PLANT-ANIMAL INTERACTIONS - ORIGINAL PAPER Ants on plants: a meta-analysis of the role of ants as plant biotic defenses Felix B. Rosumek Fernando A. O. Silveira Frederico de S. Neves Newton P. de U. Barbosa Livia Diniz Yumi Oki Flavia Pezzini G. Wilson Fernandes Tatiana Cornelissen Received: 14 May 2008 / Accepted: 4 February 2009 / Published online: 7 March 2009 Ó Springer-Verlag 2009 Abstract We reviewed the evidence on the role of ants as plant biotic defenses, by conducting meta-analyses for the effects of experimental removal of ants on plant herbivory and fitness with data pooled from 81 studies. Effects reviewed were plant herbivory, herbivore abundance, hemipteran abundance, predator abundance, plant biomass and reproduction in studies where ants were experimentally removed (n = 273 independent comparisons). Ant removal exhibited strong effects on herbivory rates, as plants without ants suffered almost twice as much damage and exhibited 50% more herbivores than plants with ants. Ants also influenced several parameters of plant fitness, as plants without ants suffered a reduction in biomass (-23.7%), leaf production (-51.8%), and reproduction (-24.3%). Effects were much stronger in tropical regions compared to tem- perate ones. Tropical plants suffered almost threefold higher herbivore damage than plants from temperate regions and exhibited three times more herbivores. Ant removal in tropical plants resulted in a decrease in plant fitness of about 59%, whereas in temperate plants this reduction was not statistically significant. Ant removal effects were also more important in obligate ant–plants (=myrmecophytes) com- pared to plants exhibiting facultative relationships with hemiptera or those plants with extrafloral nectaries and food bodies. When only tropical plants were considered and the strength of the association between ants and plants taken into account, plants with obligate association with ants exhibited almost four times higher herbivory compared to plants with facultative associations with ants, but similar reductions in plant reproduction. The removal of a single ant species increased plant herbivory by almost three times compared to the removal of several ant species. Altogether, these results suggest that ants do act as plant biotic defenses, but the effects of their presence are more pronounced in tropical systems, especially in myrmecophytic plants. Keywords Ant–plant interaction Ant–plant mutualism Formicidae Herbivory Indirect interactions Introduction Interactions between ants and plants are both ancient and widespread (Davidson and McKey 1993; Delabie et al. 2003; Rico-Gray and Oliveira 2007). The pioneering study of Janzen (1966) with Acacia trees and Pseudomyrmex ants showed that ants could act as biotic defenses, protecting plants against herbivores and parasites. In return, plants offer benefits such as shelter and food rewards. Currently, many plant species are known to engage in this ‘biological warfare’, which is often recognized as a mutualistic inter- action (Bronstein et al. 2006). Communicated by Bernhard Stadler. N. P. de U. Barbosa, L. Diniz, Y. Oki and F. Pezzini contributed equally to this work and are listed in alphabetical order. Electronic supplementary material The online version of this article (doi:10.1007/s00442-009-1309-x) contains supplementary material, which is available to authorized users. F. B. Rosumek F. A. O. Silveira F. de S. Neves N. P. de U. Barbosa L. Diniz Y. Oki F. Pezzini G. W. Fernandes Instituto de Cie ˆncias Biolo ´gicas, Universidade Federal de Minas Gerais, Belo Horizonte, MG, Brazil T. Cornelissen (&) Faculdade de Cie ˆncias Integradas do Pontal, Universidade Federal de Uberla ˆndia, Avenida Jose ´ Joa ˜o Dib, 2245 Ituiutaba, MG, Brazil e-mail: [email protected]; [email protected] 123 Oecologia (2009) 160:537–549 DOI 10.1007/s00442-009-1309-x

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

PLANT-ANIMAL INTERACTIONS - ORIGINAL PAPER
Ants on plants: a meta-analysis of the role of antsas plant biotic defenses
Felix B. Rosumek Æ Fernando A. O. Silveira Æ Frederico de S. Neves ÆNewton P. de U. Barbosa Æ Livia Diniz Æ Yumi Oki Æ Flavia Pezzini ÆG. Wilson Fernandes Æ Tatiana Cornelissen
Received: 14 May 2008 / Accepted: 4 February 2009 / Published online: 7 March 2009
� Springer-Verlag 2009
Abstract We reviewed the evidence on the role of ants as
plant biotic defenses, by conducting meta-analyses for the
effects of experimental removal of ants on plant herbivory
and fitness with data pooled from 81 studies. Effects
reviewed were plant herbivory, herbivore abundance,
hemipteran abundance, predator abundance, plant biomass
and reproduction in studies where ants were experimentally
removed (n = 273 independent comparisons). Ant removal
exhibited strong effects on herbivory rates, as plants without
ants suffered almost twice as much damage and exhibited
50% more herbivores than plants with ants. Ants also
influenced several parameters of plant fitness, as plants
without ants suffered a reduction in biomass (-23.7%), leaf
production (-51.8%), and reproduction (-24.3%). Effects
were much stronger in tropical regions compared to tem-
perate ones. Tropical plants suffered almost threefold higher
herbivore damage than plants from temperate regions and
exhibited three times more herbivores. Ant removal in
tropical plants resulted in a decrease in plant fitness of about
59%, whereas in temperate plants this reduction was not
statistically significant. Ant removal effects were also more
important in obligate ant–plants (=myrmecophytes) com-
pared to plants exhibiting facultative relationships with
hemiptera or those plants with extrafloral nectaries and food
bodies. When only tropical plants were considered and the
strength of the association between ants and plants taken into
account, plants with obligate association with ants exhibited
almost four times higher herbivory compared to plants with
facultative associations with ants, but similar reductions in
plant reproduction. The removal of a single ant species
increased plant herbivory by almost three times compared to
the removal of several ant species. Altogether, these results
suggest that ants do act as plant biotic defenses, but the
effects of their presence are more pronounced in tropical
systems, especially in myrmecophytic plants.
Keywords Ant–plant interaction � Ant–plant mutualism �Formicidae � Herbivory � Indirect interactions
Introduction
Interactions between ants and plants are both ancient and
widespread (Davidson and McKey 1993; Delabie et al.
2003; Rico-Gray and Oliveira 2007). The pioneering study
of Janzen (1966) with Acacia trees and Pseudomyrmex ants
showed that ants could act as biotic defenses, protecting
plants against herbivores and parasites. In return, plants
offer benefits such as shelter and food rewards. Currently,
many plant species are known to engage in this ‘biological
warfare’, which is often recognized as a mutualistic inter-
action (Bronstein et al. 2006).
Communicated by Bernhard Stadler.
N. P. de U. Barbosa, L. Diniz, Y. Oki and F. Pezzini contributed
equally to this work and are listed in alphabetical order.
Electronic supplementary material The online version of thisarticle (doi:10.1007/s00442-009-1309-x) contains supplementarymaterial, which is available to authorized users.
F. B. Rosumek � F. A. O. Silveira � F. de S. Neves �N. P. de U. Barbosa � L. Diniz � Y. Oki � F. Pezzini �G. W. Fernandes
Instituto de Ciencias Biologicas, Universidade Federal de Minas
Gerais, Belo Horizonte, MG, Brazil
T. Cornelissen (&)
Faculdade de Ciencias Integradas do Pontal,
Universidade Federal de Uberlandia, Avenida Jose Joao Dib,
2245 Ituiutaba, MG, Brazil
e-mail: [email protected]; [email protected]
123
Oecologia (2009) 160:537–549
DOI 10.1007/s00442-009-1309-x

Ant–plant associations occur between species in most ant
subfamilies and plant species in two fern families and
innumerous phylogenetically diverse angiosperms (David-
son and McKey 1993), having evolved independently many
times (Rico-Gray and Oliveira 2007). These ant–plant
interactions are based on the array of resources provided by
plants as rewards ranging from food bodies (Fiala et al.
1989; Dutra et al. 2006), extra-floral nectaries (=EFNs,
Oliveira 1997; Rudgers 2004; Koptur 2005) or domatia
(nesting sites), the last produced by plant species called
‘‘myrmecophytes’’ (Fonseca 1994). Indirect ant–plant
interactions can also be mediated by honeydew-producing
‘‘hemipterans’’ (Order Hemiptera: Sternorryncha and
Auchenorryncha) (Compton and Robertson 1988; Crut-
singer and Sanders 2005). In return, ants deter or prey upon
insects and vertebrate herbivores and prune encroaching
vines, increasing plant fitness. However, attractor number,
identity, function, and position on the plant both directly
and indirectly influence ant recruitment and there might be
great variation in ant protection (Table 1).
Two strategies can be distinguished within defensive
ant–plant interactions. Ant–plants, or myrmecophytes, are
continuously inhabited by ants during major parts of their
life (Webber et al. 2007). Myrmecophytic plants occur in
some species of Acacia, Cecropia, Leonardoxa, Piper,
Tococa, and Macaranga, among others, and provide nesting
sites permanently inhabited by colonies of specialized ants
which protect them in a more intimate and specific associ-
ation (Davidson and McKey 1993; Heil and McKey 2003;
Rico-Gray and Oliveira 2007). Myrmecophilic plants on
the other hand are plants that, whilst providing direct
food resources that can be utilized by ants, are not regu-
larly occupied by ant colonies (Webber et al. 2007),
Myrmecophilous species offer unspecialized rewards to
attract ants, mostly in the form of extrafloral nectar or
through indirect associations with honeydew-producing
hemipterans, and gain protection from a facultative and
opportunistic ant community. Because the costs imposed
by ant-associated hemipterans may or may not be out-
weighed by the benefits that hemipteran-tending ants confer
in protecting against non-hemipteran herbivores, hemipt-
eran-mediated ant–plant associations are among the most
facultative, opportunistic, and variable of interactions
(Rico-Gray and Oliveira 2007), with a high level of uncer-
tainty for the outcomes of tending ants to host plants
(Table 1).
Since Janzen’s (1966, 1967) studies on the role of ants as
plant biotic defenses, several works have addressed how
ants protect plants by reducing herbivory rates (Bruna et al.
2004; Del-Claro et al. 2006), herbivore abundance
(Letourneau and Barbosa 1999) and richness (Crutsinger
and Sanders 2005) and ultimately, how ants increase plant
fitness by increasing plant biomass (Messina 1981), leaf
production (Freitas et al. 2000), and flower, fruit and/or
seed production (Del-Claro et al. 1996; Letourneau 1998).
Although several studies have now suggested the generality
of the pattern described by Janzen (Heil et al. 2001;
Michelangeli 2003; Dejean et al. 2006), some other studies
have found no effect of ants as plant biotic defenses (Freitas
et al. 2000) and others have even shown that ant presence
has caused negative effects to host plants (Freitas et al. 2000;
Ruhren 2003; Renault et al. 2005; Frederickson and Gordon
2007; Mooney 2007; see also Rico-Gray and Oliveira 2007).
The effects of ants as plant defenders have been
reviewed previously (Davidson and McKey 1993; Bron-
stein 1998; Heil and McKey 2003; Del-Claro 2004;
Table 1 Expected effects of ant removal under several conditions
Comparison Stronger effects Weaker effects Underlying hypothesis
Temperate 9 tropical Tropical Temperate Higher ant and herbivore diversity in the tropics, higher
herbivore specificity and ant aggressive behavior,
higher frequency of myrmecophytic plants
Single 9 several ants Single Several Exclusion of a single ant species would be more
detrimental to plant as usually single ants are
involved in obligate relationships with plants and/or
are dominant species in ant mosaics
Obligate 9 facultative association Obligate Facultative Myrmecophytic plants (=obligate associations with
ants) rely on ants for protection, whereas non-
myrmecophytic plants benefit from the relationship,
but there is no strong dependency
Ant attractors Multiple ant attractors Hemiptera Multiple ant attractors should provide most of the
energetic needs of ants, keeping colonies patrolling
plants for longer periods of time. In association with
hemipterans, however, benefits should surpass
damage caused by sucking activities
Attractor position Leaf blade Stem Attractor position on the plant might affect the types of
tissues that ants protect and the rate of recruitment
538 Oecologia (2009) 160:537–549
123

Oliveira and Freitas 2004; Bronstein et al. 2006; Rico-Gray
and Oliveira 2007), but all these reviews have been qual-
itative in nature, limiting our ability to draw general
conclusions regarding the real role of ants as plant defenses
and to what extent they increase plant fitness. In a recent
review of the consequences of interactions between ants
and honeydew-producing insects, Styrsky and Eubanks
(2007) calculated the percentage change in plant damage
and fitness from 30 studies. This approach, although
quantitative, does not take into account differences in
sample sizes and variance among studies, limiting the use
of statistical tests to examine whether ants significantly
augment their host plant fitness. Moreover, it does not
allow statistical comparisons between categories desig-
nated by the researcher. For instance, given the enormous
latitudinal differences in both diversity and productivity
between temperate and tropical habitats, it is likely that
relationships among trophic levels may also be funda-
mentally different (Dyer and Coley 2002), with different
outcomes resulting from ant–plant interactions (Table 1) in
different regions. In this study, we used meta-analysis to
review the evidence of the role of ants as plant biotic
defenses under several circumstances.
Schemske (1982) pointed out that loose or facultative
ant–plant mutualisms usually involve several species of
ants, often from several subfamilies. In contrast, myrm-
ecophytes are often associated with a single, specialized
species with often pronounced aggressive behavior (Djieto-
Lordon et al. 2007). The latter are called plant–ants (sensu
Webber et al. 2007) and their traits include colony foun-
dation in a particular host plant; a somewhat strongly
developed host specificity; host fidelity; high occupancy
rates, and intraspecific competition for host plants. In the
best-known ant–plant systems, specialized ant species differ
in their protective effects on host plants (Rico-Gray and
Oliveira 2007). Facultative ant species provide less effec-
tive or no defense in the Acacia–Pseudomyrmex system;
levels of food body production differ between facultative
and obligatorily ant-associated Macaranga species, which
may reflect variation in the degree of specialization and
intensity of interaction with Crematogaster. Finally, obli-
gate associated Azteca are considered competitively
superior to nonobligate ants in Cecropia. Therefore, spe-
cialized plant-dwelling ants are expected to provide better
defenses when compared to opportunistic species (Table 1).
Although the effects of ants as terrestrial top predators
and the effects of predators on plant damage and biomass
have been previously reviewed through meta-analyses of
trophic cascades (Halaj and Wise 2001; Schmitz et al.
2000), to the best of our knowledge, a deeper and specific
meta-analysis of the protective role of ants on plants has
yet to be done. Therefore, we aimed to use meta-analytical
methods to: (1) determine the magnitude of the effect of
ant presence on herbivory rates and parameters of plant
fitness, (2) investigate whether there are differences in the
magnitude of those effects in tropical versus temperate
regions, (3) compare the magnitude of the effects of
removal of single versus several ant species, (4) compare
the magnitude of the effects in relationship to different ant
attractors, attractor combination and number as well as
attractor position on the plant, and (5) compare the mag-
nitude of effects on obligate ant–plants (=myrmecophytes)
and facultative (=myrmecophile) relationships between
ants and host plants, in both tropical and temperate regions.
Materials and methods
The database
This meta-analytical review was based on published stud-
ies searched electronically on the Science Citation Index
Expanded (1945–2008), using ‘‘ants’’, ‘‘herbivory’’, ‘‘ant
protection’’, ‘‘ant mutualism’’, ‘‘tri-trophic interactions’’
and ‘‘ant–plant interactions’’ as keywords. We also sur-
veyed the reference list of main reviews of the role of ants
as plant biotic defenses (Delabie 2001; Oliveira and Freitas
2004; Bronstein et al. 2006; Styrsky and Eubanks 2007)
and reviews of terrestrial trophic cascades (Halaj and Wise
2001; Schmitz et al. 2000). The literature list was finally
supplemented with studies cited in the reference lists of the
articles surveyed.
Studies reviewed
To be included in our review, some criteria had to be met
by a study, such as: (1) studies that were published in
English language, (2) studies in which ants were experi-
mentally removed from the plants, creating control (with
ants) and treatment (without ants) groups, and (3) studies
that reported data with means, sample size, and a measure
of variance (standard deviation, standard error or confi-
dence intervals) for both control and treatment groups.
Studies that reported data with median or reported statis-
tical differences between control and treatment groups by
showing only F values or P values, or studies in which
sample sizes for treatment and control group were not
clear, were excluded from this meta-analytical review.
Also, studies that manipulated only hemipteran densities
(by addition or exclusion) and/or manipulated EFN (by
removal) but did not manipulate ant densities among plants
were not included in our review. Response mean values
( �Xcontrol and �Xtreatment), standard deviations (Scontrol and
Streatment) and sample size (Ncontrol and Ntreatment) were
gathered from the text, tables and/or figures from each
study included in this review. When data were available on
Oecologia (2009) 160:537–549 539
123

figures, these were digitized, and means and measurements
of variance were obtained using the software UTHSCSA
Image Tool (University of Texas, USA) after calibrating
each picture to the nearest 0.01 mm. Measurements of
variance were all converted to standard deviations of the
mean using MetaWin Statistical Calculator (Rosenberg
et al. 2000).
When data were available for several dates (several
years, months, or seasons) or several study sites, the largest
difference between control and treatment group was used
as an independent comparison. When authors used several
treatments (e.g., removal or addition of extra-floral nec-
taries, plant fertilization, removal of pollinators and/or
other predators) but did manipulate ant densities we used
data from the lesser number of inputs.
We conducted separate meta-analyses for each one of
the following nine effects of ant removal on either herbi-
vores or plant features: herbivory; herbivore, hemipteran
and predator abundance; plant biomass; leaf, flower, fruit,
and seed production. Only effects that generated at least
five independent comparisons were included in our analy-
sis. The term ‘‘herbivory’’ encompassed several variables
described by authors, such as plant damage, number of
leaves attacked, and number of leaves lost, among others.
‘‘Herbivore abundance’’ included several variables such as
infestation level, number of eggs, herbivore density, and
number of herbivores. Predators included several arthropod
taxa, but mainly spiders. The effects listed as ‘‘plant bio-
mass’’ comprised plant size, weight, height gain, and
growth. Based on information provided by the authors, we
classified studies according to the region where the study
was done (tropical vs temperate region) and according to
the type of association between ants and plants (obligate vs
facultative). Because myrmecophytes are absent in tem-
perate regions, results could be misleading due to possible
confounding effects of the inclusion of tropical myrmeco-
phytes. So, we conducted separate analyses removing all
myrmecophytes allowing for comparisons of the effects of
ant removal in tropical versus temperate species with loose
or facultative association with ants. In order to investigate
whether there are significant differences in the effects of
ant removal between myrmecophytes versus myrmeco-
philes, analyses were also run including only tropical
species.
Comparisons were also separated according to the
number of ant species (single vs several ants), where
studies that reported two or more ant species simulta-
neously were classified as ‘several’. Single ant removal
refers to cases in which only one ant species was excluded,
whereas ‘several’ refers to the exclusion of all ant species
from plants. For association type, we used data provided by
the authors or scientific literature to classify plants as
myrmecophytic or non-myrmecophytic (sensu Webber
et al. 2007). Only one study (Gaume et al. 2005) recorded a
semi-myrmecophytic plant, and inclusion or exclusion of
this category did not change our results. For ant attractor,
we recorded data as shown by authors, such as aphids,
membracids, honeydew, EFNs, etc., and later categorized
ant attractor into function and not identity, as (1) shelter
(structures produced by plants where ants build nests such
as domatia and hollow trunks), (2) food (plant-derived
rewards such as EFN, food bodies, glandular trichomes),
(3) hemipterans (insect-derived rewards such as honeydew
secretion), and (4) unknown (ant attractor was not descri-
bed on the study and/or there was no evident ant attractor).
Food bodies comprised a variety of rewards such as pearl
bodies, Mullerian bodies, Beltian bodies, and Beccarian
bodies. In many associations, not one but a combination of
rewards may be involved (Rico-Gray and Oliveira 2007),
and we also compared whether multiple attractors were
more efficient against herbivores than a single ant attractor.
For EFN-bearing plants, the position of EFN was also
categorized as leaf blade, petiole, stipule, and stem, or
reproductive structures (Rico-Gray and Oliveira 2007). To
enable evaluation of the ant effects on plant fitness
parameters under the categorical analyses, we grouped
studies that reported flower, fruit, and seed production
under the category ‘‘plant reproduction’’.
Data analysis
We used the log of the response ratio (ln ratio) to sum-
marize the effects of ant presence on both herbivores and
plant features. The response ratio is the ratio of some
measure of outcome in the experimental group to that of
the control group (Rosenberg et al. 2000) and it has the
advantage of estimating the effect as a proportionate
change resulting from experimental manipulation. We
calculated the natural log of the response ratio for each
effect studied as lr = ln �Xwithoutants=�Xcontrolð Þ (Rosenberg
et al. 2000) and effects are reported as the proportional
change from control groups (with ants). Negative per-
centage changes indicate a decrease in the plant and/or
herbivore variable compared to plants with ants and posi-
tive values indicate an increase in the effect measured due
to ant absence. To estimate the cumulative effect size
(E??) for a sample of studies addressing the same effect,
effect sizes were combined across studies using a weighted
average in which the weight for the ith study was the
reciprocal of its sampling variance (Rosenberg et al. 2000).
We used a mixed effect model of meta-analysis, in which it
is assumed that studies within a class share a common
mean effect but that there is also random variation among
studies in a class, in addition to sampling variation. Ninety-
five percent confidence limits around the effect size were
calculated and estimates of the effect sizes were considered
540 Oecologia (2009) 160:537–549
123

significant if the confidence intervals did not overlap with
zero. All analyses were conducted using MetaWin 2.1.3.4
(Rosenberg et al. 2000).
We calculated the total heterogeneity (QT) for all effects
tested and heterogeneity within (QW) and between groups
(QB). The significance of these statistics was evaluated
using a Chi-square distribution. Because our analyses were
based only on published studies, and studies that show
large and significant effects might be more likely to be
published than studies that show weak or no effects (the
‘file-drawer problem’ sensu Rosenthal 1979) we calculated
fail-safe numbers for each effect tested. Fail-safe numbers
indicate the number of non-significant, unpublished, or
missing studies that would need to be added to the sample
in order to change its results from significant to non-sig-
nificant (Rosenberg et al. 2000). As a rule of thumb, fail-
safe results are considered robust if the fail-safe number
exceeds 5k ? 10 (Moller and Jennions 2001), where k is
the number of comparisons in the analysis. We also used
funnel plots as a graphical method to assess publication
bias as, in the absence of bias, a symmetrical ‘funnel’ shape
is formed when the effect size of each study is plotted
against sample size.
Results
Qualitative results
Eighty-one studies regarding the role of ants as biotic
defenses met our criteria for inclusion in the meta-analysis,
generating 273 independent comparisons (Appendix 1).
These studies covered a wide variety of herbaceous and
woody plant taxa, including 86 host plant species in 61
genera. From these host species, two families belonged to
non-seed plant, one is a gymnosperm and 36 are angio-
sperms. Fabaceae (19 spp.) was the most common family,
followed by Euphorbiaceae (11), Salicaceae (6) and Mel-
astomataceae (6). Twenty-seven families were represented
by a single species. Species classification and nomenclature
follow APG II (2003).
Most independent comparisons came from studies con-
ducted in tropical regions (57.9%) versus 42.1% conducted
at temperate latitudes. Among myrmecophiles, 52.5% were
in temperate areas and 47.5% in tropical areas. In tropical
areas, 70.6% of the comparisons included looser association
whereas only 29.3% consisted of obligate associations.
Natural habitats (81.7%) were more studied when compared
to managed systems (18.3%). Extra-floral nectaries were the
most frequent ant attractor, accounting for 34.8% of inde-
pendent comparisons. Hemipterans represented 27.1% of
the ant attractors, followed by a combination of EFN and
hemiptera (7.3%), domatia (6.9%). and less frequent
combinations of food and shelter. Ant attractor was unde-
termined in approximately 10% of the comparisons.
Effects of a total of 20 ant genera were reviewed and ant
genus was undetermined in only 6 comparisons (2.2%).
Nearly 53% of the comparisons addressed the removal of a
particular ant species whereas in 47% of the comparisons
several ant species were excluded. When a single ant genus
was excluded, Formica was the most common genus (25.7%
of independent comparisons), followed by Pheidole (10%),
Solenopsis (10%), and Azteca (9.3%). Other ant genera
comprised 40.5% of the comparisons. Most comparisons
dealt with native ant species (95.2%), whereas only 13
comparisons (4.8%) addressed invasive ants. The insect
herbivores belonged to seven different guilds: chewers,
flower feeder, gall-inducers, miners, sap-suckers, seed pre-
dators, and stem borers, but chewers represented 67.8% of the
independent comparisons reviewed, followed by sap-suckers
(13.6%) and all the other guilds combined represented 13.0%
of all the comparisons tested. Undetermined guilds occurred
in only 5.1% of the independent comparisons.
Amongst the 273 comparisons, more than half (56.0%)
used Tanglefoot� to exclude ants, followed by the use of
insecticides (20.1%). The other 14 mechanisms of ant
exclusion accounted for 23.9% of total comparisons.
Quantitative results
We observed strong effects of ant removal on both her-
bivory rates and plant fitness. Plants without ants suffered
nearly 97% more herbivory than plants with ants
(E?? = 0.972, CI = 0.84–1.09) and exhibited 53.1%
more herbivores than control plants (E?? = 0.5,315,
CI = 0.31–0.75; Fig. 1). We also observed effects of ants
on other predators (mainly spiders), which increased by
more than 100% in plants where ants were excluded
(E?? = 1.04, CI = 0.65–1.44). As expected, a 66.2%
reduction in ‘‘Hemiptera’’ abundance was observed on ant-
excluded plants compared to ant-inhabited plants, although
this result was not significant (CI = -2.13–0.81). Ant
presence also affected plant biomass and plant reproduc-
tion. Ant exclusion increased herbivory rates and
consequently reduced plant biomass by more than 26%
(E?? = -0.264, CI = -0.41 to -0.11) and leaf produc-
tion by more than 50% (E?? = -0.518, CI = -0.76 to
–0.27). Plants without ants produced fewer fruits
(E?? = -0.381, CI = -0.71 to -0.04) and seeds
(E?? = -0.360, CI = -0.50 to -0.22). Contrary to
expectations, a trend for increased flower production was
observed on plants in which ants were removed
(E?? = 0.184, CI = -0.004–0.38), though this result was
not statistically significant.
We observed stronger effects in most parameters of
tropical regions compared to temperate ones (Fig. 2).
Oecologia (2009) 160:537–549 541
123
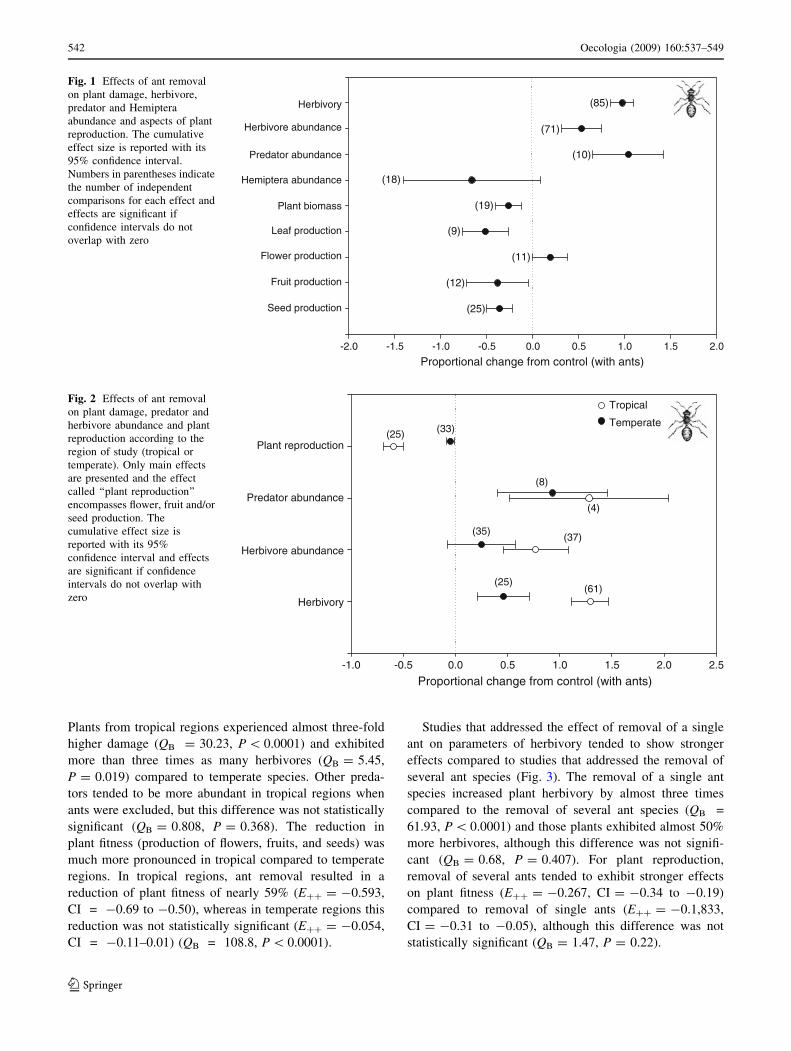
Plants from tropical regions experienced almost three-fold
higher damage (QB = 30.23, P \ 0.0001) and exhibited
more than three times as many herbivores (QB = 5.45,
P = 0.019) compared to temperate species. Other preda-
tors tended to be more abundant in tropical regions when
ants were excluded, but this difference was not statistically
significant (QB = 0.808, P = 0.368). The reduction in
plant fitness (production of flowers, fruits, and seeds) was
much more pronounced in tropical compared to temperate
regions. In tropical regions, ant removal resulted in a
reduction of plant fitness of nearly 59% (E?? = -0.593,
CI = -0.69 to -0.50), whereas in temperate regions this
reduction was not statistically significant (E?? = -0.054,
CI = -0.11–0.01) (QB = 108.8, P \ 0.0001).
Studies that addressed the effect of removal of a single
ant on parameters of herbivory tended to show stronger
effects compared to studies that addressed the removal of
several ant species (Fig. 3). The removal of a single ant
species increased plant herbivory by almost three times
compared to the removal of several ant species (QB =
61.93, P \ 0.0001) and those plants exhibited almost 50%
more herbivores, although this difference was not signifi-
cant (QB = 0.68, P = 0.407). For plant reproduction,
removal of several ants tended to exhibit stronger effects
on plant fitness (E?? = -0.267, CI = -0.34 to -0.19)
compared to removal of single ants (E?? = -0.1,833,
CI = -0.31 to -0.05), although this difference was not
statistically significant (QB = 1.47, P = 0.22).
Proportional change from control (with ants)-2.0 -1.5 -1.0 -0.5 0.0 0.5 1.0 1.5 2.0
Herbivory
Herbivore abundance
Predator abundance
Hemiptera abundance
Plant biomass
Leaf production
Flower production
Fruit production
Seed production
(85)
(71)
(10)
(18)
(19)
(9)
(11)
(25)
(12)
Fig. 1 Effects of ant removal
on plant damage, herbivore,
predator and Hemiptera
abundance and aspects of plant
reproduction. The cumulative
effect size is reported with its
95% confidence interval.
Numbers in parentheses indicate
the number of independent
comparisons for each effect and
effects are significant if
confidence intervals do not
overlap with zero
Proportional change from control (with ants)-1.0 -0.5 0.0 0.5 1.0 1.5 2.0 2.5
Herbivory
Herbivore abundance
Predator abundance
Plant reproduction
(61)(25)
(35)(37)
(8)
(4)
(25) (33)
Tropical
Temperate
Fig. 2 Effects of ant removal
on plant damage, predator and
herbivore abundance and plant
reproduction according to the
region of study (tropical or
temperate). Only main effects
are presented and the effect
called ‘‘plant reproduction’’
encompasses flower, fruit and/or
seed production. The
cumulative effect size is
reported with its 95%
confidence interval and effects
are significant if confidence
intervals do not overlap with
zero
542 Oecologia (2009) 160:537–549
123

When association type was considered, only the effects
of ants on herbivory, herbivore abundance, and plant
reproduction were supported by enough data to allow cat-
egorization. Myrmecophytic plants exhibited four times
higher herbivory after ant removal than myrmecophiles
(E??obligate = 2.32, CI = 2.06–2.57; E??facultative = 0.55,
CI = 0.41–0.69). Moreover, a reduction in plant repro-
duction of approximately 55% (E??obligate = -0.56,
CI = -0.73 to -0.39) compared to 18% in non-myrmec-
ophytic plants was also observed (E??facultative = -0.177,
CI = -0.23 to -0.11). When only facultative (= looser)
associations were considered on each region, tropical
plants without ants still exhibited a stronger increase in
herbivory compared to temperate plants (E??facultative tropical
= 0.64, CI = 0.41–0.86; E??facultative temperate = 0.46,
CI = 0.23–0.69; QB = 27.31, P \ 0.05), higher herbivore
abundance (E??facultative tropical = 0.77, CI = 0.46–1.01;
E??facultative temperate = 0.25, CI = -0.07–0.585; QB =
2.25, P = 0.108), and stronger reduction in plant reproduc-
tion (E??facultative tropical = -0.62, CI = -0.71 to -0.47;
E??facultative temperate = -0.05, CI = -0.19E??0.01;
QB = 34.19, P \ 0.05). When only tropical myrmecophytes
and myrmecophiles were contrasted, ant–plants exhibited
almost 4 times higher herbivory compared to plants
with facultative associations (E??tropical obligate = 2.31,
CI = 2.02–2.60; E??tropical facultative = 0.64, CI = 0.36–
0.88; QB = 81.23, P \ 0.001), but similar reductions in
plant reproduction (E??tropical obligate = -0.51, CI = -0.79
to –0.40; E??tropical facultative = -0.46, CI = -0.74 to
-0.18; QB = 6.51, P = 0.09).
We also considered whether ant attractor types had a
significant impact on the magnitude of the effects studied.
In relationship to attractor type (= attractor identity), plants
possessing a combination of domatia and Hemiptera
exhibited the strongest effects of ant removal on herbivore
damage (E?? = 3.32), followed by plants with domatia
only (E?? = 2.42) and food bodies (E?? = 1.14). Dif-
ferent results were observed for the abundance of
herbivores, as when the category ‘‘unknown’’ was ignored,
plants hosting sap-sucking hemipterans as the sole ant
attractor exhibited the strongest effect, with a 65.7%
increase in the abundance of other non-hemipteran herbi-
vores following ant exclusion. For plant reproduction,
when to EFN-attracted ants were excluded, the reduction
on plant fitness was nearly 36% (E?? = -0.362, CI =
-0.47 to -0.24), significantly differing from the effects of
other ant attractors (QB = 28.52, P \ 0.0001). All other
ant attractors alone or in combination exhibited non-sig-
nificant effects on plant reproduction after ant exclusion
(all with confidence intervals that overlapped with zero).
When ant attractor function instead of identity was
considered, similar results were found, as plants offering
shelter and hemipterans as ant attractors exhibited the
strongest effects on herbivory rates (E?? = 3.32,
CI = 2.51–4.12), followed by shelter only (E?? = 2.43,
CI = 2.05–2.80) (Fig. 4). Weakest effects on herbivory
were observed for plants that offered only food for ants in
the form of EFNs or food bodies (E?? = 0.48, CI = 0.30–
0.66). For plant reproduction, significant effects of ant
exclusion were observed only when ants were attracted to
food rewards, with a reduction of 36% in plant fitness
compared to control plants where ants were allowed.
Multiple ant attractors exhibited strongest effects on her-
bivory (E?? = 1.49) and plant reproduction (E?? =
-0.44) compared to single ant attractors (QB = 20.61 and
QB = 10.04, respectively; P \ 0.05). When effects of
Proportional change from control (with ants)-0.5 0.0 0.5 1.0 1.5 2.0
Herbivory
Herbivore abundance
Plant reproduction
(44)(40)
(39)
(32)
(15)
(43)
single ant species
several species
Fig. 3 Effects of ant removal
on plant damage, herbivore
abundance and plant
reproduction according to the
number of ant species excluded
(either a single species or
several species). Only main
effects are presented with its
95% confidence interval. Effects
are significant if confidence
intervals do not overlap with
zero
Oecologia (2009) 160:537–549 543
123
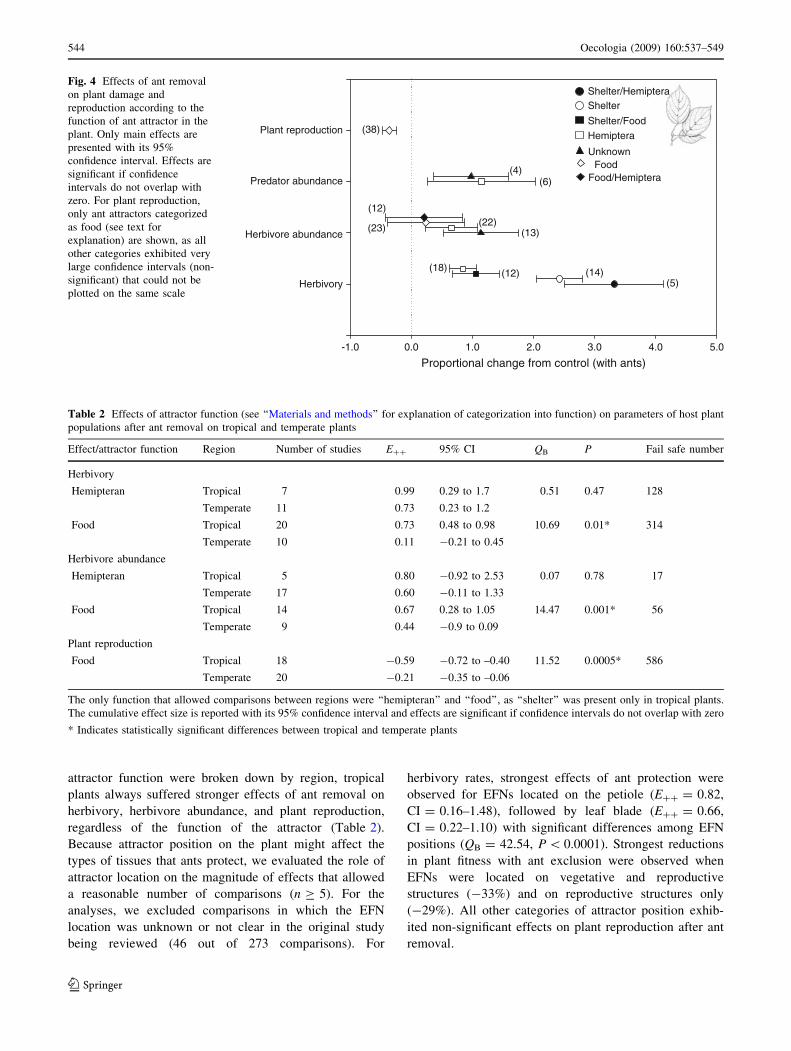
attractor function were broken down by region, tropical
plants always suffered stronger effects of ant removal on
herbivory, herbivore abundance, and plant reproduction,
regardless of the function of the attractor (Table 2).
Because attractor position on the plant might affect the
types of tissues that ants protect, we evaluated the role of
attractor location on the magnitude of effects that allowed
a reasonable number of comparisons (n C 5). For the
analyses, we excluded comparisons in which the EFN
location was unknown or not clear in the original study
being reviewed (46 out of 273 comparisons). For
herbivory rates, strongest effects of ant protection were
observed for EFNs located on the petiole (E?? = 0.82,
CI = 0.16–1.48), followed by leaf blade (E?? = 0.66,
CI = 0.22–1.10) with significant differences among EFN
positions (QB = 42.54, P \ 0.0001). Strongest reductions
in plant fitness with ant exclusion were observed when
EFNs were located on vegetative and reproductive
structures (-33%) and on reproductive structures only
(-29%). All other categories of attractor position exhib-
ited non-significant effects on plant reproduction after ant
removal.
Proportional change from control (with ants)-1.0 0.0 1.0 2.0 3.0 4.0 5.0
Herbivory
Herbivore abundance
Predator abundance
Plant reproduction
Shelter
Shelter/Food
Hemiptera
UnknownFood
Food/Hemiptera
(5)(14)(12)(18)
(13)(22)(23)
(12)
(6)(4)
(38)
Shelter/HemipteraFig. 4 Effects of ant removal
on plant damage and
reproduction according to the
function of ant attractor in the
plant. Only main effects are
presented with its 95%
confidence interval. Effects are
significant if confidence
intervals do not overlap with
zero. For plant reproduction,
only ant attractors categorized
as food (see text for
explanation) are shown, as all
other categories exhibited very
large confidence intervals (non-
significant) that could not be
plotted on the same scale
Table 2 Effects of attractor function (see ‘‘Materials and methods’’ for explanation of categorization into function) on parameters of host plant
populations after ant removal on tropical and temperate plants
Effect/attractor function Region Number of studies E?? 95% CI QB P Fail safe number
Herbivory
Hemipteran Tropical 7 0.99 0.29 to 1.7 0.51 0.47 128
Temperate 11 0.73 0.23 to 1.2
Food Tropical 20 0.73 0.48 to 0.98 10.69 0.01* 314
Temperate 10 0.11 -0.21 to 0.45
Herbivore abundance
Hemipteran Tropical 5 0.80 -0.92 to 2.53 0.07 0.78 17
Temperate 17 0.60 -0.11 to 1.33
Food Tropical 14 0.67 0.28 to 1.05 14.47 0.001* 56
Temperate 9 0.44 -0.9 to 0.09
Plant reproduction
Food Tropical 18 -0.59 -0.72 to –0.40 11.52 0.0005* 586
Temperate 20 -0.21 -0.35 to –0.06
The only function that allowed comparisons between regions were ‘‘hemipteran’’ and ‘‘food’’, as ‘‘shelter’’ was present only in tropical plants.
The cumulative effect size is reported with its 95% confidence interval and effects are significant if confidence intervals do not overlap with zero
* Indicates statistically significant differences between tropical and temperate plants
544 Oecologia (2009) 160:537–549
123

For those plants with honeydew-producing hemipterans,
stronger effects of ant exclusion on herbivory rates were
observed when ants were associated with Coccidae
(E?? = 1.18, CI = 0.24–2.11) compared to Aphididae
(E?? = 1.00, CI = 0.47–1.54) (QB = 7.04, P \ 0.05).
Due to small sample sizes for categorization, all other
effects evaluated in this study exhibited non-significant
relationships with ant exclusion according to hemipteran
family.
Assessment of publication bias
Fail-safe numbers for effects of ant removal on herbivory
rates and herbivore abundance were large (8,606 and 551
studies, respectively) relative to the number of independent
comparisons included in the meta-analysis (85 and 71,
respectively). For the effect grouped into ‘‘plant repro-
duction’’, the fail-safe number was 1,163 studies,
indicating the strength of the results found. Scatter plots of
effects size against sample size of all data exhibited a
typical funnel shape (figure not shown), indicating that
studies with low precision—generally with small sample
sizes—show a large dispersal of effect sizes around the true
effect, whereas those with large sample sizes have an effect
size close to the true value. Our results suggest that there
was little publication bias in the studies included in this
meta-analytical review.
Discussion
This meta-analytical review provides strong support for
ant–plant symbioses, although it also reveals the diversity
of ant–plant interactions with differential effects of ant
removal according to latitude, type of association between
ants and plants, number of ants involved in these associa-
tions, and type of ant attractor. The evidence presented here
reinforces the role of ants as plant biotic defenses, with
increased herbivory rates, herbivore density, and reduced
plant fitness following ant exclusion. Effects of ant
removal, however, were not homogeneous on plant
parameters investigated. Ant effects on herbivory rates
were stronger than their effects on insect herbivore density
and/or predator abundance. Ant removal increased her-
bivory by more than 95%, whereas increased herbivore
abundance by only 50%. Quantification of ant effects on
herbivory loads suffered by plants might be actually easier
to perform than quantification of herbivore abundance after
ant exclusion, as herbivory levels might be investigated
only once during a course of a study (a snapshot of effects
of ant exclusion in a season, such as number of chewed
leaves or feeding holes), whereas data collection on her-
bivore abundance encompasses direct observations and/or
comparatively higher sampling efforts. Moreover, herbi-
vores might increase residence time on plants without ants
(Suzuki et al. 2004) therefore increasing the amount of
damage on the plants they feed upon and contributing to
stronger effects of ant removal on herbivory compared to
effects on insect abundance.
The effects of ant–plant or ant–plant-hemipteran asso-
ciations on herbivory is also species-specific, with different
outcomes in partners’ fitness (Heil and McKey 2003). This
variation in strength among species is dependent upon the
vulnerability of herbivores to ant predation and/or avoid-
ance, and can lead to changes in the overall structure of
arthropod communities in the presence of ants (Fowler and
MacGarvin 1985). Styrsky and Eubanks (2007) have
shown that species richness of other sucking, non-honey-
dew-producing herbivores was reduced by 28% after ant
exclusion, whereas species richness of leaf-chewing cat-
erpillars was increased by 69% on branches without ants
compared to branches with ants. In contrast, species rich-
ness of leaf-mining caterpillars, a guild of herbivores that is
protected against ant predation, was actually 44% greater
on the trees with hemipteran-tending ants, presumably
because the ants indirectly protect the concealed caterpil-
lars from other predators (Fowler and MacGarvin 1985).
Our review has shown that, although herbivores studied
belonged to seven different guilds, chewers such as cater-
pillars and folivorous beetles accounted for 68% of the
independent comparisons reviewed, impairing analyses
regarding the effects of ant removal on herbivory rates
according to herbivore feeding mode. In addition, ant
behavior may also affect the outcome of the interaction.
Ants vary greatly in their behavior towards herbivores
(Bronstein 1998; Michelangeli 2003) and in the way they
interact with insects, therefore differing in their protective
effect on host plants (Fraser et al. 2001). The function
of ant attractant and the ultimate effects on a plant
thus depend on the array of species visiting the attractant
(Cuautle and Rico-Gray 2003).
Ecologically dominant ants have been shown to alter
arthropod communities, acting as important predators,
mutualists, competitors, and prey (Gibb 2003). The fact
that hemipteran-tending ants reduce the survival and
abundance and alter the spatial distribution of hemipterans’
natural enemies is extensively documented (Del-Claro and
Oliveira 2000; Renault et al. 2005). Our results have shown
that ant exclusion caused an increase in predator abundance
and a tendency to a decrease in the abundance of hemipt-
erans. In the 14 comparisons of changes in the density of
other predators from nine independent studies, the majority
of predators studied were generalist predators, such as
spiders. Some authors have argued that effects of ants on
other predators tend to be less consistent than effects of
ants on herbivores, with some predator taxa responding to
Oecologia (2009) 160:537–549 545
123

ant exclusion (Sipura 2002) and others not (Karhu 1998;
Gibb 2003; Offenberg et al. 2005). Our results for ant
effects on predators come from a relatively small number
of comparisons, but they do show strong effects of ant
removal on predator abundance in terrestrial communities
which, in turn, have the potential to affect the composition,
diversity, and abundance of the herbivore community on
those plants. Increase in the abundance of other predators
with ant exclusion might be explained by increased
vulnerability of ant-tended hemiptera in the absence of
ants, making them easy targets for spiders (Del-Claro and
Oliveira 2000), beetles (James et al. 1999), and syrphid
flies (Sipura 2002). Ant exclusion allows the attack of
natural enemies of hemipterans, thereby decreasing their
abundance (by 66% in our review, although not significant)
and supporting the evidence for the strong trophobiosis
between Formicidae and hemipterans (see Delabie 2001).
Some of the most central functions enabling plant sur-
vival and reproduction depend on mutualisms (Heil 2008),
and many plants rely on the third trophic level in order to
get protection from the second trophic level. Beneficial
effects of ants on plants arise when ants are capable of
reducing herbivore numbers, herbivory levels, or both,
thereby decreasing plant damage and increasing plant fit-
ness. Positive effects of ants on plant fitness were less
commonly addressed than ant effects on herbivory, the
former representing only 21.5% of the independent com-
parisons included here. Only 18% of all studies reviewed
addressed effects of ant removal on herbivory and on
components of plant fitness simultaneously (Del-Claro
et al. 1996; Letourneau 1998; Izzo and Vasconcelos 2002;
Rudgers 2004). Among these, 62% were performed in
temperate systems. Effects of ant removal on plant repro-
duction were significant, but weaker than effects of ants on
plant herbivory. Plants without ants experienced a 25%
decrease in reproduction (flower, fruit, and seed production
combined: E??= -24.2, CI = -0.30 to –0.18, n = 58) and
effects were significant for tropical systems only. Weaker
effects of ants on plant reproduction might be explained
by several reasons: herbivores might not affect plant
fitness, as plants are able to tolerate herbivory without
fitness reduction (Strauss and Agrawal 1999). Alterna-
tively, effects of increased herbivory on ant-excluded
plants might not incur in fitness reductions in the same
season, as time lags for the beneficial effects of ants on
herbivory deterrence and subsequent results on plant
fitness might occur. The majority of studies reviewed
here were short-term studies (but see Torres-Hernandez
et al. 2000; Rudgers 2004) where aspects of plant
reproduction were evaluated soon after ant exclusion.
Long-term studies might be necessary to address the
relationship between ant presence, plant herbivory, and
plant reproduction.
Protective ant–plant or ant–hemipteran interactions are
important in both temperate and tropical communities
(Bronstein 1998; Heil and McKey 2003). Ant–plant mu-
tualisms in the neotropics have received much more
attention compared to other regions (Fiala et al. 1999) even
though predatory ants are considered keystone species in
temperate and boreal woodlands due to their effects on
herbivore community composition and abundance (Sipura
2002). In a recent review, Styrsky and Eubanks (2007) did
not observe significant differences in the effects of ant–
hemipteran interactions between temperate and tropical
regions. On the other hand, this study reviewed 81 studies
of the effects of ants as plant biotic defenses, and we have
shown stronger effects of ant removal on herbivory in
tropical environments compared to temperate ones. Pre-
sumably, this effect is due to higher ant and herbivore
diversity in the tropics as well as higher herbivore speci-
ficity (Coley and Barone 1996; Dyer et al. 2007; but see
Novotny et al. 2006) and ant aggressive behavior in trop-
ical compared to temperate regions. Moreover, tropical
areas harbor both myrmecophytes and myrmecophilies,
whereas temperate vegetation lacks myrmecophytic spe-
cies. In a review of tritrophic interactions in tropical and
temperate ecosystems, Dyer and Coley (2002) observed
that tropical plants are better defended against herbivores
then temperate species and that natural enemies have
strong negative effects on herbivores at all latitudes, but the
magnitude of the effect was significantly higher in tropical
than in temperate areas. Therefore, trophic cascades differ
between tropical and temperate ecosystems, and stronger
top-down effects of predators on herbivores and of herbi-
vores on plants, are significantly stronger in the tropics.
Our results have also shown that single ants on myr-
mecophytic plants have stronger effects on herbivory rates
and herbivore abundance than several ants, reinforcing the
idea that ants better protect tropical than temperate plants.
Our results have also shown that effects of single ant
species on arthropod communities and/or herbivore dam-
age and plant fitness were stronger than the effects of
several ant species. Visitation by multiple ant species that
vary in anti-herbivore abilities may result in reduced ben-
efits, relative to an exclusive association with a high-
quality mutualist (Miller 2007). The higher effect of single
ants in tropical plants could be explained by a stronger and
specialized mutualism, or by the presence of ecologically
dominant aggressive ants. Mosaics of behaviorally domi-
nant ants, for example, have been observed in tropical
canopies, where ants represent more than 90% of the
individuals and 50% of the arthropods (Dejean and Corbara
2003). Other studies also observed that plant resources,
such as EFNs, hemipterans, or domatia, shaping these ant
mosaics (Davidson 1997; Bluthgen et al. 2000; Hossaert-
McKey et al. 2001) influence thereby the outcomes of
546 Oecologia (2009) 160:537–549
123

plant–ant mutualisms in tropical regions. Defense of
exclusive foraging territories in tropical tree canopies by
particularly abundant and aggressive hemipteran-tending
ants reduces the density and diversity of other ants,
resulting in mosaic distributions of dominant and sub-
dominant arboreal ant species (Bluthgen et al. 2000, 2004;
Djieto-Lordon et al. 2007). Single but aggressive ants, such
as Azteca (Schultz and McGlynn 2000), have the potential
to influence the strength of the interaction between ants and
plants in mutualistic associations. However, the division
between dominant and subordinate ant species was not
possible using data provided by authors of the studies
reviewed. Also, the effect for single species was stronger
when ants from obligate association were removed, rein-
forcing the first hypothesis. The fact that almost 80% of the
myrmecophytic plants reviewed here were colonized by a
single ant species and myrmecophytic plants associated
with several ant species were rare in our review (only three
studies: see Alvarez et al. 2001; Heil et al. 2001; Michel-
angeli 2003) also reinforces the hypothesis of the strong
role of ants as plant biotic defenses in obligate ant–plants in
the tropical regions. In temperate regions, on the other
hand, all ant–plant associations are facultative and half of
these associations were maintained with single ant species
and half with several ants. Therefore, it is not surprising
that specialist ants have stronger effects than opportunistic
species.
The outcome of ant–plant mutualisms can be dependent
upon several biotic and abiotic factors (Heil and McKey
2003), and the type of food rewards are among those fac-
tors that shape this interaction. In this study, domatia-
bearing plants hosting honeydew-producing hemipterans
exhibited strongest effects of ant removal compared to
other ant attractors. In myrmecophiles, ants generally
exhibit low fidelity to the food association (Kersch and
Fonseca 2005) and many species are commonly present on
a given plant over its lifetime or might switch among plants
(Beattie 1985). In fact, EFN-based ant–plant associations
tend to be more generalized while the associations
involving domatia and food bodies tend to be more spe-
cialized and specific (Rico-Gray and Oliveira 2007).
Domatia seem to offer a much more specific reward than
EFNs (Heil and McKey 2003) and plant–ants generally
show strong fidelity to their ant–plants (Rico-Gray and
Oliveira 2007). Although widespread, mutualistic systems
involving either hemipterans or EFNs seem to be looser or
less specific with uncertain outcome to the plant (Rico-
Gray and Oliveira 2007) whereas aggressive ant behavior
on domatia-bearing plants has been previously recorded
(Janzen 1966; Heil and McKey 2003). Our results provide
support for the hypothesis that identity of the ant attractor
plays an important role in the evolution of the interaction
among plants, ants, and other herbivores, shaping therefore
the outcome of mutualistic interactions, especially in
tropical regions, where plants with domatia are commonly
found (Heil and McKey 2003).
Previous studies provided compelling evidence that ant–
hemipteran and ant–plant interactions can act as ‘keystone
interactions’ that dramatically change the structure of
arthropod communities on plants. In the presence of hon-
eydew-producing hemipterans, EFNs, food bodies, or
domatia, ants alter the abundance and distribution of spe-
cialist and generalist predators and parasitoids, and
multiple species of herbivores in several feeding guilds,
resulting in changes to local species diversity (Styrsky and
Eubanks 2007). We showed here that ant presence on
plants, regardless of the type of ant attractor or geographic
location, broadly affects the local abundance and distri-
bution of predators and insect herbivores, affecting in turn
herbivory levels and plant fitness. Therefore, ant–plant or
ant–hemipteran interactions may represent ‘keystone
interactions’ in many communities. However, the effect of
the ant presence is stronger in tropical environments, and
despite of the fact that selection has only rarely favored
obligate mutualisms (Rico-Gray and Oliveira 2007), the
positive outcome is also stronger in domatia-bearing plants
associated with single dominant ant species.
Acknowledgments This study was part of the course ‘‘Topics in
Ecology––Meta-Analysis’’ of the graduate program in Ecology,
Conservation and Management at UFMG, taught by T Cornelissen.
We would like to thank all the authors that kindly sent separates or
pdfs, especially Dr. Daniel Janzen. T Cornelissen acknowledges
FAPESP (06/57881-5) for a postdoctoral fellowship and G.W.
Fernandes acknowledges CNPq for a research fellowship (30.9633/
2007-9).
References
Alvarez G, Armbrecht I, Jimenez E, Armbrecht H, Ulloa-Chacon P
(2001) Ant–plant association in two Tococa species from a
primary rain forest of Colombian Choco (Hymenoptera: Form-
icidae). Sociobiology 38:585–602
A.P.G. [= Angiosperm Phylogeny Group] II (2003) An update of
the Angiosperm Phylogeny Group classification for the orders
and families of flowering plants: APG II. Bot.J Linn Soc
141:399–436
Beattie AJ (1985) The evolutionary ecology of ant–plant mutualisms.
Cambridge University Press, Cambridge
Bluthgen N, Verhaagh M, Goitıa W, Jaffe K, Morawetz W, Barthlott
W (2000) How plants shape the ant community in the
Amazonian rainforest canopy: the key role of extrafloral
nectaries and homopteran honeydew. Oecologia 125:229–240
Bluthgen N, Stork NE, Fiedler K (2004) Bottom-up control and co-
occurrence in complex communities: honeydew and nectar
determine a rainforest ant mosaic. Oikos 106:4344–4358
Bronstein JL (1998) The contribution of ant plant protection studies to
our understanding of mutualism. Biotropica 30:150–161
Bronstein JL, Alarcon R, Geber M (2006) The evolution of plant–
insect mutualisms. New Phytol 172:412–428
Oecologia (2009) 160:537–549 547
123

Bruna EM, Lapola DM, Vasconcelos HL (2004) Interspecific
variation in the defensive responses of obligate plant–ants:
experimental tests and consequences for herbivory. Oecologia
138:558–565
Coley PD, Barone JA (1996) Herbivory and plant defenses in tropical
forests. Annu Rev Ecol Syst 27:305–335
Compton SG, Robertson HG (1988) Complex interactions between
mutualisms: ants tending homopterans protect fig seeds and
pollinators. Ecology 69:1302–1305
Crutsinger GM, Sanders NJ (2005) Aphid-tending ants affect
secondary users in leaf shelters and rates of herbivory on Salixhookeriana in a coastal dune habitat. Am Midl Nat 154:296–304
Cuautle M, Rico-Gray V (2003) The effect of wasps and ants on the
reproductive success of the extrafloral nectaried plant Turneraulmifolia (Turneraceae). Funct Ecol 17:417–423
Davidson DW (1997) The role of resource imbalances in the
evolutionary ecology of tropical arboreal ants. Biol J Linnean
Soc 61:153–181
Davidson DW, McKey D (1993) The evolutionary ecology of
symbiotic ant–plant relationships. J Hymenoptera Res 2:13–83
Dejean A, Corbara B (2003) A review of mosaics of dominant ants in
rainforests and plantations. In: Basset Y, Novotny V, Miller SE
(eds) Arthropods of tropical forests: spatio-temporal dynamics
and resource use in the canopy. Cambridge University Press,
New York, pp 341–347
Dejean A, Delabie JHC, Cerdan P, Gibernau M, Corbara B (2006)
Are myrmecophytes always better protected against herbivores
than other plants? Biol J Linn Soc 89:91–98
Delabie JHC (2001) Trophobiosis between Formicidae and Hemiptera
(Sternorrhyncha and Auchenorrhyncha): an overview. Neotrop
Entomol 30:501–516
Delabie JHC, Osprina M, Zabala G (2003) Relaciones entre hormigas
y plantas: una introduccion. In: Fernandez F (ed) Introduccion a
las Hormigas de la region Neotropical. Instituto de Investigacion
de Recursos Biologicos Alexander Von Humboldt, Bogota, pp
167–180
Del-Claro K (2004) Multitrophic relationships, conditional mutual-
isms, and the study of interaction biodiversity in tropical
savannas. Neotrop Entomol 33:665–672
Del-Claro K, Oliveira PS (2000) Conditional outcomes in a neotrop-
ical treehopper–ant association: temporal and species-specific
variation in ant protection and homopteran fecundity. Oecologia
124:156–165
Del-Claro K, Berto V, Reu W (1996) Effect of herbivore deterrence
by ants on the fruit set of an extrafloral nectary plant, Qualeamultiflora (Vochysiaceae). J Trop Ecol 12:887–892
Del-Claro K, Byke J, Yugue GM, Morato MG (2006) Conservative
benefits in an ant–hemipteran association in the Brazilian
tropical savanna. Sociobiology 47:415–421
Djieto-Lordon C, Dejean A, Ring RA, Nkongmeneck BA, Lauga J,
McKey D (2007) Ecology of an improbable association: the
Pseudomyrmecine plant–ant Tetraponera tessmanni and the
myrmecophytic liana Vitex thyrsiflora (Lamiaceae) in Camer-
oon. Biotropica 37:421–430
Dutra HP, Freitas AVL, Oliveira PS (2006) Dual ant attraction in the
neotropical shrub Urera baccifera (Urticaceae): the role of ant
visitation to pearl bodies and fruits in herbivore deterrence and
leaf longevity. Funct Ecol 20:252–260
Dyer LA, Coley PD (2002) Tritrophic interactions in tropical and
temperate communities. In: Tscharntke T, Hawkins B (eds)
Multitrophic Level Interactions. Cambridge University Press,
Cambridge, pp 67–88
Dyer LA, Singer MS, Lill JT, Stireman JO, Gentry GL, Marquis RJ,
Greeney HF, Wagner DL, Morais HC, Diniz IR, Kursar TA,
Coley PD (2007) Host specificity of Lepidoptera in tropical and
temperate forests. Nature 448:696–699
Fiala B, Maschwitz U, Pong TY, Helbig AJ (1989) Studies of a
Southeast asian ant–plant association: protection of Macarangatrees by Crematogaster borneensis. Oecologia 79:463–470
Fiala B, Jakob A, Maschwitz U (1999) Diversity, evolutionary
specialization and geographic distribution of a mutualistic ant–
plant complex: Macaranga and Crematogaster in South East
Asia. Biol J Linn Soc 66:305–331
Fonseca CR (1994) Herbivory and the long-lived leaves of an
Amazonian ant-tree. J Ecol 82:833–842
Fowler SV, MacGarvin M (1985) The impact of hairy wood ants,
Formica lugubris, on the guild structure of herbivorous insects
on birch, Betula pubescens. J Anim Ecol 54:847–855
Fraser AM, Axen AH, Pierce NE (2001) Assessing the quality of
different ant species as partners of a myrmecophilous butterfly.
Oecologia 129:452–460
Frederickson ME, Gordon DM (2007) The devil to pay: a cost of
mutualism with Myrmelachista schumanni ants in ‘devil’s
gardens’ is increased herbivory on Duroia hirsuta trees. Proc
R Soc Lond B 274:1117–1123
Freitas L, Galetto L, Bernardello G, Paoli AAS (2000) Ant exclusion
and reproduction of Croton sarcopetalus (Euphorbiaceae). Flora
195:398–402
Gaume L, Zacharias M, Grosbois V, Borges RM (2005) The fitness
consequences of bearing domatia and having the right ant
partner: experiments with protective and non-protective ants in a
semi-myrmecophyte. Oecologia 145:76–86
Gibb H (2003) Dominant meat ants affect only their specialist
predator in a complex natural system. Oecologia 136:609–615
Halaj J, Wise DH (2001) Terrestrial trophic cascades: how much do
they trickle? Am Nat 157:262–281
Heil M (2008) Indirect defense via tritrophic interactions. New Phytol
178:41–61
Heil M, McKey D (2003) Protective ant–plant interactions as model
systems in ecological and evolutionary research. Annu Rev Ecol
Evol Syst 34:425–453
Heil M, Fiala B, Maschwitz U, Linsenmair KE (2001) On benefits of
indirect defence: short- and long-term studies of antiherbivore
protection via mutualistic ants. Oecologia 126:395–403
Hossaert-McKey M, Orivel J, Labeyrie E, Pascal L, Delabie JHC,
Dejean A (2001) Differential associations with ants of three co-
occurring extrafloral nectary-bearing plants. Ecoscience 8:325–
335
Izzo TJ, Vasconcelos HL (2002) Cheating the cheater: domatia loss
minimizes the effects of ant castration in an Amazonian ant–
plant. Oecologia 133:200–205
James DG, Stevens MM, O’Malley KJ, Faulder RJ (1999) Ant
foraging reduces the abundance of beneficial and incidental
arthropods in Citrus canopies. Biol Control 14:121–126
Janzen DH (1966) Coevolution of mutualism between ants and
acacias in Central America. Evolution 20:249–275
Janzen DH (1967) Interaction of the bull’s horn Acacia (Acaciacornigera L.) with an ant inhabitant (Pseudomyrmex ferrugineaF. Smith) in Eastern Mexico. Univ Kans Sci Bull 47:315–558
Karhu KJ (1998) Effects of ant exclusion during outbreaks of a
defoliator and a sap-sucker on birch. Ecol Entomol 23:185–194
Kersch MF, Fonseca CR (2005) Abiotic factors and the conditional
outcome of an ant–plant mutualism. Ecology 86:2117–2126
Koptur S (2005) Nectar as fuel for plant protectors. In: Wackers FL,
van Rijn PCJ, Bruin J (eds) Plant-provided food for carnivorous
insects: a protective mutualism and its applications, Cambridge
University Press, Cambridge, pp 75–108
Letourneau DK (1998) Ants, stem-borers, and fungal pathogens:
Experimental tests of a fitness advantage in Piper ant–plants.
Ecology 79:593–603
Letourneau DK, Barbosa P (1999) Ants, stem borers, and pubescence
in Endospermum in Papua New Guinea. Biotropica 31:295–302
548 Oecologia (2009) 160:537–549
123

Messina FJ (1981) Plant protection as a consequence of an ant–
membracid mutualism: interactions on goldenrod (Solidago sp.).
Ecology 62:1433–1440
Michelangeli FA (2003) Ant protection against herbivory in three
species of Tococa (Melastomataceae) occupying different envi-
ronments. Biotropica 35:181–188
Miller TEX (2007) Does having multiple partners weaken the benefits
of facultative mutualism? A test with cacti and cactus-tending
ants. Oikos 116:500–512
Moller AP, Jennions MD (2001) Testing and adjusting for publication
bias. Trends Ecol Evol 16:580–586
Mooney KA (2007) Tritrophic effects of birds and ants on a canopy
food web, tree growth, and phytochemistry. Ecology 88:2005–
2014
Novotny V, Drozd P, Miller SE, Kulfan M, Janda M, Basset Y,
Weiblen GD (2006) Why are there so many species of
herbivorous insects in tropical rainforests? Science 313:1115–
1118
Offenberg J, Nielsen MG, Macintosh DJ, Havanon S, Aksornkoae S
(2005) Lack of ant attendance may induce compensatory plant
growth. Oikos 111:170–178
Oliveira PS (1997) The ecological function of extrafloral nectaries:
Herbivore deterrence by visiting ants and reproductive output in
Caryocar brasiliense (Caryocaraceae). Funct Ecol 11:323–330
Oliveira PS, Freitas AVL (2004) Ant–plant–herbivore interactions in
the Neotropical Cerrado Savanna. Naturwissenschaften 91:557–
570
Renault CK, Buffa LM, Delfino MA (2005) An aphid–ant interaction:
effects on different trophic levels. Ecol Res 20:71–74
Rico-Gray V, Oliveira P (2007) The ecology and evolution of ant–
plant interactions. University of Chicago Press, Chicago
Rosenberg MS, Adams DC, Gurevitch J (2000) MetaWin: statistical
software for meta-analysis. Version 2.0, Sinauer Associates,
Sunderland
Rosenthal R (1979) The ‘file drawer’ problem and tolerance for null
results. Psychol Bull 86:638–641
Rudgers JA (2004) Enemies of herbivores can shape plant traits:
Selection in a facultative ant–plant mutualism. Ecology 85:192–
205
Ruhren S (2003) Seed predators are undeterred by nectar-feeding ants
on Chamaecrista nictitans (Caesalpineaceae). Plant Ecol
166:189–198
Schemske DW (1982) Ecological correlates of a neotropical mutu-
alism: ant assemblages at Costus extrafloral nectaries. Ecology
63:932–941
Schmitz OJ, Hamback PA, Beckerman AP (2000) Trophic cascades in
terrestrial systems: a review of the effects of carnivore removal
on plants. Am Nat 55:141–153
Schultz TR, McGlynn TP (2000) The interaction of ants with another
organisms. In: Agosti D, Majer JD, Alonso LE, Schultz TR (eds)
Ants: standard methods for measuring and monitoring biodiver-
sity. Smithsonian Institution Press, Washington, pp 1–8
Sipura M (2002) Contrasting effects of ants on the herbivory and
growth of two willow species. Ecology 83:2680–2690
Strauss SY, Agrawal AA (1999) The ecology and evolution of plant
tolerance to herbivory. Trends Ecol Evol 14:179–185
Styrsky JD, Eubanks MD (2007) Ecological consequences of
interactions between ants and honeydew-producing insects. Proc
R Soc Lond Ser B Biol Sci 274:151–164
Suzuki N, Ogura K, Katayama N (2004) Efficiency of herbivore
exclusion by ants attracted to aphids on the vetch Viciaangustifolia L. (Leguminosae). Ecol Res 19:275–282
Torres-Hernandez L, Rico-Gray V, Castillo-Guevara C, Vergara JA
(2000) Effect of nectar-foraging ants and wasps on the repro-
ductive fitnesse ot Turnera ulmifolia (Turneraceae) in a coastal
sand dune in Mexico. Acta Zool Mex 81:13–21
Webber BL, Moog J, Curtis ASO, Woodrow IE (2007) The diversity
of ant–plant interactions in the rainforest understory tree,
Ryparosa (Achariaceae): food bodies, domatia, prostomata, and
hemipteran trophobionts. Biol J Linn Soc 154:353–371
Oecologia (2009) 160:537–549 549
123
Related Documents











