Provided for non-commercial research and educational use only. Not for reproduction, distribution or commercial use. This chapter was originally published in the book Progress in Brain Research, published by Elsevier, and the attached copy is provided by Elsevier for the author’s benefit and for the benefit of the author’s institution, for non-commercial research and educational use including without limitation use in instruction at your institution, sending it to specific colleagues who know you, and providing a copy to your institution’s administrator. All other uses, reproduction and distribution, including without limitation commercial reprints, selling or licensing copies or access, or posting on open internet sites, your personal or institution’s website or repository, are prohibited. For exceptions, permission may be sought for such use through Elsevier’s permissions site at: http://www.elsevier.com/locate/permissionusematerial From Andy J. Fong, Roland R. Roy, Ronaldo M. Ichiyama, Igor Lavrov, Grégoire Courtine, Yury Gerasimenko, Y.C. Tai, Joel Burdick and V. Reggie Edgerton, Recovery of control of posture and locomotion after a spinal cord injury: solutions staring us in the face. In: Graham Webb, editor: Progress in Brain Research, Vol 175, Neurotherapy: Progress in Restorative Neuroscience and Neurology, Joost Verhaagen, Elly M. Hol, Inge Huitinga, Jan Wijnholds, Arthur A. Bergen, Gerald J. Boer and Dick F. Swaab. The Netherlands: Elsevier, 2009, pp. 393–418. ISBN: 978-0-12-374511-8 © Copyright 2009 Elsevier BV. Elsevier

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Provided for non-commercial research and educational use only. Not for reproduction, distribution or commercial use.
This chapter was originally published in the book Progress in Brain Research , published by Elsevier, and the attached copy is provided by Elsevier for the author’s benefi t and for the benefi t of the author’s institution, for
non-commercial research and educational use including without limitation use in instruction at your institution, sending it to specifi c colleagues who know you, and providing a copy to your institution’s administrator.
All other uses, reproduction and distribution, including without limitation commercial reprints, selling or licensing copies or access, or posting on open internet sites, your personal or institution’s website or repository, are prohibited. For exceptions, permission may be sought for such use through Elsevier’s permissions site at:
http://www.elsevier.com/locate/permissionusematerial From Andy J. Fong, Roland R. Roy, Ronaldo M. Ichiyama, Igor Lavrov, Gr é goire Courtine, Yury
Gerasimenko, Y.C. Tai, Joel Burdick and V. Reggie Edgerton, Recovery of control of postureand locomotion after a spinal cord injury: solutions staring us in the face. In: Graham Webb, editor:
Progress in Brain Research, Vol 175, Neurotherapy: Progress in Restorative Neuroscience andNeurology, Joost Verhaagen, Elly M. Hol, Inge Huitinga, Jan Wijnholds, Arthur A. Bergen,
Gerald J. Boer and Dick F. Swaab. The Netherlands: Elsevier, 2009, pp. 393 – 418. ISBN: 978-0-12-374511-8
© Copyright 2009 Elsevier BV. Elsevier

J. Verhaagen et al. (Eds.)Progress in Brain Research, Vol. 175ISSN 0079-6123Copyright r 2009 Elsevier B.V. All rights reserved
CHAPTER 25
Recovery of control of posture and locomotion aftera spinal cord injury: solutions staring us in the face
Andy J. Fong7, Roland R. Roy1,3, Ronaldo M. Ichiyama4, Igor Lavrov1,Gregoire Courtine5, Yury Gerasimenko1,6, Y.C. Tai7,8, Joel Burdick7,8
and V. Reggie Edgerton1,2,3,�
1Department of Physiological Science, University of California, Los Angeles, Los Angeles, CA, USA2Department of Neurobiology, University of California, Los Angeles, Los Angeles, CA, USA
3Brain Research Institute, University of California, Los Angeles, Los Angeles, CA, USA4Institute of Membrane and Systems Biology, University of Leeds, Leeds, UK
5Neurobiology Department, University of Zurich, Zurich, Switzerland6Pavlov Institute of Physiology, St. Petersburg, Russia
7Division of Engineering, Bioengineering, California Institute of Technology, Pasadena, CA, USA8Division of Engineering, Mechanical Engineering Options, California Institute of Technology,
Pasadena, CA, USA
Abstract: Over the past 20 years, tremendous advances have been made in the field of spinal cord injuryresearch. Yet, consumed with individual pieces of the puzzle, we have failed as a community to grasp themagnitude of the sum of our findings. Our current knowledge should allow us to improve the lives ofpatients suffering from spinal cord injury. Advances in multiple areas have provided tools for pursuingeffective combination of strategies for recovering stepping and standing after a severe spinal cord injury.Muscle physiology research has provided insight into how to maintain functional muscle properties after aspinal cord injury.Understanding the role of the spinal networks in processing sensory information that is important for the
generation of motor functions has focused research on developing treatments that sharpen the sensitivity ofthe locomotor circuitry and that carefully manage the presentation of proprioceptive and cutaneous stimulito favor recovery. Pharmacological facilitation or inhibition of neurotransmitter systems, spinal cordstimulation, and rehabilitative motor training, which all function by modulating the physiological state ofthe spinal circuitry, have emerged as promising approaches. Early technological developments, such asrobotic training systems and high-density electrode arrays for stimulating the spinal cord, can significantlyenhance the precision and minimize the invasiveness of treatment after an injury.Strategies that seek out the complementary effects of combination treatments and that efficiently
integrate relevant technical advances in bioengineering represent an untapped potential and are likely tohave an immediate impact. Herein, we review key findings in each of these areas of research and present
�Corresponding author.Tel.: +310 825 1910; Fax: +310 267 2071;E-mail: [email protected]
DOI: 10.1016/S0079-6123(09)17526-X 393
Author's personal copy

a unified vision for moving forward. Much work remains, but we already have the capability, and moreimportantly, the responsibility, to help spinal cord injury patients now.
Keywords: spinal cord injury; rehabilitation; robotic motor training; pharmacological intervention;skeletal muscle adaptation; proprioception; epidural stimulation; locomotion
Why does spinal cord injury result in a lossof movement control?
Movements are defined by the combination ofmotor pools activated, the level at which they arerecruited, and the effectiveness with which thecorresponding muscles generate force. The dimin-ished level of movement that follows a spinal cordinjury has been attributed generally to an inabilityto activate motor pools. For most individuals witha spinal cord injury, however, this is less of a factorthan typically assumed. Three more salient issuesare linked to the impaired ability to recruit appro-priate ensembles of motor units in a manner thatyields effective movement. First, a significant por-tion of movement loss is attributable to functionalalterations of the spinal circuitry that disrupt thecoordination of the motor pools. Second, whena person with a severe, incomplete spinal cordlesion attempts to perform a movement, the levelof recruitment is insufficient for some motor pools,while actually exceeding normal levels for others.Finally, it is well known that chronic spinal cordinjuries lead to a progressive decline in musclefunction. All three of these impairments must beaddressed to realize the maximal potential forrecovering functional locomotion.
Aberrant synapse formation leadsto inappropriate muscle recruitmentand poor coordination
The loss of most, if not all, descending neuralcontrol after a spinal cord injury rapidly triggersadaptation of circuits in the brain and spinal cord.In particular, the neural circuitries responsible forposture and locomotion undergo major reorgani-zation, a process that can continue to evolve foryears (Humphrey et al., 2006). These adaptationsinclude the formation of new functional connec-tions. Yet while a large number of new synapses
are formed, there is overwhelming evidencethat many of these are abnormal connections thatmisdirect neurons to inappropriate downstreammotor networks. The development of such aber-rant connections (between the brain and thespinal cord for incomplete injuries, and withinthe spinal cord circuitry for complete injuries)generally results in poor coordination, unintendedmovements, and spasticity. For example, whenindividuals with a severe mid-thoracic spinalcord injury attempt to flex or extend the ankleon one side, often the entire lower limb will flexor extend, or the movement will occur bilater-ally (Maegele et al., 2002). Such stimulus-evokedactivation of abnormally large numbers of musclesis very common, and may correspond to the wide-spread synapsing of locomotor network neuronsonto multiple nonspecific targets (Calancie et al.,1993). This lack of specificity in synapse forma-tion leads to coactivation of circuits that are notnormally activated synchronously, which is amajor determinant of step failure.
Changes in the excitability of the spinallocomotor networks render some synapseshyperexcitable and others hypoexcitable
In the literature, the spasticity and other func-tional deficits associated with spinal cord injuryare often attributed to hyperexcitability of thespinal circuitry (Nance, 2003). It seems unlikely,however, that this explanation is sufficient toexplain the complex cadre of neural changes thataccompany spinal cord injury. Hyperexcitability isnot always detrimental. In some cases, a higher-than-normal level of recruitment can actuallyserve as a significant positive adaptation: coopera-tion between motor pathways, wherein hyperex-citation of one motor pool helps to compensatefor hypoexcitation of a related motor pool, canbe an important mechanism and strategy for
394
Author's personal copy

regaining motor function. Furthermore, althoughthere is an increase in the number of aberrantconnections after a complete spinal cord injury,some of the spinal circuits are hypoexcitable. Up-regulation of the inhibitory neurotransmittersystems, i.e., the GABAergic and glycinergicsystems, depresses the excitability of the spinalcircuitry after injury (Edgerton et al., 2001).Strategies targeted at reversing this hypoexcitablestate have been very effective. Within minutes,pharmacological treatment with antagonists ofthese inhibitory neurotransmitters dramaticallyenhances the locomotor capability of completespinal cats, taking them from being completelyunable to step, to being able to execute successfulweight-bearing stepping over a range of speeds(De Leon et al., 1999b). In addition, locomotortraining can reverse the depression of the spinalcircuits by reducing the number of glycine recep-tors and the level of GAD67 expression (Edgertonet al., 1991, 2001; Tillakaratne et al., 2002). Aftera spinal cord injury, pathways can be renderedeither hyper- or hypoexcitable. Successful reha-bilitation requires properly managing the levelof excitability, as necessary, of each of the criti-cal locomotor circuits. Excitatory treatments areneeded for some circuits, while inhibition isrequired for others.
Progressive deterioration of muscle propertiesdiminishes the ability to generate movements
The skeletal musculature is highly sensitive to thelevel of neuromuscular activity, i.e., the levels ofactivation and loading imposed on the muscles(Edgerton and Roy, 1996; Roy et al., 1991). Thechronic decrease in both the activation andloading levels of the muscles below the levelof a spinal cord injury results in atrophy, aconcomitant loss of force generating potential,and a decrease in fatigue resistance (Castro et al.,1999; Gerrits et al., 2002, 2003; Shields andDudley-Javoroski, 2006). In other words, themuscles become weak and easy to fatigue in theabsence of any countermeasure interventions.The functional consequence of these muscleadaptations is that the individual must recruit ahigher percentage of motor units from the muscles
involved in performing any given task. Regainingthe ability to stand or step most likely will beadversely affected if the muscles are allowed todeteriorate over any prolonged period. A numberof rehabilitative strategies have been implemen-ted in attempts to prevent muscle deteriorationassociated with a chronic decrease in muscle use.The most effective intervention has been the useof electrical stimulation under loaded conditions.With the appropriate use of this countermeasure,it seems feasible that the skeletal musculature canbe maintained in a state that will provide theoptimum conditions for regaining standing andstepping ability via epidural stimulation and/orpharmacological interventions (see below).
The motor deficits associated with spinal cordinjury arise from multiple deficiencies in theneuromuscular system. In this review, we willidentify how appropriate interventions involvingactivity-based therapies can be used to improveposture and locomotion by restoring muscleproperties and reinforcing appropriate synapticconnections.
Initiating, sustaining, and stoppingmovements: sources of control
‘‘Conscious’’ control
A common, but incorrect, assumption is thatcontrol of movement occurs almost exclusively inthe motor cortex. Likewise, it is a misconceptionthat most movements are controlled consciously.On the contrary, there is overwhelming evidencethat the details of most movements are performedroutinely, with little conscious or voluntary effort.Shik and Orlovsky (1976) proposed the concept of‘‘automaticity’’ in movement control, suggestingthat many movements are executed by parts ofthe brain and spinal cord that are not commonlyassociated with ‘‘voluntary’’ or ‘‘conscious’’ con-trol. For over a century, it has been known thateven tasks as complex as weight-bearing locomo-tion can be executed rather effectively in animalsafter the cerebral cortex is ablated (Grillner,1981). This evidence of subcortical control ofposture and locomotion suggests that there are
395
Author's personal copy

potential sites in the brainstem and spinal cordthat can generate complex movements in responseto general stimulating signals without requir-ing detailed, millisecond-to-millisecond consciouscontrol.
‘‘Brainstem’’ control
Shik et al. (1966) convincingly demonstrated indecerebrate cats that a region within the mesen-cephalon caudal to the pons (now commonlyreferred to as the mesencephalic locomotorregion) can be tonically electrically stimulated toinduce stepping. When either the voltage orfrequency of stimulation is increased, animalsstep faster. These authors recognized, however,that the speed of stepping also is regulatedstrongly by the speed of the treadmill belt. Moriet al. (1991) demonstrated that an awake, sittingcat can be induced to rise and to begin stepping bystimulating chronically implanted electrodesplaced in the ventral tegmental field of the caudalpons along its midline. In addition, cats can beinduced to stop stepping and to sit on theirhindquarters by stimulating the dorsal tegmentalfield. These results demonstrate that very complexmotions can be triggered with relatively non-specific stimulation parameters, but at specificsites within the brainstem.
‘‘Spinal’’ control
Multiple sites within the spinal cord can bestimulated to induce or facilitate stepping move-ments. Stepping can be initiated in cats by pro-viding relatively nonspecific tonic stimulationto a region of the spinal cord referred to asthe ‘‘locomotor strip’’ (Kazennikov et al., 1983).This strip is located just lateral to the dorsalboundary of the dorsal horn, lying at a depth ofapproximately 1–2 mm, and extending from C1caudally to approximately the L1 spinal cordsegment. The capability of the spinal cordto convert rather non-specific stimulating signalsinto functional motor activity is even moreremarkable than in the brainstem. For example,stimulation via epidural electrodes placed
anywhere between the T12 and the L6 spinalcord segments can induce locomotor-like andstanding-like movements in both cats and rats(Gerasimenko et al., 2002, 2003, 2008; Ichiyamaet al., 2005; Kazennikov et al., 1983). Never-theless, regional differentiation does exist, and itdictates how the spinal cord responds to electri-cal stimulation. For example, in rats, electrodesplaced along the midline of spinal cord segmentsL2 and S1 seem to be more effective in facilitatingstepping compared to electrodes placed at otherlevels (Ichiyama et al., 2005), whereas stimulationof the L2 and L5 spinal levels is most effectivein humans and cats, respectively (Gerasimenkoet al., 2002, 2008). In addition, stimulating atspinal cord segments L2 and S1 seem to be moreeffective in facilitating stepping compared tostimulating at either segmental level alone in therat (van den Brand et al., 2007).
The ability of epidural stimulation to generateeffective stepping is attributed frequently tothe activation of neural circuits in the spinalcord responsible for central pattern generation,i.e., circuits that generate coordinated alternatingflexor-extensor neuromotor patterns in theabsence of supraspinal or sensory modulation.Central pattern generation certainly has animportant role in locomotion, but when the injuryspares the locomotor circuitry and afferent inputs,sensory information, e.g., proprioceptive, cuta-neous, etc., can be equally, if not more, importantin shaping the recovered locomotor patterns.Effective execution of weight-bearing stepping inspinal subjects appears to be accomplished whenthe spinal locomotor circuits are modulated bystepping-associated sensory input. The importantpoint is that the stimulation parameters can berather nonspecific if the sensory input associatedwith stepping is available to provide the finetuning.
‘‘Sensory’’ control
The intact spinal cord has a remarkable ability toutilize cutaneous and proprioceptive sensoryinformation to adapt to different environmentalconditions during locomotion (Buford and Smith,1993; Forssberg, 1979). A series of experiments
396
Author's personal copy

reported over the last few years demonstrate thatthese capabilities are retained after spinal cordinjury (Cai et al., 2006; Cote and Gossard, 2004;Musienko et al., 2007; Timoszyk et al., 2002, 2005).For example, it is well known that complete, low-thoracic spinal cats can modify their steppingkinematics during the swing and stance phases toadapt to changes in treadmill speed, makingessentially the same adjustments as intact animals(De Leon et al., 1998). These complete spinal catscan modulate the excitatory levels of appropriatemotor pools during different levels of weightbearing, and they can even walk backwardswhen the treadmill is reversed (Musienko et al.,2007). Essentially the same responses have beenobserved in human subjects with complete spinalcord injuries when they are partially assisted towalk on a treadmill (Harkema et al., 1997). Theseexperiments demonstrate that it is the combina-tion of the intrinsic ability of the spinal circuitry toexecute rhythmic motor patterns with the acces-sibility to activity-specific sensory information thatallows the spinal cord to function with effectiveautomaticity and minimal or no control from thebrain. The realization that the locomotor circuitscan function independently from brain controlopens the door to new paradigms for recoveringposture and locomotion in individuals with severespinal cord injuries.
Treatment paradigms for preserving musclefunction after a spinal cord injury
Spinal cord injury leads to the degradationof muscle properties
One of the primary effects of a spinal cord injuryon the motor system is a loss of mass and functionof the muscles below the level of the injury. Theseeffects appear to be muscle type specific. Musclesthat function as extensors, i.e., those that areheavily involved in weight support and propulsivefunctions and show the highest daily activitylevels, are the most affected after the injury. Forexample, in chronic complete spinal cats, thesoleus and medial gastrocnemius muscles (primaryplantarflexors) show a greater loss in mass and
maximum force potential than the tibialis ante-rior (a primary dorsiflexor) (Roy et al., 1991). Inaddition, the predominantly slow soleus is affectedmore than the predominantly fast medial gastro-cnemius. Spinal cord injury also has a severeimpact on the muscle phenotype, i.e., there is ageneral shift toward an increase in the percentageof fibers having a faster phenotype within theaffected muscles (Talmadge, 2000). This is parti-cularly evident in muscles that normally not onlyhave a high percentage of type I (slow) fibers suchas the soleus, but also involve a shift to the fastestphenotypes in normally predominantly fast mus-cles such as the medial gastrocnemius and tibialisanterior. In effect, the muscles below a spinal cordlesion become smaller, weaker, and more fatigableafter the injury. Similar effects generally areobserved in human subjects after a spinal cordinjury: atrophy, loss of maximum force potential,slow to fast fiber type conversion, and increasedfatigability (Burnham et al., 1997; Castro et al.,1999; Gerrits et al., 1999; Shields, 1995).
Activity-based treatments helpto maintain muscle properties
A number of interventions have been attemptedto prevent the loss of muscle function associatedwith spinal cord injury. Several exercise modal-ities have been used with varying results. Forexample, we have shown that training completespinal cord transected cats to step on a tread-mill ameliorates, but does not prevent, the loss ofmass and force potential of the soleus muscle(Roy et al., 1998), but has a minimal effect onthese properties in the medial gastrocnemius andtibialis anterior muscles (Roy et al., 1999).Training the cats to support their weight (standtraining), on the other hand, had a positiveeffect in both the soleus (Roy et al., 1998) andthe medial gastrocnemius (Roy et al., 1999).A ‘‘passive’’ cycling exercise regime in spinal ratsalso had a muscle-specific effect: the total crosssectional area of all fiber types was maintainednear control levels in the soleus, but not in theextensor digitorum longus (a predominantly fastsynergist of the tibialis anterior for dorsiflexion)(Dupont-Versteegden et al., 1998). This same
397
Author's personal copy

cycling paradigm in combination with fetal spinalcord tissue implant was more effective in main-taining the mass of the soleus and plantaris(a predominantly fast plantar flexor) muscle thaneither intervention alone (Dupont-Versteegdenet al., 2000), highlighting the importance of com-binatory strategies for rehabilitation.
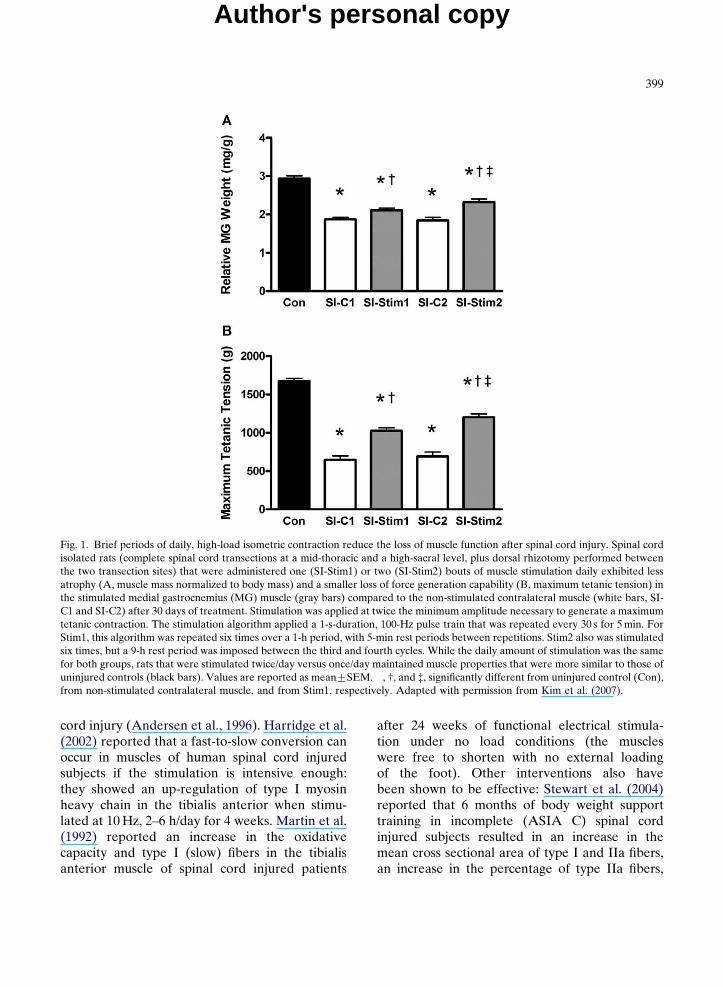
Even limited amounts of daily muscle stimula-tion can help to maintain muscle properties. Theeffectiveness of stimulation depends, in part, onhow the subjects are trained. We have used themodel of spinal cord isolation, which results inneuromuscular inactivity, to examine these issues.The advantage of the spinal cord isolation model isthat the baseline level of neuromuscular activity inthe muscle is known, i.e., the muscles are virtuallyinactive (Roy et al., 2007), and thus the effectsof imposing known patterns and/or amounts ofactivity can be determined. For example, we foundthat the same amount of activation (electricalstimulation through the lateral gastrocnemius-soleus nerve mimicking the EMG patternobserved during treadmill walking) imposed dur-ing repeated isometric contractions was moreeffective in maintaining mass and phenotype ofthe cat soleus muscle closer to normal than whenstimulating with either lengthening or shorteningcontractions (Roy et al., 2002). Using the ratmodel of spinal cord isolation, we have initiatedstudies to determine the minimum number ofcontractions required to maintain the propertiesof a muscle. As little as 1 min of brief, high-loadisometric contractions per day was sufficient tosignificantly ameliorate the loss of mass and maxi-mum tetanic tension, and the shift from slower-to-faster phenotypes in the otherwise inactive medialgastrocnemius muscle (Kim et al., 2007). Inaddition, delivering the same amount of activityduring two sessions per day was more effectivethan one session per day (Fig. 1). Furthermore, arecent study in our laboratory using the sametwo per day stimulation protocol indicates that atotal of 4 min of stimulation per day maintainedthe mass of the medial gastrocnemius at normalcontrol values. In effect, these data from animalstudies indicate that a minimal amount of high-load activity is very effective in maintainingskeletal muscle properties (Kim et al., 2008).
In humans, the most common rehabilitativestrategy used to maintain skeletal muscle massafter a spinal cord injury has been functionalelectrical stimulation, usually administered duringcycling activity. Functional electrical stimula-tion ameliorates muscle loss if the muscles areallowed to produce significant forces during thestimulation. For example, Crameri et al. (2004)highlighted the importance of the loading char-acteristics in paraplegic subjects: both fiber sizeand phenotype were maintained closer to controlvalues in the vastus lateralis (a knee extensor)using an isometric loading paradigm as opposed toa dynamic, concentric, minimally loaded para-digm. In addition, this group also showed that anisometric electrical stimulation regime couldlargely prevent the adverse effects of spinal cordinjury on the fiber size and fiber type of paralyzedhuman muscle if initiated soon after the injury(Crameri et al., 2000). Similarly, Stein et al. (1992)stimulated the tibialis anterior of spinal cordinjured subjects for 6 weeks under conditionswhere the muscle was allowed to shorten (lowload) and found an increase in fatigue resistancebut no change in tetanic force. The importance ofload in maintaining muscle properties also wasdemonstrated for the wrist extensors of tetraple-gic subjects (Hartkopp et al., 2003). A high resis-tance protocol (30 Hz, maximum load) improvedmuscle strength and fatigue resistance, whereas alow resistance protocol (15 Hz, 50% of maximumload for the same total work as the high resistanceprotocol) improved only the fatigue resistanceover a 12-week period. A potential complicationwith functional electrical stimulation, however, isthat electrically evoked isometric contractionscan cause muscle damage in long-term spinal cordinjured patients (Bickel et al., 2004). Whether ornot the muscles become more damaged ornonfunctional, however, is probably determinedby the specific parameters of the stimulationparadigm.
Functional electrical stimulation also canameliorate the adaptations in fiber phenotypeassociated with spinal cord injury. For example,long-term electrical stimulation (1 year) of thevastus lateralis can prevent the shift from slow tofast phenotypes usually associated with spinal
398
Author's personal copy
cord injury (Andersen et al., 1996). Harridge et al.(2002) reported that a fast-to-slow conversion canoccur in muscles of human spinal cord injuredsubjects if the stimulation is intensive enough:they showed an up-regulation of type I myosinheavy chain in the tibialis anterior when stimu-lated at 10 Hz, 2–6 h/day for 4 weeks. Martin et al.(1992) reported an increase in the oxidativecapacity and type I (slow) fibers in the tibialisanterior muscle of spinal cord injured patients
after 24 weeks of functional electrical stimula-tion under no load conditions (the muscleswere free to shorten with no external loadingof the foot). Other interventions also havebeen shown to be effective: Stewart et al. (2004)reported that 6 months of body weight supporttraining in incomplete (ASIA C) spinal cordinjured subjects resulted in an increase in themean cross sectional area of type I and IIa fibers,an increase in the percentage of type IIa fibers,
Fig. 1. Brief periods of daily, high-load isometric contraction reduce the loss of muscle function after spinal cord injury. Spinal cordisolated rats (complete spinal cord transections at a mid-thoracic and a high-sacral level, plus dorsal rhizotomy performed betweenthe two transection sites) that were administered one (SI-Stim1) or two (SI-Stim2) bouts of muscle stimulation daily exhibited lessatrophy (A, muscle mass normalized to body mass) and a smaller loss of force generation capability (B, maximum tetanic tension) inthe stimulated medial gastrocnemius (MG) muscle (gray bars) compared to the non-stimulated contralateral muscle (white bars, SI-C1 and SI-C2) after 30 days of treatment. Stimulation was applied at twice the minimum amplitude necessary to generate a maximumtetanic contraction. The stimulation algorithm applied a 1-s-duration, 100-Hz pulse train that was repeated every 30 s for 5 min. ForStim1, this algorithm was repeated six times over a 1-h period, with 5-min rest periods between repetitions. Stim2 also was stimulatedsix times, but a 9-h rest period was imposed between the third and fourth cycles. While the daily amount of stimulation was the samefor both groups, rats that were stimulated twice/day versus once/day maintained muscle properties that were more similar to those ofuninjured controls (black bars). Values are reported as mean7SEM. �, w, and =, significantly different from uninjured control (Con),from non-stimulated contralateral muscle, and from Stim1, respectively. Adapted with permission from Kim et al. (2007).
399
Author's personal copy

a concomitant decrease in type IIa/IIx fibers, andan increase in the oxidative capacity of the vastuslateralis muscle. There also was an improvement inambulatory capacity and fatigue resistance (timeon the treadmill). These data are particularlyintriguing and begin to address the important issueof whether the maintenance of muscle mass has apositive effect on the recovery of locomotor ability.
One of the primary deleterious effects of aspinal cord injury in humans is an increase in thefatigability of the muscles. It appears that thedecrease in fatigue resistance after an injury isprogressive: the soleus muscle of chronic paral-yzed subjects (3.772.05 years) is more fatigablethan in acute paralyzed subjects (4.671.1 weeks)(Shields, 1995). Functional electrical stimula-tion, under either loaded or unloaded conditions,has been effective in restoring or maintainingfatigue resistance. For example, Gerrits et al.(2000) reported an increase in fatigue resistancein the quadriceps muscles of motor-completespinal cord injured subjects after 6 weeks offunctional electrical stimulation cycle ergometrytraining. Subsequently, they showed that low-frequency stimulation (10 Hz) was more effec-tive than high frequency stimulation (50 Hz) inincreasing fatigue resistance while having similareffects in increasing maximum tension capability(20%) over a 12-week period (Gerrits et al.,2002). When initiated within 6 weeks of injury, aunilateral plantar flexion electrical stimulationprotocol applied under high-load conditionsresulted in the following: compared to the non-stimulated limb, the stimulated limb was lessfatigable and produced higher torques under thesame testing conditions (Shields and Dudley-Javoroski, 2006). Some possible mechanismsinvolved in regaining fatigue resistance in musclesafter a spinal cord injury include: (1) an increasein the percentage of high oxidative fibers and/orfibers containing the slow isoform of sarco(endo)-plasmic reticulum calcium ATPase (SERCA2)(Talmadge et al., 2002), (2) an improved oxidativecapacity of the muscles, e.g., increased succinatedehydrogenase activity (Gerrits et al., 2003), and(3) an increase in fiber size, thus decreasing thenumber of activated motor units required toperform a given task.
Neurotrophic factors help to maintain muscleproperties even in the absence of neuromuscularactivity
Thus far we have emphasized the role of neuro-muscular activity, i.e., the amount and pattern ofloading and activation, in maintaining the homeo-static level of skeletal muscles. It is important torealize, however, that several other factors mustbe considered. For example, in the absence ofneuromuscular activity, the presence of an intactneuromuscular connectivity has a beneficial effecton the muscle properties. After 60 days ofinactivity, the relative mass, maximum tetanictension, specific tension (tension/physiologicalcross sectional area), and fatigue resistance of therat soleus is significantly higher in spinal cordisolated (neuromuscular connectivity intact) thanin denervated (no neuromuscular connectivity)rats (see Table 1 in Roy et al., 2002). Similarly, themotoneurons associated with the affected musclesare differentially affected: even after prolongedperiods of inactivity the motoneurons in spinalcord isolated animals maintain their size andsuccinate dehydrogenase activity level near con-trol values, whereas axotomized motoneuronshave decreased succinate dehydrogenase levels(Chalmers et al., 1992; Roy et al., 2007). Thedifferences in the effects on the muscle propertiesin these two models of inactivity are most likelyrelated to the presence of activity-independentneurotrophic influences between the muscle andthe innervating motoneurons. The recent reportby Lee et al. (2007) showing that a long-termperipheral nerve graft in combination with acidicfibroblast growth factor repair (6 months) incomplete spinal transected rats was effective inpartially restoring the mass and slow phenotypecomposition of the soleus muscle is consistent witha possible beneficial role of neurotrophic (growth)factors. Spinal cord injury depresses the mRNAand/or protein levels of brain-derived neuro-trophic factor and neurotrophin-3 in the spinalcord and/or skeletal muscles, and exercise subse-quently elevates these levels to or above controlvalues (Dupont-Versteegden et al., 2004; Gomez-Pinilla et al., 2004; Ying et al., 2005). Thus, theseactivity-independent neurotrophic factors may be
400
Author's personal copy

significant contributors to the amelioration ofmuscle fiber atrophy and phenotype shifts observedwith the functional electrical stimulation and exer-cise countermeasures described above. For injuriessuch as denervation, where the endogenous supplyof neurotrophic factors is lost, exogenous adminis-tration may serve as an effective treatment formaintaining muscle function. This may be particu-larly true for cauda equina lesions where, despitemassive denervation, functional electrical stimula-tion can recover some of the lost muscle mass, evenafter prolonged periods of injury (Kern et al., 2004).
Significance of the concept of ‘‘physiologicalstate’’ of the spinal circuitry in relearning to step
Phase-dependent modulation of proprioceptiveinput during stepping
A clear example of the dynamic ability of thespinal locomotor circuitry to process and adaptto sensory information is the enhanced activa-tion of flexor motor pools in response to amechanical tripping stimulus. When an obstacleis placed in front of the paw of a spinal cat duringthe swing phase of stepping, there is enhancedflexion of the tripped limb (Forssberg, 1979). Ifthis same mechanical stimulus is applied duringthe stance phase, however, there is enhancedexcitation of the ipsilateral extensor motor pools.In other words, the same stimulus will causeopposite effects depending on the phase of thestep cycle.
The functional importance of phase-dependentmodulation of sensory input to the spinal cordhas been reinforced by recent experimentsdemonstrating a very predictable suppression orpotentiation of monosynaptic and polysynapticresponses when electrically evoked stimuli areapplied to the dorsum of the spinal cord. Theamplitudes of these responses are increasedduring the normal active bursting phases of agiven muscle and suppressed during the inter-burst intervals. In other words during the stancephase, the net effect of sensory input is poten-tiated in the extensor musculature phase, whileduring the swing phase the sensory input is
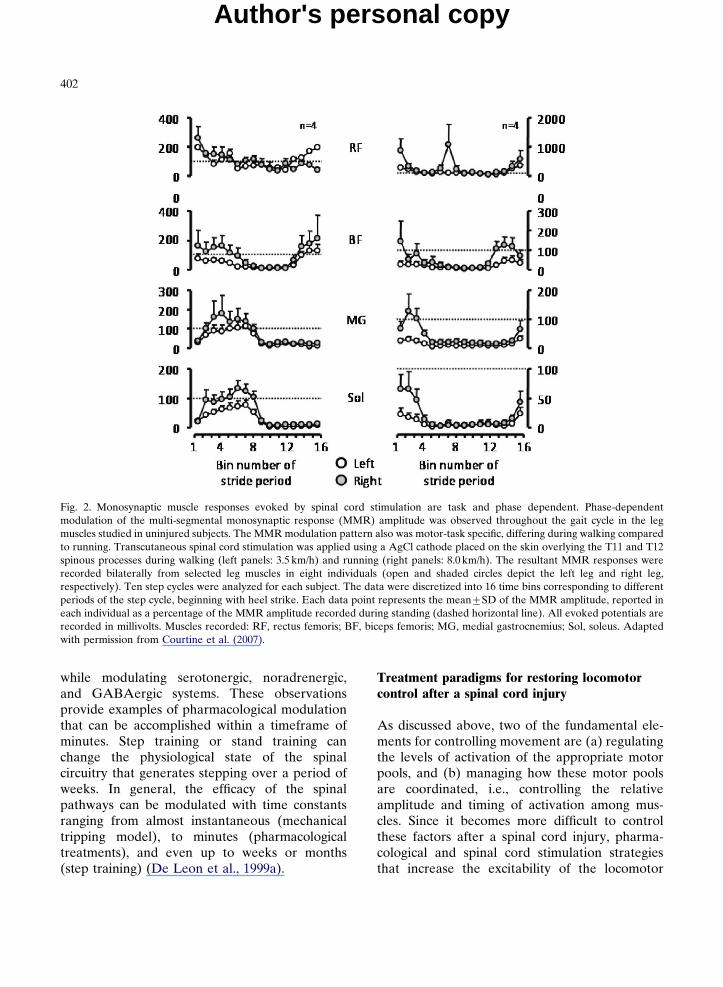
potentiated in the flexor musculature. Thistype of modulation occurs in both uninjuredand complete spinal rats, cats, and humans(Gerasimenko et al., 2007; Lavrov et al., 2006,2008). A similar modulation of responses hasbeen reported in uninjured human subjectsduring treadmill locomotion (Courtine et al.,2007) (Fig. 2). From these data, it is clear thatthe spinal circuitry processes sensory informationin a strongly cyclic, phase-dependent mannerwhether or not the spinal cord is injured.
Although the mechanisms for these dynamicresponses are unknown, the results reflect a levelof ‘‘smartness’’ and decision making capabilityof the spinal cord circuitry, and provide someinsight into how sensory information combineswith central pattern generation to generate suchremarkably effective locomotion after a spinalcord injury. For instance, these observations makeit obvious that phase-dependent processing ofproprioceptive input provides a remarkable meansof coordinating massive amounts of dynamic sen-sory information projecting to the motor poolsthat generate stepping.
More long-term changes in ‘‘state dependence’’
Another kind of state-dependent modulationof the motor output that can be achieved witha longer time constant can be mediated pharma-cologically in the spinal animals. Administrationof a number of agonists and antagonists ofeach of the neurotransmitter systems withinthe spinal circuitry can readily improve ordepress locomotor function. The direction andmagnitude of this modulation depends, of course,on dosage, but each pharmacological interven-tion will depend in large part on the func-tional state of the locomotor circuitry. Forexample, administration of a modest dose ofstrychnine, a relatively specific blocker of glycine-mediated inhibition, can have a dramatic effectin facilitating effective weight-bearing steppingwithin a matter of minutes, whereas the samedosage administered to a spinal animal thathas been trained, and that therefore can stepwell, will have minimal effect (De Leon et al.,1999b). Similar responses have been observed
401
Author's personal copy
while modulating serotonergic, noradrenergic,and GABAergic systems. These observationsprovide examples of pharmacological modulationthat can be accomplished within a timeframe ofminutes. Step training or stand training canchange the physiological state of the spinalcircuitry that generates stepping over a period ofweeks. In general, the efficacy of the spinalpathways can be modulated with time constantsranging from almost instantaneous (mechanicaltripping model), to minutes (pharmacologicaltreatments), and even up to weeks or months(step training) (De Leon et al., 1999a).
Treatment paradigms for restoring locomotorcontrol after a spinal cord injury
As discussed above, two of the fundamental ele-ments for controlling movement are (a) regulatingthe levels of activation of the appropriate motorpools, and (b) managing how these motor poolsare coordinated, i.e., controlling the relativeamplitude and timing of activation among mus-cles. Since it becomes more difficult to controlthese factors after a spinal cord injury, pharma-cological and spinal cord stimulation strategiesthat increase the excitability of the locomotor
Fig. 2. Monosynaptic muscle responses evoked by spinal cord stimulation are task and phase dependent. Phase-dependentmodulation of the multi-segmental monosynaptic response (MMR) amplitude was observed throughout the gait cycle in the legmuscles studied in uninjured subjects. The MMR modulation pattern also was motor-task specific, differing during walking comparedto running. Transcutaneous spinal cord stimulation was applied using a AgCl cathode placed on the skin overlying the T11 and T12spinous processes during walking (left panels: 3.5 km/h) and running (right panels: 8.0 km/h). The resultant MMR responses wererecorded bilaterally from selected leg muscles in eight individuals (open and shaded circles depict the left leg and right leg,respectively). Ten step cycles were analyzed for each subject. The data were discretized into 16 time bins corresponding to differentperiods of the step cycle, beginning with heel strike. Each data point represents the mean7SD of the MMR amplitude, reported ineach individual as a percentage of the MMR amplitude recorded during standing (dashed horizontal line). All evoked potentials arerecorded in millivolts. Muscles recorded: RF, rectus femoris; BF, biceps femoris; MG, medial gastrocnemius; Sol, soleus. Adaptedwith permission from Courtine et al. (2007).
402
Author's personal copy

circuits, as well as activity-based training techni-ques that reinstate functional motor pool coordina-tion, can be highly effective in helping subjectsregain the ability to step.
Pharmacological treatments
Pharmacological treatments can have an impor-tant role in restoring the chemical environmentof critical locomotor circuits after a spinal cordinjury. Many of the central nervous systemneurotransmitters, including the monoamines, aresynthesized in isolated regions of the brain (e.g.,5-HT is synthesized in the raphe nucleus) andthen transported to the spinal cord. Spinal cordinjuries that disrupt the descending flow of neuro-transmitters can severely hinder synaptic commu-nication caudal to the lesion, which translatesinto motor function loss. The damage caused bydiminished supraspinal input is aggravated by asignificant up-regulation in the inhibitory poten-tial of spinal neurons that mediate locomotion.This causes the locomotor circuitry to become lessresponsive to excitation from peripheral afferents,which normally provide important proprioceptivetriggers that can control many of the details oflocomotion.
Despite the disruption of critical neurotrans-mitter systems, the spinal locomotor circuits retainthe capability to respond to sensory-driven pre-synaptic excitation. For example, serotonergicreceptors remain functional after a spinal cordinjury and under some circumstances are even up-regulated (Fuller et al., 2005; Kim et al., 1999;Otoshi et al., 2009). Since reversing the chemicalchanges caused by spinal cord injury relates toincreasing neurotransmitter supply rather thanregenerating lost receptors, pharmacological treat-ments that supplement the spinal cord with anexogenous supply of neurotransmitter agonistscan help regulate synaptic communication andcoordinate the activation of stepping-relatedmotor pools. A number of studies have shownthat the responsiveness of the locomotor circuitryto sensory input can be readily tuned by ‘‘bathing’’the lumbosacral spinal cord with various neuro-transmitter agonists and antagonists (De Leonet al., 1999b; Edgerton et al., 1997a, b; Kiehn et al.,
2008; Rossignol and Barbeau, 1993; Rossignolet al., 1998, 2001). The effectiveness of such bluntpharmacological presentation is quite remarkable.
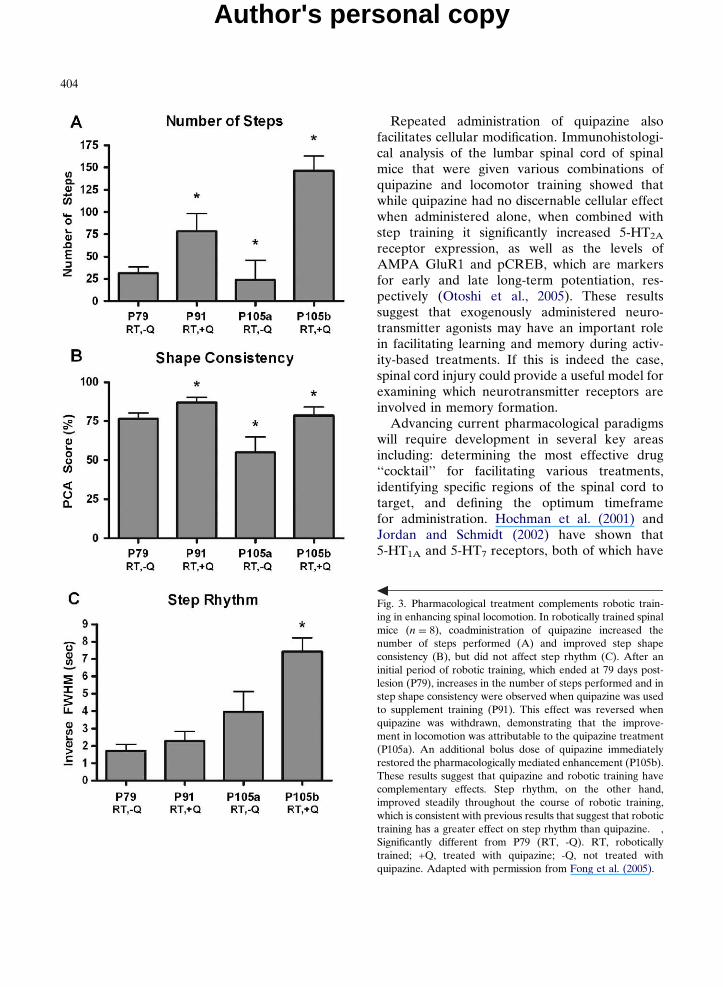
Low-dose pharmacological treatments, includ-ing monoaminergic, glycinergic, and GABAergicagonists, help to supplement the neurotransmitterenvironment of the post-injury spinal cord andthus can partially restore synaptic communica-tion (Parker, 2005; Rossignol and Barbeau, 1993).Treatments using quipazine (Antri et al., 2002;Barbeau and Rossignol, 1990; Feraboli-Lohnherret al., 1999; Fong et al., 2005; Guertin, 2004a, b),clonidine (Barbeau and Rossignol, 1991; Chauet al., 1998; Cote et al., 2003), L-DOPA (Barbeauand Rossignol, 1991; Guertin, 2004b; De Melloet al., 2004; Doyle and Roberts, 2004; McEwenand Stehouwer, 2001), strychnine (De Leonet al., 1999b; Edgerton et al., 1997a, b), and/orbicuculline (Edgerton et al., 1997a, b; Robinsonand Goldberger, 1986) have been shown to facili-tate locomotor recovery. The enhanced synaptictransmission generated by using these drugspotentiates other treatments, including spinal cordstimulation and locomotor training, by loweringthe activation threshold of the neurons associ-ated with locomotion. When quipazine, a broad-spectrum serotonin agonist, is combined withstep training, it removes the ‘‘ceiling’’ on loco-motor recovery that occurs when training isapplied alone, resulting in much higher levels ofperformance in spinal mice (Fong et al., 2005)(Fig. 3). Similarly, treatment with both quipazineand 8-OH-DPAT [8-hydroxy-2-(di-n-propylamino)-tetraline], a 5-HT1A/7 agonist, increases the step-ping performance in spinal rats above and beyondthat which can be elicited using spinal cord stimu-lation or either drug alone (Antri et al., 2005).Cells engineered to secrete serotonin also canenhance locomotion when transplanted into thelumbosacral spinal cord, presumably by makinga continuous source of the neurotransmitter avail-able during training (Feraboli-Lohnherr et al.,1997; Gimenez et al., 1998; Ribotta et al., 2000;Hains et al., 2001; Majczynski et al., 2005). Whenused to facilitate rather than directly generatelocomotion, low-dose pharmacological treatmentspotentially provide an effective solution forenhancing stepping in spinal cord injured subjects.
403
Author's personal copy
Repeated administration of quipazine alsofacilitates cellular modification. Immunohistologi-cal analysis of the lumbar spinal cord of spinalmice that were given various combinations ofquipazine and locomotor training showed thatwhile quipazine had no discernable cellular effectwhen administered alone, when combined withstep training it significantly increased 5-HT2A
receptor expression, as well as the levels ofAMPA GluR1 and pCREB, which are markersfor early and late long-term potentiation, res-pectively (Otoshi et al., 2005). These resultssuggest that exogenously administered neuro-transmitter agonists may have an important rolein facilitating learning and memory during activ-ity-based treatments. If this is indeed the case,spinal cord injury could provide a useful model forexamining which neurotransmitter receptors areinvolved in memory formation.
Advancing current pharmacological paradigmswill require development in several key areasincluding: determining the most effective drug‘‘cocktail’’ for facilitating various treatments,identifying specific regions of the spinal cord totarget, and defining the optimum timeframefor administration. Hochman et al. (2001) andJordan and Schmidt (2002) have shown that5-HT1A and 5-HT7 receptors, both of which have
Fig. 3. Pharmacological treatment complements robotic train-ing in enhancing spinal locomotion. In robotically trained spinalmice (n ¼ 8), coadministration of quipazine increased thenumber of steps performed (A) and improved step shapeconsistency (B), but did not affect step rhythm (C). After aninitial period of robotic training, which ended at 79 days post-lesion (P79), increases in the number of steps performed and instep shape consistency were observed when quipazine was usedto supplement training (P91). This effect was reversed whenquipazine was withdrawn, demonstrating that the improve-ment in locomotion was attributable to the quipazine treatment(P105a). An additional bolus dose of quipazine immediatelyrestored the pharmacologically mediated enhancement (P105b).These results suggest that quipazine and robotic training havecomplementary effects. Step rhythm, on the other hand,improved steadily throughout the course of robotic training,which is consistent with previous results that suggest that robotictraining has a greater effect on step rhythm than quipazine. �,Significantly different from P79 (RT, -Q). RT, roboticallytrained; +Q, treated with quipazine; -Q, not treated withquipazine. Adapted with permission from Fong et al. (2005).
404
Author's personal copy

been implicated in the control of stepping, arecentered in different regions of the lumbarspinal cord. Ichiyama et al. (2005) has demon-strated that activating each of these pools affectsdifferent aspects of stepping. These findingsprovide examples of the differential concentrationof neurotransmitter receptors within the spinalcord, and highlight the need for spatially specifictreatments. Furthermore, since the spinal chemi-cal environment is highly dynamic during thefirst few months after a spinal cord injury,due largely to the antagonism between recoveryprocesses and spreading secondary damage, adifferent pharmacological intervention will likelybe required at each stage of injury progression.Since synaptic communication is essential for allmotor functions, pharmacological treatments area critical area of ongoing spinal cord rehabilitationresearch.
Locomotor training
It is well established that locomotor training canenhance the recovery of stepping (Edgerton et al.,1997a, b, 2001, 2008) after a spinal cord injuryin mice (Cai et al., 2006; Fong et al., 2005), rats(Cha et al., 2007; Timoszyk et al., 2005), cats(Barbeau and Rossignol, 1987; De Leon et al.,1998, 1999a; Edgerton et al., 1991; Lovely et al.,1986, 1990) and, excitingly, even human subjects(Dietz and Harkema, 2004; Harkema et al.,1997; Van De Crommert et al., 1998; Werniget al., 1998). Engaging the spinal circuitry withsensory input associated with weight-bearingstepping is essential to activating the locomotorcircuitry so that effective locomotion can beregained. Using this information, the spinal cordlikely performs functional pruning of the manyaberrant pathways that form after a spinal cordinjury, strengthening those circuits that arerelevant to the trained stepping patterns (Ahnet al., 2006; Ichiyama et al., 2008). Traditionalmanual training involves supporting the subject ina harness over a moving treadmill while a teamof therapists/researchers repeatedly guides thelegs through a step cycle. In complete injuredindividuals remarkable levels of recovery can be
attained if training is provided persistently over aperiod of weeks to months.
Experience has helped define a set of criticalrequirements for effective step training. First, thestepping pattern used to train must have kine-matics and kinetic parameters that are stable andappropriate to the training conditions. Second, itis important for training to provide sensory stimulithat closely match normal conditions. The spinalcord circuitry is highly sensitive to proprioceptiveand cutaneous inputs: ‘‘good’’ stimuli are pro-cessed with exquisite efficiency, whereas ‘‘bad’’stimuli can lead to failure. Using the same step-ping pattern, when spinal rats are trained on anelliptical-like device that maintains continuouscontact with the hindpaw, recovery is poorer thanon a standard treadmill, where paw contact isbroken during swing (Timoszyk et al., 2003). Inthis example, even seemingly innocuous applica-tion of plantar stimulation during stance is enoughto detract from the training effect. Third, weightbearing is essential for maximizing recovery. Inaddition to helping maintain muscle properties(Roy et al., 1991; Stewart et al., 2004), subjectswho are challenged to bear increasing amounts oftheir weight are more likely to achieve betterstepping than subjects who are fully supportedduring training (Edgerton et al., 1991). Finally,although repetitive and consistent application oftraining paradigms is essential to recovery, thereshould be a small degree of variability in theparameters that are used to train to prevent loco-motor performance from becoming dependent ona single set of stimuli. A controlled amount ofvariability in training enables subjects to benefitfrom experiential learning (Cai et al., 2006). Thisimportant concept will be discussed in greaterdetail below.
Several variables affect the extent of recoverythat can be attained using step training. Recoveryis partially dependent on the severity of the injuryand the developmental stage of the subject whenit occurs. Training is particularly effective insubjects with an incomplete injury, who typicallyrecover more easily than subjects with a com-plete injury (Coleman and Geisler, 2004; Marinoet al., 1999; Waters et al., 1995). Subjects injuredat an early age exhibit better neurologic recovery
405
Author's personal copy

than those who are older at injury due to theability to develop alternative neural pathwaysduring the formative period of the central nervoussystem (Scivoletto et al., 2003). The method oftraining also affects recovery. While traditionalforms of therapist/researcher-assisted traininghave shown tremendous benefit, there are limi-tations on the extent of recovery that can beachieved using manual approaches. For example,the size disparity between human hands andsmall mouse legs makes it difficult to control thehindlimb movements of spinal mice with sufficientconsistency and precision to be effective. In astudy using spinal mice, after more than a monthof training, it was clear that a manual approachwas unable to provide efficient training. Incontrast, statistically significant improvement wasattained when training was carried out using ahigh-precision robotic system (Fong et al., 2005).
From a research perspective, an inability toreplicate training movements day-to-day, or evenstep-to-step, makes it impossible to evaluate train-ing techniques rigorously. Inconsistency and thelack of precision inherent to manual trainingwere the impetus for developing robotic trainingsystems. Diagrammed in Fig. 4, our robotic step-training system for rodents consists of two motor-driven arms (one for each leg), a weight-supportdevice, and a computer-controlled treadmill. Thefirst robotically assisted training algorithm exam-ined used the robot arms to train a single steppingpattern repetitively during alternate periods for15 min per day in mice. During the interveningperiods, robotic control was turned off to allowthe mice to step freely. Compared to the manualmethod, daily robotic training generated visibleand statistically significant improvement within2 weeks, although the quality of stepping remainedpoorer than in uninjured control mice (Fonget al., 2005). Using this technique, we observeda plateau in the level of improvement, which weinterpreted as saturation in the amount of benefitthat the spinal circuitry could extract from asingle training pattern. To overcome this, the nextset of robotically assisted training algorithmswere designed to give subjects the opportunityto experience multiple viable stepping patterns,and even to expose them to occasional failure.
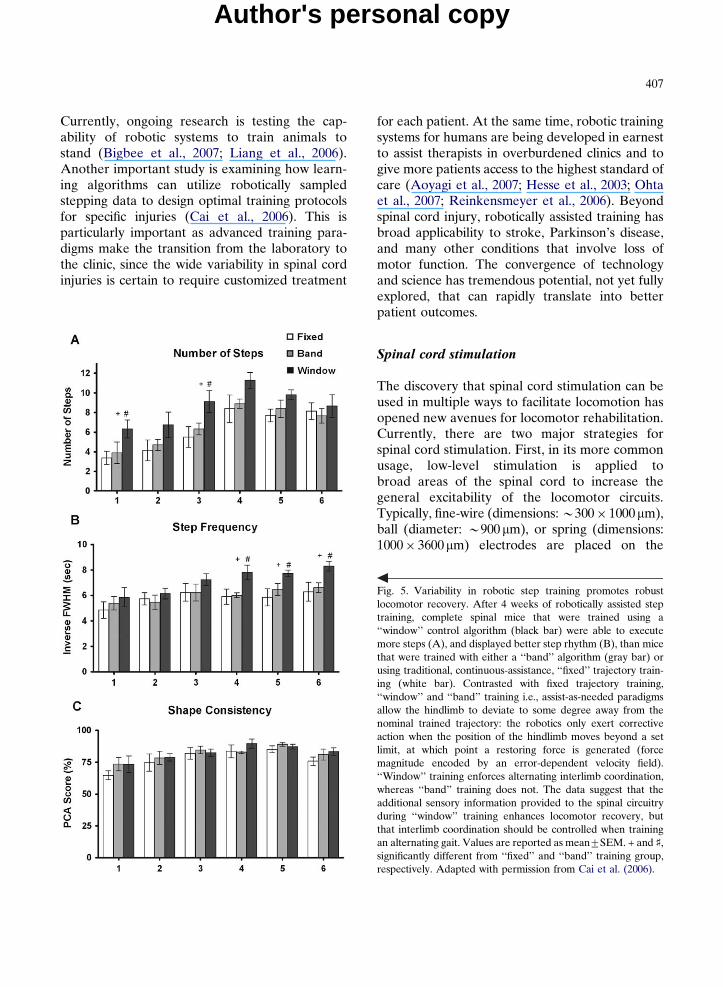
The general idea behind these ‘‘assist-as-needed’’paradigms was to provide sufficient variability toenable the subjects to learn from several good andbad stepping patterns, yet enough control toprevent catastrophic failure. The results high-light the importance of experiential learning anddemonstrate that, while it is important to enforceproper interlimb coordination, mice that areexposed to a continuum of stepping patternsrecover stepping more quickly and more robustlythan those locked into a fixed trajectory trainingpattern (Fig. 5): essentially, how you train doesmatter (Cai et al., 2006).
Technological development has provided aquantum leap in the understanding of how loco-motor training produces recovery after a spinalcord injury, and continued advancement willenable novel treatment paradigms to be con-ceived and tested (Winchester and Querry, 2006).
Fig. 4. Diagram of rodent robotic step training and evaluationsystem. The rodent robotic system consists of the followingmajor components: (A) four optical encoders, (B) four DCmotors, (C) a weight-support device, (D) two 5-bar parallelo-gram linkages, and (E) a motor-driven treadmill. When used inan active mode, the system applies step-training algorithmsthat are commanded by an external motion controller. In apassive mode, the optical encoders record the trajectories ofthe legs during free stepping. Robotics thus enables quantita-tive monitoring of both training and recovery. Adapted withpermission from Cai et al. (2005).
406
Author's personal copy
Currently, ongoing research is testing the cap-ability of robotic systems to train animals tostand (Bigbee et al., 2007; Liang et al., 2006).Another important study is examining how learn-ing algorithms can utilize robotically sampledstepping data to design optimal training protocolsfor specific injuries (Cai et al., 2006). This isparticularly important as advanced training para-digms make the transition from the laboratory tothe clinic, since the wide variability in spinal cordinjuries is certain to require customized treatment
for each patient. At the same time, robotic trainingsystems for humans are being developed in earnestto assist therapists in overburdened clinics and togive more patients access to the highest standard ofcare (Aoyagi et al., 2007; Hesse et al., 2003; Ohtaet al., 2007; Reinkensmeyer et al., 2006). Beyondspinal cord injury, robotically assisted training hasbroad applicability to stroke, Parkinson’s disease,and many other conditions that involve loss ofmotor function. The convergence of technologyand science has tremendous potential, not yet fullyexplored, that can rapidly translate into betterpatient outcomes.
Spinal cord stimulation
The discovery that spinal cord stimulation can beused in multiple ways to facilitate locomotion hasopened new avenues for locomotor rehabilitation.Currently, there are two major strategies forspinal cord stimulation. First, in its more commonusage, low-level stimulation is applied tobroad areas of the spinal cord to increase thegeneral excitability of the locomotor circuits.Typically, fine-wire (dimensions: B300� 1000mm),ball (diameter: B900mm), or spring (dimensions:1000� 3600mm) electrodes are placed on the
Fig. 5. Variability in robotic step training promotes robustlocomotor recovery. After 4 weeks of robotically assisted steptraining, complete spinal mice that were trained using a‘‘window’’ control algorithm (black bar) were able to executemore steps (A), and displayed better step rhythm (B), than micethat were trained with either a ‘‘band’’ algorithm (gray bar) orusing traditional, continuous-assistance, ‘‘fixed’’ trajectory train-ing (white bar). Contrasted with fixed trajectory training,‘‘window’’ and ‘‘band’’ training i.e., assist-as-needed paradigmsallow the hindlimb to deviate to some degree away from thenominal trained trajectory: the robotics only exert correctiveaction when the position of the hindlimb moves beyond a setlimit, at which point a restoring force is generated (forcemagnitude encoded by an error-dependent velocity field).‘‘Window’’ training enforces alternating interlimb coordination,whereas ‘‘band’’ training does not. The data suggest that theadditional sensory information provided to the spinal circuitryduring ‘‘window’’ training enhances locomotor recovery, butthat interlimb coordination should be controlled when trainingan alternating gait. Values are reported as mean7SEM. + and x,significantly different from ‘‘fixed’’ and ‘‘band’’ training group,respectively. Adapted with permission from Cai et al. (2006).
407
Author's personal copy

epidural surface of the spinal cord, and squarewave pulses (amplitude: 1–10 V or 10–200mA,duration: 100–250ms, frequency: 1–100 Hz) areapplied (Gerasimenko et al., 2003; Ichiyamaet al., 2008). Stimulation over this range of para-meters is below the threshold for direct motoractivation, but is sufficient to facilitate sensory-triggered movements. Used in this manner, theeffect of spinal cord stimulation is similar to that ofthe pharmacological treatments: by lowering theactivation threshold of locomotor neurons, spinalcord stimulation makes it easier for proprioceptiveand cutaneous signals to enable stepping. Thetype of motor output elicited by stimulation isaffected by both extrinsic and intrinsic parameters.Extrinsically, the effect of stimulation is activity-dependent: during stimulation, complete spinalcats and rats will adapt their stepping pattern,stepping forwards or backwards in accordance withtheir orientation on the treadmill (Musienko et al.,2007). Intrinsically, stimulation-evoked movementis frequency-dependent in humans, stimulation bet-ween 5 and 15 Hz preferentially induces standing,whereas stimulation between 25 and 50 Hz favorsstepping movements (Jilge et al., 2004). A secondapproach to spinal cord stimulation involves directgeneration of muscle movement by supra-thresholdstimulation of motoneurons. Rather than facili-tating locomotor circuits at the sensory or inter-neuronal levels, penetrating cylindrical electrodes(diameter: 25–30mm, height: 60–100mm) areinserted into the ventral horn at sites that are spe-cific for particular muscles (amplitude: 20–300mA,duration: 200–300ms, frequency: 1–50 Hz) (Gauntet al., 2006; Mushahwar et al., 2002, 2004). Given asufficiently large number of implanted electrodes,this approach bypasses the intrinsic circuitry andallows for external (e.g., computer-driven) controlof sequences of muscle movements. The disadvan-tages of this method are: penetrating electrodes candamage the spinal cord tissue, circumventing thelocomotor control circuitry eliminates the benefit ofintrinsic synergies that coordinate agonist andantagonist muscles, and real-time control of themany muscles necessary to generate smooth andstable locomotion is computer intensive. In general,an advantage of spinal cord stimulation over directmuscle stimulation is that it typically recruits muscle
fibers in a more normal physiological order,i.e., fatigue resistant (slow oxidative) before fastfatiguing (fast glycolytic) fibers, and thus helps tomaintain endurance (Bamford et al., 2005). Bothepidural stimulation, which facilitates the locomo-tor circuitry, and intraspinal stimulation, whichtriggers movements directly, are active areas ofresearch.
Our research in spinal cord stimulation isfocused on developing high-density epidural elec-trode arrays to extend the potential of the epiduralstimulation approach (Fig. 6). These arrays consistof platinum electrode contacts and wire lines thatare embedded in a parylene-C substrate, and arefabricated using techniques borrowed from semi-conductor and microelectromechanical systemsprocessing (Rodger et al., 2007, 2008). The highbiocompatibility of the constituent materialsmakes these arrays well suited for chronicimplantation: platinum has a long history ofbiocompatibility and meets both Tripartite andISO 10993 standards, while parylene-C is certifiedas a United States Pharmacopeia Class VI plastic.Parylene-based electrode arrays offer numerousadvantages over existing technologies. Parylenearrays allow for stable implantation: formed asthin films (B20mm thick), they are highly flexibleand conform to the spinal cord surface, resistingdisplacement. Furthermore, the close fit to thespinal cord promotes encapsulating connectivetissue growth, which further secures the arrayand effectively precludes movement. Postimplan-tation migration is the most common causeof clinical electrode failure (Barolat, 2000;LeDoux and Langford, 1993; North et al., 2005;Renard and North, 2006), in one study forcing 23%of implanted patients to have corrective, follow-up operations (Andersen, 1997). Thus, thin-filmparylene arrays provide a critical advancement.Microfabrication processes enable the design ofarrays that have novel electrode configurationsthat can be used to test advanced stimulationalgorithms. Using multilayer fabrication techni-ques, it is now technically possible to build arrayswith densities up to 1024 electrodes in a 5 mm� 6mm area (although, currently, practical appli-cation of such high densities is limited by connec-tor and stimulator technologies). Additionally,
408
Author's personal copy

electroplating can be used to increase the surfacearea of each electrode by around 40-fold, whichprovides two important advantages: it enableshigher levels of charge transfer from small electro-des, and it expands the current range over whichcharge can be transferred capacitively, reducing theoccurrence of tissue and electrode damage asso-ciated with high current, Faradaic charge transfer(Merrill et al., 2005).
Our electrode arrays provide several keybenefits over existing technologies. Access to alarge number of small electrodes makes possibleselective, high-precision stimulation of focalregions of the spinal cord. Electrode arrays areenabling the identification of specific regions ofthe spinal cord that are responsible for differentcomponents and phases of the step cycle. As ourknowledge of the somatotopic organization of thelocomotor circuitry increases, the selectivity pro-vided by electrode arrays will make it possible totarget stimulation appropriately to address the
specific deficiencies of individual injuries. Whenstimulation is targeted directly at the tissue ofinterest, less current is needed to generate adesired effect than with bulk stimulation methods.Moreover, by confining the stimulating currentto the regions that require it, high-density elec-trode arrays eliminate unnecessary stimulation ofneural tissue, which reduces the potential for long-term damage due to repeated pulsed electricalstimulation. High-density electrode arrays arethus making spinal cord stimulation more effectiveand safer. Finally, the electrode arrays can beused as a diagnostic tool to measure the proper-ties of spinal cord evoked potentials to assess thelocomotor circuitry at different stages of injuryand recovery.
While it is remarkable that relatively non-specific stimulation can promote stable, state-dependent treadmill locomotion in the absenceof supraspinal control, this represents just thebeginning of what can be achieved using spinalcord stimulation. Since the effect of spinal cordstimulation can change dramatically when theelectrode site is moved by as little as 200–300 mm(Kazennikov et al., 1983), targeted stimulationapproaches can be leveraged to control differentcomponents of stepping. In spinal rats, stimula-tion of the L2 spinal segment induces a generalenhancement of the locomotor rhythm, whilestimulation of the S1 segment activates the exten-sor muscles during stance (van den Brand et al.,2007). Stimulation applied to each of the spinalsegments between T12 and L6 produces differentlocomotor effects (Ichiyama et al., 2005). Theeffects of stimulation also vary in the medial-lateral direction, a finding that provides rationalefor pursuing 2-D array designs. As the spatialresolution of the electrode arrays continues toimprove, spinal cord stimulation will evolve fromproviding nonspecific excitation of the spinalcircuits to fine tuning very specific aspects of thelocomotor pattern. What is most exciting aboutspinal cord stimulation is that it can be usedeffectively at both of these levels.
Future stimulation approaches will examinethe implementation of biologically inspired stimu-lation patterns, as well as patterns that involvesimultaneous or sequential stimulation of multiple
Fig. 6. Photographs of spinal cord electrode arrays. (Top)Photograph of a spinal cord electrode array and 36-pin headconnector juxtaposed against a small coin for size comparison.(Bottom) Close-up photograph of the 18-electrode contacts ofa 3� 6 electrode array with physiologically determinedrostrocaudal inter-electrode spacing.
409
Author's personal copy

electrode sites. Other applications for multielec-trode spinal cord stimulation include manage-ment of chronic pain, stroke, and other conditionsinvolving motor function loss. The continueddevelopment of electrode array technology isproviding unparalleled access to the interneuronalcircuitry, and may serve as the best technique forfine-tuning gross motor behaviors after a spinalcord injury.
Integrating neuroengineering and biologicalconcepts to regain posture and locomotion
Based on the successful recovery that we haveattained using muscle stimulation, spinal cordstimulation, pharmacological interventions, andactivity-based training, the potential for enhan-cing locomotor recovery by aggressively pursuingcomplementary and synergistic strategies is clearand represents a logical direction for transla-ting some of the basic biological concepts to theclinical setting. While it cannot always be assumedthat multiple interventions will be complementary(Maier et al., 2009) with careful considerationof their interactive effects, multi-interventionapproaches truly are the obvious solutions thatare staring us in the face.
We already have observed significant positiveinteraction when multiple modes of treatment arecombined (Fig. 7). Optimal recovery of locomo-tion requires two important factors: the damagedspinal cord must be provided with adequateinformation that it can use to relearn to step, but,before that, it must be prepared to receive thatinformation. This explains why the recovery oflocomotion using robotically assisted training,which provides information on functional steppingpatterns, is significantly enhanced by coadminis-tration of pharmacological agonists that improvesynaptic signaling. In mice, e.g., while robotictraining restores gross stepping function, pharma-cological modulation with quipazine furtherimproves locomotion by facilitating the recoveryof movements that are difficult to access withtraining alone, e.g., activation of the distalextensor muscles during weight-bearing stance(Fong et al., 2005). We also have observed
substantial recovery in rats from a combinationof locomotor training, two serotonergic drugs, andmultiple-site epidural stimulation, and haveshown that selective combinations of these treat-ments lead to very different locomotor effects(van den Brand et al., 2007). The next step is tooptimize the combination treatment parametersto maximize the synergies between the constituentinterventions. All evidence suggests that engagingcomplementary approaches may result in thegreatest functional gains.
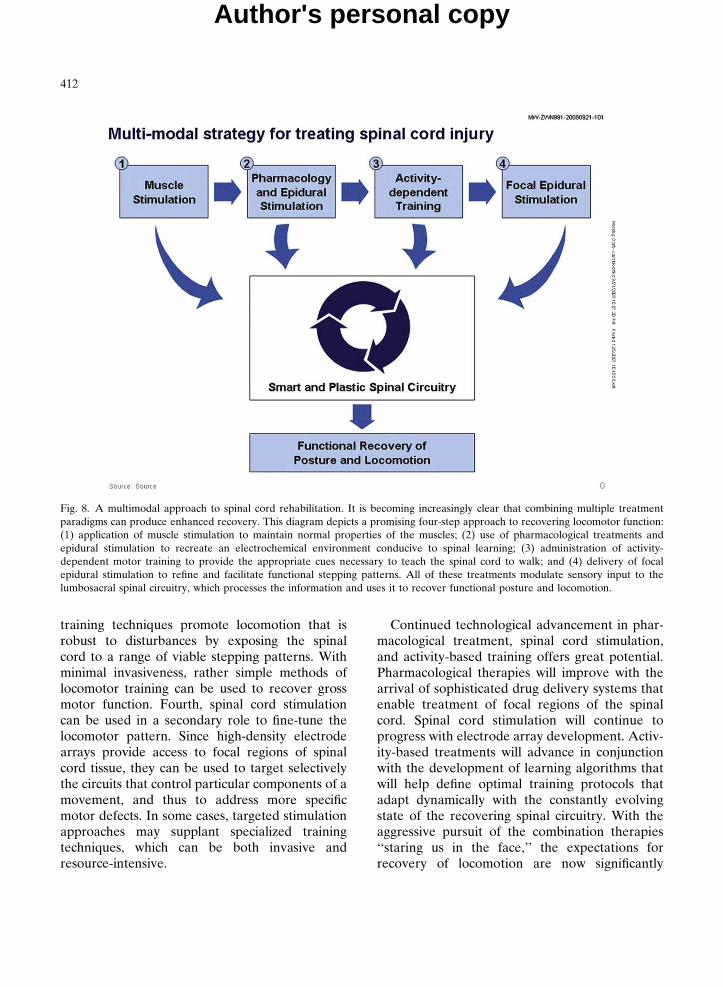
Combinations of paradigms can be effectivewhen each component treatment focuses on repai-ring a different aspect of motor function loss.Figure 8 depicts the recovery of stepping afterspinal cord injury as a multicomponent process.Although this diagram to some degree over-simplifies the complexity of the underlying mecha-nisms, it provides a didactic representation ofhow the different treatments interact to promotelocomotor recovery. First, muscle stimulationinitiated early after injury helps to maintain muscleproperties at a functional level. Insufficient muscletone and/or lack of a normal complement offatigue-resistant fibers make it more difficult forsubsequent treatments to generate appropriatemovements, increase the likelihood of injury,and may result in poor endurance. Second, bothpharmacological facilitation and spinal cord stimu-lation can be used to increase the generalexcitability of the spinal circuits and to strengthenthe efficacy of synaptic transmission. Neither low-dose drug treatment nor subthreshold spinal cordstimulation can generate movement independentlyor produce long-lasting recovery, but they cansufficiently lower the activation threshold ofrelevant sensory neurons and interneurons toenable even small amounts of stepping-associatedstimuli to trigger and sustain locomotion. Third,activity-dependent training helps to reinforce kine-matically appropriate stepping patterns. Repeti-tion of these facilitated movements improvesmuscle recruitment and coordination, and enablesthe spinal cord to learn to perform new tasks. Overtime, the persistent activation of specific spinalpathways results in changes that strengthen thosesynaptic connections and provide the structuralbasis for motor learning. The most effective
410
Author's personal copy
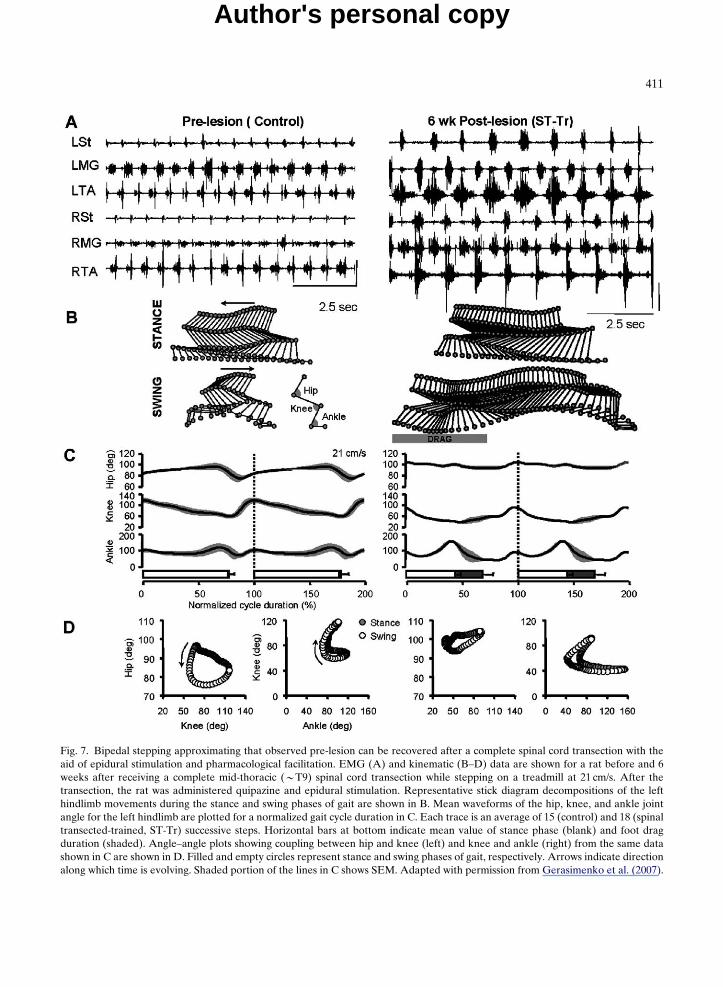
Fig. 7. Bipedal stepping approximating that observed pre-lesion can be recovered after a complete spinal cord transection with theaid of epidural stimulation and pharmacological facilitation. EMG (A) and kinematic (B–D) data are shown for a rat before and 6weeks after receiving a complete mid-thoracic (BT9) spinal cord transection while stepping on a treadmill at 21 cm/s. After thetransection, the rat was administered quipazine and epidural stimulation. Representative stick diagram decompositions of the lefthindlimb movements during the stance and swing phases of gait are shown in B. Mean waveforms of the hip, knee, and ankle jointangle for the left hindlimb are plotted for a normalized gait cycle duration in C. Each trace is an average of 15 (control) and 18 (spinaltransected-trained, ST-Tr) successive steps. Horizontal bars at bottom indicate mean value of stance phase (blank) and foot dragduration (shaded). Angle–angle plots showing coupling between hip and knee (left) and knee and ankle (right) from the same datashown in C are shown in D. Filled and empty circles represent stance and swing phases of gait, respectively. Arrows indicate directionalong which time is evolving. Shaded portion of the lines in C shows SEM. Adapted with permission from Gerasimenko et al. (2007).
411
Author's personal copy
training techniques promote locomotion that isrobust to disturbances by exposing the spinalcord to a range of viable stepping patterns. Withminimal invasiveness, rather simple methods oflocomotor training can be used to recover grossmotor function. Fourth, spinal cord stimulationcan be used in a secondary role to fine-tune thelocomotor pattern. Since high-density electrodearrays provide access to focal regions of spinalcord tissue, they can be used to target selectivelythe circuits that control particular components of amovement, and thus to address more specificmotor defects. In some cases, targeted stimulationapproaches may supplant specialized trainingtechniques, which can be both invasive andresource-intensive.
Continued technological advancement in phar-macological treatment, spinal cord stimulation,and activity-based training offers great potential.Pharmacological therapies will improve with thearrival of sophisticated drug delivery systems thatenable treatment of focal regions of the spinalcord. Spinal cord stimulation will continue toprogress with electrode array development. Activ-ity-based treatments will advance in conjunctionwith the development of learning algorithms thatwill help define optimal training protocols thatadapt dynamically with the constantly evolvingstate of the recovering spinal circuitry. With theaggressive pursuit of the combination therapies‘‘staring us in the face,’’ the expectations forrecovery of locomotion are now significantly
Fig. 8. A multimodal approach to spinal cord rehabilitation. It is becoming increasingly clear that combining multiple treatmentparadigms can produce enhanced recovery. This diagram depicts a promising four-step approach to recovering locomotor function:(1) application of muscle stimulation to maintain normal properties of the muscles; (2) use of pharmacological treatments andepidural stimulation to recreate an electrochemical environment conducive to spinal learning; (3) administration of activity-dependent motor training to provide the appropriate cues necessary to teach the spinal cord to walk; and (4) delivery of focalepidural stimulation to refine and facilitate functional stepping patterns. All of these treatments modulate sensory input to thelumbosacral spinal circuitry, which processes the information and uses it to recover functional posture and locomotion.
412
Author's personal copy

higher for individuals with spinal cord injury, theirfamily, friends, therapists, and physicians.
Acknowledgments
The work presented in this paper was supportedby the National Institutes of Health GrantsNS16333 and NS42291, the Russian Foundationfor Basic Research — US Civilian Research andDevelopment Foundation Grant 07-04-91106, theChristopher and Diana Reeve Paralysis Founda-tion, and the Roman Reed Spinal Cord InjuryResearch Fund of California.
References
Ahn, S. N., Guu, J. J., Tobin, A. J., Edgerton, V. R., &Tillakaratne, N. J. (2006). Use of c-fos to identify activity-dependent spinal neurons after stepping in intact adult rats.Spinal Cord, 44, 547–559.
Andersen, C. (1997). Complications in spinal cord stimulationfor treatment of angina pectoris. Differences in unipolar andmultipolar percutaneous inserted electrodes. Acta Cardiolo-gica, 52, 325–333.
Andersen, J. L., Mohr, T., Biering-Sorensen, F., Galbo, H., &Kjaer, M. (1996). Myosin heavy chain isoform transforma-tion in single fibres from m. vastus lateralis in spinal cordinjured individuals: effects of long-term functional electricalstimulation (FES). Pflugers Archive, 431, 513–518.
Antri, M., Barthe, J. Y., Mouffle, C., & Orsal, D. (2005). Long-lasting recovery of locomotor function in chronic spinal ratfollowing chronic combined pharmacological stimulation ofserotonergic receptors with 8-OHDPAT and quipazine.Neuroscience Letters, 384, 162–167.
Antri, M., Orsal, D., & Barthe, J. Y. (2002). Locomotorrecovery in the chronic spinal rat: effects of long-termtreatment with a 5-HT2 agonist. The European Journal ofNeurosciences, 16, 467–476.
Aoyagi, D., Ichinose, W. E., Harkema, S. J., Reinkensmeyer,D. J., & Bobrow, J. E. (2007). A robot and control algorithmthat can synchronously assist in naturalistic motion duringbody-weight-supported gait training following neurologicinjury. IEEE Transactions on Neural Systems and Rehabi-litation Engineering, 15, 387–400.
Bamford, J. A., Putman, C. T., & Mushahwar, V. K. (2005).Intraspinal microstimulation preferentially recruits fatigue-resistant muscle fibres and generates gradual force in rat.Journal of Physiology, 569, 873–884.
Barbeau, H., & Rossignol, S. (1987). Recovery of locomotionafter chronic spinalization in the adult cat. Brain Research,412, 84–95.
Barbeau, H., & Rossignol, S. (1990). The effects of serotonergicdrugs on the locomotor pattern and on cutaneous reflexes ofthe adult chronic spinal cat. Brain Research, 514, 55–67.
Barbeau, H., & Rossignol, S. (1991). Initiation and modulationof the locomotor pattern in the adult chronic spinal cat bynoradrenergic, serotonergic and dopaminergic drugs. BrainResearch, 546, 250–260.
Barolat, G. (2000). Spinal cord stimulation for chronic painmanagement. Archives of Medical Research, 31, 258–262.
Bickel, C. S., Slade, J. M., & Dudley, G. A. (2004). Long-termspinal cord injury increases susceptibility to isometriccontraction-induced muscle injury. European Journal ofApplied Physiology, 91, 308–313.
Bigbee, A. J., Crown, E. D., Ferguson, A. R., Roy, R. R.,Tillakaratne, N. J., Grau, J. W., et al. (2007). Two chronicmotor training paradigms differentially influence acuteinstrumental learning in spinally transected rats. BehaviouralBrain Research, 180, 95–101.
Buford, J. A., & Smith, J. L. (1993). Adaptive control forbackward quadrupedal walking. III. Stumbling correctivereactions and cutaneous reflex sensitivity. Journal ofNeurophysiology, 70, 1102–1114.
Burnham, R., Martin, T., Stein, R., Bell, G., Maclean, I., &Steadward, R. (1997). Skeletal muscle fibre type transforma-tion following spinal cord injury. Spinal Cord, 35, 86–91.
Cai, L. L., Fong, A. J., Otoshi, C. K., Liang, Y., Burdick, J. W.,Roy, R. R., et al. (2006). Implications of assist-as-neededrobotic step training after a complete spinal cord injury onintrinsic strategies of motor learning. The Journal ofNeuroscience, 26, 10564–10568.
Cai, L. L., Fong, A. J., Otoshi, C. K., Liang, Y. Q., Cham, J. G.,Zhong, H., et al. (2005). Effects of consistency vs. variabilityin robotically controlled training of stepping in adult spinalmice. Proceedings of International Conference on Rehabilita-tion Robotics, 575–579.
Calancie, B., Broton, J. G., Klose, K. J., Traad, M., Difini, J., &Ayyar, D. R. (1993). Evidence that alterations in presynapticinhibition contribute to segmental hypo- and hyperexcit-ability after spinal cord injury in man. Electroencephalogra-phy and Clinical Neurophysiology, 89, 177–186.
Castro, M. J., Apple, D. F., Jr., Staron, R. S., Campos, G. E., &Dudley, G. A. (1999). Influence of complete spinal cordinjury on skeletal muscle within 6 mo of injury. Journal ofApplied Physiology, 86, 350–358.
Cha, J., Heng, C., Reinkensmeyer, D. J., Roy, R. R.,Edgerton, V. R., & De Leon, R. D. (2007). Locomotorability in spinal rats is dependent on the amount of activityimposed on the hindlimbs during treadmill training. Journalof Neurotrauma, 24, 1000–1012.
Chalmers, G. R., Roy, R. R., & Edgerton, V. R. (1992).Adaptability of the oxidative capacity of motoneurons. BrainResearch, 570, 1–10.
Chau, C., Barbeau, H., & Rossignol, S. (1998). Earlylocomotor training with clonidine in spinal cats. Journal ofNeurophysiology, 79, 392–409.
Coleman, W. P., & Geisler, F. H. (2004). Injury severity asprimary predictor of outcome in acute spinal cord injury:
413
Author's personal copy

retrospective results from a large multicenter clinical trial.The Spine Journal, 4, 373–378.
Cote, M. P., & Gossard, J. P. (2004). Step training-dependentplasticity in spinal cutaneous pathways. The Journal ofNeuroscience, 24, 11317–11327.
Cote, M. P., Menard, A., & Gossard, J. P. (2003). Spinal catson the treadmill: changes in load pathways. The Journal ofNeuroscience, 23, 2789–2796.
Courtine, G., Harkema, S. J., Dy, C. J., Gerasimenko, Y. P., &Dyhre-Poulsen, P. (2007). Modulation of multisegmentalmonosynaptic responses in a variety of leg muscles duringwalking and running in humans. Journal of Physiology, 582,1125–1139.
Crameri, R. M., Cooper, P., Sinclair, P. J., Bryant, G., &Weston, A. (2004). Effect of load during electrical stimula-tion training in spinal cord injury. Muscle & Nerve, 29,104–111.
Crameri, R. M., Weston, A. R., Rutkowski, S., Middleton, J. W.,Davis, G. M., & Sutton, J. R. (2000). Effects of electricalstimulation leg training during the acute phase of spinal cordinjury: a pilot study. European Journal of Applied Physiology,83, 409–415.
De Leon, R. D., Hodgson, J. A., Roy, R. R., & Edgerton, V. R.(1998). Locomotor capacity attributable to step trainingversus spontaneous recovery after spinalization in adult cats.Journal of Neurophysiology, 79, 1329–1340.
De Leon, R. D., Hodgson, J. A., Roy, R. R., & Edgerton, V. R.(1999a). Retention of hindlimb stepping ability in adultspinal cats after the cessation of step training. Journal ofNeurophysiology, 81, 85–94.
De Leon, R. D., Tamaki, H., Hodgson, J. A., Roy, R. R., &Edgerton, V. R. (1999b). Hindlimb locomotor and post-ural training modulates glycinergic inhibition in the spinalcord of the adult spinal cat. Journal of Neurophysiology, 82,359–369.
De Mello, M. T., Esteves, A. M., & Tufik, S. (2004). Com-parison between dopaminergic agents and physical exerciseas treatment for periodic limb movements in patients withspinal cord injury. Spinal Cord, 42, 218–221.
Dietz, V., & Harkema, S. J. (2004). Locomotor activity inspinal cord-injured persons. Journal of Applied Physiology,96, 1954–1960.
Doyle, L. M., & Roberts, B. L. (2004). Functional recovery andaxonal growth following spinal cord transection is acceler-ated by sustained L-DOPA administration. European Jour-nal of Neuroscience, 20, 2008–2014.
Dupont-Versteegden, E. E., Houle, J. D., Dennis, R. A.,Zhang, J., Knox, M., Wagoner, G., et al. (2004). Exercise-induced gene expression in soleus muscle is dependenton time after spinal cord injury in rats. Muscle & Nerve, 29,73–81.
Dupont-Versteegden, E. E., Houle, J. D., Gurley, C. M., &Peterson, C. A. (1998). Early changes in muscle fiber size andgene expression in response to spinal cord transection andexercise. American Journal of Physiology, 275, C1124–C1133.
Dupont-Versteegden, E. E., Murphy, R. J., Houle, J. D.,Gurley, C. M., & Peterson, C. A. (2000). Mechanisms
leading to restoration of muscle size with exercise andtransplantation after spinal cord injury. American Journal ofPhysiology. Cell Physiology, 279, C1677–C1684.
Edgerton, V. R., Courtine, G., Gerasimenko, Y. P., Lavrov, I.,Ichiyama, R. M., Fong, A. J., et al. (2008). Training loco-motor networks. Brain Research. Brain Research Reviews,57, 241–254.
Edgerton, V. R., De Guzman, C. P., Gregor, R. J., Roy, R. R.,Hodgson, J. A., & Lovely, R. G. (1991). Trainability ofthe spinal cord to generate hindlimb stepping patternsin adult spinalized cats. In M. Shimamura, S. Grillner, &V. R. Edgerton (Eds.), Neurobiological basis of humanlocomotion. Tokyo: Japan Scientific Societies Press.
Edgerton, V. R., De Leon, R. D., Tillakaratne, N.,Recktenwald, M. R., Hodgson, J. A., & Roy, R. R.(1997a). Use-dependent plasticity in spinal stepping andstanding. Advances in Neurology, 72, 233–247.
Edgerton, V. R., Leon, R. D., Harkema, S. J., Hodgson, J. A.,London, N., Reinkensmeyer, D. J., et al. (2001). Retrainingthe injured spinal cord. Journal of Physiology, 533, 15–22.
Edgerton, V. R., & Roy, R. R. (1996). Neuromuscular adap-tations for actual and simulated spaceflight. In M. J. Fregly &C. M. Blatteis (Eds.), Environmental physiology. Bethesda,MD: American Physiological Society.
Edgerton, V. R., Roy, R. R., De Leon, R., Tillakaratne, N., &Hodgson, J. A. (1997b). Does motor learning occur in thespinal cord? Neuroscientist, 3, 294.
Feraboli-Lohnherr, D., Barthe, J. Y., & Orsal, D. (1999).Serotonin-induced activation of the network for loco-motion in adult spinal rats. Journal of NeuroscienceResearch, 55, 87–98.
Feraboli-Lohnherr, D., Orsal, D., Yakovleff, A., Gimenez, Y.,Ribotta, M., & Privat, A. (1997). Recovery of locomotoractivity in the adult chronic spinal rat after sublesional trans-plantation of embryonic nervous cells: specific role of seroto-nergic neurons. Experimental Brain Research, 113, 443–454.
Fong, A. J., Cai, L. L., Otoshi, C. K., Reinkensmeyer, D. J.,Burdick, J. W., Roy, R. R., et al. (2005). Spinal cord-transected mice learn to step in response to quipazinetreatment and robotic training. The Journal of Neuroscience,25, 11738–11747.
Forssberg, H. (1979). Stumbling corrective reaction: a phase-dependent compensatory reaction during locomotion. Jour-nal of Neurophysiology, 42, 936–953.
Fuller, D. D., Baker-Herman, T. L., Golder, F. J., Doperalski,N. J., Watters, J. J., & Mitchell, G. S. (2005). Cervical spinalcord injury upregulates ventral spinal 5-HT2A receptors.Journal of Neurotrauma, 22, 203–213.
Gaunt, R. A., Prochazka, A., Mushahwar, V. K., Guevremont,L., & Ellaway, P. H. (2006). Intraspinal microstimulationexcites multisegmental sensory afferents at lower stimuluslevels than local alpha-motoneuron responses. Journal ofNeurophysiology, 96, 2995–3005.
Gerasimenko, Y., Roy, R. R., & Edgerton, V. R. (2008).Epidural stimulation: comparison of the spinal circuits thatgenerate and control locomotion in rats, cats and humans.Experimental Neurology, 209, 417–425.
414
Author's personal copy

Gerasimenko, Y. P., Avelev, V. D., Nikitin, O. A., &Lavrov, I. A. (2003). Initiation of locomotor activity inspinal cats by epidural stimulation of the spinal cord.Neuroscience and Behavioral Physiology, 33, 247–254.
Gerasimenko, Y. P., Ichiyama, R. M., Lavrov, I. A., Courtine, G.,Cai, L., Zhong, H., et al. (2007). Epidural spinal cord stimula-tion plus quipazine administration enable stepping in completespinal adult rats. Journal of Neurophysiology, 98, 2525–2536.
Gerasimenko, Y. P., Makarovskii, A. N., & Nikitin, O. A.(2002). Control of locomotor activity in humans and animalsin the absence of supraspinal influences. Neuroscience andBehavioral Physiology, 32, 417–423.
Gerrits, H. L., De Haan, A., Hopman, M. T., Van DerWoude, L. H., Jones, D. A., & Sargeant, A. J. (1999).Contractile properties of the quadriceps muscle in indivi-duals with spinal cord injury. Muscle & Nerve, 22, 1249–1256.
Gerrits, H. L., De Haan, A., Sargeant, A. J., Dallmeijer, A., &Hopman, M. T. (2000). Altered contractile properties of thequadriceps muscle in people with spinal cord injury followingfunctional electrical stimulated cycle training. Spinal Cord,38, 214–223.
Gerrits, H. L., Hopman, M. T., Offringa, C., Engelen, B. G.,Sargeant, A. J., Jones, D. A., et al. (2003). Variability in fibreproperties in paralysed human quadriceps muscles andeffects of training. Pflugers Archive, 445, 734–740.
Gerrits, H. L., Hopman, M. T., Sargeant, A. J., Jones, D. A., &De Haan, A. (2002). Effects of training on contractileproperties of paralyzed quadriceps muscle. Muscle & Nerve,25, 559–567.
Gimenez, Y., Ribotta, M., Orsal, D., Feraboli-Lohnherr, D.,Privat, A., Provencher, J., et al. (1998). Kinematic analysis ofrecovered locomotor movements of the hindlimbs inparaplegic rats transplanted with monoaminergic embryonicneurons. Annals of the New York Academy of Sciences, 860,521–523.
Gomez-Pinilla, F., Ying, Z., Roy, R. R., Hodgson, J., &Edgerton, V. R. (2004). Afferent input modulates neuro-trophins and synaptic plasticity in the spinal cord. Journal ofNeurophysiology, 92, 3423–3432.
Grillner, S. (1981). Control of locomotion in bipeds, tetrapods,and fish. In J. M. Brookhart & V. B. Mountcastle (Eds.), Thenervous system: motor control. Bethesda, MD: AmericanPhysiological Society.
Guertin, P. A. (2004a). Role of NMDA receptor activation inserotonin agonist-induced air-stepping in paraplegic mice.Spinal Cord, 42, 185–190.
Guertin, P. A. (2004b). Synergistic activation of the centralpattern generator for locomotion by l-beta-3,4-dihydroxy-phenylalanine and quipazine in adult paraplegic mice.Neuroscience Letters, 358, 71–74.
Hains, B. C., Johnson, K. M., Mcadoo, D. J., Eaton, M. J., &Hulsebosch, C. E. (2001). Engraftment of serotonergicprecursors enhances locomotor function and attenuateschronic central pain behavior following spinal hemisectioninjury in the rat. Experimental Neurology, 171, 361–378.
Harkema, S. J., Hurley, S. L., Patel, U. K., Requejo, P. S.,Dobkin, V. R., & Edgerton, V. R. (1997). Human
lumbosacral spinal cord interprets loading during stepping.Journal of Neurophysiology, 77, 797–811.
Harridge, S. D., Andersen, J. L., Hartkopp, A., Zhou, S.,Biering-Sorensen, F., Sandri, C., et al. (2002). Training bylow-frequency stimulation of tibialis anterior in spinal cord-injured men. Muscle & Nerve, 25, 685–694.
Hartkopp, A., Harridge, S. D., Mizuno, M., Ratkevicius, A.,Quistorff, B., Kjaer, M., et al. (2003). Effect of trainingon contractile and metabolic properties of wrist extensorsin spinal cord-injured individuals. Muscle & Nerve, 27,72–80.
Hesse, S., Schmidt, H., Werner, C., & Bardeleben, A. (2003).Upper and lower extremity robotic devices for rehabilita-tion and for studying motor control. Current Opinion inNeurology, 16, 705–710.
Hochman, S., Garraway, S. M., Machacek, D. W., &Shay, B. L. (2001). 5-HT receptors and the neruomodulatorycontrol of spinal cord functions. In T. C. Cope (Ed.),Motor neurobiology of the spinal cord. Boca Raton, FL:CRC Press.
Humphrey, D. R., Mao, H., & Griffith, R. W. (2006). Brainreorganization continues for 25 years after spinal injury:changing patterns of activation in human cerebral cortexduring attempts to move. Society of Neuroscience Abstract,Program Number 88.10.
Ichiyama, R. M., Gerasimenko, Y. P., Zhong, H., Roy, R. R., &Edgerton, V. R. (2005). Hindlimb stepping movements incomplete spinal rats induced by epidural spinal cord stimula-tion. Neuroscience Letters, 383, 339–344.
Ichiyama, R. M., Courtine, G., Gerasimenko, Y. P., Yang, G. J.,van den Brand, R., Lavrov, I. A., et al. (2008). Step trainingreinforces specific spinal locomotor circuitry in adult spinalrats. The Journal of Neuroscience, 28, 7370–7375.
Jilge, B., Minassian, K., Rattay, F., & Dimitrijevic, M. R.(2004). Frequency-dependent selection of alternative spinalpathways with common periodic sensory input. BiologicalCybernetics, 91, 359–376.
Jordan, L. M., & Schmidt, B. J. (2002). Propriospinal neuronsinvolved in the control of locomotion: potential targetsfor repair strategies? In L. McKerracher, G. Doucet, &S. Rossignol (Eds.), Progress in brain research. Amsterdam:Elsevier Science, B.V.
Kazennikov, O. V., Shik, M. L., & Yakovleva, G. V. (1983).Stepping movements induced in cats by stimulation of thedorsolateral funiculus of the spinal-cord. Bulletin of experi-mental biology and medicine, 96, 1036–1039.
Kern, H., Boncompagni, S., Rossini, K., Mayr, W., Fano, G.,Zanin, M. E., et al. (2004). Long-term denervation inhumans causes degeneration of both contractile and excita-tion-contraction coupling apparatus, which is reversible byfunctional electrical stimulation (FES): a role for myofiberregeneration? Journal of neuropathology and experimentalneurology, 63, 919–931.
Kiehn, O., Quinlan, K. A., Restrepo, C. E., Lundfald, L.,Borgius, L., Talpalar, A. E., et al. (2008). Excitatory com-ponents of the mammalian locomotor CPG. Brain Research.Brain Research Reviews, 57, 56–63.
415
Author's personal copy

Kim, D., Adipudi, V., Shibayama, M., Giszter, S., Tessler, A.,Murray, M., et al. (1999). Direct agonists for serotoninreceptors enhance locomotor function in rats that receivedneural transplants after neonatal spinal transection. TheJournal of Neuroscience, 19, 6213–6224.
Kim, S. J., Roy, R. R., Kim, J. A., Zhong, H., Haddad, F.,Baldwin, K. M., et al. (2008). Gene expression duringinactivity-induced muscle atrophy: effects of brief bouts of aforceful contraction countermeasure. Journal of AppliedPhysiology, 105, 1246–1254.
Kim, S. J., Roy, R. R., Zhong, H., Suzuki, H., Ambartsumyan, L.,Haddad, F., et al. (2007). Electromechanical stimulationameliorates inactivity-induced adaptations in the medialgastrocnemius of adult rats. Journal of Applied Physiology,103, 195–205.
Lavrov, I., Dy, C. J., Fong, A. J., Gerasimenko, Y., Courtine, G.,Zhong, H., et al. (2008). Epidural stimulation inducedmodulation of spinal locomotor networks in adult spinal rats.The Journal of Neuroscience, 28, 6022–6029.
Lavrov, I., Gerasimenko, Y. P., Ichiyama, R. M., Courtine, G.,Zhong, H., Roy, R. R., et al. (2006). Plasticity of spinal cordreflexes after a complete transection in adult rats: relation-ship to stepping ability. Journal of Neurophysiology, 96,1699–1710.
Ledoux, M. S., & Langford, K. H. (1993). Spinal cordstimulation for the failed back syndrome. Spine, 18, 191–194.
Lee, Y. S., Lin, C. Y., Caiozzo, V. J., Robertson, R. T.,Yu, J., & Lin, V. W. (2007). Repair of spinal cord transec-tion and its effects on muscle mass and myosin heavy chainisoform phenotype. Journal of Applied Physiology, 103,1808–1814.
Liang, Y., Cai, L. L., Burdick, J. W., & Edgerton, V. R. (2006).A robotic training system for studies of post-SCI standrehabilitation. In: The First IEEE/RAS-EMBS InternationalConference on Biomedical Robotics and Biomechatronics,2006 (BioRob 2006).
Lovely, R. G., Gregor, R. J., Roy, R. R., & Edgerton, V. R.(1986). Effects of training on the recovery of full-weight-bearing stepping in the adult spinal cat. ExperimentalNeurology, 92, 421–435.
Lovely, R. G., Gregor, R. J., Roy, R. R., & Edgerton, V. R.(1990). Weight-bearing hindlimb stepping in treadmill-exercised adult spinal cats. Brain Research, 514, 206–218.
Maegele, M., Muller, S., Wernig, A., Edgerton, V. R., &Harkema, S. J. (2002). Recruitment of spinal motor poolsduring voluntary movements versus stepping after humanspinal cord injury. Journal of Neurotrauma, 19, 1217–1229.
Maier, I. C., Ichiyama, R. M., Courtine, G., Schnell, L.,Lavrov, I., Edgerton, V. R., et al. (2009). Differentialeffects of anti-Nogo-A antibody treatment and treadmilltraining in rats with incomplete spinal cord injury. Brain(in press).
Majczynski, H., Maleszak, K., Cabaj, A., & Slawinska, U.(2005). Serotonin-related enhancement of recovery ofhind limb motor functions in spinal rats after graftingof embryonic raphe nuclei. Journal of Neurotrauma, 22,590–604.
Marino, R. J., Ditunno, J. F., Jr., Donovan, W. H., &Maynard, F., Jr. (1999). Neurologic recovery after traumaticspinal cord injury: data from the Model Spinal Cord InjurySystems. Archives of Physical Medicine and Rehabilitation,80, 1391–1396.
Martin, T. P., Stein, R. B., Hoeppner, P. H., & Reid, D. C.(1992). Influence of electrical stimulation on the morpholo-gical and metabolic properties of paralyzed muscle. Journalof Applied Physiology, 72, 1401–1406.
Mcewen, M. L., & Stehouwer, D. J. (2001). Kinematic analysesof air-stepping of neonatal rats after mid-thoracic spinal cordcompression. Journal of Neurotrauma, 18, 1383–1397.
Merrill, D. R., Bikson, M., & Jefferys, J. G. (2005). Elec-trical stimulation of excitable tissue: design of efficaciousand safe protocols. Journal of Neuroscience Methods, 141,171–198.
Mori, S., Sakamoto, T., & Takakusaki, K. (1991). Interactionof posture and locomotion in cats: its automatic andvolitional control aspects. In M. Shimamura, S. Grillner, &V. R. Edgerton (Eds.), Neurobiological basis of humanlocomotion. Tokyo: Japan Scientific Societies Press.
Mushahwar, V. K., Aoyagi, Y., Stein, R. B., & Prochazka, A.(2004). Movements generated by intraspinal microstimula-tion in the intermediate gray matter of the anesthetized,decerebrate, and spinal cat. Canadian Journal of Physiologyand Pharmacology, 82, 702–714.
Mushahwar, V. K., Gillard, D. M., Gauthier, M. J., &Prochazka, A. (2002). Intraspinal micro stimulation gener-ates locomotor-like and feedback-controlled movements.IEEE Transactions on Neural Systems and RehabilitationEngineering, 10, 68–81.
Musienko, P. E., Bogacheva, I. N., & Gerasimenko, Y. P.(2007). Significance of peripheral feedback in the generationof stepping movements during epidural stimulation of thespinal cord. Neuroscience and Behavioral Physiology, 37,181–190.
Nance, P. W. (2003). Management of spasticity. In V. W. Lin(Ed.), Spinal cord medicine: Principles and practice. NewYork: Demos Medical Publishing.
North, R. B., Kidd, D. H., Petrucci, L., & Dorsi, M. J. (2005).Spinal cord stimulation electrode design: a prospective,randomized, controlled trial comparing percutaneous withlaminectomy electrodes: part II-clinical outcomes. Neuro-surgery, 57, 990–996.
Ohta, Y., Yano, H., Suzuki, R., Yoshida, M., Kawashima, N., &Nakazawa, K. (2007). A two-degree-of-freedom motor-powered gait orthosis for spinal cord injury patients.Proceedings of the Institution of Mechanical Engineers.Part H, Journal of Engineering in Medicine, 221, 629–639.
Otoshi, C. K., Fong, A. J., Cai, L. L., Zhong, H., Roy, R. R.,Tillakaratne, N. J. K., et al. (2005). 5-HT receptor distribu-tion after spinal cord transection: effects of chronic seroto-nergic agonist administration and robotic training. Society ofNeuroscience Abstract, Program Number 396.11.
Otoshi, C. K., Walwyn, W. M., Tillakaratne, N. J. K.,Zhong, H., Roy, R. R., & Edgerton, V. R. (2009). Distribu-tion and localization of 5-HT(1A) receptors in the rat lumbar
416
Author's personal copy

spinal cord after transection and deafferentation. Journal ofNeurotrauma, 26, 575–584.
Parker, D. (2005). Pharmacological approaches to functionalrecovery after spinal injury. Current Drug Targets. CNS andNeurological Disorders, 4, 195–210.
Reinkensmeyer, D. J., Aoyagi, D., Emken, J. L., Galvez, J. A.,Ichinose, W., Kerdanyan, G., et al. (2006). Tools for under-standing and optimizing robotic gait training. Journal ofRehabilitation Research and Development, 43, 657–670.
Renard, V. M., & North, R. B. (2006). Prevention ofpercutaneous electrode migration in spinal cord stimulationby a modification of the standard implantation technique.Journal of Neurosurgery: Spine, 4, 300–303.
Ribotta, M. G., Provencher, J., Feraboli-Lohnherr, D.,Rossignol, S., Privat, A., & Orsal, D. (2000). Activation oflocomotion in adult chronic spinal rats is achieved bytransplantation of embryonic raphe cells reinnervatinga precise lumbar level. The Journal of Neuroscience, 20,5144–5152.
Robinson, G. A., & Goldberger, M. E. (1986). The develop-ment and recovery of motor function in spinal cats. II.Pharmacological enhancement of recovery. ExperimentalBrain Research, 62, 387–400.
Rodger, D. C., Fong, A. J., Li, W., Ameri, H., Ahuja, A. K.,Gutierrez, C., et al. (2008). Flexible parylene-based multi-electrode array technology for high-density neural stimula-tion and recording. Sensors and Actuators B: Chemical, 132,449–460.
Rodger, D. C., Fong, A. J., Li, W., Ameri, H., Lavrov, I.,Zhong, H., et al. (2007). High-density flexible parylene-based multielectrode arrays for retinal and spinal cordstimulation. Solid-State Sensors, Actuators and MicrosystemsConference, 2007. Transducers, pp. 1385–1388.
Rossignol, S., & Barbeau, H. (1993). Pharmacology oflocomotion: an account of studies in spinal cats and spinalcord injured subjects. The Journal of the American Para-plegia Society, 16, 190–196.
Rossignol, S., Chau, C., Brustein, E., Giroux, N., Bouyer, L.,Barbeau, H., et al. (1998). Pharmacological activation andmodulation of the central pattern generator for locomotionin the cat. Annals of the New York Academy of Sciences, 860,346–359.
Rossignol, S., Giroux, N., Chau, C., Marcoux, J., Brustein, E., &Reader, T. A. (2001). Pharmacological aids to locomotortraining after spinal injury in the cat. Journal of Physiology,533, 65–74.
Roy, R. R., Baldwin, K. M., & Edgerton, V. R. (1991). Theplasticity of skeletal muscle: effects of neuromuscularactivity. Exercise and Sport Sciences Reviews, 19, 269–312.
Roy, R. R., Talmadge, R. J., Hodgson, J. A., Oishi, Y.,Baldwin, K. M., & Edgerton, V. R. (1999). Differentialresponse of fast hindlimb extensor and flexor muscles to exer-cise in adult spinalized cats. Muscle & Nerve, 22, 230–241.
Roy, R. R., Talmadge, R. J., Hodgson, J. A., Zhong, H.,Baldwin, K. M., & Edgerton, V. R. (1998). Training effectson soleus of cats spinal cord transected (T12-13) as adults.Muscle & Nerve, 21, 63–71.
Roy, R. R., Zhong, H., Hodgson, J. A., Grossman, E. J.,Siengthai, B., Talmadge, R. J., et al. (2002). Influencesof electromechanical events in defining skeletal muscleproperties. Muscle & Nerve, 26, 238–251.
Roy, R. R., Zhong, H., Khalili, N., Kim, S. J., Higuchi, N.,Monti, R. J., et al. (2007). Is spinal cord isolation a goodmodel of muscle disuse? Muscle & Nerve, 35, 312–321.
Scivoletto, G., Morganti, B., Ditunno, P., Ditunno, J. F., &Molinari, M. (2003). Effects on age on spinal cord lesionpatients’ rehabilitation. Spinal Cord, 41, 457–464.
Shields, R. K. (1995). Fatigability, relaxation properties, andelectromyographic responses of the human paralyzed soleusmuscle. Journal of Neurophysiology, 73, 2195–2206.
Shields, R. K., & Dudley-Javoroski, S. (2006). Musculoskeletalplasticity after acute spinal cord injury: effects of long-termneuromuscular electrical stimulation training. Journal ofNeurophysiology, 95, 2380–2390.
Shik, M. L., & Orlovsky, G. N. (1976). Neurophysiology oflocomotor automatism. Physiological Reviews, 56, 465–501.
Shik, M. L., Severin, F. V., & Orlovskii, G. N. (1966). Controlof walking and running by means of electric stimulation ofthe midbrain. Biofizika, 11, 659–666.
Stein, R. B., Gordon, T., Jefferson, J., Sharfenberger, A.,Yang, J. F., De Zepetnek, J. T., et al. (1992). Optimalstimulation of paralyzed muscle after human spinal cordinjury. Journal of Applied Physiology, 72, 1393–1400.
Stewart, B. G., Tarnopolsky, M. A., Hicks, A. L., Mccartney, N.,Mahoney, D. J., Staron, R. S., et al. (2004). Treadmilltraining-induced adaptations in muscle phenotype in personswith incomplete spinal cord injury. Muscle & Nerve, 30,61–68.
Talmadge, R. J. (2000). Myosin heavy chain isoform expressionfollowing reduced neuromuscular activity: potential regula-tory mechanisms. Muscle & Nerve, 23, 661–679.
Talmadge, R. J., Castro, M. J., Apple, D. F., Jr., & Dudley, G. A.(2002). Phenotypic adaptations in human muscle fibers 6 and24 wk after spinal cord injury. Journal of Applied Physiology,92, 147–154.
Tillakaratne, N. J., De Leon, R. D., Hoang, T. X., Roy, R. R.,Edgerton, V. R., & Tobin, A. J. (2002). Use-dependentmodulation of inhibitory capacity in the feline lumbar spinalcord. The Journal of Neuroscience, 22, 3130–3143.
Timoszyk, W. K., De Leon, R. D., London, N., Joynes, R.,Minakata, K., Roy, R. R., et al. (2003). Comparison ofvirtual and physical treadmill environments for trainingstepping after spinal cord injury. Robotica, 21, 25–32.
Timoszyk, W. K., De Leon, R. D., London, N., Roy, R. R.,Edgerton, V. R., & Reinkensmeyer, D. J. (2002). The ratlumbosacral spinal cord adapts to robotic loading appliedduring stance. Journal of Neurophysiology, 88, 3108–3117.
Timoszyk, W. K., Nessler, J. A., Acosta, C., Roy, R. R.,Edgerton, V. R., Reinkensmeyer, D. J., et al. (2005). Hind-limb loading determines stepping quantity and qualityfollowing spinal cord transection. Brain Research, 1050,180–189.
Van De Crommert, H. W., Mulder, T., & Duysens, J. (1998).Neural control of locomotion: sensory control of the central
417
Author's personal copy

pattern generator and its relation to treadmill training.Gait & Posture, 7, 251–263.
Van Den Brand, R. J., Gerasimenko, Y., Dy, C. J., Ichiyama,R. M., Lavrov, I., Zhong, H., et al. (2007). Epidural stimula-tion and pharmacological interventions facilitate neuroreh-abilitation by enabling stepping following spinal cord injury.Society of Neuroscience Abstract, Program Number 75.13.
Waters, R. L., Sie, I., Adkins, R. H., & Yakura, J. S. (1995).Injury pattern effect on motor recovery after traumaticspinal cord injury. Archives of Physical Medicine andRehabilitation, 76, 440–443.
Wernig, A., Nanassy, A., & Muller, S. (1998). Maintenance oflocomotor abilities following Laufband (treadmill) therapyin para- and tetraplegic persons: follow-up studies. SpinalCord, 36, 744–749.
Winchester, P., & Querry, R. (2006). Robotic orthoses forbody weight-supported treadmill training. Physical Medicineand Rehabilitation Clinics of North America, 17, 159–172.
Ying, Z., Roy, R. R., Edgerton, V. R., & Gomez-Pinilla, F.(2005). Exercise restores levels of neurotrophins andsynaptic plasticity following spinal cord injury. ExperimentalNeurology, 193, 411–419.
418
Author's personal copy
Related Documents











