Zhou et al, H-00557-2007.R1 0 Metabolic Response to an Acute Jump in Cardiac Workload: Effects on Malonyl-CoA, Mechanical Efficiency, and Fatty Acid Oxidation An Acute Jump in Cardiac Workload Lufang Zhou 1 , Hazel Huang 2 , Celvie L. Yuan 3 , Wendy Keung 4 , Gary D. Lopaschuk 4 and William C. Stanley 2,3,5 1 Departments of Biomedical Engineering, 2 Physiology & Biophysics, and 3 Nutrition, Case Western Reserve University, Cleveland, OH 44106; 4 Department of Pediatrics, University of Alberta, Edmonton, Canada, T6G 2S2; and 5 Division of Cardiology, Department of Medicine, University of Maryland, Baltimore, MD 21201 Short title: Cardiac Metabolism at High Workload Address for correspondence: William C. Stanley, Ph.D. Division of Cardiology Department of Medicine University of Maryland-Baltimore 20 Penn St., HSF2, Room S022 Baltimore, MD 21201, USA 410-706-3585 (Phone) 410-706-3583 (Fax) e-mail: [email protected] Page 1 of 27 Copyright Information Articles in PresS. Am J Physiol Heart Circ Physiol (December 14, 2007). doi:10.1152/ajpheart.00557.2007 Copyright © 2007 by the American Physiological Society.

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Zhou et al, H-00557-2007.R1
0
Metabolic Response to an Acute Jump in Cardiac Workload:
Effects on Malonyl-CoA, Mechanical Efficiency, and Fatty
Acid Oxidation An Acute Jump in Cardiac Workload
Lufang Zhou1, Hazel Huang2, Celvie L. Yuan3, Wendy Keung4, Gary D. Lopaschuk4 and
William C. Stanley2,3,5
1Departments of Biomedical Engineering, 2Physiology & Biophysics, and 3Nutrition, Case
Western Reserve University, Cleveland, OH 44106; 4Department of Pediatrics, University of
Alberta, Edmonton, Canada, T6G 2S2; and 5Division of Cardiology, Department of Medicine,
University of Maryland, Baltimore, MD 21201
Short title: Cardiac Metabolism at High Workload
Address for correspondence: William C. Stanley, Ph.D.
Division of Cardiology Department of MedicineUniversity of Maryland-Baltimore20 Penn St., HSF2, Room S022Baltimore, MD 21201, USA410-706-3585 (Phone)410-706-3583 (Fax)e-mail: [email protected]
Page 1 of 27
Copyright Information
Articles in PresS. Am J Physiol Heart Circ Physiol (December 14, 2007). doi:10.1152/ajpheart.00557.2007
Copyright © 2007 by the American Physiological Society.

Zhou et al, H-00557-2007.R1
1
ABSTRACT
Inhibition of myocardial fatty acid oxidation can improve left ventricular (LV) mechanical
efficiency by increasing LV power for a given rate of myocardial energy expenditure. This
phenomenon has not been assessed at high workloads in non-ischemic myocardium, so we
therefore subjected in vivo pig hearts to a high workload for 5 min, and assessed whether
blocking mitochondrial fatty acid oxidation with the carnitine palmitoyltransferase-I (CPT-I)
inhibitor oxfenicine would improve LV mechanical efficiency. In addition, the cardiac content
of malonyl-CoA, (an endogenous inhibitor of CPT-I) and acetyl-CoA carboxylase activity
(which synthesizes malonyl-CoA) were assessed. Increased workload was induced by aorta
constriction and dobutamine infusion, and LV efficiency was calculated from the LV pressure-
volume loop and LV energy expenditure. In untreated pigs the increase in LV power resulted in
a 2.5-fold increase in both fatty acid oxidation and cardiac malonyl-CoA content, but did not
affect the activation state of acetyl-CoA carboxylase (ACC). The activation state of the ACC
inhibitory kinase, AMP activated protein kinase, decreased by 40% with increased cardiac
workload. Pretreatment with oxfenicine inhibited fatty acid oxidation by 75% and had no effect
on cardiac energy expenditure, but significantly increased LV power and LV efficiency (37+5%
vs. 26+5%, respectively, p<0.05) at high workload. In conclusion 1) myocardial fatty acid
oxidation increases with a short-term increase in cardiac workload despite an increase in
malonyl-CoA concentration, and 2) inhibition of fatty acid oxidation improves LV mechanical
efficiency by increasing LV power without effecting cardiac energy expenditure.
Key words: acetyl-CoA carboxylase, AMP activated protein kinase, exercise, fatty acids, heart,
mitochondria
Page 2 of 27
Copyright Information

Zhou et al, H-00557-2007.R1
2
INTRODUCTION
One of the major determinants of myocardial oxygen consumption (MVO2) at a given
rate of left ventricular (LV) power generation is mitochondrial substrate selection (23; 44).
Under normal resting conditions, fatty acid oxidation is the predominant source of energy for
cardiac power generation (60~80%), however studies in humans(39), dogs(30; 31) and pigs(25)
in vivo, and in isolated perfused rat (3; 21) and mouse(20) hearts show that with high rates fatty
acid oxidation, the external power is reduced for a given MVO2 (3; 30; 39). In the failing heart,
or during acute ischemia and/or reperfusion, pharmacological treatment with agents that either
inhibit myocardial fatty acid oxidation (5; 6) or directly activate carbohydrate oxidation (2; 28;
41) increase LV function without affecting MVO2, and therefore improve the left ventricular
mechanical efficiency (defined as the ratio of external LV power/LV energy expenditure).
However, the effect of inhibiting fatty acid oxidation on cardiac function and MVO2 during high
cardiac workloads in the healthy heart is not known.
Under normal or ischemic conditions, fatty acid oxidation strongly inhibits the
mitochondrial enzyme pyruvate dehydrogenase, which inhibits oxidation of pyruvate, and thus
glucose and lactate uptake and oxidation(35). On the other hand, it has been shown that
inhibition of fatty acid oxidation increases pyruvate oxidation and glucose uptake and oxidation
at rest and during exercise(26; 41; 45). Fatty acid oxidation in the heart is regulated at the level
of the mitochondrial outer membrane by the activity of carnitine palmitoyltransferase I (CPTI),
which is inhibited by malonyl-CoA on the cytosolic side of the enzyme(22; 44). Several studies
have shown an inverse relationship between myocardial malonyl-CoA content and fatty acid
oxidation(9; 24; 37; 40; 43), and specifically that adrenergic stimulation corresponds with a
reciprocal increase in fatty acid oxidation and decrease in malonyl-CoA content(13-16; 24; 36).
Page 3 of 27
Copyright Information

Zhou et al, H-00557-2007.R1
3
Malonyl-CoA is produced by acetyl-CoA carboxylase (ACC), which is inhibited when
phosphorylated at serine-79 by AMP activated protein kinase (AMPK), while AMPK is activated
by phosphorylation at threonine-172 (10). We previously found that the reduction in malonyl-
CoA content that occurred when MVO2 was increased by adrenergic stimulation was not
associated with reduced ACC activity or increased AMPK activity in pigs (15), however tissue
was sampled between 15 and 30 minutes after the initiation of stimulation. It is not known if
there are changes in malonyl-CoA content and activation of ACC and AMPK during the initial
minutes of the transition from a low to a high cardiac workload.
The present study evaluated what effect inhibition of fatty acid oxidation on myocardial
LV function and mechanical efficiency. We hypothesized that a switch in myocardial energy
substrate use from fatty acid to carbohydrates would increase LV power without affecting
oxygen consumption, and therefore improve myocardial energy efficiency. The second aim of
this study was to determine if there is a decrease in malonyl-CoA content with an abrupt short-
term increase in cardiac workload. We hypothesized that with five minutes of increased cardiac
workload the myocardial content of malonyl-CoA would decrease due to activation of AMPK
and inhibition of ACC, resulting in greater fatty acid oxidation, as previously observed with a
longer duration of adrenergic stimulation (13-16; 24; 36). Studies were performed in an
established open-chest pig model, with animals subjected to an abrupt five minute increase in
cardiac workload induced by simultaneous adrenergic stimulation, parasympathetic blockade and
aortic constriction treatment (24; 38; 48). Fatty acid oxidation was inhibited at the level of
transport into the mitochondria using oxfenicine, a CPT-I inhibitor. There are numerous
approaches to inhibiting fatty acid oxidation(29; 42). In this study oxfenicine was selected
because it is devoid of cardiovascular effects under normal conditions(5; 7; 38; 48), has a rapid
Page 4 of 27
Copyright Information

Zhou et al, H-00557-2007.R1
4
onset of action(19), and consistently reduces fatty acid oxidation in the pig heart under
conditions of increased cardiac workload(24; 38; 48).
METHODS
Studies were performed in accordance with the Guide for the Care and Use of Laboratory
Animals (NIH Publication Number 85-23, revised 1996) and with prior approval of the
Institutional Animal Care and Use Committee at Case Western Reserve University. Twenty four
domestic pigs of either sex (mean weight, 35.1 ± 1.1 kg) were entered into the study. Data from
these studies have been reported separately on the regulation of pyruvate dehydrogenase activity,
NADH, glycogen concentration, and basic hemodynamics (48).
Surgical preparation
The surgical preparation has been previously described in detail (24; 38; 48). Briefly,
overnight fasted pigs were sedated with Telazol (6 mg/kg im), anesthetized with isoflurane by
mask (5%), ventilated with 100% O2,and maintained on isoflurane (0.75-1.5%) and ketamine (4
mg kg-1 min-1 iv) to keep PCO2 and pH in the normal range (PO2 >100 mmHg, PCO2 35-45
mmHg, and pH 7.35-7.45). A femoral artery and vein were catheterized for blood sampling and
infusion, respectively, and the animals were heparinized (200 U/kg bolus followed by 100 U kg-1
min-1i.v) to prevent clotting and thrombus formation. The heart was exposed via a midline
sternotomy, and the left atrium was catheterized for infusion of dobutamine and atropine. A
vascular occluder was placed around the ascending aorta and constricted during the dobutamine
treatment period. The cardiac anterior interventricular vein was catheterized for coronary venous
blood sampling, and a Doppler ultrasonic flow meter was placed around the proximal left
Page 5 of 27
Copyright Information

Zhou et al, H-00557-2007.R1
5
anterior descending coronary artery to record blood flow continuously (Transonics Inc). Four
sonimicrometry crystals were placed at midmyocardial depth in the base, apex, and septum of the
lateral wall of left ventricle to continuously measure LV volume using an online commercial
system (Sonometrics). A high fidelity pressure transducer (Millar Instruments) was positioned in
the LV, and the signal was integrated with LV volume, and the LV pressure – LV volume loop
area was calculated for each beat.
Experimental protocol
Three groups of pigs were studied: 1) an untreated group subjected to increased cardiac
workload with dobutamine infusion (DOB, n=8), 2) an oxfenicine treated group that was
subjected to increased cardiac workload (DOB + OXF, n=8), and 3) a control (CON, n=8) group
with sham instrumentation and normal cardiac workload. At the beginning of the protocol,
[9,10-3H]oleate tracer was infused (40 µCi/hr i.v.) for the measurement of fatty acid oxidation,
and the oxfenicine treatment was initiated in the DOB + OXF group (30 mg/kg iv bolus
oxfenicine followed by an infusion at 30 mg kg-1 min-1). After a 50 minute equilibrium period,
animals in the DOB and DOB + OXF were subjected to a 5 minute period of increased cardiac
work induced by constricting the aortic cuff sufficient to maintain the peak left ventricular
systolic pressure at ~190 mmHg, while simultaneously infusing dobutamine (100 µg/kg as a
bolus followed by 40 µmol kg-1 min-1) and atropine (2 mg i.v. bolus) into the left atrium to
increase heart rate and contractility. Arterial and venous blood samples for measurement of
blood glucose and lactate concentrations were taken before (-5 and -1 minutes) and during
dobutamine treatment (at 20, 45, 75 seconds, 2, 3, 4, and 5 minutes), and samples were drawn for
plasma free fatty acid and 3H2O concentrations at -5 and -1 minutes, and at 3, 4, and 5 minutes of
Page 6 of 27
Copyright Information

Zhou et al, H-00557-2007.R1
6
dobutamine infusion. After 5 minutes of increased workload a large punch biopsy (~3 g) was
rapidly excised from the LAD bed and immediately freeze-clamped on aluminum blocks pre-
cooled in liquid nitrogen and stored at –80°C for later analysis(40). The CON group received the
same infusion of [9,10-3H]oleate, and arterial and coronary venous blood samples were drawn at
50-60 minutes, followed immediately by a myocardial biopsy, as in the other two groups.
Analytic methods
Arterial and venous O2 saturation and hemoglobin were measured spectrophotometrically
with a hemoximeter (A-VOX System, San Antonio, TX), and pH, PCO2, and PO2 were measured
in a blood gas analyzer (Nova Biomedical, Waltham, MA). Subsequent biochemical analysis
was performed with the investigator blinded to treatment. Blood samples were analyzed for
concentrations of glucose and lactate, and plasma was assayed for free fatty acids, [3H] oleate
and 3H2O as previously described(24; 38). Malonyl-CoA and adenine nucleotide contents were
assayed by high-pressure liquid chromatography with UV detection as previously described (40).
All tissue concentrations were expressed per gram wet weight of tissue. The amount of total and
phosphorylated ACC and AMPK were assessed by western blot using specific antibodies for
phosphor-ACC at serine-79 and phospho-AMPK at threonine-172, as previously described (24;
27). AMPK activity was measured on myocardial homogenates as previously described (11).
Calculations
Myocardial blood flow was measured from the ultrasonic flow meter and normalized by
dividing by the weight of the heart being perfused by the LAD (34; 40). The net uptakes (µmol
kg-1 min-1) of glucose, lactate, free fatty acids and oxygen were calculated as the product of
Page 7 of 27
Copyright Information

Zhou et al, H-00557-2007.R1
7
arterial - venous difference and blood flow. The rates of exogenous fatty acid oxidation (µmol g-
1 min-1) was calculated as the product of the release of 3H2O (dpm/ml) and myocardial blood
flow, divided by the arterial specific radioactivity of free fatty acids (dpm/µmol)(38).
Stroke volume was calculated as LV end diastolic volume – LV end systolic volume, and
cardiac output as the product of stroke volume and heart rate. LV stroke work (J) was calculated
as the LV pressure (Pascals) times volume, and LV power (watts) was calculated as the product
of LV stroke work and heart rate(4). LV energy expenditure was calculated from MVO2
assuming 20.2J/ml of O2 (46), and LV mechanical efficiency as LV power/LV energy
expenditure.
Statistical analysis
All hemodynamic variables; rates of free fatty acid, glucose, and lactate uptakes; rate of
fatty acid oxidation; and tissue metabolite concentrations were compared between resting
conditions and increased cardiac work and between DOB and DOB + OXF groups using a one-
or two-way ANOVA with the Bonferroni post-hoc test for multiple comparisons, as appropriate.
Significance was set at P<0.05, and values are reported as means ± SE.
Page 8 of 27
Copyright Information

Zhou et al, H-00557-2007.R1
8
RESULTS
As recently published separately from these experiments, there was a significant increase
in heart rate, peak LV pressure, myocardial blood flow and MVO2 in the DOB and DOB + OXF
groups compared to CON, while there were no differences between the DOB and DOB +OXF
groups (see reference (48)). Cardiac output, stroke work and LV energy expenditure increased to
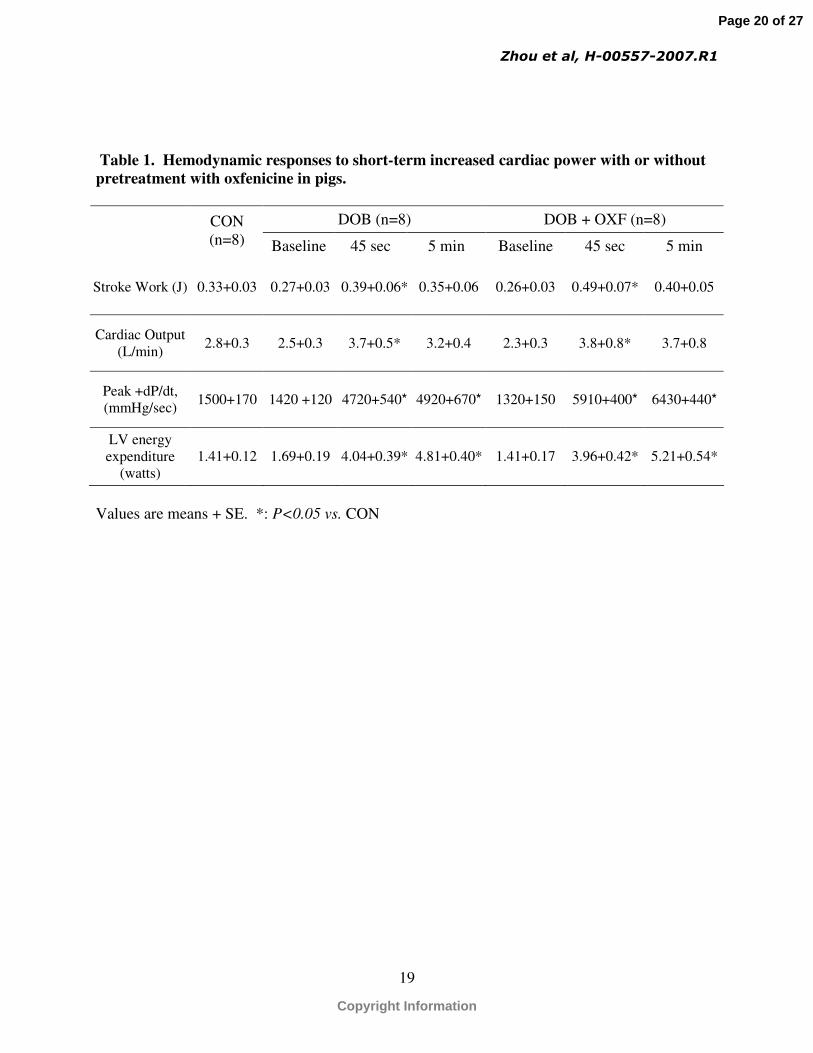
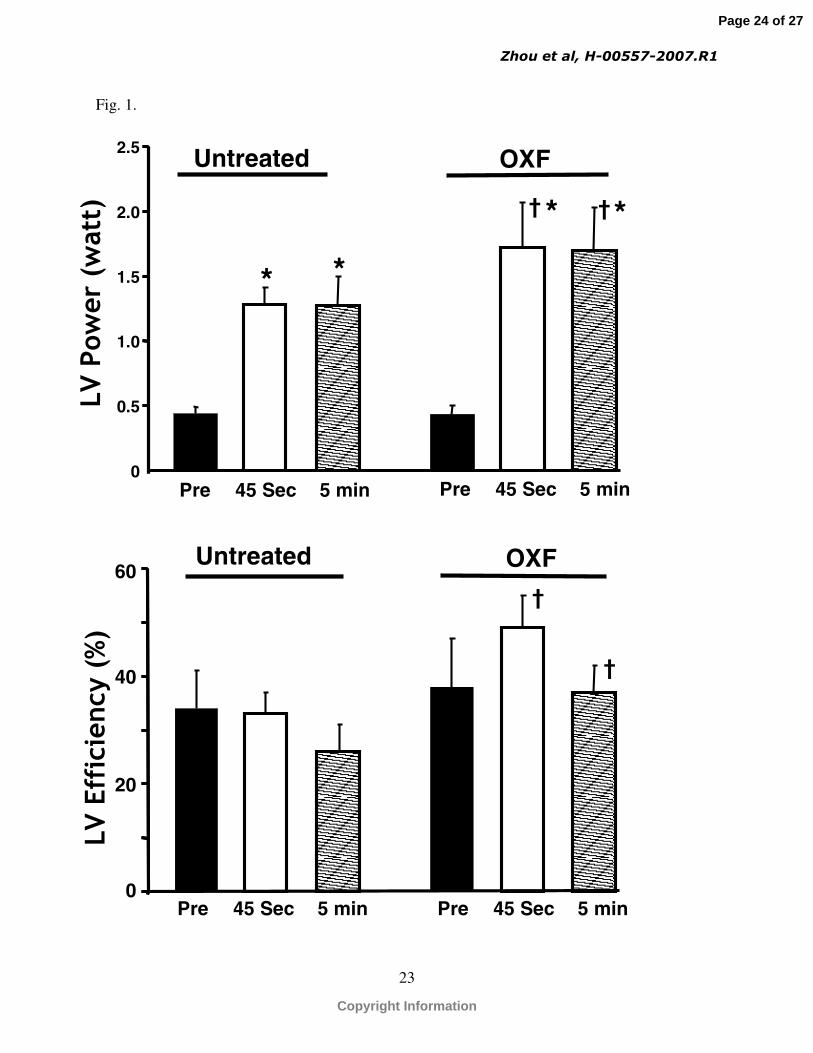
a similar extent in the DOB and DOB + OXF groups (Table 1). Treatment with oxfenicine
significantly increased LV power compared with the DOB group (1.72±0.35 vs. 1.28±0.13 at 45
seconds and 1.70±0.33 vs. 1.27±0.23 at 5 minutes, respectively) (Fig. 1) despite no effect on LV
energy expenditure (Table 1). As a result, a significant improvement in mechanical efficiency
was seen in the DOB + OXF group compared to the DOB group (Fig. 1).
The arterial concentrations of lactate, glucose, and fatty acids were unchanged over the
course of study and values were similar among the three experimental groups (data not shown).
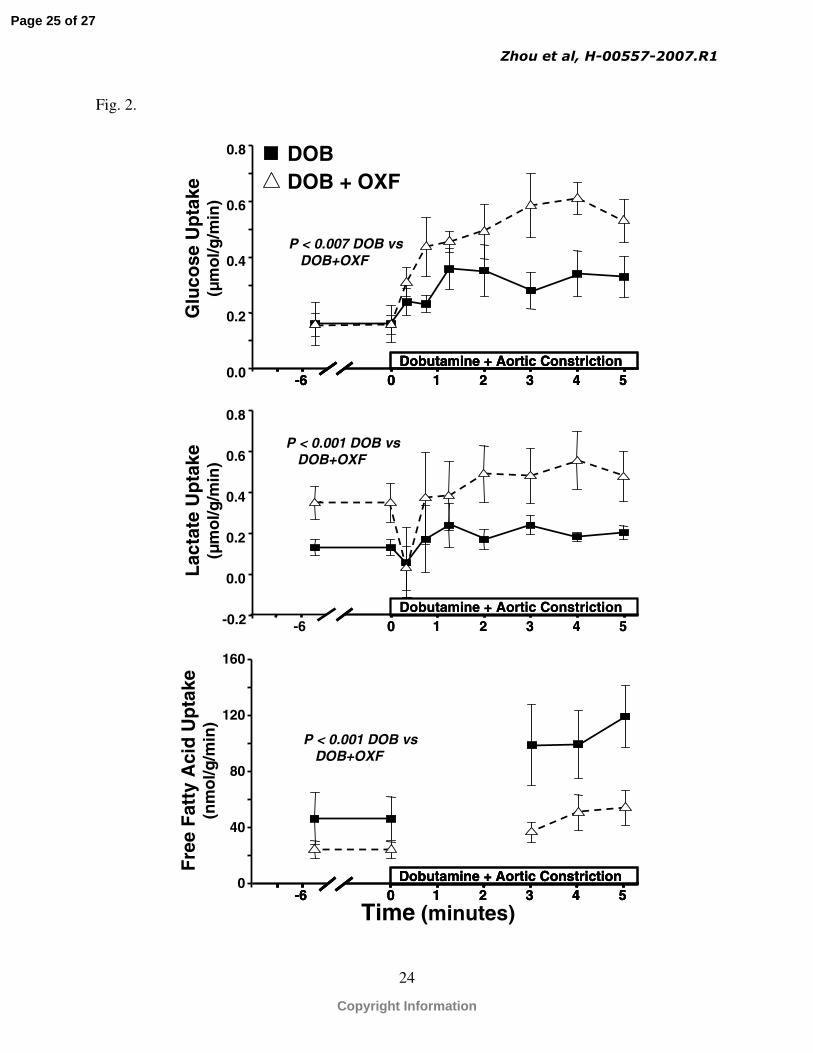
Increased cardiac work significantly increased glucose and fatty acid uptake in the DOB group
(Fig. 2). Pharmacological inhibition of CPT-1 suppressed fatty acid uptake in the DOB + OXF
group under resting and high cardiac work conditions, and further enhanced glucose and lactate
uptakes compared to the DOB group (Fig. 2). Myocardial fatty acid oxidation was also
suppressed by oxfenicine treatment under resting conditions (14±9 compared to 49±14nmol g-1
min-1; p<0.05) and at a high cardiac workload (Fig. 3). There was a decreased lactate uptake in
the DOB and DOB + OXF groups in the first minute of increased cardiac energy expenditure
(Fig. 2B), with 4 out of 8 pigs in the DOB group and 2 out of 8 pigs in the DOB + OXF group
showing a net release of lactate at 20 and/or 45 seconds.
The rate of free fatty acid oxidation was greater in the DOB group compared with the
DOB + OXF group before (data not shown) and during the dobutamine treatment period (Fig. 3).
Page 9 of 27
Copyright Information

Zhou et al, H-00557-2007.R1
9
Malonyl-CoA concentration was greater in the DOB and DOB+OXF groups compared with the
CON group (P<0.05), with no significant difference between these two groups (Fig. 3). The
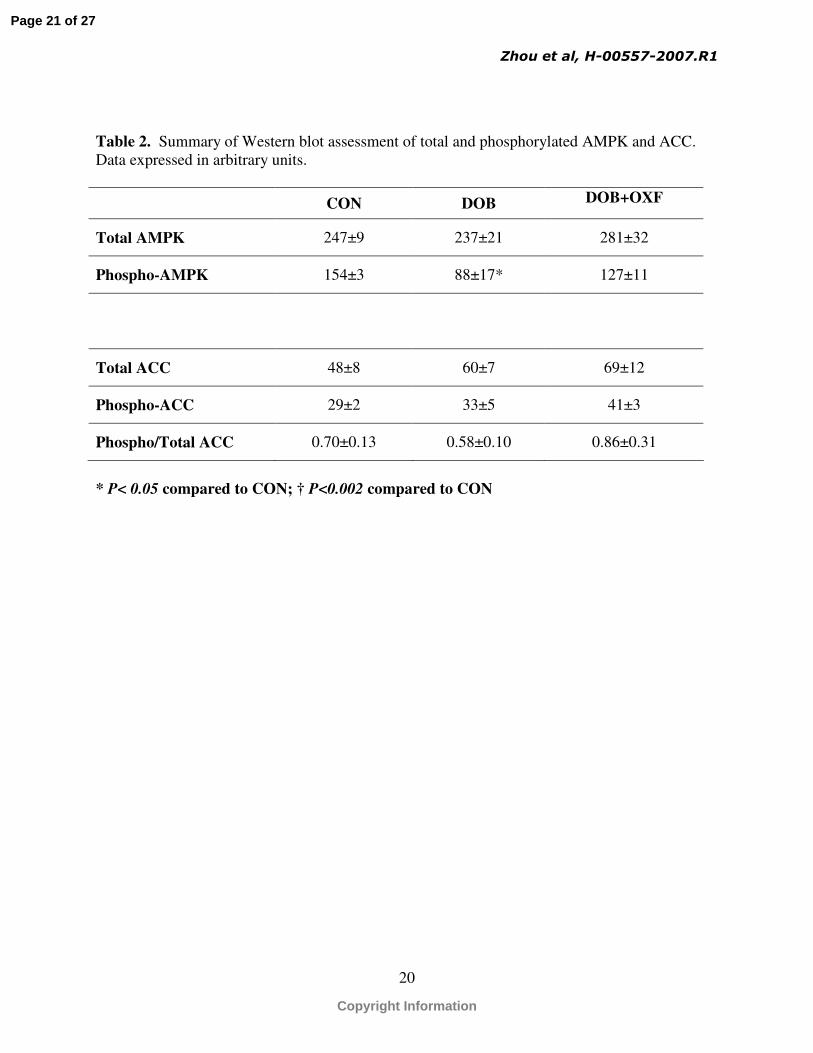
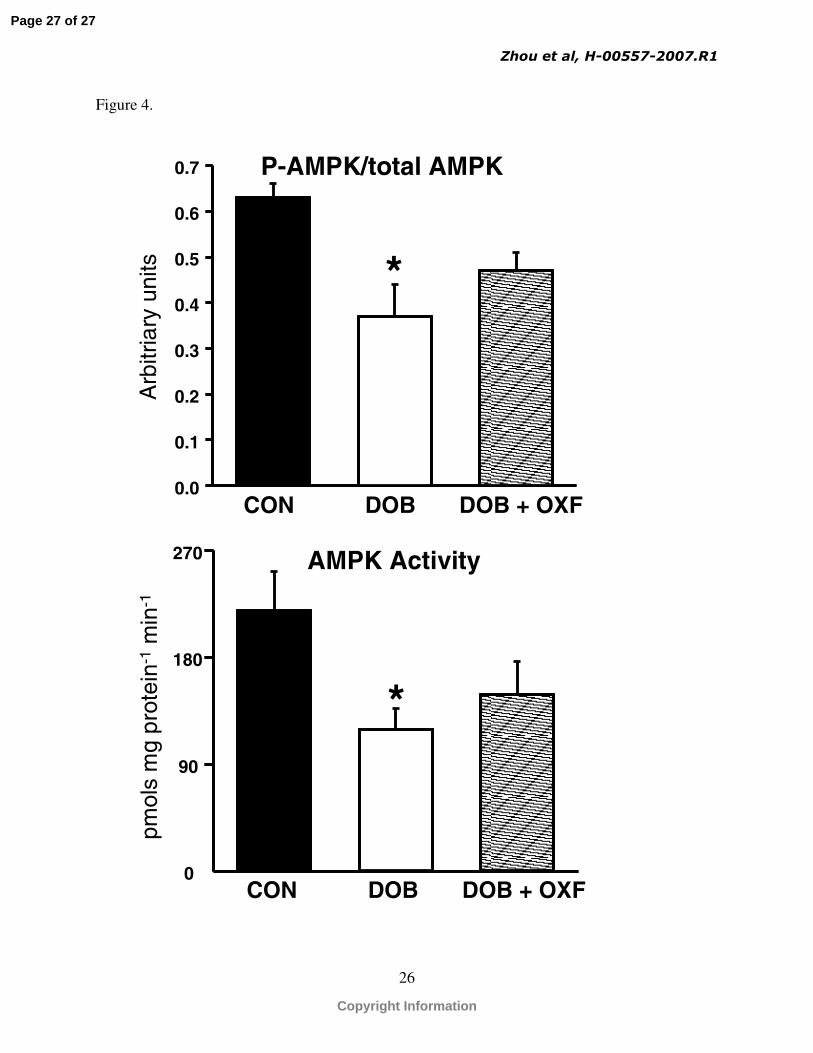
amount of total ACC and phospho-ACC were not different among groups, nor was the phospho-
ACC/total ACC ratio (Table 2). While total AMPK was not different among groups, there was a
significant reduction in phospho-AMPK and the ratio of phospho-AMPK/total AMPK in the
DOB group compared to the CON group, while the DOB + OXF group was not different from
the other two groups.
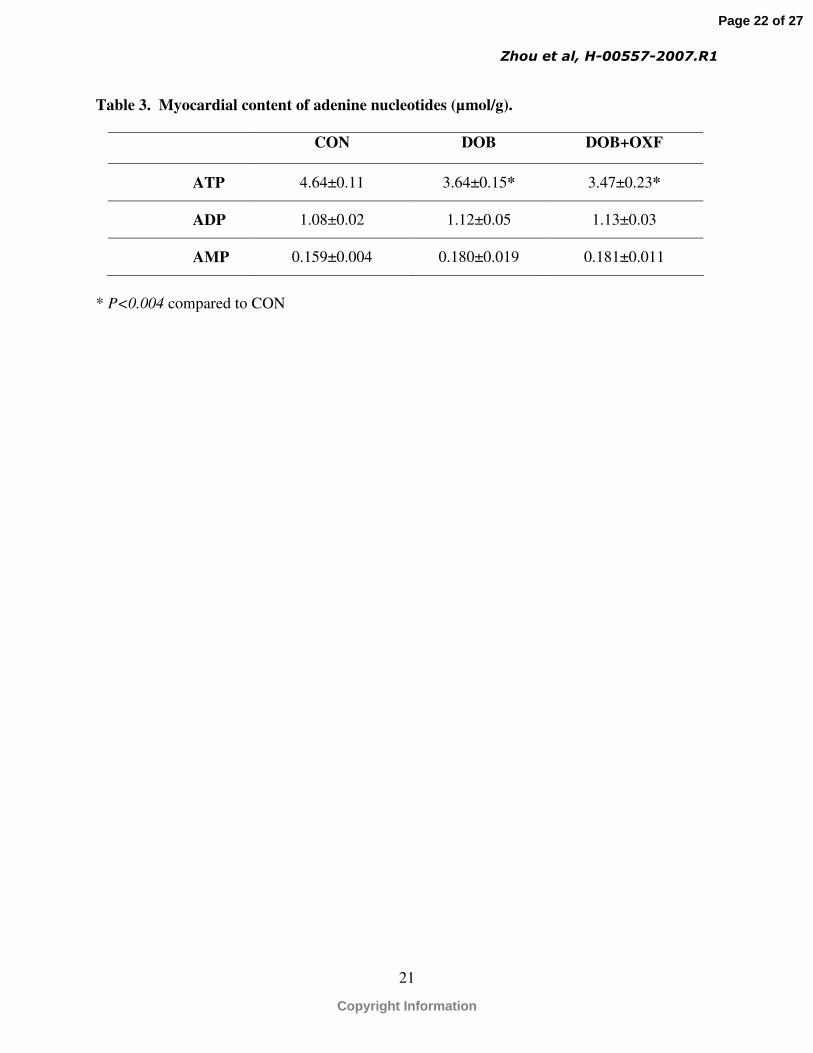
The cardiac content of AMP and ADP were not different among the groups (Table 3),
although the ATP content was 20% lower in the DOB and DOB + OXF groups compared to the
CON group. Interestingly, we previous measured ATP and ADP content on these same samples
using a less precise luciferase assay, which showed approximately twice the variability, but again
no differences among the groups(48).
Page 10 of 27
Copyright Information

Zhou et al, H-00557-2007.R1
10
DISCUSSION
The results of this study show that despite the expected increase in myocardial fatty acid
oxidation during an acute work jump there was a paradoxical increase in the tissue content of
malonyl-CoA, an established inhibitor of CPT-I and cardiac fatty acid oxidation. In addition, the
increase in malonyl-CoA was not due to activation of ACC, but did correspond to a decrease in
AMPK activation. Treatment with oxfenicine had little effect in malonyl-CoA content despite a
75% decrease in fatty acid oxidation, suggesting that the observed increase in malonyl-CoA was
largely independent from changes in fatty acid oxidation.
The second main finding of this study is that inhibition of fatty acid oxidation with
oxfenicine increases LV power without increasing cardiac energy expenditure. This
phenomenon has been observed under conditions of normal workload, during demand-induced
ischemia, and with post-ischemic reperfusion(41; 44). The present study extends this concept to
conditions of high workload similar to intense exercise in healthy people, and suggests the
provocative idea that CPT-I inhibition might improve exercise performance in short-term intense
aerobic athletic events. These observations are consistent with previous studies showing that
switching from fatty acid utilization to carbohydrate increases LV energy efficiency (2; 3; 20;
21; 25; 30; 32; 39). Compared with fatty acids, carbohydrates are more oxygen efficient (i.e., for
a given amount of ATP synthesis fatty acid oxidation requires 11% more oxygen consumption
than pyruvate), and high concentrations of fatty acids have also been shown to uncouple
oxidative phosphorylation and increase oxygen utilization in isolated mitochondria and cells(44).
Thus, the improved efficiency with oxfenicine is likely attributed to greater ATP synthesis per
oxygen consumption and/or more effective ATP use by the heart.
Page 11 of 27
Copyright Information

Zhou et al, H-00557-2007.R1
11
We have previously observed that 15 to 30 minutes of intense adrenergic stimulation in
pigs results in a reciprocal decrease in malonyl-CoA content and an increase in fatty acid uptake
and oxidation (12; 26; 38), suggesting that less malonyl-CoA inhibition of CPT-I is a primary
mechanism for the increase in cardiac fatty acid oxidation observed with physiological stresses
such as an acute bout of exercise(15; 16; 24). In contrast, the present investigation found the rate
of fatty acid oxidation increased with dobutamine treatment despite a 2.5-fold increase in tissue
malonyl-CoA concentration (Fig. 3). Since malonyl-CoA exist in both cytosol and mitochondria
(18; 22; 47), it is possible that the cytosolic malonyl-CoA decreased while mitochondrial
malonyl-CoA increased. Malonyl-CoA inhibits CPT-I on the cytosolic side of the enzyme(22;
44), and is produced in both the cytosol and mitochondrial matrix from acetyl-CoA (44). The
supply of acetyl-CoA is a major regulator of malonyl-CoA formation (36; 37). The increase in
workload in the present experiment caused a 50% increase in the acetyl-CoA concentration (48),
presumably due to the rapid stimulation of acetyl-CoA formation by pyruvate dehydrogenase,
which may have triggered a selective increase of malonyl-CoA in the mitochondrial matrix.
Thus the increased tissue malonyl-CoA content may be due to a specific increase in
mitochondrial malonyl-CoA, as previously suggested (24). It is impossible to prove this based
on current experimental results, since malonyl-CoA was measured in whole tissue without
distinguishing cytosolic and mitochondrial compartments. Future studies should rapidly separate
the cytosol and mitochondria and measure these CoAs in these two compartments, although
accurate measurements of cytosolic and mitochondrial malonyl-CoA has yet to be made due to
technical difficulties with this approach. In any case, the results of the present study clearly
indicate that a fall in total tissue malonyl-CoA content is not essential for the increase in
Page 12 of 27
Copyright Information

Zhou et al, H-00557-2007.R1
12
cardiac fatty acid oxidation that occurs in response to an acute increase in cardiac
workload.
Consistent with our previous studies in pigs (15; 24) there was no increase in the
activation of AMPK or ACC phosphorylation with increased cardiac workload. Studies in
working perfused rat hearts also found no increase AMPK activity at either 1 or 15 minutes
following a 2 to 3 fold increase in cardiac power(1). In addition, mice expressing a cardiac-
specific dominant-negative AMPKalpha2 subunit have normal ATP content and glycogen
depletion in response to acute exercise stress, stress echocardiography, and maximal exercise
capacity(33). On the other hand, 10 minutes of treadmill running in rats approximately doubled
AMPK activity and phospho-AMPK at threonine-172, and also doubled the amount of phospho-
ACC at serine-79(8). The results of the present study, and our previous work in pigs(15; 24),
consistently suggest that there is not activation of AMPK or inhibition of ACC in response to
high dose dobutanime and increased aortic pressure in pigs. While AMPK appears to play a
central role in the regulation of cardiac energy metabolism under many conditions(10), the
results of the present in vivo study shows that despite a significant decrease in phospho-AMPK
and AMPK activity (Fig 4) there is a stimulation of glucose uptake, glycogenolysis, and fatty
acid oxidation, (Table 1, Fig. 1-3)(48). Taken together, activation of AMPK is not an essential
regulatory component of metabolic response to a step increase in cardiac workload.
Cardiac ATP content, as measured by high pressure liquid chromatography, was
decreased by 20% under conditions of high workload, however AMP and ADP were not
increased, suggesting that there was a net loss of adenine nucleotides during the 5 minutes of
increased cardiac workload. This is consistent with our previous observation of an increased
adenosine production and efflux with dobutamine-induced work in pigs (17). In terms of
Page 13 of 27
Copyright Information

Zhou et al, H-00557-2007.R1
13
metabolic regulation, since there was also no increase in AMP or phospho-AMPK, it appears that
the modest fall in ATP is not a major regulator of the energetic response to an acute jump in
workload.
In conclusion, the results of the present study show that inhibition of fatty acid oxidation
improves LV mechanical efficiency by increasing LV contractile power without affecting MVO2
during an acute bout of high workload. This suggests a novel approach to improving LV
mechanical efficiency at high cardiac workloads with drugs that optimize myocardial energy
metabolism, and presents the possibility that inhibition of CPT-I could potentially enhance
performance in athletic events that are limited by cardiac pump function. In addition, we
observed a paradoxical elevation in malonyl-CoA concentration and fatty acid oxidation at high
workloads. Lastly, a significant fall in AMPK activation was observed under conditions of high
energy demand, which further illustrates that activation of AMPK is not an essential component
of the metabolic response to the increase in cardiac workload.
Page 14 of 27
Copyright Information

Zhou et al, H-00557-2007.R1
14
ACKNOWLEDGEMENTS
This work was support by NIH grants HL074237 and GM-66309, and by a grant from the
Canadian Institutes of Health Research. Gary Lopaschuk is a Medical Scientist of the Alberta
Heritage Foundation for Medical Research The authors thank Drs. Monika Duda, Isidore Okere,
and Naveen Sharma for assistance with the animal experiments.
Page 15 of 27
Copyright Information

Zhou et al, H-00557-2007.R1
15
REFERENCES
1. Beauloye C, Marsin AS, Bertrand L, Vanoverschelde JL, Rider MH and Hue L. The stimulation of heart glycolysis by increased workload does not require AMP-activated protein kinase but a wortmannin-sensitive mechanism. FEBS Lett 531: 324-328, 2002.
2. Bersin RM, Wolfe C, Kwasman M, Lau D, Klinski C, Tanaka K, Khorrami P, Henderson GN, de Marco T and Chatterjee K. Improved hemodynamic function and mechanical efficiency in congestive heart failure with sodium dichloroacetate. J Am Coll Cardiol 23: 1617-1624, 1994.
3. Burkhoff D, Weiss RG, Schulman SP, Kalil-Filho R, Wannenburg T and Gerstenblith G. Influence of metabolic substrate on rat heart function and metabolism at different coronary flows. Am J Physiol 261: H741-H750, 1991.
4. Cappola TP, Kass DA, Nelson GS, Berger RD, Rosas GO, Kobeissi ZA, Marban E and Hare JM. Allopurinol improves myocardial efficiency in patients with idiopathic dilated cardiomyopathy. Circulation 104: 2407-2411, 2001.
5. Chandler MP, Chavez PN, McElfresh TA, Huang H, Harmon CS and Stanley WC. Partial inhibition of fatty acid oxidation increases regional contractile power and efficiency during demand-induced ischemia. Cardiovasc Res 59: 143-151, 2003.
6. Chandler MP, Stanley WC, Morita H, Suzuki G, Roth BA, Blackburn B, Wolff A and Sabbah HN. Short-term treatment with ranolazine improves mechanical efficiency in dogs with chronic heart failure. Circ Res 91: 278-280, 2002.
7. Chavez PN, Stanley WC, McElfresh TA, Huang H, Sterk JP and Chandler MP. Effects of Hyperglycemia and Fatty Acid Oxidation Inhibition during Aerobic Conditions and Demand-induced Ischemia. Am J Physiol Heart Circ Physiol H1521-H1527, 2003.
8. Coven DL, Hu X, Cong L, Bergeron R, Shulman GI, Hardie DG and Young LH. Physiological role of AMP-activated protein kinase in the heart: graded activation during exercise. Am J Physiol Endocrinol Metab 285: E629-E636, 2003.
9. Dyck JR, Cheng JF, Stanley WC, Barr R, Chandler MP, Brown S, Wallace D, Arrhenius T, Harmon C, Yang G, Nadzan AM and Lopaschuk GD. Malonyl coenzyme a decarboxylase inhibition protects the ischemic heart by inhibiting fatty acid oxidation and stimulating glucose oxidation. Circ Res 94: e78-e84, 2004.
10. Dyck JR and Lopaschuk GD. AMPK alterations in cardiac physiology and pathology: enemy or ally? J Physiol 574: 95-112, 2006.
Page 16 of 27
Copyright Information

Zhou et al, H-00557-2007.R1
16
11. Folmes CD, Clanachan AS and Lopaschuk GD. Fatty acids attenuate insulin regulation of 5'-AMP-activated protein kinase and insulin cardioprotection after ischemia. Circ Res 99: 61-68, 2006.
12. Gertz EW, Wisneski JA, Stanley WC and Neese RA. Myocardial substrate utilization during exercise in humans. Dual carbon-labeled carbohydrate isotope experiments. J Clin Invest 82: 2017-2025, 1988.
13. Goodwin GW and Taegtmeyer H. Regulation of fatty acid oxidation of the heart by MCD and ACC during contractile stimulation. Am J Physiol 277: E772-E777, 1999.
14. Goodwin GW, Taylor CS and Taegtmeyer H. Regulation of energy metabolism of the heart during acute increase in heart work. J Biol Chem 273: 29530-29539, 1998.
15. Hall JL, Lopaschuk GD, Barr A, Bringas J, Pizzurro RD and Stanley WC. Increased cardiac fatty acid uptake with dobutamine infusion in swine is accompanied by a decrease in maloanyl CoA levels. Circ Res 32: 879-885, 1996.
16. Hall JL, Stanley WC, Lopaschuk GD, Wisneski JA, Pizzurro RD, Hamilton CD and McCormack JG. Impaired pyruvate osidation but normal glucose uptake in diabetic pig heart during dobutamine induced work. Am J Physiol 271: H2320-H2329, 1996.
17. Hall JL, Van Wylen DG, Pizzurro RD, Hamilton CD, Reiling CM and Stanley WC. Myocardial interstitial purine metabolites and lactate with increased work in swine. Cardiovasc Res 30: 351-356, 1995.
18. Hamilton C and Saggerson ED. Malonyl-CoA metabolism in cardiac myocytes. Biochem J350 Pt 1: 61-67, 2000.
19. Higgins AJ, Morville M, Burges RA and Blackburn KJ. Mechanism of action of oxfenicine on muscle metabolism. Biochem Biophys Res Commun 100: 291-296, 1981.
20. How OJ, Aasum E, Kunnathu S, Severson DL, Myhre ES and Larsen TS. Influence of substrate supply on cardiac efficiency, as measured by pressure-volume analysis in ex vivo mouse hearts. Am J Physiol Heart Circ Physiol 288: H2979-H2985, 2005.
21. Hutter JF, Schweickhardt C, Piper HM and Spieckermann PG. Inhibition of fatty acid oxidation and decrease of oxygen consumption of working rat heart by 4-bromocrotonic acid. J Mol Cell Cardiol 16: 105-108, 1984.
22. Kerner J and Hoppel C. Fatty acid import into mitochondria. Biochim Biophys Acta 1486: 1-17, 2000.
23. Khouri EM, Gregg DE and Rayford CR. Effect of exercise on cardiac output, left coronary flow and myocardial metabolism in the unanesthetized dog. Circ Res 17: 427-437, 1965.
Page 17 of 27
Copyright Information

Zhou et al, H-00557-2007.R1
17
24. King KL, Okere IC, Sharma N, Dyck JR, Reszko AE, McElfresh TA, Kerner J, Chandler MP, Lopaschuk GD and Stanley WC. Regulation of cardiac malonyl-CoA content and fatty acid oxidation during increased cardiac power. Am J Physiol Heart Circ Physiol 289: H1033-H1037, 2005.
25. Korvald C, Elvenes OP and Myrmel T. Myocardial substrate metabolism influences left ventricular energetics in vivo. Am J Physiol Heart Circ Physiol 278: H1345-H1351, 2000.
26. Lassers BW, Wahlqvist ML, Kaijser L and Carlson LA. Effect of nicotinic acid on myocardial metabolism in man at rest and during exercise. J Appl Physiol 33: 72-80, 1972.
27. Lei B, Matsuo K, Labinskyy V, Sharma N, Chandler MP, Ahn A, Hintze TH, Stanley WC and Recchia FA. Exogenous nitric oxide reduces glucose transporters translocation and lactate production in ischemic myocardium in vivo. Proc Natl Acad Sci U S A 102: 6966-6971, 2005.
28. Liu Q, Docherty JC, Rendell JC, Clanachan AS and Lopaschuk GD. High levels of fatty acids delay the recovery of intracellular pH and cardiac efficiency in post-ischemic hearts by inhibiting glucose oxidation. J Am Coll Cardiol 39: 718-725, 2002.
29. Lopaschuk GD and Stanley WC. Malonyl-CoA decarboxylase inhibition as a novel approach to treat ischemic heart disease. Cardiovasc Drugs Ther 20: 433-439, 2006.
30. Mjos OD. Effect of free fatty acids on myocardial function and oxygen consumption in intact dogs. J Clin Invest 50: 1386-1389, 1971.
31. Mjos OD and Kjekshus J. Increased local metabolic rate by free fatty acids in the intact dog heart. Scand J Clin Lab Invest 28: 389-393, 1971.
32. Mjos OD, Kjekshus JK and Lekven J. Importance of free fatty acids as a determinant of myocardial oxygen consumption and myocardial ischemic injury during norepinephrine infusion in dogs. J Clin Invest 53: 1290-1299, 1974.
33. Musi N, Hirshman MF, Arad M, Xing Y, Fujii N, Pomerleau J, Ahmad F, Berul CI, Seidman JG, Tian R and Goodyear LJ. Functional role of AMP-activated protein kinase in the heart during exercise. FEBS Lett 579: 2045-2050, 2005.
34. Panchal AR, Comte B, Huang H, Kerwin T, Darvish A, Des Rosiers C, Brunengraber H and Stanley WC. Partitioning of pyruvate between oxidation and anaplerosis in swine hearts. Am J Physiol Heart Circ Physiol 279: H2390-H2398, 2000.
35. Randle PJ, Garland PB, HALES CN and Newsholme EA. The glucose fatty-acid cycle. Its role in insulin sensitivity and the metabolic disturbances of diabetes mellitus. Lancet 1: 785-789, 1963.
36. Reszko AE, Kasumov T, David F, Thomas KR, Jobbins KA, Cheng JF, Lopaschuk GD, Dyck JR, Diaz M, des RC, Stanley WC and Brunengraber H. Regulation of malonyl-
Page 18 of 27
Copyright Information

Zhou et al, H-00557-2007.R1
18
CoA concentration and turnover in the normal heart. J Biol Chem 279: 34298-34301, 2004.
37. Saddik M, Gamble J, Witters LA and Lopaschuk GD. Acetyl-CoA carboxylase regulation of fatty acid oxidation in the heart. J Biol Chem 268: 25836-25845, 1993.
38. Sharma N, Okere IC, Brunengraber DZ, McElfresh TA, King KL, Sterk JP, Huang H, Chandler MP and Stanley WC. Regulation of pyruvate dehydrogenase activity and citric acid cycle intermediates during high cardiac power generation. J Physiol 562: 593-603, 2005.
39. Simonsen S and Kjekshus JK. The effect of free fatty acids on myocardial oxygen consumption during atrial pacing and catecholamine infusion in man. Circulation 58: 484-491, 1978.
40. Stanley WC, Hernandez LA, Spires D, Bringas J, Wallace S and McCormack JG. Pyruvate dehydrogenase activity and malonyl CoA levels in normal and ischemic swine myocardium: effects of dichloroacetate. J Mol Cell Cardiol 28: 905-914, 1996.
41. Stanley WC, Lopaschuk GD, Hall JL and McCormack JG. Regulation of myocardial carbohydrate metabolism under normal and ischemic conditions. Cardiovascular Research 33: 243-257, 1997.
42. Stanley WC. Partial fatty acid oxidation inhibitors for stable angina. Expert Opin Investig Drugs 11: 615-629, 2002.
43. Stanley WC, Morgan EE, Huang H, McElfresh TA, Sterk JP, Okere IC, Chandler MP, Cheng J, Dyck JR and Lopaschuk GD. Malonyl-CoA decarboxylase inhibition suppresses fatty acid oxidation and reduces lactate production during demand-induced ischemia. Am J Physiol Heart Circ Physiol 289: H2304-H2309, 2005.
44. Stanley WC, Recchia FA and Lopaschuk GD. Myocardial substrate metabolism in the normal and failing heart. Physiol Rev 85: 1093-1129, 2005.
45. Stone CK, Holden JE, Stanley W and Perlman SB. Effect of nicotinic acid on exogenous myocardial glucose utilization. J Nucl Med 36: 996-1002, 1995.
46. Suga H. Ventricular energetics. Physiol Rev 70: 247-277, 1990.
47. Zammit VA, Fraser F and Orstorphine CG. Regulation of mitochondrial outer-membrane carnitine palmitoyltransferase (CPT I): role of membrane-topology. Adv Enzyme Regul37: 295-317, 1997.
48. Zhou L, Cabrera ME, Huang H, Yuan CL, Monika DK, Sharma N, Bian F and Stanley WC. Parallel activation of mitochondrial oxidative metabolism with increased cardiac energy expenditure is not dependent on fatty acid oxidation in pigs. J Physiol 579: 811-821, 2007.
Page 19 of 27
Copyright Information
Zhou et al, H-00557-2007.R1
19
Table 1. Hemodynamic responses to short-term increased cardiac power with or without pretreatment with oxfenicine in pigs.
DOB (n=8) DOB + OXF (n=8)CON(n=8) Baseline 45 sec 5 min Baseline 45 sec 5 min
Stroke Work (J) 0.33+0.03 0.27+0.03 0.39+0.06* 0.35+0.06 0.26+0.03 0.49+0.07* 0.40+0.05
Cardiac Output (L/min)
2.8+0.3 2.5+0.3 3.7+0.5* 3.2+0.4 2.3+0.3 3.8+0.8* 3.7+0.8
Peak +dP/dt, (mmHg/sec)
1500+170 1420 +120 4720+540* 4920+670* 1320+150 5910+400* 6430+440*
LV energy expenditure
(watts)1.41+0.12 1.69+0.19 4.04+0.39* 4.81+0.40* 1.41+0.17 3.96+0.42* 5.21+0.54*
Values are means + SE. *: P<0.05 vs. CON
Page 20 of 27
Copyright Information
Zhou et al, H-00557-2007.R1
20
Table 2. Summary of Western blot assessment of total and phosphorylated AMPK and ACC. Data expressed in arbitrary units.
CON DOB DOB+OXF
Total AMPK 247±9 237±21 281±32
Phospho-AMPK 154±3 88±17* 127±11
Total ACC 48±8 60±7 69±12
Phospho-ACC 29±2 33±5 41±3
Phospho/Total ACC 0.70±0.13 0.58±0.10 0.86±0.31
* P< 0.05 compared to CON; † P<0.002 compared to CON
Page 21 of 27
Copyright Information
Zhou et al, H-00557-2007.R1
21
Table 3. Myocardial content of adenine nucleotides (µmol/g).
CON DOB DOB+OXF
ATP 4.64±0.11 3.64±0.15* 3.47±0.23*
ADP 1.08±0.02 1.12±0.05 1.13±0.03
AMP 0.159±0.004 0.180±0.019 0.181±0.011
* P<0.004 compared to CON
Page 22 of 27
Copyright Information

Zhou et al, H-00557-2007.R1
22
Figure Legend.
Fig. 1. Effect of oxfenicine on myocardial left ventricular external power (top panel) and
mechanical efficiency (lower panel) under normal conditions and at 45 seconds and 5 minutes of
the period of increased cardiac workload. *p<0.05 vs. baseline, † p<0.05 vs DOB at the same
time point.
Fig. 2. Net myocardial glucose (top), lactate (middle) and fatty acid (lower) uptake as a function
of time for DOB and DOB + OXF groups under resting condition and during increased cardiac
energy expenditure.
Fig. 3. Myocardial malonyl-CoA content and the rate of fatty acid oxidation at the end of
protocol. * p<0.05 vs. control group (CON).
Fig 4. Myocardial phospho-AMPK/total AMPK as measured by western blot (top panel), and myocardial AMPK activity (lower panel) * p<0.05 vs. control group (CON).
Page 23 of 27
Copyright Information
Zhou et al, H-00557-2007.R1
23
Fig. 1.
0
0.5
1.0
1.5
2.0
2.5
Pre 45 Sec 5 min Pre 45 Sec 5 min
LVPo
wer
(wat
t)
Untreated OXF
††
Untreated OXF
LVEf
fici
ency
(%)
0
20
40
60
Pre 45 Sec 5 min Pre 45 Sec 5 min
†
†
* *
* *
Page 24 of 27
Copyright Information
Zhou et al, H-00557-2007.R1
24
Fig. 2.
0.0
0.2
0.4
0.6
0.8 DOBDOB + OXF
-0.2
0.0
0.2
0.4
0.6
0.8
Lac
tate
Up
take
(µm
ol/g
/min
)
0
40
80
120
160
Fre
e F
atty
Aci
d U
pta
ke(n
mo
l/g/m
in)
-6 0 1 2 3 4 5Dobutamine + Aortic Constriction
-6 0 1 2 3 4 5Dobutamine + Aortic Constriction
-6 0 1 2 3 4 5Dobutamine + Aortic Constriction
-6 0 1 2 3 4 5Dobutamine + Aortic Constriction
0 1 2 3 4 5Dobutamine + Aortic Constriction
-6 0 1 2 3 4 5Dobutamine + Aortic Constriction
-6 0 1 2 3 4 5Dobutamine + Aortic Constriction
-6 0 1 2 3 4 5Dobutamine + Aortic Constriction
Time (minutes)
P < 0.007 DOB vsDOB+OXF
P < 0.001 DOB vsDOB+OXF
P < 0.001 DOB vsDOB+OXF
Glu
cose
Up
take
(µm
ol/g
/min
)
Page 25 of 27
Copyright Information

Zhou et al, H-00557-2007.R1
25
Fig. 3
0.0
0.5
1.0
1.5
2.0
2.5
CON DOB DOB + OXF
Myo
card
ial M
alo
nyl
-Co
A(µ
mo
l/g)
Myo
card
ial F
AO
(n
mo
l/g/m
in)
0
40
80
120
160
CON DOB DOB+ OXF
**
*
Page 26 of 27
Copyright Information
Zhou et al, H-00557-2007.R1
26
Figure 4.
0.0
0.1
0.2
0.3
0.4
0.5
0.6
0.7
0
90
180
270
CON DOB DOB + OXF
*
*
CON DOB DOB + OXF
P-AMPK/total AMPK
AMPK Activity
pmol
sm
g pr
otei
n-1
min
-1A
rbitr
iary
units
Page 27 of 27
Copyright Information
Related Documents











