Biosci. Biotech. Biochem., 59 (8), 1522-1527, lgg5 Trehalose Production by a Strain of Micrococcas varians Hideki KIzAwA,t Jun-ichi MIvlzlrt, Akira YororA,x Yukihiro KANEGAT,** Ken-ichiro MrvAG,r.wA, and Yoshio Sucrvlvrl Integrated Technology Laboratories, Takeda Chemical Industries, l0 Wadai, Tsukuba-shi, Ibaraki 30042, Japan *Institutefor Fermentqtion, Osaka,2-17-85 Juso-honmachi, Yodogawa-ku, Osaka 532, Japan xxTakasago Production Technology, Production Technology Department, Vitamin & Food Division, Takeda Chemical Industries, l-5 Takasago-cho, Takasago-shi, Hyogo 676, Japan Received February 15, 1995 A bacterium isolated from soil was found to accumulate abundant trehalose in its medium. The bacterium was identified as a strain of Micrococcus varians from its taxonomical characteristics and was designated M. varians strain No. 39. Several other strains of Micrococcns and Deinococcus also accumulated extracellular trehalose bat M, varians strain No. 39 produced the largest amount. Addition of manganese ions, and excess thiamine and nicotinamide, stimulated trehalose accumulation. Addition of 20 g/liter CSL led to the maximum conversion yield of trehalose in the production phase. Keeping the pH of the culture broth at 6.0 facilitated the maximum trehalose production rate. Cultivation of strain No. 39 tt 32"C during the growth phase and 34oC during the production phase resulted in maximal trehalose production. Trehalose non-assimilating mutants (strain No. 7 and 9) derived from strain No. 39 accumulated about 15olo more trehalose than the parent strain. Strain No. 7 produced 40mg/ml trehalose from 100mg/ml glucose under optimized conditions in a SJiter jar fermentor. Trehalose (O-a-o-glucopyranosyl-(1 --+ l)-a-o-glucopy- ranoside), a non-reducing disaccharide, is widespread in nature; it occurs in microorganisms and various plant and animal species.l) Trehalose was at flrst regarded only as a reserve metabolite, but recent research2'3) has focused on the role of trehalose in protection against environmental stresses such as desiccation, high osmolarity, frost, and heat. Trehalose also preserves various unstable products such as enzymes, foods, pharmaceuticals, and cosmetics during dry storage,a-e) and is becoming a preservative of practical importance. On an industrial scale, trehalose is prepared from yeast cells which store it, and the yield of trehalose extracted per unit amount of culture broth depends upon the quantity of yeast cells grown. If trehalose could be produced extracellularly, the yield might not depend on the number of cells, and increased production may be expected. A few microorganisms have so far been reported to accumulate trehalose extracellularly.ro rzl During our search for oligosaccharide-producing bac- teria, a bacterium isolated from soil was found to accu- mulate abundant extracellular trehalose. In this paper we describe the isolation and characterization of the product from the bacterium, the identification of the bacterium, the clariflcation of essential factors for the fermentative production of trehalose, the optimization of the culture conditions, and the derivation of a mutant strain with higher trehalose production. Materials and Methods Microorganisms. Strain No. 39, a trehalose producer isolated lrom soil, was the main organism used. The following strains were obtained from the Institute for Fermentation, Osaka (IFO): sixteen strains of Mioococcus (M. agilis IFO 15323, M. aurantiacus IFO 15364, M. conglomeratuslFO 15262, M. conglomeratus IFO 15263, M. conglomeraras IFO 1526'7, M. halobius IFO 15353, M. kristinae IFO 15354, M. luteus IFO 3067, M. luteus IFO 12708, M. lylae IFO 15355, M. nishinomiyaensis IFO 15356, M. roseus IFO 3768, M. sedentarius IFO 15357, M. varians IFO 3'765, M. sp. IFO 15265, and M. sp. IFO 15385), five strains of Deinococcus (D. erythromyxa IFO 15344, D. proteolyticus IFO 15345, D. radioduranslFO 15346, D. radiophilus IFO 15347, and D. radiopugnans IFO 15348), two strains of Sraplr ylococcus (Sta. aureus IFO 13276 and Sta. epidermidis IFO 12993), rwo strains of Streptococcus (Str. faecalis IFO 12580 and Slr. lactis IFO 12007), and a strain of Pediococcus (P. parvulus IFO 12234). Media and culture conditiozs. The compositions of the media used are shown in Table I. Microorganisms were maintained on Trypticase soy broth (TSB) agar slants containing 10g olglucose,30g ofTSB, and20g of agar per liter. (i) Shake flask experiments. Cells grown on the TSB agar slant were inoculated into a 200-ml conical flask containing 20 ml ol seed medium A and cultured for 18 h at 32"C on a rotary shaker (200 rpm). One milliliter of this culture was then translerred to a 200-ml conical flask containing 30ml of main medium A (or C) and was cultured for 5d at 32"C on a rotary shaker (200rpm). M. agilis IFO 15323 was cultured at24"C. For M. halobius IFO 15353, the main medium A with NaCl added to a final concentration of 60 g/liter was used. Main medium B was used as a minimum rnedium to examine the nutritional requirements lor the growth of strain No. 39 and the factors affecting trehalose accumulation by this strain. Then the inoculum of the seed medium into the main medium was reduced to 0.1m1/30m1. The medium containing casamino acid, nucleic acid base mixture, and vitamin mixture besides main medium B was used as complete medium. The nucleic acid base mixture contained 10mg each of adenine, guanine, thymine, cytosine, and uracil per liter. The vitamin mixture contained 400 pg each of thiamine'HCl, riboflavin, vitamin Bu, vitamin Brr, Ca-pantothenate, and nicotinamide, 10pg each of biotin and folic acid, 200pg of PABA, and 2mg of inositol per liter. (ii) Jar fermentor experiments. Seed culture was prepared as lor shake flask experiments except that seed medium B was used instead of seed medium A. One hundred and twenty-five milliliters ol the culture broth t Corresponding author. Present address: Discovery Research Laboratories I, Takeda Chemical Industries, 10 Wadai, Tsukuba-shi, Ibaraki 30042, Japan. Abbreviations: TSB, Trypticase soy broth; KPB, KH2PO4-K2HPO4 buffer; PABA,p-aminobenzoic acid; PMSF, phenylmethylsulfonyl fluoride; TES, lf-tris(hydroxymethyl)methyl-2-aminoethanesulflonic acid; CSL, corn steep liquor.

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Biosci. Biotech. Biochem., 59 (8), 1522-1527, lgg5
Trehalose Production by a Strain of Micrococcas varians
Hideki KIzAwA,t Jun-ichi MIvlzlrt, Akira YororA,x Yukihiro KANEGAT,** Ken-ichiro MrvAG,r.wA,and Yoshio SucrvlvrlIntegrated Technology Laboratories, Takeda Chemical Industries, l0 Wadai, Tsukuba-shi, Ibaraki 30042, Japan*Institutefor Fermentqtion, Osaka,2-17-85 Juso-honmachi, Yodogawa-ku, Osaka 532, JapanxxTakasago Production Technology, Production Technology Department, Vitamin & Food Division, Takeda ChemicalIndustries, l-5 Takasago-cho, Takasago-shi, Hyogo 676, JapanReceived February 15, 1995
A bacterium isolated from soil was found to accumulate abundant trehalose in its medium. The bacteriumwas identified as a strain of Micrococcus varians from its taxonomical characteristics and was designatedM. varians strain No. 39. Several other strains of Micrococcns and Deinococcus also accumulatedextracellular trehalose bat M, varians strain No. 39 produced the largest amount. Addition of manganeseions, and excess thiamine and nicotinamide, stimulated trehalose accumulation. Addition of 20 g/liter CSLled to the maximum conversion yield of trehalose in the production phase. Keeping the pH of the culturebroth at 6.0 facilitated the maximum trehalose production rate. Cultivation of strain No. 39 tt 32"C duringthe growth phase and 34oC during the production phase resulted in maximal trehalose production. Trehalosenon-assimilating mutants (strain No. 7 and 9) derived from strain No. 39 accumulated about 15olo moretrehalose than the parent strain. Strain No. 7 produced 40mg/ml trehalose from 100mg/ml glucose underoptimized conditions in a SJiter jar fermentor.
Trehalose (O-a-o-glucopyranosyl-(1 --+ l)-a-o-glucopy-ranoside), a non-reducing disaccharide, is widespread innature; it occurs in microorganisms and various plant andanimal species.l) Trehalose was at flrst regarded only as areserve metabolite, but recent research2'3) has focused onthe role of trehalose in protection against environmentalstresses such as desiccation, high osmolarity, frost, and heat.Trehalose also preserves various unstable products such as
enzymes, foods, pharmaceuticals, and cosmetics during drystorage,a-e) and is becoming a preservative of practicalimportance. On an industrial scale, trehalose is preparedfrom yeast cells which store it, and the yield of trehaloseextracted per unit amount of culture broth depends uponthe quantity of yeast cells grown. If trehalose could beproduced extracellularly, the yield might not depend on thenumber of cells, and increased production may be expected.A few microorganisms have so far been reported toaccumulate trehalose extracellularly.ro rzl
During our search for oligosaccharide-producing bac-teria, a bacterium isolated from soil was found to accu-mulate abundant extracellular trehalose. In this paper wedescribe the isolation and characterization of the productfrom the bacterium, the identification of the bacterium,the clariflcation of essential factors for the fermentativeproduction of trehalose, the optimization of the cultureconditions, and the derivation of a mutant strain with highertrehalose production.
Materials and MethodsMicroorganisms. Strain No. 39, a trehalose producer isolated lrom soil,
was the main organism used. The following strains were obtained fromthe Institute for Fermentation, Osaka (IFO): sixteen strains of Mioococcus
(M. agilis IFO 15323, M. aurantiacus IFO 15364, M. conglomeratuslFO15262, M. conglomeratus IFO 15263, M. conglomeraras IFO 1526'7, M.halobius IFO 15353, M. kristinae IFO 15354, M. luteus IFO 3067, M.luteus IFO 12708, M. lylae IFO 15355, M. nishinomiyaensis IFO 15356,
M. roseus IFO 3768, M. sedentarius IFO 15357, M. varians IFO 3'765, M.sp. IFO 15265, and M. sp. IFO 15385), five strains of Deinococcus (D.
erythromyxa IFO 15344, D. proteolyticus IFO 15345, D. radioduranslFO15346, D. radiophilus IFO 15347, and D. radiopugnans IFO 15348), twostrains of Sraplr ylococcus (Sta. aureus IFO 13276 and Sta. epidermidis IFO12993), rwo strains of Streptococcus (Str. faecalis IFO 12580 and Slr.lactis IFO 12007), and a strain of Pediococcus (P. parvulus IFO 12234).
Media and culture conditiozs. The compositions of the media used areshown in Table I. Microorganisms were maintained on Trypticase soybroth (TSB) agar slants containing 10g olglucose,30g ofTSB, and20gof agar per liter.
(i) Shake flask experiments. Cells grown on the TSB agar slant wereinoculated into a 200-ml conical flask containing 20 ml ol seed medium Aand cultured for 18 h at 32"C on a rotary shaker (200 rpm). One milliliterof this culture was then translerred to a 200-ml conical flask containing30ml of main medium A (or C) and was cultured for 5d at 32"C on a
rotary shaker (200rpm). M. agilis IFO 15323 was cultured at24"C. ForM. halobius IFO 15353, the main medium A with NaCl added to a finalconcentration of 60 g/liter was used.
Main medium B was used as a minimum rnedium to examine thenutritional requirements lor the growth of strain No. 39 and the factorsaffecting trehalose accumulation by this strain. Then the inoculum of theseed medium into the main medium was reduced to 0.1m1/30m1. Themedium containing casamino acid, nucleic acid base mixture, and vitaminmixture besides main medium B was used as complete medium. The nucleicacid base mixture contained 10mg each of adenine, guanine, thymine,cytosine, and uracil per liter. The vitamin mixture contained 400 pg eachof thiamine'HCl, riboflavin, vitamin Bu, vitamin Brr, Ca-pantothenate,and nicotinamide, 10pg each of biotin and folic acid, 200pg of PABA,and 2mg of inositol per liter.
(ii) Jar fermentor experiments. Seed culture was prepared as lor shake
flask experiments except that seed medium B was used instead of seed
medium A. One hundred and twenty-five milliliters ol the culture broth
t Corresponding author. Present address: Discovery Research Laboratories I, Takeda Chemical Industries, 10 Wadai, Tsukuba-shi, Ibaraki 30042,Japan.
Abbreviations: TSB, Trypticase soy broth; KPB, KH2PO4-K2HPO4 buffer; PABA,p-aminobenzoic acid; PMSF, phenylmethylsulfonyl fluoride;TES, lf-tris(hydroxymethyl)methyl-2-aminoethanesulflonic acid; CSL, corn steep liquor.
Trehalose Fermentation by M. varians
Table I. Composition of Media
1523
Seed MainComponent
DBA C
GlucoseTSBYeast extractCSLUrea(NH4)2HPO4(NH4)2SO4
KH2PO4MgSOo'7HrOCaClr'2HrOMnClr'4HrOFeClr'6HrOBiotinPABAThiamine HClNicotinamide
l0 g/liter30
20 gfiiter
8
5
2
4
I mg/iter48
8
30 g/liter
6
4
4
0.01
0.01
0.01
30 g,4iter
2
44
0.01
0.01
0.01
30 g/liter
8
I
2
4
0.01
0.01mg/liter0.2
10
20
60 gfliter
20
2.5I0.5
0.01
0.01 mg,4iter0.2
10
20
was translerred into a 5Jiter jar containing 2.5liters of main medium D.Cultivation was done at 32"C for 3d under the following conditions:aeration, l.25literlmin; agitation, 600 rpm. During cultivation, the pH waskept at 6.0 with 10% NaOH.
Measurement of trehalose. Ten milliliters ol culture broth werecentrifuged at 5000 x 9 for 15min. Trehalose preseat in the supernatantwas measured as extracellular trehalose by high performance liquidchromatography (HPLC) under the following conditions: column, ShodexSUGAR SZ 5532 (Shodex); eluent, acetonitrile-water (80:20, v/v); flowrate, I ml/min; temperature,55'C;detector, differential reflractometer. Cellswere harvested, washed once with water, and suspended in 6ml of 70%ethanol. This suspension was heated at 100'C lor 5min, cooled in a waterbath, and centriluged at 10,000xg for 5min. Trehalose in the resultantsupernatant was measured as intracellular trehalose. The amounts of bothextracellular and intracellular trehalose were recorded as the weight perml of culture broth.
Measurment of cell growth. Cell growth was measured by the opticaldensity at 600 nm, or as mg dry cell weight per ml of culture broth (DCW).To measure DCW, cells harvested lrom l0ml culture broth were washedonce with water, desiccated at 80'C in a vacuum oven for I 8 h, and weighed.
Purification of the product accumulated in the culture broth of strain No.39. The culture (700m1) of strain No. 39 with main medium A wascentriluged at 5000 x9 for 15min. The supernatant was concentrated to180m1 under reduced pressure and put on a column (3x30cm) packedwith activated charcoal (LH2C carbon, Takeda Chemical Industries, Ltd.,Japan). The column was washed with about 800m1 of distilled water andeluted with 300nr1 of l0% (vlv) ethanol. The eluate was concentrated to10 ml underreducedpressure, mixed graduallywith40 ml of 99.5o/o elhanol,and stood at 4"C overnight. The resulting crystals were collected byflltration, dissolved in 8 ml of distilled water, and then precipitated withappropriate amount of ethanol. The resultant crystals were washed witha small amount of ethanol and dried at 60'C lor 5 h.
Analysis. T\e melting point was measured by differential thermal analysiswith a SSC 5000 System (Seiko Denshi Kogyo, Tokyo). The specificrotation was measured with a Highly Sensitive Polarimeter SEPA-200(HORIBA, Tokyo). The water content was measured with a ThermalAnalysis Station TAS 100 (RIGAKU, Tokyo). The inlrared (IR) spectrumwas measured using the KBr method with a Fourier TransformationInfrared Spectrometer FT-530 (HORIBA, Tokyo).
Hydrolysis by trehalase. The reaction mixture (200p1) containing6.61 pmol of the product (calculated as the amount of trehalose), 40 pmolol citrate-sodium citrate buffer (pH 5.4), and 0.39units trehalase wasincubated at 3'7"C for 4 h. The mixture was heated in a boiling water bathfor 10min to stop the reaction, then 800p1 ofethanol was added and the
mixture was centriluged at 15,000xg for 5min. The supernatant wasanalysed by HPLC (column: Shodex SUGAR SP 0810, eluate: distilledwater, flow rate: 0.5m1/min, temperature: 80"C, detector: differentialrelractometer).
Isolation of trehalose non-assimilating mutants from strain No.39. Cellsol strain No. 39 were suspended (107 cells/ml) in 0.1 rrr KHTPO4-K2HPO4buffer (KPB) (pH 7.0), irradiated with UV light from a distance ol 30 cmto give a survival rate of lYo, and spread on an agar plate (G-plate)containing 59 of glucose, lg of (NH*)2HPO*.49 of MgSOa.THrO,0.025mg of biotin, 0.2mg olp-aminobenzoic acid (PABA), 0.4mg ofthiamine ' HCl, 20 mg ol nicotinamide, 0.1 mol of KPB (pH 6.0), and, 20 gof agar per liter. The plate was incubated at 32"C for 48 h. The coloniesappeared were transferred by a conventional replication method to anotherG-plate and T-plate containing trehalose instead of glucose in the G-plate,and the plates were incubated at 12"C for 48 h.
Assay of trehalose phosphorylase. Cells were suspended (about 250mgwet cell weight/ml) with 0.1u KPB (pH 7.0) containing 0.1muphenylmethylsulfonyl fluoride (PMSF), and sonicated (190W, 60min) at4'C. The cell debris was removed by centrifugation at 15,000 x g for 20 min.The resulting supernatant was used as the cell-flree extract.
The reaction mixture contained 200pmol trehalose, 50,umol KPB (pH7.0), 50 pmol N-Tris(hydroxymethyl)methyl-2-aminoethanesulfonic acid(TES) KOH buffer (pH 7.0), and cell-free extract (1 mg protein) in a totalvolume ol 1.0m1. The mixture was incubated at 30'C lor l5min and theglucose lormed was measured by a glucose oxidase-peroxidase method.One unit of enzyme activity was defined as the amount of enzl,rne releasingl,r^lmol of glucose.
Materials. Trehalose dihydrate, yeast extract, tnd TSB were purchasedfrom Sigma Chemical Co., Nihon Seiyaku Co., and Becton Dickinson andCompany, respectively. Trehalase (prepared from porcine kidney) waspurchased from Sigma Chemical Co. Other chemicals were purchased fromWako Pure Chemical Industries. Ltd.
ResultsIsolation and characterization of the product
After strain No. 39 had been cultivated in main mediumA, the culture flltrate was analyzed by HPLC (Fig. 1). Apeak at the same retention lime (l2.72min) as authentictrehalose was observed.
Crystallized material (4.8 g) was isolated from the culturebroth (700mI) and characterized by various analyses. [e]Dand melting point (mp) were + 175.4' and 90.4"C, re-spectively. These values almost coincided with those ([a]"+779.5", mp 87.1"C) of authentic trehalose (Sigma) and

ts24
No. 39 (A) and
those ([a]o + 178", mp 97"C) mentioned in the BiochemicalData Book.13) The specific rotation of the crystals wasclearly different from those (+70- *95 and -42- -38)of neotrehalose (O-a-o-glucopyranosyl-(1 -1)-B-o-gluco-pyranoside) and isotrehalose (O-B-o-glucopyranosyl-(l -l)-B-o-glucopyranoside). Hydrolysis of the crystals bytrehalase yielded only glucose. When the crystals weresupposed to be trehalose, the molar ratio of the consumedto the formed glucose was I : 1.9. The water content of thecrystals was found by thermogravimetric analysis to be9.640 , which agreed with the theoretical value for trehalosedihydrate (9.52%). The IR spectrum of the crystals wasconsistent with that of authentic trehalose (data not shown).These results demonstrate that the product accumulated inthe culture filtrate of strain No. 39 is trehalose (dihydrate).
Taxonomical studies on strain No. 39Strain No. 39 was taxonomically characterized and
identified according to Bergey's Manual of SystematicBacteriology, Vol. 2t+t (Table II). This strain was aGram-positive, nonmotile coccus (0.9-1 .0 pm in diameter)and formed slightly yellowish colonies. The G+C contentof the DNA was 69.9 (mol%). The cell wall peptidoglycancontained glutamic acid, lysine, and alanine with a molarratio of about I : I :6. The major menaquinone was MK-7(Hr). These chemotaxonomical studies suggest that thisstrain belongs to the gents Micrococcus and is very similarto M. varians. Further cultural, physiological, and bio-chemical characteristics were examined. The strain grew onSimmons citrate agarls) and produced acid from glucose.The cells reduced nitrate to nitrite. The strain had ureasebut not oxidase activity. From these results, strain No. 39
was identifled as a type of M. varians.
Extracellular accumulation of trehalose by Micrococcus andDeinococcus species
Trehalose productivity was investigated in sixteen strainsof Micrococcus spp and five strains of the related genus,Deinococcus spp, in flask culture with main medium A(Table III). Most strains tested accumulated extracellulartrehalose. In M. agilis IFO 15323, M. luteus IFO 3067, M.
Ktzlwt et al.
Table II. Taxonomic Characteristics of Strain No. 39
Characteristic
ABIN 1II
il lar€il Erc
111lrl"rr'ltilllilirrllllll, ;U[ lilrllul \ --lllul
llllll0 10 200 '1 0 2t
Retention time (min)
1. HPLC Profiles ol Culture Filtrate of St
hentic Trehalose (B).
Strain No. 39
I
20
f Strain
Grarn stainingMorphologyCell size
MotilityColor of cellsCatalaseOxidaseCellular fatty acids
Amino acids of peptidoglycan(Peptidoglycan type)
MenaquinoneG+C content (mol%)Urease
B-GalactosidasePhosphataseArginine dihydrolaseAcetoinNitrate reductionHydrolysis ol gelatin
StarchEsculin
Polyoxyethylenesorbitanmonooleate
Growth onSimmon's citrate agarInorganic nitrogen agarNutrient agar with
10% NaCl15% NaCl
Acid production lromo-Glucosen-Fructoseo-Galactoseo-Mannoseo-Xylosel-RhamnoseMaltoseLactoseSucroseTrehaloseGlycerolSorbitol
Temp. range lor growth ('C)Optimum temp. ('C)Strict aerobesLysozyme susceptibility
(400 pglml)(800 pglml)
+Cocci
0.9 1.0pm
Slightly yellowish+
anteiso-15 : 0 + anteiso-17 : 0
(No hydroxy acids)Glu : Lys : A1a: 1.00 : 0.97 : 5.80t--Lys-r-Ala. u (43a, 411.7)
MK-7 (Hr)69.9
++
++
+
+
+++
+
+
++
20 3'.7"C
30'c+
ResistantSensitive
Fig.Aut
luteus IFO 12708, M. varians IFO 3765, D. erythromyxaIFO 15344, D. proteolyticus IFO 15345, and D. radiopugnansIFO 15348, the amount of trehalose accumulated in theculture broth was over 2mgfml. Among these species, D.proteolyticus IFO 15345 produced the largest amount oftrehalose (8.34mg/ml). Under the same conditions, strainNo. 39 produced 1.4 times as much extracellular trehaloseas D. proteolyticus IFO 15345. These results indicate thatmany Micrococcus and Deinococczs strains are capable ofaccumulating extracellular trehalose. Further, to investigatewhether trehalose producion was a general feature ofGram-positive cocci or not, trehalose productivity in several
strains of Staphylococcus, Streptococcus, ar,d Pediococcus
spp was tested (Table III). All the strains tested, however,
did not accumulate even intracellular trehalose. Therefore,the accumulation of trehalose was thought to be a char-acteristic of strains of Micrococcus and Deinococcus spp.
Factors affecting the extracellular accumulation of trehaloseby sftain No. 39Strain No. 39 can grow in media containing yeast extract
but not in synthetic media composed only of a carbonsource, inorganic nitrogen source, and inorganic salts. Wethen examined the nutritional requirements of the strain,findingthat simultaneous supply of biotin, PABA, thiamine,and nicotinamide was very important for the growth ofstrain No. 39 (data not shown).
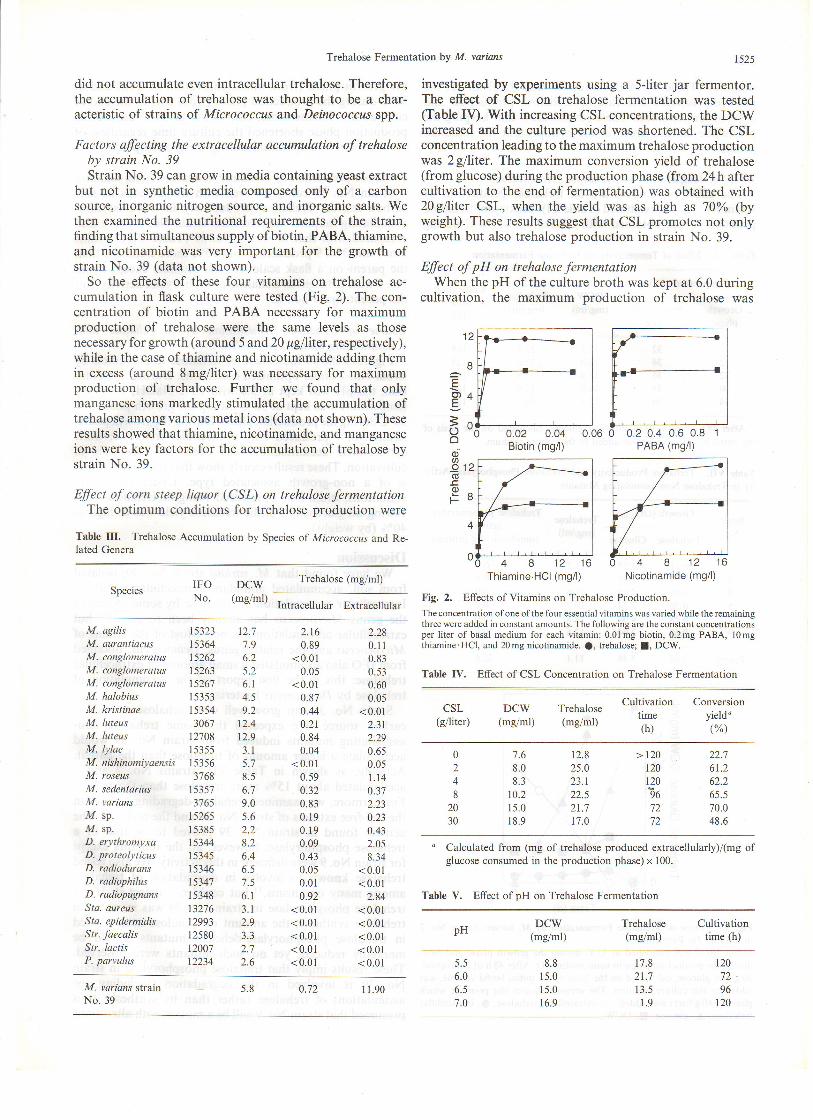
So the effects of these four vitamins on trehalose ac-cumulation in flask culture were tested (Fig. 2). The con-centration of biotin and PABA necessary for maximumproduction of trehalose were the same levels as thosenecessary for growth (around 5 and,20 1tgfiiter, respectively),while in the case of thiamine and nicotinamide adding themin excess (around 8 mg/iter) was necessary for maximumproduction of trehalose. Further we found that onlymanganese ions markedly stimulated the accumulation oftrehalose among various metal ions (data not shown). Theseresults shqwed that thiamine, nicotinamide, and manganeseions were key factors for the accumulation of trehalose bystrain No. 39.
Effect of corn steep liquor (CSL) on trehalose fermentationThe optimum conditions for trehalose production were
Table III. Trehalose Accumulation by Species of Micrococcus and Re-lated Genera
IFO DCWNo. (mglml)
Trehalose (mglml)Species
Intracellular ExtracellularFig. 2. Effects of Vitamins on Trehalose Production.
The concentration ofone of the four essential vitamins was varied while the remainingthree were added in constant amounts. The following are the constant concentrationsper liter of basal medium for each vitamin: 0.0lmg biotin, 0.2mg PABA, 10mgthiamine'HCl, and 20mg nicotinamide. a, trehalose; L DCW.
Table IV. Effect ol CSL Concentration on Trehalose Fermentation
Trehalose Fermentation by M. varians 1525
investigated by experiments using a 5-liter jar fermentor.The effect of CSL on trehalose fermentation was tested(Table IV). With increasing CSL concentrations, the DCWincreased and the culture period was shortened. The CSLconcentration leading to the maximum trehalose productionwas 2gfiiter. The maximum conversion yield of trehalose(from glucose) during the production phase (from 24 h aftercultivation to the end of fermentation) was obtained with20 glliter CSL, when the yield was as high as 10% (byweight). These results suggest that CSL promotes not onlygrowth but also trehalose production in strain No. 39.
Effect of pH on trehalose fermentationWhen the pH of the culture broth was kept at 6.0 during
cultivation, the maximum production of trehalose was
0.02 0.04Biotin (mg/l)
0.06 0 0.2 0.4 0.6 0.8 1
PABA (mg/l)
M. agilisM. aurantiacusM. conglomeratusM. conglomeratusM. conglomeratusM. halobiusM. kristinaeM. luteusM. luteusM. lylaeM. nishinomiyaensisM. roseus
M. sedentariusM. yarians
M.sp.M.sp-D. erythromyxaD. proteolyticusD. radioduransD. radiophilusD. radiopugnansSta. aureusSta. epidermidisStr. faecalisStr. lactisP. parvulus
t5323 12.7
15364 7.915262 6.215263 5.215267 6.1
15353 4.st5354 9.23067 12.4
12708 12.9
15355 3.115356 5.73768 8.5
15357 6.7376s 9.0
15265 5.615385 2.215344 8.215345 6.415346 6.5
15347 7.5
15348 6.1
13276 3.1
12993 2.912580 3.3
12007 2.712234 2.6
2.t60.89
< 0.01
0.05<0.01
0.870.440.2t0.840.04
<0.010.590.320.830.190. 19
0.090.430.050.01
0.92<0.01<0.01<0.01<0.01< 0.01
2.28
0.11
0.83
0.53
0.600.05
<0.012.31
2.290.65
0.05
1.14
0.372.23
0.23
0.43
2.058.34
<0.01<0.01
2.84<0.01<0.01<0.01<0.01<0.01
CSL(g/liter)
Cultivationtime(h)
Conversionyield'(:%)
DCW Trehalose(mg/ml) (mg/ml)
0
2
4
I20
30
7.6 12.8
8.0 25.08.3 23.1
10.2 22.515.0 21 .7
18.9 t7.0
>120t20t2096
72
72
22.761.2
62.265.5
70.048.6
' Calculated from (mg of trehalose produced extracellularly)/(mg ofglucose consumed in the production phase) x 100.
Table V. Effect of pH on Trehalose Fermentation
nH DCW' (mg/ml)
Trehalose Cultivation(me/ml) time (h)
M. varians strainNo. 39
5.5
6.06.5'7.O
8.8
15.0
15.0
16.9
t7.821.7
13.5
1.9
12072
96120
Thiamine.HCl (mg/l)
5.8 0.72 1 1.90
1526
obtained (Table V). A deviation of only 0.5 from pH 6.0markedly lowered the rate of trehalose production anddelayed fermentation. These results show that keeping thepH of the culture broth at 6.0 is important for trehaloseproduction.
Effect of temperature on trehalose fermentationThe effect of temperature on trehalose production by
strain No. 39 was examined (Table VI). Maximum trehaloseproduction was obtained with cultivation at 32"C during
Table VI. Effect of Temperature on Trehalose Fermentation
Temperature ('C)
the growth phase (from initial cultivation to 24h) and at34'C during the production phase (from 24h to the end ofcultivation). Furthermore, cultivation at 34"C during theproduction phase shortened the culture time regardless ofthe incubation temperature during the growth phase.
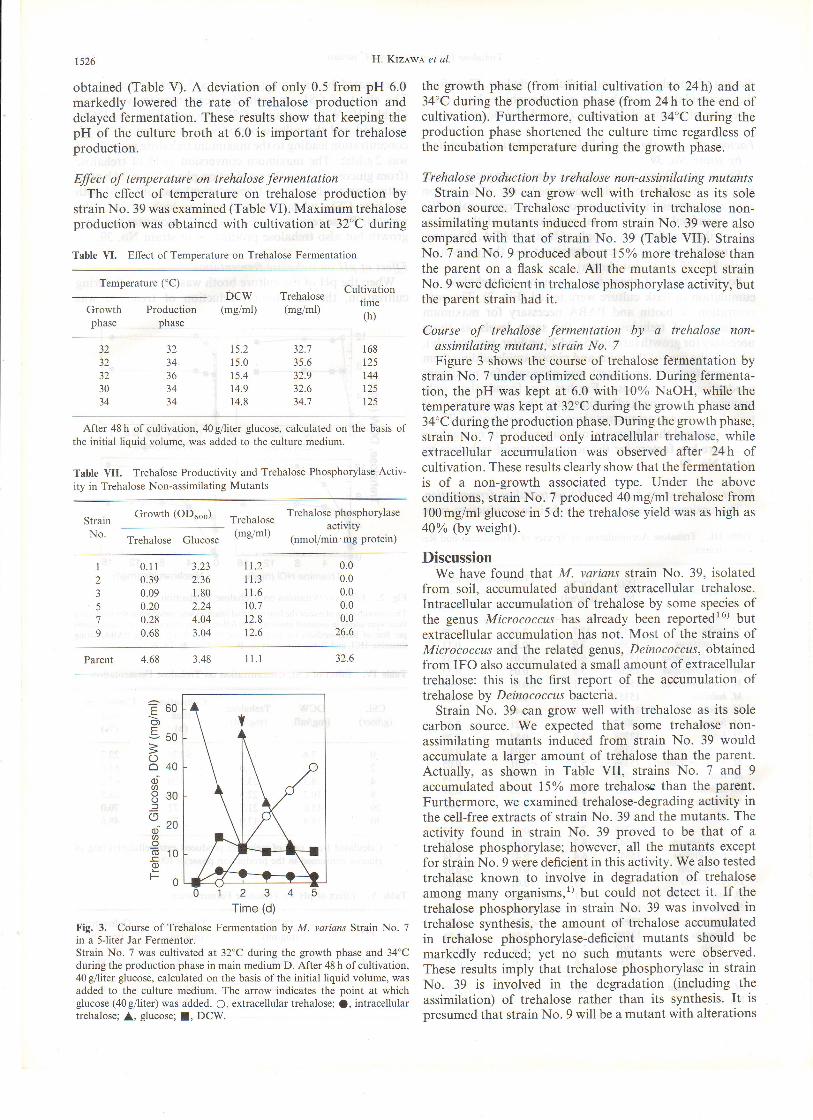
Trehqlose production by trehalose non-assimilating mutantsStrain No. 39 can grow well with trehalose as its sole
carbon source. Trehalose productivity in trehalose non-assimilating mutants induced from strain No. 39 were alsocompared with that of strain No. 39 (Table VII). StrainsNo. 7 and No. 9 produced about l5o/o morc trehalose thanthe parent on a flask scale. All the mutants except strainNo. 9 were deficient in trehalose phosphorylase activity, butthe parent strain had it.
Course of trehalose fermentation by a trehalose non-assimilating mutant, strain No. 7Figure 3 shows the course of trehalose fermentation by
strain No. 7 under optimized conditions. During fermenta-tion, the pH was kept at 6.0 with 10% NaOH, while thetemperature was kept at 32"C during the growth phase and34'C during the production phase. During the growth phase,
strain No. 7 produced only intracellular trehalose, whileextracellular accumulation was observed after 24h ofcultivation. These results clearly show that the fermentationis of a non-growth associated type. Under the aboveconditions, strain No. 7 produced 40mg/ml trehalose from100 mg/ml glucose in 5 d: the trehalose yield was as high as
a0% (by weight).
DiscussionWe have found that M. varians strain No. 39, isolated
from soil, accumulated abundant extracellular trehalose.Intracellular accumulation of trehalose by some species ofthe genus Miuococcus has already been reportedt6) butextracellular accumulation has not. Most of the strains ofMicrococcus and the related genus, Deinococcus, obtainedfrom IFO also accumulated a small amount of extracellulartrehalose: this is the first report of the accumulation oftrehalose by Deino co ccus bacteria.
Strain No. 39 can grow well with trehalose as its solecarbon source. We expected that some trehalose non-assimilating mutants induced from strain No. 39 wouldaccumulate a larger amount of trehalose than the parent.Actually, as shown in Table VII, strains No. 7 and 9
accumulated about l5oh more trehalose than the parent.Furthermore, we examined trehalose-degrading activity inthe cell-free extracts of strain No. 39 and the mutants. Theactivity found in strain No. 39 proved to be that of a
trehalose phosphorylase; however, all the mutants exceptfor strain No. 9 were deflcient in this activity. We also tested
trehalase known to involve in degradation of trehaloseamong many organisms,l) but could not detect it. If thetrehalose phosphorylase in strain No. 39 was involved intrehalose synthesis, the amount of trehalose accumulatedin trehalose phosphorylase-deficient mutants should be
markedly reduced; yet no such mutants were observed.These results imply that trehalose phosphorylase in strainNo. 39 is involved in the degradation (including theassimilation) of trehalose rather than its synthesis' It is
presumed that strain No. 9 will be a mutant with alterations
H. Krzawr et al.
Growthphase
Productionphase
DCW Trehalose(mglml) (mg/ml)
Cultivationtime(h)
JZ
32
32
30
34
32
34
36
34
J1+
15.2
15.0
t5.4t4.914.8
32;7
35.632.932.634.7
168
t25r44r25t25
After 48h of cultivation, 4lgfliter glucose, calculated on the basis ofthe initial liquid volume, was added to the culture medium.
Tabte VII. Trehalose Productivity and Trehalose Phosphorylase Activ-ity in Trehalose Non-assimilating Mutants
StrainNo.
Growth(oDuoo) Trehalose
Trehalose Glucose (mg/ml)
Trehalose phosphorylaseactivity
(nmol/min'mg protein)
I2
3
5'1
9
0.1 1
0.390.090.200.280.68
3.23
2.361.80
2.244.04
3.04
tl.2I 1.3
I t.6t0.712.8
12.6
0.00.00.00.00.0
26.6
Parent 4.68
Time (d)
Fig. 3. Course of Trehalose Fermentation by M. wrians Strain No. 7
in a Sliter Jar Fermentor.Strain No. 7 was cultivated at 32"C during the growth phase and 34"Cduring the production phase in main medium D. Alter 48 h of cultivation,40 gfirter glucose, calculated on the basis of the initial liquid volume, wasadded to the culture medium. The arrow indicates the point at whichglucose (40g/liter) was added. O, extracellular trehalose; !, intracellulartrehalose; l, glucose; l, DCW.
E60o)Fe50
=Oo40G)U)
830=(,oi 20U)o810I'0

somewhere in the steps involved in the assimilation oftrehalose. The reason why the mutant strain No. 9 producedan elevated amount of trehalose is now under investigation.We have also purified and characterized the trehalosephosphorylase in strain No. 39, and will report on this inthe near future.
Our results demonstrated that strain No. 7 produced thelargest amount of trehalose in the strains we tested: that is,it produced 40mg/ml trehalose from l0Omg/ml glucose in5 d under optimized conditions. In conclusion, M. variansstrain No. 39 and its trehalose non-assimilating mutants(especially strain No. 7) produced a superior conversionyield of carbon source into trehalose than other trehaloseproducing microorganisms such as yeasts or Brevibacteriumlactofermentr*,") and could be of potential use formanufacturing trehalose on an industrial scale.
References1) A. D. Elbein, Ady. Carbohyd. Chem. Biochem.,30,227256 (1974).2) J. H. Crowe, L. M. Crowe, and D. Chapman, Science, 223,'701-703
( 1 e84).3) G. M. Gadd, K. Chalmers, and R. H. Reed,, FEMS Mioobiol. Lett.,
Trehalose Fermentation by M. varians
4)5)
6)
7)
8)e)
10)
11)
12)
l3)
t4)
1521
48.249-254 (1987).
J. M. Thevelein, Microbiol. Rev.,48,42 59 (1984).J. H. Crowe, J. F. Carpenter, L. M. Crowe, and T. J. Anchordoguy,Cryobiology, 27, 219-231 (1990).J. H. Crowe, F. A. Hoekstra, and L. M. Crowe, Annu. Rey. Phyliol.,s4, s79 s99 (t992).I. Kaasen, P. Falkenberg, O. B. Styrvold, and A. R. Strom, J.Bacteriol., 174, 889-898 (1992).B. Roser, Trends Food Sci. Technol.,2, 16G169 (1991).C. Colaco, S. Sen, M. Thangavelu, S. Pinder, and B. Roser, .Blo.Technol., 10, 1007-101 I (1992).T. Suzuki, K. Tanaka, and S. Kinoshita, Agric. Biol. Chem.,33,190 195 (1969).K. Kunugita, K. Nakahara, H. Aoki, and H. Imanaka, Japan KokaiTokkyo Koho, 75154485 (May 31, 1914).T. Tsuchida, Y. Murakami, and Y. Nishimoto, Japan Kokai TokkyoKoho, 9321 1882 (Feb. 13, 1992).
"Biochemical Data Book I," ed. by Nihon Seikagakukai, TokyoKagaku Doujin, 1979, pp. 464.K. H. Schleifer, in "Bergey's Manual of Systematic Bacteriology,"Yol.2, ed. by P. H. A. Sneath, N. S. Mair, M. E. Sharpe, and J. G.Holt, TheWilliams &Wilkins Co., Baltimore, 1986, pp. 1003-1035.W. E. Kloos, T. G. Tornabene, and K. H. Schleifer, Int. J. Syst.Bac t eriol., U,'1 9-101 (197 4).Z.l. Ahmad, J. R. Alden, and M. D. Montague, J. Gen. Microbiol.,121, 483 486 (1980).
ls)
t6)
Related Documents











