Exp Brain Res (1992) 91:236-248 Experimental Brain Research Springer-Verlag 1992 Low-threshold, short-latency cutaneous reflexes during fictive locomotion in the "semi-chronic" spinal cat Lisa A. LaBella, Andrzej Niechaj, and Serge Rossignol Centre de Recherche en Sciences Neurologiques, Facult6 de M6decine, Universit6 de Montr6al, C.P. 6128, Succ. A, Montr6al, Qu6bec, Canada H3C 3J7 Received May 9, 1991 / Accepted April 8, 1992 Summary. Low-threshold, short-latency cutaneous re- flexes evoked in ipsilateral hindlimb motor nerves were examined during fictive locomotion. Locomotion in 11 anaemically decerebrated spinal animals (1-3 weeks after transection at T13-L1) was induced by administration of clonidine, L-dopa and nialamide; by administration of the latter two drugs only; or by exteroceptive stimulation in the absence of any drugs. The caudal and lateral cutaneous sural, caudal cutaneous femoral, saphenous and superficial peroneal nerves were stimulated at low threshold (1.5-3 T). Pooled results from all combinations of cutaneous nerves stimulated and muscle nerves re- corded show that the initial response was excitatory in 40 of 50 triceps surae and 17 of 20 semitendinosus (St) electroneurograms (ENGs). These excitatory responses occurred at latencies that ranged from 5 to 15 ms and tended to be maximal during the motor nerve's active period in the step cycle (i.e. they were modulated in a phase-dependent manner). Only three inhibitory re- sponses (9-12 ms earliest latency) were encountered in total: in two St ENGs of one animal and in one lateral gastrocnemius-soleus ENG of a different animal. In two animals a "second" excitatory response (15-25 ms laten- cy) was sometimes recorded in triceps surae and St nerves and, interestingly, could be modulated out of phase with the early response. Weak short-latency excitatory reflexes were also found in contralateral St ENGs when exam- ined. Finally, among medial gastrocnemius, lateral gas- trocnemius and soleus nerves, excitatory responses due to stimulation of any particular cutaneous nerve tended to be modulated similarly but were of consistently dif- ferent amplitude among the three. This finding, together with the general observation that excitatory reflexes produced by stimulation of a particular cutaneous nerve were modulated similarly in extensors (or flexors) of different animals, suggests that spinal circuits generating locomotion may indeed exert a stereotypic control over interneurons in specific cutaneous reflex pathways to motoneurons. The results are primarily discussed in Correspondence to: S. Rossignol terms of the existing evidence for short-latency excitatory cutaneous reflexes in extensors in a variety of locomotive and non-locomotive preparations. Key words: Cutaneous reflexes Fictive locomotion Electroneurography - Semi-chronic spinal cat Introduction Excitatory reflexes to flexors or extensors, evoked by stimulation of low-threshold cutaneous afferents, have been shown to undergo modulation during different phases of the locomotor step cycle. Such modulation has been seen in intact cats (Duysens and Stein 1978; Fors- sberg 1979; Duysens and Loeb 1980; Abraham et al. 1985; Drew and Rossignol 1987) and decerebrate ani- mals (Duysens and Pearson 1976). The demonstration of such phase dependency in both walking spinal cats (Fors- sberg et al. 1975, 1977) and acute (Andersson et al. 1978; Schomburg and Behrends 1978; Schmidt et al. 1988, 1989) or chronic (Andersson et al. 1978) spinal cats during fictive locomotion strongly suggests that, in ad- dition to certain inhibitory spinal interneurons (see Pratt and Jordan 1987), excitatory interneurons in segmental reflex pathways may be phasically driven by the spinal central pattern generator for locomotion. Evidence for such phasic control over excitatory cutaneous reflex pathways, however, may be strongest for pathways to flexor motoneurons. For example, with spinal cats walking on a treadmill, the most common observation has been excitatory responses in flexors if the stimulus is applied during swing, but a mix of excitatory and inhibi- tory responses in extensors if the stimulus is applied during stance (for review see Rossignol et al. 1988). Studies that have recorded directly from motoneurons during fictive locomotion have reported similar results. One investigation reported evidence for phase-dependent cutaneous-evoked excitatory postsynaptic potentials (EPSPs) in both flexor and extensor motoneurons but

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Exp Brain Res (1992) 91:236-248
Experimental Brain Research �9 Springer-Verlag 1992
Low-threshold, short-latency cutaneous reflexes during fictive locomotion in the "semi-chronic" spinal cat Lisa A. LaBella, Andrzej Niechaj, and Serge Rossignol
Centre de Recherche en Sciences Neurologiques, Facult6 de M6decine, Universit6 de Montr6al, C.P. 6128, Succ. A, Montr6al, Qu6bec, Canada H3C 3J7
Received May 9, 1991 / Accepted April 8, 1992
Summary. Low-threshold, short-latency cutaneous re- flexes evoked in ipsilateral hindlimb motor nerves were examined during fictive locomotion. Locomotion in 11 anaemically decerebrated spinal animals (1-3 weeks after transection at T13-L1) was induced by administration of clonidine, L-dopa and nialamide; by administration of the latter two drugs only; or by exteroceptive stimulation in the absence of any drugs. The caudal and lateral cutaneous sural, caudal cutaneous femoral, saphenous and superficial peroneal nerves were stimulated at low threshold (1.5-3 T). Pooled results from all combinations of cutaneous nerves stimulated and muscle nerves re- corded show that the initial response was excitatory in 40 of 50 triceps surae and 17 of 20 semitendinosus (St) electroneurograms (ENGs). These excitatory responses occurred at latencies that ranged from 5 to 15 ms and tended to be maximal during the motor nerve's active period in the step cycle (i.e. they were modulated in a phase-dependent manner). Only three inhibitory re- sponses (9-12 ms earliest latency) were encountered in total: in two St ENGs of one animal and in one lateral gastrocnemius-soleus E N G of a different animal. In two animals a "second" excitatory response (15-25 ms laten- cy) was sometimes recorded in triceps surae and St nerves and, interestingly, could be modulated out of phase with the early response. Weak short-latency excitatory reflexes were also found in contralateral St ENGs when exam- ined. Finally, among medial gastrocnemius, lateral gas- trocnemius and soleus nerves, excitatory responses due to stimulation of any particular cutaneous nerve tended to be modulated similarly but were of consistently dif- ferent amplitude among the three. This finding, together with the general observation that excitatory reflexes produced by stimulation of a particular cutaneous nerve were modulated similarly in extensors (or flexors) of different animals, suggests that spinal circuits generating locomotion may indeed exert a stereotypic control over interneurons in specific cutaneous reflex pathways to motoneurons. The results are primarily discussed in
Correspondence to: S. Rossignol
terms of the existing evidence for short-latency excitatory cutaneous reflexes in extensors in a variety of locomotive and non-locomotive preparations.
Key words: Cutaneous reflexes Fictive locomotion Electroneurography - Semi-chronic spinal cat
Introduction
Excitatory reflexes to flexors or extensors, evoked by stimulation of low-threshold cutaneous afferents, have been shown to undergo modulation during different phases of the locomotor step cycle. Such modulation has been seen in intact cats (Duysens and Stein 1978; Fors- sberg 1979; Duysens and Loeb 1980; Abraham et al. 1985; Drew and Rossignol 1987) and decerebrate ani- mals (Duysens and Pearson 1976). The demonstration of such phase dependency in both walking spinal cats (Fors- sberg et al. 1975, 1977) and acute (Andersson et al. 1978; Schomburg and Behrends 1978; Schmidt et al. 1988, 1989) or chronic (Andersson et al. 1978) spinal cats during fictive locomotion strongly suggests that, in ad- dition to certain inhibitory spinal interneurons (see Pratt and Jordan 1987), excitatory interneurons in segmental reflex pathways may be phasically driven by the spinal central pattern generator for locomotion. Evidence for such phasic control over excitatory cutaneous reflex pathways, however, may be strongest for pathways to flexor motoneurons. For example, with spinal cats walking on a treadmill, the most common observation has been excitatory responses in flexors if the stimulus is applied during swing, but a mix of excitatory and inhibi- tory responses in extensors if the stimulus is applied during stance (for review see Rossignol et al. 1988). Studies that have recorded directly from motoneurons during fictive locomotion have reported similar results. One investigation reported evidence for phase-dependent cutaneous-evoked excitatory postsynaptic potentials (EPSPs) in both flexor and extensor motoneurons but

237
found tha t the m o d u l a t i o n in extensors was less p r o n o u n c e d and tha t pa t t e rn s o f m o d u l a t i o n "va r i e d subs tan t i a l ly be tween different cells o f the same species" ( A n d e r s s o n et al. 1978). Two o the r s tudies r e p o r t e d clear evidence p r i m a r i l y for f lexor m o t o n e u r o n s , wi th m o r e va r i ab le resul ts ( S c h o m b u r g and Behrends 1978) or lit t le evidence (Schmid t et al. 1989) for phase d e p e n d e n c y in ex tensor m o t o n e u r o n s . A m o r e recent s tudy ( M o s - chovak i s et al. 1991) has shown tha t two sho r t - l a t ency cu t aneous p a t h w a y s giving exc i t a t ion to f lexor d i g i t o r u m longus ( F D L ) m o t o n e u r o n s were con t ro l l ed different ia l - ly in the oppos i t e phase o f the fictive l o c o m o t o r cycle. The superf icial pe ronea l (SP) nerve gave exc i ta t ion to F D L du r ing f lexion when it was n o r m a l l y active, while the med ia l p l a n t a r nerve exci ted F D L m o t o n e u r o n s on ly when it also d i s c h a r g e d dur ing the ex tensor phase .
The a p p a r e n t incons is tencies in responses o f ex tensors to cu t aneous s t imu la t ion du r ing l o c o m o t i o n is puzzl ing , since cu t aneous p ro jec t ions to ex tensor m o t o n e u r o n s have long been descr ibed (Wi l son 1963; C o l e b a t c h and Gil l ies 1979). Two recent in t race l lu la r s tudies us ing non- l o c o m o t i n g p r e p a r a t i o n s s u p p o r t the ear ly sugges t ion o f H a g b a r t h (1952) tha t h i n d l i m b ex tensor m o t o r nuclei are p re fe ren t ia l ly exci ted b y cu t aneous nerves ar i s ing f rom m o r e c i r cumsc r ibed sk in regions t han is the case for f lexor m o t o n e u r o n pools . F o r example , in the first o f these studies, t r iceps surae m o t o r nuclei - despi te be ing close func t iona l synergis ts - were shown to receive dif- ferent ia l exc i t a to ry i npu t f rom each o f the two cu t aneous sural nerves (LaBel la et al. 1989). In the second s tudy, in which five h i n d l i m b cu t aneous nerves were tested, med ia l ga s t rocnemius m o t o n e u r o n s were shown to receive p o t e n t and conve rgen t exc i t a to ry i n p u t f rom two nerves wi th n e a r b y or ove r l app ing recept ive fields ( the cauda l cu t aneous sural (CCS) a n d cauda l cu t aneous f emora l ( C C F ) nerves ; LaBel la and M c C r e a 1990). The ques t ion presen t ly add res sed is whe the r or no t a c lear (i.e. consis- tent) phase d e p e n d e n c y o f exc i t a to ry cu t aneous reflexes in ex tensors du r ing fictive l o c o m o t i o n will emerge i f we focus our s tudy on t r iceps surae and s t imula te cu t aneous nerves which are k n o w n to p r o d u c e l ow- th re sho ld exci ta- t ion in t r iceps surae m o t o n e u r o n s (see LaBel la et al. 1989 and LaBe l l a and M c C r e a 1990).
As this r e p o r t describes, in a d d i t i o n to exc i t a to ry cu t aneous reflexes r eco rded in h i n d l i m b flexor nerves o f spinal cats (1 3 weeks pos t t ransec t ion) , those evoked in the ank le ex tensor nerves also u n d e r g o cons i s t en t phase- d e p e n d e n t m o d u l a t i o n dur ing the fictive s tep cycle. F u r - t he rmore , wi th on ly some except ions , these reflexes are enhanced dur ing the phase o f the cycle in which the nerves are typ ica l ly act ive ( A n d e r s s o n et al. 1978; S c h o m b u r g and Behrends 1978), a l t h o u g h the p e r i o d o f increased reflex respons iveness does n o t s t r ic t ly las t t h r o u g h o u t the pe r iod o f l o c o m o t o r act ivi ty.
Materials and methods
The aim of the experiments was to characterize short-latency re- sponses to cutaneous nerve stimulation during fictive locomotion in the spinal cat. Normally, the adequate subject for such studies would be the acute spinal cat injected with nialamide and L-dopa
(Grillner and Zangger 1979; Schmidt et al. 1989). We have recently shown, in cats spinalized 1-2 weeks prior to the acute experiment, that fictive locomotion was sometimes evoked without drugs and also that, with clonidine only or L-dopa, locomotion was altogether easier to evoke and had a fi'equency more similar to the normal than in acutely spinalized cats (Pearson and Rossignol 1991). We have also shown that, as a function of time after spinalization, the fictive locomotor pattern becomes progressively more complex and ex- presses more features characteristic of the normal locomotor pat- tern. These facts led us to use animals (n = 11 ; mean weight 3.2 kg) that had been spinalized 1-3 weeks prior to the acute experiments.
Spinalization procedure
Spinal cords were transected using sterile techniques and with the animals under pentobarbital sodium anaesthesia (32 mg/kg in- traperitoneally or intravenously). Rostral and caudal cord segments were completely separated by dissection with fine forceps under a dissecting microscope. Penicillin G was administered during the first postoperative week (100000 IU/day). Bladders were manually expressed daily. Although the animals were not exercised on a treadmill during the postspinalization period, they were free to move around in individual large cages covered with absorbent cardboard, which was changed regularly. Cats were examined daily, and all cats had recovered brisk hindlimb reflexes when pressing on the foot or air stepping at the time of the acute experiment.
Preparation for the acute experiment
Animals were surgically prepared under steroid anaesthesia, using intravenous Saffan 12 mg/kg (alphaxalone 9 mg/kg and alpha* dolone acetate 3 mg/kg). A tracheotomy was performed and one carotid artery cannulated to monitor blood pressure. An intrave- nous cannula was placed in either a forelimb or jugular vein for drug and fluid administration as well as for slow infusion of a glucose/ bicarbonate buffer. Atropine (0.12 mg) was given subcutaneously, prednisolone sodium succinate (20 mg every few hours) intramus- cularly. Animals were then anaemically decerebrated by ligating the remaining intact carotid artery and the basilar artery (rostral to the posterior inferior cerebellar artery), which was reached by opening the skull ventrally at the level of the tympanic bulla (Pollock and Davis 1923). This was usually followed by a marked rigidity of the forelimbs and in a few cases the animal had to be ventilated. Given the residual effects of the anaesthetics, there was usually some time to initiate the dissection of hindlimb nerves before paralysis [pan- curonium bromide (Pavuton) 0.3 mg/kg] and automatic ventilation. Previous experiments using Saffan for minor surgery in otherwise intact cats showed that cats were wide awake and walking around about 3 h after a single dose. In the present experiments, recordings started more than 3 h after injection of Saffan.
Several nerves of the left (ipsilateral) hindlimb were cut and dissected free of surrounding tissue for subsequent stimulation (cutaneous nerves) or recording (motor nerves) on bipolar hook electrodes or cuff electrodes made of polysyloxane polymer and used in a monopolar configuration (Julien and Rossignol 1982; Pearson and Rossignol 1991). Semitendinosus (St), medial gas- trocnemius (MG) and lateral gastrocnemius-soleus (LGS) nerves were placed in cuff electrodes. In some experiments the soleus (Sol) branch of the LGS nerve was dissected away to allow independent recording from lateral gastrocnemius (LG) and Sol efferents (in cuff electrodes or on bipolar hooks). In some experiments the contra- lateral St (coSt) nerve was also recorded through a monopotar cuff electrode. Cutaneous nerves stimulated for the production of reflex effects in ipsitateral motor nerves include the CCS and lateral cutaneous sural (LCS) nerves; the long distal branch of the CCF nerve; the saphenous nerve (Saph), taken approximately 1 cm above the level of the knee; and the SP nerve dissected at the ankle. CCS, LCS and CCF were stimulated on bipolar hook electrodes; Saph and SP were stimulated through monopolar cuff electrodes.

238
A I I I
23
B
C
STIMULI t 6ms~ /k, ~15rns T
= 0.06 , I / \ 1 / i / \ t
. . . . . . . . . . . . t _ I . . . . . . .L . . . . . . _ 4 , i I I I
k N = 39 t T I
n I do = 0.06 u i INT = ,328 U [-~ 4 1 n m J !
D C O N T R O L S N=1o2 I T
qb = 0.06 , INT = 146 U ,
E SUBTRACTED , R E C O R D S m I STIM CCS / ~
5 ms
Fig. 1A-G. Method for measuring size, latency and durat ion of cutaneous reflexes in ENGs of motor nerves. A samples of raw ENGs from M G nerve during a locomotor sequence with stimuli indicated by ticks above the ENGs, which occur at every 2 cycles in this case. The control cycle is defined by the period between the onset of two successive M G bursts in which there was no stimulus. B Thirty-nine stimuli occurring within the first 10% of the control cycle (mean occurrence 6% or q~=0.06). The two horizontal lines before the artefact (dotted line) and at the end of the record re- present two standard deviations (SDs) above and below the mean level of activity. The two arrows prolonged by dotted lines delineate an area where the signal exceeds 2 SDs. C The bins enclosed within the two arrows in B are blackened and summed. INT value is the
F ~b= 0.06 N=39 _ ~ 1 4 6 7 / _ 8 2 _ I
qb-= 0.14 N=18 , 297- 114 = ! . . . . . . . . . . . 'sa T qb = 024 N=24 n 2 6 6 - 1 0 6 = 1 6 0 1
qf_=~_= 0.36 N_~_ =_ 1 ~ / _ - _110--_172=i
= 0.64 N=10 , 147- 79 = 68 1"
qb = 0.75 N=19 ~ 40 -30 = 10 $
qb = 0.85 N=15
qb = 0,97 N=9
G MG
100 - LU (,9 Z O (3- 6O
~c50 x <
0
, 0-0=0 I
, 9 2 - 6 7 = 2 5 1" I I
5 ms
I ii 14 i!i1--~ 070690
J 0 0.5 1.0
PHASE OF STIMULATION
sum of these bins in arbitrary units. D The mean activity in 102 control cycles at the period corresponding to the response area defined in C. E The bins in C minus the corresponding bins in D. The midpoint of each bin is used to display the subtracted record, as was the case for B. F All responses in each of the ten groups of one sequence. The values to the left represent the mean ~p value of all stimuli (N) occurring within that group. The three values to the right of each group represent, respectively, the integrated area of the response, the integrated area of the equivalent control period and the difference between the two. G The average response in each group is plotted as a percentage of the maximal response. The rectangle represents the mean burst duration of 20 cycles ( + 1 SD). Data taken from cat 1
Only the head of the supine animal was rigidly fixed to a steel table frame, and a mineral oil pool, made by fixing the skin to a sturdy circular rod above the popliteal fossa, bathed the dissected nerves of the extended left hindlimb. In other studies, it was shown that extension of the hindlimb is actually favourable at the develop- ment of the fictive rhythm, contrary to flexion (Pearson and Rossig- nol 1991). Both pool and internal body temperatures were mon- itored and regulated by a heat lamp. Arterial blood pressure and end tidal expired carbon dioxide were continuously monitored.
Data are taken from 7 of the 11 animals operated on, all of which maintained a strong, stable locomotor rhythm throughout the experiment. In 5 of the 7, fictive locomotion was induced by intravenous injection of clonidine (500 gg/kg) followed by injection of nialamide (50 mg/kg) and L-dopa (80 mg/kg); in 1, no clonidine
was administered; and in the remaining cat no drugs at all were administered. The animal which displayed fictive locomotion with- out drugs had the longest post-transection period (3 full weeks). Gentle manipulat ion of the perineal or right hindpaw region was sometimes used to assist in developing or maintaining the loco- motor rhythm in all animals.
St imulat ion and recording
Once the fictive locomotor rhythm was induced in each animal, the "sciatic threshold" (T) of the CCS, LCS and SP nerves was deter- mined by measuring the amount of current (delivered in constant current pulses of 0.2 ms duration) needed to produce the first

239
detectable volley in the sciatic nerve recording. This recording was made th rough a bipolar cuff electrode, placed as proximally on the sciatic nerve as possible, and was displayed on an oscilloscope along with selected ENGs . St imulat ion o f S P , C C S and LCS to produce reflex effects was then applied at current s t rengths that were multi- ples o f T. For Saph and C C F st imulation, T was de termined by measur ing the amoun t o f current needed to just p roduce a reflex effect in any o f the m o t o r nerves as moni tored on the oscilloscope. Thus, T current s t rengths applied to these two cutaneous nerves are less reliable and are probably somewhat greater than those applied to the o ther cutaneous nerves.
In each experiment , series of single-shock stimuli (and in one exper iment shown in Fig. 6, triple shocks) were applied to each dissected cutaneous nerve (at different times) at a current s t rength between 1.5 and 3T, usually at 2T. There were two kinds o f series per formed on each cutaneous nerve. In the "delayed-s t imulat ion" type, the delivery o f current was restricted to approximately every second step cycle. St was normal ly used to trigger the st imulat ion because it usually has a clean signal to noise ratio. Fo r this, the St signal was hardware rectified and filtered and fed into a Schmitt trigger. Thus, a s tandard pulse was generated at every burs t and every second or third pulse was used to trigger a Grass $88 stimula- tor. By adjust ing the delay of the stimulus relative to the onset o f St (in steps o f 100 ms), the stimuli were applied at different times within the cycle. Usual ly 1 0-40 stimuli were given at any one delay. This type o f series typically covered a 15- to 20-min per iod o f fictive locomotion. In "cont inuous-s t imula t ion" series, however, stimuli were simply applied at either 4 or 5 Hz th roughou t the recorded period o f locomot ion (Perret and Cabelguen 1980). This type o f series typically lasted 1.5-2.5 min. Al though the polar i ty o f the responses (excitatory or inhibitory) was the same with bo th types o f st imulation, the effects on the overall cycle structure or durat ion, and hence the dis t r ibut ion o f the responses within the normal ized cycle, could differ in bo th cases.
E N G s (filtered at a bandwid th o f 300 H b l 0 kHz), stimulus mon i to r and voice log were recorded on a seven-channel F M tape recorder moving at a speed o f 19 cm/s and having a frequency response o f 0-5000 Hz.
Data analysis
Data were played back onto electrostatic paper (Gould printer, ES-1000) to help in the selection o f appropr ia te series o f stimula- tion. The signals were low-pass filtered th rough a hardware filter set at 1.5 kHz and were digitized at 1 kHz with a P D P 11/34 computer . Cus tom-made software (Zomlefer et al. 1984) was then used to select per iods o f s teady locomot ion, mark the beginning and end of individual step cycles (period between the onset o f two successive extensor M G bursts) and tag cycles as either st imulated (S) or cont ro l (C; see Fig. 1A).
The time o f occurrence of the stimulus after the onset o f the cycle (i.e. onset o f M G ) was used to classify each stimulus in one of ten groups (referred to in Fig. 1 by the q) symbol) o f the mean control cycle obta ined by averaging several cycles ( n ~ 100) dis- persed th roughout the st imulat ion sequence. All responses within a group ( n ~ 10~40 responses) were then compute r rectified and averaged (Fig. 1B, C and F). No te that the phase values on the lef t -hand side of Fig. 1F represent the mean value (expressed in units o f phase) of all the stimuli given in each group. Signals extending above (excitatory responses) or below (inhibitory re- sponses) two s tandard deviat ions o f baseline activity taken f rom a 35-ms prest imulus per iod (see Fig. 1 F) were integrated by summing the appropr ia te bins. Figure IB shows the average o f 39 responses occurring at mean q~ value o f 0.06. The arrows at 6 and 15 ms define an interval where the signal exceeds two s tandard deviations (SDs), represented by the two hor izontal lines to the left o f the vertical
Table 1. Summary o f variations in ampli tude of reflexes produced in muscle nerves with stimulation o f the various cutaneous nerves
Cutaneous Muscle Cat 1 Cat 2 Cat 3 Cat 4 Cat 5 Cat 6 Cat 7 nerve nerve
CCS St 280 - 57 190 360 - 120 - M G - 180 170 190 - 250 - LG . . . . . 180 Sol - - - 60 - LGS - 130 30 50 - -
LCS St 260 - 63 130 40 60 - M G 140 310 40 0 - 90 - LG . . . . 0 Sol . . . . . 0 LGS 410 0 - 6 0 - - -
Saph St 710 1290 400 - - 180 140 M G 0 60 70 - 100 80 70 LG . . . . 530 70 170 Sol . . . . 100 0 80 LGS - 100 220 . . . .
C C F St . . . . . 90 0 M G . . . . 730 470 120 LG . . . . 570 200 90 Sol - - 190 90 90 LGS . . . . . .
SP St 330 270 100 M G - 60 - - - 50 70 LG . . . . . 170 0 Sol . . . . . 0 0 LGS - 60 . . . . .
Fo r each animal (cat 1 to cat 7), the largest average early response for each st imulat ion sequence is expressed as a percentage of back- g round activity in contro l cycles. Negative values represent inhibi-
tory responses. Zeros signify that there were no response in the recorded nerve. The minus sign ( - - ) signifies that the nerve was not recorded in a given st imulat ion sequence

240
dotted line which is the beginning of the stimulus. In Fig. 1C, the same average response is displayed in histogram form, and the bins contained within the interval (blackened bins) defined by the two arrows in Fig. 1B have been summed. The value of this sum, referred to as the integrated value, is expressed in arbitrary units (i.e. 328 U). The equivalent interval at the same mean ~p value was integrated in the control cycles (146 U in Fig. 1D) and subtracted from the response to give a subtracted record (Fig. 1E) representing a pure response above background activity. This process is repeated for each group (see Fig. 1F). The difference between the integrated area of the response and the corresponding control area is given for each phase at the right of Fig. 1F. All these values are then nor- malized to the largest response (in this example, the integrated response of 190 U at q~ 0.45 was set to equal 100%) and plotted as in Fig. 1G. The horizontal rectangle above the phase plot represents the average ENG burst (n ~ 20 ___ 1 SD).
The background activity has not been subtracted from the average responses given as examples in subsequent figures, because this background activity illustrates very well the magnitude of the responses above the normal locomotor activity in each phase (see Fig. 1F). However, all the phase plots are constructed from sub- tracted values, as explained above. The maximal response in one muscle nerve for stimuli of one particular cutaneous nerve might thus be different from the maximal response obtained when stim- ulating another cutaneous nerve. In order to compare the mag- nitude of the responses in one muscle nerve evoked by different cutaneous nerves, the value of the largest response in a sequence (i.e. 190 in Fig. 1F) was expressed as a percentage of the corresponding control value (see Table 1 : second value in "Cat 2" column, 170% ; i.e. 190/112=170%). The percentages given in Table 1 thus represent the amplitude of the maximal responses above back- ground activity.
Note that latencies and durations of responses given in the text are taken from records such as Fig. 1B. Thus, the individual values in the latency ranges and mean/median durations given are them- selves averages of the 10-40 responses in each phase. It should also be remembered that because of the digitization rate (1 kHz), the error in measurement can be as large as 1 ms.
Because there were no unstimulated cycles for series of the continuous-stimulation type, "controls" were derived from av- erages of ENG signals containing randomly delivered stimuli. However, phase plots in all figures of the present report are from series of the delayed-stimulation type. Also note that in illustra- tions of ENG records taken during continuous-stimulation series (Figs. 2A, 3A, 4A and 6A) the artefacts have been partially suppressed for illustration purposes only.
Results
Reflexes described in the present repor t are those result- ing f rom low-threshold (1 .5-3T; usually 2T) single (or in one experiment, triple) shock s t imulat ion o f var ious ip- silateral, h indl imb cutaneous nerves. D a t a are taken f rom a total o f seven animals. Of the 50 triceps surae (MG, LG, Sol or LGS) and 20 St E N G s in all (numbers include all combina t ions o f cu taneous nerves st imulated and muscle nerves recorded), exci tatory reflexes were the first effect o f s t imulat ion in 40 and 17, respectively. Only three incidences o f inhibi tory reflexes as the first effect o f s t imulat ion were encountered: in one St and one LGS nerve, with s t imulat ion o f two and one cu taneous nerves, respectively. In addit ion, because no t all cu taneous mus- cle nerve combina t ions were tested in each animal, the no ta t ion "4/5 St E N G s " to describe, for example, the exci tatory CCS reflexes means that in a total o f five St nerve s t imulat ion sequences, exci tatory responses were
the first effect in four ; and a l though cutaneous nerves were st imulated in two different modes in each animal (see Materials and methods) , the quali ty o f the reflex (excitatory or inhibitory) never changed with mode o f stimulation. Hence, for each cutaneous nerve tested in a given animal only one type o f response (whether excitat- ory or inhibitory) per m o t o r nerve has been counted in the overall incidence. N o t e tha t latencies and durat ions o f responses are reported for flexor and extensor nerves collectively, since with the present sample sizes for each cutaneous nerve no differences in these two groups were observed.
Contra la tera l cu taneous reflexes are briefly described (for coSt nerves), but as they were no t systematically examined their incidence is no t reported. In addit ion, occasional observat ions o f "second" exci tatory responses to cu taneous s t imulat ion (occurring at latencies between 15 and 25 ms) are also described when approriate , i.e. with Saph reflexes.
CCS reflexes
Short- la tency excitatory reflexes were the first effect o f low-threshold CCS st imulat ion in nine o f nine triceps surae and four o f five St ENGs . They occurred at laten- cies ranging f rom 6 to 9 ms (measured f rom the onset o f the stimulus artefact) with mean and median durat ions each o f 5 ms (n - -89 averaged responses; see Materials and Methods) . However , in one experiment low- threshold CCS st imulat ion p roduced an inhibi tory re- sponse (12 ms earliest latency) in St as the first occurr ing effect.
Figure 2 depicts the results o f an experiment in which exci tatory CCS reflexes were evoked in all flexor and extensor nerves tested. Figure 2A illustrates a sample o f rectified E N G s recorded dur ing 4 - H z st imulat ion o f the CCS nerve at 2T and reveals the exci tatory responses in coSt, St, M G and M G S nerves (latency o f excitatory responses in c o S t = 9 - 1 0 ms). N o t e tha t in each o f the 4 E N G s , CCS reflexes appear largest dur ing the m o t o r nerve's active phase in the step cycle. Also note that the ampli tude o f the responses above the l ocomoto r E N G s is no t uniform, bu t rather, is larger in M G and St than in M G S and coSt. Fo r all experiments in which CCS reflexes were examined, the size o f the responses ex- pressed as a percentage o f the backg round l ocomoto r activity in E N G recordings o f extensors was always larg- est in M G (Table 1). Fig. 2C, D illustrates, on a faster time base, the average o f several responses taken during the period o f extensor (Fig. 2C) and flexor activity (Fig. 2D). N o t e in Fig. 2C the ampli tude o f the responses above backg round activity compared with the small re- sponse in LGS (see also Table 1).
The phase plots o f CCS reflexes in Fig. 2B are f rom the same experiment depicted in Fig. 2A but f rom a delayed-st imulat ion series (2T stimuli delivered ap- proximate ly every second step cycle). N o t e that, for M G and St in particular, the CCS reflexes are larger dur ing the m o t o r nerve 's active phase o f the step cycle and smaller dur ing its inactive phase (note the modu la ted

A
st . . . . . . . . . .
LGS
l s
B ~iiiiiiiiiiiiiiiiiiiii~iiiiiiiiiii~iiiiiiiiii1ii~iiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiii~ MG
100
o /
St
0 0.5 1 .O
1 0 0
o
~ S t . 0.5 1.0
PHASE OF STIMULATION
C (I) 0 .4 - 0 .5
I
I
coSt I I
st j ~ A I
L MG T-- -~ . . . . . . . . . . . .
LGS ~ . . . . . . . . . . . I
5 m s
D (I) 0 .8 - 0 .9
I
coSt . . ~ _ _ ~ I
s, I I
I I I A
LGS --f- ~ . __ I
070690
241
Fig. 2A-D. Responses to stimuli delivered to the CCS nerve during fictive locomo- tion in an animal 17 days after spinalization (cat 3). A Rec- tified/filtered (0.005 s time constant) ENGs showing ex- citatory responses in four flexor and extensor nerves during 4~Hz stimulation of the CCS nerve at 2T (9 gA/0.2 ms pulses). B The phase plots show modulation of CCS re- flexes during a delayed- stimulation series at 2T (9 gA pulse applied approximately every second step cycle). Shaded rectangles depict burst duration (+ 1 SD) of extensor (MG) and flexor (St) ENGs during the averaged un- stimulated step cycle. C and D Average of 21 and 17 respon- ses, respectively, in two dif- ferent phases of the step cycle
amplitude of the unaveraged responses within individual M G bursts with 4-Hz stimulation in Fig. 2A). However, in the case of LGS and coSt, the response peaks at a particular point mid-extension and is then decreased throughout flexion and early extension on the ipsilateral side. In general, the pat tern of modula t ion of excitatory CCS reflexes seen in Fig. 2B for St, coSt and M G is typical for all experiments, in both delayed- and con- t inuous-stimulation series. However, pat terns for LGS (or L G and Sol) often resembled that shown for M G as well (i.e. a more maintained amplitude of the reflex throughout extension).
CCF reflexes
As with CCS stimulation, short-latency excitatory re- flexes were the first effect of low-threshold CCF stimula- tion in all nine triceps surae E N G s and in one of two St ENGs. These reflexes (n= 88) had latencies as short as 5 ms (range 5-14 ms; n = 88) and were frequently of longer duration (mean 7 ms, median 5 ms) than CCS reflexes recorded in the same nerves. No inhibitory CCF reflexes were encountered in flexor or extensor nerves. One St nerve had no response, al though it did have excitatory
responses to stimulation of two other cutaneous nerves in the same experiment. Figure 3 depicts results f rom the experiment in which there was no measureable effect of CCF stimulation upon St, whereas excitatory reflexes were recorded in all three triceps surae nerves. Figure 3A illustrates rectified ENGs recorded during 4-Hz stimula- tion of the CCF nerve at 2T and shows large responses in M G and L G records and smaller responses in the Sol ENG. Figure 3B shows the similar modulat ion pattern of CCF reflexes produced in the different triceps surae nerves of this experiment throughout the averaged step cycle. These phase plots are f rom the same experiment depicted in Fig. 3A, but f rom a series with delayed stim- ulation of CCF at 2T. That the excitatory CCF reflexes gNtlargest during each extensor nerve's active phase of the step cycle is evident in both Fig. 3A and B. The pat tern of modulat ion for CCF reflexes in triceps surae was generally as in Fig. 3B for all experiments: a gradual at tenuation of amplitude during extension, followed by a rapid at tenuation at the transition to flexion where the reflex remained attenuated. The one excitatory reflex recorded in an St nerve of a different experiment (not illustrated) slowly increased throughout extension to peak in mid-flexion.
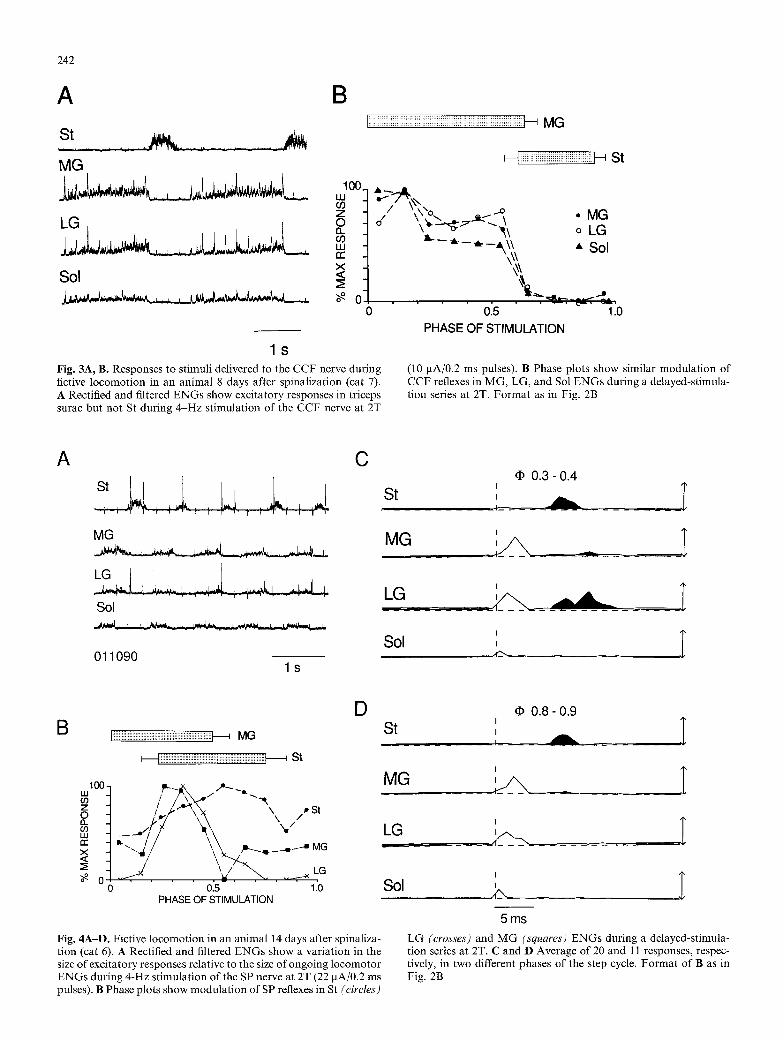
242
A �84 B
st MG
2 oo (/) z
t r
x sol .<
~iiiiiiiii~;iiiiiiiii~;~i~i~iiiiiiiiiiiii~i~i~iii~ii~ii~i~i~ MG
I~iiiiiiiiiiiiiil;i;iiiiiiiiiiiiiiiiiiiii~ St
/ / " @ 2 \ .......o~, , M G \ ' ~ - = ' ~ - ~ " ~\ o LG
o
~ - � 9 �9 Sol \,:
0.5 1.0 PHASE OF STIMULATION
l S
Fig. 3A, B. Responses to stimuli delivered to the CCF nerve during flctive locomotion in an animal 8 days after spinalization (cat 7). A Rectified and filtered ENGs show excitatory responses in triceps surae but not St during 4-Hz stimulation of the CCF nerve at 2T
(10 gA/0.2 ms pulses). B Phase plots show similar modulation of CCF reflexes in MG, LG, and Sol ENGs during a delayed-stimula- tion series at 2T. Format as in Fig. 2B
A C
st !4 i ,,,,L MG
i
Sol
011090 l s
q~ 0.3- 0.4
St , . .
' T LG _,Pf'2"b~_ -- A
' I Sol
B Iiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiii;i~ MG
~iiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiiill i St
100- w (D Z o 0- W ~C X ~s
/ r ", , s t
. j--- '~ "~/ ~ . .._._..-x LG
o o~5- ' ' 1;o PHASE OF STIMULATION
D
Fig. 4A-D. Fictive locomotion in an animal 14 days after spinaliza- tion (cat 6). A Rectified and filtered ENGs show a variation in the size of excitatory responses relative to the size of ongoing locomotor ENGs during 4-Hz stimulation of the SP nerve at 2T (22 gA/0.2 ms pulses). B Phase plots show modulat ion of SP reflexes in St (circles)
St 0 . 8 - 0 . 9
I
I i
MG
I
L G ~_ _/~-z,_ . . . . . . . . . . .
Sol I I
5 ms LG (crosses) and M G (squares) ENGs during a delayed-stimula- tion series at 2T. C and D Average of 20 and 11 responses, respec- tively, in two different phases of the step cycle. Format of B as in Fig. 2B
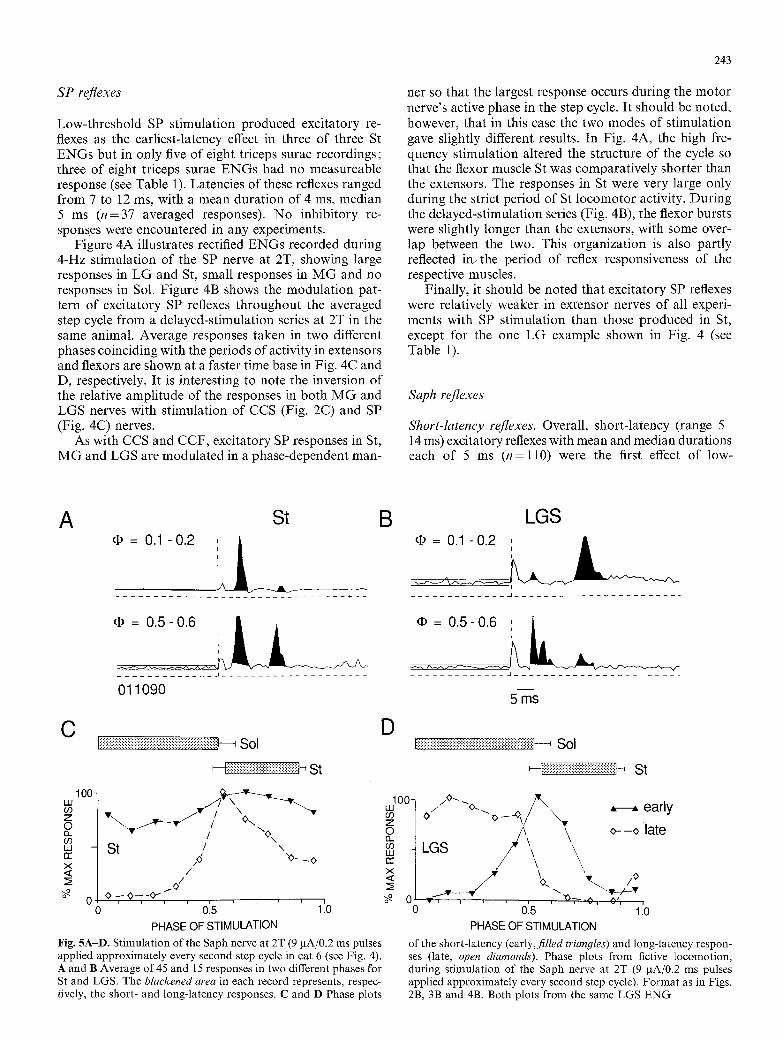
SP reflexes
Low-threshold SP stimulation produced excitatory re- flexes as the earliest-latency effect in three of three St ENGs but in only five of eight triceps surae recordings; three of eight triceps surae ENGs had no measureable response (see Table 1). Latencies of these reflexes ranged from 7 to 12 ms, with a mean duration of 4 ms, median 5 ms (n=37 averaged responses). No inhibitory re- sponses were encountered in any experiments.
Figure 4A illustrates rectified ENGs recorded during 4-Hz stimulation of the SP nerve at 2T, showing large responses in LG and St, small responses in M G and no responses in Sol. Figure 4B shows the modulation pat- tern of excitatory SP reflexes throughout the averaged step cycle from a delayed-stimulation series at 2T in the same animal. Average responses taken in two different phases coinciding with the periods of activity in extensors and flexors are shown at a faster time base in Fig. 4C and D, respectively. It is interesting to note the inversion of the relative amplitude of the responses in both MG and LGS nerves with stimulation of CCS (Fig. 2C) and SP (Fig. 4C) nerves.
As with CCS and CCF, excitatory SP responses in St, MG and LGS are modulated in a phase-dependent man-
243
net so that the largest response occurs during the motor nerve's active phase in the step cycle. It should be noted, however, that in this case the two modes of stimulation gave slightly different results. In Fig. 4A, the high fre- quency stimulation altered the structure of the cycle so that the flexor muscle St was comparatively shorter than the extensors. The responses in St were very large only during the strict period of St locomotor activity. During the delayed-stimulation series (Fig. 4B), the flexor bursts were slightly longer than the extensors, with some over- lap between the two. This organization is also partly reflected in~ the period of reflex responsiveness of the respective muscles.
Finally, it should be noted that excitatory SP reflexes were relatively weaker in extensor nerves of all experi- ments with SP stimulation than those produced in St, except for the one LG example shown in Fig. 4 (see Table 1).
Saph reflexes
Short-latency reflexes. Overall, short-latency (range 5 14 ms) excitatory reflexes with mean and median durations each of 5 ms ( n = l l 0 ) were the first effect of low-
A St
�9 = 0.1 -0.2 : k
�9 = 0.5- 0.6
. . . . . . . . . . . . . . .A . . . . . . . . . . . . . . . . . . . .
011090
C i iii ! iiiiiii!ii!iiii!i!ii iiii7iiiiiiiiiiiii i!iiiii !ii iiiiiiiiiiiiiiii!i , Sol
o o O3
oZ V ~ . . ~ I , ~ ~. ~- / o\ "' - S t / -, r r , X>._ O x "<> < /
:~ / o / 0 o.- o - -o--
o . . . . 0:5 ' ' 1:o PHASE OF STIMULATION
Fig. 5A-D. Stimulation of the Saph nerve at 2T (9 IxA/0.2 ms pulses applied approximately every second step cycle in cat 6 (see Fig. 4). A and B Average of 45 and 15 responses in two different phases for St and LGS. The blackened area in each record represents, respec- tively, the short- and long-latency responses. C and D Phase plots
B
D
100" UJ oo z O cL o3 UJ cc x <
L G S
_1 . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . .
* = i i . . . . . . . . . . . . . ~1 . . . . . . . . . . . . . . . . . . . . . .
5 ms
f~i~!~!~!~!~!:i:i:i:i:iCi~i~!~!~i~i~!~!~i~i~iC!~i~iCi~i~i~i~i~i~i~i~i~i~i~--~ Sol
~-f~i~i~!~!~!~!~!i!~!~!~!~i:!~!~i~i:!~i~i{i~ii~ St
/ ~o-.,, ~ �9 .ear ly \ , ,
LGS \ --oiate
0... ~v.~ /O
v , ~ ~ - ^ , O', o 0:5 , v 11o
PHASE OF STIMULATION
of the short-latency (early,filled triangles) and long-latency respon- ses (late, open diamonds). Phase plots from fictive locomotion, during stimulation of the Saph nerve at 2T (9 IxA/0.2 ms pulses applied approximately every second step cycle). Format as in Figs. 2B, 3B and 4B. Both plots from the same LGS ENG
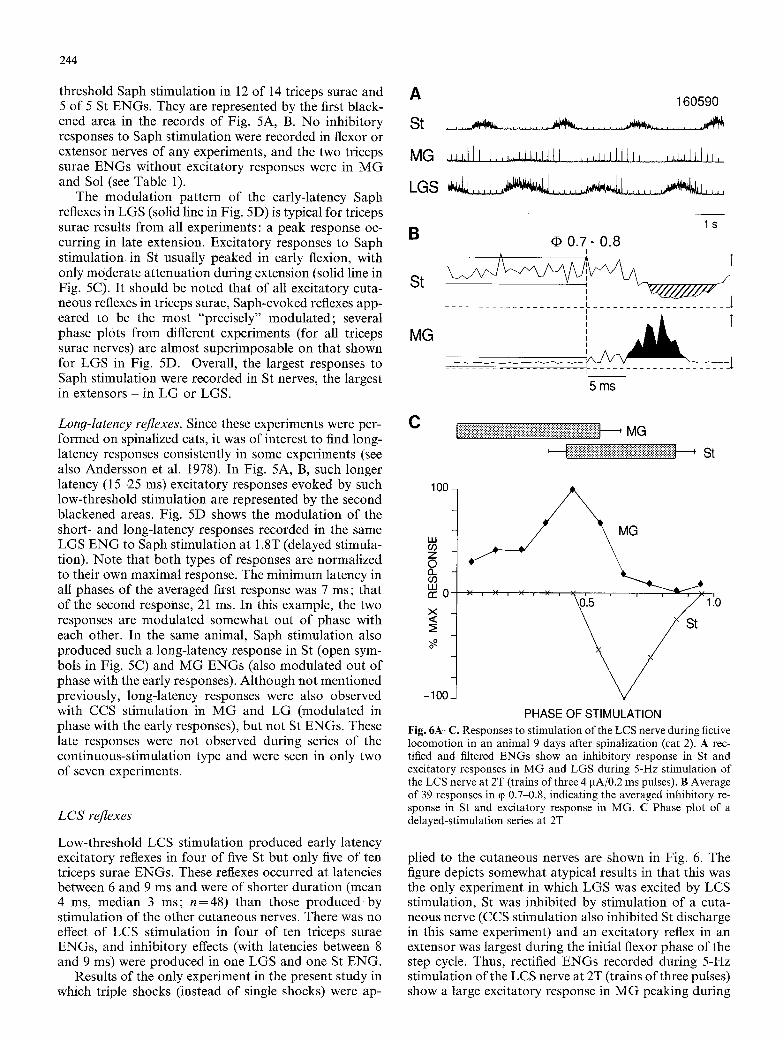
244
threshold Saph stimulation in 12 of 14 triceps surae and 5 of 5 St ENGs. They are represented by the first black- ened area in the records of Fig. 5A, B. No inhibitory responses to Saph stimulation were recorded in flexor or extensor nerves of any experiments, and the two triceps surae ENGs without excitatory responses were in M G and Sol (see Table 1).
The modulation pattern of the early-latency Saph reflexes in LGS (solid line in Fig. 5D) is typical for triceps surae results from all experiments: a peak response oc- curring in late extension. Excitatory responses to Saph st imulation in St usually peaked in early flexion, with only moderate attenuation during extension (solid line in Fig. 5C). It should be noted that of all excitatory cuta- neous reflexes in triceps surae, Saph-evoked reflexes app- eared to be the most "precisely" modulated; several phase plots from different experiments (for all triceps surae nerves) are almost superimposable on that shown for LGS in Fig. 5D. Overall, the largest responses to Saph stimulation were recorded in St nerves, the largest in extensors - in LG or LGS.
Long-latency reflexes. Since these experiments were per- formed on spinalized cats, it was of interest to find long- latency responses consistently in some experiments (see also Andersson et al. 1978). In Fig. 5A, B, such longer latency (15-25 ms) excitatory responses evoked by such low-threshold stimulation are represented by the second blackened areas. Fig. 5D shows the modulat ion of the short- and long-latency responses recorded in the same LGS E N G to Saph stimulation at 1.8T (delayed stimula- tion). Note that both types of responses are normalized to their own maximal response. The minimum latency in all phases of the averaged first response was 7 ms; that of the second response, 21 ms. In this example, the two responses are modulated somewhat out of phase with each other. In the same animal, Saph stimulation also produced such a long-latency response in St (open sym- bols in Fig. 5C) and MG ENGs (also modulated out of phase with the early responses). Although not mentioned previously, long-latency responses were also observed with CCS stimulation in M G and LG (modulated in phase with the early responses), but not St ENGs. These late responses were not observed during series of the continuous-stimulation type and were seen in only two of seven experiments.
LCS reflexes
Low-threshold LCS stimulation produced early latency excitatory reflexes in four of five St but only five of ten triceps surae ENGs. These reflexes occurred at latencies between 6 and 9 ms and were of shorter duration (mean 4 ms, median 3 ms; n--48) than those produced by stimulation of the other cutaneous nerves. There was no effect of LCS stimulation in four of ten triceps surae ENGs, and inhibitory effects (with latencies between 8 and 9 ms) were produced in one LGS and one St ENG.
Results of the only experiment in the present study in which triple shocks (instead of single shocks) were ap-
A
St
MG
LGS
160590
. . . . . . . . . . . . . . . . . . . . . .
B
St
MG
l S
d# 0.7- 0.8
J I
I
r
.....................
5 ms
C Eiiiiiiiiiiiiiiiiiii•ii!i•i•iiiiiiiiiiiiiiiii•iii•iiiiiiiiiiiiiiiiiiiiii•ii•i•i•iiiiiiiiiiiiiiiiiii•i•ii•i••• , MG
' IiilNilNi!ilNiiiNiiiiiiiiiiiiiiiiii~iiiiiiiiiiiii~i~iil , St
100
LU GO Z
2 O9 I.U co0 . . . . . . . . . . ~ ' ~ ' ' ' ' / '110 X <
-I0(
PHASE OF STIMULATION Fig. 6A-C. Responses to stimulation of the LCS nerve during fictive locomotion in an animal 9 days after spinalization (cat 2). A rec- tified and filtered ENGs show an inhibitory response in St and excitatory responses in MG and LGS during 5-Hz stimulation of the LCS nerve at 2T (trains of three 4 pA/0.2 ms pulses). B Average of 39 responses in (p 0.7-0.8, indicating the averaged inhibitory re- sponse in St and excitatory response in MG. C Phase plot of a delayed-stimulation series at 2T
plied to the cutaneous nerves are shown in Fig. 6. The figure depicts somewhat atypical results in that this was the only experiment in which LGS was excited by LCS stimulation, St was inhibited by stimulation of a cuta- neous nerve (CCS stimulation also inhibited St discharge in this same experiment) and an excitatory reflex in an extensor was largest during the initial flexor phase of the step cycle. Thus, rectified ENGs recorded during 5-Hz stimulation of the LCS nerve at 2T (trains of three pulses) show a large excitatory response in MG peaking during

245
the eary part of flexion (Fig. 6A, C), a large response in LGS during late extension (Fig. 6A) and an inhibition of St discharge during flexion (Fig. 6C). The inhibitory responses in St are indicated by the hatched area in Fig. 6B, whereas the excitatory responses in MG are illu- strated by the blackened area in Fig. 6B. No effect of stimulation was seen in a coSt recording from this animal (not illustrated).
Although the method of stimulation is slightly dif- ferent in this one experiment, in experiments overall, stimulation of the LCS nerve produced the most variable responses in flexor and extensor nerves in terms of both polarity (excitatory or inhibitory) and pattern of mod- ulation.
Discussion
The present results show that in spinal cats that are 1-3 weeks post-transection, low-threshold electrical stimula- tion of several ipsilateral hindlimb cutaneous nerves may produce short-latency excitatory reflexes in both flexor (St) and extensor (triceps surae) motor nerves, which are modulated throughout the active step cycle. With only one or two exceptions these reflexes were largest during the active phase of the motor nerve recorded (i.e. they displayed a phase dependency); and reflexes in extensors or flexors had more or less similar patterns of modulation in different animals. Occasionally, second excitatory re- sponses at longer latencies, or inhibitory responses as the first effect of stimulation were also encountered, and weak crossed excitatory responses were sometimes ob- served in coSt when its activity was recorded. Finally, variations in the amplitude of excitatory reflexes produced among MG, LG and Sol (or LGS) efferents by stimulation of particular cutaneous nerves, as sum- marized in Table 1, resembled trends previously reported for the size of EPSPs in MG, LG and Sol motoneurons (LaBella et al. 1989; LaBella and McCrea 1990).
Methodological considerations
There were three main reasons for opting to use short- survival-time "semi-chronic" spinal animals in the present study. The first reason concerns the preparation dependency of excitatory cutaneous reflexes in extensor motoneurons (our main focus), since studies in non-loco- moting preparations have clearly shown that polysynap- tic effects in motoneurons are very dependent upon ex- perimental conditions (Eccles and Lundberg 1959a). For example, effects in extensor motoneurons of acute spinal preparations tend to be characterized by a "flexor reflex- like" predominance of polysynaptic inhibition (Eccles and Lundberg t 959a), while in decerebrate preparations both polysynaptic excitation and inhibition may be sub- stantially suppressed (Eccles and Lundberg 1959b; see also Lundberg 1982). Chronic spinal animals (LaBella and McCrea, unpublished observations) as well as cord- intact cats (LaBella et al, 1989; LaBella and McCrea 1990) show a consistent presence and pattern of low-
threshold cutaneous EPSPs in triceps surae motoneu- rons. Furthermore, electrical stimulation of the dorsum of the foot of chronic spinal animals evokes phase-depen- dent cutaneous reflexes during treadmill locomotion, i.e. excitatory reflexes in flexors during swing and excitatory reflexes in extensors during stance (Forssberg et al. 1975, 1977). Thus, the chronic spinal preparation appeared to have two potential advantages over both acute spinal and decerebrate fictive preparations, although such prepara- tions were successfully used by others, albeit with less consistent responses in extensors (see Schmidt et al. 1989). In order to evoke fictive locomotion with L-dopa (Jankowska et al. 1967; Grillner and Zangger 1979), it was necessary to use a sufficiently short post-transection period (1-3 weeks) to allow transmitter release from severed, descending noradrenergic terminals (And6n et al. 1966b).
A third (retrospective) advantage of using semi- chronic spinal animals is our higher success rate of ob- taining L-dopa-induced locomotion in these than in acute spinal or decerebrate animals (unpublished observa- tions). In our hands, the anaemically decerebrated, semi- chronic spinal preparation is in much better cardiovas- cular condition and appears to maintain a high level of spinal cord excitability. While this may be due to a combination of factors, a simple explanation may be the relative lack of surgical trauma incurred by these animals at the time of the acute experiment, such as acute spi- nalization and acute mechanical section of the brain stem.
It was noted in Materials and methods that the animal with the longest post-transection survival time (3 weeks) displayed fictive locomotion without L-dopa but did re- quire continuous perineal or hindpaw stimulation to maintain a locomotor rhythm (see also Pearson and Rossignol 1991). It should be mentioned that in all other animals brief exteroceptive stimuli were only occasion- ally applied - if at all - during tests of cutaneous nerve stimulation. In addition, although both clonidine (Bar- beau et al. 1987) and L-dopa (And6n et al. 1966a) have been reported to reduce or abolish transmission in short- latency cutaneous reflex pathways, this did not appear to be a factor in the present study, in agreement with others (Schmidt et al. 1988, 1989). Not only were results com- parable between the L-dopa cats and the one non- drugged animal, but reflexes were evoked in all animals with low-threshold stimuli (1.5-3T).
Latencies of excitatory reflexes
Although sample sizes do not permit a comparison among St and triceps surae populations, minimum latencies of the excitatory responses in all motor nerves (from stim- ulus artefact to the onset of averaged responses) were 5, 5, 6, 6 and 7 ms for those produced by CCF, Saph, CCS, LCS and SP stimulation, respectively. As for the brief minimum latencies occurring with CCF and Saph stim- ulation, it is interesting to note that these are more compatible with central latencies (measured from the cord dorsum potential instead of stimulus artefact) of

246
responses recorded in motoneurons of 6-week chronic spinal animals (3.0 ms) than spinal cord-intact animals (4.8 ms; non-locomotive preparations anaesthetized with chloralose; LaBella and McCrea, unpublished observa- tions). This suggests that transmission through oligosyn- aptic cutaneous reflex pathways in semi-chronic spinal animals resembles that seen after longer post-transection periods, although differential effects upon transmission by anaesthesia and anaemic decerebration cannot be excluded (see Discussion, LaBella and McCrea 1989).
Acknowledging that measurements taken from av- eraged responses in motor nerves can only be used to obtain approximate latencies, it is clear that a few excit- atory responses in the present study fall into a longer- latency category (15-25 ms). EPSPs in St and triceps surae motoneurons evoked by low-threshold cutaneous nerve stimulation at roughly comparable latencies (ap- proximately 20 30 ms) have been previously observed (LaBella and McCrea 1990, unpublished observations); and in walking intact or decerebrate animals late excita- tion (latency 25-35 ms) has been observed in extensors with weak electrical stimulation of the paw dorsum during stance, although these responses usually occurred subsequent to early-latency inhibition (latency 10 15 ms; Duysens and Stein 1978; Prochazka et al. 1978; Forss- berg 1979; Duysens and Loeb 1980). However, late ex- citation (25 ms) subsequent to early excitatory responses was frequently seen in the St muscle of intact animals when the stimulus was applied during swing (Forssberg 1979); and strong excitatory responses were also seen in extensors of walking chronic spinal kittens (Forssberg et al. 1977), although these had latencies intermediate (10-15 ms) to the early and late excitatory periods repor- ted here for adult, fictive preparations. Considering the variety of preparation types used - including differences in the integrity of the brain and spinal cord, as well as the type and/or strength of stimulation - it is difficult to make a direct comparison of results. Nonetheless, in the present experiments long-latency excitatory responses were sometimes seen, and these occurred subsequent to, rather than exclusive of, the shorter-latency excitatory responses (And6n et al. 1966a). It is interesting to note that we did not see the late responses with low frequency repetitive stimulation (4-5 Hz) even when they were produced by stimulation of the same cutaneous nerves with delayed stimulation (see Materials and methods). The present experiments in spinalized animals rule out any supraspinal contribution to these long-latency re- flexes.
Also, the results presented by others (Schmidt et al. 1989) concerned only the initial component (first 1-2 ms) in complex intracellular recorded postsynaptic potentials (PSPs) that included large modulations in later poly- synaptic components (their Fig. 2). The present work integrates responses over 5-8 ms and is thus not directly comparable.
Excitatory cutaneous reflexes
As mentioned previously, a recent intracellular study using a non-locomoting preparation showed that the
three motor nuclei comprising triceps surae receive a differential distribution of excitatory and inhibitory syn- aptic input from at least some cutaneous afferents (LaBella et al. 1989). Thus, low-threshold CCS afferents were found to preferentially excite the MG portion of triceps surae, whereas low-threshold LCS afferents produced no effect or predominantly inhibition in all triceps surae, with some excitation which occurred main- ly in MG. A subsequent study then tested for segmental convergence on the CCS excitatory pathway to MG (LaBella and McCrea 1990), and the results support the hypothesis that a motor nucleus is preferentially excited by cutaneous nerves arising from particular skin regions, such excitatory regions being much smaller for extensor motor nuclei than for flexors (Hagbarth 1952).
Of interest here is that under very different experi- mental conditions the pattern of reflexes seen in triceps surae ENGs during fictive locomotion bears a relation- ship to these previous intracellular results, although a differential input from cutaneous nerves is suggested by variations in reflex amplitude. By comparing the size of reflexes produced in the different motor nerves (both directly and by comparing their percentage increase over ENG burst size in unstimulated cycles; see Table 1), several qualitative statements can be made: (1) CCS excitatory responses were larger in MG motor nerves than in LG, Sol or LGS (see LaBella et al. 1989, Fig. 2 and Table 1); (2) LCS excitatory responses in triceps surae were fewer in number, but four of the five seen were in MG (see LaBella et al. 1989, Fig. 2); (3) CCF excitato- ry responses were larger in MG motor nerves than in LG, Sol or LGS (consistent with the previous report of a potent excitatory pathway to MG from low-threshold CCF afferents; LaBella and McCrea 1990); (4) Saph excitatory responses were larger in LG or LGS motor nerves than in MG; and (5) the one substantial excitatory SP response (at least 100% increase in amplitude over unstimulated control) recorded in an extensor was in an LG motor nerve. These Saph and SP results are consis- tent with the preliminary finding that these two cuta- neous nerves may preferentially excite the LG portion of triceps surae (LaBella and McCrea 1990). Taken togeth- er, the results of the present and previous reports are highly suggestive of a degree of "hard wiring" between specific hindlimb cutaneous nerves and their extensor (or flexor) motor targets.
Inhibitory cutaneous reflexes
Few inhibitory responses were seen in the present study, observed only with stimulation of CCS and LCS in the St nerve of one animal and with LCS stimulation in the LGS nerve of a second animal. Inhibitory response of the ankle flexor tibialis anterior was recently reported by Yang and Stein (1990) with stimulation of the tibial or sural nerves and by Duysens et al. (1990) with stimula- tion of sural nerve in humans.
With the present experimental approach, inhibitory responses can only be observed when ongoing locomotor activity is suppressed or reduced by the stimulation.

However, as recently noted by Schmidt and colleagues, cutaneous nerve stimulation usually produces a mix of excitatory and inhibitory postsynaptic potentials (EPSPs and IPSPs, respectively) in motoneurons whereby the excitatory components occur first (see Schmidt et al. 1989). Indeed, of the five cutaneous nerves used in the present investigation, low-threshold stimulation ( < 2 T ) of LCS alone produced 1PSPs as the typical first effect in M G motoneurons of non-locomotive animals (LaBella and McCrea 1990; see above). In triceps surae motoneu- rons overall, this was also true when LCS and CCS were tested (LaBella and McCrea 1989; see above). Thus, at least for the ankle extensors, it appears that short-latency reflexes during fictive locomotion correspond to the ini- tial (i.e. excitatory) postsynaptic events recorded intra- motoneuronal ly in non-locomotive animals. Of course, the unknown admixture of depolarizing and hyper- polarizing potentials contributes to the determination of both the size and durat ion of excitatory responses in individual motoneurons , as well as the probabil i ty of increased recruitment across moto r pools.
Phase-dependent modulation of cutaneous reflexes durin 9 locomotion
Although latencies and durations of averaged cutaneous reflexes were briefly described, the emphasis of this report has been on the incidence of excitatory reflexes recorded in hindlimb extensor nerves and whether or not such reflexes are modulated during the fictive step cycle. At least with triceps surae as the extensor target, several excitatory reflex responses evoked by cutaneous afferents appear to be modulated during fictive locomotion in the semi-chronic spinal cat in particular f rom CCS and CCF nerves, which produced excitatory responses in all triceps surae nerves studied. This confirms results of earlier studies in chronic spinal kittens, which had shown that stimulation of the dorsum of the foot with silver plates fixed to the skin or by electrical stimulation through wires inserted under the skin could evoke ex- citatory responses in flexors during swing and excitatory responses in extensors during stance (Forssberg et al. 1975, 1977 ;Forssberg 1979). Since CCS and CCF nerves were identified by Hagbar th as innervating an "excitat- ory skin region" for the ankle extensors (1952), this supports the idea that reliable testing of excitatory re- flexes in extensors during different types of locomotion must take cutaneous nerve-target specificity into ac- count, as also suggested by Schmidt et al. (1989). The finding that CCS stimulation induces an excitatory res- ponse in M G is consistent with the result obtained by Abraham et al. (1985) in the intact cat. However, our finding that smaller but consistent excitatory responses in L G in fictive conditions differ f rom the observations in intact cats in which inhibitory responses are dominant .
Another aspect made clear by the present study is that the period of reflex responsiveness in a given muscle nerve does not strictly overlap with the period of loco- motor activity in that nerve (see Figs. 2 and 4). This again suggests an elaborate interneuronal control system that
247
controls the excitability of cutaneous pathways not only to different muscles but also possibly to different sub- phases of activity of a muscle within a given phase.
This conforms with the observations that muscles can be activated reflexly out of phase with their period of locomotor activity (Drew and Rossignol 1987) or that the cutaneous pathways f rom different nerves to the same muscle may be differentially controlled in various phases of the locomotor cycle (Moschovakis et al. 1991).
Given this complexity, the reduced, spinal fictive preparat ion will be impor tant yet for the consideration of many issues. For example, very little is known about the modula t ion of responses f rom cutaneous afferents innervating proximal hindlimb regions during rhythmic locomotor activity; and the technique of spatial facilita- tion during intracellular recording (Lundberg 1975) could be used in combinat ion with the methods presently employed to see if reflex pathways converging onto com- mon interneurons display phase dependency during walking. This would be an impor tant observation, since multisensory convergence upon spinal interneurons may play an important role in the ability of central control mechanisms to facilitate reflexes in one set of muscles during one phase of movement but then facilitate those produced in an antagonistic set during a different phase.
Acknowledgements. This work was supported by a grant to the Group in Neurological Sciences at Universit6 de Montr6al given by the Medical Research Council (MRC) of Canada. L. Labella was supported by an MRC post-doctoral fellowship, while A. Niechaj was supported by grants from the Fonds de la Recherche en Sant6 du Qu6bec, the Faculty of Medicine and the Centre de recherche en Sciences Neurologiques of Universit~ de Montreal. Our sincere appreciation to the following persons for their technical help: J. Provencher, S. Doucet, R. Bouchoux and D. Cyr.
References
Abraham LD, Marks WB, Loeb GE (1985) The distal hindlimb musculature of the cat. Cutaneous reflexes during locomotion. Exp Brain Res 58:594-603
And6n N-E, Jukes MGM, Lundberg A, Vyklick) L (1966a) The effect of DOPA on the spinal cord. 1. Influence on transmission from primary afferents. Acta Physiol Scand 67:373-386
And6n N-E, Jukes MGM, Lundberg A (1966b) The effect of DOPA on the spinal cord. 2. A pharmacological analysis. Acta Physiol Scand 67:387-397
Andersson O, Forssberg H, Grillner S, Lindquist M (1978) Phasic gain control of the transmission in cutaneous reflex pathways to motoneurones during "fictive" locomotion. Brain Res 149 : 503-507
Barbeau H, Julien C, Rossignol S (1987) The effects of clonidine and yohimbine on locomotion and cutaneous reflexes in the adult chronic spinal cat. Brain Res 437:83-96
Colebatch JG, Gillies JD (1979) Sural nerve effects on medial gastrocnemius motoneurones in the cat. J Physiol (Lond) 288:401~410
Drew T, Rossignol S (1987) A kinematic and electromyographic study of cutaneous reflexes evoked from the forelimb of unre- strained walking cats. J Neurophysiol 57:1160-1184
Duysens J, Loeb GE (1980) Modulation of ipsi- and contralateral reflex responses in unrestrained walking cats. J Neurophysiol 44:1024-1037

248
Duysens J, Pearson KG (1976) The role of cutaneous afferents from the distal hindlimb in the regulation of the step cycle of thalamic cats. Exp Brain Res 24:245-255
Duysens J, Stein RB (1978) Reflexes induced by nerve stimulation in walking cats with implanted cuff electrodes. Exp Brain Res 32 : 213-224
Duysens J, Trippel M, Horstmann GA, Dietz V (1990) Gating and reversal of reflexes in ankle muscles during human walking. Exp Brain Res 82:351 358
Eccles RM, Lundberg A (1959a) Synaptic actions in motoneurones by afferents which may evoke the flexion reflex. Arch Ital Biol 97:199-221
Eccles RM, Lundberg A (1959b) Supraspinal control of inter- neurones mediating spinal reflexes. J Physiol (Lond) 147:565-584
Forssberg H (1979) Stumbling corrective reaction: a phase-depen- dent compensatory reaction during locomotion. J Neurophysiol 42:936-953
Forssberg H, Grillner S, Rossignol S (1975) Phase dependent reflex reversai during walking in chronic spinal cats. Brain Res 85:103-107
Forssberg H, Grillner S, Rossignol S (1977) Phasic gain control of reflexes from the dorsum of the paw during spinal locomotion. Brain Res 132:121-139
Grillner S, Zangger P (1979) On the central generation of locomo- tion in the low spinal cat. Exp Brain Res 34:241 261
Hagbarth KE (1952) Excitatory and inhibitory skin areas for flexor and extensor motoneurones. Acta Physiol Scand 26 [Suppl 94] : 1-58
Jankowska E, Jukes MGM, Lund S, Lundberg A (1967) The effect of DOPA on the spinal cord. 5. Reciprocal organization of pathways transmitting excitatory action to alpha motoneurones of flexors and extensors. Acta Physiol Scand 70:369-388
Julien C, Rossignol S (1982) Electroneurographic recordings with polymer cuff electrodes in paralyzed cats. J Neurosci Methods 5 : 267-272
LaBella LA, McCrea DA (1990) Evidence for restricted central convergence of cutaneous afferents on an excitatory reflex path- way to medial gastrocnemius motoneurons. J Neurophysiol 64:403~412
LaBella LA, Kehler JP, McCrea DA (1989) A differential synaptic input to the motor nuclei of triceps surae from the caudal and lateral cutaneous sural nerves. J Neurophysiol 61 : 291-301
Lundberg A (1975) Control of spinal mechanisms from the brain. In: Tower DB (ed) The nervous system (The basic neurosci- ences, vol. 1 Raven Press, New York), pp 253-265
Lundberg A (1982) Inhibitory control from the brain stem of trans- mission from primary afferents to motoneurons, primary af- ferent terminals and ascending pathways. In : Sj61und B, Bj6rk- lund A (eds) Brain stem control of spinal mechanisms. Elsevier Biomedical, Amsterdam, pp 179-224
Moschovakis AK, Sholomenko GN, Burke RE (1991) Differential control of short latency cutaneous excitation in cat FDL moto- neurons during fictive locomotion. Exp Brain Res 83:489-501
Pearson KG, Rossignol S (1991) Fictive motor patterns in chronic spinal cats. J Neurophysiol 66:1874-1887
Perret C, Cabelguen J-M (1980) Main characteristics of the hind- limb locomotor cycle in the decorticate cat with special reference to bifunctional muscles. Brain Res 187 : 333-352
Pollock LJ, Davis LE (1923) Studies in decerebration I. A method of decerebration. Arch Neurol Psychiatry 10:391-398
Pratt CA, Jordan L (1987) Ia inhibitory interneurons and Renshaw cells as contributors to the spinal mechanisms of fictive locomo- tion. J Neurophysiol 57:56 71
Prochazka A, Sontag KH, Wand P (1978) Motor reactions to perturbations of gait: proprioceptive and somesthetie involve- ment. Neurosci Lett 7:35 39
Rossignol S, Lund JP, Drew T (1988) The role of sensory inputs in regulating patterns of rhythmical movements in higher verte- brates. In: Cohen A, Rossignol S, Grillner S (eds) Neural control of rhythmic movements in vertebrates. Wiley, New York, pp 201-283
Schmidt B J, Meyers DER, Fleshman JW, Tokuriki M, Burke RE (1988) Phasic modulation of short latency cutaneous excitation in flexor digitorum longus motoneurons during fictive locomo- tion. Exp Brain Res 71:568-578
Schmidt BJ, Meyers DER, Tokuriki M, Burke RE (1989) Modula- tion of short latency cutaneous excitation in flexor and extensor motoneurons during fictive locomotion in the cat. Exp Brain Res 77:57-68
Schomburg ED, Behrends HB (1978) Phasic control of the trans- mission in the excitatory and inhibitory reflex pathways from cutaneous afferents to c~-motoneurones during fictive locomo- tion in cats. Neurosci Lett 8:277-282
Wilson VJ (1963) Ipsilateral excitation of extensor motoneurones. Nature 198 : 290-291
Yang F J, Stein RB (1990) Phase-dependent reflex reversal in human leg muscles during walking. J Neurophysiol 63:1109-1117
Zomlefer MR, Provencher J, Blanchette G, Rossignol S (1984) Electromyographic study of lumbar back muscles during loco- motion in acute high decerebrate and in low spinal cats. Brain Res 290 : 249-260
Related Documents











