Ž . Brain Research 782 1998 126–135 Research report Spontaneous long-term remyelination after traumatic spinal cord injury in rats Hermelinda Salgado-Ceballos a,b, ) , Gabriel Guizar-Sahagun a,b , Alfredo Feria-Velasco c , Israel Grijalva a,b , Laura Espitia b , Antonio Ibarra a,b , Ignacio Madrazo b a Unit of Medical Research in Neurological Diseases, Instituto Mexicano del Seguro Social, Mexico, DF, C.P. 06720, Mexico ´ ´ b Proyecto Camina A.C., Mexico DF, C.P. 14050, Mexico ´ ´ c DiÕision of Experimental Pathology and Biotechnology, CIATEJ, Guadalajara, Jalisco, Mexico ´ Accepted 14 October 1997 Abstract The capability of the central nervous system to remyelinate axons after a lesion has been well documented, even though it had been Ž . described as an abortive and incomplete process. At present there are no long- term morphometric studies to assess the spinal cord SC remyelinative capability. With the purpose to understand this phenomenon better, the SC of seven lesionless rats and the SC of 21 rats subjected to a severe weight-drop contusion injury were evaluated at 1, 2, 4, 6, and 12 months after injury. The axonal diameter and the Ž . myelination index MI saxolemmal perimeter divided by myelinated fiber perimeter were registered in the outer rim of the cord at T9 SC level using a transmission electron microscope and a digitizing computer system. The average myelinated fiber loss was 95.1%. One month after the SC, 64% of the surviving fibers were demyelinated while 12 months later, only 30% of the fibers had no myelin sheath. Ž . The MI in the control group was 0.72 "0.07 X "S.D. . In the experimental groups, the greatest demyelination was observed two Ž . Ž . months after the lesion MI s0.90 "0.03 , while the greatest myelination was observed 12 months after the injury MI s0.83 "0.02 . Ž . There was a statistical difference p -0.02 in MI between 2 and 12 months which means that remyelination had taken place. Ž . Remyelination was mainly achieved because of Schwann cells. The proportion of small fibers diameter s0.5 mm or less considered as axon collaterals, increased from 18.45% at 1 month to 27.66% a year after the contusion. Results suggest that remyelination is not an abortive phenomenon but in fact a slow process occurring parallel to other tissue plastic phenomena, such as the emission of axon collaterals. q 1998 Elsevier Science B.V. Keywords: Demyelination; Myelination index; Schwann cell; Neural plasticity; Spinal cord contusion; Paraplegia 1. Introduction Ž . Mechanical trauma to the spinal cord SC produces axonal destruction and extensive demyelination by damag- ing, not only axons, but also oligodendrocytes and astro- w x cytes 8,9,26 . As a result, axons that survive the injury lose their myelin sheath and cannot be remyelinated, while axons or collaterals, resulting from the plastic mechanisms w x of the SC, are not myelinated 8,9,21,61 . Myelin is essential for normal nervous system function w x 57 so, loss of the axon myelin sheath has important repercussions on nerve physiology as may be a decrease in conduction velocity or overt nerve conduction block that may play a major role in the neurological deficits after SC ) Corresponding author. Calzada de Tlalpan 4430, Colonia Toriello Guerra C.P. 14050, Mexico DF, Mexico. Fax: q52-5-5-73-55-45. ´ ´ w x injury 61 . As a consequence, functional recovery of this pathology depends at least in part on nervous system w x remyelinating ability 40,44,60,61 . Research over the past several decades has demon- strated that myelin repair in the central nervous system Ž . CNS is a normal physiological response to myelin injury w x 13,38,49 . However, some authors regard remyelination in this system as an abortive and incomplete phenomenon w x 3,5,6,11,18,25,31–33,39,47,48,56 . The availability of numerous models of CNS demyeli- nation amenable to experimental manipulation w x 20,30,37,41,45,54,62 has allowed a broad range of ap- proaches to develop and for evaluating therapies designed w x to promote CNS remyelination 19,42–45,55 . However, with the aim to design rational therapeutic strategies in different pathologic processes related to SC injuries, it is necessary to clearly understand the spontaneous plastic 0006-8993r98r$19.00 q 1998 Elsevier Science B.V. All rights reserved.

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Ž .Brain Research 782 1998 126–135
Research report
Spontaneous long-term remyelination after traumatic spinal cord injury inrats
Hermelinda Salgado-Ceballos a,b,), Gabriel Guizar-Sahagun a,b, Alfredo Feria-Velasco c,Israel Grijalva a,b, Laura Espitia b, Antonio Ibarra a,b, Ignacio Madrazo b
a Unit of Medical Research in Neurological Diseases, Instituto Mexicano del Seguro Social, Mexico, DF, C.P. 06720, Mexico´ ´b Proyecto Camina A.C., Mexico DF, C.P. 14050, Mexico´ ´
c DiÕision of Experimental Pathology and Biotechnology, CIATEJ, Guadalajara, Jalisco, Mexico´
Accepted 14 October 1997
Abstract
The capability of the central nervous system to remyelinate axons after a lesion has been well documented, even though it had beenŽ .described as an abortive and incomplete process. At present there are no long- term morphometric studies to assess the spinal cord SC
remyelinative capability. With the purpose to understand this phenomenon better, the SC of seven lesionless rats and the SC of 21 ratssubjected to a severe weight-drop contusion injury were evaluated at 1, 2, 4, 6, and 12 months after injury. The axonal diameter and the
Ž .myelination index MIsaxolemmal perimeter divided by myelinated fiber perimeter were registered in the outer rim of the cord at T9SC level using a transmission electron microscope and a digitizing computer system. The average myelinated fiber loss was 95.1%. Onemonth after the SC, 64% of the surviving fibers were demyelinated while 12 months later, only 30% of the fibers had no myelin sheath.
Ž .The MI in the control group was 0.72"0.07 X"S.D. . In the experimental groups, the greatest demyelination was observed twoŽ . Ž .months after the lesion MIs0.90"0.03 , while the greatest myelination was observed 12 months after the injury MIs0.83"0.02 .
Ž .There was a statistical difference p-0.02 in MI between 2 and 12 months which means that remyelination had taken place.Ž .Remyelination was mainly achieved because of Schwann cells. The proportion of small fibers diameters0.5 mm or less considered as
axon collaterals, increased from 18.45% at 1 month to 27.66% a year after the contusion. Results suggest that remyelination is not anabortive phenomenon but in fact a slow process occurring parallel to other tissue plastic phenomena, such as the emission of axoncollaterals. q 1998 Elsevier Science B.V.
Keywords: Demyelination; Myelination index; Schwann cell; Neural plasticity; Spinal cord contusion; Paraplegia
1. Introduction
Ž .Mechanical trauma to the spinal cord SC producesaxonal destruction and extensive demyelination by damag-ing, not only axons, but also oligodendrocytes and astro-
w xcytes 8,9,26 . As a result, axons that survive the injurylose their myelin sheath and cannot be remyelinated, whileaxons or collaterals, resulting from the plastic mechanisms
w xof the SC, are not myelinated 8,9,21,61 .Myelin is essential for normal nervous system function
w x57 so, loss of the axon myelin sheath has importantrepercussions on nerve physiology as may be a decrease inconduction velocity or overt nerve conduction block thatmay play a major role in the neurological deficits after SC
) Corresponding author. Calzada de Tlalpan 4430, Colonia TorielloGuerra C.P. 14050, Mexico DF, Mexico. Fax: q52-5-5-73-55-45.´ ´
w xinjury 61 . As a consequence, functional recovery of thispathology depends at least in part on nervous system
w xremyelinating ability 40,44,60,61 .Research over the past several decades has demon-
strated that myelin repair in the central nervous systemŽ .CNS is a normal physiological response to myelin injuryw x13,38,49 . However, some authors regard remyelination inthis system as an abortive and incomplete phenomenonw x3,5,6,11,18,25,31–33,39,47,48,56 .
The availability of numerous models of CNS demyeli-nation amenable to experimental manipulationw x20,30,37,41,45,54,62 has allowed a broad range of ap-proaches to develop and for evaluating therapies designed
w xto promote CNS remyelination 19,42–45,55 . However,with the aim to design rational therapeutic strategies indifferent pathologic processes related to SC injuries, it isnecessary to clearly understand the spontaneous plastic
0006-8993r98r$19.00 q 1998 Elsevier Science B.V. All rights reserved.Ž .PII S0006-8993 97 01252-3

( )H. Salgado-Ceballos et al.rBrain Research 782 1998 126–135 127
mechanisms subjacent in each one of those processes andnot to negatively interfere with them.
The long-term morphometric effect on the axon diame-ters and axonal myelination of the adult rat SC fibers afteran experimental weight-drop contusion injury was ana-lyzed in order to better understand the CNS remyelinativecapacity and knowing whether remyelination constitutes anabortive and incomplete process in the biology of the SC.
2. Materials and methods
2.1. Surgery and post-surgical care
Twenty-eight Long-Evans female rats 14 to 16 weeks ofage, weighing 240 to 260 g, were anesthetized with an
Ž . Ž .intramuscular IM mixture of ketamine 77.5 mgrkg bwŽ .and xylacine 12.5 mgrkg bw . Benzatin penicillin
Ž .240,000 units was administered IM, as a single dose,once the rat was anesthetized.
The protocol was accepted by the hospital’s EthicsCommittee while considering the proper handling, hous-ing, surgical and postsurgical care of the animals. Underaseptic conditions, each rat was subjected to a one-level
Ž .laminectomy T9 , exposing the dorsal surface of the SC,without disturbing the meninges. Under stereotactic con-
Ž .trol, a severe spinal cord contusion SCC was causedw xusing the weight-drop method described by Allen 1 and
w xmodified by our group 28 . For a brief moment, a 15 gstainless steel cylinder with a flat 2 mm diameter tip, wasdropped from a height of 10 cm, through a guide tube ontothe exposed dura mater. The formation of a hematoma inthe impacted area was corroborated and, finally, the mus-cle aponeurosis and the skin were separately sutured overthe laminectomy site with 5-0 nylon.
After the surgical procedure, the rats were placed in aŽcritical care unit for small animals Schroer Manufactur-
.ing, Kansas City, KA until their recovery from the anes-thesia. The rats were fed ad libitum and caged separately.Neurogenic bladder and intestine were manually managedtwice a day until automatism was recovered.
2.2. Experimental groups
The time course of the axonal diameter and myelinsheath morphometric alterations were recorded in 21 rats,
Ž .divided in 5 different subgroups studied at 1 ns4 , 2Ž . Ž . Ž . Ž .ns5 , 4 ns5 , 6 ns4 and 12 ns3 months afterthe severe SCC. As a control group, seven animals werestudied without lesion of the SC.
2.3. Electron microscopy
Rats were anesthetized as described above and perfusedintra-aortically at 30 mlrmin and at room temperature
Žwith 100 ml of 0.15 molrl sodium cacodylate buffer pH.7.4 plus 1000 units of heparin, followed by 500 ml of a
fixative solution containing 2.5% glutaraldehyde, 2% para-formaldehyde, 0.05% CaCl and 0.025 molrl sucrose2
Ž .solution in 0.1 molrl cacodylate buffer pH 7.4 . Afterperfusion, the SC corresponding from T8 to T10 levelswas dissected, removed and sectioned transversely at the
Žepicenter zone highest tissue destruction transverse area.of the SC . The blocks obtained were placed in the same
fixative used for perfusion for 2 h and then in 0.1 molrlcacodylate buffer for 14 to 16 h. The blocks of approxi-mately 1 mm3 were post-fixed for an hour with aqueous0.1% osmium tetroxide solution. They were then dehy-drated in graded ethanol, and embedded in EPON 812resin. Semithin coronal sections 1 mm thick were stainedwith toluidine blue and observed under a light microscopein order to identify the area corresponding to the peripheralsubpial rim of white matter with preserved axons at theepicenter injured zone. The material was resectioned usingan ultramicrotome, then sections of approximately 70 nmthick were obtained which were collected on copper grids,contrasted with uranyl acetate and lead citrate and ob-served under a Zeiss EM-10 transmission electron micro-scope. The negatives were obtained at =3150 and 6300 ona 35 mm format.
2.4. Image analysis
The negatives were projected on a digitizing tabletŽ .Hewlett Packard 9874A connected to a computer systemŽ .Hewlett Packard 9825B . Digitization of fibers was madetracing their perimeters with an electronic pen. Repetitivemeasurements obtained by digitizing the same fibers weredone achieving a variation of 0.38% between two differentinvestigators and 0.32% by the same investigator. Theaxon diameters were calculated from perimeter measure-ments assuming the perimeter to be that of a perfect circleto avoid errors due to fixation changes in axon shape. Inorder to obtain the relative thickness of the myelin sheathsŽ . Ž .myelination index or MI , the inner axon and outerŽ .axonqmyelin perimeters of each fiber were calculatedand then the inner perimeter was divided by the outerperimeter.
All data were stored in a AcerMate DX4 computer andanalyzed using the Lotus 1-2-3 program. The mean MI ofeach group was obtained. If the inner perimeter divided bythe outer perimeter representing the MI was equal to 1, theaxons had no myelin and the farther away from 1, thethicker the myelin sheath was.
2.5. Statistical analysis
Data were analyzed by the Number Cruncher StatisticalSystem. The sesgo and kurtosis did not have a normaldistribution. Therefore the nonparametric Mann–WhitneyU-test was used with the aim to compare the control MIwith the corresponding MI in the injured groups. TheSpearman’s Rho test was used to analyze the relationship
( )H. Salgado-Ceballos et al.rBrain Research 782 1998 126–135128
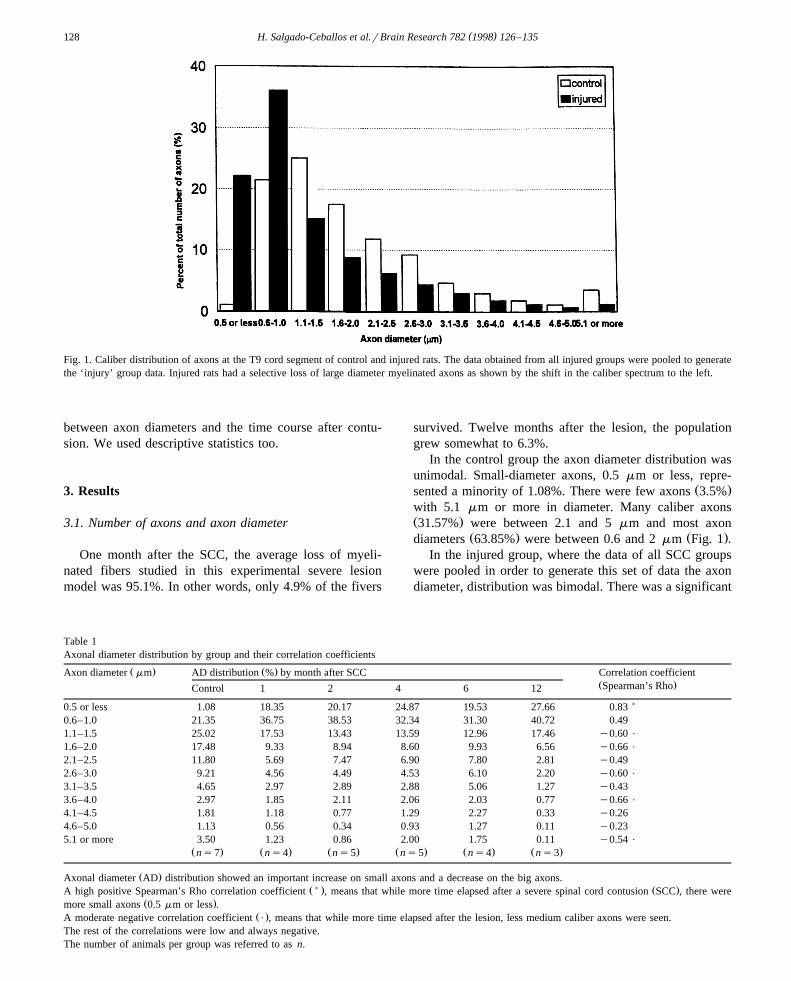
Fig. 1. Caliber distribution of axons at the T9 cord segment of control and injured rats. The data obtained from all injured groups were pooled to generatethe ‘injury’ group data. Injured rats had a selective loss of large diameter myelinated axons as shown by the shift in the caliber spectrum to the left.
between axon diameters and the time course after contu-sion. We used descriptive statistics too.
3. Results
3.1. Number of axons and axon diameter
One month after the SCC, the average loss of myeli-nated fibers studied in this experimental severe lesionmodel was 95.1%. In other words, only 4.9% of the fivers
survived. Twelve months after the lesion, the populationgrew somewhat to 6.3%.
In the control group the axon diameter distribution wasunimodal. Small-diameter axons, 0.5 mm or less, repre-
Ž .sented a minority of 1.08%. There were few axons 3.5%with 5.1 mm or more in diameter. Many caliber axonsŽ .31.57% were between 2.1 and 5 mm and most axon
Ž . Ž .diameters 63.85% were between 0.6 and 2 mm Fig. 1 .In the injured group, where the data of all SCC groups
were pooled in order to generate this set of data the axondiameter, distribution was bimodal. There was a significant
Table 1Axonal diameter distribution by group and their correlation coefficients
Ž . Ž .Axon diameter mm AD distribution % by month after SCC Correlation coefficientŽ .Spearman’s RhoControl 1 2 4 6 12
)0.5 or less 1.08 18.35 20.17 24.87 19.53 27.66 0.830.6–1.0 21.35 36.75 38.53 32.34 31.30 40.72 0.491.1–1.5 25.02 17.53 13.43 13.59 12.96 17.46 y0.60 P
1.6–2.0 17.48 9.33 8.94 8.60 9.93 6.56 y0.66 P
2.1–2.5 11.80 5.69 7.47 6.90 7.80 2.81 y0.492.6–3.0 9.21 4.56 4.49 4.53 6.10 2.20 y0.60 P
3.1–3.5 4.65 2.97 2.89 2.88 5.06 1.27 y0.433.6–4.0 2.97 1.85 2.11 2.06 2.03 0.77 y0.66 P
4.1–4.5 1.81 1.18 0.77 1.29 2.27 0.33 y0.264.6–5.0 1.13 0.56 0.34 0.93 1.27 0.11 y0.235.1 or more 3.50 1.23 0.86 2.00 1.75 0.11 y0.54 P
Ž . Ž . Ž . Ž . Ž . Ž .ns7 ns4 ns5 ns5 ns4 ns3
Ž .Axonal diameter AD distribution showed an important increase on small axons and a decrease on the big axons.Ž) . Ž .A high positive Spearman’s Rho correlation coefficient , means that while more time elapsed after a severe spinal cord contusion SCC , there were
Ž .more small axons 0.5 mm or less .Ž .A moderate negative correlation coefficient P , means that while more time elapsed after the lesion, less medium caliber axons were seen.
The rest of the correlations were low and always negative.The number of animals per group was referred to as n.
( )H. Salgado-Ceballos et al.rBrain Research 782 1998 126–135 129
Table 2Percentage of axons surviving after a spinal cord contusion and the corresponding percentage of myelinated and nonmyelinated axons
Groups % of axons surviving % of total population % of myelinated population % of nonmyelinated population
Axons Sproutings Axons Sproutings Total Axons Sproutings Total
Control 100.0 98.9 1.1 98.6 0.4 99.0 0.3 0.7 1.01 month 4.9 81.6 18.4 43.2 0.9 44.1 38.3 17.6 55.92 months 5.3 79.8 20.2 34.3 1.6 35.9 45.5 18.6 64.14 months 5.3 75.2 24.8 42.7 4.6 47.3 32.5 20.2 52.76 months 5.8 80.5 19.5 51.7 0.2 51.9 28.8 19.3 48.11 year 6.3 72.3 27.7 62.1 7.8 69.9 10.3 19.8 30.1
Percentage of axons surviving after spinal cord contusion derived from the total axon population in the control group in the area corresponding a peripheralring of white matter from the spinal cord, and the fibers surviving the lesion at this area in the experimental groups. Axons were identified when fiberdiameters )0.5 mm.Axon collaterals were identified when fiber diameterss or -0.5 mm.
increase in the proportion of the small caliber axons withan important loss of the largest. Small-diameter axons 0.5mm or less represented 22.12%, while 1.1 mm or morediameter axons diminished in different proportions withrespect to the control group even though the majority of
Ž . Žcaliber axons 59.7% were between 0.6 and 2 mm Fig..1 .
Ž .The proportion of small fibers 0.5 mm or less in-creased from 1.08% in the control group, to 27.66% at 1 yrafter the SCC. These fibers were considered as axoncollaterals or sproutings. There was a high correlationŽ .Spearman’s Rho correlation coefficients0.83 betweenthe time course after the SCC and the increase of thesesmall axons. The increase on 0.6 to 1 mm caliber axons
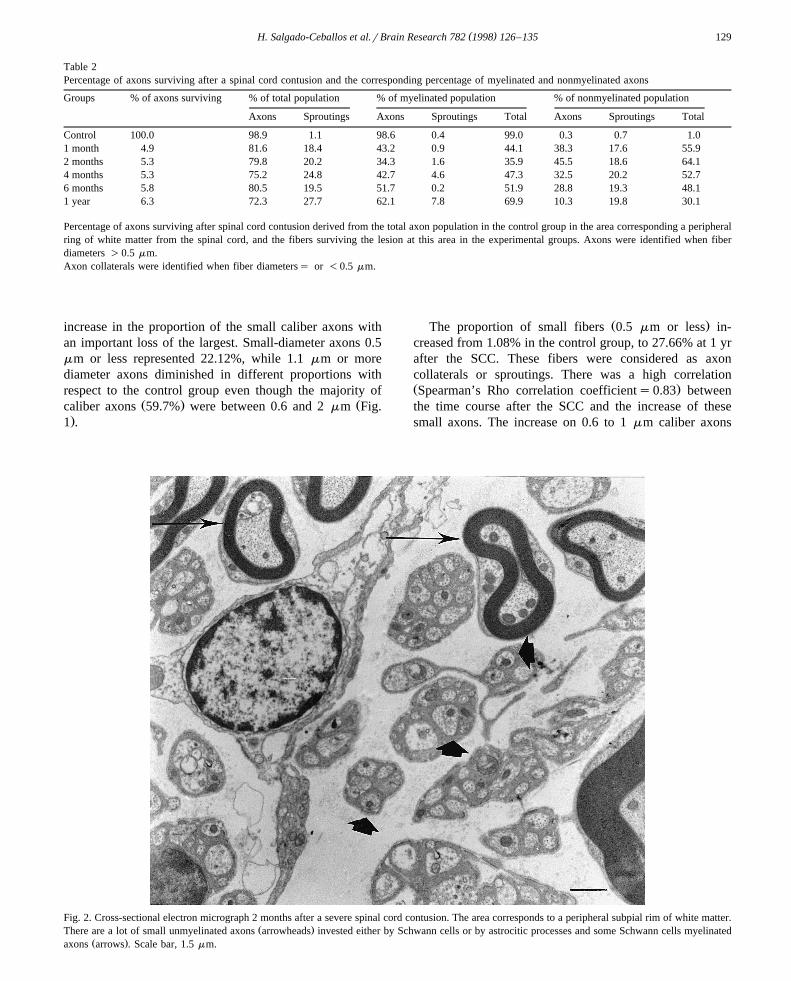
Fig. 2. Cross-sectional electron micrograph 2 months after a severe spinal cord contusion. The area corresponds to a peripheral subpial rim of white matter.Ž .There are a lot of small unmyelinated axons arrowheads invested either by Schwann cells or by astrocitic processes and some Schwann cells myelinated
Ž .axons arrows . Scale bar, 1.5 mm.
( )H. Salgado-Ceballos et al.rBrain Research 782 1998 126–135130
from 21.35% in the control group to 40.72% at 12 months,Žwas moderately significant Spearman’s Rho correlation
.coefficients0.49 . Axons with 1.1 mm or more decreasedŽin the injured groups Spearman’s Rho correlation coeffi-
.cient between y0.54 to y0.66 and showed a goodŽ .negative correlation Table 1 .
3.2. Myelin
Two months after the SCC, about 36% of the fiberswere myelinated and 64% had no myelin sheath. This is incontrast with the observations over 12 months when 70%of the fibers were myelinated and only 30% nonmyelinatedŽ .Table 2 .
One and two months after the lesion, several demyeli-nated axons were seen through the lesion site, eitherisolated or in small clusters sheathed by Schwann cell
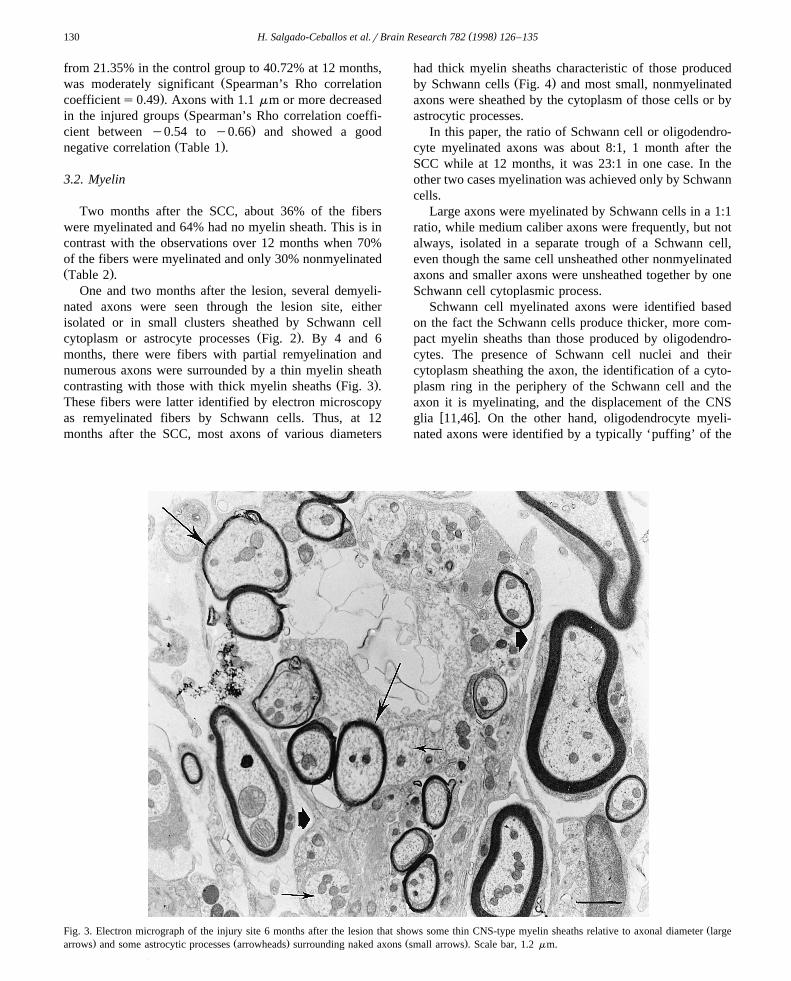
Ž .cytoplasm or astrocyte processes Fig. 2 . By 4 and 6months, there were fibers with partial remyelination andnumerous axons were surrounded by a thin myelin sheath
Ž .contrasting with those with thick myelin sheaths Fig. 3 .These fibers were latter identified by electron microscopyas remyelinated fibers by Schwann cells. Thus, at 12months after the SCC, most axons of various diameters
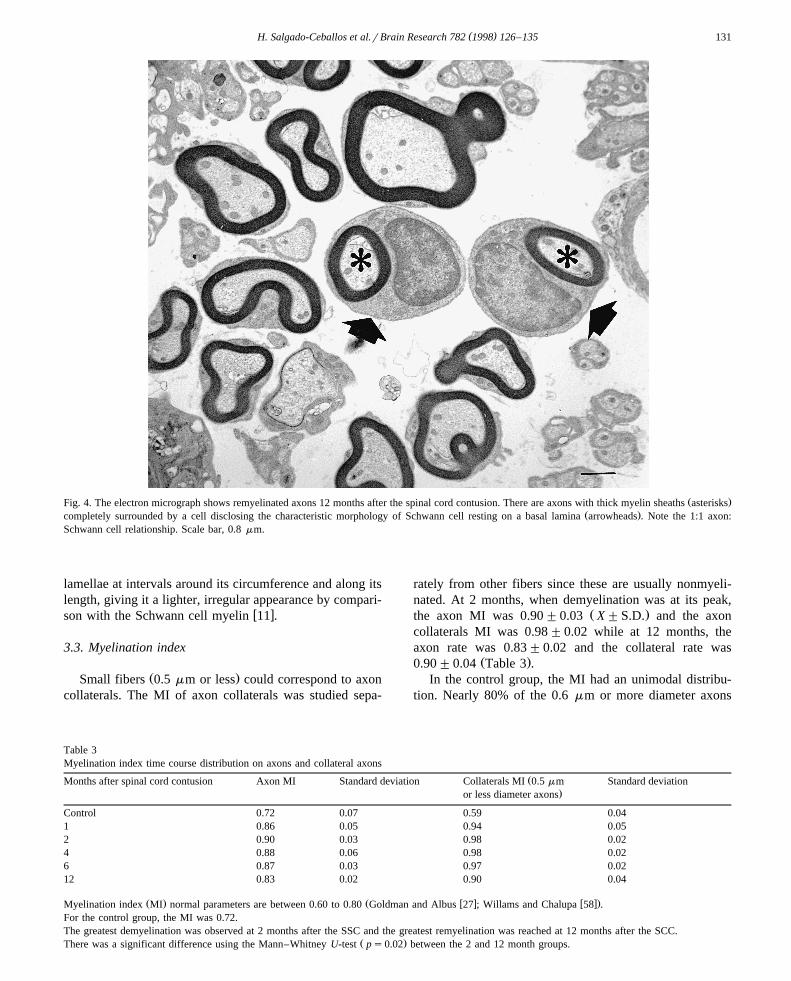
had thick myelin sheaths characteristic of those producedŽ .by Schwann cells Fig. 4 and most small, nonmyelinated
axons were sheathed by the cytoplasm of those cells or byastrocytic processes.
In this paper, the ratio of Schwann cell or oligodendro-cyte myelinated axons was about 8:1, 1 month after theSCC while at 12 months, it was 23:1 in one case. In theother two cases myelination was achieved only by Schwanncells.
Large axons were myelinated by Schwann cells in a 1:1ratio, while medium caliber axons were frequently, but notalways, isolated in a separate trough of a Schwann cell,even though the same cell unsheathed other nonmyelinatedaxons and smaller axons were unsheathed together by oneSchwann cell cytoplasmic process.
Schwann cell myelinated axons were identified basedon the fact the Schwann cells produce thicker, more com-pact myelin sheaths than those produced by oligodendro-cytes. The presence of Schwann cell nuclei and theircytoplasm sheathing the axon, the identification of a cyto-plasm ring in the periphery of the Schwann cell and theaxon it is myelinating, and the displacement of the CNS
w xglia 11,46 . On the other hand, oligodendrocyte myeli-nated axons were identified by a typically ‘puffing’ of the
ŽFig. 3. Electron micrograph of the injury site 6 months after the lesion that shows some thin CNS-type myelin sheaths relative to axonal diameter large. Ž . Ž .arrows and some astrocytic processes arrowheads surrounding naked axons small arrows . Scale bar, 1.2 mm.
( )H. Salgado-Ceballos et al.rBrain Research 782 1998 126–135 131
Ž .Fig. 4. The electron micrograph shows remyelinated axons 12 months after the spinal cord contusion. There are axons with thick myelin sheaths asterisksŽ .completely surrounded by a cell disclosing the characteristic morphology of Schwann cell resting on a basal lamina arrowheads . Note the 1:1 axon:
Schwann cell relationship. Scale bar, 0.8 mm.
lamellae at intervals around its circumference and along itslength, giving it a lighter, irregular appearance by compari-
w xson with the Schwann cell myelin 11 .
3.3. Myelination index
Ž .Small fibers 0.5 mm or less could correspond to axoncollaterals. The MI of axon collaterals was studied sepa-
rately from other fibers since these are usually nonmyeli-nated. At 2 months, when demyelination was at its peak,
Ž .the axon MI was 0.90"0.03 X"S.D. and the axoncollaterals MI was 0.98"0.02 while at 12 months, theaxon rate was 0.83"0.02 and the collateral rate was
Ž .0.90"0.04 Table 3 .In the control group, the MI had an unimodal distribu-
tion. Nearly 80% of the 0.6 mm or more diameter axons
Table 3Myelination index time course distribution on axons and collateral axons
ŽMonths after spinal cord contusion Axon MI Standard deviation Collaterals MI 0.5 mm Standard deviation.or less diameter axons
Control 0.72 0.07 0.59 0.041 0.86 0.05 0.94 0.052 0.90 0.03 0.98 0.024 0.88 0.06 0.98 0.026 0.87 0.03 0.97 0.0212 0.83 0.02 0.90 0.04
Ž . Ž w x w x.Myelination index MI normal parameters are between 0.60 to 0.80 Goldman and Albus 27 ; Willams and Chalupa 58 .For the control group, the MI was 0.72.The greatest demyelination was observed at 2 months after the SSC and the greatest remyelination was reached at 12 months after the SCC.
Ž .There was a significant difference using the Mann–Whitney U-test ps0.02 between the 2 and 12 month groups.
( )H. Salgado-Ceballos et al.rBrain Research 782 1998 126–135132
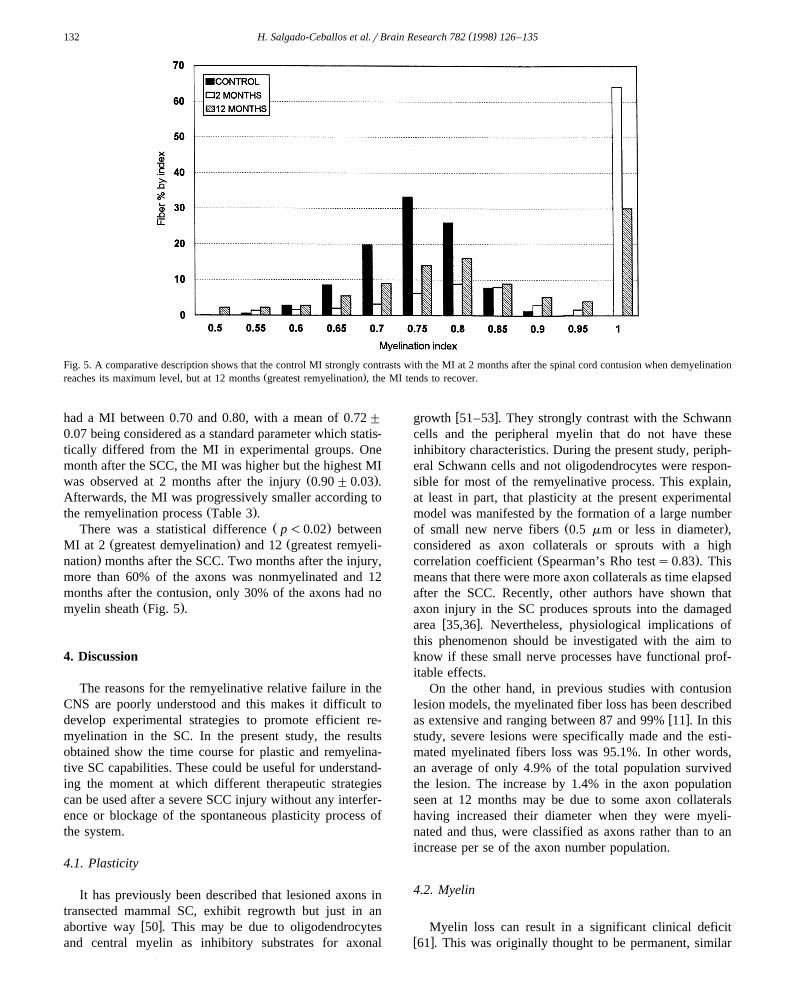
Fig. 5. A comparative description shows that the control MI strongly contrasts with the MI at 2 months after the spinal cord contusion when demyelinationŽ .reaches its maximum level, but at 12 months greatest remyelination , the MI tends to recover.
had a MI between 0.70 and 0.80, with a mean of 0.72"
0.07 being considered as a standard parameter which statis-tically differed from the MI in experimental groups. Onemonth after the SCC, the MI was higher but the highest MI
Ž .was observed at 2 months after the injury 0.90"0.03 .Afterwards, the MI was progressively smaller according to
Ž .the remyelination process Table 3 .Ž .There was a statistical difference p-0.02 between
Ž . ŽMI at 2 greatest demyelination and 12 greatest remyeli-.nation months after the SCC. Two months after the injury,
more than 60% of the axons was nonmyelinated and 12months after the contusion, only 30% of the axons had no
Ž .myelin sheath Fig. 5 .
4. Discussion
The reasons for the remyelinative relative failure in theCNS are poorly understood and this makes it difficult todevelop experimental strategies to promote efficient re-myelination in the SC. In the present study, the resultsobtained show the time course for plastic and remyelina-tive SC capabilities. These could be useful for understand-ing the moment at which different therapeutic strategiescan be used after a severe SCC injury without any interfer-ence or blockage of the spontaneous plasticity process ofthe system.
4.1. Plasticity
It has previously been described that lesioned axons intransected mammal SC, exhibit regrowth but just in an
w xabortive way 50 . This may be due to oligodendrocytesand central myelin as inhibitory substrates for axonal
w xgrowth 51–53 . They strongly contrast with the Schwanncells and the peripheral myelin that do not have theseinhibitory characteristics. During the present study, periph-eral Schwann cells and not oligodendrocytes were respon-sible for most of the remyelinative process. This explain,at least in part, that plasticity at the present experimentalmodel was manifested by the formation of a large number
Ž .of small new nerve fibers 0.5 mm or less in diameter ,considered as axon collaterals or sprouts with a high
Ž .correlation coefficient Spearman’s Rho tests0.83 . Thismeans that there were more axon collaterals as time elapsedafter the SCC. Recently, other authors have shown thataxon injury in the SC produces sprouts into the damaged
w xarea 35,36 . Nevertheless, physiological implications ofthis phenomenon should be investigated with the aim toknow if these small nerve processes have functional prof-itable effects.
On the other hand, in previous studies with contusionlesion models, the myelinated fiber loss has been described
w xas extensive and ranging between 87 and 99% 11 . In thisstudy, severe lesions were specifically made and the esti-mated myelinated fibers loss was 95.1%. In other words,an average of only 4.9% of the total population survivedthe lesion. The increase by 1.4% in the axon populationseen at 12 months may be due to some axon collateralshaving increased their diameter when they were myeli-nated and thus, were classified as axons rather than to anincrease per se of the axon number population.
4.2. Myelin
Myelin loss can result in a significant clinical deficitw x61 . This was originally thought to be permanent, similar

( )H. Salgado-Ceballos et al.rBrain Research 782 1998 126–135 133
to axonal destruction. Currently, it is well know that thew xCNS retains its capacity to remyelinate axons 19,39,44
and some workers in the field sustain that even in severeSC injury, some recovery is the rule rather than the
w xexception 16,17,61 which could involve myelin regenera-w xtion 60 . However, the process requires a long period ofŽ .time at least a year in the present study and it may be due
to either the presence of inhibitory elements preventingmyelin repair or the absence of cells or factors necessary
w xfor new myelin synthesis 14,39,44,45,48 .In the peripheral nervous system, remyelination by
Schwann cells takes place rapidly and efficiently, stronglycontrasting with the inability of the CNS oligodendrocytes
w xto remyelinate under most circumstances 11,34 . AlthoughSchwann cells are able to remyelinate central axons, it iswell known that their migration is restricted by astrocytesw x12 , and eventually astrocytes seem to be responsible for
w xtheir expulsion from the CNS parenchyma 2,7,20 . How-ever, during the current study the most frequently myelina-tion pattern observed was characteristic of that producedby Schwann cells. Furthermore, remyelination by Schwanncells took place throughout all the experimental time courseand even at 12 months after the injury. This may be due tothe fact that the lesion was so severe that the breakage ofthe blood brain barrier cannot be totally repaired by theastrocytes, which leads to a continue Schwann cells migra-tion to the SC.
The Schwann cell myelination pattern was identified bythe presence of a thin cytoplasmic rim around the entireperimeter of the ensheathed axon, the presence of thenucleus and cytoplasm in the sheath surrounding the axon
w xand a basal lamina around the Schwann cells 46 . Further-more, Schwann cell myelinated fibers were usually sur-rounded by a clear extracellular space contrasting witholigodendrocytes myelinated fibers usually embedded in a
w xmore or less dense astrocytic field 11 .
4.3. Myelination index
The MI has been measured in different nerve structuresof several species and it appears that the myelinated fibershave an appropriate conduction when the MI is between0.60 and 0.80 with specific variations between differentspecies and between different nerve structures in the same
w xspecies 27,58 . In the present study, the MI obtained fromthe control group was 0.72"0.07. Two months after thelesion when the greatest demyelination was observed, theMI was far from the standard parameters. However, at 12months when remyelination by Schwann cells was impor-tant, the MI obtained was in close proximity to the MInecessary for appropriate nerve conduction.
On the other hand, it has been shown that the thicknessof the myelin sheath in animals without lesions increaseswith the enlargement of the axon diameter, maintaining a
wrelatively constant MI for fibers of all dimensions 22–
x24,29 . However, we found that some small-diameter ax-ons had myelin sheaths as thick as those seen in somelarge axons. This could be explained by the fact thatSchwann cells produce myelin sheaths thicker than oligo-dendrocytes do and in the present study most of the axonswere myelinated by Schwann cells. Furthermore, severallarge diameter axons had thin sheaths of myelin character-istic of smaller fibers which may be explained in termsthat the remyelination at this specific moment could havebeen partial. In the same way, the decrease in the propor-tion of small axons 6 months after the SCC could beexplained because when an axon is myelinated, its diame-
w xter increases 59 .According to our results, we do not feel the MI recov-
ery trend is due to an increase in the total number of axonsor in their corresponding MI. As previously discussed,1.4% of the axon population growth seen at 12 monthsmay be due to some myelinated axon collaterals increasedtheir diameter and thus being classified as axons ratherthan an increase per se in the number of axons. Neitherdoes it seem likely that this recovery has directly dependedon the increased myelin sheath thickness without increas-ing the number of myelinated axons since in axon popula-tions, very similar in number, the percentage of demyeli-nated fibers 2 months after the SCC was 64% and at 12months it was 30%. Hence, and because the rate ofSchwann cell myelinated axons was higher by far than therate of oligodendrocyte myelinated axons, and in somecases, only Schwann cells were seen to be the cause of thisprocess, we feel the increased number of Schwann cellmyelinated axons was responsible for the MI recoverytrend.
The results obtained in this study differ from those ofother authors that assert that remyelination in the CNS isabortive and limited. The differences can be explained onthe basis that some studies had rather subjective demyeli-
w xnation and remyelination evaluations 4,38 , while othersw xhad short experimental course times 17,25 , were carried
w xout in different species models 10,37 or evaluated differ-w xent pathologic processes 15,18,47,56 . Nonetheless, these
are CNS demyelinating pathologies too.In fact, functional recovery after a CNS lesion requires
w xfor the myelin to regenerate 60 , so with the aim to designrationale therapeutic strategies, we have to clearly under-stand the spontaneous regenerative mechanisms of the SCand not negatively interfere with them.
Currently, there are many experimental therapeutic pro-posals for the therapeutic SC injury bandling. Nonetheless,there is a need to develop strategies designed to hastenmyelin repair. One could consist in diminishing the initialdamage to the myelin sheaths with pharmacological pro-tecting treatments and the other, could be in increasing theinherent remyelinative capability of the system using en-riched transplants with the intention of repairing the dam-aged myelin sheaths, but considering the spontaneous plas-tic and regenerative processes of the SC.

( )H. Salgado-Ceballos et al.rBrain Research 782 1998 126–135134
Acknowledgements
The authors wish to thank Armando Zepeda, ElenaOliva, Adrian Rondan, Silvia Antuna and Gerardo Torresfor their help with the electron microscope, Manuel Perez´for his help with computational programs and Tomas Cruz´and Mario Garcıa for their photographic work. Hermelinda´Salgado-Ceballos holds a scholarship from The National
Ž .Council of Science and Technology CONACYT , contractnumber 80959.
References
w x1 A.R. Allen, Surgery of experimental lesion of spinal cord equivalentto crush injury of fracture dislocation of spinal column, J. Am. Med.
Ž .Assoc. 57 911 878–880.w x2 A. Baron-Van Evercooren, A. Gansmuller, E. Clerin-Duhamel, F.¨
Pascal, M. Gumpel, Repair of a myelin lesion by Schwann cellstransplanted in the adult mouse spinal cord, J. Neuroimmunol. 40Ž .1992 234–242.
w x3 W.F. Blakemore, Remyelination by Schwann cells of axons de-myelinated by intraspinal injection of 6-aminonicotinamide in the
Ž .rat, J. Neurocytol. 4 1975 745–757.w x4 W.F. Blakemore, Invasion of Schwann cells into the spinal cord of
the rat following local injections of lysolecithin, Neuropathol. Appl.Ž .Neurobiol. 2 1976 21–39.
w x5 W.F. Blakemore, Remyelination of CNS axons by Schwann cellsŽ . Ž .transplanted from the sciatic nerve, Nature London 266 1977
68–69.w x6 W.F. Blakemore, A.J. Crang, The use of cultured autologous
Schwann cells to remyelinate areas of persistent demyelination inŽ .the central nervous system, J. Neurol. Sci. 70 1985 207–223.
w x7 W.F. Blakemore, A.J. Crang, R. Curtis, The interaction of Schwanncells with CNS axons in regions containing normal astrocytes, Acta
Ž . Ž .Neuropathol. Berlin 71 1986 295–300.w x8 A.R. Blight, Cellular morphology of chronic spinal cord injury in the
cat: analysis of myelinated axons by line sampling, Neuroscience 10Ž .1983 521–543.
w x9 A.R. Blight, Delayed demyelination and macrophage invasion: acandidate for secondary cell damage in spinal cord injury, Cent.
Ž .Nerv. Syst. Trauma. 2 1985 299–315.w x10 A.R. Blight, V. Decrescito, Morphometric analysis of experimental
spinal cord injury in the cat: the relation of injury intensity toŽ .survival of myelinated axons, Neuroscience 19 1986 321–341.
w x11 A.R. Blight, W. Young, Central axons in injured cat spinal cordrecover electrophysiological function following remyelination by
Ž .Schwann cells, J. Neurol. Sci. 91 1989 15–34.w x12 M.B. Bunge, V.R. Holets, M.L. Bates, T.S. Clarke, B.D. Watson,
Characterization of photochemically induced spinal cord injury in ratŽ .by light and electron microscopy, Exp. Neurol. 127 1994 76–93.
w x13 M.C. Dal Canto, R.L. Barbano, Remyelination during remission inŽ .Theiler’s virus infection, Am. J. Pathol. 116 1984 30–45.
w x14 M.C. Dal Canto, H.L. Lipton, Schwann cell remyelination andrecurrent demyelination in the central nervous system of miceinfected with attenuated Theiler’s virus, J. Neuropathol. Exp. Neu-
Ž .rol. 101 1980 101–109.w x15 J.C. De La Torre, P.K. Hill, M. Gonzalez-Carvajal, J.C. Parker,
Evaluation of transected spinal cord regeneration in the rat, Exp.Ž .Neurol. 84 1984 188–206.
w x16 T.B. Ducker, J.T. Lucas, T.A. Wallace, Recovery from spinal cordŽ .injury, Clin. Neurosurg. 30 1983 495–513.
w x Ž .17 T.B. Ducker, C.A. Walleck Eds. , Recovery from cord injury, in:Central Nervous System Trauma Status Report-1985, Bethesda,NIH-NINCDS, 1986, pp. 369–374.
w x18 I.D. Duncan, A.J. Aguayo, R.P. Bunge, P.M. Wood, Transplantationcord: transplantation of rat Schwann cells grown in tissue culture
Ž .into the mouse spinal cord, J. Neurol. Sci. 49 1981 241–252.w x19 I.D. Duncan, E.A. Milward, Glial cell transplants: experimental
Ž .therapies of myelin diseases, Brain Pathol. 5 1995 301–310.w x20 I. Dusart, S. Marty, M. Peschanski, Demyelination and remyelina-
tion by Schwann cells and oligodendrocytes after kainate inducedneuronal depletion in the central nervous system, Neuroscience 51Ž .1992 137–148.
w x21 I. Dusart, M.E. Schwab, Secondary cell death and the inflammatoryreaction after dorsal hemisection of the rat spinal cord, Eur. J.
Ž .Neurosci. 6 1994 712–724.w x22 J.P. Fraher, The growth and myelination of central and peripheral
segments of ventral motor neuron axons. A quantitative ultrastruc-Ž .tural study, Brain Res. 105 1976 193–211.
w x23 J.P. Fraher, D. O’Leary, M.A. Moran, M. Cole, R.H.M. King, P.K.Thomas, Relative growth and maturation of axon size and myelin
Ž .thickness in the tibial nerve of the rat, Acta Neuropathol. 79 1990364–374.
w x24 R.L. Friede, W. Beuche, A new approach toward analyzing periph-eral nerve fiber populations: I. Variance in sheath thickness corre-sponds to different geometric proportions of the internodes, J.
Ž .Neuropathol. Exp. Neurol. 44 1985 60–72.w x25 J. Gilson, W.F. Blakemore, Failure of remyelination in areas of
demyelination produced in the spinal cord of old rats, Neuropathol.Ž .Appl. Neurobiol. 19 1993 173–181.
w x26 R.F. Gledhill, B.M. Harrison, W.L. Mc Donald, Demyelination andremyelination after acute spinal cord compression, Exp. Neurol. 38Ž .1973 472–487.
w x27 L. Goldman, J.S. Albus, Computation of impulse conduction inmyelinated fibers: theoretical basis of the velocity–diameter relation,
Ž .Biophys. J. 8 1968 596–607.w x28 G. Guizar-Sahagun, I. Grijalva, I. Madrazo, R. Franco-Bourland, H.
Salgado, A. Ibarra, E. Oliva, A. Zepeda, Development of post-traumatic cyst in the spinal cord of rats subjected to severe spinal
Ž .cord contusion, Surg. Neurol. 41 1994 241–249.w x29 J. Guy, A.E. Ellis, K. Kelley, G.M. Hope, Spectra of G ratio, myelin
sheath thickness, and axon and fiber diameter in the guinea pig opticŽ .nerve, J. Comp. Neurol. 287 1989 446–454.
w x Ž .30 A.F. Hahn, Experimental allergic neuritis EAN as a model for theŽ .immune-mediated demyelinating neuropathies, Rev. Neurol. Paris
Ž .152 1996 328–332.w x31 B.M. Harrison, Remyelination by cells introduced into a stable
Ž .demyelinating lesion in the CNS, J. Neurol. Sci. 46 1980 63–81.w x32 A. Hirano, Review of the morphological aspects of remyelination,
Ž .Dev. Neurosci. 11 1989 112–117.w x33 Y. Itoyama, A. Ohnishi, J. Tateishi, Y. Koroiwa, H.D. Webster,
Spinal cord multiple sclerosis lesions in japanese patients: schwanncell remyelination occurs in areas that lack glial fibrillary acid
Ž . Ž . Ž .protein GFAP , Acta Neuropathol. Berlin 65 1985 217–223.w x34 P.W. Lampert Jr., S.S. Schochet, Demyelination and remyelination
in lead neuropathy: electron microscopic studies, J. Neuropathol.Ž .Exp. Neurol. 27 1968 527.
w x35 I. Li, G. Raisman, Schwann cells induce sprouting in motor andŽ .sensory axons in the adult rat spinal cord, J. Neurosci. 14 1994
4050–4063.w x36 I. Li, G. Raisman, Sprouts from cut corticospinal axons persist in the
presence of astrocytic scarring in long-term lesions of the adult ratŽ .spinal cord, Exp. Neurol. 134 1995 102–111.
w x37 S.K. Ludwin, Central nervous system demyelination and remyelina-tion in the mouse. An ultraestructural study of cuprizone toxicity,
Ž .Lab. Invest. 39 1978 597–612.w x38 S.K. Ludwin, Chronic demyelination inhibits remyelination in the
central nervous system. An analysis of contributing factors, LabŽ .Invest. 43 1980 382–387.
w x39 S.K. Ludwin, Remyelination in the central nervous system and theŽ .peripheral nervous system, Adv. Neurol. 47 1988 215–254.

( )H. Salgado-Ceballos et al.rBrain Research 782 1998 126–135 135
w x40 S.K. Ludwin, Evolving concepts and issues in remyelination, Dev.Ž .Neurosci. 11 1989 140–148.
w x41 K.F. Lunn, M.L. Fanarraga, I.D. Duncan, Myelin mutants: newŽ .models and new observations, Microscopy Res. Tech. 32 1995
183–203.w x42 F.A. McMorris, R.D. McKinron, Regulation of oligodendrocyte
development and CNS myelination by growth factors: prospects forŽ .therapy of demyelinating disease, Brain Pathol. 6 1996 313–329.
w x43 D.J. Miller, K.S. Sanborn, J.A. Katzmann, M. Rodriguez, Mono-clonal autoantibodies promote central nervous system repair in an
Ž .animal model of multiple sclerosis, J. Neurosci. 14 1994 6230–6238.
w x44 D.J. Miller, K. Asakura, M. Rodriguez, Central nervous systemremyelination. Clinica application of basic neuroscience principles,
Ž .Brain Pathol. 6 1995 331–344.w x45 D.J. Miller, K. Asakura, M. Rodriguez, Experimental strategies to
promote central nervous system remyelination in multiple sclerosis:insights gained from the Theiler’s virus model system, J. Neurosci.
Ž .Res. 41 1995 291–296.w x46 A. Peters, S.C. Palay, H. de F. Webster, Oligodendrocytes in the
Fine Structure of the Nervous System, 3rd edn., Oxford Univ. Press,New York, 1991, pp. 295–302.
w x47 J.W. Prineas, The neuropathology of multiple sclerosis, in: J.C.Ž .Koetsier Ed. , Handbook of Clinical Neurology. Demyelinating
Diseases, Elsevier, Amsterdam, 1985, pp. 213–257.w x48 C.S. Raine, Morphology of myelin and myelination, in: P. Morell
Ž .Ed. , Myelin, Plenum, New York, 1984, pp. 1–41.w x49 C.S. Raine, E. Wu, Multiple sclerosis: remyelination in acute le-
Ž .sions, J. Neuropathol. Exp. Neurol. 52 1993 199–204.w x50 S. Ramon y Cajal, Degeneration and regeneration of the nervous´
Ž .system, in: R.M. May Ed. , Oxford Univ. Press, London, 1928.w x51 M.E. Schwab, P. Carori, Oligodendrocytes and CNS myelin are
nonpermissive substrates for neurite growth and fibroblast spreadingŽ .in vitro, J. Neurosci. 8 1988 2381–2393.
w x52 M.E. Schwab, Myelin-associated inhibitors of neurite growth, Exp.Ž .Neurol. 109 1990 2–5.
w x53 M.E. Schwab, J.P. Kapfhammer, C.E. Bandtlow, Inhibitors of neu-Ž .rite growth, Annu. Rev. Neurosci. 16 1993 565–595.
w x54 M. Taniike, K. Suzuki, Proliferative capacity of oligodendrocytes inŽ .the demyelinating twitcher spinal cord, J. Neurosci. Res. 40 1995
325–332.w x55 L. Vignais, B. Nait Ouesmar, F. Mellouk, O. Gout, G. Labourdette,
A. Baron- Van Evercooren, M. Gumpel, Transplantation of oligo-dendrocyte precursors in the adult demyelinated spinal cord: migra-
Ž .tion and remyelination, Int. J. Dev. Neurosci. 11 1993 603–612.w x56 S.G. Waxman, Membranes, myelin, and the pathophysiology of
Ž .multiple sclerosis, New Engl. J. Med. 306 1982 1529–1533.w x Ž .57 H. de P. Webster, Myelin injury and repair, Adv. Neurol. 59 1993
67–73.w x58 R.W. Williams, L.M. Chalupa, An analysis of axon caliber within
the optic nerve of the cat: evidence of size groupings and regionalŽ .organization, J. Neurosci. 3 1983 1554–1564.
w x59 A.J. Windebank, P. Wood, R.P. Bunge, P.J. Dyck, Myelinationdetermines the caliber of dorsal root ganglion neurons in culture, J.
Ž .Neurosci. 5 1985 1563–1569.w x60 D. Xie, R.L. Schultz, E.F. Writter, The oligodendroglial reaction to
brain stab wounds: an immunohistochemical study, J. Neurocytol. 24Ž .1995 435–448.
w x61 W. Young, Recovery mechanisms in spinal cord injury: implicationsŽ .for regenerative therapy, in: F.J. Seil Ed. , Neural Regeneration and
Transplantation, A.R. Liss, New York, 1989, pp. 157–169.w x62 Z. Zhang, M. Fujiki, L. Guth, O. Steward, Genetic influences on
cellular reactions to spinal cord injury:a wound-healing responsepresent in normal mice is impaired in mice carrying a mutationŽ s.Wld that causes delayed wallerian degeneration, J. Comp. Neurol.
Ž .371 1996 485–495.
Related Documents











