General and Comparative Endocrinology 142 (2005) 186–192 www.elsevier.com/locate/ygcen 0016-6480/$ - see front matter 2005 Elsevier Inc. All rights reserved. doi:10.1016/j.ygcen.2004.12.013 Ontogeny of the thyroid hormones and cortisol in the gilthead sea bream, Sparus aurata V. Szisch a , N. Papandroulakis b , E. Fanouraki a , M. Pavlidis a,b,¤ a Department of Biology, University of Crete, P.O. Box 2208, GR-71409 Heraklion, Crete, Greece b Hellenic Centre for Marine Research, Institute of Aquaculture, P.O. Box 2214, GR-71003 Heraklion, Crete, Greece Received 24 September 2004; accepted 20 December 2004 Available online 1 February 2005 Abstract The aim of the study was to elucidate the pattern of changes in the whole body concentrations of thyroid hormones (TH) and cor- tisol in eggs, pre-larvae, and larvae in the gilthead sea bream, Sparus aurata. The ontogeny of these hormones was related to speciWc morphological characteristics that characterize early development. TH and cortisol were detected in substantial amounts in the eggs of sea bream. T 3 concentration in eggs was found to be higher than that of T 4 (T 3 , 4.02 § 0.38 ng g ¡1 ; T 4 , 1.63 § 0.28 ng g ¡1 ), as it is often observed in marine teleosts. T 4 decreased until hatching (T 4 , 1.01 § 0.58 ng g ¡1 ), while T 3 reached a minimum at the time of Wrst feeding (T 3 , 0.19 § 0.02 ng g ¡1 ). As development proceeded, both hormones displayed a similar pattern of changes with increasing concentrations until the completion of metamorphosis (T 3 , 15.84 § 2.92 ng g ¡1 ; T 4 , 9.78 § 2.39 ng g ¡1 ), indicating that the hypothal- amo–adenohypophysial–thyroid axis begins to function soon after Wrst feeding. The pattern of changes of cortisol (F) concentration was almost parallel to that of T 4 , starting from 0.83 ng g ¡1 in eggs. Minimum F concentration was observed at hatching (0.39 § 0.03 ng g ¡1 ) and maximum at melanophores and scale formation (14.82 § 2.71 and 14.82 § 3.12 ng g ¡1 , respectively). Results provide data for the Wrst time on the ontogeny of thyroid hormones and cortisol during the early development in sea bream and are in agreement with results in other Wsh species for an important action of TH during early development. 2005 Elsevier Inc. All rights reserved. Keywords: Cortisol; Gilthead sea bream; Larvae; Ontogeny; Sparus aurata; Thyroid hormones 1. Introduction The thyroid gland produces and secretes two thyroid hormones (TH), namely triiodothyronine (T 3 ) and thy- roxine (T 4 ), which regulates growth, development and diVerentiation, metabolism, and the maintenance of homeostasis in vertebrates. Today it is well known that thyroid hormones play an important role during embryogenesis and organogenesis in all vertebrates and that most vertebrates are unable to grow and reach their normal adult form without them (Liu et al., 2000; Oppenheimer and Schwartz, 1997; Power et al., 2001). Compared to other vertebrates, anuran development is perhaps the most dependent on TH, since metamorpho- sis in amphibians is regulated mainly by TH action and is blocked by thyroidectomy (Jones et al., 2002; Sachs et al., 2000; Wright, 2002). Despite the diversity of teleost Wsh, most undergo similar developmental phases, which can be divided generally into larva, juvenile, and adult. Larvae are dis- tinct both morphologically and physiologically from juveniles and adults. The transformation of larvae into juveniles is frequently termed metamorphosis, the most dramatic manifestation of which is the classic XatWsh ¤ Corresponding author. Fax: +30 2810 394408. E-mail address: [email protected] (M. Pavlidis).

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

General and Comparative Endocrinology 142 (2005) 186–192
www.elsevier.com/locate/ygcen
Ontogeny of the thyroid hormones and cortisol in the gilthead sea bream, Sparus aurata
V. Szischa, N. Papandroulakisb, E. Fanourakia, M. Pavlidisa,b,¤
a Department of Biology, University of Crete, P.O. Box 2208, GR-71409 Heraklion, Crete, Greeceb Hellenic Centre for Marine Research, Institute of Aquaculture, P.O. Box 2214, GR-71003 Heraklion, Crete, Greece
Received 24 September 2004; accepted 20 December 2004Available online 1 February 2005
Abstract
The aim of the study was to elucidate the pattern of changes in the whole body concentrations of thyroid hormones (TH) and cor-tisol in eggs, pre-larvae, and larvae in the gilthead sea bream, Sparus aurata. The ontogeny of these hormones was related to speciWcmorphological characteristics that characterize early development. TH and cortisol were detected in substantial amounts in the eggsof sea bream. T3 concentration in eggs was found to be higher than that of T4 (T3, 4.02 § 0.38 ng g¡1; T4, 1.63 § 0.28 ng g¡1), as it isoften observed in marine teleosts. T4 decreased until hatching (T4, 1.01 § 0.58 ng g¡1), while T3 reached a minimum at the time of Wrstfeeding (T3, 0.19 § 0.02 ng g¡1). As development proceeded, both hormones displayed a similar pattern of changes with increasingconcentrations until the completion of metamorphosis (T3, 15.84 § 2.92 ng g¡1; T4, 9.78 § 2.39 ng g¡1), indicating that the hypothal-amo–adenohypophysial–thyroid axis begins to function soon after Wrst feeding. The pattern of changes of cortisol (F) concentrationwas almost parallel to that of T4, starting from 0.83 ng g¡1 in eggs. Minimum F concentration was observed at hatching(0.39 § 0.03 ng g¡1) and maximum at melanophores and scale formation (14.82 § 2.71 and 14.82 § 3.12 ng g¡1, respectively). Resultsprovide data for the Wrst time on the ontogeny of thyroid hormones and cortisol during the early development in sea bream and arein agreement with results in other Wsh species for an important action of TH during early development. 2005 Elsevier Inc. All rights reserved.
Keywords: Cortisol; Gilthead sea bream; Larvae; Ontogeny; Sparus aurata; Thyroid hormones
1. Introduction
The thyroid gland produces and secretes two thyroidhormones (TH), namely triiodothyronine (T3) and thy-roxine (T4), which regulates growth, development anddiVerentiation, metabolism, and the maintenance ofhomeostasis in vertebrates. Today it is well known thatthyroid hormones play an important role duringembryogenesis and organogenesis in all vertebrates andthat most vertebrates are unable to grow and reach their
¤ Corresponding author. Fax: +30 2810 394408.E-mail address: [email protected] (M. Pavlidis).
0016-6480/$ - see front matter 2005 Elsevier Inc. All rights reserved.doi:10.1016/j.ygcen.2004.12.013
normal adult form without them (Liu et al., 2000;Oppenheimer and Schwartz, 1997; Power et al., 2001).Compared to other vertebrates, anuran development isperhaps the most dependent on TH, since metamorpho-sis in amphibians is regulated mainly by TH action andis blocked by thyroidectomy (Jones et al., 2002; Sachs etal., 2000; Wright, 2002).
Despite the diversity of teleost Wsh, most undergosimilar developmental phases, which can be dividedgenerally into larva, juvenile, and adult. Larvae are dis-tinct both morphologically and physiologically fromjuveniles and adults. The transformation of larvae intojuveniles is frequently termed metamorphosis, the mostdramatic manifestation of which is the classic XatWsh

V. Szisch et al. / General and Comparative Endocrinology 142 (2005) 186–192 187
metamorphosis (De Jesus et al., 1998; Huang et al.,1998; Inui et al., 1995; Pittman et al., 1998; Yamanoet al., 1991). This developmental process appears to beregulated by TH in a way comparable to amphibianmetamorphosis (Sachs et al., 2000; Tata, 1999; Wrightand Alves, 2001). Beside the direct action of TH, theyare often also required in a permissive role, assistingthe action of other hormones, as growth hormone, pro-lactin, and the corticosteroid cortisol, in the processesinvolved. Peripheral thyroid–corticoid interactions arealso well established in the regulation of amphibianmetamorphosis (Brown and Kim, 1995).
The presence of thyroid hormones and cortisol in tel-eost eggs, prior to hatching, has been reported for manyfreshwater, marine, and diadromous Wsh species, andthey are considered to be of maternal origin (Berry et al.,1995; De Jesus and Hirano, 1992; De Jesus et al., 1991;Hwang et al., 1992; Lam, 1994, 1995; Mylonas et al.,1994; Tagawa and Hirano, 1987).
The aim of this study was to determine the ontogenyof thyroid hormones and cortisol in the gilthead seabream (Sparus aurata), a model organism for Mediter-ranean mariculture, and to correlate the hormone con-centrations with the speciWc morphologicalcharacteristics of each developmental phase investi-gated in order to elucidate their function in early devel-opment in teleosts.
2. Materials and methods
2.1. Larval culture
Fertilized eggs were obtained from spontaneousspawning from three diVerent broodstocks held in cap-tivity and transferred to the intensive hatchery of theInstitute of Aquaculture of the Hellenic Center forMarine Research (Heraklion, Crete). Larval rearingwas performed applying the pseudo-green-water tech-nique (Papandroulakis et al., 2001) which comprisestwo phases: (1) the initial phase which lasted 40 days, in500-L cylindro-conical tanks, with daily supplementa-tion of phytoplankton and zooplankton and (2) thepre-weaning phase (days 41–75 post-fertilization) in2000-L cylindro-conical tanks, during which a progres-sive transition to clear water was implemented. Duringthe initial phase, larvae were fed exclusively on enrichedrotifers until day 15 post-fertilization and on a mixeddiet of rotifers and Artemia until the end of the initialphase. During the pre-weaning phase feeding was ini-tially based on Artemia and after day 65 larvae wereweaned onto artiWcial feeds. A sample of 20 larvae wastaken every day for determining the morphologicalcharacteristics and total length. For thyroid hormoneand cortisol determination 4–5 samples per diVerentdevelopmental phase were taken, the wet weight was
measured and samples were stored in ¡20 °C until theday of analysis.
2.2. Thyroid hormone analysis
For thyroid hormone extraction the protocol of Tag-awa and Hirano (1987) was modiWed as follows: 0.5–1 gof larval samples was homogenized in 5 ml ice-cold meth-anolic ammonia (99:1 v/v) containing 1 mM PTU andshaken in horizontal position for 10 min at 4 °C. Thehomogenate was then centrifuged for 15 min at 2000g,4 °C and the extraction was repeated twice. The superna-tants were pooled and evaporated in a vacuum dryer(Savant AES 2010) overnight at 45 °C. The residue wasre-suspended in 3.5 ml methanolic ammonia-PTU con-taining chloroform (1:6 v/v) and 0.5 barbital buVer(50 mM, pH 8.6). Samples were shaken in horizontal posi-tion for 10 min in room temperature and the upper layer,which contained the thyroid hormones, was isolated andevaporated for 4 h at 45 °C. Dried extracts were dissolvedin barbital buVer containing 0.1% BSA. Aliquots of 50�for T3 and 25� for T4 determination were taken for anal-ysis. The total body concentration of T3, T4 was deter-mined using coated tube radioimmunoassay kits (T3Spectria, Orion Diagnostica; Canine T4, DPC). Recoverywas 94.3 § 6.9% for T3 and 60.1 § 13.9% for T4. Dilutionseries of extract from pooled samples (n D 3; egg, Wrstfeeding, Xexion) showed a very good parallelism (T3,r2 D 0.9958; T4, r2 D 0.9968). Results are presented inng g¡1 larval wet mass, following recovery corrections.
2.3. Cortisol analysis
Cortisol extraction was performed according to DeJesus et al. (1991). RIA was performed following theprotocol of WHO Reagent Programme (SuW et al., 1994)following modiWcation. BrieXy, aliquots of 100� wereassayed with 100� 3H tracer solution (Amersham), 100�of antiserum solution (rabbit anti-cortisol, Chemicon)and 400� of buVer S. Dextran-charcoal was used for sep-aration of free from antibody-bound hormone. Recov-ery was 86% and pooled samples (n D 3; egg, Wrstfeeding, Xexion) showed good parallelism (r2 D 0.9874).Results are presented in ng g¡1 larval wet mass, follow-ing recovery corrections.
2.4. Statistics
All statistic analyses were performed with SigmaStat2.0 (Jandel ScientiWc). Statistical errors were expressedas the standard error of mean (SEM). Data were ana-lysed for normality and homogeneity. Statistical com-parisons of each respective hormone concentrationbetween the diVerent developmental phases were madeusing one-way ANOVA. The signiWcance level used wasP < 0.05.
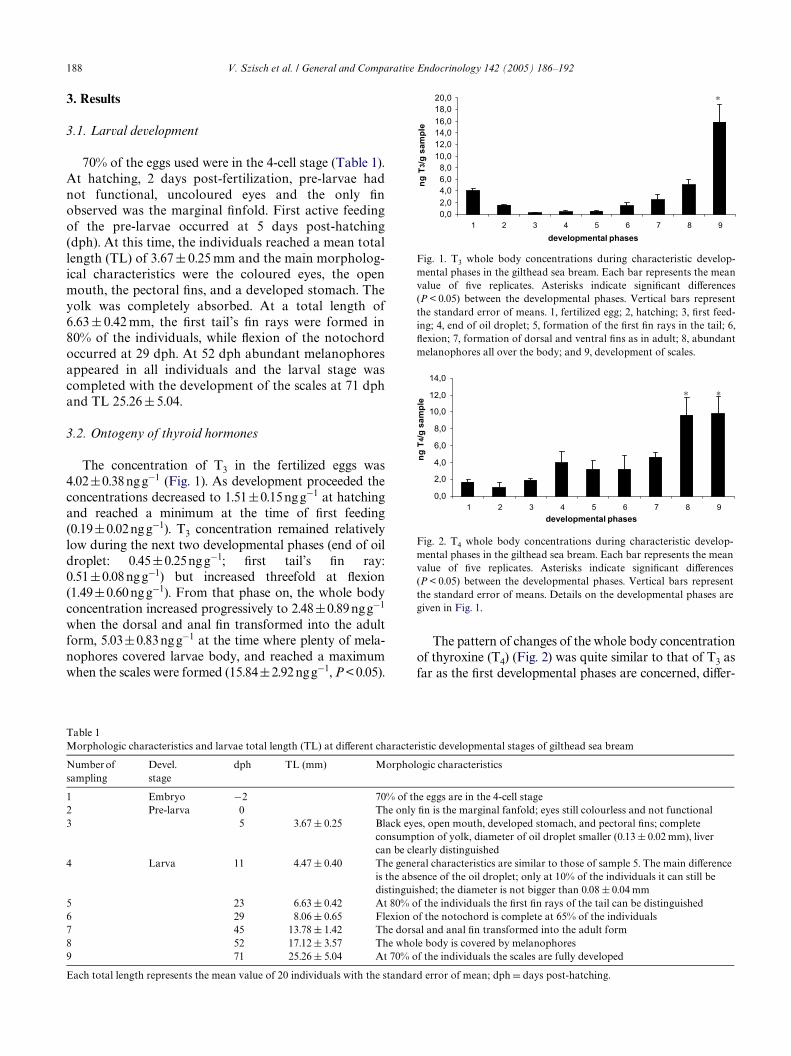
188 V. Szisch et al. / General and Comparative Endocrinology 142 (2005) 186–192
3. Results
3.1. Larval development
70% of the eggs used were in the 4-cell stage (Table 1).At hatching, 2 days post-fertilization, pre-larvae hadnot functional, uncoloured eyes and the only Wnobserved was the marginal Wnfold. First active feedingof the pre-larvae occurred at 5 days post-hatching(dph). At this time, the individuals reached a mean totallength (TL) of 3.67 § 0.25 mm and the main morpholog-ical characteristics were the coloured eyes, the openmouth, the pectoral Wns, and a developed stomach. Theyolk was completely absorbed. At a total length of6.63 § 0.42 mm, the Wrst tail’s Wn rays were formed in80% of the individuals, while Xexion of the notochordoccurred at 29 dph. At 52 dph abundant melanophoresappeared in all individuals and the larval stage wascompleted with the development of the scales at 71 dphand TL 25.26 § 5.04.
3.2. Ontogeny of thyroid hormones
The concentration of T3 in the fertilized eggs was4.02§0.38 ng g¡1 (Fig. 1). As development proceeded theconcentrations decreased to 1.51§0.15 ng g¡1 at hatchingand reached a minimum at the time of Wrst feeding(0.19§0.02 ng g¡1). T3 concentration remained relativelylow during the next two developmental phases (end of oildroplet: 0.45§0.25ng g¡1; Wrst tail’s Wn ray:0.51§0.08 ng g¡1) but increased threefold at Xexion(1.49§0.60 ng g¡1). From that phase on, the whole bodyconcentration increased progressively to 2.48§0.89 ngg¡1
when the dorsal and anal Wn transformed into the adultform, 5.03§0.83 ng g¡1 at the time where plenty of mela-nophores covered larvae body, and reached a maximumwhen the scales were formed (15.84§2.92 ngg¡1, P< 0.05).
The pattern of changes of the whole body concentrationof thyroxine (T4) (Fig. 2) was quite similar to that of T3 asfar as the Wrst developmental phases are concerned, diVer-
Fig. 1. T3 whole body concentrations during characteristic develop-mental phases in the gilthead sea bream. Each bar represents the meanvalue of Wve replicates. Asterisks indicate signiWcant diVerences(P < 0.05) between the developmental phases. Vertical bars representthe standard error of means. 1, fertilized egg; 2, hatching; 3, Wrst feed-ing; 4, end of oil droplet; 5, formation of the Wrst Wn rays in the tail; 6,Xexion; 7, formation of dorsal and ventral Wns as in adult; 8, abundantmelanophores all over the body; and 9, development of scales.
Fig. 2. T4 whole body concentrations during characteristic develop-mental phases in the gilthead sea bream. Each bar represents the meanvalue of Wve replicates. Asterisks indicate signiWcant diVerences(P < 0.05) between the developmental phases. Vertical bars representthe standard error of means. Details on the developmental phases aregiven in Fig. 1.
Table 1Morphologic characteristics and larvae total length (TL) at diVerent characteristic developmental stages of gilthead sea bream
Each total length represents the mean value of 20 individuals with the standard error of mean; dph D days post-hatching.
Number of sampling
Devel. stage
dph TL (mm) Morphologic characteristics
1 Embryo ¡2 70% of the eggs are in the 4-cell stage2 Pre-larva 0 The only Wn is the marginal fanfold; eyes still colourless and not functional3 5 3.67 § 0.25 Black eyes, open mouth, developed stomach, and pectoral Wns; complete
consumption of yolk, diameter of oil droplet smaller (0.13 § 0.02 mm), liver can be clearly distinguished
4 Larva 11 4.47 § 0.40 The general characteristics are similar to those of sample 5. The main diVerence is the absence of the oil droplet; only at 10% of the individuals it can still be distinguished; the diameter is not bigger than 0.08 § 0.04 mm
5 23 6.63 § 0.42 At 80% of the individuals the Wrst Wn rays of the tail can be distinguished6 29 8.06 § 0.65 Flexion of the notochord is complete at 65% of the individuals7 45 13.78 § 1.42 The dorsal and anal Wn transformed into the adult form8 52 17.12 § 3.57 The whole body is covered by melanophores9 71 25.26 § 5.04 At 70% of the individuals the scales are fully developed

V. Szisch et al. / General and Comparative Endocrinology 142 (2005) 186–192 189
ing only in the timing of the Wrst increase. In particular,starting from 1.63§0.28ngg¡1 in eggs the concentrationdecreased to a minimum at hatching (1.01§0.58ngg¡1). Inopposite to the pattern of T3 changes, the concentration ofthyroxine in larvae started to increase at Wrst feeding(1.86§0.25ngg¡1), until the complete consumption of theoil droplet where a concentration of 4.00§1.36ngg¡1 wasfound. T4 remained quite stable during the next two devel-opmental phases; appearance of the tail’s Wrst Wn ray(3.20§0.98ngg¡1) and Xexion (3.21§1.66ngg¡1). AfterXexion of the notochord the whole body concentration ofT4 raised constantly until the time when the dorsal andanal Wn transformed into the adult form (4.65§0.57ngg¡1). As the development of the larvae proceeded,T4 continued to rise reaching a concentration twice as high(9.59§2.17ngg¡1) when abundant melanophores occurredall over the body. At the last phase (formation of scales) T4remained high (9.78§2.39ngg¡1). The thyroxine concen-trations of the last two phases diVered signiWcantly fromthe others (P<0.05).
3.3. Ontogeny of cortisol
The pattern of changes of cortisol concentrations(Fig. 3) was almost parallel to that of T4. In fertilizedeggs, cortisol was 0.83 § 0.02 ng g¡1. Cortisol (F)decreased at hatching (0.39 § 0.03 ng g¡1) and increasedsigniWcantly to 3.40 § 0.19 ng g¡1 at Wrst feeding. Wholebody F concentration continued to rise, reaching a pla-teau at the phases where the Wrst Wn rays (7.38 §1.16 ng g¡1) and the Xexion (7.53 § 0.92 ng g¡1) of thenotochord occurred. Cortisol raised to a twofold con-centration already at the phase where the dorsal andanal Wns transform into the adult form (14.55 §2.38 ng g¡1). High concentrations were also found at thelast two phases (presence of abundant melanophores allover the body, 14.82 § 2.71 ng g¡1; formation of scales,14.82 § 3.12 ng g¡1).
Fig. 3. Cortisol whole body concentrations during characteristic devel-opmental phases in the gilthead sea bream. Each bar represents themean value of three replicates. Means with diVerent letters diVer sig-niWcantly from one another (P < 0.001). Vertical bars represent thestandard error of means. Details on the developmental phases aregiven in Fig. 1.
4. Discussion
The present results show for the Wrst time that thyroidhormones and cortisol are present in signiWcant quanti-ties in all early developmental stages of the gilthead seabream, S. aurata. Obviously, TH and cortisol content inegg is of maternal origin, since no functional thyroid tis-sue and interrenal cells are developed at embryogenesis.In general, there are signiWcant diVerences among diVer-ent species in the T4 and T3 concentrations in earlydevelopmental stages and in marine Wsh the level of T3 isusually higher than that of T4 while the opposite occursin freshwater species (Ayson and Lam, 1993; De Jesusand Hirano, 1992; De Jesus et al., 1991; Greenblatt et al.,1989; Huang et al., 1998; Hwang et al., 1992; Lam, 1994;Leatherland and Lin, 1989a,b; Mylonas et al., 1994;Reddy and Lam, 1992; Tagawa and Hirano, 1987; Tag-awa et al., 1990; Yamano et al., 1994). The fact that ingilthead sea bream the concentration of T3 is higher thanthat of T4 in fertilized eggs is in agreement with thisobservation.
The underlying mechanisms of the start of TH endog-enous synthesis, which takes place during the Wrst daysafter hatching in sea bream, seem to be connected withthe Wrst feeding of the larvae. At this stage for the Wrsttime an increase of the concentrations of thyroxine andcortisol is observed. It is worth to mention that generallythe start of active feeding coincides with the completeddevelopment of a functional mouth, stomach, gastroin-testinal tract, and functional eyes. In a recent study it hasbeen shown that the elevation of thyroid hormones inthe Wrst developmental stages of the silver sea bream,Sparus sarba, is closely related to the development of afunctional stomach and the complete yolk absorption(Deane and Woo, 2003). Other studies also support thiscorrelation by showing that immersion of larvae in T4solution promoted the diVerentiation of the gastric glandas well as the production of pepsinogen (Miwa et al.,1992). In addition, experiments where treatment withthiourea was applied showed the opposite results(Huang et al., 1998). In many species, the production ofTH coincides also with the colouration of the eyes,which happens in most of the teleost after hatching andright before Wrst feeding (Tanaka et al., 1995), as in thepresent study. Alexander et al. (2001) assumed that incoho salmon (Oncorhynchus kisutch) T3 has a directaction on the production of retinoid derivates and there-fore on the visual system function for successful switch-ing from endogenous to exogenous energy sources. Theclose relation between the onset of TH production andthe completed development of the gastrointestinal tractand functional eyes could explain the presence of mater-nal TH in the eggs, which could be utilized for the devel-opment of these systems. Former studies concluded thatin tilapia and trout the activity of the deiodinases, theenzymes which convert T4 to T3, seems to start a few
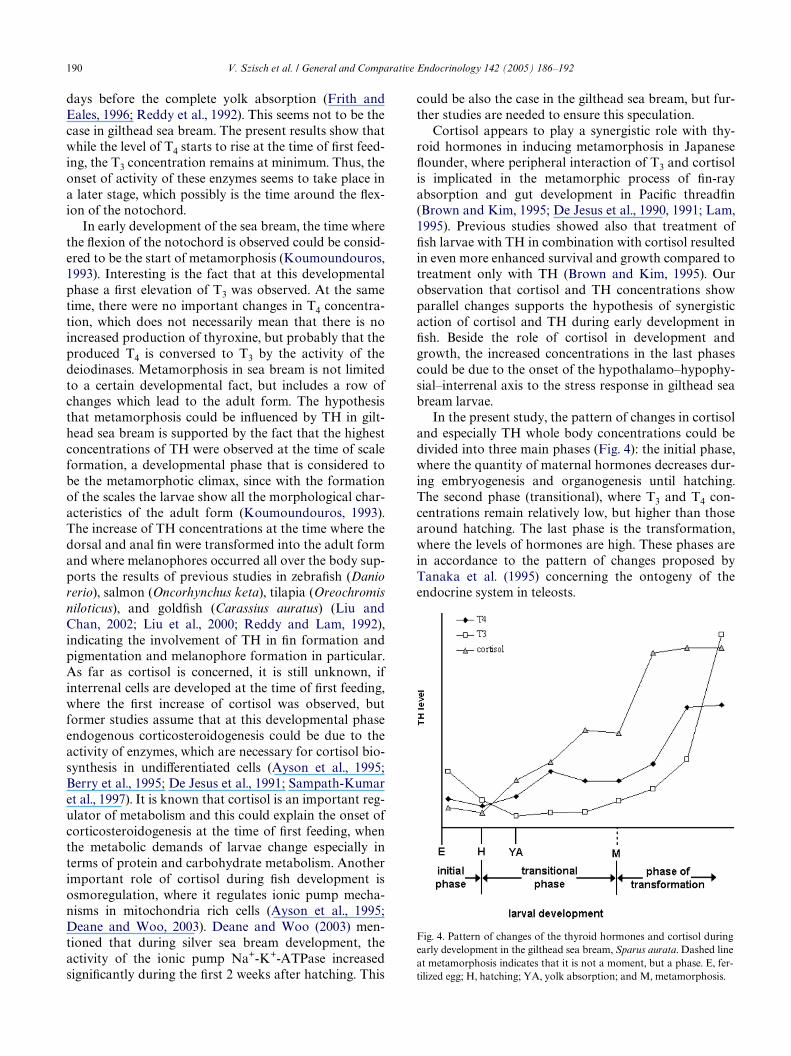
190 V. Szisch et al. / General and Comparative Endocrinology 142 (2005) 186–192
days before the complete yolk absorption (Frith andEales, 1996; Reddy et al., 1992). This seems not to be thecase in gilthead sea bream. The present results show thatwhile the level of T4 starts to rise at the time of Wrst feed-ing, the T3 concentration remains at minimum. Thus, theonset of activity of these enzymes seems to take place ina later stage, which possibly is the time around the Xex-ion of the notochord.
In early development of the sea bream, the time wherethe Xexion of the notochord is observed could be consid-ered to be the start of metamorphosis (Koumoundouros,1993). Interesting is the fact that at this developmentalphase a Wrst elevation of T3 was observed. At the sametime, there were no important changes in T4 concentra-tion, which does not necessarily mean that there is noincreased production of thyroxine, but probably that theproduced T4 is conversed to T3 by the activity of thedeiodinases. Metamorphosis in sea bream is not limitedto a certain developmental fact, but includes a row ofchanges which lead to the adult form. The hypothesisthat metamorphosis could be inXuenced by TH in gilt-head sea bream is supported by the fact that the highestconcentrations of TH were observed at the time of scaleformation, a developmental phase that is considered tobe the metamorphotic climax, since with the formationof the scales the larvae show all the morphological char-acteristics of the adult form (Koumoundouros, 1993).The increase of TH concentrations at the time where thedorsal and anal Wn were transformed into the adult formand where melanophores occurred all over the body sup-ports the results of previous studies in zebraWsh (Daniorerio), salmon (Oncorhynchus keta), tilapia (Oreochromisniloticus), and goldWsh (Carassius auratus) (Liu andChan, 2002; Liu et al., 2000; Reddy and Lam, 1992),indicating the involvement of TH in Wn formation andpigmentation and melanophore formation in particular.As far as cortisol is concerned, it is still unknown, ifinterrenal cells are developed at the time of Wrst feeding,where the Wrst increase of cortisol was observed, butformer studies assume that at this developmental phaseendogenous corticosteroidogenesis could be due to theactivity of enzymes, which are necessary for cortisol bio-synthesis in undiVerentiated cells (Ayson et al., 1995;Berry et al., 1995; De Jesus et al., 1991; Sampath-Kumaret al., 1997). It is known that cortisol is an important reg-ulator of metabolism and this could explain the onset ofcorticosteroidogenesis at the time of Wrst feeding, whenthe metabolic demands of larvae change especially interms of protein and carbohydrate metabolism. Anotherimportant role of cortisol during Wsh development isosmoregulation, where it regulates ionic pump mecha-nisms in mitochondria rich cells (Ayson et al., 1995;Deane and Woo, 2003). Deane and Woo (2003) men-tioned that during silver sea bream development, theactivity of the ionic pump Na+-K+-ATPase increasedsigniWcantly during the Wrst 2 weeks after hatching. This
could be also the case in the gilthead sea bream, but fur-ther studies are needed to ensure this speculation.
Cortisol appears to play a synergistic role with thy-roid hormones in inducing metamorphosis in JapaneseXounder, where peripheral interaction of T3 and cortisolis implicated in the metamorphic process of Wn-rayabsorption and gut development in PaciWc threadWn(Brown and Kim, 1995; De Jesus et al., 1990, 1991; Lam,1995). Previous studies showed also that treatment ofWsh larvae with TH in combination with cortisol resultedin even more enhanced survival and growth compared totreatment only with TH (Brown and Kim, 1995). Ourobservation that cortisol and TH concentrations showparallel changes supports the hypothesis of synergisticaction of cortisol and TH during early development inWsh. Beside the role of cortisol in development andgrowth, the increased concentrations in the last phasescould be due to the onset of the hypothalamo–hypophy-sial–interrenal axis to the stress response in gilthead seabream larvae.
In the present study, the pattern of changes in cortisoland especially TH whole body concentrations could bedivided into three main phases (Fig. 4): the initial phase,where the quantity of maternal hormones decreases dur-ing embryogenesis and organogenesis until hatching.The second phase (transitional), where T3 and T4 con-centrations remain relatively low, but higher than thosearound hatching. The last phase is the transformation,where the levels of hormones are high. These phases arein accordance to the pattern of changes proposed byTanaka et al. (1995) concerning the ontogeny of theendocrine system in teleosts.
Fig. 4. Pattern of changes of the thyroid hormones and cortisol duringearly development in the gilthead sea bream, Sparus aurata. Dashed lineat metamorphosis indicates that it is not a moment, but a phase. E, fer-tilized egg; H, hatching; YA, yolk absorption; and M, metamorphosis.

V. Szisch et al. / General and Comparative Endocrinology 142 (2005) 186–192 191
In conclusion, the present study presents for the Wrsttime the pattern of changes in thyroid hormone and cor-tisol concentrations during early development of the gilt-head sea bream and correlates the pattern of thesechanges with characteristic developmental phases and inparticular with metamorphosis.
Acknowledgments
Sincere thanks are due to Mr. S. Stefanakis and I.Papadakis for assistance during larval rearing and toMrs. C. Kokkari for technical help during samplings.
References
Alexander, G., Sweeting, R., Mckeown, A., 2001. The eVects of 3,4,3�-triiodo-L-thyronine on didehydroretinol synthesis by isolated Cohosalmon retinal pigment epithelial cells. General and ComparativeEndocrinology 123, 192–202.
Ayson, J.A., Lam, T.J., 1993. Thyroxine injection of female rabbitWsh(Siganus guttatus) broodstock: changes in thyroid hormone levelsin plasma, eggs, and yolk-sac larvae, and its eVect on larval growthand survival. Aquaculture 109, 83–93.
Ayson, F.G., Kaneko, T., Hasegawa, S., Hirano, T., 1995. Cortisol stim-ulates the size and number of mitochondrion rich cells in the yolk-sac membrane of embryos and larvae of tilapia (Oreochromis mos-sambicus) in vitro and in vivo. Journal of Experimental Zoology272, 419–425.
Berry, T.P., Malison, J.A., Held, J.A., Parrish, J.J., 1995. Ontogeny ofthe cortisol stress response in larval rainbow trout. General andComparative Endocrinology 97, 57–65.
Brown, C.L., Kim, B.G., 1995. Combined application of cortisol andtriiodothyronine in the culture of larval marine WnWsh. Aquaculture135, 79–86.
Deane, E.E., Woo, N.Y.S., 2003. Ontogeny of thyroid hormones, corti-sol, hsp70 and hsp90 during silver sea bream larval development.Life Sciences 72, 805–818.
De Jesus, E.G., Hirano, T., 1992. Changes in whole body concentra-tions of cortisol, thyroid hormones, and sex steroids during larvaldevelopment of the chum salmon, Oncorhynchus keta. General andComparative Endocrinology 85, 55–61.
De Jesus, E.G., Inui, Y., Hirano, T., 1990. Cortisol enhances the stimu-lating action of thyroid hormones on dorsal Wn ray resorption ofXounder larvae in vitro. General and Comparative Endocrinology79, 167–173.
De Jesus, E.G., Hirano, T., Inui, Y., 1991. Changes in cortisol and thy-roid hormone concentrations during early development and meta-morphosis in Japanese Xounder, Paralichthys olivaceus. Generaland Comparative Endocrinology 82, 369–376.
De Jesus, E.G., Toledo, J.D., Simpas, M.S., 1998. Thyroid hormonespromote early metamorphosis in grouper (Epinephelus coioides)larvae. General and Comparative Endocrinology 111, 123–140.
Frith, S.D., Eales, J.G., 1996. Thyroid hormone deiodination pathwaysin brain and liver of rainbow trout, Oncorhynchus mykiss. Generaland Comparative Endocrinology 101, 323–332.
Greenblatt, M., Brown, C.L., Lee, M., Dauder, S., Bern, H.A., 1989.Changes in thyroid hormone levels in eggs and larvae and in iodineuptake by eggs of coho and Chinook salmon Oncorhynchus kisutchand O. tschawytscha. Fish Physiology and Biochemistry 6, 261–278.
Huang, L., Schreiber, A.M., SoYentino, B., Bengston, D.A., Specker,J.L., 1998. Metamorphosis of summer Xounder (Paralichthys denta-
tus): thyroid status and the timing of gastric gland formation. Jour-nal of Experimental Zoology 280, 413–420.
Hwang, P.P., Wu, S.M., Lin, J.H., Wu, L.S., 1992. Cortisol content ofeggs and larvae of teleosts. General and Comparative Endocrinol-ogy 86, 189–196.
Inui, Y., Yamano, K., Miwa, S., 1995. The role of thyroid hormones intissue development in metamorphosing Xounder. Aquaculture 135,87–98.
Jones, I., Rogers, S.A., Kille, P., Sweeney, G.E., 2002. Molecular cloningand expression of thyroid hormone receptor alpha during salmoniddevelopment. General and Comparative Endocrinology 125, 226–235.
Koumoundouros, G., 1993. Biology of the development of the giltheadsea bream (Sparus aurata, Linnaeus, 1758, Percoidea, Sparidae) inintensive culture. M.Sc. Thesis, Department of Biology, Universityof Crete, pp. 1–84.
Lam, T.J., 1994. Hormones and egg/larval quality in Wsh. Journal of theWorld Aquaculture Society 25, 2–12.
Lam, T.J., 1995. Possible roles of hormones in the control of egg overri-pening and embryonic and larval development in Wsh. Asian Fish-eries Society Special Publication 10, 29–39.
Leatherland, J.F., Lin, L., 1989a. Thyroid hormone content of eggs andearly developmental stages of Wve Oncorhynchus species. CanadianJournal of Fish Aquaculture Science 46, 2140–2145.
Leatherland, J.F., Lin, L., 1989b. Thyroid hormone content of eggs andearly developmental stages of three stocks of Goitred coho salmon(Oncorhynchus kisutch) from the great lakes of North America, anda comparison with a stock from British Columbia. Canadian Jour-nal of Fish Aquaculture Science 46, 2146–2152.
Liu, Y.-W., Chan, W.-K., 2002. Thyroid hormones are important forembryonic to larval transitory phase in zebraWsh. DiVerentiation70, 36–45.
Liu, Y.W., Lo, L.J., Chan, W.K., 2000. Temporal expression and T3induction of thyroid hormone receptors �1 and �1 during earlyembryonic and larval development in zebraWsh. Molecular and Cel-lular Endocrinology 159, 187–195.
Miwa, S., Yamano, K., Inui, Y., 1992. Thyroid hormone stimulates gas-tric development in Xounder larvae during metamorphosis. Journalof Experimental Zoology 261, 424–430.
Mylonas, C.C., Sullivan, C.V., Hinshaw, J.M., 1994. Thyroid hormonesin brown trout (Salmo trutta) reproduction and early development.Fish Physiology and Biochemistry 13, 485–493.
Oppenheimer, J.H., Schwartz, H.L., 1997. Molecular basis of thyroidhormone-dependent brain development. Endocrine Reviews 18,462–475.
Papandroulakis, N., Divanach, P., Anastasiadis, P., Kentouri, M., 2001.The pseudo-green water technique for intensive rearing of seabream (Sparus aurata) larvae. Aquaculture International 9, 205–216.
Pittman, K., Jelmert, A., Næss, T., Harboe, T., Watanabe, K., 1998.Plasticity of viable postmetamorphic forms of farmed Atlantic hal-ibut, Hippoglossus hippoglossus, L. Aquaculture Research 29, 949–954.
Power, D.M., Llewellyn, L., Faustino, M., Nowell, M.A., Bjornsson,B.Th., Einarsdottir, I.E., Canario, A.V.M., Sweeney, G.E., 2001.Thyroid hormones in growth and development of Wsh. Compara-tive Biochemistry and Physiology (C) 130, 447–459.
Reddy, P.K., Lam, T.J., 1992. EVect of thyroid hormones on morpho-genesis and growth of larvae and fry of telescopic-eye black gold-Wsh, Carassius auratus. Aquaculture 107, 383–394.
Reddy, P.K., Brown, C.L., Leatherland, J.F., Lam, T.J., 1992. Role ofthyroid hormones in tilapia larvae (Oreochromis mossambicus):II. Changes in the hormones and 5’-monodeiodinase activityduring development. Fish Physiology and Biochemistry 9, 487–496.
Sachs, L.M., Damjanovski, S., Jones, P.L., Li, Q., Amano, T., Ueda, S.,Shi, Y.B., Ishizuya-Oka, A., 2000. Metamorphosis—dual functions

192 V. Szisch et al. / General and Comparative Endocrinology 142 (2005) 186–192
of thyroid hormone receptors during Xenopus development. Com-parative Biochemistry and Physiology (C) 126, 199–211.
Sampath-Kumar, R., Lee, S.T.L., Tan, C.H., Munro, A.D., Lam, T.J.,1997. Biosynthesis in vivo and excretion of cortisol by Wsh larvae.Journal of Experimental Zoology 277, 337–344.
SuW, S.B., Donaldson, A., JeVcoate, S.L., 1994. World Health Organiza-tion (WHO), Matched Reagent Programme Method Manual, 18thed.
Tagawa, M., Hirano, T., 1987. Presence of thyroxine in eggs andchanges in its content during early development of chum salmon,Oncorhynchus keta. General and Comparative Endocrinology 68,129–135.
Tagawa, M., Tanaka, M., Matsumoto, S., Hirano, T., 1990. Thyroidhormones in eggs of various freshwater, marine and diadromousteleosts and their changes during egg development. Fish Physiologyand Biochemistry 8, 515–520.
Tanaka, M., Tanangonan, J.B., Tagawa, M., de Jesus, E.G., Nishida, H.,Isaka, M., Kimura, R., Hirano, T., 1995. Development of the pitui-tary, thyroid and interrenal glands and application of endocrinol-
ogy to the improved rearing of marine Wsh larvae. Aquaculture 135,111–126.
Tata, J.R., 1999. Amphibian metamorphosis as a model for studyingthe developmental actions of thyroid hormone. Biochimie 81, 359–366.
Wright, M.L., 2002. Melatonin, diel rhythms, and metamorphosis inanuran amphibians. General and Comparative Endocrinology 126,251–254.
Wright, M.L., Alves, C.D., 2001. The decrease in plasma melatonin atmetamorphic climax in Rana catesbeiana bullfrog tadpoles isinduced by thyroxine. Comparative Biochemistry and PhysiologyPart A 129, 653–663.
Yamano, K., Tagawa, M., de Jesus, E.G., Hirano, T., Miwa, S., Inui, Y.,1991. Changes in whole body concentrations of thyroid hormonesand cortisol in metamorphosing conger eel. Journal of Compara-tive Physiology (B) 161, 371–375.
Yamano, K., Araki, K., Sekiwawa, K., Inui, Y., 1994. Cloning of thy-roid hormone receptor genes expressed in metamorphosing Xoun-der. Developmental Genetics 15, 378–382.
Related Documents











