DNA RESEARCH 3, 55-64 (:.996) Unique Telomeric Expression Site of Major-Surface-Glycoprotein Genes of Pneumocystis carinii Miki WADA and Yoshikazu NAKAMURA* Department of Tumor Biology, The Institute of Medical Science, The University of Tokyo, P.O. Takanawa, Tokyo 108, Japan (Received 4 April 1996) Abstract Major cell surface glycoproteins (MSG) of Pneumocystis carinii play a crucial role in the host-parasite interactions involved in P. carinii pneumonia in AIDS patients. Genes encoding MSGs are repeated, highly polymorphic, and distributed among all of the 14-15 chromosomes. Here we show, by BAL-31 exonuclease cleavage and DNA cloning experiments, that the unique expression site (previously termed UCS) of MSG genes located in the 500-kb chromosome is telomeric. The 11-kb genomic UCS fragment isolated and sequenced in this study contained one MSG coding sequence (termed msglO5), subtelomeric repetitive sequences and telomere-specific tandem repeats of TTAGGG oriented 5' to 3' towards the DNA end. Despite the N-terminal polymorphism, the C-terminal one-third sequence of MSG105 was identical to one of the known MSG-cDNAs, suggesting homologous recombination within the MSG coding sequences. These features closely resemble the Variant Surface Glycoprotein system of the protozoan parasite Trypanosoma brucei, suggesting that the genetic heterogeneity of MSGs is generated by recombination between the UCS expression site and multiple MSG genes by means of reciprocal exchange or gene conversion. Key words: Pneumocystis carinii; major surface glycoprotein; expression site; telomere 1. Introduction Pneumocystis carinii is an opportunistic pathogen that often causes fatal pneumonia in patients under immuno- suppressed or immunodeficient conditions such as AIDS, cancer chemotherapy or organ transplantation. More than 60% of AIDS patients suffer from P. carinii pneu- monia at some time during the course of the disease. 1 P. carinii is a fungus 2 ' 3 that is coated by abundant het- erogeneous major surface glycoproteins (MSGs) called MSG, 4 - 5 P115, 6 - 7 gpl20 8 or gpA. 9 ' 10 The MSGs are thought to play a crucial role in the host-parasite inter- action during P. carinii pneumonia because of the par- tial protection conferred by passive immunization with an anti-MSG monoclonal antibody against the progres- sion of P. carinii pneumonia, 11 the attachment of the or- ganism via interaction between MSGs and fibronectin on alveolar epithelial cells and macrophages, 12 ' 13 a specific T-cell response to MSGs after immunization and natu- ral infection, 14 and the uptake of P. carinii by alveolar macrophages mediated by the mannose receptor that tar- Communicated by Mituru Takanami * To whom correspondence shculd be addressed. Tel. +81- 3-5449-5307, Fax. +81-3-5449-5415, E-mail: [email protected] tokyo.ac.jp f The nucleotide sequence data reported in this paper will ap- pear in the DDBJ, GenBank said EMBL nucleotide sequence databases with the accession number D82031. gets the mannose moiety of MSG. 15 MSG is highly glycosylated with mannose and its molecular mass has been reported to be from 100 to 125 kDa. 6 ' 8 ' 16 It accounts for three-quarters of the total cel- lular protein when isolated from rats. 6 Genes encoding MSGs are repeated, highly polymorphic, and distributed among all of the 14-15 chromosomes. 4 ' 5 ' 7 ' 9 - 10 ' 17 ' 18 We showed that MSG gene expression is mediated by a DNA element termed the upstream conserved sequence, UCS. 19 The UCS element maps to a single chromosome of about 500 kb, is attached to expressed MSG genes, and encodes the sequence found at the 5' ends of most MSG mRNAs. The UCS is not highly repeated, but P. carinii populations contain many different MSG genes attached to the UCS, suggesting that the genetic heterogeneity of MSGs is generated by UCS-MSG recombination. 19 Here we show that the UCS locus is telomeric, suggesting that UCS-MSG recombination occurs by recombination be- tween the UCS expression site and multiple MSG genes by means of reciprocal exchange or gene conversion. 2. Materials and Methods 2.1. Organisms Pathogen-free athymic (rnu/rnu) nude rats were in- fected with P. carinii as described, 6 and P. carinii or- ganisms were prepared from bronchoalveolar lavage as Downloaded from https://academic.oup.com/dnaresearch/article-abstract/3/2/55/481488 by guest on 18 March 2018

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

DNA RESEARCH 3, 55-64 (:.996)
Unique Telomeric Expression Site of Major-Surface-GlycoproteinGenes of Pneumocystis carinii
Miki WADA and Yoshikazu NAKAMURA*Department of Tumor Biology, The Institute of Medical Science, The University of Tokyo,
P.O. Takanawa, Tokyo 108, Japan
(Received 4 April 1996)
Abstract
Major cell surface glycoproteins (MSG) of Pneumocystis carinii play a crucial role in the host-parasiteinteractions involved in P. carinii pneumonia in AIDS patients. Genes encoding MSGs are repeated, highlypolymorphic, and distributed among all of the 14-15 chromosomes. Here we show, by BAL-31 exonucleasecleavage and DNA cloning experiments, that the unique expression site (previously termed UCS) of MSGgenes located in the 500-kb chromosome is telomeric. The 11-kb genomic UCS fragment isolated andsequenced in this study contained one MSG coding sequence (termed msglO5), subtelomeric repetitivesequences and telomere-specific tandem repeats of TTAGGG oriented 5' to 3' towards the DNA end.Despite the N-terminal polymorphism, the C-terminal one-third sequence of MSG105 was identical to oneof the known MSG-cDNAs, suggesting homologous recombination within the MSG coding sequences. Thesefeatures closely resemble the Variant Surface Glycoprotein system of the protozoan parasite Trypanosomabrucei, suggesting that the genetic heterogeneity of MSGs is generated by recombination between the UCSexpression site and multiple MSG genes by means of reciprocal exchange or gene conversion.
Key words: Pneumocystis carinii; major surface glycoprotein; expression site; telomere
1. Introduction
Pneumocystis carinii is an opportunistic pathogen thatoften causes fatal pneumonia in patients under immuno-suppressed or immunodeficient conditions such as AIDS,cancer chemotherapy or organ transplantation. Morethan 60% of AIDS patients suffer from P. carinii pneu-monia at some time during the course of the disease.1 P.carinii is a fungus2'3 that is coated by abundant het-erogeneous major surface glycoproteins (MSGs) calledMSG,4-5 P115,6-7 gpl208 or gpA.9'10 The MSGs arethought to play a crucial role in the host-parasite inter-action during P. carinii pneumonia because of the par-tial protection conferred by passive immunization withan anti-MSG monoclonal antibody against the progres-sion of P. carinii pneumonia,11 the attachment of the or-ganism via interaction between MSGs and fibronectin onalveolar epithelial cells and macrophages,12'13 a specificT-cell response to MSGs after immunization and natu-ral infection,14 and the uptake of P. carinii by alveolarmacrophages mediated by the mannose receptor that tar-
Communicated by Mituru Takanami
* To whom correspondence shculd be addressed. Tel. +81-3-5449-5307, Fax. +81-3-5449-5415, E-mail: [email protected]
f The nucleotide sequence data reported in this paper will ap-pear in the DDBJ, GenBank said EMBL nucleotide sequencedatabases with the accession number D82031.
gets the mannose moiety of MSG.15
MSG is highly glycosylated with mannose and itsmolecular mass has been reported to be from 100 to 125kDa.6'8'16 It accounts for three-quarters of the total cel-lular protein when isolated from rats.6 Genes encodingMSGs are repeated, highly polymorphic, and distributedamong all of the 14-15 chromosomes.4'5'7'9-10'17'18 Weshowed that MSG gene expression is mediated by aDNA element termed the upstream conserved sequence,UCS.19 The UCS element maps to a single chromosomeof about 500 kb, is attached to expressed MSG genes, andencodes the sequence found at the 5' ends of most MSGmRNAs. The UCS is not highly repeated, but P. cariniipopulations contain many different MSG genes attachedto the UCS, suggesting that the genetic heterogeneity ofMSGs is generated by UCS-MSG recombination.19 Herewe show that the UCS locus is telomeric, suggesting thatUCS-MSG recombination occurs by recombination be-tween the UCS expression site and multiple MSG genesby means of reciprocal exchange or gene conversion.
2. Materials and Methods
2.1. OrganismsPathogen-free athymic (rnu/rnu) nude rats were in-
fected with P. carinii as described,6 and P. carinii or-ganisms were prepared from bronchoalveolar lavage as
Downloaded from https://academic.oup.com/dnaresearch/article-abstract/3/2/55/481488by gueston 18 March 2018

56 Telomeric Expression Site of P. carinii [Vol. 3,
described.6'19 The purity of P. carinii organisms as as-sessed by the relative intensity of ribosomal RNA bandsafter agarose gel electrophoresis was about 95%.6'19
2.2. Genomic libraryGenomic DNA was prepared from P. carinii organ-
isms as described.6'19 P. carinii DNA samples (3 fig)were digested with 10 units of Mung Bean Nuclease(Takara Shuzo, Kyoto. Japan), followed by BamHI. Af-ter the fill-in reaction with the Klenow fragment, BamHIlinkers were added, followed by cleavage by BamHI. tofacilitate cloning into AEMBL3 DNA. The DNA wasthen ligated into the BamHI site of AEMBL3, pack-aged with Gigapack Gold Plus packaging extract (Strata-gene, La Jolla, CA), plated on Escherichia coli LE392cells,20 and screened by plaque hybridization with theUCS probe. The probe was a 505-bp UCS segment am-plified by the polymerase chain reaction (PCR) fromgenomic DNA using the UCS primers (sense primer,5'-TGTGCAATAATGACCATTGC-3'; antisense primer,5'-CCTCGATCAAATGACCTCCA-3'). Detection afterblot hybridization was performed using the ECL DirectNucleic Acid Labelling and Detection Systems (Amer-sham, Buckinghamshire, UK). Probe DNAs were labelledwith the enzyme horseradish peroxidase, hybridized toDNA blots and detected by enhanced chemiluminescenceusing the Hyperfihn™-ECL (Amersham) according tothe manufacturer's instructions. Two positive cloneswere selected from 9 x 104 recombinant plaques and thenucleotide sequence of one recombinant clone, AMW124,was determined.
2.3. BAL-31 analysisThe P. carinii DNA (2.5 /j,g) was digested with 1.8
units of BAL-31 at 30°C for 0, 30 or 60 min in 200 filof BAL-31 buffer (20 mM Tris-HCl, pH 8.0, 600 mMNaCl, 12 mM CaCl2, 12 mM MgCl2, 1 mM EDTA).The reaction was stopped by adding ethyleneglycol-fris-(/3-aminoethyl)-AT,iV,./V',./V/-tetraacetic acid (EGTA) toa final concentration of 15 mM. The DNA was re-solved by agarose-gel electrophoresis after digestion withBamHI, blotted onto Hybond™-N+ membranes (Amer-sham), and hybridized to the UCS or the translationfactor EF3-coding21 probe DNA using the ECL DirectNucleic Acid Labelling and Detection Systems (Amer-sham). The EF3-coding sequence was amplified by PCRfrom genomic DNA using the probes (sense primers, 5'-CCGAATTCCCGGTTGAGCAACACATATG-3'; anti-sense primer, 5'-CCGAATTCCTATATGACATCTGGC-ACGG-3'), digested at the internal BamHI site and the3' fragment (377 bp) was used as the EF3 probe.
2.4- Karyotype blot hybridizationThe membrane blot of P. carinii chromosomes re-
solved by field inversion gel electrophoresis was pre-pared as described.19 The membrane was hybridized tothe (TTAGGG)5-oligonuclectide probe labeled with the3'ECL oligolabeling system (Amersham). After strippingthe Southern blots as described by Sambrook et al.,22 thesame karyotype blot was hybridized to subtelomeric re-gion II (330-bp Hindi, positions 10306-10636) and regionIV (650-bp EcoRl, positions 7604-8253) probes generatedfrom AMW124 DNA using t i e ECL Direct Nucleic AcidLabelling and Detection Systems (Amersham).
3. Results
We isolated three UCS-MSG clones of 2-6 kb from a P.carinii genomic library consisting of BamHI fragments.19
Since Southern blotting predominantly detected an 11- to13-kb segment hybridizable to the UCS probe when theP. carinii DNA was cleaved with BamHI,19 these threeisolates might have represented minor sets of UCS-MSGsequences among P. carinii populations that incidentallycontained BamHI site(s) within the polymorphic codingregion. Further trials to isolate a UCS clone having the11- to 13-kb fragment from the conventional library wereunsuccessful even after screening more than a half mil-lion plaques. This suggested that the 11- to 13-kb UCSfragment is telomeric and that it is not susceptible toconventional cloning using restriction endonucleases.
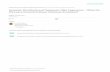
To determine whether the UCS is telomeric, P. cariniiDNA was digested with BAL-31 exonuclease for 0, 30 and60 min, then with BamHI. The samples were resolvedby agarose gel electrophoresis for Southern hybridizationto the UCS and EF3 probes. The EF3 gene fragmentwas not sensitive to BAL-31 (Fig. 1A, right). On theother hand, the size of the BamHI fragments that hy-bridized to the UCS probe slightly, though significantly,decreased depending on the time periods (Fig. 1A, left).Image analysis using the NIH Image 1.52 computer pro-gram confirmed the finding (Fig. IB). A minor UCS frag-ment that migrated faster than the others was sensitiveto BAL-31. These results suggest that most, if not all,of the UCS of the MSG genes are telomeric.
A P. carinii DNA library containing the telomeric re-gions was constructed by digesting with Mung Bean Nu-clease and BamHI, then ligating with BamHI linker af-ter the fill-in reaction with the Klenow fragment (seeMaterials and Methods). The recombinant phages werescreened by plaque hybridization using the UCS probe,and two A clones carrying an 11-kb fragment were iso-lated. Sequence analysis of one of them, AMW124,showed that it contained the UCS and one MSG-codingsequence, but no other significant open reading frames(Fig. 2). The MSG gene in AMW124, named msglO5, is
Downloaded from https://academic.oup.com/dnaresearch/article-abstract/3/2/55/481488by gueston 18 March 2018
No. 21 M. Wada and Y. Nakamura m
ucs EF3
30 60 0 30 60 (min)
(kb)
2 3 . 1 -
9 . 4 "
6 . 6 -
4 .4 —
B
0 min
30 min
60 min
c
Q
J.J
JVL.
iii
Den
sity
/
/
j
I\
Pixels 49 Pixels 49
Figure 1. Sensitivity of the UCS and EF3 genes to BAL-31 digestion. (A) P. carinii DNA was digested with BAL-31 for 0, 30 and60 min, followed by BamHl, then fragments were resolved by 0.8% agarose-gel electrophoresis and Southern blotted with the UCS(left) and EF3 (right) probes. Experimental procedures and conditions are described in Materials and Methods. Hindlll digests ofA DNA served as size markers shown on the left. (B) Densitometry scanning of Southern blots using the NIH Image 1.52 program.The solid and dotted arrows (left panel) indicate the positions of the major and minor UCS bands, respectively. The solid line (rightpanel) indicates the position of the EF3 bands.
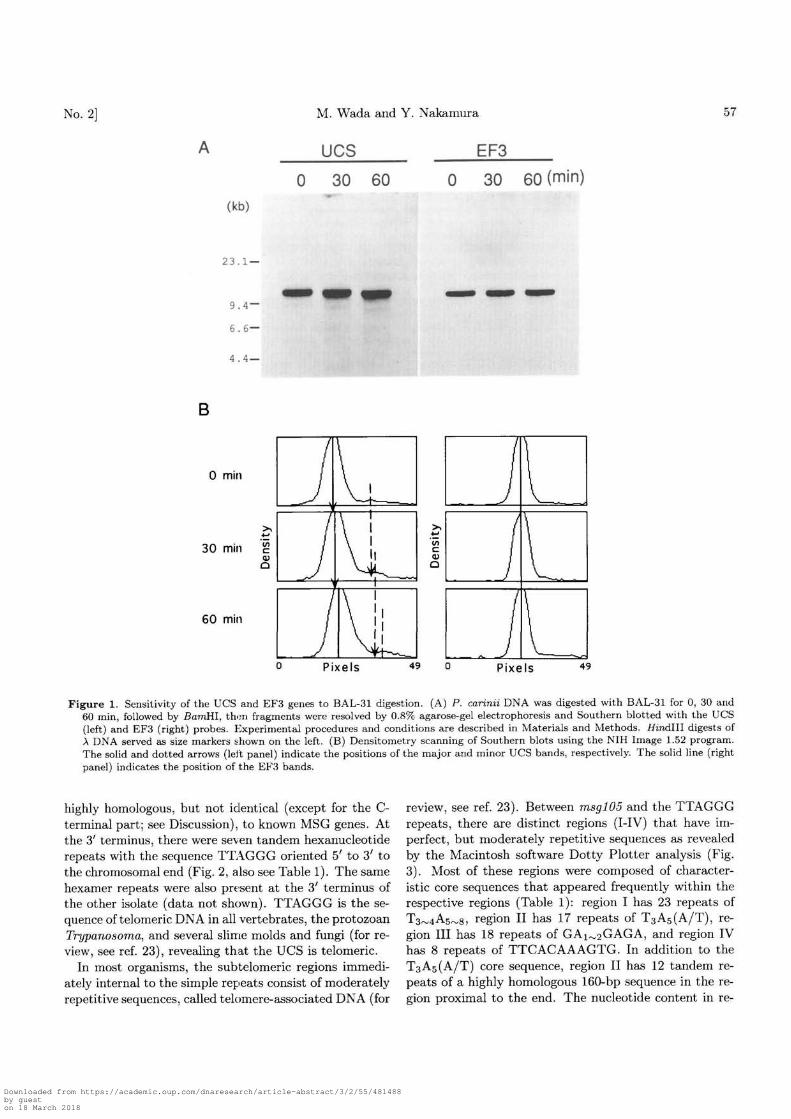
highly homologous, but not identical (except for the C-terminal part; see Discussion), to known MSG genes. Atthe 3' terminus, there were seven tandem hexanucleotiderepeats with the sequence TTAGGG oriented 5' to 3' tothe chromosomal end (Fig. 2, also see Table 1). The samehexamer repeats were also present at the 3' terminus ofthe other isolate (data not shown). TTAGGG is the se-quence of telomeric DNA in all vertebrates, the protozoanTrypanosoma, and several slime molds and fungi (for re-view, see ref. 23), revealing that the UCS is telomeric.
In most organisms, the subtelomeric regions immedi-ately internal to the simple repieats consist of moderatelyrepetitive sequences, called telomere-associated DNA (for
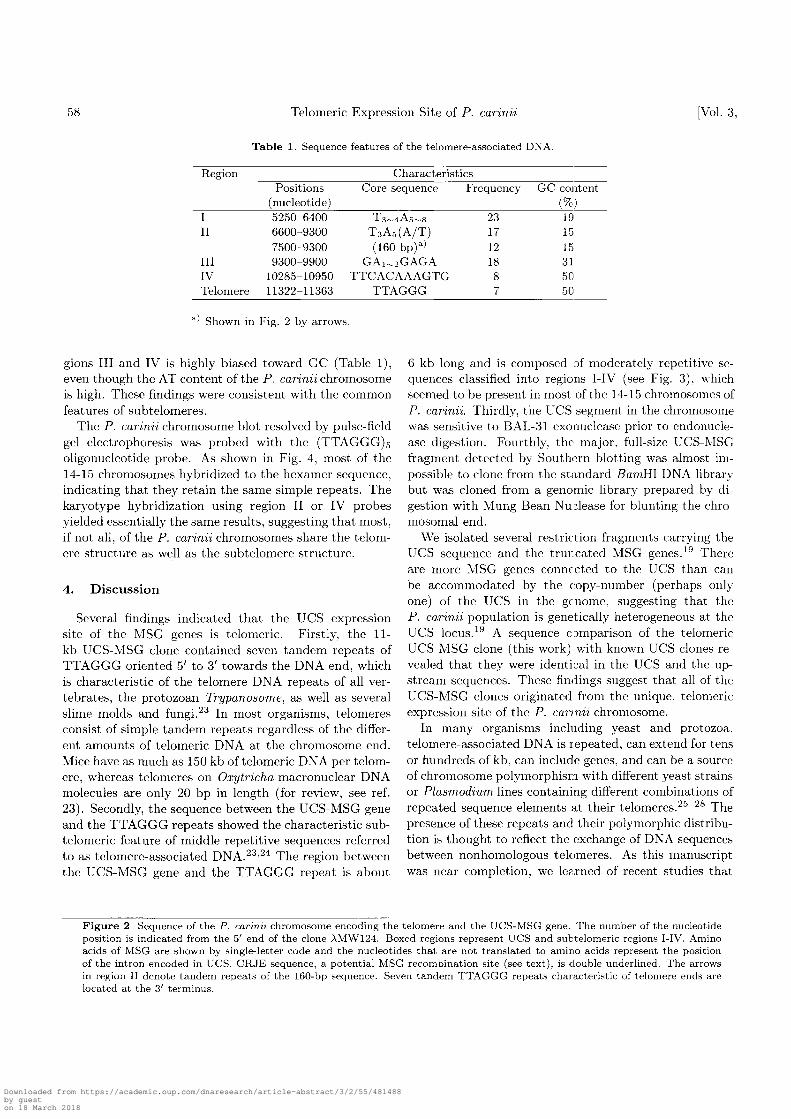
review, see ref. 23). Between msglO5 and the TTAGGGrepeats, there are distinct regions (I-IV) that have im-perfect, but moderately repetitive sequences as revealedby the Macintosh software Dotty Plotter analysis (Fig.3). Most of these regions were composed of character-istic core sequences that appeared frequently within therespective regions (Table 1): region I has 23 repeats ofT3^4A5^s, region II has 17 repeats of TaA5(A/T), re-gion III has 18 repeats of GAx^GAGA, and region IVhas 8 repeats of TTCACAAAGTG. In addition to theT3A5(A/T) core sequence, region II has 12 tandem re-peats of a highly homologous 160-bp sequence in the re-gion proximal to the end. The nucleotide content in re-
Downloaded from https://academic.oup.com/dnaresearch/article-abstract/3/2/55/481488by gueston 18 March 2018
58 Telomeric Expression Site of P. carinii [Vol. 3,
Table 1. Sequence features of the telomere-associated DNA.
Region
III
IIIIVTelomere
Positions(nucleotide)5250-64006600-93007500-93009300-9900
10285-1095011322-11363
CharacteristicsCore sequence
T3~4A5~8T3A5(A/T)(160 bp)a)
GAi^2GAGATTCACAAAGTG
TTAGGG
Frequency
2317121887
GC content(%)191515315050
Shown in Fig. 2 by arrows.
gions III and IV is highly biased toward GC (Table 1),even though the AT content of the P. carinii chromosomeis high. These findings were consistent with the commonfeatures of subtelomeres.
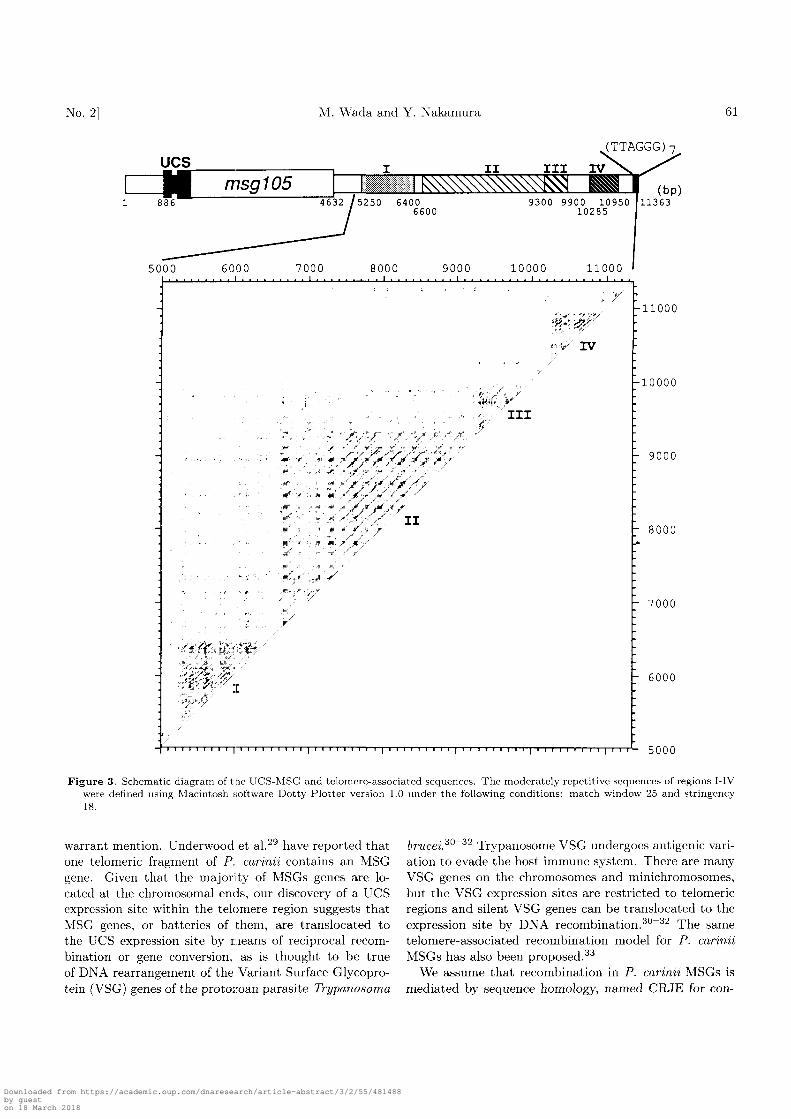
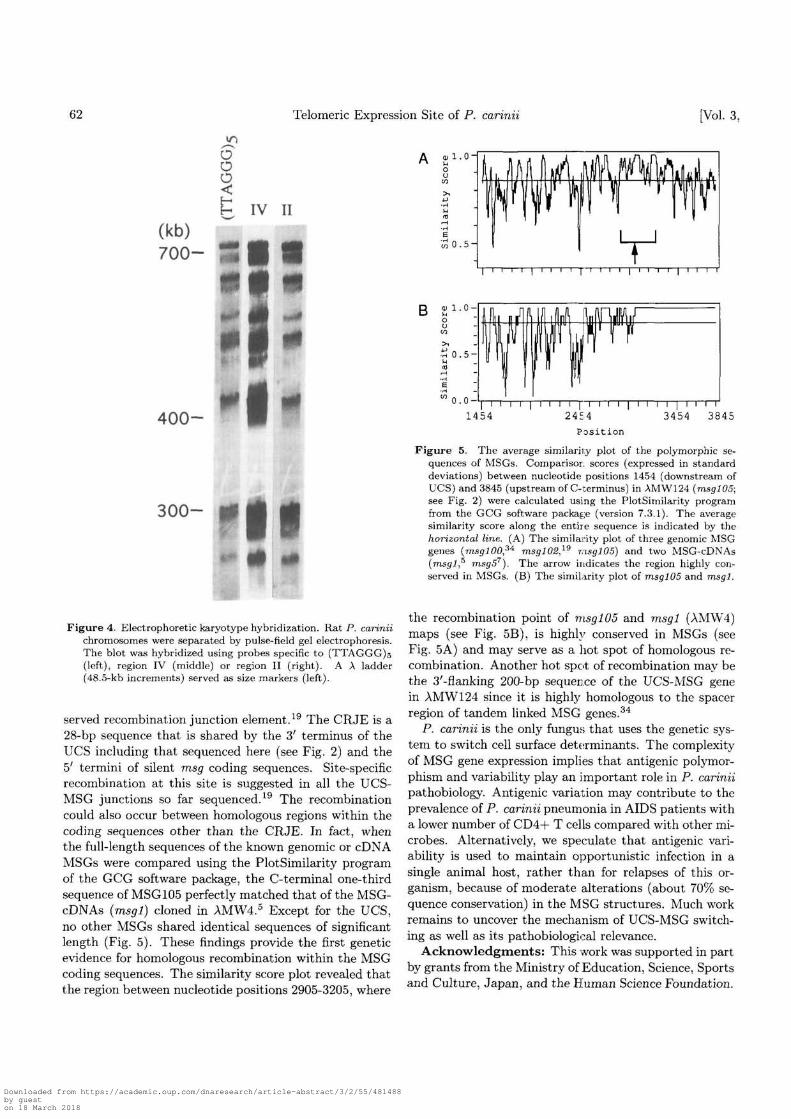
The P. carinii chromosome blot resolved by pulse-fieldgel electrophoresis was probed with the (TTAGGG)soligonucleotide probe. As shown in Fig. 4, most of the14-15 chromosomes hybridized to the hexamer sequence,indicating that they retain the same simple repeats. Thekaryotype hybridization using region II or IV probesyielded essentially the same results, suggesting that most,if not all, of the P. carinii chromosomes share the telom-ere structure as well as the subtelomere structure.
4. Discussion
Several findings indicated that the UCS expressionsite of the MSG genes is telomeric. Firstly, the 11-kb UCS-MSG clone contained seven tandem repeats ofTTAGGG oriented 5' to 3' towards the DNA end, whichis characteristic of the telomere DNA repeats of all ver-tebrates, the protozoan Trypanosome, as well as severalslime molds and fungi.23 In most organisms, telomeresconsist of simple tandem repeats regardless of the differ-ent amounts of telomeric DNA at the chromosome end.Mice have as much as 150 kb of telomeric DNA per telom-ere, whereas telomeres on Oxytricha macronuclear DNAmolecules are only 20 bp in length (for review, see ref.23). Secondly, the sequence between the UCS-MSG geneand the TTAGGG repeats showed the characteristic sub-telomeric feature of middle repetitive sequences referredto as telomere-associated DNA.23'24 The region betweenthe UCS-MSG gene and the TTAGGG repeat is about
6 kb long and is composed of moderately repetitive se-quences classified into regions I-IV (see Fig. 3), whichseemed to be present in most of the 14-15 chromosomes ofP. carinii. Thirdly, the UCS segment in the chromosomewas sensitive to BAL-31 exonuclease prior to endonucle-ase digestion. Fourthly, the major, full-size UCS-MSGfragment detected by Southern blotting was almost im-possible to clone from the standard BarriHl DNA librarybut was cloned from a genomic library prepared by di-gestion with Mung Bean Nuclease for blunting the chro-mosomal end.
We isolated several restriction fragments carrying theUCS sequence and the truncated MSG genes.19 Thereare more MSG genes connected to the UCS than canbe accommodated by the copy-number (perhaps onlyone) of the UCS in the genome, suggesting that theP. carinii population is genetically heterogeneous at theUCS locus.19 A sequence comparison of the telomericUCS-MSG clone (this work) with known UCS clones re-vealed that they were identical in the UCS and the up-stream sequences. These findings suggest that all of theUCS-MSG clones originated from the unique, telomericexpression site of the P. carinii chromosome.
In many organisms including yeast and protozoa,telomere-associated DNA is repeated, can extend for tensor hundreds of kb, can include genes, and can be a sourceof chromosome polymorphism with different yeast strainsor Plasmodium lines containing different combinations ofrepeated sequence elements at their telomeres.25~28 Thepresence of these repeats and their polymorphic distribu-tion is thought to reflect the exchange of DNA sequencesbetween nonhomologous telomeres. As this manuscriptwas near completion, we learned of recent studies that
Figure 2 Sequence of the P. carinii chromosome encoding the telomere and the UCS-MSG gene. The number of the nucleotideposition is indicated from the 5' end of the clone AMW124. Boxed regions represent UCS and subtelomeric regions I-IV. Aminoacids of MSG are shown by single-letter code and the nucleotides that are not translated to amino acids represent the positionof the intron encoded in UCS. CRJE sequence, a potential MSG recombination site (see text), is double underlined. The arrowsin region II denote tandem repeats of the 160-bp sequence. Seven tandem TTAGGG repeats characteristic of telomere ends arelocated at the 3' terminus.
Downloaded from https://academic.oup.com/dnaresearch/article-abstract/3/2/55/481488by gueston 18 March 2018

No. 2] M. Wada and Y. Nakamura 59
GGATCCGTTTTTATTATAAAATTTCATGrTTTATGTATAAAACTTTCTTTTTTTTATTTTTACTTGGTCCCTTTCTTTCCCTCATCATACTTGGAATTTTCTGATATATTACAATAGATG 120
GTTCATTCCTCTACTCACTTTTAAAATCrTATTTAACTGACCATTCATGCTATTTTTCCCCTATTTTTTATAAAAAGACTCGTCGCTTATATATAGAATGGCGTATACGTAAAAATAGAA 240
ACGTGTATGCGTGAAATAGAATGGTGCTrTTAAATTTAGAAACTTTATTCAAATTAGGAAATCTTTTTTTCCTTTTTAGTTTTGGAATCCATTATTCTTCTATACAAAGTATTTCTTTTT 360
TTCCTAACATGACTATTTATACTTTATArTTGGAAACCTATGCATATACTCGATATCACTCTGACTTTGCATATTCTACTCCCTCCTTTCCTTTCACCCCCTCTGGGACACTCCCCTGTC 480
CGGACACACCCCGGACAGCTCTTGGCTGrCCCTCCCTCGACCCTTCTATACCCATCTATCCATCTATCCATCTATATATACACCGTTTTTACTCATTTCTATCCTATAAAAATTGACTTG 600
TAATTTTGGATATATTCCTTCTATATGCATTTTTAGGACTGGCTAATATTTTAAGGTCTAGCTCTTTTTATTTCCGGCTAGTCATCCCCCCATCCGGAAATTGGCTTTTCCTTTTTCAGC 720
GTTTTCTTCGATTCATCCGTAAATTTCC3TCAATATTTAAGGGTCTTTGCATATTAAGGGGACTGTTTCTGGACTGTATTTTTACCATACCCGTTGCTTCCTTTAGCGCTTGGAAAATAT 840
ATTTTTTCTi|GATATCCGTCTCGTCTTCAGTTTGTTTGTGCAATAATGAGGATTGCATTTTTTGCGCTTTTTGCGCAACTTAGTTGTATTTTAGrTTATTCAATAGCAGAAAGGGATTTC | 960
M R I A F F A L F A Q L S C I L V Y S I A E R D F
ATGTCArTAGATGAAATATATGGTGAGTrTTTrTTCCTTAATTTCCTGrTGTATCATTTTrTACACGTCTTTTTTTACrCGrCTTTrACTCATCTTTTTACrCATCTTTCTATTCTCTCC
M S L D E I Y
TCTTCTTTTGGAAATCTCTTTTTCCTTTTATGGTTTTACTCATGTTTTCTAGAAGGAGGCGATATAAGTTTTGATCATGAAAAACTCGAATTTAACGAATATAATCAAGTTTTACAAATG
E G G D I S F D H E K L E F N E Y N Q V L Q M
CTTGAAAAGGCAAAAAAATTGGGAACCGGCTTTGTTGATAGAACCAAAGATTTTTCTAATAGACGATATGAAGGGAGAATTGAGTTAAATCATrTGGGGAGACGCCCAGGAGTCGACTAT
L E K A K K L G T G F V D R T K D F S N R R Y E G R I E L N H L G R R P G V D Y
TTTAGGAAAGGTGGGGATGTTTTTJ '
F R K G G D V F T D G Y P R G G H L I E D E L S E E V A M A R P V K R Q
3CACCACAAGCA
^ P Q A
GCACCAGCAGCGGATGGGATTAAGGAGG.\ACACCTTTTGGCTTTCATTCTGAAGGAGAAATATAAAAACGGATGCAAAGATGAGCTTGAAAAATATTGTAAAGAGTTGAAGGAAGCAGAT
A P A A D G I K E E H L L A F I L K E K Y K N G C K D E L E K Y C K E L K E A D
AAAAATCTAGAGAAAGTGAATGATAAAGTTAAAGGATTCTGTGAAGATGGAAAACAAGACGAAAAATGCAAAGGCCTGAAAGAGAAAGTTGAACAAGAATTGGACACTTTTGATGCAGAA
K N L E K V N D K V K G F C E D G K Q D E K C K G L K E K V E Q E L D T F D A E
CTTCCAGATGATTTAGGAAATCTAAAAG.MGAAGAATGTGAAAAACATGAAGAAAAATGTATCCTTTTAGAAGAGACGGGTTATAATGATGATATTAAGAAGAAGTGTGTCAACTTGAGG
L P D D L G N L K D E E C E K H E E K C I L L E E T G Y N D D I K K K C V N L R
GAAGGATGTTACAAATTGAAGCGTGAAA,^GGTGGCAGAGGAGCTCCTTTTGAGGGCGCTCGGAAAGGAAGCTAAAGAAAAAAACACATGTGAACTAAAGATGAAAACTGTTTGCCCAGTG
E G C Y K L K R E K V A E E L L L R A L G K E A K E K N T C E L K M K T V C P V
TTAAGCCGAGAAAGCGACGAATTGATGTTTTTCTGCCTTGATCCGACTAAAACATGTGGAGATATGAAAACAAAATTGGAAGAAAATTGCAAGCCTTTAAAAGAAAAGCTTAAAGATAAC
L S R E S D E L M F F C L D P T K T C G D M K T K L E E N C K P L K E K L K D N
GAATTAGAGGGAAAGTGTCATGAAAGACTTGAGAAATGTCATTrTTACGGAGAAGCGTGTGCTGATGCGAAATGTAAGGAGGATAAGACGAAATGCGAGGGAAAAGGATTCACATATAAA
E L E G K C H E R L E K C H F Y G E A C A D A K C K E D K T K C E G K G F T Y K
ACGCCAGAATCCGATTTTAGTCCGGTGAJ^ACCGAAGGCGTCGTTGTTGAGAAGTATTGGGTTGGATGATGTGTATAAAAACGCAGAAAAAGATGGAATTATTATTGGAAAAGCAGGAGTG
T P E S D F S P V K P K A S L L R S I G L D D V Y K N A E K D G I I I G K A G V
GATCTACCAAGGAAGTCGGGTACAAAATTTCTGCAAGATCTCTTGCTAGTGTTGAGCAGAGATGAGAATGATAAGAATACAGATAAAAAGTGCAAAAAAGCGTTAGAAAAATGTGGTGCT
D L P R K S G T K F L Q D L L L V L S R D E N D K N T D K K C K K A L E K C G A
TCTGAGTATTTGGATGCTGATTTGAAAA^^GTTATGCAAAGATGGTGATAAACAAGAGAAATGCAAAGCATTACTAGATGTAAATGTAAAAGAAAGATGTACAAATCTCAAATTAAAACTT
S E Y L D A D L K K L C K D G D K Q E K C K A L L D V N V K E R C T N L K L K L
TATCTGAAAGGATTGTCTACGGAATATD^TGATCAAGAATCAGATCCTTTATCGTGGGGACAGCRTCCAACATTATTTACGAAGGGAGAGTGRGCAGAACTTGAGTCGGAATGTTTCTAT
Y L K G L S T E Y D D Q E S D P L S W G Q L P T L F T K G E C A E L E S E C F Y
TTAGAAAATGCGTGTAAGGATAATAAGATTGATGAAGCATGTCAAAATGCAAGAGCAGCGTGCTATAAAAAGGGACAAGACAGGATGTTGAATAAGTTCTTTCAAAAGGAATTGAGGGGA
L E N A C K D N K I D E A C Q N A R A A C Y K K G Q D R M L N K F F Q K E L R G
AATCTTGGTCTTGTAAGATTTTATAGCG/1TCCTGAAGAATGCAAAAAATCTGTGGTAGGAAACTGTACAAAACTTAAAGAAGATAGTAGATACCTTTCAAAATGTCTTTATCCTAAAGAA
N L G L V R F Y S D P E E C K K S V V G N C T K L K E D S R Y L S K C L Y P K E
TTATGTTATGCGCTTTCAAATGATATTTTTCTTCAATCCAAAGAGTTAAGTTCGCTTTTGGATGATCAAAGGGATTTTCCATTAGAAAAGGATTGTCTTGAATTGGTGGAGAAGTGTGAT
L C Y A L S N D I F L Q S K E L S S L L D D Q R D F P L E K D C L E L V E K C D
GAACTTAGTAGTGATTCATTATTGAATTTAGAAAAGTGTATAACATTGAAAAGACGCTGTGAATACTTTAAGGTTACAGAGGGATTTAGAAACGTATTTTTAGAAAAAAAGGATGATTCG
E L S S D S L L N L E K C I T L K R R C E Y F K V T E G F R N V F L E K K D D S
TTAATGACTCAGGAAAACTGTACAAAGGCATTGCATGAGAAATGTAATGCTTTATCTAGGAGGAGAAAGAATTCATTTAGTGTRTCATGTGCTTTACCAGAAGAAACATGTAGTTATATG
L M T Q E N C T K A L H E K C N A L S R R R K N S F S V S C A L P E E T C S Y M
GTATTCCACACAAGTCAAGATTGTAAATATTTAAAAGAAAACATCAAGAATGGAGGAATTGTAGGAAAAATTGGGGGAGCAAATGGAAATGAAGCAACACTCGAAGAACTCTGCACAACA
V F H T S Q D C K Y L K E N I K N G G I V G K I G G A N G N E A T L E E L C T T
TGGGGCCAACATTGTCACCAACTTGTGG;\GAATTGTCCAGATCAGTTGAAAAAAGAGAATGGCAATAARCATAACTGRGATGAACTCGATGAAAAATGCAGTGATACCRTTAAAAGGTTG
W G Q H C H Q L V E N C P D Q L K K E N G N N H N C D E L D E K C S D T F K R L
AAATTAGAGGAGGAGCTGACTCATCTGT"GAAAGGCAGTTTAAAGAAGGATGATCAATGTACAGAAGCATTAGGAAAGAATTGCACTGACTTGCAAAAGAATGAAACATTCAAAATTCTG
K L E E E L T H L L K G S L K K D D Q C T E A L G K N C T D L Q K N E T F K I L
CTTAGTGATTGTGAAAAAGATAAGCAAA;VTGTTTGCACAAAATTAGTTAAAAAAGTACAAGAGAGATGTCCTACTTTAAAAGCTGATCTGGATAAAGCGAAAGATGAGTTGACAAAGATG
L S D C E K D K Q N V C T K L V K K V Q E R C P T L K A D L D K A K D E L T K M
AAGAATGAGTACGAAAAGGCAAAAGAGG;\GGCAGAAAAATCTACAAATAAAGCTAGCTTATTGCTATCAAAGTCTGGAAAAGCCGCAATGCCAAGTGGACAGGATAGCAATGGCTCTGTA
K N E Y E K A K E E A E K S T N K A S L L L S K S G K A A M P S G Q D S N G S V
P A P P A E P A P G T L P P P P S T D G T V N T P A G G S G T S A G G S G T P S
AGTGGAACGACAGACTCTGCAAAACTTGGACTCGTTAAAAGAGCGTARGTAGAAGGAGGAGTATCAGAAGTAGAGGTAAAAGCATTTGATGCAACGACGGTAGCATTGGAATTGTATTTG
S G T T D S A K L G L V K R A Y V E G G V S E V E V K A F D A T T V A L E L Y L
GAATTGAAAGAGGAATGCAATGCTTTASlACTAGATTGCGGTTTTAAAGAGGATTGTTCAGATCTTCAAGTTTGCGGAGAAATAGACACGTTATGCGGAGGAATAAAACCATTAGAAATT
E L K E E C N A L E L D C G F K E D C S D L Q V C G E I D T L C G G I K P L E I
AAGCCTCATCATACAGAGACAATAACAAJiTAAGGTGACGGAAACGAAGACGGAAACAAAGACAGAAACAAAGACTGATGACAAGGCTGATGAGAAGACCGGTACGAAAACTGTTACAGAA
K P H H T E T I T N K V T E T K T E T K T E T K T D D K A D E K T G T K T V T E
ucs
Downloaded from https://academic.oup.com/dnaresearch/article-abstract/3/2/55/481488by gueston 18 March 2018

60 Telomeric Expression Site of P. carinii [Vol. 3.
ACCAAGACAATAGGTGGAGGAAAAGTAACAGAAGAGTGTACAATGATACAAACAACAGATACATGGGTAACACGTACATCATTGCATACGAGTACGACAACGAGTACGTCAACGGTGACG
T K T I G G G K V T E E C T M I Q T T D T W V T R T S L H T S T T T S T S T V T
TCGACAGTGACGTTGACGTCGATGCGCAAGTGCAAGCCTACCAAATGTACTACTGATTCAAGCAGAGAGACAGATAAAGGAGAAGAAGGAAAAGAAGGAGAAGAACATGTAAAACCGAAT
S T V T L T S M R K C K P T K C T T D S S R E T D K G E E G K E G E E D V K P N
GATGGGATGAAAATAAGAGTCCCTGATATGATTAAAATAATATTGTTGGGAGTGATTGTTATGGGGATGATGTAAATGAATGAAAAAAATGTTAATAGATTGAAAJTGTGCATATATCCA
D G M K I R V P D M I K I I L L G V I V M G M M
TTGTTTATATATAATAGAAATCTAAATGAATGAAATGAAGTTTTAATTATTTTAACACACCATCTTAGTTCGAAACTATGCTTTATCGGGTAAAGTTTAAGCTTATCGAAGAACTAAAGT
TTATGATAAGAAGATGTTTAAATTTGTTGTAAAAGCACTTGAATGAATGGATTATCGTGAAAAAAAGAGAGTTCTGCGTATCTTAAAGAATTAGTAGGGGTTATaiTTATTTTAAAAAAA
AATCCCACTAACCAGAGAAAGAAGTTArCTTGAACCCATTAAAAGACGATTTGAAArCGAGAGCTTTATGATCATCAAAAAGAGAAACATTAGArCGATTGAAAATGGAATGATCATGAA
AAGTTGTAACCCTAACCCTAACCCTAAAGTGACTAACCAGATCACCCGACTCTTCTAAAACCACTAGAATCTCATTGCAAAATCAGTCAkcrGAATGTCCATTrwAAAGAAGTACrACG
CTTTATGATTATTTTAAAAAAACTAGTGAAATCTATTGTTTATTTAAAAAAATTAGTAAAATCCAAGTTTGTTGTAAAAAAATTAACTCTACGCTAATTACAAAA/AACTAGGAAATCTA
TGTAAAAAACTAGTGAAATCCAAGTTTGTTATAAAAAACCACTAGAATGATTGTAAAAGAACTAATAAACTTrATGATTACTTTAAAAAAAATAAAATCTGTTGA"TATTTCAAAAAAAC
ATCTAAArTTTATGArTACTCCAAAGAArTAGTAGAATCTATrGTTTArTTTAAAAAAATCAGTAAACrCTACGCTAATTACAAAAGAACTAGTGAAATCAATTG"TTATTTAAAAAAAA
TTACTAAAATCCAAGTTTGTTGTAAAAAACTAGGAAATCTATTGATTATTTCAAAAAAATTAATAAACTCTAAAATCATTGAAAAAAACCAGTGAAATCCAAGTT"ATTGTAAAAACGAT
TAGAATAATTATAAAAGAAGTAATAAACTTTATGATTATTTAAAAAAAAAGAGAAAAGTTTATGATTACTTTAAAAAAAATAGTCAAATCCAAGTTTGTTGTAAA/jmAATATTTAACTC
ATITCTTATTTAAAAAAATAGTCAAATCCAAGTTTGTTGltlAAAAACCAGTAAAATGATTGAAAAAAAAGTAGTGAAATCTATTGTTTATCAAAAAGAGTCTATaTTATGATAAAAAAA
GAGTTAGACATTGGTTATCGTGAAAAAAAACGAATCCArCTGTrTAGCAAAAATTAGTCTGTCGTTAGGGCGAAGCTAGTGGTGATTTAATAAAAGTAAGCTATTCAATAAGGTTGAAlf
TAAAAAAAATTGTTAATCATTAATCAAATAAAAAAGAATTGAATTTTCATTGATAAATATAAGAAATGAAAGCATAGACTAGGTTTTATTAAAAAAAAGATGTTTATTTAAAAATATCTT
TAAAAAAATTTGAAGAGATTTTAAAATTAAAAATGAAAATCAAAAATAAAAATGATATCTAGAAAAATTTATTGAATAGATCAAGTAAGATTGAATTAATTTTTG/vTAGTAAAAGAAAAG
ATTAGAAGGAGGAGTrTTATTAAAAAGAAGAAAAAAAGAGATTGATTTGAGAGATATTTTTATTTAAAAATAACTTGAAAAGAAAGGATAGAGAATAGGTTTTAT"AAAAAGAAGAAGAG
ATTGATTTAAGAGATATTTTTATTTAAAAATAATTTAAAAAAAAAGAAAAGATTTTAAAATGAAATTTAAAAATGAAAATAAGAAAAAAAAAATGAATAAATTGGACAAGTTGAGTTGAA
TTTTCATTGATAAATATAAGAAATGAAAGGATAAAGTAGGTTTTATTAAAAAAAAGATGTCCATTTAAAAATATCTTTAATTTAAAAAAAAAAAGATTTTAAAAT"AAAAATGAAAATGA
AAAATAAAAATGAAATCTAAAAAAATTTGTTAAATAGATTAAATAAGATTGAATTGATTTTTGATAGTAAAGATGAGAAATGAAGGGTTAGAGTAGGTTTTATTACyiAGTACAATTGAAT
GAAATTTAAAAGGATTTTAAAAATGAAGATGAAATTAAAAGTGAAATTGAAATGTAAAAAAAATTGTTCACACTTGATCAAATGAAATAGAATTGATTTTTTATTCATTTAAATAAGAAT
TTAGAAATATTTTTGTTAAAAGTACAATTTTTTGTTAAAAGTAGAATTCAATGAAATTTAAAATTAAAAATGAAATCTAAAAAAAAAATCAAATAIAAAAAAAAT-'GTTAACAGTTGATC
AGATGAAAATGGATTGATTTTTTATTGATTTAAATAAGAATTTAGAAATAATTTTGTTAGAAGTACAATTAAATCAAATTTAAAAGGATTTTTAAAATAAAAATTAAAATAAAAAAATAA
AAATAAAATAT;'AA»AAA.TTGTTAACAGTTGATCAAATGAAATAGAATTGATTTTTTATCAACTAAAAAGAGATAAGAATTTAGAAATCTCTTTGTTAAAAGTAGAATTAGATGAAATAT
GAAAGGATTTTGArTAAAATAAAAAATGAAAATGAAAATTAAAATGAAAGTGAAATTCAAAATGAAATGTAAAAAAAAA"TTGTTGACAGTTGATCAAATGAAGTA<»ATTGATTTTGTAT
TAAGTAAAAAAGATAAGAATTTAGAAATAATTTTGTTAGAAGTAAAATTAGATGAAATATAAAAGGATTTTAAAAATGAAGATGAAATTAAAAATGAAATAT,AAAAAAA T̂TTGTTGACAG
TTGATTATATGAAGTAGAATTGATTTTTTATTGATTTAAATAATTTAGAAATATTTTTGTTAAAAGTACAATTTTTTGTTAAAAGTAGAATTCAATGAAATTTAAAATTAAAAATAAAAT
TAAAATAAAAAATSAAATGTiAAAAAAAT
AATTTAAAAGGATtTTTAAAATAAAAATrAAAATAAAAAAArAAAAArAAAATATi•AAAAAJiA'•TTGTTAACAGTTGATCAAATGAAATAGAATTGATTTTTTATCAACTAAAAAGAGATAA
GAATTTAGAAATATTTTTGTTAGAAGTAGAATTTTTTATTAAAAGTAGAATTCAATGAAATTTAAAAGGATTTTAAAAATGAAAATTAATGAAATATi-AAAAAAAA1.rTGTTAACAGTTGAT
CAAATAAAGTAGAATTGATTTTGTATTAAGTAAAAAGGATAAGAATTTAGAGATAATTTTGTTAAAAGTAGAATTAGATGAAATTTAAAAGGATTTTTATTTAAWlATTAGAAAATGAAA
TTAAAATTAAAATGAAATATiGAAAAAA'L T T A T T T A G C A G T C A A T T A T A T A A A A A T G A A T T G A T T T T T A T T A G T A G A A A T G A G A A T T T A G A A A T A A T T C T G T T A G A A G T A G A A T T T T T T G T T
AAAACTAGAATTAGATGAAATTTTAAAATAAAAATTAAAATAAAAAAATAAAAATGAAATCTi•AAAAAAA'.TTTGTTCACAGTTGATCAAATGAAGTAGAATTGATT'-TTTATTGACTTAAT
AAGAATTTAGAAATAATTTTGTTAAAAGTAAAATTAGATGAAATTTAAAAGGATTTTTATTTAAAAATTAGAAAGTGAAATTAAAATTAAAATGAAATATAAAAAiJiTTATTTAGCACTC
AATTATATGAAAATGAATTGATTTTTCATTAGTAAGATGAAAATTTAGAAAJATTTTTAACAATTGATCAAATGAAGTAGAATTAAITTTTTATTGACTAAAAAAAATAAGAATTTAAAA
ATAATTTTGTTAAAAGTAGATTTTTTTGTTAAAAGTAGAATTAGATGAAAATTAAAGT^J^AAGAAGAGAGAAGAGAGTAGAGTGAGAGATATTTTTATTAATAAAAGGAAGAGAGTAG
AGAAACGTCAATTGAATAGATCTTCTGTTCGTGAGAAGAAGAAGAGAGTAGGATGAGAAGAAGAGAGTAGAGGGAGCTTTAAAAGAAGAGATTAGAGTGAGTTTT/WAAAGAAGAGATTA
TAGGAGATACTTTTGTTTTAAATAAAAAGGAATGAATTATTTTAAAAGAGAAGAGAGTAA[:GTCAATTGAATAGTTTTTTTCTTCGTAAAAAGGAGGCAGAGTAG(^TGTGAGATCATTT
GTTTTAAAAAGGGGCAGCGTCGAATGGAATGGCACTACCGAGGAATGCGACTACCGAGGAATGGCACTTCCGAGGAATGGTGCGTCCGAGGAATGTGCGTAGGAGTATAAATAGGAGGGA
AGGAAATAGGGTGGGTGGGGCGGTGGTTGGTGGTGGTTAGAAAGGGTTAGGAAGAAGTAGGGGGGGTGGGGGAGAGAGGAGAGGGGGGGGTCGAAAGTCACGTGACGAAGGACGAGGTCA
AG^ACCGGCTTAACGTCTGTTCA
TAAAGTGCTAGTACTTGGTGGACGTAAAAAGAAGTGCTAGAAAGAGGTGAACGTCAATTCACGAAGTGCTACTTCTGGGTGAACGTAAGTAGGAAAGGTGAGCGTMGTAGGAAAGGTGG
GCGTAAGTAGrACTCGGTGGGCGTAAGTAGTACTCGGTGGACGTCTATTCACAAAGTGCTCCGGGTAGGTGAACGCAGAATGTGAGTCGGTCGAGGGACGCAGTAGTCGGTCGGGGAAGT
GCTCCGGGTAGGCGGACGTAAGTAGGAGTTAGGTGGGTGTAGAAGTACTAGGTGGGCGCAGAAGTACTGGGTTAACGTCAATTCACAAAGTGCTCCGGGTAGGTGI^ICGCAAGTAGAGAA
GGGTTAACGTCTATTCACAAAGTGCTCCGGGTAGGCGGGCGTAGAAGTACTAGGTGAACGCCAATTCACAAAGTGCTCCGGGTAGGTGGGCGTAAGTAGTAGTACTAGGTGGACGTCTAT
TCACAAAGTGCTAGrGGTAGGTGGACGTCTGTTCACAAAGTGCTAGTACTGGGTGAGGTAAGTAAGAAAAGGTGGGCGTAAGTAGGGTTAGGTGACGTCTGTTCACAAAGTGCTCGGGGT
AGGTAACGCAAAGTGAAGTGCTATGAAGGTAAGG1JCAAAGTCACGTGACCAGTAAAAGTCGAGTAAGAGAGTTGTAGTACGTAGGTTCGAGTCGTAGTAGGTGAG"TCGAGTCTTCAGGT
TCACCTAGTCAAACGGTTAGGGGGGTTAGGTGGTTAGCACGTTAGGGTTAGGGTTAGGGTTAGGGTTAGGGTTAGGGTTAGGG 11363
4800
4 920
5040
5160
5280
5400
5520
5640
5760
5880
6000
6120
6240
636.0
6480
6600
6720
6840
6960
7080
7200
7320
7440
7560
7680
7800
7920
8040
8160
8280
8400
8520
8640
8760
8880
9000
9120
9240
9360
9480
9600
9720
9840
9960
10080
10200
10320
10440
10560
10680 I V
10800
10920
11040
11160
11280
II
III
Figure 2. Continued.
Downloaded from https://academic.oup.com/dnaresearch/article-abstract/3/2/55/481488by gueston 18 March 2018
No. 2 M. Wada and Y. Nakamura 61
ucs(TTAGGG)7
5000
4632 /5250 64006600
8000 9000
9300 9900 1095010285
10000 11000. . . I I . .
(bp)11363
I V
wm. /'/•/.
"t ' •|.rV rf'V'V ^"• _ ; _ - : ' . / . / ; . - • '
, .s1. .::f jt- .->•'
n
-11000
-10000
- 9000
- 8000
- 7000
- 6000
5000
Figure 3. Schematic diagram of the UCS-MSG and telomere-associated sequences. The moderately repetitive sequences of regions I-IVwere defined using Macintosh software Dotty Plotter version 1.0 under the following conditions: match window 25 and stringency18.
warrant mention. Underwood et al.29 have reported thatone telomeric fragment of P. carinii contains an MSGgene. Given that the majority of MSGs genes are lo-cated at the chromosomal ends, our discovery of a UCSexpression site within the telomere region suggests thatMSG genes, or batteries of them, are translocated tothe UCS expression site by means of reciprocal recom-bination or gene conversion, as is thought to be trueof DNA rearrangement of the Variant Surface Glycopro-tein (VSG) genes of the protozoan parasite Trypanosoma
brucei.30 32 Trypanosome VSG undergoes antigenic vari-ation to evade the host immune system. There are manyVSG genes on the chromosomes and minichromosomes,but the VSG expression sites are restricted to telomericregions and silent VSG genes can be translocated to theexpression site by DNA recombination.30^32 The sametelomere-associated recombination model for P. cariniiMSGs has also been proposed.33
We assume that recombination in P. carinii MSGs ismediated by sequence homology, named CRJE for con-
Downloaded from https://academic.oup.com/dnaresearch/article-abstract/3/2/55/481488by gueston 18 March 2018
•62 Telomeric Expression Site of P. carinii [Vol. 3,
i . o -
IV II(kb)700-
400-
300 -
m «•
Figure 4. Electrophoretic karyotype hybridization. Rat P. cariniichromosomes were separated by pulse-field gel electrophoresis.The blot was hybridized using probes specific to (TTAGGG)s(left), region IV (middle) or region II (right). A A ladder(48.5-kb increments) served as size markers (left).
served recombination junction element.19 The CRJE is a28-bp sequence that is shared by the 3' terminus of theUCS including that sequenced here (see Fig. 2) and the5' termini of silent msg coding sequences. Site-specificrecombination at this site is suggested in all the UCS-MSG junctions so far sequenced.19 The recombinationcould also occur between homologous regions within thecoding sequences other than the CRJE. In fact, whenthe full-length sequences of the known genomic or cDNAMSGs were compared using the PlotSimilarity programof the GCG software package, the C-terminal one-thirdsequence of MSG105 perfectly matched that of the MSG-cDNAs {msgl) cloned in AMW4.5 Except for the UCS,no other MSGs shared identical sequences of significantlength (Fig. 5). These findings provide the first geneticevidence for homologous recombination within the MSGcoding sequences. The similarity score plot revealed thatthe region between nucleotide positions 2905-3205, where
! o . s -
B <D 1 . 0 -
0 . 5 -
0 . 0 -
1454 3454 3845Position
Figure 5. The average similarity plot of the polymorphic se-quences of MSGs. Comparison scores (expressed in standarddeviations) between nucleotide positions 1454 (downstream ofUCS) and 3845 (upstream of C-terminus) in AMW124 (msglOS;see Fig. 2) were calculated using the PlotSimilarity programfrom the GCG software package (version 7.3.1). The averagesimilarity score along the entire sequence is indicated by thehorizontal line, (A) The similarity plot of three genomic MSGgenes (msglOO,34 msglO2,19 r.isglOS) and two MSG-cDNAs(msgl,5 msg57). The arrow indicates the region highly con-served in MSGs. (B) The similarity plot of msglO5 and msgl.
the recombination point of msg 105 and msgl (AMW4)maps (see Fig. 5B), is highly conserved in MSGs (seeFig. 5A) and may serve as a hot spot of homologous re-combination. Another hot spot of recombination may bethe 3'-flanking 200-bp sequence of the UCS-MSG genein AMW124 since it is highly homologous to the spacerregion of tandem linked MSG genes.34
P. carinii is the only fungus that uses the genetic sys-tem to switch cell surface determinants. The complexityof MSG gene expression implies that antigenic polymor-phism and variability play an important role in P. cariniipathobiology. Antigenic variation may contribute to theprevalence of P. carinii pneumonia in AIDS patients witha lower number of CD4+ T cells compared with other mi-crobes. Alternatively, we speculate that antigenic vari-ability is used to maintain opportunistic infection in asingle animal host, rather than for relapses of this or-ganism, because of moderate alterations (about 70% se-quence conservation) in the MSG structures. Much workremains to uncover the mechanism of UCS-MSG switch-ing as well as its pathobiological relevance.
Acknowledgments: This work was supported in partby grants from the Ministry of Education, Science, Sportsand Culture, Japan, and the Human Science Foundation.
Downloaded from https://academic.oup.com/dnaresearch/article-abstract/3/2/55/481488by gueston 18 March 2018

No. 2] M. Wada and Y. Nakamura 63
References
1. Selik, R. M., Starcher, E. T., and Curran, J. W. 1987,Opportunistic diseases reported in AIDS patients: fre-quencies, associations, and trends, AIDS, 1, 175-182.
2. Edman, J. C , Kovacs, J. A., Masur, H., Santi, D. V.,Elwood, H. J., and Sogin, M. L. 1988, Ribosomal RNAsequence shows Pneumocystis camnii to be a member ofthe Fungi, Nature (London), 334, 519-522.
3. Watanabe, J., Hori, H., Tanabe, K., and Nakamura, Y.1989, Phylogenetic association of Pneumocystis cariniiwith the 'Rhizopoda/Myxomycota/Zygomycota group'indicated by comparison of 5S ribosomal RNA sequences,Mol. Biochem. Parasitol, 32, 163-168.
4. Kovacs, J. A., Powell, F., Edman, J. C. et al. 1993, Multi-ple genes encode the major surface glycoprotein of Pneu-mocystis carinii, J. Biol. Chem., 268, 6034-6040.
5. Wada, M., Kitada, K., Saito, M., Egawa, K., andNakamura, Y. 1993, cDNA sequence diversity and ge-nomic clusters of major surface glycoprotein genes ofPneumocystis carinii, J. Infect. Dis., 168, 979-985.
6. Tanabe, K., Takasaki, S., Watanabe, J., Kobata, A.,Egawa, K., and Nakamura, Y. 1989, Glycoproteins com-posed of major surface immunodeterminants of Pneumo-cystis carinii, Infect. Imraun., 57, 1363-1368.
7. Kitada, K., Wada, M., and Nakamura, Y. 1994, Multi-gene family of major surface glycoproteins of Pneumocys-tis carinii: full-size cDNA cloning and expression, DNARes., 1, 57-66.
8. Radding, J. A., Armstrang, M. Y. K., UUu, E., andRichards, F. F. 1989, Identification and isolation of amajor cell surface glycop>rotein of Pneumocystis carinii,Infect. Immun., 57, 2149-2157.
9. Haidaris, P. J., Wright, T. W., Gigliotti, F., and Haidaris,C. G. 1992, Expression and characterization of a cDNAclone encoding an immunodeterminant surface glycopro-tein of Pneumocystis carinii, J. Infect. Dis., 166, 1113-1123.
10. Wright, T. W., Simpson-Haidaris, P. J., Gigliotti, F.,Harmsen, A. G., and Haidaris, C. G. 1994, Conserved se-quence homology of cysteine-rich regions in genes encod-ing glycoprotein A in Pneumocystis carinii derived fromdifferent host species, Infect. Immun., 62, 1513-1519.
11. Gigliotti, F. and Hughes, W. T. 1988, Passive immuno-prophylaxis with specific monoclonal antibody conferspartial protection against Pneumocystis carinii pneu-monitis in animal models. J. Clin. Invest., 81 , 1666-1668.
12. Pottratz, S. T. and Martin II, W. J. 1990, Mechanism ofPneumocystis carinii attachment to cultured rat alveolarmacrophages, J. Clin. Invest, 86, 1678-1683.
13. Pottratz, S. T., Paulsrud, J., Smith, J. S., and Martin II,W. J. 1991, Pneumocystis carinii attachment to culturedlung cells by Pneumocystis gpl20, a fibronectin bindingprotein, J. Clin. Invest., 88, 403-407.
14. Fisher, D. J., Gigliotti, F., Zauderer, M., and Harmsen,A. G. 1991, Specific T-cell response to a Pneumocystiscarinii surface glycoprotein (gpl20) after immunizationand natural infection, Infect. Immun., 59, 3372-3376.
15. Ezekowitz, R. A. B., Williams, D. J., Koziel, H. et al.1991, Uptake of Pneumocystis carinii mediated by the
macrophage mannose receptor, Nature (London), 351,155-158.
16. Gigliotti, F. 1992, Host species-specific antigenic varia-tion of a mannosylated surface glycoprotein of Pneumo-cystis carinii, J. Infect. Dis., 165, 329-336.
17. Linke, M. J., Smulian, A. G., Stringer, J. R., and Walzer,P. D. 1994, Characterization of multiple unique cDNAsencoding the major surface glycoprotein of rat-derivedPneumocystis carinii, Parasitol. Res., 80, 478-486.
18. Garbe, T. R. and Stringer, J. R. 1994, Molecular charac-terization of clustered variants of genes encoding majorsurface antigens of human Pneumocystis carinii. Infect.Immun., 62, 3092-3101.
19. Wada, M., Sunkin, S. M., Stringer, J. R., and Nakamura,Y. 1995, Antigenic variation by positional control of ma-jor surface glycoprotein gene expression in Pneumocystiscarinii, J. Infect. Dis., 171, 1563-1568.
20. Frischauf, A. M., Lehrach, H., Poustka, A., and Murray,N. 1983, Lambda replacement vectors carrying polylinkersequences, J. Mol. Bioi, 170, 827-842.
21. Ypma-Wong, M. F., Fonzi, W. A., and Sypherd, P.S. 1992, Fungus-specific translation elongation factor 3gene present in Pneumocystis carinii, Infect. Immun., 60,4140-4145.
22. Sambrook, J., Fritsch, E. F., and Maniatis, T. 1989,Molecular Cloning: A Laboratory Manual, 2nd ed., ColdSpring Harbor Laboratory, Cold Spring Harbor, NewYork.
23. Zakian, V. A. 1995, Telomeres: begtnning to understandthe end, Science, 270, 1601-1607.
24. Blackburn, E. H. and Szostak, J. W. 1984, The molecu-lar structure of centromeres and telomeres, Annu. Rev.Biochem., 53, 163-194.
25. Charron, M. J., Read, E., Haut, S. R., and Michels, C.A. 1988, Molecular evolution of the telomere associatedMAL loci of Saccharomyces, Genetics, 122, 307-316.
26. Corcoran, L. M., Thompson, J. K., Walliker, D., andKemp, D. J. 1988, Homologous recombination withinsubtelomeric repeat sequences generates chromosome sizepolymorphisms in P. falciparum, Cell, 53, 807-813.
27. Zakian, V. A. and Blanton, H. M. 1988, Distribution oftelomere associated sequences on natural chromosomes inyeast Saccharomyces cerevisiae, Mol. Cell Biol., 8, 2257-2260.
28. Brown, W. R. A., MacKinnon, P. J., Villasante, A.,Spurr, N., Buckle, V. J., and Dobson, M. J. 1990, Struc-ture and polymorphism of human telomere-associatedDNA, Cell, 63, 119-132.
29. Underwood, A. P., Louis, E. J., Borts, R. H., Stringer,J. R., and Wakefield, A. E. 1996, Pneumocystis cariniitelomere repeats are composed of TTAGGG and the sub-telomeric sequence contains a gene encoding the majorsurface glycoprotein, Mol. Microbioi, 19, 273-281.
30. Borst, P. 1991, Molecular genetics of antigenic variation,Immunol. Today, 12, A29-33.
31. Pays, E. and Steinert, M. 1988, Control of antigen geneexpression in African trypanosome, Annu. Rev. Genet,22, 107-126.
32. Pays, E., Vanhamme, L., and Berberof, M. 1994, Geneticcontrols for the expression of surface antigens in African
Downloaded from https://academic.oup.com/dnaresearch/article-abstract/3/2/55/481488by gueston 18 March 2018

64 Telomeric Expression Site of P. carinii [Vol. 3,
trypanosomes, Annu. Rev. Microbiol., 48, 25-52. 295.33. Sunkin, S. M. and Stringer, J. R. 1996, Translocation of 34. Wada, M. and Nakamura, Y. 1994, MSG gene cluster
surface antigen genes to a unique telomeric expression encoding major cell surface glycoproteins of rat Pneumo-site in Pneumocystis carinii, Mol. Microbiol., 19, 283- cystis carinii, DNA Res., 1, 163-168.
Downloaded from https://academic.oup.com/dnaresearch/article-abstract/3/2/55/481488by gueston 18 March 2018
Related Documents