FACULTY OF HEALTH AND MEDICAL SCIENCES UNIVERSITY OF COPENHAGEN PhD thesis Julie Svalø K v 7 channels Expression, regulation and function in the urinary bladder

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

FA C U LT Y O F H E A LT H A N D M E D I C A L S C I E N C E S !U N I V E R S I T Y O F C O P E N H A G E N
PhD thesis Julie Svalø
!Kv7 channels Expression, regulation and function in the urinary bladder

!

!PhD Thesis !
!!!
Kv7 channels Expression, regulation and function in the urinary bladder !
!Julie Svalø, M.Sc.
!!
Smooth Muscle Research Center Copenhagen University Hospital, Køge !Faculty of Health and Medical Sciences
University of Copenhagen !

Title: Kv7 channels: Expression, regulation and function in the urinary bladder
!Author: Julie Svalø, M.Sc. Pharmaceutical Sciences
!Department: Smooth Muscle Research Center, Copenhagen University Hospital, Køge,
Denmark
Institution: Faculty of Health and Medical Sciences, University of Copenhagen, Denmark
Submitted: September 19, 2014
!!Academic advisors:
Associate Professor Majid Sheykhzade, Department of Drug Design and Pharmacology,
University of Copenhagen, Denmark
Medical Director Pierre N. Bouchelouche, Smooth Muscle Research Center, Department of
Clinical Biochemistry, Copenhagen University Hospital, Køge, Denmark
Professor Jørgen Nordling, Department of Urology, Copenhagen University Hospital, Herlev,
Denmark
!!Evaluating Committee:
Associate Professor Uffe Kristiansen, Department of Drug Design and Pharmacology,
University of Copenhagen, Denmark
Professor Axel Forman, Department of Clinical Medicine, University of Aarhus, Denmark
Professor Iain Greenwood, Biomedical Sciences, St. George’s, University of London, United
Kingdom

Contents!PREFACE 1 ACKNOWLEDGEMENTS 3 SUMMARY 4 RESUME 5 ABBREVIATIONS 6 INTRODUCTION 7
THE URINARY BLADDER 8 PHYSIOLOGY 8 OVERACTIVE BLADDER SYNDROME 11 POTASSIUM CHANNELS AS A PUTATIVE TARGET 12
KV7 CHANNEL PHYSIOLOGY 13 STRUCTURE 13 REGULATION 14 PHARMACOLOGY 16 TISSUE DISTRIBUTION 18
AIM 20 EXPERIMENTAL METHODS 21
ANIMALS AND PATIENTS 21 GENE EXPRESSION 21
RNA ISOLATION 22 RT-QPCR 23
MYOGRAPHY 24 TISSUE HANDLING 24 LENGTH-TENSION RELATION 25 CONTRACTILE RESPONSES 26 DENERVATION PROTOCOL 27
STATISTICAL ANALYSIS 28
RESULTS AND DISCUSSION 29 KCNQ AND KCNE EXPRESSION WITHIN THE URINARY BLADDER 29
EXPRESSION LEVELS IN NORMAL BLADDER 29 EXPRESSIONAL CHANGES IN OUTFLOW OBSTRUCTED BLADDER 30
KV7 CHANNEL FUNCTIONALITY IN THE DETRUSOR. 32 MEDIATORS OF RESTING TONE 32 KV7 CHANNEL SUBTYPE DIFFERENTIATION 33
REGULATION OF KV7 CHANNELS IN THE DETRUSOR. 35 FUNCTIONAL EVALUATION OF LOCALISATION 35 PIP2 DEPENDENCY 37 INVOLVEMENT OF KV7 CHANNELS IN THE GS PATHWAY 37
CLINICAL IMPACT OF OUR FINDINGS 40
CONCLUSION AND PERSPECTIVES 41 BIBLIOGRAPHY 42 APPENDICES 58

Preface
This thesis is based on the following original manuscripts made by Julie Svalø during her
three year PhD program at the Faculty of Health and Medical Sciences, University of
Copenhagen. The work was conducted at the Smooth Muscle Research Center at Copenhagen
University Hospital at Køge in collaboration with the Department of Drug Design and
Pharmacology, University of Copenhagen, Denmark and the Departments of Urology at
Herlev, Skejby and Næstved Hospitals.
!In the text the manuscripts are referred to by their Roman numerals:
!I. Svalø J, Nordling J, Bouchelouche K, Andersson KE, Korstanje C, Bouchelouche
P (2013). The novel β3-adrenoceptor agonist mirabegron reduces carbachol-induced
contractile activity in detrusor tissue from patients with bladder outflow obstruction
with or without detrusor overactivity. Eur. J. Pharmacol. 699: 101-105.
!II. Svalø J, Bille M, Parameswaran Theepakaran N, Sheykhzade M, Nordling J,
Bouchelouche P (2013). Bladder contractility is modulated by Kv7 channels in pig
detrusor. Eur. J. Pharmacol. 715: 312-320.
!III. Svalø J, Sheykhzade M, Nordling J, Matras C, Bouchelouche P. Functional and
molecular evidence for Kv7 channel subtypes in human detrusor from patients with
and without bladder outflow obstruction. Resubmitted to PLOS ONE, august 2014.
!!!!!!!!!
"1

The following original and peer-reviewed papers are related to the work described in the
thesis at hand, but not included in the thesis, and are referred to as regular references:
!Rode F*, Svalø J*, Sheykhzade M, Rønn LC (2010). Functional effects of the KCNQ
modulators retigabine and XE991 in the rat urinary bladder. Eur. J. Pharmacol. 638:121-127. *First author
Svalø J, Hansen HH, Rønn LC, Sheykhzade M, Munro G, Rode F (2011). Kv7 positive
modulators reduce detrusor overactivity and increase bladder capacity in rats. Basic Clin.
Pharmacol. Toxicol. 110:145-153.
!!The following peer-reviewed papers are related to smooth muscle cell function and were also
prepared during the PhD period, but not included in the thesis:
!Rosenbaum ST, Svalø J, Nielsen K, Larsen T, Joergensen J, Bouchelouche P (2012).
Immunolocalization and expression of small-conductance calcium-activated potassium
channels in human myometrium. J. Cell. Mol. Med. 16: 3001-3008.
!Rosenbaum ST, Svalø J, Larsen T, Joergensen J, Bouchelouche P (2013). Relaxant Effect of
Cyppa In Vitro and Expression Analysis of SK3 Splice Variants in the Myometrium of
Pregnant and Non-Pregnant Women. Gynecol. Obstet. 3:171.
!!During her PhD studies Julie Svalø has been the academic supervisor for master students
Michala B. Nielsen and Neeraja P. Theepakaran, who wrote the master thesis entitled:
Characterisation of the voltage-gated potassium channel Kv7 in pig detrusor muscle, in 2012.
"2

Acknowledgements
I would like to express my sincere gratitude to my supervisors: Majid Sheykhzade for your
deep commitment in my research. Jørgen Nordling for introducing me to the clinic and for
making sure that we could obtain biopsies. Pierre N. Bouchelouche for providing a state-of-
the-art laboratory and for giving me the chance to evolve as a scientist by believing in me.
A special thank to present and former colleagues at the Smooth Muscle Research Center:
master students Neeraja P. Theepakaran and Michala Bille for being dedicated in the
laboratory and for being joyful company; PhD student Sofia Rosenbaum for interesting
scientific discussions, and for including me in her research; Technician Majken Jansen for
highly qualified technical support and for being a humorous and ever supporting friend; and
PhD student Grith Eng for highly qualified feedback on presentations. I would like to thank
Sonja Jensen for always being helpful and of course all my other colleagues at the Department
of Clinical Biochemistry for creating a warm working atmosphere.
The studies on human bladder specimens could not have been conducted without the deep
insight into the literature by my co-authors Cees Korstanje and Karl-Erik Andersson; the
commitment by the doctors and nurses at the Urological Departments at Herlev, Næstved and
Skejby Hospitals, especially Christina Matras and Jørgen Bjerggaard Jensen; and the patients
for their interest in donating tissue for the studies. The experiments on pig bladder specimens
were only possible due to the help from The Danish Meat Trade College, Roskilde and the
Department of Basic Animal and Veterinary Sciences, University of Copenhagen.
I would like to thank my former colleagues at NeuroSearch A/S, Frederik Rode and Lars
Christian Rønn, for introducing me to potassium channels in the urinary bladder and for
giving me the initial boost to become a scientist. I would like to thank: Vanderbilt University,
USA and NeuroSearch A/S, Denmark for providing compounds, and: Astellas Pharma Europe
B.V., Aase og Ejnar Danielsens Fond, A.P. Møller Fonde, Region Sjællands
Sundhedvidenskabelige Forskningsfond, Pharmadanmark and especially the Department of
Clinical Biochemistry at Køge Hospital for supporting the project financially.
Finally, I would like to thank my parents for teaching me to set high standards. A special
thank to my husband Theis for your crucial support and to our sweet son Storm for reminding
me that there is a life worth living outside of the laboratory.
"3

SUMMARY
Summary The urinary bladder is a highly innervated organ whose anatomy supports both urine collection and elimination. As a result of this complexity, more than 10% of the adult population suffers from overactive bladder syndrome (OAB), characterised by a dysfunctional storage of urine. OAB is often associated with bladder outflow obstruction (BOO) and the syndrome is characterised by urgency, frequency, nocturia and incontinence. Following on from this, it is clear that OAB has a huge negative impact on the patients’ quality of life, especially since present treatment is poor. Potassium channels are important regulators of smooth muscle cells, in the bladder as elsewhere in the body. Clinical studies on the Kv7.2-7.5 potassium channel activator retigabine have shown that the opening of Kv7 channels leads to urinary retention, presumably through relaxation of the bladder musculature. Therefore, the aims of this thesis are to determine whether Kv7 channels are putative targets for the treatment of OAB. This is accomplished by exploring the expression, regulation and function of Kv7 channels in the urinary bladder. The β3-adrenoceptor agonist mirabegron has recently been approved for the treatment of OAB, therefore, we aim at investigating whether Kv7 channels interplay with β3-adrenoceptors. The experiments are performed on bladder biopsies from both patients with normal bladder function and patients with BOO. We also examine pig bladder specimens in order to determine whether, or not, pigs constitute an appropriate model for human bladder function with regards to Kv7 channels. We show that the gene KCNQ1, encoding Kv7.1 channels, is up-regulated in bladder tissue from patients with BOO compared to that seen in patients with a normal bladder function. This suggests that Kv7.1 channels are up-regulated as a compensatory mechanism in order to avoid detrusor overactivity. We also show, both functionally and molecularly, that Kv7.4 and/or Kv7.5 channels are important for the function of the human and pig urinary bladder. Hence, there is an overlap in Kv7 channel functionality between pig and human urinary bladders. Following on from this, we studied the relaxation induced by the β3-adrenoceptor agonist mirabegron in human bladder samples. We end up confirming the proof-of-principle that β3-adrenoceptor agonists constitute promising pharmaceutical agents for the treatment of OAB; moreover, we show that the relaxation mediated via Kv7 channels and that brought about by β-adrenoceptors are independent mechanisms. We can therefore conclude that Kv7 channel activation provides a novel pathway for reducing urinary bladder contractility. In turn, one can imply from this, that positive Kv7 channels modulators could provide a novel treatment of OAB.
"4

RESUME
Resume Urinblæren er et komplekst organ, der er rigt innerveret og har en anatomi der understøtter både opsamling og ladning af urin. På grund af denne kompleksitet, lider mere end 10 % af den voksne befolkning af overaktiv blæresyndrom (OAB), som er kendetegnet ved at blærens reservoirfunktion er dysfunktionel. OAB ses ofte i forbindelse med blæreobstruktion (BO) og syndromet er karakteriseret ved bydende vandladningstrang, hyppig vandladning, nykturi og inkontinens. OAB har stor indvirkning på de ramte patienters livskvalitet og effekten af den nuværende behandling er beskeden. Kalium kanaler er vigtige regulatorer af glatte muskelceller, også i urinblæren. Kliniske studier med stoffet retigabine, som aktiverer Kv7.2-7.5 kalium kanaler, har vist at åbning af Kv7 kanaler medfører urinretention, formentlig via afslapning af blæremuskulaturen. Formålet med denne afhandling er at belyse hvorvidt Kv7 kanaler er et muligt farmakologisk angrebsmål for nye lægemidler til behandling af OAB. I forsøgene undersøges udtrykket, reguleringen og funktionen af Kv7 kanaler i urinblæren. β3-adrenoceptor agonisten, mirabegron, er for nyligt blevet godkendt til behandling af OAB, derfor undersøger vi om Kv7 kanaler interagerer med β3-adrenoceptorer. Forsøgene er udført på blærebiopsier fra patienter med en normal blærefunktion og fra patienter med BO. Derudover er væv fra grises urinblærer undersøgt for at bestemme hvorvidt grisen er en velegnet model for den menneskelige blærefunktion i relation til Kv7 kanaler. Forsøgene viser at genet KCNQ1, der koder for Kv7.1 kanalen, er opreguleret i blærebiopsier fra patienter med BO i forhold til gruppen af patienter med normal blærefunktion. Dette tyder på at Kv7.1 kanaler opreguleres som en kompensatorisk mekanisme for at undgå overaktivitet af blæremuskulaturen. Vores forsøg viser også, både funktionelt og molekylært, at Kv7.4 og/eller Kv7.5 er vigtige for funktionen af både menneskets og grisens urinblære. Dermed er der overlap mellem Kv7 kanalers funktion i menneskers og grises urinblærer. Vi bekræfter også, at β3-receptor agonisten mirabegron har en afslappende effekt på blæremuskulatur fra mennesker og at β3-adrenoceptor agonister er lovende terapeutiske midler til behandling af OAB. Derudover viser vi at den afslappende effekt medieret via Kv7 kanaler er uafhængig af afslapning medieret via β-adrenoceptorer. Vi har vist at aktivering af Kv7 kanaler er en nyopdaget mekanisme til at reducere blæremuskulaturens sammentrækning. Dette betyder at aktivatorer af Kv7 kanaler er potentielle lægemidler til behandling af OAB. !!!
"5

ABBREVIATIONS
Abbreviations ACh acetylcholine ACTB actin, beta 4-AP 4-aminopyridine ATP adenosine triphosphate ATP5G ATP synthase BKCa large-conductance calcium-activated potassium channels B2M beta-2-microglobulin BOO bladder outflow obstruction cAMP cyclic adenosine monophosphate Cav voltage-gated calcium channels cDNA complementary deoxyribonucleic acid DO detrusor overactivity eEF-1 eukaryotic elongation factor 1 gamma-like protein GABAA gamma-aminobutyric acid type A receptor GAPDH glyceraldehyde-3-phosphate dehydrogenase GPXI glutathione peroxidase 1 GPI glucose phosphate isomerase GSR glutathione reductase H2S hydrogen sulphide IP3 inositol trisphosphate IPSS international prostate symptom score KATP ATP-sensitive potassium channel KPSS potassium physiological saline solution Kv7 voltage-gated potassium channel M muscarinic µM micromolar mM millimolar mN millinewton mRNA messenger ribonucleic acid N number of strips n number of patients ng nanogram OAB overactive bladder Oligo(dt) short sequence deoxy-thymine nucleotides PIP2 phosphatidylinositol-4,5-biphosphate PSS physiological saline solution RT-qPCR reverse transcription quantitative real-time polymerase chain reaction 18S 18S rRNA gene SEM standard error of the mean SF3A1 splicing factor 3A1 SDHA succinate dehydrogenase complex SHAS2 hyaluronan synthase 2 SKCa small-conductance calcium-activated potassium channels TOP1 topoisomerase DNA I UBC polyubiquitin
"6

INTRODUCTION
Introduction
Awoken by accident while I completed by bachelor studies in medicinal chemistry at the
University of Aarhus, my fascination with the urinary bladder led to it becoming my main
research focus. My bachelor project involved the organocatalytic synthesis of the
antimuscarinic drug, darifenacin, in Professor Karl-Anker Jørgensen’s laboratory. While
writing my bachelor thesis my attention was drawn towards the physiology of this unique
organ - the urinary bladder. My growing interest in physiology and especially pharmacology
inspired me to move to Copenhagen to study pharmaceutical sciences. At the Faculty of
Pharmaceutical Sciences, Associate Professor Majid Sheykhzade introduced me to a master
thesis project at NeuroSearch A/S. It was a well-known fact, that a large proportion of the
adult human population suffers from overactive bladder syndrome (OAB) (>10%) and that the
use of antimuscarinics does not provide sufficient alleviation from the symptoms. In addition,
compliance was very poor due to side effects. Accordingly, there was a desperate need for
novel targets in the bladder. I therefore continued my studies on urinary bladder function, this
time focussing on voltage-gated potassium (Kv7) channels in rats, under the supervision of
Research Scientist Frederik Rode. Together, we found evidence that these channels were
indeed important for urinary bladder function, both in normal bladders and in models of OAB
(Rode et al., 2010; Svalø et al., 2011). As a part of my studies, I spend a few weeks at the
Smooth Muscle Research Center at Køge Hospital, under the supervision of Medical Director
Pierre N. Bouchelouche. Here, preliminary data from pigs confirmed our previous findings in
rats. Having obtained my Masters Degree I continued to study Kv7 channel function, this time
in the urinary bladder of humans at Køge Hospital in collaboration with Professor Jørgen
Nordling (Department of Urology, Herlev Hospital), and Associate Professor Majid
Sheykhzade (Department of Drug Design and Pharmacology, Copenhagen University). The
primary focus of the thesis at hand was to describe the expression, regulation and function of
Kv7 channels in human and pig urinary bladder (Paper I-III) with a view to elaborate on
previous findings. The aim of the following chapter is to provide the reader with background
information and an overview of the field at hand. To achieve this, I have included a short
introduction to the physiology and pathophysiology of the bladder, followed by a section
focusing on Kv7 channel structure, regulation and pharmacology.
"7

INTRODUCTION
The Urinary Bladder
Physiology
The urinary bladder is a hollow organ that is responsible for the collection and voiding of
urine (Fig. 1). The bladder wall is composed of an external layer of peritoneum followed by
three inner layers of smooth muscle, collectively known as the detrusor. Whereas the outer
and inner muscle layers of the detrusor are primarily longitudinally oriented, the middle layer
is circularly oriented (McCrea, 1926). The inner wall of the bladder is covered by urothelium
which acts as a barrier between the urine and the muscles. Equally important is the fact that
the urothelium releases, amongst other, adenosine triphosphate (ATP) (Ferguson et al., 1997)
and nitric oxide (Birder et al., 1998). Hence, the urothelium acts as a sensor in relation to its
surroundings, in addition to which it possibly transduces information to the afferent nerves
and smooth muscle cells (Birder et al., 2010).
Urine is led from the kidneys through the ureters to the bladder. Previous studies have shown
that as animal bladders are filling, the sympathetic efferents release noradrenalin, which in
"8
Figure 1. Physiology of the urinary bladder. The ureters leads urine to the bladder. The smooth muscle layer, detrusor, of the bladder is covered by peritoneum. The inner wall of the bladder is lined with urothelium. (Downloaded for free at http://cnx.org/content/col11496/latest/.)

INTRODUCTION
turn activates β-adrenoceptors - keeping the detrusor relaxed in the process (Elliott, 1907).
The controversial question at this point is whether, or not, the sympathetic nervous system is
an important mediator of resting tone in normal human urinary bladder, given that resection of
the sympathetic chain has been found to have little effect on bladder storage (Learmonth,
1931; Nordling et al., 1981). However, it has also been shown that the application of β-
adrenoceptor agonists reduces contractility in both normal (Igawa et al., 2001; Biers et al.,
2006; Yamanishi et al., 2006; Badawi et al., 2007; Otsuka et al., 2008; Kanie et al., 2012) and
neurogenic human bladders (Igawa et al., 2001). Hence, β-adrenoceptors seems important for
relaxation of the human urinary bladder, though they do not appear to be important during the
filling phase. In humans, β3-adrenoceptors are the main β-adrenoceptors in the bladder, both
at mRNA level (Nomiya and Yamaguchi, 2003) and at functional level (Igawa et al., 1998;
Igawa et al., 1999; Takeda et al., 1999; Kanie et al., 2012). Accordingly, β3-adrenoceptors
seem promising as a therapeutic target in the bladder.
In contrast, spontaneous contractile activity of the smooth muscle cells underlies human
bladder tone during filling (Turner and Brading, 1997). Electrophysiological recordings have
demonstrated that these contractions are action potential initiated by calcium via L-type
calcium channels (Hashitani and Brading, 2003). While interstitial cells form a dense network
in-between the muscle layers and lamina propria, cells in both locations exhibit ionic currents.
Since intramuscular interstitial cells have a calcium wave pattern different to that of the
smooth muscles, intramuscular interstitial cells are unlikely to act as pacemakers (Hashitani et
al., 2004; Johnston et al., 2008; Gray et al., 2013). Instead, it has been proposed that
intramuscular interstitial cells act as a brake to avoid a coordinated contraction of the bladder
wall during filling, and that lamina propria-located interstitial cells are important for the
positive modulation of smooth muscle cell contraction (Andersson and McCloskey, 2014).
When the bladder is full, distension of the bladder produces signals that are conveyed to
higher brain levels via the afferent Aδ-fibres (Fowler et al., 2008). The resulting activation of
the parasympathetic nervous system leads to the activation of muscarinic receptors through
the release of ACh (Fig. 2). M3-receptors in the detrusor mediate contraction by activating
phospholipase C, which in turn cleaves phosphatidyl inositol 4,5-bisphosphate (PIP2) into
inositol triphosphate (IP3) and diacylglycerol. IP3 activates the IP3 receptors of the
"9
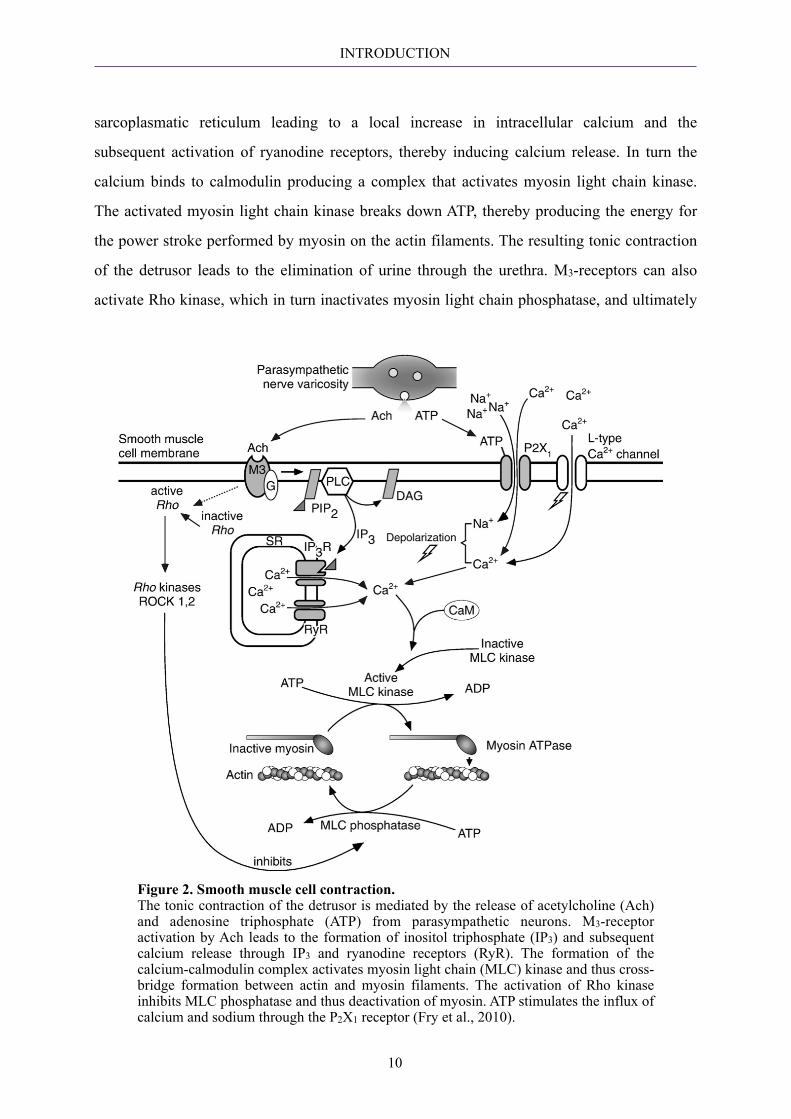
INTRODUCTION
sarcoplasmatic reticulum leading to a local increase in intracellular calcium and the
subsequent activation of ryanodine receptors, thereby inducing calcium release. In turn the
calcium binds to calmodulin producing a complex that activates myosin light chain kinase.
The activated myosin light chain kinase breaks down ATP, thereby producing the energy for
the power stroke performed by myosin on the actin filaments. The resulting tonic contraction
of the detrusor leads to the elimination of urine through the urethra. M3-receptors can also
activate Rho kinase, which in turn inactivates myosin light chain phosphatase, and ultimately
"10
Figure 2. Smooth muscle cell contraction. The tonic contraction of the detrusor is mediated by the release of acetylcholine (Ach) and adenosine triphosphate (ATP) from parasympathetic neurons. M3-receptor activation by Ach leads to the formation of inositol triphosphate (IP3) and subsequent calcium release through IP3 and ryanodine receptors (RyR). The formation of the calcium-calmodulin complex activates myosin light chain (MLC) kinase and thus cross-bridge formation between actin and myosin filaments. The activation of Rho kinase inhibits MLC phosphatase and thus deactivation of myosin. ATP stimulates the influx of calcium and sodium through the P2X1 receptor (Fry et al., 2010).

INTRODUCTION
leads to a reduced dephosphorylation of myosin light chain and hence to the sustained
contraction of the musculature.
In bladder pathologies, the release of ATP from parasympathetic efferents stimulates P2X1
receptors with a resultant influx of calcium and sodium. However, in normal bladders ACh
seems to be the sole contributor to smooth muscle cell contraction (Sibley, 1984). (Andersson
and Arner, 2004)
Overactive bladder syndrome
Since the bladder is highly innervated and the interplay between storage and elimination of
urine is a complex matter, a number of symptoms can arise from the lower urinary tract
system. One of the symptom complexes linked to the storage of urine is known as OAB. In
OAB, the symptoms are due to detrusor overactivity. The symptoms have been defined as
urgency, frequency, nocturia and incontinence by the International Continence Society
(Abrams et al., 2003). This means that patients experience an overwhelming sensation of
needing to visit the restroom. Furthermore, they have a high voiding frequency, both night and
day, defined as more than eight times during the day and two or more times per night. The
symptoms have a tremendous impact on the patients’ quality of life (Stewart et al., 2003). It is
also worth noting that OAB is highly prevalent within the general population. From
population-based surveys it has been estimated that more than 10% of the adult population
suffers from OAB and that its prevalence increases with age (Irwin et al., 2006; Coyne et al.,
2009; Coyne et al., 2013). A number of individuals develop OAB without any clear
underlying cause, and accordingly the symptom complex can be idiopathic (Hanna-Mitchell
et al., 2014). However, the development of OAB can also be brought about by a spinal cord
injury; neurodegeneration as in patients with Alzheimer's disease; diabetes; menopause; or
benign prostatic hyperplasia, which can lead to BOO. In fact, 60% BOO patients suffer from
detrusor overactivity (Abrams et al., 1979; Ameda et al., 1994). It is unknown why these
individuals develop OAB, but a decrease in large-conductance calcium-activated potassium
channel (BKCa) transcript levels has been observed in both humans with BOO (Chang et al.,
2010) and animal models of BOO (Aydin et al., 2012).
While the symptoms of OAB can be alleviated in some cases by exercising the pelvic floor
muscles, pharmacological treatment is often necessary. The first-in-line treatment consists of
"11

INTRODUCTION
muscarinic receptor antagonists that block the input from the parasympathetic nervous system
to the bladder. Unfortunately, in this case patients suffer from side effects such as dry mouth
and dizziness. Accordingly, more than 50% discontinue the treatment within the first six
months (Sexton et al., 2011). Therefore, great effort has been put into finding relevant new
pharmacological targets for the treatment of OAB. Recently, the β3-adrenoceptor agonist
mirabegron was approved for the treatment of OAB, with clinical studies implying that the
use of mirabegron decreases the OAB symptoms with a low degree of side effects (Chapple et
al., 2013; Herschorn et al., 2013; Khullar et al., 2013; Nitti et al., 2013; Yamaguchi et al.,
2014). A number of other possible pharmaceutical agents have also been studied: e.g.
purinoceptor antagonists; endothelin-receptor antagonists; phosphatase diesterase inhibitors;
and Rho-kinase inhibitors (Andersson, 2007). However, none of them have been approved for
the treatment of OAB at this point of time. Thus, the need for novel pharmaceutical drugs for
the treatment of OAB still exists.
Potassium channels as a putative target
With regards to the human bladder, the following potassium channels have been found and
studied in more or less detail: small-conductance calcium-activated (SKCa) (Darblade et al.,
2006; Nielsen et al., 2011; Afeli et al., 2012b; Soder et al., 2013; La Fuente et al., 2014); and
BKCa (Darblade et al., 2006; Hristov et al., 2011; Hristov et al., 2012b; Afeli et al., 2013b;
Hristov et al., 2013; La Fuente et al., 2014); ATP-sensitive (KATP)(de Moura et al., 1993;
Trivedi et al., 1995a; Trivedi et al., 1995b; Buckner et al., 2000; Aishima et al., 2006;
Darblade et al., 2006; Badawi et al., 2008; Kajioka et al., 2011; Ekman et al., 2012); stretch-
activated (TREK-1)(Lei et al., 2014); and voltage-gated potassium channels (Kv1.3, Kv1.6,
Kv2 and Kv9.3) (Davies et al., 2002; Hristov et al., 2012a). In fact, the KATP-sensitive
potassium channel opener ZD0947 has been clinically tested as a potential OAB treatment,
unfortunately, without showing any significant effect (Chapple et al., 2006). In contrast,
clinical trials on retigabine, a Kv7 channel activator used in the treatment of epilepsy, have
shown that the activation of Kv7 channels leads to urinary retention (Brickel et al., 2012).
Furthermore, in vivo and in vitro studies in rats (Streng et al., 2004; Rode et al., 2010; Svalø et
al., 2011) and guinea pigs (Anderson et al., 2009; Afeli et al., 2013a; Anderson et al., 2013)
have shown that Kv7 channels are functionally important in the bladder. As to the side effects
"12

INTRODUCTION
of positive Kv7 channel modulators, clinical studies have demonstrated that flupirtine and
retigabine do not change the blood pressure or heart rate (Herrmann et al., 1987; French et al.,
2011), which are the main concern in relation to treatments based on Kv7 channel modulators.
These studies led us to the conviction that Kv7 channels might be important regulators of
urinary bladder function. Consequently, the expression and function of these channels in
human and pig urinary bladder are the main focus of this thesis. In saying this, previous
findings in rat and guinea pig bladder will also be described and a detailed description of the
structure, function and tissue distribution of Kv7 channels will be presented below.
Kv7 Channel Physiology
Structure
There are five different subtypes of Kv7 channels, Kv7.1-7.5, all of which are encoded by the
KCNQ1-5 genes. The functional channels are formed either by hetero- or by homotetramers,
each of which are composed by four subunits. In turn, each subunit has six transmembrane
segments (S1-S6), with the result that both the N- and C-terminals are located intracellularly.
The trafficking, conductance, gating, ion selectivity, regulation and pharmacology of the
channels can be modulated by their non-pore-forming β-subunits KCNE1-5, which are
encoded by the KCNE1-5 genes (Abbott, 2014). The importance of the channels in the human
body has become clear as mutations of the KCNQ1-4 genes have been linked to a number of
human diseases. Currently known: long QT syndrome causing cardiac arrhythmia (Q Wang et
al., 1996); benign familial neonatal convulsions (Biervert, 1998); and nonsyndromic
autosomal dominant deafness (Kubisch, 1999). Furthermore, it seems that single-nucleotide
polymorphisms in KCNQ1 are associated with an increased susceptibility to develop type 2
diabetes (Unoki et al., 2008).
The Kv7 channels are gated by voltage and act as important regulators of membrane
excitability. Kv7.1 channels are found in the heart where they are responsible for the
repolarising IKs current of the cardiac action potential (Barhanin et al., 1996). In neurons, a
muscarine-sensitive potassium current, known as the M-current (Brown and Adams, 1980), is
mediated by Kv7.2, Kv7.3 and Kv7.5 channels (H Wang, 1998; Selyanko et al., 1999; Lerche
et al., 2000). In contrast, Kv7.4 channels were first discovered in sensory outer hair cells of the
"13

INTRODUCTION
inner ear where they influence electrical excitability (Kubisch, 1999). In the stomach and
intestines, Kv7 channels are involved in the secretion of gastric acid (Grahammer et al., 2001)
and chloride (Warth et al., 2002), respectively. Due to the electrochemical gradient, opening
the various Kv7 channels leads to potassium efflux, and hence to hyperpolarisation. The
channels activate at sub-threshold potentials with half-maximum activation constants ranging
from -40 to -20 mV depending on the Kv7 channel subtype. The kinetics of the various
subtypes differ from one another. In general, however, Kv7 channels are responsible for
repolarising the action potential. This is accomplished by their slow activation during
depolarisation and slow deactivation during hyperpolarisation, along with incomplete
inactivation (Soldovieri et al., 2011). The specific regulation of the respective channels is
further modulated by regulatory subunits as described below.
Regulation
Kv7 channels are regulated in a number of ways as illustrated in Figure 3. To produce
functional Kv7.2/7.3 channels requires the binding of calmodulin to the C-terminus (Wen and
Levitan, 2002; Yus-Najera et al., 2002). In addition, calmodulin modulates the membrane
trafficking of Kv7.2/Kv7.3 channels (Etxeberria et al., 2008; W Liu and Devaux, 2014).
Interestingly, calmodulin binds to Kv7.2/7.3, Kv7.4 and Kv7.5 channels, where it acts as a
calcium-sensor leading to reduced currents through the channels (Gamper and Shapiro, 2003;
Gamper et al., 2005).
The activation of Kv7 channels can be modulated by PIP2. The Kv7.2/Kv7.3 channels underlie
the M-current since the channels are inhibited by the activation of the muscarinic receptors
(Brown and Adams, 1980; H Wang, 1998; Selyanko et al., 1999). It is now known that this
inhibition is due to the depletion of PIP2 from the Kv7 channels as the compound is cleaved
into IP3 and diacylglycerol by phospholipase C subsequent to muscarinic receptor activation
(Li et al., 2005; Suh et al., 2006; Zhang et al., 2010; Linley et al., 2012). Interestingly,
homomeric Kv7.2 and heteromeric Kv7.2/Kv7.3 channels are not the only ones that are
regulated by PIP2, this is also the case for Kv7.4 channels (Leitner et al., 2011) and Kv7.1/
KCNE1 channels (Loussouarn et al., 2003), which suggests that this may be a general feature
of Kv7 channels.
"14

INTRODUCTION
More recently, another regulator of Kv7 channels has been described in the literature, namely
hydrogen sulphide (H2S), which is an endogenously produced gaseous substance. Hydrogen
sulphide has been found to reduce contractility via Kv7 channels in rat (Schleifenbaum et al.,
2010; Martelli et al., 2013) and porcine (Hedegaard et al., 2014) blood vessels. In the latter
publication, Kv7 channel inhibition diminished H2S- and hypoxia-induced relaxation
(Hedegaard et al., 2014). Similarly, the relaxant effect of H2S in the bladder has been
investigated in trout (Dombkowski et al., 2006), rat (Patacchini et al., 2004; Gai et al., 2013)
and human (Gai et al., 2013). While the response to H2S was insensitive to a number of K+-
channel blockers, including 4-aminopyridine (4-AP) and glibenclamide (Dombkowski et al.,
2006) in trout bladders, in human and rat bladders the effect of H2S could be diminished by
the KATP blocker glibenclamide (Gai et al., 2013). However, neither Dombkowski et al. (2006)
nor Gai et al. (2013) tested Kv7 channel inhibitors. As Kv7.1, Kv7.2, Kv7.3, Kv7.2/7.3 and
"15
Figure 3. Schematic representation of Kv7 channels and their interaction sites. The interaction sites with calmodulin (CaM), phosphatidylinositol 4,5-bisphosphate (PIP2), A-kinase anchoring protein and the auxiliary β subunit KCNE1 on a general model of Kv7 channels (Barros et al., 2012)

INTRODUCTION
Kv7.4 channels are 4-AP insensitive (Kubisch, 1999; Robbins, 2001), further studies are
needed if we are to clarify if the response to H2S is dependent upon the presence of Kv7
channels in the bladder.
In smooth muscle cells, the activation of Kv7 channels mediates relaxation, which is also the
case for β-adrenoceptor agonists. In addition, BKCa channels have been shown to be important
for β-adrenoceptor mediated relaxation in the bladder of rats and guinea pigs (Kobayashi et
al., 2000; Frazier et al., 2005; Uchida et al., 2005). Furthermore, that Kv7 channels interplay
with β-adrenoceptors in rat renal arteries has been demonstrated recently (Chadha et al.,
2012). Chadha et al. (2012) found that Kv7.4 was the main contributor to Kv7 channel
mediated relaxation, and that the blockade of Kv7 channels decreased the response to the β-
adrenoceptor agonist isoprenaline. Following on from this it has been shown, that β-
adrenoceptors interact with Kv7 channels, and that this seemingly occurs via βγ subunit
translocation and protein kinase A activation (Stott et al., 2014). However, whether β-
adrenoceptors and Kv7 channels interplay in the urinary bladder still remains to be explored.
Pharmacology
Besides the endogenous regulators of Kv7 channels a number of compounds have been
developed with varying selectivity for the different channel subtypes. Retigabine and
flupirtine are some of the best characterised Kv7 channel activators, with the compounds
activating subtypes 2 through 5 (Main et al., 2000; Wickenden et al., 2000; Schrøder et al.,
2001; Tatulian et al., 2001; Dupuis et al., 2002). Hence, not only does retigabine activate Kv7
channels, it also inhibits native Kv7 currents at potentials positive to ∼ 0 mV in murine hepatic
portal vein myocytes leading to a bell-shaped activation curve (Yeung et al., 2008). Yeung et
al. (2008) has suggested that this bimodal effect might be explained by the expression of
heteromeric, and thus not only homomeric, Kv7.1, Kv7.4 and Kv7.5 channels in murine
myocytes (Yeung et al., 2008). The compounds are not merely tool compounds, since
retigabine and flupirtine have already been approved for the treatment of epilepsy in 2010 and
for pain relief in 1984, respectively. Retigabine is generally accepted as a Kv7 selective
compound, being 100-fold more potent at activating Kv7 channels compared to potentiating
gamma aminobutyric acid type A (GABAA) receptors (Rundfeldt, 1999; Rundfeldt and
Netzer, 2000). However, its central effects on the urinary bladder (Streng et al., 2004) may in
"16

INTRODUCTION
part be mediated by the tonic inhibition of micturition via GABAA receptors. Furthermore,
retigabine is known to block L-type calcium channels (Mani et al., 2013), thereby decreasing
smooth muscle cell relaxation. Due to the above mentioned limitations of retigabine, care
must be taken when interpreting its effects. Moving on, the acrylamide, S-1, has been found to
inhibit Kv7.1 channels and activate Kv7.2-7.5 channels, by binding to the same tryptophan
residue as retigabine (Bentzen et al., 2006). Since interest in Kv7 channels has been on the
increase in recent years, the number of subtype selective compounds has also increased. The
Vanderbilt Center for Accelerated Probe Development has developed two positive modulators,
i.e. ML213 and ML277, that are Kv7.2/Kv7.4 and Kv7.1 channel selective, respectively (Yu et
al., 2011; Mattmann et al., 2012). That the development of subtype selective compounds is
difficult is a well-known fact. Unfortunately, ML213 has recently been described as an
activator of both Kv7.4/7.5 heteromers, and Kv7.5 homomers (Brueggemann et al., 2014),
which means that it cannot facilitate the functional discrimination between these subtypes.
Another compound that is selective for Kv7.4 channels is SMB-1. This compound is also an
inhibitor of Kv7.2 channels (Blom et al., 2014). With regards to the activation of the Kv7
channel subtypes e.g. in neurons, ICA-27243 activates Kv7.2/7.3 channels with a higher
potency than Kv7.4 channels and Kv7.3/7.5 channels (Wickenden et al., 2008; Blom et al.,
2010), while meclofenamic acid is an activator of Kv7.2/3 channels (Peretz et al., 2005). To
study the Kv7 channel subtypes located e.g. in the heart, L-364373 is appropriate as it
activates Kv7.1 channels (Salata et al., 1998). When evaluating Kv7.1-7.5 channel blockers, it
is clear that XE991 and linopirdine (DuP 996) (Robbins, 2001) are the most frequently used
tool compounds. Unfortunately, in addition to inhibiting Kv7 channels, XE991 has been found
to inhibit recombinant Kv1.2/Kv1.5 and Kv2.1/Kv9.3 channels (Zhong et al., 2010). Similarly,
while chromanol 293B provides the opportunity to block Kv7.1 channels selectively, it has
also been proven a potent inhibitor of Kv1.5 channels (Yang et al., 2000). With regards to
these unspecific effects, especially the inhibition of Kv2.1/Kv9.3 channels is concerning, since
these channels are functionally important in human detrusor (Hristov et al., 2012a). The use of
linopirdine instead of XE991 has some advantages as this compound does not inhibit Kv2.1/
Kv9.3 channels (Zhong et al., 2010). The effect of chromanol 293B depends upon the β-
subunits bound to Kv7.1 as it has been shown that KCNE3 increases the potency of Kv7.1
"17

INTRODUCTION
channels 30-fold compared to that of KCNE1 (Bett et al., 2006). Please consult Appendix IV
for a more detailed overview of the potency, selectivity and IUPAC names of the commonly
used Kv7 channel modulators.
Tissue distribution
In recent years, the expression and functionality of Kv7 channels in a number of smooth
muscle tissues: vascular (Ohya et al., 2003; Yeung and Greenwood, 2005; Joshi et al., 2006;
Yeung et al., 2007; Mackie et al., 2008; Yeung et al., 2008; Joshi et al., 2009; Zhong et al.,
2010; Jepps et al., 2011; Mani et al., 2011; Ng et al., 2011; Chadha et al., 2012; Chadha et al.,
2014; Hedegaard et al., 2014), colon (Jepps et al., 2009), stomach (Grahammer et al., 2001;
Ohya et al., 2002; Ipavec et al., 2011), airway (Brueggemann et al., 2012) and myometrium
(McCallum et al., 2009; McCallum et al., 2011) have been examined, and confirmed. The
specific findings have been carefully reviewed by Jepps el al. (2013) and will therefore not be
described in further detail in the present thesis.
As previously mentioned, there is growing evidence for the importance of Kv7 channels, not
only in smooth muscle cells of the cardiovascular system, but also in the urinary bladder.
More specifically, functional evidence for Kv7 channels has previously been achieved using
retigabine, flupirtine and XE991 in the bladder of rats (Rode et al., 2010; Svalø et al., 2011;
Baysting and McCloskey, 2014). Moreover, studies have shown that mRNA could be detected
for KCNQ5, and especially KCNQ4, whereas KCNQ1-3 were expressed at very low levels
(Svalø et al., 2011) or completely undetectable (Ohya et al., 2002) in rat bladder. Accordingly,
it must be assumed that homomeric Kv7.4 and/or Kv7.5 channels and/or heteromeric Kv7.4/7.5
channels constitute important regulators of urinary bladder function in rats. With regards to
the β-subunits of Kv7 channels, KCNE1 and KCNE2 were detected by end-point RT-PCR in
rat bladder (Ohya et al., 2002). However, to the best of our knowledge, this is the only
description of the KCNE subtypes in bladder tissue. Accordingly, it needs to be elaborated
upon by examining all five subtypes.
Interestingly, a broader analysis of the Kv7 channel subtype distribution has been performed
in guinea pigs. This is the case at both protein and functional level (Afeli et al., 2013a;
Anderson et al., 2013), Anderson et al. (2013) showed the expression of all subtypes of the
Kv7 channels. Interestingly, a number of ionic currents, including Kv7 currents, are also
"18

INTRODUCTION
exhibited in intramuscular interstitial cells (Anderson et al., 2009). However, as the regulatory
role of interstitial cells has not yet been fully elucidated, how they affect detrusor function
remains to be determined.
Spontaneously hypertensive rats are known to exhibit detrusor overactivity and increased
voiding frequency (Clemow et al., 1998; Spitsbergen et al., 1998). Interestingly, KCNQ4 was
3.7-fold down-regulated which has been demonstrated and confirmed by a 50% reduction in
band intensity for Kv7.4 protein in arteries from spontaneously hypertensive rats (Jepps et al.,
2011). Furthermore, a decreased contractility in response to linopirdine and S-1 has been
demonstrated in renal arteries (Chadha et al., 2012), not to mention to XE991 (inhibitor of
Kv7.1-7.5) in the urinary bladder (Baysting and McCloskey, 2014), when compared to
normotensive rats. These findings support the notion that Kv7.4 channels play a functional
role in the urinary bladder of rats. How this relates to human bladder disease is unknown.
However, gene transcript levels seem to be important in choosing the right treatment.
In addition to the in vitro studies, Kv7 channel function has been examined in vivo in rats
using cystometry in saline infused bladders (Streng et al., 2004), acetic acid-induced
overactivity (Svalø et al., 2011) and capsaicin-induced overactivity (Streng et al., 2004; Svalø
et al., 2011). In this case it was observed that the activation of Kv7 channels led to an
increased voiding interval. Interestingly, the interval was only partially increased when the
animals were orally administered with retigabine and the bladder irritation induced by
capsaicin (Svalø et al., 2011). Accordingly, the observed in vitro effects have been supported
by the in vivo data. Combined with the clinical finding showing that retigabine leads to
urinary retention (Brickel et al., 2012), we speculate that positive modulators of Kv7 channels
could prove useful for the treatment of OAB by decreasing the number of voidings per day.
However, a deeper understanding of Kv7 channel function in human bladders is needed before
this can be concluded.
!
"19

AIM
Aim
We hypothesise that Kv7 channels are important regulators of urinary bladder function and
could be potential targets for the treatment of OAB. The overall aim of this thesis is to
characterise the expression, regulation and function of Kv7 channels in isolated urinary
bladders of humans and pigs. Since the β3-adrenoceptor agonist mirabegron recently has been
approved for the treatment of OAB, we aim at investigating whether, or not, Kv7 channel
activation interplays with that of β3-adrenoceptors or if it provides a novel pathway for
detrusor relaxation.
!Specifically, the aims are:
!• To determine KCNQ and KCNE expression in the urinary bladder of humans, and to
compare it to the transcript level in pigs. This is done on biopsies from patients with
normal bladder function and patients with BOO.
!• To study the functionality of Kv7 channel subtypes in the urinary bladder of pigs and
humans.
!• To investigate the mechanisms underlying Kv7 channel mediated relaxation in the non-
pathological urinary bladder of pigs and humans. Among others, to explore if there is an
interaction between Kv7 channels and β-adrenoceptors in the human urinary bladder.
!!
"20

EXPERIMENTAL METHODS
Experimental Methods
This thesis is based on the original research presented in Paper I-III. In this section, the
applied methods will be described in short with emphasis on the methodological aspects. All
protocols are described in detail in the Materials and Methods section of the respective papers
and can be found in Appendix I-III.
Animals and Patients
Paper II is based on pig bladder tissue. The purpose of using pig detrusor was both to
investigate whether pigs are an appropriate model for Kv7 channel function in the human
bladder in addition to economise the human bladder tissue. The urodynamic and functional
characteristics of the pig urinary bladder is similar and hence comparable to that of humans
(Crowe and Burnstock, 1989). Therefore the use of pig bladders, rather than those of small
laboratory animals, is more appropriate when studying clinically relevant effects.
Paper I and III are based on experiments using human bladder specimens. Whole bladder wall
samples and deep muscle biopsies were collected from the bladder dome at cystectomy or
cystoscopy, respectively. Patients, with a normal bladder function, having their bladder
completely removed (cystectomy) due to malignancy, were included in the ‘normal’ group. In
contrast, biopsies for the ‘BOO’ and ‘BOO+DO’ group were obtained at cystoscopy during
transurethral resection of the prostate due to benign prostatic hyperplasia from patients with or
without detrusor overactivity (DO).
The study was approved by the Copenhagen County Ethics Committee (SJ-219) and detrusor
tissue was obtained following informed written consent. The number of patients to be
included for each myograph study was determined by a power analysis showing that n=4 per
group would be necessary to detect differences of at least 5% between control and treatment
group.
Gene expression
In Paper II and III we performed reverse transcription quantitative real-time polymerase chain
reaction (RT-qPCR) to analyse the mRNA levels of KCNQ1-5 and KCNE1-5 which are the
genes encoding the Kv7 channels and their β-subunits KCNE1-5. Due to a very limited
"21

EXPERIMENTAL METHODS
amount of sample material we did not aim to analyse the level of β1-3-adrenoceptor transcript,
though this would have added valuable information to Paper I.
RNA isolation
To ensure high quality RNA the bladder specimens were immediately fixed after removal
from the body. With regard to human bladder specimens the biopsies were obtained at
cystoscopy and fixed in RNAlater as our collaborator at Skejby Hospital, surgeon Jørgen
Bjerggaard Jensen has formerly found that the RNA starts to degrade as soon as the blood
supply is cut off. Therefore, it is not suitable to use mRNA obtained at cystectomy since blood
perfusion is stopped for 15-45 minutes before removal of the bladder specimens (Jensen JB,
unpublished). RNAlater was chosen for fixation purposes of human biopsies as only nontoxic
chemicals were allowed in the operating theatre and as this compound is suitable for long-
term storage of tissue at -20 ○C, which was the freezer type available at all of the involved
hospitals. The tissue was collected from 2012-2013. The pig specimens for RT-qPCR were
chosen to be collected at the Faculty of Science, instead of at the Danish Meat Trade College,
since they are experienced in collecting tissue immediately after blood flow has been
terminated.
The human biopsies were, as mentioned previously, deep muscle biopsies and contained both
urothelium and detrusor tissue, whereas the pig bladder specimens were whole bladder wall
samples and therefore contained epithelial cells from the peritoneum in addition to the other
cell types that are present in bladder biopsies. Furthermore, the human biopsies used for RT-
qPCR were collected at the Departments of Urology at Skejby and Næstved Hospital and
therefore there was a risk of the biopsies containing different ratios of urothelium and detrusor
tissue. To eliminate this difference we carefully analysed which genes were most appropriate
as reference genes using the geNorm analysis (qBasePlus, Biogazelle, Netherlands) (Fig. 4).
The tissue was ground in liquid nitrogen and further homogenised by a rotor-stator
homogeniser after which the RNA was extracted using the Trizol method. This method is
based on a ready-to-use monophasic solution of phenol and guanidine isothiocyanate leading
to the isolation of high quality RNA after cell lysis. Addition of bromochloropropane led to
phase separation and RNA could then be precipitated from the aqueous phase using isopropyl
alcohol.
"22

EXPERIMENTAL METHODS
In Paper II the quality of RNA was checked by gel electrophoresis using the ethidium
bromide staining technique whereas a Bioanalyzer 2100 was used for the sample acquisition
in Paper III. The reason for this was that the human bladder biopsies were only 50-150 mg in
size and therefore the amount of total RNA was limited. The Bioanalyzer 2100 is also a gel
electrophoresis based technique, however the gels are cast in microchips and therefore it is
only necessary to apply 50 ng of RNA whereas 500-1000 ng are needed for our standard gel
system. Furthermore, the Bioanalyzer 2100 provided a more detailed description of the RNA
quality depicted by the electropherogram; in addition it calculated the RNA quantity, thus
omitting the need for using the Nanodrop spectrophotometer for quantification of RNA in
Paper III.
RT-qPCR
DNase treatment was conducted to ensure that genomic DNA was not amplified during the
RT-qPCR analysis and the efficiency of the DNase treatment was controlled by the inclusion
of no-reverse-transcription controls. Another way to avoid the amplification of genomic DNA
would be to design primers that span the exon-exon boundary. However, this can be difficult
if multiple genes look alike and all splice variants are intended to be amplified by a single
primer set. Therefore we did not choose to do this. We determined quality and quantity of
RNA both before and after DNase treatment.
"23
Figure 4. Reference genes in pig and human detrusor. The expression of an array of genes were examined for their stability properties in human (n=8) (A) and pig (n=10) (B) detrusor. The following genes were evaluated as reference genes: 18S (18S rRNA gene), SF3A1 (splicing factor 3A1), B2M (beta-2-microglobulin), TOP1 (topoisomerase DNA I), ACTB (beta actin), GAPDH (glyceraldehyde-3-phosphate dehydrogenase), GPXI (glutathione peroxidase 1), SDHA (succinate dehydrogenase complex), ATP5G (ATP synthase), GSR (glutathione reductase), SHAS2 (hyaluronan synthase 2), GPI (glucose phosphate isomerase), UBC (poly-ubiquitin) and eEF-1 (eukariotic elongation factor 1 gamma-like protein).
A B

EXPERIMENTAL METHODS
RNA was reversely transcribed into complementary deoxyribonucleic acid (cDNA) using
oligo(dt) primers and reverse transcriptase. Oligo(dt) primers favours the amplification of
mRNA as they bind to the poly-A tail, compared to random primers which is a more
appropriate choice when analysing degraded samples as it provides a higher cDNA yield. The
following qPCR was performed using SYBR green which fluoresces when bound to double-
stranded DNA. The relative quantity of DNA was calculated from the amplification curve as
the cycle at which the amplification reached a software determined threshold. Accordingly,
this method is not a copy number analysis and therefore only allows for the determination of
relative expressional changes within a given gene.
All primers for the genes of interest were designed using the free software Primer3 (NCBI)
and sequences can be found in Table 1 of the relevant papers. To ensure primer specificity, the
melting curve was evaluated for primer dimers and sequence analysis of amplicons was
performed for all the studied genes.
Normalisation using the qBasePlus software was performed by calculating the geometric
mean of the reference genes. The number of reference genes was also determined by the
software and depends on the homogeneity of the samples. In our case two reference genes
were necessary for normalisation of both pig and human bladder samples.
Myography
In the present thesis, organ bath studies were included in Paper I-III to evaluate tissue
responses to Kv7 channel modulators (Paper II and III) and β-adrenoceptor agonists (Paper I
and III).
Tissue handling
Myography is an in vitro method at which small pieces of muscle, are mounted in a chamber
filled with physiological buffer at 37 ○C. The detrusor strips were cut 1 mm in width, 1 mm in
girth and 3 mm in length. In our experiments, the pH of the buffer was controlled by sodium
hydrogen carbonate and therefore the equilibrium could be maintained at pH 7.4 by bobbling
the solution with a mixture of oxygen (95%) and carbon dioxide (5%). In contrast, when
transporting the bladder specimens from either the pig slaughterhouse or the operating theatre,
the tissue was kept in Tyrode’s salt solution added 20 mM HEPES (4-(2-hydroxyethyl)-1-
"24

EXPERIMENTAL METHODS
piperazineethanesulfonic acid) to maintain the pH at 7.4. Thereby we could avoid bobbling
the solution during transportation (Paper II and III). Our laboratory has formerly transported
the tissue in MEM-d-val medium as the biopsies were also used for culturing of smooth
muscle cells. This medium was used in the study presented in Paper I. However, during our
studies we found that buffer containing HEPES was more efficient in keeping the pH at 7.4
and accordingly, exchanging the transport solution increased the success rate of obtaining
viable specimens for the myograph experiments.
The tissue was mainly used on the day of collection of the material but also after 20 hours
cold storage in Tyrode’s salt solution with 20 mM HEPES. It has been shown that the
responses to contractile agents decreases after 24 hours cold storage in Kreb’s solution with/
without oxygenation (Riehmann et al., 1997; G Liu and Daneshgari, 2006). However, when
evaluating the tissue responses of human detrusor to different contractile agents we did not
observe any differences between day one and day two (baseline tension: P=0.51, n=8, N=107;
carbachol: P=0.36, n=8, N=51; 40 mM high potassium physiological saline solution (KPSS):
P=0.73, n=8, N=50; 20 mM KPSS: P=0.29, n=2, N=8).
The tissue was mounted directly onto two stainless steel pins of a DMT myograph (700MO,
Danish Myo Technology, Aarhus, Denmark). One of the pins was connected to a force
transducer which reported the force in volts. Connecting the myograph to a powerlab allowed
the data to be transformed into millinewton (mN) which were collected by LabChart 7
software (AD instruments, DMT, Aarhus, Denmark) for data analysis.
Length-tension relation
Pig detrusor strips were used for length-tension investigations. The tissue was stretched to a
baseline tension of 0 to 10 mN in PSS and the length of the strip was measured. The passive
tension was measured in Ca2+-free PSS whereas the active tension was measured in 125 mM
KPSS by subtracting the passive tension from the total tension. The active tension curve was
best fitted to a Gaussian distribution (R2=0.30). The maximal obtained active tension was at
2.9 mm increase in length corresponding to Lo (optimal length). Accordingly, stretching the
tissue by 2.5 mm provided 85% of the maximal obtained active tension to 125 mM KPSS
(Fig. 5). This increase in length was used for all myograph experiments presented in Paper II
and III. It would have been optimal to perform length-tension analysis on detrusor tissue from
"25
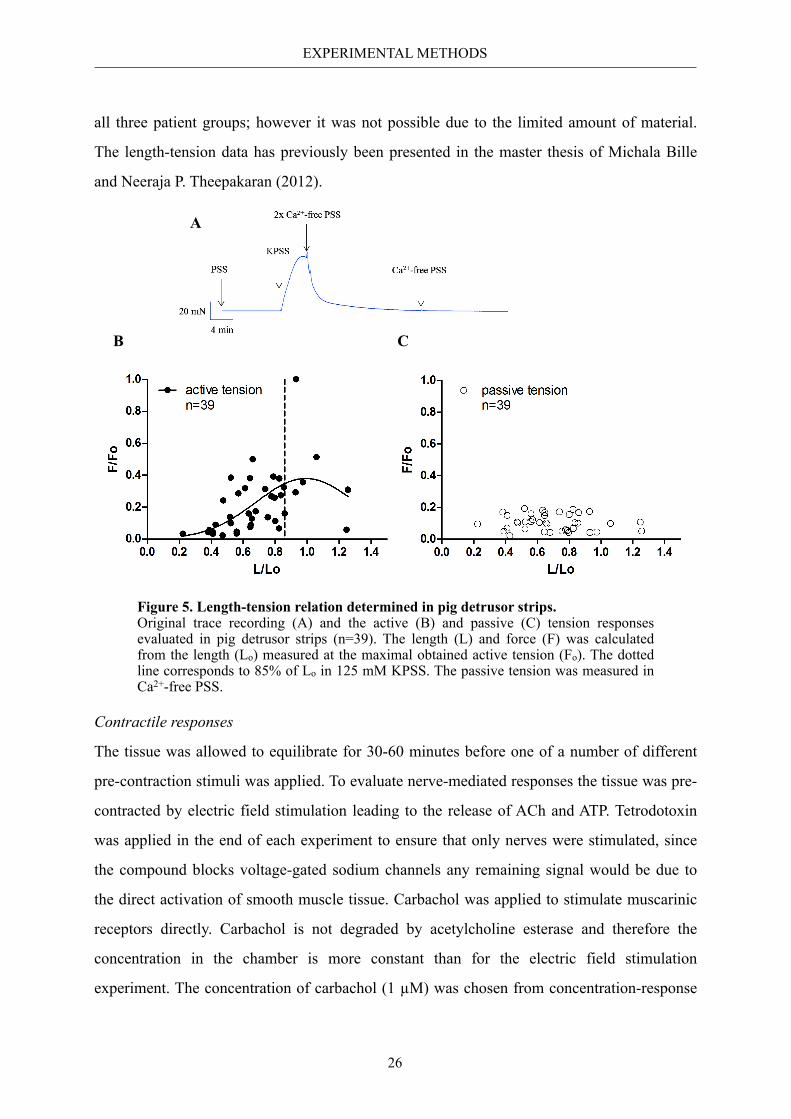
EXPERIMENTAL METHODS
all three patient groups; however it was not possible due to the limited amount of material.
The length-tension data has previously been presented in the master thesis of Michala Bille
and Neeraja P. Theepakaran (2012).
Contractile responses
The tissue was allowed to equilibrate for 30-60 minutes before one of a number of different
pre-contraction stimuli was applied. To evaluate nerve-mediated responses the tissue was pre-
contracted by electric field stimulation leading to the release of ACh and ATP. Tetrodotoxin
was applied in the end of each experiment to ensure that only nerves were stimulated, since
the compound blocks voltage-gated sodium channels any remaining signal would be due to
the direct activation of smooth muscle tissue. Carbachol was applied to stimulate muscarinic
receptors directly. Carbachol is not degraded by acetylcholine esterase and therefore the
concentration in the chamber is more constant than for the electric field stimulation
experiment. The concentration of carbachol (1 µM) was chosen from concentration-response
"26
Figure 5. Length-tension relation determined in pig detrusor strips. Original trace recording (A) and the active (B) and passive (C) tension responses evaluated in pig detrusor strips (n=39). The length (L) and force (F) was calculated from the length (Lo) measured at the maximal obtained active tension (Fo). The dotted line corresponds to 85% of Lo in 125 mM KPSS. The passive tension was measured in Ca2+-free PSS.
A
!!!B C

EXPERIMENTAL METHODS
curves as this caused approximately 60% of the maximal obtainable contractility with
carbachol in all three patient groups (Fig. 6).
To induce depolarisation of the tissue, different concentrations (20 or 40 mM) of potassium
were applied. Sodium was exchanged with potassium on an equimolar basis to maintain the
osmolarity of the KPSS. Thereafter the strips were treated with a combination of compounds
either as a single concentration or as cumulative concentrations. With regard to the tissue
stimulated by 20 mM KPSS, only a single concentration was applied since the phasic
contractions did not persist for more than one hour. It can be argued that it is not optimal to
use potassium for depolarisation purposes when evaluating a potassium channel. However, as
the tissue did not contract spontaneously, and as we also used carbachol and electric field
stimulation protocols, a broad description of the effects of the channels could be obtained. In
addition, we could ascertain that the effect was indeed mediated through potassium channels
as we have formerly observed that the potency of Kv7 channel modulators decreased by
increasing the concentration of potassium (Rode et al., 2010). For more details regarding the
used compounds please consult the Materials and Drugs section of the relevant papers.
Denervation protocol
In Paper II, we applied a denervation protocol in which KPSS (125 mM) was exchanged with
PSS in the myograph chamber repeatedly until the KPSS peaks reached steady-state. This has
been shown to deplete neuronal stores (Hubbard and Kwanbunbumpen, 1968; Fouda et al.,
"27
Figure 6. Contractile responses to carbachol in human detrusor strips. Original trace recording of the response to carbachol in normal detrusor strips (A). The contractile response to carbachol was evaluated in people with normal bladder function (n=7), bladder outflow obstruction with (n=2) or without (n=6) detrusor overactivity (B).
A B

EXPERIMENTAL METHODS
1991). When the contractions were stable, tetrodotoxin (0.1 µM) was applied to strips in PSS
to block voltage-gated sodium channels and thereby the propagation of nerve signals. Time-
matched vehicle controls were run in parallel for comparison. Strips were then pre-contracted
by carbachol (1 µM) after which the strips were treated with either retigabine (10 µM),
XE991 (10 µM) or vehicle control. The data were normalised to the pre-contraction level of
the same strip and compared to the time-matched vehicle control.
Statistical analysis
The statistics in this section mainly covers those used in the section Results & Discussion of
the present thesis. For details regarding the data presented in Paper I-III, please consult
Appendices I-III.
The day to day variations to different contractile agents and the effects of ML277 and
chromanol 293B were analysed using student’s t-test. In both cases we used unpaired t-tests
as the strips were divided into two groups and thus one strip did not receive both treatments
which would have led to the use of a paired t-test. The active-tension curve of pig detrusor
strip responses was prepared using a Gaussian distribution to determine the optimal length of
the strips. Concentration-response curves were prepared by non-linear regression curve fit to
the sigmoidal equation: Y=Bottom+(Top-Bottom)/1+10^(LogEC50-X)*HillSlope)); where Y
is the response to modulator/agonist that decreases as X increases, normalised to pre-
contraction agent; X is the log of modulator/agonist concentration; Top and Bottom are the
plateaus of the concentration-response curve; Emax is the maximum obtained effect and
logEC50 is the log molar concentration at which 50% of the maximum response is obtained.
Compound effects were compared to that of vehicle control by two-way repeated measures
ANOVA followed by Bonferroni post hoc test. Using repeated measures, takes into account
that the concentration-responses are performed as cumulative additions of compound and not
single concentrations to different strips. In all statistical tests, P<0.05 was considered
statistically significant. All standard errors are presented as standard error of the mean (SEM),
n equals the number of patients, whereas N equals the total number of strips. All statistics and
graphical presentations were prepared using GraphPad Prism 5.04 (GraphPad Software, CA,
USA).
"28

RESULTS AND DISCUSSION
Results and Discussion
KCNQ and KCNE expression within the urinary bladder
Expression levels in normal bladder
Kv7 channels and their β-subunits, KCNE1-5, are encoded by the KCNQ and KCNE gene,
respectively. Following from this we have studied their relative expression levels in pig and
human urinary bladders (Paper II and III). Our findings show that KCNQ1 and KCNQ3-5 are
expressed in human bladder (see Fig. 1 in Paper III) and that KCNQ3-5 are expressed in pig
bladder (see Fig. 1 in Paper II). Unfortunately, we were unable to determine the presence of
the remaining gene transcripts, KCNQ1-2, in pig bladder as the genes in question have not yet
been described in the literature.
KCNQ transcript levels have already been described in rodent bladders, i.e. rats and guinea
pigs. While we have previously identified all known subtypes in rats (Svalø et al., 2011),
KCNQ1-3 were very low (Svalø et al., 2011) and were not even detected by Ohya et al.
(2002). In addition, while we could confirm the presence of Kv7.4 protein by using western
blot analysis, we did not detect the presence of Kv7.2 (Svalø et al., 2011). Interestingly, all
subtypes have been detected in guinea pig bladder, both at mRNA and protein level (Afeli et
al., 2013a; Anderson et al., 2013). This was achieved by using end-point PCR on dispersed
detrusor cells and immunohistochemistry, respectively. Of interest is the fact that Afeli et al.
(2013a) did not detect Kv7.4 channels when staining slices of bladder tissue. Whether, or not,
this was due to the use of poor antibodies is unknown. Since Anderson et al. (2013) was
succesful in locating Kv7.4 channel protein in guinea pig bladder, we agree that this subtype is
also important in this species. Despite the fact that there were small species-specific
differences in KCNQ expression, KCNQ channels were expressed in all of the studied species
leading us to the conviction that it was important that further studies be carried out on Kv7
channels within the urinary bladder.
!The β-subunits, KCNE1-5, provide a means of fine-tuning the function of Kv7 channels. We
therefore studied all five subtypes of the ancillary β-subunit KCNE, which interestingly
enough we also found to be present in human urinary bladder biopsies (see Fig. 2 in Paper
"29

RESULTS AND DISCUSSION
III). The expression of KCNE1-2 has also been found in rat bladder (Ohya et al., 2002).
Similarly, all KCNE subtypes have been identified within both human heart (Bendahhou et al.,
2005) and myometrium (McCallum et al., 2011). Hence we can safely conclude that KCNE
transcripts are widely expressed.
Even though we did not compare the gene transcripts of different genes directly to one
another, we did observe that KCNQ4 and KCNE4 were the most abundantly expressed
subtypes, in terms of their Cq raw values. This led us to expect the presence of functional
Kv7.4 channels. In Xenopus oocytes KCNE4 has been shown to increase the voltage-
dependency of Kv7.4 channels (Strutz-Seebohm et al., 2006). While not confirmed in isolated
cells, the above findings point towards the existence of a Kv7.4-KCNE4 complex. It is worth
noting that we did not explore which of the β-subunits combined with the Kv7.4 channel in
the human detrusor. Also, to the best of our knowledge, whether, or not, a Kv7.4-KCNE4
complex exists remains to be described.
Expressional changes in outflow obstructed bladder
As mentioned in the introductory section, we wished to find out if Kv7 channels are expressed
in diseased bladders as well as in normal bladders. We therefore proceeded to determine the
expression of KCNQ and KCNE in bladder biopsies from patients with normal bladder
function and compared these to their expressed levels in bladder biopsies from patients with
an enlarged prostate (Paper III). We found that KCNQ1 was up-regulated 3.4-fold in patients
with BOO compared to those with normal bladder levels of KCNQ1 (P<0.05) (see Fig. 1
Paper III). We also observed a tendency towards a small change in KCNE expression between
the two groups. Namely, a 1.3-fold up-regulation of KCNE3 mRNA (P=0.38) and a 1.9-fold
down-regulation of KCNE1 (P=0.40) in patients with BOO compared to that observed in
patients with normal bladder function. If indeed there is an up-regulation of the KCNE3 β-
subunit; it is a highly interesting finding, given that the Kv7.1-KCNE3 complex has been
described to be constitutively active (Schroeder et al., 2000). We would like to avail of the
opportunity to speculate, that KCNQ1 is both up-regulated and turned into a constitutively
active channel as a compensatory mechanism in order to avoid detrusor overactivity in
patients with BOO. However, since a direct association between mRNA level and protein
"30

RESULTS AND DISCUSSION
expression does not necessarily exist, the results need to be confirmed at protein level and in
an increased sample set, and as such should only be considered as preliminary results.
!At present, KCNE research has mainly focused on the role of KCNE subtypes in relation to
Kv7.1 channel function as they are important in the heart due to the fact that a number of
mutations in KCNE leads to cardiac rhythm channelopathies (Splawski et al., 1997; Bianchi et
al., 1999; Isbrandt et al., 2002; Lundby et al., 2008; Ravn et al., 2008). However, KCNE
subtypes have also been confirmed in other tissues. KCNE4 and KCNQ1 are the most
abundantly expressed subtypes in murine and human myometrium (McCallum et al., 2009;
McCallum et al., 2011); furthermore, no apparent functional effect of Kv7.1 channels has been
observed since chromanol 293B failed to increase contractility in mice (McCallum et al.,
2009). They believed this was achieved by the inhibitory effect of KCNE4 on Kv7.1 channels
presumably by disruption of Kv7.1-calmodulin interactions (Grunnet et al., 2002; Grunnet et
al., 2005; Manderfield et al., 2009; Ciampa et al., 2011). Similarly, Kv7.1-KCNE4 complexes
might be formed in the bladder since we did not observe an effect of chromanol 293B on
bladder contractility (Fig. 7).
In addition to modulating Kv7.1 channels, KCNE3 has been shown to inhibit Kv7.5 channels
(Roura-Ferrer et al., 2009). Since we observed an increase in the level of KCNE3 mRNA and
found KCNQ5 transcript in patients with BOO, it would be interesting to evaluate selective
Kv7.5 channel modulators with a view to observe whether, or not, the effect of such
modulators is reduced in patients with BOO. This would, in turn, reveal whether, or not,
positive Kv7.5 channel modulators could prove useful for the treatment of bladder disorders in
these patients. In short, further studies are required if we are to understand the implications of
the different KCNE subtypes for Kv7 channel function in vivo.
Another potassium channel that is regulated during disease states is the BKCa channel. It has
been shown that the effect of BKCa channel inhibition is more pronounced in normal bladders
compared to that observed in bladders from patients with neurogenic bladders (Hristov et al.,
2013) and patients with benign prostatic hyperplasia (La Fuente et al., 2014). This could be
due to the 1.7-fold down-regulation of KCNMA1 (BKCa α-subunit) mRNA observed in a rat
model of BOO (Aydin et al., 2012), which was confirmed by a 15.8-fold down-regulation
"31

RESULTS AND DISCUSSION
observed in neurogenic human bladders (Hristov et al., 2013). It was thus possible to link
KCNMA1 expression with detrusor overactivity. Accordingly, BKCa channels and Kv7.1
channels are regulated oppositely to one another, in patients with BOO. However,
pharmacological studies in urinary bladder strips from male organ donors with BOO have
shown that BKCa channel activation can inhibit contractions induced by electric field
stimulation (La Fuente et al., 2014). Therefore, the expressional changes do not appear to
exclude BKCa channels as pharmacological targets for the treatment of overactive bladder.
Kv7 channel functionality in the detrusor.
Since both KCNQ and KCNE are expressed in the urinary bladder (Paper II and III) we went
on to investigate the functional implications of these channels with the help of myography.
Mediators of resting tone
In Paper II we studied the function of Kv7 channels in pig detrusor strips, demonstrating in the
process how, in contrast to XE991, chromanol 293B (Kv7.1 channel blocker), does not
increase the contractility of isolated detrusor strips at low depolarisation (20 mM KPSS)(Fig.
7 and see Fig. 2 in Paper II). However, this was not the case for muscarinic receptor mediated
pre-contraction induced either by electric field stimulation or by carbachol directly (see Fig.
3-5 in Paper II). Similarly, while XE991 (10 µM) had no effect on contractile function in
"32
Figure 7. Effect of negative Kv7 channel modulators on 20 mM KPSS stimulated pig detrusor strips. Original trace recordings of the responses to the negative modulators XE991 (10 µM) and chromanol 293B (10 µM) on pig detrusor strips.

RESULTS AND DISCUSSION
human detrusor strips that had been pre-contracted by carbachol or 40 mM KPSS, XE991
increased the contractility of 20 mM KPSS-stimulated detrusor strips (see Table 2 in Paper
III). Following on from this, it seems that one or more Kv7 channels are responsible for
maintaining the tone in pig and human bladder, though we now know that in pig detrusor this
is not Kv7.1 channels. Similar to pig detrusor, Kv7.1 channels have been shown not to regulate
the resting tone of human arteries (Ng et al., 2011). In contrast, in guinea pig detrusor strips
chromanol 293B increased the amplitude of spontaneous contractions indicating that Kv7.1
channels may be important for the resting tension in this species (Anderson et al., 2013).
Kv7 channel subtype differentiation
To elucidate which Kv7 channel subtypes are important in the urinary bladder we applied
positive Kv7 channel modulators to pig and human detrusor strips (Fig. 8, Paper II and III).
We subsequently discovered that retigabine decreased the mean tone of the detrusor strips.
This was the case in both pig (electric field stimulated and 1 µM carbachol-induced
contractions, see Fig. 3-5 Paper II), and human (1 µM carbachol-, 40 mM and 20 mM KPSS-
induced contractions, see Fig. 3 and Table 2 in Paper III) detrusor strips. Since retigabine is a
Kv7.2-7.5 channel activator the use of this compound allowed us to discriminate between
subtypes Kv7.1 and Kv7.2-7.5. With a view to explore the functionality of Kv7 channel
subtypes further we applied the Kv7.2/Kv7.4/Kv7.5 selective compound ML213, thereby
omitting the contribution of Kv7.3 channels.
We only detected KCNQ2 mRNA in one out of ten analysed biopsies (Paper III) and we
therefore believe that the effect of ML213 does not occur through this channel subtype, but
instead is due to the activation of homo- and/or heteromeric Kv7.4 and/or Kv7.5 channels.
However, we cannot rule out that Kv7.2 channels play a role in pig urinary bladder, as the
KCNQ2 gene has not been described in pigs and therefore the level of KCNQ2 transcript has
not yet been analysed. We found that ML213 was as efficacious as retigabine on 1 µM
carbachol-induced pre-contraction in both pig and human detrusor strips. Kv7.4/Kv7.5
channels are therefore likely to be implicated in the Kv7 channel mediated relaxation
following muscarinic receptor mediated pre-contraction (Paper II and III). In Paper II, we
confirmed the Kv7 channel selectivity of both retigabine and ML213 by attenuating the
maximum response upon the pre-incubation with XE991 (10 µM). At present, as it has been
"33

RESULTS AND DISCUSSION
determined that the potency of ML213 does not differ between homomeric Kv7.4 and Kv7.5
channels and heteromeric Kv7.4/Kv7.5 channels (Brueggemann et al., 2014), we cannot
discriminate further between the functionality of the different subtypes within the urinary
bladder.
In Paper III, we describe how we used the Kv7.1 selective activator ML277 on human and pig
detrusor strips (see Table 2 and Fig. S1 in Paper III) since the use of retigabine did not provide
us with information regarding the Kv7.1 channels. In human detrusor, ML277 decreased the
mean tone of carbachol and 40 mM KPSS pre-contracted strips while strips pre-contracted by
20 mM KPSS did not respond to ML277 at all (Table 2 in Paper III). We can therefore
conclude that Kv7.1 channels in the human urinary bladder can be pharmacologically
"34
A B
!!!!!!!C
!
Figure 8. Retigabine and ML213 decrease the pre-contraction tone of human and pig detrusor strips. Cumulative concentration-response curves for retigabine and ML213 on 1 µM carbachol pre-contracted bladder was studied on human (A) and pig (B) detrusor strips. Original trace recording of the effect of retigabine and ML213 in human detrusor strips (C). Retigabine decreased mean tone with a lower potency compared to ML213. *P<0.05, ***P<0.001, compared to vehicle control.

RESULTS AND DISCUSSION
modulated. In contrast, ML277 did not affect any of the pre-contraction agents used in pigs
(see Fig. S1 in Paper III) and we can therefore conclude that Kv7.1 channels are not important
in the case of pig urinary bladders. This is in keeping with our previous findings, i.e. that
chromanol 293B does not increase contractility in pig detrusor (Fig. 7 and Fig. 2-5 in Paper
II). However, at present this conclusion cannot be supported by mRNA transcript levels.
Therefore, care must be taken when assuming that ML277 can bind to and activate Kv7.1
channels of pig origin as the homology of the channels has not been compared to that of
human Kv7.1 channels. ML277 has been reported as having an EC50 of 0.26 µM on Kv7.1
channels and produce less than 25% activation of Kv7.4 channels at 30 µM (Mattmann et al.,
2012). Accordingly, it is very likely that the effect of 10 µM ML277 is indeed mediated by
Kv7.1 channels. Similarly, the Kv7.1 channel activator, L-364373, decreased the muscle force
of bladder strips in guinea pigs (Afeli et al., 2013a), while chromanol 293B increased the
contractility of spontaneous contractions (Anderson et al., 2013). Thus, Kv7.1 channels would
appear to be important for urinary bladder function in both guinea pigs and humans. Kv7.4/
Kv7.5 selective activators and blockers, on the other hand, have not been used on guinea pig
and rat bladder. Therefore, it remains to be elucidated whether, or not, Kv7.4/Kv7.5 channels
are functionally important in these species. Fortunately, pigs seem to constitute promising
animal models for the evaluation of Kv7 channel function in human urinary bladders.
Regulation of Kv7 channels in the detrusor.
In summary, we found that Kv7 channels are important regulators of detrusor contractility in
both pigs and humans, findings that led us to further study the pathways underlying Kv7
channel mediated relaxation.
Functional evaluation of localisation
Kv7 channels have been identified as being present in neurons as well as in smooth muscle
cells. However, at present, the localisation of Kv7 channels in the urinary bladder has only
been studied in relation to guinea pigs (Afeli et al., 2013a; Anderson et al., 2013). In our
second experimental setup (Paper II), we performed a series of functional studies using a
denervation detrusor strip protocol to which we had added KPSS and tetrodotoxin on pig
detrusor strips. The aim of the experiment was to test whether, or not, the response to Kv7
"35

RESULTS AND DISCUSSION
channel modulators depended upon Kv7 channels being present in nerves, or solely upon
those located in smooth muscle. These experiments revealed that the Kv7 channel mediated
response on strip contractility was nerve-independent, thereby proving that Kv7 channels in
smooth muscle cells mediate the observed responses in isolated detrusor strips (Fig. 9).
Similar results were obtained with rat detrusor (Svalø et al., 2011). While we cannot
completely rule out the fact that the inhibition of Kv7 channels by XE991 induces
depolarisation, Ca2+ influx and transmitter release due to the XE991 induced blockade of Kv7
channels, we did not observe any difference in response between that of vehicle and XE991.
Furthermore, our findings are supported by studies carried out on guinea pig bladders. These
studies showed that Kv7 channels are localised in interstitial cells of Cajal and in smooth
muscle cells (Anderson et al., 2009; Afeli et al., 2013a; Anderson et al., 2013), but not in
neurons.
"36
125 mM KPSS 1 µM carbachol 10 µM retigabine 1 µM carbachol 10 µM retigabine
Figure 9. Denervation of pig detrusor strips. Original trace recording of the effect of retigabine (10 µM) or vehicle on 125 mM KPSS + TTX (A) or control (B) treated detrusor strips. Bar chart of the responses to retigabine (10 µM), XE991 (10 µM) or vehicle normalised to 1 µM pre-contraction tension (C).
A B
!!!
!
C

RESULTS AND DISCUSSION
PIP2 dependency
The potassium current passing through Kv7.2 and Kv7.3 channels, originally named the M-
current, was first described in neurons. As previously mentioned the inhibition of Kv7
channels by XE991 was only seen to be linked to increased tone at low membrane
depolarisation (Fig. 7) and not at muscarinic receptor stimulation (Paper II and III). This
could be explained by Kv7 channels being inhibited by PIP2 depletion via the activation of
muscarinic M3-receptors (Li et al., 2005; Suh et al., 2006; Linley et al., 2012). This process
has been observed, in the case of both Kv7.2/Kv7.3 channels and that of Kv7.4 channels
(Hernandez et al., 2008; Leitner et al., 2011). Although these are preliminary results, we
believe that PIP2 depletion may account for the lack of response to XE991.
Involvement of Kv7 channels in the Gs pathway
β-adrenoceptors are Gs protein coupled receptors, with the activation of the receptors leading
to the activation of adenylyl cyclase and subsequently to cyclic adenosine monophosphate
(cAMP) synthesis followed by protein kinase A activation. However, this pathway is currently
being reinvestigated. Interestingly, the activation of β3-adrenoceptors has been shown to
induce detrusor relaxation in humans (Morita et al., 2000; Igawa et al., 2001; Wuest et al.,
2009). Moreover, in rat renal arteries β-adrenoceptor mediated relaxation has been shown to
depend upon Kv7.4 channels (Chadha et al., 2012). We therefore, started out by studying the
role of β-adrenoceptors in human detrusor, both that of patients with normal bladder function
(Fig. 10) and that of patients with BOO (with and without detrusor overactivity) (Paper I). We
then went on to investigate whether, or not, an interplay exists between β-adrenoceptors and
Kv7 channels (Paper III).
In Paper I we found that while the β3-adrenoceptor agonist, mirabegron, had a lower potency
(P<0.01), it also had a larger efficacy (P>0.05) compared to that of isoprenaline within all
three patient groups. Furthermore, the efficacy of the two compounds did not differ amongst
the patient groups (P>0.05). Accordingly, since mirabegron is at least as efficacious as
isoprenaline, β3-adrenoceptors accounts for the β-adrenoceptor mediated relaxation in all
three patient groups. We confirmed our findings in Paper III, in which the efficacy of
isoprenaline on 1 µM carbachol-induced contractions did not differ from that described in
Paper I in relation to normal bladder specimens (P=0.45). This supports the observation by
"37

RESULTS AND DISCUSSION
others that β3-adrenoceptors are the main subtype in the urinary bladder of humans (Morita et
al., 2000; Igawa et al., 2001; Wuest et al., 2009). With regard to the animal species in which
Kv7 channel function has been evaluated, β-adrenoceptor-induced relaxation is mediated by
both β1-3-adrenoceptors in the case of rats (Longhurst and Levendusky, 1999) and β2-3-
adrenoceptors in the case of pigs (Yamanishi et al., 2002). In contrast, in guinea pig bladder
the β3-adrenoceptors have been shown to be expressed, though only playing a minor role for
the functionality of detrusor strips (Afeli et al., 2012a). Thus, it is evident that guinea pigs do
not constitute a promising model of human bladder β-adrenoceptor function.
β-adrenoceptor activation in the urinary bladder has been shown not necessarily to lead to
cAMP production, but instead to BKCa channel activation (Kobayashi et al., 2000; Frazier et
al., 2005; Uchida et al., 2005). We tested the adenylyl cyclase activator, forskolin, and found
that during muscarinic receptor stimulation (1 µM) and direct membrane depolarisation (40
mM and 20 mM KPSS) the adenylyl cyclase pathway could be stimulated. We also found that
inhibition of protein kinase A by Rp-cAMPS (Rp-Adenosine 3′,5′-cyclic Monophos-
phorothioate, sodium salt) did not decrease the response to isoprenaline in pig detrusor strips
(see Fig. S2 in Paper III). Thus, we did not find evidence for a cAMP independent pathway in
pre-contracted pig detrusor strips. These findings are similar to those made by Frazier et al.
(2005) and Uchida et al. (2005) in rats using 50 mM and 40 mM KPSS, respectively.
"38
Figure 10. Traces of functional in vitro studies of β-adrenoceptor agonists. Representative traces of 1 µM carbachol-induced contractions in normal human detrusor strips. Strips were treated with mirabegron (1 nM - 10 µM, upper) or vehicle (DMSO, lower) (A) or isoprenaline + 0.1 mM ascorbic acid (1 nM - 10 µM, upper) or vehicle (DMSO + 0.1 mM ascorbic acid, lower) (B).
Vehicle Vehicle

RESULTS AND DISCUSSION
However, Frazier et al. (2005) and Uchida et al. (2005) demonstrated that at passive tension,
SQ22536 (i.e. an adenylyl cyclase inhibitor) and H7 (i.e. a protein kinase A inhibitor)
decreased the potency of isoprenaline; hence, isoprenaline mediated relaxation was cAMP
dependent under these experimental settings. Therefore, in our opinion, the experiments
performed in Paper III should be repeated in human detrusor strips at passive tension before
reaching a final conclusion regarding cAMP pathways in human detrusor strips.
As previously described, Frazier et al. (2005) and Uchida et al. (2005) have demonstrated that
isoprenaline mediated relaxation was dependent upon SKCa and BKCa channel activation in
50/40 mM KPSS pre-contracted rat bladder strips. In connection with this, we investigated
whether Kv7 channels are important for the Gs pathway in the human urinary bladder (Paper
III). We did this by pre-incubating the Kv7 channel modulators, retigabine (i.e. positive
modulator of Kv7.2-7.5, 10 µM) and XE991 (i.e. negative modulator of Kv7.1-7.5, 10 µM)
before the cumulative addition of either isoprenaline or forskolin. We subsequently found that
neither of the compounds affected the β-adrenoceptor adenylyl cyclase mediated relaxation of
human detrusor strips (see Fig. 5-7 in Paper III). We therefore concluded that there does not
seem to be an interplay between Kv7 channels and β-adrenoceptors under these conditions in
human urinary bladder. Thus, SKCa and BKCa channels might be the only potassium channels
relevant for pre-contracted β-adrenoceptor mediated relaxation in the bladder, though this
needs to be confirmed in human detrusor. Furthermore, even though we found the effect of
mirabegron and isoprenaline not to differ amongst the three patient groups, it would be
preferable to confirm our findings in Paper III in all three patient groups. The study on Kv7.4
channels in rat renal arteries did not discriminate between the various β-adrenoceptor
subtypes (β1, β2 and β3) (Chadha et al., 2012), and to the best of our knowledge, the
expression of β3-adrenoceptors in renal arteries has not been studied. Preliminary data
presented by Jahan et al. (2014) described how β1 and β2-adrenoceptors are important
regulators of renal artery contractility in rats. As β3-adrenoceptors predominate in the human
urinary bladder, variations in subtype distribution might be the reason why the β-adrenergic
pathway is independent on Kv7 channels in the bladder but leads to the activation of Kv7.4
channels in renal arteries (Chadha et al., 2012). Interestingly, if Kv7 channels are not involved
in β-adrenoceptor mediated relaxation the former may be seen as providing an independent
"39

RESULTS AND DISCUSSION
mechanism for the relaxation of the detrusor, and as such could provide a novel target for the
treatment of bladder disorders.
Clinical impact of our findings
Our novel findings in human and pig bladder have shown that Kv7 channels are functionally
important in vitro (Paper II and III). Our results point towards Kv7.4 and/or Kv7.5 channels
being the main contributors of Kv7 channel mediated relaxation of the detrusor. Previous in
vivo studies on rats (Streng et al., 2004; Svalø et al., 2011) and clinical studies in humans
(Brickel et al., 2012) using retigabine, have shown promising features of Kv7 channel
activation in vivo with regard to the treatment of OAB.
When considering the risk of side effects, clinical studies with retigabine and flupirtine have
not reported any cardiovascular side effects (Herrmann et al., 1987; French et al., 2011).
Therefore, it seems possible to target Kv7 channels in the bladder without affecting the
cardiovascular system. Even though KCNQ4 (Kharkovets et al., 2000; Hansen et al., 2006)
and KCNQ5 (Jensen et al., 2005; Caminos et al., 2007) are also present in brain tissue, as we
have also shown in pig tissue (Paper II), we argue that the development of Kv7.4/Kv7.5
selective compounds, which cannot pass the blood-brain-barrier, would prove useful for the
treatment of OAB. In addition, different KCNQ-KCNE complexes have different
pharmacologies and the knowledge of KCNQ and KCNE expression in other human tissues
would additionally aid the development of bladder specific compounds.
"40

CONCLUSION AND PERSPECTIVES
Conclusion and Perspectives
In connection with Papers II and III, we found that there was a profound overlap between Kv7
channel subtypes in pig bladder tissue compared to those in human bladder tissue. We also
found evidence for a PIP2 dependent activation of the Kv7 channels in both species.
Unfortunately, the genes encoding Kv7.1-7.2 channels have not yet been determined in pigs
and this need to be done before we can determine if pigs are indeed a good model of human
urinary bladder function in relation to Kv7 channels.
In connection with Papers I and III, our functional studies have underlined the fact that Kv7
channels and β-adrenoceptors are efficient mediators of urinary bladder relaxation in vitro.
For β-adrenoceptor agonists this was furthermore determined in patients with BOO with or
without detrusor overactivity, which supports the proof-of-principle that the β3-adrenoceptor
agonist, mirabegron, is appropriate for the treatment of OAB. This has led us to believe that
these patients will experience an improved quality of life when they commence this novel
treatment. In addition, we discovered that Kv7 channels do not seem to be involved in the β-
adrenergic mediated relaxation, although we did not assess the effect of β-adrenoceptor
agonists and Kv7 channel modulators on spontaneous contractions. However, it seems that
Kv7 channel activation is an independent mechanism for mediating urinary bladder relaxation.
In Paper III we discovered an up-regulation of KCNQ1 and KCNE3 in patients with BOO in
combination with a down-regulation of KCNE1. However, due to the low number of biopsies
in the present study the changes in KCNE expression were not significantly different from the
levels found in patients with a normal, functioning, bladder. Whether, or not, these changes
were in fact dependent upon the patient group needs to be further studied by increasing the
number of bladder specimens and performing electrophysiological recordings. This will help
to reveal whether, or not, a constitutively active channel is formed as a compensatory
mechanism in patients with BOO. Furthermore, western blot analysis and immunohisto-
chemical studies are needed to determine the amount and cellular localisation of the various
Kv7 channel subtypes and their β-subunits.
Overall, our studies have revealed that Kv7 channels are not only important for urinary
bladder function in smaller mammals, but also in pigs and humans. These findings have led us
to believe that the use of Kv7 channel activators could prove useful for the treatment of OAB.
"41

BIBLIOGRAPHY
Bibliography
Abbott, GW (2014). Biology of the KCNQ1 Potassium Channel. New Journal of Science 2014: 1-26.
Abrams, P, Cardozo, L, Fall, M, Griffiths, D, Rosier, P, Ulmsten, U, Van Kerrebroeck, P, Victor, A and Wein, A (2003). The standardisation of terminology in lower urinary tract function: report from the standardisation sub-committee of the International Continence Society. Urology 61(1): 37-49.
Abrams, PH, Farrar, DJ, Turner-Warwick, RT, Whiteside, CG and Feneley, RC (1979). The results of prostatectomy: a symptomatic and urodynamic analysis of 152 patients. J Urol 121(5): 640-642.
Afeli, SA, Hristov, KL and Petkov, GV (2012a). Do beta3-adrenergic receptors play a role in guinea pig detrusor smooth muscle excitability and contractility? Am J Physiol Renal Physiol 302(2): F251-263.
Afeli, SA, Malysz, J and Petkov, GV (2013a). Molecular expression and pharmacological evidence for a functional role of kv7 channel subtypes in Guinea pig urinary bladder smooth muscle. PLoS One 8(9): e75875.
Afeli, SA, Rovner, ES and Petkov, GV (2012b). SK but not IK channels regulate human detrusor smooth muscle spontaneous and nerve-evoked contractions. Am J Physiol Renal Physiol 303(4): F559-568.
Afeli, SA, Rovner, ES and Petkov, GV (2013b). BRL37344, a beta3-adrenergic receptor agonist, decreases nerve-evoked contractions in human detrusor smooth muscle isolated strips: role of BK channels. Urology 82(3): 744 e741-747.
Aishima, M, Tomoda, T, Yunoki, T, Nakano, T, Seki, N, Yonemitsu, Y, Sueishi, K, Naito, S, Ito, Y and Teramoto, N (2006). Actions of ZD0947, a novel ATP-sensitive K+ channel opener, on membrane currents in human detrusor myocytes. Br J Pharmacol 149(5): 542-550.
Ameda, K, Koyanagi, T, Nantani, M, Taniguchi, K and Matsuno, T (1994). The relevance of preoperative cystometrography in patients with benign prostatic hyperplasia: correlating the findings with clinical features and outcome after prostatectomy. J Urol 152(2 Pt 1): 443-447.
Anderson, UA, Carson, C, Johnston, L, Joshi, S, Gurney, AM and McCloskey, KD (2013). Functional expression of KCNQ (Kv 7) channels in guinea pig bladder smooth muscle and their contribution to spontaneous activity. Br J Pharmacol 169(6): 1290-1304.
Anderson, UA, Carson, C and McCloskey, KD (2009). KCNQ currents and their contribution to resting membrane potential and the excitability of interstitial cells of Cajal from the guinea pig bladder. J Urol 182(1): 330-336.
"42

BIBLIOGRAPHY
Andersson, KE (2007). LUTS treatment: future treatment options. Neurourol Urodyn 26(6 Suppl): 934-947.
Andersson, KE and Arner, A (2004). Urinary bladder contraction and relaxation: physiology and pathophysiology. Physiol Rev 84(3): 935-986.
Andersson, KE and McCloskey, KD (2014). Lamina propria: the functional center of the bladder? Neurourol Urodyn 33(1): 9-16.
Aydin, M, Wang, HZ, Zhang, X, Chua, R, Downing, K, Melman, A and DiSanto, ME (2012). Large-conductance calcium-activated potassium channel activity, as determined by whole-cell patch clamp recording, is decreased in urinary bladder smooth muscle cells from male rats with partial urethral obstruction. BJU Int 110(8 Pt B): E402-408.
Badawi, JK, Ding, A and Bross, S (2008). Inhibitory effects of different ATP-sensitive potassium channel openers on electrically generated and carbachol-induced contractions of porcine and human detrusor muscle. Fundam Clin Pharmacol 22(1): 75-86.
Badawi, JK, Seja, T, Uecelehan, H, Honeck, P, Kwon, ST, Bross, S and Langbein, S (2007). Relaxation of human detrusor muscle by selective beta-2 and beta-3 agonists and endogenous catecholamines. Urology 69(4): 785-790.
Barhanin, J, Lesage, F, Guillemare, E, Fink, M, Lazdunski, M and Romey, G (1996). K(V)LQT1 and lsK (minK) proteins associate to form the I(Ks) cardiac potassium current. nature 384(6604): 78-80.
Baysting, L and McCloskey, KD (2014). Investigation of Kv7 (KCNQ) modulators on contractility of normal and overactive rat bladder. Physiology 2014, London, UK, Physiological Society. PCB187.
Bendahhou, S, Marionneau, C, Haurogne, K, Larroque, MM, Derand, R, Szuts, V, Escande, D, Demolombe, S and Barhanin, J (2005). In vitro molecular interactions and distribution of KCNE family with KCNQ1 in the human heart. Cardiovasc Res 67(3): 529-538.
Bentzen, BH, Schmitt, N, Calloe, K, Dalby Brown, W, Grunnet, M and Olesen, SP (2006). The acrylamide (S)-1 differentially affects Kv7 (KCNQ) potassium channels. Neuropharmacology 51(6): 1068-1077.
Bett, GC, Morales, MJ, Beahm, DL, Duffey, ME and Rasmusson, RL (2006). Ancillary subunits and stimulation frequency determine the potency of chromanol 293B block of the KCNQ1 potassium channel. J Physiol 576(Pt 3): 755-767.
Bianchi, L, Shen, Z, Dennis, AT, Priori, SG, Napolitano, C, Ronchetti, E, Bryskin, R, Schwartz, PJ and Brown, AM (1999). Cellular dysfunction of LQT5-minK mutants: abnormalities of IKs, IKr and trafficking in long QT syndrome. Hum Mol Genet 8(8): 1499-1507.
"43

BIBLIOGRAPHY
Biers, SM, Reynard, JM and Brading, AF (2006). The effects of a new selective beta3-adrenoceptor agonist (GW427353) on spontaneous activity and detrusor relaxation in human bladder. BJU Int 98(6): 1310-1314.
Biervert, C (1998). A Potassium Channel Mutation in Neonatal Human Epilepsy. Science 279(5349): 403-406.
Bille, M and Parameswaran Theepakaran, N (2012). Characterisation of the voltage-gated potassium channel Kv7 in pig detrusor muscle. (Master), University of Copenhagen.
Birder, LA, Apodaca, G, De Groat, WC and Kanai, AJ (1998). Adrenergic- and capsaicin-evoked nitric oxide release from urothelium and afferent nerves in urinary bladder. Am J Physiol 275(2 Pt 2): F226-229.
Birder, LA, Kanai, AJ, Cruz, F, Moore, K and Fry, CH (2010). Is the urothelium intelligent? Neurourol Urodyn 29(4): 598-602.
Blom, SM, Rottlander, M, Kehler, J, Bundgaard, C, Schmitt, N and Jensen, HS (2014). From pan-reactive KV7 channel opener to subtype selective opener/inhibitor by addition of a methyl group. PLoS One 9(6): e100209.
Blom, SM, Schmitt, N and Jensen, HS (2010). Differential effects of ICA-27243 on cloned K(V)7 channels. Pharmacology 86(3): 174-181.
Brickel, N, Gandhi, P, VanLandingham, K, Hammond, J and DeRossett, S (2012). The urinary safety profile and secondary renal effects of retigabine (ezogabine): a first-in-class antiepileptic drug that targets KCNQ (K(v)7) potassium channels. Epilepsia 53(4): 606-612.
Brown, DA and Adams, PR (1980). Muscarinic suppression of a novel voltage-sensitive K+ current in a vertebrate neurone. nature 283(5748): 673-676.
Brueggemann, LI, Haick, JM, Cribbs, LL and Byron, KL (2014). Differential Activation of Vascular Smooth Muscle Kv7.4, Kv7.5, and Kv7.4/7.5 Channels by ML213 and ICA-069673. Mol Pharmacol, 10.1124/mol.114.093799.
Brueggemann, LI, Kakad, PP, Love, RB, Solway, J, Dowell, ML, Cribbs, LL and Byron, KL (2012). Kv7 potassium channels in airway smooth muscle cells: signal transduction intermediates and pharmacological targets for bronchodilator therapy. Am J Physiol Lung Cell Mol Physiol 302(1): L120-132.
Buckner, SA, Milicic, I, Daza, A, Davis-Taber, R, Scott, VE, Sullivan, JP and Brioni, JD (2000). Pharmacological and molecular analysis of ATP-sensitive K(+) channels in the pig and human detrusor. Eur J Pharmacol 400(2-3): 287-295.
"44

BIBLIOGRAPHY
Caminos, E, Garcia-Pino, E, Martinez-Galan, JR and Juiz, JM (2007). The potassium channel KCNQ5/Kv7.5 is localized in synaptic endings of auditory brainstem nuclei of the rat. J Comp Neurol 505(4): 363-378.
Chadha, PS, Jepps, TA, Carr, G, Stott, JB, Zhu, HL, Cole, WC and Greenwood, IA (2014). Contribution of kv7.4/kv7.5 heteromers to intrinsic and calcitonin gene-related peptide-induced cerebral reactivity. Arterioscler Thromb Vasc Biol 34(4): 887-893.
Chadha, PS, Zunke, F, Zhu, HL, Davis, AJ, Jepps, TA, Olesen, SP, Cole, WC, Moffatt, JD and Greenwood, IA (2012). Reduced KCNQ4-encoded voltage-dependent potassium channel activity underlies impaired beta-adrenoceptor-mediated relaxation of renal arteries in hypertension. Hypertension 59(4): 877-884.
Chang, S, Gomes, CM, Hypolite, JA, Marx, J, Alanzi, J, Zderic, SA, Malkowicz, B, Wein, AJ and Chacko, S (2010). Detrusor overactivity is associated with downregulation of large-conductance calcium- and voltage-activated potassium channel protein. Am J Physiol Renal Physiol 298(6): F1416-1423.
Chapple, CR, Kaplan, SA, Mitcheson, D, Klecka, J, Cummings, J, Drogendijk, T, Dorrepaal, C and Martin, N (2013). Randomized double-blind, active-controlled phase 3 study to assess 12-month safety and efficacy of mirabegron, a beta(3)-adrenoceptor agonist, in overactive bladder. Eur Urol 63(2): 296-305.
Chapple, CR, Patroneva, A and Raines, SR (2006). Effect of an ATP-sensitive potassium channel opener in subjects with overactive bladder: a randomized, double-blind, placebo-controlled study (ZD0947IL/0004). Eur Urol 49(5): 879-886.
Ciampa, EJ, Welch, RC, Vanoye, CG and George, AL, Jr. (2011). KCNE4 juxtamembrane region is required for interaction with calmodulin and for functional suppression of KCNQ1. J Biol Chem 286(6): 4141-4149.
Clemow, DB, Steers, WD, McCarty, R and Tuttle, JB (1998). Altered regulation of bladder nerve growth factor and neurally mediated hyperactive voiding. Am J Physiol 275(4 Pt 2): R1279-1286.
Coyne, KS, Sexton, CC, Bell, JA, Thompson, CL, Dmochowski, R, Bavendam, T, Chen, CI and Quentin Clemens, J (2013). The prevalence of lower urinary tract symptoms (LUTS) and overactive bladder (OAB) by racial/ethnic group and age: results from OAB-POLL. Neurourol Urodyn 32(3): 230-237.
Coyne, KS, Sexton, CC, Thompson, CL, Milsom, I, Irwin, D, Kopp, ZS, Chapple, CR, Kaplan, S, Tubaro, A, Aiyer, LP and Wein, AJ (2009). The prevalence of lower urinary tract symptoms (LUTS) in the USA, the UK and Sweden: results from the Epidemiology of LUTS (EpiLUTS) study. BJU Int 104(3): 352-360.
"45

BIBLIOGRAPHY
Crowe, R and Burnstock, G (1989). A histochemical and immunohistochemical study of the autonomic innervation of the lower urinary tract of the female pig. Is the pig a good model for the human bladder and urethra? J Urol 141(2): 414-422.
Darblade, B, Behr-Roussel, D, Oger, S, Hieble, JP, Lebret, T, Gorny, D, Benoit, G, Alexandre, L and Giuliano, F (2006). Effects of potassium channel modulators on human detrusor smooth muscle myogenic phasic contractile activity: potential therapeutic targets for overactive bladder. Urology 68(2): 442-448.
Davies, AM, Batchelor, TJ, Eardley, I and Beech, DJ (2002). Potassium channel KV alpha1 subunit expression and function in human detrusor muscle. J Urol 167(4): 1881-1886.
de Moura, RS, de Mello, RF and D'Aguinaga, S (1993). Inhibitory effect of cromakalim in human detrusor muscle is mediated by glibenclamide-sensitive potassium channels. J Urol 149(5): 1174-1177.
Dombkowski, RA, Doellman, MM, Head, SK and Olson, KR (2006). Hydrogen sulfide mediates hypoxia-induced relaxation of trout urinary bladder smooth muscle. J Exp Biol 209(Pt 16): 3234-3240.
Dupuis, DS, Schrøder, RL, Jespersen, T, Christensen, JK, Christophersen, P, Jensen, BS and Olesen, SP (2002). Activation of KCNQ5 channels stably expressed in HEK293 cells by BMS-204352. European Journal of Pharmacology 437(3): 129-137.
Ekman, M, Rippe, C, Sadegh, MK, Dabestani, S, Morgelin, M, Uvelius, B and Sward, K (2012). Association of muscarinic M(3) receptors and Kir6.1 with caveolae in human detrusor muscle. Eur J Pharmacol 683(1-3): 238-245.
Elliott, TR (1907). The innervation of the bladder and urethra. J Physiol 35(5-6): 367-445.
Etxeberria, A, Aivar, P, Rodriguez-Alfaro, JA, Alaimo, A, Villace, P, Gomez-Posada, JC, Areso, P and Villarroel, A (2008). Calmodulin regulates the trafficking of KCNQ2 potassium channels. FASEB J 22(4): 1135-1143.
Ferguson, DR, Kennedy, I and Burton, TJ (1997). ATP is released from rabbit urinary bladder epithelial cells by hydrostatic pressure changes--a possible sensory mechanism? J Physiol 505 ( Pt 2): 503-511.
Fouda, AK, Kaufmann, A, Thorin, E, Henrion, D, Capdeville-Atkinson, C and Atkinson, J (1991). The role of endogenous norepinephrine release in potassium-evoked vasoconstriction of the rat tail artery. Eur J Pharmacol 205(1): 63-72.
Fowler, CJ, Griffiths, D and de Groat, WC (2008). The neural control of micturition. Nat Rev Neurosci 9(6): 453-466.
Frazier, EP, Mathy, MJ, Peters, SL and Michel, MC (2005). Does cyclic AMP mediate rat urinary bladder relaxation by isoproterenol? J Pharmacol Exp Ther 313(1): 260-267.
"46

BIBLIOGRAPHY
French, JA, Abou-Khalil, BW, Leroy, RF, Yacubian, EM, Shin, P, Hall, S, Mansbach, H, Nohria, V and Investigators, RS (2011). Randomized, double-blind, placebo-controlled trial of ezogabine (retigabine) in partial epilepsy. Neurology 76(18): 1555-1563.
Gai, JW, Wahafu, W, Guo, H, Liu, M, Wang, XC, Xiao, YX, Zhang, L, Xin, ZC and Jin, J (2013). Further evidence of endogenous hydrogen sulphide as a mediator of relaxation in human and rat bladder. Asian J Androl 15(5): 692-696.
Gamper, N, Li, Y and Shapiro, MS (2005). Structural requirements for differential sensitivity of KCNQ K+ channels to modulation by Ca2+/calmodulin. Mol Biol Cell 16(8): 3538-3551.
Gamper, N and Shapiro, MS (2003). Calmodulin mediates Ca2+-dependent modulation of M-type K+ channels. J Gen Physiol 122(1): 17-31.
Grahammer, F, Wittekindt, OH, Nitschke, R, Herling, AW, Lang, HJ, Bleich, M, Schmitt–Gräff, A, Barhanin, J and Warth, R (2001). The cardiac K+ channel KCNQ1 is essential for gastric acid secretion. Gastroenterology 120(6): 1363-1371.
Gray, SM, McGeown, JG, McMurray, G and McCloskey, KD (2013). Functional innervation of Guinea-pig bladder interstitial cells of cajal subtypes: neurogenic stimulation evokes in situ calcium transients. PLoS One 8(1): e53423.
Grunnet, M, Jespersen, T, Rasmussen, HB, Ljungstrom, T, Jorgensen, NK, Olesen, SP and Klaerke, DA (2002). KCNE4 is an inhibitory subunit to the KCNQ1 channel. J Physiol 542(Pt 1): 119-130.
Grunnet, M, Olesen, SP, Klaerke, DA and Jespersen, T (2005). hKCNE4 inhibits the hKCNQ1 potassium current without affecting the activation kinetics. Biochem Biophys Res Commun 328(4): 1146-1153.
Hanna-Mitchell, AT, Kashyap, M, Chan, WV, Andersson, KE and Tannenbaum, C (2014). Pathophysiology of idiopathic overactive bladder and the success of treatment: a systematic review from ICI-RS 2013. Neurourol Urodyn 33(5): 611-617.
Hansen, HH, Ebbesen, C, Mathiesen, C, Weikop, P, Ronn, LC, Waroux, O, Scuvee-Moreau, J, Seutin, V and Mikkelsen, JD (2006). The KCNQ channel opener retigabine inhibits the activity of mesencephalic dopaminergic systems of the rat. J Pharmacol Exp Ther 318(3): 1006-1019.
Hashitani, H and Brading, AF (2003). Electrical properties of detrusor smooth muscles from the pig and human urinary bladder. Br J Pharmacol 140(1): 146-158.
Hashitani, H, Yanai, Y and Suzuki, H (2004). Role of interstitial cells and gap junctions in the transmission of spontaneous Ca2+ signals in detrusor smooth muscles of the guinea-pig urinary bladder. J Physiol 559(Pt 2): 567-581.
"47

BIBLIOGRAPHY
Hedegaard, ER, Nielsen, BD, Kun, A, Hughes, AD, Kroigaard, C, Mogensen, S, Matchkov, VV, Frobert, O and Simonsen, U (2014). KV 7 channels are involved in hypoxia-induced vasodilatation of porcine coronary arteries. Br J Pharmacol 171(1): 69-82.
Hernandez, CC, Zaika, O and Shapiro, MS (2008). A carboxy-terminal inter-helix linker as the site of phosphatidylinositol 4,5-bisphosphate action on Kv7 (M-type) K+ channels. J Gen Physiol 132(3): 361-381.
Herrmann, WM, Kern, U and Aigner, M (1987). On the adverse reactions and efficacy of long-term treatment with flupirtine: preliminary results of an ongoing twelve-month study with 200 patients suffering from chronic pain states in arthrosis or arthritis. Postgrad Med J 63 Suppl 3: 87-103.
Herschorn, S, Barkin, J, Castro-Diaz, D, Frankel, JM, Espuna-Pons, M, Gousse, AE, Stolzel, M, Martin, N, Gunther, A and Van Kerrebroeck, P (2013). A phase III, randomized, double-blind, parallel-group, placebo-controlled, multicentre study to assess the efficacy and safety of the beta(3) adrenoceptor agonist, mirabegron, in patients with symptoms of overactive bladder. Urology 82(2): 313-320.
Hristov, KL, Afeli, SA, Parajuli, SP, Cheng, Q, Rovner, ES and Petkov, GV (2013). Neurogenic detrusor overactivity is associated with decreased expression and function of the large conductance voltage- and Ca(2+)-activated K(+) channels. PLoS One 8(7): e68052.
Hristov, KL, Chen, M, Afeli, SA, Cheng, Q, Rovner, ES and Petkov, GV (2012a). Expression and function of K(V)2-containing channels in human urinary bladder smooth muscle. Am J Physiol Cell Physiol 302(11): C1599-1608.
Hristov, KL, Chen, M, Kellett, WF, Rovner, ES and Petkov, GV (2011). Large-conductance voltage- and Ca2+-activated K+ channels regulate human detrusor smooth muscle function. Am J Physiol Cell Physiol 301(4): C903-912.
Hristov, KL, Parajuli, SP, Soder, RP, Cheng, Q, Rovner, ES and Petkov, GV (2012b). Suppression of human detrusor smooth muscle excitability and contractility via pharmacological activation of large conductance Ca2+-activated K+ channels. Am J Physiol Cell Physiol 302(11): C1632-1641.
Hubbard, JI and Kwanbunbumpen, S (1968). Evidence for the vesicle hypothesis. J Physiol 194(2): 407-420.
Igawa, Y, Yamazaki, Y, Takeda, H, Akahane, M, Ajisawa, Y, Yoneyama, T and Nishizawa, O (1998). Possible beta 3-adrenoceptor-mediated relaxation of the human detrusor. Acta Physiol Scand 164(1): 117-118.
Igawa, Y, Yamazaki, Y, Takeda, H, Hayakawa, K, Akahane, M, Ajisawa, Y, Yoneyama, T, Nishizawa, O and Andersson, KE (1999). Functional and molecular biological evidence
"48

BIBLIOGRAPHY
for a possible beta3-adrenoceptor in the human detrusor muscle. Br J Pharmacol 126(3): 819-825.
Igawa, Y, Yamazaki, Y, Takeda, H, Kaidoh, K, Akahane, M, Ajisawa, Y, Yoneyama, T, Nishizawa, O and Andersson, KE (2001). Relaxant effects of isoproterenol and selective beta3-adrenoceptor agonists on normal, low compliant and hyperreflexic human bladders. J Urol 165(1): 240-244.
Ipavec, V, Martire, M, Barrese, V, Taglialatela, M and Curro, D (2011). KV7 channels regulate muscle tone and nonadrenergic noncholinergic relaxation of the rat gastric fundus. Pharmacol Res 64(4): 397-409.
Irwin, DE, Milsom, I, Hunskaar, S, Reilly, K, Kopp, Z, Herschorn, S, Coyne, K, Kelleher, C, Hampel, C, Artibani, W and Abrams, P (2006). Population-based survey of urinary incontinence, overactive bladder, and other lower urinary tract symptoms in five countries: results of the EPIC study. Eur Urol 50(6): 1306-1314; discussion 1314-1305.
Isbrandt, D, Friederich, P, Solth, A, Haverkamp, W, Ebneth, A, Borggrefe, M, Funke, H, Sauter, K, Breithardt, G, Pongs, O and Schulze-Bahr, E (2002). Identification and functional characterization of a novel KCNE2 (MiRP1) mutation that alters HERG channel kinetics. J Mol Med (Berl) 80(8): 524-532.
Jahan, KS, Stott, JB and Greenwood, IA (2014). Further investigations into the role of Kv7 channels in beta-adrenoceptor mediated relaxation of renal arteries. Physiology 2014, London, UK, PCB186.
Jensen, HS, Callo, K, Jespersen, T, Jensen, BS and Olesen, SP (2005). The KCNQ5 potassium channel from mouse: a broadly expressed M-current like potassium channel modulated by zinc, pH, and volume changes. Brain Res Mol Brain Res 139(1): 52-62.
Jepps, TA, Chadha, PS, Davis, AJ, Harhun, MI, Cockerill, GW, Olesen, SP, Hansen, RS and Greenwood, IA (2011). Downregulation of Kv7.4 channel activity in primary and secondary hypertension. Circulation 124(5): 602-611.
Jepps, TA, Greenwood, IA, Moffatt, JD, Sanders, KM and Ohya, S (2009). Molecular and functional characterization of Kv7 K+ channel in murine gastrointestinal smooth muscles. Am J Physiol Gastrointest Liver Physiol 297(1): G107-115.
Jepps, TA, Olesen, SP and Greenwood, IA (2013). One man's side effect is another man's therapeutic opportunity: targeting Kv7 channels in smooth muscle disorders. Br J Pharmacol 168(1): 19-27.
Johnston, L, Carson, C, Lyons, AD, Davidson, RA and McCloskey, KD (2008). Cholinergic-induced Ca2+ signaling in interstitial cells of Cajal from the guinea pig bladder. Am J Physiol Renal Physiol 294(3): F645-655.
"49

BIBLIOGRAPHY
Joshi, S, Balan, P and Gurney, AM (2006). Pulmonary vasoconstrictor action of KCNQ potassium channel blockers. Respir Res 7: 31.
Joshi, S, Sedivy, V, Hodyc, D, Herget, J and Gurney, AM (2009). KCNQ modulators reveal a key role for KCNQ potassium channels in regulating the tone of rat pulmonary artery smooth muscle. J Pharmacol Exp Ther 329(1): 368-376.
Kajioka, S, Shahab, N, Asano, H, Morita, H, Sugihara, M, Takahashi-Yanaga, F, Yoshihara, T, Nakayama, S, Seki, N and Naito, S (2011). Diphosphate regulation of adenosine triphosphate sensitive potassium channel in human bladder smooth muscle cells. J Urol 186(2): 736-744.
Kanie, S, Otsuka, A, Yoshikawa, S, Morimoto, T, Hareyama, N, Okazaki, S, Kobayashi, R, Hasebe, K, Nakao, K, Hayashi, R, Mochizuki, H, Matsumoto, R and Ozono, S (2012). Pharmacological effect of TRK-380, a novel selective human beta3-adrenoceptor agonist, on mammalian detrusor strips. Urology 79(3): 744 e741-747.
Kharkovets, T, Hardelin, JP, Safieddine, S, Schweizer, M, El-Amraoui, A, Petit, C and Jentsch, TJ (2000). KCNQ4, a K+ channel mutated in a form of dominant deafness, is expressed in the inner ear and the central auditory pathway. Proc Natl Acad Sci U S A 97(8): 4333-4338.
Khullar, V, Amarenco, G, Angulo, JC, Cambronero, J, Hoye, K, Milsom, I, Radziszewski, P, Rechberger, T, Boerrigter, P, Drogendijk, T, Wooning, M and Chapple, C (2013). Efficacy and tolerability of mirabegron, a beta(3)-adrenoceptor agonist, in patients with overactive bladder: results from a randomised European-Australian phase 3 trial. Eur Urol 63(2): 283-295.
Kobayashi, H, Adachi-Akahane, S and Nagao, T (2000). Involvement of BKCa channels in the relaxation of detrusor muscle via β-adrenoceptors. European Journal of Pharmacology 404(1-2): 231-238.
Kubisch, C (1999). KCNQ4, a novel potassium channel expressed in sensory outer hair cells, is mutated in dominant deafness. Cell 96: 437-446.
La Fuente, JM, Fernandez, A, Cuevas, P, Gonzalez-Corrochano, R, Chen, MX and Angulo, J (2014). Stimulation of large-conductance calcium-activated potassium channels inhibits neurogenic contraction of human bladder from patients with urinary symptoms and reverses acetic acid-induced bladder hyperactivity in rats. Eur J Pharmacol 735: 68-76.
Learmonth, JR (1931). A Contribution to the Neurophysiology of the Urinary Bladder in Man. Brain 54(2): 147-176.
Lei, Q, Pan, XQ, Chang, S, Malkowicz, SB, Guzzo, TJ and Malykhina, AP (2014). Response of the human detrusor to stretch is regulated by TREK-1, a two-pore-domain (K2P) mechano-gated potassium channel. J Physiol 592(Pt 14): 3013-3030.
"50

BIBLIOGRAPHY
Leitner, MG, Halaszovich, CR and Oliver, D (2011). Aminoglycosides inhibit KCNQ4 c h a n n e l s i n c o c h l e a r o u t e r h a i r c e l l s v i a d e p l e t i o n o f phosphatidylinositol(4,5)bisphosphate. Mol Pharmacol 79(1): 51-60.
Lerche, C, Scherer, CR, Seebohm, G, Derst, C, Wei, AD, Busch, AE and Steinmeyer, K (2000). Molecular cloning and functional expression of KCNQ5, a potassium channel subunit that may contribute to neuronal M-current diversity. J Biol Chem 275(29): 22395-22400.
Li, Y, Gamper, N, Hilgemann, DW and Shapiro, MS (2005). Regulation of Kv7 (KCNQ) K+ channel open probability by phosphatidylinositol 4,5-bisphosphate. J Neurosci 25(43): 9825-9835.
Linley, JE, Pettinger, L, Huang, D and Gamper, N (2012). M channel enhancers and physiological M channel block. J Physiol 590(Pt 4): 793-807.
Liu, G and Daneshgari, F (2006). Effects of cold storage on the function and morphology of isolated urinary bladder in rat. Neurourol Urodyn 25(4): 381-387.
Liu, W and Devaux, JJ (2014). Calmodulin orchestrates the heteromeric assembly and the trafficking of KCNQ2/3 (Kv7.2/3) channels in neurons. Mol Cell Neurosci 58: 40-52.
Longhurst, PA and Levendusky, M (1999). Pharmacological characterization of beta-adrenoceptors mediating relaxation of the rat urinary bladder in vitro. Br J Pharmacol 127(7): 1744-1750.
Loussouarn, G, Park, KH, Bellocq, C, Baro, I, Charpentier, F and Escande, D (2003). Phosphatidylinositol-4,5-bisphosphate, PIP2, controls KCNQ1/KCNE1 voltage-gated potassium channels: a functional homology between voltage-gated and inward rectifier K+ channels. EMBO J 22(20): 5412-5421.
Lundby, A, Ravn, LS, Svendsen, JH, Hauns, S, Olesen, SP and Schmitt, N (2008). KCNE3 mutation V17M identified in a patient with lone atrial fibrillation. Cell Physiol Biochem 21(1-3): 47-54.
Mackie, AR, Brueggemann, LI, Henderson, KK, Shiels, AJ, Cribbs, LL, Scrogin, KE and Byron, KL (2008). Vascular KCNQ potassium channels as novel targets for the control of mesenteric artery constriction by vasopressin, based on studies in single cells, pressurized arteries, and in vivo measurements of mesenteric vascular resistance. J Pharmacol Exp Ther 325(2): 475-483.
Main, MJ, Cryan, JE, Dupere, JR, Cox, B, Clare, JJ and Burbidge, SA (2000). Modulation of KCNQ2/3 potassium channels by the novel anticonvulsant retigabine. Mol Pharmacol 58(2): 253-262.
Manderfield, LJ, Daniels, MA, Vanoye, CG and George, AL, Jr. (2009). KCNE4 domains required for inhibition of KCNQ1. J Physiol 587(Pt 2): 303-314.
"51

BIBLIOGRAPHY
Mani, BK, Brueggemann, LI, Cribbs, LL and Byron, KL (2011). Activation of vascular KCNQ (Kv7) potassium channels reverses spasmogen-induced constrictor responses in rat basilar artery. Br J Pharmacol 164(2): 237-249.
Mani, BK, O'Dowd, J, Kumar, L, Brueggemann, LI, Ross, M and Byron, KL (2013). Vascular KCNQ (Kv7) potassium channels as common signaling intermediates and therapeutic targets in cerebral vasospasm. J Cardiovasc Pharmacol 61(1): 51-62.
Martelli, A, Testai, L, Breschi, MC, Lawson, K, McKay, NG, Miceli, F, Taglialatela, M and Calderone, V (2013). Vasorelaxation by hydrogen sulphide involves activation of Kv7 potassium channels. Pharmacol Res 70(1): 27-34.
Mattmann, ME, Yu, H, Lin, Z, Xu, K, Huang, X, Long, S, Wu, M, McManus, OB, Engers, DW, Le, UM, Li, M, Lindsley, CW and Hopkins, CR (2012). Identification of (R)-N-(4-(4-methoxyphenyl)thiazol-2-yl)-1-tosylpiperidine-2-carboxamide, ML277, as a novel, potent and selective K(v)7.1 (KCNQ1) potassium channel activator. Bioorg Med Chem Lett 22(18): 5936-5941.
McCallum, LA, Greenwood, IA and Tribe, RM (2009). Expression and function of K(v)7 channels in murine myometrium throughout oestrous cycle. Pflugers Arch 457(5): 1111-1120.
McCallum, LA, Pierce, SL, England, SK, Greenwood, IA and Tribe, RM (2011). The contribution of Kv7 channels to pregnant mouse and human myometrial contractility. J Cell Mol Med 15(3): 577-586.
McCrea, ED (1926). The Musculature of the Bladder. Proc R Soc Med 19(Sect Urol): 35-43.
Morita, T, Iizuka, H, Iwata, T and Kondo, S (2000). Function and distribution of beta3-adrenoceptors in rat, rabbit and human urinary bladder and external urethral sphincter. J Smooth Muscle Res 36(1): 21-32.
Ng, FL, Davis, AJ, Jepps, TA, Harhun, MI, Yeung, SY, Wan, A, Reddy, M, Melville, D, Nardi, A, Khong, TK and Greenwood, IA (2011). Expression and function of the K+ channel KCNQ genes in human arteries. Br J Pharmacol 162(1): 42-53.
Nielsen, JS, Rode, F, Rahbek, M, Andersson, KE, Ronn, LC, Bouchelouche, K, Nordling, J and Bouchelouche, P (2011). Effect of the SK/IK channel modulator 4,5-dichloro-1,3-diethyl-1,3-dihydro-benzoimidazol-2-one (NS4591) on contractile force in rat, pig and human detrusor smooth muscle. BJU Int 108(5): 771-777.
Nitti, VW, Auerbach, S, Martin, N, Calhoun, A, Lee, M and Herschorn, S (2013). Results of a randomized phase III trial of mirabegron in patients with overactive bladder. J Urol 189(4): 1388-1395.
"52

BIBLIOGRAPHY
Nomiya, M and Yamaguchi, O (2003). A quantitative analysis of mRNA expression of alpha 1 and beta-adrenoceptor subtypes and their functional roles in human normal and obstructed bladders. J Urol 170(2 Pt 1): 649-653.
Nordling, J, Meyhoff, HH and Hald, T (1981). Neuromuscular dysfunction of the lower urinary tract with special reference to the influence of the sympathetic nervous system. Scand J Urol Nephrol 15(1): 7-19.
Ohya, S, Asakura, K, Muraki, K, Watanabe, M and Imaizumi, Y (2002). Molecular and functional characterization of ERG, KCNQ, and KCNE subtypes in rat stomach smooth muscle. Am J Physiol Gastrointest Liver Physiol 282(2): G277-287.
Ohya, S, Sergeant, GP, Greenwood, IA and Horowitz, B (2003). Molecular variants of KCNQ channels expressed in murine portal vein myocytes: a role in delayed rectifier current. Circ Res 92(9): 1016-1023.
Otsuka, A, Shinbo, H, Matsumoto, R, Kurita, Y and Ozono, S (2008). Expression and functional role of beta-adrenoceptors in the human urinary bladder urothelium. Naunyn Schmiedebergs Arch Pharmacol 377(4-6): 473-481.
Patacchini, R, Santicioli, P, Giuliani, S and Maggi, CA (2004). Hydrogen sulfide (H2S) stimulates capsaicin-sensitive primary afferent neurons in the rat urinary bladder. Br J Pharmacol 142(1): 31-34.
Peretz, A, Degani, N, Nachman, R, Uziyel, Y, Gibor, G, Shabat, D and Attali, B (2005). Meclofenamic acid and diclofenac, novel templates of KCNQ2/Q3 potassium channel openers, depress cortical neuron activity and exhibit anticonvulsant properties. Mol Pharmacol 67(4): 1053-1066.
Ravn, LS, Aizawa, Y, Pollevick, GD, Hofman-Bang, J, Cordeiro, JM, Dixen, U, Jensen, G, Wu, Y, Burashnikov, E, Haunso, S, Guerchicoff, A, Hu, D, Svendsen, JH, Christiansen, M and Antzelevitch, C (2008). Gain of function in IKs secondary to a mutation in KCNE5 associated with atrial fibrillation. Heart Rhythm 5(3): 427-435.
Riehmann, M, Rhodes, PR, Ametani, MS, Sall, M and Bruskewitz, RC (1997). Preservation of bladder tissue in different solutions for varying times. Urology 50(1): 142-149.
Robbins, J (2001). KCNQ potassium channels: physiology, pathophysiology, and pharmacology. Pharmacol Ther 90(1): 1-19.
Rode, F, Svalø, J, Sheykhzade, M and Ronn, LC (2010). Functional effects of the KCNQ modulators retigabine and XE991 in the rat urinary bladder. Eur J Pharmacol 638(1-3): 121-127.
Roura-Ferrer, M, Etxebarria, A, Sole, L, Oliveras, A, Comes, N, Villarroel, A and Felipe, A (2009). Functional implications of KCNE subunit expression for the Kv7.5 (KCNQ5) channel. Cell Physiol Biochem 24(5-6): 325-334.
"53

BIBLIOGRAPHY
Rundfeldt, C (1999). Characterization of the K+ channel opening effect of the anticonvulsant retigabine in PC12 cells. Epilepsy Res 35(2): 99-107.
Rundfeldt, C and Netzer, R (2000). Investigations into the mechanism of action of the new anticonvulsant retigabine. arzneimittelforschung 50(12): 1063-1070.
Salata, JJ, Jurkiewicz, NK, Wang, J, Evans, BE, Orme, HT and Sanguinetti, MC (1998). A novel benzodiazepine that activates cardiac slow delayed rectifier K+ currents. Mol Pharmacol 54(1): 220-230.
Schleifenbaum, J, Kohn, C, Voblova, N, Dubrovska, G, Zavarirskaya, O, Gloe, T, Crean, CS, Luft, FC, Huang, Y, Schubert, R and Gollasch, M (2010). Systemic peripheral artery relaxation by KCNQ channel openers and hydrogen sulfide. J Hypertens 28(9): 1875-1882.
Schroeder, BC, Waldegger, S, Fehr, S, Bleich, M, Warth, R, Greger, R and Jentsch, TJ (2000). A constitutively open potassium channel formed by KCNQ1 and KCNE3. nature 403(6766): 196-199.
Schrøder, RL, Jespersen, T, Christophersen, P, Strøbæk, D, Jensen, BS and Olesen, S-P (2001). KCNQ4 channel activation by BMS-204352 and retigabine. Neuropharmacology 40(7): 888-898.
Selyanko, AA, Hadley, JK, Wood, IC, Abogadie, FC, Delmas, P, Buckley, NJ, London, B and Brown, DA (1999). Two types of K(+) channel subunit, Erg1 and KCNQ2/3, contribute to the M-like current in a mammalian neuronal cell. J Neurosci 19(18): 7742-7756.
Sexton, CC, Notte, SM, Maroulis, C, Dmochowski, RR, Cardozo, L, Subramanian, D and Coyne, KS (2011). Persistence and adherence in the treatment of overactive bladder syndrome with anticholinergic therapy: a systematic review of the literature. Int J Clin Pract 65(5): 567-585.
Sibley, GN (1984). A comparison of spontaneous and nerve-mediated activity in bladder muscle from man, pig and rabbit. J Physiol 354: 431-443.
Soder, RP, Parajuli, SP, Hristov, KL, Rovner, ES and Petkov, GV (2013). SK channel-selective opening by SKA-31 induces hyperpolarization and decreases contractility in human urinary bladder smooth muscle. Am J Physiol Regul Integr Comp Physiol 304(2): R155-163.
Soldovieri, MV, Miceli, F and Taglialatela, M (2011). Driving with no brakes: molecular pathophysiology of Kv7 potassium channels. Physiology (Bethesda) 26(5): 365-376.
Spitsbergen, JM, Clemow, DB, McCarty, R, Steers, WD and Tuttle, JB (1998). Neurally mediated hyperactive voiding in spontaneously hypertensive rats. Brain Res 790(1-2): 151-159.
"54

BIBLIOGRAPHY
Splawski, I, Tristani-Firouzi, M, Lehmann, MH, Sanguinetti, MC and Keating, MT (1997). Mutations in the hminK gene cause long QT syndrome and suppress IKs function. Nat Genet 17(3): 338-340.
Stewart, WF, Van Rooyen, JB, Cundiff, GW, Abrams, P, Herzog, AR, Corey, R, Hunt, TL and Wein, AJ (2003). Prevalence and burden of overactive bladder in the United States. World J Urol 20(6): 327-336.
Stott, JB, Povstyan, OV and Greenwood, IA (2014). Electrophysiological effects of G protein beta gamma subunits on Kv7.4 channels. Physiology (Bethesda), London, Uk, PCA199.
Streng, T, Christoph, T and Andersson, K-E (2004). Urodynamic Effects of the K+ Channel (Kcnq) Opener Retigabine in Freely Moving, Conscious Rats. J Urol 172(5): 2054-2058.
Strutz-Seebohm, N, Seebohm, G, Fedorenko, O, Baltaev, R, Engel, J, Knirsch, M and Lang, F (2006). Functional coassembly of KCNQ4 with KCNE-beta- subunits in Xenopus oocytes. Cell Physiol Biochem 18(1-3): 57-66.
Suh, BC, Inoue, T, Meyer, T and Hille, B (2006). Rapid chemically induced changes of PtdIns(4,5)P2 gate KCNQ ion channels. Science 314(5804): 1454-1457.
Svalø, J, Hansen, HH, Rønn, LC, Sheykhzade, M, Munro, G and Rode, F (2011). K(v) 7 Positive Modulators Reduce Detrusor Overactivity and Increase Bladder Capacity in Rats. Basic Clin Pharmacol Toxicol 110(2): 145-153.
Takeda, M, Obara, K, Mizusawa, T, Tomita, Y, Arai, K, Tsutsui, T, Hatano, A, Takahashi, K and Nomura, S (1999). Evidence for beta3-adrenoceptor subtypes in relaxation of the human urinary bladder detrusor: analysis by molecular biological and pharmacological methods. J Pharmacol Exp Ther 288(3): 1367-1373.
Tatulian, L, Delmas, P, Abogadie, FC and Brown, DA (2001). Activation of expressed KCNQ potassium currents and native neuronal M-type potassium currents by the anti-convulsant drug retigabine. J Neurosci 21(15): 5535-5545.
Trivedi, S, Potter-Lee, L, McConville, MW, Li, JH, Ohnmacht, CJ, Trainor, DA and Kau, ST (1995a). K-channel opening activity of dihydropyridine ZM244085: effect on 86Rb efflux and 3H-P1075 binding in urinary bladder smooth muscle. Res Commun Mol Pathol Pharmacol 88(2): 137-151.
Trivedi, S, Stetz, SL, Potter-Lee, L, McConville, M, Li, JH, Empfield, J, Ohnmacht, CJ, Russell, K, Brown, FJ, Trainor, DA and Kau, ST (1995b). K-channel opening activity of ZD6169 and its analogs: effect on 86Rb efflux and 3H-P1075 binding in bladder smooth muscle. Pharmacology 50(6): 388-397.
Turner, WH and Brading, AF (1997). Smooth muscle of the bladder in the normal and the diseased state: Pathophysiology, diagnosis and treatment. Pharmacology & Therapeutics 75(2): 77-110.
"55

BIBLIOGRAPHY
Uchida, H, Shishido, K, Nomiya, M and Yamaguchi, O (2005). Involvement of cyclic AMP-dependent and -independent mechanisms in the relaxation of rat detrusor muscle via beta-adrenoceptors. Eur J Pharmacol 518(2-3): 195-202.
Unoki, H, Takahashi, A, Kawaguchi, T, Hara, K, Horikoshi, M, Andersen, G, Ng, DP, Holmkvist, J, Borch-Johnsen, K, Jorgensen, T, Sandbaek, A, Lauritzen, T, Hansen, T, Nurbaya, S, Tsunoda, T, Kubo, M, Babazono, T, Hirose, H, Hayashi, M, Iwamoto, Y, Kashiwagi, A, Kaku, K, Kawamori, R, Tai, ES, Pedersen, O, Kamatani, N, Kadowaki, T, Kikkawa, R, Nakamura, Y and Maeda, S (2008). SNPs in KCNQ1 are associated with susceptibility to type 2 diabetes in East Asian and European populations. Nat Genet 40(9): 1098-1102.
Wang, H (1998). KCNQ2 and KCNQ3 Potassium Channel Subunits: Molecular Correlates of the M-Channel. Science 282(5395): 1890-1893.
Wang, Q, Curran, ME, Splawski, I, Burn, TC, Millholland, JM, VanRaay, TJ, Shen, J, Timothy, KW, Vincent, GM, de Jager, T, Schwartz, PJ, Toubin, JA, Moss, AJ, Atkinson, DL, Landes, GM, Connors, TD and Keating, MT (1996). Positional cloning of a novel potassium channel gene: KVLQT1 mutations cause cardiac arrhythmias. Nat Genet 12(1): 17-23.
Warth, R, Garcia Alzamora, M, Kim, JK, Zdebik, A, Nitschke, R, Bleich, M, Gerlach, U, Barhanin, J and Kim, SJ (2002). The role of KCNQ1/KCNE1 K(+) channels in intestine and pancreas: lessons from the KCNE1 knockout mouse. Pflugers Arch 443(5-6): 822-828.
Wen, H and Levitan, IB (2002). Calmodulin is an auxiliary subunit of KCNQ2/3 potassium channels. J Neurosci 22(18): 7991-8001.
Wickenden, AD, Krajewski, JL, London, B, Wagoner, PK, Wilson, WA, Clark, S, Roeloffs, R, McNaughton-Smith, G and Rigdon, GC (2008). N-(6-chloro-pyridin-3-yl)-3,4-difluoro-benzamide (ICA-27243): a novel, selective KCNQ2/Q3 potassium channel activator. Mol Pharmacol 73(3): 977-986.
Wickenden, AD, Yu, W, Zou, A, Jegla, T and Wagoner, PK (2000). Retigabine, a novel anti-convulsant, enhances activation of KCNQ2/Q3 potassium channels. Mol Pharmacol 58(3): 591-600.
Wuest, M, Eichhorn, B, Grimm, MO, Wirth, MP, Ravens, U and Kaumann, AJ (2009). Catecholamines relax detrusor through beta 2-adrenoceptors in mouse and beta 3-adrenoceptors in man. J Pharmacol Exp Ther 328(1): 213-222.
Yamaguchi, O, Marui, E, Kakizaki, H, Homma, Y, Igawa, Y, Takeda, M, Nishizawa, O, Gotoh, M, Yoshida, M, Yokoyama, O, Seki, N, Ikeda, Y and Ohkawa, S (2014). Phase III, randomised, double-blind, placebo-controlled study of the beta3-adrenoceptor agonist
"56

BIBLIOGRAPHY
mirabegron, 50 mg once daily, in Japanese patients with overactive bladder. BJU Int 113(6): 951-960.
Yamanishi, T, Chapple, CR, Yasuda, K, Yoshida, K and Chess-Williams, R (2002). Identification of beta-adrenoceptor subtypes in lower urinary tract of the female pig. J Urol 168(6): 2706-2710.
Yamanishi, T, Yasuda, K, Kitahara, S, Nakai, H, Yoshida, K and Iizuka, H (2006). Effects of 138-355, a beta3-adrenoceptor selective agonist, on relaxation of the human detrusor muscle in vitro. Neurourol Urodyn 25(7): 815-819.
Yang, IC, Scherz, MW, Bahinski, A, Bennett, PB and Murray, KT (2000). Stereoselective interactions of the enantiomers of chromanol 293B with human voltage-gated potassium channels. J Pharmacol Exp Ther 294(3): 955-962.
Yeung, SY and Greenwood, IA (2005). Electrophysiological and functional effects of the KCNQ channel blocker XE991 on murine portal vein smooth muscle cells. Br J Pharmacol 146(4): 585-595.
Yeung, SY, Pucovsky, V, Moffatt, JD, Saldanha, L, Schwake, M, Ohya, S and Greenwood, IA (2007). Molecular expression and pharmacological identification of a role for K(v)7 channels in murine vascular reactivity. Br J Pharmacol 151(6): 758-770.
Yeung, SY, Schwake, M, Pucovsky, V and Greenwood, I (2008). Bimodal effects of the Kv7 channel activator retigabine on vascular K+ currents. Br J Pharmacol 155(1): 62-72.
Yu, H, Wu, M, Townsend, SD, Zou, B, Long, S, Daniels, JS, McManus, OB, Li, M, Lindsley, CW and Hopkins, CR (2011). Discovery, Synthesis, and Structure Activity Relationship of a Series of N-Aryl- bicyclo[2.2.1]heptane-2-carboxamides: Characterization of ML213 as a Novel KCNQ2 and KCNQ4 Potassium Channel Opener. ACS Chem Neurosci 2(10): 572-577.
Yus-Najera, E, Santana-Castro, I and Villarroel, A (2002). The identification and characterization of a noncontinuous calmodulin-binding site in noninactivating voltage-dependent KCNQ potassium channels. J Biol Chem 277(32): 28545-28553.
Zhang, X, Chen, X, Jia, C, Geng, X, Du, X and Zhang, H (2010). Depolarization increases phosphatidylinositol (PI) 4,5-bisphosphate level and KCNQ currents through PI 4-kinase mechanisms. J Biol Chem 285(13): 9402-9409.
Zhong, XZ, Harhun, MI, Olesen, SP, Ohya, S, Moffatt, JD, Cole, WC and Greenwood, IA (2010). Participation of KCNQ (Kv7) potassium channels in myogenic control of cerebral arterial diameter. J Physiol 588(Pt 17): 3277-3293.
!!
"57

APPENDICES
Appendices
Appendices I-III can be retrieved at the respective journals.
I. Svalø J, Nordling J, Bouchelouche K, Andersson KE, Korstanje C, Bouchelouche
P (2013). The novel beta3-adrenoceptor agonist mirabegron reduces carbachol-induced
contractile activity in detrusor tissue from patients with bladder outflow obstruction
with or without detrusor overactivity. Eur. J. Pharmacol. 699: 101-5
II. Svalø J, Bille M, Parameswaran Theepakaran N, Sheykhzade M, Nordling J,
Bouchelouche P (2013). Bladder contractility is modulated by Kv7 channels in pig
detrusor. Eur. J. Pharmacol. 715: 312-20
III. Svalø J, Sheykhzade M, Nordling J, Matras C, Bouchelouche P. Functional and
molecular evidence for Kv7 channel subtypes in human detrusor from patients with and
without bladder outflow obstruction. PLoS ONE. 10:e0117350
IV. Table listing Kv7 channel modulators
!58

APPENDIX IV
Overview of the potency of positive (+) and negative (-) Kv7 channel modulators
Target Potency (µM)
Reference
Retigabine (N-(2-amino-4-[fluorobenzylamino]-phenyl) carbamic acid)
+ Kv7.2 + Kv7.2/3 + Kv7.3 + Kv7.4 + Kv7.5
2.5 1.6-3.6
0.6 1.4-5.2 N.D.
Main et al. 2000, Wickenden et al. 2000, Schrøder et al. 2001, Tatulian et al. 2001
Flupirtine (2-amino-3-(ethoxycarbonylamino)-6-(4-fluorobenzylamino)pyridine)
+ Kv7.2-7.5 N.D.
S-1 (S)-N-[1-(3-morpholin-4-yl-phenyl)-ethyl]-3-phenyl-acrylamide)
+ Kv7.2 + Kv7.3 + Kv7.4 + Kv7.5 - Kv7.1 - Kv7.1/E1
N.D. N.D. 10.4 N.D. 18 27
Bentzen et al. 2006
ML277 ((R)-N-(4-(4-methoxyphenyl)thiazol-2-yl)-1-tosylpiperidine-2-carboxamide)
+ Kv7.1 0.26 Mattmann et al. 2012
L-364373 (5-(2-fluorophenyl)-1,3-dihydro-3-(1H-indol-3-ylmethyl)-1-methyl-2H-1,4-benzodiazepin-2-one)
+ Kv7.1 N.D.
ML213 (N-mesitylbicyclo[2,2,1]heptane-2-carboxamide)
+ Kv7.2 + Kv7.2/3 + Kv7.4 + Kv7.4/7.5 + Kv7.5
0.33 0.37
0.51-0.8 1.1 0.7
Yu et al. 2011, Brueggemann et al. 2014
Meclofenamic acid (2-[(2,6-dichloro-3-methylphenyl) amino] benzoic acid)
+ Kv7.2/7.3 25 Peretz et al. 2005
ICA-27243 (N-(6-chloropyridin-3-yl)-3,4-difluorobenzamide)
+ Kv7.2/7.3 + Kv7.4 + Kv7.3/5 - Kv7.1/E1
0.4 9.7 >10
40 (#)
Blom et al. 2010, Wickenden et al. 2008
SMB-1 ((E)-N-[(S)-1-(4-cyclopropylmethyl-3,4-dihydro-2H-1,4-benzoxazin-6-yl)-ethyl]-3-(2-fluoro-phenyl)-N-methyl-acrylamide)
+ Kv7.4 - Kv7.2
5.7 7.4
Blom et al. 2014
XE991 (10,10-bis(4-pyridinylmethyl)-9(10H)-anthracenone dihydrochloride)
- Kv7.1 - Kv7.2 - Kv7.2/3 - Kv7.3/4 - Kv7.4 - Kv7.5
0.8 0.7-1.5 0.6-0.9
1.4 1.2 65
Robbins 2001, Schrøder et al. 2001
Overview of the potency of positive (+) and negative (-) Kv7 channel modulators

APPENDIX IV
Linopirdine (1,3-dihydro-1-phenyl-3,3-bis(4-pyridinylmethyl)-2H-indol-2-one dihydrochloride)
- Kv7.1 - Kv7.2 - Kv7.2/3 - Kv7.3 - Kv7.3/4 - Kv7.3/5 - Kv7.4 - Kv7.5
8.1 3.4-4.8 2.7-10
4.8 1.4 15 1.7 16
Robbins 2001, Schrøder et al. 2001
Chromanol 293B N-[(3R,4S)-6-cyano-3,4-dihydro-3-hydroxy-2,2-dimethyl-2H-1-benzopyran-4-yl]-N-methylethanesulfonamide
- Kv7.1 - Kv7.1/E1 - Kv7.1/E3
65.4 1.36-15.1
0.54
Yang et al. 2000, Bett et al. 2006
Potencies (EC50/IC50) values were determined by electrophysiological recordings except othervise noted, whereas (86)Rb efflux studies are denoted by (#). Voltage-gated potassium channels and not determined values are denoted by Kv and N.D., respectively.
Overview of the potency of positive (+) and negative (-) Kv7 channel modulators
Target Potency (µM)
Reference
Overview of the potency of positive (+) and negative (-) Kv7 channel modulators
Related Documents











