Characterization of novel bacteriophagephiC119 capable of lysing multidrug-resistant Shiga toxin-producingEscherichia coli O157:H7
Luis Amarillas1,2, Cristobal Chaidez3, Arturo Gonzalez-Robles4,Yadira Lugo-Melchor5 and Josefina Leon-Felix1
1 Laboratorio de Biologıa Molecular y Genomica Funcional, Centro de Investigacion en
Alimentacion y Desarrollo, A. C., Culiacan, Sinaloa, Mexico2 Laboratorio de Genetica, Instituto de Investigacion Lightbourn, A. C., Cd. Jimenez, Chihuahua,
Mexico3 Inocuidad Alimentaria, Centro de Investigacion en Alimentacion y Desarrollo, A. C., Culiacan,
Sinaloa, Mexico4 Departamento de Infectomica y Patogenesis Molecular, Centro de Investigacion y de Estudios
Avanzados, Instituto Politecnico Nacional, Ciudad de Mexico, Mexico5 Laboratorio de Biologıa Molecular de la Unidad de Servicios Analıticos y Metrologicos, Centro
de Investigacion y Asistencia en Tecnologıa y Diseno del Estado de Jalisco A. C., Guadalajara,
Jalisco, Mexico
ABSTRACTBackground: Shiga toxin-producing Escherichia coli (STEC) is one of the most
common and widely distributed foodborne pathogens that has been frequently
implicated in gastrointestinal and urinary tract infections. Moreover, high rates
of multiple antibiotic-resistant E. coli strains have been reported worldwide. Due
to the emergence of antibiotic-resistant strains, bacteriophages are considered an
attractive alternative to biocontrol pathogenic bacteria. Characterization is a
preliminary step towards designing a phage for biocontrol.
Methods: In this study, we describe the characterization of a bacteriophage
designated phiC119, which can infect and lyse several multidrug-resistant STEC
strains and some Salmonella strains. The phage genome was screened to detect
the stx-genes using PCR, morphological analysis, host range was determined, and
genome sequencing were carried out, as well as an analysis of the cohesive ends
and identification of the type of genetic material through enzymatic digestion of
the genome.
Results: Analysis of the bacteriophage particles by transmission electron microscopy
showed that it had an icosahedral head and a long tail, characteristic of the family
Siphoviridae. The phage exhibits broad host range against multidrug-resistant and
highly virulent E. coli isolates. One-step growth experiments revealed that the
phiC119 phage presented a large burst size (210 PFU/cell) and a latent period of
20 min. Based on genomic analysis, the phage contains a linear double-stranded
DNA genome with a size of 47,319 bp. The phage encodes 75 putative proteins,
but lysogeny and virulence genes were not found in the phiC119 genome.
Conclusion: These results suggest that phage phiC119 may be a good biological
control agent. However, further studies are required to ensure its control of STEC
and to confirm the safety of phage use.
How to cite this article Amarillas et al. (2016), Characterization of novel bacteriophage phiC119 capable of lysing multidrug-resistant
Shiga toxin-producing Escherichia coli O157:H7. PeerJ 4:e2423; DOI 10.7717/peerj.2423
Submitted 15 April 2016Accepted 9 August 2016Published 13 September 2016
Corresponding authorJosefina Leon-Felix,
Academic editorBlanca Landa
Additional Information andDeclarations can be found onpage 17
DOI 10.7717/peerj.2423
Copyright2016 Amarillas et al.
Distributed underCreative Commons CC-BY 4.0

Subjects Microbiology, Molecular Biology, Virology
Keywords Shiga toxin, Phage phiC119, Genome analysis, Siphoviridae, Biocontrol applications,
Phage group relationships
INTRODUCTIONEscherichia coli is an innocuous commensal of the gastrointestinal tract; however,
pathogenic E. coli, including Shiga toxin-producing E. coli (STEC), particularly serotype
O157:H7, has been identified as one of the major pathogens causing foodborne diseases
(Farfan & Torres, 2012). The Centers for Disease Control and Prevention (Centers for
Disease Control Prevention, 2015) estimate that approximately 265,000 illnesses and
approximately 4,000 hospitalizations in the United States occur every year due to
infections caused by STEC; in developing countries, the situation is often much worse.
Northwestern Mexico is a region that is heavily involved in the production and
commercialization of agricultural exports to the US and other countries. Recently, several
resistant STEC O157:H7 strains have been isolated from domestic animals on rural farms
in this region. The commonality between these strains was multidrug resistance and
virulence-encoding genes (Amezquita-Lopez et al., 2012; Canizalez-Roman et al., 2013),
which may have potential health risks to humans in the region (Belanger et al., 2011), as it
has been widely documented that several E. coli outbreaks worldwide had a zoonotic
origin (Jakobsen et al., 2012; Pierard et al., 2012).
Furthermore, antibiotic treatment is contraindicated for STEC infection due to
potential worsening of the infection, and alternatives are therefore needed. Implementing
strategies to control pathogenic E. coli and other foodborne pathogens is a critical step to
strengthen food safety in the region. In this regard, among the potential antimicrobial
agents, bacteriophages (also called phages) are promising and sustainable agents that
can be used against pathogenic bacteria (Mahony et al., 2011; Guenther et al., 2012;
Hungaro et al., 2013).
In recent years, interest in the concept of bacteriophages as biocontrol agents has
significantly increased. Bacteriophages are viruses that infect bacteria and cause bacterial
lysis and are thus considered biocontrol agents for pathogenic bacteria. Desirable
candidate phages used for biocontrol should be strictly lytic because they always cause
bacterial lysis and release progeny virions (Hagens & Loessner, 2010). Moreover, virulent
phages must not integrate their DNA into the host DNA and should display a minimal
transduction frequency (negligible rates of transduction); therefore, non-integrating
bacteriophages will be the most effective as biocontrol agents. Phages potentially used
for biocontrol should be capable of infecting many strains (broad host range) (Chan,
Abedon & Loc-Carrillo, 2013; Akhtar, Viazis & Diez-Gonzalez, 2014).
For safety reasons, candidates for biocontrol should not have genes encoding
pathogenicity or allergy-triggering proteins. For example, Shiga toxins (Stxs) are encoded
in the genome of some bacteriophages, and the genetic information encoding Stxs can be
integrated into the host chromosome (Yan et al., 2011). This type of bacteriophage should
be discarded for the purposes of biocontrol because it is possible that the phage could
transfer genetic material to the host bacteria. Therefore, a detailed characterization of the
Amarillas et al. (2016), PeerJ, DOI 10.7717/peerj.2423 2/22

bacteriophages is required to provide useful information to determine their potential
as biocontrol agents.
Lysogeny-associated, virulence-related and/or antibiotic-resistance genes should be
absent in the genome of the bacteriophage, making genome sequencing essential for
assessing the safety of a phage (Jun et al., 2015).
Phages have been used by many researchers to biocontrol E. coli and other types of
bacteria. In all cases, none of the phages reported have been able to lyse all strains.
Therefore, it is very important to continue isolating and characterizing novel
bacteriophages with broad host ranges against drug-resistant E. coli strains prevalent
in a given region, which may involve local phage isolation.
In this regard, the new bacteriophage phiC119 isolated in northwestern Mexico
(Castro del Campo et al., 2011), exhibited strong in vitro lytic activity against STEC strains,
indicating that it could be a candidate biological control agent. However, information on
this phage is limited. Therefore, to extend our understanding of the phage characteristics,
we describe in this study the characterization of phiC119, providing data that are
critical in determining whether it can potentially be used as a biological control agent.
MATERIALS AND METHODSBacteriophage, bacterial strain and culture conditionsBacteriophage phiC119 was previously isolated from horse feces in Sinaloa, Mexico with
an enrichment technique. The bacteriophage was isolated from horse feces collected
from five different farms located in the region of located in Northwestern Mexico.
Briefly, 5 g of horse feces was diluted 1:10 in sterile distilled water (pH 7.0) and gently
mixed by inversion. The mixture was cleared by low-speed centrifugation at 6,500 g
for 20 min and filtered through a cellulose acetate syringe filter (0.45 mm pore size,
GVS filter technology, USA). The 1 mL of filtered supernatant was then mixed with
20 mL exponential phase bacterial culture, and incubated at 37 �C for 18–24 h. After
incubation, the bacterial cells were centrifuged and the supernatant was filtered through
a 0.22 mm pore size cellulose acetate syringe filter (GVS filter technology, IN, USA).
Then, 100 ml of filtrate and 1 mL of the host strain were mixed with soft agar and poured
onto an TSA agar plate. After 24 h incubation at 37 �C, plates were checked for a clear
zone of bacterial lysis. Single plaques were picked with a sterile glass Pasteur pipette and
suspended in 1 mL of sterile distilled water, and each individual plaque was re-isolated
three times to ensure the purity of the phage isolate. The phage was stored at -20 �C in
tryptic soy broth (TSB, Bioxon, Mexico) containing 30% (v/v) glycerol for further
characterization. E. coli O157 EC-48 (63-Fv18-1) was previously isolated from fecal
samples from domestic animals collected from farms located in the Culiacan Valley and
was used as the host for phage propagation in this study. Bacterial strains and phage
stocks were obtained from the culture collection maintained by the Food Safety National
Research Laboratory (LANIIA) at the Research Center in Food & Development (CIAD),
Culiacan station. E. coli was grown on TSB at 37 �C; the overnight culture was usedin the assays described below.
Amarillas et al. (2016), PeerJ, DOI 10.7717/peerj.2423 3/22

Host rangeThe host range of phage phiC119 was determined with a spotting assay using strains
previously described as pathogenic in mammalian cells (Amezquita-Lopez et al., 2014).
Additionally, 44 environmental Salmonella strains were also included in the study
(Jimenez et al., 2014; Estrada-Acosta et al., 2014) (Table 1). On the surface of TSA plates
(TSA media with 1.2% agar), 1 mL of overnight culture of each strain and 3 mL of soft
agar (TSA media with 0.4% agar) were poured and allowed to solidify. Then, a 10 mL
aliquot of several phage dilutions were spotted onto each bacterial overlay and incubated
at 37 �C for 18–24 h. After incubation, the presence of phage lysis zones was evaluated
in the drops. All testing was performed in triplicate. Bacterial strains used for the
bacteriophage host-range investigation were obtained from the LANIIA at the CIAD.
One-step growth curveE. coli O157 EC-48 was inoculated into 40 mLTSB broth medium and incubated at 37 �Cwith shaking to reach an OD600 of 0.5. The phage and host cells were mixed with a MOI
of 0.01 and allowed to adsorb for 2 min at room temperature. After incubation, the
mixture was harvested by centrifugation at 10,000 � g for 1 min at 4 �C. Subsequently,the supernatant was discarded to remove the free phages. The pellet containing infected
host cells was gently re-suspended in equal volume of pre-warmed TSB and shake culture
at 37 �C. Samples were taken at 5 min intervals (up to 60 min), and phage titer was
calculated by double agar plates. The experiment was carried out in triplicated to estimate
burst size and latency.
Bacteriophage propagation and DNA extractionBacteriophage propagation was performed using the double-layer plaque technique
described by Carey-Smith et al. (2006). Briefly, 100 mL of phage stock was mixed with
1 mL of overnight cultured E. coli (CECT 4076) and 2.8 mL of TSB agar (0.4%) preheated
to 50 �C. The mixture was poured onto tryptic soy agar (TSA, Bioxon, Mexico) plates
(100� 15 mm Petri dishes) and incubated for 18–24 h at 37 �C under aerobic conditions.
Six milliliters of sterile SM buffer (100 mm NaCl, 25 mm Tris-HCl (pH = 7.5), 8 mm
MgSO4 and 0.01% (w/v) gelatin) was added to the surface of each plate, and the top agar
was recovered using a sterile loop. Then, the eluate was centrifuged at 4,500� g for 10 min
at 4 �C, and the supernatant was recovered; the procedure was repeated twice. The final
pooled supernatant was filtered through a cellulose acetate syringe filter with a 0.45 mm
pore size (GVS filter technology, IN, USA). The phage filtrate was concentrated by
centrifugation at 40,000 � g for 2 h, and then the pellet was gently resuspended by
pipetting in 10 mL of SM buffer and filtered using a cellulose acetate syringe filter with a
0.20 mm pore size. The bacteriophage titer was determined by a double-layer plaque
technique with serial decimal dilutions of phage concentrate. The final purified phages
were stored at 4 �C.One milliliter of purified phage suspension (approximately 1 � 1012 plaque forming
units (PFU) per mL) was incubated with 10 mL of DNase I/RNase A (10 mg/mL) (Sigma-
Aldrich, MO, USA) for 1 h at 37 �C. Phage DNA was extracted using SDS-proteinase
Amarillas et al. (2016), PeerJ, DOI 10.7717/peerj.2423 4/22

Table 1 Bacterial strains used in the host range spectrum of the bacteriophage phiC119. Phage was
assessed for host range by spot testing.
Bacterial Strain Bacterial lysis
E. coli O157:H7 HC14-1 +
E. coli O157:H7 HE7-1 +
E. coli O157:H7 HC14-2 +
E. coli O157:H7 AC6-1 +
E. coli O157:H7 HE10-1 -E. coli O157:H7 AR7-2 -E. coli O157:H7 AR17-2 -E. coli O157:H7 AC6-1 -E. coli O157:H7 AR15-1 -E. coli O157:H7 AR17-1 -E. coli O157:H7 RM8744 +
E. coli O157:H7 RM8753 +
E. coli O157:H7 RM8754 +
E. coli O157:H7 RM8759 +
E. coli O157:H7 RM8767 +
E. coli O157:H7 RM8768 +
E. coli O157:H7 RM8769 +
E. coli O157:H7 RM8781 +
E. coli O157:H7 RM8920 +
E. coli O157:H7 RM8921 +
E. coli O157:H7 RM8922 +
E. coli O157:H7 RM8927 +
E. coli O157:H7 RM8928 -E. coli O157:H7 RM9450 +
E. coli O157:H7 RM9451 +
E. coli O157:H7 RM9452 +
E. coli O157:H7 RM9453 +
E. coli O157:H7 RM9455 +
E. coli O157:H7 RM9457 +
E. coli O157:H7 RM9458 +
E. coli O157:H7 RM9459 +
E. coli O157:H7 RM9462 -E. coli O157:H7 RM9463 +
Salmonella Weltevreden AC2-039 -Salmonella Oranienburg AC2-041 -Salmonella Saintpaul AC2-046 -Salmonella Minnesota AC2-070 +
Salmonella Anatum AC2-079 -Salmonella Oranienburg AC2-100 -Salmonella Montevideo CM-02 -Salmonella Saintpaul AC2-137 -
(Continued)
Amarillas et al. (2016), PeerJ, DOI 10.7717/peerj.2423 5/22

K method as previously described (Sambrook & Russell, 2001). Phage DNA was stored at
4 �C until use. The nucleic acid extract was subjected to digestion with DNase I and RNase
according to the manufacturer’s instructions.
Table 1 (continued).
Bacterial Strain Bacterial lysis
Salmonella Oranienburg AC2-142 -Salmonella Luciana AC2-240 +
Salmonella Anatum CM-50 -Salmonella Minnesota CM-51 -Salmonella Montevideo CM-52 -Salmonella Agona AC2-346 -Salmonella Muenster CM-08 -Salmonella Muenster AC2-366 -Salmonella Montevideo AC2-370 -Salmonella Weltevreden CM-08 -Salmonella Poona CM-18 -Salmonella Oranienburg CM-21 -Salmonella Saintpaul CM-25 -Salmonella Give CM-31 -Salmonella Saintpaul AC2-098 -Salmonella Oranienburg AC2-026 +
Salmonella Pomona AC2-248 -Salmonella Oranienburg HC2-2 -Salmonella Oranienburg HC2-1 -Salmonella Oranienburg HC2-3 -Salmonella Give HB4-2 -Salmonella Saintpaul HE4-1 -Salmonella Give HB4-1 -Salmonella Give HB4-1 -Salmonella Weltevreden HD4-2 -Salmonella Give HB4-3 -Salmonella Saintpaul HE4-3 -Salmonella Weltevreden HD4-3 -Salmonella Agona HD5-1 +
Salmonella Give HD6-3 -Salmonella Oranienburg HD5-2 -Salmonella Oranienburg HE6-1 -Salmonella Sandiego HF6-3 -Salmonella Montevideo S-188 -Salmonella Oranienburg S-190 +
Salmonella Oranienburg S-228 -Notes:
+, indicate positive sensitivity to phage lysis.-, indicate negative sensitivity to phage lysis.
Amarillas et al. (2016), PeerJ, DOI 10.7717/peerj.2423 6/22

Transmission electron microscopy and plaque characteristicsThirty microliters of purified phage suspension was adsorbed to carbon-coated copper
grids (400-mesh) in a vacuum evaporator (JEE400, JEOL Ltd. Tokyo, Japan), allowed
to air dry and then negatively stained with 2% phosphotungstic acid (pH 7.2). The
excess solution was absorbed with filter paper, and samples were observed with a
transmission electron microscope (JEM-1011, JEOL Ltd. Tokyo, Japan) operating at
80 kV (Lopez-Cuevas et al., 2011).
Bacteriophage plaques formed on a TSA plate during the process of propagation
(using dilutions that generated 15–30 plaques per plate) were analyzed according to
the procedure described by Gallet, Kannoly & Wang (2011) with minor modifications.
Briefly, images of ten plates were captured by a supersensitive high-resolution 16-bit
camera that was deeply cooled for faint image detection (Bio-Rad Laboratories), and the
image of five plaques for each plate were displayed with the ImageJ software (developed
at the National Institutes of Health, Bethesda, Maryland). The plates were then incubated
for 18–24 h at 37 �C before plaque size determination. To calculate the surface area
(expressed in square millimeters) corresponding to each pixel, a graticule of 1 mm2 was
used as the reference scale for the simplified measurement of the lysis plaques. According
to the analysis, each pixel corresponded to 0.5 mm2.
PCR to identify stx1 and stx2 encoding bacteriophageMultiplex PCR using a GoTaq� PCR Core System I (Promega, WI, USA) was performed
to determine the presence of the stx1 and stx2 genes in the genome of phage phiC119. PCR
assays were performed using the protocol previously described by Paton & Paton (1998).
In addition, E. coli O157:H7 (CECT 4076) DNA was included in the PCR screen as a
positive control. All primers used in the PCR assays were commercially synthesized
by Sigma–Aldrich (Toluca, Mexico).
Genome size estimation and analysis of the cohesive endsThe genome ends were determined as described by Casjens & Gilcrease (2009). Briefly,
1 mg of phage genetic material was digested with the restriction enzyme EcoRV
according to the manufacturer’s specifications, followed by heating for 15 min at 75 �C.Subsequently, the reaction mixture was divided into two equal parts. One was rapidly
cooled by immersion into an ice-water bath for 10 min, and the other was cooled to
room temperature prior to electrophoresis on a 1% agarose gel at a voltage of 75 V for
90 min. They were then stained with ethidium bromide (1 mL mL-1), and images were
captured using a ChemiDocTM MP imaging system with Image LabTM software (Bio-Rad
Laboratories). The lambda phage DNA was used as a positive control. Lambda DNA
digested with the HindIII endonuclease was used as a standard molecular weight marker
(Promega, WI, USA).
Genome sequencing and annotationDNA sequencing was performed at the National Laboratory of Genomics for Biodiversity
(LANGEBIO) using theMiSeq sequencing system (Illumina, Inc.) (150-bp single-end reads).
Amarillas et al. (2016), PeerJ, DOI 10.7717/peerj.2423 7/22

In total, 4,832,127 reads were generated and assembled into one contig using
Geneious v8.1.2 (the final sequence coverage was approximately 50�). The sequence
assembly was validated by a comparative restriction profile (Promega, WI, USA).
Potential open reading frames (ORFs) longer than 100 bp were predicted by GeneMark
(http://exon.gatech.edu/) and ORF Finder (http://www.ncbi.nlm.nih.gov/gorf/gorf.
html). The putative ORFs were analyzed by BLAST at the National Center for
Biotechnology Information (NCBI) (http://blast.ncbi.nlm.nih.gov/Blast.cgi) against
the database of non-redundant protein sequences using a significant E-value of 10-3.
Moreover, all identified ORFs were compared against the virulence factor database
(http://www.mgc.ac.cn/VFs/) (Chen et al., 2012) and the ResFinder database (http://cge.
cbs.dtu.dk/services/ResFinder/) (Kleinheinz, Joensen & Larsen, 2014). The predicted
phage protein sequences were searched to identify proteins that were potentially
allergenic using tools available at http://www.allergenonline.com from the Food Allergy
Research. This analysis was complemented with a search for conserved protein domains
using InterProScan, HMMER, Prosite, Motif Search and SMART. Hypothetical
isoelectric points and the molecular weights of putative proteins were predicted using
the ExPASy server (http://us.expasy.org/tools/protparam.html). Potential tRNA genes
in the genome sequence were predicted using tRNAscan-SE and ARAGORN. Promoters
and potential rho-independent terminators were identified using the Neural Network
Promoter Prediction tool of the Berkeley Drosophila Genome Project (http://www.
fruitfly.org/seq_tools/promoter.html) and the FindTerm program (http://linux1.
softberry.com/berry.phtml?topic=findterm&group=programs&subgroup=gfinb)
(energy threshold value: -11), respectively. The nucleotide genome sequence of
phage phiC119 has been deposited in the GenBank database under accession
number KT825490.
The lifestyle of the phages was predicted using the PHACTS program (http://www.
phantome.org/PHACTS/upload.php). Statistical analysis was performed using Minitab
statistical software version 14 (Minitab Inc., State College, PA, USA). Hierarchical
clustering analysis was used to determine the relationship between genome size, gene
density, and lifestyle.
Furthermore, the amino acid sequences of terminase large subunits of phiC119 and
others phages were obtained from GenBank. Twelve bacteriophages, including the
phiC119, were selected for phylogenetic analysis, these phages were selected as being the
most well-known representatives of each important family of phages. The amino-acid-
sequences were aligned using the program ClustalW, and the neighbor-joining
phylogenetic tree was generated using Geneious v8.1.2.
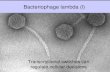
RESULTSBacteriophage, bacterial strain and culture conditionsElectron microscopic analysis revealed that phage phiC119 was non-enveloped with an
icosahedral capsid of approximately 43–45 nm in diameter and a tail of 168–172 nm
in length and 7–9 nm in width. These characteristics suggest that phage phiC119 is a
member of the Siphoviridae family. The flexibility and the uniformity of the tail
Amarillas et al. (2016), PeerJ, DOI 10.7717/peerj.2423 8/22

lengths indicated that it was non-contractile (Fig. 1). Phage phiC119 produced very
large (1.0–1.5 mm in diameter), clear and uniform-sized plaques after 18–24 h incubation
at 37 �C with E. coli O157:H7 EC-48 (63-104 Fv18-1) using the double-agar overlay
technique.
Host rangeThe bacteriophage phiC119 was recently isolated by our lab from horse feces and
to determine the susceptibility of bacterial strains to lysis by phage, thirty-three
environmental isolates of E. coli, previously isolated at the CIAD, were used for determine
the host range of phage phiC119 (Table 1). A high proportion (75.75%, n = 25) of E. coli
strains were sensitive to phage phiC119, which formed plaqueson a broad spectrum of
E. coli serogroups O157, including Stx-producing E. coli. These E. coli isolates were
previously characterized as highly virulent because they exhibit toxicity against
mammalian cells and have high levels of antibiotic resistance (Amezquita-Lopez et al.,
2014; Amezquita-Lopez et al., 2016).
Additionally, we determined the host range of the phage phiC119 with a collection
of 44 Salmonella strains. Interestingly, the phage was also able to infect only some strains
of certain Salmonella serotypes (Oranienburg, Agona, Luciana, andMinnesota). However,
the phage was not able to lyse the other bacterial species used in this study.
One-step growth curveOne-step growth curve experiment was performed to determine the latent time
period and burst size of the phage, as these are two of the most important
characteristics of phage infection process. According to the results obtained, the entire
phiC119 life cycle takes about 60 min to complete. phiC119 had approximately
20 min of latent period and the average burst size is 210 phage particles per infected
cell after 55 min at 37 �C (Fig. 2).
Figure 1 Transmission electron micrograph of phage phiC119 negatively stained with 2% unanyl
acetate. Phage phiC119 showing typical Siphoviridae morphology, which exhibit a noncontractile tail
with a length of 168–172 nm. The icosahedral head of phiC119 has a length of 43–45 nm and a width of
7–9 nm. The bar indicates 100 nm.
Amarillas et al. (2016), PeerJ, DOI 10.7717/peerj.2423 9/22

Detection of the stx genesThe phage was tested for the presence of the stx1 and stx2 genes (Fig. 3). PCR screening
for the stx genes using DNA isolated from bacteriophage phiC119 was negative. However,
other virulence factors may be encoded in the bacteriophage genome, and therefore,
genome sequencing and in silico analyses are required to ensure the absence of virulence,
antibiotic resistance or lysogenic genes because lysogenic conversion can increase
the pathogenic potential of the bacteria towards their hosts. Hence, bacteriophages
suitable for biocontrol purposes should not encode virulence genes or potential
immunoreactive allergens.
Analysis of the cohesive endsThe nucleic acid of phage phiC119 was resistant to RNase, sensitive to DNase and digested
by restriction enzymes. These results indicate that the phage genome is double-stranded
DNA and is approximately 47 kb in size (genome size estimated from the digested
fragments). Moreover, enzymatic digestion of the genome suggested that phage phiC119
utilizes the pac-mechanism of DNA packaging because heating/cooling of DNA after
enzymatic digestion did not alter the restriction patterns (Casjens & Gilcrease, 2009)
(Fig. 4). There, was no evidence for the existence of cohesive ends in the bacteriophage
genome. In addition, the analysis revealed a close a phylogenetic relationship between the
phagephiC119 and other pac-type phages (Fig. 5).
Bacteriophage genome featuresOverall, the bacteriophage genome contained 75 putative ORFs (90.4% of the genome
consists of a coding region) (Fig. 6), 21 of which are transcribed from the complementary
Figure 2 One-step growth curve of phage phiC119 on E. coli at 37 �C. The parameters of phage growth
are indicated in the figure, showing the latent period (20 min) and the average burst size (210 viral
particles per host cell). Means ± standard error from three independent experiments are shown. Some of
the error bars were too small to be visible.
Amarillas et al. (2016), PeerJ, DOI 10.7717/peerj.2423 10/22

strand. Based on sequence similarities and protein domains/motifs and BLAST
searches, 42 genes were assigned to conserved sequences and 33 were sorted into known
functional categories. Furthermore, bioinformatics analysis revealed an organization of the
phage genome into four functional modules, coding for structural proteins, DNA packaging,
replication and host lysis (a detailed description of gene functions is shown in Table S1).
The genome sequence of phiC119 consisted of 47,319 bp with an average GC content
of 44.20%, which is significantly lower than that of E. coli (average 50%). Furthermore,
a tRNA gene was identified (Arg-tRNA (anti-codon CCT)) between positions
42,465–42,540 in an adjacent region to the morphogenetic cluster, indicating probable
involvement in phage morphogenesis.
Bacteriophage phiC119 genomepossesses a high gene density (1.60 genes per kilobase), it
contains a large proportion of genes that overlap with coding regions of neighboring genes.
Similarly, different authors have indicated that the genes of coliphages (bacteriophages
that infect E. coli hosts) are usually tightly packed together with small intergenic regions
and a high gene density (Miller et al., 2003; Santos & Bicalho, 2011). Moreover, the genome
of phage phiC119 contains several overlapping sets of genes; 20 ORFs overlap with an
adjacent ORF, thus generating an increase in the density of genetic information.
Genomic analysis showed that phage phiC119 does not have lysogenic genes, such as
integrase and repressor genes. In addition, lifestyle prediction using the PHACT program
Figure 3 Agarose gel electrophoresis of PCR products amplified from DNA extracted from phage
phiC119. PCR was performed to detect the presence of stx1 and stx2 genes in phage genome. The
size of stx1 and stx2 amplicon corresponds to the 180 and 255 bp band, respectively. Lane M; 100 bp
DNA ladder (Promega), Lane 1; negative control, Lane 2; Bacteriophage phiC119 sample, Lane 3;
positive control.
Amarillas et al. (2016), PeerJ, DOI 10.7717/peerj.2423 11/22

suggested that phiC119 is a virulent bacteriophage. Furthermore, the bioinformatics
analysis of the phiC119 phage did not find any undesired genes in its genome, indicating
the lack of known genes coding for potential allergens and virulence genes. Therefore,
bacteriophage phiC119 has two of the desirable features of candidate phages used for
biocontrol.
Morphology moduleGenomic analyses revealed that at least 18 ORFs are involved in the morphogenesis of
bacteriophage phiC119. The products of putative ORFs 1 and 75 shared identity with
prohead proteases, suggesting that these ORFs are necessary for capsid morphogenesis.
Moreover, phage phiC119 possesses a potential major capsid protein encoded by ORF 4.
The tail proteins were identified as ORFs 10, 11, 14, 15, 17, 18, and 19. Additionally, ORFs
encoding tailspike and two tail fiber proteins were found. According to Yamashita et al.
(2011), these structures are required for specific recognition and binding to the host
receptor and were identified as ORF 16 and ORF 20 and 29, respectively. Phage phiC119
encodes two tape measure chaperone proteins (ORFs 12 and 13).
Nucleotide metabolism moduleWe also identified ORFs involved in nucleotide metabolism including ORF 3, which
encodes DNA polymerase I, an enzyme used during DNA replication of the
bacteriophage. The product of ORF 23 encodes a regulatory protein, which is an essential
enzyme for DNA transcription.
Figure 4 Endonuclease digestion analysis of phage phiC119 genomic DNA. Phage genomic DNAwas
digested with the restriction enzyme EcoRV. The digested DNA fragments were separated by 1% agarose
gel electrophoresis. HindIII-digested lambda DNA was used as a positive control to detect annealing of
cohesive ends (Lane 1 and 2) and phiC119 DNA digested with EcoRV (Lane 3 and 4). After digestion,
lines 1 and 3 were rapid cooling by immersion into an ice-water bath for 10 min, and the lines 2 and 4
cooled to room temperature. The arrows indicate fragments that bind to be cohesive in positive control.
Amarillas et al. (2016), PeerJ, DOI 10.7717/peerj.2423 12/22

ORFs 25, 26 and 27 encode, respectively, exodeoxyribonuclease VIII, a recombination
protein and an endonuclease. Assays performed by Pickard et al. (2008) have shown that
these proteins are essential for proper DNA packaging, and therefore, these proteins may
have comparable roles in phage phiC119.
The putative ORF 28 encodes a protein with conserved motifs associated with a single-
stranded DNA binding protein. Single-stranded DNA-binding proteins promote the
integration of components of the DNA replication complex (Hollis et al., 2001). This
protein is likely essential for DNA replication of phage phiC119. Phage phiC119 contains
an alpha replication protein, a putative transcriptional regulator and an ATP-dependent
helicase (ORFs 30, 31 and 32), which are all proteins involved in DNA replication
(Hua et al., 2014). ORF 41 is closely related to an HNH endonuclease that participates
in phage DNA repair (Moodley, Maxwell & Kanelis, 2012).
Figure 5 Phylogenetic analysis of the terminase large subunits of phage phiC119 and other large
terminase genes from diverse phage genomes. Numbers on the branches are bootstrap values.
Amarillas et al. (2016), PeerJ, DOI 10.7717/peerj.2423 13/22

Lysis moduleA total of three putative ORFs encoding proteins associated with the lysis of the host
were found; we determined that ORF 45 encodes holin, a protein that permeabilizes
the inner membrane, oligomerizes in the host cell membrane and forms large pores that
are utilized as transport channels for endolysin to access and degrade the peptidoglycan
layer (Shin et al., 2014). Moreover, the lysis module includes ORF 46, which encodes
a protein sharing 84% identity with an endolysin. Analysis of ORF 46 revealed one
conserved motif with lysozyme. The presence of this motif suggests that this protein is
probably an enzyme involved in peptidoglycan cleavage (Xu et al., 2015). The product of
ORF 47 shared over 94% identity with a spanin, a small lipoprotein that is required for
disruption of the outer membrane (Berry et al., 2012).
DISCUSSIONPhages have been used by many researchers to biocontrol E. coli and others types of
bacteria. In all cases, none of the phages reported have been able to lyse all strains. The
present study describes a new bacteriophage, designated phiC119, including a description
of its morphology, host range, analysis of the cohesive ends and genome sequence.
Transmission electron microscopy revealed that the bacteriophage phiC119 belongs to
the order Caudovirales as a member of the Siphoviridae family according to classifications
proposed by the International Committee on Taxonomy of Virus. These results are
consistent with previous reports on bacteriophages because approximately 95% of phage
Figure 6 Graphic representation of genome organization of the phage phiC119. Putative ORFs are indicated as arrows, the orientation of which
shows the direction of transcription. The colors were assigned according to the possible function of each ORF. Morphogenesis (blue), DNA
replication (yellow), lisis (black), DNA packaging (pink), and hypothetical genes with unknown function (gray).
Amarillas et al. (2016), PeerJ, DOI 10.7717/peerj.2423 14/22

isolates are classified in the order Caudovirales (Swanson et al., 2012). Furthermore, within
approximately 4 h, phage phiC119 formed large and clear plaques, which is associated
with phages that possess a lytic cycle (Kwiatek et al., 2015). Previous research suggested
that bacteriophages that produce larger plaques generally have a larger burst size,
indicative of lytic phages (Abedon & Culler, 2007).
Phage phiC119 has strong lytic activity against the E. coli strains used in this study.
Many of the E. coli strains are multidrug resistant and pathogenic in mammalian cells
(Amezquita-Lopez et al., 2012; Amezquita-Lopez et al., 2014). Moreover, the phage was able
to lyse some strains of Salmonella serotypes such as Minnesota, Luciana, Oranienburg,
and Agona, suggesting that phage phiC119 can be considered a broad host range phage
and may be an effective biocontrol agent, as phages with broad host range activity against
STEC strains are advantageous in biocontrol (Niu et al., 2012). The potential for lysis of
the highest number of strains is important for the potential use of bacteriophages in
biocontrol of the bacterial pathogens (Eyer et al., 2007). Therefore, based on broad host
range against STEC strain, we suggest that phiC119 should be considered a good
candidate for biocontrol.
Biological characterization of the phage revealed that phiC119 has an average burst size
of 210 PFU per infected cell with an average latent period of 20 min, indicating that
phiC119 has strong lysis. Phages with high burst sizes are more effective to biocontrol and
phage therapy (Abedon, Herschler & Stopar, 2001). According to the one-step growth curve
results, phiC119 can be considered as a candidate for biocontrol evaluation.
Genetic analyses suggest that the bacteriophage genome is organized into functional
modules. This modular organization allows genes that are involved in the same
biological process to be clustered in the same module, which is common in most tailed
bacteriophages (Haddad et al., 2014; Teng et al., 2015). Furthermore, the phage does
not have cohesive ends. In this regard, Casjens & Gilcrease (2009) argued that phages
with the pac-mechanism (called headful packaging) are able to produce transduction.
However, most new viral particles generated in such process are expected to be
nonviable with defective replication functions and are eliminated by natural selection
(Krupovic et al., 2011). In contrast, recent reports suggested that cos-type phages
represent a novel mechanism of horizontal gene transfer, although at a lower frequency
than pac-type phages.
The restriction profiles indicated the absence of cohesive ends in phiC119 phage
genome. To determine the most probable packaging strategy used by this phage,
phylogenetic tree was constructed by comparing the amino acid sequences of terminase
proteins of the most well-known representatives of each important family of phages,
including the phiC119.
The terminase in phage phiC119 showed 62.1% sequence identity with that of
Enterobacteriophage T1, this phage packages its DNAvia a headful packaging mechanism
(Roberts, Martin & Kropinski, 2004). Considering that terminase determines the DNA-
packaging strategy of the phage (Casjens & Gilcrease, 2009), phylogenetic analysis suggests
that the phage phiC119 packages DNA by a headful mechanism similar to that of T1.
This is in agreement with the restriction endonuclesae digestion analysis.
Amarillas et al. (2016), PeerJ, DOI 10.7717/peerj.2423 15/22

The genome sequence of phiC119 consisted of 47,319 bp with a GC content of 44.20%,
a value lower than that of its hosts. This observation is consistent with previous
reports showing that virulent phages are on average 4% poorer in GC content than
their hosts, while in temperate bacteriophages, the guanine content is usually very close to
the host (Rocha & Danchin, 2002). The low GC content of phage genome suggests that
phage phiC119 might have acquired the ability to infect E. coli strains over a long period of
time (Kwan et al., 2006; Jin et al., 2014). Additionally, genome size is an important
biological property of the virus, as the genome size determines the numbers of proteins
encoded by the phage and is correlated with virion complexity, although there are some
exceptions (Abedon, 2011). These results suggest that phiC119 is a bacteriophage with low
structural complexity; this is consistent with transmission electron microscopic
observations of the phiC119 bacteriophage.
Phage phiC119 has overlapping ORFs, overlapping genes is a common phenomenon
in phage genomes, which is a tactic to minimize genome size. Thus, this represents the
compression of a large amount of genetic information into short nucleotide sequences
without a loss of protein function (Pavesi, 2006). This strategy also plays a fundamental
role in transcriptional and translational regulation of gene expression (Johnson &
Chisholm, 2004).
It is possible that phage phiC119 expresses structural proteins in a more efficient
way because phages encoding tRNAs can overcome possible differences in codon usage
between the phage and the host (Samson & Moineau, 2010). The presence of tRNAs is
common in strictly virulent or lytic phages (Santos et al., 2011). From a biological point
of view, the existence of tRNAs in the phage genome suggests that phiC119 may have a
short latent period and a large burst size because a previous study revealed that tRNAs
enable phages to improve propagation and increase the kinetics of viral replication, as
tRNAs are related to optimal codon usage (Jun et al., 2014).
Comparative analysis of genes at the amino acid sequences using the BLASTP program
revealed that the tail fiber proteins of phiC119 (protein_id = ALJ98900.1 and ALJ98909.1)
are homologous to tail fiber proteins of phages that infect the members of the bacterial
family Enterobacteriaceae, including phage that infect Salmonella and E. coli. Phage
specificity is largely determined by the tail fiber’s ability to bind to specific structures on
the surface of bacteria. The similarities of the tail fiber proteins could imply that these
phages in general have the same host range (Haggard-Ljungquist, Halling & Calendar,
1992). This may be the main reason for the polyvalent activity on Salmonella and E. coli
O157:H7 by phiC119.
Analysis of the genome sequence of bacteriophages considered for use as a biocontrol
agent is essential. This is to ensure that the phage is strictly lytic and does not encode
any phage lysogeny factors, virulence-related genes and/or antibiotic resistance genes
(Endersen et al., 2015). The complete genomic sequence analysis of bacteriophage phiC119
revealed the absence of virulence-encoding genes, potential immunoreactive allergens,
and lysogeny genes.
In conclusion, transmission electron microscopy revealed that phage phiC119 belongs
to the Siphoviridae family. Furthermore, phage phiC119 exhibited a broad host range.
Amarillas et al. (2016), PeerJ, DOI 10.7717/peerj.2423 16/22

Genomic analysis suggests that phage phiC119 does not establish a lysogenic state and
has no known toxic genes, potential allergens or integrases. These results indicate that
phage phiC119 exhibits a number of properties suitable for application as a biocontrol
agent for STEC strains. However, further toxicity studies are required to ensure the
safety of the phage. Therefore, our future research will be aimed at characterizing this
phage for a better understanding of its potential as a biocontrol agent.
ACKNOWLEDGEMENTSWe thank the Food Safety National Research Laboratory (LANIIA) at the Research
Center in Food & Development (CIAD) for experimental support. The authors are
thankful to QFB Lucıa Margarita Rubı Rangel, QFB Sergio Juan Manuel Gonzalez de
Leon, and QFB Jesus Hector Carrillo Yanez for critical technical assistance. We would like
to thank MC Mitzi Dayanira Estrada Acosta for critical reading of manuscript.
ADDITIONAL INFORMATION AND DECLARATIONS
FundingThis investigation was partially supported by Fundacion Produce Sinaloa. The funders
had no role in study design, data collection and analysis, decision to publish, or
preparation of the manuscript.
Competing InterestsThe authors declare that they have no competing interests.
Author Contributions� Luis Amarillas conceived and designed the experiments, performed the experiments,
analyzed the data, wrote the paper, prepared figures and/or tables, reviewed drafts
of the paper.
� Cristobal Chaidez analyzed the data, contributed reagents/materials/analysis tools,
reviewed drafts of the paper.
� Arturo Gonzalez-Robles performed the experiments, analyzed the data, contributed
reagents/materials/analysis tools, reviewed drafts of the paper.
� Yadira Lugo-Melchor analyzed the data, reviewed drafts of the paper.
� Josefina Leon-Felix conceived and designed the experiments, performed the
experiments, analyzed the data, contributed reagents/materials/analysis tools, wrote the
paper, prepared figures and/or tables, reviewed drafts of the paper.
DNA DepositionThe following information was supplied regarding the deposition of DNA sequences:
GenBank: accession number KT825490.
Data DepositionThe following information was supplied regarding data availability:
The raw data has been supplied as Supplemental Dataset Files.
Amarillas et al. (2016), PeerJ, DOI 10.7717/peerj.2423 17/22

Supplemental InformationSupplemental information for this article can be found online at http://dx.doi.org/
10.7717/peerj.2423#supplemental-information.
REFERENCESAbedon ST. 2011. Size does matter-distinguishing bacteriophages by genome length (and
‘breadth’). Microbiology Australia 32(2):95–96.
Abedon ST, Culler RR. 2007. Optimizing bacteriophage plaque fecundity. Journal of
Theoretical Biology 249(3):582–592 DOI 10.1016/j.jtbi.2007.08.006.
Abedon ST, Herschler TD, Stopar D. 2001. Bacteriophage latent-period evolution as a response
to resource availability. Applied and Environmental Microbiology 67(9):4233–4241
DOI 10.1128/AEM.67.9.4233-4241.2001.
Akhtar M, Viazis S, Diez-Gonzalez F. 2014. Isolation, identification and characterization of
lytic, wide host range bacteriophages from waste effluents against Salmonella enterica serovars.
Food Control 38:67–74 DOI 10.1016/j.foodcont.2013.09.064.
Amezquita-Lopez BA, Quinones B, Cooley MB, Leon-Felix J, Castro-del Campo N,
Mandrell RE, Jimenez M, Chaidez C. 2012. Genotypic analyses of Shiga toxin-producing
Escherichia coli O157 and non-O157 rcovered from feces of domestic animals on rural farms
in Mexico. PLoS ONE 7(12):e51565 DOI 10.1371/journal.pone.0051565.
Amezquita-Lopez BA, Quinones B, Lee BG, Chaidez C. 2014. Virulence profiling of Shiga toxin-
producing Escherichia coli recovered from domestic farm animals in Northwestern Mexico.
Frontiers in Cellular and Infection Microbiology 4:7 DOI 10.3389/fcimb.2014.00007.
Amezquita-Lopez BA, Quinones B, Soto-Beltran M, Lee BG, Yambao JC, Lugo-Melchor OY,
Chaidez C. 2016. Antimicrobial resistance profiles of Shiga toxin-producing Escherichia coli
O157 and non-O157 recovered from domestic farm animals in rural communities in
Northwestern Mexico. Antimicrobial Resistance and Infection Control 5(1):1
DOI 10.1186/s13756-015-0100-5.
Belanger L, Garenaux A, Harel J, Boulianne M, Nadeau E, Dozois CM. 2011. Escherichia coli
from animal reservoirs as a potential source of human extraintestinal pathogenic E. coli.
FEMS Immunology & Medical Microbiology 62(1):1–10
DOI 10.1111/j.1574-695X.2011.00797.x.
Berry J, Rajaure M, Pang T, Young R. 2012. The Spanin complex is essential for Lambda lysis.
Journal of Bacteriology 194(20):5667–5674 DOI 10.1128/JB.01245-12.
Canizalez-Roman A, Gonzalez-Nunez E, Vidal JE, Flores-Villasenor H, Leon-Sicairos N.
2013. Prevalence and antibiotic resistance profiles of diarrheagenic Escherichia coli strains
isolated from food items in northwestern Mexico. International Journal of Food Microbiology
164(1):36–45 DOI 10.1016/j.ijfoodmicro.2013.03.020.
Carey-Smith GV, Billington C, Cornelius AJ, Hudson JA, Heinemann JA. 2006. Isolation and
characterization of bacteriophages infecting Salmonella spp. FEMS Microbiology Letters
258(2):182–186 DOI 10.1111/j.1574-6968.2006.00217.x.
Casjens SR, Gilcrease EB. 2009. Determining DNA packaging strategy by analysis of the
termini of the chromosomes in tailed-bacteriophage virions. Methods and Protocols, Volume 2,
Molecular and Applied Aspects. Vol. 502. Humana Press, 91–111.
Castro del CampoN, Amarillas Bueno LA, Garcıa CamarenaMG, Chaidez Quiroz C, Leon Felix J,
Martınez Rodrıguez CI. 2011. Presencia de Salmonella y Escherichia coli O157:H7 en la
Amarillas et al. (2016), PeerJ, DOI 10.7717/peerj.2423 18/22

zona centro del estado de Sinaloa y su control biologico mediante el uso de bacteriofagos
[abstract no. C39]. Congreso Internacional de Inocuidad de Alimentos 8:165–168.
Centers for Disease Control Prevention. 2015. Available at http://www.cdc.gov/ecoli/general/
index.html/ (accessed 3 March 2016).
Chan BK, Abedon ST, Loc-Carrillo C. 2013. Phage cocktails and the future of phage therapy.
Future Microbiology 8(6):769–783 DOI 10.2217/fmb.13.47.
Chen L, Xiong Z, Sun L, Yang J, Jin Q. 2012. VFDB 2012 update: toward the genetic diversity and
molecular evolution of bacterial virulence factors. Nucleic Acids Research 40(D1):D641–D645
DOI 10.1093/nar/gkr989.
Endersen L, Guinane CM, Johnston C, Neve H, Coffey A, Ross RP, McAuliffe O, O’Mahony J.
2015. Genome analysis of Cronobacter phage vB_CsaP_Ss1 reveals an endolysin with
potential for biocontrol of Gram-negative bacterial pathogens. Journal of General Virology
96(2):463–477 DOI 10.1099/vir.0.068494-0.
Estrada-Acosta M, Jimenez M, Chaidez C, Leon-Felix J, Castro-del Campo N. 2014.
Irrigation water quality and the benefits of implementing good agricultural practices during
tomato (Lycopersicum esculentum) production. Environmental Monitoring and Assessment
186(7):4323–4330 DOI 10.1007/s10661-014-3701-1.
Eyer L, Pantucek R, Zdrahal Z, Konecna H, Kasparek P, Ruzickova V, Hernychova L, Preisler J,
Doska�r J. 2007. Structural protein analysis of the polyvalent staphylococcal bacteriophage 812.
Proteomics 7(1):64–72 DOI 10.1002/(ISSN)1615-9861.
Farfan MJ, Torres AG. 2012. Molecular mechanisms that mediate colonization of Shiga
toxin-producing Escherichia coli strains. Infection and Immunity 80(3):903–913
DOI 10.1128/IAI.05907-11.
Gallet R, Kannoly S, Wang I-N. 2011. Effects of bacteriophage traits on plaque formation.
BMC Microbiology 11(1):181 DOI 10.1186/1471-2180-11-181.
Guenther S, Herzig O, Fieseler L, Klumpp J, Loessner MJ. 2012. Biocontrol of Salmonella
Typhimurium in RTE foods with the virulent bacteriophage FO1-E2. International Journal of
Food Microbiology 154(1–2):66–72 DOI 10.1016/j.ijfoodmicro.2011.12.023.
Haddad LE, Abdallah NB, Plante P-L, Dumaresq J, Katsarava R, Labrie S, Corbeil J, St-Gelais D,
Moineau S. 2014. Improving the safety of Staphylococcus aureus polyvalent phages by their
production on a Staphylococcus xylosus strain. PLoS ONE 9(7):e102600
DOI 10.1371/journal.pone.0102600.
Hagens S, Loessner MJ. 2010. Bacteriophage for biocontrol of foodborne pathogens:
calculations and considerations. Current Opinion in Biotechnology 11(1):58–68
DOI 10.2174/138920110790725429.
Haggard-Ljungquist E, Halling C, Calendar R. 1992. DNA sequences of the tail fiber genes of
bacteriophage P2: evidence for horizontal transfer of tail fiber genes among unrelated
bacteriophages. Journal of Bacteriology 174(5):1462–1477.
Hollis T, Stattel JM, Walther DS, Richardson CC, Ellenberger TE. 2001. Crystal structure of
gp2.5, a single-stranded DNA binding protein encoded by bacteriophage T7. Proceedings of the
National Academy of Sciences of the United States of America 98(17):9557–9562
DOI 10.1073/pnas.171317698.
Hua Y, An X, Pei G, Li S, Wang W, Xu X, Fan H, Huang Y, Zhang Z, Mi Z, Chen J, Li J,
Zhang F, Tong Y. 2014. Characterization of the morphology and genome of an
Escherichia coli podovirus. Archives of Virology 159(12):3249–3256
DOI 10.1007/s00705-014-2189-x.
Amarillas et al. (2016), PeerJ, DOI 10.7717/peerj.2423 19/22

Hungaro HM, Mendonca RCS, Gouvea DM, Vanetti MCD, de Oliveira Pinto CL. 2013. Use
of bacteriophages to reduce Salmonella in chicken skin in comparison with chemical agents.
Food Research International 52(1):75–81 DOI 10.1016/j.foodres.2013.02.032.
Jakobsen L, Garneau P, Bruant G, Harel J, Olsen SS, Porsbo LJ, Hammerum AM,
Frimodt-Møller N. 2012. Is Escherichia coli urinary tract infection a zoonosis? Proof of
direct link with production animals and meat. European Journal of Clinical Microbiology &
Infectious Diseases 31(6):1121–1129 DOI 10.1007/s10096-011-1417-5.
Jimenez M, Martinez-Urtaza J, Rodriguez-Alvarez MX, Leon-Felix J, Chaidez C. 2014.
Prevalence and genetic diversity of Salmonella spp. in a river in a tropical environment in
Mexico. Journal of Water and Health 12(4):874–884 DOI 10.2166/wh.2014.051.
Jin J, Li Z-J, Wang S-W, Wang S-M, Chen S-J, Huang D-H, Zhang G, Li Y-H, Wang X-T, Wang J,
Zhao G-Q. 2014. Genome organisation of the Acinetobacter lytic phage ZZ1 and
comparison with other T4-like Acinetobacter phages. BMC Genomics 15:793
DOI 10.1186/1471-2164-15-793.
Johnson ZI, Chisholm SW. 2004. Properties of overlapping genes are conserved across microbial
genomes. Genome Research 14(11):2268–2272 DOI 10.1101/gr.2433104.
Jun JW, Kim HJ, Yun SK, Chai JY, Park SC. 2015. Genomic structure of the Aeromonas
bacteriophage pAh6-C and its comparative genomic analysis. Archives of Virology
160(2):561–564 DOI 10.1007/s00705-014-2221-1.
Jun JW, Yun SK, Kim HJ, Chai JY, Park SC. 2014. Characterization and complete genome
sequence of a novel N4-like bacteriophage, pSb-1 infecting Shigella boydii. Research in
Microbiology 165(8):671–678 DOI 10.1016/j.resmic.2014.09.006.
Kleinheinz KA, Joensen KG, Larsen MV. 2014. Applying the ResFinder and VirulenceFinder
web-services for easy identification of acquired antibiotic resistance and E. coli virulence
genes in bacteriophage and prophage nucleotide sequences. Bacteriophage 4(2):e27943
DOI 10.4161/bact.27943.
Krupovic M, Prangishvili D, Hendrix RW, Bamford DH. 2011. Genomics of bacterial and
archaeal viruses: dynamics within the prokaryotic virosphere. Microbiology and Molecular
Biology Reviews 75(4):610–635 DOI 10.1128/MMBR.00011-11.
Kwan T, Liu J, DuBow M, Gros P, Pelletier J. 2006. Comparative Genomic Analysis of 18
Pseudomonas aeruginosa Bacteriophages. Journal of Bacteriology 188(3):1184–1187
DOI 10.1128/JB.188.3.1184-1187.2006.
Kwiatek M, Mizak L, Parasion S, Gryko R, Olender A, Niemcewicz M. 2015. Characterization
of five newly isolated bacteriophages active against Pseudomonas aeruginosa clinical strains.
Folia Microbiologica 60(1):7–14 DOI 10.1007/s12223-014-0333-3.
Lopez-Cuevas O, Castro-del Campo N, Leon-Felix J, Gonzalez-Robles A, Chaidez C. 2011.
Characterization of bacteriophages with a lytic effect on various Salmonella serotypes and
Escherichia coli O157:H7. Canadian Journal of Microbiology 57(12):1042–1051
DOI 10.1139/w11-099.
Mahony J, McAuliffe O, Ross RP, van Sinderen D. 2011. Bacteriophages as biocontrol
agents of food pathogens. Current Opinion in Biotechnology 22(2):157–163
DOI 10.1016/j.copbio.2010.10.008.
Miller ES, Kutter E, Mosig G, Arisaka F, Kunisawa T, Ruger W. 2003. Bacteriophage T4
genome. Microbiology and Molecular Biology Reviews 67(1):86–156
DOI 10.1128/MMBR.67.1.86-156.2003.
Moodley S, Maxwell K, Kanelis V. 2012. The protein gp74 from the bacteriophage HK97
functions as a HNH endonuclease. Protein Science 21(6):809–818 DOI 10.1002/pro.2064.
Amarillas et al. (2016), PeerJ, DOI 10.7717/peerj.2423 20/22

Niu YD, Stanford K, Kropinski AM, Ackermann H-W, Johnson RP, She Y-M, McAllister TA.
2012. Genomic, proteomic and physiological characterization of a T5-like bacteriophage for
control of Shiga toxin-producing Escherichia coli O157: H7. PLoS ONE 7(4):e34585
DOI 10.1371/journal.pone.0034585.
Paton AW, Paton JC. 1998. Detection and characterization of shiga toxigenic Escherichia coli by
using multiplex PCR assays for stx1, stx2, eaea, enterohaemorrhagic E. coli hlya, rfbO111,
and rfbO157. Journal of Clinical Microbiology 36(2):598–602.
Pavesi A. 2006. Origin and evolution of overlapping genes in the family Microviridae. Journal of
General Virology 87(4):1013–1017 DOI 10.1099/vir.0.81375-0.
Pickard D, Thomson NR, Baker S, Wain J, Pardo M, Thomson NR, Baker S, Wain J, Pickard D,
Goulding D, Hamlin N, Choudhary J, Threfall J, Dougan G. 2008.Molecular characterization
of the Salmonella enterica serovar Typhi Vi-typing bacteriophage E1. Journal of Bacteriology
190(7):2580–2587 DOI 10.1128/JB.01654-07.
Pierard D, De Greve H, Haesebrouck F, Mainil J. 2012. O157:H7 and O104:H4 Vero/Shiga toxin-
producing Escherichia coli outbreaks: respective role of cattle and humans. Veterinary Research
43(1):13 DOI 10.1186/1297-9716-43-13.
Roberts MD, Martin NL, Kropinski AM. 2004. The genome and proteome of coliphage T1.
Virology 318(1):245–266 DOI 10.1016/j.virol.2003.09.020.
Rocha EPC, Danchin A. 2002. Base composition bias might result from competition for metabolic
resources. Trends in Genetics 18(6):291–294 DOI 10.1016/S0168-9525(02)02690-2.
Sambrook J, Russell DW. 2001. Molecular Cloning: A Laboratory Manual. Third edition.
New York: Cold Spring Harbor Laboratory Press.
Samson JE, Moineau S. 2010. Characterization of Lactococcus lactis phage 949 and comparison
with other Lactococcal phages. Applied and Environmental Microbiology 76(20):6843–6852
DOI 10.1128/AEM.00796-10.
Santos SB, Kropinski AM, Ceyssens P-J, Ackermann H-W, Villegas A, Lavigne R, Krylov VN,
Carvalho CM, Ferreira EC, Azeredo J. 2011. Genomic and proteomic characterization of
ghe broad-host-range Salmonella phage PVP-SE1: creation of a new phage genus. Journal of
Virology 85(21):11265–11273 DOI 10.1128/JVI.01769-10.
Santos TMA, Bicalho RC. 2011. Complete genome sequence of vB_EcoM_ECO1230-10: a
coliphage with therapeutic potential for bovine metritis. Veterinary Microbiology
148(2–4):267–275 DOI 10.1016/j.vetmic.2010.08.034.
Shin H, Lee J-H, Yoon H, Kang D-H, Ryu S. 2014. Genomic investigation of lysogen formation
and host lysis systems of the Salmonella temperate bacteriophage SPN9CC. Applied and
Environmental Microbiology 80(1):374–384 DOI 10.1128/AEM.02279-13.
Swanson MM, Reavy B, Makarova KS, Cock PJ, Hopkins DW, Torrance L, Koonin EV,
Taliansky M. 2012.Novel bacteriophages containing a genome of another bacteriophage within
their genomes. PLoS ONE 7(7):e40683 DOI 10.1371/journal.pone.0040683.
Teng T, Yu J, Yang H, Wei H. 2015. Isolation and complete genome sequence of a
novel virulent mycobacteriophage, CASbig. Virologica Sinica 30(1):76–79
DOI 10.1007/s12250-014-3545-4.
Xu Y, Ma Y, Yao S, Jiang Z, Pei J, Cheng C. 2015. Characterization, genome sequence, and analysis
of Escherichia phage CICC 80001, a bacteriophage infecting an efficient L-aspartic acid
producing Escherichia coli. Food and Environmental Virology 8(1):18–26
DOI 10.1007/s12560-015-9218-0.
Yamashita E, Nakagawa A, Takahashi J, Tsunoda K, Yamada S, Takeda S. 2011. The host-binding
domain of the P2 phage tail spike reveals a trimeric iron-binding structure. Acta
Amarillas et al. (2016), PeerJ, DOI 10.7717/peerj.2423 21/22

Crystallographica Section F Structural Biology and Crystallization Communications
67(Pt 8):837–841 DOI 10.1107/S1744309111005999.
Yan Y, Shi Y, Cao D, Meng X, Xia L, Sun J. 2011. Prevalence of Stx phages in environments of a pig
farm and lysogenic infection of the field E. coli O157 isolates with a recombinant converting
phage. Current Microbiology 62(2):458–464 DOI 10.1007/s00284-010-9729-8.
Amarillas et al. (2016), PeerJ, DOI 10.7717/peerj.2423 22/22





















![BACTERIOPHAGE-RESISTANT AND BACTERIOPHAGE-SENSITIVE ...halsmith/phagemutantsubmitted_2.pdf · BACTERIOPHAGE-RESISTANT AND BACTERIOPHAGE-SENSITIVE BACTERIA IN A CHEMOSTAT ... [22],](https://static.cupdf.com/doc/110x72/5b3839687f8b9a5a518d2ce1/bacteriophage-resistant-and-bacteriophage-sensitive-halsmithphagemutantsubmitted2pdf.jpg)