Mechanisms of Spontaneous Activity in the Developing Spinal Cord and Their Relevance to Locomotion M. J. O’DONOVAN, a,c P. WENNER, a N. CHUB, a J. TABAK, a AND J. RINZEL b a Laboratory of Neural Control, National Institute of Neurological Disorders and Stroke, National Institutes of Health, Bethesda, Maryland 20892, USA. b Center for Neural Science and Courant Institute of Mathematical Sciences,New York University, New York, New York 10003, USA. ABSTRACT: The isolated lumbosacral cord of the chick embryo generates spontaneous episodes of rhythmic activity. Muscle nerve recordings show that the discharge of sar- torius (flexor) and femorotibialis (extensor) motoneurons alternates even though the motoneurons are depolarized simultaneously during each cycle. The alternation occurs because sartorius motoneuron firing is shunted or voltage-clamped by its synaptic drive at the time of peak femorotibialis discharge. Ablation experiments have identified a region dorsomedial to the lateral motor column that may be required for the alternation of sartorius and femorotibialis motoneurons. This region overlaps the location of interneurons activated by ventral root stimulation. Whole- cell recordings from interneurons receiving short latency ventral root input indicate that they fire at an appropriate time to contribute to the cyclical pause in firing of sar- torius motoneurons. Spontaneous activity was modeled by the interaction of three variables: network activity and two activity-dependent forms of network depression. A “slow” depression which regulates the occurrence of episodes and a “fast” depres- sion that controls cycling during an episode. The model successfully predicts several aspects of spinal network behavior including spontaneous rhythmic activity and the recovery of network activity following blockade of excitatory synaptic transmission. N etworks in the isolated spinal cord of the chick embryo are spontaneously active. The activity comprises recurring episodes in which motoneurons and interneurons are dri- ven into rhythmic bursting. During such bursts flexor and extensor motoneurons can alter- nate their discharge suggesting that spontaneous embryonic activity engages some of the same circuitry used later in development for locomotion. 1,2 During the past few years sig- nificant progress has been made in identifying the mechanisms responsible for the timing of motoneuron activity and the occurrence of spontaneous episodes in the chick spinal cord. This chapter will review our current understanding of these processes. PATTERNING OF DISCHARGE IN FLEXOR AND EXTENSOR MOTONEURONS DURING EPISODES OF RHYTHMIC ACTIVITY Our findings about the alternation of flexor and extensor motoneurons have been sur- prising because the mechanism appears to be quite different from that generally assumed 130 c Address correspondence to: Michael J. O’Donovan, Laboratory of Neural Control, Bldg. 49, Room 3A50, NINDS, NIH, Bethesda, MD 20892. E-mail: [email protected]

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Mechanisms of Spontaneous Activity inthe Developing Spinal Cord and TheirRelevance to Locomotion
M. J. O’DONOVAN,a,c P. WENNER,a N. CHUB,a J. TABAK,a AND
J. RINZELb
aLaboratory of Neural Control, National Institute of NeurologicalDisorders and Stroke, National Institutes of Health, Bethesda, Maryland20892, USA.bCenter for Neural Science and Courant Institute of MathematicalSciences, New York University, New York, New York 10003, USA.
ABSTRACT: The isolated lumbosacral cord of the chick embryo generates spontaneousepisodes of rhythmic activity. Muscle nerve recordings show that the discharge of sar-torius (flexor) and femorotibialis (extensor) motoneurons alternates even though themotoneurons are depolarized simultaneously during each cycle. The alternationoccurs because sartorius motoneuron firing is shunted or voltage-clamped by itssynaptic drive at the time of peak femorotibialis discharge. Ablation experimentshave identified a region dorsomedial to the lateral motor column that may berequired for the alternation of sartorius and femorotibialis motoneurons. This regionoverlaps the location of interneurons activated by ventral root stimulation. Whole-cell recordings from interneurons receiving short latency ventral root input indicatethat they fire at an appropriate time to contribute to the cyclical pause in firing of sar-torius motoneurons. Spontaneous activity was modeled by the interaction of threevariables: network activity and two activity-dependent forms of network depression.A “slow” depression which regulates the occurrence of episodes and a “fast” depres-sion that controls cycling during an episode. The model successfully predicts severalaspects of spinal network behavior including spontaneous rhythmic activity and therecovery of network activity following blockade of excitatory synaptic transmission.
Networks in the isolated spinal cord of the chick embryo are spontaneously active. Theactivity comprises recurring episodes in which motoneurons and interneurons are dri-
ven into rhythmic bursting. During such bursts flexor and extensor motoneurons can alter-nate their discharge suggesting that spontaneous embryonic activity engages some of thesame circuitry used later in development for locomotion.1,2 During the past few years sig-nificant progress has been made in identifying the mechanisms responsible for the timingof motoneuron activity and the occurrence of spontaneous episodes in the chick spinalcord. This chapter will review our current understanding of these processes.
PATTERNING OF DISCHARGE IN FLEXOR AND EXTENSORMOTONEURONS DURING EPISODES OF RHYTHMIC ACTIVITY
Our findings about the alternation of flexor and extensor motoneurons have been sur-prising because the mechanism appears to be quite different from that generally assumed
130
cAddress correspondence to: Michael J. O’Donovan, Laboratory of Neural Control, Bldg. 49,Room 3A50, NINDS, NIH, Bethesda, MD 20892. E-mail: [email protected]

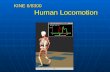
to operate during locomotion of adult animals. In adult animals, alternation of antagonistmotoneurons is believed to be produced by an alternating hyperpolarizing and depolariz-ing synaptic drive to flexor and extensor motoneurons.3,4 In the chick spinal cord rhythmi-cally active motoneurons are not subject to an alternating depolarizing and hyperpolarizingsynaptic drive even though they discharge out of phase (FIG.1A).
Perhaps our most important insight into the timing of motoneuron activity in the chickcord has been the realization that each episode is produced by the repetition of largelyequivalent cycles. Intracellular recordings have shown that flexor and extensor motoneu-rons are depolarized synchronously during each cycle.2,5 Knee extensor, femorotibialismotoneurons discharge at the peak of their depolarization whereas the hip flexor, sartoriusmotoneurons stop firing when they are maximally depolarized and then resume as thedepolarization decays. In sartorius motoneurons, this leads to a double burst in each cycleof activity—a brief burst of impulses on the rising part of the depolarization, followed bymore prolonged discharge on the falling phase.2,5 Whole-cell recordings and pharmacolog-ical experiments have revealed that the depolarization in each class of motoneuron is com-plex comprising NMDA, AMPA/kainate, cholinergic, GABAergic, and glycinergiccomponents.5,6
FIGURE 1A illustrates two cycles of sartorius and femorotibialis motoneuron activityrecorded from the cut muscle nerves of an isolated spinal cord superfused with normalTyrode’s solution. It can be seen that the peak femorotibialis discharge is coincident with thepause in firing of the flexor sartorius. A similar alternating pattern of discharge can berecorded when spontaneous activity recovers in the presence of a mixture of various excita-tory transmitter antagonists (NMDA, non-NMDA, and nicotinic cholinergic) indicating thatthis pattern can be produced by circuits of GABAergic and glycinergic neurons.7 The abilityof GABA/glycinergic networks to sustain rhythmic activity presumably occurs becauseGABA and glycine produce depolarizing synaptic potentials during development.6,8-10
In sartorius motoneurons, under control conditions, the pause is largely determined bythe GABAergic component of their rhythmic synaptic drive because it is abolished byejection of the GABAA antagonist bicuculline over (or into) the sartorius pool of motoneu-rons;6 (cf. FIG.1A and B). Application of bicuculline within the femorotibialis motoneuronpool has little effect on the extensor’s discharge pattern. GABAergic drive is present onfemorotibialis motoneurons, but it does not block action potential generation as it does insartorius motoneurons.
The sartorius pause can also be influenced by ejection of several transmitter antagonistsonto sartorius motoneurons although, with the exception of curare which may act onGABA
A receptors, none is as effective as the GABAA antagonists bicuculline or picrotoxin.6
However, the pause persists, and actually lengthens, in the presence of bath-applied bicu-culline, indicating that GABAergic drive is not required for the expression of the pause indisinhibited spinal networks. In addition, a pause is induced in the extensor femorotibialis(FIG.1C). In the disinhibited cord the sartorius pause can be abolished by local applicationof excitatory amino acid antagonists into the sartorius motoneuron pool, indicating that it ismediated largely by a glutamatergic synaptic drive (FIG.1D, see also ref. 6).
These observations have led us to propose that the pause is determined by a combina-tion of the impedance change and the voltage clamping effect of synaptic conductances thatact near the soma.2,6 We hypothesize that when bicuculline and strychnine are bath-appliedto spinal networks they produce disinhibition, thereby increasing the excitatory synapticdrive and the associated conductance change in both sartorius and femorotibialis motoneu-rons. This type of experiment illustrates an important aspect of GABA and glycinergicfunction in developing networks: GABA and glycine can be both excitatory and inhibitory.The conditions under which the transmitters express either inhibitory or excitatory actionare not understood. One possibility is that their action depends upon where their synapticinputs are located. Somatically located synapses may be more effective than dendritic ones
O’DONOVAN et al.: DEVELOPMENT OF MOTOR ACTIVITY 131
at producing inhibition by a conductance shunt. By contrast, dendritic inputs may deliverdepolarizations to the soma without a substantial somal conductance shunt and should,therefore, be excitatory.
132 ANNALS NEW YORK ACADEMY OF SCIENCES
FIGURE 1. Temporal relationship between discharge recorded from the sartorius (sart, gray) and thefemorotibialis (fem, black) muscle nerves under several conditions. (A) Control recordings in normalTyrode’s solution showing two cycles of activity. The boxes in A delineate the pause in firing of thesartorius nerve. (B) Local application of the GABAA antagonist bicuculline over LS1 abolishes thepause in sartorius discharge. The boxes are the same width as in A and positioned at approximatelythe same part of the cycle as in A (recordings from the same cord are shown in A and B). (C)Recordings of three cycles of activity from the muscle nerves when the cord is bathed in theinhibitory amino acid antagonists bicuculline and strychnine. The drugs induce a pause in each cycleof femorotibialis activity coincident with the pause in sartorius. The boxes delineate the duration ofthe sartorius pause. (D) When the cord is in bicuculline and strychnine, local application of the exci-tatory amino antagonist AP5 within LS1 abolishes the pause in sartorius discharge (the same cord isshown in C and D). Under these conditions the firing behavior of the nerves is reversed when com-pared to the control conditions. The boxes in D are the same width as in C and positioned at approx-imately the same part of the cycle. (Data modified from Sernagor et al.6) The schematic at the top ofthe figure shows a diagram of the lumbosacral cord (segments T7–LS3) illustrating the position ofthe drug ejection electrode and the sartorius (gray) and femorotibialis (black) motoneurons pools.

INTERNEURONS INVOLVED IN FLEXOR AND EXTENSOR ALTERNATION
Under conditions of normal perfusion, in the absence of drugs, the pause of sartoriusdischarge appears to be determined primarily by a depolarizing GABAergic input. At pre-sent the identity and location of the GABAergic interneurons responsible for this inputare unknown. Ablation experiments have identified a region dorsomedial to the lateralmotor column that seems to be required for the sartorius pause and the alternation of flex-ors and extensors.11 Lesions that encroach upon this area result in abolition of the sarto-rius pause and in the synchronization of sartorius and femorotibialis discharge (FIG. 2A,middle panel). It is important to emphasize, however, that the severe lesions necessary toabolish the sartorius pause are likely to also be associated with a significant decrease inthe synaptic drive and the accompanying conductance change experienced by motoneu-rons. It could be this reduction that is responsible for the abolition of the pause in sarto-rius discharge.
However, we have also found that this dorsomedial region encompasses the locationof a population of GABAergic interneurons activated by stimulation of the ventral roots(FIG. 2B). This was established using an optical method to identify the interneurons acti-vated by ventral root stimulation (see ref. 12). For this purpose we backlabeled interneu-rons that projected into the lateral motor column and the ventrolateral funiculus withcalcium-sensitive dyes. We then monitored the fluorescence changes in the transverseface of the cord following stimulation of the ventral roots (FIG. 2B, control). The opticalsignals from these interneurons were abolished in the presence of the nicotinic, choliner-gic antagonist mecamylamine, indicating that they depend upon the activity of motoneu-ron recurrent collaterals (FIG. 2B, mecamylamine). This and other evidence havesuggested that these interneurons may be the avian equivalent of the mammalianRenshaw cell.13 The finding that their location overlaps with a region that appears neces-sary for the alternation of flexor and extensor discharge raises the possibility that thesecells may contribute some of the GABAergic synaptic drive to sartorius motoneurons. Toaddress this issue further, we recorded intracellularly from individual interneurons thatreceived short latency synaptic input from the ventral roots. In some of the cells we wereable to show that they projected directly to motoneurons using spike-triggered averagingof ventral root potentials (see FIG. 1 and ref. 12). Such potentials were prolonged, lasting50–100 ms and were blocked by bicuculline suggesting that the interneurons wereGABAergic. In FIGURE 2C we illustrate the firing of a putative Renshaw cell and com-pare it to the rhythmic slow potential recorded from the ventral roots. This particularinterneuron fired on the rising phase of its rhythmic depolarization just before the peakof the ventral root potential—a time which would allow it to contribute a GABAergicpotential to the sartorius pause. At present, we have no evidence that these interneuronsproject to sartorius motoneurons. In future experiments it will be of particular interest toestablish whether they do, and if so, whether the projection is stronger than to the exten-sor femorotibialis motoneurons.
The existence of a population of GABAergic interneurons in the ventral part of the corddiffers from the situation in adult animals where GABA-immunoreactive interneurons arerestricted to the dorsal horn. However, in both the chick embryo14 and the rat fetus,15 a tran-sient population of GABA-immunoreactive interneurons can be detected in the ventral partof the cord that disappears later in development. The fate of these neurons is unknown butthey may switch their transmitter from GABA to glycine because the number of glycine-immunoreactive neurons in the ventral horn increases in parallel with the decline of theGABA-immunoreactive population.16 A switch from GABA to glycine neurotransmissionhas been documented to occur in the developing auditory system of the gerbil17 and wouldbe consistent with the predominantly glycinergic inhibition of the adult spinal cord.
O’DONOVAN et al.: DEVELOPMENT OF MOTOR ACTIVITY 133

134 ANNALS NEW YORK ACADEMY OF SCIENCES
MECHANISMS OF NETWORK ACTIVITY
In two previous reviews we discussed the mechanisms we believe operate to producespontaneous episodes in the spinal cord.18,19 In this section we briefly discuss these ideas anddescribe a mathematical model that accounts for several features of the spontaneous activ-ity generated by the isolated spinal cord. We have proposed that the general mechanism ofspontaneous activity depends on the existence of recurrent excitatory connections within
FIGURE 2. (A) The left panel shows a diagram of the transverse face of the cord showing a regionidentified by lesion experiments that appears to be necessary for the pause in sartorius discharge. Themiddle panels show recordings from the sartorius (sart) and femorotibialis (fem) muscle nerves fol-lowing a removal of the dorsal and intermediate part of the spinal cord over 4–5 segments fromT7–LS4, leaving only a ventral strip which is shown in the diagram to the right of the traces(Modified from Ho and O’Donovan.11) (B) Optical identification of interneurons activated by stimu-lation the ventral roots. The videomicrographs show difference images (activity–control) of the trans-verse face of the spinal cord at LS4 during application of a brief train of stimuli to the ipsilateralventral root as shown in the diagram of the cord to the right of the micrographs. The effects of ven-tral root stimulation are shown before (control) and after (mecamylamine) application of the nico-tinic, cholinergic antagonist mecamylamine (50 µM). (C) Illustration of the firing behavior of aninterneuron that received direct motoneuron input and projected to motoneurons as demonstrated byspike-triggered averaging (“Renshaw” cell). The intracellular recording was obtained simultaneouslywith rhythmic ventral root potentials recorded from the same segment as the cell. The recordingarrangement is shown to the right of the traces.

developing spinal networks coupled with activity-dependent depression of network func-tion that recovers with a time constant of several minutes. Developing networks are hyper-excitable because the classically inhibitory transmitters GABA and glycine aredepolarizing. Activity-dependent network depression is mediated by several factors includ-ing post-episode hyperpolarization of motoneurons and interneurons that recovers as adepolarizing ramp between episodes,20 post-episode depression of synaptic transmissionand spontaneously occurring synaptic events that recover in the interval between episodes.21
We have occasionally recorded from interneurons that appear to exhibit voltage-dependent oscillations in the quiescent periods between episodes, but we have no evidencethat they play a crucial role in rhythmogenesis. Moreover, the excitatory amino acid ago-nist NMDA can induce TTX-resistant oscillations in chick spinal neurons, but several linesof evidence indicate that they do not underlie the spontaneously occurring rhythmic drive22
(see also ref. 23).We have modeled spontaneous activity as the interaction of three variables: the “activ-
ity” of the network (a) which can be considered as the average firing rate, synaptic drive,or the number of neurons discharging; a fast depression variable (d) which limits the fir-ing within each cycle of an episode, and a slow depression variable (s) subsuming severalslow activity-dependent depressors of network activity (see also ref. 24).
The three corresponding equations are:
τa da/dt = a∞ (s*d*a) – a (1)
τd dd/dt = d∞ (a) – d (2)
τs ds/dt = s∞ (a) – s (3)
Each of the variables a, s, and d are fractional levels and range between zero and one.Note that both s and d are functions of a.
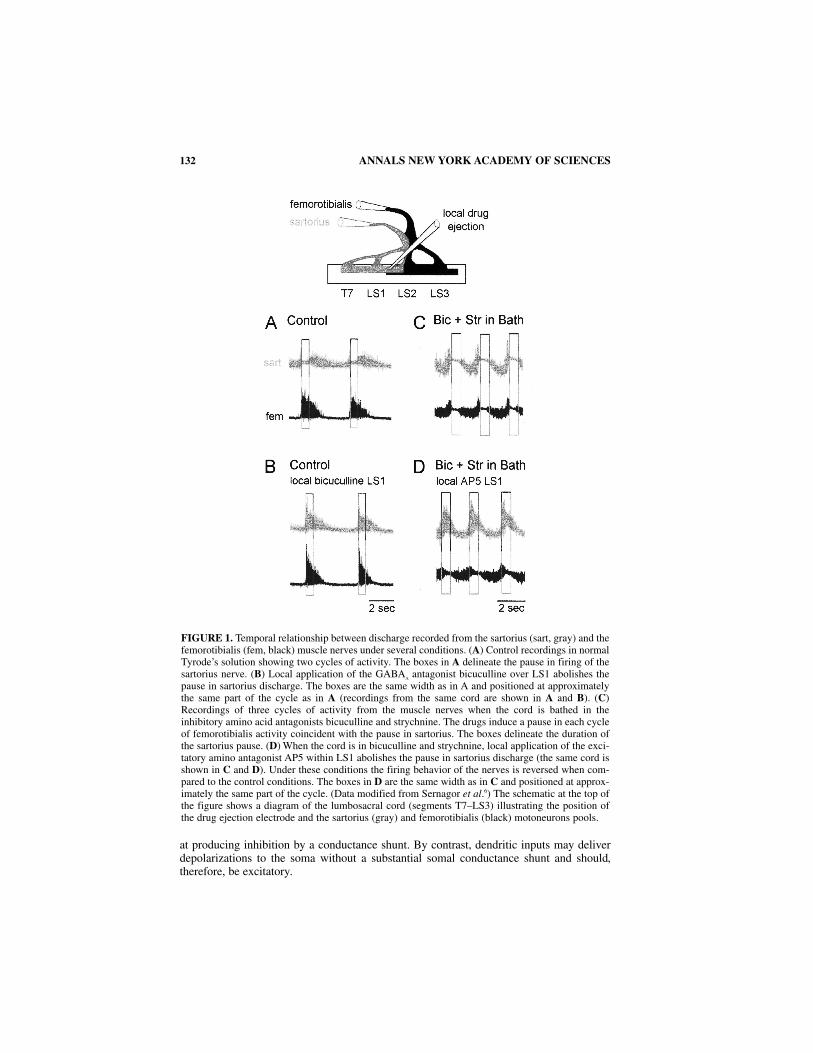
To understand this model, consider first the simplest case where there is no depression(s = 1, d = 1). The network’s dynamics are described by the single equation ta da/dt = a∞(a) – a that can be interpreted in the following way. The function a∞(a) describes therecruitment of the network as a function of a. As illustrated in FIGURE 3A (left panel), thisis a sigmoidal increasing function of a so that a∞(a) is close to 0 for small a and close to1 for large a. The variable a can be considered as the input to the network and a∞(a) asthe output the network approaches with a time constant τa. Thus the function describesthe steady state input-output relation of the network—the network’s steady activity levelas a function of its input, as generated by its own cell firings and synaptic conductances.This is easily visualized in a recurrently connected excitatory network in which activityin any particular region maintains the network in a state of excitation by virtue of itsrecurrent projections to other parts of the network.
For self-consistency then we must have, at steady state a∞(a) = a. Interpreting this graphi-cally in FIGURE 3A means finding intersections of the diagonal line of slope one (a = a) withthe curve a∞(a) versus a. This network, with depolarizing, nondepressing synapses, is bistable.The two open squares identify the network’s rest state (a near zero) and fully active state (a nearone); the open circle defines the threshold so that for a < threshold the network returns to therest state, and for a > threshold the network becomes fully active. Steady states are approachedwith time constant, τa.
If we now include some form of activity-dependent depression (the multiplicative fac-tors s and d in equation 1, then activity will not be maintained but will decrease or termi-nate. Initially, we consider the case where s =1 and d is a dynamic variable that tendstoward d∞ (a) with time constant τd. At high levels of activity the value of d∞(a) will besmall, thereby intensifying the effective depression and limiting activity (a) in the network[d∞(a) is close to 1 for small a, close to 0 for large a]. Such a system with this underlyingbistability can express oscillations when d is a dynamic variable that is slow relative to a, i.e.,
O’DONOVAN et al.: DEVELOPMENT OF MOTOR ACTIVITY 135
136 ANNALS NEW YORK ACADEMY OF SCIENCES
with d approximately 1–10 times a. A discrete model, similar to this, has been introduced bySenn and his colleagues25 to account for spontaneous oscillations observed in cultured spinalnetworks.26 Notice that if the synapses become too depressed (s*d small), then curve a∞(a)
FIGURE 3. Mathematical model of network activity. (A) The middle panel shows the network at threedifferent stages: quiescent (black), just at threshold (green) and fully active (red). The lines relate thesenetwork configurations to the corresponding values of the network activation function a∞(a) (left-hand panel) and the activity of the network as a function of time (right-hand panel). Theleft-hand panel shows the function a∞(a) for two values of the variables s and d. When s*d =1, thecurve intersects the line of identity (a = a) at three points. Stable states are enclosed by the opensquares and the threshold, which is unstable, is shown in the green open circle. A second curve (blue)is also shown for s*d = 0.3. Notice that this curve only intersects the line of identity at the low activ-ity state. (B) Output of the model showing the activity (a) and slow network depression (s) as a func-tion of time. Note that as s increases the “slow” depression recovers. In A and B, suprathresholdactivity is shown in red and subthreshold behavior in black.

shifts to the right and no longer intersects the diagonal line of identity at the high-activitystate. As a result the network evolves into the quiescent state (in FIG. 3A, the blue curve fors = 1, d = 0.3).
If we now include a second “slow” network depression variable s with a time constant1–3 orders of magnitude slower than d, then—depending on the value of s—the networkcan express episodes of rhythmic activity by alternating between the low and high activitystates (FIG. 3B). When activity is low, the slow depression recovers and s increases [s∞(a)is close to 1 for small a, close to 0 for large a]. When recovery is sufficient, the networkwill trigger and jump to the high activity state. During the episode, the slow depressionwill gradually evolve until the value of s reaches a critical level which cannot support net-work activity. At this point the episode stops and, in the absence of network activity, theslow depression variable starts to recover. Thus, the occurrence and termination ofepisodes is controlled by the slow depression, whereas the cycling within an episode is dueto the interaction of activity and the “fast” depression.
It is possible to start an episode before reaching the value of s which is critical for thespontaneous re-initiation of activity. This can be accomplished by electrically stimulatingthe cord before a spontaneous episode would have occurred. Under such conditions, it willtake less time to reach the level of depression at which network activity stops. In otherwords, if the interval before an episode is shortened, then the duration of the followingepisode is also shortened—a correlation that is observed experimentally (see ref. 27).
THE EFFECTS OF CHANGES IN CONNECTIVITY ON NETWORK BEHAVIOR
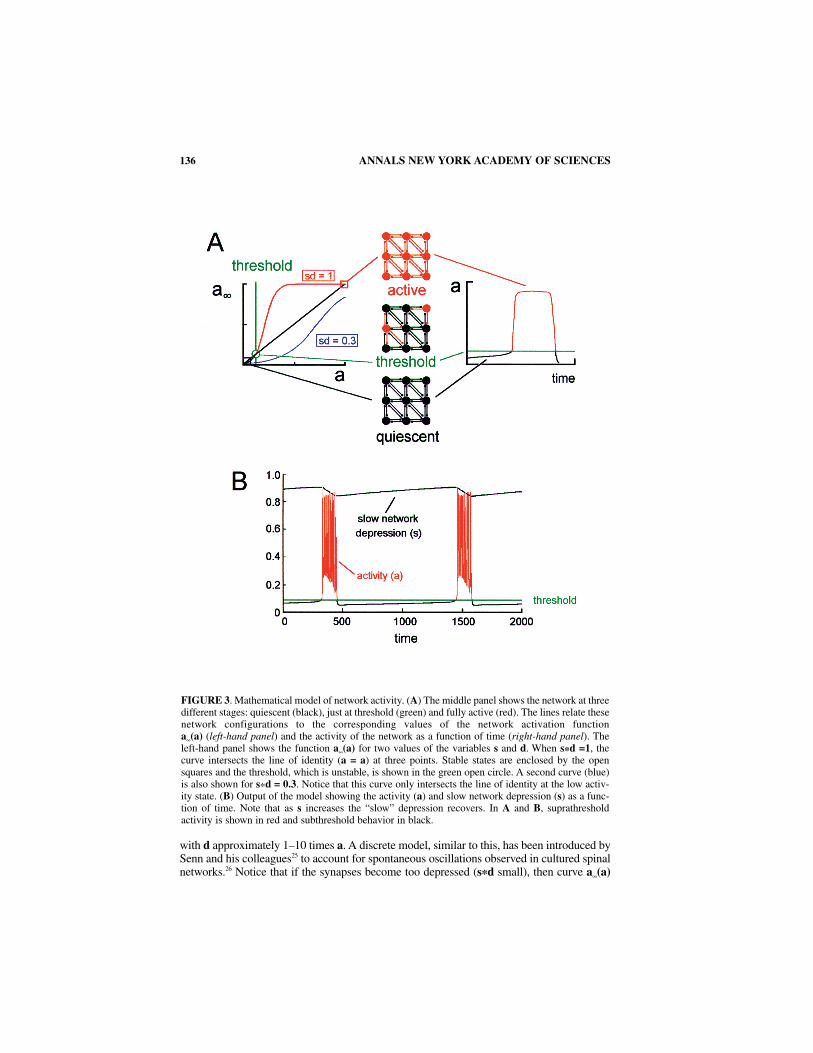
Spontaneous rhythmic episodes recover in the presence of excitatory antagonists or inthe presence of bicuculline and strychnine.6,7,28 Such networks have fewer effective con-nections because the antagonists remain functional during the recovery.7 We have shownthat network activity resumes after a prolonged quiescent interval following either com-plete or partial excitatory blockade and then stabilizes at a slightly lower frequency ofepisodes than before the drugs were added.7,28 By contrast, when network activity recoversin the presence of bath-applied bicuculline and strychnine, the interepisode intervalslengthen considerably and become erratic.7
We found that the model network could mimic the effects of excitatory blockade (FIG.4) if a network connectivity factor (η) was included and if this parameter was suddenlyreduced. It is important to emphasize that the model considers the excitatory drive derivedfrom glutamatergic and GABA/glycine inputs to be functionally equivalent so that block-ade of glutamatergic transmission can be considered simply as a reduction of connectivity.
The extended description of the network becomes
τa da/dt = a∞ (η*s*d*a) – a (1′)
Decreasing η can be interpreted as a reduction in the synaptic weights of existingconnections or a reduction in the number of functional connections. In the model a sud-den reduction of η produces a very long interval following which the network reestab-lishes its prior levels of high and low oscillating activity (FIG. 4A, model). Theimportant insight provided by the model was that for lower “connectivity” (η < 1) thenetwork reestablishes activity in a range of reduced slow depression (higher s). Thislower level of network depression arises in the following way. The “effective functionalconnectivity” of the network is the argument of a∞ which is (η*s*d). In the model thenetwork can only become active at a particular value of η*s. Therefore, if η is reduceds must increase to reach the appropriate value of η*s. In physiological terms this meansif the connectivity is reduced then the remaining connections must compensate by
O’DONOVAN et al.: DEVELOPMENT OF MOTOR ACTIVITY 137
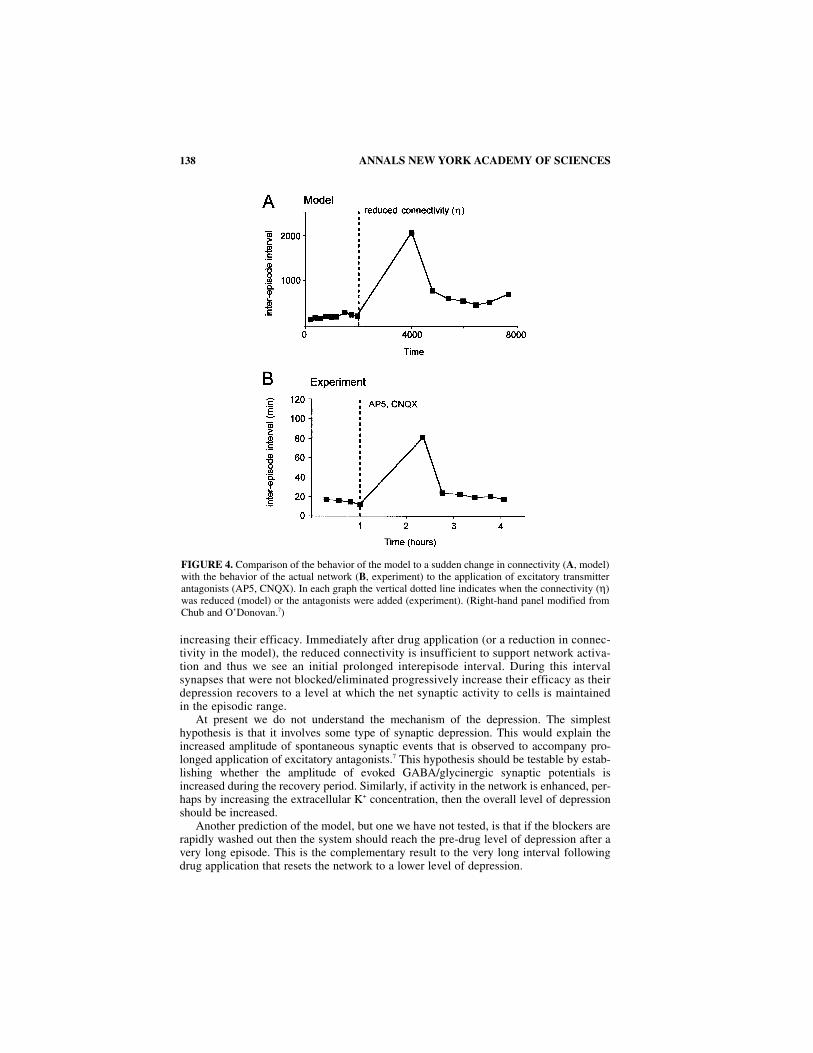
increasing their efficacy. Immediately after drug application (or a reduction in connec-tivity in the model), the reduced connectivity is insufficient to support network activa-tion and thus we see an initial prolonged interepisode interval. During this intervalsynapses that were not blocked/eliminated progressively increase their efficacy as theirdepression recovers to a level at which the net synaptic activity to cells is maintainedin the episodic range.
At present we do not understand the mechanism of the depression. The simplesthypothesis is that it involves some type of synaptic depression. This would explain theincreased amplitude of spontaneous synaptic events that is observed to accompany pro-longed application of excitatory antagonists.7 This hypothesis should be testable by estab-lishing whether the amplitude of evoked GABA/glycinergic synaptic potentials isincreased during the recovery period. Similarly, if activity in the network is enhanced, per-haps by increasing the extracellular K+ concentration, then the overall level of depressionshould be increased.
Another prediction of the model, but one we have not tested, is that if the blockers arerapidly washed out then the system should reach the pre-drug level of depression after avery long episode. This is the complementary result to the very long interval followingdrug application that resets the network to a lower level of depression.
138 ANNALS NEW YORK ACADEMY OF SCIENCES
FIGURE 4. Comparison of the behavior of the model to a sudden change in connectivity (A, model)with the behavior of the actual network (B, experiment) to the application of excitatory transmitterantagonists (AP5, CNQX). In each graph the vertical dotted line indicates when the connectivity (η)was reduced (model) or the antagonists were added (experiment). (Right-hand panel modified fromChub and O’Donovan.7)

It is important to emphasize that there are several aspects of network behavior that arenot predicted by the model. For example, in the model, network depression is at a maxi-mum immediately after the episode. However, experimentally synaptic noise and evokedpotentials do not reach a minimum until about 1–2 min after an episode has ended. In addi-tion, when network activity recovers in the presence of the NMDA receptor antagonistAPV, it first exhibits a long interval (as predicted by the model) but then subsequent inter-vals oscillate (not predicted by the model) before stabilizing at a new level.28 Finally, whenactivity recovers in the presence of bicuculline and strychnine, the interepisode intervalsslow and become unstable. We do not know what accounts for this difference in the behav-ior of networks under excitatory and GABA/glycine blockade. One possibility is that thedepression mechanisms operate differently on GABA/glycine and glutamatergic synapses.This is testable by comparing the amplitude and frequency dependence of glutamatergicand GABA/glycine synapses to alterations in overall network activity.
CONCLUDING REMARKS AND SPECULATIONS
Our work has begun to reveal the mechanisms responsible for patterned rhythmic activ-ity expressed by developing spinal networks in the chick spinal cord. The evidence sug-gests that rhythmic activity is an emergent property of recurrently connected, functionallyexcitatory networks coupled with activity-dependent network depression. Further workwill be required to define the mechanisms for the depression of network activity and toestablish if they act presynaptically, postsynaptically or a combination of both. In theremaining part of this chapter we will highlight some of the mechanisms, that appear tofunction in developing spinal networks and consider to what extent they could operate inthe locomotor circuits of adult animals.
We have identified several mechanisms in developing networks that have not yet beendescribed in the rhythmogenesis or pattern formation of adult animals. The first is a selec-tive shunting of action potential generation in flexor motoneurons, such as the sartorius, atthe time of extensor activity. We do not know how the shunting mechanism becomes trans-formed into the adult mechanisms responsible for the timing of motoneuron discharge dur-ing locomotion. It is unlikely that a depolarizing shunt contributes to the alternation ofadult flexor and extensor discharge because of the developmental emergence of hyperpo-larizing GABA and glycine synaptic potentials. However, hyperpolarizing IPSPs have anassociated conductance change that could influence motoneuron discharge particularly ifthe inhibitory synapses are located close to the soma. This is an issue which has not beenthoroughly investigated in adult animals.
In the embryonic cord, sartorius motoneurons are more susceptible than femorotibialismotoneurons to action potential blockade by their synaptic inputs. This could occurbecause of differences in their intrinsic membrane properties or alternatively in thestrength, number or location of their synaptic inputs. Whatever the mechanism, it will beof great interest to establish whether any of these hypothesized differences are preservedin the adult cord, and if so, whether they contribute to the timing of motoneuron discharge.
The second mechanism we have identified in the developing spinal cord is the explo-sive recruitment of networks which occurs because of positive feedback arising from therecurrent excitatory connections. Such a pattern of activation might be termed auto-recruitment because a small suprathreshold input can recruit the whole network. Withdevelopment and the acquisition of hyperpolarizing IPSPs, networks become less excitableand lose their spontaneous activity. However, auto-recruitment of networks affords a veryefficient mechanism for activating a population of neurons with a small control signal thatmight operate in the adult cord. Hyperpolarizing IPSPs could provide an important regu-latory mechanism for limiting or shaping network recruitment. Although no evidence
O’DONOVAN et al.: DEVELOPMENT OF MOTOR ACTIVITY 139

140 ANNALS NEW YORK ACADEMY OF SCIENCES
exists for such a mechanism operating in mature locomotor networks, it may become man-ifest in the adult nervous system in pathological conditions such as epilepsy.29
Finally, we have argued that network activity is terminated by some form of networkor synaptic depression. At present there is no evidence to support this idea in adult loco-motor networks although the issue has not been thoroughly investigated. Synaptic depres-sion can be detected in the connections of muscle spindle afferents to motoneurons in thespinal cord of the adult cat but it is dominated by post-tetanic potentiation.30 It is possiblethat the degree of depression and potentiation could be independently regulated at partic-ular times or at particular synapses. Synaptic depression is a feature of adult corticalsynapses31,32 and has been postulated to play a role in the genesis of spontaneous synchro-nized activity in cultured cortical neurons.33
REFERENCES
1. BEKOFF, A. 1976. Ontogeny of leg motor output in the chick embryo: A neural analysis. BrainRes. 106: 271–291.
2. O’DONOVAN, M. J. 1989. Motor activity in the isolated spinal cord of the chick embryo: Synapticdrive and firing pattern of single motoneurons. J. Neurosci. 9: 943–958.
3. PERRET, C. 1983. Centrally generated pattern of motoneuron activity during locomotion in thecat. Symp. Soc. Exp. Biol. 37: 405–422.
4. ORSAL, D., C. PERRET & J. M. CABELGUEN. 1986. Evidence of rhythmic inhibitory synaptic influ-ences in hindlimb motoneurons during fictive locomotion in the thalamic cat. Exp. Brain Res.64: 217–224.
5. SERNAGOR, E. & M. J. O’DONOVAN. 1991. Whole cell patch clamp of rhythmically activemotoneurons in the isolated spinal cord of the chick embryo. Neurosci. Lett. 128: 211–216.
6. SERNAGOR, E., N. CHUB, A. RITTER & M. J. O’DONOVAN. 1995. Pharmacological characteriza-tion of the rhythmic synaptic drive onto lumbosacral motoneurons in the chick embryo spinalcord. J. Neuroscience 15: 7452–7464.
7. CHUB, N. & M. J. O’DONOVAN. 1998. Blockade and recovery of spontaneous rhythmic activityfollowing application of neurotransmitter antagonists to spinal networks of the chick embryo.J. Neuroscience 18: 294–306.
8. OBATA, K., M. OIDE & H. TANAKA. 1978. Excitatory and inhibitory actions of glycine on embry-onic chick spinal neurons in culture. Brain Res. 144: 179–184.
9. WU, W., L. ZISKIND-CONHAIM & M. A. SWEET. 1992. Early development of glycine- and GABA-mediated synapses in rat spinal cord. J. Neurosci. 12: 3935–3945.
10. NISHIMARU, H., M. IIZUKA, S. OZAKI & N. KUDO. 1996. Spontaneous motoneuronal activitymediated by glycine and GABA in the spinal cord of rat fetuses in vitro. J. Physiol. 497:131–143.
11. HO, S. & M. J. O’DONOVAN. 1993. Regionalization and inter-segmental coordination of rhythmgenerating networks in the spinal cord of the chick embryo. J. Neurosci. 13: 1345–1371.
12. WENNER, P., M. MATISE, A. JOYNER & M. J. O’DONOVAN. 1998. Physiological and molecu-lar characterization of interneurons in the developing spinal cord. Ann. N. Y. Acad. Sci.This volume.
13. WENNER, P., M. MATISE & M. J. O’DONOVAN. 1997. Physiological and molecular characteriza-tion of interneurons in the developing spinal cord. Soc. Neurosci. Abstr. 23: 1978.
14. ANTAL, M., A. CS. BERKI, L. HORVATH & M. J. O’DONOVAN. 1994. Developmental changes in thedistribution of GABA-immunoreactive neurons in the embryonic chick lumbosacral spinalcord. J. Comp. Neurol. 341: 1–9.
15. MA, W., T. BEHAR & J. L. BARKER. 1992. Transient expression of GABA immunoreactivity inthe developing rat spinal cord. J. Comp. Neurol. 325: 271–290.
16. BERKI, A. C., M. J. O’DONOVAN & M. ANTAL. 1995. Developmental expression of glycine-immunoreactivity and its colocalization with GABA in the embryonic chick lumbosacralspinal cord. J. Comp. Neurol. 363: 1–14.
17. KOTAK, V. C., S. KORADA, I. R. SCHWARTZ & D. H. SANES. 1998. A developmental shift fromGABAergic to glycinergic transmission in the central auditory system. J. Neurosci. 18:4646–4655.

18. O’DONOVAN, M. J. & N. CHUB. 1997. Population behavior and self-organization in the genesisof spontaneous rhythmic activity by developing spinal networks. Semin. Cell Dev. Biol. 8:21–28.
19. O’DONOVAN, M. J., N. CHUB & P. WENNER. 1998. Mechanisms of spontaneous activity in devel-oping spinal networks. J. Neurobiol. In Press.
20. CHUB, N. & M. J. O’DONOVAN. 1995. Multi-functional actions of GABA in the regulation ofspontaneous activity in the isolated spinal cord of the chick embryo. Soc. Neurosci. Abstr. 21:688.
21. FEDIRCHUK, B. & M. J. O’DONOVAN. 1996. Evoked potentials are transiently depressed follow-ing rhythmic activity in the embryonic chick spinal cord. Soc. Neurosci. Abstr. 22: 1377
22. CHUB, N., L. E. MOORE & M. J. O’DONOVAN. 1997. Comparison of NMDA-induced membranepotential oscillations and spontaneous rhythmic activity in the chick spinal cord. Soc.Neurosci. Abstr. 23: 1978.
23. CHUB, N., L. E. MOORE & M. J. O’DONOVAN. 1997. Comparison of NMDA-induced membranepotential oscillations and spontaneous rhythmic activity in the chick spinal cord. Ann. N. Y.Acad. Sci. This volume.
24. RINZEL, J. & M. J. O’DONOVAN. 1997. A model based on synaptic depression for populationrhythmogenesis. Abstract for CNS*97 meeting.
25. SENN, W., K. WYLER, J. STREIT, M. LARKUM, H-R. LUSCHER, H. MEY, L. MULLER, D.STAINHAUSER, K. VOGT & TH. WANNIER. 1996. Dynamics of a random neural network withsynaptic depression. Neural Networks 9: 575–588.
26. STREIT, J., C. LUSCHER & H-R. LUSCHER. 1992. Depression of postsynaptic potentials by high-frequency stimulation in embryonic motoneurons grown in spinal cord slice cultures. J.Neurophysiol. 68: 1793–1803.
27. TABAK, J. & M. J. O’DONOVAN. 1998. Statistical analysis and intersegmental delays reveal pos-sible roles of network depression in the generation of spontaneous activity in the chick embryospinal cord. Ann. N. Y. Acad. Sci. This volume.
28. BARRY, M. & M. J. O’DONOVAN. 1987. The effects of excitatory amino acids and their antago-nists on the generation of motor activity in the isolated chick cord. Dev. Brain. Res. 36:271–276.
29. WONG, R. K., R. D. TRAUB & R. MILES. 1986. Cellular basis of neuronal synchrony in epilepsy.Adv. Neurol. 44: 583–592.
30. LEV-TOV, A., M. J. PINTER & R. E. BURKE. 1983. Posttetanic potentiation of group Ia EPSPs:Possible mechanisms for differential distribution among medial gastrocnemius motoneurons.J. Neurophysiol. 50: 379–398.
31. DEISZ, R.A. & D. A. PRINCE. 1989. Frequency-dependent depression of inhibition in guinea-pigneocortex in vitro by GABAB receptor feed-back on GABA release. J. Physiol. 412: 513–541.
32. MARKRAM, H. & M. TSODYKS. 1996. Redistribution of synaptic efficacy between neocorticalpyramidal neurons. Nature 882: 807–810.
33. MAEDA, E., H. P. ROBINSON & A. KAWANA. 1995. The mechanisms of generation and propaga-tion of synchronized bursting in developing networks of cortical neurons. J. Neurosci. 15:6834–6845.
O’DONOVAN et al.: DEVELOPMENT OF MOTOR ACTIVITY 141
Related Documents