Mkfobiology (1997), 143, 3417-3429 Printed in Great Britain Sequence analysis and characterization of #01205, a temperate bacteriophage infecting Streptococcus thermophilus CNRZI205 Elizabeth StanIey,’n2 Gerald F. Fitzgerald,’i2 Claire Le Marrec,’t Blandine Fayard’+ and Douwe van Sinderen’’’ Author for correspondence: Douwe van Sinderen. Tel: +353 21 902811. Fax: +353 21 903101. e-mail : [email protected] Department of Microbiology’ and National Food Biotechnology Centre*, University College, Cork, Ireland ~ The complete nucleotidesequence of Wl205, a temperate bacteriophage infecting Streptococcus thermophilus strain CNRZ1205, was determined. The phage genome has a unit length of 43075 bp and appears to be packaged by the so-called headful mechanism. The genomic organization and structure of #01205 resemble those of several temperate lactococcal phages that display a life-cycle-specific organization, where ORFs believedto be involved in the lysogenic life-cycleare clustered and arranged in an orientation opposite to the ORFs supposedly involved in the lytic life-cycle.Database searches revealed putative functions for several identified ORFs and further indicated that #01205 is genetically related to a particular group of lactococcal phages. Three genes encoding the major structural proteins were identified on the Wl205 genome. The phage attachment site attP, the bacterial attachment site attB, and the two phagekhromosome junctions attL and attR were identified and found to contain a 40 bp common core sequence. Keywords : Streptococcus thermophilus, bacteriophage, attachment site, structural proteins, sequence analysis INTRODUCTION Bacteriophage infection of starter cultures has been recognized as a significant problem in the dairy industry since the 1930s. Phages can cause slow lactic acid fermentations, yielding products of inferior quality. Many studies have been undertaken to characterize phages infecting mesophilic lactococci and as a result, a substantial body of knowledge has been generated with respect to the phage-host interactions as well as to molecular aspects of these phages (for reviews see, among others, Mata & Ritzenthaler, 1988 ; Jarvis et al., t Present address: lnstitut des Sciences et Techniques des Aliments, Universite Bordeaux 1, Talenc. France. +Present address: Department of Genetics, Groningen Biomolecular Sciences and BiotechnologyInstitute, University of Groningen, Haren, The Netherlands. The GenBank accession number for the complete sequence of 401205 is U88974. The nucleotide sequence of the attB region of 5. thermophilus CNRZ1205 as well as information on the core region of 5. thermophilus CNRZ385 and 5. thermophilus ND16 can be found under accession number U88973. 1991; Klaenhammer & Fitzgerald, 1994; Garvey et al., 1995).Until recently, however, there have been relatively few scientific studies concerning phages infecting thermophilic lactic acid bacteria and these have been limited primarily to the analysis of phage-host inter- actions. Streptococcus thermophilus phages have assumed a greater significance to the dairy industry due to an increase in large-scale commercial use of thermo- philic starter cultures in products such as yogurt and Mozzarella cheese. S. thermophilus is also used in the manufacture of short-method Cheddar cheese (Czulak et al., 1954), where it may become susceptible to phage attack during large-scale intensive production processes. The genomes of all characterized virulent and temperate S. thermophilus phages exhibit homology to each other in a modular fashion, where non-homologous regions are flanked by homologous DNA sections (Mercenier, 1990). Several studies have been undertaken to charac- terize and type both lytic and lysogenic S. thermophilus phages (Krusch et al., 1987; Prevots et al., 1989; Neve et al., 1989; Benbadis et al., 1990; Larbi et al., 1990; Carminati & Giraffa, 1992; Sebastiani & Jager, 1992; Fayard et al., 1993; Brussow et al., 1994a, b ; Brussow & 0002-1832 0 1997 SGM 341 7

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Mkfobiology (1 997), 143, 341 7-3429 Printed in Great Britain
Sequence analysis and characterization of #01205, a temperate bacteriophage infecting Streptococcus thermophilus CNRZI 205
Elizabeth StanIey,’n2 Gerald F. Fitzgerald,’i2 Claire Le Marrec,’t Blandine Fayard’+ and Douwe van Sinderen’’’
Author for correspondence: Douwe van Sinderen. Tel: +353 21 902811. Fax: +353 21 903101. e-mail : [email protected]
Department of Microbiology’ and National Food Biotechnology Centre*, University College, Cork, Ireland
~
The complete nucleotide sequence of Wl205, a temperate bacteriophage infecting Streptococcus thermophilus strain CNRZ1205, was determined. The phage genome has a unit length of 43075 bp and appears to be packaged by the so-called headful mechanism. The genomic organization and structure of #01205 resemble those of several temperate lactococcal phages that display a life-cycle-specific organization, where ORFs believed to be involved in the lysogenic life-cycle are clustered and arranged in an orientation opposite to the ORFs supposedly involved in the lytic life-cycle. Database searches revealed putative functions for several identified ORFs and further indicated that #01205 is genetically related to a particular group of lactococcal phages. Three genes encoding the major structural proteins were identified on the W l 2 0 5 genome. The phage attachment site attP, the bacterial attachment site attB, and the two phagekhromosome junctions attL and attR were identified and found to contain a 40 bp common core sequence.
Keywords : Streptococcus thermophilus, bacteriophage, attachment site, structural proteins, sequence analysis
INTRODUCTION
Bacteriophage infection of starter cultures has been recognized as a significant problem in the dairy industry since the 1930s. Phages can cause slow lactic acid fermentations, yielding products of inferior quality. Many studies have been undertaken to characterize phages infecting mesophilic lactococci and as a result, a substantial body of knowledge has been generated with respect to the phage-host interactions as well as to molecular aspects of these phages (for reviews see, among others, Mata & Ritzenthaler, 1988 ; Jarvis et al.,
t Present address: lnstitut des Sciences et Techniques des Aliments, Universite Bordeaux 1, Talenc. France.
+Present address: Department of Genetics, Groningen Biomolecular Sciences and Biotechnology Institute, University of Groningen, Haren, The Netherlands.
The GenBank accession number for the complete sequence of 401205 is U88974. The nucleotide sequence of the attB region of 5. thermophilus CNRZ1205 as well as information on the core region of 5. thermophilus CNRZ385 and 5. thermophilus ND16 can be found under accession number U88973.
1991; Klaenhammer & Fitzgerald, 1994; Garvey et al., 1995). Until recently, however, there have been relatively few scientific studies concerning phages infecting thermophilic lactic acid bacteria and these have been limited primarily to the analysis of phage-host inter- actions. Streptococcus thermophilus phages have assumed a greater significance to the dairy industry due to an increase in large-scale commercial use of thermo- philic starter cultures in products such as yogurt and Mozzarella cheese. S. thermophilus is also used in the manufacture of short-method Cheddar cheese (Czulak et al., 1954), where it may become susceptible to phage attack during large-scale intensive production processes.
The genomes of all characterized virulent and temperate S. thermophilus phages exhibit homology to each other in a modular fashion, where non-homologous regions are flanked by homologous DNA sections (Mercenier, 1990). Several studies have been undertaken to charac- terize and type both lytic and lysogenic S . thermophilus phages (Krusch et al., 1987; Prevots et al., 1989; Neve et al., 1989; Benbadis et al., 1990; Larbi et al., 1990; Carminati & Giraffa, 1992; Sebastiani & Jager, 1992; Fayard et al., 1993; Brussow et al., 1994a, b ; Brussow &
0002-1832 0 1997 SGM 341 7

E. S T A N L E Y a n d OTHERS
Bruttin, 1995; Bruttin & Brussow, 1996). So far only two reports have included actual sequence information on S. thermophilus phages (Briissow et al., 1994b; Bruttin & Brussow, 1996). The first published S. thermophilus phage D N A sequence described a 2-2 kb EcoRI fragment which appeared to be present in approximately 60 '/o of the phages encountered (Briissow et al., 1994b). Two ORFs were identified within this DNA fragment but no function could be ascribed to any of these putative genes. The second report on an S. thermophilus phage sequence concerned an X6aI fragment which was shown to contain sequences suffering from spontaneous deletions and possibly involved in life-cycle control (Bruttin & Brussow, 1996).
This study represents the initial phase of a project designed to obtain fundamental knowledge on the molecular biology of phages infecting S. thermophilus. We report the complete nucleotide sequence and mol- ecular characterization of the temperate phage 01205, which is resident as a prophage in the S. tbermopbilus strain CNRZ120.5.
Bacterial strains, plasmids, phage, and growth media. Phage, bacterial strains and plasmids used in this study are listed in Table 1. S. thermophilus CNRZ120.5 was grown at 43 "C in either Elliker medium (Difco) supplemented with 10 g beef extract 1-1 (Difco) or M17 medium (Terzaghi & Sandine, 1975) supplemented with 10 g lactose 1-I (LM17). Escherichia coli was cultivated in Luria-Bertani broth as describ5d by Sambrook et al. (1989). Ampicillin and tetracycline were used to select for E . coli transformants at concentrations of 100 pg ml-I and 12.5 pg rnl-', respectively. IPTG and X-Gal were used, where appropriate, at concentrations of 0.5 mM and 40 pg ml-', respectively.
Induction of #01205. 401205 was induced from its lysogenic host S. thermophilus CNRZ1205 with mitomycin C (Sigma). After removal of the cell debris the lysate was treated with RNase (10 pg ml-l), DNase (10 pg ml-l) and MgCl, (10 mM) to ensure degradation of host nucleic acids. NaCl (0.5 M) and PEG 6000 (10 '/o ) were added to the supernatant and phage particles were allowed to precipitate during overnight in- cubation at 4 "C. Phage particles were subsequently collected by centrifugation and concentrated by caesium chloride density-gradient centrifugation as described by Sambrook et al. (1989).
Electron microscopy. 4 0 1205 was negatively stained with 2 '/o
uranyl acetate and examined using a JEOL 1200 EX trans- mission electron microscope a t an accelerating voltage of 80 kV.
Molecular cloning and DNA isolations. Restriction enzymes, calf intestinal phosphatase and T4 DNA ligase were purchased from Boehringer Mannheim and used according to the supplier's instructions.
Phage genomic DNA was isolated from concentrated phage by performing a number of salt-saturated phenol extractions followed by a single chloroform/isoamyl alcohol (24: 1, v/v) extraction. DNA was precipitated with 100 '/o ethanol at
-20 "C in the presence of 0.3 M sodium acetate (pH 5-0). The DNA pellet was washed with 80% ethanol, dried and resuspended in TE (10 mM Tris, 1 mM EDTA, p H 8.0) buffer. Plasmid DNA was isolated by the method of Birnboim & Doly (1979) or by using the QIAprep Spin Plasmid Miniprep kit (QIAGEN). Chromosomal DNA was isolated essentially according to the method of Slos et al. (1991) with the following modifications. Following cell lysis, 0.5 vol. 5 M NaCl and 1.0 vol. chloroform were added and incubation was continued for a further 30 min at room temperature. The solution was subsequently centrifuged, the aqueous phase collected and the DNA precipitated by adding an equal volume of 2-propanol followed by incubation at room temperature for 30 min. The DNA pellet was isolated by centrifugation and resuspended in TE buffer. The chromosomal DNA was further purified by caesium chloride/ethidium bromide density-gradient centri- fugation as described by Sambrook et al. (1989).
The E . coli vector pBluescript I1 SK( - ) (Stratagene) was used for shotgun cloning of a variety of restriction-enzyme- generated fragments of the 401205 genome. Ligation mixtures were used to transform electrocompetent E. coli XL1 Blue (Stratagene) using ampicillin selection. Positive selection of recombinant plasmids was performed using blue/white screening on plates containing X-Gal and IPTG.
DNA sequence analysis. DNA sequence determination was performed with an Applied Biosystems 373A automated DNA sequencer, using synthetic oligonucleotides (Oligo 1000M, Beckman Instruments) as primers. 401205 sequences were obtained by sequencing cloned phage DNA fragments and primer walking. Assembly of sequences was performed using the Seqman program of the DNASTAR software package. Database searches were performed using the FASTA (Pearson & Lipman, 1988), BLASTN and TBLASTN (Altschul et d, 1990) programs with sequences present in the following databases : SWISSPROT (release 30) ; NBRF-PIR (release 42) ; GenBank translated (release 86) and EMBL (release 38). Sequence alignments were performed using the Clustal method of the MEGALIGN program of the DNASTAR software package.
DNA transfer and hybridization. After digested DNA had been separated on an agarose gel the fragments were trans- ferred to a Hybond-N+ nucleic transfer membrane (Amersham) by Southern blotting (Southern, 1975). The enhanced chemiluminescence (ECL) kit was used to label DNA for use as a probe according to conditions specified by the supplier (Amersham).
Inverse PCR. Inverse PCR (Ochman et al., 1990) was per- formed on digested and religated DNA using the Expand Long Template PCR System (Boehringer Mannheim) . SDS- PAGE and determination of the N-terminal amino acid sequence of the major structural phage proteins. SDS-PAGE was performed as described by Laemmli (1970). Fifteen micrograms of CsC1-purified phage was boiled for 20 min with an equal volume of SDS-PAGE loading buffer (50 mM Tris/HCl, 3 '/o SDS, 1 '/o /?-mercaptoethanol, 20 YO glycerol, 0.7% bromophenol blue, pH 6.8) and applied to an SDS- containing polyacrylamide gel (10 O/O or 12.5 %) using a mini- Protean I1 system (Bio-Rad). After electrophoresis the gels were either stained with Coomassie brilliant blue R-250 (Sigma) to visualize proteins, or transferred to a PVDF protein transfer membrane (Applied Biosystems), using CAPS (Sigma) transfer buffer. The blot was subsequently stained with Coomassie brilliant blue R-250 and clearly visible bands were excised. Their N-terminal amino acid sequence was deter- mined using an Applied Biosystems 477A protein sequencer.
341 8
S. thermophilus phage 01205
Table 1. Bacterial strains, phage and plasmid
Bacterium, phage Relevant characteristics Source or reference or plasmid
S . thermophilus CNRZ120.5
CNRZ38.5
ND16
E . coli XL1-Blue
S . thermophilus
#01205 Plasmid pBluescript I1 SK( - )
phage
Traditional strain used in yogurt production ;
Highly proteolytic and phage-resistant strain
Highly proteolytic and phage-resistant strain
Fayard et al. (1993)
M. Desmazeaud"
Dr S. Sannabhadtit
lysogenic for #01205
isolated from yogurt
isolated from dahi (an Indian fermented milk)
recAl endAl gyrA96 thi-1 hsdRl7 supE44 Stratagene relAl lac [F' proAB lacZqZAM1.5Tn10(Tet')]
Temperate phage induced from CNRZ1205 Fayard et al. (1993)
Amp', lacZ a complementation Stratagene
'* INRA, Jouy-en- Josas, France. t University of Anand, Province of Gujarat, India.
RESULTS
Electron microscopy of #01205
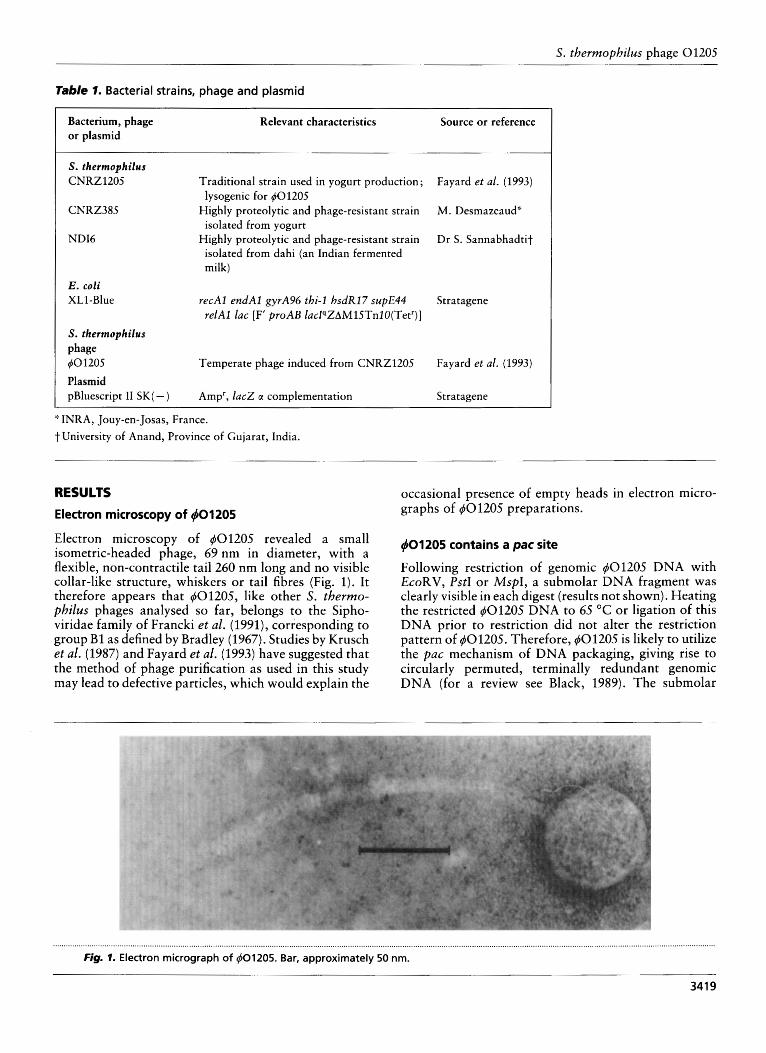
Electron microscopy of 4 0 1205 revealed a small isometric-headed phage, 69 nm in diameter, with a flexible, non-contractile tail 260 nm long and no visible collar-like structure, whiskers or tail fibres (Fig. 1). It therefore appears that 401205, like other S. thermo- philus phages analysed so far, belongs to the Sipho- viridae family of Francki et al. (1991), corresponding to group B1 as defined by Bradley (1967). Studies by Krusch et al. (1987) and Fayard et al. (1993) have suggested that the method of phage purification as used in this study may lead to defective particles, which would explain the
occasional presence of empty heads in electron micro- graphs of 401205 preparations.
#01205 contains a pac site
Following restriction of genomic 401205 DNA with EcoRV, PstI or MspI , a submolar DNA fragment was clearly visible in each digest (results not shown). Heating the restricted #01205 DNA to 65 "C or ligation of this DNA prior to restriction did not alter the restriction pattern of 40120.5. Therefore, 401205 is likely to utilize the pac mechanism of DNA packaging, giving rise to circularly permuted, terminally redundant genomic DNA (for a review see Black, 1989). The submolar
Fig. 7. Electron micrograph of 401205. Bar, approximately 50 nm.
341 9

E. S T A N L E Y a n d OTHERS
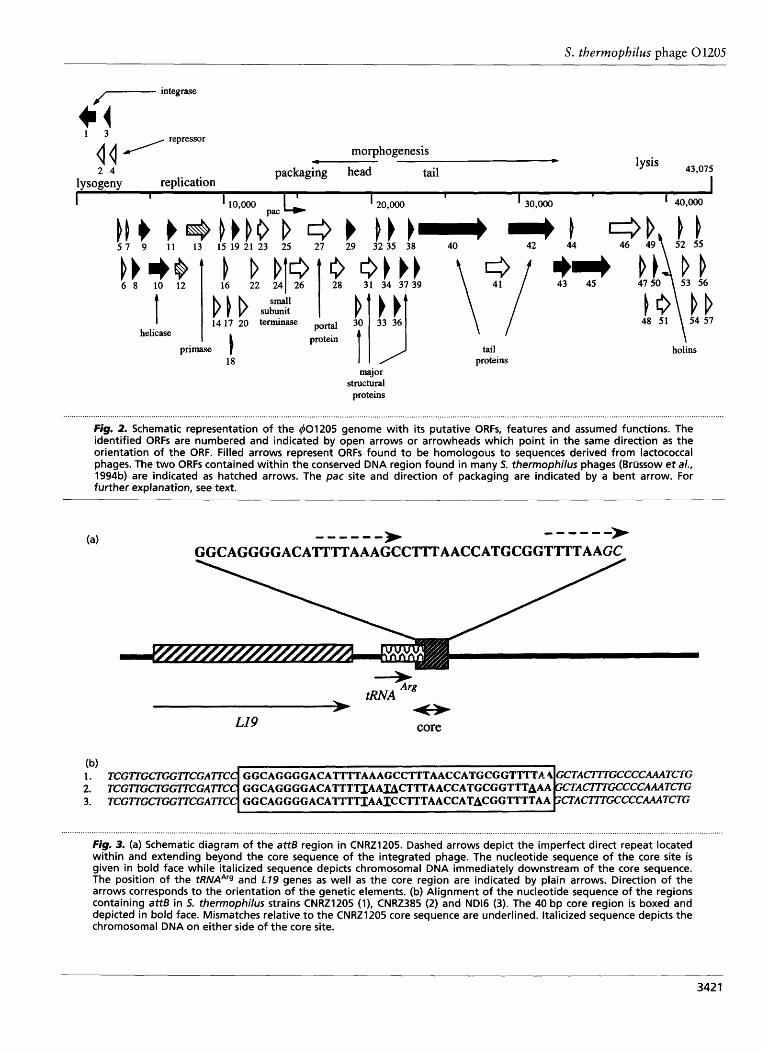
fragment observed in many restriction digests is the product of cleavage at the pac site at one end of the submolar fragment by the terminase and restriction at the recognition site of the restriction endonuclease at the other end. The direction of packaging is therefore from the pac site towards the restriction enzyme site. The size and identity of single submolar fragments derived from various restriction enzyme digests of 40120.5 genomic DNA was compared to their corresponding intact DNA fragments on the restriction map derived from the sequence data. These comparisons allowed the delin- eation of the pac site as well as the determination of the direction of packaging (see Fig. 2). The location of pac was determined to be within the coding region of ORF2.5, which specifies a putative small subunit of the terminase (see below). The direction of packaging was deduced to be the same as the orientation of this ORF.
Localization and analysis of the DNA region containing attP, attL attR and attB
In order to identify the DNA fragments carrying the attP, attL, attR and attB sites, 401205 genomic DNA and chromosomal DNA from its lysogenic host, CNRZ120.5, were digested with various enzymes and analysed in a Southern hybridization experiment using labelled 401205 DNA as a probe. This hybridization experiment indicated that only one integration site was present and that the attP site was localized on a 1-5 kb M6oI fragment or an 800 bp EcoRV fragment of the phage genome (data not shown). Sequence analysis of the fragment containing the attP site allowed the design of primers which were used to amplify the attL- and attR-containing regions through inverse PCR using appropriately digested and ligated chromosomal DNA of strain CNRZ120.5 as a template. Sequencing of these inverse PCR products, and comparison of the sequences obtained with that of the DNA region which contained attP, identified the chromosomal sequences from CNRZ120.5 flanking the attL and attR sites. Additional chromosomal sequences on either side of the attachment region were obtained through further rounds of inverse PCR. The attB-containing region was determined using primers designed on the chromosomal DNA sequence upstream from the attR region and downstream from the attL region. It is interesting that attB could be amplified in CNRZ1205, indicating that prophage-free chromosomes are also present in the lysogenic host, presumably due to a low frequency of spontaneous excision of the phage. Similar observations were made for the lysogenic Lactococcus lactis strains IMN-C3 (Lillehaug & Birkeland, 1993) and UC.509 (Van de Guchte et al., 1994). Inspection of the attL and attR junction points and the attP- and attB-containing regions revealed the presence of a common 40 bp region (Fig. 3). This common DNA sequence probably represents the so-called core region (Campbell, 1992) which denotes the site of recombination between 40120.5 and the host chromosome. The primers used to amplify and sequence the attB region in S. thermophilus CNRZ120.5 were used in a similar experiment with the non-lysogenic strains
CNRZ385 and ND16 in order to determine whether the putative core region was present in other strains of S. thermophilus. Comparison of the DNA sequence obtained from the PCR products revealed the presence of a slightly different core sequence in each of the strains tested. Four nucleotide substitutions were found in the core region of CNRZ385, while three base-pair sub- stitutions were observed in the corresponding region of NDI6. Whether these differences affect the capacity of 401205 to lysogenize these strains could not be de- termined since CNRZ38.5 and NDI6 are not sensitive to infection by 401205. Sequence analysis of the DNA region surrounding the attL and attR sites revealed that 401205 is integrated at the 3' end of a gene almost identical to a tRNAArg gene (anticodon CCU) from E. coli (Kiesewetter et al., 1987; Komine et al., 1990) and that the integration event does not affect the integrity of the tRNAArg. A second ORF upstream from the putative tRNAArg (Fig. 3) shows significant similarity to the ribosomal protein L19 from a number of genera, e.g. Haemophilus influenzae (accession number L44846), Salmonella typhimurium (accession number X74933), Serratia marcescens (Jin & Benedik, 1994) and Synecho- cystis sp. (Schmidt & Subramanian, 1993).
Nucleotide sequence determination of the 401 205 genome
A shotgun cloning strategy was used to clone various restriction enzyme fragments of 4 0 1205 genomic DNA into appropriately restricted E. coli cloning vector pBluescript I1 SK( - ). Randomly selected clones were sequenced and the resulting sequence information was subsequently used to design primers in order to sequence the remaining gaps using either existing clones or genomic 401205 DNA as a template. In this manner both DNA strands were sequenced with an approximate fourfold redundancy, revealing a linear genome with a unit length of 43075 bp. Predicted restriction maps of the sequence matched experimentally determined re- striction maps, which indicated that no obvious as- sembly errors or cloning rearrangements had occurred. The total G + C content of the phage is 38 mol ' /o, which is similar to that of the S. thermophilus host chromo- some (Kilpper-Balz et al., 1982).
Identification and organization of ORFs
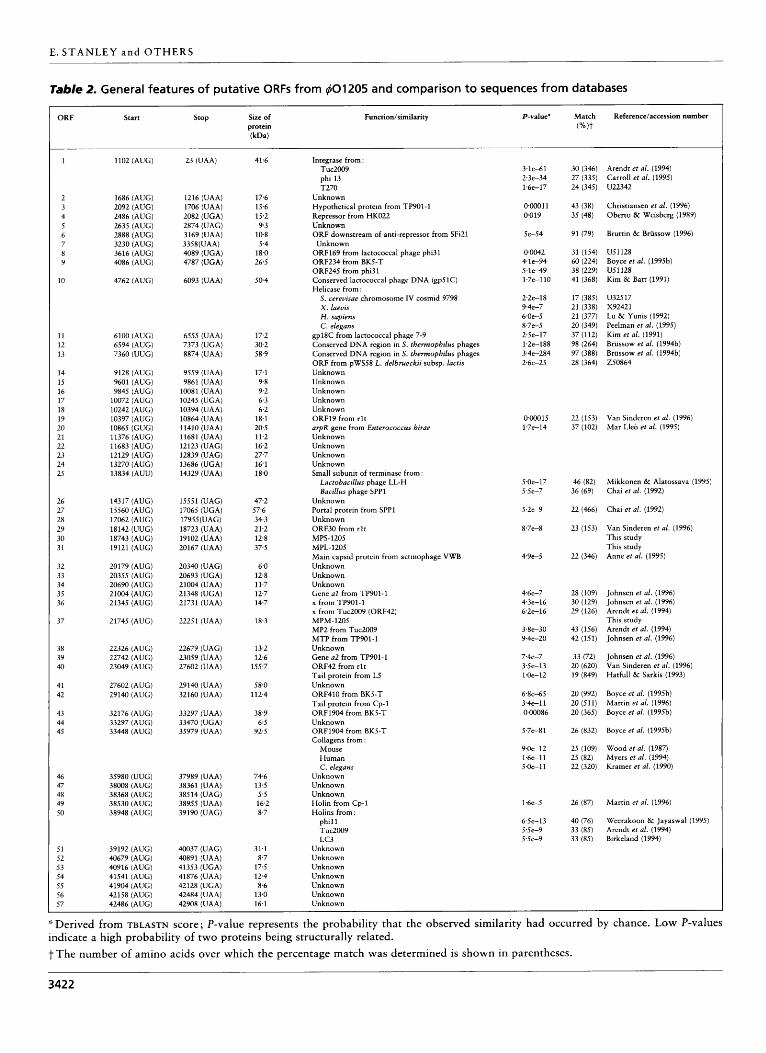
Analysis of the phage genome revealed the presence of 57 putative ORFs (Table 2). These were identified based on the adopted criteria that an ORF consists of at least 40 codons preceded by a potential Shine-Dalgarno sequence at an appropriate distance (6-15 bp) from one of the commonly used initiation codons (AUG, UUG, GUG) (Moran et al., 1982; Harley & Reynolds, 1987). One exception to this rule appears to be ORF2.5, which is preceded by a strong ribosome-binding site followed by the unusual initiation codon AUU (Sacerdot et al., 1996). The ORFs identified on the 40120.5 genome appear to be organized in two oppositely orientated clusters, consisting of four and 53 ORFs (Fig. 2). These two clusters are separated from each other on one side
3420
S. thermophilus phage 01205
1. TcGTTGCTCGTTCGAll'CC GCCACCCGACATTTTAAAGCCTTTAACCATCCATGCCGT~TA. 4 2. TCGZ'TGCTGGlTCGATTCC GGCAGGGGACATTTTIAAUCTTTAACCATGCGGTTTAAA 3. TCG7TGC7'GGITCGATTCC GG CAGG GG ACATTTTXA AXC CTTTAACCATAC GG TTTTA A
d T 1 7 integme
GCTACTlTGcCCCAAATCTG GCTAcT1TGcCCCAAATCTG f CTACTITGCCCCAAA TCTG
1 J
a 4 repressor morphogenesis
head tail *
lySis 43,075
I 2' 4 packaging
1 y sogen y replication I I I I I ' 20,000 ' 30,000 ' 4aoO0
'lo*OOO Dac L 5 7 9 b W W P 11 13 15 19 21 23 25 9 27 b 29 3235 IW- 38 40 42 44 I eD, 46 49 52 1 1 55 \ e 41/ I). 43 45 4 7 5 0 \ 5 ~ ~ 7 I)) b t )
48 10 51 b D p r i m e 1 tail holins
proteins major
structural proteins
6 8 10 12
helicase
18
Fig. 2. Schematic representation of the 401205 genome with i t s putative ORFs, features and assumed functions. The identified ORFs are numbered and indicated by open arrows or arrowheads which point in the same direction as the orientation of the ORF. Filled arrows represent ORFs found to be homologous to sequences derived from lactococcal phages. The two ORFs contained within the conserved DNA region found in many S. thermophilus phages (Brilssow eta/., 1994b) are indicated as hatched arrows. The pac site and direction of packaging are indicated by a bent arrow. For further explanation, see text.
-.---- --1.11 * >
GCCAGCGGACATTTTAAAGCCTTT AACCATGCCCTTTTAAGC
+ tRNA -
core L19
Figrn 3. (a) Schematic diagram of the attB region in CNRZ1205. Dashed arrows depict the imperfect direct repeat located within and extending beyond the core sequence of the integrated phage. The nucleotide sequence of the core site is given in bold face while italicized sequence depicts chromosomal DNA immediately downstream of the core sequence. The position of the tRNAArg and L79 genes as well as the core region are indicated by plain arrows. Direction of the arrows corresponds to the orientation of the genetic elements. (b) Alignment of the nucleotide sequence of the regions containing attB in 5. thermophilus strains CNRZ1205 (l), CNRZ385 (2) and ND16 (3). The 40 bp core region is boxed and depicted in bold face. Mismatches relative to the CNRZ1205 core sequence are underlined. Italicized sequence depicts the chromosomal DNA on either side of the core site.
342 1
E. S T A N L E Y a n d OTHERS
Table 2. General features of putative ORFs from 401205 and comparison to sequences from databases
ORF Start StOD Size of
- __
1
2 3 4 5 6 7 8 9
10
11 12 13
14 15 16 17 18 19 20 21 22 23 24 25
26 27 28 29 30 31
32 33 34 35 36
37
38 39 40
41 42
43 44 45
46 47 48 49 50
51 52 53 54 55 56 57
Function/similarity P-value' Match Reference/accession number ("/OH
1102 (AUG)
1686 (AUG) 2092 (AUG) 2486 (AUG) 2635 (AUG) 2888 (AUG) 3230 (AUG) 3616 (AUG) 4086 (AUG)
4762 (AUG)
6100 (AUG) 6594 (AUG) 7360 (UUG)
9128 (AUG) 9601 (AUG) 9845 (AUG)
10072 (AUG) 10242 (AUG) 10397 (AUG) 10865 (GUG) 11376 (AUG) 11683 (AUG) 12129 (AUG) 13270(AUG) 13834 (AUU)
14317(AUG) 15560 (AUG) 17062 (AUG) 18142 (UUG) 18743(AUG) 19121 (AUG)
20179 (AUG) 20355 (AUG) 20690 (AUG) 21004 (AUG) 21345(AUG)
21745(AUG)
22326(AUG) 22742 (AUG) 23049 (AUG)
27602(AUG) 29140 (AUG)
32176 (AUG) 33297 (AUG) 33448 (AUG)
35980 (UUC) 38008 (AUG) 38368 (AUG) 38530 (AUG) 38948 (AUG)
39192 (AUG) 40679 (AUG) 40916 (AUG) 41541 (AUG) 41904 (AUG) 42158 (AUG) 42486 (AUG)
23 (UAA)
1216 (UAA) 1706 (UAA) 2082 (UGA) 2874 (UAG) 3169 (UAA) 3358(UAA) 4089 (UGA) 4787 (UGA)
6093 (UAA)
6555 (UAA) 7373 (UGA) 8874 (UAA)
9559 (UAA) 9861 (UAA)
10081(UAA) 10245(UGA) 10394 (UAA) 10864(UAA) 11410(UAA) 11681 (UAA) 12123 (UAG) 12839 (UAG) 13686(UGA) 14329 (UAA)
15551 (UAG) 17065(UGA) 17955(UAG) 18723 (UAA) 19102(UAA) 20167(UAA)
20340 (UAG) 20693 (UGA) 21004 (UAA) 21348(UGA) 21731(UAA)
22251 (UAA)
22679(UAG) 23059 (UAA) 27602(UAA)
29140 (UAA) 32160 (UAA)
33297(UAA) 33470(UGA) 35979 (UAA)
37989 (UAA) 38361(UAA) 38514 (UAG) 38955(UAA) 39190 (UAG)
40037 (UAG) 40891 (UAA) 41353(UGA) 41876(UAA) 42128(UGA) 42484(UAA) 42908(UAA)
41.6
17.6 15.6 15.2 9 3
10.8 5.4
18.0 26.5
50.4
17.2 302 58.9
17.1 9.8 9.2 6.3 6.2
18.1 205 11.2 16.2 277 16.1 18.0
472 57.6 343 21.2 128 37.5
6 0 12.8 11.7 12.7 14.7
18.3
13.2 12.6
155.7
58.0 112.4
38.9 6.5
92.5
74.6 13.5 5.5 16.2 8.7
31.1 8.7
175 124 8.6
13.0 16.1
Integrase from : Tuc2009 phi 13 T270
Unknown Hypothetical protein from TP901-1 Repressor from HK022 Unknown ORF downstream of anti-repressor from SFi21
ORF169 from lactococcal phage phi31 ORF234 from BK5-T ORF245 from phi31 Conserved lactococcal phage DNA (gp51C) Helicase from :
S. cereuisae chromosome IV cosmid 9798 X. laeuis H. sapiens C. elegans
Unknown
gpl8C from lactococcal phage 7-9 Conserved DNA region in S. thermophilus phages Conserved DNA region in S. thermophilus phages ORF from pWS58 L. delbrueckii subsp. lactis Unknown Unknown Unknown Unknown Unknown ORF19 from r l t arpR gene from Enterococcus hirae Unknown Unknown Unknown Unknown Small subunit of terminase from :
Lactobacillus phage LL-H Bacillus phage SPPl
Unknown Portal protein from SPPl Unknown ORF30 from r l t MPS- 1205
Main capsid protein from actinophage VWB Unknown Unknown Unknown Gene a1 from TP901-1 x from TP901-1 x from Tuc2009 (ORF42)
MP2 from Tuc2009 MTP from TP901-1 Unknown Gene a2 from TP901-1 ORF42 from r l t Tail protein from L5 Unknown ORF410 from BK5-T Tail protein from Cp-1 ORF1904 from BK5-T Unknown ORF1904 from BK5-T Collagens from :
MPL- 1205
MPM- 1205
Mouse Human C. elegans
Unknown Unknown Unknown Holin from Cp-1 Holins from :
phi11 Tuc2009 LC3
Unknown Unknown Unknown Unknown Unknown Unknown Unknown
3.1e-61 2.3e-34 1.6e-17
0.00011 0019
5-54
00042 41e-94 5 , 1 4 9 1.7-110
2.2e-18 9.4e-7 6.0-5 8.7e-5 25e-17 1.2-188 3.4-284 2.6-25
000015 1.7e-14
5.0e-17 5.5e-7
5.2e-9
8.7-8
49e-5
46e-7 43e-16 6.2e-16
3.8e-30 94e-20
74e-7 35-13 1.0e-12
6 , 8 6 5 3.4e-11 000086
5.7-81
9.0-12 1.6-11 5.0-11
1.6-5
6.5e-13 5.5-9 5.5e-9
30 (346) 27 (335) 24(345) U22342
43 (38) 35 (48)
91 (79)
31 (154) US1128 60 (224) 38 (229) US1128 41 (368)
Arendt et al. (1994) Carroll et al. (1995)
Christiansen et al. (1996) Oberto & Weisberg (1989)
Bruttin & Briissow (1996)
Boyce et al. (1995b)
Kim & Batt (1991)
17 (385) 21 (338) 21 (377) 20 (349) 37 (112) 98 (264) 97 (388) 28 (364)
U32.517 X92421 Lu & Yunis (1992) Peelman et al. (1995) Kim et al. (1991) Briissow et al. (1994b) Briissow et al. (1994b) 250864
22 (153) 37 (102)
Van Sinderen et al. (1996) Mar Lleo et al. (1995)
46 (82) 36 (69)
22 (466)
Mikkonen & Alatossava (1995) Chai et al. (1992)
Chai et al. (1992)
23 (153) Van Sinderen et al. (1996) This study This study Anne et al. (1995) 22 (346)
28 (109) 30 (129) 29 (126)
43 (156) 42 (151)
33 (72) 20 (620) 19 (849)
20 (992) 20 (511) 20 (365)
26 (832)
25 (109) 25 (82) 22 (320)
Johnsen et al. (1996) Johnsen et al. (1996) Arendt et al. (1994) This study Arendt et al. (1994) Johnsen et al. (1996)
Johnsen et al. (1996) Van Sinderen et al. (1996) Hatfull & Sarkis (1993)
Boyce et al. (1995b) Martin et al. (1996) Boyce et al. (1995b)
Boyce et al. (1995b)
Wood et al. (1987) Myers et a!. (1994) Kramer et al. (1990)
26 (87)
40 (76) 33 (85) 33 (85) Birkeland (1994)
Martin et a[. (1996)
Weerakoon & Jayaswal (1995) Arendt et al. (1994)
'' Derived from TBLASTN score; P-value represents the probability that the observed similarity had occurred by chance. Low P-values indicate a high probability of two proteins being structurally related. ?The number of amino acids over which the percentage match was determined is shown in parentheses.
3422

S . thermophilus phage 01205
Fig. 4. Alignment of parts of the putative integrase from 401205 (indicated here as 1205-1) and the integrases from T270 (accession number U22342), phi 13 (Carroll et a/., 1995) and Tuc2009 (Van de Guchte et a/., 1994) (top to bottom, respectively). The four invariant amino acids are indicated by asterisks. Identical amino acids are boxed. Numbers at the left hand margin refer to the amino acid position in each of the depicted proteins.
HK022 29 QADLAVRLKVTPKAISKWFNGESIPRKDKMESLASVL GTT AAYLHGYADDDGITVNHLSRSNDYYRVDVLDVQASAGPGT~SNEFIEKIR-AIEYTTEQ 127
1205-4 37 QKELGIKL--GKSALSQWSGKSIPDQDKLVLLSKTL GVS EAWLMGYDTNESTDVKKESIDLSNLREKVVMFDGKPLSDDD~K-IEQIIRLSIEVTGDE 133 Q :L:::L A:S: :G SIP :DK: L: L G : A L GY ::: V: S R V: :: : :: V : : : 1R :IE T ::
=a====== +++ ========
helix turn h e l i x
Fig. 5. Alignment of the N-terminal region of the repressor from the lambdoid phage HK022 (Oberto & Weisberg, 1989) and ORF4 from 401205 (indicated here as 1205-4). Amino acid identities are indicated by the common amino acid and conservative replacements are indicated by colons (:). The potential helix-turn-helix motif is indicated. Numbers refer to the amino acid position.
by the attachment site located between ORFl and ORF57, and on the other side by what is believed to be a non-coding region encompassing regulatory elements which control the genetic switch and is located between ORF4 and ORF.5 (see below)(Fig. 2).
Similarity searches of the 401205 nucleotide sequence and deduced proteins with sequence data bases
The nucleotide sequences and the deduced amino acid sequences of the putative ORFs from 401205 were compared to the contents of the available databases (see Methods). Twenty-seven of the 57 deduced proteins showed significant similarity with sequences in the databanks. Deduced proteins which contain interesting motifs or showing similarity to proteins to which a function could be assigned are discussed below (see also Table 2). ORFI. The deduced protein specified by this ORF shows significant similarity to members of the integrase family of recombinases, e.g. the integrases from the Lacto- coccus lactis phage Tuc2009 (Arendt et al., 1994), the staphylococcal phage phi 13 (Carroll et al., 1995), and T270, a phage from Streptococcus pyogenes (accession number U22342). Similarity was particularly high in the C-terminal region, which is believed to contain the catalytic site of this family of integrases and includes four highly conserved amino acids (Abremski & Hoess, 1992) (Fig. 4).
ORF4. The deduced product of ORF4 exhibits limited similarity to the N-terminal part of the 243 amino acid repressor of the lambdoid phage HK022 (Oberto & Weisberg, 1989). This region of similarity includes a putative helix-turn-helix motif (Dodd & Egan, 1990) , which suggests that it may be involved in DNA binding (Fig. 5) . This similarity and the overall organization of 401205 would favour the assumption that ORF4 specifies a repressor involved in the genetic switch of 401205. ORF4 is the first in a cluster of four ORFs which also includes the putative integrase gene, having an orientation opposite to that of the remaining ORFs. This arrangement suggests a life-cycle-specific organization reminiscent of that found in several tem- perate lactococcal small isometric-headed phages, e.g. r l t (Van Sinderen et al., 1996; Nauta et al., 1996), Tuc2009 (D. van Sinderen, unpublished) and BK.5-T (Boyce et al., 1995b). The putative repressor of 401205 differs from other known repressors, e.g. Tuc2009 (Van de Guchte et al., 1994), P22 (Sauer et al., 1981), and A (Horii et al., 1981) in that it does not contain the highly conserved motifs which are present in proteins exhibiting RecA-mediated autocleavage (Horii et al., 1981; Sauer et al., 1981). The absence of such motifs suggests that the genetic switch in 401205 is controlled by a mechanism different from that of most other phages and may even be RecA-independent.
ORF9. The protein specified by ORF9 exhibits a signifi- cant resemblance to the products of ORF234 from the
3423

E. S T A N L E Y a n d OTHERS
+ * r e * * + *
Fig. 6. Alignment of ORF9 from $01205 (indicated here as 1205-9), ORF234 from BK5-T (Boyce et a/., 1995b) and ORF245 from phi31 (accession number U51128) (depicted top to bottom, respectively). The amino acids which form a putative nucleotide- binding site (Walker A motif) are indicated by asterisks. Identical amino acids are boxed. Numbers refer to amino acid position.
M I C. elegans T P C R D E A D
A R C D
P R I G R
ORFlO 001205 s. cerevisiae x. laevis C. el-
Fig. 7. Alignment of parts of ORFlO from $01205, the derived amino acid sequence of a region similar to the DEAD box family of helicases from the Saccharomyces cerevisae chromosome IV cosmid 9798 (accession number U32517), the RNA helicase p54 from Xenopus laevis (accession number X92421) and a putative RNA helicase from Caenorhabditis elegans (Peelman et a/., 1995) (depicted top to bottom, respectively). Identical amino acids are boxed and the eight conserved motifs (Schmid & Linder, 1992) are indicated by bold letters below the corresponding aligned sequence. Numbers refer to amino acid positions.
lactococcal phage BK5-T (Boyce et al., 1995b) and ORF245 from the lactococcal phage phi31 (accession number U51128) (Fig. 6). No function has been assigned to either of these ORFs. Interestingly, however, all these ORFs contain a putative ATP/GTP-binding site motif A (Walker et al., 1982) (Fig. 6), indicating that ATP may be essential for their activity.
ORFlO and ORF11. The DNA region encompassing these ORFs shows significant homology to a conserved region of the lactococcal phage 7-9 (Kim & Batt, 1991; Kim et al., 1991), which has been suggested to play a role in regulating phage replication (Kim & Batt, 1991). In addition, the deduced product of ORFlO exhibits significant similarity to the protein specified by an ORF
from the Saccbaromyces cerevisae chromosome IV cosmid 9798 (accession number U32517), as well as proteins from a number of other eukaroytes, e.g. Xenopus laevis (accession number X92421), Homo sapiens (Lu & Yunis, 1992) and Caenorbabditis elegans (Peelman et al., 1995), all of which are thought to belong to the DEAD-box-containing family of DNA/RNA helicases (for a review see Schmid & Linder, 1992). Examination of ORFlO revealed the presence of most of the sequence motifs characteristic of the superfamily I1 of DNA and RNA helicases (Fig. 7). The protein product of ORFlO may therefore possess helicase activity.
ORFl2 and ORF13. The DNA region harbouring ORF12 and ORF13 is almost identical to a conserved DNA
3424

S . thermophilus phage 01205
region found in a large percentage of phages infecting S. thermophilus (Brussow et al., 1994b). In addition, the protein specified by ORF13 exhibits significant simi- larity to the putative primase from the cryptic plasmid pWS58 from Lactobacillus delbrueckii subsp. lactis (accession number 250864). However, this similarity is limited to the C-terminal region and ORF13 does not appear to contain the conserved motifs found in the N- terminal region of many primases (Versalovic & Lupski, 1993).
ORF25. The product of this ORF shows significant similarity to the small subunit of the terminase protein complex from a number of phages, including the Bacillus subtilis phage SPPl (Chai et al., 1992) and the Lacto- bacillus delbrueckii phage LL-H (Mikkonen & Alatossava, 1995).
ORF27. The product of this ORF exhibits significant similarity to the portal protein of the Bacillus subtilis phage SPPl (Chai et al., 1992), which is a structural component of the phage responsible for forming the entrance to the head and determining the amount of DNA to be packaged (Dube et al., 1993).
ORF3EORF39. The DNA region encoding ORF35- ORF39 exhibits significant homology to a region of the lactococcal small isometric-headed phages TP901-1 and Tuc2009 (Table 2). ORF37 encodes the major structural protein MPM-1205 (see below) and is similar to the structural proteins MTP and MP2 specified by TP901-1 (Johnsen et al., 1996) and Tuc2009 (Arendt et al., 1994), respectively, which are believed to encode major tail proteins (Johnsen et al., 1996). When compared to the completely sequenced Tuc2009, the region of homology is seen to extend from ORF33 to ORF40 (D. van Sinderen, unpublished).
ORF49 and ORF50. The product specified by ORF49 is similar to the holin from the streptococcal phage Cp-1 (Martin et al., 1996), while the protein product of ORF5O is similar to the holin of the staphylococcal phage phi11 (Weerakoon & Jayaswal, 1995) and the lactococcal phages Tuc2009 (Arendt et al., 1994) and LC3 (Birkeland, 1994). Holins are usually small proteins of 60-145 amino acids in size and have a hydrophilic N- terminal, a highly polar, charged C-terminal and two membrane-spanning domains separated by a p-turn (Young & Blasi, 1995). The deduced proteins specified by both ORF49 and ORF50 appear to have all of these characteristics, thus corroborating the assumption that they may function as holins.
Structural components of #01205
SDS-PAGE of the phage identified three major and several minor structural proteins. The estimated mol- ecular masses of the large (MPL-1205), medium (MPM- 1205) and small (MPS-1205) proteins as deduced from their electrophoretic mobility each were 41 kDa, 25 kDa and 13 kDa, respectively. N-terminal amino acid sequencing of the three major proteins revealed the
following (N-termini on the left) : MPL-1205, GLIYDKVTASNIAGYFNAL ; MPM-1205, ADNNEA- LLGKDKILM; MPS-1205 : PTTTIFDTSNIVRSL. The deduced N-terminal sequences of the deduced proteins specified by ORFs 30, 31 and 37 exactly matched the determined N-terminal sequences of MPS- 1205, MPL-1205 and MPM-120.5, respectively. The molecular masses of the three major proteins as deduced by sequence analysis were 37.5 kDa (MPL-1205), 18 kDa (MPM-1205) and 12.8 kDa (MPS-1205). The rather large discrepancy between the molecular mass of MPM-1205 as predicted by SDS-PAGE (i.e. 25 kDa) and that determined by sequence data (i.e. 18 kDa) is not easily explained by abnormal physical properties of this protein and may be due to covalent linkage to another protein. Based on similarities discussed above, ORF31 is likely to encode a capsid protein whereas ORF37 probably encodes a major tail protein.
DISCUSSION
401205 is the first phage infecting S. thermophilus for which the complete nucleotide sequence has been determined. The genome of 401205 appears to contain 57 ORFs and only a small portion of the genome appears to be non-coding. The genomic organization of the 401205 indicates that this phage may have a life-cycle- specific organization similar to that observed for the temperate small isometric-headed lactococcal phages r l t (Van Sinderen et al., 1996; Nauta et al., 1996), Tuc2009 (D. van Sinderen, unpublished) and probably also the partially sequenced BKS-T (Boyce et al., 1995b). Consistent with this life-cycle-specific organization is the finding that ORFl of 401205 is likely to specify the integrase, whereas the protein specified by ORF4 is a probable candidate for the repressor of 401205. Lysogenization by 401205 is assumed to have resulted from integration of the phage genome into the bacterial chromosome via a site-specific recombination event mediated by the putative integrase, encoded by ORF1. The attP site is located close to the 3’ end of the putative integrase gene, which is the typical organization of this type of protein (Leong et al., 1986; Goodman & Scocca, 1989 ; Lillehaug & Birkeland, 1993 ; Boyce et al., 1995b ; Van de Guchte et al., 1994). Analysis of the sequence of the DNA regions containing attP, attL, attR and attB allowed the identification of a 40 bp common sequence which is expected to represent the core region. The size of the common core sequence necessary for recom- bination to occur varies from a 182 bp region in the temperate phage HP1 from Haemophilus infiuenzae (Goodman & Scocca, 1989) to a 3 bp core in the Streptomyces temperate phage 4C31 (Rausch & Lehmann, 1991). The phage was found to integrate via the 3’ end of a putative tRNAArg. Genes encoding tRNAs are frequently used as insertion sites in phages (for some examples see Reiter et al., 1989; Hauser & Scocca, 1990; Campbell, 1992). The region within which the attachment site is located is often AT-rich and contains inverted repeats (Landy & Ross, 1977; Raya et
3425

E. STANLEY a n d OTHERS
al., 1992; Lillehaug & Birkeland, 1993 ; Christiansen et al., 1994). This is also true for the attP region of 40120.5, which exhibited an AT content of 75 mol YO as opposed to an overall value for the phage genome of 62 molY0. Analysis of the sequence revealed an imperfect direct repeat located within and extending beyond the 40 bp sequence into the integrated phage (Fig. 3a) as well as a number of direct and inverted repeats upstream and downstream from attP.
Two ORFs involved in replication have tentatively been identified in 401205. ORFlO contains some charac- teristics common to helicases and ORF13 may encode a primase. Directly downstream of the potential primase, a 250 bp non-coding region rich in direct and inverted repeats is observed and this may constitute the rep- lication origin of the phage. Work is in progress to confirm this hypothesis.
The pac site of 401205 was shown to be located within ORF25, which encodes the putative small subunit of the terminase protein complex. A similar position for pac has been observed for other phages, e.g. P22 (Black, 1989) and SPPl (Chai et al., 1992). The terminase enzyme is responsible for the initial cut at the pac site in the first round of packaging. It is commonly composed of two subunits: the small subunit is approximately 160-190 amino acids long and the large subunit is 420-650 residues in length. The small subunit of the terminase is thought to bind to the pac site and also to bind and hydrolyse ATP. The large terminase subunit, in complex with the small subunit, appears to bind to the prohead and may be involved in cutting the concatemeric phage molecules (for a review see Black, 1989). The genes encoding the small and large subunits of the terminase are frequently genetically linked (Black, 1989; Chai et al., 1992), which would make ORF26 a possible candidate to encode the large subunit because of its size and its downstream position from the putative small-subunit-encoding ORF25.
The lysis cassette of 40120.5 seems to consist of two putative holins (ORF49 and ORF50). A situation where two holins are required for the lysis of one phage has not previously been described in the literature. As yet, similarity searches have not identified the lysin of #01205. However, the lysin gene is often located immediately downstream of the holin, indicating that ORF51 may encode the lysin of 40120.5.
DNA hybridization studies by Fayard et al. (1993) and Briissow & Bruttin (1995) indicate that q50120.5 is closely related to other S. thermophilus phages and is therefore a suitable representative with which to study phage biology of ' thermophilus ' phages. 401205 displays remarkable homologies to a number of small isometric- headed lactococcal phages belonging to the P335 species (Jarvis et al., 1991), including phi31, phage 7-9, and the temperate phages Tuc2009, r l t and TP901-1. In ad- dition, 401205 exhibits homology to the temperate lactococcal small isometric-headed phage BK5-T, which has been grouped as a separate phage species, although
it contains DNA regions homologous to temperate phages of the P335 group (Boyce et al., 1995a). An interesting homology is found throughout a cluster of ORFs which are likely to be involved in phage mor- phogenesis in 401205, Tuc2009 and TP901-1. The conserved gene order between ORF33 and ORF40 from 401205 and between ORF39 and ORF46 from Tuc2009 suggests that 40120.5 may be closely related to the P335 and BK5-T groups. S. thermophilus and L. lactis share a common growth medium (milk) and although inter- generic cross-infection has not been described it is possible that horizontal gene transfer may have occurred between some phages of streptococcal and lactococcal origin. Alternatively, a common ancestor phage infecting one species may have acquired the capacity to infect the other bacterial species. Such intergeneric gene transfer events may not be unique, as this phenomenon may also explain the presence of very homologous IS sequences found in various lactic acid bacteria (Guedon et al., 1995).
ACKNOWLEDGEMENTS
We wish to extend our gratitude to Mary Moryis-Quinn and Sinead Geary for DNA sequencing. We thank Aine Healy for the synthesis of oligonucleotides and the N-terminal sequence determinations. We are grateful to M. Desmazeaud for providing S. thermophilus CNRZ385 and Dr S. Sannabhadti for S. thermophilus NDI6. We would like to thank Sylvain Moineau and Aat Ledeboer for critically reading the manu- script.
This work was partly financed by Quest International and also by European Community Biotechnology grants (contracts Bio4-CT96-0402 and Bio4-CT96-0439) to G. F. F. and D. V. S. D.V.S is a recipient of a long-term EMBO fellowship (ALTF 43 1-1995).
REFERENCES
Abremski, K. E. & Hoess, R. H. (1992). Evidence for a second conserved arginine residue in the integrase family of recom- bination proteins. Protein Eng 5 , 87-91. Altschul, 5. F., Gish, W., Miller, W., Myers, E. W. & Lipman, D. 1. (1990). Basic local alignment search tool. J Mol Biol215,403410. Anne, J., Fiten, P., Van Mellaert, L., Joris, B., Opdenakker, G. & Eyssen, H. (1995). Analysis of the open reading frames of the main capsid proteins of actinophage VWB. Arch Viroll40,1033-1047. Arendt, E. K., Daly, C., Fitzgerald, G. F. & Van de Guchte, M. (1994). Molecular characterization of lactococcal bacteriophage Tuc2009 and identification and analysis of genes encoding lysin, a putative holin, and two structural proteins. Appl Environ Microbiol60, -2875-1883. Benbadis, L., Faelen, M., Slos, P., Fazel, A. & Mercenier, A. (1990). Characterization and comparison of virulent bacteriophages of Streptococcus thermophilus isolated from yogurt. Biochemie 72,
Birkeland, N. K. (1994). Cloning, molecular characterization, and expression of the genes encoding the lytic functions of the lactococcal bacteriophage 4LC3 : a dual lysis system of modular design. Can J Microbiol40, 658-665. Birnboim, H. C. & Doly, 1. (1979). A rapid alkaline extraction
855-862.
3426

S. therrnophilus phage 01205
procedure for screening recombinant plasmid DNA. Nucleic Acids Res 7 , 1513-1523.
Black, L. W. (1989). DNA packaging in ds DNA bacteriophages. Annu Rev Microbiol43, 267-292.
Boyce, 1. D., Davidson, B. E. & Hillier, A. J. (1995a). Sequence analysis of the Lactococcus lactis temperate bacteriophage BK5- T and demonstration that the phage DNA has cohesive ends. Appl Environ Microbiol 61, 4089-4098.
Boyce, J. D., Davidson, 6. E. & Hillier., A. J. (1995b). Spontaneous deletion mutants of the Lactococcus lactis temperate bacterio- phage BK5-T and localization of the BK5-T attP site. Appl Environ Microbiol61,41054109.
Bradley, D. E. (1967). A review : ultrastructure of bacteriophage and bacteriocins. Bacteriol Rev 31, 230-314.
BrUssow, H. & Bruttin, A. (1995). Characterization of a temperate Streptococcus thermophilus bacteriophage and its genetic re- lationship with lytic phages. Virology 212, 632-640.
BrUssow, H., Frbmont, M., Bruttin, A., Sidoti, J., Constable, A. & Fryder, V. (1994a). Detection and classification of Streptococcus thermophilus bacteriophages isolated from industrial milk fer- mentation. Appl Environ Microbiol60, 4537-4543.
BrUssow, H., Probst, A., Frbmont, M. & Sidoti, 1. (1994b). Distinct Streptococcus thermophilus bacteriophages share an extremely conserved DNA fragment. Virology 200, 854-857.
Bruttin, A. & BrUssow, H. (1996). Site-specific spontaneous deletions in three genome regions of a temperate Streptococcus thermophilus phage. Virology 219, 96-104.
Campbell, A. M. (1992). Chromosomal insertion sites for phages and plasmids. J Bacteriol 174,7495-7499.
Carminati, D. & Giraffa, G. (1992). Evidence and characterization of temperate bacteriophage in Streptococcus salivarius subsp. thermophilus St18. J Dairy Res 59, 71-79.
Carroll, D., Kehoe, M. A,, Cavanagh, D. & Coleman, D. C. (1995). Novel organization of the site-specific integration and excision recombination functions of the Staphylococcus aureus serotype F virulence-converting phages 413 and 442. Mol Microbiol 16,
Chai, S., Bravo, A., LUder, G., Nedlin, A., Trautner, T. A. & Alonso, J. C. (1992). Molecular analysis of the Bacillus subtilis bac- teriophage S P P l region encompassing genes 1 to 6 . J Mol Biol224,
Christiansen, B., Johnsen, M. G., Stenby, E., Vogensen, F. K. & Hammer, K. (1994). Characterization of the lactococcal temperate phage TP901-1 and its site specific integration. ] Bacteriol 176,
Christiansen, B., Brendsted, L., Vogensen, F. K. & Hammer, K. (1996). A resolvase-like protein is required for the site-specific integration of the temperate lactococcal bacteriophage TP901-1. ] Bacteriol 178, 5164-5173.
Czulak, J., Hammond, L. A. & Meharry, H. J. (1954). A new method for Cheddar cheese manufacture. Aust Dairy Rev 22, 18-22.
Dodd, 1. B. & Egan, J. B. (1990). Improved detection of helix- turn-helix DNA-binding motifs in protein sequences. Nucleic Acids Re5 18, 5019-5026.
Dube, P., Tavares, P., Lurz, R. & van Heel, M. (1993). The portal protein of bacteriophage S P P 1 : a DNA pump with 13-fold symmetry. EMBO J 12, 1303-1309.
Fayard, B., Haeflinger, M. & Accolas, I.-P. (1993). Interaction of temperate bacteriophage of Streptococcus salivarius subsp. thermophilus with lysogenic indicators affect phage DNA re- striction patterns on host ranges. J Dairy Res 60, 385-399.
877-893.
87-102.
1069-1 076.
Francki, R. 1. B., Fauquet, C. M., Knudson, D. L. & Brown, F. (1991). Classification and nomenclature of viruses. Fifth report of the International Committee on Taxonomy of Viruses. Arch Virol2, 1-450.
Garvey, P., Van Sinderen, D., Twomey, D. P., Hill, C. & Fitzgerald, G. F. (1995). Molecular genetics of bacteriophage and natural phage defence systems in the genus Lactococcus. Znt Dairy J 5 ,
Goodman, 5. D. & Scocca, J. J. (1989). Nucleotide sequence and expression of the gene for the site-specific integration protein from bacteriophage HP1 of Haemophilus infiuenzae. J Bacteriol 171,4232-4240. GuBdon, G., Bourgoin, F., PBbay, M., Roussel, Y., Colmin, C., Simonet, 1. M. & Decaris, B. (1995). Characterization and dis- tribution of two insertion sequences, IS1191 and iso-IS981, in Streptococcus thermophilus : does intergeneric transfer of in- sertion sequences occur in lactic acid bacteria co-cultures ? Mol Microbiol 16, 69-78.
Harley, C. B. & Reynolds, R. P. (1987). Analysis of E . coli promoter sequences. Nucleic Acids Res 15, 2343-2361.
Hatfull, G . F. & Sarkis, G. J. (1993). DNA sequence, structure, and gene expression of mycobacteriophage L.5: a phage system for mycobacterial genetics. Mol Microbiol7, 395405.
Hauser, M. A. & Scocca, 1.1. (1990). Location of the host attachment site for phage H P 1 within a cluster of Haemophilus influenzae tRNA genes. Nucleic Acids Res 18, 5305.
Horii, T., Ogawa, T., Nakatani, T., Hase, T., Matsubara, H. & Ogawa, H. (1981). Regulation of SOS functions: purification of E . coli LexA protein and determination of its specific site cleaved by the RecA protein. Cell 27, 515-522.
Jarvis, A. W., Fitzgerald, G. F., Mata, M., Mercenier, A., Neve, H., Powell, I. B., Ronda, C., Saxelin, M. & Teuber., M. (1991). Species and type phages of lactococcal bacteriophages. Zntervirology 32,
Jin, 5. & Benedik, M. J. (1994). Sequences of the Serratia marcescens rplS and trmD genes. Gene 145, 147-148.
Johnsen, M. G., Appel, K. F., Madsen, P. L., Vogensen, F. K., Hammer, K. & Arnau, J. (1996). A genomic region of the lactococcal temperate bacteriophage TP901-1 encoding major virion proteins. Virology 218, 306-315.
Kiesewetter, S., Fischer, W. & Sprinzl, M. (1987). Sequence of three minor tRNAArg from E . coli. Nucleic Acids Res 15, 3184.
Kilpper-Bllz, R., Lathe, G. & Lecocq, 1. P. (1982). Nucleic acid hybridization of Group N and group D streptococci. Curr Microbiol7, 245-250.
Kim, S.G. & Batt, C.A. (1991). Identification of a nucleotide sequence conserved in Lactococcus lactis bacteriophages. Gene
Kim, 5. G., Bor, Y.-C. & Batt, C. A. (1991). Bacteriophage resistance in Lactococcus lactis ssp. lactis using antisense ri bonucleic acid. ] Dairy Sci 75, 1761-1767.
Klaenhammer, T. R. & Fitzgerald, G. F. (1994). Bacteriophages and bacteriophage resistance. In Genetics and Biotechnology of Lactic Acid Bacteria, pp. 106-168. Edited by M. J. Gasson & W. de Vos. Glasgow : Blackie. Komine, Y., Adachi, T., Inokuchi, H. & Ozeki, H. (1990). Genomic organization and physical mapping of the transfer RNA genes in Escherichia coli K12. J Mol Biol212, 579-598.
Kramer, J. M., French, R. P., Park, E. C. &Johnson, 1. J. (1990). The Caenorhabditis elegans rol-6 gene, which interacts with the sgt-1 collagen gene to determine organismal morphology, encodes a collagen. Mol Cell Biol 10, 2081-2089.
905-947.
2-9.
98,95-100.
3427

E. S T A N L E Y a n d OTHERS
Krusch, U., Neve, H., Luschei, B. & Teuber, M. (1987). Charac- terization of virulent bacteriophage of Streptococcus salivarius subsp. thermophilus by host specificity and electron microscopy. Kiel Milchwirtsch Forschungsber 39, 155-167. Laemmli, U. K. (1970). Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 227, 680-685. Landy, A. & Ross, W. (1977). Viral integration and excision. Structure of the A att sites. Science 197, 1147-1160. Larbi, D., Colmin, C., Rouselle, L., Decaris, B. & Simonet, I.-M. (1 990). Genetic and biological characterization of nine Strepto- coccus salivarius subsp. thermophilus bacteriophages. Lait 70,
Leong, J. M., Nunes-DUby, 5. E., Oser, A. B., Lesser, C. F., Youderian, P., Susskind, M. M. & Landy, A. (1986). Structural and regulatory divergence among site-specific recombination genes of lambdoid phage. J Mol Biol 189, 603-613. Lillehaug, D. & Birkeland, N.-K. (1993). Characterization of genetic elements required for site-specific integration of the temperate lactococcal bacteriophage 4LC3 and construction of integration- negative 4LC3 mutants. J Bacteriol 175, 1745-1755. Lu, D. & Yunis, 1. J. (1992). Cloning, expression and localization of an RNA helicase gene from a human lymphoid cell line with chromosomal breakpoint 11q23.3. Nucleic Acids Res 20,
Mar Lleb, M., Fontana, R. & Solioz, M. (1995). Identification of a gene (arpU) controlling muramidase-2 export in Enterococcus hirae. J Bacteriol 177, 5912-5917. Martin, A. C., Lopez, R. & Garcia, P. (1996). Analysis of the complete nucleotide sequence and functional organization of the genome of Streptococcus pneumoniae bacteriophage Cp-1. J Virol70,3678-3687. Mata, M. & Ritzenthaler, P. (1988). Present state of lactic acid bacteria phage taxonomy. Biochimie 70, 395-399. Mercenier, A. (1 990). Molecular genetics of Streptococcus thermo- philus. FEMS Microbiol Rev 87, 61-78. Mikkonen, M. & Alatossava, T. (1995). A group I intron in the terminase gene of Lactobacillus delbrueckii subsp. lactis phage LL-H. Microbiology 141, 2183-2190. Moran, C. P., Lang, N., le Grice, S. F. J., Lee, G., Stephens, M., Sonenshein, A. L., Pero, J. & Losick, R. (1982). Nucleotide sequences that signal the initiation of transcription and trans- lation in Bacillus subtilis. Mol Gen Genet 186, 339-346. Myers, J. C., Yang, H., D'lppolito, 1. A,, Presente, A., Miller, M. K. & Dion, A. 5. (1994). The triple-helicase region of human type XIX collagen consists of multiple collagenous subdomains and exhibits limited sequence homology to alpha I (XVI). ] Biol Chem
Nauta, A., Van Sinderen, D., Karsens, H., Smit, E., Venema, G. & Kok, 1. (1996). Inducible gene expression mediated by a repressor-operator system isolated from Lactococcus lactis bac- teriophage rlt. Mol Micro 19, 1331-1341. Neve, H., Krush, U. & Teuber, M. (1989). Classification of virulent bacteriophage of Streptococcus salivarius subsp. thermophilus isolated from yoghurt and Swiss-type cheese. Appl Microbiol Biotechnol30, 624-629. Oberto, J. & Weisberg, R. A. (1989). Structure and function of the nun gene and the immunity region of the lambdoid phage HK022. J Mol Biol207, 675-693. Ochman, H., Medhora, M. M., Garza, D. & Hartl, D. L. (1990). Amplification of flanking sequence by inverse PCR. In PCR Protocols: a Guide to Methods and Applications, pp. 219-227. New York: Academic Press.
107-1 16.
1967-1972.
269, 18546-18557.
Pearson, W. R. & Lipman, D. 1. (1988). Improved tools for biological sequence comparison. Proc Natl Acad Sci USA 85, 2444-2448. Peelman, L. 1.. Chardon, P.8 Nunes, M., Renard, C., Geffrotin, C., Vaiman, M., Van Zeveren, A., Coppieters, W., Van de Weghe, A. & Bouquet, Y. (1995). The BAT1 gene in the M H C encodes an evolutionary conserved putative nuclear RNA helicase of the DEAD family. Genomics 26, 210-218. Prevots, F., Relano, P., Mata, M. & Ritzenthaler, P. (1989). Close relationship of virulent bacteriophages of Streptococcus salivarius subsp. thermophilus at both the protein and the DNA level. J Gen Microbiol 135,3337-3344. Rausch, H. & Lehmann, M. (1991). Structural analysis of the actinophage 4C31 attachment site. Nucleic Acids Res 19,
Raya, R. R., Frernaux, C., De Antoni, G. L. & Klaenhammer, T. R. (1992). Site-specific integration of the temperate bacteriophage 4adh into the Lactobacillus gasseri chromosome and molecular characterization of the phage (attp) and bacterial (attB) at- tachment sites. J Bacteriol 174, 55844592. Reiter, W. D., Palm, P. & Yeats, 5. (1989). Transfer RNA genes frequently serve as integration sites for prokaryotic genetic elements. Nucleic Acids Res 17, 1907-1914. Sacerdot, C., Chiaruttini, C., Engst, K., Graffe, M., Millet, M., Mathy, N., Dondon, 1. & Springer, M. (1996). The role of the AUU initiation codon in the negative feedback regulation of the gene for translation factor IF3 in Escherichia coli. Mol Microbiol21,
Sambrook, J., Fritsch, E. F. & Maniatis, T. (1989). In Molecular Cloning: a Laboratory Manual, 2nd edn. Cold Spring Harbor, NY: Cold Spring Harbor Laboratory. Sauer, R. T., Pan, J., Hopper, P., Hehir, K., Brown, J. & Poteete, A. R. (1981). Primary structure of the phage P22 repressor and the gene c2. Biochemistry 20, 3591-3598. Sebastiani, H. & Jlger, H. (1 992). Bacteriophages of Streptococcus salivarius subsp. thermophilus : characterization of hereditary relationships and determination of virus-host interaction. Milchwissenschaft 48, 25-29. Schmid, 5. R. & Linder, P. (1992). D-E-A-D protein family of putative RNA helicases. Mol Microbiol6, 283-292. Schmidt, 1. & Subramanian, A. R. (1993). Sequence of the cyanobacterial tRNA-W in synechocystis PCC 6803 : requirement of 3' CCA attachment to the acceptor stem. Nucleic Acids Res 21, 2519. Slos, P., Bourquin, J.-C., Lemoine, Y. & Mercenier, A. (1991). Isolation and characterization of chromosomal promoters of Streptococcus salivarius subsp. thermophilus. Appl Environ Microbiol57, 1333-1339. Southern, E. M. (1975). Detection of specific sequences among DNA fragments separated by gel electrophoresis. J Mol Biol98,
Terzaghi, B. E. & Sandine, W. E. (1975). Improved media for lactic streptococci and their bacteriophages. Appl Microbiol 29,
Van de Guchte, M., Daly, C., Fitzgerald, G. F. & Arendt, E. K. (1994). Identification of int and attP on the genome of the lactococcal bacteriophage Tuc2009 and their use for site-specific plasmid integration in the chromosome of Tuc2009-resistant Lactococcus lactis MG1363. Appl Environ Microbiol 60, 2324-2329. Van Sinderen, D., Karsens, H., Kok, J., Terpstra, P., Ruiters, M. H. J., Venema, G. & Nauta, A. (1996). Sequence analysis and
5 1 87-5 189.
331-346.
503-517.
807-813.
3428

S. thermophilus phage 01205
molecular characterization of the temperate lactococcal bac- teriophage rl t . Mol Microbiol 19, 1343-1355. Venalovic, J. & Lupski, J. R. (1993). The Haemophilus influenza dnaG sequence and conserved bacterial primase motifs. Gene 136,
Walker, J. E., Sarasate, M., Runswick, M. J. & Gay, N. J. (1982). Distantly related sequences in the a- and /?-subunits of ATP synthase, myosin, kinases and other ATP-requiring enzymes and a common nucleotide binding fold. EMBO J 1, 945-951. Weerakoon, L. K. & Jayaswal, R. K. (1995). Sequence analysis of
281-286.
the region upstream of a peptidoglycan hydrolase-encoding gene from bacteriophage phi 11 of Staphylococcus aureus. FEMS Microbiol Lett 133, 9-15. Wood, L., Thericult, N. & Vogeli, G. (1987). Complete nucleotide sequence of the N-terminal domains of the murine alpha-1 type I11 collagen chain. Gene 61, 225-230. Young, R. & Bllsi, U. (1995). Holins: form and function in bacteriophage lysis. FEMS Microbiol Rev 17, 191-205.
Received 6 May 1997; revised 1 July 1997; accepted 7 July 1997.
3429
Related Documents











