UNIVERSITY OF AMSTERDAM – BRAIN AND COGNITIVE SCIENCE – COGNITIVE TRACK The roles of serotonin and dopamine in reactive and proactive aggression A literature thesis in partial fulfilment of the requirements for the degree of Master of Science Jonathan Krikeb, BSc, 10065180 12 June 2015 Supervisor: Co-assessor: dhr. prof. dr. C.K.W. de Dreu dhr. dr. M.P. Lebreton

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

UNIVERSITY OF AMSTERDAM – BRAIN AND COGNITIVE SCIENCE – COGNITIVE TRACK
The roles of serotonin anddopamine in reactive and
proactive aggressionA literature thesis in partial fulfilment of the
requirements for the degree of Master ofScience
Jonathan Krikeb, BSc, 1006518012 June 2015
Supervisor: Co-assessor:
dhr. prof. dr. C.K.W. de Dreu dhr. dr. M.P. Lebreton

Krikeb, J., Serotonin and dopamine in predator-prey aggression
Abstract:
Aggression is often linked to violence but this is not a necessary connection. Aggression could
also be motivating choices for economic-decision making. The question of what leads to aggression
is what this paper will address as it discusses the bi-modal classification of aggression: proactive and
reactive. These two classes will be linked to a new predator-prey research paradigm that separates
the greed and its proactive tendencies, from the fear and its reactive actions. This, as well as a few
other economic games, will be linked to the wide scope of research into aggressive violent
behaviour, that is mostly based on clinical cases, as well as decision-making research that is founded
on the idea that focuses on impulsive behaviour as it has been linked to aggression in the past. These
past findings have also found correlations between serotonin hypoactivity, and also dopamine
hyperactivity, in cases of irregular aggressive behaviour. We will attempt to establish how activities
of the serotonergic and the dopaminergic circuitries parallel aggression in predator and prey type of
interactions.
1

Krikeb, J., Serotonin and dopamine in predator-prey aggression
Table of Contents
1. Introduction.......................................................................................................................................4
1.1. Aggressive behaviour.................................................................................................................4
1.2. Bi-modal classification of aggression.........................................................................................6
1.3. Aggression networks..................................................................................................................8
2. Experimental paradigms..................................................................................................................10
2.1. Aggressive behaviour in economic games...............................................................................10
2.2. The predator prey game..........................................................................................................11
2.2.1. Greed and calculated aggression in the predator............................................................14
2.2.2. PFC and goal-oriented behaviour....................................................................................14
2.2.3. Fear and reactive aggression in the prey.........................................................................16
2.2.4. Amygdala and fearful behaviour......................................................................................16
2.3. Aggressive behaviour in current research on neurotransmitters............................................18
2.4. Impulsive behaviour in experiments........................................................................................20
2.5. Experimental paradigms of impulsivity....................................................................................21
2.5.1. 5-CSRT..............................................................................................................................22
2.5.2. Go/no-go.........................................................................................................................22
2.5.3. SSRT.................................................................................................................................23
2.5.4. Reversal learning.............................................................................................................23
2.5.5. Delayed reward...............................................................................................................23
3. Serotonin.........................................................................................................................................23
3.1.1. Molecule..........................................................................................................................24
3.1.2. Different receptors in different brain regions..................................................................25
3.1.3. Tryptophan depletion......................................................................................................27
3.1.4. Tryptophan supplementation..........................................................................................28
2

Krikeb, J., Serotonin and dopamine in predator-prey aggression
3.1.5. Serotonin knockouts........................................................................................................28
3.1.6. Specific agonists/antagonists and neurotoxins................................................................29
3.2. Serotonin and impulsivity........................................................................................................29
3.3. Link between serotonin research to predator and prey behaviours........................................30
4. Dopamine........................................................................................................................................35
4.1.1. Molecule..........................................................................................................................35
4.1.2. Different receptors in different brain regions..................................................................35
4.1.3. Parkinson’s and L-dopa....................................................................................................37
4.1.4. Other manipulations to dopamine..................................................................................37
4.2. Dopamine and impulsivity.......................................................................................................38
4.3. Link between dopamine research to predator and prey behaviours.......................................38
5. Interactions between serotonin and dopamine...............................................................................39
6. Discussion........................................................................................................................................40
3

Krikeb, J., Serotonin and dopamine in predator-prey aggression
1. IntroductionAn attack to gain more resources by someone who has an abundance of them does not have the
same motivation as the defensive reaction of the prey, in this same scenario, that defends its limited
resources. In this sort of predator against prey interaction, two types of aggression come into play:
proactive and reactive. Reactive aggression has been widely studied, often in context of impulsive
aggression, while the literature on proactive aggression is more scarce. In light of the research into
the motives behind these behaviours, and how this is reflected in terms of neurotransmitters, this
paper will follow past studies and look at how the key neurotransmitters serotonin and dopamine
interact in these two different types of aggression, namely predator and prey.
1.1. Aggressive behaviour
Aggression traditionally requires a conflict between at least two parties that may compete for
the same object, physical or otherwise (Nelson & Trainor, 2007).Since aggression and violence often
go hand-in-hand, the results of aggressive behaviour often lead to damage, which is often physical,
and therefore aggression is dangerous and not always the best choice in conflict circumstances. It
does have an evolutionary role in food, or mate, acquisition, or protection, as well as inner
motivations such as fear, greed, anger or even pleasure. When these motives lead to aggressive acts
that hurt, or injure others, then we consider aggression as unaccepted in our current human society
– war or criminal acts such as robbery or battery (de Almeida, Ferrari, Parmigiani, & Miczek, 2005).
There is a wide selection of literature available on aggression, much of it is composed of
psychological research focusing on clinical and criminal cases such as: workplace aggression (Hills &
Joyce, 2013; Piquero, Piquero, Craig, & Clipper, 2013; S. F. Smith & Lilienfeld, 2013), domestic
aggression (George et al., 2001; Soler, Vinayak, & Quadagno, 2000), alcohol and drug related (Anholt
4

Krikeb, J., Serotonin and dopamine in predator-prey aggression
& Mackay, 2012; Badawy, 2003; de Almeida et al., 2005; George et al., 2001; Gowin, Swann, Moeller,
& Lane, 2010; Skara et al., 2008), and arson (Linnoila, Virkkunen, & Scheinin, 1983). All this research
shows how ingrained in human society aggression is and how destructive it could get, thus
necessitating a deeper understanding of the motives behind it.
In order to better understand aggression and its motives, different classification systems were
established. In this paper, as the introduction suggests, we will focus on a binary classification
system. Two other systems were suggested in previous studies as well. First, as described both by
Siegel and Victoroff (2009) and Umukoro, Aladeokin, and Eduviere (2013) where they divide
aggression into seven separate motivations it may originate from: fear -induced, maternal, irritable,
inter-male, sex-related, predatory, and territorial. However, in the case of humans, these are not as
clear-cut cases as they are in the animal world.
A third classification is offered by separating the motives into four categories: stress and fear-
induced, anger and frustration-induced, instrumental offence, and pleasure motivated. However, in
both cases of the alternative classifications, we could collapse them into a bimodal classification
system that is easier to apply for humans, where the study of aggression is complex as it is. An
example used by Siegel and Victoroff (2009) is a gang fight where the mix of planning, inter-male
dominance, and emotions of anger or fear interact to spark the fighting. In such a bimodal
classification, one side would be predatory, goal-oriented, calculated, instrumental, proactive,
premeditated aggression. This would be contrasted with a reactive, impulsive, protective, defensive,
hostile aggression (Malone et al., 1998; McEllistrem, 2004; Nelson & Trainor, 2007; Siegel &
Victoroff, 2009; Umukoro et al., 2013; Weinshenker & Siegel, 2002). Whereas the former has very
little literature looking into it, the latter sort is more related to pathological aggression, and often
associated in research with reduced levels of the neurotransmitter 5-hydroxytryptamine (5-HT or
5

Krikeb, J., Serotonin and dopamine in predator-prey aggression
serotonin) (Anholt & Mackay, 2012; Badawy, 2003; Berman, Tracy, & Coccaro, 1997; Booij et al.,
2010; Brown et al., 1982; Brown, Goodwin, & Ballenger, 1979; Crockett, Clark, Lieberman, Tabibnia,
& Robbins, 2010; Crockett, Clark, & Robbins, 2009; Crockett, Clark, Tabibnia, Lieberman, & Robbins,
2008; Daw, Kakade, & Dayan, 2002; de Boer & Koolhaas, 2005; Linnoila et al., 1983; Perez, 2012;
Wetzler, Kahn, Asnis, Korn, & van Praag, 1991). These studies relate many pathologies related to
impulse control including drug and alcohol addiction, pyromancy, suicidal tendencies, as well as
repeated violent crimes, to observed low levels of serotonin. While it is important to notice that
these studies find correlation, and not causation, we will examine throughout this paper the role of
serotonin in aggression and impulsive behaviour.
Fig. 1. Bi-modal classification of aggression and where neurotransmitter research has been involved so far.
1.2. Bi-modal classification of aggression
As suggested above, this paper will focus on a bimodal classification of aggression, namely:
predatory, and reactive. This bimodal classification is discussed in in depth in a few articles
6

Krikeb, J., Serotonin and dopamine in predator-prey aggression
(Chichinadze, Chichinadze, & Lazarashvili, 2011; Malone et al., 1998; McEllistrem, 2004; Siegel &
Victoroff, 2009; Umukoro et al., 2013; Weinshenker & Siegel, 2002). Reactive aggression is closely
related in the literature to affective defence, or aggression (McEllistrem, 2004; Weinshenker &
Siegel, 2002). This type of aggression is much more studied in comparison to the other, predatory
type (Siegel & Victoroff, 2009). In animal models, it is easily classified since it is expected in cases of
invasion to personal space or ingression on food reserves. This aggression is also easily measured in
terms of strong sympathetic nervous system activation (Weinshenker & Siegel, 2002). This suggests
that this type of aggression is instinctive, and therefore impulsive – there is no consideration of the
long-term results, only immediate elimination of the current threat. Implicitly this links this
aggression to emotions: anger, anxiety, and fear, and thus to the limbic system. The impulsive aspect
will be discussed further since it is a core concept in criminal and clinical studies of aggression.
The other type of aggression of interest in this paper is offensive, predatory aggression. Amongst
animals this is usually the manner in which a predatory animal hunts and consumes a prey animal.
However, our focus should be, in order to compare to human cases, on intraspecies aggression such
as climbing up the social hierarchy amongst groups of monkeys where one aims to dominate
(Chichinadze et al., 2011). A human example could be, for instance, a burglary – ingression into
another’s property in order to obtain gain illegal possession of property. On a larger scale this could
be the preying of a strong nation on a weaker, less resourceful one. This type is under-studied and
while it may interact with the reactive aggression, does not rely on the same mechanisms
(McEllistrem, 2004; Umukoro et al., 2013; Weinshenker & Siegel, 2002). For starters, there is a lack
of sympathetic arousal in the predatory aggressors. Additionally, feelings, if they play a role at all,
lead to pleasure or satisfaction, as opposed to fear or anger involved in defensive acts. In this
manner, it is possible to observe the most striking difference between the types of aggression:
reactive defence is unmeasured, it lashes out (Weinshenker & Siegel, 2002). In economic terms, the
7

Krikeb, J., Serotonin and dopamine in predator-prey aggression
investment would be un-proportional to the risk. In contrast, predatory, calculated, aggression, as
the latter term suggests, involves very thought-out allocation of resources – both manner and
magnitude become significant, whereas in defence they are not.
Weinshenker and Siegel (2002) discuss the advantage, for the sake of research, of distinguishing
the psychopaths. These individuals, who have difficulty relating to emotions, comprise a large part of
prison populations and their motivations are more alike to predatory in nature, as opposed to
impulsive violent offenders. Interestingly, instrumental aggression is also linked to dominance
(Chichinadze et al., 2011). The brain mechanisms involved in planning and executive function, are
the ones that would also be instrumental in predatory action. This coincides with data linking
advantage of serotonin enhanced performance leading to domination among monkeys whereas
depletion of the neurotransmitter, associated with impulsivity, does not have the same effect
(Berman et al., 1997).
In the following sections we will follow the research paradigms implemented in impulsive
behaviour and aggression research in economic decision-making. This will lead to the predator-prey
paradigm, devised by de Dreu, Scholte, van Winden, and Ridderinkhof (2014), that establishes, using
an asymmetrical game, the roles of proactive and reactive aggression.
1.3. Aggression networks
Aggressive behaviour is closely linked to social behaviour, both because of the context in which
aggression takes place, and also due to the brain network involved in aggressive behaviour. This
circuitry involves: the amygdala, and the rest of the limbic system; the periaqueductal gray (PAG);
the hypothalamus; and sections of the prefrontal cortex (PFC) (Nelson & Trainor, 2007). These
regions are also innervated by the raphe nuclei where serotonin is produced in the brain
(Martinowich & Lu, 2008). The connection with serotonin has been also observed in the clinic where
8
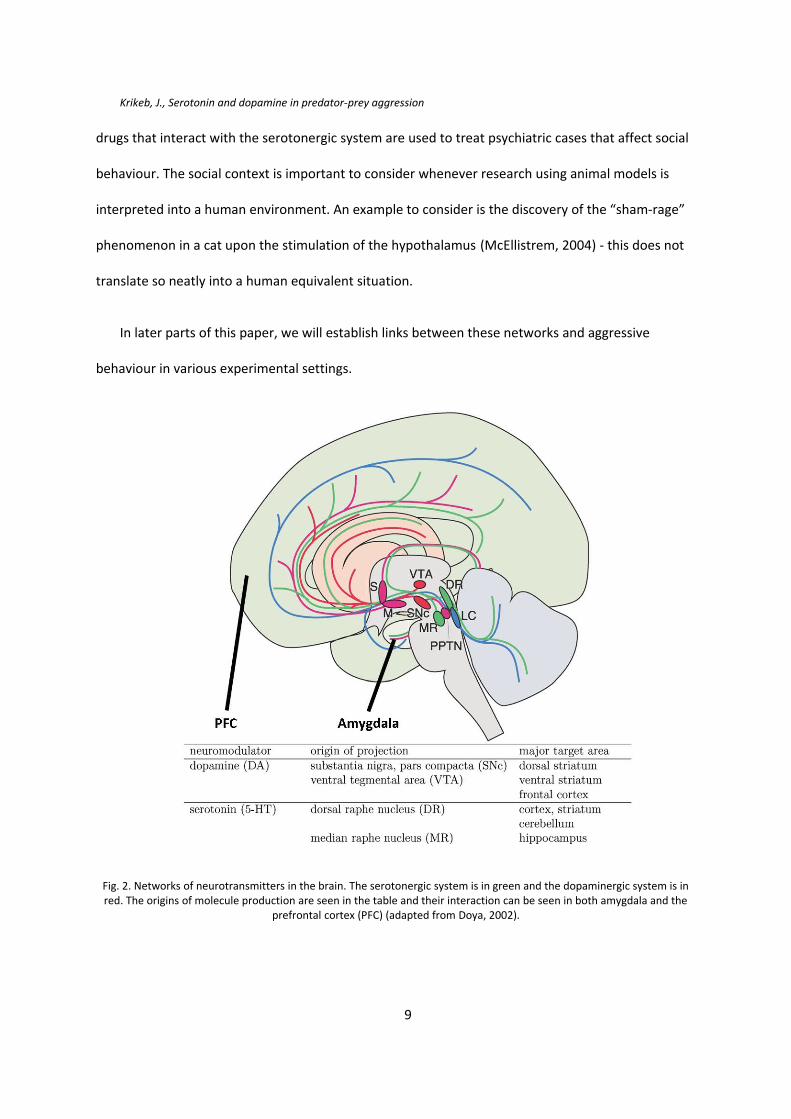
Krikeb, J., Serotonin and dopamine in predator-prey aggression
drugs that interact with the serotonergic system are used to treat psychiatric cases that affect social
behaviour. The social context is important to consider whenever research using animal models is
interpreted into a human environment. An example to consider is the discovery of the “sham-rage”
phenomenon in a cat upon the stimulation of the hypothalamus (McEllistrem, 2004) - this does not
translate so neatly into a human equivalent situation.
In later parts of this paper, we will establish links between these networks and aggressive
behaviour in various experimental settings.
Fig. 2. Networks of neurotransmitters in the brain. The serotonergic system is in green and the dopaminergic system is inred. The origins of molecule production are seen in the table and their interaction can be seen in both amygdala and the
prefrontal cortex (PFC) (adapted from Doya, 2002).
9

Krikeb, J., Serotonin and dopamine in predator-prey aggression
2. Experimental paradigms
2.1. Aggressive behaviour in economic games
In economic settings, an aggressive investor puts more money into play and takes bigger risks
(Afza & Nazir, 2007; Nazir & Afza, 2009). Investments of this nature seem more impulsive. However,
aggressive behaviour could also translate into offensive behaviour; initiating purchases and trying to
increase assets. This behaviour is a type of calculated aggression. This highlights the difficulty in
separating the two types of aggression in economic decision-making.
In economic experiments, using different games to model decision-making, defection or
punishment is often conceptualised as aggression (Crockett, 2009). For instance, in the prisoner’s
dilemma game, the peaceful solution would be cooperation where both sides gain together
maximally, however, the aggressive solution, where both defect, is the Nash Equilibrium. These two
behaviour choices alternate depending on the type and frequency of interaction between the parties
(Kassinove, Roth, Owens, & Fuller, 2002; Martin, Juvina, Lebiere, & Gonzalez, 2013). This applies
similarly to the ultimatum game. In this game, one side decides how to share an initial sum and the
other side can decide whether to reject the offer, thereby leaving both participants with nothing, or
accept it, and share as agreed. If one is aggressive, he would reject an unfair (or perhaps even fair)
offer. If the player is cooperative, he would accept the offer, as long as he gets something (Mehta &
Beer, 2010). Rejecting an offer in the ultimatum game can be perceived as a type of punishment.
This however, was challenged by Crockett et al. (2008), as we will further discuss later. Finally, it
would be of the biggest interest to separate instrumental (predatory) aggression, of the form where
one side wishes to break the status-quo and increase its winnings, from reactive (prey) aggression,
where one side fends off the predator, at cost to itself, in order to maintain the current equilibrium.
We can relate both of these behaviours to market, and social, behaviours such as a buy-out of a
10

Krikeb, J., Serotonin and dopamine in predator-prey aggression
company or going bankrupt so as not to be bought-off. We have found two experimental paradigms
that separate these different aggressive behaviours and elucidate their underlying mechanisms
(Crockett, Clark, and Robbins, 2009; and de Dreu, Scholte, van Winden, and Ridderinkhof, 2014).
2.2. The predator prey game
Ideally we would like a game that creates a distinction between the two groups of aggressors:
goal-oriented, predatory aggressors, versus the prey-like, reactive (defensive) aggressors. This in
addition to manipulations of subjects in terms of neurotransmitters and stress, as well as tasks
involving impulse control and aggression questionnaires; all of which will be addressed in a following
section.
We need to first question the motives for being aggressive as either predator, or prey. We will
start with the prey. We can look at an individual, or a group, as belonging to a prey category when
they wish to maintain a certain status-quo – wishing to keep a job, or ownership of a company or a
piece of land. Therefore, for someone in that situation to turn to aggression, to attack or invest
money and resources, implies that they are compelled by fear of someone altering the current state
of things. In lab conditions there are various games to notice this reaction. Looking at a prisoners’
dilemma game as an example, this fear of defection of the other party could motivate one to defect
as well. This is a reactive defence behaviour. In an alternative version of the prisoners’ dilemma, a
withdrawal option is added to the game (Insko, Schopler, Hoyle, Dardis, & Graetz, 1990). This option
can distinguish between two fear reactions: those who turn to impulsivity and defect, or those who
are calculated, maintain their calm, and withdraw.
11

Krikeb, J., Serotonin and dopamine in predator-prey aggression
Table 1. Prisoner's dilemma standard payoff matrix (adapted from Wood et al., 2006).
Table 2. Prisoner's dilemma alternative payoff matrix (adapted from Insko et al., 1990).
In contrast, the predator, a greedy, goal-driven, individual or group, would wish to revisit the
current situation. This party would like to earn more than it currently does, or own another’s land or
property. It is important when comparing to animal models to consider only intraspecies preying, as
it is in humans. For that purpose of increasing winnings the predator would invest, and risk, its
resources. Therefore seemingly it is greed that motivates the predator in its choices when it decides
to aggress against a docile counterpart. Similar to the case described for the prey, the predator
would also defect in the prisoners’ dilemma game. Unlike the prey, the predator would do so in the
hope that the other party cooperates, and thus the predator will maximise its personal gain.
The two psychological motives have been taken apart in different paradigms in order to study
only fear, or only greed, and what decisions they would lead to. The chicken game uses a similar
payoff scheme to the prisoners’ dilemma, only in this scenario a tie is a loss thus only defection
would lead to a win and fear is dominant (Bornstein & Gilula, 2003). The assurance game, in
contrast, encourages cooperation since that would lead to the biggest payoff to both parties and
therefore is meant to exemplify only greed (Bornstein & Gilula, 2003).
12
A\B Coop DefectCoop 20\20 0\30Defect 30\0 10\10
A\B Coop Withdraw DefectCoop 57\57 48\48 33\66
Withdraw 48\48 48\48 48\48Defect 66\33 48\48 39\39

Krikeb, J., Serotonin and dopamine in predator-prey aggression
Table 3. The assurance game group payoff matrix with the independent variable being the number of investors inevery group (adapted from Bornstein & Gilula, 2003).
Table 4. The chicken game group payoff matrix with the independent variable being the number of investors in everygroup (adapted from Bornstein & Gilula, 2003).
To contrast these games, a predator-prey game captured both motives in an asymmetrical game
and thus allowed both types of aggressive behaviours to come into play (De Dreu et al., 2014). In this
game, one party, the predator, wins by investing more than the prey, thereby taking all the prey’s
leftover sum after investments. During the same investment, the other party, the prey, can only
keep their leftover sum if they amassed a sufficient defence – they invested equal to or more than
the predator (De Dreu et al., 2014). This design, in its intra-group version (De Dreu, Giffin, Krikeb,
Prochazkova, & Columbus, 2015), is easy to equate to warfare between nations over territory or
resources. One side, with predatory motives, is aggressing by taking action and investing in order to
win what resources the prey party has. In reaction to this aggression, the other side defends itself,
investing of its own resources to protect the remaining resources. As we will discuss later, this
defence could be entirely impulsive and therefore likely to be exaggerated, or it could be also
calculated, with an added long-term effect view. Accordingly, brain regions involved in greed and
fear are different and so it has been observed that the two parties have different brain activations:
13
A\B 0 1 2 30 135\135 45\120 45\105 45\901 120\45 120\120 30\105 30\902 105\45 105\30 105\105 15\903 90\45 90\30 90\15 90\90
A\B 0 1 2 30 45\45 45\120 45\105 45\901 120\45 30\30 30\105 30\902 105\45 105\30 15\15 15\903 90\45 90\30 90\15 0\0

Krikeb, J., Serotonin and dopamine in predator-prey aggression
the amygdala and superior frontal gyrus (anatomically part of the PFC) activations in the predators
were dampened by oxytocin whereas the prey’s amygdala was active regardless of condition (De
Dreu et al., 2014). Following this, we would expect a different pattern of neurotransmitter activity to
support these activations.
2.2.1. Greed and calculated aggression in the predator
In a predator-prey game, any predator that plays the game, instead of simply keeping their initial
endowment, becomes an aggressor. It is this type of aggression that is under-studied and we would
like to better highlight what sort of neural mechanism is potentially motivating it, as opposed to the
impulsive aggression that is under the main study focus in relation to reduced levels of 5-HT (de
Almeida et al., 2005; Takahashi, Quadros, de Almeida, & Miczek, 2011). In a cat, predatory behaviour
has been initiated by stimulation of the perifornical lateral hypothalamus, the ventral part of the
PAG, and the ventral-tegmental area (VTA) where dopamine is produced (Siegel & Victoroff, 2009).
The goal of this aggression, being something with a benefit past the act itself, is in contrast to
the reactive defence which lacks foresight. As Siegel and Victoroff (2009) point out, this planning
leads to the base assumption that the cerebral cortex is involved in the predatory aggression,
whereas it is not necessary for prey defence.
2.2.2. PFC and goal-oriented behaviour
To go further in depth into the decision-making of the predator, we need to examine some brain
regions that are associated with planning. First, the prefrontal cortex (PFC) is mainly associated with
top-down cognitive function, goal-oriented behaviour, temporal control, and attention control
(Agnoli & Carli, 2012; Narayanan, Land, Solder, Deisseroth, & DiLeone, 2012; Winstanley, Theobald,
Dalley, Cardinal, & Robbins, 2006). It is therefore connected to impulse-control and conditions
involving its control such as suicidal tendencies, attention deficit hyperactivity disorder, and
14

Krikeb, J., Serotonin and dopamine in predator-prey aggression
obsessive compulsive disorder (OCD). The PFC also has projections from the thalamus via the basal
ganglia, where dopamine is a key neurotransmitter (Agnoli & Carli, 2012). This aspect has been
studied using the 5 choice serial reaction time task (5-CSRT) with rats where it was shown that a
bilateral lesion of the medial PFC or dorsomedial (including the superior frontal gyrus) lead to
increased impulsivity. The aspect of motor inhibition, associated with action planning, was studied
by Rubia et al. (2005) using a go/no-go task. In their paper, they found decreased activation in the
right orbital and the inferior PFC following tryptophan depletion in humans using functional
magnetic resonance imaging (fMRI). Their results also indicate no real change in mood nor any
actual effect on the inhibitory control during the task (Rubia et al., 2005). Impulse-control is also
related to loss-aversion, that has also been a proposed task of the PFC (Murphy et al., 2009).
Further properties of the PFC we would need to inspect are the type of neurons to be found and
which neurotransmitters activate them. Pyramidal neurons, as well as GABAergic ones, in the medial
PFC (mPFC) express 5-HT1A receptors. While serotonin endogenously inhibits mPFC pyramidal
excitation, systemic 5-HT agonists administered to rats tend to excite the VTA projecting cells (Lladó-
Pelfort, Santana, Ghisi, Artigas, & Celada, 2012). The VTA, in the midbrain, also projects back to the
PFC using dopamine as input (Narayanan et al., 2012). Both D1 and D2 receptors are expressed in the
PFC. Narayanan et al. (2012) showed that D1 disruption interferes with temporal control while its
stimulation enhances efficiency in a fixed-interval timing task. These dopaminergic neurons could be
innervated by serotonergic projections coming from the raphe nuclei to both VTA and PFC where in
addition to 5-HT1A, also 5-HT2A receptors are present (Pehek, Nocjar, Roth, Byrd, & Mabrouk, 2006).
Pehek et al. (2006) showed that 5-HT2A blocking in the PFC results in dopamine blocking thus indeed
linking the dopaminergic to the serotonergic system. Both these neurotransmitter systems will come
into further focus later on and it is important that it first understood that they play a significant role
in the PFC’s activity.
15

Krikeb, J., Serotonin and dopamine in predator-prey aggression
2.2.3. Fear and reactive aggression in the prey
A prey in the game, similar to a mother defending her offspring, would resort to an affective
defence behaviour (Siegel & Victoroff, 2009). This defence behaviour is observed despite the
anonymity of the players (De Dreu et al., 2015). This fear of danger leads to a reaction whose
purpose is eliminating the threat. It is for this reason that this action, often aggressive, is impulsive;
it does not require any calculation beyond the threat and therefore likely to be out of proportion for
the assurance of its success. Additionally, strong impulses can induce fear (Apter et al., 1990),
showing us that also in this mechanism there may be a feedback loop. The fear reaction is strongly
based in the limbic system (Post et al., 1998; Yoon, Fitzgerald, Angstadt, McCarron, & Phan, 2007).
Moreover, research into defensive rage found that by direct stimulation of the medial hypothalamus
or the dorsolateral region of the PAG, this defensive reaction could be initiated (Nelson & Trainor,
2007; Siegel & Victoroff, 2009).
In terms of neurotransmitters, impulsive behaviour is mainly linked to serotonergic hypoactivity
(de Almeida et al., 2005; McEllistrem, 2004; Siegel & Victoroff, 2009; Takahashi et al., 2011;
Weinshenker & Siegel, 2002) as well as irregularities in the dopaminergic system (Rogers, 2011; E. S.
Smith, Geissler, Schallert, & Lee, 2013). It is important to realise that the reactive aggression, largely
based on sympathetic activation, could be accompanied by calculated aggression, which would
mitigate the impulsive reaction and allow planning to take place beyond the immediate effect of
removing the threat (McEllistrem, 2004; Weinshenker & Siegel, 2002). This could be compared to a
territorial war where the defender has realised that conquest of some of the attacker’s territory
would be beneficial.
2.2.4. Amygdala and fearful behaviour
16

Krikeb, J., Serotonin and dopamine in predator-prey aggression
The limbic system in general, and the amygdala specifically as part of it, include the brain
structures classically associated with emotions (Post et al., 1998; Yoon et al., 2007). This system is
also believed to be a part that belongs to the early stages of brain evolution. Of highest relevance to
this paper, it is associated with fear and anxiety (De Dreu, 2012; Saha, Gamboa-Esteves, & Batten,
2010). Irregular activation of the amygdala is associated with many anxiety disorders, such as PTSD
(Nardo et al., 2010; Pagani et al., 2012) and different phobias (see: Caseras et al., 2010; Klumpp,
Angstadt, Nathan, & Phan, 2010; Yoon et al., 2007). Crucially for this paper is its relation to decisions
instructed by fear as the dominant emotion. As de Dreu and colleagues (2014) found in their study
using the predator-prey game, the amygdala was more strongly activated among the prey,
contrasted with the predator, when making their investment decisions. This, in addition to the faster
reaction time, indicates the decision was more instinctive.
At this point, we need to explore the projections of the amygdala in order to understand what
the roles of dopamine and serotonin, as neurotransmitters in that region, may be. First, the
amygdala, as part of the limbic system, projects to the hypothalamus, and the PAG (Siegel &
Victoroff, 2009). This links the amygdala to the aggressions that we know could be initiated by the
hypothalamus.
Serotonin, produced in the raphe nuclei, influences the amygdala through many projections
from the dorsal raphe nuclei. Past studies also showed that 5-HT1A and 5-HT1B receptors are
associated with anxiety and depression (Saha et al., 2010). Saha et al. (2010) further elaborated on
this by showing the large number of neurons marked for 5-HT1B receptors and relatively low number
with 5-HT1A receptors in the amygdala. This will become more significant later when we discuss the
receptors’ separate roles.
17

Krikeb, J., Serotonin and dopamine in predator-prey aggression
In addition to serotonergic function, also dopamine is instrumental in fear response. There are
many efferent fibres to the amygdala from the VTA (Rezayof, Hosseini, & Zarrindast, 2009) as well as
bidirectional interaction with the substantia-nigra (E. S. Smith et al., 2013). These two areas are
both part of the dopamine-dominant reward-system. The effects of dopamine are quite substantial
on the amygdala. For instance, dopamine depletion prevents memory formation related to fear
through amygdala function, whereas specific restoration of dopamine to the VTA –amygdala
pathway reverses the effect (Li, Dabrowska, Hazra, & Rainnie, 2011). Specifically D1, and not D2,
receptors are crucial for this process. D1 receptor antagonist SCH23390 has been shown to eliminate
learning entirely by blocking the long-term potentiation (LTP) process from occurring in
glutamatergic neurons (Li et al., 2011). Also the role of dopamine will be further explored later in
this paper.
2.3. Aggressive behaviour in current research on neurotransmitters
While this paper is focused on the decisions that derive from both reactive, as well as predatory
aggression, there is little research that focuses on the neurotransmitters that are involved in both of
these decisions in humans. In several studies, based mostly on the ultimatum game, the type of
aggression that matches punishment was correlated to serotonin levels using a tryptophan depletion
experiment (Crockett et al., 2010, 2009, 2008; Crockett, 2009). In other research, aggressive
behaviour is measured in other ways. The lifetime history of aggression scale is used by health
workers to establish an individual’s trait aggression based on interviews and clinical history (Nelson
& Trainor, 2007). Better suited to this paper is the aggression questionnaire the was developed by
Vitiello, Behar, Hunt, Stoff, and Ricciuti (1990) that separates the questions to predatory and
affective classifications. In their unique study they found that almost every subject had a mixture of
the two characteristics. The more predatory individuals tended to have a higher IQ score, while the
18

Krikeb, J., Serotonin and dopamine in predator-prey aggression
reactive children had a higher prevalence of schizophrenia. Malone et al. (1998) have used this same
questionnaire, as well as the Overt Aggression Scale and the Global Clinical Judgements Scale and
had similar findings, yet not with the same significance.
Alternatively, the choice of violent offenders as subjects eliminates the need for objectivity
amongst the subjects (Linnoila et al., 1983). This research also led to the idea that it is not aggression
that is indicated by lower serotonin levels but impulsivity. This has been the dominant dogma for a
long period and has been the focus of many studies (Anholt & Mackay, 2012; Badawy, 2003; Berman
et al., 1997; Booij et al., 2010; Brown et al., 1982, 1979; Crockett et al., 2010, 2009, 2008; Daw et al.,
2002; de Boer & Koolhaas, 2005; Linnoila et al., 1983; Perez, 2012; Wetzler et al., 1991). Another
major data source for the 5-HT impulsivity theory comes from suicide cases; both from autopsies,
and from CSF of failed attempts. Low levels of serotonin, or its metabolite, 5-HIAA, in the latter case,
have been discovered in past studies (Dalley & Roiser, 2012). People who attempted suicide also
testify through questionnaires on possessing more impulsive tendencies.
In animal models there are different paradigms. Research focusing on the hypothalamus and the
“sham rage” phenomenon in cats was pioneering the way for future research on violence and
aggression (McEllistrem, 2004; Weinshenker & Siegel, 2002). Studies involved direct stimulation of
brain areas believed to be involved in aggression, introduction of a trespasser, or inter-male violence
in competition for dominance or a female. In these studies they either manipulate or measure levels
of different neurotransmitters. These are mostly measures of violence, as well as being in lab
settings. This conditions make it difficult to translate to human environment (Nelson & Trainor,
2007).
Also studied, albeit with a little less focus, are the increased levels of dopamine in aggression
(Boureau & Dayan, 2011; Daw et al., 2002; de Almeida et al., 2005; Seo, Patrick, & Kennealy, 2008;
19

Krikeb, J., Serotonin and dopamine in predator-prey aggression
Vukhac, Sankoorikal, & Wang, 2001) and norepinephrine (NE, also called noradrenaline) (Anholt &
Mackay, 2012; Chichinadze et al., 2011; Higley et al., 1996; Perez, 2012), and the effects of gamma-
Aminobutyric acid (GABA) (Anholt & Mackay, 2012; de Almeida et al., 2005; Gowin et al., 2010; Seo
et al., 2008). However, as previously written, this paper will restrict itself to serotonin and dopamine.
2.4. Impulsive behaviour in experiments
In a large part of available research on aggression, the focus is on criminal and clinical cases, and
not on aggressive choices. These cases come under the impulsive sort of aggression in a large part of
the cases (exception of psychopaths who show no emotion and therefore seem to be more
predatory (Perez, 2012)). For instance, Linnoila, Virkkunen, and Scheinin (1983) found a link between
serotonin and impulsive aggression amongst violent criminals. They went on to suggest that what
serotonin hypoactivity indicates is impulsive behaviour, and not aggression in general.
Dalley et al. (2011) define impulsive behaviour as “the tendency to act prematurely without
foresight”. They also clearly distinguish between impulsive and compulsive behaviour, despite the
oftentimes seen confusion of the terms. Yet since their definition for compulsive behaviour focuses
on the undesired consequences of the actions (p. 680), we could bundle both behaviours as having
unforeseen (negative) results. Therefore, we would follow from studies that examine impulsivity to
conclusions concerning rash decisions that lead to negative results.
A majority of the clinical data on human impulsive behaviour comes from suicide cases, a case of
extreme impulsivity (Dalley & Roiser, 2012). This link is also explored in many other studies (Crockett
et al., 2010; Dalley, Everitt, & Robbins, 2011; Higley et al., 1996; Malone et al., 1998; Mehta & Beer,
2010; Umukoro et al., 2013). The problem with many such studies is that the focus is on clinical or
criminal cases which pre-assigns the subjects and places them under a certain category – impulsive
individuals. A better scenario is using healthy individuals, or animals, to test impulsivity under
20

Krikeb, J., Serotonin and dopamine in predator-prey aggression
certain conditions. The conditions could be induced stress, hormonal manipulations, or
neurotransmitter manipulation, as some examples. The manner in which the impulsivity, or
aggression, could be tested is using questionnaires, such as the lifetime history of aggression scale,
or decisions made in tests such as the 5-choice serial reaction time task, delay discounting task, stop-
signal task, or go/no-go task. In questionnaires, individuals must testify about their own behaviour.
In the tasks, impulsivity is decided based on the ability to learn where waiting can lead to reward.
Using these methods it is possible to observe how healthy subjects react to manipulations and
therefore allow for determination of causation. We will review these tasks later as they will be used
to draw some conclusions concerning the roles of serotonin and dopamine in impulsive behaviour
among healthy subjects.
2.5. Experimental paradigms of impulsivity
What we learn from past experiments on aggression is that it is necessary to extract the
cognitive mechanism leading to it, which is, to a large extent impulsive behaviour (Dalley, Mar,
Economidou, & Robbins, 2008; Homberg, 2012; Kiser, Steemers, Branchi, & Homberg, 2012). It is a
type of behaviour that is more conspicuous in clinical cases related to gambling, alcoholism, drug
abuse, depression, and schizophrenia (Badawy, 2003; Booij et al., 2010; Dalley et al., 2011; Daw et
al., 2002; Doya, 2002; Linnoila et al., 1983; Okai, Samuel, Askey-Jones, David, & Brown, 2011; Rogers,
2011; Scholes et al., 2007; Umukoro et al., 2013) but while it does enlighten us as to some
particularities of the system, we are not interested in the pathological case, but in the healthy. It is
healthy people who mostly interact in predator-prey situations.
Impulsivity could be examined using questionnaires or different tasks. Questionnaires include:
the Barratt impulsiveness scale, the urgency, premeditation, perseverance, and sensation seeking
impulsive behaviour scale, Behaviour Scale, the Impulsiveness Venturesomeness and Empathy
21

Krikeb, J., Serotonin and dopamine in predator-prey aggression
Questionnaire, and the Lifetime History of Impulsive Behaviours (Dalley et al., 2011; Dalley & Roiser,
2012). There are several tasks that are often used in the lab, some only fit for animals, and some for
both animals and humans, that are used to examine impulsive behaviour. Even though action
suppression is mostly the manner of study, and impulsive action does not use the exact same
mechanism as impulsive choice, it is a useful study tool since the mechanisms interact along with the
serotonergic and dopaminergic systems (Dalley et al., 2011). We will now briefly introduce some of
the more common methods.
2.5.1. 5-CSRT
The 5-choice serial reaction time task involves the animal initiating the trial and then waiting for
one of five cues to light up. A press before the light has turned on is deemed impulsive and results a
5 second timeout. A press on the button under the correct light, once it has turned on, results in a
reward food-item. An incorrect press, or no choice at all, are incorrect choices and result in a
timeout as well. The goal of this task is to learn to suppress a response in order to gain a reward
(Dalley et al., 2011; Dalley & Roiser, 2012).
2.5.2. Go/no-go
A Go/no-go, or stop-signal, task can have a human or an animal version to it; in the human
version the reward is often simply a smiley face while in the animal model version the reward is a
food item. In this task the subject must press the button on ‘go’ trials upon the presentation of a
cue. In ‘no-go’ trials the subject must avoid pressing the button when the cue is given (Dalley &
Roiser, 2012). This task combines the effects of learning – subjects learn during the task itself how to
respond to the cue, and relearning; thereby fighting the impulse to act on what has been learnt
previously for the ‘go’ trials in order to succeed in the ‘no-go’ trials.
22

Krikeb, J., Serotonin and dopamine in predator-prey aggression
2.5.3. SSRT
The stop signal reaction time task models inhibition of motor control – the ability to stop an
action that has already been initiated. It is similar to the go/no-go task only here all trials are ‘go’
trials until a ‘stop’ signal is given at which point subjects must stop responding (Dalley et al., 2011).
Crucially, reaction times for the responses are taken so it can be compared between trials after a
‘stop’ signal has been given and thus observe readjustment. Despite the similarities in the tasks,
different manipulations affect the results of SSRT and go/no-go differently (Dalley et al., 2011).
2.5.4. Reversal learning
In a reversal learning task subjects are conditioned to respond to a specific conditioned stimulus
and ignore a second stimulus. Once this conditioning is established, the subjects are asked to switch
the reactions between the stimuli – act on the second and ignore the first (Homberg, 2012). This task
can be practised both with humans, as well as with animals, even though, according to Homberg
(2012), it is not entirely clear whether the two employ the same brain mechanism for the task. This
is another paradigm of learning and suppression of action.
2.5.5. Delayed reward
A delayed reward, or delayed discounting task, constructs a situation where the subjects face a
choice between a small reward now and a larger reward later. The manipulations could be either on
the size of the reward or the length of the delay (Dalley & Roiser, 2012). In animals, the delay is
measured in seconds and the reward is food. In humans, the design involves a hypothetical choice of
monetary reward that spans between minutes and years. This paradigm has a clear focus on
impulsive choice since the strictly logical choice is always the larger reward.
3. Serotonin
23

Krikeb, J., Serotonin and dopamine in predator-prey aggression
Serotonin, or 5-HT, is a well-studied neurotransmitter that has been associated with mood,
survival, social behaviour, sleep, brain plasticity, impulsivity, and mental illness (Benningfield &
Cowan, 2013; Blier & El Mansari, 2013; Dalley et al., 2011, 2008; Dalley & Roiser, 2012; Gellynck et
al., 2013; Homberg, 2012; Jouvet, 1999; Kiser et al., 2012; Lovinger, 2010; Martinowich & Lu, 2008;
Navailles & De Deurwaerdère, 2011; Roberts, 2011; Rogers, 2011; Scholes et al., 2007; Takahashi et
al., 2011). Our focus in this paper is on impulsive (choice) and aggressive behaviour as they are
linked to altered serotonin levels. Brown et al. (1979) are the first to find a negative correlation
between serotonin, based on 5-HIAA concentrations in CSF, and aggressive behaviour in humans. In
another study, both aggression and low levels of serotonin are correlated with suicide attempts
(Brown et al., 1982). In another lane of research, Linnoila et al. (1983) suggest, based on their results
that separate impulsive offenders and those who were conscious and calculated in their actions, that
low levels of serotonin are an indication of impulsive behaviour. A problem with this conclusion is
that they decided according to the offence itself who was premeditated and who was not (Berman
et al., 1997). It seems however, that despite the problems this study may present, studies that
examined the influence of alcohol on aggression, do find that certain individuals with a tendency for
violence will be more prone to it following the ingestion of alcohol and this could stem from the
alcohol’s effect on brain serotonin levels – i.e. it depletes them (Badawy, 2003). These findings
correlate with the idea that the correct food supplementation could eliminate, or reduce, aggressive
behaviour (Badawy, 2003).
3.1.1. Molecule
5-HT is produced in the raphe nuclei that sit in the mesencephalon (brainstem) (Martinowich &
Lu, 2008). It is a simple monoamine molecule, much like many other neurotransmitters such as:
dopamine, melatonin, epinephrine, and norepinephrine. 5-HT is derived from L-tryptophan, one of
the basic amino acids (Folk & Long, 1988; Kiser et al., 2012; McEllistrem, 2004). 5-HT has a major
24

Krikeb, J., Serotonin and dopamine in predator-prey aggression
central metabolite, 5-hydroxyindoleactic acid (5-HIAA), and the most reliable method to examine the
presence of serotonin in the human brain is looking for it in a sample of cerebrospinal fluid (CSF)
(Brown et al., 1979).
3.1.2. Different receptors in different brain regions
Serotonin seems to have appeared early in our evolution and thus, being an important molecule,
it presents the highest number of receptors in the brain. These divide into ionotropic, which allow
ion transfer upon ligand binding, and metabotropic, which start a signalling chain within the cells
when the ligand binds. Accordingly they have different functions (Gellynck et al., 2013; Kiser et al.,
2012; Martinowich & Lu, 2008).
Serotonin is infamous for its effects following the ingestion of 3,4-
methylenedioxymethamphetamine (MDMA), which inhibits the reuptake of the neurotransmitter
back into the presynaptic neuron from the synaptic cleft by competing with it, and other
monoamines, for their reuptake transporters (SERT – serotonin reuptake transporter, and DAT –
dopamine reuptake transporter) (Benningfield & Cowan, 2013; Rubia et al., 2005). It also inserts
itself into the presynaptic neuron and induces release of more serotonin into the synaptic cleft. This
leads to social disinhibition and therefore MDMA is a very popular for recreational use. These effects
are similar to those that form the basis of antidepressant drugs, that also have the same effect of
saturating the synapses with serotonin, either through blocking reuptake or through inducing
increased production (Blier & El Mansari, 2013; Martinowich & Lu, 2008). The effects on social
behaviour are the important effects we need to keep in mind in order to understand the role of
serotonin.
Selective serotonin reuptake inhibitors (SSRIs) are the current antidepressants of choice among
clinicians (Blier & El Mansari, 2013; Crockett et al., 2009; Dalley & Roiser, 2012; Kiser et al., 2012;
25

Krikeb, J., Serotonin and dopamine in predator-prey aggression
Martinowich & Lu, 2008). SSRI drugs are agonists of the 5-HT1 receptors family (Takahashi et al.,
2011). The higher concentrations of 5-HT are believed to have beneficial effects on social behaviour
generally even though that in the case of major depression, a common mental disorder, the low
levels of serotonin are not necessarily the cause (Blier & El Mansari, 2013). However, even with only
partial support linking the low levels of serotonin to aggression and major depression, increasing the
levels of 5-HT is often beneficial.
To counteract the effects of aggression, similar to antidepressants, there are drugs that activate
only a subset of the serotonin receptors. There are 16 different genes encoding 5-HT receptors (Kiser
et al., 2012) and various drugs work with different specificity on these receptors. These drugs are
tested mostly in experiments using animal models to examine their effects. De Boer and Koolhaas
(2005) examine some compounds that act as agonists and antagonists to 5-HT1A and 5-HT1B family
receptors. The former acts as an autoreceptor on 5-HT neurons, as well as a postsynaptic receptor
on pyramidal and GABAergic (inter)neurons (Lladó-Pelfort et al., 2012), that reacts differently to
different drugs according to its location. For example, S-15535, a benzodioxopiperazine, acts as an
agonist to 5-HT1A when it is on the presynaptic neuron and as an antagonist when it is on the
postsynaptic one (de Boer & Koolhaas, 2005). It has a similar effect as other pharmacological agents
in reducing aggressive behaviour, for instance: repinotan, 8-OHDPAT, ipsapirone, and some others.
Unlike these other compounds, S-15535 does not affect other aspects of non-aggressive motor
behaviour (de Boer & Koolhaas, 2005). These other effects could occur due to several reasons: a
higher dose is needed, or the non-selective activation of other receptors. This sort of study highlights
the difficulty in assessing the specific role of a specific receptor. Despite this, there are many other
studies doing so and we will look into some more of them.
26

Krikeb, J., Serotonin and dopamine in predator-prey aggression
Selective antagonists of 5-HT1A and 5-HT1B indeed demonstrate blocking of the attenuating
aggression effects if given alongside S-15525, while having no effect by themselves. Specifically
related to the amygdala function, the effects of 5-HT1A receptor deficit seems to lead to anxiety
whereas its over-expression leads to aggressive behaviour (Saha et al., 2010). This is contrasted with
5-HT1B receptors whose over-expression restricts aggressive behaviour while their rat knockout
models show increased aggression (Saha et al., 2010). To contrast the actions of 5-HT1A from 5-HT2
and 5-HT3, it was shown by Stein, Davidowa, and Albrecht (2000) that a 5-HT1A agonist decreases the
firing rate of neurons in the amygdala while 5-HT2 and 5-HT3 agonists resulted an increase in firing
rate. Moreover, research on the different 5-HT2 receptors leads to different results based on the
specific location of action, as opposed to global infusion of a drug, and that is in addition to the
effect of the drug being agonist or antagonist (Homberg, 2012). In that review, Homberg (2012)
states that 5-HT6 “univocally contribute[s]” to impulsivity (p. 230) however it receives little mention
elsewhere. In addition to the amygdala, also the presence of 5-HT1A receptors in the PFC has been
addressed. Additionally, there are functional links between the OFC and the basolateral amygdala
that influence decision making (Winstanley, Theobald, Cardinal, & Robbins, 2004).
Interestingly, there is no hypersensitivity of 5-HT receptors observed in aggressive individuals
(Wetzler et al., 1991). However, aggressive individuals do have different ratios of 5-HT receptors
expressed in the brain (de Almeida et al., 2005).
3.1.3. Tryptophan depletion
One manipulation that can be done in studies specifically oriented at the effects of 5-HT is using
an acute tryptophan depletion paradigm. In the studies using this paradigm, participants drink a
liquid that contains a concoction of different concentrations of amino acids besides tryptophan
(control group drinks the same concoction that includes tryptophan as well). Based on previous
27

Krikeb, J., Serotonin and dopamine in predator-prey aggression
studies analysing rat brain tissue, this leads to a sharp reduction in serotonin production (Crockett et
al., 2009). Five hours after the intake of said drink, participants normally are controlled for changes
in mood, and then are given the task and/or commence the game (Crockett et al., 2009; Wood,
Rilling, Sanfey, Bhagwagar, & Rogers, 2006). This form of research allows for a contrast between
normal and decreased systemic concentrations of serotonin to exemplify its behavioural effects.
3.1.4. Tryptophan supplementation
Similar to depletions studies, there are also tryptophan supplementation studies performed.
Tryptophan could be supplemented in an acute study (Scarnà, McTavish, Cowen, Goodwin, &
Rogers, 2005) or over a duration of a couple of weeks (Murphy et al., 2009). In either case, the
dietary supplement consists of a high level or tryptophan that should keep the enzyme tryptophan
hydroxylase, the rate-limiting enzyme in the 5-HT production, close to saturation (Murphy et al.,
2009). The increased levels of tryptophan should in turn lead to increased 5-HT levels that alter
social behaviour (Murphy et al., 2009), similar to effects of other drugs that interact with the
serotonergic system that have been previously touched upon. Other effects on mood, memory, or
attention are then controlled for before the main task is initiated.
3.1.5. Serotonin knockouts
Another manipulation that can be performed in the lab using animal models is knockout a
specific gene using viral agents and breeding. In this paper our highlight is the knockout of specific 5-
HT receptor or enzyme genes. In such knockout models the role of a specific receptor in the entire
mechanism can be assumed from the changes observed in behaviour in the knockout animals, as
opposed to the wild-type healthy controls. Examples include knockout of the 5-HT1A or 5-HT1B
receptor (Saha et al., 2010), 5-HT transporter (5-HTT) (Kiser et al., 2012), or monoamine oxidase A
28

Krikeb, J., Serotonin and dopamine in predator-prey aggression
(MAOA) which is a main serotonin catabolising enzyme (McDermott, Tingley, Cowden, Frazzetto, &
Johnson, 2009).
3.1.6. Specific agonists/antagonists and neurotoxins
Other paradigms that involve mature animals are the use of neurotoxins or specific agonists and
antagonists. For example, 5,7-dihydroxytryptamine (5,7-DHT) injected directly to the brain removes
all serotonin (Dalley & Roiser, 2012).
Some examples of agonists and antagonists are: 8-OH-DPAT is a specific 5-HT1A agonist, (±)-1-
(2,5-dimethoxy-4-iodo- phenyl)-2-aminopropan (DOI) is a 5-HT2A/2C agonist while SER082 is an
antagonist for these receptors, SB242084 is a 5-HT2C antagonist and M100907 is an antagonist
specific for 5-HT2A (Dalley & Roiser, 2012). These are just some examples of molecules used to
examine the role of specific receptors or the interactions between them. Some of these molecules
can be injected also very accurately to specific regions, while others are given globally.
3.2. Serotonin and impulsivity
Since we discussed serotonin as a key player in impulsive behaviour, we will now look at some
evidence from the lab. Dalley and Roiser (2012) address some of the past results using various
agonists and antagonists for different 5-HT receptors. For instance: a systematic administration of
DOI increases impulsivity in a delay discounting task relying on 5-HT2A mechanism. A similar result
was achieved in the 5-CSRTT using the 5-HT2C antagonist SB242084 (Dalley & Roiser, 2012). SER082, a
5-HT2A/2C antagonist, did not affect 5-CSRTT behaviour but did impact the delay discounting task by
reducing impulsive response. 8-OH-DPAT, a 5-HT1A agonist had positive results for the choice
reaction time task however it negatively affected delay discounting. 5-HT reuptake inhibitors yielded
positive results for both tasks (Dalley & Roiser, 2012). M100907, a 5-HT2A antagonist injected to the
29

Krikeb, J., Serotonin and dopamine in predator-prey aggression
PFC, and the 5-HT1A receptor agonist 8-OHDPAT, both result in blocking impulsivity in the 5-CSRTT
(Dalley et al., 2011).
These results exemplify the role serotonin has in different types of impulsive behaviour and
would suggest therefore that it is a good candidate to maintain when examining human impulsive
decision-making.
3.3. Link between serotonin research to predator and prey behaviours
Apter et al. (1990), in their paper, discuss separating impulsivity, as a character trait, from the
different psychological diagnoses available, since it seems that this is what 5-HIAA measurements
indicate. Berman et al. (1997) suggest in their discussion that a distinction between impulsive and
non-impulsive aggression be made in further research and that potential mediating factors, such as
environmental factors, be examined. This suits our bimodal classification system.
In brain circuitry terms, this goes in line with the fact that the raphe nuclei project to the
orbitofrontal cortex (OFC), and also project back to it, thus creating a feedback loop with the
serotonergic system (Roberts, 2011). The OFC is associated with depression and obsessive
compulsive disorder but additionally, along with the PFC, it is associated with top-down control,
planning, and decision-making (Dalley et al., 2011). This decision-making could be impulsive and
more leaning towards risk, while it could be more risk-aversive. It has been found that tryptophan
supplemented diet reduced risk-taking in the form of the reflection effect as well as influencing loss-
aversion (Murphy et al., 2009, in Rogers 2011). Additionally, carriers of the ss (short) allele of the 5-
HTTPLR were more sensitive to the framing effect (Tversky & Kahneman, 1981), and could be
supposed to have been more emotional in their rationalisation since a stronger interaction between
the amygdala and the PFC was observed (Roiser et al., 2009, in Rogers 2011).
30

Krikeb, J., Serotonin and dopamine in predator-prey aggression
A link between decision-making and impulsivity could be made based on the study that found
that only delayed bad outcomes were retarded by tryptophan depletion (Blair et al., 2008, and
Tanaka et al., 2009, in Rogers 2011). This delayed learning would invite further impulsive socially
inappropriate behaviour. This is also discussed by Homberg (2012), detailing the discounting of
future reward, as well as past punishment, following tryptophan depletion. The social implications of
low levels of serotonin are also evident through the iterated prisoner’s dilemma game where
tryptophan depleted subjects were less cooperative, even while playing against the quite forgiving
tit-for-tat strategy (Wood et al., 2006).
Also according to Kiser and colleagues (2012), impulsive aggression correlates with low levels of
serotonin and calculated aggression correlates with higher levels. This works well with the idea that
higher 5-HT levels are expected in socially capable individuals and therefore they are able to
perceive hostile behaviour towards them and react accordingly. This would apply in economic terms
as well when a person, or group, should feel threatened it would enact decisions to protect itself,
however, in a measured manner.
An example for the effects of increased serotonin levels was found in monkeys. In some studies
of monkeys it was shown that tryptophan supplementation led to both better social interaction, as
well as strategic aggression the led to a rise in the hierarchy (Kiser et al., 2012).
As we have described, impulsive decisions, besides violence (a poor choice of behaviour in
current society), lead to choices that have no or little foresight. Let as move from there to economic
decisions. Thus, we would expect people with lower concentrations of 5-HT to act impulsively not
only in the standard prisoners’ dilemma game, but also in the iterated version (Wood et al., 2006; Yi,
Johnson, & Bickel, 2005). These people will choose to defect also when repeated interaction plays a
role in the eventual outcome of the game and therefore we would term them as being selfish, or
31

Krikeb, J., Serotonin and dopamine in predator-prey aggression
aggressive players. The outcomes in Yi et al. (2005) show that delayed discounting correlated with
(the successful) tit-for-tat strategy, and not with random play, in the iterated game. This behaviour
indicates an inability to see the mutual benefits of cooperation as the model of reciprocal altruism of
the iterated prisoners’ dilemma suggests, especially under the tit-for-tat strategy. Interestingly, in
Wood and colleagues’ (2006) work, trait aggression is not a consequence of tryptophan depletion.
Furthermore, the results were not replicated in the second day of the study; an effect related to the
role serotonin plays in social learning – possibly after learning the game on the first day, being
depleted on the second day did not induce as strong an of an effect.
Fig. 3. Ultimatum game decision-making (based on results of Corckett et al., 2008, 2010).
An additional point was made by Crockett et al. (2008) in their study of altruistic punishment as
a tool for social behaviour. Altruistic punishment, being a punishment where one has to make an
effort, or invest of their own funds, in order to inflict (financial) pain on others, has shown success in
experimental conditions at maintaining a social norm and is believed to serve a reciprocal social
system (Fehr & Rockenbach, 2004). According to other studies, that link serotonergic activity to it,
32

Krikeb, J., Serotonin and dopamine in predator-prey aggression
altruistic punishment negatively correlates to serotonin levels following tryptophan depletion,
suggesting that it is impulsive behaviour rather than calculated (Crockett et al., 2010, 2008). Crockett
and colleagues (2010, 2008), used the ultimatum game as a tool for measuring altruistic punishment.
In this game, participants could choose whether to accept offers of splitting a sum or reject them. By
accepting, both sides split the sum as agreed, while rejection means that neither side receives
anything, thus in fact being an altruistic punishment. While it is commonly believed that rejection of
unfair offers is perceived as a calculated “teaching” strategy on behalf of the responder, according to
their findings, Crockett et al. (2010, 2008) conclude that it is in fact an impulsive choice. This is based
on the significant increase in rejection of unfair offers following tryptophan depletion. They further
support this by showing worse choices made in a delay discounting task (Crockett et al., 2010). This
task demands people to choose between a small reward now or a bigger reward later (with the size
of reward and length of time delay as variables) and is amongst the common tests used to measure
impulsivity since it makes little sense to opt for the immediate reward. In Crockett and colleagues’
(2010) study, tryptophan depletion also resulted in increase in number of instant rewards chosen.
Additionally, in their discussion, Crockett et al. (2010) connect the dorsal striatum, part of the
reward system and mostly dopamine dependant, to the enjoyment of punishment (p.860).
Protective, or reactive, aggression, is most likely related to fear and therefore to the amygdala
and the rest of the limbic system. This is based on de Dreu and colleagues’ (2014) work where they
found that the subjects acting as prey made a quicker choice in their investment. This decision was
accompanied by activation of the amygdala and therefore we can call this decision a more instinctive
one. Since the amygdala carries serotonin projections as well (Homberg, 2012; Kiser et al., 2012;
Saha et al., 2010), excitement in the amygdala is likely to involve, amongst other neurotransmitters,
also serotonin action. If we can observe increased activation then we can also assume a higher firing
rate of the 5-HT2 as well as 5-HT3 (Stein et al., 2000). This can lead also 5-HT1B activation but that
33

Krikeb, J., Serotonin and dopamine in predator-prey aggression
should reduce aggression (Saha et al., 2010). Therefore, our expectation would be 5-HT1A activation,
that we would expect to correlate with anxiety, but that should decrease activity levels across the
amygdala (Stein et al., 2000). Once again we are confronted with the intricacy of the system.
Siegel and Victoroff (2009) discuss the suppressing effect serotonin has on reactive aggression
through 5-HT1A activation in the medial hypothalamus or in the PAG. However, they indicate that
through the 5-HT2 receptor activation in the PAG, limbic system, and the hypothalamus, reactive
aggression is initiated. This links fear and aggression through the serotonergic system.
Increased activity in the superior frontal gyrus is associated with impulse control as well as
calculated behaviour (De Dreu et al., 2014). Decreased levels of activity follow tryptophan depletion
in healthy humans (Rubia et al., 2005). These two findings interact neatly to form a picture of
calculated decision-making in the PFC.
From a conscious logical perspective it is interesting to wonder why would the predator
aggressor attempt an offence to begin with since generally speaking, the defender increases its
levels of aggression often to the extent that the predator loses the conflict and therefore from the
logical standpoint, the foresight should be that initiating an offensive is the wrong move. Rogers
(2011) indicates that there is little research looking into risky choices that lead to bigger rewards and
serotonin but part of it is related to delay discounting being disturbed by tryptophan depletion that
has been previously discussed. Interestingly, probabilistic discounting is not diminished by the
depletion of tryptophan. These are both examples of behaviour that could be paralleled to predator
behaviour. To further support this effect of 5-HT deficiency, an experiment with macaques
demonstrated that switching from risky choice to safe choice demands a lower variance for the pay-
offs following tryptophan depletion, further suggesting that calculated aggression is not as
premeditated as we assume.
34

Krikeb, J., Serotonin and dopamine in predator-prey aggression
All the evidence from the research on serotonin suggests that it is a necessary component of
intelligent decision making. We can hypothesise that a potential prey will become one when they
are with low levels of serotonin and one that is already a prey will invest excessively as it is the
impulsive choice. We can further hypothesise that a predator with low serotonin levels does not
exist.
4. Dopamine
4.1.1. Molecule
Dopamine, another monoamine neurotransmitter, is often studied for its effects on fine
procedural memory, working memory, reward circuit, addiction, and motor skills in relation to
Parkinson’s disease (Doya, 2002; Lovinger, 2010; Navailles & De Deurwaerdère, 2011; Okai et al.,
2011; Segura-Aguilar et al., 2014; Wise, 2009). It is also the major focus of research into the reward
circuitry in the brain and how it affects both addiction and reinforcement learning (Daw et al., 2002;
Glimcher, 2011; Volkow, Wang, Fowler, Tomasi, & Telang, 2011). It is produced from tyrosine in
dopaminergic neurons of the VTA and the substantia-nigra (SN) that project to the striatum mainly
(Brichta, Greengard, & Flajolet, 2013; Dalley & Roiser, 2012; Ikemoto, 2010; Scholes et al., 2007;
Wise, 2009). The former is related to the reward mesolimbic system and the latter the nigrostriatal
projections that are key for motor control (Wise, 2009).
4.1.2. Different receptors in different brain regions
Dopaminergic input, besides the striatum, project from the VTA to the PFC (Doya, 2002). Its
effect is mediated through five different metabotropic G protein-coupled receptors (Beaulieu &
Gainetdinov, 2011) that are differentially functional as well as differentially spread out in the brain.
The D1 and D2 receptors are significantly present in the amygdala whereas the D5 receptors are
35

Krikeb, J., Serotonin and dopamine in predator-prey aggression
present in a high concentration in the pyramidal neurons of the PFC (Beaulieu & Gainetdinov, 2011).
They are all present in other regions, in addition to the other two receptors; however, it is less
significant for this paper.
Some examples for the rewarding function of dopamine will be given here since they interaction
of this system with social behaviour is obvious, even if it is not directly related to aggression or
impulsivity. Dopamine is rewarding when there is a spike in its activity and certain dopamine
receptors, such as D4, have different polymorphisms that allow levels of sensitivity and stronger or
weaker good feeling that is very important for the sort of social behaviour humans present (Bachner-
Melman et al., 2005). The prediction errors for reward, or lack thereof, have been correlated to
spikes in dopamine activity in the midbrain following unexpected reward, or alternatively, a dip
following a lack of an expected reward (Schultz, 2004, 2007, in Rogers, 2011).
Other dopamine receptors are studied in other contexts. Of relevance for our paper is the
interventions performed on the dopaminergic systems in order to reduce aggression. For instance,
haloperidol has been used for many years as a treatment for psychotic patients. Haloperidol acts as
an antagonistic agent on the dopamine D2 receptors (de Almeida et al., 2005). In other research it
has been found that D2 receptor antagonists also hinder people from detecting angry faces (Seo et
al., 2008). D2 receptors also have two isoforms that have been studied in mouse models: D2L, long
form, and D2S, the short one, and in the mouse brain the long form is more abundant, with an overall
D2 density reduction with old age (Vukhac et al., 2001). It has been observed that older mice are
significantly less aggressive, moreover, mice lacking the long isoform, D2L, are significantly less
aggressive, without losing overall D2 density (Vukhac et al., 2001). D2 receptor’s role is deeper than
aggression though since it could be characterised as working to regulate compulsions and impulsive
36

Krikeb, J., Serotonin and dopamine in predator-prey aggression
behaviour as has been shown with the effects on gambling behaviour with people who take D2
affecting drugs (Rogers, 2011).
4.1.3. Parkinson’s and L-dopa
Since dopamine is produced in the substantia-nigra that is the main target of Parkinson’s
disease, a large amount of research is focused in that area. The loss of dopamine producing neurons
leads to loss of fine motor control as well as impulsive behaviour such as compulsive gambling and
binge eating (Brichta et al., 2013; Dalley & Roiser, 2012; Okai et al., 2011). The standard treatment is
using L-dopa, an in-between molecule in the process of converting tyrosine to dopamine, in drug
form (Brichta et al., 2013; Segura-Aguilar et al., 2014). Thanks to this treatment, tests can be
performed comparing patients in their “on” and “off” states. For instance, in “off” state, patients
show impatience in a delayed discounting task (Dalley & Roiser, 2012). This is implicating both L-
dopa, thus dopamine in general, as well as D2/D3 specific agonists that are also sometimes used as
drugs. The D2 receptor is also associated with learning as another Parkinson’s study shows us.
Patients in the “off” mode learnt better from negative outcomes than from positive outcomes, while
this was reversed in the “on” mode (Frank et al., 2004, in Rogers, 2011).
Further data on the D1 and D2 function in the amygdala is given by Smith, Geissler, Schallert, and
Lee (2013) using a Parkinson’s disease animal model. The study focused on a cognitive attention
switching behaviour that is also deficient in patients. The importance of the dopamine projections
from the substantia-nigra to the amygdala are stressed in this study and D1 importance in a
switching as well as a 5-CSRT task.
4.1.4. Other manipulations to dopamine
Administering L-dopa is a similar treatment to tryptophan supplementation in order to examine
higher levels of serotonin. Also specific agonists can be used in the case of the different dopamine
37

Krikeb, J., Serotonin and dopamine in predator-prey aggression
receptors, as has been described for some of the Parkinson’s disease studies. Additionally, targeting
neurons using toxin injections or knockout models is used in dopamine research as it is used in 5-HT
research.
4.2. Dopamine and impulsivity
The D2 receptor is associated with impulsivity and aggression, and D1 is associated with decision-
making (Rogers, 2011). A few studies described in his review indicate an interaction taking place in
the anterior cingulate cortex (ACC) and the nucleus accumbens that seem to reflect cost-benefit
calculations. This has been surmised from rats being observed willing to exert more effort for larger
rewards. Rogers further describes the distribution of the D1 and D2 receptors as being instrumental in
the balance between the direct and the indirect striatal pathways (2011). These two pathways are
believed to dominate the Go and No-Go competing actions respectively. While there is empirical
support of this hypothesis, affirming these assumptions with humans is at the moment tricky due to
drug selectivity and location of action. D1 receptors, and not D2, are associated with fear learning as
well as with mechanisms that involve the decisions relevant for the 5-CSRT (E. S. Smith et al., 2013).
Impulsive behaviour is often observed in substance abusers. Rogers (2011) discusses the
deficiencies substance abusers have in probabilistic decision making that would be caused by
damage to the fronto-striatal systems. However, as he further remarks, it could be that the drug
seeking, impulsive, behaviour is the cause of an innate irregularity in the system that affects
decision-making.
4.3. Link between dopamine research to predator and prey behaviours
How to link dopamine effects to predator and prey behaviour becomes a bit tricky, despite the
effects it has on decision-making and the close link to the serotonin system. McEllistrem (2004)
discusses some evidence found, most of it from research that is a little dated. In cats, dopamine
38

Krikeb, J., Serotonin and dopamine in predator-prey aggression
agonists reduced hissing and reduced reactive aggression while antagonists have had the opposite
effect. This is in line with evidence from clinical cases where haloperidol, a dopamine antagonist, is
used to treat violent cases. Additionally, as has been previously discussed, the D2 receptor role in
aggression and impulsivity, as well as the D1 role in fear learning, which leads to a certain expected
interaction between the two in prey reaction to a threat.
Dopamine seems to be a potentiator, through D2 receptor activation, of both predator and prey
types of aggression (Siegel & Victoroff, 2009). There are dopaminergic projections connecting the
VTA to the hypothalamus and the limbic system through which this effect takes place. Siegel and
Victoroff (2009, p. 211) also suggest that dopamine is “permissive” in aggression, rather than
initiating it. Thus in essence, the dopamine system’s interaction with other systems is how it exerts
its effect and this is what we are now going to examine.
5. Interactions between serotonin and dopamineThere are only a few studies that touch on the interaction between the serotonergic and
dopaminergic systems. For instance, these interactions between 5-HT and dopamine have been
found in rats under the influence of d-amphetamine (Dalley & Roiser, 2012). Amphetamine is a
stimulant that increases the levels of dopamine and thus is believed to reduce impulsive action in
the delay-discounting task. However, rats that have been depleted of serotonin do not experience
this improvement in decision-making (Dalley & Roiser, 2012). Rogers (2011) in his review also
suggests that serotonin and dopamine have complementary roles. In the article another evidence to
point it out is that tryptophan depletion removes the rewarding effect of cocaine in humans, while
SSRIs reinforce the effects of cocaine in rats.
39

Krikeb, J., Serotonin and dopamine in predator-prey aggression
An interesting finding concerning both the PFC and the striatum and attentional control involves
a study including healthy individuals who ingest a tyrosine and/or tryptophan deficient diet in order
to cause an acute serotonin and/or dopamine deficiency (Scholes et al., 2007). In this research,
Scholes et al. (2007) found that individual decreases of these two neurotransmitters improves
attentional control as it is measured by the Stroop task (depletion of both dopamine and serotonin
was insignificant). Also this experiment indicates a close link between the two systems, in this case,
on attentional control. This is further supported by a study using 8-OHDPAT to excite 5-HT1A neurons
(Lladó-Pelfort et al., 2012). It was shown that the excitation of these neurons in the hypothalamus or
raphe nuclei does not lead to increased pyramidal neuron firing while in the medial PFC this same
agonist activates preferentially GABAergic neurons that lead to a disinhibition of pyramidal neurons
that in turn affect downstream dopaminergic neurons.
6. DiscussionWe first started with a definition of aggression. If we would ask what the purpose of aggression
is in evolutionary terms, it is “as solutions […] to a host of distinct adaptive problems, such as
resource procurement, intrasexual competition, hierarchy negotiation, and mate retention” (Buss &
Shackelford, 1997). It is difficult to place predatory aggression on this scale. As Siegel and Victoroff
(2009) question, what is the function of this type of aggression. Additionally, they point out, there is
much more research into the defensive type than the predatory one.
Definition of impulsivity in the lab is still unclear since it seems to depend on the test performed
or the questionnaire given. Dalley and Roiser (2012) present that the D2/3 antagonist eticlopride,
when it was infused into the nucleus accumbens, resulted improved performance in the 5-CSRTT. In
that same paper, they also indicate that D1/2 antagonists, systematically administered, lead to
40

Krikeb, J., Serotonin and dopamine in predator-prey aggression
increased impulsivity in delay-discounting tasks. We can question the role of the different locations
of the receptors in the brain, or across the synaptic cleft, but it would be wiser to first question the
task and ask what sort of conclusions it leads us to.
Another point of discussion is brought by de Almeida et al. (2005). In their paper they
questioned the underlying assumption that sampling of CSF at a certain instant indeed exemplifying
the regular levels rather than what it is, a single moment’s sample.
A true confounder in the data comes from Murphy et al. (2009) who investigated the reflection
effect. This effect demonstrates the misguided ideas concerning gains and losses people have. Given
the choice between a certain win, a 50/50 chance of double that amount, or no win at all, people
tend to go for the certain win. This risk-aversive choice is reversed when the same choices are given
with the option to lose a certain amount. Thus, when the option is to lose money, people are more
willing to seek more risk. This risk-seeking versus risk-aversive behaviour is modified by tryptophan
supplementation. The participants who were on an added tryptophan diet took a longer time to
reach their decisions, both for wins and even more so for losses (Murphy et al., 2009). Additionally,
they seemed to be more risk-seeking for the wins while being more risk-aversive for the losses. This
reversal is difficult to explain but perhaps could suggest that the increased levels of 5-HT allowed for
more learning and therefore the decisions made were made within a broader frame of all the losses
and wins in the experiment.
Aaldering and de Dreu's (2012) hawkish and dovish paradigm of research placed some
participants as hawks and others as doves thus artificially endowing them with character.
Interestingly, this empowerment, or pacification, indeed was reflected in their behaviour. Booij et
al.'s (2010) longitudinal study followed children with a history of aggression over 21 years and found
that following that period, even the ones with low 5-HT concentrations do not present the
41

Krikeb, J., Serotonin and dopamine in predator-prey aggression
heightened levels of aggression. Both of these studies conclusions suggest together that perhaps the
most important thing is the environment and the external input, and not the anatomy or
neurotransmitters; there may be a tendency, more in some than in others, but eventually this
reflects what is imposed on it. This is in a similar vein with (De Dreu, 2012; Israel, Weisel, Ebstein, &
Bornstein, 2012) concerning the effect of oxytocin, which interacts with the serotonergic system, in
different contexts. This also follows from a line of research on the effects of chronic stress on the
alteration of monoamine levels in the brain (Roberts, 2011).
Bottom line, we shouldn’t forget that we are trying to model very complex behaviour based on
evidence from very specific brain circuits and experiments that look at them and this is never going
to be one-to-one. This entire paper attempted to draw connections post-hoc between many studies
performed on aggressiveness and impulsivity and the predator-prey situation which has only one
published study based on that paradigm (De Dreu et al., 2014). There are bound to be some
suggestions for links that should be scrutinized and executed in a specialised research. For instance:
the specific 5-HT receptors that activate in the amygdala during a fearful reaction and examine the
causation of amygdala-hypothalamus interaction. It is to be hoped that the outline given for two
differently motivated aggressive behaviours was sufficient to convince that they are indeed rooted
differently in the brain circuitry and neurotransmitter systems that dominate them. Future research
should examine the motivations for intraspecies predatory behaviour in order to better create this
construct of aggression.
Acknowledgements:
The author would like to thank Joëlle Lafeber and Simon Columbus for their feedback on writing.
42

Krikeb, J., Serotonin and dopamine in predator-prey aggression
7. Bibliography
Aaldering, H., & De Dreu, C. K. W. (2012). Why hawks fly higher than doves: Intragroup conflict in representative negotiation. Group Processes & Intergroup Relations, 15(6), 713–724. doi:10.1177/1368430212441638
Afza, T., & Nazir, M. (2007). Is it better to be aggressive or conservative in managing working capital. Journal of Quality and Technology Management, 3(2), 11–21. Retrieved from http://www.ciitlahore.edu.pk/Papers/Abstracts/146-8588087935136570808.pdf
Agnoli, L., & Carli, M. (2012). Dorsal-striatal 5-HT2A and 5-HT2C receptors control impulsivity and perseverative responding in the 5-choice serial reaction time task. Psychopharmacology, 219(2), 633–645. doi:10.1007/s00213-011-2581-0
Anholt, R. R. H., & Mackay, T. F. C. (2012). Genetics of aggression. Annual Review of Genetics, 46, 145–164. doi:10.1146/annurev-genet-110711-155514
Apter, A., van Praag, H. M., Plutchik, R., Sevy, S., Korn, M., & Brown, S.-L. (1990). Interrelationships among anxiety, aggression, impulsivity, and mood: A serotonergically linked cluster? PsychiatryResearch, 32(2), 191–199. doi:10.1016/0165-1781(90)90086-K
Bachner-Melman, R., Gritsenko, I., Nemanov, L., Zohar, a H., Dina, C., & Ebstein, R. P. (2005). Dopaminergic polymorphisms associated with self-report measures of human altruism: a fresh phenotype for the dopamine D4 receptor. Molecular Psychiatry, 10(4), 333–335. doi:10.1038/sj.mp.4001635
Badawy, A. (2003). Alcohol and violence and the possible role of serotonin. Criminal Behaviour and Mental Health, 13(1998), 31–44. Retrieved from http://onlinelibrary.wiley.com/doi/10.1002/cbm.529/full
Beaulieu, J.-M., & Gainetdinov, R. R. (2011). The physiology, signaling, and pharmacology of dopamine receptors. Pharmacological Reviews, 63(1), 182–217. doi:10.1124/pr.110.002642.182
Benningfield, M. M., & Cowan, R. L. (2013). Brain serotonin function in MDMA (ecstasy) users: evidence for persisting neurotoxicity. Neuropsychopharmacology : Official Publication of the American College of Neuropsychopharmacology, 38(1), 253–5. doi:10.1038/npp.2012.178
Berman, M., Tracy, J., & Coccaro, E. (1997). The serotonin hypothesis of aggression revisited. Clinical Psychology Review, 17(6), 651–665. Retrieved from http://www.sciencedirect.com/science/article/pii/S0272735897000391
Blier, P., & El Mansari, M. (2013). Serotonin and beyond: therapeutics for major depression. Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences, 368(1615), 20120536. doi:10.1098/rstb.2012.0536
43

Krikeb, J., Serotonin and dopamine in predator-prey aggression
Booij, L., Tremblay, R. E., Leyton, M., Séguin, J. R., Vitaro, F., Gravel, P., … Benkelfat, C. (2010). Brain serotonin synthesis in adult males characterized by physical aggression during childhood: a 21-year longitudinal study. PloS One, 5(6), e11255. doi:10.1371/journal.pone.0011255
Bornstein, G., & Gilula, Z. (2003). Between-Group Communication and Conflict Resolution in Assurance and Chicken Games. Journal of Conflict Resolution, 47(3), 326–339. doi:10.1177/0022002703252367
Boureau, Y.-L., & Dayan, P. (2011). Opponency revisited: competition and cooperation between dopamine and serotonin. Neuropsychopharmacology, 36(1), 74–97. doi:10.1038/npp.2010.151
Brichta, L., Greengard, P., & Flajolet, M. (2013). Advances in the pharmacological treatment of Parkinson’s disease: targeting neurotransmitter systems. Trends in Neurosciences, 36(9), 543–554. doi:10.1016/j.tins.2013.06.003
Brown, G., Ebert, M., Goyer, P., Jimerson, D., Klein, W., Bunney, W., & Goodwin, F. (1982). Aggression, suicide, and serotonin: Relationships of CSF amine metabolites. The American Journal of Psychiatry, 139(6), 741–746. Retrieved from http://psycnet.apa.org/psycinfo/1982-25902-001
Brown, G., Goodwin, F., & Ballenger, J. (1979). Aggression in humans correlates with cerebrospinal fluid amine metabolites. Psychiatry Research, 1, 131–139. Retrieved from http://www.sciencedirect.com/science/article/pii/0165178179900532
Buss, D. M., & Shackelford, T. K. (1997). Human aggression in evolutionary psychological perspective.Clinical Psychology Review, 17(6), 605–619. doi:10.1016/S0272-7358(97)00037-8
Caseras, X., Mataix-Cols, D., Trasovares, M. V, López-Solà, M., Ortriz, H., Pujol, J., … Torrubia, R. (2010). Dynamics of brain responses to phobic-related stimulation in specific phobia subtypes. The European Journal of Neuroscience, 32(8), 1414–22. doi:10.1111/j.1460-9568.2010.07424.x
Chichinadze, K., Chichinadze, N., & Lazarashvili, A. (2011). Hormonal and neurochemical mechanismsof aggression and a new classification of aggressive behavior. Aggression and Violent Behavior, 16(6), 461–471. doi:10.1016/j.avb.2011.03.002
Crockett, M. J. (2009). The neurochemistry of fairness: clarifying the link between serotonin and prosocial behavior. Annals of the New York Academy of Sciences, 1167, 76–86. doi:10.1111/j.1749-6632.2009.04506.x
Crockett, M. J., Clark, L., Lieberman, M. D., Tabibnia, G., & Robbins, T. W. (2010). Impulsive choice and altruistic punishment are correlated and increase in tandem with serotonin depletion. Emotion (Washington, D.C.), 10(6), 855–862. doi:10.1037/a0019861
Crockett, M. J., Clark, L., & Robbins, T. W. (2009). Reconciling the role of serotonin in behavioral inhibition and aversion: acute tryptophan depletion abolishes punishment-induced inhibition inhumans. The Journal of Neuroscience : The Official Journal of the Society for Neuroscience, 29(38), 11993–9. doi:10.1523/JNEUROSCI.2513-09.2009
44

Krikeb, J., Serotonin and dopamine in predator-prey aggression
Crockett, M. J., Clark, L., Tabibnia, G., Lieberman, M. D., & Robbins, T. W. (2008). Serotonin modulates behavioral reactions to unfairness. Science, 320(27), 2008. Retrieved from http://www.sciencemag.org/content/320/5884/1739.short
Dalley, J. W., Everitt, B. J., & Robbins, T. W. (2011). Impulsivity, compulsivity, and top-down cognitivecontrol. Neuron, 69(4), 680–694. doi:10.1016/j.neuron.2011.01.020
Dalley, J. W., Mar, A. C., Economidou, D., & Robbins, T. W. (2008). Neurobehavioral mechanisms of impulsivity: fronto-striatal systems and functional neurochemistry. Pharmacology, Biochemistry, and Behavior, 90(2), 250–260. doi:10.1016/j.pbb.2007.12.021
Dalley, J. W., & Roiser, J. P. (2012). Dopamine, serotonin and impulsivity. Neuroscience, 215, 42–58. doi:10.1016/j.neuroscience.2012.03.065
Daw, N. D., Kakade, S., & Dayan, P. (2002). Opponent interactions between serotonin and dopamine.Neural Networks, 15(4-6), 603–616. doi:10.1016/S0893-6080(02)00052-7
De Almeida, R. M. M., Ferrari, P. F., Parmigiani, S., & Miczek, K. a. (2005). Escalated aggressive behavior: dopamine, serotonin and GABA. European Journal of Pharmacology, 526(1-3), 51–64.doi:10.1016/j.ejphar.2005.10.004
De Boer, S. F., & Koolhaas, J. M. (2005). 5-HT1A and 5-HT1B receptor agonists and aggression: a pharmacological challenge of the serotonin deficiency hypothesis. European Journal of Pharmacology, 526(1-3), 125–39. doi:10.1016/j.ejphar.2005.09.065
De Dreu, C. K. W. (2012). Oxytocin modulates cooperation within and competition between groups: an integrative review and research agenda. Hormones and Behavior, 61(3), 419–28. doi:10.1016/j.yhbeh.2011.12.009
De Dreu, C. K. W., Giffin, M., Krikeb, J., Prochazkova, E., & Columbus, S. (2015). No Parochial AltruismEvolves to Defend against Predation and Needs No Norm Enforcement. Proceedings of the National Academy of Sciences, Under revi.
De Dreu, C. K. W., Scholte, H. S., van Winden, F. A. A. M., & Ridderinkhof, K. R. (2014). Oxytocin tempers calculated greed but not impulsive defense in predator–prey contests. Social Cognitiveand Affective Neuroscience. doi:10.1093/scan/nsu109
Doya, K. (2002). Metalearning and neuromodulation. Neural Networks, 15(4-6), 495–506. doi:10.1016/S0893-6080(02)00044-8
Fehr, E., & Rockenbach, B. (2004). Human altruism: economic, neural, and evolutionary perspectives.Current Opinion in Neurobiology, 14(6), 784–90. doi:10.1016/j.conb.2004.10.007
Folk, G. E. J., & Long, J. P. (1988). Serotonin as a neurotransmitter: a review. Comparative Biochemistry and Physiology, 91(1), 251–257. Retrieved from http://www.sciencedirect.com/science/article/pii/0742841388901934
45

Krikeb, J., Serotonin and dopamine in predator-prey aggression
Gellynck, E., Heyninck, K., Andressen, K. W., Haegeman, G., Levy, F. O., Vanhoenacker, P., & Van Craenenbroeck, K. (2013). The serotonin 5-HT7 receptors: two decades of research. Experimental Brain Research, 230(4), 555–68. doi:10.1007/s00221-013-3694-y
George, D. T., Umhau, J. C., Phillips, M. J., Emmela, D., Ragan, P. W., Shoaf, S. E., & Rawlings, R. R. (2001). Serotonin, testosterone and alcohol in the etiology of domestic violence. Psychiatry Research, 104(1), 27–37. doi:10.1016/S0165-1781(01)00292-X
Glimcher, P. W. (2011). Understanding dopamine and reinforcement learning: the dopamine reward prediction error hypothesis. Proceedings of the National Academy of Sciences of the United States of America, 108(suppl 3), 15647–15654. doi:10.1073/pnas.1115170108
Gowin, J. L., Swann, A. C., Moeller, F. G., & Lane, S. D. (2010). Zolmitriptan and human aggression: interaction with alcohol. Psychopharmacology, 210(4), 521–31. doi:10.1007/s00213-010-1851-6
Higley, J., King, Jr, S., Hasert, M., Champoux, M., Suomi, S., & Linnoila, M. (1996). Stability of interindividual differences in serotonin function and its relationship to severe aggression and competent social behavior in rhesus macaque females. Neuropsychopharmachology, 14(1), 67–76. Retrieved from http://www.sciencedirect.com/science/article/pii/S0893133X96800601
Hills, D., & Joyce, C. (2013). A review of research on the prevalence, antecedents, consequences and prevention of workplace aggression in clinical medical practice. Aggression and Violent Behavior, 18(5), 554–569. doi:10.1016/j.avb.2013.07.014
Homberg, J. R. (2012). Serotonin and decision making processes. Neuroscience and Biobehavioral Reviews, 36(1), 218–236. doi:10.1016/j.neubiorev.2011.06.001
Ikemoto, S. (2010). Brain reward circuitry beyond the mesolimbic dopamine system: a neurobiological theory. Neuroscience and Biobehavioral Reviews, 35(2), 129–50. doi:10.1016/j.neubiorev.2010.02.001
Insko, C. A., Schopler, J., Hoyle, R. H., Dardis, G. J., & Graetz, K. A. (1990). Individual-group discontinuity as a function of fear and greed. Journal of Personality and Social Psychology, 58(1), 68–79. doi:10.1037/0022-3514.58.1.68
Israel, S., Weisel, O., Ebstein, R. P., & Bornstein, G. (2012). Oxytocin, but not vasopressin, increases both parochial and universal altruism. Psychoneuroendocrinology, 37(8), 1341–4. doi:10.1016/j.psyneuen.2012.02.001
Jouvet, M. (1999). Sleep and serotonin: an unfinished story. Neuropsychopharmacology : Official Publication of the American College of Neuropsychopharmacology, 21(2 Suppl), 24S–27S. doi:10.1016/S0893-133X(99)00009-3
Kassinove, H., Roth, D., Owens, S. G., & Fuller, J. R. (2002). Effects of trait anger and anger expressionstyle on competitive attack responses in a wartime prisoner’s dilemma game. Aggressive Behavior, 28(2), 117–125. doi:10.1002/ab.90013
46

Krikeb, J., Serotonin and dopamine in predator-prey aggression
Kiser, D., Steemers, B., Branchi, I., & Homberg, J. R. (2012). The reciprocal interaction between serotonin and social behaviour. Neuroscience and Biobehavioral Reviews, 36(2), 786–798. doi:10.1016/j.neubiorev.2011.12.009
Klumpp, H., Angstadt, M., Nathan, P. J., & Phan, K. L. (2010). Amygdala reactivity to faces at varying intensities of threat in generalized social phobia: an event-related functional MRI study. Psychiatry Research, 183(2), 167–9. doi:10.1016/j.pscychresns.2010.05.001
Li, C., Dabrowska, J., Hazra, R., & Rainnie, D. G. (2011). Synergistic activation of dopamine D1 and TrkB receptors mediate gain control of synaptic plasticity in the basolateral amygdala. PloS One, 6(10), e26065. doi:10.1371/journal.pone.0026065
Linnoila, M., Virkkunen, M., & Scheinin, M. (1983). Low cerebrospinal fluid 5-hydroxyindoleacetic acid concentration differentiates impulsive from nonimpulsive violent behavior. Life Sciences, 33(26), 2609–2614. Retrieved from http://www.sciencedirect.com/science/article/pii/0024320583903442
Lladó-Pelfort, L., Santana, N., Ghisi, V., Artigas, F., & Celada, P. (2012). 5-HT1A receptor agonists enhance pyramidal cell firing in prefrontal cortex through a preferential action on GABA interneurons. Cerebral Cortex (New York, N.Y. : 1991), 22(7), 1487–1497. doi:10.1093/cercor/bhr220
Lovinger, D. M. (2010). Neurotransmitter roles in synaptic modulation, plasticity and learning in the dorsal striatum. Neuropharmacology, 58(7), 951–961. doi:10.1016/j.neuropharm.2010.01.008
Malone, R. P., Bennett, D. S., Luebbert, J. F., Rowan, A. B., Biesecker, K. A., Blaney, B. L., & Delaney, M. A. (1998). Aggression classification and treatment response. Psychopharmacology Bulletin, 34(1), 41–45. Retrieved from http://europepmc.org/abstract/med/9564197
Martin, J. M., Juvina, I., Lebiere, C., & Gonzalez, C. (2013). The effects of individual and context on aggression in repeated social interaction. Applied Ergonomics, 44(5), 710–718. doi:10.1016/j.apergo.2012.04.014
Martinowich, K., & Lu, B. (2008). Interaction between BDNF and serotonin: role in mood disorders. Neuropsychopharmacology : Official Publication of the American College of Neuropsychopharmacology, 33(1), 73–83. doi:10.1038/sj.npp.1301571
McDermott, R., Tingley, D., Cowden, J., Frazzetto, G., & Johnson, D. D. P. (2009). Monoamine oxidaseA gene (MAOA) predicts behavioral aggression following provocation. Proceedings of the National Academy of Sciences of the United States of America, 106(7), 2118–2123. doi:10.1073/pnas.0808376106
McEllistrem, J. E. (2004). Affective and predatory violence: A bimodal classification system of human aggression and violence. Aggression and Violent Behavior, 10(1), 1–30. doi:10.1016/j.avb.2003.06.002
47

Krikeb, J., Serotonin and dopamine in predator-prey aggression
Mehta, P. H., & Beer, J. (2010). Neural mechanisms of the testosterone–aggression relation: The roleof orbitofrontal cortex. Journal of Cognitive Neuroscience, 22(10), 2357–2368. Retrieved from http://www.mitpressjournals.org/doi/abs/10.1162/jocn.2009.21389
Murphy, S. E., Longhitano, C., Ayres, R. E., Cowen, P. J., Harmer, C. J., & Rogers, R. D. (2009). The Roleof Serotonin in Nonnormative Risky Choice: The Effects of Tryptophan Supplements on the “‘Reflection Effect’” in Healthy Adult Volunteers. Journal of Cognitive Neuroscience, 21(9), 1709–1719.
Narayanan, N. S., Land, B. B., Solder, J. E., Deisseroth, K., & DiLeone, R. J. (2012). Prefrontal D1 dopamine signaling is required for temporal control. Proceedings of the National Academy of Sciences of the United States of America, 109(50), 20726–31. doi:10.1073/pnas.1211258109
Nardo, D., Högberg, G., Looi, J. C. L., Larsson, S., Hällström, T., & Pagani, M. (2010). Gray matter density in limbic and paralimbic cortices is associated with trauma load and EMDR outcome in PTSD patients. Journal of Psychiatric Research, 44(7), 477–85. doi:10.1016/j.jpsychires.2009.10.014
Navailles, S., & De Deurwaerdère, P. (2011). Presynaptic control of serotonin on striatal dopamine function. Psychopharmacology, 213(2-3), 213–242. doi:10.1007/s00213-010-2029-y
Nazir, M., & Afza, T. (2009). Impact of aggressive working capital management policy on firms’ profitability. The IUP Journal of Applied Finance, 15(8), 19–31. Retrieved from http://www.stieykpn.ac.id/images/artikel/Presentasi 5 - Impact of Aggresive Working Capital Management Policy on Firms Profitability.pdf
Nelson, R. J., & Trainor, B. C. (2007). Neural mechanisms of aggression. Nature Reviews Neuroscience, 8(July), 536–546. doi:10.1038/nrn2174
Okai, D., Samuel, M., Askey-Jones, S., David, a S., & Brown, R. G. (2011). Impulse control disorders and dopamine dysregulation in Parkinson’s disease: a broader conceptual framework. European Journal of Neurology : The Official Journal of the European Federation of NeurologicalSocieties, 18(12), 1379–1383. doi:10.1111/j.1468-1331.2011.03432.x
Pagani, M., Di Lorenzo, G., Verardo, A. R., Nicolais, G., Monaco, L., Lauretti, G., … Siracusano, A. (2012). Neurobiological correlates of EMDR monitoring - an EEG study. PloS One, 7(9), e45753. doi:10.1371/journal.pone.0045753
Pehek, E. a, Nocjar, C., Roth, B. L., Byrd, T. a, & Mabrouk, O. S. (2006). Evidence for the preferential involvement of 5-HT2A serotonin receptors in stress- and drug-induced dopamine release in the rat medial prefrontal cortex. Neuropsychopharmacology : Official Publication of the American College of Neuropsychopharmacology, 31(2), 265–277. doi:10.1038/sj.npp.1300819
Perez, P. R. (2012). The etiology of psychopathy: A neuropsychological perspective. Aggression and Violent Behavior, 17(6), 519–522. doi:10.1016/j.avb.2012.07.006
48

Krikeb, J., Serotonin and dopamine in predator-prey aggression
Piquero, N. L., Piquero, A. R., Craig, J. M., & Clipper, S. J. (2013). Assessing research on workplace violence, 2000–2012. Aggression and Violent Behavior, 18(3), 383–394. doi:10.1016/j.avb.2013.03.001
Post, R. M., Weiss, S. R., Li, H., Smith, M. a, Zhang, L. X., Xing, G., … McCann, U. D. (1998). Neural plasticity and emotional memory. Development and Psychopathology, 10(4), 829–55. Retrievedfrom http://www.ncbi.nlm.nih.gov/pubmed/21204425
Rezayof, A., Hosseini, S.-S., & Zarrindast, M.-R. (2009). Effects of morphine on rat behaviour in the elevated plus maze: the role of central amygdala dopamine receptors. Behavioural Brain Research, 202(2), 171–178. doi:10.1016/j.bbr.2009.03.030
Roberts, A. C. (2011). The importance of serotonin for orbitofrontal function. Biological Psychiatry, 69(12), 1185–1191. doi:10.1016/j.biopsych.2010.12.037
Rogers, R. D. (2011). The roles of dopamine and serotonin in decision making: evidence from pharmacological experiments in humans. Neuropsychopharmacology : Official Publication of the American College of Neuropsychopharmacology, 36(1), 114–132. doi:10.1038/npp.2010.165
Rubia, K., Lee, F., Cleare, A. J., Tunstall, N., Fu, C. H. Y., Brammer, M., & McGuire, P. (2005). Tryptophan depletion reduces right inferior prefrontal activation during response inhibition in fast, event-related fMRI. Psychopharmacology, 179(4), 791–803. doi:10.1007/s00213-004-2116-z
Saha, S., Gamboa-Esteves, F. O., & Batten, T. F. C. (2010). Differential distribution of 5-HT 1A and 5-HT 1B-like immunoreactivities in rat central nucleus of the amygdala neurones projecting to thecaudal dorsomedial medulla oblongata. Brain Research, 1330, 20–30. doi:10.1016/j.brainres.2010.03.009
Scarnà, A., McTavish, S. F. B., Cowen, P. J., Goodwin, G. M., & Rogers, R. D. (2005). The effects of a branched chain amino acid mixture supplemented with tryptophan on biochemical indices of neurotransmitter function and decision-making. Psychopharmacology, 179, 761–768. doi:10.1007/s00213-004-2105-2
Scholes, K. E., Harrison, B. J., O’Neill, B. V, Leung, S., Croft, R. J., Pipingas, A., … Nathan, P. J. (2007). Acute serotonin and dopamine depletion improves attentional control: findings from the stroop task. Neuropsychopharmacology : Official Publication of the American College of Neuropsychopharmacology, 32(7), 1600–1610. doi:10.1038/sj.npp.1301262
Segura-Aguilar, J., Paris, I., Muñoz, P., Ferrari, E., Zecca, L., & Zucca, F. a. (2014). Protective and toxic roles of dopamine in Parkinson’s disease. Journal of Neurochemistry, 129(6), 898–915. doi:10.1111/jnc.12686
Seo, D., Patrick, C. J., & Kennealy, P. J. (2008). Role of Serotonin and Dopamine System Interactions in the Neurobiology of Impulsive Aggression and its Comorbidity with other Clinical Disorders. Aggression and Violent Behavior, 13(5), 383–395. doi:10.1016/j.avb.2008.06.003
49

Krikeb, J., Serotonin and dopamine in predator-prey aggression
Siegel, A., & Victoroff, J. (2009). Understanding human aggression: New insights from neuroscience. International Journal of Law and Psychiatry, 32(4), 209–215. doi:10.1016/j.ijlp.2009.06.001
Skara, S., Pokhrel, P., Weiner, M. D., Sun, P., Dent, C. W., & Sussman, S. (2008). Physical and relational aggression as predictors of drug use: gender differences among high school students.Addictive Behaviors, 33(12), 1507–15. doi:10.1016/j.addbeh.2008.05.014
Smith, E. S., Geissler, S. a, Schallert, T., & Lee, H. J. (2013). The role of central amygdala dopamine in disengagement behavior. Behavioral Neuroscience, 127(2), 164–174. doi:10.1037/a0031043
Smith, S. F., & Lilienfeld, S. O. (2013). Psychopathy in the workplace: The knowns and unknowns. Aggression and Violent Behavior, 18(2), 204–218. doi:10.1016/j.avb.2012.11.007
Soler, H., Vinayak, P., & Quadagno, D. (2000). Biosocial aspects of domestic violence. Psychoneuroendocrinology, 25(7), 721–739. doi:10.1016/S0306-4530(00)00022-6
Stein, C., Davidowa, H., & Albrecht, D. (2000). 5-HT1A Receptor-Mediated Inhibition and 5-HT2 As Well As 5-HT3 Receptor-Mediated Excitation in Different Subdivisions of the Rat Amygdala. Synapse, 38, 328–337.
Takahashi, A., Quadros, I. M., de Almeida, R. M. M., & Miczek, K. a. (2011). Brain serotonin receptors and transporters: initiation vs. termination of escalated aggression. Psychopharmacology, 213(2-3), 183–212. doi:10.1007/s00213-010-2000-y
Tversky, A., & Kahneman, D. (1981). The framing of decisions and the psychology of choice. Science , 211 (4481 ), 453–458. doi:10.1126/science.7455683
Umukoro, S., Aladeokin, A. C., & Eduviere, A. T. (2013). Aggressive behavior: A comprehensive review of its neurochemical mechanisms and management. Aggression and Violent Behavior, 18(2), 195–203. doi:10.1016/j.avb.2012.11.002
Vitiello, B., Behar, D., Hunt, J., Stoff, D., & Ricciuti, A. (1990). Subtyping Aggression in Children and Adolescents. Journal of Neuropsychiatry, 2(2), 189–192.
Volkow, N. D., Wang, G.-J., Fowler, J. S., Tomasi, D., & Telang, F. (2011). Addiction: beyond dopaminereward circuitry. Proceedings of the National Academy of Sciences of the United States of America, 108(37), 15037–42. doi:10.1073/pnas.1010654108
Vukhac, K., Sankoorikal, E., & Wang, Y. (2001). Dopamine D2L receptor-and age-related reduction in offensive aggression. Neuroreport, 12(5), 1035–1038. Retrieved from http://journals.lww.com/neuroreport/Abstract/2001/04170/Dopamine_D2L_receptor__and_age_related_reduction.34.aspx
Weinshenker, N. J., & Siegel, A. (2002). Bimodal classification of aggression: affective defense and predatory attack. Aggression and Violent Behavior, 7(3), 237–250. doi:10.1016/S1359-1789(01)00042-8
50

Krikeb, J., Serotonin and dopamine in predator-prey aggression
Wetzler, S., Kahn, R., Asnis, G. M., Korn, M., & van Praag, H. M. (1991). Serotonin receptor sensitivityand aggression. Psychiatry Research, 37(3), 271–279. doi:10.1016/0165-1781(91)90063-U
Winstanley, C. a, Theobald, D. E. H., Cardinal, R. N., & Robbins, T. W. (2004). Contrasting roles of basolateral amygdala and orbitofrontal cortex in impulsive choice. The Journal of Neuroscience : The Official Journal of the Society for Neuroscience, 24(20), 4718–4722. doi:10.1523/JNEUROSCI.5606-03.2004
Winstanley, C. a., Theobald, D. E. H., Dalley, J. W., Cardinal, R. N., & Robbins, T. W. (2006). Double dissociation between serotonergic and dopaminergic modulation of medial prefrontal and orbitofrontal cortex during a test of impulsive choice. Cerebral Cortex, 16(January), 106–114. doi:10.1093/cercor/bhi088
Wise, R. a. (2009). Roles for nigrostriatal--not just mesocorticolimbic--dopamine in reward and addiction. Trends in Neurosciences, 32(10), 517–24. doi:10.1016/j.tins.2009.06.004
Wood, R. M., Rilling, J. K., Sanfey, A. G., Bhagwagar, Z., & Rogers, R. D. (2006). Effects of tryptophan depletion on the performance of an iterated Prisoner’s Dilemma game in healthy adults. Neuropsychopharmacology : Official Publication of the American College of Neuropsychopharmacology, 31(5), 1075–1084. doi:10.1038/sj.npp.1300932
Yi, R., Johnson, M. W., & Bickel, W. K. (2005). Relationship between cooperation in an iterated prisoner’s dilemma game and the discounting of hypothetical outcomes. Learning & Behavior, 33(3), 324–336. doi:10.3758/BF03192861
Yoon, K. L., Fitzgerald, D. a, Angstadt, M., McCarron, R. a, & Phan, K. L. (2007). Amygdala reactivity toemotional faces at high and low intensity in generalized social phobia: a 4-Tesla functional MRI study. Psychiatry Research, 154(1), 93–8. doi:10.1016/j.pscychresns.2006.05.004
51
Related Documents











