincreases the basal level of eating, and 2) a phasic or cyclicinhibition, whose loss leads to acyclicity.
Early demonstrations that estradiol-treated ovariectomizedrats ate normal amounts and maintained normal body weightindicated that estrogens are the crucial link between HPG-axisfunction and eating in rats (191, 589, 726, 764, 766, 771) (forreviews see Refs. 18, 102, 203, and 253). Most importantly, anear-physiological cyclic regimen of estradiol treatment wassufficient to maintain both the tonic and the phasic controlsof eating, normal spontaneous meal patterns, and normal bodyweight in ovariectomized rats (15) (Fig. 6, B and C). In contrast,near-physiological doses of progesterone did not affect eating orbody weight in rats, although pharmacological doses may do so(259, 294, 764, 766). The efficacy of estradiol treatment tonormalize body weight may depend on when treatment is begun
because estradiol was markedly less effective in reversing ovari-ectomy-induced weight gain than in preventing it (726).
As discussed in the previous section, the cyclic inhibition ofeating during estrus is likely to result from the increase inestrogen secretion 24–48 h earlier during diestrus 2. Thisexplanation, however, leaves unclear why the even higherestrogen level during proestrus fails to further decrease eatingduring diestrus 1. This unexpected pattern may result from thecentral processing of the estrogenic signal. It does not involveanother ovarian control because it occurs in estradiol-treatedovariectomized rats as well (15).
The duration of exogenous estradiol’s eating-inhibitory ef-fect suggests that estrogen secretion during diestrus 2 andproestrus is sufficient to produce the tonic inhibition of eatingthroughout the cycle: 1) Asarian and Geary (16) observed thatin ovariectomized rats maintained on weekly cyclic treatmentwith 2 �g estradiol benzoate, food intake returned to the levelof untreated rats 5 days after estradiol treatment. Because ofthe rapid esterification of estradiol benzoate to estradiol and therapid clearance of estradiol from the circulation (417, 724),estradiol levels return to basal within 2 days of injection.2) Similarly, Gray and Greenwood (287) reported that ovari-ectomized rats’ food intake was still decreased 7 days afterinjection of 2 �g estradiol benzoate. 3) Tarttelin and Gorski(727) reported that if rats spontaneously entered pseudopreg-nancy, during which there is little estrogen secretion, intakeincreased above the diestrous level after �5 days. These datasuggest that the eating-inhibitory effect of estrogens secretedduring diestrus and proestrus persists �4–7 days, more thanlong enough to explain the tonic inhibition of eating during thecycle.
GnRH, LH, FSH, and prolactin do not appear to mediate theeffects of estradiol on eating because exogenous estradiol stillinhibited eating in hypophysectomized rats (771). Hypophy-sectomy also decreases the secretion of estrogens, however, sothe observation that hypophysectomy did not increase eating orbody weight in the same study (771) appears paradoxical. Itmay be that capacity of hypophysectomized rats to gain weightis impaired. Consistent with this idea, a more selective lesion,transgenic deletion of FSH receptors, prevented ovarian folli-cle development and produced a typical estrogen-deficiencysyndrome in female mice, including increased body weight andadiposity; unfortunately eating was not measured (162).
The effects of ovariectomy and of estradiol treatment oneating in rats, like the estrous decrease in eating, are expressedsolely as changes in spontaneous meal size; i.e., increases anddecreases, respectively (15, 77, 107, 376). Meal frequencyusually decreases slowly after ovariectomy and increases withestradiol treatment, but not enough to balance the meal sizeeffects, at least for several weeks after ovariectomy. Femalemice also decrease meal size during estrus (547). As reviewedbelow, this specificity has been a useful clue for investigationsof the underlying mechanisms. In contrast, the mechanism forthe slowly developing, apparently compensatory decrease inmeal frequency that limits the effect of ovariectomy on bodyadiposity has received no attention.
Finally, there appears to be a species difference in the effectof ovariectomy on eating in mice and rats. In mice, ovariec-tomy usually (350, 604, 796), but not always (75, 130),increases body weight and adiposity without affecting eating.This is presumably due to the many effects of estrogens on
Review
R1224 SEX DIFFERENCES IN THE PHYSIOLOGY OF EATING
AJP-Regul Integr Comp Physiol • doi:10.1152/ajpregu.00446.2012 • www.ajpregu.org
by 10.220.32.247 on Septem
ber 18, 2016http://ajpregu.physiology.org/
Dow
nloaded from
physical and metabolic energy expenditure, energy metabo-lism, and adipose tissue physiology (40, 361, 462, 463, 729,767). It would be interesting to determine whether this speciesdifference is related to one or more of the neural mechanismsunderlying the divergent controls of eating and energy expen-diture described in male rats (e.g., Refs. 36, 500, 673).
Pregnancy and lactation. As described in Wade and Sch-neider’s expert reviews (645, 770), animals respond in a varietyof ways to the energetic challenges of pregnancy and lactation.Rats and mice eat more and select different micronutrients andmacronutrients during pregnancy and lactation (26, 81, 133,197, 214, 639, 691, 779). The underlying neuroendocrine
controls are not well understood. Part of the cause may besimply the release from the estrogenic inhibition of eating, assuggested by the pseudopregnancy and ovariectomy data re-viewed above. Other factors must also be involved, however,because rats eat more during the later stages of pregnancy andduring lactation than after ovariectomy. Both oral andpostingestive factors may contribute. Bowen (80) found anincrease in the intake of a sweet food in pregnant rats, sug-gesting that the phenomenon in pregnant women describedbelow is, at least in part, physiological. The increase in intakeof sweet foods may be specific because, in another study (691),pregnant rats increased intakes of a 55% high-fat diet and of
Fig. 6. Cyclic estradiol benzoate (EB) treatment models theendogenous cycle and maintains normal patterns of bodyweight gain, daily food intake, and spontaneous meal sizein ovariectomized (OVX) rats. A: OVX increased andcyclic estradiol treatment normalized body weight. Data toleft of the solid vertical lines are from the last ovarian cyclebefore OVX (x-axis labels appear in panel B: Preovx; D1,diestrus 1, D2 diestrus 2; P, proestrus; E, estrus), and datato the right of the solid vertical lines are sham-operatedintact rats (solid circles), OVX rats treated with EB (trian-gles), and OVX rats treated with the oil vehicle (opencircles); dashed vertical lines divide the numbered 4-daytreatment cycles (days 1–4), which are aligned so that thelast day of each cycle is the second day after EB injection,the day that models estrus. B: OVX increased and cyclicestradiol treatment normalized daily food intake (x-axislabels explained above). Note 1) that OVX elevated, andEB normalized, the basal level of daily food intake (tonicestrogenic inhibition of eating; tested on day 2) and 2) thatOVX eliminated, and EB reinstated, the drop in food intakeduring estrus in intact rats and on cycle day 4 in OVX rats(phasic or cyclic estrogenic inhibition of eating). C: OVXincreased and cyclic estradiol treatment normalized noctur-nal spontaneous meal size. Triangles indicate mean mealsizes during the last cycle Preovx (abbreviations as above);solid circles indicate mean Postovx meal sizes duringcycles 2–7 of cyclic EB treatment (injection time indicatedby arrow); and open circles indicate mean Postovx mealsizes in control rats treated with the oil vehicle. �Signifi-cantly different from intact rats and EB-treated rats on day2; *Significantly different from diestrus 2 (intact group) orday 2 (OVX group). Meal frequency was not increased byOVX or decreased by EB (data not shown). Reprinted fromHormones and Behavior, Cyclic estradiol treatment nor-malizes body weight and restores physiological patterns ofspontaneous feeding and sexual receptivity in ovariecto-mized rats, 42: 461–471, 2002; republished with permis-sion from Elsevier; from Asarian and Geary (15).
Review
R1225SEX DIFFERENCES IN THE PHYSIOLOGY OF EATING
AJP-Regul Integr Comp Physiol • doi:10.1152/ajpregu.00446.2012 • www.ajpregu.org
by 10.220.32.247 on Septem
ber 18, 2016http://ajpregu.physiology.org/
Dow
nloaded from

chow similarly. Potential roles for CCK and leptin signaling inthe increase in eating during pregnancy are reviewed below(please see subsections Cholecystokinin and Leptin).
Lactational hyperphagia again highlights the primacy ofmeal size in the HPG control of eating in rats discussed above.That is, rat dams increase spontaneous meal size early inlactation and increase meal frequency only later on and ifnursing larger litters (227, 702). Lactational hyperphagia is notdependent on ovarian function because it was not affected bypostpartum ovariectomy (227). It may result from increases inNPY in the dorsomedial hypothalamus driven by increasedprolactin secretion and by downregulation of an inhibitory�MSH input (119, 802). The reductions in basal insulin andleptin that accompany lactation do not cause the hyperphagiabecause normalization of insulin and leptin levels did not af-fect it (811).
Contemporary investigators have not pursued a classicalobservation by Curt Richter on dietary self-selection by preg-nant and lactating rats (251). Richter (590) showed that if ratscould obtain certain micronutrients, such as sodium and cal-cium, from sources other than the source of dietary energy,energy intake during pregnancy and lactation was markedlyreduced. Thus, the appetites for micronutrients, not energy,drive much of the hyperphagia in chow-fed rats during preg-nancy and lactation. Clearly, the mechanisms underlying theseeffects should be investigated in situations that permit the ratsto regulate micronutrient homeostasis and energy homeostasisseparately.
Androgens. Androgens have activational effects on eating inrats in addition to the organizational effects described above.Adult orchiectomy decreases rats’ daily food intake and bodyweight, and androgen treatment, usually with testosterone pro-pionate, normalizes both (115, 266, 402, 516, 518, 622, 764,767, 776). Testosterone increased eating similarly in one studyin mice (546), but not another (495). In both species, the eatingeffects were due to changes in meal frequencies, with meal sizemoving in the opposite direction (115, 546). The mechanismsthrough which orchiectomy and androgen treatment affecteating have been studied far less than those of ovariectomy andestrogen treatment. Androgens, like estrogens, have manymetabolic effects that can lead to changes in body weight andcomposition in the absence of changes in eating (40, 361, 462,463, 625, 729, 767), and orchiectomy and androgen treatmentseem to affect body weight and body composition more reli-ably than they do eating. Transgenic mice lacking androgenreceptors also increased adiposity without increasing eating(216).
The effects of androgens on eating may be related in part toaromatization to estrogens. In some (288, 519, 664), but not all(199, 266, 622), studies, treatment with relatively high doses oftestosterone propionate, which can be aromatized to estrogens,increased eating more potently than similar doses of nonaro-matizable androgens, such as of 5-alpha-dihydrotestosteronepropionate. It remains unclear, however, whether physiologicalandrogen doses would produce such an effect.
Sex Differences in Eating in Anthropoid Primates
Male-female differences. Although males and females mayeat differently from a very young age, most sex differences inhuman eating do not appear to be physiologically based (171,
188, 234, 565, 605, 783, 788). For example, Wardle et al.(783), in an analysis of data from 23 countries, found thatwomen chose fewer high-fat foods and more fruits and high-fiber foods than men, but that health beliefs explained �50%of the effects and dieting status as much as 20%. Nevertheless,there are at least four apparently physiological sex differencesin human eating. 1) Men, who are generally larger than women, eatmore than women and, as in rats, increases in meal size ratherthan in meal frequency produced this difference (172). 2) Menwere more responsive than women to the negative-feedbackeffects of oral nutrient loads on eating in several situations(170, 544, 605). 3) Men were more responsive than women tothe eating-stimulatory effect of food deprivation (820), paral-leling the rat and mouse phenomena described above. 4)Finally, again as in rats and mice, normal homeostatic eatingmaintains a significantly higher body adiposity in women thanin men; at a “normal” BMI of 22–23 kg/m2, women had 26%fat as a percent of body weight vs. 13% in men (245).Understanding the mechanisms underlying this sex differencemay have important ramifications for the general understand-ing of energy homeostasis.
A number of epidemiological studies done in several West-ern societies that involved different ethnicities and socialstrata, included adults and children as young as 2–5 yr of age,and assessed intakes of solid foods, carbonated beverages, andfruit juices failed to detect male-female differences in sugarintake, expressed as a percentage of total energy intake (78,539, 733, 782). Sex differences in food selection did appear,however, in surveys of obese persons. Drewnowski et al. (194)reported that obese men identified high-fat, high-protein foodsamong their favorites, whereas obese women identified, high-fat, high-carbohydrate foods, especially high-sugar foods, amongtheir favorites. Macdiarmid et al. (451) showed that this dif-ference was reflected in food intake: obese women ate morehigh-sugar, high-fat foods (median intake �146 g/day ofcakes, chocolate, etc.) than did obese men (�103 g/day),leading to a higher sugar intake (21 vs. 17% of daily energy);nonobese persons did not show this difference. As discussedbelow, these differences may result from sex differences inflavor hedonics. It is important to determine whether theydevelop prior to obesity or are consequences of obesity (e.g.,Ref. 696). Finally, recent data suggest that the trend towardovereating in the United States is stronger in women, whoincreased daily energy intake 22% between 1971 and 2004,than men, who increased only 10% (437). In that overeatingappears to be the primary cause of the obesity epidemic (195,496, 716), this difference could contribute to the sex differ-ences in obesity prevalence mentioned above (226).
Three findings support the view that a physiological sexdifference affects sweet preference in anthropoid primates:1) Among the Hadza of Tanzania, hunter-gatherers who derive�90% of their energy from wild food, although honey was themost preferred food in both sexes, women preferred sweetberries more than meat, whereas men preferred meat more thanberries (61). 2) A field study of wild Borneo orangutans(Pongo pygmaeus) indicated that when sweet fruits were inseason, males increased their intake about twofold (from 3,800to 8,400 kcal/day), whereas females increased their intakeabout four-fold (from 1,800 to 7,400 kcal/day) (401). 3) In alaboratory study in which savanna baboons (Papio cynoceph-
Review
R1226 SEX DIFFERENCES IN THE PHYSIOLOGY OF EATING
AJP-Regul Integr Comp Physiol • doi:10.1152/ajpregu.00446.2012 • www.ajpregu.org
by 10.220.32.247 on Septem
ber 18, 2016http://ajpregu.physiology.org/
Dow
nloaded from

alus) were offered 75% sucrose fruit-flavored candy and chowpellets, females ate relatively more sugar than males (228).
Ovarian cycle. CYCLIC CHANGES IN EATING. We know of threewithin-subjects studies of women’s eating throughout the ovar-ian cycle (229, 278, 450). These involved a total of 53 women.Food intake was measured by weighing the food and convert-ing these data to energy equivalents, and cycling was charac-terized by urinary LH assays and reports of menses. As shownin Fig. 7A, these studies demonstrated that eating decreasesthrough the follicular phase to a minimum during the periovu-latory phase and is similarly high during the early-follicularand luteal phases. The weighted mean differences in foodintake between the luteal and periovulatory phases was 275
kcal/day, and that between the mid-follicular and luteal phaseswas 228 kcal/day. Significant differences in food intake be-tween the midfollicular and midluteal phases were detected in9 of 10 similarly designed studies in which only those twocycle phases were tested (n � 192, mean effect, 218 kcal/day)(38, 353, 431, 436, 458, 544, 558, 561, 725, 809); the negativeresult occurred in the smallest study (n � 9) (809). Similarcyclic changes in food intake also were found in many studiesreviewed by Buffenstein et al. (92) and by Dye and Blundell(198), which used less sensitive methods, such as use of bodytemperature to monitor cycling and use of food diaries tomeasure eating (789). Those studies also provided evidencethat the cyclic changes in eating do not occur during anovula-tory cycles (38, 596).
None of the studies reviewed above determined whether thecyclic changes observed were due to changes in spontaneousmeal size, as in rats, or changes in meal number. In two studiesinvolving test meals, however, meal size was significantly lessin the midfollicular phase than in the luteal phase (85, 561),consistent with the idea that in women as in rats, cyclicchanges in eating are expressed as changes in spontaneousmeal size.
Cyclic changes in eating across ovarian cycle similar to thatseen in women were found in several studies of rhesus ma-caques (153, 154, 156, 373, 615) (Fig. 7B) and in both captiveand wild chacma baboons (Papio ursinus) (70). The monkeystudies demonstrate that eating decreases continuously duringthe follicular phase and is maintained at a relatively constanthigh level during the luteal phase. In wild baboons, the cycliceffect was similar in females that were in consort with malesand those that were not in consort, indicating that reproductivebehavior did not influence eating.
All of these data, together with the apparently similar peri-ovulatory decrease in eating in rats and mice, indicate thatcyclic changes in women’s eating are biological sex differ-ences under the control of HPG axis function. The magnitudesof the effects are more than large enough to be relevant to bodyweight regulation; recent estimates suggest that consistentimbalances of only 50–100 kcal/day are sufficient to cause thegradual development of obesity (437, 496).
HPG-AXIS MEDIATION. Eating during the follicular phase inanthropoid primates is closely associated with changes inestrogen levels. From a neuroendocrine perspective, the phaseof maximal intake in rats, diestrus, better parallels the early-follicular phase than the luteal phase. Thus, if estrogens doinhibit eating during the follicular phase, then food intakeshould be maximal during the early follicular phase, whenestrogen levels are lowest, and then decrease progressivelyuntil the periovulatory phase, when estrogen levels are highest.Although this hypothesis has not been investigated quantita-tively, the available data in women (229, 278, 450) and rhesusmonkeys (154, 156, 756) are consistent with it (Fig. 7).
In contrast, the contribution of gonadal steroids to thecontrol of eating during the luteal phase is problematic. Estro-gen levels are higher in the mid-luteal phase than the mid-follicular phase, although food intake is less during the mid-follicular phase. This pattern is inconsistent with a simpleestrogenic inhibitory effect. Progestins do not appear to pro-vide a solution. Progestins are secreted during the luteal phase,and exogenous progesterone can reduce the eating-inhibitoryeffect of estrogens, but this appears to be a pharmacological
Fig. 7. Daily food intake during the ovarian cycle in women (A) and rhesusmacaques (B). Note the progressive decreases in food intake during thefollicular phase in both women and monkeys and the high, constant levels offood intake during most of the luteal phase in monkeys (women’s data wereaveraged across the entire luteal phase). Women’s data (kilocalories eaten perday; values are expressed as means SE) are calculated from three studies inwhich food intake was measured by weighing, and the cycle phase wasmonitored with urinary LH and reports of menses in a total of 34 women. Ineach study, data were averaged across the early-follicular (eF; 4 day), midfol-licular (mF; �9 day), periovulatory (PO; 4 day), and luteal (L; �11 day)phases. *Significantly different from luteal phase. Adapted from Am. J. Clin.Nutr. (1993; 57: 43–46), American Society for Nutrition (229); Am. J. Clin.Nutr. (1989; 49: 252–258), American Society for Nutrition (278); and Am. J.Clin. Nutr. (1989; 49: 1164–1168), American Society for Nutrition (450).Monkey data are plasma LH concentrations (open circles, means SE) anddaily food intakes (solid bars, means SE) averaged across consecutive 3-dayintervals relative to the LH peak (day 0) in 7 monkeys. *Significantly differentfrom day 2/0. Reprinted with permission from Human Reproduction Update,Brain imaging studies of appetite in the context of obesity and the menstrualcycle, Dean A. Van Vugt, 16: 276–292, 2010 (756).
Review
R1227SEX DIFFERENCES IN THE PHYSIOLOGY OF EATING
AJP-Regul Integr Comp Physiol • doi:10.1152/ajpregu.00446.2012 • www.ajpregu.org
by 10.220.32.247 on Septem
ber 18, 2016http://ajpregu.physiology.org/
Dow
nloaded from

rather than a physiological effect in rats (259, 764, 766) andmonkeys (70, 154, 372). For example, using estradiol andprogesterone doses that led to changes in plasma levels thatwere within the range occurring during the menstrual cycle,Czaja (154) failed to detect any effect of either acute or chronicprogesterone treatment on eating in ovariectomized rhesusmacaques. The effect of progestins in the absence of endoge-nous gonadal steroids has not been studied in women. Pelkmanet al. (544), however, failed to detect any effect of depotmedroxyprogesterone on food intake in an adequately pow-ered, prospective, placebo-controlled study in cycling women.That result extends an earlier study in which a low-doseestrogen (35 �g/day ethinyl estradiol) together with escalatingprogestin doses (0.5–1.0 mg/day norethindrone) failed to de-tectably affect food intake measured with diet records in cyclingwomen (201). These studies suggest, but do not prove, thatsome factor other than estrogens and progestins controls eatingduring the luteal phase. Further work is required to identify thatfactor.
MACRONUTRIENT INTAKE. Dye and Blundell (198) reviewed15 studies in which macronutrient intakes were tabulatedacross the ovarian cycle; none found a significant effect.Nevertheless, there may be a cyclic change in the intake ofsweet foods. Bowen and Grunberg (82) found that women atemore of three sweet test foods during the luteal phase thanduring the mid-follicular phase, whereas intakes of salty and ofbland foods did not vary. Similarly, Fong and Kretch (229)reported that women consumed significantly more sugar-con-taining carbonated drinks (typically about 0.3 M sucrose)during the luteal phase (417 27 ml/day; mean SE) thanduring the midfollicular, periovulatory, or menses phases (335 31, 359 26, and 370 26 ml/day, respectively). Kemnitz etal. (373), however, failed to find a cyclic change in rhesusmacaques’ intake of 0.5 M sucrose that was offered 3 days/wk,2 h/day. The possible contributions of cyclic changes in sen-sory and hedonic processing of sweets is discussed below.
INTERACTION WITH COGNITIVE CONTROLS OF EATING. The fol-licular and periovulatory decrease in eating can be modulatedby non biological factors. For example, the decrease is appar-ently blunted or absent in women with high levels of dietaryrestraint (92, 198), a psychological trait related to the ability tolimit eating cognitively that is an important control of eating inmany young women (322, 328, 794). In an interesting study, Liet al. (431) found that the difference in energy intake betweenthe follicular and luteal phases in a group of university studentswas �10% when weekday data were used and �23% whenweekend data were used. This weekend effect was not attrib-utable to changes in alcohol intake, in frequency of restaurantvisits, or in the daily level of energy intake, which did not varysignificantly across the week. One possibility is that it was dueto the relaxation of cognitive restraint on weekends, but thiswas not measured.
Food cravings, i.e., recurring intense desires for specificfoods, are a common and stable aspect of human eating,distinct from the palatability [i.e., craved foods are not simplythose an individual finds most palatable (786, 813)]. In both theUnited States and Spain, women crave sweet foods mostfrequently, whereas men crave mainly savory foods (543, 623,823). This sex difference may be related to the sex differencein sweet hedonics discussed below.
Chocolate is the single most craved food among women inthe United States, United Kingdom, Canada, and Spain, andmany women report cyclic changes in craving, especially forchocolate, with a maximum in the late luteal and mensesphases (738, 822). This cyclic effect does not seem to behormonally based: 1) neither frequency of cravings nor thetype of food craved were significantly related to circulatingestrogen levels (597); 2) progesterone administration did notaffect cravings (480); 3) premenopausal and postmenopausalwomen who were chocolate cravers had similar frequencies ofchocolate craving (338); and 4) there was no cyclic change inchocolate cravings in Spanish women whose chocolate crav-ings were as intense as those reported by U.S. Americanwomen who did crave cyclically (822).
Carbohydrate craving may represent a form of self-medica-tion for dysphoria (137). Early reports linked carbohydratecraving to serotonergic (640) and opioidergic (192, 193) func-tion, but neither lead has been pursued. Carbohydrate cravingis often discussed in relation to premenstrual dysphoria andbulimia nervosa, but whether they are, indeed, linked remainscontroversial (132, 296, 752, 813).
Loss of ovarian function and hormone treatment. Meno-pause increases adiposity independent of aging (336, 429, 535,550, 714, 739, 741), and some data indicate that high-estrogenhormone-replacement treatment (HRT) prevents this (297, 312,419, 628, 686). Whether ovariectomy (or oophorectomy),menopause, or exogenous estrogens affect eating in women,however, remains in doubt, perhaps due to the inadequacies inthe designs used so far. For example, Lovejoy et al. (443) hadsubjects complete 4-day food diaries annually and failed todetect any change in total energy intake between 3 yr premeno-pause and 2 yr postmenopause. The power of this study,however, was such that changes of �30% were required forsignificance. We know of three reports in which HRT failed toaffect eating (585, 587, 672). None of these studies, however,was strongly designed. For example, in one (587), only a singleday’s food intake was estimated by interview, and in another(585), a between-subjects design with only 6 and 9 women wasused. In the third study (672), normal-weight women who hadhad natural menopause 0.5–5 yr previously were randomlyassigned either an HRT regimen or to placebo in a double-blinddesign (n � 14 each), and eating was estimated by food diariesfor 3-day periods at the onset and after 2 yr of treatment. HRTwas associated with a decrease in daily intake of 212 kcal/day(12%), compared with no change in untreated women, but noneof the differences was statistically significant. The power of thefood diary method to detect differences and the potentialinfluence of the time since menopause were not assessed. Inaddition, HRT was a relatively high-progestin regimen (6.25mg/day oral conjugated estrogens plus 2.5 mg/day medroxy-progesterone), which may have masked an effect of estrogenson eating, as occurs in rats (766).
In contrast, experiments with monkeys provide strongersupport for the hypothesis that loss of ovarian function in-creases eating due to release from estrogenic inhibition. Sulli-van et al. (705) showed rhesus macaques’ food intake in-creased 29% and body weight 3% during the first 2 mo afterovariectomy compared with preovariectomy baselines. In afollow-up study (707), they found that 3 mo of treatment withthe selective estrogen-receptor modulator GSK232802A de-creased body weight 4.6% in a group of ovariectomized rhesus
Review
R1228 SEX DIFFERENCES IN THE PHYSIOLOGY OF EATING
AJP-Regul Integr Comp Physiol • doi:10.1152/ajpregu.00446.2012 • www.ajpregu.org
by 10.220.32.247 on Septem
ber 18, 2016http://ajpregu.physiology.org/
Dow
nloaded from

macaques whose body weights previously increased 4.8% afterovariectomy. GSK232802A decreased food intake �5–10%during the treatment period, and food intake also correlatednegatively (r � 0.52) with weight loss during this period.Finally, GSK232802A increased spontaneous physical activ-ity, measured with triaxial accelerometry. Interestingly, thefattest, least active monkeys showed the largest activity in-creases and weight losses. They also previously reported thatspontaneous activity levels, but not food intake levels, wereinversely correlated with weight gain over a 3-mo period inintact female rhesus macaques (706). These data, together withLovejoy et al.’s (443) report that spontaneous activity de-creases across the menopausal transition, identify physicalactivity and eating as important targets for future menopauseresearch.
Pregnancy and lactation. Women increase energy intakeduring pregnancy and lactation in a way that appears to meettheir increased energetic costs (47, 71, 569). Part of themechanism appears to be related to altered food preferences.Most women increase intake of craved foods during the thirdthrough the seventh month of pregnancy (58, 81, 337, 564).Sweet foods, including fruits and fruit juices, dairy foods, andchocolate are most frequently craved. Increased food cravingsduring pregnancy are also thought to contribute to excessivegestational weight gain, which occurs in about one third ofpregnant women and increases a variety of health risks both formother and child (144, 526, 642, 654). For this reason andbecause the results of current intervention strategies are incon-sistent (721), the physiological controls of increased eatingduring pregnancy are an important research challenge.
Some women also develop aversions, often to foods that areusually liked. Because food aversions and nausea develop mostfrequently around the third month of pregnancy, it is possiblethat they are produced by the increases in human chorionicgonadotropin that occur at this time (244, 540). A moderatedegree of nausea and vomiting is considered normal duringpregnancy and may improve pregnancy outcomes (244, 540).
Androgens. The role of androgens in eating in anthropoidprimates is unknown. In rhesus monkeys, testosterone treat-ment failed to significantly affect eating despite producinglarge (�50%) increases in body weight and lean body mass in7–9 yr-old monkeys that had been orchiectomized at 0–3 mo ofage (375). The potential influences of the early orchiectomyand the long orchiectomy-treatment delay on the results werenot assessed. We know of no studies of androgens and eatingin healthy men. Male hypogonadism has a variety of causes,but often appears as men’s testosterone levels decrease duringaging (161, 553, 711). It is associated with increased adiposity.Changes in appetite have not been reported to our knowledge.Hypogonadism is treated with testosterone, which suppressesthe HPG axis and leads to infertility, or, if fertility is desired,by stimulating the HPG axis with either gonadotropins or theestrogen-receptor modulator clomiphene citrate, which de-crease estrogenic negative feedback (161, 553, 711). Thus,hypogonadal men seem to present a potentially interestingmodel for investigation of androgenic influences on eating.
ER Mechanisms Controlling Eating
Two types of nuclear ER, ER�, and ER�, were identified inthe 1990s (178, 512). These are encoded by separate genes,
ESR1 and ESR2 in humans and Er� and Er� in mice. The twogenes have overlapping but distinct expression patterns in thebrain and periphery, and Er� and Er� knockout mice displaydistinctive phenotypes. The classical mode of action of nuclearER involves binding to estrogen-response elements (ERE),which, in turn, bind to DNA and affect gene transcription,usually over the course of hours to days (125, 326). Morerecently, it has been found that ER� and ER�, as well as avariety of novel ER, are expressed outside the nucleus, usuallyon cell membranes (428, 475, 548, 602, 759). In addition togenomic effects via non-ERE transcription promotors, mem-brane ER have rapid (i.e., within seconds) nongenomic effects.These rapid effects are increasingly recognized to be crucial formany neural actions of estrogens.
ER� in rats and mice. The phenotypes of transgenic micewith null mutations affecting estrogen signaling via ER� dem-onstrate a necessary role for ER� role in weight regulation, butdo not clearly disclose their role in the control of eating. Heineet al. (324) characterized the obesity phenotype of the originalline of Er�/ mice. Both male and female Er�/ mice hadexcess body weight and body fat, especially intra-abdominalfat, signs of metabolic syndrome, and decreased energy expen-diture. Food intake, tested continuously beginning at 60 days ofage, was unchanged in male Er�/ mice; again females werenot tested. Ohlsson et al. (523) found a similar adiposityphenotype in another line of Er�/ mice, but did not measureeating. Jones et al. (356) tested transgenic aromatase-deficientmice, which cannot synthesize estrogens, and found that foodintake was reduced in females beginning before 5 wk of age;males were not tested. Xu et al. (812) reported that femaletransgenic mice lacking ER� only in the brain ate more thanwild-type mice at 7 wk of age, but they did not test other ages.Finally, Park et al. (537) described a strain of Er�/ mice inwhich non-ERE estrogen signaling was transgenically rescued,Er�/AA mice. Eating tests beginning at 3 wk of age failed toreveal any difference between female wild-type mice andeither Er�/ or Er�/AA mice in intake of chow or of a 45%high-fat diet. We conclude that there is no apparent reason toexpect disruption of peripheral ER� signaling to obscure aneffect of brain ER� lesion on eating and, therefore, that untilXu et al.’s (812) solitary report of increased eating in trans-genic mice with disrupted ER� is extended, the Er�/ phe-notype fails to provide clear evidence that ER� signaling isnecessary for the normal control of eating. Given the difficultyin interpreting negative transgenic null-mutation data and themany positive indications that ER� is involved in the control ofeating that we review below, however, we do not consider thisa crucial failure.
To better establish whether activational effects of estrogenscontrol eating and adiposity, Geary et al. (257) tested theeffects of estradiol treatment in Er�/ mice that were ovari-ectomized after puberty. Estradiol reduced daily food intake,weight gain, and fat gain in wild-type, but not Er�/ mice(Fig. 8). Estradiol also increased CCK satiation in wild-type,but not Er�/, mice (please see Cholecystokinin). BecauseCCK is involved in the phasic control of eating by estrogens, thesedata indicate that an activational effect of estrogen signaling viaER� is involved in both the phasic and the tonic estrogenic controlof eating. This does not preclude the possibility that organizationaleffects mediated by ER� are also important for these controls.
Review
R1229SEX DIFFERENCES IN THE PHYSIOLOGY OF EATING
AJP-Regul Integr Comp Physiol • doi:10.1152/ajpregu.00446.2012 • www.ajpregu.org
by 10.220.32.247 on Septem
ber 18, 2016http://ajpregu.physiology.org/
Dow
nloaded from
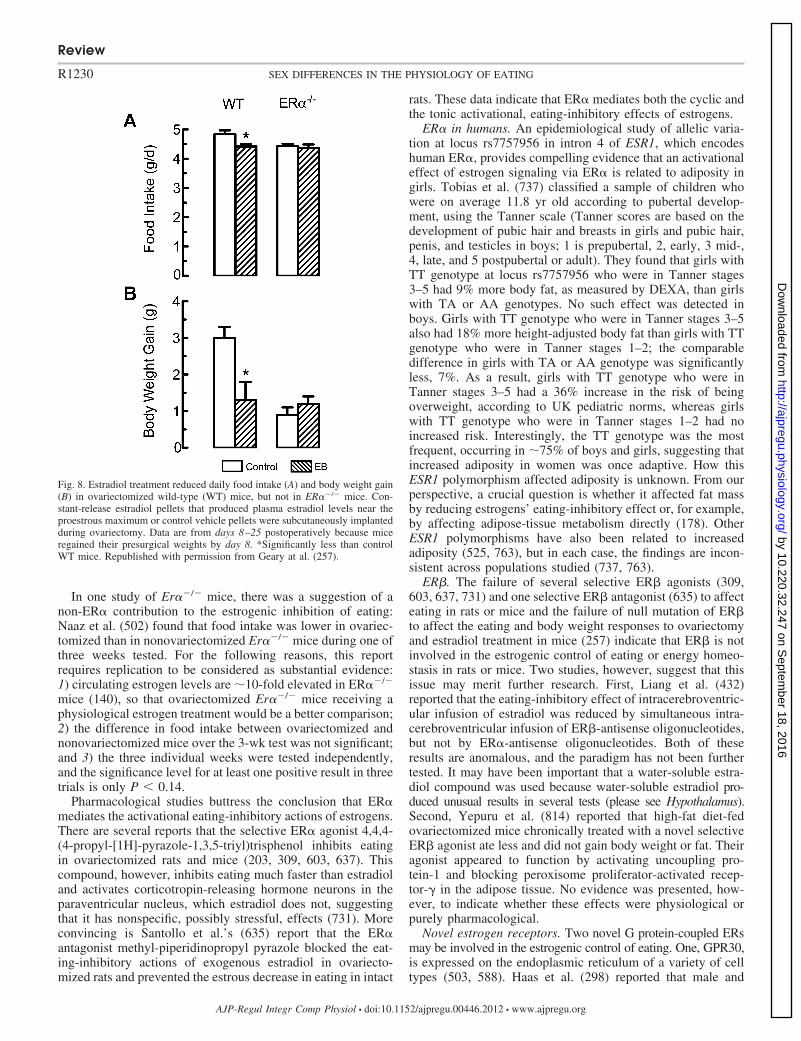
In one study of Er�/ mice, there was a suggestion of anon-ER� contribution to the estrogenic inhibition of eating:Naaz et al. (502) found that food intake was lower in ovariec-tomized than in nonovariectomized Er�/ mice during one ofthree weeks tested. For the following reasons, this reportrequires replication to be considered as substantial evidence:1) circulating estrogen levels are �10-fold elevated in ER�/
mice (140), so that ovariectomized Er�/ mice receiving aphysiological estrogen treatment would be a better comparison;2) the difference in food intake between ovariectomized andnonovariectomized mice over the 3-wk test was not significant;and 3) the three individual weeks were tested independently,and the significance level for at least one positive result in threetrials is only P � 0.14.
Pharmacological studies buttress the conclusion that ER�mediates the activational eating-inhibitory actions of estrogens.There are several reports that the selective ER� agonist 4,4,4-(4-propyl-[1H]-pyrazole-1,3,5-triyl)trisphenol inhibits eatingin ovariectomized rats and mice (203, 309, 603, 637). Thiscompound, however, inhibits eating much faster than estradioland activates corticotropin-releasing hormone neurons in theparaventricular nucleus, which estradiol does not, suggestingthat it has nonspecific, possibly stressful, effects (731). Moreconvincing is Santollo et al.’s (635) report that the ER�antagonist methyl-piperidinopropyl pyrazole blocked the eat-ing-inhibitory actions of exogenous estradiol in ovariecto-mized rats and prevented the estrous decrease in eating in intact
rats. These data indicate that ER� mediates both the cyclic andthe tonic activational, eating-inhibitory effects of estrogens.
ER� in humans. An epidemiological study of allelic varia-tion at locus rs7757956 in intron 4 of ESR1, which encodeshuman ER�, provides compelling evidence that an activationaleffect of estrogen signaling via ER� is related to adiposity ingirls. Tobias et al. (737) classified a sample of children whowere on average 11.8 yr old according to pubertal develop-ment, using the Tanner scale (Tanner scores are based on thedevelopment of pubic hair and breasts in girls and pubic hair,penis, and testicles in boys; 1 is prepubertal, 2, early, 3 mid-,4, late, and 5 postpubertal or adult). They found that girls withTT genotype at locus rs7757956 who were in Tanner stages3–5 had 9% more body fat, as measured by DEXA, than girlswith TA or AA genotypes. No such effect was detected inboys. Girls with TT genotype who were in Tanner stages 3–5also had 18% more height-adjusted body fat than girls with TTgenotype who were in Tanner stages 1–2; the comparabledifference in girls with TA or AA genotype was significantlyless, 7%. As a result, girls with TT genotype who were inTanner stages 3–5 had a 36% increase in the risk of beingoverweight, according to UK pediatric norms, whereas girlswith TT genotype who were in Tanner stages 1–2 had noincreased risk. Interestingly, the TT genotype was the mostfrequent, occurring in �75% of boys and girls, suggesting thatincreased adiposity in women was once adaptive. How thisESR1 polymorphism affected adiposity is unknown. From ourperspective, a crucial question is whether it affected fat massby reducing estrogens’ eating-inhibitory effect or, for example,by affecting adipose-tissue metabolism directly (178). OtherESR1 polymorphisms have also been related to increasedadiposity (525, 763), but in each case, the findings are incon-sistent across populations studied (737, 763).
ER�. The failure of several selective ER� agonists (309,603, 637, 731) and one selective ER� antagonist (635) to affecteating in rats or mice and the failure of null mutation of ER�to affect the eating and body weight responses to ovariectomyand estradiol treatment in mice (257) indicate that ER� is notinvolved in the estrogenic control of eating or energy homeo-stasis in rats or mice. Two studies, however, suggest that thisissue may merit further research. First, Liang et al. (432)reported that the eating-inhibitory effect of intracerebroventric-ular infusion of estradiol was reduced by simultaneous intra-cerebroventricular infusion of ER�-antisense oligonucleotides,but not by ER�-antisense oligonucleotides. Both of theseresults are anomalous, and the paradigm has not been furthertested. It may have been important that a water-soluble estra-diol compound was used because water-soluble estradiol pro-duced unusual results in several tests (please see Hypothalamus).Second, Yepuru et al. (814) reported that high-fat diet-fedovariectomized mice chronically treated with a novel selectiveER� agonist ate less and did not gain body weight or fat. Theiragonist appeared to function by activating uncoupling pro-tein-1 and blocking peroxisome proliferator-activated recep-tor-� in the adipose tissue. No evidence was presented, how-ever, to indicate whether these effects were physiological orpurely pharmacological.
Novel estrogen receptors. Two novel G protein-coupled ERsmay be involved in the estrogenic control of eating. One, GPR30,is expressed on the endoplasmic reticulum of a variety of celltypes (503, 588). Haas et al. (298) reported that male and
Fig. 8. Estradiol treatment reduced daily food intake (A) and body weight gain(B) in ovariectomized wild-type (WT) mice, but not in ER�/ mice. Con-stant-release estradiol pellets that produced plasma estradiol levels near theproestrous maximum or control vehicle pellets were subcutaneously implantedduring ovariectomy. Data are from days 8–25 postoperatively because miceregained their presurgical weights by day 8. *Significantly less than controlWT mice. Republished with permission from Geary at al. (257).
Review
R1230 SEX DIFFERENCES IN THE PHYSIOLOGY OF EATING
AJP-Regul Integr Comp Physiol • doi:10.1152/ajpregu.00446.2012 • www.ajpregu.org
by 10.220.32.247 on Septem
ber 18, 2016http://ajpregu.physiology.org/
Dow
nloaded from

female mice with null mutations of GPR30 (GPR30/ mice)developed marked intra-abdominal obesity, as well as a num-ber of other abnormalities. The effects of ovariectomy andestradiol treatment, however, did not differ between wild-typemice and a different strain of GPR30/ mice (795). A secondnovel ER, Gq-mER, is expressed on the cell membranes ofproopiomelanocortin (POMC), dopamine, and GABA neuronsin the arcuate nucleus of the hypothalamus (576, 601, 602).Gq-mER reduces the activation of potassium channels onGABAB and �-opioid receptors and affects the expression ofNPY and other neuropeptides involved in the control of energyhomeostasis. The Gq-mER-selective agonist STX reducedfood intake and body weight gain in ovariectomized guineapigs under several conditions (576, 599, 600). Importantly,however, doses of STX and estradiol that had similar effects ontotal food intake had opposite effects on spontaneous mealpatterns: estradiol decreased meal frequency, whereas STXdecreased mainly meal size (600). This suggests that STX isunlikely to mimic the tonic effects of endogenous estrogens oneating in guinea pigs. Unfortunately, whether the estrous de-crease in eating in guinea pigs (156) is produced by changes inmeal size or meal frequency is unknown.
More generally, the apparent necessity of ER� for the estro-genic inhibition of eating in rats and mice and the �2-day latencyof this effect both indicate that the rapid actions of novel mem-brane ER are unlikely to be sufficient for the estrogenic inhibitionof eating in these species (12, 731). They may, however, interactwith slower, ER�-mediated effects to control eating. Kow andPfaff (404) described just such an interaction in the control of thelordosis reflex. They found that maximal lordosis was induced bytwo brief pulses of estradiol, separated by 5 h, delivered into theVMH, and that this occurred even if only one pulse was estradioland the other pulse was a complex of estradiol bound to albumincomplex, which does not penetrate cell membranes and, therefore,activated membrane ER.
Site of ER Controlling Eating
Eckel (203) and Rivera and Eckel (592) provided pharma-cological evidence that activation of ER in the brain is neces-sary and sufficient for the estrogenic inhibition of eating bycomparing the eating-inhibitory effects of peripheral and cen-
tral injections of ICI-182,780, a pure antiestrogen that appar-ently does not cross the blood-brain barrier. Infusion of ICI-182,780 into the lateral ventricle reversed the eating-inhibitoryeffect of peripheral estradiol injection in ovariectomized rats,but subcutaneous ICI-182,780 injections did not. Conversely,peripheral, but not central, ICI-182,780 treatment blockedestradiol’s effects on uterine weight and vaginal cytology. Xuet al.’s (812) report that transgenic mice with a brain-specificdeletion of ER� are hyperphagic and obese is also consistentwith Rivera and Eckel’s (592) data, although a developmentaldefect appeared to account for most of their effect (i.e., hy-perphagia seemed to appear before puberty), and the effects ofovariectomy and estrogenic stimulation were not tested.
Investigations of brain sites of ER that control eating usinglocal estradiol administration and brain lesions are summarizedin Table 1 (studies using central doses greater than �300 ngare not included for the reasons described in Hormone treat-ment regimens). As Table 1 and the discussion below indicate,there is no consistent evidence for a role of any hypothalamicER population in the estrogenic control of eating in rats ormice. In contrast, several approaches implicate ER� in thecaudal medial nucleus of the solitary tract (cmNTS) in theestrogenic inhibition of eating.
cmNTS. The first suggestion that the nucleus of the solitarytract (NTS) might be involved in the estrogenic control ofeating came from our finding that the NTS was one of the brainareas in which estradiol treatment increased the number ofneurons expressing c-Fos after food ingestion or CCK injectionin ovariectomized rats and mice (205, 206, 257, 688) (theeffects of estrogens on CCK satiation are reviewed in Chole-cystokinin). The next c-Fos finding was more interesting.Estradiol increased the numbers of cells expressing c-Fos inresponse to intraduodenal infusions of fat only in one brainarea, the cmNTS just posterior to the area postrema. The effectwas CCK-dependent, and, most importantly, many of theneurons expressing c-Fos also expressed ER� (17). The cmNTS hasthe densest population of ER�-expressing neurons in the NTS(17, 643, 663, 668, 688, 689, 757).
We then showed that local administration of estradiol to thesurface of the hindbrain over the cmNTS was sufficient toreduce eating and to increase the number of ER�-expressing
Table 1. Status of central ER populations in the control of eating
Site Sufficient? Necessary? Comment
Brain stemcmNTS Yes (732) Yes (19, 20) ER�/c-Fos colocalizationDR Yes (636) Latency
HypothalamusVMH Yes (636) Latency
No (103, 533) No (75, 374, 499, 734, 812)MPOA Yes (158, 636) Latency
No (103, 343) No (381)PVN Yes (103, 104, 108) Yes (106)
No (343) No (159)
Sufficiency is indicated by a decrease in eating following administration of small amounts of estradiol in the brain area indicated in ovariectomized rats;necessity is indicated by a diminished or absent effect of peripheral estradiol on eating or weight gain following electrolytic, chemical, or molecular lesion ofthe brain area indicated in ovariectomized rats or, in the case of the ventromedial hypothalamus (VMH) in mice. VMH data include both the ventromedial nucleus(VMN) and the arcuate nucleus (Arc). References are shown in parentheses. DR, dorsal raphe nucleus; cmNTS, caudal medial nucleus of the solitary tract;MPOA, medial preoptic area; PVN, paraventricular nucleus. “Latency” indicates that the eating-inhibitory effect was much faster (i.e., a few hours) than thatof peripherally administered estradiol (i.e., �2 days), as described in text. “ER�/c-Fos colocalization” indicates that food stimuli elicited neural activity, asindicated by c-Fos immunocytochemistry, in ER�-expressing neurons.
Review
R1231SEX DIFFERENCES IN THE PHYSIOLOGY OF EATING
AJP-Regul Integr Comp Physiol • doi:10.1152/ajpregu.00446.2012 • www.ajpregu.org
by 10.220.32.247 on Septem
ber 18, 2016http://ajpregu.physiology.org/
Dow
nloaded from
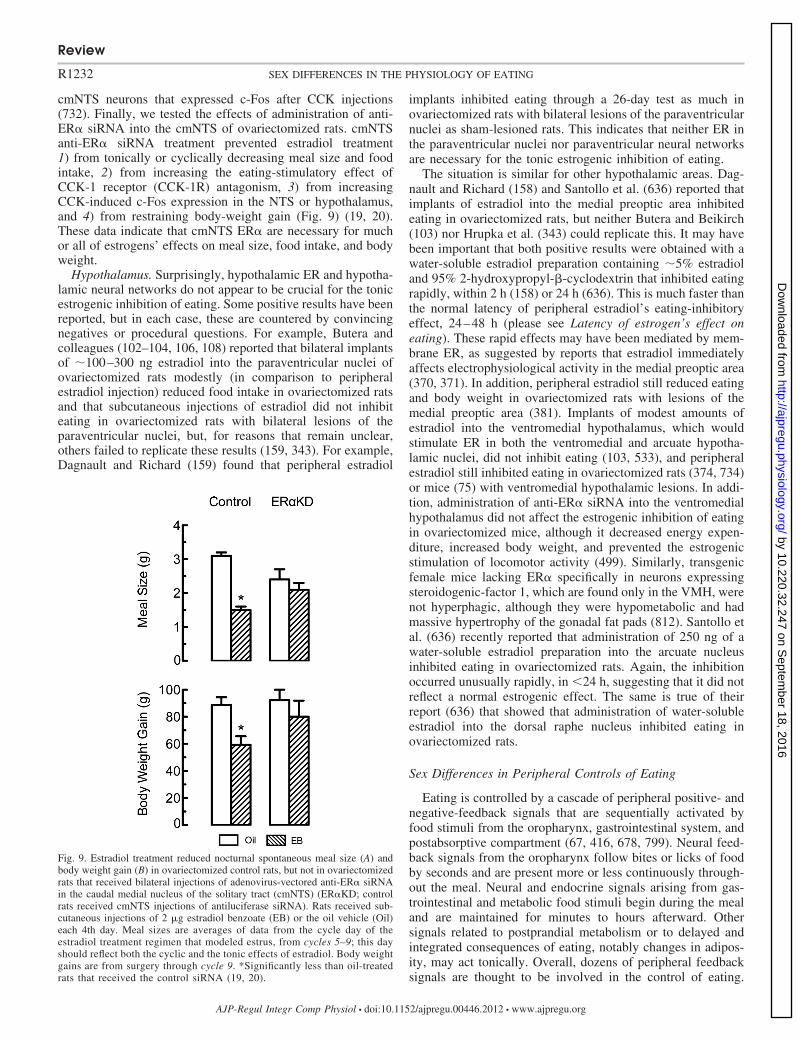
cmNTS neurons that expressed c-Fos after CCK injections(732). Finally, we tested the effects of administration of anti-ER� siRNA into the cmNTS of ovariectomized rats. cmNTSanti-ER� siRNA treatment prevented estradiol treatment1) from tonically or cyclically decreasing meal size and foodintake, 2) from increasing the eating-stimulatory effect ofCCK-1 receptor (CCK-1R) antagonism, 3) from increasingCCK-induced c-Fos expression in the NTS or hypothalamus,and 4) from restraining body-weight gain (Fig. 9) (19, 20).These data indicate that cmNTS ER� are necessary for muchor all of estrogens’ effects on meal size, food intake, and bodyweight.
Hypothalamus. Surprisingly, hypothalamic ER and hypotha-lamic neural networks do not appear to be crucial for the tonicestrogenic inhibition of eating. Some positive results have beenreported, but in each case, these are countered by convincingnegatives or procedural questions. For example, Butera andcolleagues (102–104, 106, 108) reported that bilateral implantsof �100–300 ng estradiol into the paraventricular nuclei ofovariectomized rats modestly (in comparison to peripheralestradiol injection) reduced food intake in ovariectomized ratsand that subcutaneous injections of estradiol did not inhibiteating in ovariectomized rats with bilateral lesions of theparaventricular nuclei, but, for reasons that remain unclear,others failed to replicate these results (159, 343). For example,Dagnault and Richard (159) found that peripheral estradiol
implants inhibited eating through a 26-day test as much inovariectomized rats with bilateral lesions of the paraventricularnuclei as sham-lesioned rats. This indicates that neither ER inthe paraventricular nuclei nor paraventricular neural networksare necessary for the tonic estrogenic inhibition of eating.
The situation is similar for other hypothalamic areas. Dag-nault and Richard (158) and Santollo et al. (636) reported thatimplants of estradiol into the medial preoptic area inhibitedeating in ovariectomized rats, but neither Butera and Beikirch(103) nor Hrupka et al. (343) could replicate this. It may havebeen important that both positive results were obtained with awater-soluble estradiol preparation containing �5% estradioland 95% 2-hydroxypropyl-�-cyclodextrin that inhibited eatingrapidly, within 2 h (158) or 24 h (636). This is much faster thanthe normal latency of peripheral estradiol’s eating-inhibitoryeffect, 24–48 h (please see Latency of estrogen’s effect oneating). These rapid effects may have been mediated by mem-brane ER, as suggested by reports that estradiol immediatelyaffects electrophysiological activity in the medial preoptic area(370, 371). In addition, peripheral estradiol still reduced eatingand body weight in ovariectomized rats with lesions of themedial preoptic area (381). Implants of modest amounts ofestradiol into the ventromedial hypothalamus, which wouldstimulate ER in both the ventromedial and arcuate hypotha-lamic nuclei, did not inhibit eating (103, 533), and peripheralestradiol still inhibited eating in ovariectomized rats (374, 734)or mice (75) with ventromedial hypothalamic lesions. In addi-tion, administration of anti-ER� siRNA into the ventromedialhypothalamus did not affect the estrogenic inhibition of eatingin ovariectomized mice, although it decreased energy expen-diture, increased body weight, and prevented the estrogenicstimulation of locomotor activity (499). Similarly, transgenicfemale mice lacking ER� specifically in neurons expressingsteroidogenic-factor 1, which are found only in the VMH, werenot hyperphagic, although they were hypometabolic and hadmassive hypertrophy of the gonadal fat pads (812). Santollo etal. (636) recently reported that administration of 250 ng of awater-soluble estradiol preparation into the arcuate nucleusinhibited eating in ovariectomized rats. Again, the inhibitionoccurred unusually rapidly, in �24 h, suggesting that it did notreflect a normal estrogenic effect. The same is true of theirreport (636) that showed that administration of water-solubleestradiol into the dorsal raphe nucleus inhibited eating inovariectomized rats.
Sex Differences in Peripheral Controls of Eating
Eating is controlled by a cascade of peripheral positive- andnegative-feedback signals that are sequentially activated byfood stimuli from the oropharynx, gastrointestinal system, andpostabsorptive compartment (67, 416, 678, 799). Neural feed-back signals from the oropharynx follow bites or licks of foodby seconds and are present more or less continuously through-out the meal. Neural and endocrine signals arising from gas-trointestinal and metabolic food stimuli begin during the mealand are maintained for minutes to hours afterward. Othersignals related to postprandial metabolism or to delayed andintegrated consequences of eating, notably changes in adipos-ity, may act tonically. Overall, dozens of peripheral feedbacksignals are thought to be involved in the control of eating.
Fig. 9. Estradiol treatment reduced nocturnal spontaneous meal size (A) andbody weight gain (B) in ovariectomized control rats, but not in ovariectomizedrats that received bilateral injections of adenovirus-vectored anti-ER� siRNAin the caudal medial nucleus of the solitary tract (cmNTS) (ER�KD; controlrats received cmNTS injections of antiluciferase siRNA). Rats received sub-cutaneous injections of 2 �g estradiol benzoate (EB) or the oil vehicle (Oil)each 4th day. Meal sizes are averages of data from the cycle day of theestradiol treatment regimen that modeled estrus, from cycles 5–9; this dayshould reflect both the cyclic and the tonic effects of estradiol. Body weightgains are from surgery through cycle 9. *Significantly less than oil-treatedrats that received the control siRNA (19, 20).
Review
R1232 SEX DIFFERENCES IN THE PHYSIOLOGY OF EATING
AJP-Regul Integr Comp Physiol • doi:10.1152/ajpregu.00446.2012 • www.ajpregu.org
by 10.220.32.247 on Septem
ber 18, 2016http://ajpregu.physiology.org/
Dow
nloaded from
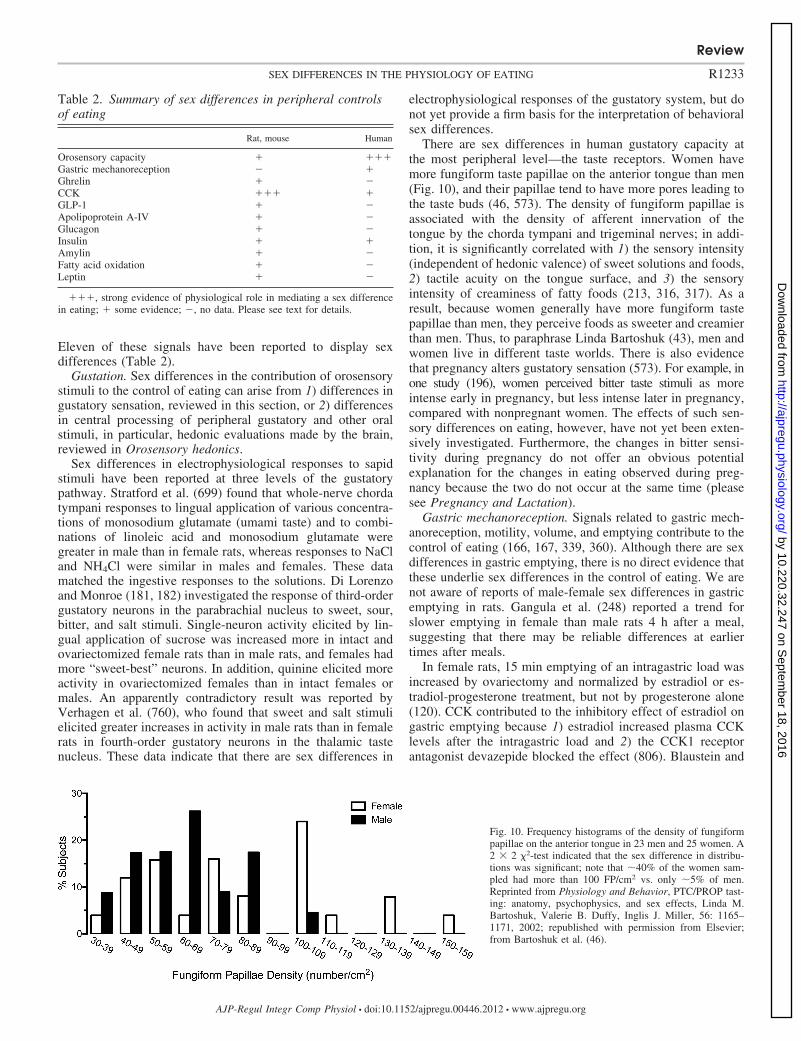
Eleven of these signals have been reported to display sexdifferences (Table 2).
Gustation. Sex differences in the contribution of orosensorystimuli to the control of eating can arise from 1) differences ingustatory sensation, reviewed in this section, or 2) differencesin central processing of peripheral gustatory and other oralstimuli, in particular, hedonic evaluations made by the brain,reviewed in Orosensory hedonics.
Sex differences in electrophysiological responses to sapidstimuli have been reported at three levels of the gustatorypathway. Stratford et al. (699) found that whole-nerve chordatympani responses to lingual application of various concentra-tions of monosodium glutamate (umami taste) and to combi-nations of linoleic acid and monosodium glutamate weregreater in male than in female rats, whereas responses to NaCland NH4Cl were similar in males and females. These datamatched the ingestive responses to the solutions. Di Lorenzoand Monroe (181, 182) investigated the response of third-ordergustatory neurons in the parabrachial nucleus to sweet, sour,bitter, and salt stimuli. Single-neuron activity elicited by lin-gual application of sucrose was increased more in intact andovariectomized female rats than in male rats, and females hadmore “sweet-best” neurons. In addition, quinine elicited moreactivity in ovariectomized females than in intact females ormales. An apparently contradictory result was reported byVerhagen et al. (760), who found that sweet and salt stimulielicited greater increases in activity in male rats than in femalerats in fourth-order gustatory neurons in the thalamic tastenucleus. These data indicate that there are sex differences in
electrophysiological responses of the gustatory system, but donot yet provide a firm basis for the interpretation of behavioralsex differences.
There are sex differences in human gustatory capacity atthe most peripheral level—the taste receptors. Women havemore fungiform taste papillae on the anterior tongue than men(Fig. 10), and their papillae tend to have more pores leading tothe taste buds (46, 573). The density of fungiform papillae isassociated with the density of afferent innervation of thetongue by the chorda tympani and trigeminal nerves; in addi-tion, it is significantly correlated with 1) the sensory intensity(independent of hedonic valence) of sweet solutions and foods,2) tactile acuity on the tongue surface, and 3) the sensoryintensity of creaminess of fatty foods (213, 316, 317). As aresult, because women generally have more fungiform tastepapillae than men, they perceive foods as sweeter and creamierthan men. Thus, to paraphrase Linda Bartoshuk (43), men andwomen live in different taste worlds. There is also evidencethat pregnancy alters gustatory sensation (573). For example, inone study (196), women perceived bitter taste stimuli as moreintense early in pregnancy, but less intense later in pregnancy,compared with nonpregnant women. The effects of such sen-sory differences on eating, however, have not yet been exten-sively investigated. Furthermore, the changes in bitter sensi-tivity during pregnancy do not offer an obvious potentialexplanation for the changes in eating observed during preg-nancy because the two do not occur at the same time (pleasesee Pregnancy and Lactation).
Gastric mechanoreception. Signals related to gastric mech-anoreception, motility, volume, and emptying contribute to thecontrol of eating (166, 167, 339, 360). Although there are sexdifferences in gastric emptying, there is no direct evidence thatthese underlie sex differences in the control of eating. We arenot aware of reports of male-female sex differences in gastricemptying in rats. Gangula et al. (248) reported a trend forslower emptying in female than male rats 4 h after a meal,suggesting that there may be reliable differences at earliertimes after meals.
In female rats, 15 min emptying of an intragastric load wasincreased by ovariectomy and normalized by estradiol or es-tradiol-progesterone treatment, but not by progesterone alone(120). CCK contributed to the inhibitory effect of estradiol ongastric emptying because 1) estradiol increased plasma CCKlevels after the intragastric load and 2) the CCK1 receptorantagonist devazepide blocked the effect (806). Blaustein and
Table 2. Summary of sex differences in peripheral controlsof eating
Rat, mouse Human
Orosensory capacity � ���Gastric mechanoreception �Ghrelin � CCK ��� �GLP-1 � Apolipoprotein A-IV � Glucagon � Insulin � �Amylin � Fatty acid oxidation � Leptin �
���, strong evidence of physiological role in mediating a sex differencein eating; � some evidence; , no data. Please see text for details.
Fig. 10. Frequency histograms of the density of fungiformpapillae on the anterior tongue in 23 men and 25 women. A2 � 2 2-test indicated that the sex difference in distribu-tions was significant; note that �40% of the women sam-pled had more than 100 FP/cm2 vs. only �5% of men.Reprinted from Physiology and Behavior, PTC/PROP tast-ing: anatomy, psychophysics, and sex effects, Linda M.Bartoshuk, Valerie B. Duffy, Inglis J. Miller, 56: 1165–1171, 2002; republished with permission from Elsevier;from Bartoshuk et al. (46).
Review
R1233SEX DIFFERENCES IN THE PHYSIOLOGY OF EATING
AJP-Regul Integr Comp Physiol • doi:10.1152/ajpregu.00446.2012 • www.ajpregu.org
by 10.220.32.247 on Septem
ber 18, 2016http://ajpregu.physiology.org/
Dow
nloaded from

Wade (76) did not see an effect of estradiol on gastric empty-ing; whether this was because they tested anesthetized rats oranother procedural difference is not clear. Gastric emptyingwas also increased in lactating rats (121). In one study, neitherorchiectomy nor testosterone treatment affected gastric emp-tying in male rats (120).
In contrast to the lack of male-female sex differences ingastric emptying in rats, women empty both solids and liquidsfrom the stomach significantly more slowly than men (60, 163,349, 399). Differences often appear soon enough (i.e., �30min) to potentially affect within-meal control of eating. In onestudy (349), a similar male-female sex difference in gastricemptying of solid food was found for postmenopausal womenwho took estrogen-progestin hormone treatments, but not forpostmenopausal women who did not. Whether the ovariancycle affects emptying is not clear: 1) Gill et al. (270) foundthat solid food, but not liquid food, emptied from the stomachfaster during follicular phase than the luteal phase; 2) Brennanet al. (85) found that liquid food emptied from the stomachfaster during the follicular phase; and 3) Horowitz et al. (340)failed to find cycle differences in emptying of either solid orliquid food. Eating was measured in only one of these tests.Brennan et al. (85) reported that oral loads of 50 g glucose in300 ml water emptied from the stomach �15–20% moreslowly and that intake during a meal 90 min later was �700 kJless during the midfollicular phase than the midluteal phase innormal-weight women. If the food ingested during these mealsalso emptied more slowly, the decrease in meal size is unlikelyto have resulted from feedback effects of postgastric foodstimuli. Thus, these data suggest that the potency of satiationsignals related to gastric mechanoreception varies during theovarian cycle in women.
Ghrelin. Ghrelin is unique among gut peptides in that itssecretion increases during intermeal intervals and that ghrelinadministration increases eating, apparently by acting centrally(383). Clegg et al. (130) reported that both intraperitoneal andintraventricular (third ventricle) ghrelin administration in-creased intake significantly more in intact male rats and un-treated ovariectomized female rats than intact female rats orestradiol-treated ovariectomized rats. Furthermore, the eating-stimulatory effect of both peripheral and central ghrelin ad-
ministration varied cyclically in both intact, cycling rats andovariectomized rats cyclically treated with estradiol, such thatghrelin was more efficacious during diestrus 1 and 2 thanduring proestrus or estrus (Fig. 11). Butera (102) found similarchanges in the eating-stimulatory potency of peripheral ghre-lin. These data suggest that ghrelin may be necessary for boththe tonic and the cyclic estrogenic controls of eating. Oneaspect of the ghrelin’s effect in ovariectomized rats, however,is inconsistent with this conclusion and requires resolution.Ghrelin’s effects in ovariectomized rats were due to changes inmeal frequency, not in meal size (102, 130), whereas estrogensaffect meal size, not meal frequency (please see Ovariectomyand hormone treatment).
Sex differences in ghrelin’s eating-stimulatory effect may bemediated by changes in ghrelin secretion, as well as changes inghrelin sensitivity. Female rat gastric mucosal cells that areimmunopositive for ghrelin also express ER�, and ovariec-tomy temporarily increases both the number of cells immuno-positive for ghrelin and plasma ghrelin level (130, 461, 626).The time courses of the postovariectomy increases in plasmaghrelin level and in eating were strikingly similar (130), againconsistent with a tonic estrogenic effect of ghrelin on eating inrats. Ghrelin levels have not been determined through theestrous cycle, and no changes in plasma levels of active ghrelinwere detected in normally cycling women (157).
How estrogens influence ghrelin’s eating-stimulatory effect inmice is not clear. Clegg et al. (130) reported that ovariectomy didnot increase food intake or body weight in transgenic mice withnull mutations of the ghrelin receptor, suggesting a necessary rolefor estrogens. Maletínská et al. (453), however, failed to detectany influence of estradiol treatment on the eating-inhibitory andweight-reducing effects of twice daily subcutaneous injections ofthe ghrelin antagonist [D-Lys3]GHRP-6 in ovariectomized micefed chow or high-fat diet. A parsimonious potential resolution ofthese data is that estrogens act during early development to reducethe eating-stimulatory effect of ghrelin in adult mice.
Ghrelin levels are positively associated with testosteronelevels in men and estrogen levels in women, and ghrelin levelsincreased both in hypogonadal men treated with testosterone(532) and postmenopausal women treated with estrogens (369,541). These findings, however, have not yet been linked to thecontrol of eating.
Cholecystokinin. CCK is an important controller of meal-ending satiation in animals and humans (52, 416, 591, 677).CCK secretion increases during meals, and premeal adminis-tration of CCK-1R antagonists increased meal size in bothmale rats and men. CCK-1R antagonism also reduces oreliminates the satiating action of intraduodenal infusions ofseveral nutrients, including, in male rats, fats, proteins, andsome carbohydrates and, in humans, fats. CCK acts locally inthe abdomen to initiate a vagal afferent signal that is transmit-ted to the NTS.
RATS AND MICE. There is extensive evidence that an activa-tional effect of estrogens increases CCK satiation in female rats(16–18, 102, 204, 345, 435, 455, 774). 1) In intact female rats,intraperitoneal CCK injection inhibited eating during proestrusor estrus, but not during diestrus, and the desatiating effect ofCCK-1R antagonism was larger during estrus than duringdiestrus. 2) The de-satiating effect of CCK-1R antagonism wasevident only after puberty in intact females. In ovariectomizedrats, estradiol treatment 3) increased the satiating effect of
Fig. 11. Cyclic changes in the eating-stimulatory effect of ghrelin in rats. Theestrous cycle was monitored in intact rats (D1, diestrus 1; D2, diestrus 2; P,proestrus; E, estrus), and third cerebroventricular injections of 0.1 nmol ofghrelin or saline were tested on each cycle day. Ghrelin stimulated eating onlyon D1 and D2. *Significantly different from saline, same cycle day. Reprintedwith permission from the American Diabetes Association from Diabetes,Deborah J. Clegg, Lynda M. Brown, Jeffrey M. Zigman, Christopher J. Kemp,April D. Strader, Stephen C. Benoit, Stephen C. Woods, Michela Mangia-racina, and Nori Geary, 56: April 2007; from Clegg et al. (130).
Review
R1234 SEX DIFFERENCES IN THE PHYSIOLOGY OF EATING
AJP-Regul Integr Comp Physiol • doi:10.1152/ajpregu.00446.2012 • www.ajpregu.org
by 10.220.32.247 on Septem
ber 18, 2016http://ajpregu.physiology.org/
Dow
nloaded from
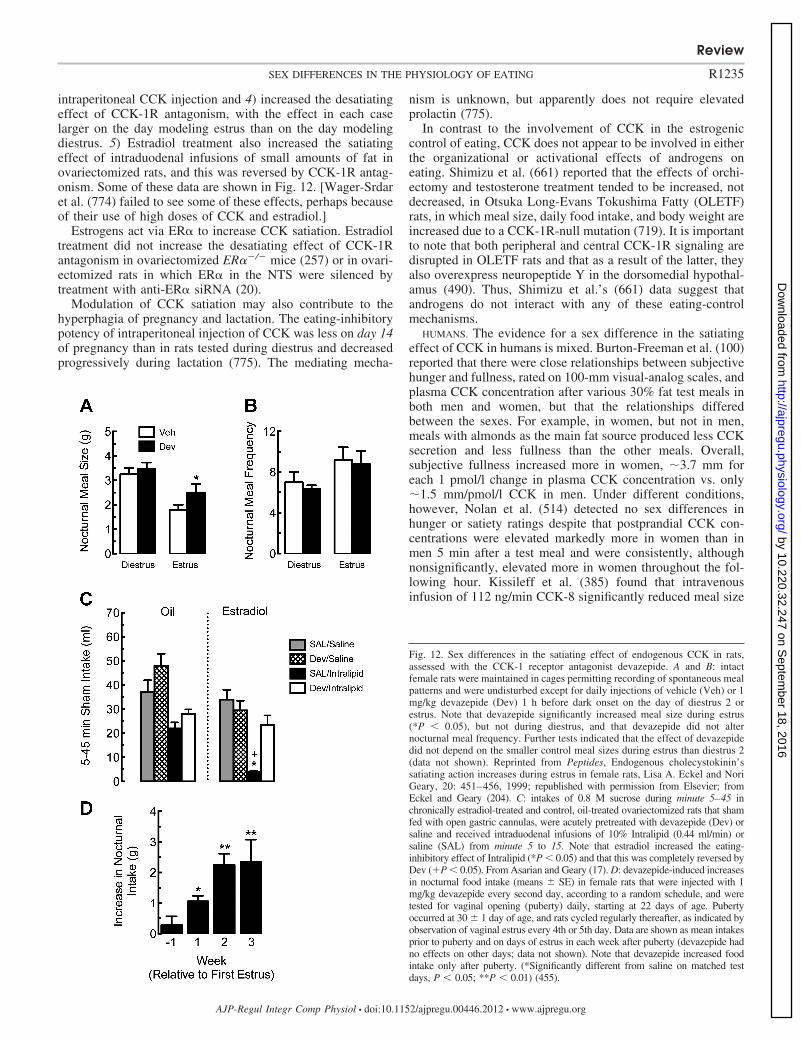
intraperitoneal CCK injection and 4) increased the desatiatingeffect of CCK-1R antagonism, with the effect in each caselarger on the day modeling estrus than on the day modelingdiestrus. 5) Estradiol treatment also increased the satiatingeffect of intraduodenal infusions of small amounts of fat inovariectomized rats, and this was reversed by CCK-1R antag-onism. Some of these data are shown in Fig. 12. [Wager-Srdaret al. (774) failed to see some of these effects, perhaps becauseof their use of high doses of CCK and estradiol.]
Estrogens act via ER� to increase CCK satiation. Estradioltreatment did not increase the desatiating effect of CCK-1Rantagonism in ovariectomized ER�/ mice (257) or in ovari-ectomized rats in which ER� in the NTS were silenced bytreatment with anti-ER� siRNA (20).
Modulation of CCK satiation may also contribute to thehyperphagia of pregnancy and lactation. The eating-inhibitorypotency of intraperitoneal injection of CCK was less on day 14of pregnancy than in rats tested during diestrus and decreasedprogressively during lactation (775). The mediating mecha-
nism is unknown, but apparently does not require elevatedprolactin (775).
In contrast to the involvement of CCK in the estrogeniccontrol of eating, CCK does not appear to be involved in eitherthe organizational or activational effects of androgens oneating. Shimizu et al. (661) reported that the effects of orchi-ectomy and testosterone treatment tended to be increased, notdecreased, in Otsuka Long-Evans Tokushima Fatty (OLETF)rats, in which meal size, daily food intake, and body weight areincreased due to a CCK-1R-null mutation (719). It is importantto note that both peripheral and central CCK-1R signaling aredisrupted in OLETF rats and that as a result of the latter, theyalso overexpress neuropeptide Y in the dorsomedial hypothal-amus (490). Thus, Shimizu et al.’s (661) data suggest thatandrogens do not interact with any of these eating-controlmechanisms.
HUMANS. The evidence for a sex difference in the satiatingeffect of CCK in humans is mixed. Burton-Freeman et al. (100)reported that there were close relationships between subjectivehunger and fullness, rated on 100-mm visual-analog scales, andplasma CCK concentration after various 30% fat test meals inboth men and women, but that the relationships differedbetween the sexes. For example, in women, but not in men,meals with almonds as the main fat source produced less CCKsecretion and less fullness than the other meals. Overall,subjective fullness increased more in women, �3.7 mm foreach 1 pmol/l change in plasma CCK concentration vs. only�1.5 mm/pmol/l CCK in men. Under different conditions,however, Nolan et al. (514) detected no sex differences inhunger or satiety ratings despite that postprandial CCK con-centrations were elevated markedly more in women than inmen 5 min after a test meal and were consistently, althoughnonsignificantly, elevated more in women throughout the fol-lowing hour. Kissileff et al. (385) found that intravenousinfusion of 112 ng/min CCK-8 significantly reduced meal size
Fig. 12. Sex differences in the satiating effect of endogenous CCK in rats,assessed with the CCK-1 receptor antagonist devazepide. A and B: intactfemale rats were maintained in cages permitting recording of spontaneous mealpatterns and were undisturbed except for daily injections of vehicle (Veh) or 1mg/kg devazepide (Dev) 1 h before dark onset on the day of diestrus 2 orestrus. Note that devazepide significantly increased meal size during estrus(*P � 0.05), but not during diestrus, and that devazepide did not alternocturnal meal frequency. Further tests indicated that the effect of devazepidedid not depend on the smaller control meal sizes during estrus than diestrus 2(data not shown). Reprinted from Peptides, Endogenous cholecystokinin’ssatiating action increases during estrus in female rats, Lisa A. Eckel and NoriGeary, 20: 451–456, 1999; republished with permission from Elsevier; fromEckel and Geary (204). C: intakes of 0.8 M sucrose during minute 5–45 inchronically estradiol-treated and control, oil-treated ovariectomized rats that shamfed with open gastric cannulas, were acutely pretreated with devazepide (Dev) orsaline and received intraduodenal infusions of 10% Intralipid (0.44 ml/min) orsaline (SAL) from minute 5 to 15. Note that estradiol increased the eating-inhibitory effect of Intralipid (*P � 0.05) and that this was completely reversed byDev (�P � 0.05). From Asarian and Geary (17). D: devazepide-induced increasesin nocturnal food intake (means SE) in female rats that were injected with 1mg/kg devazepide every second day, according to a random schedule, and weretested for vaginal opening (puberty) daily, starting at 22 days of age. Pubertyoccurred at 30 1 day of age, and rats cycled regularly thereafter, as indicated byobservation of vaginal estrus every 4th or 5th day. Data are shown as mean intakesprior to puberty and on days of estrus in each week after puberty (devazepide hadno effects on other days; data not shown). Note that devazepide increased foodintake only after puberty. (*Significantly different from saline on matched testdays, P � 0.05; **P � 0.01) (455).
Review
R1235SEX DIFFERENCES IN THE PHYSIOLOGY OF EATING
AJP-Regul Integr Comp Physiol • doi:10.1152/ajpregu.00446.2012 • www.ajpregu.org
by 10.220.32.247 on Septem
ber 18, 2016http://ajpregu.physiology.org/
Dow
nloaded from

in women (�120 g), but not in men (�70 g) but, again, the sexdifference was not significant. It is possible that the apparentsex difference in this study was due to differential effects of theCCK infusions on plasma CCK levels because identical doseswere given to all subjects, although women weighed less (59vs. 72 kg). Unfortunately, none of these three studies moni-tored menstrual cycling, which, as the studies discussed belowsuggest, may have added significant variability to the results.
Two attempts to detect cyclic changes in endogenous CCKsatiation in women produced encouraging data. Pohle-Krauzaet al. (561) had women ingest 10 g L-phenylalanine, which is apotent CCK secretogogue in humans (although not in rats), 20min prior to lunch and dinner buffet meals in the midfollicularand the midluteal phases. In 11 subjects with low rigid-restraint[i.e., scores of 0 or 1 in the rigid-restraint subscale of thethree-factor eating questionnaire (703)], L-phenylalanine re-duced total daily food intake significantly more in the follicularphase than in the luteal phase. Surprisingly, just the oppositeoccurred in the 9 women with high rigid-restraint scores. Thesedata suggest that CCK, ovarian cycling, and cognitive restraintmay interact in the control of eating in women. Whether theseeffects were related to differences in CCK secretion was notdetermined. Brennan et al. (85), in the study reviewed justabove (please see Gastric mechanoreception), found no cyclicchange in postprandial CCK levels after a test meal. As gastricemptying was significantly reduced during the follicular phase,however, it is possible that intraintestinal nutrient stimulistimulated CCK secretion more effectively during the follicularphase.
VAGAL MECHANISMS. That estradiol increases the satiatingpotency of exogenous CCK in ovariectomized rats (105, 259)indicates that estrogens change the neural processing of CCKafferent signaling to modulate CCK satiation rather than in-creasing CCK secretion. Such altered processing could occurdue to changes in CCK-1 receptor function in vagal afferentterminals in the abdomen. Because most CCK receptors in theNTS are expressed on the central terminals of vagal afferents(136, 491), we estimated changes in abdominal vagal CCK-1receptor function by measuring estradiol’s effects on CCK-1receptor function in the NTS (258). Neither CCK-1 receptornumber nor affinity was altered. Thus, estrogen signalingapparently either 1) increases transduction of CCK-1 receptoractivation into a vagal afferent signal (hypothesis 1) or2) modulates processing of this signal in the NTS or furtherdownstream (hypothesis 2). The findings that female rats have�50% more myelinated vagal afferent fibers than males due tothe presence of a specific low-threshold fiber type and that theexcitability of these fibers is estrogen-dependent (430, 575) areconsistent with hypothesis 1. On the other hand, the necessityof ER� expression in the NTS for the estrogenic modulation ofCCK satiation (please see cmNTS) strongly supports hypothesis2. Perhaps both mechanisms contribute.
Because lesion of abdominal vagal afferents eliminates thesatiating actions of exogenous and endogenous CCK in malerats, vagotomy should also interfere with the estrogenic inhi-bition of eating. This has been tested once, and the result wasnegative. That is, ovariectomy still increased body weight, andsubsequent estradiol treatment still decreased body weight, inrats with total subdiaphragmatic vagotomies (212) The reasonfor this apparent discrepancy is unclear. It may have beenimportant that neither meal patterns nor food intake was
measured in this study or that, because of the debilitatingeffects of total vagotomy (405), all the vagotomized ratsmaintained body weights well below those of control rats. It isalso possible that the apparent discrepancy is due to compen-satory effects of other estrogen-related but vagally independentsignals.
Glucagon-like peptide-1. Glucagon-like peptide-1 (GLP-1)acting in peripheral paracrine or endocrine and central neuro-crine modes appears to contribute to the physiological controlof meal size in rats and humans (39, 53, 318, 319, 791).Estradiol increased both the satiating potency of intraperitonealinjections of GLP-1 (Asarian, L, unpublished data) and thedesatiating potency of intraperitoneal injections of the GLP-1receptor-antagonist exendin-9 in ovariectomized rats (13). Weassume that this is related to the satiating action of peripheralGLP-1 because, although central GLP-1 neurons are located inthe cmNTS, they do not express ER� and do not express c-Fosin response to intraduodenal infusions of Intralipid (Asarian, L,unpublished data). As GLP-1 agonists are attractive candidatesfor obesity therapy (21, 798), these initial findings certainlymerit pursuit.
Apolipoprotein A-IV. Apolipoprotein A-IV is synthesized inthe brain, as well as in the intestines, and appears to actphysiologically to inhibit eating (744). A study by Shen et al.(657) indicates that apolipoprotein A-IV signaling in the NTSmay contribute to the estrogenic inhibition of eating: 1) ovari-ectomy increased eating and weight gain more in mice withnull mutations of the apolipoprotein A-IV gene than in wild-type mice; 2) cyclic estradiol treatment increased the eating-inhibitory effect of an acute injection of apolipoprotein A-IVinto the fourth cerebral ventricle of ovariectomized rats; and3) ovariectomy decreased expression of apolipoprotein A-IVmRNA in the NTS of rats. In addition, apolipoprotein A-IVappeared to interact with CCK signaling to produce satiation inmale rats (438), suggesting the interesting hypothesis that thesame NTS estrogen-signaling pathway may modulate bothCCK and apolipoprotein A-IV satiation.
Glucagon. Glucagon is released from the pancreas duringmeals, apparently largely due to cephalic phase reflexes, andcontributes to the physiological control of meal size in rats andperhaps in humans (300, 801). In ovariectomized rats, estradioltreatment increased both the satiating potency of intramealhepatic portal infusions of glucagon and the desatiating po-tency of antagonism of endogenous glucagon by hepatic portalinfusion of glucagon antibodies (256). Whether this reflected acyclic or a tonic effect of estradiol, and whether similarphenomena occur in cycling rats was not determined. There isalso a sex difference in glucagon secretion in humans that maybe relevant to eating. Both basal plasma glucagon concentra-tion and prandial glucagon concentrations were approximatelytwofold higher in normal-weight or obese men than in normal-weight or obese women (112). These data indicate that therenewed interest in glucagon as a pharmacological tool in thetreatment of obesity (300) should include studies of sex dif-ferences in its eating-inhibitory actions.
Insulin. Basal plasma levels of insulin correlate with bodyfat mass, and tonic insulin signaling is hypothesized to act as anegative-feedback signal linking adiposity to the central ner-vous system control of energy homeostasis, i.e., to be anadiposity signal (91, 801), although the hypothesis is increas-ingly controversial (275, 311, 331, 799).
Review
R1236 SEX DIFFERENCES IN THE PHYSIOLOGY OF EATING
AJP-Regul Integr Comp Physiol • doi:10.1152/ajpregu.00446.2012 • www.ajpregu.org
by 10.220.32.247 on Septem
ber 18, 2016http://ajpregu.physiology.org/
Dow
nloaded from

RATS AND MICE. Clegg et al. (129, 131) reported a male-female sex difference in the eating-inhibitory effect of insulin;i.e., that injection of 1 or 4 mIU insulin into the third cerebralventricle inhibited eating more in male rats than in age- orweight-matched females. Keen-Rhinehart et al. (368), how-ever, saw no such sex difference in tests of peripubertal maleand female rats, which they speculated may have been due tolower endogenous estrogen levels in the young rats. To add tothe uncertainty, Brüning et al. (91) reported that female, but notmale, transgenic mice with brain-specific-null mutations of theinsulin receptor were hyperphagic. This effect may have beensecondary to different degrees of disruptions of the HPG axis:female knockout mice had a �90% reduction in plasma LHconcentration, vs. a �40% reduction in males.
Clegg et al. (129) also reported that both central (20 ngestradiol benzoate once each 4th day into the third ventricle)and subcutaneous (2 �g estradiol benzoate once each 4th day)estradiol treatments decreased the inhibition of eating producedby central injection of 8 mIU insulin in ovariectomized rats.These surprising data suggest that an activational influence ofestrogens dampens insulin’s anorectic potency; i.e., that insulinopposes the physiological estrogenic inhibition of eating.Where in the brain the injected hormones acted was notdetermined.
HUMANS. Hallschmid and colleagues (59, 305) reported amale-female sex difference in insulin’s effects in humanssimilar to that reported by Clegg et al. (129, 131) in rats. In onestudy (59), normal-weight (mean BMI, �21–23) subjects self-administered 160 IU regular human insulin or the vehicleintranasally before eating a buffet breakfast; women usedestrogen-dominant contraceptives and were tested with aninterval of 28 days to control for cyclic effects on eating. Insulinsignificantly decreased food intake in men, from 1,351 105 to1,159 96 kcal, but not in women, increasing from 769 44to 787 41 kcal, and the sex difference was significant.Interestingly, performance in a memory task was improved byinsulin in women but not in men. This was followed by an 8wk, placebo-controlled trial of four daily intranasal insulintreatments, once before each meal and again before bedtime(305). Insulin significantly reduced body fat mass, estimated bybioelectric impedance, �1.4 kg in men and had no detectableeffect in women. A variety of neuroendocrine, cardiovascular,metabolic, and subjective measures suggested that these effectswere not secondary to deleterious side effects, that the subjectscould not detect whether they had received insulin or placebo,and that insulin selectively reduced premeal hunger. A simi-larly designed trial in obese men indicated that althoughchronic insulin treatment had the same cognitive effects inobese men as had been found in the normal-weight men, itfailed to reduce body weight, suggesting that obesity mayinduce a kind of insulin resistance in the neural networksregulating body weight (304).
Amylin. Amylin, a peptide cosecreted with insulin from thepancreatic �-cells, has a physiological eating-inhibitory action,and amylin agonists are promising antiobesity agents (446,447, 618, 619, 801). Two tests of the estrogenic modulation ofamylin’s eating-inhibitory effects produced opposite results.Trevaskis et al. (742) reported that cyclic estradiol treatmentmarkedly reduced the potency of chronic subcutaneous infu-sion of 50 �g·kg1·day1 amylin to reduce eating and bodyweight in ovariectomized rats. On the other hand, Asarian et al.
(14) found that in acute eating tests, estradiol cyclically in-creased the satiating potency of exogenous amylin and thedesatiating potency of amylin antagonism. Estradiol also in-creased the number of dopamine �-hydroxylase-positive cellsin the area postrema that expressed c-Fos after amylin injec-tion, consistent with the catecholaminergic mediation of amy-lin’s satiating effect. Especially in view of amylin’s physio-logical role and therapeutic potential, it is important to resolvethe contrasting outcomes of these studies.
Metabolic signals. A distributed network of metabolite- andhormone-sensing neurons in the brain and periphery contrib-utes to energy homeostasis, in part via the control of eating (69,416, 427). A relatively small amount of data suggests that thereare sex differences in the metabolic control of eating. Wadeand Gray (767) proposed in 1979 that estrogens control eatingindirectly by increasing the flux of energy metabolites from theadipose tissue into the circulation, from which they reachhepatic or brain metabolic receptors. Attempts to provideexperimental support for this hypothesis failed (287, 517, 533,582, 583), however, and they soon abandoned it (768). Morerecent experiments with metabolic inhibitors have producedinconsistent results. Swithers et al. (717, 718) found thatmercaptoacetate, which antagonizes fatty-acid oxidation, in-creased eating in male rats, but not females, whereas Sandovalet al. (632) found that mercaptoacetate was more effective infemales than males. The cause for this discrepancy is unclear.Sandoval et al. (632) also found a sex difference related toglucose metabolism: 2-deoxy-D-glucose (2DG), which inhibitsglycolysis, elicited more eating in male than female rats.Unexpectedly, both estradiol-treated and untreated ovariecto-mized females were more sensitive to 2DG than intact females(632), which contrasts with an earlier report (469) that estradioltreatment decreased the eating-stimulatory effect of 2DG inovariectomized rats. Thus, more research is required to deter-mine whether there are sex differences in metabolic controls ofeating, and if so, whether they are estrogen-regulated.
Leptin. Basal plasma levels of leptin, like those of insulin,are tightly correlated with body fat mass and are thought tosignal adiposity status to the brain and to initiate compensatorychanges in eating and energy expenditure, especially in thecase of reduced adiposity (218, 239, 422, 501). Dissecting thenormal role of endogenous leptin in energy homeostasis hasbeen far more difficult than was expected on the basis of theexaggerated obesity phenotype exhibited by mice, rats orhumans with null mutations in the genes encoding leptin or itssignaling receptor, leptin receptor-b (252, 331, 799). Never-theless, there are now several lines of evidence that leptintonically inhibits eating in mice, rats, and humans (1, 301, 310,400, 418, 459, 614, 755, 824). It does not appear that thiscontrol is sexually differentiated. Although there are clear sexdifferences in leptin secretion, there is little evidence that theseare related to sex differences in eating.
LEPTIN SECRETION. Basal plasma leptin levels are linearlyrelated to fat mass in both men and women, with obese andnever-obese persons falling on the same line (612, 613). Menand women have different relationships, however, with plasmaconcentrations of leptin per kilogram body fat mass markedlyhigher in women. Postmenopausal women have less basalplasma leptin per kilogram fat mass than premenopausalwomen, but more than men. Genetic differences, differences inandrogen and estrogen levels, and regional fat distribution all
Review
R1237SEX DIFFERENCES IN THE PHYSIOLOGY OF EATING
AJP-Regul Integr Comp Physiol • doi:10.1152/ajpregu.00446.2012 • www.ajpregu.org
by 10.220.32.247 on Septem
ber 18, 2016http://ajpregu.physiology.org/
Dow
nloaded from

appear to contribute to these sex differences in basal plasmaleptin levels, with no single factor alone sufficient (488, 598,612, 613).
Basal plasma leptin concentration is correlated with fat massin rats as well, but in contrast to humans, postpubertal male ratshave higher basal plasma leptin levels than postpubertal fe-males (415, 497, 683). The implication of this for leptinsignaling is not clear, however, because plasma concentrationof the soluble leptin receptor-e, which acts as a plasma bindingprotein that reduces leptin signaling, also is much higher inpostpubertal male rats than postpubertal female rats (681).
LEPTIN AND SEX DIFFERENCES IN EATING. No role for the sexdifferences in basal plasma leptin concentration describedabove in eating has been established. Acute injections of leptininto the third cerebral ventricle, however, inhibited eating morein female rats than male rats (2, 129, 131). This sex differencemay be related to activational effects of gonadal steroids. Inovariectomized rats, estradiol treatment increased leptin’s eat-ing-inhibitory potency (2, 129, 131) and upregulated leptinreceptor-b, the signaling form, in the hypothalamus (474, 595).The physiological relevance of these effects for leptin’s normaltonic eating-inhibitory effect, however, is uncertain. 1) Plasmaestradiol levels did not influence the effect of chronic leptintreatment on body fat mass in female mice (545) or rats (124).2) The eating-inhibitory potency of chronic leptin treatment didnot vary over the ovarian cycle in intact rats (208). 3) Normal-weight women ate less during the periovulatory phase thanduring the luteal phase, but leptin levels were higher in theluteal phase and were not correlated with eating during anyphase (536).
Androgens may affect leptin’s eating-inhibitory potency inthe opposite manner as estrogens. This is because exogenousleptin had smaller effects on eating and on nuclear transloca-tion of leptin-signal transducer and activator of transcription 3in Arc neurons in transgenic mice lacking AR than in malewild-type mice (215). Basal levels of food intake in theAR-knockout and in wild-type mice, however, were not dif-ferent (215).
Pregnant and lactating rats are both hyperleptinemic and, interms of the inhibition of eating, leptin resistant. This resis-tance may be due to increased prolactin levels (412, 743); i.e.,release from the tonic inhibitory effect of leptin on eating mayincrease food intake during pregnancy and lactation. In supportof this, Grattan and colleagues (25, 26, 285, 412, 413) showedthat 1) pregnancy-induced leptin resistance was associated withelevated neuropeptide Y mRNA and decreased POMC mRNAin the arcuate nucleus during pregnancy, 2) leptin receptormRNA and leptin-induced phosphorylated-signal transducerand activator of transcription-3 protein decreased in the VMHduring pregnancy, and 3) injection of leptin into the thirdcerebral ventricle that reduced daily food intake by �20% innonpregnant rats had no effect in pregnant rats or in pseudo-pregnant rats after intracerebroventricular infusions of prolac-tin that were designed to mimic the pattern of placentallactogen secretion that occurs during pregnancy. The develop-ment of leptin resistance during lactation and lack of effect ofleptin on eating during lactation were recently replicated bySuzuki et al. (713). A useful further step would be to verify thatprogressive decreases in endogenous leptin signaling are nec-essary for the progressively increased food intake during preg-nancy and lactation, as could be done, for example, using leptin
antagonism (459, 824). Changes in NPY and melanocortinsignaling in the dorsomedial hypothalamus were also impli-cated in pregnancy-induced hyperphagia (119).
Finally, the many physiological interactions between leptinand gonadal steroids, for example, in the early development ofneural architecture controlling adult behavior (493, 667, 805),in pubertal development (187, 563), and in nutritional infertil-ity (645, 769, 787), certainly justify further investigation oftheir interactions in the control of normal eating and adiposity(see Ref. 567 for further discussion).
Sex Differences in Central Mechanisms Controlling Eating
Neurochemical controls of eating. Neuropharmacologicalstudies, including molecular genetic manipulations of neuro-chemicals, implicate several brain signaling molecules in sexdifferences in eating. We review some of the most interestingdata. An unfortunate aspect of this literature is that, as yet, noconcrete links have been forged between the neurochemicalfindings reviewed here and the progress in identifying eitherthe sites of ER or the peripheral mechanisms mediating sexdifferences in eating reviewed above.
SEROTONIN (5-HYDROXYTRYPTAMINE, 5HT). Serotonergic neu-rotransmission plays integral roles in both satiation and foodreward (302, 315, 414, 416). Whether there are male-femaledifferences in the serotonergic control of eating is not clear. Inone study, fenfluramine inhibited eating in female rats morethan in male rats (209), whereas in another study, it did not(150). In contrast, there are several indications that estrogensaffect serotonin-induced eating. Eckel et al. (209) found thatthe eating-inhibitory effect of the 5HT agonist fenfluraminewas 1) larger in intact, cycling rats during estrus than duringdiestrus and 2) increased by estradiol treatment in ovariecto-mized rats (593). In addition, they found that intraperitonealand lateral cerebroventricular injections of meta-chlorophe-nylpiperazine, a selective antagonist of 5HT2C receptors, in-creased eating more in estradiol-treated than control ovariec-tomized rats (594). Souquet and Rowland (687), however,found no difference in the effect of chronic fenfluraminetreatment in estradiol-treated than control ovariectomized rats.Taken together, these data suggest that 5HT-5HT2C signalingis involved in estrogens’ cyclic, periovulatory eating-inhibitoryeffect, but not its tonic eating-inhibitory effect. The role of5HT in the estrogenic control of eating may also be diet-specific because cyclic variations in the eating-inhibitory effectof 5HT were not detected when rats were fed more palatablefoods or offered different macronutrient sources (325, 538).
5HT1A autoreceptors also may be involved in the periovu-latory decrease in eating because 8-hydroxy-2–9(di-n-propyl-amino)tetralin (8-OH-DPAT), which decreases 5HT signalingby stimulating 5HT1A autoreceptors, stimulated eating less1) in estradiol-treated than in control ovariectomized rats(627), 2) in intact females rats than in ovariectomized rats(150), and 3) during proestrus and estrus than during diestrus incycling rats (749).
Some of this estrogen-serotonin interaction may involve theamygdala. This is because 1) Eckel et al. (205, 206) found thatestradiol increased eating and CCK-induced expression ofc-Fos in the central nucleus of the amygdala (205, 206), and2) Parker et al. (538) reported there were cyclic changes in theeating-stimulatory effect of injections of the 5HT2A/2C recep-
Review
R1238 SEX DIFFERENCES IN THE PHYSIOLOGY OF EATING
AJP-Regul Integr Comp Physiol • doi:10.1152/ajpregu.00446.2012 • www.ajpregu.org
by 10.220.32.247 on Septem
ber 18, 2016http://ajpregu.physiology.org/
Dow
nloaded from
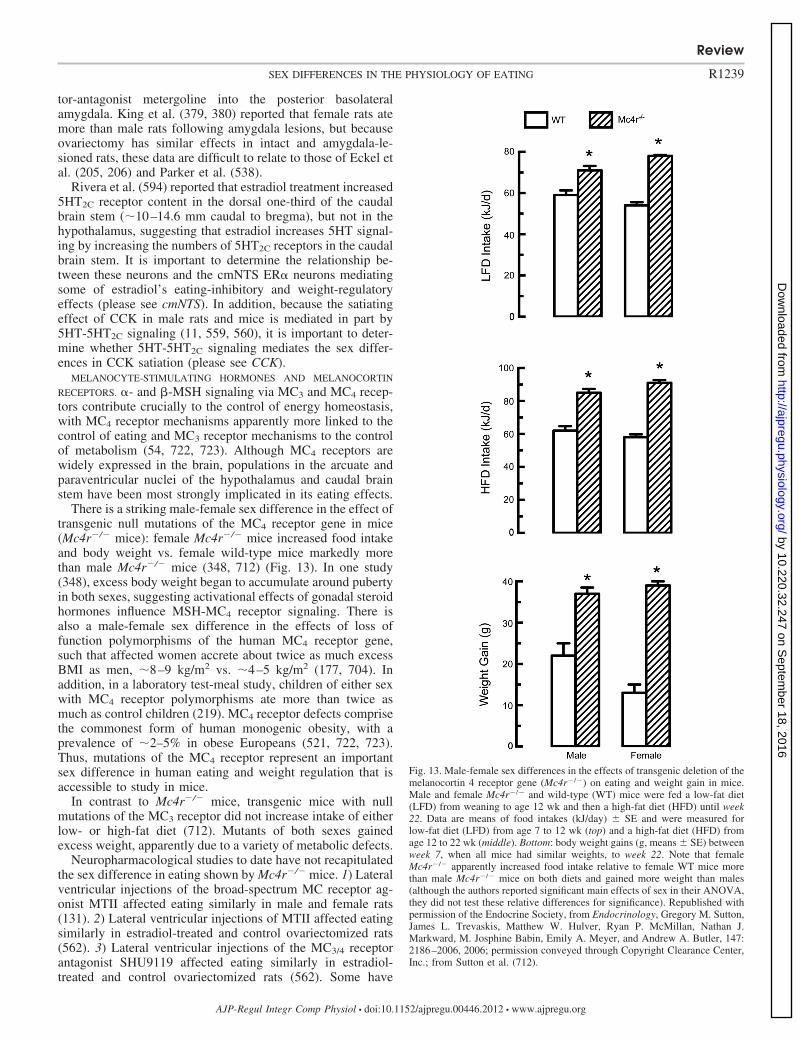
tor-antagonist metergoline into the posterior basolateralamygdala. King et al. (379, 380) reported that female rats atemore than male rats following amygdala lesions, but becauseovariectomy has similar effects in intact and amygdala-le-sioned rats, these data are difficult to relate to those of Eckel etal. (205, 206) and Parker et al. (538).
Rivera et al. (594) reported that estradiol treatment increased5HT2C receptor content in the dorsal one-third of the caudalbrain stem (�10–14.6 mm caudal to bregma), but not in thehypothalamus, suggesting that estradiol increases 5HT signal-ing by increasing the numbers of 5HT2C receptors in the caudalbrain stem. It is important to determine the relationship be-tween these neurons and the cmNTS ER� neurons mediatingsome of estradiol’s eating-inhibitory and weight-regulatoryeffects (please see cmNTS). In addition, because the satiatingeffect of CCK in male rats and mice is mediated in part by5HT-5HT2C signaling (11, 559, 560), it is important to deter-mine whether 5HT-5HT2C signaling mediates the sex differ-ences in CCK satiation (please see CCK).
MELANOCYTE-STIMULATING HORMONES AND MELANOCORTIN
RECEPTORS. �- and �-MSH signaling via MC3 and MC4 recep-tors contribute crucially to the control of energy homeostasis,with MC4 receptor mechanisms apparently more linked to thecontrol of eating and MC3 receptor mechanisms to the controlof metabolism (54, 722, 723). Although MC4 receptors arewidely expressed in the brain, populations in the arcuate andparaventricular nuclei of the hypothalamus and caudal brainstem have been most strongly implicated in its eating effects.
There is a striking male-female sex difference in the effect oftransgenic null mutations of the MC4 receptor gene in mice(Mc4r/ mice): female Mc4r/ mice increased food intakeand body weight vs. female wild-type mice markedly morethan male Mc4r/ mice (348, 712) (Fig. 13). In one study(348), excess body weight began to accumulate around pubertyin both sexes, suggesting activational effects of gonadal steroidhormones influence MSH-MC4 receptor signaling. There isalso a male-female sex difference in the effects of loss offunction polymorphisms of the human MC4 receptor gene,such that affected women accrete about twice as much excessBMI as men, �8–9 kg/m2 vs. �4–5 kg/m2 (177, 704). Inaddition, in a laboratory test-meal study, children of either sexwith MC4 receptor polymorphisms ate more than twice asmuch as control children (219). MC4 receptor defects comprisethe commonest form of human monogenic obesity, with aprevalence of �2–5% in obese Europeans (521, 722, 723).Thus, mutations of the MC4 receptor represent an importantsex difference in human eating and weight regulation that isaccessible to study in mice.
In contrast to Mc4r/ mice, transgenic mice with nullmutations of the MC3 receptor did not increase intake of eitherlow- or high-fat diet (712). Mutants of both sexes gainedexcess weight, apparently due to a variety of metabolic defects.
Neuropharmacological studies to date have not recapitulatedthe sex difference in eating shown by Mc4r/ mice. 1) Lateralventricular injections of the broad-spectrum MC receptor ag-onist MTII affected eating similarly in male and female rats(131). 2) Lateral ventricular injections of MTII affected eatingsimilarly in estradiol-treated and control ovariectomized rats(562). 3) Lateral ventricular injections of the MC3/4 receptorantagonist SHU9119 affected eating similarly in estradiol-treated and control ovariectomized rats (562). Some have
Fig. 13. Male-female sex differences in the effects of transgenic deletion of themelanocortin 4 receptor gene (Mc4r/) on eating and weight gain in mice.Male and female Mc4r/ and wild-type (WT) mice were fed a low-fat diet(LFD) from weaning to age 12 wk and then a high-fat diet (HFD) until week22. Data are means of food intakes (kJ/day) SE and were measured forlow-fat diet (LFD) from age 7 to 12 wk (top) and a high-fat diet (HFD) fromage 12 to 22 wk (middle). Bottom: body weight gains (g, means SE) betweenweek 7, when all mice had similar weights, to week 22. Note that femaleMc4r/ apparently increased food intake relative to female WT mice morethan male Mc4r/ mice on both diets and gained more weight than males(although the authors reported significant main effects of sex in their ANOVA,they did not test these relative differences for significance). Republished withpermission of the Endocrine Society, from Endocrinology, Gregory M. Sutton,James L. Trevaskis, Matthew W. Hulver, Ryan P. McMillan, Nathan J.Markward, M. Josphine Babin, Emily A. Meyer, and Andrew A. Butler, 147:2186–2006, 2006; permission conveyed through Copyright Clearance Center,Inc.; from Sutton et al. (712).
Review
R1239SEX DIFFERENCES IN THE PHYSIOLOGY OF EATING
AJP-Regul Integr Comp Physiol • doi:10.1152/ajpregu.00446.2012 • www.ajpregu.org
by 10.220.32.247 on Septem
ber 18, 2016http://ajpregu.physiology.org/
Dow
nloaded from

claimed that following SHU9119 treatment in this study (562),the estrogenic inhibition of eating was “not detected” [(599), p7] or estradiol was “unable to induce anorexia” [(463), p 5].This is incorrect. SHU9119 did not change the effect ofestradiol in tests when it occurred. For example, estradioldecreased 24-h food intake �5 g both in rats treated with 500pmol SHU9119 and in untreated rats [Fig. 1 in (562)].
One possible explanation for the apparent discrepancy be-tween the transgenic and neuropharmacological data is thatMc4r/ mutation might disrupt early development of theHPG axis and that females may be relatively more sensitive tothe disruption than males. A recent study, however, seemsinconsistent with this hypothesis. That is, Xu et al. (812) foundthat female transgenic mice in which ER� was deleted fromneurons expressing Pomc, the gene encoding MSH, becamehyperphagic beginning around puberty, suggestive of disrup-tion of an activational rather than an organizational effect ofestrogens. In this study, however, the weight gain was limitedto lean body mass, with no effect on fat mass. Because ovariec-tomy increases fat mass more that lean mass, the significance ofthese data for the normal estrogenic regulation of eating andenergy homeostasis is unclear.
Agouti-related peptide and neuropeptide Y. Neurons ex-pressing Agouti-related peptide (AgRP), an inverse agonist ofMC4 receptors, coexpress neuropeptide Y (NPY) and are foundsolely in the arcuate nucleus of the hypothalamus; NPY neu-rons are more widespread (87, 416). NPY and AgRP/NPYneurons project to a variety of forebrain and brain stem sitesinvolved in the control of eating and body weight, and centraladministration of either peptide potently stimulates eating (22,87, 416, 693). Although transgenic deletion of the Npy gene orthe Agrp gene or silencing them in early development did notaffect adult eating or body weight, silencing them in adulthoodled to pronounced anorexia (293, 445, 534, 574). Whetherthere were sex differences in these effects was not reported.Central injections of AgRP elicited similar effects on foodintake and body weight in intact and gonadectomized male andfemale rats (131, 281), however, suggesting that the male-female sex difference in the effect of MC4 receptor mutationsdescribed above is not related to AgRP function.
A study by Olofsson et al. (527) indicates that AgRP/NPYneurons play a selective role in the periovulatory decrease ineating in mice. They reported that 1) hypothalamic Agrp andNpy gene expression decreased during estrus and 2) mice withmassive degeneration of AgRP/NPY neurons due to transgenicdeletion of mitochondrial transcription factor-A in AgRP neu-rons did not reduce food intake during estrus (Fig. 14), al-though they were fertile and vaginal estrous cycling wasnormal. AgRP/NPY neuron-deficient mice displayed normalaverage levels of food intake and normal body weight, indi-cating that these neurons selectively control the cyclic, but notthe tonic, eating-inhibitory effect of estrogens. This effect wasindirect in that hypothalamic AgRP/NPY neurons did notexpress ER�. The authors also investigated the effects ofestradiol treatment, but these data are difficult to interpretphysiologically because of the high doses used (2 �g centrallyand 150 �g peripherally, please see Hormone treatment regi-mens). Whether AgRP/NPY neurons are involved in the peri-ovulatory decrease in eating in other species has not beentested, although ovariectomy increased hypothalamic NPY andAgRP mRNA contents in rats (130) and estradiol reduced the
concentration of AgRP in the cerebrospinal fluid of ovariecto-mized rhesus monkeys (810). Central injections of AgRP hadsimilar effects in estradiol-treated and control ovariectomizedrats (562, 633), which, together with Olofsson et al.’s (527)data, suggests that estrogens may modulate AgRP synthesisand release rather than MC4 receptor function or downstreammechanisms. In contrast to the lack of effect of estradiol onAgRP’s eating-stimulatory effect, estradiol did increase NPY-induced eating in one study (633), suggesting that estrogensmay modulate NPY receptor function.
MELANIN-CONCENTRATING HORMONE. Melanin-concentratinghormone (MCH) is a hypothalamic neuropeptide that stimu-lates eating and may contribute to sex differences in eating.Santollo and Eckel (634) reported that the eating-stimulatoryeffect of lateral cerebroventricular injections of MCH was1) greater during diestrus than during estrus, 2) reduced byestradiol treatment in ovariectomized rats, and 3) greater inmale rats than in estradiol-treated ovariectomized females.
Orosensory hedonics. Orosensory hedonic or palatabilityresponses result from evaluative processes in the brain thatproduce subjective experiences (pleasure, surfeit, disgust, etc.)and controls of eating related to them (66, 68, 416, 607). Avariety of data suggest that there are important physiologicalsex differences in these processes.
RATS. The hedonic control of eating can be assessed behav-iorally in animals in situations that minimize the postoropha-ryngeal effects of food. This can be done by measuring lickrates or intake at just the onset of the meal, in brief-access tests,or in sham-eating tests, during which open gastric cannulasprevent ingested food from accumulating in the stomach orentering the intestines (679). An elegant apparatus for brief-access tests offers animals repeated 10-s exposures to as manyas eight different test fluids in random order in the samesession, so that the postoropharyngeal food stimuli that de-velop during the session do not differ for the different testfluids (680). Using this method, Curtis et al. (152) found that1) male rats licked approximately twofold more 0.025 or 0.05M sucrose than did females, 2) males and females lickedsimilarly during exposure to 0.1–0.4 M sucrose (Fig. 15A) and
Fig. 14. Agouti-related peptide/neuropeptide Y (AgRP/NPY) neurons arenecessary for the periovulatory decrease in eating in mice. Mice in whichAgRP and NPY neurons were transgenically deleted (solid squares) did notshow an estrous decrease in daily food intake under conditions in whichwild-type control mice did (open symbols). M, diestrus 1; D, diestrus 2; P,proestrus; E estrus. From Olofsson et al. (527).
Review
R1240 SEX DIFFERENCES IN THE PHYSIOLOGY OF EATING
AJP-Regul Integr Comp Physiol • doi:10.1152/ajpregu.00446.2012 • www.ajpregu.org
by 10.220.32.247 on Septem
ber 18, 2016http://ajpregu.physiology.org/
Dow
nloaded from
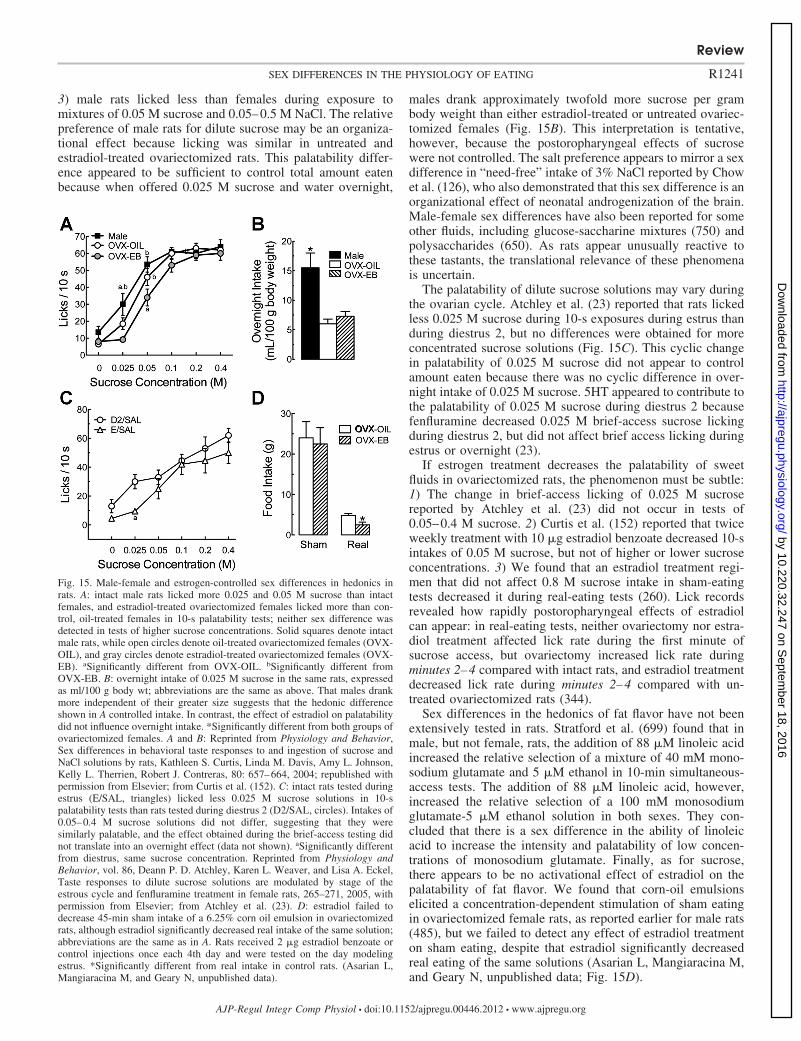
3) male rats licked less than females during exposure tomixtures of 0.05 M sucrose and 0.05–0.5 M NaCl. The relativepreference of male rats for dilute sucrose may be an organiza-tional effect because licking was similar in untreated andestradiol-treated ovariectomized rats. This palatability differ-ence appeared to be sufficient to control total amount eatenbecause when offered 0.025 M sucrose and water overnight,
males drank approximately twofold more sucrose per grambody weight than either estradiol-treated or untreated ovariec-tomized females (Fig. 15B). This interpretation is tentative,however, because the postoropharyngeal effects of sucrosewere not controlled. The salt preference appears to mirror a sexdifference in “need-free” intake of 3% NaCl reported by Chowet al. (126), who also demonstrated that this sex difference is anorganizational effect of neonatal androgenization of the brain.Male-female sex differences have also been reported for someother fluids, including glucose-saccharine mixtures (750) andpolysaccharides (650). As rats appear unusually reactive tothese tastants, the translational relevance of these phenomenais uncertain.
The palatability of dilute sucrose solutions may vary duringthe ovarian cycle. Atchley et al. (23) reported that rats lickedless 0.025 M sucrose during 10-s exposures during estrus thanduring diestrus 2, but no differences were obtained for moreconcentrated sucrose solutions (Fig. 15C). This cyclic changein palatability of 0.025 M sucrose did not appear to controlamount eaten because there was no cyclic difference in over-night intake of 0.025 M sucrose. 5HT appeared to contribute tothe palatability of 0.025 M sucrose during diestrus 2 becausefenfluramine decreased 0.025 M brief-access sucrose lickingduring diestrus 2, but did not affect brief access licking duringestrus or overnight (23).
If estrogen treatment decreases the palatability of sweetfluids in ovariectomized rats, the phenomenon must be subtle:1) The change in brief-access licking of 0.025 M sucrosereported by Atchley et al. (23) did not occur in tests of0.05–0.4 M sucrose. 2) Curtis et al. (152) reported that twiceweekly treatment with 10 �g estradiol benzoate decreased 10-sintakes of 0.05 M sucrose, but not of higher or lower sucroseconcentrations. 3) We found that an estradiol treatment regi-men that did not affect 0.8 M sucrose intake in sham-eatingtests decreased it during real-eating tests (260). Lick recordsrevealed how rapidly postoropharyngeal effects of estradiolcan appear: in real-eating tests, neither ovariectomy nor estra-diol treatment affected lick rate during the first minute ofsucrose access, but ovariectomy increased lick rate duringminutes 2–4 compared with intact rats, and estradiol treatmentdecreased lick rate during minutes 2–4 compared with un-treated ovariectomized rats (344).
Sex differences in the hedonics of fat flavor have not beenextensively tested in rats. Stratford et al. (699) found that inmale, but not female, rats, the addition of 88 �M linoleic acidincreased the relative selection of a mixture of 40 mM mono-sodium glutamate and 5 �M ethanol in 10-min simultaneous-access tests. The addition of 88 �M linoleic acid, however,increased the relative selection of a 100 mM monosodiumglutamate-5 �M ethanol solution in both sexes. They con-cluded that there is a sex difference in the ability of linoleicacid to increase the intensity and palatability of low concen-trations of monosodium glutamate. Finally, as for sucrose,there appears to be no activational effect of estradiol on thepalatability of fat flavor. We found that corn-oil emulsionselicited a concentration-dependent stimulation of sham eatingin ovariectomized female rats, as reported earlier for male rats(485), but we failed to detect any effect of estradiol treatmenton sham eating, despite that estradiol significantly decreasedreal eating of the same solutions (Asarian L, Mangiaracina M,and Geary N, unpublished data; Fig. 15D).
Fig. 15. Male-female and estrogen-controlled sex differences in hedonics inrats. A: intact male rats licked more 0.025 and 0.05 M sucrose than intactfemales, and estradiol-treated ovariectomized females licked more than con-trol, oil-treated females in 10-s palatability tests; neither sex difference wasdetected in tests of higher sucrose concentrations. Solid squares denote intactmale rats, while open circles denote oil-treated ovariectomized females (OVX-OIL), and gray circles denote estradiol-treated ovariectomized females (OVX-EB). aSignificantly different from OVX-OIL. bSignificantly different fromOVX-EB. B: overnight intake of 0.025 M sucrose in the same rats, expressedas ml/100 g body wt; abbreviations are the same as above. That males drankmore independent of their greater size suggests that the hedonic differenceshown in A controlled intake. In contrast, the effect of estradiol on palatabilitydid not influence overnight intake. *Significantly different from both groups ofovariectomized females. A and B: Reprinted from Physiology and Behavior,Sex differences in behavioral taste responses to and ingestion of sucrose andNaCl solutions by rats, Kathleen S. Curtis, Linda M. Davis, Amy L. Johnson,Kelly L. Therrien, Robert J. Contreras, 80: 657–664, 2004; republished withpermission from Elsevier; from Curtis et al. (152). C: intact rats tested duringestrus (E/SAL, triangles) licked less 0.025 M sucrose solutions in 10-spalatability tests than rats tested during diestrus 2 (D2/SAL, circles). Intakes of0.05–0.4 M sucrose solutions did not differ, suggesting that they weresimilarly palatable, and the effect obtained during the brief-access testing didnot translate into an overnight effect (data not shown). aSignificantly differentfrom diestrus, same sucrose concentration. Reprinted from Physiology andBehavior, vol. 86, Deann P. D. Atchley, Karen L. Weaver, and Lisa A. Eckel,Taste responses to dilute sucrose solutions are modulated by stage of theestrous cycle and fenfluramine treatment in female rats, 265–271, 2005, withpermission from Elsevier; from Atchley et al. (23). D: estradiol failed todecrease 45-min sham intake of a 6.25% corn oil emulsion in ovariectomizedrats, although estradiol significantly decreased real intake of the same solution;abbreviations are the same as in A. Rats received 2 �g estradiol benzoate orcontrol injections once each 4th day and were tested on the day modelingestrus. *Significantly different from real intake in control rats. (Asarian L,Mangiaracina M, and Geary N, unpublished data).
Review
R1241SEX DIFFERENCES IN THE PHYSIOLOGY OF EATING
AJP-Regul Integr Comp Physiol • doi:10.1152/ajpregu.00446.2012 • www.ajpregu.org
by 10.220.32.247 on Septem
ber 18, 2016http://ajpregu.physiology.org/
Dow
nloaded from

HUMANS: PSYCHOPHYSICS. Determination of sex differences inflavor hedonics in humans requires comparisons of psycho-physical ratings between different types of subjects, which is aclassical problem in psychophysics. The general labeled-mag-nitude scale is the most valid tool available for this and isreshaping understanding of the relationships among hedonics,sex, and obesity (44, 45, 685). For example, Hayes and Duffy(316) reported a complex interaction between the density offungiform papillae on the tongue and the pleasantness ofsucrose-cream mixtures. As described above (please see Gus-tation), greater densities of fungiform papillae on the tongueare associated with increases in the perceived intensity ofsweetness and creaminess of food, independent of the hedonicvalence of the stimuli. In this study, Hayes and Duffy (316)found a sex difference in the relationship between fungiformpapillae density and hedonic ratings. Women with fewer fun-giform papillae and men with more fungiform papillae foundmost pleasant high-creamy, high-sweet flavors, whereaswomen with more fungiform papillae and men with fewerfungiform papillae found most pleasant medium-creamy, me-dium-sweet flavors. The effects were large: women and menwho preferred most high-creamy, high-sweet flavors endorsedabsolute intensities of liking that were approximately twofoldlarger than men and women who preferred most medium-creamy, medium-sweet flavors (Fig. 16). Given that absoluteliking ratings of the most-preferred foods are positively corre-lated with BMI (45) and that obese women prefer high-fat,high-sugar foods (194, 451), these data suggest that a relativelack of fungiform papillae would increase a woman’s risk forobesity. How the data relate to obesity risk in men is moredifficult to predict, given that obese men prefer high-fat,high-protein foods to high-fat, high-sugar foods (194, 451).The complexity of interactions of sensory and hedonic flavorresponses undoubtedly contributes to the variable findings onchanges in food preferences over the menstrual cycle (e.g.,241, 359, 472) and in functional brain imaging studies (nextsection).
HUMAN FUNCTIONAL BRAIN IMAGING: INTRODUCTION. The neu-ral bases of normal and disordered human eating, eating-related behavior, cognition, and affect can now be investigatedwith functional brain imaging, which indirectly measures lev-els of neuronal activity in circumscribed brain areas (111, 141,173, 230, 410, 509, 753, 756). Progress in this exciting area isdifficult for a number of reasons: 1) Images of brain responsesrelated to cognition and affect are extraordinarily sensitive towithin- and between-subjects variations in arousal, mood, hab-its, life experiences, food-related memories, social context, andother difficult-to-control factors. 2) Images of brain responsesare only indirect measures of actual neural activity. They arebased on indirect detection of cerebral glucose metabolism(positron-emission tomography) or, more frequently, indirectdetection of cerebral blood flow (functional magnetic reso-nance imaging). Such measures cannot distinguish inhibitionfrom excitation. Changes in blood flow or metabolism mayarise from excitatory or inhibitory neural activation. In addi-tion, differences in response intensity across regions mighthave more to do with differences in the neural architecture ofmicrocircuits within the regions than with differential involve-ment in processing the stimuli tested. 3) Detection of brainactivation depends both on the physical sensitivity of theimaging machinery and the information-processing algorithms
used. For example, the present limit of spatial resolution ofbrief scans is not sufficient to differentiate activity in adjacenthypothalamic areas that have different functions. In addition,the temporal resolution is too poor to enable inferences aboutthe sequence of information processing. 4) Images of largertelencephalic areas often differentiate activity within subareasfor which no functional differences are known. 5) Psycholog-ical functions cannot be assigned unambiguously to brainareas. For example, the orbital frontal cortex (OFC), whichmany data suggest is the most important telencephalic node inthe neural network mediating food hedonics, is clearly alsoinvolved in cognitive processes, such as decision making, riskevaluation, and expectation (66, 403, 409, 410, 530, 609).Furthermore, there are sex differences in some of these cogni-tive processes (403, 530).
Despite these difficulties, pictures of foods and tasting foodreliably increase brain activation in a number of areas (111,173, 410, 509, 753, 754, 756). Increased flavor intensity,independent of hedonic evaluation, usually increases activationin the middle insular cortex, cerebellum, and amygdala. In-creased hedonic intensity usually increases activation in theOFC, anteroventral striatum, and nucleus accumbens (NAc),anterior insular cortex (AIC), and anterior cingulate cortex(ACC). The dorsolateral prefrontal (dlPFC) and parietal corti-ces are also often activated, which is usually interpreted to beassociated with cognitive or “executive-control” aspects of theexperience. These areas tend to be more activated in peoplewith higher levels of dietary restraint (dietary restraint isdiscussed in Interaction with cognitive controls of eating).When the stimuli are pictures of foods, the fusiform gyrus, ahigher-order visual processing area, is usually activated.
The involvement of the amygdala and NAc in human hedo-nics, as well as studies of the involvement of dopaminergicneurotransmission that we did not review (694, 695), parallelstudies of flavor reward in rats (35, 515, 651). Similarly,studies of the responses of individual neurons in both subcor-tical and cortical brain areas in rhesus macaques to taste stimulisuggest that there is substantial overlap in the neural structuresmediating flavor hedonics in monkeys and in humans (284,606, 608). Thus, in addition to providing unique insights abouthuman hedonics, neuroimaging data also support the use ofanimal models. Nevertheless, some species differences are tobe expected. For example, neuroimaging and clinical dataindicate that the AIC is necessary for normal affect related tobodily states, if not for all affect, and monkeys and rodents donot have a homologous structure (142, 143).
An important discovery is that food stimuli affect brainactivation differently in normal-weight and obese persons(111, 173, 410, 509, 694, 753, 756). For example, obesepersons typically display greater differential activation in re-sponse to pictures of high vs. low energy-density foods in theNAc, AIC, ACC, and OFC, suggesting that increased hedonicevaluation of food may contribute to the pathophysiology ofobesity. A study by Stice et al. (696) supported this hypothesis.They found that palatable food and monetary rewards in-creased brain activation in several brain areas associated withhedonic evaluations in normal-weight children at high familialrisk for obesity (by virtue of having two overweight or obeseparents) compared with low-risk children (two normal-weightparents).
Review
R1242 SEX DIFFERENCES IN THE PHYSIOLOGY OF EATING
AJP-Regul Integr Comp Physiol • doi:10.1152/ajpregu.00446.2012 • www.ajpregu.org
by 10.220.32.247 on Septem
ber 18, 2016http://ajpregu.physiology.org/
Dow
nloaded from
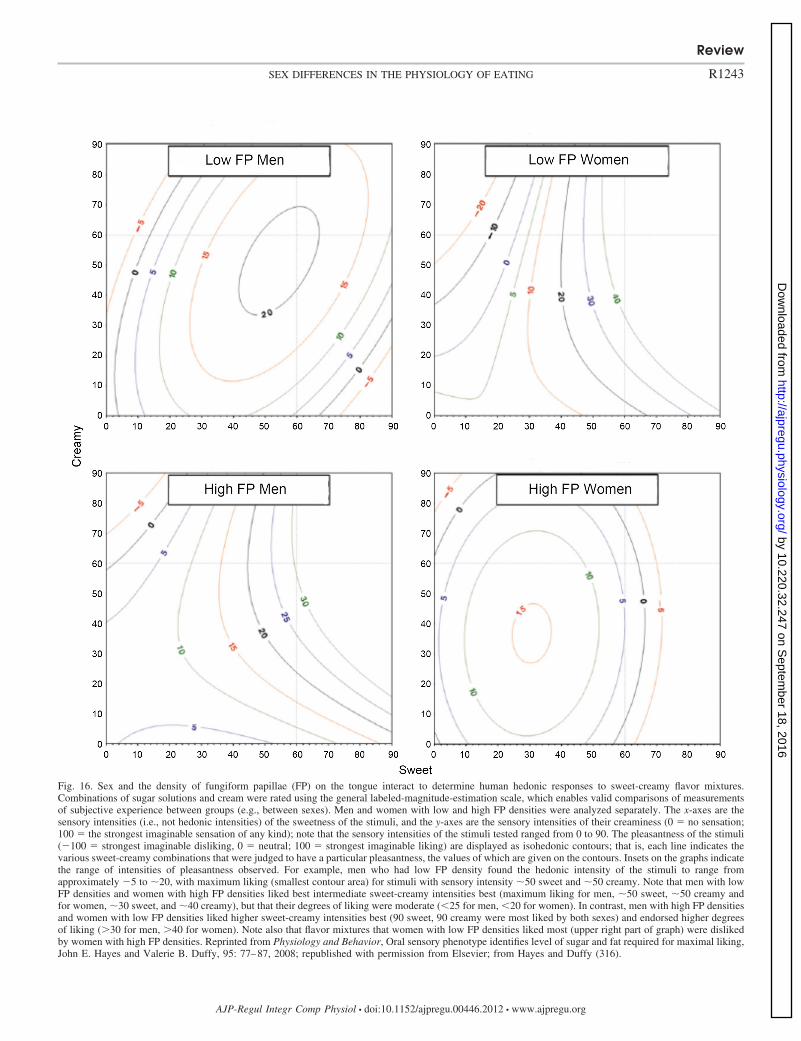
Fig. 16. Sex and the density of fungiform papillae (FP) on the tongue interact to determine human hedonic responses to sweet-creamy flavor mixtures.Combinations of sugar solutions and cream were rated using the general labeled-magnitude-estimation scale, which enables valid comparisons of measurementsof subjective experience between groups (e.g., between sexes). Men and women with low and high FP densities were analyzed separately. The x-axes are thesensory intensities (i.e., not hedonic intensities) of the sweetness of the stimuli, and the y-axes are the sensory intensities of their creaminess (0 � no sensation;100 � the strongest imaginable sensation of any kind); note that the sensory intensities of the stimuli tested ranged from 0 to 90. The pleasantness of the stimuli(100 � strongest imaginable disliking, 0 � neutral; 100 � strongest imaginable liking) are displayed as isohedonic contours; that is, each line indicates thevarious sweet-creamy combinations that were judged to have a particular pleasantness, the values of which are given on the contours. Insets on the graphs indicatethe range of intensities of pleasantness observed. For example, men who had low FP density found the hedonic intensity of the stimuli to range fromapproximately 5 to �20, with maximum liking (smallest contour area) for stimuli with sensory intensity �50 sweet and �50 creamy. Note that men with lowFP densities and women with high FP densities liked best intermediate sweet-creamy intensities best (maximum liking for men, �50 sweet, �50 creamy andfor women, �30 sweet, and �40 creamy), but that their degrees of liking were moderate (�25 for men, �20 for women). In contrast, men with high FP densitiesand women with low FP densities liked higher sweet-creamy intensities best (90 sweet, 90 creamy were most liked by both sexes) and endorsed higher degreesof liking (�30 for men, �40 for women). Note also that flavor mixtures that women with low FP densities liked most (upper right part of graph) were dislikedby women with high FP densities. Reprinted from Physiology and Behavior, Oral sensory phenotype identifies level of sugar and fat required for maximal liking,John E. Hayes and Valerie B. Duffy, 95: 77–87, 2008; republished with permission from Elsevier; from Hayes and Duffy (316).
Review
R1243SEX DIFFERENCES IN THE PHYSIOLOGY OF EATING
AJP-Regul Integr Comp Physiol • doi:10.1152/ajpregu.00446.2012 • www.ajpregu.org
by 10.220.32.247 on Septem
ber 18, 2016http://ajpregu.physiology.org/
Dow
nloaded from

HUMAN FUNCTIONAL BRAIN IMAGING: SEX DIFFERENCES. Male-female sex differences in brain activation in response to theflavors of normal foods (176, 675), to pure tastes (299), and topictures of food (135, 231, 264, 378, 748) have been reported.In a recent review of this literature, Geliebter et al. (264)concluded that if brain areas are classed by their predominatefunctions, as described above, pictures of food stimuli elicitmore activation in areas related to planning and executingbehaviors in men and more activation in areas related tocognitive and affective processes in women. This conclusion,although certainly generally correct, belies the complexity ofthe data obtained in these studies.
The complex effects of sex on imaging responses is clearlyreflected in Geliebter et al.’s (264) study. Subjects wereweight-stable, healthy obese men and women. They ate a 1,000kcal dinner, fasted 12 h, then ate either 750 ml liquid diet(�740 kcal, fed state) or 750 ml water (fasted state), and weretested 95 min later. The difference in response between highand low energy-density foods was larger in men than womenduring the fed state in five brain areas and during the fastedstate in five different brain areas. The high-low difference waslarger in women than men in five brain areas during the fedstate and during the fasted state in four different areas, as wellas two of the same areas, the insula and the dlPFC. The insularesponses occurred on the left side in both men and women, butnot in similar subregions [Montreal Neurological Institutestandard brain coordinates (right-left, anterior-posterior, supe-rior-inferior), 32, 34, 20 and 28, 0, 30, respectively]. Inthe dlPFC, differences were detected in similar subregions, butoccurred in the right hemisphere during the fed state and in theleft hemisphere during the fasted state (coordinates 30, 40, 34,and 34, 38, and 32, respectively). Furthermore, in two otherparts of the dlPFC, women showed smaller rather than largerresponses than men in the fed state (coordinates 42, 34, 16, and30, 22, 34). The only other area in which brain responsesoccurred in more than one condition was the right fusiformgyrus, where the high-low difference was larger in men thanwomen in the fed state and larger in women than men in thefasted state. Aside from the lack of menstrual-cycle phase data,this is a well-planned and analyzed study. Nevertheless, currentunderstanding of human brain function simply does not permita coherent interpretation of most of the effects obtained.
It is possible that studies of brain responses to receipt of foodmight lead to simpler patterns of data. As far as we know,however, only two such studies have appeared. Haase at al.(299) studied brain responses to intraoral infusions of 0.3 ml0.04 M caffeine (bitter), 0.01 M citric acid (sour), 0.64 Msucrose (sweet) or 0.16 M NaCl (salty) in normal-weight menand women during fed and fasted states [using a methodsimilar to that of Geliebter et al. (264) described above].Indeed, only two sex differences in brain responses to sucrosebetween the fed and fasted states were detected—the differen-tial responses were larger in men than women in the insula andcerebellum (sex differences for other tastes occurred in severalother areas). Smeets et al. (675) did a more naturalistic study.Brain responses to chocolate-flavored milk, administered insmall amounts by an undescribed method, were tested innormal-weight men and women, first in a fasted state and thenafter eating solid bittersweet chocolate and water to satiety.Sex differences in the effect of satiation were detected only inthree areas: men showed greater increases in activation in the
ventral striatum and greater decreases in the medial PFC, whilewomen showed greater decreases in the hypothalamus.
We are aware of two reports that women’s brain activationin response to pictures of foods varies across the menstrualcycle. Frank et al. (232) reported that the differential effect ofpictures of high energy-density foods vs. nonfoods was largerin the follicular phase than in the luteal phase in the NAc,amygdala, and hippocampus; the effect of low energy-densityfoods vs. nonfoods was significant only in the hippocampus.Alonso-Alonso et al. (4) tested the effects of pictures of foodbefore and after a standard meal designed to contain 20% oftheir daily energy need; this was done once early (days 3–6after the onset of menstruation) and once late (days 10–13) inthe follicular phase of the menstrual cycle. The differentialeffect of fed vs. fasted states of activation in response topictures of food was greater in the late vs. the early follicularphase in the inferior frontal gyrus and fusiform gyrus. Theinferior frontal gyrus is considered to be an “executive-control”area and, as mentioned above, the fusiform gyrus is a higher-order visual-processing area; thus, neither is likely to bedirectly involved in the control of eating. Nevertheless, it isintriguing that, unique among cortical areas, the volume of thefusiform gyrus-parahippocampal gyrus area, as well as perfor-mance on hippocampal-dependent memory tasks, variedthroughout the menstrual cycle (557, 572).
Physiological Sex Differences in Disordered Human Eating
Psychiatric eating disorders. The current view is that thepsychiatric eating disorders anorexia nervosa, bulimia nervosa,binge-eating disorder, and “eating disorders not otherwisespecified” (which accounts for about half of eating-disorderpatients) are caused by complex interactions among cultural,social, familial, and biological factors (5, 30, 365, 387, 460,740, 777). Perhaps the strongest evidence that biological fac-tors are important in psychiatric eating disorders is their highheritability (heritability is the fraction of phenotypic varianceaccounted for by genetic variation). Twin studies, for example,suggest that genetic variability accounts for �50–80% of therisks for anorexia nervosa and bulimia nervosa (365, 736, 740).
The various influences on psychiatric eating disorders arethought to interact dynamically, so that precipitating vulnera-bilities spiral into vicious cycles involving increasing numbersof causal factors. Sex is clearly an important variable. Thelifetime prevalence of psychiatric eating disorders (346, 351)and various symptoms of disordered eating (700) are aboutthree-fold higher in women than in men in community sam-ples, and these differences emerge around the onset of puberty(27, 631, 715). Subclinical symptoms of disordered eating arealso more prevalent in females than males (329, 700) and werehighly heritable in pubertal and postpubertal girls, but not inprepubertal girls (148). In addition, recovery from eating dis-orders is slower in women than in men, and rates of remissionare higher (698).
That the pathogenesis of eating disorders involves biologicalfactors naturally encourages studies of potential biologicalbases of these sex differences, but as our review reflects, fewhuman studies have done so. Similarly, although the utility ofanimal models of disordered eating is increasingly emphasized(113, 138, 139, 362, 363, 665, 676), the contributions of
Review
R1244 SEX DIFFERENCES IN THE PHYSIOLOGY OF EATING
AJP-Regul Integr Comp Physiol • doi:10.1152/ajpregu.00446.2012 • www.ajpregu.org
by 10.220.32.247 on Septem
ber 18, 2016http://ajpregu.physiology.org/
Dow
nloaded from

physiological sex differences are not a major component ofthese research efforts.
Eating disorders affect gonadal-steroid hormone levels.Women who are acutely ill with anorexia nervosa have abnor-mally low gonadotropin, estrogen and progestin levels andusually amenorrhea, and women with bulimia nervosa fre-quently report menstrual dysfunction (30, 332, 334, 483, 566).In addition, testosterone levels are decreased in symptomaticwomen with anorexia nervosa and increased in symptomaticwomen with bulimia nervosa (30, 504, 709). Changes ingonadal steroid hormone levels, however, are consequences ofweight loss and extreme dieting rather than being causes ofdisordered eating (7, 30, 277, 483, 586). Nevertheless, phar-macological treatments that affect gonadal hormone levels maybe useful in the treatment of disordered eating, as least inselected patients (30, 63, 334, 366, 483, 504). For example,initial results suggest that antiandrogenic treatment may reducebinge eating in women with bulimia nervosa (504, 710).
ANOREXIA NERVOSA: HUMAN STUDIES. Twin studies indicatethat genetic factors account for up to 74 (395) to 88% (96) ofthe variance in anorexia nervosa. The genes accounting for thishigh heritability and their contribution to sex differences inanorexia nervosa are unknown. One potential clue comes froma family-based association study involving 321 French families(761) that suggested that polymorphisms in ESR1, the geneencoding ER�, may contribute. In this study, overtransmissionof three ESR1 single-nucleotide polymorphisms (rs726281,rs3798577, and rs2295193), as well as a haplotype of ESR1involving these and five additional single-nucleotide polymor-phisms were associated with restricting-type anorexia nervosa(i.e., energy intake controlled by dieting rather than purging).This study (761) also failed to detect the previously reported(200, 616) association between the single nucleotide polymor-phism rs1256049 in ESR2, which encodes ER�, and anorexianervosa. Further work is required to determine whether theseeffects can be replicated and extended to other populations.This has not yet succeeded. Rather, genome-wide associationstudies to date, although yielding some candidates for furtherresearch, have not identified genome-wide significance forvariants in ESR1 or any other genes in anorexia nervosa (128,354, 773, 781).
Klump and colleagues (146, 389) presented two lines ofsupport for their hypothesis that disordered human eating isinfluenced by early organizational effects of gonadal steroidhormones similar to those discovered in animals (please seeDevelopment). 1) In normal-weight women with either male orfemale twins, higher ratios of the lengths of the second andfourth fingers, indicative of less prenatal androgen exposure,were significantly correlated with increased scores on a ques-tionnaire measuring subclinical disordered-eating symptoms(389). 2) Disordered-eating scores were significantly higher ingirls with female twins than girls with male twins, who pre-sumably had greater prenatal androgen exposure (146). Fur-thermore, this difference in disordered eating scores emergedduring mid-late puberty (147), suggesting that the developmen-tal effect forms a substrate for a later activational effect ofgonadal steroids (as described below). These associations,however, were detected in only one (577) of several (31, 95,449, 571, 581) subsequent studies involving patients witheating disorders. Thus, Klump’s organizational hypothesis re-quires further research.
The increased incidence of anorexia nervosa around pubertysuggests that an activational effect of estrogens may causeanorexia nervosa, as originally hypothesized by Young (815,816). Reports (389, 579) of significant correlations betweensalivary estradiol levels and subclinical eating-disorder symp-toms across the menstrual cycle are consistent with this hy-pothesis. Whether any of these women progressed to anorexianervosa, however, was not established. Misra et al. (486)performed a randomized, double-blind study in which femalepatients with anorexia nervosa (n � 110; mean age, 16 yr;mean BMI, 17.4 kg/m2; minimum duration of amenorrhea, 3mo) received either placebo or a physiological regimen ofestrogen treatment in addition to their ongoing treatment reg-imens. Estrogen treatment consisted of a fixed dose of estradiolin older patients and an escalating dose schedule that wasdesigned to mimic peripubertal plasma estradiol levels inyounger patients. After 18 mo, estradiol did not significantlyaffect body weight gain (4.6 1.0 kg in estradiol-treated vs.4.2 1.0 kg in control patients) or BMI gain (1.6 0.4 kg/m2
vs. 1.5 0.4 kg/m2) but did significantly improve spine andhip bone mineral densities. These data suggest that estrogensdid not exacerbate the symptoms of anorexia nervosa in thesepatients, but because eating was not measured and may havebeen too low in these patients to detect decreases, they are notconclusive.
ANOREXIA NERVOSA: ANIMAL MODELS. The most popular ani-mal model of anorexia nervosa is Routtenberg’s (620, 621)activity-based anorexia model, which is self-starvation broughtabout in mice and rats by providing ad libitum access to anactivity wheel and limiting food access to �1–2 h/day. Veryfew activity-based anorexia studies involve females. Doerrieset al. (186) found that female Holtzman Sprague-Dawley ratsran more than weight-matched males, but ate more and lostweight slower. Female rats may not develop activity-basedanorexia as reliably as males; in one study, females did notincrease activity (79) and in another they increased activity butdid not decrease eating (185).
Activity-based anorexia rapidly suppresses ovarian cyclingin rats, paralleling the endocrine disturbances of anorexianervosa (554) and other situations of insufficient energy intake(441, 442, 645, 770, 792). This activity-based-anorexia effectis due to an interaction of high activity and reduced food intakebecause neither alone was sufficient for it (184, 185, 784). Asimilar synergistic effect on ovarian cycling occurred in re-sponse to combinations of food restriction and psychosocialstress in cynomolgus monkeys (792). Increased activity anddecreased eating combine to inhibit kisspeptin or GnRH neu-rons in the hypothalamus, which leads to reductions in LH andestrogens (175, 456, 666). Whether the reduced estrogen se-cretion is related to the weaker anorectic effect in female ratscompared with male rats (185) is unknown.
There may also be a sex difference in the development ofvulnerability to activity-based anorexia in rats. Hancock andGrant (307) reported that early, acute, maternal separation ledto greater increases in activity and more profound anorexia infemales than males when tested as adolescents, but more inmales than females when tested as adults. These data suggestboth 1) that early-life stress has complex effects on suscepti-bility to activity-based anorexia later in life and 2) that sex mayaffect the susceptibility to activity-based anorexia indirectly,
Review
R1245SEX DIFFERENCES IN THE PHYSIOLOGY OF EATING
AJP-Regul Integr Comp Physiol • doi:10.1152/ajpregu.00446.2012 • www.ajpregu.org
by 10.220.32.247 on Septem
ber 18, 2016http://ajpregu.physiology.org/
Dow
nloaded from
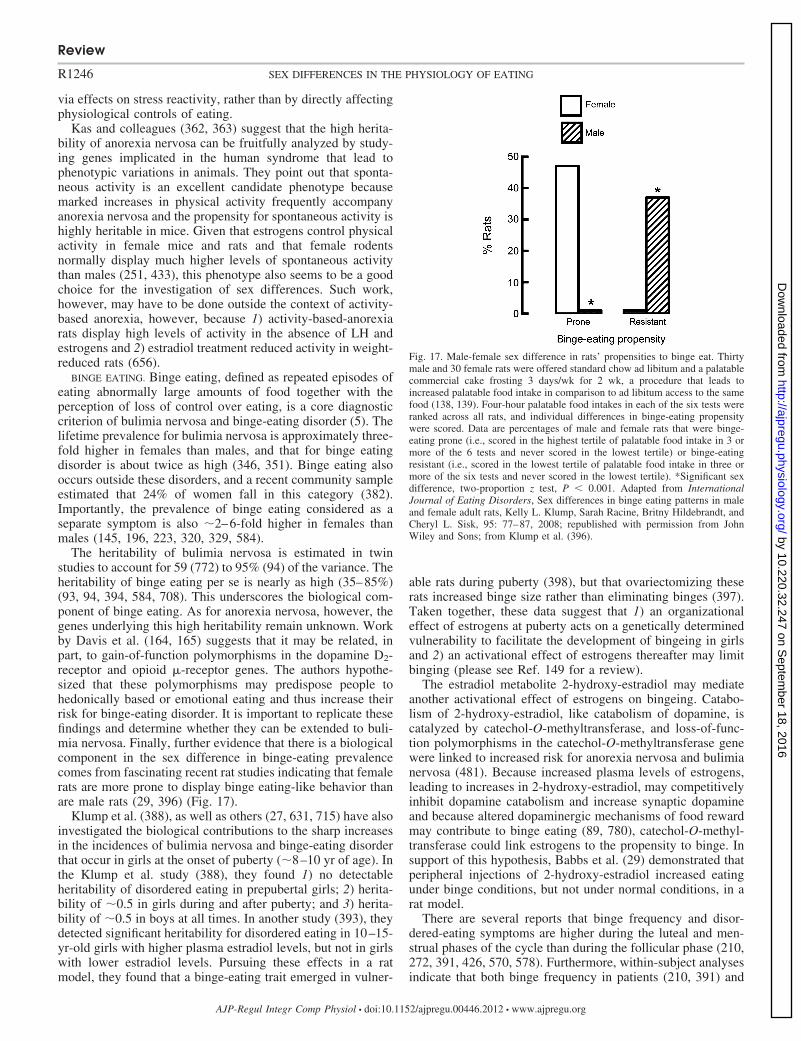
via effects on stress reactivity, rather than by directly affectingphysiological controls of eating.
Kas and colleagues (362, 363) suggest that the high herita-bility of anorexia nervosa can be fruitfully analyzed by study-ing genes implicated in the human syndrome that lead tophenotypic variations in animals. They point out that sponta-neous activity is an excellent candidate phenotype becausemarked increases in physical activity frequently accompanyanorexia nervosa and the propensity for spontaneous activity ishighly heritable in mice. Given that estrogens control physicalactivity in female mice and rats and that female rodentsnormally display much higher levels of spontaneous activitythan males (251, 433), this phenotype also seems to be a goodchoice for the investigation of sex differences. Such work,however, may have to be done outside the context of activity-based anorexia, however, because 1) activity-based-anorexiarats display high levels of activity in the absence of LH andestrogens and 2) estradiol treatment reduced activity in weight-reduced rats (656).
BINGE EATING. Binge eating, defined as repeated episodes ofeating abnormally large amounts of food together with theperception of loss of control over eating, is a core diagnosticcriterion of bulimia nervosa and binge-eating disorder (5). Thelifetime prevalence for bulimia nervosa is approximately three-fold higher in females than males, and that for binge eatingdisorder is about twice as high (346, 351). Binge eating alsooccurs outside these disorders, and a recent community sampleestimated that 24% of women fall in this category (382).Importantly, the prevalence of binge eating considered as aseparate symptom is also �2–6-fold higher in females thanmales (145, 196, 223, 320, 329, 584).
The heritability of bulimia nervosa is estimated in twinstudies to account for 59 (772) to 95% (94) of the variance. Theheritability of binge eating per se is nearly as high (35–85%)(93, 94, 394, 584, 708). This underscores the biological com-ponent of binge eating. As for anorexia nervosa, however, thegenes underlying this high heritability remain unknown. Workby Davis et al. (164, 165) suggests that it may be related, inpart, to gain-of-function polymorphisms in the dopamine D2-receptor and opioid �-receptor genes. The authors hypothe-sized that these polymorphisms may predispose people tohedonically based or emotional eating and thus increase theirrisk for binge-eating disorder. It is important to replicate thesefindings and determine whether they can be extended to buli-mia nervosa. Finally, further evidence that there is a biologicalcomponent in the sex difference in binge-eating prevalencecomes from fascinating recent rat studies indicating that femalerats are more prone to display binge eating-like behavior thanare male rats (29, 396) (Fig. 17).
Klump et al. (388), as well as others (27, 631, 715) have alsoinvestigated the biological contributions to the sharp increasesin the incidences of bulimia nervosa and binge-eating disorderthat occur in girls at the onset of puberty (�8–10 yr of age). Inthe Klump et al. study (388), they found 1) no detectableheritability of disordered eating in prepubertal girls; 2) herita-bility of �0.5 in girls during and after puberty; and 3) herita-bility of �0.5 in boys at all times. In another study (393), theydetected significant heritability for disordered eating in 10–15-yr-old girls with higher plasma estradiol levels, but not in girlswith lower estradiol levels. Pursuing these effects in a ratmodel, they found that a binge-eating trait emerged in vulner-
able rats during puberty (398), but that ovariectomizing theserats increased binge size rather than eliminating binges (397).Taken together, these data suggest that 1) an organizationaleffect of estrogens at puberty acts on a genetically determinedvulnerability to facilitate the development of bingeing in girlsand 2) an activational effect of estrogens thereafter may limitbinging (please see Ref. 149 for a review).
The estradiol metabolite 2-hydroxy-estradiol may mediateanother activational effect of estrogens on bingeing. Catabo-lism of 2-hydroxy-estradiol, like catabolism of dopamine, iscatalyzed by catechol-O-methyltransferase, and loss-of-func-tion polymorphisms in the catechol-O-methyltransferase genewere linked to increased risk for anorexia nervosa and bulimianervosa (481). Because increased plasma levels of estrogens,leading to increases in 2-hydroxy-estradiol, may competitivelyinhibit dopamine catabolism and increase synaptic dopamineand because altered dopaminergic mechanisms of food rewardmay contribute to binge eating (89, 780), catechol-O-methyl-transferase could link estrogens to the propensity to binge. Insupport of this hypothesis, Babbs et al. (29) demonstrated thatperipheral injections of 2-hydroxy-estradiol increased eatingunder binge conditions, but not under normal conditions, in arat model.
There are several reports that binge frequency and disor-dered-eating symptoms are higher during the luteal and men-strual phases of the cycle than during the follicular phase (210,272, 391, 426, 570, 578). Furthermore, within-subject analysesindicate that both binge frequency in patients (210, 391) and
Fig. 17. Male-female sex difference in rats’ propensities to binge eat. Thirtymale and 30 female rats were offered standard chow ad libitum and a palatablecommercial cake frosting 3 days/wk for 2 wk, a procedure that leads toincreased palatable food intake in comparison to ad libitum access to the samefood (138, 139). Four-hour palatable food intakes in each of the six tests wereranked across all rats, and individual differences in binge-eating propensitywere scored. Data are percentages of male and female rats that were binge-eating prone (i.e., scored in the highest tertile of palatable food intake in 3 ormore of the 6 tests and never scored in the lowest tertile) or binge-eatingresistant (i.e., scored in the lowest tertile of palatable food intake in three ormore of the six tests and never scored in the lowest tertile). *Significant sexdifference, two-proportion z test, P � 0.001. Adapted from InternationalJournal of Eating Disorders, Sex differences in binge eating patterns in maleand female adult rats, Kelly L. Klump, Sarah Racine, Britny Hildebrandt, andCheryl L. Sisk, 95: 77–87, 2008; republished with permission from JohnWiley and Sons; from Klump et al. (396).
Review
R1246 SEX DIFFERENCES IN THE PHYSIOLOGY OF EATING
AJP-Regul Integr Comp Physiol • doi:10.1152/ajpregu.00446.2012 • www.ajpregu.org
by 10.220.32.247 on Septem
ber 18, 2016http://ajpregu.physiology.org/
Dow
nloaded from

disordered-eating symptoms in community samples (390, 392,578, 580) are positively correlated with preceding progesteronelevels and negatively correlated with preceding estradiol lev-els, suggesting possibly causal relationships. Furthermore, thecorrelations between estradiol and progesterone levels anddisordered eating were evident across a range of patient BMI,levels of dietary restraint, and levels of impulsivity (390, 580).Clearly, mechanistic work on the relationships between acti-vational effects of estrogens and progestins and binge eating iswarranted. Two issues that require further research in thiscontext are whether binge size, in addition to frequency,varies across the cycle and whether cyclic effects on binge-ing are related to the cyclic changes in normal eating (pleasesee Ovarian cycle).
The hormonal control of binge size, but not of binge fre-quency, has been investigated in rat binge-eating models basedon intermittent access to palatable food (138, 139). Binge size1) did not vary across the ovarian cycle in intact rats (57),2) was tonically, but not cyclically, decreased during a cyclicestradiol treatment regimen that both tonically and cyclicallydecreased normal eating in ovariectomized rats (817, 818), and3) was not affected by cyclic treatment with a near-physiolog-ical progesterone dose, either alone or in combination withestradiol (817). Thus, how closely cyclic changes in bingefrequency in women can be modeled in rats is not yet clear.
Finally, test-meal studies indicate that patients who binge eatdisplay decreased perceived satiation together with decreasedgastric emptying or increased gastric capacity (180, 263, 265),decreased postprandial plasma ghrelin drops (262), de-creased postprandial plasma CCK increases (180, 269, 367,555), and decreased postprandial plasma GLP-1 increases(506). Whether these are adaptations to binge eating orcontribute causally to binge size is not certain. Nevertheless,in view of the sex differences in these mechanisms reviewedabove, they merit testing as potential sources of sex differ-ences in binge eating.
Obesity. As reviewed above, the effects of loss of ovarianfunction and estrogen treatment on adiposity are clearly linkedto changes in eating in mice and rats and may be so in women.Here, we discuss two further links among HPG function,eating, and obesity.
POLYCYSTIC OVARY SYNDROME. Polycystic ovary syndrome isan endocrine disorder that includes abnormal ovarian morphol-ogy, hyperandrogenism (e.g., hirsutism) and oligomenorrhea oramenorrhea after puberty (211, 280, 334, 785). It affects �10%of reproductive-aged women. The majority of women withpolycystic ovary syndrome are also obese, with atypical re-gional adipose-tissue distribution, i.e., predominately abdomi-nal rather than gluteo-femoral adiposity. Because obesity, es-pecially abdominal obesity, disrupts ovarian cycling and leadsto signs of hyperandrogenism, obesity is presumed to cause orexacerbate polycystic ovary disorder (37, 211, 785). Progres-sive development of insulin resistance, hyperinsulinemia, andinsulin-mediated inhibition of estrogen secretion may alsorelease eating from estrogenic inhibition. In addition, someeating abnormalities that are associated with polycystic ovarysyndrome are similar to those of bulimia nervosa, suggestingthat increased androgen levels may underlie both (334, 335,434, 505).
Meal-related gastrointestinal-hormone secretion is disorderedin polycystic ovary syndrome: test meals decreased plasma ghre-
lin levels less (489) and increased plasma CCK level less (335) inpatients with polycystic ovary syndrome than in weight-matchedcontrol women. Basal ghrelin secretion is also reduced in womenwith polycystic ovary syndrome, and increased during anti-androgen therapy in one small study (246). More work is requiredto determine the causes of these changes and whether they arerelated to disordered eating.
BARIATRIC SURGERY. The most effective treatment for obesity isbariatric surgery. Because prevalence of morbid obesity in theUSA is approximately twofold higher in women than men (226)and because women appear to suffer more from these disorders interms of quality of life (24, 84, 273, 531, 762), �80% of bariatricsurgery patients in the United States are women (568, 630). Bothanimal and human data suggest that physiological sex differencesaffect bariatric surgery outcome. 1) A retrospective analysis of�1,300 obese women who underwent either Roux-en-Y gastricbypass surgery or gastric banding indicated that presumptivelypremenopausal women (i.e., women age 20–45 yr) lost signifi-cantly more weight than presumptively postmenopausal women(i.e., age 55–65 yr of age) (522). This is consistent with thehypothesis that estrogens affect weight even in women in whombariatric surgery has massively altered gastrointestinal food han-dling. 2) Estradiol treatment further decreased eating and bodyweight after Roux-en-Y gastric bypass surgery in ovariectomizedrats (13) (Fig. 18). Thus, physiological sex differences in eatingmay be important issues in the surgical treatment of obesity.
Conclusions
We reviewed the variety and complexity of physiologicalsex differences in eating in rats, mice and anthropoid primates,including phenomenology, peripheral and central neuroendo-crine mechanisms, and pathophysiology. We discussed physi-ological sex differences in early development, puberty, and reproduc-tive adulthood and senescence. The prominence and variety ofphysiological sex differences in rats, mice, and anthropoidprimates with potential relevance for understanding normal anddisordered human eating convince us of the importance of thisarea. For example 1) the physiological sex difference in intakeof sweets that occurs in animals and women has clear potentialimplications for the development of countermeasures forworldwide epidemic of overeating and obesity, and 2) evidencefor sex-linked physiological contributions to the vulnerabilityto psychiatric eating disorders may facilitate development ofmore efficacious treatment of these disorders. Furthermore, thenumerous mechanistic approaches that already have beenbrought to bear on these and other sex differences in eating thatwe reviewed underscore their tractability to physiological andmolecular analyses in humans, as well as in animals. Finally,powerful new methods are likely to accelerate progress inseveral aspects of physiological sex differences in eating. Thisis reflected, for example, in work under way on 1) the devel-opment of HPG-axis controls of eating, 2) links between genevariations and eating, and 3) functional brain imaging.
The greatest amount of data presently available concerns theactivational effects of estrogens on eating in rats and mice. Onthe basis of these data, as well as the more limited results inanthropoid primates, we hypothesize that the population ofER�-expressing neurons in the cmNTS is an essential compo-nent of a local neuronal network that integrates a variety ofperipheral afferent signals, as well as descending diencephalic
Review
R1247SEX DIFFERENCES IN THE PHYSIOLOGY OF EATING
AJP-Regul Integr Comp Physiol • doi:10.1152/ajpregu.00446.2012 • www.ajpregu.org
by 10.220.32.247 on Septem
ber 18, 2016http://ajpregu.physiology.org/
Dow
nloaded from

and telencephalic influences to control meal size, food intake,and body weight in rats and mice, and probably in women.Fig. 19 diagrams this hypothesis and emphasizes its specula-tive nature. For example, we conclude that at present 1) CCK,ghrelin, and gustation are firmly established estrogen-sensitiveperipheral controls, and, of these, only CCK satiation is certainto be influenced by cmNTS ER�, and 2) 5HT, AgRP, andorosensory hedonics are firmly established estrogen-sensitivecentral controls, and whether they are affected by cmNTS ER�or provide descending inputs to the local neuronal networks inwhich cmNTS ER� reside, is wholly speculative. As we alsoreviewed, however, these disparate data suggest several poten-tial toeholds for efforts to test this and related hypotheses. Forexample, the apparent similarity of the roles of CCK, 5HT, and
AgRP on the phasic estrogenic inhibition of eating during theovarian cycle presents one such opportunity. Figure 19 alsomakes clear that there is an androgenic control of meal size,food intake, and body weight and that nothing is known aboutthe central mechanisms mediating it. Finally, available dataindicate that progestins do not contribute to the control ofeating.
We emphasize that the hypothesis that physiological sexdifferences in the controls of eating play causal roles in bothnormal and disordered human eating does not imply either1) that physiological sex differences in eating alone are suffi-cient to cause disordered eating or 2) that physiological sexdifferences not directly involved in eating are not important indisordered eating, for example, variants in ESR1 that arerelated to personality differences in women (482). Rather, ourview is that disordered eating results from dynamic interactionsamong numerous cultural, social, familial, and biological fac-tors to which physiological sex differences, including sexdifferences in the physiology of eating, may contribute. Fur-thermore, we believe that physiological sex differences may berelevant to the treatment of disordered eating even if they arenot causal. For example, a particular physiologically basedtherapy may be more or less efficacious in various states ofHPG function, such as in premenopausal or postmenopausalwomen.
Despite the significant progress being made on the study ofsex differences in the physiology of eating, many aspects of theproblem remain nearly untouched. Examples of such specificissues include 1) how eating is released from estrogenic inhi-bition during the luteal phase in anthropoid primates, 2) howthe activational effects of androgens on eating are mediated,3) whether and how puberty and reproductive senescenceaffect eating, and 4) how basic physiological controls of eatingcontribute to disordered human eating.
In addition to these relatively specific issues, our review alsoindicates that more integrative approaches are called for. Thereare opportunities for integrative sex-difference studies of atleast five kinds. 1) Almost no studies to date have addressedthe neuroendocrine integration of peripheral and central eating-control mechanisms, such as the integration of brain stem andtelencephalic neural mechanisms. This type of integrationcomprises one of the most exciting frontiers in the contempo-rary physiology of eating (291, 416, 494, 648, 800) and is anunfortunate lacuna in sex-difference research. 2) How hypo-thalamic and pituitary HPG-axis mechanisms, such as thosemediated by kisspeptin or GnIH, are integrated with controls ofeating mediated by gonadal steroids requires research. 3)Studies of physiological sex differences in eating must bebetter integrated with physiologies of growth, energy metabo-lism, and adiposity. Although there are certainly a number ofefforts in this direction, particularly at the molecular geneticlevel, these should be better linked to the physiology ofindividual meals, where, in our opinion, sex difference studiesare most advanced. 4) There are opportunities for more trans-lational sex-difference studies integrating research advances inanimal models and in humans. These include sex differencesin peripheral and central controls of eating, in flavor hedonics,in binge eating, and in eating-disorder susceptibility genes, aswell as many other phenomena that we reviewed. 5) Mood,anxiety, impulsivity, stress reactions, and other psychologicalprocesses often involve changes in eating and food-related
Fig. 18. Estradiol-treated ovariectomized rats (RYGB-E2) eat less (B) and gainless weight (A) after Roux-en-Y gastric-bypass surgery (RYGB) than untreatedovariectomized rats (RYGB-OIL), and these effects of estradiol were similar tothose in rats with sham RYGB surgeries (SHAM-E2 and SHAM-OIL). Ratswere fed solid, high-energy diet for 3 wk preoperatively (phase 1) and for 4 wkpostoperatively (phase 2); they were then fed Ensure Plus liquid diet for 3 wk(phase 3). #SHAM-OIL significantly different from RYGB-OIL; *RYGB-OILsignificantly different from RYGB-E2; �SHAM-OIL significantly differentfrom RYGB-E2, all P � 0.05. Reprinted from Gastroenterology, Estradiolincreases body weight loss and gut-peptide satiation after Roux-en-Y gastricbypass in ovariectomized rats, Lori Asarian, Kathrin Abegg, Nori Geary, MarcSchiesser, Thomas A. Lutz, and Marco Bueter, 143: 325–327.e2, 2012;republished with permission from Elsevier; from Asarian et al. (13).
Review
R1248 SEX DIFFERENCES IN THE PHYSIOLOGY OF EATING
AJP-Regul Integr Comp Physiol • doi:10.1152/ajpregu.00446.2012 • www.ajpregu.org
by 10.220.32.247 on Septem
ber 18, 2016http://ajpregu.physiology.org/
Dow
nloaded from

cognitions and affect. These processes also display a variety ofsex differences, and there are well-developed methods toassess them in animals and humans (6, 49, 50, 62, 183, 225,238, 542, 624, 644, 653, 658, 703, 720). The study of interac-tions among such psychological processes and physiologicalsex differences in eating seem to us to be profitable avenues ofinvestigation.
All of the problems outlined above have to be tackled tocreate an integrative regulatory physiology of HPG influenceson eating. Viewed from the perspective of these challenges,what has been accomplished seems uneven and fragmentary.Sex is inappropriately neglected in the physiology of eating, asit is in most other branches of physiology and medicine (51, 64,73, 109, 110, 466, 797, 826). Our review will have succeededif it illuminates opportunities for meaningful research andattracts more attention to them.
We close by acknowledging several limitations of the re-view. 1) We reviewed only the “consummatory phase” ofeating, i.e., eating per se, and not the “appetitive phase,” inwhich important sex differences are also to be expected. 2) Asmentioned above, our focus on biological sex differences is notmeant to suggest that there are sharp borders between purelybiological and nonbiological causes of sex differences in be-havior, and future work needs to address these gray zones.3) Our emphasis on the role of eating in determining sexdifferences in adiposity should not be taken to discount the
potential importance of sex differences in metabolism andenergy expenditure. 4) Although rat and mouse models dom-inate the study of sex differences in eating and adiposity, it isnot clear if these are always the most appropriate model speciesfor humans. For example, as we discussed, these animals’ovarian cycles do not include a period comparable to thehuman luteal phase, and mice often fail to overeat after ovari-ectomy. Furthermore, the relative importance of ER� in thecontrol of eating apparently differs among rodent species.Thus, future work should focus more on identifying whichanimal species provide the most appropriate models of partic-ular human sex differences.
ACKNOWLEDGMENTS
The authors acknowledge the invaluable intellectual and technical contri-butions of the many colleagues who have assisted us in our research: col-leagues whose helpful discussions improved our review, including Art Arnold(University of California, Los Angeles), Lisa Eckel (Florida State University),Richard Foltin (Columbia University Medical Center), Kelly Klump (MichiganState University), Lance Kriegsfeld (University of California, Berkeley),Timothy Moran (Johns Hopkins University School of Medicine), and JanetPolivy (Universtiy of Toronto). We also thank Barry E. Levin, AssociateEditor, American Journal of Physiology—Regulatory, Integrative andComparative Physiology, for the invitation to prepare this review, as wellas the three anonymous reviewers for their very helpful critiques of theinitial submission.
Fig. 19. Schematic summary of the activational effects ofgonadal steroid hormones on eating, emphasizing hypothe-sized neural mechanisms and their integration. The diagramis based on our review of rat, mouse, and anthropoidprimate, including human, data. It is superimposed on aschematic midsagittal section of a rat brain, although mostnamed structures are lateral to the midline. Red arrows andtext boxes indicate estrogenic mechanisms; question marksand black font identify less well-established mechanisms.Estrogens acting on ER� on neurons in the cmNTS (filledoval) affect the neural processing of peripheral CCK signals(solid red arrow) so as to reduce meal size, food intake, andbody weight; the same ER� neurons are hypothesized to beinvolved in the processing of a variety of other peripheralsignals, especially ghrelin and gustatory signals, which areapparently affected by estrogens (dashed red arrows). ER�in the dorsal raphe and several hypothalamic areas are notstrongly implicated in the control of eating (open red oval).A number of forebrain signaling molecules, especially 5HTand AgRP, as well as flavor hedonics contribute to theestrogenic control of eating, probably via hypothalamic andtelencephalic mechanisms (dashed red arrows). Neural pro-cessing in these areas is presumably linked bidirectionallyto the cmNTS and other brain stem areas (double-headeddashed red arrow), so that cmNTS ER� may also mediatethese effects; non-ER�-expressing AgRP neurons in theArc are the strongest candidates. Androgens acting on ARin unknown sites increase meal frequency, food intake, andbody weight (green arrows and text boxes). In contrast,progestins appear not to have physiological effects oneating (blue, dashed text box). Challenges for future mech-anistic studies of sex differences in eating include 1) estab-lishing the physiological and pathophysiological roles ofthe estrogenic mechanisms shown and 2) identifying theandrogenic mechanisms affecting eating.
Review
R1249SEX DIFFERENCES IN THE PHYSIOLOGY OF EATING
AJP-Regul Integr Comp Physiol • doi:10.1152/ajpregu.00446.2012 • www.ajpregu.org
by 10.220.32.247 on Septem
ber 18, 2016http://ajpregu.physiology.org/
Dow
nloaded from

GRANTS
The authors gratefully acknowledge research funding for our work from theU.S. National Institutes of Health Grants MH-51135 (to N. Geary), MH-65024(to N. Geary), DK-54523 (to L. Asarian and N. Geary), and DK-092608 (to L.Asarian), as well as Grant 3100-122567 from the Swiss National ScienceFoundation (to N. Geary), from the Society for Women’s Health Research (toN. Geary), and from the Novartis Foundation (to L. Asarian).
DISCLOSURES
No conflicts of interest, financial or otherwise, are declared by the authors.
AUTHOR CONTRIBUTIONS
Author contributions: L.A. and N.G. conception and design of research;L.A. and N.G. interpreted results of experiments; L.A. and N.G. preparedfigures; L.A. and N.G. edited and revised manuscript; L.A. and N.G. approvedfinal version of manuscript; N.G. drafted manuscript.
REFERENCES
1. Ahima RS, Kelly J, Elmquist JK, Flier JS. Distinct physiologic andneuronal responses to decreased leptin and mild hyperleptinemia. Endo-crinology 140: 4923–4931, 1999.
2. Ainslie DA, Morris MJ, Wittert G, Turnbull H, Proietto J, ThorburnAW. Estrogen deficiency causes central leptin insensitivity and increasedhypothalamic neuropeptide Y. Int J Obes Relat Metab Disord 25:1680–1688, 2001.
3. al-Dahan MI, Thalmann RH. Progesterone regulates gamma-aminobu-tyric acid B (GABAB) receptors in the neocortex of female rats. BrainRes 727: 40–48, 1996.
4. Alonso-Alonso M, Ziemke F, Magkos F, Barrios FA, Brinkoetter M,Boyd I, Rifkin-Graboi A, Yannakoulia M, Rojas R, Pascual-Leone A,Mantzoros CS. Brain responses to food images during the early and latefollicular phase of the menstrual cycle in healthy young women: relationto fasting and feeding. Am J Clin Nutr 94: 377–384, 2011.
5. American Psychiatric Association. Diagnostic and Statistical Manualof Mental Disorders-5. Arlington, VA: American Psychiatric Associa-tion, 2013.
6. Anestis MD, Peterson CB, Bardone-Cone AM, Klein MH, MitchellJE, Crosby RD, Wonderlich SA, Crow SJ, le Grange D, Joiner TE.Affective lability and impulsivity in a clinical sample of women withbulimia nervosa: the role of affect in severely dysregulated behavior. IntJ Eat Disord 42: 259–266, 2009.
7. Arimura C, Nozaki T, Takakura S, Kawai K, Takii M, Sudo N, KuboC. Predictors of menstrual resumption by patients with anorexia nervosa.Eat Weight Disord 15: e226–e233, 2010.
8. Arnold AP. The organizational-activational hypothesis as the foundationfor a unified theory of sexual differentiation of all mammalian tissues.Horm Behav 55: 570–578, 2009.
9. Arnold AP, Chen X, Link JC, Itoh Y, Reue K. Cell-autonomous sexdetermination outside of the gonad. Dev Dyn 242: 371–379, 2013.
10. Arnold AP, Gorski RA. Gonadal steroid induction of structural sexdifferences in the central nervous system. Annu Rev Neurosci 7: 413–442, 1984.
11. Asarian L. Loss of cholecystokinin and glucagon-like peptide-1-inducedsatiation in mice lacking serotonin 2C receptors. Am J Physiol RegulIntegr Comp Physiol 296: R51–R56, 2009.
12. Asarian L. Membrane estrogen receptors and energy homeostasis. JNeurosci 26: 11255–11256, 2006.
13. Asarian L, Abegg K, Geary N, Schiesser M, Lutz TA, Bueter M.Estradiol increases body weight loss and gut-peptide satiation afterRoux-en-Y gastric bypass in ovariectomized rats. Gastroenterology 143:325–7.e2, 2012.
14. Asarian L, Boyle CN, Lutz TA. Estradiol increases the acute eating-inhibitory effect of amlyin in ovariectomized rats (Abstract). Appetite 57:S2, 2011.
15. Asarian L, Geary N. Cyclic estradiol treatment normalizes body weightand restores physiological patterns of spontaneous feeding and sexualreceptivity in ovariectomized rats. Horm Behav 42: 461–471, 2002.
16. Asarian L, Geary N. Cyclic estradiol treatment phasically potentiatesendogenous cholecystokinin’s satiating action in ovariectomized rats.Peptides 20: 445–450, 1999.
17. Asarian L, Geary N. Estradiol enhances cholecystokinin-dependentlipid-induced satiation and activates estrogen receptor-alpha-expressing
cells in the nucleus tractus solitarius of ovariectomized rats. Endocrinol-ogy 148: 5656–5666, 2007.
18. Asarian L, Geary N. Modulation of appetite by gonadal steroid hor-mones. Philos Trans R Soc Lond B Biol Sci 361: 1251–1263, 2006.
19. Asarian L, Thammacharoen S, Geary N, Clegg DJ, Ogawa S, LutzTA. Knock-down of estrogen receptor-� neurons in the nucleus tractussolitarii eliminates CCK-induced c-Fos expression in the paraventricularnucleus of the hypothalamus (Abstract). Appetite 54: 633, 2010.
20. Asarian L, Thammacharoen S, Lutz TA, Geary N. Selective RNAiknockdown of estrogen-receptor-� neurons in the nuceus tractus solitariieliminates estradiol’s inhibitory effects on food intake in ovareictomizedrats (Abstract). Appetite 52: 817, 2009.
21. Astrup A, Rossner S, Van Gaal L, Rissanen A, Niskanen L, AlHakim M, Madsen J, Rasmussen MF, Lean ME. Effects of liraglutidein the treatment of obesity: a randomised, double-blind, placebo-con-trolled study. Lancet 374: 1606–1616, 2009.
22. Atasoy D, Betley JN, Su HH, Sternson SM. Deconstruction of a neuralcircuit for hunger. Nature 488: 172–177, 2012.
23. Atchley DP, Weaver KL, Eckel LA. Taste responses to dilute sucrosesolutions are modulated by stage of the estrous cycle and fenfluraminetreatment in female rats. Physiol Behav 86: 265–271, 2005.
24. Atlantis E, Baker M. Obesity effects on depression: systematic reviewof epidemiological studies. Int J Obes (Lond) 32: 881–891, 2008.
25. Augustine RA, Grattan DR. Induction of central leptin resistance inhyperphagic pseudopregnant rats by chronic prolactin infusion. Endocri-nology 149: 1049–1055, 2008.
26. Augustine RA, Ladyman SR, Grattan DR. From feeding one tofeeding many: hormone-induced changes in bodyweight homeostasisduring pregnancy. J Physiol 586: 387–397, 2008.
27. Austin SB, Ziyadeh NJ, Forman S, Prokop LA, Keliher A, Jacobs D.Screening high school students for eating disorders: results of a nationalinitiative. Prev Chronic Dis 5: A114, 2008.
28. Aydin ZD. Determinants of age at natural menopause in the IspartaMenopause and Health Study: premenopausal body mass index gain rateand episodic weight loss. Menopause 17: 494–505, 2010.
29. Babbs RK, Wojnicki FH, Corwin RL. Effect of 2-hydroxyestradiol onbinge intake in rats. Physiol Behav 103: 508–512, 2011.
30. Baker JH, Girdler SS, Bulik CM. The role of reproductive hormones inthe development and maintenance of eating disorders. Expert Rev ObstetGynecol 7: 573–583, 2012.
31. Baker JH, Maes HH, Lissner L, Aggen SH, Lichtenstein P, KendlerKS. Genetic risk factors for disordered eating in adolescent males andfemales. J Abnorm Psychol 118: 576–586, 2009.
32. Bakker J, Baum MJ. Role for estradiol in female-typical brain andbehavioral sexual differentiation. Front Neuroendocrinol 29: 1–16, 2008.
33. Bakker J, De Mees C, Douhard Q, Balthazart J, Gabant P, SzpirerJ, Szpirer C. Alpha-fetoprotein protects the developing female mousebrain from masculinization and defeminization by estrogens. Nat Neu-rosci 9: 220–226, 2006.
34. Bakker J, Honda S, Harada N, Balthazart J. The aromatase knock-outmouse provides new evidence that estradiol is required during develop-ment in the female for the expression of sociosexual behaviors inadulthood. J Neurosci 22: 9104–9112, 2002.
35. Baldo BA, Kelley AE. Discrete neurochemical coding of distinguishablemotivational processes: insights from nucleus accumbens control offeeding. Psychopharmacology (Berl) 191: 439–459, 2007.
36. Balthasar N, Dalgaard LT, Lee CE, Yu J, Funahashi H, Williams T,Ferreira M, Tang V, McGovern RA, Kenny CD, Christiansen LM,Edelstein E, Choi B, Boss O, Aschkenasi C, Zhang CY, Mountjoy K,Kishi T, Elmquist JK, Lowell BB. Divergence of melanocortin path-ways in the control of food intake and energy expenditure. Cell 123:493–505, 2005.
37. Barber TM, McCarthy MI, Wass JA, Franks S. Obesity and polycys-tic ovary syndrome. Clin Endocrinol (Oxf) 65: 137–145, 2006.
38. Barr SI, Janelle KC, Prior JC. Energy intakes are higher during theluteal phase of ovulatory menstrual cycles. Am J Clin Nutr 61: 39–43,1995.
39. Barrera JG, Jones KR, Herman JP, D’Alessio DA, Woods SC, SeeleyRJ. Hyperphagia and increased fat accumulation in two models ofchronic CNS glucagon-like peptide-1 loss of function. J Neurosci 31:3904–3913, 2011.
40. Barros RP, Gustafsson JA. Estrogen receptors and the metabolicnetwork. Cell Metab 14: 289–299, 2011.
Review
R1250 SEX DIFFERENCES IN THE PHYSIOLOGY OF EATING
AJP-Regul Integr Comp Physiol • doi:10.1152/ajpregu.00446.2012 • www.ajpregu.org
by 10.220.32.247 on Septem
ber 18, 2016http://ajpregu.physiology.org/
Dow
nloaded from

41. Bartness TJ, Keen-Rhinehart E, Dailey MJ, Teubner BJ. Neural andhormonal control of food hoarding. Am J Physiol Regul Integr CompPhysiol 301: R641–R655, 2011.
42. Bartness TJ, Waldbillig RJ. Dietary self-selection in intact, ovariecto-mized, and estradiol-treated female rats. Behav Neurosci 98: 125–137,1984.
43. Bartoshuk LM. The psychophysics of taste. Am J Clin Nutr 31:1068–1077, 1978.
44. Bartoshuk LM, Duffy VB, Green BG, Hoffman HJ, Ko CW, Luc-china LA, Marks LE, Snyder DJ, Weiffenbach JM. Valid across-group comparisons with labeled scales: the gLMS versus magnitudematching. Physiol Behav 82: 109–114, 2004.
45. Bartoshuk LM, Duffy VB, Hayes JE, Moskowitz HR, Snyder DJ.Psychophysics of sweet and fat perception in obesity: problems, solutionsand new perspectives. Philos Trans R Soc Lond B Biol Sci 361: 1137–1148, 2006.
46. Bartoshuk LM, Duffy VB, Miller IJ. PTC/PROP tasting: anatomy,psychophysics, and sex effects. Physiol Behav 56: 1165–1171, 1994.
47. Beal VA. Nutritional studies during pregnancy. I. Changes in intakes ofcalories, carbohydrates, fat, protein, and calcium. J Am Diet Assoc 58:312–320, 1971.
48. Becker JB, Arnold AP, Berkley KJ, Blaustein JD, Eckel LA, Hamp-son E, Herman JP, Marts S, Sadee W, Steiner M, Taylor J, Young E.Strategies and methods for research on sex differences in brain andbehavior. Endocrinology 146: 1650–1673, 2005.
49. Becker JB, Hu M. Sex differences in drug abuse. Front Neuroendocri-nol 29: 36–47, 2008.
50. Becker JB, Perry AN, Westenbroek C. Sex differences in the neuralmechanisms mediating addiction: a new synthesis and hypothesis. BiolSex Differ 3: 14, 2012.
51. Beery AK, Zucker I. Sex bias in neuroscience and biomedical research.Neurosci Biobehav Rev 35: 565–572, 2011.
52. Beglinger C, Degen L. Fat in the intestine as a regulator of appetite—role of CCK. Physiol Behav 83: 617–621, 2004.
53. Beglinger C, Degen L. Gastrointestinal satiety signals in humans—physiological roles for GLP-1 and PYY? Physiol Behav 89: 460–464,2006.
54. Begriche K, Sutton GM, Butler AA. Homeostastic and non-homeo-static functions of melanocortin-3 receptors in the control of energybalance and metabolism. Physiol Behav 104: 546–554, 2011.
55. Behre HM, Kuhlage J, Gassner C, Sonntag B, Schem C, SchneiderHP, Nieschlag E. Prediction of ovulation by urinary hormone measure-ments with the home use ClearPlan Fertility Monitor: comparison withtransvaginal ultrasound scans and serum hormone measurements. HumReprod 15: 2478–2482, 2000.
56. Bell DD, Zucker I. Sex differences in body weight and eating: organi-zation and activation by gonadal hormones in the rat. Physiol Behav 7:27–34, 1971.
57. Bello NT, Patinkin ZW, Moran TH. Opioidergic consequences ofdietary-induced binge eating. Physiol Behav 104: 98–104, 2011.
58. Belzer LM, Smulian JC, Lu SE, Tepper BJ. Food cravings and intakeof sweet foods in healthy pregnancy and mild gestational diabetesmellitus. A prospective study. Appetite 55: 609–615, 2010.
59. Benedict C, Kern W, Schultes B, Born J, Hallschmid M. Differentialsensitivity of men and women to anorexigenic and memory-improvingeffects of intranasal insulin. J Clin Endocrinol Metab 93: 1339–1344,2008.
60. Bennink R, Peeters M, Van den Maegdenbergh V, Geypens B,Rutgeerts P, De Roo M, Mortelmans L. Comparison of total andcompartmental gastric emptying and antral motility between healthy menand women. Eur J Nucl Med 25: 1293–1299, 1998.
61. Berbesque JC, Marlowe FW. Sex differences in food preferences ofHazda hunter-gatherers. Evol Psychol 7: 601–616, 2009.
62. Berg KC, Crosby RD, Cao L, Peterson CB, Engel SG, Mitchell JE,Wonderlich SA. Facets of negative affect prior to and following binge-only, purge-only, and binge/purge events in women with bulimia ner-vosa. J Abnorm Psychol 122: 111–118, 2013.
63. Bergman L, Eriksson E. Marked symptom reduction in two womenwith bulimia nervosa treated with the testosterone receptor antagonistflutamide. Acta Psychiatr Scand 94: 137–139, 1996.
64. Berkley KJ. Vive la difference! Trends Neurosci 15: 331–332, 1992.65. Bernstein IL. Taste aversion learning: a contemporary perspective.
Nutrition 15: 229–234, 1999.
66. Berridge KC, Kringelbach ML. Affective neuroscience of pleasure:reward in humans and animals. Psychopharmacology (Berl) 199: 457–480, 2008.
67. Berthoud HR. Multiple neural systems controlling food intake and bodyweight. Neurosci Biobehav Rev 26: 393–428, 2002.
68. Berthoud HR, Lenard NR, Shin AC. Food reward, hyperphagia, andobesity Am J Physiol Regul Integr Comp Physiol 300: R1266–R1277,2011.
69. Berthoud HR, Munzberg H, Richards BK, Morrison CD. Neural andmetabolic regulation of macronutrient intake and selection. Proc NutrSoc 71: 390–400, 2012.
70. Bielert C, Busse C. Influences of ovarian hormones on the food intakeand feeding of captive and wild female chacma baboons (Papio ursinus).Physiol Behav 30: 103–111, 1983.
71. Black AE, Wiles SJ, Paul AA. The nutrient intakes of pregnant andlactating mothers of good socio-economic status in Cambridge, UK:some implications for recommended daily allowances of minor nutrients.Br J Nutr 56: 59–72, 1986.
72. Blandau JR, Boling JL, Young WG. The length of heat in the albino ratas determined by the copulatory response. Anat Rec 79: 453–463, 1941.
73. Blaustein JD. Animals have a sex, and so should titles and methodssections of articles in endocrinology. Endocrinology 153: 2539–2540,2012.
74. Blaustein JD. An estrogen by any other name. Endocrinology 149:2697–2698, 2008.
75. Blaustein JD, Gentry RT, Roy EJ, Wade GN. Effects of ovariectomyand estradiol on body weight and food intake in gold thioglucose-treatedmice. Physiol Behav 17: 1027–1030, 1976.
76. Blaustein JD, Wade GN. Ovarian hormones and meal patterns in rats:effects of progesterone and role of gastrointestinal transit. Physiol Behav19: 23–27, 1977.
77. Blaustein JD, Wade GN. Ovarian influences on the meal patterns offemale rats. Physiol Behav 17: 201–208, 1976.
78. Bleich SN, Wang YC, Wang Y, Gortmaker SL. Increasing consump-tion of sugar-sweetened beverages among US adults: 1988–1994 to1999–2004. Am J Clin Nutr 89: 372–381, 2009.
79. Boakes RA, Mills KJ, Single JP. Sex differences in the relationshipbetween activity and weight loss in the rat. Behav Neurosci 113: 1080–1089, 1999.
80. Bowen DJ. Possible explanations for excess weight gains in pregnancy:an animal model. Physiol Behav 46: 935–939, 1989.
81. Bowen DJ. Taste and food preference changes across the course ofpregnancy. Appetite 19: 233–242, 1992.
82. Bowen DJ, Grunberg NE. Variations in food preference and consump-tion across the menstrual cycle. Physiol Behav 47: 287–291, 1990.
83. Brawer JR, Beaudet A, Desjardins GC, Schipper HM. Pathologiceffect of estradiol on the hypothalamus. Biol Reprod 49: 647–652, 1993.
84. Bray GA. Medical consequences of obesity. J Clin Endocrinol Metab89: 2583–2589, 2004.
85. Brennan IM, Feltrin KL, Nair NS, Hausken T, Little TJ, GentilcoreD, Wishart JM, Jones KL, Horowitz M, Feinle-Bisset C. Effects of thephases of the menstrual cycle on gastric emptying, glycemia, plasmaGLP-1 and insulin, and energy intake in healthy lean women. Am JPhysiol Gastrointest Liver Physiol 297: G602–G610, 2009.
86. Broberg DJ, Dorsa DM, Bernstein IL. Nausea in bulimic women inresponse to a palatable food. J Abnorm Psychol 99: 183–188, 1990.
87. Broberger C, Johansen J, Johansson C, Schalling M, Hokfelt T. Theneuropeptide Y/agouti gene-related protein (AGRP) brain circuitry innormal, anorectic, and monosodium glutamate-treated mice. Proc NatlAcad Sci USA 95: 15043–15048, 1998.
88. Brock O, Baum MJ, Bakker J. The development of female sexualbehavior requires prepubertal estradiol. J Neurosci 31: 5574–5578, 2011.
89. Broft AI, Berner LA, Martinez D, Walsh BT. Bulimia nervosa andevidence for striatal dopamine dysregulation: a conceptual review.Physiol Behav 104: 122–127, 2011.
90. Brown-Grant K, Exley D, Naftolin F. Peripheral plasma oestradiol andluteinizing hormone concentrations during the oestrous cycle of the rat.J Endocrinol 48: 295–296, 1970.
91. Brüning JC, Gautam D, Burks DJ, Gillette J, Schubert M, OrbanPC, Klein R, Krone W, Müller-Wieland D, Kahn CR. Role of braininsulin receptor in control of body weight and reproduction. Science 289:2122–2125, 2000.
Review
R1251SEX DIFFERENCES IN THE PHYSIOLOGY OF EATING
AJP-Regul Integr Comp Physiol • doi:10.1152/ajpregu.00446.2012 • www.ajpregu.org
by 10.220.32.247 on Septem
ber 18, 2016http://ajpregu.physiology.org/
Dow
nloaded from

92. Buffenstein R, Poppitt SD, McDevitt RM, Prentice AM. Food intakeand the menstrual cycle: a retrospective analysis, with implications forappetite research. Physiol Behav 58: 1067–1077, 1995.
93. Bulik CM, Sullivan PF, Kendler KS. Genetic and environmentalcontributions to obesity and binge eating. Int J Eat Disord 33: 293–298,2003.
94. Bulik CM, Sullivan PF, Kendler KS. Heritability of binge-eating andbroadly defined bulimia nervosa. Biol Psychiatry 44: 1210–1218, 1998.
95. Bulik CM, Sullivan PF, Tozzi F, Furberg H, Lichtenstein P, PedersenNL. Prevalence, heritability, and prospective risk factors for anorexianervosa. Arch Gen Psychiatry 63: 305–312, 2006.
96. Bulik CM, Sullivan PF, Wade TD, Kendler KS. Twin studies of eatingdisorders: a review. Int J Eat Disord 27: 1–20, 2000.
97. Bulun SE, Adashi EY. The physiology and pathophysiology of thefemale reproductive tract. In: Williams Textbook of Endocrinology (10thed.), edited by Larsen PR, Kronenberg HM, Melmed S, and PolonskyKS. Philadelphia, PA: Elsevier/Saunders, 2003, p. 587–664.
98. Burger H. The menopausal transition—endocrinology. J Sex Med 5:2266–2273, 2008.
99. Burger HG, Hale GE, Dennerstein L, Robertson DM. Cycle andhormone changes during perimenopause: the key role of ovarian func-tion. Menopause 15: 603–612, 2008.
100. Burton-Freeman B, Davis PA, Schneeman BO. Interaction of fatavailability and sex on postprandial satiety and cholecystokinin aftermixed-food meals. Am J Clin Nutr 80: 1207–1214, 2004.
101. Butcher RL, Collins WE, Fugo NW. Plasma concentration of LH, FSH,prolactin, progesterone and estradiol-17� throughout the 4-day estrouscycle of the rat. Endocrinology 94: 1704–1708, 1974.
102. Butera PC. Estradiol and the control of food intake. Physiol Behav 99:175–180, 2010.
103. Butera PC, Beikirch RJ. Central implants of diluted estradiol: indepen-dent effects on ingestive and reproductive behaviors of ovariectomizedrats. Brain Res 491: 266–273, 1989.
104. Butera PC, Beikirch RJ, Willard DM. Changes in ingestive behaviorsand body weight following intracranial application of 17�-estradiol.Physiol Behav 47: 1291–1293, 1990.
105. Butera PC, Bradway DM, Cataldo NJ. Modulation of the satiety effectof cholecystokinin by estradiol. Physiol Behav 53: 1235–1238, 1993.
106. Butera PC, Willard DM, Raymond SA. Effects of PVN lesions on theresponsiveness of female rats to estradiol. Brain Res 576: 304–310,1992.
107. Butera PC, Wojcik DM, Clough SJ. Effects of estradiol on food intakeand meal patterns for diets that differ in flavor and fat content. PhysiolBehav 99: 142–145, 2010.
108. Butera PC, Xiong M, Davis RJ, Platania SP. Central implants of diluteestradiol enhance the satiety effect of CCK-8. Behav Neurosci 110:823–830, 1996.
109. Cahill L. A half-truth is a whole lie: on the necessity of investigating sexinfluences on the brain. Endocrinology 153: 2541–2543, 2012.
110. Cahill L. Why sex matters for neuroscience. Nat Rev Neurosci 7:477–484, 2006.
111. Carnell S, Gibson C, Benson L, Ochner CN, Geliebter A. Neuroim-aging and obesity: current knowledge and future directions. Obes Rev 13:43–56, 2012.
112. Carroll JF, Kaiser KA, Franks SF, Deere C, Caffrey JL. Influence ofBMI and gender on postprandial hormone responses. Obesity (SilverSpring) 15: 2974–2983, 2007.
113. Casper RC, Sullivan EL, Tecott L. Relevance of animal models tohuman eating disorders and obesity. Psychopharmacology (Berl) 199:313–329, 2008.
114. Cecchini DJ, Chattoraj SC, Fanous AS, Panda SK, Brennan TF,Edelin KC. Radioimmunoassay of 2-hydroxyestrone in plasma duringthe estrous cycle of the rat: interrelationships with estradiol, progester-one, and the gonadotropins. Endocrinology 112: 1122–1126, 1983.
115. Chai JK, Blaha V, Meguid MM, Laviano A, Yang ZJ, Varma M. Useof orchiectomy and testosterone replacement to explore meal number-to-meal size relationship in male rats. Am J Physiol Regul Integr CompPhysiol 276: R1366–R1373, 1999.
116. Chambers KC. Hormonal influences on sexual dimorphism in rate ofextinction of a conditioned taste aversion in rats. J Comp Physiol Psychol90: 851–856, 1976.
117. Chen H, Hardy MP, Zirkin BR. Age-related decreases in Leydig celltestosterone production are not restored by exposure to LH in vitro.Endocrinology 143: 1637–1642, 2002.
118. Chen H, Zirkin BR. Long-term suppression of Leydig cell steroidogen-esis prevents Leydig cell aging. Proc Natl Acad Sci USA 96: 14877–14881, 1999.
119. Chen P, Williams SM, Grove KL, Smith MS. Melanocortin 4 receptor-mediated hyperphagia and activation of neuropeptide Y expression in thedorsomedial hypothalamus during lactation. J Neurosci 24: 5091–5100,2004.
120. Chen TS, Doong ML, Chang FY, Lee SD, Wang PS. Effects of sexsteroid hormones on gastric emptying and gastrointestinal transit in rats.Am J Physiol Gastrointest Liver Physiol 268: G171–G176, 1995.
121. Chen TS, Doong ML, Wang SW, Tsai SC, Lu CC, Shih HC, ChenYH, Chang FY, Lee SD, Wang PS. Gastric emptying and gastrointes-tinal transit during lactation in rats. Am J Physiol Gastrointest LiverPhysiol 272: G626–G631, 1997.
122. Chen X, McClusky R, Chen J, Beaven SW, Tontonoz P, Arnold AP,Reue K. The number of X chromosomes causes sex differences inadiposity in mice. PLoS Genet 8: e1002709, 2012.
123. Chen X, McClusky R, Itoh Y, Reue K, Arnold AP. X and Ychromosome complement influence adiposity and metabolism in mice.Endocrinology 154: 1092–1104, 2013.
124. Chen Y, Heiman ML. Increased weight gain after ovariectomy is not aconsequence of leptin resistance. Am J Physiol Endocrinol Metab 280:E315–E322, 2001.
125. Cheskis BJ, Greger JG, Nagpal S, Freedman LP. Signaling byestrogens. J Cell Physiol 213: 610–617, 2007.
126. Chow SY, Sakai RR, Witcher JA, Adler NT, Epstein AN. Sex andsodium intake in the rat. Behav Neurosci 106: 172–180, 1992.
127. Clarke IJ, Smith JT, Henry BA, Oldfield BJ, Stefanidis A, Millar RP,Sari IP, Chng K, Fabre-Nys C, Caraty A, Ang BT, Chan L, FraleyGS. Gonadotropin-inhibitory hormone is a hypothalamic peptide thatprovides a molecular switch between reproduction and feeding. Neuroen-docrinology 95: 305–316, 2012.
128. Clarke TK, Weiss AR, Berrettini WH. The genetics of anorexianervosa. Clin Pharmacol Ther 91: 181–188, 2012.
129. Clegg DJ, Brown LM, Woods SC, Benoit SC. Gonadal hormonesdetermine sensitivity to central leptin and insulin. Diabetes 55: 978–987,2006.
130. Clegg DJ, Brown LM, Zigman JM, Kemp CJ, Strader AD, BenoitSC, Woods SC, Mangiaracina M, Geary N. Estradiol-dependent de-crease in the orexigenic potency of ghrelin in female rats. Diabetes 56:1051–1058, 2007.
131. Clegg DJ, Riedy CA, Smith KA, Benoit SC, Woods SC. Differentialsensitivity to central leptin and insulin in male and female rats. Diabetes52: 682–687, 2003.
132. Cohen IT, Sherwin BB, Fleming AS. Food cravings, mood, and themenstrual cycle. Horm Behav 21: 457–470, 1987.
133. Cohen LR, Woodside BC. Self-selection of protein during pregnancyand lactation in rats. Appetite 12: 119–136, 1989.
134. Cone RD, Low MJ, Elmquist JE, Cameron JL. Neuroendocrinology.In: William’s Textbook of Endocrinology (10th ed.), edited by Larsen PR,Kronenberg, HM, Melmed S, and Polonsky KS. Philadephia, PA:Elsevier/Saunders, 2003, p. 81–176.
135. Cornier MA, Salzberg AK, Endly DC, Bessesen DH, Tregellas JR.Sex-based differences in the behavioral and neuronal responses to food.Physiol Behav 99: 538–543, 2010.
136. Corp ES, McQuade J, Moran TH, Smith GP. Characterization of typeA and type B CCK receptor binding sites in rat vagus nerve. Brain Res623: 161–166, 1993.
137. Corsica JA, Spring BJ. Carbohydrate craving: a double-blind, placebo-controlled test of the self-medication hypothesis. Eat Behav 9: 447–454,2008.
138. Corwin RL, Avena NM, Boggiano MM. Feeding and reward: perspec-tives from three rat models of binge eating. Physiol Behav 104: 87–97,2011.
139. Corwin RL, Buda-Levin A. Behavioral models of binge-type eating.Physiol Behav 82: 123–130, 2004.
140. Couse JF, Korach KS. Estrogen receptor null mice: what have welearned and where will they lead us? Endocr Rev 20: 358–417, 1999.
141. Craig AD. How do you feel—now? The anterior insula and humanawareness. Nat Rev Neurosci 10: 59–70, 2009.
142. Craig AD. A rat is not a monkey is not a human: comment on Mogil[Nature Rev. Neurosci 10, 283–294 (2009)] Nat Rev Neurosci 10: 466,2009.
Review
R1252 SEX DIFFERENCES IN THE PHYSIOLOGY OF EATING
AJP-Regul Integr Comp Physiol • doi:10.1152/ajpregu.00446.2012 • www.ajpregu.org
by 10.220.32.247 on Septem
ber 18, 2016http://ajpregu.physiology.org/
Dow
nloaded from

143. Craig AD. Significance of the insula for the evolution of human aware-ness of feelings from the body. Ann NY Acad Sci 1225: 72–82, 2011.
144. Crane JM, White J, Murphy P, Burrage L, Hutchens D. The effect ofgestational weight gain by body mass index on maternal and neonataloutcomes. J Obstet Gynaecol Can 31: 28–35, 2009.
145. Croll J, Neumark-Sztainer D, Story M, Ireland M. Prevalence andrisk and protective factors related to disordered eating behaviors amongadolescents: relationship to gender and ethnicity. J Adolesc Health 31:166–175, 2002.
146. Culbert KM, Breedlove SM, Burt SA, Klump KL. Prenatal hormoneexposure and risk for eating disorders: a comparison of opposite-sex andsame-sex twins. Arch Gen Psychiatry 65: 329–336, 2008.
147. Culbert KM, Breedlove SM, Sisk CL, Burt A, Klump KL. Theemergence of sex differences in risk for disordered eating attitudes duringpuberty: A role for prenatal testosterone exposure. J Abnorm Psychol122: 420–432, 2013.
148. Culbert KM, Burt SA, McGue M, Iacono WG, Klump KL. Pubertyand the genetic diathesis of disordered eating attitudes and behaviors. JAbnorm Psychol 118: 788–796, 2009.
149. Culbert KM, Racine SE, Klump KL. The influence of gender andpuberty on the heritability of disordered eating symptoms. Curr TopBehav Neurosci 6: 177–185, 2011.
150. Currie PJ, Braver M, Mirza A, Sricharoon K. Sex differences in thereversal of fluoxetine-induced anorexia following raphe injections of8-OH-DPAT. Psychopharmacology (Berl) 172: 359–364, 2004.
151. Curtis KS. Estrogen and the central control of body fluid balance.Physiol Behav 97: 180–192, 2009.
152. Curtis KS, Davis LM, Johnson AL, Therrien KL, Contreras RJ. Sexdifferences in behavioral taste responses to and ingestion of sucrose andNaCl solutions by rats. Physiol Behav 80: 657–664, 2004.
153. Czaja JA. Food rejection by female rhesus monkeys during the men-strual cycle and early pregnancy. Physiol Behav 14: 579–587, 1975.
154. Czaja JA. Ovarian influences on primate food intake: assessment ofprogesterone actions. Physiol Behav 21: 923–928, 1978.
155. Czaja JA, Butera PC, McCaffrey TA. Independent effects of estradiolon water and food intake. Behav Neurosci 97: 210–220, 1983.
156. Czaja JA, Goy RW. Ovarian hormones and food intake in female guineapigs and rhesus monkeys. Horm Behav 6: 329–349, 1975.
157. Dafopoulos K, Sourlas D, Kallitsaris A, Pournaras S, Messinis IE.Blood ghrelin, resistin, and adiponectin concentrations during the normalmenstrual cycle. Fertil Steril 92: 1389–1394, 2009.
158. Dagnault A, Richard D. Involvement of the medial preoptic area in theanorectic action of estrogens. Am J Physiol Regul Integr Comp Physiol272: R311–R317, 1997.
159. Dagnault A, Richard D. Lesions of hypothalamic paraventricular nucleido not prevent the effect of estradiol on energy and fat balance. Am JPhysiol Endocrinol Metab 267: E32–E38, 1994.
160. Dalla C, Shors TJ. Sex differences in learning processes of classical andoperant conditioning. Physiol Behav 97: 229–238, 2009.
161. Dandona P, Dhindsa S. Update: Hypogonadotropic hypogonadism intype 2 diabetes and obesity. J Clin Endocrinol Metab 96: 2643–2651,2011.
162. Danilovich N, Babu PS, Xing W, Gerdes M, Krishnamurthy H,Sairam MR. Estrogen deficiency, obesity, and skeletal abnormalities infollicle-stimulating hormone receptor knockout (FORKO) female mice.Endocrinology 141: 4295–4308, 2000.
163. Datz FL, Christian PE, Moore J. Gender-related differences in gastricemptying. J Nucl Med 28: 1204–1207, 1987.
164. Davis C, Levitan RD, Yilmaz Z, Kaplan AS, Carter JC, Kennedy JL.Binge eating disorder and the dopamine D2 receptor: genotypes andsub-phenotypes. Prog Neuropsychopharmacol Biol Psychiatry 38: 328–335, 2012.
165. Davis CA, Levitan RD, Reid C, Carter JC, Kaplan AS, Patte KA,King N, Curtis C, Kennedy JL. Dopamine for “wanting” and opioidsfor “liking”: a comparison of obese adults with and without binge eating.Obesity (Silver Spring) 17: 1220–1225, 2009.
166. Davis JD, Smith GP, Miesner J. Postpyloric stimuli are necessary forthe normal control of meal size in real feeding and sham feeding rats. AmJ Physiol Regul Integr Comp Physiol 265: R888–R895, 1993.
167. Davis JD, Smith GP, Sayler JL. Reduction of intake in the rat due togastric filling. Am J Physiol Regul Integr Comp Physiol 272: R1599–R1605, 1997.
168. Davis PG, Krieger MS, Barfield RJ, McEwen BS, Pfaff DW. The siteof action of intrahypothalamic estrogen implants in feminine sexual
behavior: an autoradiographic analysis. Endocrinology 111: 1581–1586,1982.
169. Davis PG, McEwen BS, Pfaff DW. Localized behavioral effects oftritiated estradiol implants in the ventromedial hypothalamus of femalerats. Endocrinology 104: 898–903, 1979.
170. Davy BM, Van Walleghen EL, Orr JS. Sex differences in acute energyintake regulation. Appetite 49: 141–147, 2007.
171. Davy SR, Benes BA, Driskell JA. Sex differences in dieting trends,eating habits, and nutrition beliefs of a group of midwestern collegestudents. J Am Diet Assoc 106: 1673–1677, 2006.
172. de Castro JM, Kreitzman SM. A microregulatory analysis of sponta-neous human feeding patterns. Physiol Behav 35: 329–335, 1985.
173. De Silva A, Salem V, Matthews PM, Dhillo WS. The use of functionalMRI to study appetite control in the CNS. Exp Diabetes Res 2012:764017, 2012.
174. De Vries GJ, Rissman EF, Simerly RB, Yang LY, Scordalakes EM,Auger CJ, Swain A, Lovell-Badge R, Burgoyne PS, Arnold AP. Amodel system for study of sex chromosome effects on sexually dimorphicneural and behavioral traits. J Neurosci 22: 9005–9014, 2002.
175. Dedes I. Kisspeptins and the control of gonadotrophin secretion. SystBiol Reprod Med 58: 121–128, 2012.
176. Del Parigi A, Chen K, Gautier JF, Salbe AD, Pratley RE, RavussinE, Reiman EM, Tataranni PA. Sex differences in the human brain’sresponse to hunger and satiation. Am J Clin Nutr 75: 1017–1022, 2002.
177. Dempfle A, Hinney A, Heinzel-Gutenbrunner M, Raab M, Geller F,Gudermann T, Schafer H, Hebebrand J. Large quantitative effect ofmelanocortin-4 receptor gene mutations on body mass index. J MedGenet 41: 795–800, 2004.
178. Deroo BJ, Korach KS. Estrogen receptors and human disease. J ClinInvest 116: 561–570, 2006.
179. Desjardins GC, Brawer JR, Beaudet A. Estradiol is selectively neu-rotoxic to hypothalamic �-endorphin neurons. Endocrinology 132: 86–93, 1993.
180. Devlin MJ, Walsh BT, Guss JL, Kissileff HR, Liddle RA, Petkova E.Postprandial cholecystokinin release and gastric emptying in patientswith bulimia nervosa. Am J Clin Nutr 65: 114–120, 1997.
181. Di Lorenzo PM, Monroe S. Taste responses in the parabrachial pons ofmale, female and pregnant rats. Brain Res Bull 23: 219–227, 1989.
182. Di Lorenzo PM, Monroe S. Taste responses in the parabrachial pons ofovariectomized rats. Brain Res Bull 25: 741–748, 1990.
183. DiLeone RJ, Taylor JR, Picciotto MR. The drive to eat: comparisonsand distinctions between mechanisms of food reward and drug addiction.Nat Neurosci 15: 1330–1335, 2012.
184. Dimarco NM, Dart L, Sanborn CB. Modified activity-stress paradigmin an animal model of the female athlete triad. J Appl Physiol 103:1469–1478, 2007.
185. Dixon DP, Ackert AM, Eckel LA. Development of, and recovery from,activity-based anorexia in female rats. Physiol Behav 80: 273–279, 2003.
186. Doerries LE, Stanley EZ, Aravich PF. Activity-based anorexia: rela-tionship to gender and activity-stress ulcers. Physiol Behav 50: 945–949,1991.
187. Donato J Jr, Cravo RM, Frazao R, Elias CF. Hypothalamic sites ofleptin action linking metabolism and reproduction. Neuroendocrinology93: 9–18, 2011.
188. Dovey TM, Staples PA, Gibson EL, Halford JC. Food neophobia and‘picky/fussy’ eating in children: a review. Appetite 50: 181–193, 2008.
189. Downs JL, Wise PM. The role of the brain in female reproductive aging.Mol Cell Endocrinol 299: 32–38, 2009.
190. Drewett RF. The meal patterns of the oestrous cycle and their motiva-tional significance. Q J Exp Psychol 26: 489–494, 1974.
191. Drewett RF. Oestrous and dioestrous components of the ovarian inhi-bition on hunger in the rat. Anim Behav 21: 772–780, 1973.
192. Drewnowski A, Krahn DD, Demitrack MA, Nairn K, Gosnell BA.Naloxone, an opiate blocker, reduces the consumption of sweet high-fatfoods in obese and lean female binge eaters. Am J Clin Nutr 61:1206–1212, 1995.
193. Drewnowski A, Krahn DD, Demitrack MA, Nairn K, Gosnell BA.Taste responses and preferences for sweet high-fat foods: evidence foropioid involvement. Physiol Behav 51: 371–379, 1992.
194. Drewnowski A, Kurth C, Holden-Wiltse J, Saari J. Food preferencesin human obesity: carbohydrates versus fats. Appetite 18: 207–221, 1992.
195. Duffey KJ, Popkin BM. Energy density, portion size, and eatingoccasions: contributions to increased energy intake in the United States,1977–2006. PLoS Med 8: e1001050, 2011.
Review
R1253SEX DIFFERENCES IN THE PHYSIOLOGY OF EATING
AJP-Regul Integr Comp Physiol • doi:10.1152/ajpregu.00446.2012 • www.ajpregu.org
by 10.220.32.247 on Septem
ber 18, 2016http://ajpregu.physiology.org/
Dow
nloaded from

196. Duffy VB, Bartoshuk LM, Striegel-Moore R, Rodin J. Taste changesacross pregnancy. Ann NY Acad Sci 855: 805–809, 1998.
197. Dufour DL, Sauther ML. Comparative and evolutionary dimensions ofthe energetics of human pregnancy and lactation. Am J Hum Biol 14:584–602, 2002.
198. Dye L, Blundell JE. Menstrual cycle and appetite control: implicationsfor weight regulation. Hum Reprod 12: 1142–1151, 1997.
199. Earley CJ, Leonard BE. Androgens, estrogens and their anti-hormones:effects on body weight and food consumption. Pharmacol BiochemBehav 11: 211–214, 1979.
200. Eastwood H, Brown KM, Markovic D, Pieri LF. Variation in theESR1 and ESR2 genes and genetic susceptibility to anorexia nervosa.Mol Psychiatry 7: 86–89, 2002.
201. Eck LH, Bennett AG, Egan BM, Ray JW, Mitchell CO, Smith MA,Klesges RC. Differences in macronutrient selections in users and non-users of an oral contraceptive. Am J Clin Nutr 65: 419–424, 1997.
202. Eckel LA. Estradiol: a rhythmic, inhibitory, indirect control of meal size.Physiol Behav 82: 35–41, 2004.
203. Eckel LA. The ovarian hormone estradiol plays a crucial role in thecontrol of food intake in females. Physiol Behav 104: 517–524, 2011.
204. Eckel LA, Geary N. Endogenous cholecystokinin’s satiating actionincreases during estrus in female rats. Peptides 20: 451–456, 1999.
205. Eckel LA, Geary N. Estradiol treatment increases feeding-induced c-Fosexpression in the brains of ovariectomized rats. Am J Physiol RegulIntegr Comp Physiol 281: R738–R746, 2001.
206. Eckel LA, Houpt TA, Geary N. Estradiol treatment increases CCK-induced c-Fos expression in the brains of ovariectomized rats. Am JPhysiol Regul Integr Comp Physiol 283: R1378–R1385, 2002.
207. Eckel LA, Houpt TA, Geary N. Spontaneous meal patterns in femalerats with and without access to running wheels. Physiol Behav 70:397–405, 2000.
208. Eckel LA, Langhans W, Kahler A, Campfield LA, Smith FJ, GearyN. Chronic administration of OB protein decreases food intake byselectively reducing meal size in female rats. Am J Physiol Regul IntegrComp Physiol 275: R186–R193, 1998.
209. Eckel LA, Rivera HM, Atchley DP. The anorectic effect of fenflu-ramine is influenced by sex and stage of the estrous cycle in rats. Am JPhysiol Regul Integr Comp Physiol 288: R1486–R1491, 2005.
210. Edler C, Lipson SF, Keel PK. Ovarian hormones and binge eating inbulimia nervosa. Psychol Med 37: 131–141, 2007.
211. Ehrmann DA. Polycystic ovary syndrome. N Engl J Med 352: 1223–1236, 2005.
212. Eng R, Gold RM, Wade GN. Ovariectomy-induced obesity is notprevented by subdiaphragmatic vagotomy in rats. Physiol Behav 22:353–356, 1979.
213. Essick GK, Chopra A, Guest S, McGlone F. Lingual tactile acuity,taste perception, and the density and diameter of fungiform papillae infemale subjects. Physiol Behav 80: 289–302, 2003.
214. Faas MM, Melgert BN, de Vos P. A brief review on how pregnancy andsex hormones interfere with taste and food intake. Chemosens Percept 3:51–56, 2010.
215. Fan W, Yanase T, Nishi Y, Chiba S, Okabe T, Nomura M, Yoshi-matsu H, Kato S, Takayanagi R, Nawata H. Functional potentiation ofleptin-signal transducer and activator of transcription 3 signaling by theandrogen receptor. Endocrinology 149: 6028–6036, 2008.
216. Fan W, Yanase T, Nomura M, Okabe T, Goto K, Sato T, Kawano H,Kato S, Nawata H. Androgen receptor null male mice develop late-onsetobesity caused by decreased energy expenditure and lipolytic activity butshow normal insulin sensitivity with high adiponectin secretion. Diabetes54: 1000–1008, 2005.
217. Fantino M, Brinnel H. Body weight set-point changes during theovarian cycle: experimental study of rats using hoarding behavior.Physiol Behav 36: 991–996, 1986.
218. Farooqi IS, O’Rahilly S. Leptin: a pivotal regulator of human energyhomeostasis. Am J Clin Nutr 89: 980S–984S, 2009.
219. Farooqi IS, Yeo GS, Keogh JM, Aminian S, Jebb SA, Butler G,Cheetham T, O’Rahilly S. Dominant and recessive inheritance ofmorbid obesity associated with melanocortin 4 receptor deficiency. J ClinInvest 106: 271–279, 2000.
220. Feldman HA, Longcope C, Derby CA, Johannes CB, Araujo AB,Coviello AD, Bremner WJ, McKinlay JB. Age trends in the level ofserum testosterone and other hormones in middle-aged men: longitudinalresults from the Massachusetts male aging study. J Clin EndocrinolMetab 87: 589–598, 2002.
221. Ferin M, Tempone A, Zimmering PE, Van de Wiele RL. Effect ofantibodies to 17�-estradiol and progesterone on the estrous cycle of therat. Endocrinology 85: 1070–1078, 1969.
222. Fessler DM. No time to eat: an adaptationist account of periovulatorybehavioral changes. Q Rev Biol 78: 3–21, 2003.
223. Field AE, Colditz GA, Peterson KE. Racial/ethnic and gender differ-ences in concern with weight and in bulimic behaviors among adoles-cents. Obes Res 5: 447–454, 1997.
224. Findlay AL, Fitzsimons JT, Kucharczyk J. Dependence of spontane-ous and angiotensin-induced drinking in the rat upon the oestrous cycleand ovarian hormones. J Endocrinol 82: 215–225, 1979.
225. Fischer S, Smith GT, Cyders MA. Another look at impulsivity: ameta-analytic review comparing specific dispositions to rash action intheir relationship to bulimic symptoms. Clin Psychol Rev 28: 1413–1425,2008.
226. Flegal KM, Carroll MD, Ogden CL, Curtin LR. Prevalence and trendsin obesity among US adults, 1999–2008. JAMA 303: 235–241, 2010.
227. Fleming AS. Ovarian influences on food intake and body weight regu-lation in lactating rats. Physiol Behav 17: 969–978, 1976.
228. Foltin RW, Woolverton WL, Schuster CR. Effects of psychomotorstimulants, alone and in pairs, on milk drinking in the rat after intraperi-toneal and intragastric administration. J Pharmacol Exp Ther 226:411–418, 1983.
229. Fong AK, Kretsch MJ. Changes in dietary intake, urinary nitrogen, andurinary volume across the menstrual cycle. Am J Clin Nutr 57: 43–46,1993.
230. Frank GK, Kaye WH. Current status of functional imaging in eatingdisorders. Int J Eat Disord 45: 723–736, 2012.
231. Frank S, Laharnar N, Kullmann S, Veit R, Canova C, Hegner YL,Fritsche A, Preissl H. Processing of food pictures: influence of hunger,gender and calorie content. Brain Res 1350: 159–166, 2010.
232. Frank TC, Kim GL, Krzemien A, Van Vugt DA. Effect of menstrualcycle phase on corticolimbic brain activation by visual food cues. BrainRes 1363: 81–92, 2010.
233. Frazao R, Cravo RM, Donato J Jr, Ratra DV, Clegg DJ, ElmquistJK, Zigman JM, Williams KW, Elias CF. Shift in Kiss1 cell activityrequires estrogen receptor alpha. J Neurosci 33: 2807–2820, 2013.
234. Freedman LS, Guenther PM, Krebs-Smith SM, Dodd KW, Midt-hune D. A population’s distribution of Healthy Eating Index-2005component scores can be estimated when more than one 24-hour recall isavailable. J Nutr 140: 1529–1534, 2010.
235. Freeman EW, Sammel MD, Lin H, Gracia CR. Anti-mullerian hor-mone as a predictor of time to menopause in late reproductive agewomen. J Clin Endocrinol Metab 97: 1673–1680, 2012.
236. Freeman ME. Neuroendocrine control of the ovarian cycle of the rat. In:Knobil and Neill’s Physiology of Reproduction (3rd ed.), edited by NeillJS. Amsterdam, The Netherlands: Elsevier/Academic, 2006, p. 2327–2388.
237. Fregly MJ. Effect of chronic treatment with estrogen on the dipsogenicresponse of rats to angiotensin. Pharmacol Biochem Behav 12: 131–136,1980.
238. French SA, Epstein LH, Jeffery RW, Blundell JE, Wardle J. Eatingbehavior dimensions. Associations with energy intake and body weight.A review. Appetite 59: 541–549, 2012.
239. Friedman JM. Leptin at 14 y of age: an ongoing story. Am J Clin Nutr89: 973S–979S, 2009.
240. Frye CA. Neurosteroids’ effects and mechanisms for social, cognitive,emotional, and physical functions. Psychoneuroendocrinology 34 Suppl1: S143–S161, 2009.
241. Frye CA, Crystal S, Ward KD, Kanarek RB. Menstrual cycle anddietary restraint influence taste preferences in young women. PhysiolBehav 55: 561–567, 1994.
242. Frye CA, Paris JJ. Progesterone turnover to its 5�-reduced metabolitesin the ventral tegmental area of the midbrain is essential for initiatingsocial and affective behavior and progesterone metabolism in female rats.J Endocrinol Invest 34: e188–e199, 2011.
243. Fu LY, van den Pol AN. Kisspeptin directly excites anorexigenicproopiomelanocortin neurons but inhibits orexigenic neuropeptide Ycells by an indirect synaptic mechanism. J Neurosci 30: 10205–10219,2010.
244. Furneaux EC, Langley-Evans AJ, Langley-Evans SC. Nausea andvomiting of pregnancy: endocrine basis and contribution to pregnancyoutcome. Obstet Gynecol Surv 56: 775–782, 2001.
Review
R1254 SEX DIFFERENCES IN THE PHYSIOLOGY OF EATING
AJP-Regul Integr Comp Physiol • doi:10.1152/ajpregu.00446.2012 • www.ajpregu.org
by 10.220.32.247 on Septem
ber 18, 2016http://ajpregu.physiology.org/
Dow
nloaded from

245. Gallagher D, Visser M, Sepulveda D, Pierson RN, Harris T, Heyms-field SB. How useful is body mass index for comparison of body fatnessacross age, sex, and ethnic groups? Am J Epidemiol 143: 228–239, 1996.
246. Gambineri A, Pagotto U, Tschop M, Vicennati V, Manicardi E,Carcello A, Cacciari M, De Iasio R, Pasquali R. Anti-androgentreatment increases circulating ghrelin levels in obese women withpolycystic ovary syndrome. J Endocrinol Invest 26: 629–634, 2003.
247. Gangestad SW, Thornhill R. Human oestrus. Proc Biol Sci 275:991–1000, 2008.
248. Gangula PR, Maner WL, Micci MA, Garfield RE, Pasricha PJ.Diabetes induces sex-dependent changes in neuronal nitric oxide syn-thase dimerization and function in the rat gastric antrum. Am J PhysiolGastrointest Liver Physiol 292: G725–G733, 2007.
249. Garland HO, Atherton JC, Baylis C, Morgan MR, Milne CM.Hormone profiles for progesterone, oestradiol, prolactin, plasma reninactivity, aldosterone and corticosterone during pregnancy and pseudo-pregnancy in two strains of rat: correlation with renal studies. J Endo-crinol 113: 435–444, 1987.
250. Geary N. Counterpoint: physiologists should not distinguish “sex” and“gender”. Am J Physiol Regul Integr Comp Physiol 298: R1702–R1704,2010.
251. Geary N. Curt Richter and the female rat. Appetite 49: 376–387, 2007.252. Geary N. Endocrine controls of eating: CCK, leptin, and ghrelin. Physiol
Behav 81: 719–733, 2004.253. Geary N. The estrogenic inhibition of eating. In: Handbook of Behav-
ioral Neurobiology (2nd ed.), edited by Stricker EM, Woods SC. NewYork, NY: Kluwer Academic/Plenum Publishers, 2004, p. 307–345.
254. Geary N. Sex differences in disease anorexia. Nutrition 17: 499–507,2001.
255. Geary N, Asarian L. Cyclic estradiol treatment normalizes body weightand test meal size in ovariectomized rats. Physiol Behav 67: 141–147,1999.
256. Geary N, Asarian L. Estradiol increases glucagon’s satiating potency inovariectomized rats. Am J Physiol Regul Integr Comp Physiol 281:R1290–R1294, 2001.
257. Geary N, Asarian L, Korach KS, Pfaff DW, Ogawa S. Deficits inE2-dependent control of feeding, weight gain, and cholecystokinin sati-ation in ER-� null mice. Endocrinology 142: 4751–4757, 2001.
258. Geary N, Smith GP, Corp ES. The increased satiating potency ofCCK-8 by estradiol is not mediated by upregulation of NTS CCKreceptors. Brain Res 719: 179–186, 1996.
259. Geary N, Trace D, McEwen B, Smith GP. Cyclic estradiol replacementincreases the satiety effect of CCK-8 in ovariectomized rats. PhysiolBehav 56: 281–289, 1994.
260. Geary N, Trace D, Smith GP. Estradiol interacts with gastric orpostgastric food stimuli to decrease sucrose ingestion in ovariectomizedrats. Physiol Behav 57: 155–158, 1995.
261. Geiselman PJ, Martin JR, Vanderweele DA, Novin D. Dietary self-selection in cycling and neonatally ovariectomized rats. Appetite 2:87–101, 1981.
262. Geliebter A, Hashim SA, Gluck ME. Appetite-related gut peptides,ghrelin, PYY, and GLP-1 in obese women with and without binge eatingdisorder (BED). Physiol Behav 94: 696–699, 2008.
263. Geliebter A, Melton PM, McCray RS, Gallagher DR, Gage D,Hashim SA. Gastric capacity, gastric emptying, and test-meal intake innormal and bulimic women. Am J Clin Nutr 56: 656–661, 1992.
264. Geliebter A, Pantazatos SP, McOuatt H, Puma L, Gibson CD,Atalayer D. Sex-based fMRI differences in obese humans in response tohigh- vs. low-energy food cues. Behav Brain Res 243: 91–96, 2013.
265. Geliebter A, Yahav EK, Gluck ME, Hashim SA. Gastric capacity, testmeal intake, and appetitive hormones in binge eating disorder. PhysiolBehav 81: 735–740, 2004.
266. Gentry RT, Wade GN. Androgenic control of food intake and bodyweight in male rats. J Comp Physiol Psychol 90: 18–25, 1976.
267. Gentry RT, Wade GN. Sex differences in sensitivity of food intake,body weight, and running-wheel activity to ovarian steroids in rats. JComp Physiol Psychol 90: 747–754, 1976.
268. George JT, Seminara SB. Kisspeptin and the hypothalamic control ofreproduction: lessons from the human. Endocrinology 153: 5130–5136,2012.
269. Geracioti TD Jr., Liddle RA. Impaired cholecystokinin secretion inbulimia nervosa. N Engl J Med 319: 683–688, 1988.
270. Gill RC, Murphy PD, Hooper HR, Bowes KL, Kingma YJ. Effect ofthe menstrual cycle on gastric emptying. Digestion 36: 168–174, 1987.
271. Gillies GE, McArthur S. Estrogen actions in the brain and the basis fordifferential action in men and women: a case for sex-specific medicines.Pharmacol Rev 62: 155–198, 2010.
272. Gladis MM, Walsh BT. Premenstrual exacerbation of binge eating inbulimia. Am J Psychiatry 144: 1592–1595, 1987.
273. Glass CM, Haas SA, Reither EN. The skinny on success: body mass,gender and occupational standing across the life course. Soc Forces 88:1777–1806, 2010.
274. Gloy V, Langhans W, Hillebrand JJ, Geary N, Asarian L. Ovariec-tomy and overeating palatable, energy-dense food increase subcutaneousadipose tissue more than intra-abdominal adipose tissue in rats. Biol SexDiffer 2: 6, 2011.
275. Gloy VL, Lutz TA, Langhans W, Geary N, Hillebrand JJ. Basalplasma levels of insulin, leptin, ghrelin, and amylin do not signaladiposity in rats recovering from forced overweight. Endocrinology 151:4280–4288, 2010.
276. Gold EB, Bromberger J, Crawford S, Samuels S, Greendale GA,Harlow SD, Skurnick J. Factors associated with age at natural meno-pause in a multiethnic sample of midlife women. Am J Epidemiol 153:865–874, 2001.
277. Golden NH, Jacobson MS, Schebendach J, Solanto MV, Hertz SM,Shenker IR. Resumption of menses in anorexia nervosa. Arch PediatrAdolesc Med 151: 16–21, 1997.
278. Gong EJ, Garrel D, Calloway DH. Menstrual cycle and voluntary foodintake. Am J Clin Nutr 49: 252–258, 1989.
279. Gonzalez-Martinez D, De Mees C, Douhard Q, Szpirer C, Bakker J.Absence of gonadotropin-releasing hormone 1 and Kiss1 activation inalpha-fetoprotein knockout mice: prenatal estrogens defeminize the po-tential to show preovulatory luteinizing hormone surges. Endocrinology149: 2333–2340, 2008.
280. Goodarzi MO, Dumesic DA, Chazenbalk G, Azziz R. Polycystic ovarysyndrome: etiology, pathogenesis and diagnosis. Nat Rev Endocrinol 7:219–231, 2011.
281. Goodin SZ, Kiechler AR, Smith M, Wendt D, Strader AD. Effect ofgonadectomy on AgRP-induced weight gain in rats. Am J Physiol RegulIntegr Comp Physiol 295: R1747–R1753, 2008.
282. Goodman RL. A quantitative analysis of the physiological role ofestradiol and progesterone in the control of tonic and surge secretion ofluteinizing hormone in the rat. Endocrinology 102: 142–150, 1978.
283. Goodman RL, Lehman MN. Kisspeptin neurons from mice to men:similarities and differences. Endocrinology 153: 5105–5118, 2012.
284. Grabenhorst F, Rolls ET, Parris BA, d’Souza AA. How the brainrepresents the reward value of fat in the mouth. Cereb Cortex 20:1082–1091, 2010.
285. Grattan DR, Ladyman SR, Augustine RA. Hormonal induction ofleptin resistance during pregnancy. Physiol Behav 91: 366–374, 2007.
286. Graves NS, Hayes H, Fan L, Curtis KS. Time course of behavioral,physiological, and morphological changes after estradiol treatment ofovariectomized rats. Physiol Behav 103: 261–267, 2011.
287. Gray JM, Greenwood MR. Time course of effects of ovarian hormoneson food intake and metabolism. Am J Physiol Endocrinol Metab 243:E407–E412, 1982.
288. Gray JM, Nunez AA, Siegel LI, Wade GN. Effects of testosterone onbody weight and adipose tissue: role of aromatization. Physiol Behav 23:465–469, 1979.
289. Griffen JE, Ojeda SR. Textbook of Endocrine Physiology (6th ed.). NewYork, NY: Oxford Universtiy Press, 2012.
290. Griffin JE, Wilson JD. Disorders of the testes and the male reproductivetract In: Williams Textbook of Endocrinology (10th ed.), edited by LarsenPR, Kronenberg, HM, Melmed S, Polonsky KS. Philadelphia, PA:Elsevier/Saunders, 2003, p. 709–769.
291. Grill HJ. Leptin and the systems neuroscience of meal size control.Front Neuroendocrinol 31: 61–78, 2010.
292. Grilo CM, Masheb RM, Brody M, Burke-Martindale CH, Roth-schild BS. Binge eating and self-esteem predict body image dissatisfac-tion among obese men and women seeking bariatric surgery. Int J EatDisord 37: 347–351, 2005.
293. Gropp E, Shanabrough M, Borok E, Xu AW, Janoschek R, Buch T,Plum L, Balthasar N, Hampel B, Waisman A, Barsh GS, HorvathTL, Bruning JC. Agouti-related peptide-expressing neurons are man-datory for feeding. Nat Neurosci 8: 1289–1291, 2005.
294. Grueso E, Rocha M, Puerta M. Plasma and cerebrospinal fluid leptinlevels are maintained despite enhanced food intake in progesterone-treated rats. Eur J Endocrinol 144: 659–665, 2001.
Review
R1255SEX DIFFERENCES IN THE PHYSIOLOGY OF EATING
AJP-Regul Integr Comp Physiol • doi:10.1152/ajpregu.00446.2012 • www.ajpregu.org
by 10.220.32.247 on Septem
ber 18, 2016http://ajpregu.physiology.org/
Dow
nloaded from

295. Gueguen N. The receptivity of women to courtship solicitation acrossthe menstrual cycle: a field experiment. Biol Psychol 80: 321–324, 2009.
296. Guertin TL. Eating behavior of bulimics, self-identified binge eaters,and non-eating-disordered individuals: what differentiates these popula-tions? Clin Psychol Rev 19: 1–23, 1999.
297. Haarbo J, Marslew U, Gotfredsen A, Christiansen C. Postmenopausalhormone replacement therapy prevents central distribution of body fatafter menopause. Metabolism 40: 1323–1326, 1991.
298. Haas E, Bhattacharya I, Brailoiu E, Damjanovic M, Brailoiu GC,Gao X, Mueller-Guerre L, Marjon NA, Gut A, Minotti R, MeyerMR, Amann K, Ammann E, Perez-Dominguez A, Genoni M, CleggDJ, Dun NJ, Resta TC, Prossnitz ER, Barton M. Regulatory role of Gprotein-coupled estrogen receptor for vascular function and obesity. CircRes 104: 288–291, 2009.
299. Haase L, Green E, Murphy C. Males and females show differentialbrain activation to taste when hungry and sated in gustatory and rewardareas. Appetite 57: 421–434, 2011.
300. Habegger KM, Heppner KM, Geary N, Bartness TJ, DiMarchi R,Tschop MH. The metabolic actions of glucagon revisited. Nat RevEndocrinol 6: 689–697, 2010.
301. Halaas JL, Boozer C, Blair-West J, Fidahusein N, Denton DA,Friedman JM. Physiological response to long-term peripheral andcentral leptin infusion in lean and obese mice. Proc Natl Acad Sci USA94: 8878–8883, 1997.
302. Halford JC, Harrold JA. 5-HT(2C) receptor agonists and the control ofappetite. Handb Exp Pharmacol 209: 349–356, 2012.
303. Hall JE. Neuroendocrine changes with reproductive aging in women.Semin Reprod Med 25: 344–351, 2007.
304. Hallschmid M, Benedict C, Schultes B, Born J, Kern W. Obese menrespond to cognitive but not to catabolic brain insulin signaling. Int JObes (Lond) 32: 275–282, 2008.
305. Hallschmid M, Benedict C, Schultes B, Fehm HL, Born J, Kern W.Intranasal insulin reduces body fat in men but not in women. Diabetes53: 3024–3029, 2004.
306. Hammond GL. Diverse roles for sex hormone-binding globulin inreproduction. Biol Reprod 85: 431–441, 2011.
307. Hancock SD, Grant VL. Sexually dimorphic effects of postnatal treat-ment on the development of activity-based anorexia in adolescent andadult rats. Dev Psychobiol 51: 679–695, 2009.
308. Handelsman DJ. Aging in the hypothalamic-pituitary-testcular axis In:Knobil and Neill’s Physiology of Reproduction (3rd ed.), edited by NeillJD. St. Louis, MO: Elsevier, 2006, p. 2697–2728.
309. Harris HA, Katzenellenbogen JA, Katzenellenbogen BS. Character-ization of the biological roles of the estrogen receptors, ER� and ER�, inestrogen target tissues in vivo through the use of an ER�-selective ligand.Endocrinology 143: 4172–4177, 2002.
310. Harris RB, Bartness TJ, Grill HJ. Leptin responsiveness in chronicallydecerebrate rats. Endocrinology 148: 4623–4633, 2007.
311. Harris RB, Kasser TR, Martin RJ. Dynamics of recovery of bodycomposition after overfeeding, food restriction or starvation of maturefemale rats. J Nutr 116: 2536–2546, 1986.
312. Hartmann B, Kirchengast S, Albrecht A, Laml T, Bikas D, Huber J.Effects of hormone replacement therapy on growth hormone secretionpatterns in correlation to somatometric parameters in healthy postmeno-pausal women. Maturitas 22: 239–246, 1995.
313. Hashimoto I, Henricks DM, Anderson LL, Melampy RM. Progester-one and pregn-4-en-20 alpha-ol-3-one in ovarian venous blood duringvarious reproductive states in the rat. Endocrinology 82: 333–341, 1968.
314. Hashimoto I, Wiest WG. Correlation of the secretion of ovarian steroidswith function of a single generation of corpora lutea in the immature rat.Endocrinology 84: 873–885, 1969.
315. Hayes DJ, Greenshaw AJ. 5-HT receptors and reward-related behav-iour: a review. Neurosci Biobehav Rev 35: 1419–1449, 2011.
316. Hayes JE, Duffy VB. Oral sensory phenotype identifies level of sugarand fat required for maximal liking. Physiol Behav 95: 77–87, 2008.
317. Hayes JE, Duffy VB. Revisiting sugar-fat mixtures: sweetness andcreaminess vary with phenotypic markers of oral sensation. Chem Senses32: 225–236, 2007.
318. Hayes MR. Neuronal and intracellular signaling pathways mediatingGLP-1 energy balance and glycemic effects. Physiol Behav 106: 413–416, 2012.
319. Hayes MR, Bradley L, Grill HJ. Endogenous hindbrain glucagon-likepeptide-1 receptor activation contributes to the control of food intake by
mediating gastric satiation signaling. Endocrinology 150: 2654–2659,2009.
320. Healy K, Conroy RM, Walsh N. The prevalence of binge-eating andbulimia in 1063 college students. J Psychiatr Res 19: 161–166, 1985.
321. Heape W. The “sexual season” of mammals and the relation of the“pre-oestrum” to menstruation. Q J Microscopic Sci 44: 1–70, 1900.
322. Heatherton TF, Herman CP, Polivy J, King GA, McGree ST. The(mis)measurement of restraint: an analysis of conceptual and psychomet-ric issues. J Abnorm Psychol 97: 19–28, 1988.
323. Heil SH. Activational and organizational actions of gonadal hormonesand the sex-specific effects of prolactin on food intake by rats. DevPsychobiol 35: 61–67, 1999.
324. Heine PA, Taylor JA, Iwamoto GA, Lubahn DB, Cooke PS. Increasedadipose tissue in male and female estrogen receptor-alpha knockoutmice. Proc Natl Acad Sci USA 97: 12729–12734, 2000.
325. Heisler LK, Kanarek RB, Homoleski B. Reduction of fat and proteinintakes but not carbohydrate intake following acute and chronic fluox-etine in female rats. Pharmacol Biochem Behav 63: 377–385, 1999.
326. Heldring N, Pike A, Andersson S, Matthews J, Cheng G, Hartman J,Tujague M, Strom A, Treuter E, Warner M, Gustafsson JA. Estrogenreceptors: how do they signal and what are their targets. Physiol Rev 87:905–931, 2007.
327. Henderson KD, Bernstein L, Henderson B, Kolonel L, Pike MC.Predictors of the timing of natural menopause in the Multiethnic CohortStudy. Am J Epidemiol 167: 1287–1294, 2008.
328. Herman CP, Polivy J. Anxiety, restraint, and eating behavior. J AbnormPsychol 84: 66–72, 1975.
329. Hilbert A, de Zwaan M, Braehler E. How frequent are eating distur-bances in the population? Norms of the eating disorder examination-questionnaire. PLoS One 7: e29125, 2012.
330. Hill JW, Elmquist JK, Elias CF. Hypothalamic pathways linkingenergy balance and reproduction. Am J Physiol Endocrinol Metab 294:E827–E832, 2008.
331. Hillebrand JJ, Geary N. Do leptin and insulin signal adiposity? ForumNutr 63: 111–122, 2010.
332. Himmerich H, Schonknecht P, Heitmann S, Sheldrick AJ. Laboratoryparameters and appetite regulators in patients with anorexia nervosa. JPsychiatr Pract 16: 82–92, 2010.
333. Hintiryan H, Foster NN, Chambers KC. Dissociating the conditioningand the anorectic effects of estradiol in female rats. Behav Neurosci 123:1226–1237, 2009.
334. Hirschberg AL. Sex hormones, appetite and eating behaviour in women.Maturitas 71: 248–256, 2012.
335. Hirschberg AL, Naessen S, Stridsberg M, Bystrom B, Holtet J.Impaired cholecystokinin secretion and disturbed appetite regulation inwomen with polycystic ovary syndrome. Gynecol Endocrinol 19: 79–87,2004.
336. Ho SC, Wu S, Chan SG, Sham A. Menopausal transition and changesof body composition: a prospective study in Chinese perimenopausalwomen. Int J Obes (Lond) 34: 1265–1274, 2010.
337. Hook EB. Dietary cravings and aversions during pregnancy. Am J ClinNutr 31: 1355–1362, 1978.
338. Hormes JM, Rozin P. Perimenstrual chocolate craving. What happensafter menopause? Appetite 53: 256–259, 2009.
339. Horner KM, Byrne NM, Cleghorn GJ, Naslund E, King NA. Theeffects of weight loss strategies on gastric emptying and appetite control.Obes Rev 12: 935–951, 2011.
340. Horowitz M, Maddern GJ, Chatterton BE, Collins PJ, Petrucco OM,Seamark R, Shearman DJ. The normal menstrual cycle has no effect ongastric emptying. Br J Obstet Gynaecol 92: 743–746, 1985.
341. Houpt K. Domestic Animal Behavior for Veterinarians and AnimalScientists. Ames, IA: Blackwell, 2004.
342. Howards PP, Schisterman EF, Wactawski-Wende J, Reschke JE,Frazer AA, Hovey KM. Timing clinic visits to phases of the menstrualcycle by using a fertility monitor: the BioCycle Study. Am J Epidemiol169: 105–112, 2009.
343. Hrupka BJ, Smith GP, Geary N. Hypothalamic implants of diluteestradiol fail to reduce feeding in ovariectomized rats. Physiol Behav 77:233–241, 2002.
344. Hrupka BJ, Smith GP, Geary N. Ovariectomy and estradiol affectpostingestive controls of sucrose licking. Physiol Behav 61: 243–247,1997.
345. Huang YS, Doi R, Chowdhury P, Pasley JN, Nishikawa M, HuangTJ, Rayford PL. Effect of cholecystokinin on food intake at different
Review
R1256 SEX DIFFERENCES IN THE PHYSIOLOGY OF EATING
AJP-Regul Integr Comp Physiol • doi:10.1152/ajpregu.00446.2012 • www.ajpregu.org
by 10.220.32.247 on Septem
ber 18, 2016http://ajpregu.physiology.org/
Dow
nloaded from

stages of the estrous cycle in female rats. J Assoc Acad Minor Phys 4:56–58, 1993.
346. Hudson JI, Hiripi E, Pope HG Jr, Kessler RC. The prevalence andcorrelates of eating disorders in the National Comorbidity Survey Rep-lication. Biol Psychiatry 61: 348–358, 2007.
347. Hung AJ, Stanbury MG, Shanabrough M, Horvath TL, Garcia-Segura LM, Naftolin F. Estrogen, synaptic plasticity and hypothalamicreproductive aging. Exp Gerontol 38: 53–59, 2003.
348. Huszar D, Lynch CA, Fairchild-Huntress V, Dunmore JH, Fang Q,Berkemeier LR, Gu W, Kesterson RA, Boston BA, Cone RD, SmithFJ, Campfield LA, Burn P, Lee F. Targeted disruption of the melano-cortin-4 receptor results in obesity in mice. Cell 88: 131–141, 1997.
349. Hutson WR, Roehrkasse RL, Wald A. Influence of gender and meno-pause on gastric emptying and motility. Gastroenterology 96: 11–17,1989.
350. Isken F, Pfeiffer AF, Nogueiras R, Osterhoff MA, Ristow M, ThorensB, Tschop MH, Weickert MO. Deficiency of glucose-dependent insuli-notropic polypeptide receptor prevents ovariectomy-induced obesity inmice. Am J Physiol Endocrinol Metab 295: E350–E355, 2008.
351. Jacobi F, Wittchen HU, Holting C, Hofler M, Pfister H, Muller N,Lieb R. Prevalence, co-morbidity and correlates of mental disorders inthe general population: results from the German Health Interview andExamination Survey (GHS). Psychol Med 34: 597–611, 2004.
352. Johnson MA, Tsutsui K, Fraley GS. Rat RFamide-related peptide-3stimulates GH secretion, inhibits LH secretion, and has variable effectson sex behavior in the adult male rat. Horm Behav 51: 171–180, 2007.
353. Johnson WG, Corrigan SA, Lemmon CR, Bergeron KB, Crusco AH.Energy regulation over the menstrual cycle. Physiol Behav 56: 523–527,1994.
354. Jonassaint CR, Szatkiewicz JP, Bulik CM, Thornton LM, Bloss C,Berrettini WH, Kaye WH, Bergen AW, Magistretti P, Strober M,Keel PK, Brandt H, Crawford S, Crow S, Fichter MM, Goldman D,Halmi KA, Johnson C, Kaplan AS, Klump KL, La Via M, MitchellJE, Rotondo A, Treasure J, Woodside DB. Absence of associationbetween specific common variants of the obesity-related FTO gene andpsychological and behavioral eating disorder phenotypes. Am J MedGenet B Neuropsychiatr Genet 156B: 454–461, 2011.
355. Jones AB, Curtis KS. Differential effects of estradiol on drinking byovariectomized rats in response to hypertonic NaCl or isoproterenol:Implications for hyper- vs. hypo-osmotic stimuli for water intake. PhysiolBehav 98: 421–426, 2009.
356. Jones ME, Thorburn AW, Britt KL, Hewitt KN, Wreford NG,Proietto J, Oz OK, Leury BJ, Robertson KM, Yao S, Simpson ER.Aromatase-deficient (ArKO) mice have a phenotype of increased adipos-ity. Proc Natl Acad Sci USA 97: 12735–12740, 2000.
357. Jungheim ES, Moley KH. Current knowledge of obesity’s effects in thepre- and periconceptional periods and avenues for future research. Am JObstet Gynecol 203: 525–530, 2010.
358. Kanarek RB, Marks-Kaufman R, Lipeles BJ. Increased carbohydrateintake as a function of insulin administration in rats. Physiol Behav 25:779–782, 1980.
359. Kanarek RB, Ryu M, Przypek J. Preferences for foods with varyinglevels of salt and fat differ as a function of dietary restraint and exercisebut not menstrual cycle. Physiol Behav 57: 821–826, 1995.
360. Kaplan JM, Moran TH. Gastrointestinal signaling in the control of foodintake. In: Handbook of Behavioral Neurobiology (2nd ed.), edited byStricker E, Woods, SC. New York: Kluwer Academic/Plenum Publish-ers, 2004, p. 275–306.
361. Karastergiou K, Smith SR, Greenberg AS, Fried SK. Sex differencesin human adipose tissues—the biology of pear shape. Biol Sex Differ 3:13, 2012.
362. Kas MJ, Adan RA. Animal models of eating disorder traits. Curr TopBehav Neurosci 6: 209–227, 2011.
363. Kas MJ, Kaye WH, Foulds Mathes W, Bulik CM. Interspeciesgenetics of eating disorder traits. Am J Med Genet B NeuropsychiatrGenet 150B: 318–327, 2009.
364. Kauffman AS, Rissman EF. Neuroendocrine control of mating-inducedovulation. In: Knobil and Neill’s Physiology of Reproduction (3rd ed.),edited by Neill JD. St. Louis, MO: Elsevier/Academic, 2006, p. 2283–2326.
365. Kaye W. Neurobiology of anorexia and bulimia nervosa. Physiol Behav94: 121–135, 2008.
366. Keating C, Tilbrook A, Kulkarni J. Oestrogen: an overlooked mediatorin the neuropsychopharmacology of treatment response? Int J Neuropsy-chopharmacol 14: 553–566, 2011.
367. Keel PK, Wolfe BE, Liddle RA, De Young KP, Jimerson DC. Clinicalfeatures and physiological response to a test meal in purging disorder andbulimia nervosa. Arch Gen Psychiatry 64: 1058–1066, 2007.
368. Keen-Rhinehart E, Desai M, Ross MG. Central insulin sensitivity inmale and female juvenile rats. Horm Behav 56: 275–280, 2009.
369. Kellokoski E, Poykko SM, Karjalainen AH, Ukkola O, Heikkinen J,Kesaniemi YA, Horkko S. Estrogen replacement therapy increasesplasma ghrelin levels. J Clin Endocrinol Metab 90: 2954–2963, 2005.
370. Kelly MJ, Moss RL, Dudley CA. The effects of microelectrophoreti-cally applied estrogen, cortisol and acetylcholine on medial preoptic-septal unit activity throughout the estrous cycle of the female rat. ExpBrain Res 30: 53–64, 1977.
371. Kelly MJ, Moss RL, Dudley CA, Fawcett CP. The specificity of theresponse of preoptic-septal area neurons to estrogen: 17�-estradiol versus17�-estradiol and the response of extrahypothalamic neurons. Exp BrainRes 30: 43–52, 1977.
372. Kemnitz JW, Gibber JR, Lindsay KA, Eisele SG. Effects of ovarianhormones on eating behaviors, body weight, and glucoregulation inrhesus monkeys. Horm Behav 23: 235–250, 1989.
373. Kemnitz JW, Gibber JR, Eisele SG, Lindsay KA. Relationship ofreproductive condition to food intake and sucrose consumption of femalerhesus monkeys. In: Current Perspectives in Primate Social Dynamics,edited by Taub DM, King FA. New York: Van Nostrand Reinhold, 1986,p. 274–286.
374. Kemnitz JW, Goy RW, Keesey RE. Effects of gonadectomy on hypo-thalamic obesity in male and female rats. Int J Obes (Lond) 1: 259–270,1977.
375. Kemnitz JW, Sladky KK, Flitsch TJ, Pomerantz SM, Goy RW.Androgenic influences on body size and composition of adult rhesusmonkeys. Am J Physiol Endocrinol Metab 255: E857–E864, 1988.
376. Kenney NJ, Mook DG. Effects of ovariectomy on meal pattern in thealbino rat. J Comp Physiol Psychol 87: 302–309, 1974.
377. Khan AR, Kauffman AS. The role of kisspeptin and RFamide-relatedpeptide-3 neurones in the circadian-timed preovulatory luteinising hor-mone surge. J Neuroendocrinol 24: 131–143, 2012.
378. Killgore WD, Yurgelun-Todd DA Sex differences in cerebral responsesto images of high versus low-calorie food. Neuroreport 21: 354–358,2010.
379. King BM, Rollins BL, Grundmann SJ, Olivier LG. Excessive weightgains in female rats with transections of the stria terminalis. PhysiolBehav 78: 563–568, 2003.
380. King BM, Rollins BL, Stines SG, Cassis SA, McGuire HB, LagardeML. Sex differences in body weight gains following amygdaloid lesionsin rats. Am J Physiol Regul Integr Comp Physiol 277: R975–R980, 1999.
381. King JM. Effects of lesions of the amygdala, preoptic area, and hypo-thalamus on estradiol-induced activity in the female rat. J Comp PhysiolPsychol 93: 360–367, 1979.
382. Kinzl JF, Traweger C, Trefalt E, Mangweth B, Biebl W. Binge eatingdisorder in females: a population-based investigation. Int J Eat Disord25: 287–292, 1999.
383. Kirchner H, Heppner KM, Tschop MH. The role of ghrelin in thecontrol of energy balance. Handb Exp Pharmacol: 161–184, 2012.
384. Kisley LR, Sakai RR, Ma LY, Fluharty SJ. Ovarian steroid regulationof angiotensin II-induced water intake in the rat. Am J Physiol RegulIntegr Comp Physiol 276: R90–R96, 1999.
385. Kissileff HR, Carretta JC, Geliebter A, Pi-Sunyer FX. Cholecysto-kinin and stomach distension combine to reduce food intake in humans.Am J Physiol Regul Integr Comp Physiol 285: R992–R998, 2003.
386. Klingerman CM, Krishnamoorthy K, Patel K, Spiro AB, Struby C,Patel A, Schneider JE. Energetic challenges unmask the role of ovarianhormones in orchestrating ingestive and sex behaviors. Horm Behav 58:563–574, 2010.
387. Klump KL, Bulik CM, Kaye WH, Treasure J, Tyson E. Academy foreating disorders position paper: eating disorders are serious mentalillnesses. Int J Eat Disord 42: 97–103, 2009.
388. Klump KL, Culbert KM, Slane JD, Burt SA, Sisk CL, Nigg JT. Theeffects of puberty on genetic risk for disordered eating: evidence for a sexdifference. Psychol Med 42: 627–637, 2012.
389. Klump KL, Gobrogge KL, Perkins PS, Thorne D, Sisk CL, Breed-love SM. Preliminary evidence that gonadal hormones organize andactivate disordered eating. Psychol Med 36: 539–546, 2006.
Review
R1257SEX DIFFERENCES IN THE PHYSIOLOGY OF EATING
AJP-Regul Integr Comp Physiol • doi:10.1152/ajpregu.00446.2012 • www.ajpregu.org
by 10.220.32.247 on Septem
ber 18, 2016http://ajpregu.physiology.org/
Dow
nloaded from

390. Klump KL, Keel PK, Burt SA, Racine SE, Neale MC, Sisk CL, BokerS. Ovarian hormones and emotional eating associations across the men-strual cycle: an examination of the potential moderating effects of bodymass index and dietary restraint. Int J Eat Disord 46: 256–263, 2013.
391. Klump KL, Keel PK, Culbert KM, Edler C. Ovarian hormones andbinge eating: exploring associations in community samples. Psychol Med38: 1749–1757, 2008.
392. Klump KL, Keel PK, Racine SE, Burt SA, Neale M, Sisk CL, BokerS, Hu JY. The interactive effects of estrogen and progesterone onchanges in emotional eating across the menstrual cycle. J AbnormPsychol 122: 131–137, 2013.
393. Klump KL, Keel PK, Sisk C, Burt SA. Preliminary evidence thatestradiol moderates genetic influences on disordered eating attitudes andbehaviors during puberty. Psychol Med 40: 1745–1753, 2010.
394. Klump KL, McGue M, Iacono WG. Age differences in genetic andenvironmental influences on eating attitudes and behaviors in preadoles-cent and adolescent female twins. J Abnorm Psychol 109: 239–251,2000.
395. Klump KL, Miller KB, Keel PK, McGue M, Iacono WG. Genetic andenvironmental influences on anorexia nervosa syndromes in a popula-tion-based twin sample. Psychol Med 31: 737–740, 2001.
396. Klump KL, Racine S, Hildebrandt B, Sisk CL. Sex differences inbinge eating patterns in male and female adult rats. Int J Eat Disord Inpress.
397. Klump KL, Suisman JL, Culbert KM, Kashy DA, Keel PK, Sisk CL.The effects of ovariectomy on binge eating proneness in adult femalerats. Horm Behav 59: 585–593, 2011.
398. Klump KL, Suisman JL, Culbert KM, Kashy DA, Sisk CL. Bingeeating proneness emerges during puberty in female rats: a longitudinalstudy. J Abnorm Psychol 120: 948–955, 2011.
399. Knight LC, Parkman HP, Brown KL, Miller MA, Trate DM, Mau-rer AH, Fisher RS. Delayed gastric emptying and decreased antralcontractility in normal premenopausal women compared with men. Am JGastroenterol 92: 968–975, 1997.
400. Knight WD, Seth R, Boron J, Overton JM. Short-term physiologicalhyperleptinemia decreases arterial blood pressure. Regul Pept 154: 60–68, 2009.
401. Knott CD. Changes in orangutan caloric intake, energy homeostasis andketones in response to fluctuating fruit availability. Intl. J. Primatol. 19:1061–1079, 1999.
402. Kochakian CD, Webster JA. Effect of testosterone propionate on theappetite, body weight and composition of the normal rat. Endocrinology63: 737–742, 1958.
403. Koscik T, Bechara A, Tranel D. Sex-related functional asymmetry inthe limbic brain. Neuropsychopharmacology 35: 340–341, 2010.
404. Kow LM, Pfaff DW. The membrane actions of estrogens can potentiatetheir lordosis behavior—facilitating genomic actions. Proc Natl Acad SciUSA 101: 12354–12357, 2004.
405. Kraly FS, Jerome C, Smith GP. Specific postoperative syndromes aftertotal and selective vagotomies in the rat. Appetite 7: 1–17, 1986.
406. Krause EG, Curtis KS, Davis LM, Stowe JR, Contreras RJ. Estrogeninfluences stimulated water intake by ovariectomized female rats. PhysiolBehav 79: 267–274, 2003.
407. Kriegsfeld LJ, Gibson EM, Williams WP, 3rd Zhao S, Mason AO,Bentley GE, Tsutsui K. The roles of RFamide-related peptide-3 inmammalian reproductive function and behaviour. J Neuroendocrinol 22:692–700, 2010.
408. Kriegsfeld LJ, Mei DF, Bentley GE, Ubuka T, Mason AO, Inoue K,Ukena K, Tsutsui K, Silver R. Identification and characterization of agonadotropin-inhibitory system in the brains of mammals. Proc NatlAcad Sci USA 103: 2410–2415, 2006.
409. Kringelbach ML. The human orbitofrontal cortex: linking reward tohedonic experience. Nat Rev Neurosci 6: 691–702, 2005.
410. Kringelbach ML, Stein A. Cortical mechanisms of human eating.Forum Nutr 63: 164–175, 2010.
411. Labrie F, Luu-The V, Labrie C, Belanger A, Simard J, Lin SX,Pelletier G. Endocrine and intracrine sources of androgens in women:inhibition of breast cancer and other roles of androgens and theirprecursor dehydroepiandrosterone. Endocr Rev 24: 152–182, 2003.
412. Ladyman SR, Augustine RA, Grattan DR. Hormone interactionsregulating energy balance during pregnancy. J Neuroendocrinol 22:805–817, 2010.
413. Ladyman SR, Tups A, Augustine RA, Swahn-Azavedo A, Kokay IC,Grattan DR. Loss of hypothalamic response to leptin during pregnancy
associated with development of melanocortin resistance. J Neuroendo-crinol 21: 449–456, 2009.
414. Lam DD, Garfield AS, Marston OJ, Shaw J, Heisler LK. Brainserotonin system in the coordination of food intake and body weight.Pharmacol Biochem Behav 97: 84–91, 2010.
415. Landt M, Gingerich RL, Havel PJ, Mueller WM, Schoner B, HaleJE, Heiman ML. Radioimmunoassay of rat leptin: sexual dimorphismreversed from humans. Clin Chem 44: 565–570, 1998.
416. Langhans W, Geary N. Overview of the physiological control of eating.Forum Nutr 63: 9–53, 2010.
417. Larner JM, Hochberg RB. The clearance and metabolism of estradioland estradiol-17-esters in the rat. Endocrinology 117: 1209–1214, 1985.
418. Larsson H, Elmstahl S, Berglund G, Ahren B. Evidence for leptinregulation of food intake in humans. J Clin Endocrinol Metab 83:4382–4385, 1998.
419. Lee CC, Kasa-Vubu JZ, Supiano MA. Differential effects of raloxifeneand estrogen on insulin sensitivity in postmenopausal women. J AmGeriatr Soc 51: 683–688, 2003.
420. Lee JS, Ettinger B, Stanczyk FZ, Vittinghoff E, Hanes V, Cauley JA,Chandler W, Settlage J, Beattie MS, Folkerd E, Dowsett M, GradyD, Cummings SR. Comparison of methods to measure low serumestradiol levels in postmenopausal women. J Clin Endocrinol Metab 91:3791–3797, 2006.
421. Lee TM, McClintock MK. Female rats in a laboratory display seasonalvariation in fecundity. J Reprod Fertil 77: 51–59, 1986.
422. Leibel RL. Molecular physiology of weight regulation in mice andhumans. Int J Obes (Lond) 32 Suppl 7: S98–S108, 2008.
423. Leibowitz SF, Akabayashi A, Alexander JT, Wang J. Gonadal ste-roids and hypothalamic galanin and neuropeptide Y: role in eatingbehavior and body weight control in female rats. Endocrinology 139:1771–1780, 1998.
424. Leidy LE. Timing of menopause in relation to body size and weightchange. Hum Biol 68: 967–982, 1996.
425. Lenz KM, Nugent BM, McCarthy MM. Sexual differentiation of therodent brain: dogma and beyond. Front Neurosci 6: 26, 2012.
426. Lester NA, Keel PK, Lipson SF. Symptom fluctuation in bulimianervosa: relation to menstrual-cycle phase and cortisol levels. PsycholMed 33: 51–60, 2003.
427. Levin BE, Magnan C, Dunn-Meynell A, Le Foll C. Metabolic sensingand the brain: who, what, where, and how? Endocrinology 152: 2552–2557, 2011.
428. Levin ER. Minireview: Extranuclear steroid receptors: roles in modula-tion of cell functions. Mol Endocrinol 25: 377–384, 2011.
429. Ley CJ, Lees B, Stevenson JC. Sex- and menopause-associated changesin body-fat distribution. Am J Clin Nutr 55: 950–954, 1992.
430. Li BY, Qiao GF, Feng B, Zhao RB, Lu YJ, Schild JH. Electrophys-iological and neuroanatomical evidence of sexual dimorphism in aorticbaroreceptor and vagal afferents in rat. Am J Physiol Regul Integr CompPhysiol 295: R1301–R1310, 2008.
431. Li ET, Tsang LB, Lui SS. Menstrual cycle and voluntary food intake inyoung Chinese women. Appetite 33: 109–118, 1999.
432. Liang YQ, Akishita M, Kim S, Ako J, Hashimoto M, Iijima K, OhikeY, Watanabe T, Sudoh N, Toba K, Yoshizumi M, Ouchi Y. Estrogenreceptor � is involved in the anorectic action of estrogen. Int J Obes RelatMetab Disord 26: 1103–1109, 2002.
433. Lightfoot JT. Sex hormones’ regulation of rodent physical activity: areview. Int J Biol Sci 4: 126–132, 2008.
434. Lim SS, Norman RJ, Clifton PM, Noakes M. Hyperandrogenemia,psychological distress, and food cravings in young women. PhysiolBehav 98: 276–280, 2009.
435. Linden A, Uvnas-Moberg K, Forsberg G, Bednar I, Sodersten P.Involvement of cholecystokinin in food intake: III. oestradiol potentiatesthe inhibitory effect of cholecystokinin octapeptide on food intake inovariectomized rats. J Neuroendocrinol 2: 797–801, 1990.
436. Lissner L, Stevens J, Levitsky DA, Rasmussen KM, Strupp BJ.Variation in energy intake during the menstrual cycle: implications forfood-intake research. Am J Clin Nutr 48: 956–962, 1988.
437. Lloyd-Jones D, Adams RJ, Brown TM, Carnethon M, Dai S, DeSimone G, Ferguson TB, Ford E, Furie K, Gillespie C, Go A,Greenlund K, Haase N, Hailpern S, Ho PM, Howard V, Kissela B,Kittner S, Lackland D, Lisabeth L, Marelli A, McDermott MM,Meigs J, Mozaffarian D, Mussolino M, Nichol G, Roger VL, Rosa-mond W, Sacco R, Sorlie P, Thom T, Wasserthiel-Smoller S, WongND, Wylie-Rosett J. Heart disease and stroke statistics—2010 update: a
Review
R1258 SEX DIFFERENCES IN THE PHYSIOLOGY OF EATING
AJP-Regul Integr Comp Physiol • doi:10.1152/ajpregu.00446.2012 • www.ajpregu.org
by 10.220.32.247 on Septem
ber 18, 2016http://ajpregu.physiology.org/
Dow
nloaded from

report from the American Heart Association. Circulation 121: e46–e215,2010.
438. Lo CM, Zhang DM, Pearson K, Ma L, Sun W, Sakai RR, DavidsonWS, Liu M, Raybould HE, Woods SC, Tso P. Interaction of apolipo-protein AIV with cholecystokinin on the control of food intake. Am JPhysiol Regul Integr Comp Physiol 293: R1490–R1494, 2007.
439. Lomniczi A, Loche A, Castellano JM, Ronnekleiv OK, Bosch M,Kaidar G, Knoll JG, Wright H, Pfeifer GP, Ojeda SR. Epigeneticcontrol of female puberty. Nat Neurosci 16: 281–289, 2013.
440. Long JA, Evans ME. The oestrous cycle of the rat and its associatedphenomena. Mem Univ Calif 6: 1–103, 1922.
441. Loucks AB. The response of luteinizing hormone pulsatility to 5 days oflow-energy availability disappears by 14 years of gynecological age. JClin Endocrinol Metab 91: 3158–3164, 2006.
442. Loucks AB, Redman LM. The effect of stress on menstrual function.Trends Endocrinol Metab 15: 466–471, 2004.
443. Lovejoy JC, Champagne CM, de Jonge L, Xie H, Smith SR. Increasedvisceral fat and decreased energy expenditure during the menopausaltransition. Int J Obes (Lond) 32: 949–958, 2008.
444. Lu JK, LaPolt PS, Nass TE, Matt DW, Judd HL. Relation ofcirculating estradiol and progesterone to gonadotropin secretion andestrous cyclicity in aging female rats. Endocrinology 116: 1953–1959,1985.
445. Luquet S, Perez FA, Hnasko TS, Palmiter RD. NPY/AgRP neuronsare essential for feeding in adult mice but can be ablated in neonates.Science 310: 683–685, 2005.
446. Lutz TA. Effects of amylin on eating and adiposity. Handb Exp Phar-macol: 231–250, 2012.
447. Lutz TA. The role of amylin in the control of energy homeostasis. Am JPhysiol Regul Integr Comp Physiol 298: R1475–R1484, 2010.
448. Luu-The V, Labrie F. The intracrine sex steroid biosynthesis pathways.Prog Brain Res 181: 177–192, 2010.
449. Lydecker JA, Pisetsky EM, Mitchell KS, Thornton LM, Kendler KS,Reichborn-Kjennerud T, Lichtenstein P, Bulik CM, Mazzeo SE.Association between co-twin sex and eating disorders in opposite sextwin pairs: Evaluations in North American, Norwegian, and Swedishsamples. J Psychosom Res 72: 73–77, 2012.
450. Lyons PM, Truswell AS, Mira M, Vizzard J, Abraham SF. Reductionof food intake in the ovulatory phase of the menstrual cycle. Am J ClinNutr 49: 1164–1168, 1989.
451. Macdiarmid JI, Vail A, Cade JE, Blundell JE. The sugar-fat relation-ship revisited: differences in consumption between men and women ofvarying BMI. Int J Obes Relat Metab Disord 22: 1053–1061, 1998.
452. Madrid JA, Lopez-Bote C, Martin E. Effect of neonatal androgeniza-tion on the circadian rhythm of feeding behavior in rats. Physiol Behav53: 329–335, 1993.
453. Maletínská L, Matyšková R, Maixnerová J, Sykora D, Pychová M,Spolcová A, Blechová M, Drápalová J, Lacinová Z, Haluzik M,Zelezná B. The peptidic GHS-R antagonist [D-Lys(3)]GHRP-6 markedlyimproves adiposity and related metabolic abnormalities in a mouse modelof postmenopausal obesity. Mol Cell Endocrinol 343: 55–62, 2011.
454. Malpaux B. Seasonal reguation of reproduction in mammals. In: Knobiland Neill’s Physiology of Reproduction (3rd ed.), edited by Neill JD. St.Louis, MO: Elsevier/Academic, 2006, p. 2231–2281.
455. Mangiaracina M, Wolfe A, Azzara A, Schwartz GJ, Walsh BT,Geary N. The satiating potency of endogenous CCK increases afterpuberty in rats (Abstract). Appetite 42: 382, 2004.
456. Manning JM, Bronson FH. Suppression of puberty in rats by exercise:effects on hormone levels and reversal with GnRH infusion. Am JPhysiol Regul Integr Comp Physiol 260: R717–R723, 1991.
457. Marco A, Schroeder M, Weller A. Microstructural pattern of palatablefood intake from weaning to adulthood in male and female OLETF rats.Behav Neurosci 123: 1251–1260, 2009.
458. Martini MC, Lampe JW, Slavin JL, Kurzer MS. Effect of themenstrual cycle on energy and nutrient intake. Am J Clin Nutr 60:895–899, 1994.
459. Matheny M, Zhang Y, Shapiro A, Tumer N, Scarpace PJ. Centraloverexpression of leptin antagonist reduces wheel running and under-scores importance of endogenous leptin receptor activity in energyhomeostasis. Am J Physiol Regul Integr Comp Physiol 297: R1254–R1261, 2009.
460. Mathes WF, Brownley KA, Mo X, Bulik CM. The biology of bingeeating. Appetite 52: 545–553, 2009.
461. Matsubara M, Sakata I, Wada R, Yamazaki M, Inoue K, Sakai T.Estrogen modulates ghrelin expression in the female rat stomach. Pep-tides 25: 289–297, 2004.
462. Mauvais-Jarvis F. Estrogen and androgen receptors: regulators of fuelhomeostasis and emerging targets for diabetes and obesity. TrendsEndocrinol Metab 22: 24–33, 2011.
463. Mauvais-Jarvis F, Clegg DJ, Hevener AL. The role of estrogens incontrol of energy balance and glucose homeostasis. Endocr Rev 34:309–338, 2013.
464. Mayer C, Acosta-Martinez M, Dubois SL, Wolfe A, Radovick S,Boehm U, Levine JE. Timing and completion of puberty in female micedepend on estrogen receptor �-signaling in kisspeptin neurons. Proc NatlAcad Sci USA 107: 22693–22698, 2010.
465. Mazzeo SE, Saunders R, Mitchell KS. Gender and binge eating amongbariatric surgery candidates. Eat Behav 7: 47–52, 2006.
466. McCarthy MM, Arnold AP, Ball GF, Blaustein JD, De Vries GJ. Sexdifferences in the brain: the not so inconvenient truth. J Neurosci 32:2241–2247, 2012.
467. McCarthy MM, Caba M, Komisaruk BR, Beyer C. Modulation byestrogen and progesterone of the effect of muscimol on nociception in thespinal cord. Pharmacol Biochem Behav 37: 123–128, 1990.
468. McCarthy MM, Coirini H, Schumacher M, Pfaff DW, McEwen BS,Schwartz-Giblin S. Ovarian steroid modulation of [3H]muscimol bind-ing in the spinal cord of the rat. Brain Res 556: 321–323, 1991.
469. McDermott LJ, Jorgensen DE, Byers DJ. Estradiol and progesteronesuppress feeding induced by 2-deoxy-D-glucose. Physiol Behav 32:731–736, 1984.
470. McEwen B. Estrogen actions throughout the brain. Recent Prog HormRes 57: 357–384, 2002.
471. McEwen BS, Alves SE. Estrogen actions in the central nervous system.Endocr Rev 20: 279–307, 1999.
472. McVay MA, Copeland AL, Newman HS, Geiselman PJ. Food crav-ings and food cue responding across the menstrual cycle in a non-eatingdisordered sample. Appetite 59: 591–600, 2012.
473. Melhorn SJ, Krause EG, Scott KA, Mooney MR, Johnson JD,Woods SC, Sakai RR. Meal patterns and hypothalamic NPY expressionduring chronic social stress and recovery. Am J Physiol Regul IntegrComp Physiol 299: R813–R822, 2010.
474. Meli R, Pacilio M, Raso GM, Esposito E, Coppola A, Nasti A, DiCarlo C, Nappi C, Di Carlo R. Estrogen and raloxifene modulate leptinand its receptor in hypothalamus and adipose tissue from ovariectomizedrats. Endocrinology 145: 3115–3121, 2004.
475. Mendelsohn ME, Karas RH. Rapid progress for non-nuclear estrogenreceptor signaling. J Clin Invest 120: 2277–2279, 2010.
476. Micevych P, Eckersell CB, Holland K, Smith A. Induction of CCKmRNA levels in the limbic-hypothalamic circuit: time course and site-specific effects of estrogen. J Neurobiol 30: 465–479, 1996.
477. Micevych P, Sinchak K. Estradiol regulation of progesterone synthesisin the brain. Mol Cell Endocrinol 290: 44–50, 2008.
478. Micevych P, Sinchak K. The neurosteroid progesterone underlies estro-gen positive feedback of the LH surge. Front Endocrinol (Lausanne) 2:90, 2011.
479. Micevych PE, Kelly MJ. Membrane estrogen receptor regulation ofhypothalamic function. Neuroendocrinology 96: 103–110, 2012.
480. Michener W, Rozin P, Freeman E, Gale L. The role of low proges-terone and tension as triggers of perimenstrual chocolate and sweetscraving: some negative experimental evidence. Physiol Behav 67: 417–420, 1999.
481. Mikolajczyk E, Grzywacz A, Samochowiec J. The association ofcatechol-O-methyltransferase genotype with the phenotype of womenwith eating disorders. Brain Res 1307: 142–148, 2010.
482. Miller A, Vo H, Huo L, Roca C, Schmidt PJ, Rubinow DR. Estrogenreceptor alpha (ESR-1) associations with psychological traits in womenwith PMDD and controls. J Psychiatr Res 44: 788–794, 2010.
483. Miller KK. Endocrine dysregulation in anorexia nervosa update. J ClinEndocrinol Metab 96: 2939–2949, 2011.
484. Miller WL, Auchus RJ. The molecular biology, biochemistry, andphysiology of human steroidogenesis and its disorders. Endocr Rev 32:81–151, 2011.
485. Mindell S, Smith GP, Greenberg D. Corn oil and mineral oil stimulatesham feeding in rats. Physiol Behav 48: 283–287, 1990.
486. Misra M, Katzman D, Miller KK, Mendes N, Snelgrove D, RussellM, Goldstein MA, Ebrahimi S, Clauss L, Weigel T, Mickley D,Schoenfeld DA, Herzog DB, Klibanski A. Physiological estrogen
Review
R1259SEX DIFFERENCES IN THE PHYSIOLOGY OF EATING
AJP-Regul Integr Comp Physiol • doi:10.1152/ajpregu.00446.2012 • www.ajpregu.org
by 10.220.32.247 on Septem
ber 18, 2016http://ajpregu.physiology.org/
Dow
nloaded from

replacement increases bone density in adolescent girls with anorexianervosa. J Bone Miner Res 26: 2430–2438, 2011.
487. Mittelman-Smith MA, Williams H, Krajewski-Hall SJ, Lai J, Ciofi P,McMullen NT, Rance NE. Arcuate kisspeptin/neurokinin B/dynorphin(KNDy) neurons mediate the estrogen suppression of gonadotropinsecretion and body weight. Endocrinology 153: 2800–2812, 2012.
488. Montague CT, Prins JB, Sanders L, Digby JE, O’Rahilly S. Depot-and sex-specific differences in human leptin mRNA expression: impli-cations for the control of regional fat distribution. Diabetes 46: 342–347,1997.
489. Moran LJ, Noakes M, Clifton PM, Wittert GA, Tomlinson L, Gal-letly C, Luscombe ND, Norman RJ. Ghrelin and measures of satiety arealtered in polycystic ovary syndrome but not differentially affected bydiet composition. J Clin Endocrinol Metab 89: 3337–3344, 2004.
490. Moran TH, Bi S. Hyperphagia and obesity in OLETF rats lackingCCK-1 receptors. Philos Trans R Soc Lond B Biol Sci 361: 1211–1218,2006.
491. Moran TH, Norgren R, Crosby RJ, McHugh PR. Central and periph-eral vagal transport of cholecystokinin binding sites occurs in afferentfibers. Brain Res 526: 95–102, 1990.
492. Morris DH, Jones ME, Schoemaker MJ, McFadden E, Ashworth A,Swerdlow AJ. Body mass index, exercise, and other lifestyle factors inrelation to age at natural menopause: analyses from the breakthroughgenerations study. Am J Epidemiol 175: 998–1005, 2012.
493. Morris JA, Jordan CL, Breedlove SM. Sexual differentiation of thevertebrate nervous system. Nat Neurosci 7: 1034–1039, 2004.
494. Morton GJ, Cummings DE, Baskin DG, Barsh GS, Schwartz MW.Central nervous system control of food intake and body weight. Nature443: 289–295, 2006.
495. Moverare-Skrtic S, Venken K, Andersson N, Lindberg MK, Svens-son J, Swanson C, Vanderschueren D, Oscarsson J, Gustafsson JA,Ohlsson C. Dihydrotestosterone treatment results in obesity and alteredlipid metabolism in orchidectomized mice. Obesity (Silver Spring) 14:662–672, 2006.
496. Mozaffarian D, Hao T, Rimm EB, Willett WC, Hu FB. Changes indiet and lifestyle and long-term weight gain in women and men. N EnglJ Med 364: 2392–2404, 2011.
497. Mulet T, Pico C, Oliver P, Palou A. Blood leptin homeostasis: sex-associated differences in circulating leptin levels in rats are independentof tissue leptin expression. Int J Biochem Cell Biol 35: 104–110, 2003.
498. Murakami M, Matsuzaki T, Iwasa T, Yasui T, Irahara M, Osugi T,Tsutsui K. Hypophysiotropic role of RFamide-related peptide-3 in theinhibition of LH secretion in female rats. J Endocrinol 199: 105–112,2008.
499. Musatov S, Chen W, Pfaff DW, Mobbs CV, Yang XJ, Clegg DJ,Kaplitt MG, Ogawa S. Silencing of estrogen receptor alpha in theventromedial nucleus of hypothalamus leads to metabolic syndrome.Proc Natl Acad Sci USA 104: 2501–2506, 2007.
500. Myers MG, Cowley MA, Munzberg H. Mechanisms of leptin actionand leptin resistance. Annu Rev Physiol 70: 537–556, 2008.
501. Myers MG Jr, Munzberg H, Leinninger GM, Leshan RL. Thegeometry of leptin action in the brain: more complicated than a simpleARC. Cell Metab 9: 117–123, 2009.
502. Naaz A, Zakroczymski M, Heine P, Taylor J, Saunders P, Lubahn D,Cooke PS. Effect of ovariectomy on adipose tissue of mice in theabsence of estrogen receptor � (ER�): a potential role for estrogenreceptor beta (ER�). Horm Metab Res 34: 758–763, 2002.
503. Nadal A, Ropero AB, Laribi O, Maillet M, Fuentes E, Soria B.Nongenomic actions of estrogens and xenoestrogens by binding at aplasma membrane receptor unrelated to estrogen receptor alpha andestrogen receptor �. Proc Natl Acad Sci USA 97: 11603–11608, 2000.
504. Naessen S, Carlstrom K, Bystrom B, Pierre Y, Hirschberg AL.Effects of an antiandrogenic oral contraceptive on appetite and eatingbehavior in bulimic women. Psychoneuroendocrinology 32: 548–554,2007.
505. Naessen S, Carlstrom K, Garoff L, Glant R, Hirschberg AL. Poly-cystic ovary syndrome in bulimic women—an evaluation based on thenew diagnostic criteria. Gynecol Endocrinol 22: 388–394, 2006.
506. Naessen S, Carlstrom K, Holst JJ, Hellstrom PM, Hirschberg AL.Women with bulimia nervosa exhibit attenuated secretion of glucagon-like peptide 1, pancreatic polypeptide, and insulin in response to a meal.Am J Clin Nutr 94: 967–972, 2011.
507. Nance DM, Gorski RA. Neurohormonal determinants of sex differencesin the hypothalamic regulation of feeding behavior and body weight inthe rat. Pharmacol Biochem Behav 3: 155–162, 1975.
508. Neal-Perry G, Santoro NF. Aging in the hypothalamic-pituitary-ovarianaxis In: Knobil and Neill’s Physiology of Reproduction (3rd ed.), editedby Neill JD. St. Louis, MO: Elsevier, 2006, p. 2729–2755.
509. Neary MT, Batterham RL. Gaining new insights into food reward withfunctional neuroimaging. Forum Nutr 63: 152–163, 2010.
510. Neill JD, Freeman ME, Tillson SA. Control of the proestrus surge ofprolactin and luteinizing hormone secretion by estrogens in the rat.Endocrinology 89: 1448–1453, 1971.
511. Nequin LG, Alvarez J, Schwartz NB. Measurement of serum steroidand gonadotropin levels and uterine and ovarian variables throughout 4day and 5 day estrous cycles in the rat. Biol Reprod 20: 659–670, 1979.
512. Nilsson S, Gustafsson JA. Estrogen receptors: therapies targeted toreceptor subtypes. Clin Pharmacol Ther 89: 44–55, 2011.
513. Nohara K, Zhang Y, Waraich RS, Laque A, Tiano JP, Tong J,Munzberg H, Mauvais-Jarvis F. Early-life exposure to testosteroneprograms the hypothalamic melanocortin system. Endocrinology 152:1661–1669, 2011.
514. Nolan LJ, Guss JL, Liddle RA, Pi-Sunyer FX, Kissileff HR. Elevatedplasma cholecystokinin and appetitive ratings after consumption of aliquid meal in humans. Nutrition 19: 553–557, 2003.
515. Norgren R, Hajnal A, Mungarndee SS. Gustatory reward and thenucleus accumbens. Physiol Behav 89: 531–535, 2006.
516. Nunez AA. Dose-dependent effects of testosterone on feeding and bodyweight in male rats. Behav Neural Biol 34: 445–449, 1982.
517. Nunez AA, Gray JM, Wade GN. Food intake and adipose tissuelipoprotein lipase activity after hypothalamic estradiol benzoate implantsin rats. Physiol Behav 25: 595–598, 1980.
518. Nunez AA, Grundman M. Testosterone affects food intake and bodyweight of weanling male rats. Pharmacol Biochem Behav 16: 933–936,1982.
519. Nunez AA, Siegel LI, Wade GN. Central effects of testosterone on foodintake in male rats. Physiol Behav 24: 469–471, 1980.
520. O’Donnell L, Meachem SJ, Stanton PG, McLachlan RI. Endocrineregulation of spermatogenesis. In: Knobil and Neill’s Physiology ofReproduction (3rd ed.), edited by Neill JD. St. Louis, MO: Elsevier/Academic, 2006, p. 1017–1069.
521. O’Rahilly S, Farooqi IS. Genetics of obesity. Philos Trans R Soc LondB Biol Sci 361: 1095–1105, 2006.
522. Ochner C, Teixeira J, Geary N, Asarian L. Greater short-term weightloss in women 20–45 versus 55–65 years of age following bariatricsurgery. Obes Surg 23: 1650–1654, 2013.
523. Ohlsson C, Hellberg N, Parini P, Vidal O, Bohlooly YM, Rudling M,Lindberg MK, Warner M, Angelin B, Gustafsson JA. Obesity anddisturbed lipoprotein profile in estrogen receptor-alpha-deficient malemice. Biochem Biophys Res Commun 278: 640–645, 2000.
524. Oka K, Hirano T, Noguchi M. Changes in the concentration oftestosterone in serum during the menstrual cycle, as determined by liquidchromatography. Clin Chem 34: 557–560, 1988.
525. Okura T, Koda M, Ando F, Niino N, Ohta S, Shimokata H. Associ-ation of polymorphisms in the estrogen receptor alpha gene with body fatdistribution. Int J Obes Relat Metab Disord 27: 1020–1027, 2003.
526. Olafsdottir AS, Skuladottir GV, Thorsdottir I, Hauksson A, Stein-grimsdottir L. Maternal diet in early and late pregnancy in relation toweight gain. Int J Obes (Lond) 30: 492–499, 2006.
527. Olofsson LE, Pierce AA, Xu AW. Functional requirement of AgRP andNPY neurons in ovarian cycle-dependent regulation of food intake. ProcNatl Acad Sci USA 106: 15932–15937, 2009.
528. Olster DH, Blaustein JD. Biochemical and immunocytochemical as-sessment of neural progestin receptors following estradiol treatments thateliminate the sex difference in progesterone-facilitated lordosis in guineapigs. J Neuroendocrinol 2: 79–86, 1990.
529. Olster DH, Blaustein JD. Estradiol pulses induce progestin receptorsselectively in substance P-immunoreactive neurons in the ventrolateralhypothalamus of female guinea pigs. J Neurobiol 23: 293–301, 1992.
530. Overman WH. Sex differences in early childhood, adolescence, andadulthood on cognitive tasks that rely on orbital prefrontal cortex. BrainCogn 55: 134–147, 2004.
531. Paeratakul S, White MA, Williamson DA, Ryan DH, Bray GA. Sex,race/ethnicity, socioeconomic status, and BMI in relation to self-percep-tion of overweight. Obes Res 10: 345–350, 2002.
Review
R1260 SEX DIFFERENCES IN THE PHYSIOLOGY OF EATING
AJP-Regul Integr Comp Physiol • doi:10.1152/ajpregu.00446.2012 • www.ajpregu.org
by 10.220.32.247 on Septem
ber 18, 2016http://ajpregu.physiology.org/
Dow
nloaded from

532. Pagotto U, Gambineri A, Pelusi C, Genghini S, Cacciari M, Otto B,Castaneda T, Tschop M, Pasquali R. Testosterone replacement therapyrestores normal ghrelin in hypogonadal men. J Clin Endocrinol Metab88: 4139–4143, 2003.
533. Palmer K, Gray Central vs JM. peripheral effects of estrogen on foodintake and lipoprotein lipase activity in ovariectomized rats. PhysiolBehav 37: 187–189, 1986.
534. Palmiter RD, Erickson JC, Hollopeter G, Baraban SC, SchwartzMW. Life without neuropeptide Y. Recent Prog Horm Res 53: 163–199,1998.
535. Panotopoulos G, Ruiz JC, Raison J, Guy-Grand B, Basdevant A.Menopause, fat and lean distribution in obese women. Maturitas 25:11–19, 1996.
536. Paolisso G, Rizzo MR, Mazziotti G, Rotondi M, Tagliamonte MR,Varricchio G, Carella C, Varricchio M. Lack of association betweenchanges in plasma leptin concentration and in food intake during themenstrual cycle. Eur J Clin Invest 29: 490–495, 1999.
537. Park CJ, Zhao Z, Glidewell-Kenney C, Lazic M, Chambon P, KrustA, Weiss J, Clegg DJ, Dunaif A, Jameson JL, Levine JE. Geneticrescue of nonclassical ER� signaling normalizes energy balance in obeseER�-null mutant mice. J Clin Invest 121: 604–612, 2011.
538. Parker GC, Bishop C, Coscina DV. Estrous cycle and food availabilityaffect feeding induced by amygdala 5-HT receptor blockade. PharmacolBiochem Behav 71: 701–707, 2002.
539. Parnell W, Wilson N, Alexander D, Wohlers M, Williden M, MannJ, Gray A. Exploring the relationship between sugars and obesity. PublicHealth Nutr 11: 860–866, 2008.
540. Patil CL, Abrams ET, Steinmetz AR, Young SL. Appetite sensationsand nausea and vomiting in pregnancy: an overview of the explanations.Ecol Food Nutr 51: 394–417, 2012.
541. Paulo RC, Brundage R, Cosma M, Mielke KL, Bowers CY, VeldhuisJD. Estrogen elevates the peak overnight production rate of acylatedghrelin. J Clin Endocrinol Metab 93: 4440–4447, 2008.
542. Pearson CM, Combs JL, Zapolski TC, Smith GT. A longitudinaltransactional risk model for early eating disorder onset. J AbnormPsychol 121: 707–718, 2012.
543. Pelchat ML. Food cravings in young and elderly adults. Appetite 28:103–113, 1997.
544. Pelkman CL, Chow M, Heinbach RA, Rolls BJ. Short-term effects ofa progestational contraceptive drug on food intake, resting energy ex-penditure, and body weight in young women. Am J Clin Nutr 73: 19–26,2001.
545. Pelleymounter MA, Baker MB, McCaleb M. Does estradiol mediateleptin’s effects on adiposity and body weight? Am J Physiol EndocrinolMetab 276: E955–E963, 1999.
546. Petersen S. Effects of testosterone upon feeding in male mice. AnimBehav 26: 945–952, 1978.
547. Petersen S. The temporal pattern of feeding over the oestrous cycle ofthe mouse. Anim Behav 24: 939–955, 1976.
548. Pfaff D, Waters E, Khan Q, Zhang X, Numan M. Minireview:estrogen receptor-initiated mechanisms causal to mammalian reproduc-tive behaviors. Endocrinology 152: 1209–1217, 2011.
549. Pfaff DW, Sakuma Y, Kow LM, Lee WL, Easton A. Hormonal,neural, and genomic mechanisms for female reproductive behaviors,motivation, and arousal. In: Knobil and Neill’s Physiology of Reproduc-tion (3rd ed.), edited by Neill JD. Amsterdam, The Netherlands: Elsevier/Academic, 2006, p. 1825–1920.
550. Phillips GB, Jing T, Heymsfield SB. Does insulin resistance, visceraladiposity, or a sex hormone alteration underlie the metabolic syndrome?Studies in women. Metabolism 57: 838–844, 2008.
551. Phoenix CH, Goy RW, Gerall AA, Young WC. Organizing action ofprenatally administered testosterone propionate on the tissues mediatingmating behavior in the female guinea pig. Endocrinology 65: 369–382,1959.
552. Pinilla L, Aguilar E, Dieguez C, Millar RP, Tena-Sempere M.Kisspeptins and reproduction: physiological roles and regulatory mech-anisms. Physiol Rev 92: 1235–1316, 2012.
553. Pinsky MR, Hellstrom WJ. Hypogonadism, ADAM, and hormonereplacement. Ther Adv Urol 2: 99–104, 2010.
554. Pirke KM, Broocks A, Wilckens T, Marquard R, Schweiger U.Starvation-induced hyperactivity in the rat: the role of endocrine andneurotransmitter changes. Neurosci Biobehav Rev 17: 287–294, 1993.
555. Pirke KM, Kellner MB, Friess E, Krieg JC, Fichter MM. Satiety andcholecystokinin. Int J Eat Disord 15: 63–69, 1994.
556. Plant TM. A comparison of the neuroendocrine mechanisms underlyingthe initiation of the preovulatory LH surge in the human, Old Worldmonkey and rodent. Front Neuroendocrinol 33: 160–168, 2012.
557. Pletzer B, Kronbichler M, Aichhorn M, Bergmann J, Ladurner G,Kerschbaum HH. Menstrual cycle and hormonal contraceptive usemodulate human brain structure. Brain Res 1348: 55–62, 2010.
558. Pliner P, Fleming AS. Food intake, body weight, and sweetness pref-erences over the menstrual cycle in humans. Physiol Behav 30: 663–666,1983.
559. Poeschla B, Gibbs J, Simansky KJ, Greenberg D, Smith GP. Chole-cystokinin-induced satiety depends on activation of 5-HT1C receptors.Am J Physiol Regul Integr Comp Physiol 264: R62–R64, 1993.
560. Poeschla B, Gibbs J, Simansky KJ, Smith GP. The 5-HT1A agonist8-OH-DPAT attenuates the satiating action of cholecystokinin. Pharma-col Biochem Behav 42: 541–543, 1992.
561. Pohle-Krauza RJ, Carey KH, Pelkman CL. Dietary restraint andmenstrual cycle phase modulated L-phenylalanine-induced satiety.Physiol Behav 93: 851–861, 2008.
562. Polidori C, Geary N. Estradiol treatment fails to affect the feedingresponses to melanocortin-3/4 receptor agonism or antagonism in ovari-ectomized rats. Peptides 23: 1697–1700, 2002.
563. Popa SM, Clifton DK, Steiner RA. The role of kisspeptins and GPR54in the neuroendocrine regulation of reproduction. Annu Rev Physiol 70:213–238, 2008.
564. Pope JF, Skinner JD, Carruth BR. Cravings and aversions of pregnantadolescents. J Am Diet Assoc 92: 1479–1482, 1992.
565. Powell MR, Hendricks B. Body schema, gender, and other correlates innonclinical populations. Genet Soc Gen Psychol Monogr 125: 333–412,1999.
566. Poyastro Pinheiro A, Thornton LM, Plotonicov KH, Tozzi F, KlumpKL, Berrettini WH, Brandt H, Crawford S, Crow S, Fichter MM,Goldman D, Halmi KA, Johnson C, Kaplan AS, Keel P, LaVia M,Mitchell J, Rotondo A, Strober M, Treasure J, Woodside DB, VonHolle A, Hamer R, Kaye WH, Bulik CM. Patterns of menstrualdisturbance in eating disorders. Int J Eat Disord 40: 424–434, 2007.
567. Pralong FP. Insulin and NPY pathways and the control of GnRHfunction and puberty onset. Mol Cell Endocrinol 324: 82–86, 2010.
568. Pratt GM, Learn CA, Hughes GD, Clark BL, Warthen M, Pories W.Demographics and outcomes at American Society for Metabolic andBariatric Surgery Centers of Excellence. Surg Endosc 23: 795–799,2009.
569. Prentice AM, Prentice A. Energy costs of lactation. Annu Rev Nutr 8:63–79, 1988.
570. Price WA, Torem MS, DiMarzio LR. Premenstrual exacerbation ofbulimia. Psychosomatics 28: 378–379, 1987.
571. Procopio M, Marriott P. Intrauterine hormonal environment and risk ofdeveloping anorexia nervosa. Arch Gen Psychiatry 64: 1402–1407, 2007.
572. Protopopescu X, Butler T, Pan H, Root J, Altemus M, Polanecsky M,McEwen B, Silbersweig D, Stern E. Hippocampal structural changesacross the menstrual cycle. Hippocampus 18: 985–988, 2008.
573. Prutkin J, Fisher EM, Etter L, Fast K, Gardner E, Lucchina LA,Snyder DJ, Tie K, Weiffenbach J, Bartoshuk LM. Genetic variationand inferences about perceived taste intensity in mice and men. PhysiolBehav 69: 161–173, 2000.
574. Qian S, Chen H, Weingarth D, Trumbauer ME, Novi DE, Guan X,Yu H, Shen Z, Feng Y, Frazier E, Chen A, Camacho RE, ShearmanLP, Gopal-Truter S, MacNeil DJ, Van der Ploeg LH, Marsh DJ.Neither agouti-related protein nor neuropeptide Y is critically requiredfor the regulation of energy homeostasis in mice. Mol Cell Biol 22:5027–5035, 2002.
575. Qiao GF, Li BY, Lu YJ, Fu YL, Schild JH. 17�-estradiol restoresexcitability of a sexually dimorphic subset of myelinated vagal afferentsin ovariectomized rats. Am J Physiol Cell Physiol 297: C654–C664,2009.
576. Qiu J, Bosch MA, Tobias SC, Krust A, Graham SM, Murphy SJ,Korach KS, Chambon P, Scanlan TS, Ronnekleiv OK, Kelly MJ. AG-protein-coupled estrogen receptor is involved in hypothalamic controlof energy homeostasis. J Neurosci 26: 5649–5655, 2006.
577. Quinton SJ, Smith AR, Joiner T. The 2 to 4 digit ratio (2D:4D) andeating disorder diagnosis in women. Pers Individ Dif 51: 402–405, 2011.
578. Racine SE, Culbert KM, Keel PK, Sisk CL, Alexandra Burt S,Klump KL. Differential associations between ovarian hormones anddisordered eating symptoms across the menstrual cycle in women. Int JEat Disord 45: 333–344, 2012.
Review
R1261SEX DIFFERENCES IN THE PHYSIOLOGY OF EATING
AJP-Regul Integr Comp Physiol • doi:10.1152/ajpregu.00446.2012 • www.ajpregu.org
by 10.220.32.247 on Septem
ber 18, 2016http://ajpregu.physiology.org/
Dow
nloaded from

579. Racine SE, Culbert KM, Keel PK, Sisk CL, Burt SA, Klump KL.Differential associations between ovarian hormones and disordered eat-ing symptoms across the menstrual cycle in women. Int J Eat Disord 45:333–344, 2012.
580. Racine SE, Keel PK, Burt SA, Sisk CL, Neale M, Boker S, KlumpKL. Individual differences in the relationship between ovarian hormonesand emotional eating across the menstrual cycle: a role for personality?Eat Behav 14: 161–166, 2013.
581. Raevuori A, Kaprio J, Hoek HW, Sihvola E, Rissanen A, Keski-Rahkonen A. Anorexia and bulimia nervosa in same-sex and opposite-sex twins: lack of association with twin type in a nationwide study ofFinnish twins. Am J Psychiatry 165: 1604–1610, 2008.
582. Ramirez I. Estradiol-induced changes in lipoprotein lipase, eating, andbody weight in rats. Am J Physiol Endocrinol Metab 240: E533–E538,1981.
583. Ramirez I. Relation between estrogen-induced hyperlipemia and foodintake and body weight in rats. Physiol Behav 25: 511–518, 1980.
584. Reichborn-Kjennerud T, Bulik CM, Kendler KS, Roysamb E, MaesH, Tambs K, Harris JR. Gender differences in binge-eating: a popula-tion-based twin study. Acta Psychiatr Scand 108: 196–202, 2003.
585. Reimer RA, Debert CT, House JL, Poulin MJ. Dietary and metabolicdifferences in pre- versus postmenopausal women taking or not takinghormone replacement therapy. Physiol Behav 84: 303–312, 2005.
586. Resch M, Szendei G, Haasz P. Bulimia from a gynecological view:hormonal changes. J Obstet Gynaecol 24: 907–910, 2004.
587. Reubinoff BE, Wurtman J, Rojansky N, Adler D, Stein P, SchenkerJG, Brzezinski A. Effects of hormone replacement therapy on weight,body composition, fat distribution, and food intake in early postmeno-pausal women: a prospective study. Fertil Steril 64: 963–968, 1995.
588. Revankar CM, Cimino DF, Sklar LA, Arterburn JB, Prossnitz ER.A transmembrane intracellular estrogen receptor mediates rapid cellsignaling. Science 307: 1625–1630, 2005.
589. Richard D. Effects of ovarian hormones on energy balance and brownadipose tissue thermogenesis. Am J Physiol Regul Integr Comp Physiol250: R245–R249, 1986.
590. Richter CP. Total self-regulatory functions in animals and humanbeings. Harvey Lecture Series 38: 63–103, 1942–1943.
591. Ritter RC. Gastrointestinal mechanisms of satiation for food. PhysiolBehav 81: 249–273, 2004.
592. Rivera HM, Eckel LA. Activation of central, but not peripheral, estro-gen receptors is necessary for estradiol’s anorexigenic effect in ovariec-tomized rats. Endocrinology 151: 5680–5688, 2010.
593. Rivera HM, Eckel LA. The anorectic effect of fenfluramine is increasedby estradiol treatment in ovariectomized rats. Physiol Behav 86: 331–337, 2005.
594. Rivera HM, Santollo J, Nikonova LV, Eckel LA. Estradiol increasesthe anorexia associated with increased 5-HT(2C) receptor activation inovariectomized rats. Physiol Behav 105: 188–194, 2011.
595. Rocha M, Bing C, Williams G, Puerta M. Physiologic estradiol levelsenhance hypothalamic expression of the long form of the leptin receptorin intact rats. J Nutr Biochem 15: 328–334, 2004.
596. Rock CL, Gorenflo DW, Drewnowski A, Demitrack MA. Nutritionalcharacteristics, eating pathology, and hormonal status in young women.Am J Clin Nutr 64: 566–571, 1996.
597. Rodin J, Mancuso J, Granger J, Nelbach E. Food cravings in relationto body mass index, restraint and estradiol levels: a repeated-measuresstudy in healthy women. Appetite 17: 177–185, 1991.
598. Roemmich JN, Clark PA, Berr SS, Mai V, Mantzoros CS, Flier JS,Weltman A, Rogol AD. Gender differences in leptin levels duringpuberty are related to the subcutaneous fat depot and sex steroids. Am JPhysiol Endocrinol Metab 275: E543–E551, 1998.
599. Roepke TA. Oestrogen modulates hypothalamic control of energy ho-meostasis through multiple mechanisms. J Neuroendocrinol 21: 141–150, 2009.
600. Roepke TA, Bosch MA, Rick EA, Lee B, Wagner EJ, Seidlova-Wuttke D, Wuttke W, Scanlan TS, Ronnekleiv OK, Kelly MJ.Contribution of a membrane estrogen receptor to the estrogenic regula-tion of body temperature and energy homeostasis. Endocrinology 151:4926–4937, 2010.
601. Roepke TA, Malyala A, Bosch MA, Kelly MJ, Ronnekleiv OK.Estrogen regulation of genes important for K� channel signaling in thearcuate nucleus. Endocrinology 148: 4937–4951, 2007.
602. Roepke TA, Ronnekleiv OK, Kelly MJ. Physiological consequences ofmembrane-initiated estrogen signaling in the brain. Front Biosci 16:1560–1573, 2011.
603. Roesch DM. Effects of selective estrogen receptor agonists on foodintake and body weight gain in rats. Physiol Behav 87: 39–44, 2006.
604. Rogers NH, Perfield JW, 2nd Strissel KJ, Obin MS, Greenberg AS.Reduced energy expenditure and increased inflammation are early eventsin the development of ovariectomy-induced obesity. Endocrinology 150:2161–2168, 2009.
605. Rolls BJ, Fedoroff IC, Guthrie JF. Gender differences in eatingbehavior and body weight regulation. Health Psychol 10: 133–142, 1991.
606. Rolls ET. Brain mechanisms underlying flavour and appetite. PhilosTrans R Soc Lond B Biol Sci 361: 1123–1136, 2006.
607. Rolls ET. Taste, olfactory and food texture reward processing in thebrain and obesity. Int J Obes (Lond) 35: 550–561, 2011.
608. Rolls ET. Taste, olfactory and food texture reward processing in thebrain and the control of appetite. Proc Nutr Soc 71: 488–501, 2012.
609. Rolls ET, Grabenhorst F. The orbitofrontal cortex and beyond: fromaffect to decision-making. Prog Neurobiol 86: 216–244, 2008.
610. Rometo AM, Krajewski SJ, Voytko ML, Rance NE. Hypertrophy andincreased kisspeptin gene expression in the hypothalamic infundibularnucleus of postmenopausal women and ovariectomized monkeys. J ClinEndocrinol Metab 92: 2744–2750, 2007.
611. Roselli CE, Liu M, Hurn PD. Brain aromatization: classic roles andnew perspectives. Semin Reprod Med 27: 207–217, 2009.
612. Rosenbaum M, Leibel RL. Clinical review 107: Role of gonadalsteroids in the sexual dimorphisms in body composition and circulatingconcentrations of leptin. J Clin Endocrinol Metab 84: 1784–1789, 1999.
613. Rosenbaum M, Nicolson M, Hirsch J, Heymsfield SB, Gallagher D,Chu F, Leibel RL. Effects of gender, body composition, and menopauseon plasma concentrations of leptin. J Clin Endocrinol Metab 81: 3424–3427, 1996.
614. Rosenbaum M, Sy M, Pavlovich K, Leibel RL, Hirsch J. Leptinreverses weight loss-induced changes in regional neural activity re-sponses to visual food stimuli. J Clin Invest 118: 2583–2591, 2008.
615. Rosenblatt H, Dyrenfurth I, Ferin M, vande Wiele RL. Food intakeand the menstrual cycle in rhesus monkeys. Physiol Behav 24: 447–449,1980.
616. Rosenkranz K, Hinney A, Ziegler A, Hermann H, Fichter M, MayerH, Siegfried W, Young JK, Remschmidt H, Hebebrand J. Systematicmutation screening of the estrogen receptor � gene in probands ofdifferent weight extremes: identification of several genetic variants. JClin Endocrinol Metab 83: 4524–4527, 1998.
617. Ross GT, Cargille CM, Lipsett MB, Rayford PL, Marshall JR, StrottCA, Rodbard D. Pituitary and gonadal hormones in women duringspontaneous and induced ovulatory cycles. Recent Prog Horm Res 26:1–62, 1970.
618. Roth JD, Erickson MR, Chen S, Parkes DG. GLP-1R and amylinagonism in metabolic disease: complementary mechanisms and futureopportunities. Br J Pharmacol 166: 121–136, 2012.
619. Roth JD, Roland BL, Cole RL, Trevaskis JL, Weyer C, Koda JE,Anderson CM, Parkes DG, Baron AD. Leptin responsiveness restoredby amylin agonism in diet-induced obesity: evidence from nonclinicaland clinical studies. Proc Natl Acad Sci USA 105: 7257–7262, 2008.
620. Routtenberg A. “Self-starvation” of rats living in activity wheels:adaptation effects. J Comp Physiol Psychol 66: 234–238, 1968.
621. Routtenberg A, Kuznesof AW. Self-starvation of rats living in activitywheels on a restricted feeding schedule. J Comp Physiol Psychol 64:414–421, 1967.
622. Rowland DL, Perrings TS, Thommes JA. Comparison of androgeniceffects on food intake and body weight in adult rats. Physiol Behav 24:205–209, 1980.
623. Rozin P, Levine E, Stoess C. Chocolate craving and liking. Appetite 17:199–212, 1991.
624. Rubinow DR, Schmidt PJ. Gonadal steroid regulation of mood: thelessons of premenstrual syndrome. Front Neuroendocrinol 27: 210–216,2006.
625. Saad F, Aversa A, Isidori AM, Gooren LJ. Testosterone as potentialeffective therapy in treatment of obesity in men with testosterone defi-ciency: a review. Curr Diabetes Rev 8: 131–143, 2012.
626. Sakata I, Tanaka T, Yamazaki M, Tanizaki T, Zheng Z, Sakai T.Gastric estrogen directly induces ghrelin expression and production in therat stomach. J Endocrinol 190: 749–757, 2006.
Review
R1262 SEX DIFFERENCES IN THE PHYSIOLOGY OF EATING
AJP-Regul Integr Comp Physiol • doi:10.1152/ajpregu.00446.2012 • www.ajpregu.org
by 10.220.32.247 on Septem
ber 18, 2016http://ajpregu.physiology.org/
Dow
nloaded from

627. Salamanca S, Uphouse L. Estradiol modulation of the hyperphagiainduced by the 5-HT1A agonist, 8-OH-DPAT. Pharmacol BiochemBehav 43: 953–955, 1992.
628. Salpeter SR, Walsh JM, Ormiston TM, Greyber E, Buckley NS,Salpeter EE. Meta-analysis: effect of hormone-replacement therapy oncomponents of the metabolic syndrome in postmenopausal women.Diabetes Obes Metab 8: 538–554, 2006.
629. Sammel MD, Freeman EW, Liu Z, Lin H, Guo W. Factors thatinfluence entry into stages of the menopausal transition. Menopause 16:1218–1227, 2009.
630. Samuel I, Mason EE, Renquist KE, Huang YH, Zimmerman MB,Jamal M. Bariatric surgery trends: an 18-year report from the Interna-tional Bariatric Surgery Registry. Am J Surg 192: 657–662, 2006.
631. Sancho C, Arija MV, Asorey O, Canals J. Epidemiology of eatingdisorders: a two year follow up in an early adolescent school population.Eur Child Adolesc Psychiatry 16: 495–504, 2007.
632. Sandoval DA, Ryan KK, de Kloet AD, Woods SC, Seeley RJ. Femalerats are relatively more sensitive to reduced lipid versus reduced carbo-hydrate availability. Nutr Diabetes 2: e27, 2012.
633. Santollo J, Eckel LA. Estradiol decreases the orexigenic effect ofneuropeptide Y, but not agouti-related protein, in ovariectomized rats.Behav Brain Res 191: 173–177, 2008.
634. Santollo J, Eckel LA. The orexigenic effect of melanin-concentratinghormone (MCH) is influenced by sex and stage of the estrous cycle.Physiol Behav 93: 842–850, 2008.
635. Santollo J, Katzenellenbogen BS, Katzenellenbogen JA, Eckel LA.Activation of ER� is necessary for estradiol’s anorexigenic effect infemale rats. Horm Behav 58: 872–877, 2010.
636. Santollo J, Torregrossa AM, Eckel LA. Estradiol acts in the medialpreoptic area, arcuate nucleus, and dorsal raphe nucleus to reduce foodintake in ovariectomized rats. Horm Behav 60: 86–93, 2011.
637. Santollo J, Wiley MD, Eckel LA. Acute activation of ER� decreasesfood intake, meal size, and body weight in ovariectomized rats. Am JPhysiol Regul Integr Comp Physiol 293: R2194–R2201, 2007.
638. Santoro N, Adel T, Skurnick JH. Decreased inhibin tone and increasedactivin A secretion characterize reproductive aging in women. FertilSteril 71: 658–662, 1999.
639. Savard R, Palmer JE, Greenwood MR. Effects of exercise training onregional adipose tissue metabolism in pregnant rats. Am J Physiol RegulIntegr Comp Physiol 250: R837–R844, 1986.
640. Sayegh R, Schiff I, Wurtman J, Spiers P, McDermott J, Wurtman R.The effect of a carbohydrate-rich beverage on mood, appetite, andcognitive function in women with premenstrual syndrome. Obstet Gyne-col 86: 520–528, 1995.
641. Scalera G. Effects of conditioned food aversions on nutritional behaviorin humans. Nutr Neurosci 5: 159–188, 2002.
642. Schieve LA, Cogswell ME, Scanlon KS. Trends in pregnancy weightgain within and outside ranges recommended by the Institute of Medicinein a WIC population. Matern Child Health J 2: 111–116, 1998.
643. Schlenker EH, Hansen SN. Sex-specific densities of estrogen receptorsalpha and beta in the subnuclei of the nucleus tractus solitarius, hypo-glossal nucleus and dorsal vagal motor nucleus weanling rats. Brain Res1123: 89–100, 2006.
644. Schmidt PJ, Rubinow DR. Sex hormones and mood in the perimeno-pause. Ann NY Acad Sci 1179: 70–85, 2009.
645. Schneider JE. Energy balance and reproduction. Physiol Behav 81:289–317, 2004.
646. Schneider JE, Klingerman CM, Abdulhay A. Sense and nonsense inmetabolic control of reproduction. Front Endocrinol (Lausanne) 3: 26,2012.
647. Schumacher M, Coirini H, Pfaff DW, McEwen BS. Light-dark differ-ences in behavioral sensitivity to oxytocin. Behav Neurosci 105: 487–492, 1991.
648. Schwartz GJ. Integrative capacity of the caudal brainstem in the controlof food intake. Philos Trans R Soc Lond B Biol Sci 361: 1275–1280,2006.
649. Schwartz SM, Wade GN. Effects of estradiol and progesterone on foodintake, body weight, and carcass adiposity in weanling rats. Am J PhysiolEndocrinol Metab 240: E499–E503, 1981.
650. Sclafani A, Hertwig H, Vigorito M, Feigin MB. Sex differences inpolysaccharide and sugar preferences in rats. Neurosci Biobehav Rev 11:241–251, 1987.
651. Sclafani A, Touzani K, Bodnar RJ. Dopamine and learned foodpreferences. Physiol Behav 104: 64–68, 2011.
652. Scott HM, Mason JI, Sharpe RM. Steroidogenesis in the fetal testis andits susceptibility to disruption by exogenous compounds. Endocr Rev 30:883–925, 2009.
653. Scott KA, Melhorn SJ, Sakai RR. Effects of chronic social stress onobesity. Curr Obes Rep 1: 16–25, 2012.
654. Sen S, Carpenter AH, Hochstadt J, Huddleston JY, Kustanovich V,Reynolds AA, Roberts S. Nutrition, weight gain and eating behavior inpregnancy: a review of experimental evidence for long-term effects onthe risk of obesity in offspring. Physiol Behav 107: 138–145, 2012.
655. Shaikh AA. Estrone and estradiol levels in the ovarian venous bloodfrom rats during the estrous cycle and pregnancy. Biol Reprod 5:297–307, 1971.
656. Shelley DN, Dwyer E, Johnson C, Wittkowski KM, Pfaff DW.Interactions between estrogen effects and hunger effects in ovariecto-mized female mice. I. Measures of arousal. Horm Behav 52: 546–553,2007.
657. Shen L, Wang DQ, Lo CM, Tso P, Davidson WS, Woods SC, Liu M.Estradiol increases the anorectic effect of central apolipoprotein A-IV.Endocrinology 151: 3163–3168, 2010.
658. Shepard KN, Michopoulos V, Toufexis DJ, Wilson ME. Genetic,epigenetic and environmental impact on sex differences in social behav-ior. Physiol Behav 97: 157–170, 2009.
659. Shetty G, Wilson G, Hardy MP, Niu E, Huhtaniemi I, Meistrich ML.Inhibition of recovery of spermatogenesis in irradiated rats by differentandrogens. Endocrinology 143: 3385–3396, 2002.
660. Shi H, Strader AD, Woods SC, Seeley RJ. Sexually dimorphic re-sponses to fat loss after caloric restriction or surgical lipectomy. Am JPhysiol Endocrinol Metab 293: E316–E326, 2007.
661. Shimizu H, Ohtani KI, Uehara Y, Abe Y, Takahashi H, Tsuchiya T,Sato N, Ibuki Y, Mori M. Orchiectomy and response to testosterone inthe development of obesity in young Otsuka-Long-Evans-TokushimaFatty (OLETF) rats. Int J Obes Relat Metab Disord 22: 318–324, 1998.
662. Shimomura Y, Shimizu H, Takahashi M, Sato N, Uehara Y, FukatsuA, Negishi M, Kobayashi I, Kobayashi S. The significance of decreasedambulatory activity during the generation by long-term observation ofobesity in ovariectomized rats. Physiol Behav 47: 155–159, 1990.
663. Shughrue PJ, Lane MV, Merchenthaler I. Comparative distribution ofestrogen receptor-� and -� mRNA in the rat central nervous system. JComp Neurol 388: 507–525, 1997.
664. Siegel LI, Nunez AA, Wade GN. Effects of androgens on dietaryself-selection and carcass composition in male rats. J Comp PhysiolPsychol 95: 529–539, 1981.
665. Siegfried Z, Berry EM, Hao S, Avraham Y. Animal models in theinvestigation of anorexia. Physiol Behav 79: 39–45, 2003.
666. Silveira LF, Latronico AC. Approach to the patient with hypogonado-tropic hypogonadism. J Clin Endocrinol Metab 98: 1781–1788, 2013.
667. Simerly RB. Wired on hormones: endocrine regulation of hypothalamicdevelopment. Curr Opin Neurobiol 15: 81–85, 2005.
668. Simonian SX, Herbison AE. Differential expression of estrogen recep-tor and neuropeptide Y by brainstem A1 and A2 noradrenaline neurons.Neuroscience 76: 517–529, 1997.
669. Simpson ER, Misso M, Hewitt KN, Hill RA, Boon WC, Jones ME,Kovacic A, Zhou J, Clyne CD. Estrogen—the good, the bad, and theunexpected. Endocr Rev 26: 322–330, 2005.
670. Sisk CL, Foster DL. The neural basis of puberty and adolescence. NatNeurosci 7: 1040–1047, 2004.
671. Sisk CL, Zehr JL. Pubertal hormones organize the adolescent brain andbehavior. Front Neuroendocrinol 26: 163–174, 2005.
672. Sites CK, L’Hommedieu GD, Toth MJ, Brochu M, Cooper BC,Fairhurst PA. The effect of hormone replacement therapy on bodycomposition, body fat distribution, and insulin sensitivity in menopausalwomen: a randomized, double-blind, placebo-controlled trial. J ClinEndocrinol Metab 90: 2701–2707, 2005.
673. Skibicka KP, Grill HJ. Hypothalamic and hindbrain melanocortinreceptors contribute to the feeding, thermogenic, and cardiovascularaction of melanocortins. Endocrinology 150: 5351–5361, 2009.
674. Slonaker JR. The effect of copulation, pregnancy, pseudopregnancy,and lactation on the voluntary activity and food consumption of thealbino rat. Am J Physiol 71: 362–394, 1925.
675. Smeets PA, de Graaf C, Stafleu A, van Osch MJ, Nievelstein RA, vander Grond J. Effect of satiety on brain activation during chocolatetasting in men and women. Am J Clin Nutr 83: 1297–1305, 2006.
676. Smith GP. Animal models of human eating disorders. Ann NY Acad Sci575: 63–72; discussion 72–64, 1989.
Review
R1263SEX DIFFERENCES IN THE PHYSIOLOGY OF EATING
AJP-Regul Integr Comp Physiol • doi:10.1152/ajpregu.00446.2012 • www.ajpregu.org
by 10.220.32.247 on Septem
ber 18, 2016http://ajpregu.physiology.org/
Dow
nloaded from

677. Smith GP. Cholecystokinin and treatment of meal size: proof of princi-ple. Obesity (Silver Spring) 14 Suppl 4: 168S–170S, 2006.
678. Smith GP. The controls of eating: a shift from nutritional homeostasis tobehavioral neuroscience. Nutrition 16: 814–820, 2000.
679. Smith GP. Sham feeding in rats with chronic, reversible gastric fistulas.Curr Protoc Neurosci Chap. 8: Unit 8.6D, 2001.
680. Smith JC. The history of the “Davis Rig”. Appetite 36: 93–98, 2001.681. Smith JT, Mark PJ, Waddell BJ. Developmental increases in plasma
leptin binding activity and tissue Ob-Re mRNA expression in the rat. JEndocrinol 184: 535–541, 2005.
682. Smith JT, Shahab M, Pereira A, Pau KY, Clarke IJ. Hypothalamicexpression of KISS1 and gonadotropin inhibitory hormone genes duringthe menstrual cycle of a non-human primate. Biol Reprod 83: 568–577,2010.
683. Smith JT, Waddell BJ. Developmental changes in plasma leptin andhypothalamic leptin receptor expression in the rat: peripubertal changesand the emergence of sex differences. J Endocrinol 176: 313–319, 2003.
684. Smith MS, Freeman ME, Neill JD. The control of progesterone secre-tion during the estrous cycle and early pseudopregnancy in the rat:prolactin, gonadotropin and steroid levels associated with rescue of thecorpus luteum of pseudopregnancy. Endocrinology 96: 219–226, 1975.
685. Snyder DJ, Bartoshuk LM. Epidemiological studies of taste function:discussion and perspectives. Ann NY Acad Sci 1170: 574–580, 2009.
686. Sorensen MB, Rasmussen V, Jensen G, Ottesen B. Temporal changesin clinic and ambulatory blood pressure during cyclic post-menopausalhormone replacement therapy. J Hypertens 18: 1387–1391, 2000.
687. Souquet AM, Rowland NE. Dexfenfluramine: action with estradiol onfood intake and body weight in ovariectomized rats. Am J Physiol RegulIntegr Comp Physiol 258: R211–R215, 1990.
688. Spary EJ, Maqbool A, Batten TF. Changes in oestrogen receptor alphaexpression in the nucleus of the solitary tract of the rat over the oestrouscycle and following ovariectomy. J Neuroendocrinol 22: 492–502, 2010.
689. Spary EJ, Maqbool A, Batten TF. Oestrogen receptors in the centralnervous system and evidence for their role in the control of cardiovas-cular function. J Chem Neuroanat 38: 185–196, 2009.
690. Spiteri T, Musatov S, Ogawa S, Ribeiro A, Pfaff DW, Agmo A.Estrogen-induced sexual incentive motivation, proceptivity and receptiv-ity depend on a functional estrogen receptor alpha in the ventromedialnucleus of the hypothalamus but not in the amygdala. Neuroendocrinol-ogy 91: 142–154, 2010.
691. Steingrimsdottir L, Greenwood MR, Brasel JA. Effect of pregnancy,lactation and a high-fat diet on adipose tissue in Osborne-Mendel rats. JNutr 110: 600–609, 1980.
692. Stengel A, Wang L, Goebel-Stengel M, Tache Y. Centrally injectedkisspeptin reduces food intake by increasing meal intervals in mice.Neuroreport 22: 253–257, 2011.
693. Sternson SM. Hypothalamic survival circuits: blueprints for purposivebehaviors. Neuron 77: 810–824, 2013.
694. Stice E, Dagher A. Genetic variation in dopaminergic reward in humans.Forum Nutr 63: 176–185, 2010.
695. Stice E, Spoor S, Bohon C, Small DM. Relation between obesity andblunted striatal response to food is moderated by TaqIA A1 allele.Science 322: 449–452, 2008.
696. Stice E, Yokum S, Burger KS, Epstein LH, Small DM. Youth at riskfor obesity show greater activation of striatal and somatosensory regionsto food. J Neurosci 31: 4360–4366, 2011.
697. Stotsenburg JM. The effect of spaying and semi-spaying young albinorats (Mus norvegicus albinus) on the growth in body weight and bodylength. Anat Rec 7: 183–194, 1913.
698. Stoving RK, Andries A, Brixen K, Bilenberg N, Horder K. Genderdifferences in outcome of eating disorders: a retrospective cohort study.Psychiatry Res 186: 362–366, 2011.
699. Stratford JM, Curtis KS, Contreras RJ. Linoleic acid increases chordatympani nerve responses to and behavioral preferences for monosodiumglutamate by male and female rats. Am J Physiol Regul Integr CompPhysiol 295: R764–R772, 2008.
700. Striegel-Moore RH, Rosselli F, Perrin N, DeBar L, Wilson GT, MayA, Kraemer HC. Gender difference in the prevalence of eating disordersymptoms. Int J Eat Disord 42: 471–474, 2009.
701. Strohmayer AJ, Smith GP. The meal pattern of genetically obese(ob/ob) mice. Appetite 8: 111–123, 1987.
702. Strubbe JH, Gorissen J. Meal patterning in the lactating rat. PhysiolBehav 25: 775–777, 1980.
703. Stunkard AJ, Messick S. The three-factor eating questionnaire tomeasure dietary restraint, disinhibition and hunger. J Psychosom Res 29:71–83, 1985.
704. Stutzmann F, Tan K, Vatin V, Dina C, Jouret B, Tichet J, Balkau B,Potoczna N, Horber F, O’Rahilly S, Farooqi IS, Froguel P, Meyre D.Prevalence of melanocortin-4 receptor deficiency in Europeans and theirage-dependent penetrance in multigenerational pedigrees. Diabetes 57:2511–2518, 2008.
705. Sullivan EL, Daniels AJ, Koegler FH, Cameron JL. Evidence infemale rhesus monkeys (Macaca mulatta) that nighttime caloric intake isnot associated with weight gain. Obes Res 13: 2072–2080, 2005.
706. Sullivan EL, Koegler FH, Cameron JL. Individual differences inphysical activity are closely associated with changes in body weight inadult female rhesus monkeys (Macaca mulatta). Am J Physiol RegulIntegr Comp Physiol 291: R633–R642, 2006.
707. Sullivan EL, Shearin J, Koegler FH, Cameron JL. Selective estrogenreceptor modulator promotes weight loss in ovariectomized female rhe-sus monkeys (Macaca mulatta) by decreasing food intake and increasingactivity. Am J Physiol Endocrinol Metab 302: E759–E767, 2012.
708. Sullivan PF, Bulik CM, Kendler KS. Genetic epidemiology of bingingand vomiting. Br J Psychiatry 173: 75–79, 1998.
709. Sundblad C, Bergman L, Eriksson E. High levels of free testosteronein women with bulimia nervosa. Acta Psychiatr Scand 90: 397–398,1994.
710. Sundblad C, Landen M, Eriksson T, Bergman L, Eriksson E. Effectsof the androgen antagonist flutamide and the serotonin reuptake inhibitorcitalopram in bulimia nervosa: a placebo-controlled pilot study. J ClinPsychopharmacol 25: 85–88, 2005.
711. Surampudi PN, Wang C, Swerdloff R. Hypogonadism in the agingmale diagnosis, potential benefits, and risks of testosterone replacementtherapy. Int J Endocrinol 2012: 625434, 2012.
712. Sutton GM, Trevaskis JL, Hulver MW, McMillan RP, MarkwardNJ, Babin MJ, Meyer EA, Butler AA. Diet-genotype interactions in thedevelopment of the obese, insulin-resistant phenotype of C57BL/6J micelacking melanocortin-3 or -4 receptors. Endocrinology 147: 2183–2196,2006.
713. Suzuki Y, Kurose Y, Takahashi H, Asakuma S, Azuma Y, KobayashiS. The differences in feeding-inhibitory responses to peripheral andcentral leptin between non-lactating and lactating rats. J Endocrinol 207:105–111, 2010.
714. Svendsen OL, Hassager C, Christiansen C. Age- and menopause-associated variations in body composition and fat distribution in healthywomen as measured by dual-energy X-ray absorptiometry. Metabolism44: 369–373, 1995.
715. Swanson SA, Crow SJ, Le Grange D, Swendsen J, Merikangas KR.Prevalence and correlates of eating disorders in adolescents. Results fromthe national comorbidity survey replication adolescent supplement. ArchGen Psychiatry 68: 714–723, 2011.
716. Swinburn B, Sacks G, Ravussin E. Increased food energy supply ismore than sufficient to explain the US epidemic of obesity. Am J ClinNutr 90: 1453–1456, 2009.
717. Swithers SE, McCurley M, Hamilton E, Doerflinger A. Influence ofovarian hormones on development of ingestive responding to alterationsin fatty acid oxidation in female rats. Horm Behav 54: 471–477, 2008.
718. Swithers SE, McCurley M, Scheibler A, Doerflinger A. Differentialeffects of lipoprivation and food deprivation on chow and milk intake in25- and 30-day-old rats. Appetite 45: 86–93, 2005.
719. Takiguchi S, Takata Y, Funakoshi A, Miyasaka K, Kataoka K,Fujimura Y, Goto T, Kono A. Disrupted cholecystokinin type-Areceptor (CCKAR) gene in OLETF rats. Gene 197: 169–175, 1997.
720. Tamashiro KL, Sakai RR, Shively CA, Karatsoreos IN, Reagan LP.Chronic stress, metabolism, and metabolic syndrome. Stress 14: 468–474, 2011.
721. Tanentsapf I, Heitmann BL, Adegboye AR. Systematic review ofclinical trials on dietary interventions to prevent excessive weight gainduring pregnancy among normal weight, overweight and obese women.BMC Pregnancy Childbirth 11: 81, 2011.
722. Tao YX. The melanocortin-4 receptor: pharmacology, pathophysiology,and physiology Endocr Rev 31: 506–543, 2010.
723. Tao YX. Mutations in melanocortin-4 receptor and human obesity. ProgMol Biol Transl Sci 88: 173–204, 2009.
724. Tapper CM, Brown-Grant K. The secretion and metabolic clearancerates of oestradiol in the rat. J Endocrinol 64: 215–227, 1975.
Review
R1264 SEX DIFFERENCES IN THE PHYSIOLOGY OF EATING
AJP-Regul Integr Comp Physiol • doi:10.1152/ajpregu.00446.2012 • www.ajpregu.org
by 10.220.32.247 on Septem
ber 18, 2016http://ajpregu.physiology.org/
Dow
nloaded from

725. Tarasuk V, Beaton GH. Menstrual-cycle patterns in energy and macro-nutrient intake. Am J Clin Nutr 53: 442–447, 1991.
726. Tarttelin MF, Gorski RA. The effects of ovarian steroids on food andwater intake and body weight in the female rat. Acta Endocrinol (Co-penh) 72: 551–568, 1973.
727. Tarttelin MF, Gorski RA. Variations in food and water intake in thenormal and acyclic female rat. Physiol Behav 7: 847–852, 1971.
728. Taya K, Komura H, Watanabe G, Sasamoto S. Peripheral blood levelsof immunoreactive inhibin during pseudopregnancy, pregnancy and lac-tation in the rat. J Endocrinol 121: 545–552, 1989.
729. Tchernof A, Despres JP. Pathophysiology of human visceral obesity: anupdate. Physiol Rev 93: 359–404, 2013.
730. Tehrani FR, Solaymani-Dodaran M, Tohidi M, Gohari MR, Azizi F.Modeling age at menopause using serum concentration of anti-Mullerianhormone. J Clin Endocrinol Metab 98: 729–735, 2013.
731. Thammacharoen S, Geary N, Lutz TA, Ogawa S, Asarian L. Diver-gent effects of estradiol and the estrogen receptor-alpha agonist PPT oneating and activation of PVN CRH neurons in ovariectomized rats andmice. Brain Res 1268: 88–96, 2009.
732. Thammacharoen S, Lutz TA, Geary N, Asarian L. Hindbrain admin-istration of estradiol inhibits feeding and activates estrogen receptor-alpha-expressing cells in the nucleus tractus solitarius of ovariectomizedrats. Endocrinology 149: 1609–1617, 2008.
733. Thompson FE, McNeel TS, Dowling EC, Midthune D, MorrissetteM, Zeruto CA. Interrelationships of added sugars intake, socioeconomicstatus, and race/ethnicity in adults in the United States: National HealthInterview Survey, 2005. J Am Diet Assoc 109: 1376–1383, 2009.
734. Thompson ME, Cox VC. The effects of estradiol on body weight andfood intake in normal weight VMH-lesioned rats. Physiol Behav 22:627–629, 1979.
735. Thorneycroft IH, Mishell DR Jr, Stone SC, Kharma KM, NakamuraRM. The relation of serum 17-hydroxyprogesterone and estradiol-17�levels during the human menstrual cycle. Am J Obstet Gynecol 111:947–951, 1971.
736. Thornton LM, Mazzeo SE, Bulik CM. The heritability of eatingdisorders: methods and current findings. Curr Top Behav Neurosci 6:141–156, 2011.
737. Tobias JH, Steer CD, Vilarino-Guell C, Brown MA. Effect of anestrogen receptor-alpha intron 4 polymorphism on fat mass in 11-year-old children. J Clin Endocrinol Metab 92: 2286–2291, 2007.
738. Tomelleri R, Grunewald KK. Menstrual cycle and food cravings inyoung college women. J Am Diet Assoc 87: 311–315, 1987.
739. Toth MJ, Tchernof A, Sites CK, Poehlman ET. Effect of menopausalstatus on body composition and abdominal fat distribution. Int J ObesRelat Metab Disord 24: 226–231, 2000.
740. Treasure J, Claudino AM, Zucker N. Eating disorders. Lancet 375:583–593, 2010.
741. Tremollieres FA, Pouilles JM, Ribot CA. Relative influence of age andmenopause on total and regional body composition changes in postmeno-pausal women. Am J Obstet Gynecol 175: 1594–1600, 1996.
742. Trevaskis JL, Turek VF, Wittmer C, Griffin PS, Wilson JK, Reyn-olds JM, Zhao Y, Mack CM, Parkes DG, Roth JD. Enhanced amylin-mediated body weight loss in estradiol-deficient diet-induced obese rats.Endocrinology 151: 5657–5668, 2010.
743. Trujillo ML, Spuch C, Carro E, Senaris R. Hyperphagia and centralmechanisms for leptin resistance during pregnancy. Endocrinology 152:1355–1365, 2011.
744. Tso P, Liu Apolipoprotein AIV M, food intake, obesity. Physiol Behav83: 631–643, 2004.
745. Tsutsui K, Ubuka T, Bentley GE, Kriegsfeld LJ. Gonadotropin-inhibitory hormone (GnIH): discovery, progress and prospect. Gen CompEndocrinol 177: 305–314, 2012.
746. Ubuka T, Lai H, Kitani M, Suzuuchi A, Pham V, Cadigan PA, WangA, Chowdhury VS, Tsutsui K, Bentley GE. Gonadotropin-inhibitoryhormone identification, cDNA cloning, and distribution in rhesus ma-caque brain. J Comp Neurol 517: 841–855, 2009.
747. Ubuka T, Son YL, Tobari Y, Tsutsui K. Gonadotropin-inhibitoryhormone action in the brain and pituitary. Front Endocrinol (Lausanne)3: 148, 2012.
748. Uher R, Treasure J, Heining M, Brammer MJ, Campbell IC. Cere-bral processing of food-related stimuli: effects of fasting and gender.Behav Brain Res 169: 111–119, 2006.
749. Uphouse L, Salamanca S,. and Caldarola-Pastuszka M. Gender andestrous cycle differences in the response to the 5-HT1A agonist 8-OH-DPAT. Pharmacol Biochem Behav 40: 901–906, 1991.
750. Valenstein ES, Kakolewski JW, Cox VC. Sex differences in tastepreference for glucose and saccharin solutions. Science 156: 942–943,1967.
751. Valle A, Catala-Niell A, Colom B, Garcia-Palmer FJ, Oliver J, RocaP. Sex-related differences in energy balance in response to caloricrestriction. Am J Physiol Endocrinol Metab 289: E15–E22, 2005.
752. Van den Eynde F, Koskina A, Syrad H, Guillaume S, Broadbent H,Campbell IC, Schmidt U. State and trait food craving in people withbulimic eating disorders. Eat Behav 13: 414–417, 2012.
753. Van den Eynde F, Treasure J. Neuroimaging in eating disorders andobesity: implications for research. Child Adolesc Psychiatr Clin N Am18: 95–115, 2009.
754. van der Laan LN, de Ridder DT, Viergever MA, Smeets PA. The firsttaste is always with the eyes: a meta-analysis on the neural correlates ofprocessing visual food cues. Neuroimage 55: 296–303, 2011.
755. Van Heek M, Compton DS, France CF, Tedesco RP, Fawzi AB,Graziano MP, Sybertz EJ, Strader CD, Davis HR Jr. Diet-inducedobese mice develop peripheral, but not central, resistance to leptin. J ClinInvest 99: 385–390, 1997.
756. Van Vugt DA. Brain imaging studies of appetite in the context of obesityand the menstrual cycle. Hum Reprod Update 16: 276–292, 2010.
757. Vanderhorst VG, Gustafsson JA, Ulfhake B. Estrogen receptor-alphaand -beta immunoreactive neurons in the brainstem and spinal cord ofmale and female mice: relationships to monoaminergic, cholinergic, andspinal projection systems. J Comp Neurol 488: 152–179, 2005.
758. Varma M, Chai JK, Meguid MM, Laviano A, Gleason JR, Yang ZJ,Blaha V. Effect of estradiol and progesterone on daily rhythm in foodintake and feeding patterns in Fischer rats. Physiol Behav 68: 99–107,1999.
759. Vasudevan N, Pfaff DW. Membrane-initiated actions of estrogens inneuroendocrinology: emerging principles. Endocr Rev 28: 1–19, 2007.
760. Verhagen JV, Giza BK, Scott TR. Effect of amiloride on gustatoryresponses in the ventroposteromedial nucleus of the thalamus in rats. JNeurophysiol 93: 157–166, 2005.
761. Versini A, Ramoz N, Le Strat Y, Scherag S, Ehrlich S, Boni C,Hinney A, Hebebrand J, Romo L, Guelfi JD, Gorwood P. Estrogenreceptor 1 gene (ESR1) is associated with restrictive anorexia nervosa.Neuropsychopharmacology 35: 1818–1825, 2010.
762. Viner RM, Cole TJ. Adult socioeconomic, educational, social, andpsychological outcomes of childhood obesity: a national birth cohortstudy. BMJ 330: 1354, 2005.
763. Voorhoeve PG, van Mechelen W, Uitterlinden AG, Delemarre-vande Waal HA, Lamberts SW. Estrogen receptor-alpha gene polymor-phisms and body composition in children and adolescents. Horm ResPaediatr 76: 86–92, 2011.
764. Wade GN. Gonadal hormones and behavioral regulation of body weight.Physiol Behav 8: 523–534, 1972.
765. Wade GN. Interaction between estradiol-17 beta and growth hormone incontrol of food intake in weanling rats. J Comp Physiol Psychol 86:359–362, 1974.
766. Wade GN. Some effects of ovarian hormones on food intake and bodyweight in female rats. J Comp Physiol Psychol 88: 183–193, 1975.
767. Wade GN, Gray JM. Gonadal effects on food intake and adiposity: ametabolic hypothesis. Physiol Behav 22: 583–593, 1979.
768. Wade GN, Gray JM, Bartness TJ. Gonadal influences on adiposity. IntJ Obes (Lond) 9 Suppl 1: 83–92, 1985.
769. Wade GN, Jones JE. Neuroendocrinology of nutritional infertility. AmJ Physiol Regul Integr Comp Physiol 287: R1277–R1296, 2004.
770. Wade GN, Schneider JE. Metabolic fuels and reproduction in femalemammals. Neurosci Biobehav Rev 16: 235–272, 1992.
771. Wade GN, Zucker I. Development of hormonal control over food intakeand body weight in female rats. J Comp Physiol Psychol 70: 213–220,1970.
772. Wade T, Martin NG, Neale MC, Tiggemann M, Treloar SA, BucholzKK, Madden PA, Heath AC. The structure of genetic and environmen-tal risk factors for three measures of disordered eating. Psychol Med 29:925–934, 1999.
773. Wade TD, Gordon S, Medland S, Bulik CM, Heath AC, MontgomeryGW, Martin NG. Genetic variants associated with disordered eating. IntJ Eat Disord, 2013.
Review
R1265SEX DIFFERENCES IN THE PHYSIOLOGY OF EATING
AJP-Regul Integr Comp Physiol • doi:10.1152/ajpregu.00446.2012 • www.ajpregu.org
by 10.220.32.247 on Septem
ber 18, 2016http://ajpregu.physiology.org/
Dow
nloaded from

774. Wager-Srdar SA, Gannon M, Levine AS. The effect of cholecysto-kinin on food intake in gonadectomized and intact rats: the influence ofsex hormones. Physiol Behav 40: 25–28, 1987.
775. Wager-Srdar SA, Levine AS. The effect of cholecystokinin-octapeptideon food intake and consummatory behavior in lactating rats. PhysiolBehav 50: 331–336, 1991.
776. Wallen WJ, Belanger MP, Wittnich C. Sex hormones and the selectiveestrogen receptor modulator tamoxifen modulate weekly body weightsand food intakes in adolescent and adult rats. J Nutr 131: 2351–2357,2001.
777. Walsh BT, Devlin MJ. Eating disorders: progress and problems. Science280: 1387–1390, 1998.
778. Wang GH. Age and sex differences in the amount of daily food-intakeof the albino rat. Am J Physiol 71: 729–773, 1925.
779. Wang GH. The changes in the amount of daily food-intake in the albinorat during pergnancy and lactation. Am J Physiol 71: 736–741, 1925.
780. Wang GJ, Geliebter A, Volkow ND, Telang FW, Logan J, Jayne MC,Galanti K, Selig PA, Han H, Zhu W, Wong CT, Fowler JS. Enhancedstriatal dopamine release during food stimulation in binge eating disor-der. Obesity (Silver Spring) 19: 1601–1608, 2011.
781. Wang K, Zhang H, Bloss CS, Duvvuri V, Kaye W, Schork NJ,Berrettini W, Hakonarson H. A genome-wide association study oncommon SNPs and rare CNVs in anorexia nervosa. Mol Psychiatry 16:949–959, 2011.
782. Wang YC, Bleich SN, Gortmaker SL. Increasing caloric contributionfrom sugar-sweetened beverages and 100% fruit juices among US chil-dren and adolescents, 1988–2004. Pediatrics 121: e1604–1614, 2008.
783. Wardle J, Haase AM, Steptoe A, Nillapun M, Jonwutiwes K, BellisleF. Gender differences in food choice: the contribution of health beliefsand dieting. Ann Behav Med 27: 107–116, 2004.
784. Watanabe K, Hara C, Ogawa N. Feeding conditions and estrous cycleof female rats under the activity-stress procedure from aspects of an-orexia nervosa. Physiol Behav 51: 827–832, 1992.
785. Weaver JU. Classical endocrine diseases causing obesity. Front HormRes 36: 212–228, 2008.
786. Weingarten HP, Elston D. The phenomenology of food cravings.Appetite 15: 231–246, 1990.
787. Welt CK, Chan JL, Bullen J, Murphy R, Smith P, DePaoli AM,Karalis A, Mantzoros CS. Recombinant human leptin in women withhypothalamic amenorrhea. N Engl J Med 351: 987–997, 2004.
788. Westenhoefer J. Age and gender dependent profile of food choice.Forum Nutr: 44–51, 2005.
789. Westerterp KR, Goris AH. Validity of the assessment of dietary intake:problems of misreporting. Curr Opin Clin Nutr Metab Care 5: 489–493,2002.
790. Whitaker KW, Totoki K, Reyes TM. Metabolic adaptations to earlylife protein restriction differ by offspring sex and post-weaning diet in themouse. Nutr Metab Cardiovasc Dis, 2011.
791. Williams DL. Minireview: finding the sweet spot: peripheral versuscentral glucagon-like peptide 1 action in feeding and glucose homeosta-sis. Endocrinology 150: 2997–3001, 2009.
792. Williams NI, Berga SL, Cameron JL. Synergism between psychosocialand metabolic stressors: impact on reproductive function in cynomolgusmonkeys. Am J Physiol Endocrinol Metab 293: E270–E276, 2007.
793. Williams WP, 3rd, Kriegsfeld LJ. Circadian control of neuroendocrinecircuits regulating female reproductive function. Front Endocrinol (Lau-sanne) 3: 60, 2012.
794. Williamson DA, Martin CK, York-Crowe E, Anton SD, RedmanLM, Han H, Ravussin E. Measurement of dietary restraint: validity testsof four questionnaires. Appetite 48: 183–192, 2007.
795. Windahl SH, Andersson N, Chagin AS, Martensson UE, Carlsten H,Olde B, Swanson C, Moverare-Skrtic S, Savendahl L, LagerquistMK, Leeb-Lundberg LM, Ohlsson C. The role of the G protein-coupled receptor GPR30 in the effects of estrogen in ovariectomizedmice. Am J Physiol Endocrinol Metab 296: E490–E496, 2009.
796. Witte MM, Resuehr D, Chandler AR, Mehle AK, Overton JM.Female mice and rats exhibit species-specific metabolic and behavioralresponses to ovariectomy. Gen Comp Endocrinol 166: 520–528, 2010.
797. Wizemann TM, Pardue ML. Exploring the biological contributions tohuman health: Does sex matter? Washington, DC: National AcademyPress, 2001.
798. Wolnerhanssen B, Beglinger C. Therapeutic potential of gut peptides.Forum Nutr 63: 54–63, 2010.
799. Woods SC. The control of food intake: behavioral versus molecularperspectives. Cell Metab 9: 489–498, 2009.
800. Woods SC, D’Alessio DA. Central control of body weight and appetite.J Clin Endocrinol Metab 93: S37–50, 2008.
801. Woods SC, Lutz TA, Geary N, Langhans W. Pancreatic signalscontrolling food intake; insulin, glucagon and amylin. Philos Trans R SocLond B Biol Sci 361: 1219–1235, 2006.
802. Woodside B. Prolactin and the hyperphagia of lactation. Physiol Behav91: 375–382, 2007.
803. Woodward CJ, Emery PW. Energy balance in rats given chronichormone treatment. 2. Effects of corticosterone. Br J Nutr 61: 445–452,1989.
804. Woolley CS, McEwen BS. Roles of estradiol and progesterone inregulation of hippocampal dendritic spine density during the estrouscycle in the rat. J Comp Neurol 336: 293–306, 1993.
805. Wright CL, Schwarz JS, Dean SL, McCarthy MM. Cellular mecha-nisms of estradiol-mediated sexual differentiation of the brain. TrendsEndocrinol Metab 21: 553–561, 2010.
806. Wu CL, Hung CR, Chang FY, Pau KY, Wang PS. Involvement ofcholecystokinin receptor in the inhibition of gastrointestinal motility byestradiol in ovariectomized rats. Scand J Gastroenterol 37: 1133–1139,2002.
807. Wu MV, Manoli DS, Fraser EJ, Coats JK, Tollkuhn J, Honda S,Harada N, Shah NM. Estrogen masculinizes neural pathways andsex-specific behaviors. Cell 139: 61–72, 2009.
808. Wurtman JJ, Baum MJ. Estrogen reduces total food and carbohydrateintake, but not protein intake, in female rats. Physiol Behav 24: 823–827,1980.
809. Wurtman JJ, Brzezinski A, Wurtman RJ, Laferrere B. Effect ofnutrient intake on premenstrual depression. Am J Obstet Gynecol 161:1228–1234, 1989.
810. Xiao E, Kim AJ, Dutia R, Conwell I, Ferin M, Wardlaw SL.Effects of estradiol on cerebrospinal fluid levels of agouti-relatedprotein in ovariectomized rhesus monkeys. Endocrinology 151: 1002–1009, 2010.
811. Xu J, Kirigiti MA, Grove KL, Smith MS. Regulation of food intakeand gonadotropin-releasing hormone/luteinizing hormone during lac-tation: role of insulin and leptin. Endocrinology 150: 4231–4240,2009.
812. Xu Y, Nedungadi TP, Zhu L, Sobhani N, Irani BG, Davis KE,Zhang X, Zou F, Gent LM, Hahner LD, Khan SA, Elias CF,Elmquist JK, Clegg DJ. Distinct hypothalamic neurons mediateestrogenic effects on energy homeostasis and reproduction. CellMetab 14: 453–465, 2011.
813. Yanovski S. Sugar and fat: cravings and aversions. J Nutr 133: 835S–837S, 2003.
814. Yepuru M, Eswaraka J, Kearbey JD, Barrett CM, Raghow S,Veverka KA, Miller DD, Dalton JT, Narayanan R. Estrogen receptor-�-selective ligands alleviate high-fat diet- and ovariectomy-inducedobesity in mice. J Biol Chem 285: 31292–31303, 2010.
815. Young JK. Anorexia nervosa and estrogen: current status of the hypoth-esis. Neurosci Biobehav Rev 34: 1195–1200, 2010.
816. Young JK. Estrogen and the etiology of anorexia nervosa. NeurosciBiobehav Rev 15: 327–331, 1991.
817. Yu Z, Geary N, Corwin RL. Individual effects of estradiol and proges-terone on food intake and body weight in ovariectomized binge rats.Physiol Behav 104: 687–693, 2011.
818. Yu Z, Geary N, Corwin RL. Ovarian hormones inhibit fat intake underbinge-type conditions in ovariectomized rats. Physiol Behav 95: 501–507, 2008.
819. Yuan DL, Chambers KC. Estradiol accelerates extinction of a condi-tioned taste aversion in female and male rats. Horm Behav 36: 1–16,1999.
820. Zandian M, Ioakimidis I, Bergh C, Leon M, Sodersten P. A sexdifference in the response to fasting. Physiol Behav 103: 530–534, 2011.
821. Zeleznik AJ, Pohl CR. Control of follicular development, corpusluteum function, the maternal recognition of pregnancy, and theneuroendocrine regulation of the menstrual cycle in higher primates.In: Knobil and Neill’s Physiology of Reproduction (3rd ed.), edited byNeill JD. Amsterdam, The Netherlands: Elsevier/Academic, 2006, p.2449 –2510.
Review
R1266 SEX DIFFERENCES IN THE PHYSIOLOGY OF EATING
AJP-Regul Integr Comp Physiol • doi:10.1152/ajpregu.00446.2012 • www.ajpregu.org
by 10.220.32.247 on Septem
ber 18, 2016http://ajpregu.physiology.org/
Dow
nloaded from

822. Zellner DA, Garriga-Trillo A, Centeno S, Wadsworth E. Chocolatecraving and the menstrual cycle. Appetite 42: 119–121, 2004.
823. Zellner DA, Garriga-Trillo A, Rohm E, Centeno S, Parker S. Foodliking and craving: A cross-cultural approach. Appetite 33: 61–70, 1999.
824. Zhang J, Matheny MK, Tumer N, Mitchell MK, Scarpace PJ. Leptinantagonist reveals that the normalization of caloric intake and the thermic
effect of food after high-fat feeding are leptin dependent. Am J PhysiolRegul Integr Comp Physiol 292: R868–R874, 2007.
825. Zucker I. Progesterone in the experimental control of the behaviouralsex cycle in the female rat. J Endocrinol 38: 269–277, 1967.
826. Zucker I, Beery AK. Males still dominate animal studies. Nature 465:690, 2010.
Review
R1267SEX DIFFERENCES IN THE PHYSIOLOGY OF EATING
AJP-Regul Integr Comp Physiol • doi:10.1152/ajpregu.00446.2012 • www.ajpregu.org
by 10.220.32.247 on Septem
ber 18, 2016http://ajpregu.physiology.org/
Dow
nloaded from
Related Documents











