Original Article Tamoxifen-Induced Anorexia Is Associated With Fatty Acid Synthase Inhibition in the Ventromedial Nucleus of the Hypothalamus and Accumulation of Malonyl-CoA Miguel Lo ´ pez, 1 Christopher J. Lelliott, 1 Sulay Tovar, 2 Wendy Kimber, 1 Rosalı ´a Gallego, 3 Sam Virtue, 1 Margaret Blount, 1 Maria J. Va ´ zquez, 2 Nick Finer, 1 Trevor J. Powles, 4 Stephen O’Rahilly, 1 Asish K. Saha, 5 Carlos Die ´ guez, 2 and Antonio J. Vidal-Puig 1 Fatty acid metabolism in the hypothalamus has recently been shown to regulate feeding. The selective estrogen receptor modulator tamoxifen (TMX) exerts a potent an- orectic effect. Here, we show that the anorectic effect of TMX is associated with the accumulation of malonyl-CoA in the hypothalamus and inhibition of fatty acid synthase (FAS) expression specifically in the ventromedial nucleus of the hypothalamus (VMN). Furthermore, we demonstrate that FAS mRNA expression is physiologically regulated by fasting and refeeding in the VMN but not in other hypotha- lamic nuclei. Thus, the VMN appears to be the hypothalamic site where regulation of FAS and feeding converge. Sup- porting the potential clinical relevance of these observa- tions, reanalysis of a primary breast cancer prevention study showed that obese women treated with TMX gained significantly less body weight over a 6-year period than obese women given placebo. The finding that TMX can modulate appetite through alterations in FAS expression and malonyl-CoA levels suggests a link between hypotha- lamic sex steroid receptors, fatty acid metabolism, and feeding behavior. Diabetes 55:1327–1336, 2006 T he clinical use of drugs designed to treat specific diseases occasionally reveals unexpected effects of potential therapeutic significance. It has been documented that certain antipsychotic (1) or antidepressant (2) drugs also have effects in food intake and body weight. Although it is unlikely that these drugs might one day be used as treatment for obesity, the discovery of their mechanisms of action may identify important pathways involved in energy homeostasis that could be targeted for antiobesity drug development. Following a similar rationale, we investigated the mo- lecular mechanisms mediating the anorectic effects of tamoxifen (TMX), a selective estrogen receptor modulator widely used for the treatment of breast cancers (3,4). The effect of TMX on energy homeostasis has been studied in rodents and has been shown to markedly decrease food intake and body weight (5,6). In spite of this, the molecular mechanism for such anorectic effects has not been identified. Feeding is regulated at hypothalamic levels by orexi- genic and anorexigenic neuropeptides. Neuropeptide Y, agouti-related protein, and melanin-concentrating hor- mone exert orexigenic actions, whereas -melanocyte– stimulating hormone and cocaine- and amphetamine- regulated transcript (CART) peptides are the main anorexigenic signals. The expression of these neuropep- tides is under the control of hormonal and nutritional signals (7–12). More recently, it has been suggested that the precursors and break-down products of the fatty acid synthesis may be important metabolic signals regulating hypothalamic neuropeptides (10,13,14). The pathway of lipogenesis de novo and the hypothalamic levels of malo- nyl-CoA have been also identified as a key intermediate controlling food intake. The cellular level of malonyl-CoA depends on its rate of synthesis, catalyzed by acetyl-CoA carboxylase (ACC), relative to its rate of utilization, cata- lyzed by fatty acid synthase (FAS), and degradation, catalyzed by malonyl-CoA decarboxylase (15,16). While it is well established that the lipogenic enzymes are ex- pressed in the arcuate (ARC), dorsomedial, paraventricu- lar (PVN), and ventromedial (VMN) hypothalamic nuclei (17,18), their nuclei-specific regulation in response to nutrition has not previously been studied. Recent data have linked hypothalamic FAS to feeding regulation. It has been reported that administration of the FAS inhibitors cerulenin and C75 inhibits food intake and induces weight From the 1 Department of Clinical Biochemistry, University of Cambridge, Addenbrooke’s Hospital, Cambridge, U.K.; the 2 Department of Physiology, School of Medicine, University of Santiago de Compostela, Santiago de Compostela, Spain; the 3 Department of Morphological Sciences, School of Medicine, University of Santiago de Compostela, Santiago de Compostela, Spain; the 4 Parkside Oncology Clinic, London, U.K.; and the 5 Diabetes Research Unit, EBRC-827, Boston Medical Centre, Boston, Massachusetts. Address correspondence and reprint requests to Antonio J. Vidal-Puig, PhD, MD, Department of Clinical Biochemistry, University of Cambridge, Adden- brooke’s Hospital, Hills Road Cambridge, CB2 2QR, U.K. E-mail: ajv22@ cam.ac.uk. Received for publication 18 October 2005 and accepted in revised form 1 February 2006. M.L. and C.J.L. contributed equally to this work. Additional information for this article can be found in an online appendix at http://diabetes.diabetesjournals.org. ACC, acetyl-CoA carboxylase; AMPK, AMP-activated protein kinase; ARC, arcuate nucleus of the hypothalamus; CART, cocaine- and amphetamine- regulated transcript; FAS, fatty acid synthase; LHA, lateral hypothalamic area; POMC, proopiomelanocortin; PVN, paraventricular nucleus of the hypothala- mus; RMH, Royal Marsden Hospital; TMX, tamoxifen; TOFA, 5-(tetradecy- loxy)-2-furoic acid; VMN, ventromedial nucleus of the hypothalamus. DOI: 10.2337/db05-1356 © 2006 by the American Diabetes Association. The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “advertisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact. DIABETES, VOL. 55, MAY 2006 1327

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Original Article
Tamoxifen-Induced Anorexia Is Associated With FattyAcid Synthase Inhibition in the Ventromedial Nucleus ofthe Hypothalamus and Accumulation of Malonyl-CoAMiguel Lopez,
1Christopher J. Lelliott,
1Sulay Tovar,
2Wendy Kimber,
1Rosalıa Gallego,
3Sam Virtue,
1
Margaret Blount,1
Maria J. Vazquez,2
Nick Finer,1
Trevor J. Powles,4
Stephen O’Rahilly,1
Asish K. Saha,5
Carlos Dieguez,2
and Antonio J. Vidal-Puig1
Fatty acid metabolism in the hypothalamus has recently
been shown to regulate feeding. The selective estrogen
receptor modulator tamoxifen (TMX) exerts a potent an-
orectic effect. Here, we show that the anorectic effect of
TMX is associated with the accumulation of malonyl-CoA in
the hypothalamus and inhibition of fatty acid synthase
(FAS) expression specifically in the ventromedial nucleus
of the hypothalamus (VMN). Furthermore, we demonstrate
that FAS mRNA expression is physiologically regulated by
fasting and refeeding in the VMN but not in other hypotha-
lamic nuclei. Thus, the VMN appears to be the hypothalamic
site where regulation of FAS and feeding converge. Sup-
porting the potential clinical relevance of these observa-
tions, reanalysis of a primary breast cancer prevention
study showed that obese women treated with TMX gained
significantly less body weight over a 6-year period than
obese women given placebo. The finding that TMX can
modulate appetite through alterations in FAS expression
and malonyl-CoA levels suggests a link between hypotha-
lamic sex steroid receptors, fatty acid metabolism, and
feeding behavior. Diabetes 55:1327–1336, 2006
The clinical use of drugs designed to treat specificdiseases occasionally reveals unexpected effectsof potential therapeutic significance. It has beendocumented that certain antipsychotic (1) or
antidepressant (2) drugs also have effects in food intakeand body weight. Although it is unlikely that these drugsmight one day be used as treatment for obesity, thediscovery of their mechanisms of action may identifyimportant pathways involved in energy homeostasis thatcould be targeted for antiobesity drug development.
Following a similar rationale, we investigated the mo-lecular mechanisms mediating the anorectic effects oftamoxifen (TMX), a selective estrogen receptor modulatorwidely used for the treatment of breast cancers (3,4). Theeffect of TMX on energy homeostasis has been studied inrodents and has been shown to markedly decrease foodintake and body weight (5,6). In spite of this, the molecularmechanism for such anorectic effects has not beenidentified.
Feeding is regulated at hypothalamic levels by orexi-genic and anorexigenic neuropeptides. Neuropeptide Y,agouti-related protein, and melanin-concentrating hor-mone exert orexigenic actions, whereas �-melanocyte–stimulating hormone and cocaine- and amphetamine-regulated transcript (CART) peptides are the mainanorexigenic signals. The expression of these neuropep-tides is under the control of hormonal and nutritionalsignals (7–12). More recently, it has been suggested thatthe precursors and break-down products of the fatty acidsynthesis may be important metabolic signals regulatinghypothalamic neuropeptides (10,13,14). The pathway oflipogenesis de novo and the hypothalamic levels of malo-nyl-CoA have been also identified as a key intermediatecontrolling food intake. The cellular level of malonyl-CoAdepends on its rate of synthesis, catalyzed by acetyl-CoAcarboxylase (ACC), relative to its rate of utilization, cata-lyzed by fatty acid synthase (FAS), and degradation,catalyzed by malonyl-CoA decarboxylase (15,16). While itis well established that the lipogenic enzymes are ex-pressed in the arcuate (ARC), dorsomedial, paraventricu-lar (PVN), and ventromedial (VMN) hypothalamic nuclei(17,18), their nuclei-specific regulation in response tonutrition has not previously been studied. Recent datahave linked hypothalamic FAS to feeding regulation. It hasbeen reported that administration of the FAS inhibitorscerulenin and C75 inhibits food intake and induces weight
From the 1Department of Clinical Biochemistry, University of Cambridge,Addenbrooke’s Hospital, Cambridge, U.K.; the 2Department of Physiology,School of Medicine, University of Santiago de Compostela, Santiago deCompostela, Spain; the 3Department of Morphological Sciences, School ofMedicine, University of Santiago de Compostela, Santiago de Compostela,Spain; the 4Parkside Oncology Clinic, London, U.K.; and the 5DiabetesResearch Unit, EBRC-827, Boston Medical Centre, Boston, Massachusetts.
Address correspondence and reprint requests to Antonio J. Vidal-Puig, PhD,MD, Department of Clinical Biochemistry, University of Cambridge, Adden-brooke’s Hospital, Hills Road Cambridge, CB2 2QR, U.K. E-mail: [email protected].
Received for publication 18 October 2005 and accepted in revised form 1February 2006.
M.L. and C.J.L. contributed equally to this work.Additional information for this article can be found in an online appendix at
http://diabetes.diabetesjournals.org.ACC, acetyl-CoA carboxylase; AMPK, AMP-activated protein kinase; ARC,
arcuate nucleus of the hypothalamus; CART, cocaine- and amphetamine-regulated transcript; FAS, fatty acid synthase; LHA, lateral hypothalamic area;POMC, proopiomelanocortin; PVN, paraventricular nucleus of the hypothala-mus; RMH, Royal Marsden Hospital; TMX, tamoxifen; TOFA, 5-(tetradecy-loxy)-2-furoic acid; VMN, ventromedial nucleus of the hypothalamus.
DOI: 10.2337/db05-1356© 2006 by the American Diabetes Association.The costs of publication of this article were defrayed in part by the payment of page
charges. This article must therefore be hereby marked “advertisement” in accordance
with 18 U.S.C. Section 1734 solely to indicate this fact.
DIABETES, VOL. 55, MAY 2006 1327
loss (13,19,20). Interestingly, this anorectic effect requiresmalonyl-CoA accumulation (14). Although it is not likelythat these drugs become treatments for obesity, the evi-dence that selective inhibition of FAS can control feedingraises questions about the physiological involvement offatty acid biosynthesis controlling food intake.
Using a systems biology approach, we have recentlyidentified a primary action of TMX selectively decreasingthe expression and activity of FAS in liver (21). Byanalogy, we studied whether the anorectic effect of TMXmay be mediated through the inhibition of FAS in thehypothalamus. We also investigated the potential rele-vance of these findings to humans by reanalyzing datafrom the Royal Marsden Hospital (RMH) trial (3).
RESEARCH DESIGN AND METHODS
Human studies. We investigated body weight changes in the RMH trial (3)(online appendix [available at http://diabetes.diabetesjournals.org]). The ef-fects of TMX or placebo on body weight were analyzed in 2,329 subjects forwhom BMI data were available and who did not develop cancer during thestudy. The trial was approved by the RMH Ethics Committee (3).Rat studies. We used Wistar rats (450–500 g; Charles River Laboratories),lean Zucker rats [Crl:(ZUC)-faBR, 300–350 g; The Jackson Laboratories], andobese Zucker rats [Crl:(ZUC)-faBR, 475–525 g; The Jackson Laboratories]age-matched (11–12 weeks) with lean Zucker rats. Rats were housed in atemperature-controlled room, with a 12-h light/dark cycle (lights on at 8:00A.M. and off at 8:00 P.M.). All experiments were conducted in accordance withHome Office Guidelines and the Ethics Committee of the University ofSantiago de Compostela. Male rats were used to exclude the confoundingeffects of the estrous cycle on food intake (22,23).Subcutaneous TMX treatment. TMX (Sigma) was dissolved in sesame oilcontaining 1% benzyl alcohol (5). The control group received a daily subcu-taneous injection of vehicle and was provided standard rodent diet ad libitum.The TMX group received daily subcutaneous injections of TMX and was alsoprovided ad libitum food. To monitor the effects of food intake, a pair-fed
group was also included; for 5 days, the pair-fed group received the averageamount of food eaten by the TMX-treated rats and received vehicle injections.For the hypothalamic studies, a dose of 0.5 mg � kg�1 � day�1 TMX wasadministered for 5 days. On day 5, rats were killed 4 h after TMX treatment.Zucker lean and obese rats (n � 10–12 animals/group) were also treatedsubcutaneously with TMX (0.5 mg � kg�1 � day�1) for 5 days. On the final dayof the protocol, rats were given a final dose of TMX (9:00 A.M.) and killed 4 hlater. To test the effect of TMX treatment on refeeding, rats were fasted for24 h before TMX administration. Next, the rats (n � 9–10 animals/group) weretreated with vehicle or TMX (0.5 mg/kg) and allowed to eat ad libitum. In allthe protocols, TMX was administered subcutaneously at 9:00 A.M.Intracerebroventricular TMX citrate treatment. TMX citrate was usedfor intracerebroventricular experiments because it can be dissolved in aque-ous solutions. Chronic (intracerebroventricular) cannulae were stereotaxi-cally implanted under ketamine-xylacine anesthesia (50 mg/kg i.p.) in thelateral ventricle as previously described (24). Correct localization of thecannulae was confirmed by histological analysis. Before TMX citrate admin-istration, rats (n � 9 animals/group) were fasted for 24 h in order to adjust therats to the same pretreatment levels of stimulated appetite response. Ratsreceived either a single administration of TMX citrate (10 �g in 10 �l2-hydroxypropyl-�-cyclodextrin; Sigma) or vehicle (10 �l, control rats). Sincethere are no reported studies about intracerebroventricular treatments withTMX, we selected this dose on the basis of previous literature based uponintracerebroventricular administration of other estrogenic compounds (25). Athird group of animals was treated with an equimolar dose of citrate (Sigma)to that of the TMX citrate animals.Conditioned taste aversion. Five days before the test, rats were allowed 2 hdaytime access to water. On the day of the experiment, rats (n � 16rats/group) were given access to 0.15% sodium saccharin rather than water for30 min. Immediately afterward, one experimental group was injected intra-peritoneally with 0.15 mol/l lithium chloride (LiCl) in saline (26,27) andsubcutaneously with sesame oil containing 1% benzyl alcohol (vehicle). Thesecond experimental group was injected intraperitoneally with saline andsubcutaneously with TMX (0.5 mg/kg). A third experimental group (controlgroup) was injected intraperitoneally with saline and subcutaneously withvehicle. Twenty-four hours later, rats were given 2 h access to a two-bottlechoice test of 0.15% saccharin versus water. Data were expressed as percent
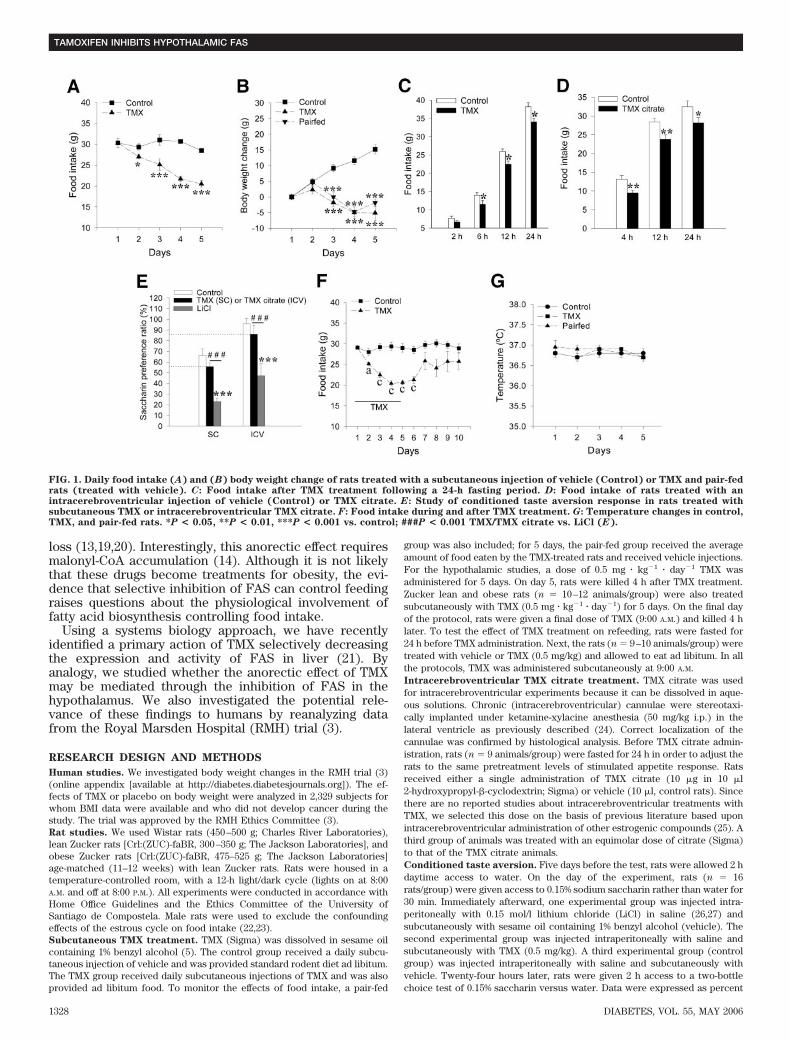
FIG. 1. Daily food intake (A) and (B) body weight change of rats treated with a subcutaneous injection of vehicle (Control) or TMX and pair-fedrats (treated with vehicle). C: Food intake after TMX treatment following a 24-h fasting period. D: Food intake of rats treated with anintracerebroventricular injection of vehicle (Control) or TMX citrate. E: Study of conditioned taste aversion response in rats treated withsubcutaneous TMX or intracerebroventricular TMX citrate. F: Food intake during and after TMX treatment. G: Temperature changes in control,TMX, and pair-fed rats. *P < 0.05, **P < 0.01, ***P < 0.001 vs. control; ###P < 0.001 TMX/TMX citrate vs. LiCl (E).
TAMOXIFEN INHIBITS HYPOTHALAMIC FAS
1328 DIABETES, VOL. 55, MAY 2006
saccharin preference ratio (100 � saccharin intake/[saccharin intake � waterintake]).
For the intracerebroventricular-conditioned taste aversion studies, onegroup was injected intraperitoneally with 0.15 mol/l LiCl in saline andintracerebroventricularlly with vehicle (10% 2-hydroxypropyl-�-cyclodextrin);the second group was injected intraperitoneally with saline and intracerebrov-entricularlly with TMX citrate (10 �g). A third group (control group) wasinjected intraperitoneally with saline and intracerebroventricularlly withvehicle. The experiment proceeded as the subcutaneous treatment (n � 16rats/group).Intracerebroventricular SHU9119 and 5-(tetradecyloxy)-2-furoic acid
treatments. Rats were fasted for 24 h (n � 8–12 animals/group). To evaluatethe effect of melanocortin receptors on the anorectic actions of TMX, the ratsreceived a single intracerebroventricular administration of saline or themelanocortin-3/-4 receptors antagonist SHU9119 (3 nmol in saline; Phoenix)(28,29). To test the effect of the inhibition of ACC on the anorectic actions of
TMX, the rats were treated with a single intracerebroventricular administra-tion of vehicle (DMSO) or an ACC inhibitor: 5-(tetradecyloxy)-2-furoic acid(TOFA; 10 �g) (14). Subsequently, these rats received either vehicle or TMX(0.5 mg/kg) administered subcutaneously.Blood biochemistry. Kits for the measurement of cholesterol, glucose, andtriglycerides were obtainded from Roche (21,30). Free fatty acids weremeasured using the NEFA-C colorimetric kit (Wako Chemicals) (21,30).Plasma leptin and insulin levels were measured by ELISA kits (Crystal)(21,30).Real-time quantitative PCR. The mRNA levels of estrogen receptor � andprogesterone receptor were studied by using real-time PCR (TaqMan) (onlineappendix Table 1 and online appendix methods) as previously described (30).We used six to eight rats per group.In situ hybridization. Coronal brain sections (16 �m) were probed withspecific antisense oligos (online appendix Table 2). In situ hybridizations wereperformed as previously published (24,31,32) (online appendix methods). Weused 10–19 rats per group.Immunohistochemistry. Diaminobenzidine immunohistochemistry was per-formed as described (33) (online appendix methods) using a c-FOS antibody(Santa Cruz). We used six to eight rats per group.Western blotting. Hypothalamus total protein lysates were subjected toSDS-PAGE, electrotransferred on a polyvinylidine fluoride membrane, andprobed with the indicated antibodies: ACC, pACC-Ser79, AMP-activated pro-tein kinase (AMPK)�1, and AMPK�2 (Upstate); pAMPK-Thr172 (Cell Signal-ing); and �-actin (Abcam). For protein detection, we used horseradishperoxidase–conjugated secondary antibodies and chemiluminescence (Amer-sham) (online appendix methods). We used 8–12 rats per group.Kinase assay. Hypothalamic protein extracts were incubated at 4°C with thecorresponding antibody conjugated to protein G-Sepharose (34). Phospho-transferase activity toward the AMARA-peptide for AMPK activity (35) orLKB1tide-peptide (Upstate) was then measured (36) (online appendix meth-ods). We used 8–12 rats/group.Malonyl-CoA assay. Malonyl-CoA was measured by radioisotopic method inneutralized perchloric acid filtrates (37,38) (online appendix methods). Weused 8–10 rats per group.Statistical analysis. Data were expressed as means � SE and analyzed byusing StatView 4.57 (Abacus Concepts). Statistical significance was deter-mined by ANOVA and post hoc Bonferroni test. P 0.05 was consideredsignificant. Protein and mRNA levels were presented as percentage change inrelation to control group (vehicle treated or fed ad libitum).
RESULTS
Subcutaneous administration of TMX reduces foodintake and body weight. Subcutaneous administration ofTMX to rats markedly decreased food intake (Fig. 1A andonline appendix Fig. 1A) and body weight (Fig. 1B andonline appendix Fig. 1B). To determine whether the an-orexigenic effect of TMX was dependent on feeding state,subcutaneous TMX was administered to 24-h fasted rats.TMX significantly attenuated refeeding at 6, 12, and 24 h(Fig. 1C).
We detect no differences in plasma glucose, triglycer-ides, and free fatty acids among the experimental groups(online appendix Table 3). Both TMX-treated and pair-fedrats showed significant decreases in insulin and leptinwhen compared with vehicle-treated rats (online appendixTable 3). Moreover, as it has been previously demon-strated (21,39), serum total, HDL, and LDL cholesterolwere all reduced in the TMX group when compared withthe ad libitum and pair-fed groups (online appendix Table3).Intracerebroventricular administration of TMX re-duces food intake. Since TMX is insoluble in an aqueoussolution, TMX citrate was used for intracerebroventricularexperiments. We confirmed that TMX citrate subcutane-ous administration, at an equimolar dose to TMX (0.5 mg �kg�1 � day�1), recapitulated the effects observed with TMX(online appendix Fig. 1C and D). Intracerebroventricularadministration of TMX citrate to 24-h fasted rats decreasedrefeeding at 4, 12, and 24 h after treatment (Fig. 1D). Thisanorectic effect was specific for TMX because administra-
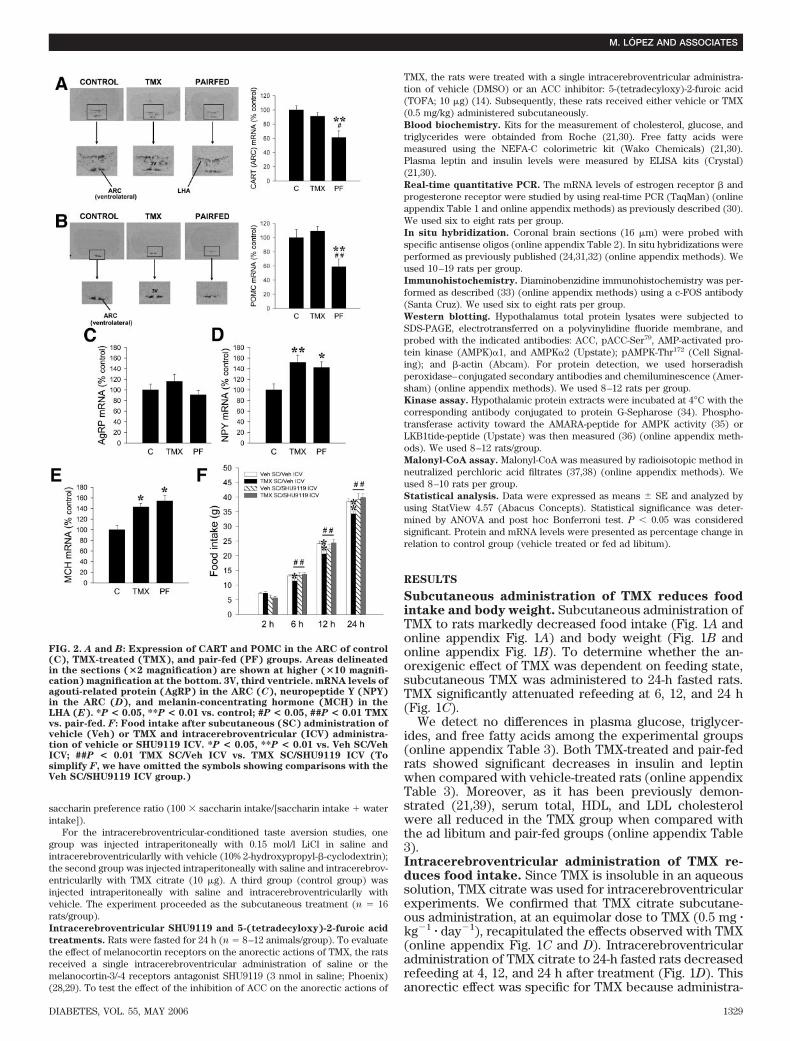
FIG. 2. A and B: Expression of CART and POMC in the ARC of control(C), TMX-treated (TMX), and pair-fed (PF) groups. Areas delineatedin the sections (�2 magnification) are shown at higher (�10 magnifi-cation) magnification at the bottom. 3V, third ventricle. mRNA levels ofagouti-related protein (AgRP) in the ARC (C), neuropeptide Y (NPY)in the ARC (D), and melanin-concentrating hormone (MCH) in theLHA (E). *P < 0.05, **P < 0.01 vs. control; #P < 0.05, ##P < 0.01 TMXvs. pair-fed. F: Food intake after subcutaneous (SC) administration ofvehicle (Veh) or TMX and intracerebroventricular (ICV) administra-tion of vehicle or SHU9119 ICV. *P < 0.05, **P < 0.01 vs. Veh SC/VehICV; ##P < 0.01 TMX SC/Veh ICV vs. TMX SC/SHU9119 ICV (Tosimplify F, we have omitted the symbols showing comparisons with theVeh SC/SHU9119 ICV group.)
M. LOPEZ AND ASSOCIATES
DIABETES, VOL. 55, MAY 2006 1329
tion of citrate alone induced no changes in food intake (4h: control, 14.2 � 0.9 vs. citrate, 14.1 � 1.2 g). Moreover,intracerebroventricular administration of TMX citrate didnot affect plasma glucose, insulin, or leptin levels (onlineappendix Table 4), suggesting that the anorectic effect ofTMX was not the result of peripheral secondary effects ofTMX. Furthermore, we showed that subcutaneous admin-istration of TMX at an equivalent dose to the intracerebro-ventricular treatment (10 �g i.c.v. � 25 �g � kg�1 � day�1
s.c.) does not affect feeding, body weight (online appendixFig. 1E and F), plasma glucose, insulin, or leptin levels(online appendix Table 5). These data make any peripheraleffect of centrally administered TMX diffusing from thecentral nervous system unlikely.
To confirm that TMX was bioactive at hypothalamiclevels, we studied the mRNA levels of estrogen receptor �and progesterone receptor of TMX-treated rats. As previ-ously reported, the mRNA expression of estrogen receptor� (40) and progesterone receptor (41) was increased in thehypothalamus after TMX treatment (online appendix Fig.2). These results indicate that the anorexic action ofintracerebroventricular TMX citrate is mediated by centralmechanisms.
The anorectic effect of TMX is not related to aversive
effects. We evaluated whether the anorectic effect of TMXwas mediated by conditioned taste aversion (Fig. 1E). Asexpected, LiCl-treated rats drank significantly less saccha-rin solution (LiCl-paired flavor), indicating that they devel-oped conditioned taste aversion (26,27,42). Rats treatedsubcutaneously with TMX or intracerebroventricularlywith TMX citrate showed significantly increased saccha-rine preference ratios compared with LiCl-treated rats.The preference ratios for subcutaneous TMX (55.0 � 4.4)and intracerebroventricular TMX citrate (86.1 � 8.5) didnot show significant differences when compared with theirrespective controls (control for the subcutaneous experi-ment: 65.4 � 6.0; control for the intracerebroventricularexperiment: 96.0 � 5.1). Additionally, the preference ratiosfor TMX and TMX citrate were 50%, indicating theabsence of aversive effects (26,42).
TMX-treated rats recovered their normal feeding patternafter treatment was discontinued (Fig. 1F), indicating thatTMX did not induce a permanent state of wasting (42).Furthermore, we did not observe changes in other com-monly used parameters of health status: temperature (Fig.1G), skin aspect, stool consistency, or obvious abnormal
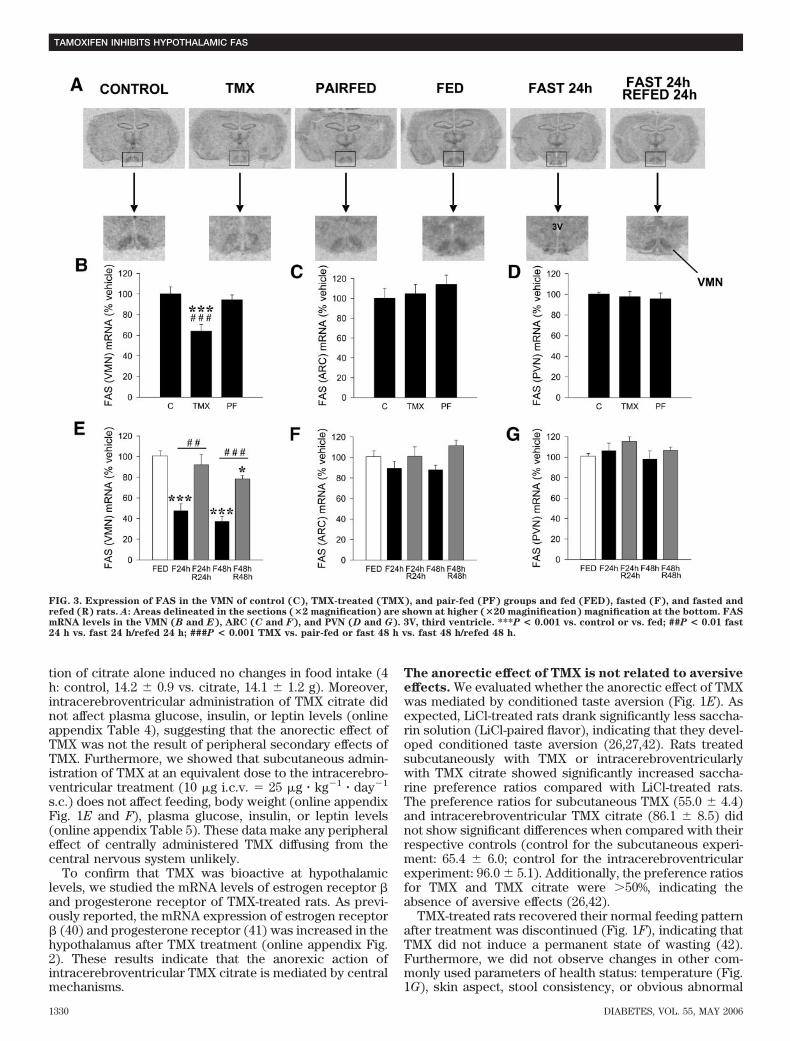
FIG. 3. Expression of FAS in the VMN of control (C), TMX-treated (TMX), and pair-fed (PF) groups and fed (FED), fasted (F), and fasted andrefed (R) rats. A: Areas delineated in the sections (�2 magnification) are shown at higher (�20 maginification) magnification at the bottom. FASmRNA levels in the VMN (B and E), ARC (C and F), and PVN (D and G). 3V, third ventricle. ***P < 0.001 vs. control or vs. fed; ##P < 0.01 fast24 h vs. fast 24 h/refed 24 h; ###P < 0.001 TMX vs. pair-fed or fast 48 h vs. fast 48 h/refed 48 h.
TAMOXIFEN INHIBITS HYPOTHALAMIC FAS
1330 DIABETES, VOL. 55, MAY 2006
behavior, as was evident in the LiCl-treated animals. Thesedata suggest that the powerful anorectic effect of TMX,when administered intracerebroventricularlly or subcuta-neously, was not related to aversive effects, illness, ormalaise.TMX reverses hyperphagia induced by defective lep-tin receptors. We examined whether the anorexigeniceffect of TMX could overcome the hyperphagic stateassociated with a defective leptin receptor signaling.Zucker rats were treated with TMX (0.5 mg � kg�1 � day�1)for 5 days. TMX markedly decreased food intake in obeseZucker rats (which lack a functional long form of leptinreceptor) and their lean littermates (online appendix Fig.3A and B) and prevented body weight gain in both animalmodels (online appendix Fig. 3C and D).TMX prevents the food restriction–induced decreasein proopiomelanocortin and CART expression. Thelevels of CART and proopiomelanocortin (POMC) mRNAsin the ARC were increased in TMX-treated rats whencompared with pair-fed animals (Fig. 2A and B). The
mRNA levels of both neuropeptides were similar to thelevels in untreated fed animals, suggesting that theseanorexigenic signals were inappropriately elevated for thelevel of feeding in TMX-treated animals. Agouti-relatedprotein mRNA was unaffected (Fig. 2C). We detectedincreased levels of neuropeptide Y mRNA in the ARC (Fig.2D) and melanin-concentrating hormone mRNA levels inthe lateral hypothalamic area (LHA) (Fig. 2E) in theTMX-treated animals. These data suggest a compensatoryupregulation of orexigenic neuropeptides against the re-duction in feeding induced by TMX. Of note, these effectsof TMX were specific since no changes were detected inthe mRNA levels of CART in the PVN and in the LHA,corticotrophin-releasing hormone in the PVN, and thyrotro-phin-releasing hormone in the PVN (online appendix Fig. 4).Intracerebroventricular administration of SHU9119attenuates the anorectic effect of subcutaneous TMXtreatment. Our data suggested that TMX could inhibitfeeding through a mechanism involving the POMC neuronsin the ARC. To test this hypothesis, we used SHU9119, a
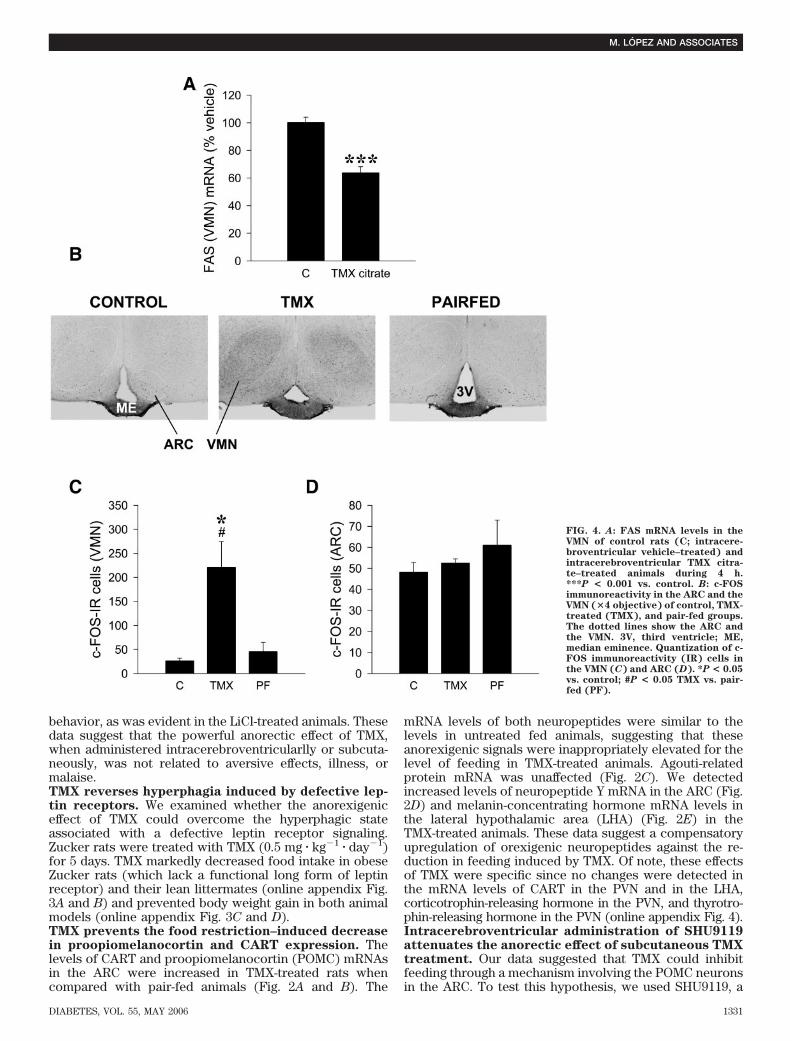
FIG. 4. A: FAS mRNA levels in theVMN of control rats (C; intracere-broventricular vehicle–treated) andintracerebroventricular TMX citra-te–treated animals during 4 h.***P < 0.001 vs. control. B: c-FOSimmunoreactivity in the ARC and theVMN (�4 objective) of control, TMX-treated (TMX), and pair-fed groups.The dotted lines show the ARC andthe VMN. 3V, third ventricle; ME,median eminence. Quantization of c-FOS immunoreactivity (IR) cells inthe VMN (C) and ARC (D). *P < 0.05vs. control; #P < 0.05 TMX vs. pair-fed (PF).
M. LOPEZ AND ASSOCIATES
DIABETES, VOL. 55, MAY 2006 1331
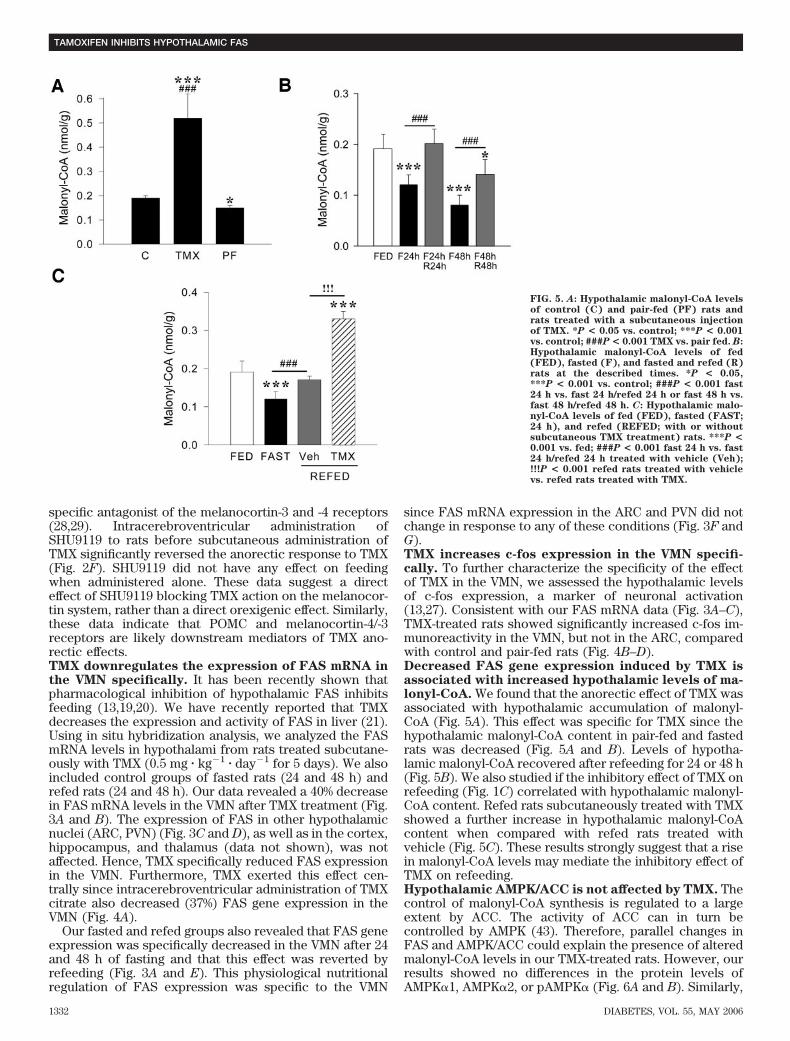
specific antagonist of the melanocortin-3 and -4 receptors(28,29). Intracerebroventricular administration ofSHU9119 to rats before subcutaneous administration ofTMX significantly reversed the anorectic response to TMX(Fig. 2F). SHU9119 did not have any effect on feedingwhen administered alone. These data suggest a directeffect of SHU9119 blocking TMX action on the melanocor-tin system, rather than a direct orexigenic effect. Similarly,these data indicate that POMC and melanocortin-4/-3receptors are likely downstream mediators of TMX ano-rectic effects.TMX downregulates the expression of FAS mRNA inthe VMN specifically. It has been recently shown thatpharmacological inhibition of hypothalamic FAS inhibitsfeeding (13,19,20). We have recently reported that TMXdecreases the expression and activity of FAS in liver (21).Using in situ hybridization analysis, we analyzed the FASmRNA levels in hypothalami from rats treated subcutane-ously with TMX (0.5 mg � kg�1 � day�1 for 5 days). We alsoincluded control groups of fasted rats (24 and 48 h) andrefed rats (24 and 48 h). Our data revealed a 40% decreasein FAS mRNA levels in the VMN after TMX treatment (Fig.3A and B). The expression of FAS in other hypothalamicnuclei (ARC, PVN) (Fig. 3C and D), as well as in the cortex,hippocampus, and thalamus (data not shown), was notaffected. Hence, TMX specifically reduced FAS expressionin the VMN. Furthermore, TMX exerted this effect cen-trally since intracerebroventricular administration of TMXcitrate also decreased (37%) FAS gene expression in theVMN (Fig. 4A).
Our fasted and refed groups also revealed that FAS geneexpression was specifically decreased in the VMN after 24and 48 h of fasting and that this effect was reverted byrefeeding (Fig. 3A and E). This physiological nutritionalregulation of FAS expression was specific to the VMN
since FAS mRNA expression in the ARC and PVN did notchange in response to any of these conditions (Fig. 3F andG).TMX increases c-fos expression in the VMN specifi-cally. To further characterize the specificity of the effectof TMX in the VMN, we assessed the hypothalamic levelsof c-fos expression, a marker of neuronal activation(13,27). Consistent with our FAS mRNA data (Fig. 3A–C),TMX-treated rats showed significantly increased c-fos im-munoreactivity in the VMN, but not in the ARC, comparedwith control and pair-fed rats (Fig. 4B–D).Decreased FAS gene expression induced by TMX isassociated with increased hypothalamic levels of ma-lonyl-CoA. We found that the anorectic effect of TMX wasassociated with hypothalamic accumulation of malonyl-CoA (Fig. 5A). This effect was specific for TMX since thehypothalamic malonyl-CoA content in pair-fed and fastedrats was decreased (Fig. 5A and B). Levels of hypotha-lamic malonyl-CoA recovered after refeeding for 24 or 48 h(Fig. 5B). We also studied if the inhibitory effect of TMX onrefeeding (Fig. 1C) correlated with hypothalamic malonyl-CoA content. Refed rats subcutaneously treated with TMXshowed a further increase in hypothalamic malonyl-CoAcontent when compared with refed rats treated withvehicle (Fig. 5C). These results strongly suggest that a risein malonyl-CoA levels may mediate the inhibitory effect ofTMX on refeeding.Hypothalamic AMPK/ACC is not affected by TMX. Thecontrol of malonyl-CoA synthesis is regulated to a largeextent by ACC. The activity of ACC can in turn becontrolled by AMPK (43). Therefore, parallel changes inFAS and AMPK/ACC could explain the presence of alteredmalonyl-CoA levels in our TMX-treated rats. However, ourresults showed no differences in the protein levels ofAMPK�1, AMPK�2, or pAMPK� (Fig. 6A and B). Similarly,
FIG. 5. A: Hypothalamic malonyl-CoA levelsof control (C) and pair-fed (PF) rats andrats treated with a subcutaneous injectionof TMX. *P < 0.05 vs. control; ***P < 0.001vs. control; ###P < 0.001 TMX vs. pair fed. B:Hypothalamic malonyl-CoA levels of fed(FED), fasted (F), and fasted and refed (R)rats at the described times. *P < 0.05,***P < 0.001 vs. control; ###P < 0.001 fast24 h vs. fast 24 h/refed 24 h or fast 48 h vs.fast 48 h/refed 48 h. C: Hypothalamic malo-nyl-CoA levels of fed (FED), fasted (FAST;24 h), and refed (REFED; with or withoutsubcutaneous TMX treatment) rats. ***P <0.001 vs. fed; ###P < 0.001 fast 24 h vs. fast24 h/refed 24 h treated with vehicle (Veh);!!!P < 0.001 refed rats treated with vehiclevs. refed rats treated with TMX.
TAMOXIFEN INHIBITS HYPOTHALAMIC FAS
1332 DIABETES, VOL. 55, MAY 2006
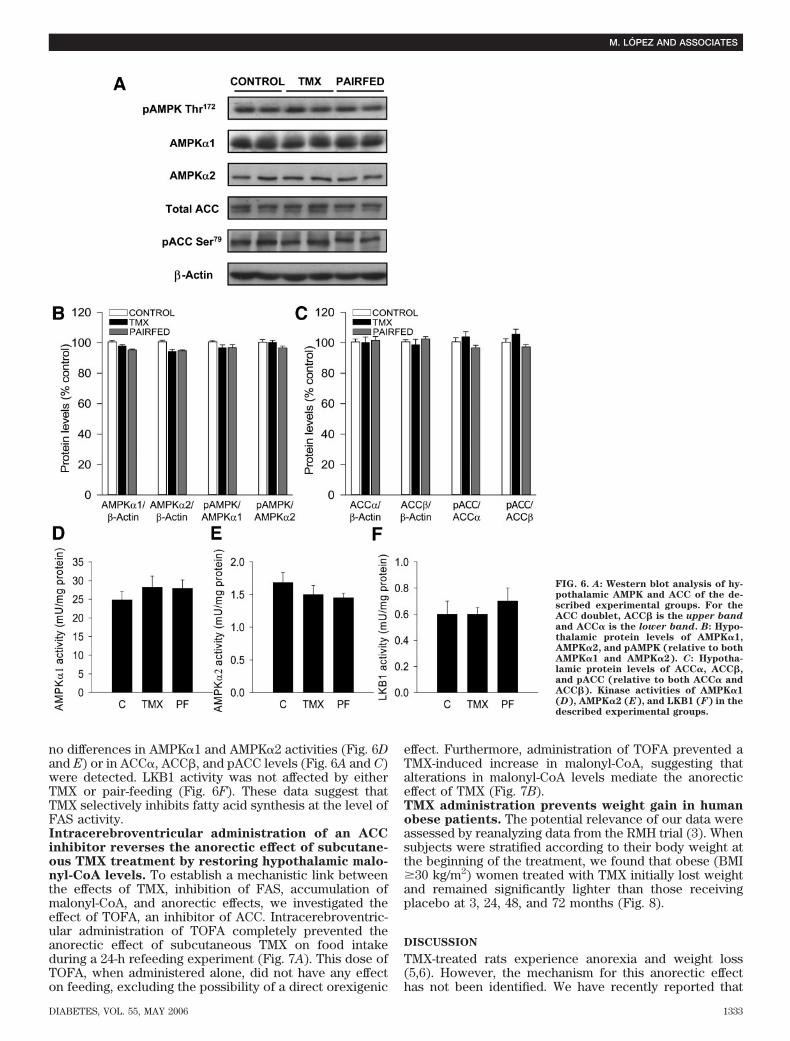
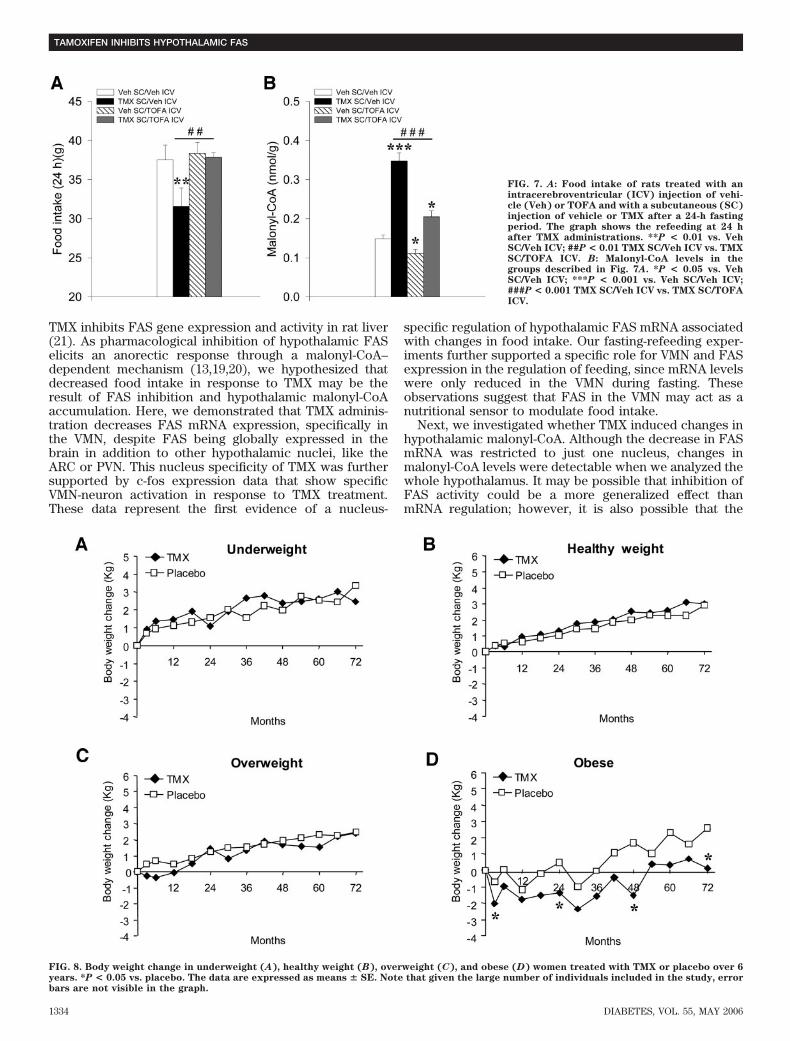
no differences in AMPK�1 and AMPK�2 activities (Fig. 6Dand E) or in ACC�, ACC�, and pACC levels (Fig. 6A and C)were detected. LKB1 activity was not affected by eitherTMX or pair-feeding (Fig. 6F). These data suggest thatTMX selectively inhibits fatty acid synthesis at the level ofFAS activity.Intracerebroventricular administration of an ACCinhibitor reverses the anorectic effect of subcutane-ous TMX treatment by restoring hypothalamic malo-nyl-CoA levels. To establish a mechanistic link betweenthe effects of TMX, inhibition of FAS, accumulation ofmalonyl-CoA, and anorectic effects, we investigated theeffect of TOFA, an inhibitor of ACC. Intracerebroventric-ular administration of TOFA completely prevented theanorectic effect of subcutaneous TMX on food intakeduring a 24-h refeeding experiment (Fig. 7A). This dose ofTOFA, when administered alone, did not have any effecton feeding, excluding the possibility of a direct orexigenic
effect. Furthermore, administration of TOFA prevented aTMX-induced increase in malonyl-CoA, suggesting thatalterations in malonyl-CoA levels mediate the anorecticeffect of TMX (Fig. 7B).TMX administration prevents weight gain in humanobese patients. The potential relevance of our data wereassessed by reanalyzing data from the RMH trial (3). Whensubjects were stratified according to their body weight atthe beginning of the treatment, we found that obese (BMI�30 kg/m2) women treated with TMX initially lost weightand remained significantly lighter than those receivingplacebo at 3, 24, 48, and 72 months (Fig. 8).
DISCUSSION
TMX-treated rats experience anorexia and weight loss(5,6). However, the mechanism for this anorectic effecthas not been identified. We have recently reported that
FIG. 6. A: Western blot analysis of hy-pothalamic AMPK and ACC of the de-scribed experimental groups. For theACC doublet, ACC� is the upper band
and ACC� is the lower band. B: Hypo-thalamic protein levels of AMPK�1,AMPK�2, and pAMPK (relative to bothAMPK�1 and AMPK�2). C: Hypotha-lamic protein levels of ACC�, ACC�,and pACC (relative to both ACC� andACC�). Kinase activities of AMPK�1(D), AMPK�2 (E), and LKB1 (F) in thedescribed experimental groups.
M. LOPEZ AND ASSOCIATES
DIABETES, VOL. 55, MAY 2006 1333
TMX inhibits FAS gene expression and activity in rat liver(21). As pharmacological inhibition of hypothalamic FASelicits an anorectic response through a malonyl-CoA–dependent mechanism (13,19,20), we hypothesized thatdecreased food intake in response to TMX may be theresult of FAS inhibition and hypothalamic malonyl-CoAaccumulation. Here, we demonstrated that TMX adminis-tration decreases FAS mRNA expression, specifically inthe VMN, despite FAS being globally expressed in thebrain in addition to other hypothalamic nuclei, like theARC or PVN. This nucleus specificity of TMX was furthersupported by c-fos expression data that show specificVMN-neuron activation in response to TMX treatment.These data represent the first evidence of a nucleus-
specific regulation of hypothalamic FAS mRNA associatedwith changes in food intake. Our fasting-refeeding exper-iments further supported a specific role for VMN and FASexpression in the regulation of feeding, since mRNA levelswere only reduced in the VMN during fasting. Theseobservations suggest that FAS in the VMN may act as anutritional sensor to modulate food intake.
Next, we investigated whether TMX induced changes inhypothalamic malonyl-CoA. Although the decrease in FASmRNA was restricted to just one nucleus, changes inmalonyl-CoA levels were detectable when we analyzed thewhole hypothalamus. It may be possible that inhibition ofFAS activity could be a more generalized effect thanmRNA regulation; however, it is also possible that the
FIG. 7. A: Food intake of rats treated with anintracerebroventricular (ICV) injection of vehi-cle (Veh) or TOFA and with a subcutaneous (SC)injection of vehicle or TMX after a 24-h fastingperiod. The graph shows the refeeding at 24 hafter TMX administrations. **P < 0.01 vs. VehSC/Veh ICV; ##P < 0.01 TMX SC/Veh ICV vs. TMXSC/TOFA ICV. B: Malonyl-CoA levels in thegroups described in Fig. 7A. *P < 0.05 vs. VehSC/Veh ICV; ***P < 0.001 vs. Veh SC/Veh ICV;###P < 0.001 TMX SC/Veh ICV vs. TMX SC/TOFAICV.
FIG. 8. Body weight change in underweight (A), healthy weight (B), overweight (C), and obese (D) women treated with TMX or placebo over 6years. *P < 0.05 vs. placebo. The data are expressed as means � SE. Note that given the large number of individuals included in the study, errorbars are not visible in the graph.
TAMOXIFEN INHIBITS HYPOTHALAMIC FAS
1334 DIABETES, VOL. 55, MAY 2006

changes in malonyl-CoA levels in the VMN are significantenough to be detectable when the whole hypothalamus isassayed.
If FAS expression within the VMN has a role in regulat-ing food intake, then the fact that FAS mRNA levelsdecrease in response to both fasting and TMX treatment isapparently contradictory. The explanation for this appar-ent paradox is found by examining malonyl-CoA levels. Wedemonstrate that the anorectic effect induced by TMX issimilar to that of C75 in that it requires the accumulationof hypothalamic malonyl-CoA (14). The anorectic effect ofTMX was reverted by pharmacological inhibition of ACCthat prevented accumulation of malonyl-CoA. Thus, spe-cific inhibition of FAS activity by TMX in the presence ofmaintained ACC activity (see below) facilitates accumula-tion of malonyl-CoA. Conversely, during fasting the de-crease in FAS mRNA expression is associated withdecreased levels of malonyl-CoA, suggesting that earlysteps of the hypothalamic lipogenic pathway are physio-logically inhibited during fasting. Supporting this argu-ment are recent reports demonstrating that during fasting,activation of hypothalamic AMPK phosphorylates andinhibits ACC (44,45).
We also investigated the effects of TMX on other com-ponents of the fatty acid biosynthetic pathway. AMPKplays a critical role in the hypothalamic regulation offeeding through its effects on the lipogenic pathway andneuropeptides (44,45). In situations of energy demandsuch as fasting, AMPK is phosphorylated by LKB1 at Thr172
(and then activated), resulting in phosphorylation andsubsequent inhibition of ACC, thereby inactivating thefatty acid biosynthetic pathway (43). Under conditions ofenergy surplus, ACC is not phosphorylated, facilitatingproduction of malonyl-CoA (43,45). Our data showed thatFAS inhibition induced by TMX is not associated withchanges in either AMPK protein levels/activity or phos-phorylated ACC levels. These data suggest that the hypo-thalamic AMPK/ACC axis is not affected by TMX. Hence,the accumulation of malonyl-CoA induced by TMX is theresult of specific FAS inhibition in the presence of normalACC activity (online appendix Fig. 5).
Food restriction is associated with reduction in themRNA levels of CART and POMC in the ARC (7–9,12). Thisexpected reduction in CART and POMC levels wasblocked by TMX. Furthermore, pharmacological inhibitionof melanocortin-3/-4 receptors by SHU9119 prevented theTMX anorectic effects, indicating that inhibition of feedingby TMX involves the melanocortin system. CART/POMCneurons receive neural projections and excitatory inputsfrom neurons in the VMN (46,47); therefore, changes inFAS/malonyl-CoA in the VMN might be linked to CART/POMC changes in the ARC. However, this mechanism stillneeds to be clarified.
TMX is a drug widely used for the treatment of estrogenreceptor–positive breast cancers (3,4). Despite its wide-spread use, the action of TMX on human body weight iscontroversial. Data from the WHEL (Women’s HealthyEating and Living) clinical trial show that obese womenwith breast cancer under TMX treatment had a decreasedBMI (48). However, measurement of BMI in this studyoccurred at varying times after breast cancer diagnosisand thus did not necessarily reflect prediagnosis BMI. Theresults of this study may also have been confounded by theconcurrent diagnosis of cancer. Hence, we reanalyzed thebody weight data from the RMH trial (3). Since thesesubjects were free of cancer during the study, changes in
body weight are more likely to be a direct effect of TMX.We found that obese (BMI �30 kg/m2) women treated withTMX were significantly lighter than those receiving pla-cebo. Although this is a post hoc analysis of the data, wesuggest that the large size of the cohort studied stronglysupports the finding that low-dose TMX can induce asustainable body weight decrease in obese humans. Al-though the RMH trial was not an obesity prevention/treatment trial, the finding that the obese women lostweight initially for 4 years and on average had maintainedtheir initial weight by the end of the study is a highlysignificant deviation from the expected (49,50).
In conclusion, this study demonstrates that the anorec-tic actions of TMX involve a specific decrease in FASmRNA levels in the VMN and hypothalamic malonyl-CoAaccumulation. Specific regulation of FAS in the VMN byTMX or fasting suggests that fatty acid metabolism in thisnucleus may play a physiological role in feeding control.Altogether, our data indicate that altering FAS in the VMNmay be a suitable strategy for altering food intake andbody weight homeostasis in experimental animals andobese subjects.
ACKNOWLEDGMENTS
This work has been supported by grants from the MedicalResearch Council (to A.J.V.-P.), the Welcome Trust (toA.J.V.-P.), the Spanish Ministry of Education (to C.D.), theXunta de Galicia (to C.D.), and the European Union(LSHM-CT-2003-503041) (to C.D.). A.K.S. is funded by theU.S. Public Health Service (DK 19514) and a grant from theJuvenile Diabetes Research Foundation. M.L. is funded bythe Marie Curie Program (QLK6-CT-2002-51671).
We are grateful to Keith Burling, Mark Campbell, andJanice Carter for their technical assistance, to Dr. AnthonyP. Coll (University of Cambridge, Cambridge, U.K.), to Dr.Benjamin Challis (University of Cambridge, Cambridge,U.K.), to Prof. Manuel Ros (University Rey Juan Carlos,Madrid, Spain), and to Prof. Manuel Tena-Sempere (Uni-versity of Cordoba, Cordoba, Spain) for their useful com-ments and criticism. We thank Prof. Len Storlien(AstraZeneca, Mondal, Sweden) for supplying us TOFAand Prof. Dario Alessi (University of Dundee, Dundee,U.K.) for the LKB1 antibody.
REFERENCES
1. Wirshing DA: Schizophrenia and obesity: impact of antipsychotic medica-tions. J Clin Psychiatry 65 (Suppl. 18):13–26, 2004
2. Korner J, Aronne LJ: Pharmacological approaches to weight reduction:therapeutic targets. J Clin Endocrinol Metab 89:2616–2621, 2004
3. Powles T, Eeles R, Ashley S, Easton D, Chang J, Dowsett M, Tidy A, ViggersJ, Davey J: Interim analysis of the incidence of breast cancer in the RoyalMarsden Hospital tamoxifen randomised chemoprevention trial. Lancet
352:98–101, 19984. Jordan VC: Selective estrogen receptor modulation: concept and conse-
quences in cancer. Cancer Cell 5:207–213, 20045. Wade GN, Heller HW: Tamoxifen mimics the effects of estradiol on food
intake, body weight, and body composition in rats. Am J Physiol 264:R1219–R1223, 1993
6. Gray JM, Schrock S, Bishop M: Estrogens and antiestrogens: actions andinteractions with fluphenazine on food intake and body weight in rats.Am J Physiol 264:R1214–R1218, 1993
7. Meister B: Control of food intake via leptin receptors in the hypothalamus.Vitam Horm 59:265–304, 2000
8. Schwartz MW, Woods SC, Porte D Jr, Seeley RJ, Baskin DG: Centralnervous system control of food intake. Nature 404:661–671, 2000
9. Saper CB, Chou TC, Elmquist JK: The need to feed: homeostatic andhedonic control of eating. Neuron 36:199–211, 2002
10. Obici S, Feng Z, Morgan K, Stein D, Karkanias G, Rossetti L: Central
M. LOPEZ AND ASSOCIATES
DIABETES, VOL. 55, MAY 2006 1335

administration of oleic acid inhibits glucose production and food intake.Diabetes 51:271–275, 2002
11. Cota D, Marsicano G, Lutz B, Vicennati V, Stalla GK, Pasquali R, Pagotto U:Endogenous cannabinoid system as a modulator of food intake. Int J Obes
Relat Metab Disord 27:289–301, 200312. Flier JS: Obesity wars: molecular progress confronts an expanding epi-
demic. Cell 116:337–350, 200413. Gao S, Lane MD: Effect of the anorectic fatty acid synthase inhibitor C75
on neuronal activity in the hypothalamus and brainstem. Proc Natl Acad
Sci U S A 100:5628–5633, 200314. Hu Z, Cha SH, Chohnan S, Lane MD: Hypothalamic malonyl-CoA as a
mediator of feeding behavior. Proc Natl Acad Sci U S A 100:12624–12629,2003
15. Ruderman NB, Saha AK, Kraegen EW: Minireview: malonyl CoA, AMP-activated protein kinase, and adiposity. Endocrinology 144:5166–5171,2003
16. Lam TK, Schwartz GJ, Rossetti L: Hypothalamic sensing of fatty acids. Nat
Neurosci 8:579–584, 200517. Kim EK, Miller I, Landree LE, Borisy-Rudin FF, Brown P, Tihan T,
Townsend CA, Witters LA, Moran TH, Kuhajda FP, Ronnett GV: Expressionof FAS within hypothalamic neurons: a model for decreased food intakeafter C75 treatment. Am J Physiol Endocrinol Metab 283:E867–E879, 2002
18. Sorensen A, Travers MT, Vernon RG, Price NT, Barber MC: Localization ofmessenger RNAs encoding enzymes associated with malonyl-CoA metab-olism in mouse brain. Brain Res Gene Expr Patterns 1:167–173, 2002
19. Loftus TM, Jaworsky DE, Frehywot GL, Townsend CA, Ronnett GV, LaneMD, Kuhajda FP: Reduced food intake and body weight in mice treatedwith fatty acid synthase inhibitors. Science 288:2379–2381, 2000
20. Clegg DJ, Wortman MD, Benoit SC, McOsker CC, Seeley RJ: Comparison ofcentral and peripheral administration of C75 on food intake, body weight,and conditioned taste aversion. Diabetes 51:3196–3201, 2002
21. Lelliott CJ, Lopez M, Curtis RK, Parker N, Laudes M, Yeo G, Jimenez-LinanM, Grosse J, Saha AK, Wiggins D, Hauton D, Brand MD, O’Rahilly S, GriffinJL, Gibbons GF, Vidal-Puig A: Transcript and metabolite analysis of theeffects of tamoxifen in rat liver reveals inhibition of fatty acid synthesis inthe presence of hepatic steatosis. FASEB J 19:1108–1119, 2005
22. Mystkowski P, Seeley RJ, Hahn TM, Baskin DG, Havel PJ, Matsumoto AM,Wilkinson CW, Peacock-Kinzig K, Blake KA, Schwartz MW: Hypothalamicmelanin-concentrating hormone and estrogen-induced weight loss. J Neu-
rosci 20:8637–8642, 200023. Tritos NA, Segal-Lieberman G, Vezeridis PS, Maratos-Flier E: Estradiol-
induced anorexia is independent of leptin and melanin-concentratinghormone. Obes Res 12:716–724, 2004
24. Lopez M, Seoane LM, Tovar S, Nogueiras R, Dieguez C, Senarıs R: Orexin-Aregulates growth hormone releasing hormone mRNA content in a nucleusspecific manner and somatostatin mRNA content in a growth hormone-dependent fashion in the rat hypothalamus. Eur J Neurosci 19:2080–2088,2004
25. Liang YQ, Akishita M, Kim S, Ako J, Hashimoto M, Iijima K, Ohike Y,Watanabe T, Sudoh N, Toba K, Yoshizumi M, Ouchi Y: Estrogen receptorbeta is involved in the anorectic action of estrogen. Int J Obes Relat Metab
Disord 26:1103–1109, 200226. Kelly JF, Elias CF, Lee CE, Ahima RS, Seeley RJ, Bjorbaek C, Oka T, Saper
CB, Flier JS, Elmquist JK: Ciliary neurotrophic factor and leptin inducedistinct patterns of immediate early gene expression in the brain. Diabetes
53:911–920, 200427. Halatchev IG, Cone RD: Peripheral administration of PYY3-36 produces
conditioned taste aversion in mice. Cell Metab 1:159–168, 200528. Kask A, Mutulis F, Muceniece R, Pahkla R, Mutule I, Wikberg JE, Rago L,
Schioth HB: Discovery of a novel superpotent and selective melanocortin-4receptor antagonist (HS024): evaluation in vitro and in vivo. Endocrinol-
ogy 139:5006–5014, 199829. Heisler LK, Cowley MA, Tecott LH, Fan W, Low MJ, Smart JL, Rubinstein
M, Tatro JB, Marcus JN, Holstege H, Lee CE, Cone RD, Elmquist JK:Activation of central melanocortin pathways by fenfluramine. Science
297:609–611, 200230. Medina-Gomez G, Virtue S, Lelliott C, Boiani R, Campbell M, Christodou-
lides C, Perrin C, Jimenez-Linan M, Blount M, Dixon J, Zahn D, ThresherRR, Aparicio S, Carlton M, Colledge WH, Kettunen MI, Seppanen-Laakso T,Sethi JK, O’Rahilly S, Brindle K, Cinti S, Oresic M, Burcelin R, Vidal-Puig A:The link between nutritional status and insulin sensitivity is dependent on
the adipocyte-specific peroxisome proliferator-activated receptor-�2 iso-form. Diabetes 54:1706–1716, 2005
31. Coll AP, Challis BG, Lopez M, Piper S, Yeo GS, O’Rahilly S: Proopiomela-nocortin-deficient mice are hypersensitive to the adverse metabolic effectsof glucocorticoids. Diabetes 54:2269–2276, 2005
32. Lopez M, Seoane LM, Tovar S, Garcıa MC, Nogueiras R, Dieguez C, SeñarısRM: A possible role of neuropeptide Y, agouti-related protein and leptinreceptor isoforms in hypothalamic programming by perinatal feeding inthe rat. Diabetologia 48:140–148, 2005
33. Nogueiras R, Gallego R, Gualillo O, Caminos JE, Garcıa-Caballero T,Casanueva FF, Dieguez C: Resistin is expressed in different rat tissues andis regulated in a tissue- and gender-specific manner. FEBS Lett 548:21–27,2003
34. Sakamoto K, Goransson O, Hardie DG, Alessi DR: Activity of LKB1 andAMPK-related kinases in skeletal muscle: effects of contraction, phen-formin, and AICAR. Am J Physiol Endocrinol Metab 287:E310–E317, 2004
35. Dale S, Wilson WA, Edelman AM, Hardie DG: Similar substrate recognitionmotifs for mammalian AMP-activated protein kinase, higher plant HMG-CoA reductase kinase-A, yeast SNF1, and mammalian calmodulin-depen-dent protein kinase I. FEBS Lett 361:191–195, 1995
36. Lizcano JM, Goransson O, Toth R, Deak M, Morrice NA, Boudeau J,Hawley SA, Udd L, Makela TP, Hardie DG, Alessi DR: LKB1 is a masterkinase that activates 13 kinases of the AMPK subfamily, including MARK/PAR-1. EMBO J 23:833–843, 2004
37. McGarry JD, Stark MJ, Foster DW: Hepatic malonyl-CoA levels of fed,fasted and diabetic rats as measured using a simple radioisotopic assay.J Biol Chem 253:8291–8293, 1978
38. Saha AK, Kurowski TG, Ruderman NB: A malonyl-CoA fuel-sensingmechanism in muscle: effects of insulin, glucose, and denervation. Am J
Physiol 269:E283–E289, 199539. Gold E, Stapley S, Goulding A: Tamoxifen and norethisterone: effects on
plasma cholesterol and total body calcium content in the estrogen-deficient rat. Horm Metab Res 26:100–103, 1994
40. Patisaul HB, Aultman EA, Bielsky IF, Young LJ, Wilson ME: Immediate andresidual effects of tamoxifen and ethynylestradiol in the female rathypothalamus. Brain Res 978:185–193, 2003
41. Tena-Sempere M, Navarro VM, Mayen A, Bellido C, Sanchez-Criado JE:Regulation of estrogen receptor (ER) isoform messenger RNA expressionby different ER ligands in female rat pituitary. Biol Reprod 70:671–678,2004
42. Kim MS, Park JY, Namkoong C, Jang PG, Ryu JW, Song HS, Yun JY,Namgoong IS, Ha J, Park IS, Lee IK, Viollet B, Youn JH, Lee HK, Lee KU:Anti-obesity effects of alpha-lipoic acid mediated by suppression ofhypothalamic AMP-activated protein kinase. Nat Med 10:727–733, 2004
43. Kahn BB, Alquier T, Carling D, Hardie DG: AMP-activated protein kinase:ancient energy gauge provides clues to modern understanding of metab-olism. Cell Metab 1:15–25, 2005
44. Minokoshi Y, Alquier T, Furukawa N, Kim YB, Lee A, Xue B, Mu J, FoufelleF, Ferre P, Birnbaum MJ, Stuck BJ, Kahn BB: AMP-kinase regulates foodintake by responding to hormonal and nutrient signals in the hypothala-mus. Nature 428:569–574, 2004
45. Andersson U, Filipsson K, Abbott CR, Woods A, Smith K, Bloom SR,Carling D, Small CJ: AMP-activated protein kinase plays a role in thecontrol of food intake. J Biol Chem 279:12005–12008, 2004
46. Pinto S, Roseberry AG, Liu H, Diano S, Shanabrough M, Cai X, FriedmanJM, Horvath TL: Rapid rewiring of ARC nucleus feeding circuits by leptin.Science 304:110–115, 2004
47. Sternson SM, Shepherd GM, Friedman JM: Topographic mapping of VMH3 ARC nucleus microcircuits and their reorganization by fasting. Nat
Neurosci 8:1356–1363, 200548. Wasserman L, Flatt SW, Natarajan L, Laughlin G, Matusalem M, Faerber S,
Rock CL, Barrett-Connor E, Pierce JP: Correlates of obesity in postmeno-pausal women with breast cancer: comparison of genetic, demographic,disease-related, life history and dietary factors. Int J Obes Relat Metab
Disord 28:49–56, 200449. Finer N: Pharmacotherapy of obesity. Best Pract Res Clin Endocrinol
Metab 16:717–742, 200250. Li Z, Maglione M, Tu W, Mojica W, Arterburn D, Shugarman LR, Hilton L,
Suttorp M, Solomon V, Shekelle PG, Morton SC: Meta-analysis: pharmaco-logic treatment of obesity. Ann Intern Med 142:532–546, 2005
TAMOXIFEN INHIBITS HYPOTHALAMIC FAS
1336 DIABETES, VOL. 55, MAY 2006
Related Documents











