




EK-11
ANKARA ÜNİVERSİTESİ
BİLİMSEL ARAŞTIRMA PROJESİ KESİN RAPORU
Tutuklanmış L.Kefir ile Enantiyomerik Saflıkta 1- Fenil Etanol Üretiminin Dolgulu Kolon
Biyoreaktörde İncelenmesi
Proje Yürütücüsünün İsmi: Prof. Dr. Ülkü MEHMETOĞLU
Yardımcı Araştırmacıların İsmi: Prof. Dr. Emine BAYRAKTAR Araş. Gör. Özlem AYDOĞAN
Proje Numarası: 09B4343010 Başlama Tarihi: Mart 2009
Bitiş Tarihi: Mart 2011 Rapor Tarihi: Nisan 2011
Ankara Üniversitesi Bilimsel Araştırma Projeleri Ankara - " 2011 "

I. Projenin Türkçe ve İngilizce Adı ve Özetleri Tutuklanmış L.Kefir ile Enantiyomerik Saflıkta 1-Fenil Etanol Üretiminin Dolgulu Kolon Biyoreaktörde İncelenmesi Enantiyomerik saflıktaki kiral alkoller pek çok farmokolojik ürünün çıkış maddesidir. Bu projede ilaç
etken maddesi olarak geniş kullanım alanına sahip olan 1-fenil etanolün üretimi, asetofenonun alkol
dehidrojenaz (ADH) enzimi katalizli asimetrik indirgenme tepkimesiyle gerçekleştirilmiştir. Bu
çalışmada, mikroorganizmaların, enzimlere göre daha ekonomik olması nedeniyle ADH aktivitesine
sahip, Lactobacillus kefir NRRL B-1839 mikroorganizması kullanılmıştır. Bir prokiral keton olan
asetofenonun asimetrik indirgenmesiyle enantiyomerik saflıkta 1-fenil etanolün üretimi tutuklanmış
Lactobacillus kefir ile sürekli işletilen dolgulu kolon biyoreaktörde gerçekleştirilmiştir.
Çalışmanın ilk aşamasında, serbest Lactobacillus kefir hücreleriyle çalışılmış ve bu mikroorganizmanın
R-enantiyomer için yüksek enantiyomerik aşırılık verdiği (> % 99) görülmüştür. Özellikle farmokolojik
ürün üretiminde yüksek enantiyomerik aşırılık istenen bir durumdur. Ancak ulaşılan dönüşüm
değerlerinin; düşük substrat derişimlerinde (Co=1 mM) bile, düşük (% 80) olduğu görülmüştür.
Asimetrik indirgeme tepkimelerini oksidoredüktaz sınıfından, alkol dehidrojenaz (ADH) enzimi
katalizler ve bu enzim kofaktöre ihtiyaç duyarlar. Düşük substrat derişimlerinde bile dönüşümün düşük
olması nedenini enzimin kofaktörünün tepkime sırasında tükenmesidir. Bu yüzden tepkime esnasında
kofaktör ilavesi veya kofaktör rejenerasyonunun gerekli olduğunu göstermiştir. Kofaktörler pahalı
olduklarından ve tepkime sonucu tükendiklerinden, kofaktör rejenerasyonunun yapılması daha
ekonomik olmaktadır. Kofaktör rejenerasyonu, ucuz karbon kaynakları kullanılarak gerçekleştirilebilir.
Bu çalışmada, kofaktör rejenerasyonu için üç farklı yol izlenmiş ve en iyi sonuçlar glikoz ile yapılan
kofaktör rejenerasyonu ile elde edilmiştir. Diğer rejenerasyon türleri olarak, 2-propanolün kullanılması
ve gen klonlama ile alkol dehidrojenaz ve laktat dehidrojenaz enzimlerinin Escherichia coli’ ye
aktarılarak, iki enzim ile kofaktör rejenerasyonudur. 2-propanol ile 20 mM başlangıç substrat
derişiminde gerçekleştirilen deneylerde, hem enantiyomerik aşırılık (% 97) hemde dönüşüm açısından
(% 95), glikoz sonuçlarına göre (enantiyomerik aşırılık >% 99 ve dönüşüm % 100) daha düşük değerler
elde edilmiştir. Alkol dehidrojenaz (ADH) ve laktat dehidrojenaz (LDH) enzimlerini içeren
rekombinant Escherichia coli de, LDH enzimi aktivite gösterirken, ADH aktivite göstermemiştir. Bu
nedenle çalışmanın ilerleyen kısımlarında, glikoz ile kofaktör rejenerasyonu sağlanmıştır.

R-1-fenil etanol üretimini, sürekli sistemde gerçekleştirmeden önce, tutuklanmış mikroorganizma ile
kesikli sistemde çalışılmıştır. En uygun koşullar, sıcaklık, 35 oC, pH 8 Tris tamponu, kofaktör
rejenerasyonu için % 2 w/v glikoz, 4 h olarak bulunmuştur. Bu koşullar belirlendikten sonra, 1,7 cm iç
çapında, 27 cm dolgu yüksekliğinde pyrex camdan yapılmış, dolgulu kolon biyoraktörde 1-fenil etanol
üretimi sürekli sistemde gerçekleştirilmiştir.
Sürekli sistemde, asetofenon derişimi 15 mM, 25 mM ve 40 mM aralığında incelenmiştir. Substrat
derişimi etkisi incelenirken, diğer parametreler, glukoz derişimi (0.02 g/mL), kalma süresi (4.5 h),
sıcaklık (35 oC) sabit tutulmuştur. Saat başı örnekler alınarak 9 saat boyunca, R-1-feniletanol derişimi,
dönüşüm ve enantiyomerik aşırılığın değişimi izlenmiştir. Girdi derişimi arttıkça, ürün derişimide
artmıştır. 40 mM başlangıç girdi derişiminde, 12.8 mM R-1-feniletanol, 15 mM başlangıç girdi
derişiminde ise 11.2 mM R-1-feniletanol (% 79 dönüşüm) elde edilmiştir.
Kiral alkollerin endüstriyel üretiminde, yüksek ürün derişimi, yüksek verimlilik istenmektedir. 40 mM
başlangıç girdi derişiminde, verimlilik 2.82 mmolL-1h-1, 15 mM başlangıç girdi derişiminde ise 2.50
mmolL-1h-1 verimliliğe ulaşılmıştır. Substrat derişiminin artmasının, dönüşümü olumsuz yönde
etkilediği görülmektedir. Bunun nedeni, daha önceden serbest hücrelerle yapılan çalışmalar sonucu
görülen substrat inhibisyonunun olmasıdır. Ancak çalışılan tüm asetofenon derişimlerinde,
enantiyomerik aşırılık > % 99 elde edilmiştir.
Sürekli sistemde çalışırken yine, kofaktör rejenerasyonunda kullanılan glikoz miktarının etkisi
incelenmiştir. Glikoz derişimi 0.01, 0.02 ve 0.03 g/mL aralığında incelenmiş, diğer parametreler,
substrat derişimi (15 mM), kalma süresi (4.5 h) ve sıcaklık (35 oC) sabit tutulmuştur. Glikoz derişimi,
0.01 g/mL den 0.02 g/mL ye artırıldığında, dönüşüm % 46 dan % 79 e artmıştır. Glukoz derişiminin
0.03 g/mL ye çıkarılması ile dönüşüm % 34’e düşmüştür Burada, aşırı glukoz derişiminin inhibisyona
neden olduğu görülmüştür.
Sürekli sistemde, farklı akış hızlarında çalışılmış, buna bağlı olarak biyoreaktörede kalma süreleri
sırasıyla, 2.25 saat, 3 saat, 4.5 saat ve 9 saat olarak bulunmuştur. Kalma süresi arttıkça yatışkın koşula
gelme süresinin de arttığı gözlenmiştir. Düşük akış hızlarında 7 ila 8 saat arasında yatışkın koşula
ulaşılmıştır. Yüksek akış hızlarında ise daha kısa sürede (4 h) yatışkın koşula ulaşılmıştır. Kalma
süresinin 9 saat olduğu durumda % 86 dönüşüm ve % 99 enantiyomerik aşırılığa ulaşılmıştır.

Kalma süresinin 4.5 saat olduğu durumda ise % 79 dönüşüm ve % 99 enantiyomerik aşırılığa
ulaşılmıştır. Çalışılan en düşük kalma süresinde (2.25 saat) yani akış hızının çok yüksek olduğu
durumlarda dönüşüm % 55’ e düşmüştür. Dönüşümdeki bu düşüş enzim-substrat etkileşim süresinin
çok kısa olmasından yani dönüşüm için yeterli zaman olmamasından kaynaklanmaktadır.
Enantiyomerik aşırılık kalma süresinden etkilenmemektedir. Çalışılan tüm kalma sürelerinde, % 99
enantiyomerik aşırılık elde edilmiştir. Kalma süresinin verimlilik ve R-1-Feniletanol derişimi üzerine
etkisi incelenmiş, kalma süresi arttıkça, ürün derişimi artmış, ancak verimlilik azalmıştır. Yüksek
verimlilik ve yüksek ürün derişimi elde edebilmek için 3.75 h kalma süresinin ekonomik olduğu
bulunmuştur.
Sürekli sistemler, kesikli sistemlere göre sürekli işletilmeleri nedeniyle tercih edilmektedirler.
Çalışmada, dolgulu kolon reaktörün ne kadar süre işletilebileceği incelenmiştir. Tutuklanmış hücreler,
5 gün işletim süresince aktivetisinin % 90’ ını, 12 gün işletim süresince aktivetisinin % 50’ isini
korumaktadır.
Sonuç olarak, enantiyomerik saflıkta (> % 99) R-1fenil etanol üretimi, laboratuar ölçekte, tutuklanmış
mikroorganizma ile sürekli sistemde başarı ile gerçekleştirilmiştir.

Production of enantiomerically pure 1-phenylethanol using immobilized L.Kefir in a packed –bed reactor The conversion of substituted acetophenones to their corresponding optically active alcohols
(phenylethanols) is one of the most common reactions in organic chemistry. (R)-1-phenylethanol or
(S)-1-phenylethanol have a number of potential applications and used as building blocks for the
synthesis of bioactive compounds such as pharmaceuticals, agrochemicals and natural products. In this
project, alcohol dehydrogenases was used for bioreduction of acetophenone. ADHs catalyze the
enantioselective reduction of ketones for the synthesis of chiral alcohols with the help of nicotinamide
cofactors (NADH or NADPH). Whole cells rather than isolated enzymes were used preferentially to
avoid enzyme purification and cofactor addition or the requirement for an associate system for cofactor
regeneration, since bioreduction using isolated ADHs often require stoichiometric amounts of NADH
or NADPH . The most important advantage of using whole cells is that all of the reaction system for
cofactor regeneration is present within the cells themselves. Simple and cheap compounds such as
glucose or ethanol can be used as an energy source for cofactor regeneration when whole cells are used.
Above all, whole cells are easier to obtain and more cheaper than isolated enzymes. Therefore,
Lactobacillus kefir NRRL B-1839 was used for an enzyme source in our study. Continuous production
of R-1-phenylethanol was accomplished using immobilized Lactobacillus kefir continous packed bed
bioreactor.
Firstly, free Lactobacillus kefir was used in the experiments and results show that enantiomerically
pure (R)-1-Phenylethanol (ee > 99 % ) was produced. However, conversion values were low even if at
lower susbtrate concentartions (Co=1 mM, conversion % 80). Because of the lack of cofactor during
bioreduction, the conversion values were low. For solving cofactor regeneration problem, thre diffrent
cofactor regenation systems were used. These were: regenation using glucose, regenation using 2-
propanol and regenation using recombinant microorganism. The most effecient regenation systems was
found with glucose. When 2-propanol was used at 20 mM initial substrate concentration, conversion
and enantioselctivity was obtained as 95 % and 97% whereas using glucose, 100% and > 99 % was
obtained. For regeneration, lactate dehydrogenase show activity, however alcohol dehydrogenase did
not show activity in recombinant Escherichia coli. For this reason, glucose was used for cofactor
rejenaration for further experiments.

Before studying in continuous bioreactor, immobilized cells were used in a batch system for
determining the optimum conditions to produce (R)-1-phenylethanol. These were found as:
Temperature, 35 oC, pH 8 Tris buffer, % 2 w/v glucose for cofactor regeenration, time, 4 h. After than,
a pyrex glass column reactor 27 cm in length and 1.7 cm inner diameter with a water cooling jacket
was employed.
Several levels of substrate concentrations 15 mM, 25 mM and 40 mM were presented to determine
their effects on the conversion and enantiomeric excess and concentration of (R)-1-phenylethanol. The
concentration of (R)-1-phenylethanol was followed during 9 h along. At all studied substrate
concentrations, steady-state was reached approximately at 7 hours. Concentration of (R)-1-
phenylethanol increased with increasing substrate concentrations. At 4.5 h residence time, 12.8 mM
(R)-1-phenylethanol was obtained at 40 mM initial substrate concentration, whereas 11.2 mM (R)-1-
phenylethanol was produced at 15 mM initial substrate concentration (conversion 79%).
For industrial applications of asymmetric bioreduction, high product concentration and high
productivity are important. The productivity was 2.82 mmolL-1h-1 at 40 mM acetophenone
concentration whereas it was 2.50 mmolL-1h-1 at 15 mM acetophenone concentration. That means
higher productivity was obtained with increasing substrate concentration. On the contrary, conversion
decreased because of substrate inhibition. However, enantiomeric excess (> 99 % ) did not change with
substrate concentration.
During continuous reduction, the effect of glucose concentration on conversion and enantiomeric
excess and concentration of (R)-1-phenylethanol was also studied.Asymmetric bioreduction was
investigated at three different glucose concentration, 0.01, 0.02 and 0.03 g/mL. In this cases, the molar
ratios of glucose to acetophenone were obtained as 3.5, 7 and 11, respectively. The highest conversion
(79 %) was obtained at glucose concentration 0.02 g glucose /mL. When glucose concentration 0.03 g
glucose /mL, conversion decreased (34 %) significantly. It means that higher glucose concentration
caused inhibition on bioreduction. However, enantiomeric excess did not change with glucose
concentration.

The effect of residence time in continuous bioreactor was studied as 2.25 h, 3 h, 4.5 h and 9 h. High
enantiomeric excess (>99 %) was obtained at all studided residence times. The conversion increased
with increasing residence time at steady-state conditions. At 9 h residence time, conversion (86 %) ,
enantioselectivity (>%99) was obtained in our system. At 4.5 h residence time, conversion (79 %) ,
enantioselectivity (>%99) was obtained. The shortest residence time was 2.25 h in this study and the
conversion was %55. The conversion increased with increasing residence time at steady-state
conditions. When residence time increased, high (R)-1-phenylethanol concentration was obtained. This
causes a rise in the conversion because of the much more reaction time for substrate with the
biocatalysts in the column. When final conversions were compared between 4.5 h and 9 h residence
time, the values of conversions were close in range. Because of that experiments did not carried out
above 9 h residence time. It was obtained that product concentration increased with increasing
residence time. However, productivity decreased at the same conditions. Therefore, not only
conversion, but also productivity is important for industrial application. To reach both high
productivity and high concentration, 3.75 h was economically feasible residence time.
Continuous processes have a number of advantages over batch processes, including maximum reaction
rates, minimum nutrient depletion and product inhibition, higher space-time yields, low down-times for
the reactor, studying under steady-state conditions. In this study, the bioreactor operation time was
investigated. The immobilized cells was retain its 90 % activity duruing 5 days and 50 % activity
during 12days.
In conclusion, in laboratory scale, production of (R)-1-phenylethanol was achieved in a good
conversion and enantioselectivity (> % 99) in packed bed reactor.

II. Amaç ve Kapsam
Enantiyomerik saflıktaki kiral alkoller özellikle farmasötik sanayinde önemli ilaç etken maddesidir. Bu
çalışmada bir sekonder alkol olan 1-fenil 1-etanolün asimetrik indirgenme ile enantiyomerik saflıkta
elde edilmesi ilk kez sürekli çalışan dolgulu kolon biyoreaktörde gerçekleştirilmiştir. Enantiyomerik
saflıktaki 1-fenil etanol ve asetat türevleri farmasötik açıdan ilgi görmektedir.
(http://reference.md/files/D010/mD010626.html). Optik saflıktaki 1-fenil etanol, değerli kimyasalların
(fine chemicals) üretiminde kiral yapıtaşı olarak kullanılmaktadır. Örneğin, (R)-1-fenil etanol oftalmik
(gözle ilgili) koruyucu, kolestrol inhibitörü, solvatokromik boya olarak kullanılmaktadır (Suan vd.,
2004).
Çalışmada üretim yöntemlerinden, asimetrik indirgeme kullanılmıştır çünkü dönüşüm % 100
olabilmektedir. Steroseçimli asimetrik indirgeme tepkimelerinde, alkol dehidrojenazlar birçok
fonksiyonel grup üzerinde etkili olduğundan bu tip tepkimelerde tercih edilen enzimlerdir (Mertens vd.,
2003). Alkol dehidrojenazlar, geniş substrat aralığı (aldehitler, ketonlar, esterler) ve yüksek
stereoseçiciliğe sahiptirler. Organik çözücüde az çözünen substratlara etki etmezler. Alkol
dehidrojenazlar kofaktöre ihtiyaç duyarlar ve kofaktörler oldukça pahalı maddelerdir, enzim ve
kofaktörü organik çözücü ortamında veya su-organik ara yüzeyinde çok kararsızdır. Mikrobiyal
hücreler enzim kaynağı (ADH) olarak kullanıldığında, hücre içindeki kofaktör yeterli olabileceği gibi,
kofaktör rejenerasyonu ekonomik olarak gerçekleşebilmektedir (Villela Filho vd. 2003).
Bu proje kapsamında enantiyomerik saflıkta 1-fenil etanol üretimi ilk kez mikroorganizmanın
tutuklanarak enzim kaynağı olarak kullanıldığı sistemde gerçekleştirilmiştir.
Şekil 1. Asetofenonun Lactobacillus kefir ile asimetrik indirgenmesi tepkimesi

Dolgulu kolon biyoraktörde tutuklanmış Lactobacillus kefir NRRL B-1839 ile üretimin
gerçekleştirilmesinde en önemli avantaj enantiyoseçimliliğin ve % dönüşümün yüksek olmasıdır.
Çalışmada, öncelikle serbest ve tutuklanmış hücrelerle kesikli sistemde optimum koşullar (pH, tepkime
ortamı tasarımı, sıcaklık, süre) belirlenmiştir. Kesikli sistemde elde edilen optimum koşullar; dolgulu
kolonda sürekli üretime uygulanmış ve sürekli üretim için işletme koşullarının (akış hızı, substrat
derişimi, işletme süresi ve kofaktör rejenerasyonu için glikoz miktarı) enantiyoseçimlilik, dönüşüm ve
verimliliğe etkisi araştırılmıştır.
Yapılan çalışmalar sonucu, yüksek enantiyomerik saflıkta R-1-feniletanol üretimi, sürekli işetilen
biyoreaktörde başarı ile gerçekleştirilmiştir. Laboratuar ölçekte gerçekleştirilen bu çalışma, endüstriyel
üretim içinde ışık tutmaktadır.

III. Materyal ve Yöntem
Mikroorganizma ve Üretim Koşulları Bu çalışmada, asetofenonun asimetrik biyoindirgenmesi için alkol dehidrojenaz kaynağı olarak
Lactobacillus kefir NRRL B-1839 mikroorganizması olarak kullanılmıştır. Mikroorganizma United
States Department of Agriculture (US)’ dan temin edilmiştir. Mikroorganizma üreme ortamının
bileşimi Tablo 2 de görülmekte ve pH’ sı 0.5 M H2SO4 ile 7’ e ayarlanan ortama 1/10 oranında aşılanan
Lactobacillus kefir, 24 saat boyunca 35 oC’ de orbital çalkalayıcıda karıştırılmıştır.
Tablo 1. Lactobacillus kefir NRRL B-1839’ a ait üreme ortamının bileşimi
Kimyasal Miktarı, g/L Glukoz·H2O 22 Kazein pepton 10 Sodyum asetat 5 Et özütü 10 Maya özütü 5 Tween 80 1 K2HPO4 2 MgSO4 ·7H2O 0.2 MnSO4 ·H2O 0.05
Alkol dehidrojenaz, kofaktör varlığında aktivite gösterebilen bir enzimdir. Kofaktörler çok pahalı
olduklarından stokiyometrik miktarda kullanılmaları ekonomik değildir. Bu nedenle kofaktör
rejenerasyonu gerekli olmaktadır. Çalışmada, kofaktör rejenerasyonu için laktat dehidrojenaz geni
içeren, rekombinant Escherichia coli ile kofaktör rejenerasyonu gerçekleştirilmeye çalışılmıştır.
Rekombinant Escherichia coli’ deki, laktat dehidrojenaz enzimi de kofaktör ile aktivite
gösterebilmektedir. Ancak, laktat dehidrojenaz enzimi, ADH’ ın kullandığı kofaktörü kullanmakta ve
tekrar ADH’ ın kullanabileceği forma dönüştürmektedir. Özetlersek, ADH enzimi ile yükseltgenen
kofaktör (S1 + NADH � Ü1 + NAD+), LDH enzimi tarafından kullanılacak ve kofaktör
yükseltgenecektir (S2 + NAD+ � Ü2 + NADH). İndirgenen kofaktör, tekrar ADH enzimi ile
yükseltgenecek ve böylelikle kofaktör rejenerasyonu sağlanacaktır. Bu sebeble, Gebze Yüksek
Teknoloji Enstitüsü’ nden temin eldilen, laktat dehidrojenaz aktivitesine sahip, rekombinant
Escherichia coli kullanılmıştır. Rekombinant E.coli, bileşimi Tablo 3 de verilen 1 M NaOH ile pH’ sı
7’ ye ayarlanan üreme ortamında üretilmiştir. Steril ortama mikroorganizma aşılanmadan önce 100
µg/mL amfisilin ilave edilmiştir. 1/10 oranında aşılanan E. Coli 16 saat boyunca 37 oC’ de, 100 rpm
karıştırma hızında orbital çalkalayıcıda çoğaltılmıştır.

Tablo 2. Rekombinant Escherichia coli’ ye ait üreme ortamının bileşimi (Lurai broth ortamı)
Kimyasal Miktarı, g/L Pepton 10 Maya özütü 5 Sodyum klorür 10
ADH Aktivitesinin Belirlenmesi Bu çalışmada Lactobacillus kefir NRRL B-1839 mikroorganizması enzim kaynağı olarak
kullanılmıştır. Alkol dehidrojenaz hücre içi bir enzimdir. Aktivitesini ölçebilmek için enzimin hücre
dışına alınması gerekmektedir. Alkol dehidrojenaz enzim aktivitesini belirlemek için, mikroorganizma
çoğaltıldıktan sonra, santrifüjlenmiş, pH 7 olan tamponda, ultrasonikatör (10 dakika ses ötesi dalga
verilerek, 30 saniye ses ötesi dalga, 30 saniye bekleme) ile hücreler parçalanmıştır. Sıvı kısım, aktivite
belirlemek için kullanılmıştır. Aktivite belirlemek için, etanolün, asetaldehide, kofaktör varlığında
oksidasyon tepkimesi UV spektrofotometrede takip edilmiştir. Kofaktör (NAD, NADH, NADP veya
NADPH), 340 nm de absorbans vermektedir. Bu tepkimeye göre, absorbansda artış takip edilmiştir.
ADH
Etanol + β-NAD � Asetaldehit + β-NADH LDH Aktivitesinin Belirlenmesi Bu çalışmada rekombinant Escherichia coli mikroorganizması laktat dehidrojenaz enzim kaynağı
olarak kullanılmıştır. Laktat dehidrojenaz hücre içi bir enzimdir. Aktivitesini ölçebilmek için enzimin
hücre dışına alınması gerekmektedir. Enzim aktivitesini belirlemek için, mikroorganizma çoğaltıldıktan
sonra, santrifüjlenmiş, pH 7 olan tamponda, ultrasonikatör (10 dakika ses ötesi dalga verilerek, 30
saniye ses ötesi dalga, 30 saniye bekleme) ile hücreler parçalanmıştır. Sıvı kısım, aktivite belirlemek
için kullanılmıştır. Aktivite belirlemek için, laktatın, pirüvata, kofaktör varlığında oksidasyon
tepkimesinin absorbansı 340 nm de UV spektrofotometrede takip edilmiştir.
LDH
L-laktat + β-NAD � Pirüvat + β-NADH

Kofaktör rejenerasyonu için ADH ve LDH genlerinin Escherichia coli ye klonlanması
Lactobacillus kefir mikroorganizmasındaki ADH geni ile Escherichia coli’ deki LDH geni bu
mikroorganizmalardan izole edilmiş ve Escherichia coli’ ye klonlanarak enzimlerin tek bir
mikroorganizma içinde aktivite göstermesi amaçlanmıştır. Böylece kofaktör rejenerasyonu
sağlanabilecektir. Bu amaçla, Gebze Yüksek Teknoloji Enstitüsü Çevre Mühendisliği Bölümü öğretim
üyelerinden Doç. Dr. Melek Özkan ile birlikte klonlama gerçekleştirilmiştir. Öncelikle,
mikroorganizmalardan (Lactobacillus kefir ve Escherichia coli) genomik DNA izolasyonu aşağıda
anlatıldığı gibi gerçekleştirilmiştir.
Lactobacillus kefir ve Escherichia coli den Genomik DNA İzolasyonu
ADH genini içeren Lactobacillus kefir, tablo 1 da verilen üreme ortamında üretildikten sonra,
santrifüjlenerek ayrılmıştır. Daha sonra, aşağıdaki adımlar yapılarak genomik DNA izole edilmiştir.
1. Bir öze yardımı ile mikroorganizma eppendorf tüpüne alınır ve üzerine üzerine Tris-EDTA
tamponu, (pH 7) (0,5mL)/lizozim(5 mg/mL olacak şekilde) eklenir ve süspansiyon haline
getirilir.
2. Bu süspansiyona 2-3 µl proteinaz K ve 100 µl % 10’ luk sodyumdodesilsülfat eklenir ve 37 oC
de 15 dakika bekletilir. Tüp alt üst edilerek karıştırılır.
3. 100 µl, 5 M NaCl eklenir ve iyice karıştırılır.
4. 100 µl, % 10 luk heksadesiltrimetil aminobromür/0,7 M NaCl karışımından ilave edilir. İyice
karıştırılır. Daha sonra 65 oC de 10 dakika bekletilir.
5. Üzerine eşit hacimde kloroform/izoamil alkol (24/1) eklenir ve alt üst edilerek iyice karıştırılır.
Karışım daha sonra 10000 rpm de 5 dakika kadar santrifüj edilir.
6. Üst faz başka bir tüpe alınır ve üzerine eşit oranda fenol/kloroform/izoamilalkol (24:24:1)
eklenir. 10000 rpm de 10 dakika santrifüjlenir, süpernatant atılır ve tüp hemen ters çevrilerek
oda sıcaklığında kurutulması sağlanır.
7. Oluşan pelet üzerine 100 µl Tris-EDTA tamponu konur ve iyice süspansiyon haline gelmesi
sağlanır. İzole edilmiş olan bu DNA -20 oC de saklanır.

LDH genini içeren Escherichia coli tablo 2 da verilen üreme ortamında üretildikten sonra, aşağıdaki
adımlar yapılarak genomik DNA izole edilmiştir.
1. Escherichia coli içeren kültür eşit oranlarda 1,5 mL’ lik eppendorf tüplere aktarılır ve 6000
rpm’ de 3 dakika santrifüj edilir. Santrifüj sonunda süpernatant kısım dökülür.
2. Pelet, 200 µl GET solüsyonunda (50 mM Glukoz, 25 mM Tris-HCl, 10 mM EDTA, pH 8.0)
çözülür. Karışımın homojen hale gelmesi sağlanır ve +4 oC de bekletilir.
3. 200 µl SDS/NaOH (0.2 M NaOH + 1% SDS) çözeltisinden ilave dilir. İyice karıştırılır ve oda
sıcaklığında 5 dakika tutulur.
4. 200 µl Potasyum asetat/formik asit ilave edilir ve 2-3 dakika kadar karıştırılır. Oda sıcaklığında
10 dakika kadar bekletilir.
5. Süspansiyon 13000 rpm’ de 8 dakika kadar santrifüjlenir. Süpernatant (plazmid DNA’ nın
bulunduğu kısım) dikkatli bir şekilde temiz bir tüpe aktarılır. Proteinlerden oluşan beyaz renkli
peletin, plazmid DNA’ yı ihtiva eden supernatanta karıştırılmamasına özen gösterilmelidir.
6. Elde edilen DNA süspansiyonunun % 10 u kadar sodyum asetat ve 2.5 katı kadarda etanol ilave
edilerek -35 oC de 30-60 dakika kadar tutulur.
7. Plazmid DNA solüsyonu 13000 rpm de 10 dakika kadar santrifüj edilir ve süpernatant dökülür
(pelete dokunulmamalı)
8. Kurutulan pelet 250 µl steril su veya Tris-EDTA (10 mM Tris-HCl, 0.1 mM EDTA, pH 7.6) ile
çözülür ve -20 oC de saklanır.
Agaroz Jel Elektroforezi
DNA moleküllerinin izolasyonlarında ve DNA’ nın görüntülenmesinde genellikle, agaroz jel
elektroforezi kullanılmaktadır. Elektroforezde, küçük molekül yapısına sahip DNA bantları ağır
olanlardan daha hızlı bir şekilde hareket ederek farklılaşır. Hazırlanan jeldeki agar oranı % 1 w/v dir.
Elektroforez için uygulanan adımlar aşağıda verilmiştir.

1. Sıvı solüsyona (50 mM Tris borat, 1 mM EDTA, 0.5µg/mL Etidyum bromür), agar (1 g/100
mL) ilave edildikten sonra agarozun çözülmesi için solüsyon kapağı gevşek bir şişe içerisinde
kaynar su banyosunda kaynatılır.
2. Daha sonra, 50 oC ya kadar soğutulduktan sonra agar tankına dökülür.
3. Jelin bir tarafına elektroforez tarağı yerleştirilir. 25-35 dakika sonra katılaşan jelden tarak
dikkatlice çıkarılır ve jel tanktan alınarak sıvı içeren elektroforez tankına yerleştirilir.
4. Tarak dişlerinin oluşturduğu yuvalara incelenecek DNA örnekleri 10 µl olarak aktarılır. Bu
arada molekül büyüklükleri bilinen marker DNA’ larda jelin bir yuvasına yerleştirilir. DNA
süspansiyonunun elektroforez uygulaması sırasında örneklerin jelin sonuna doğru hareket edip
etmediğini anlamak için örneklere bir miktar boya maddesi ilave edilmiştir. Bu boya çözeltisi %
0,25 bromofenol mavisi (metilen mavisi) % 30 gliserol, %0,25 xylene cyanolden oluşmaktadır.
Böylece agaroz jel içerisindeki DNA bantları görünür hale gelir.
6. Güç kaynağından elektroforez cihazına uygun miktarda akım vererek DNA moleküllerinin
hareketi sağlanır.
7. Jeldeki yürütme işlemi, bromofenol mavisi uygun uzaklığa ilerleyene kadar sürdürülür.
8. Jeldeki DNA bantlarının aydınlatılması ultraviyole ışık kaynağı transillüminatör kullanılarak
yapılır.
Polimeraz Zincir Reaksiyonu
Polimeraz zincir reaksiyonu elde edilen DNA’ nın çoğaltılması amacıyla gerçekleştirilir. Burada çift
sarmallı DNA’ nın ısıtılarak iplikçiklerin birbirinden ayrılması sağlanır. Reaksiyon, eppendorf bir tüp
(0,5 mL’ lik) içersinde toplam 50 µL lik bir solüsyon olacak şekilde 1.25 ünite Taq DNA polimeraz,
0,5 ng kaynak DNA, 0,1 ng sentetik primerler, 200 µM her bir dNTP ve 10 mM Tris-HCl, 50 mM KCl,
2 mM MgCl2, 35 döngü olacak şekilde (94 oC de 1 dakika, 55 oC de 30 saniye ve 72 oC de 1 dakika)
tamamlanmıştır. Reaksiyonda kullanılan MgCl2 polimeraz enziminin aktivitesinde görev almaktadır.
PCR tamamlandıktan sonra tekrar agaroz jel elektroforezi gerçekleştirilmiştir. Sonra istenilen kDa
karşılık gelen bantlar kesilerek -20 oC de saklanmıştır.

Alıcı (Competent) Hücrenin Hazırlanması
1. Bu amaçla, E.coli’nin DH5 α, suşu kullanılmıştır.
2. Stok E.coli kültüründen katı Luria agar besiyerine aşılama yapılır.
3. Aşılanmış petri kutuları 12-16 saat kadar 37 oC de inkübasyona bırakılır.
4. Gelişen kolonilerden biri, 5 mL Lurai broth (LB) sıvı besiyerine aşılandıktan sonra 5-6 saat
kadar 37 oC de inkübasyona bırakılır.
5. Yukarıda belirtilen süre sonucunda oluşan kültürden 0,1 mL alınıp 50 mL’ lik LB besiyerine
aşılanır ve 37 oC de optik densitesi (OD 550 nm) 0.2-0.4 ulaşıncaya kadar inkübasyona
bırakılır. Buradan alınan kültür 10-20 dakika kadar buz üzerinde bekletilir.
6. Kültür örneğinden eşit oranlarda santrifüj tüplerine aktarılır ve 6000 rpm’ de 5 dakika kadar
santrifüj edilir. Santrifüj sonucunda oluşan supernatant dökülür, hücrelerden ibaret olan pelet
2.5 mL CT solüsyonunda (CaCl2/Tris) çözülür ve yaklaşık 45 dakika kadar +4 oC de bekletilir.
Bu durumdaki hücreye competent hücre adı verilir ve transformasyon için hazırdırlar.
7. Örnekler 6000 rpm de 5 dakika santrifüj edildikten sonra supernatant dökülür ve pelet 0.5 mL
CT içerisinde hafif bir şekilde çözülür.
8. Hazırlanan bu hücre süspansiyonunun 100 µL sine, 10µL DNA eklenir, nazikçe çalkalanır.
Hücreler, 30 – 40 dakika buzda bekletilerek, DNA’ nın hücre içine girmesi beklenir.
9. Hücreler, ısı şokuna tutulur, 42oC su banyosunda 45-60 saniye tutulur ve 1 dakida buz üzerinde
bekletirilir.
10. Hücrelerin üzerine, 1 mL LB ortamı ilave edilir ve 37oC 1-2 saat hafifçe çalkalanarak inkübe
edilir.
11. Daha sonra Lurai-agar ortama ekilerek, 37oC 1 gece boyunca kloni oluşumu izlenir.

Mikroorganizmanın Ayrılması
Üretim ortamında üreyen mikroorganizmalar yüksek hızda (9500 rpm) 15 dakika, -4oC de
santrifüjlenerek ayrılmıştır.
Tutuklama Yöntemi (κ-Karragenan jel taneciklerinin oluşturulması)
Hücreler, κ-Karragenan jeline tutuklanmıştır. κ-Karragenan-hücre karışımı KCl çözeltisine damlatıldığı
zaman hücreler jel içerisine hapsolmaktadır (Wang, W. Y., Hetter, D.J. 1982. Cell Immobilization in k-
carragenenan with Tricalcium Phosphate, Biotechnology and Bioengineering 24: 1827-1838). % 1’ lik
κ-Karragenan çözeltisi, 50 oC de, damıtık suda çözülerek hazırlanmış ve içerisine istenilen oranda (16
gram) yaş hücre eklenerek homojenlik sağlanıncaya kadar karıştırılmıştır. Karışım, devamlı
karıştırılarak 1 M KCl çözeltisine 1 mL’ lik pipet ucundan peristaltik pompa kullanılarak
damlatılmıştır. Tutuklanmış hücre içeren κ-Karragenan jeli taneciklerinin tam olarak olgunlaşması için
2 saat 0.1 M KCl çözeltisinde bekletilmiştir. Pelletler daha sonra kullanılmak üzere 0.1 M KCl
çözeltisine alınmış ve -4oC de saklanmıştır.
Hücreler, kalsiyum-alginat jeline tutuklandığında ise; % 1’ lik Na-alginat çözeltisi, 50 oC de, damıtık
suda çözülerek hazırlanmış ve içerisine istenilen oranda (12 gram) yaş hücre eklenerek homojenlik
sağlanıncaya kadar karıştırılmıştır. Hücre ve Na-alginat karışımı devamlı karıştırılarak 0.27 M CaCl2
çözeltisine oda sıcaklığında 1 mL’ lik pipet ucundan peristaltik pompa kullanılarak damlatılmıştır.
Tutuklanmış hücre içeren Ca-alginat jeli taneciklerinin tam olarak olgunlaşması için 2 saat 0.2 M CaCl2
çözeltisinde bekletilmiştir. Pelletler daha sonra kullanılmak üzere süzülerek 0.027 M CaCl2 çözeltisine
alınmış ve -4oC de saklanmıştır.

Şekil 2: κ-Karragenana tutuklanmış Lactobacillus kefir hücrelerinin oluşturulması
Tepkime
Şekil 3. Asetofenonun asimetrik indirgenmesiyle optikçe saf 1-feniletanol üretimi
Sıvı üreme ortamı
Santrifüj
Mikroorganizma κ-Karragenan çözeltisi
κ-Karragenan -mikroorganizma
çözeltisi
Pompa
1 M KCl çözeltisi
Tutuklanmış hücre

Analiz Koşulları
Tepkime ortamından alınan örnekler, eş hacim metil tersiyer bütil eter ile ekstrakte edilmiştir.
Organik ve sulu fazlar ayrıldıktan sonra organik faz yüksek basınç sıvı kromatografi ile, analiz
edilmiştir. Taşıyıcı faz olarak, hegzan/2-propanol (95/05), akış hızı 0.90 mL/dak, 10 µL enjeksiyon
hacmi, 30 oC’ de Chiralcell OB kolonu (4.6 mm x 50 mm, Daicel Chemical Ind. Ltd. France) 254
nm UV dedektör ile analizlenmiştir. Yüksek Basınçlı Sıvı Kromatografi (HPLC) sistemi Şekil 4 ve
yukarıdaki koşullarda gerçekleştirilen analize ait kromatogram IV. Analiz ve Bulgular bölümünde
verilmiştir.
Şekil 4. Yüksek Basınçlı Sıvı Kromatografi (HPLC) sistemi

Sürekli sistemde (R)-1-feniletanol üretimi
Enantiyomerik saflıkta R-1-feniletanol üretimi boyutları 1.7 cm x 27 cm olan cam kolon biyoreaktörde,
3 mm tanecik çapında κ-Karragenana tutuklanmış L. kefir hücreleriyle gerçekleştirilmiştir. Yüksek
dönüşüm ve enantiyomerik aşırılık elde etmek amacıyla, substrat derişimi, glukoz derişimi, kolonda
kalma süresi etkisi incelenmiştir. Tutuklanmış Lactobacillius kefir deneye başlamadan önce kolona
doldurulmuş ve kolon gözenekliliğine bakılmıştır. Kolon gözenekliliği tespit edildikten sonra incelenen
kalma sürelerinde akış hızları belirlenmiştir. Besleme çözeltisi; substrat (50 mM pH 8 Tris tamponu
içinde), glukoz (kofaktör rejenerasyonu için) ve KCl (0.05 mM tutuklanmış mikroorganizmanın
mekanik kararlılığı için) karışımı kolona alttan bir peristaltik pompa yardımıyla beslenmiştir. Ürün ise
kolonun üst çıkışından alınmıştır. Kolonda istenen sıcaklık ceketten geçen su yardımıyla sağlanmıştır.
Her saat başı zaman aralıklarında alınan örnekler HPLC ile analiz edilmiştir. Sürekli sisteme ait deney
düzeneği şekil 5 de verilmiştir.
Şekil 5.Dolgulu kolon biyoreaktör deney düzeneği

Kolonda kalma süresinin belirlenmesi
Dolgulu kolon biyoreaktörde çalışmaya başlamadan önce istenen kalma süresinde akış hızlarının tespiti
için kolon gözenekliliğine bakılmıştır. Bu amaçla kolon içindeki katı (tutuklanmış mikroorganizma),
hacmi belli olan bir miktar suyun içine boşaltılır ve suyun hacmindeki artış belirlenir. Aşağıdaki formül
yardımıyla kolon gözenekliliği hesaplanmıştır.
o
ko
V
VVε
−=
ε : Kolon gözenekliliği Vo: Kolon hacmi, mL Vk: kolon içindeki katı (tutuklanmış mikroorganizma), mL Kolon gözenekliliği tespit edildikten sonra tepkime hacmi (Eşitlik ) bulunmuş ve bu değer yardımıyla
kalma süresine (Eşitlik) geçilmiştir.
V=Vo* ε V: Tepkime hacmi, mL
Q
Vτ =
τ : Kalma süresi, dak Q: Akış hızı, mL/dak

IV. Analiz ve Bulgular
Asimetrik biyoindirgeme ile asetofenonun biyoindirgenmesi sonucu R-1-feniletanol üretimiştir.
Çalışmada aşağıdaki şekilde gerçekleştirilmiştir.
1. Lactobacillus kefir hücrelerinin (serbest hücre) üretildiği besi ortamında (hücreler büyüdükten
sonra) gerçekleştirilen deneyler
2. Lactobacillus kefir hücrelerinin (serbest hücre) üretildiği besi ortamına glukoz ilavesi (kofaktör
rejenerasyonu için) yapıldıktan sonra gerçekleştirilen deneyler
3. Lactobacillus kefir hücrelerinin (serbest hücre) üretildiği besi ortamına 2-propanol ilavesi
(kofaktör rejenerasyonu için) yapıldıktan sonra gerçekleştirilen deneyler
4. Tutuklanmış Lactobacillus kefir hücreleriyle kesikli sistemde gerçekleştirilen deneyler
5. Lactobacillus kefir ve Escherichia coli ile kofaktör rejenerasyonu
6. Tutuklanmış Lactobacillus kefir hücreleriyle sürekli sistemde gerçekleştirilen deneyler
Lactobacillus kefir hücrelerinin (serbest hücre) üretildiği besi ortamında (hücreler büyüdükten
sonra) gerçekleştirilen deneyler
İlk olarak asimetrik biyoindirgeme deneyleri; Lactobacillus kefir hücrelerinin 24 saat üretildiği besi
ortamında (bileşimi Tablo da verilen üreme ortamı), gerçekleştirilmiştir. Burada, hücreler üretildikten
sonra (24 saat sonunda) ortama substrat (asetofenon) istenilen derişimde olacak şekilde ilave edilmiştir.
Hücrelerin üretildiği ortam, zengin ortam olarak adlandırılmıştır. Bu ortamda, başlıca karbon kaynağı
olarak; glukoz, diğer besin maddeleri (pepton, et özütü, vb.) ve diğer tuzları (Na+, K+, Mg+2 ve Mn+2,
vb.) içermektedir. Yapılan bu deneylerde, öncelikle farklı başlangıç substrat derişimlerinde (1 mM, 5
mM, 10 mM, 15 mM, 20 mM, 25 mM) çalışılmıştır. Sıvı üreme ortamında (50 mL) üretilen
mikroorganizma üstel üreme bölgesine ulaştıktan sonra (24 saat), ortama dimetil sülfoksitde çözülen
substrat (asetofenon) ilave edilmiştir. Burada, üreme ortamındaki mikroorganizma derişimi, 0.01g / mL
dir. Substrat ilave edildikten sonra, 35 oC sıcaklık ve 150 rpm karıştırma hızında, deneyler
gerçekleştirilmiştir. Ulaşılan % dönüşüm ve % ee değerleri Şekil 6 da verilmiştir.

0
20
40
60
80
100
1 5 10 15 20 25
Substrat derişimi, mM
% e
e v
e %
dö
nü
şü
m
% dönüşüm
% ee
Şekil 6. Serbest hücrelerle asimetrik biyoindirgemeye substrat derişimi etkisi ( Cx=0.01g / mL T= 35 oC, N=150 rpm)
Lactobacillus kefir hücrelerinin, bu tepkime için yüksek enantiyoseçimlilik gösterdiği ve R-
enantiyomer için yüksek enantiyomerik aşırılık verdiği şekilden görülmektedir. Burada, substrat
derişimi arttıkça dönüşümde artış olmamıştır. Çalışılan en küçük substrat derişimi olan 1 mM bile
dönüşüm % 100 değildir. Bu dönüşümü artırmak için, sıcaklık etkisi 10 mM başlangıç substrat
derişiminde, farklı sıcaklıklarda tekrar gerçekleştirilmiştir. Her enzimin aktif olduğu bir optimum
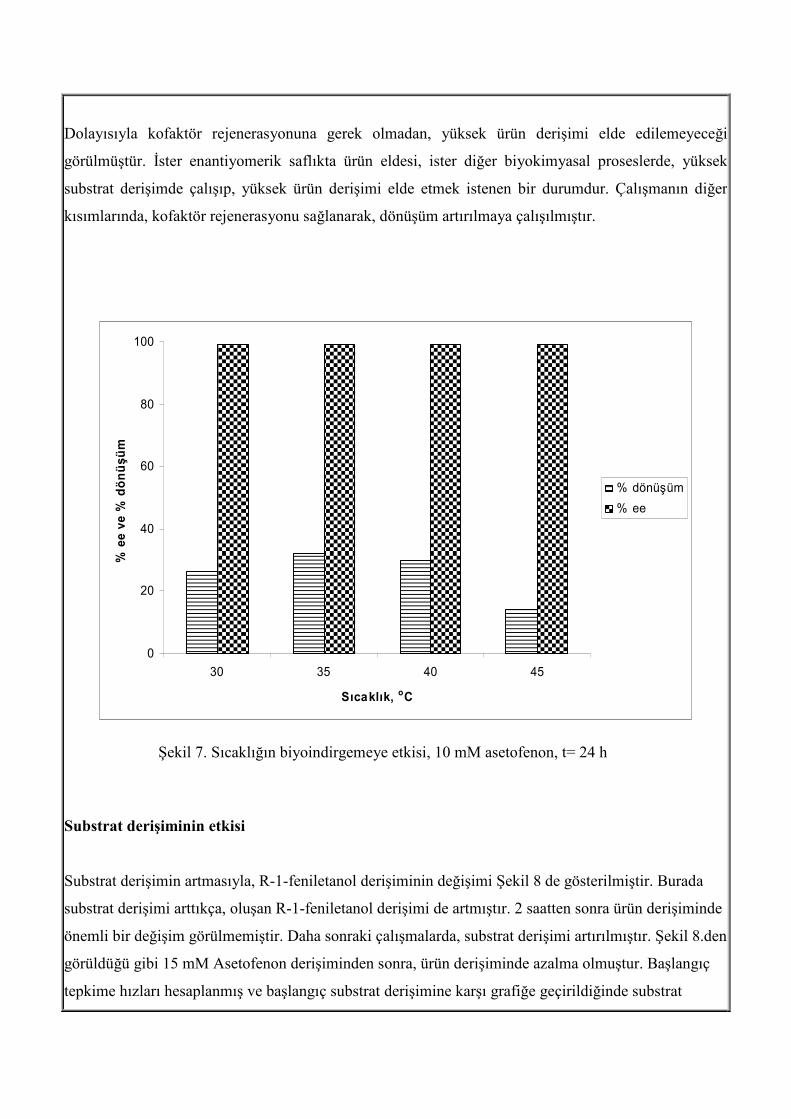
sıcaklık aralığı vardır. Sıcaklık, genel olarak tepkime hızını artırır. Ancak yüksek sıcaklıklar, enzimi
deaktive eder. Bu yüzden, sıcalık aralığı olarak, 30 oC, 35 oC 40 oC ve 45 oC aralığı incelenmiştir (şekil
7). Enantiyomerik aşırılık değerleri her sıcaklıkta yüksekken, dönüşüm değerleri farklıdır. Burada sabit
substrat derişiminde (10 mM) çalışılmıştır. Literatürde, biyoindirgeme tepkimeleri genellikle 30 oC ila
35 oC tercih edilmektedir. Asetofenonun indirgenmesinde, 30 oC ila 40 oC arasında ulaşılan ürün
derişim ve % dönüşüm değerleri birbirlerine yakın, fakat 35 oC’ nin diğer sıcaklıklara göre daha yüksek
dönüşüm verdiği görülmüştür. Ancak, dönüşüm, % 36 da kalmıştır. Öncelikle, saf enzim değilde,
mikroorganizma kullanıldığında, büyüme ortamında bulunan hücrelerin, kofaktör rejenerasyonunu
kendi, kendine yapabilmesi düşünülmüştür. Bu yüzden ek bir rejenerasyon yapılmasına gerek
duyulmamıştır. Fakat görülmüştür ki, burada, % dönüşümü artırmak mümkün olmamıştır.
Dolayısıyla kofaktör rejenerasyonuna gerek olmadan, yüksek ürün derişimi elde edilemeyeceği
görülmüştür. İster enantiyomerik saflıkta ürün eldesi, ister diğer biyokimyasal proseslerde, yüksek
substrat derişimde çalışıp, yüksek ürün derişimi elde etmek istenen bir durumdur. Çalışmanın diğer
kısımlarında, kofaktör rejenerasyonu sağlanarak, dönüşüm artırılmaya çalışılmıştır.
0
20
40
60
80
100
30 35 40 45
Sıcaklık, oC
% e
e v
e %
dö
nü
şü
m
% dönüşüm
% ee
Şekil 7. Sıcaklığın biyoindirgemeye etkisi, 10 mM asetofenon, t= 24 h
Substrat derişiminin etkisi Substrat derişimin artmasıyla, R-1-feniletanol derişiminin değişimi Şekil 8 de gösterilmiştir. Burada
substrat derişimi arttıkça, oluşan R-1-feniletanol derişimi de artmıştır. 2 saatten sonra ürün derişiminde
önemli bir değişim görülmemiştir. Daha sonraki çalışmalarda, substrat derişimi artırılmıştır. Şekil 8.den
görüldüğü gibi 15 mM Asetofenon derişiminden sonra, ürün derişiminde azalma olmuştur. Başlangıç
tepkime hızları hesaplanmış ve başlangıç substrat derişimine karşı grafiğe geçirildiğinde substrat

inhibisyonu olduğu görülmüştür (Şekil 8.). Model sabitlerinin bulunması için Lineweaver Burk
doğrusallaştırması yapılmıştır.
0
0,5
1
1,5
2
2,5
3
3,5
4
0 10 20 30 40 50 60 70 80
süre, h
CR
-1fe
nil
eta
no
l
1 mM
5 mM
10 mM
15 mM
20 mM
25 mM
Şekil 8. Substrat derişimi ile R-1-feniletanol derişiminin değişimi
Başlangıç tepkime hızları aşağıda verilen hız tanımından hesaplanmıştır (Fogler 1992).
o
FERo dt
dCr
= −
Şekil 8 de başlangıçtan geçen teğetin eğimi, başlangıç tepkime hızını verecektir.
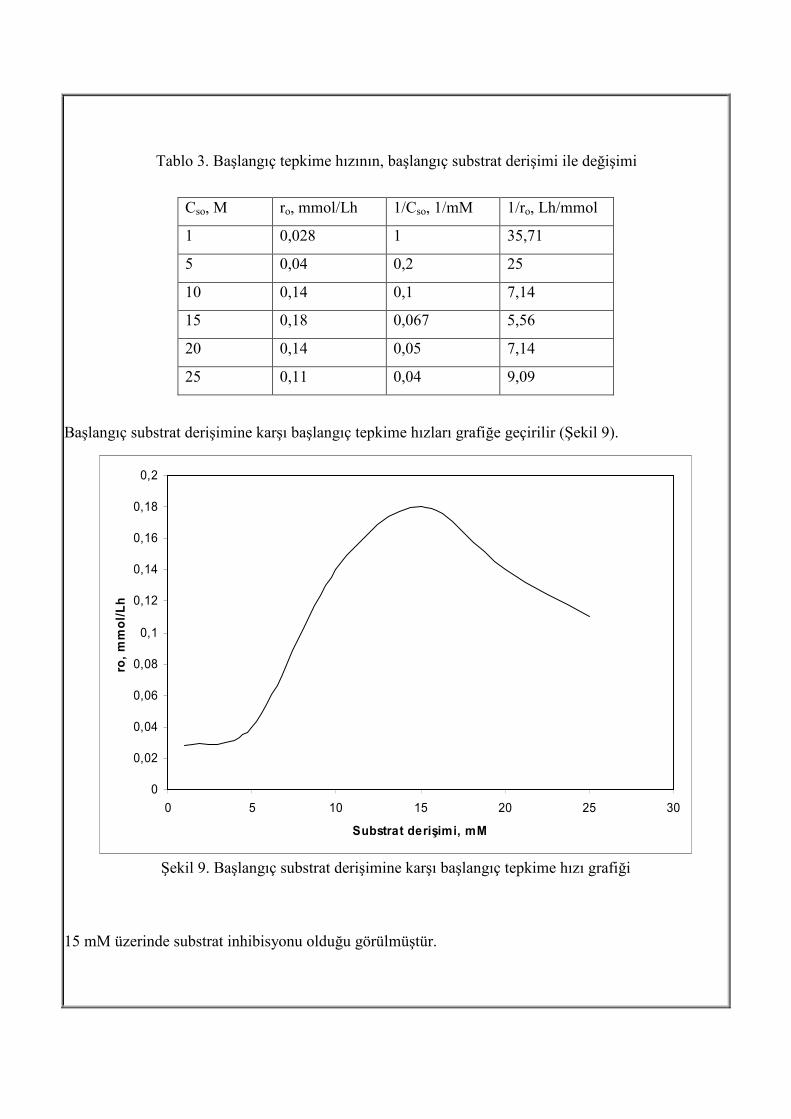
Tablo 3. Başlangıç tepkime hızının, başlangıç substrat derişimi ile değişimi
Cso, M ro, mmol/Lh 1/Cso, 1/mM 1/ro, Lh/mmol
1 0,028 1 35,71
5 0,04 0,2 25
10 0,14 0,1 7,14
15 0,18 0,067 5,56
20 0,14 0,05 7,14
25 0,11 0,04 9,09
Başlangıç substrat derişimine karşı başlangıç tepkime hızları grafiğe geçirilir (Şekil 9).
0
0,02
0,04
0,06
0,08
0,1
0,12
0,14
0,16
0,18
0,2
0 5 10 15 20 25 30
Substrat derişimi, mM
ro,
mm
ol/
Lh
Şekil 9. Başlangıç substrat derişimine karşı başlangıç tepkime hızı grafiği
15 mM üzerinde substrat inhibisyonu olduğu görülmüştür.
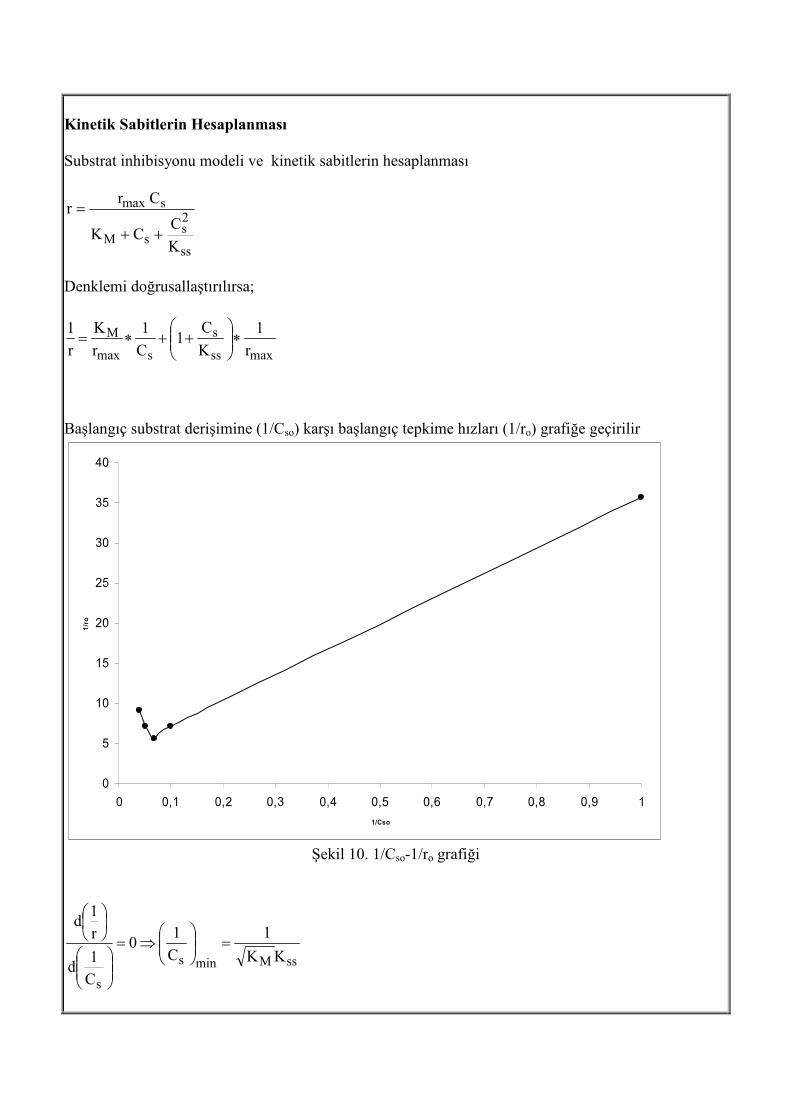
Kinetik Sabitlerin Hesaplanması Substrat inhibisyonu modeli ve kinetik sabitlerin hesaplanması
ss
2s
sM
smax
K
CCK
Crr
++
=
Denklemi doğrusallaştırılırsa;
maxss
s
smax
M
r
1
K
C1
C
1
r
K
r
1∗
++∗=
Başlangıç substrat derişimine (1/Cso) karşı başlangıç tepkime hızları (1/ro) grafiğe geçirilir
0
5
10
15
20
25
30
35
40
0 0,1 0,2 0,3 0,4 0,5 0,6 0,7 0,8 0,9 1
1/Cso
1/r
o
Şekil 10. 1/Cso-1/ro grafiği
ssMmins
s
KK
1
C
10
C
1d
r
1d
=
⇒=

Şekil 10 ‘den 065,0=C
1
mins
okunur.
Düşük substrat derişimlerinde ss
s
K
C<< 1 olduğundan;
maxsmax
M
r
1
C
1
r
K
r
1+∗= olur.
Düşük substrat derişimlerine (1/Cso) karşı başlangıç tepkime hızları (1/ro) grafiğe geçirilir. Elde edilen
doğrunun kayması rmax’ ı, eğimi ise KM’ yi verir (Şekil 11).
1/r = 32,05(1/Cs)x + 3,6691
R2 = 0,9998
0
5
10
15
20
25
30
35
40
0 0,2 0,4 0,6 0,8 1 1,2
1/Cso
1/r
o
Şekil 11. Düşük substrat derişimlerine (1/Cso) karşı başlangıç tepkime hızları (1/ro)
66,3=r
1=Kayma
max
hL/mmol27,0=rmax ’ dır.

Eğim 05,32=r
K=
max
M
L/mmol65,8=KM ’ dır.
ssssMmins K5,2
1=065,0ise
KK
1=
C
1
L/molm36,27=K ss ’ dır.
Kinetik sabitler model denkleminde yerine yazılır.
36,27
C+C+65,8
C27,0=r 2
ss
s
Lactobacillus kefir hücrelerinin (serbest hücre) üretildiği besi ortamına glukoz ilavesi (kofaktör
rejenerasyonu için) yapıldıktan sonra gerçekleştirilen deneyler
Lactobacillus kefir hücrelerinin (serbest hücre) üretildiği besi ortamında biyoindirgeme
gerçekleştirildiğinde, düşük başlangıç substrat derişimlerinde bile dönüşüm kısıtlı olmaktadır. Alkol
dehidrojenazlar, kofaktör gerektiren enzimlerdir. Kofaktör tükendiğinde veya rejenerasyon
olmadığında tepkime yürümemektedir. Bu yüzden, glukoz ilavesi (kofaktör rejenerasyonu için) ile
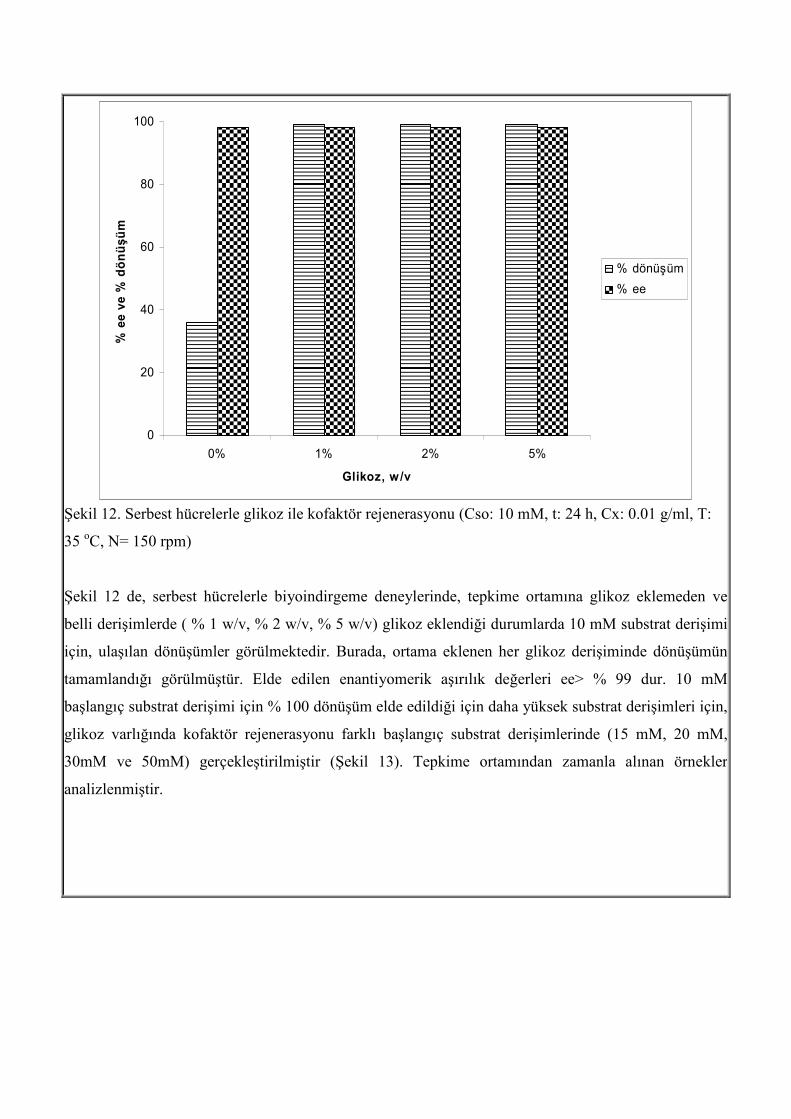
dönüşümün artırılması incelenmiştir. Burada kofaktör rejenerasyonu, glikoz ile sağlanmıştır. Serbest
Lactobacillus kefir hücrelerinin bulunduğu ortama %1 w/v, % 2 w/v, %5 w/v glikoz eklenmiş ve
tepkimeler gerçekleştirilmiştir.
0
20
40
60
80
100
0% 1% 2% 5%
Glikoz, w/v
% e
e v
e %
dö
nü
şü
m
% dönüşüm
% ee
Şekil 12. Serbest hücrelerle glikoz ile kofaktör rejenerasyonu (Cso: 10 mM, t: 24 h, Cx: 0.01 g/ml, T:
35 oC, N= 150 rpm)
Şekil 12 de, serbest hücrelerle biyoindirgeme deneylerinde, tepkime ortamına glikoz eklemeden ve
belli derişimlerde ( % 1 w/v, % 2 w/v, % 5 w/v) glikoz eklendiği durumlarda 10 mM substrat derişimi
için, ulaşılan dönüşümler görülmektedir. Burada, ortama eklenen her glikoz derişiminde dönüşümün
tamamlandığı görülmüştür. Elde edilen enantiyomerik aşırılık değerleri ee> % 99 dur. 10 mM
başlangıç substrat derişimi için % 100 dönüşüm elde edildiği için daha yüksek substrat derişimleri için,
glikoz varlığında kofaktör rejenerasyonu farklı başlangıç substrat derişimlerinde (15 mM, 20 mM,
30mM ve 50mM) gerçekleştirilmiştir (Şekil 13). Tepkime ortamından zamanla alınan örnekler
analizlenmiştir.

0
20
40
60
80
100
120
0 5 10 15 20 25 30
Süre, h
% d
ön
üş
üm
ve
% e
e
15 mM
20 mM
30 mM
50 mM
% ee
Şekil 13. Farklı substrat derişimlerinde serbest hücrelerle glikoz ile kofaktör rejenerasyonu (Cglikoz: %
2 w/v, Cx: 0.01 g/ml, T: 35 oC, N=150 rpm)
Burada glikoz ilave edildiği durumda, kofaktör rejenerasyonunun gerçekleştiği ve dönüşümün,
glikozsuz orama oranla örneğin, 15 ve 20 mM başlangıç substrat derişimleri için % 29 ve dan % 100’ e
arttığı ve % 16’ dan % 96’ ya arttığı görülmektedir (Şekil 14).
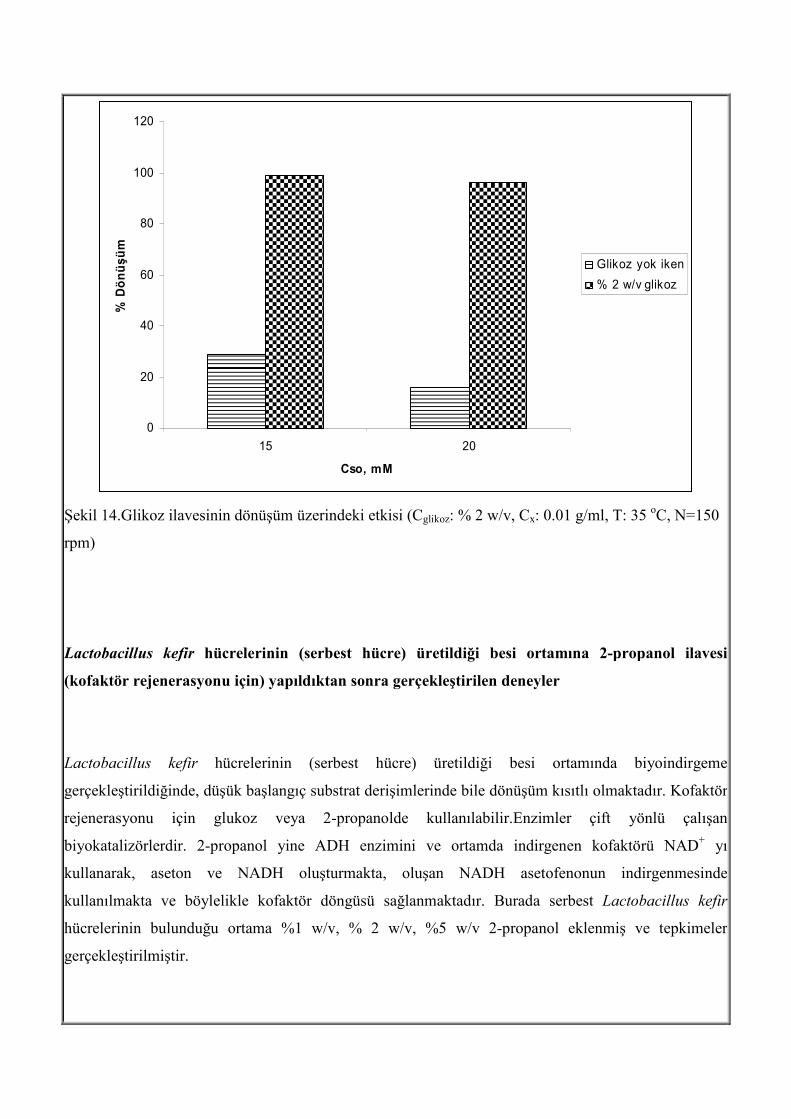
0
20
40
60
80
100
120
15 20
Cso, mM
% D
ön
üş
üm
Glikoz yok iken
% 2 w/v glikoz
Şekil 14.Glikoz ilavesinin dönüşüm üzerindeki etkisi (Cglikoz: % 2 w/v, Cx: 0.01 g/ml, T: 35 oC, N=150
rpm)
Lactobacillus kefir hücrelerinin (serbest hücre) üretildiği besi ortamına 2-propanol ilavesi
(kofaktör rejenerasyonu için) yapıldıktan sonra gerçekleştirilen deneyler
Lactobacillus kefir hücrelerinin (serbest hücre) üretildiği besi ortamında biyoindirgeme
gerçekleştirildiğinde, düşük başlangıç substrat derişimlerinde bile dönüşüm kısıtlı olmaktadır. Kofaktör
rejenerasyonu için glukoz veya 2-propanolde kullanılabilir.Enzimler çift yönlü çalışan
biyokatalizörlerdir. 2-propanol yine ADH enzimini ve ortamda indirgenen kofaktörü NAD+ yı
kullanarak, aseton ve NADH oluşturmakta, oluşan NADH asetofenonun indirgenmesinde
kullanılmakta ve böylelikle kofaktör döngüsü sağlanmaktadır. Burada serbest Lactobacillus kefir
hücrelerinin bulunduğu ortama %1 w/v, % 2 w/v, %5 w/v 2-propanol eklenmiş ve tepkimeler
gerçekleştirilmiştir.
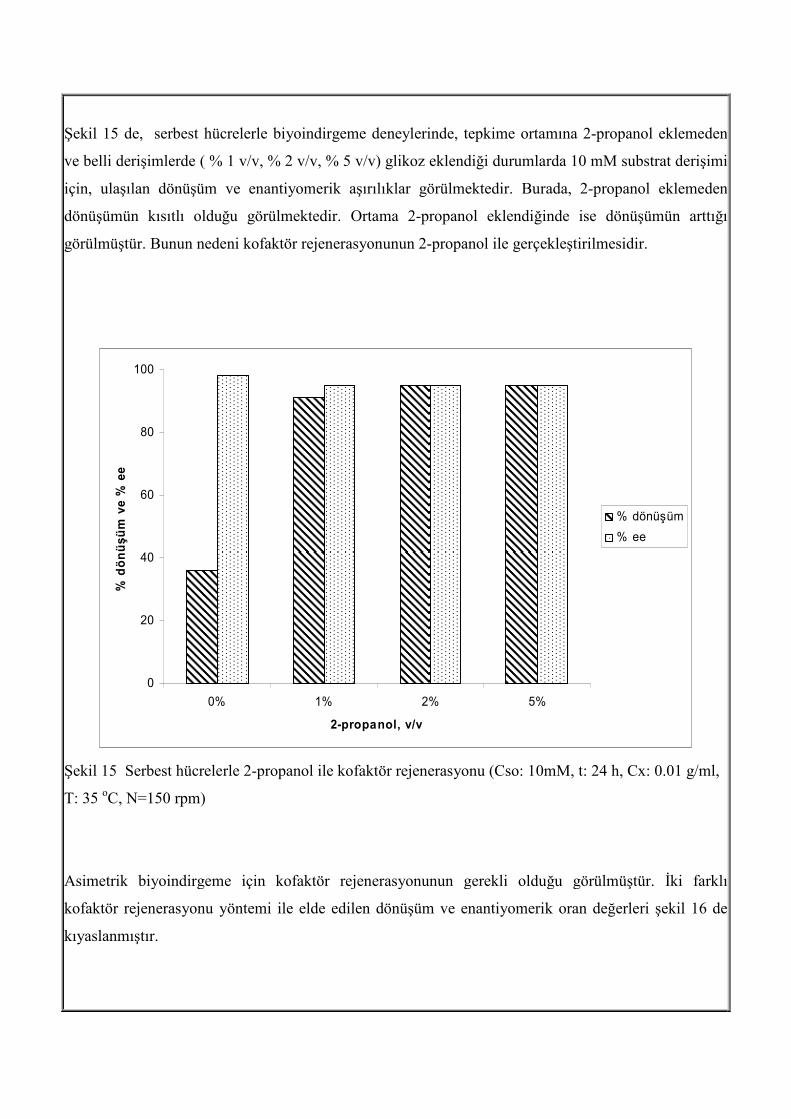
Şekil 15 de, serbest hücrelerle biyoindirgeme deneylerinde, tepkime ortamına 2-propanol eklemeden
ve belli derişimlerde ( % 1 v/v, % 2 v/v, % 5 v/v) glikoz eklendiği durumlarda 10 mM substrat derişimi
için, ulaşılan dönüşüm ve enantiyomerik aşırılıklar görülmektedir. Burada, 2-propanol eklemeden
dönüşümün kısıtlı olduğu görülmektedir. Ortama 2-propanol eklendiğinde ise dönüşümün arttığı
görülmüştür. Bunun nedeni kofaktör rejenerasyonunun 2-propanol ile gerçekleştirilmesidir.
0
20
40
60
80
100
0% 1% 2% 5%
2-propanol, v/v
% d
ön
üş
üm
ve
% e
e
% dönüşüm
% ee
Şekil 15 Serbest hücrelerle 2-propanol ile kofaktör rejenerasyonu (Cso: 10mM, t: 24 h, Cx: 0.01 g/ml,
T: 35 oC, N=150 rpm)
Asimetrik biyoindirgeme için kofaktör rejenerasyonunun gerekli olduğu görülmüştür. İki farklı
kofaktör rejenerasyonu yöntemi ile elde edilen dönüşüm ve enantiyomerik oran değerleri şekil 16 de
kıyaslanmıştır.

0
20
40
60
80
100
1% 2% 5%
% d
ön
üş
üm
Glikoz için % dönüşüm
2-propanol için % dönüşüm
Şekil 16. Serbest hücrelerle glikoz veya 2-propanol ile kofaktör rejenerasyonun dönüşüm açısından
karşılaştırılması(Cso: 10mM, t: 24 h, Cx: 0.01 g/ml, T: 35 oC, N=150 rpm)
2-propanol ile rejenerasyonda ulaşılan dönüşüm değerleri glikoz ile karşılaştırıldığında daha düşüktür.
Glikozun rejenerasyon için dönüşüm açısından daha uygun olduğu görülmektedir. Glikoz,
mikroorganizma üremesi için karbon kaynağıdır ve 2-propanol den daha az toksik olması nedeniyle,
rejenerasyon için daha uygun bir girdidir. Şekil 17 de her iki durumda elde edilen % enantiyomerik
aşırılık değerleri karşılaştırılmıştır.
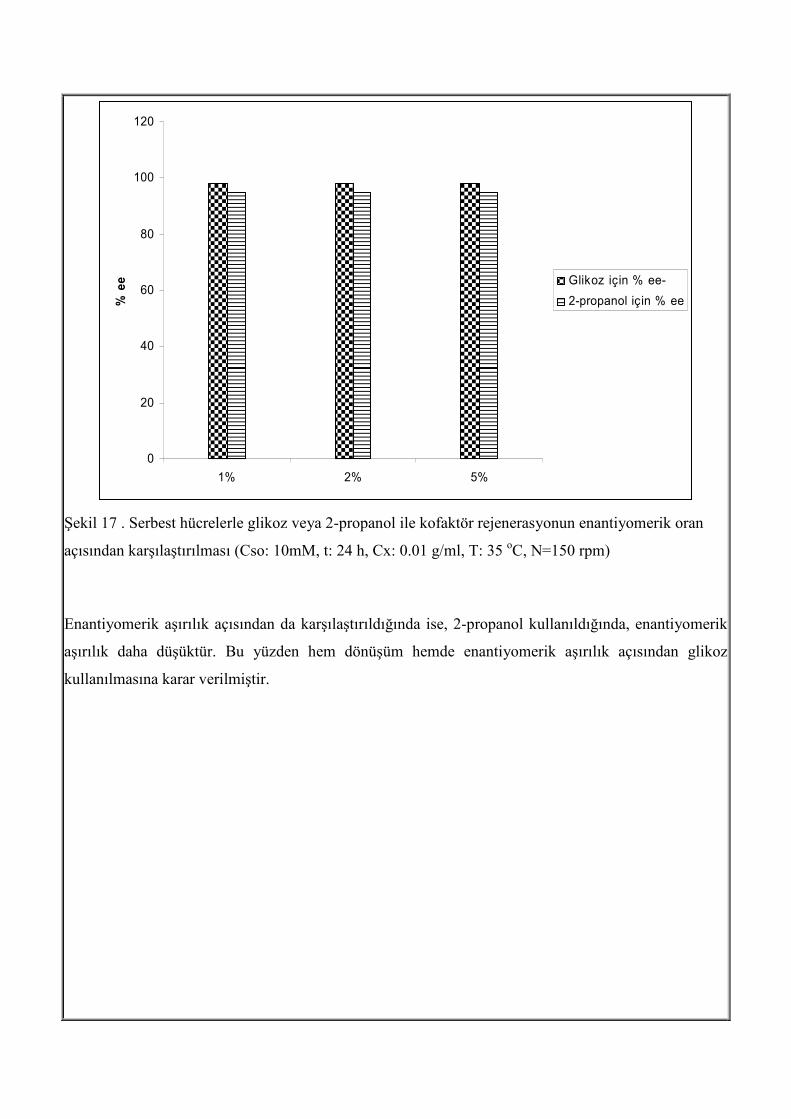
0
20
40
60
80
100
120
1% 2% 5%
% e
e
Glikoz için % ee-
2-propanol için % ee
Şekil 17 . Serbest hücrelerle glikoz veya 2-propanol ile kofaktör rejenerasyonun enantiyomerik oran
açısından karşılaştırılması (Cso: 10mM, t: 24 h, Cx: 0.01 g/ml, T: 35 oC, N=150 rpm)
Enantiyomerik aşırılık açısından da karşılaştırıldığında ise, 2-propanol kullanıldığında, enantiyomerik
aşırılık daha düşüktür. Bu yüzden hem dönüşüm hemde enantiyomerik aşırılık açısından glikoz
kullanılmasına karar verilmiştir.

Tutuklanmış Lactobacillus kefir hücreleriyle kesikli sistemde gerçekleştirilen deneyler
Burada, Lactobacillus kefir hücreleri, kalsiyum aljinat jeline tutuklanmıştır. Kofaktör rejenerasyonu
için % 2 glikoz içeren ve bileşimi Tablo1’ de verilen zengin ortam adını verdiğimiz ortam
kullanılmıştır. Zengin ortam aynı zamanda Lactobacillus kefir in üreme ortamıdır. Tutuklanmış
mikroorganizmayla çalıştığımız için, asimetrik biyoindirgemeye etki eden parametreler incelenmiştir.
Bunlar, pH etkisi, sıcaklık etkisi, Substrat derişiminin etkisi, tampon türü etkisidir.
pH etkisi
Biyoindirgeme deneyleri kalsiyum aljinat tutuklanmış Lactobacillus kefir hücreleriyle, % 2 w/v glikoz
içeren, zengin ortamda gerçekleştirilmiştir. pH etkisi incelenirken, zengin ortamın pH sı 0.1 M H2SO4
ve 0.1 M NaOH ile 6, 6.5, 7 ve 7.5 e ayarlandıktan sonra 2 gram tutuklanmış hücre ile tepkime
gerçekleştirilmiştir. Tüm pH değerlerinde enantiyomerik aşırılık > % 99 iken, dönüşüm pH 7 de en
yüksektir.

0
20
40
60
80
100
5,5 6 6,5 7 7,5 8
pH
% d
ön
üş
üm
ve
% e
e
% dönüşüm
% ee
Şekil 18. Cso=10 mM, Cx: 2 g tutuklanmış hücre, Cx: 0.4 g/ml tutuklanmış hücre, t: 24 h, T=35oC Dönüşüm açısından en uygun pH değeri 7 dir. Enantiymerik aşırılık > % 99 dur ve pH dan etkilenmemektedir. Substrat derişiminin etkisi Tutuklanmış mikroorganizma ile çalışıldığında, substrat derişiminin etkisi yine zengin ortamda incelenmiştir. Burada 5 mM, 10 mM, 20 mM ve 50 mM derişimlerinde 2 gram tutuklanmış hücre ile tepkime gerçekleştirilmiştir. Enantiyomerik aşırılık > % 99 iken, substrat derişimi arttıkça dönüşüm azalmaktadır.
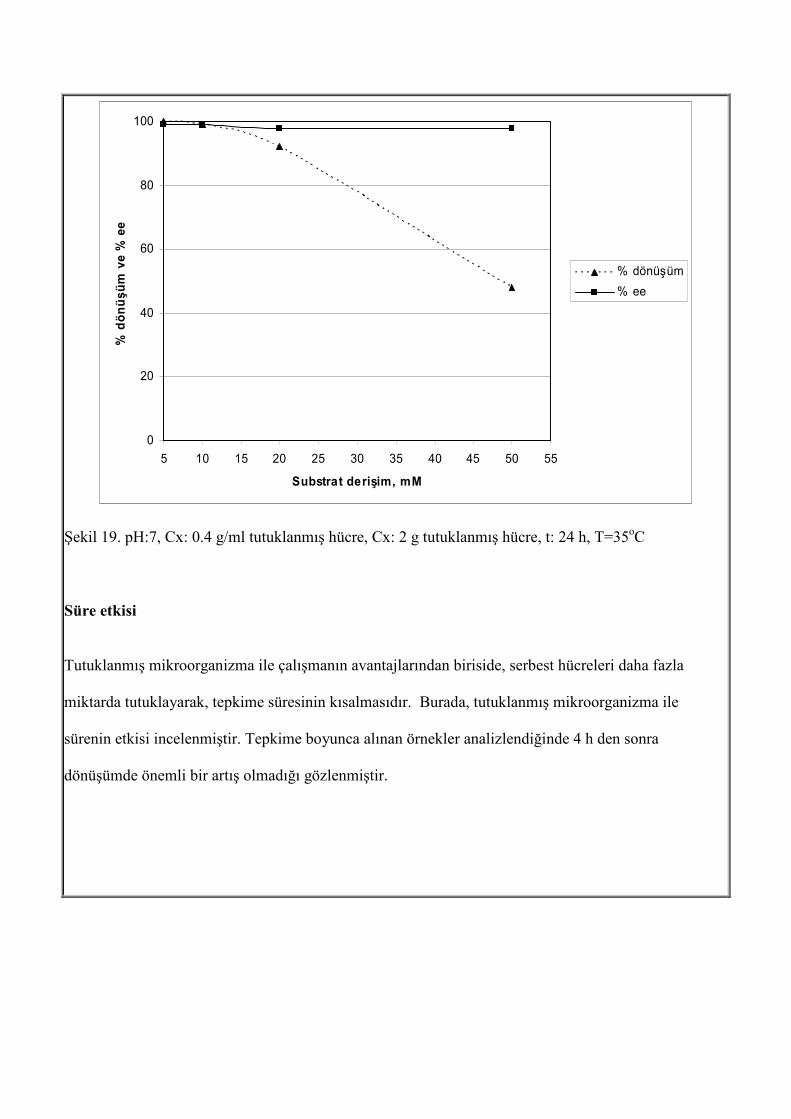
0
20
40
60
80
100
5 10 15 20 25 30 35 40 45 50 55
Substrat derişim, mM
% d
ön
üş
üm
ve
% e
e
% dönüşüm
% ee
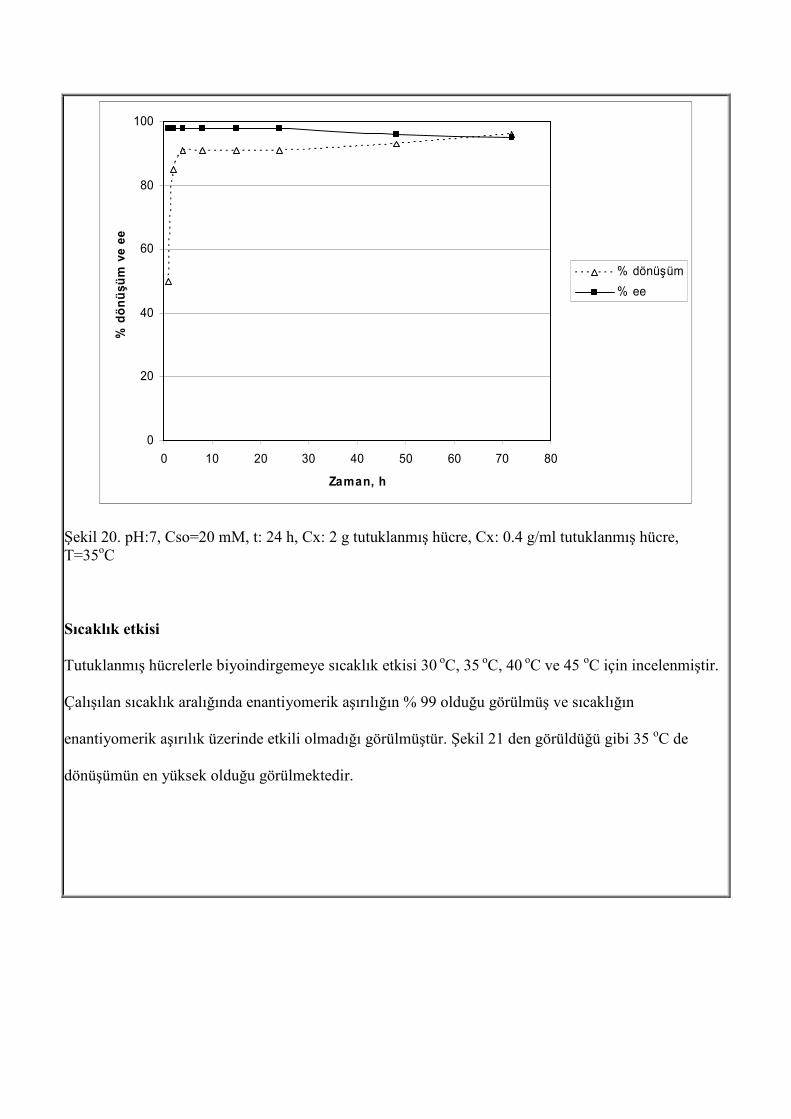
Şekil 19. pH:7, Cx: 0.4 g/ml tutuklanmış hücre, Cx: 2 g tutuklanmış hücre, t: 24 h, T=35oC Süre etkisi Tutuklanmış mikroorganizma ile çalışmanın avantajlarından biriside, serbest hücreleri daha fazla miktarda tutuklayarak, tepkime süresinin kısalmasıdır. Burada, tutuklanmış mikroorganizma ile sürenin etkisi incelenmiştir. Tepkime boyunca alınan örnekler analizlendiğinde 4 h den sonra dönüşümde önemli bir artış olmadığı gözlenmiştir.
0
20
40
60
80
100
0 10 20 30 40 50 60 70 80
Zaman, h
% d
ön
üş
üm
ve
ee
% dönüşüm
% ee
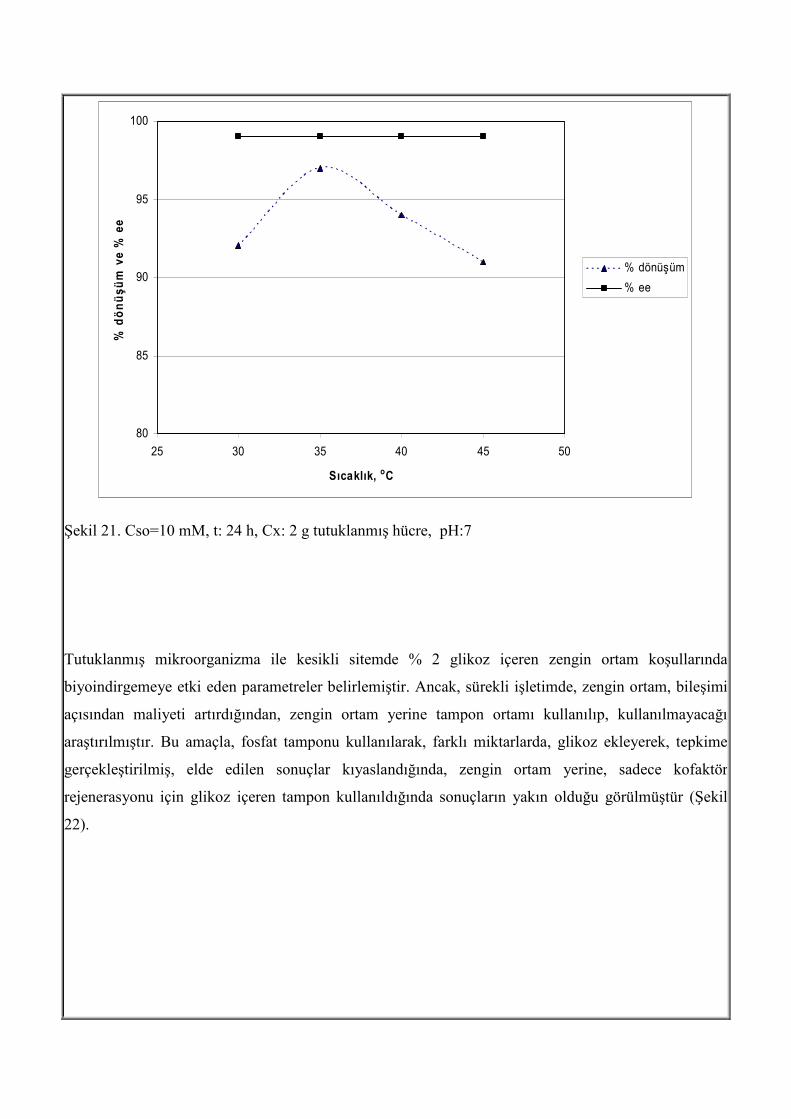
Şekil 20. pH:7, Cso=20 mM, t: 24 h, Cx: 2 g tutuklanmış hücre, Cx: 0.4 g/ml tutuklanmış hücre, T=35oC Sıcaklık etkisi Tutuklanmış hücrelerle biyoindirgemeye sıcaklık etkisi 30 oC, 35 oC, 40 oC ve 45 oC için incelenmiştir. Çalışılan sıcaklık aralığında enantiyomerik aşırılığın % 99 olduğu görülmüş ve sıcaklığın enantiyomerik aşırılık üzerinde etkili olmadığı görülmüştür. Şekil 21 den görüldüğü gibi 35 oC de dönüşümün en yüksek olduğu görülmektedir.
80
85
90
95
100
25 30 35 40 45 50
Sıcaklık, oC
% d
ön
üş
üm
ve
% e
e
% dönüşüm
% ee
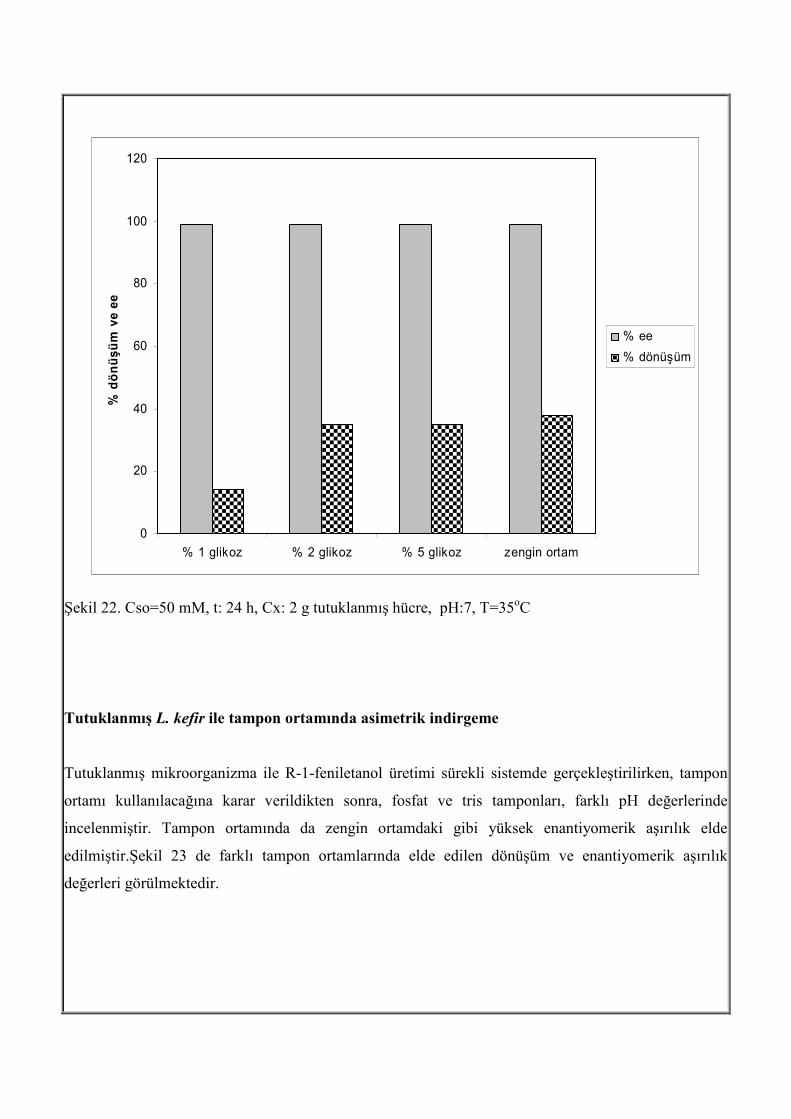
Şekil 21. Cso=10 mM, t: 24 h, Cx: 2 g tutuklanmış hücre, pH:7 Tutuklanmış mikroorganizma ile kesikli sitemde % 2 glikoz içeren zengin ortam koşullarında
biyoindirgemeye etki eden parametreler belirlemiştir. Ancak, sürekli işletimde, zengin ortam, bileşimi
açısından maliyeti artırdığından, zengin ortam yerine tampon ortamı kullanılıp, kullanılmayacağı
araştırılmıştır. Bu amaçla, fosfat tamponu kullanılarak, farklı miktarlarda, glikoz ekleyerek, tepkime
gerçekleştirilmiş, elde edilen sonuçlar kıyaslandığında, zengin ortam yerine, sadece kofaktör
rejenerasyonu için glikoz içeren tampon kullanıldığında sonuçların yakın olduğu görülmüştür (Şekil
22).
0
20
40
60
80
100
120
% 1 glikoz % 2 glikoz % 5 glikoz zengin ortam
% d
ön
üş
üm
ve
ee
% ee
% dönüşüm
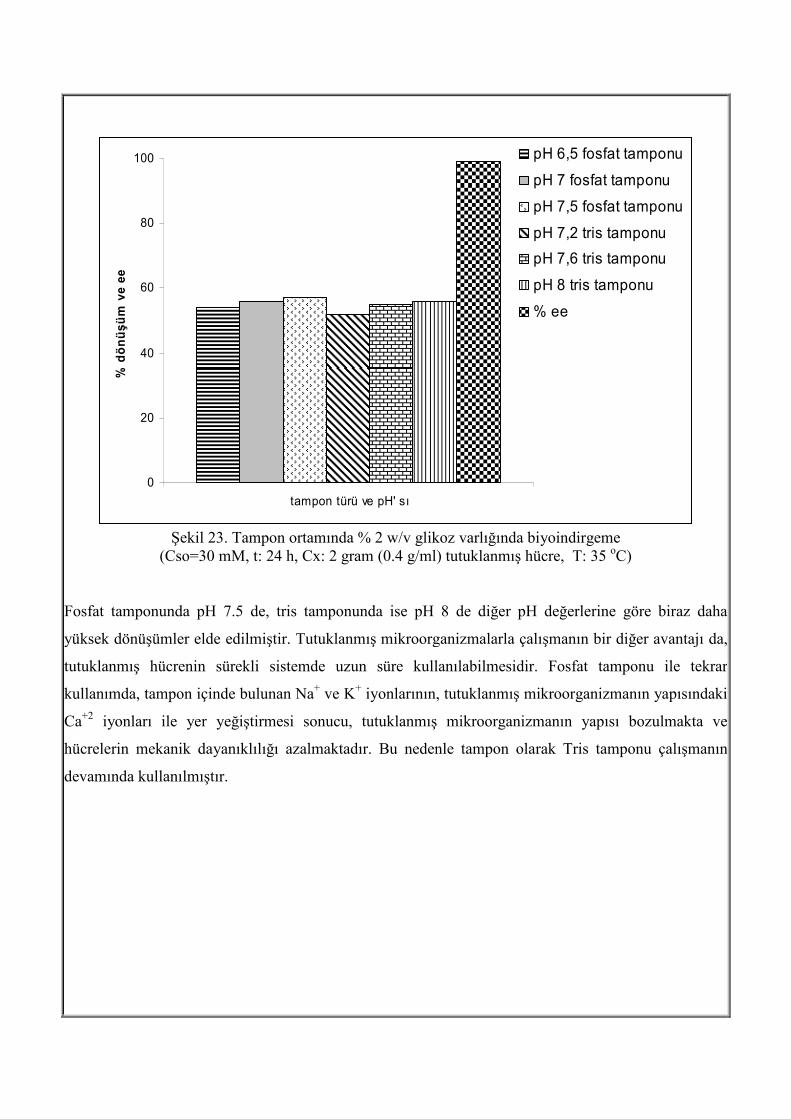
Şekil 22. Cso=50 mM, t: 24 h, Cx: 2 g tutuklanmış hücre, pH:7, T=35oC Tutuklanmış L. kefir ile tampon ortamında asimetrik indirgeme Tutuklanmış mikroorganizma ile R-1-feniletanol üretimi sürekli sistemde gerçekleştirilirken, tampon
ortamı kullanılacağına karar verildikten sonra, fosfat ve tris tamponları, farklı pH değerlerinde
incelenmiştir. Tampon ortamında da zengin ortamdaki gibi yüksek enantiyomerik aşırılık elde
edilmiştir.Şekil 23 de farklı tampon ortamlarında elde edilen dönüşüm ve enantiyomerik aşırılık
değerleri görülmektedir.
0
20
40
60
80
100
tampon türü ve pH' sı
% d
ön
üş
üm
ve
ee
pH 6,5 fosfat tamponu
pH 7 fosfat tamponu
pH 7,5 fosfat tamponu
pH 7,2 tris tamponu
pH 7,6 tris tamponu
pH 8 tris tamponu
% ee
Şekil 23. Tampon ortamında % 2 w/v glikoz varlığında biyoindirgeme
(Cso=30 mM, t: 24 h, Cx: 2 gram (0.4 g/ml) tutuklanmış hücre, T: 35 oC) Fosfat tamponunda pH 7.5 de, tris tamponunda ise pH 8 de diğer pH değerlerine göre biraz daha
yüksek dönüşümler elde edilmiştir. Tutuklanmış mikroorganizmalarla çalışmanın bir diğer avantajı da,
tutuklanmış hücrenin sürekli sistemde uzun süre kullanılabilmesidir. Fosfat tamponu ile tekrar
kullanımda, tampon içinde bulunan Na+ ve K+ iyonlarının, tutuklanmış mikroorganizmanın yapısındaki
Ca+2 iyonları ile yer yeğiştirmesi sonucu, tutuklanmış mikroorganizmanın yapısı bozulmakta ve
hücrelerin mekanik dayanıklılığı azalmaktadır. Bu nedenle tampon olarak Tris tamponu çalışmanın
devamında kullanılmıştır.

Lactobacillus kefir ve Escherichia coli ile kofaktör rejenerasyonu
Biyoindirgeme tepkimesinde Lactobacillus kefir hücreleri alkol dehidrojenaz kaynağı olarak
kullanılmaktadır ve kofaktör olarak NAD(P)H kullanmaktadır. Tepkime sonucu; asetofenon, R-1-
feniletanole indirgenirken, NAD(P)H da NAD(P)+ ya yükseltgenmektedir (Şekil 24). NADP+ nın tekrar
NAD(P)H ya dönüştürülmesi işlemi kofaktör rejenerasyonudur.
Şekil 24. Asetofenonun ADH biyokatalizörlüğünde biyoindirgenmesi
Kofaktör rejenerasyonu için Gebze Yüksek Teknoloji Enstitüsü’ nden temin eldilen, laktat
dehidrojenaz aktivitesine sahip, rekombinant Escherichia coli kullanılmıştır. Laktat dehidrojenaz,
laktatı, pirüvata dönüştürüken, NAD+ yı kofaktör olarak kullanmaktadır. Tepkime sonucu, laktat
pirüvata yükseltgenirken, NAD+ NADH ya indirgenmektedir.
Şekil 24. Laktatın LDH biyokatalizörlüğünde yükseltgenmesi
Şekil 23 ve Şekil 24 deki bu iki bağımsız tepkime, kofaktör döngüsü sağlanmaya çalışılmıştır.Kofaktör
rejenerasyonu, tepkime sistemi Şekil 25. de görülmektedir.

Şekil 25. Kofaktör rejenerasyon sistemi
Laktattan pirüvat oluşumu tepkimesi 340 nm de UV spektrofotometrede takip edilmiştir. Laktat,
pirüvata yükseltgenirken, NAD+, NADH’ ya indirgenmektedir. NAD+, NADH a dönüşürken
absorbansta artış olmaktadır. Absorbanstaki artış; NAD+ dan NADH oluşumunu göstermektedir (Tablo
1).
Tablo 4. Laktatın LDH biyokatalizli yükseltgenmesi
Zaman, dakika Absorbans (340 nm) 0 0.150 1 0.220 2 0,296 3 0,320 4 0,341 5 0,362 20 0,634 53 1,00
Absorbans artışından da görüldüğü gibi recombinant Escherichia coli laktat dehidrojenaz aktivitesine
(LDH) sahiptir ve tepkimeyi katalizlemektedir. Daha sonra her iki enzim ile çalışılmışlardır. Hem
ADH, hemde LDH hücre içi enzimler olduğu için, Lactobacillus kefir ve Escherichia coli, ayrı ayrı
homojenizatörde parçalandıktan sonra, 9500 rpm de +4 oC de 15 dakika santrifüjlenmiş ve supernatant
ile çalışılarak kofaktör rejenerasyonu UV spektrofotometrede incelenmiştir. ADH’ ın katalizlediği
tepkime ile NADH dan NAD+ oluşmakta ve absorbans azalmaktadır. Burada kör ve diğer küvetin her
ikisinede LDH ve laktat ilave edildiği için bu tepkimeden absorbans etkilenmemektedir.

Tablo 5. ADH ve LDH varlığında kofaktör rejenerasyonu
Tablo 5. ADH ve LDH varlığında kofaktör rejenerasyonu (devam)
Zaman, dakika Absorbans (340 nm) 0 0,146 1 0,092 2 0,045 3 -0,022 4 -0,096 5 -0,403 6 -0,550 8 -0,602 10 -0,602
Burada absorbanstaki azalma sadece NADH ın NAD+ ya yükseltgenmesinden kaynaklanmaktadır.Her
iki enzim birlikte çalıştığı için, bu enzimleri kodlayan genlerin, tek bir mikroorganizma içinde
çalışması için klonlama çalışmaları gerçekleştirilmiştir.
Madde Kör Tepkime
NAD+ 10 mM 10 mM
Na-laktat 50 mM 50 mM
LDH- crude extract 100 µl 100 µl
ADH- crude extract 300 µl 300 µl
Asetofenon - - - 10 mM
Toplam çalışma hacmi 2 ml 2 ml

ADH ve LDH genlerinin klonlanması
Lactobacillus kefir mikroorganizmasındaki ADH geni ile Escherichia coli deki LDH geni bu
mikroorganizmalardan izole edilmiş ve Escherichia coli ye klonlanarak kofaktör rejenerasyonu
gerçekleştirilmek amaçlanmıştır. Bu amaçla, Gebze Yüksek Teknoloji Enstitüsü, Çevre Mühendisliği
Bölümü öğretim üyelerinden Doç.Dr. Melek Özkan ile birlikte klonlama çalışmaları
gerçekleştirilmiştir. Öncelikle Lactobacillus kefirden genomik DNA izolasyonu “Lactobacillus kefir ve
Escherichia coli den Genomik DNA İzolasyonu” bölümünde anlatıldığı gibi gerçekleştirilmiştir.
İzolasyon sonucu ADH geninin izole edilip edilmediğini anlamak için agaroz jel elektroforezi
gerçekleştirilmiştir. ADH genin molekül kütlesi (759 bp) (Weckbecker A. 2006 ) ile markerin molekül
kütlesi karşılaştırılmış ve genomik izolasyonun olduğu görülmüştür.
ADH ADH Marker 759 bp �
Şekil 26. ADH geninin isolasyon sonrası agaroz jel eletroforezi görüntüsü

Daha sonra polimeraz zincir reaksiyonu (PCR) ile elde edilen DNA çoğaltılmıştır. Polimeraz zincir reaksiyonu ile “Polimeraz Zincir Reaksiyonu“bölümün de anlatıldığı şekilde gerçekleştirilmiştir.PCR tamamlandıktan sonra tekrar agaroz jel elektroforezi gerçekleştirilmiştir. Sonra istenilen kDa karşılık gelen bantlar kesilerek -20 oC de saklanmıştır. Marker ADH ADH ADH 759 bp �
Şekil 27.Polimeraz zincir reaksiyon ürünü ADH ın agaroz jel elektroforezi
Lactobacillus kefirden DNA izolasyonu ve PCR başarı ile gerçekleştirildikten sonra, Escherichia coliden izolasyonu, “Lactobacillus kefir ve Escherichia coli den Genomik DNA İzolasyonu” bölümünde anlatıldığı gibi gerçekleştirilmiştir. Daha sonra agaroz jel elektroforezi yapılarak, LDH nın izole edilip, edilmediği incelenmiştir. Şekil 28. den görüldüğü gibi, LDH da izole edilmiştir.

Marker LDH ADH
Şekil 28. LDH ve ADH geni agaroz jel elektroforezi
Elde edilen bu genler (LDH ve ADH geni) klonlanmadan önce Sal-I ve Sma-I enzimleri ile ayrı ayrı
kesilmişlerdir. Kesilen bu enzimler, ligasyon yapılarak arka arkaya birleştirilmiş, daha sonrada “Alıcı
(Competent) Hücrenin Hazırlanması” bölümünde anlatıldığı gibi CaCl2 transformasyonu ile
Escherichia coli’nin DH5 α, suşu alıcı hücre olarak kullanılmış ve transformasyon gerçekleştirilmiştir.
Transformasyon sonucu 17 kloni oluşumu görülmüştür. Her bir kloni, “Mikroorganizma ve Üretim
Koşulları” bölümünde anlatıldığı gibi üretilmiş ve ultrasonikasyon ile enzimler hücre dışına çıkarılmış
ve Escherichia coli’nin DH5-α suşu kontrol olarak kullanılarak, LDH ve ADH aktiviteleri, UV
spektrofotometre de ölçülmüştür. Ancak, kontrol ile kıyaslandığında, aktivite, kontrol göre farklılık
göstermemiştir. Escherichia coli’nin DH5-α suşlarında LDH aktivitesi görülmüş fakat ADH aktivitesi
gözlenmemiştir. Sonuçlardan emin olabilmek için, 12 mM asetofenonon, 15 mM NADH ve homojenat
içeren karışım ile 24 saat boyunca, tepkime gerçekleştirilmiş fakat, hiçbirindeR-1-feniletanol oluşumu
gözlenmemiştir. Bu sonuç, genleri içeren Escherichia coli’nin DH5-α suşunda, ADH’ ın aktivitesi
olmadığını göstermiştir. Bunun üzerinde, kofaktör rejenerasyonu yine glikoz ile yapılarak tutuklanmış
L.kefir ile sürekli sistemde üretime geçilmiştir.

Tutuklanmış Lactobacillus kefir hücreleriyle sürekli sistemde gerçekleştirilen deneyler
Sürekli sistemde biyoindirgemeyi etkileyen parametrelerin incelenmesi
Sürekli sistemde asetofenonun mikrobiyal biyoindirgenmesi tepkimesi ile enantiyomerik saflıkta (R)-1-
feniletanol üretimi incelenmiştir. Katalizörlerin etkin ıslanmasından ve dönüşümün yüksek olmasından
dolayı yukarı akışlı dolgulu biyoreaktörde çalışılmıştır. Çalışmada, tepkime karışımı (asetofenon,
glukoz, KCl ve tampon) kolonun altından bir pompa yardımıyla gönderilmiştir. Çalışmalar, 35°C’ de
gerçekleştirilmiştir. Örnekler, kolonun üstünden ayrılan akımdan 1 saat aralıklarla numuneler alınmış
ve analizlenmiştir.
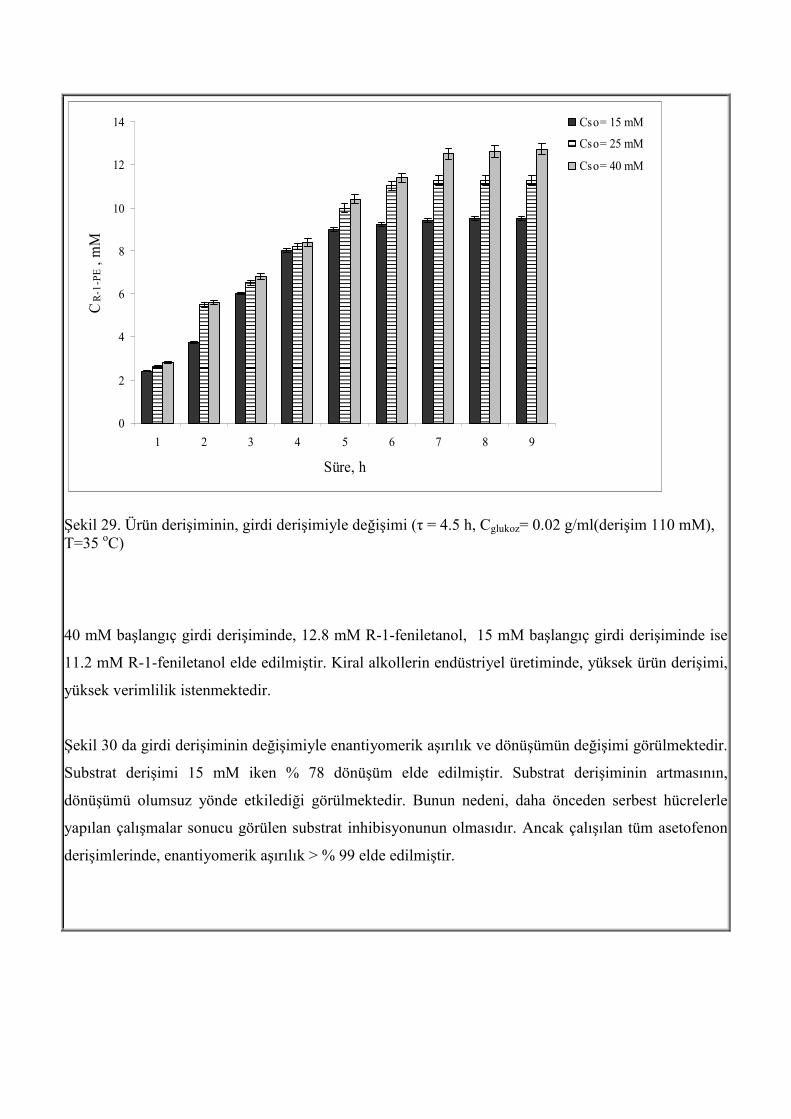
Substrat Derişimi Etkisi Şekil 29 de substrat derişiminin biyoindirgemeye etkisini göstermektedir. Kalma süresi, 4.5 saat iken
çalışılmıştır. Saat başı örnekler alınarak 9 saat boyunca, R-1-feniletanol derişimi, dönüşüm ve
enantiyomerik aşırılığın değişimi izlenmiştir. Asetofenon derişimi 15, 25 ve 40 mM aralığında
incelenmiştir. Substrat derişimi etkisi incelenirken, diğer parametreler, glukoz derişimi (0.02 g/mL),
kalma süresi (4.5 h), sıcaklık (35 oC) sabit tutulmuştur. Şekil 29 de farklı girdi derişimlerinde, üretilen
R-1-feniletanol derişiminin zamanla değişimi görülmektedir. Sistem yatışkın hale ulaştıktan (7 saat)
sonra ürün derişimindeki değişim sabit kalmıştır. Girdi derişimi arttıkça, ürün derişimide artmıştır.
0
2
4
6
8
10
12
14
1 2 3 4 5 6 7 8 9
Süre, h
CR-1
-PE, m
M
Cso= 15 mM
Cso= 25 mM
Cso= 40 mM
Şekil 29. Ürün derişiminin, girdi derişimiyle değişimi (τ = 4.5 h, Cglukoz= 0.02 g/ml(derişim 110 mM), T=35 oC) 40 mM başlangıç girdi derişiminde, 12.8 mM R-1-feniletanol, 15 mM başlangıç girdi derişiminde ise
11.2 mM R-1-feniletanol elde edilmiştir. Kiral alkollerin endüstriyel üretiminde, yüksek ürün derişimi,
yüksek verimlilik istenmektedir.
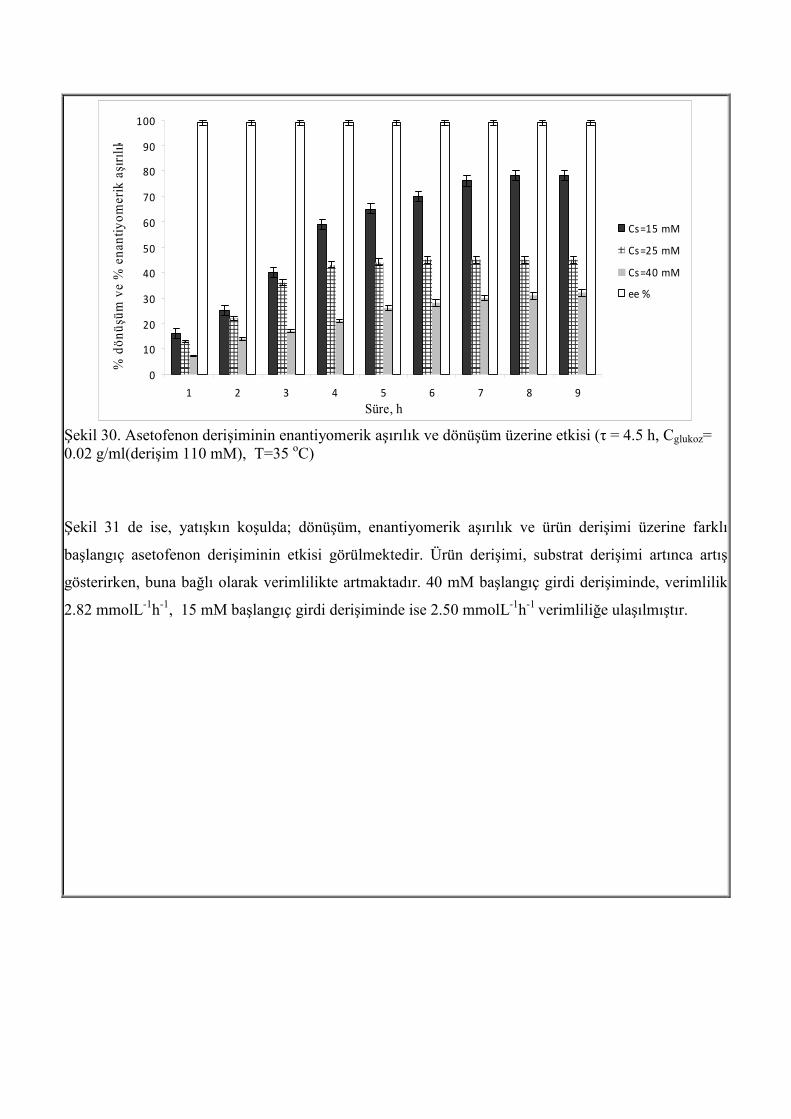
Şekil 30 da girdi derişiminin değişimiyle enantiyomerik aşırılık ve dönüşümün değişimi görülmektedir.
Substrat derişimi 15 mM iken % 78 dönüşüm elde edilmiştir. Substrat derişiminin artmasının,
dönüşümü olumsuz yönde etkilediği görülmektedir. Bunun nedeni, daha önceden serbest hücrelerle
yapılan çalışmalar sonucu görülen substrat inhibisyonunun olmasıdır. Ancak çalışılan tüm asetofenon
derişimlerinde, enantiyomerik aşırılık > % 99 elde edilmiştir.
0
10
20
30
40
50
60
70
80
90
100
1 2 3 4 5 6 7 8 9
Süre, h
% d
önüş
üm v
e %
ena
ntiy
omer
ik a
şırılık
Cs=15 mM
Cs=25 mM
Cs=40 mM
ee %
Şekil 30. Asetofenon derişiminin enantiyomerik aşırılık ve dönüşüm üzerine etkisi (τ = 4.5 h, Cglukoz= 0.02 g/ml(derişim 110 mM), T=35 oC) Şekil 31 de ise, yatışkın koşulda; dönüşüm, enantiyomerik aşırılık ve ürün derişimi üzerine farklı
başlangıç asetofenon derişiminin etkisi görülmektedir. Ürün derişimi, substrat derişimi artınca artış
gösterirken, buna bağlı olarak verimlilikte artmaktadır. 40 mM başlangıç girdi derişiminde, verimlilik
2.82 mmolL-1h-1, 15 mM başlangıç girdi derişiminde ise 2.50 mmolL-1h-1 verimliliğe ulaşılmıştır.

0
20
40
60
80
100
15 25 40
Cso, mM
% d
önüş
üm v
e %
ee
5
7
9
11
13
15
CR-1
FPE, m
M
dönüşüm, %
ee, %
(R)-1-FEderişimi, mM
Şekil 31. Asetofenon derişiminin biyoindirgemeye etkisi (τ = 4.5 h, Cglukoz= 0.02 g/ml(derişim 110 mM), T=35 oC) Glukoz Derişimi Etkisi Karbonil grubu içeren akiral ketonların saf enzim biyokatalizörlüğünde indirgenmesi için stokiyometrik
miktarda NADH veya NADPH kofaktörlerine ihtiyaç vardır. Ancak kofaktörler oldukça pahalıdır. Saf
enzim yerine tüm hücre kullanmak, kofaktör gereksinimini ortadan kaldırır. Tüm hücre kullanarak,
kofaktör rejenerasyonu fermente edilebilir karbonhidratlar sayesinde hücre içinde kendiliğinden
gerçekleştirilebilir. Bu amaçla, basit şekerler, en çok glukoz, kofaktör rejenerasyonu için kullanılır.
Mikroorganizmanın kullanıldığı proseslerde; glukoz, elektron verici rol oynayarak kofaktör
rejenerasyonunu sağlamaktadır. Glukozun tepkime ortamında gereğinden az olması, tepkimenin
yürümesini olumsuz yönde etkiler. Ancak bu moleküllerin ortamda gereğinden fazla olmasıda
inhibisyona neden olmaktadır. Bundan dolayı, glukoz derişimi, biyoindirgeme tepkimelerinde önemli
olmaktadır. Şekil 32 ve şekil 33 da glukoz derişiminin biyoindirgemeye etkisini görülmektedir. Glukoz
derişimi 0.01, 0.02 ve 0.03 g/mL aralığında incelenmiş, diğer parametreler, substrat derişimi (15 mM),
kalma süresi (4.5 h) ve sıcaklık (35 oC) sabit tutulmuştur. Glukozun girdi derişimine oranı, 3.5, 7 ve 11
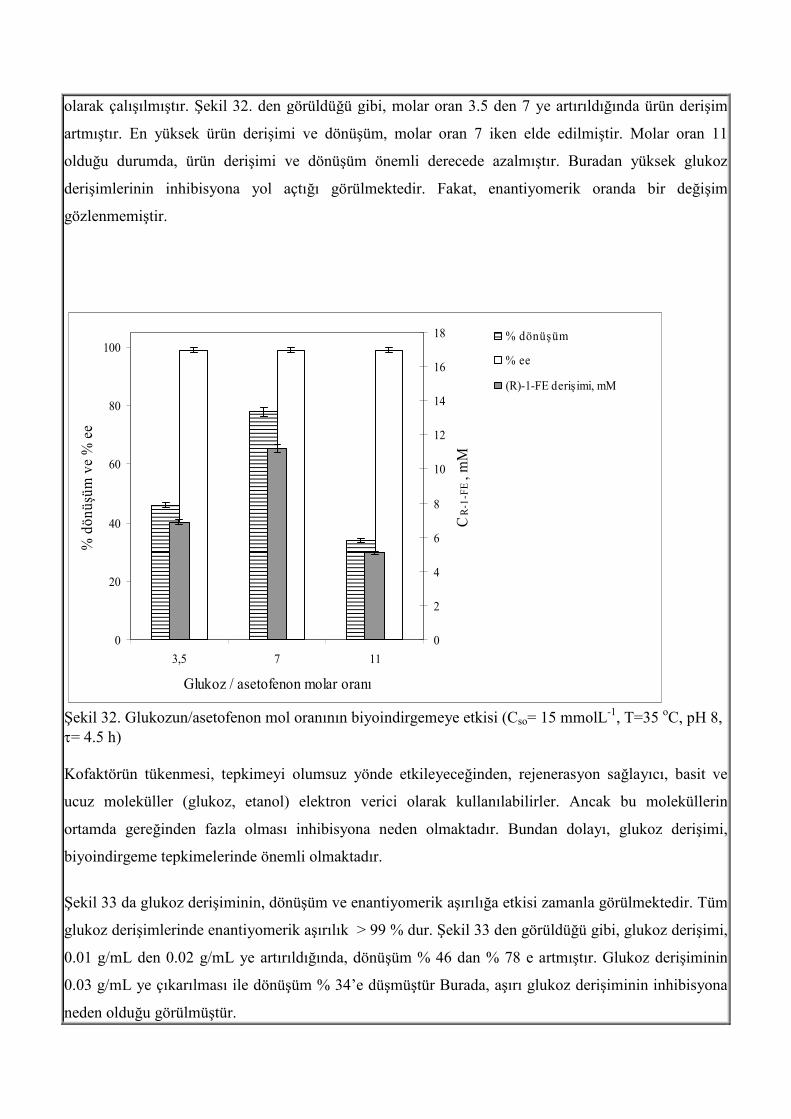
olarak çalışılmıştır. Şekil 32. den görüldüğü gibi, molar oran 3.5 den 7 ye artırıldığında ürün derişim
artmıştır. En yüksek ürün derişimi ve dönüşüm, molar oran 7 iken elde edilmiştir. Molar oran 11
olduğu durumda, ürün derişimi ve dönüşüm önemli derecede azalmıştır. Buradan yüksek glukoz
derişimlerinin inhibisyona yol açtığı görülmektedir. Fakat, enantiyomerik oranda bir değişim
gözlenmemiştir.
0
20
40
60
80
100
3,5 7 11
Glukoz / asetofenon molar oranı
% d
önüş
üm v
e %
ee
0
2
4
6
8
10
12
14
16
18
CR-1
-FE, m
M
% dönüşüm
% ee
(R)-1-FE derişimi, mM
Şekil 32. Glukozun/asetofenon mol oranının biyoindirgemeye etkisi (Cso= 15 mmolL-1, T=35 oC, pH 8, τ= 4.5 h) Kofaktörün tükenmesi, tepkimeyi olumsuz yönde etkileyeceğinden, rejenerasyon sağlayıcı, basit ve
ucuz moleküller (glukoz, etanol) elektron verici olarak kullanılabilirler. Ancak bu moleküllerin
ortamda gereğinden fazla olması inhibisyona neden olmaktadır. Bundan dolayı, glukoz derişimi,
biyoindirgeme tepkimelerinde önemli olmaktadır.
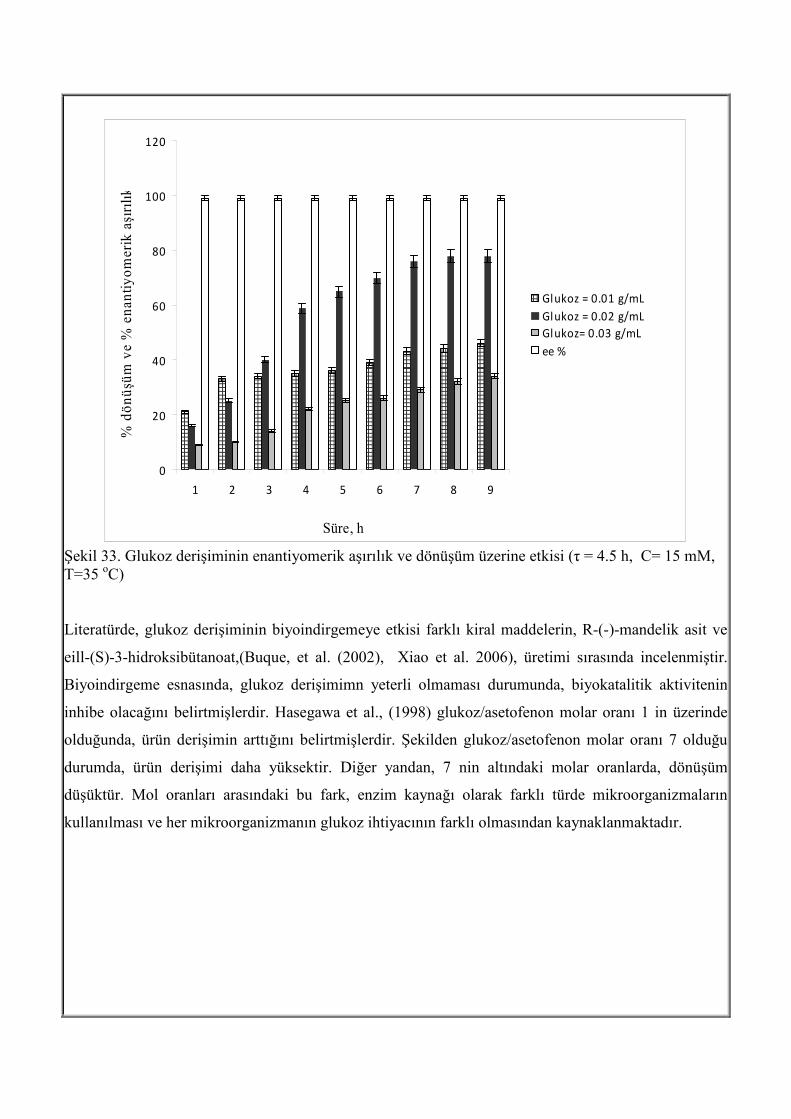
Şekil 33 da glukoz derişiminin, dönüşüm ve enantiyomerik aşırılığa etkisi zamanla görülmektedir. Tüm
glukoz derişimlerinde enantiyomerik aşırılık > 99 % dur. Şekil 33 den görüldüğü gibi, glukoz derişimi,
0.01 g/mL den 0.02 g/mL ye artırıldığında, dönüşüm % 46 dan % 78 e artmıştır. Glukoz derişiminin
0.03 g/mL ye çıkarılması ile dönüşüm % 34’e düşmüştür Burada, aşırı glukoz derişiminin inhibisyona
neden olduğu görülmüştür.
0
20
40
60
80
100
120
1 2 3 4 5 6 7 8 9
Süre, h
% d
önüş
üm v
e %
ena
ntiy
omer
ik aşırılık
Glukoz = 0.01 g/mL
Glukoz = 0.02 g/mL
Glukoz= 0.03 g/mL
ee %
Şekil 33. Glukoz derişiminin enantiyomerik aşırılık ve dönüşüm üzerine etkisi (τ = 4.5 h, C= 15 mM, T=35 oC) Literatürde, glukoz derişiminin biyoindirgemeye etkisi farklı kiral maddelerin, R-(-)-mandelik asit ve
eill-(S)-3-hidroksibütanoat,(Buque, et al. (2002), Xiao et al. 2006), üretimi sırasında incelenmiştir.
Biyoindirgeme esnasında, glukoz derişimimn yeterli olmaması durumunda, biyokatalitik aktivitenin
inhibe olacağını belirtmişlerdir. Hasegawa et al., (1998) glukoz/asetofenon molar oranı 1 in üzerinde
olduğunda, ürün derişimin arttığını belirtmişlerdir. Şekilden glukoz/asetofenon molar oranı 7 olduğu
durumda, ürün derişimi daha yüksektir. Diğer yandan, 7 nin altındaki molar oranlarda, dönüşüm
düşüktür. Mol oranları arasındaki bu fark, enzim kaynağı olarak farklı türde mikroorganizmaların
kullanılması ve her mikroorganizmanın glukoz ihtiyacının farklı olmasından kaynaklanmaktadır.

Kolonda kalma süresinin etkisi
Kalma süresinin etkisini incelemek amacıyla, öncelikle kolon gözenekliliği tespit edilmiştir ve 0.45
olarak bulunmuştur. Farklı akış hızlarında çalışılmış, kolon gözenekliliğinden yararlanarak, kalma
süreleri belirlenmiştir. İncelenen kalma süreleri sırasıyla, 2.25 saat, 3 saat, 4.5 saat ve 9 saattir.
Tepkime, Şekil 5 de şekli verilen biyoreaktörde gerçekleştirilmiştir. Şekiller (34, 35, 36 ve 37)’ den
görüldüğü gibi farklı kalma sürelerinde çalışılırken, sürekli sistemde saat başı örnek alınmış ve yatışkın
koşula gelme zamanı tespit edilmeye çalışılmıştır. Kalma süresi etkisi incelenirken, diğer parametreler,
başlangıç substrat derişimi (15 mM), glukoz derişimi (0.02 g/mL), sıcaklık (35 oC) sabit tutulmuştur.
Kalma süresi arttıkça yatışkın koşula gelme süresinin de arttığı gözlenmiştir. Düşük akış hızlarında 7
ila 8 saat arasında yatışkın koşula ulaşılmıştır. Yüksek akış hızlarında ise daha kısa sürede (4 h)
yatışkın koşula ulaşılmıştır. Her farklı kalma süresinde elde edilen % dönüşüm ve % enantiyomerik
aşırılığın zamanla değişimi “Teknik ve Bilimsel Ayrıntılar” bölümünde verilmiştir. İncelenen kalma
sürelerinde % dönüşüm ve % enantiyomerik aşırılığın değişimi, yatışkın hale ulaştıktan sonra Şekil 38
de görülmektedir. Kalma süresinin 9 saat olduğu durumda % 86 dönüşüm ve % 99 enantiyomerik
aşırılığa ulaşılmıştır. Kalma süresinin 4.5 saat olduğu durumda ise % 79 dönüşüm ve % 99
enantiyomerik aşırılığa ulaşılmıştır. Çalışılan en düşük kalma süresinde (2.25 saat) yani akış hızının
çok yüksek olduğu durumlarda dönüşüm % 55’ e düşmüştür. Dönüşümdeki bu düşüş enzim-substrat
etkileşim süresinin çok kısa olmasından yani dönüşüm için yeterli zaman olmamasından
kaynaklanmaktadır. Enantiyomerik aşırılık kalma süresinden etkilenmemektedir. Çalışılan tüm kalma
sürelerinde, % 99 enantiyomerik aşırılık elde edilmiştir.

0
10
20
30
40
50
60
70
80
90
100
1 2 3 4 5 6 7 8 9 10
Süre , saat
% d
ön
üşü
m v
e %
ee
%dönüşüm
% ee
Şekil 34. Kalma süresinin enantiyomerik aşırılık ve dönüşüm üzerine etkisi (τ = 9h, Cglukoz= 0.02 g/ml(derişim 110 mM), Cso=15 mM, T=35 oC)
0
10
20
30
40
50
60
70
80
90
100
1 2 3 4 5 6 7 8 9 10
Süre, saat
% d
ön
üşü
m
ve
%
ee
%dönüşüm
% ee
Şekil 35. Kalma süresinin enantiyomerik aşırılık ve dönüşüm üzerine etkisi (τ = 4.5 h, Cglukoz= 0.02 g/ml(derişim 110 mM), Cso=15 mM, T=35 oC)
0
10
20
30
40
50
60
70
80
90
100
1 2 3 4 5 6 7 8 9 10
Süre, saat
% d
ön
üşü
m v
e %
ee
% dönüşüm
% ee
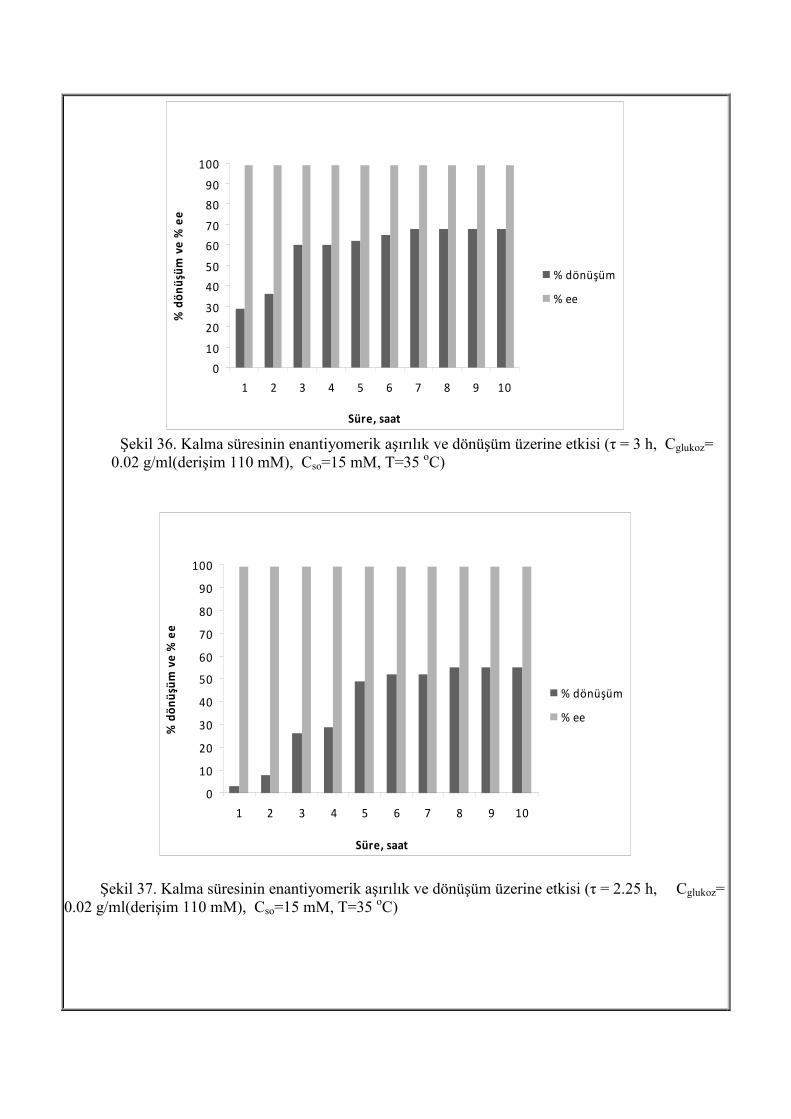
Şekil 36. Kalma süresinin enantiyomerik aşırılık ve dönüşüm üzerine etkisi (τ = 3 h, Cglukoz=
0.02 g/ml(derişim 110 mM), Cso=15 mM, T=35 oC)
0
10
20
30
40
50
60
70
80
90
100
1 2 3 4 5 6 7 8 9 10
Süre, saat
% d
ön
üşü
m v
e %
ee
% dönüşüm
% ee
Şekil 37. Kalma süresinin enantiyomerik aşırılık ve dönüşüm üzerine etkisi (τ = 2.25 h, Cglukoz= 0.02 g/ml(derişim 110 mM), Cso=15 mM, T=35 oC)

Şekil 38, farklı kalma sürelerinde yatışkın koşulda, ulaşılan, dönüşüm ve enantiyomerik aşırılık değerlerini göstermektedir. Buradan görüldüğü gibi, kalma süresi 4.5 saatten 9 saate artırıldığında, % dönüşümdeki çok önemli bir atış olmadığından 9 saatten fazla kalma sürelerinde çalışılmamıştır.
0
20
40
60
80
100
120
2,25 3 4,5 9
Kalma süres i, h
% d
önüş
üm v
e %
ee
c %
ee %
Şekil 38.Yatışkın halde farklı kalma sürelerinde % dönüşüm ve % enantiyomerik aşırılık değerleri (Cso=15 mM, Cglukoz= 0.02 g/ml(derişim 110 mM), T=35 oC) Şekil 39, kalma süresinin verimlilik ve R-1-Feniletanol derişmi üzerine etkisini göstermektedir. Kalma
süresi arttıkça, ürün derişmi artmış, ancak verimlilik azalmıştır. Yüksek verimlilik ve yüksek ürün
derişimi elde edebilmek için 3.75 h kalma süresinin ekonomik olduğu görülmüştür.

1
1,5
2
2,5
3
3,5
4
2,25 3 4,5 9
Kalma süresi, h
Ver
imlil
ik, m
mol
/L/h
1
3
5
7
9
11
13
15
CR-1
-PE, m
M
verimlilik, mmol/L/h
R-1-FE derişimi, mM
Şekil 39. Kalma süresinin verimlilik ve (R)-1-FE derişimine etkisi (Cso=15 mM, Cglukoz= 0.02 g/mL, T=35 oC, pH 8)
Sürekli sistemler, kesikli sistemlere göre daha uzun işletme ömrü olmaları nedeniyle tercih
edilmektedirler. Burada dolgulu kolon reaktörün, aktivitesini ne kadar süre koruduğu incelenmiştir.
Bunu için hücreler, taze tutuklandıktan sonra, biyoindirgeme gerçekleştirilmiş, daha sonra kolon,
günler boyunca, işletilmiştir (Şekil 40 ).

0
10
20
30
40
50
60
70
80
90
100
110
1 5 9 13 17 21 25
gün
% D
ön
üş
üm
Şekil 40. Dolgulu kolon biyoreaktörün aktivitesinin zamanla değişimi (τ = 4.5 h, Cso=20 mM, Cglukoz= 0.02 g/ml(derişim 110 mM), T=35 oC, pH 8) Sürekli sistemler, kesikli sistemlere göre sürekli işletilmeleri nedeniyle tercih edilmektedirler. Şekilden
görüldüğü gibi, dolgulu kolon reaktörün ne kadar süre işletilebileceği incelenmiştir. Çalışmada,
Tutuklanmış hücreler, 5 gün işletim süresince aktivetisinin % 90’ ını, 12 gün işletim süresince
aktivetisinin % 50’ isini korumaktadır.

IV. Sonuç ve Öneriler
Bu çalışmada üretim yöntemlerinden, asimetrik indirgeme kullanılmıştır ancak literatürde sürekli
işletimde asimetrik biyoindirgeme üzerine çok az çalışma bulunmaktadır. Sürekli sistemler, kesikli
sistemlere göre sürekli işletilmeleri nedeniyle tercih edilmektedirler. Dolayısıyla, bu çalışmada
literatürdeki bu boşluğu doldurmaktadır. Asetofenonun, tutuklanmış mikroorganizma ile asimetrik
biyoindirgenmesi sonucu > 99 % enantiyomerik saflıkta R-1-fenil etanol üretimi sürekli sistemde
başarı ile gerçekleştirilmiştir. Yüksek verimlilik ve yüksek ürün derişimi elde edebilmek için 3.75 h
kalma süresinin ekonomik olduğu bulunmuştur. Tutuklanmış hücreler, 5 gün işletim süresince
aktivetisinin % 90’ ını, 12 gün işletim süresince aktivetisinin % 50’ isini korumaktadırlar. 40 mM
başlangıç girdi derişiminde, 12.8 mM R-1-feniletanol üretilmiş, verimlilik 2.82 mmolL-1h-1dur. 15
mM başlangıç girdi derişiminde ise 11.2 mM R-1-feniletanol (% 79 dönüşüm) elde edilmiş, 2.50
mmolL-1h-1 verimliliğe ulaşılmıştır. Laboratuar ölçekte, gerçekleştirilen bu çalışamanın sonuçları
kullanılarak, endüstriyel ölçekte üretim yapmak mümkün olacaktır.

V. Kaynaklar
Anonim. 2008. Web sitesi: http://tr.wikipedia.org/wiki/Enzim. Erişim Tarihi: 30/04/2008. Atkins, R.C., Carey, F.A. 2003.Organik Kimya. Bilim Yayıncılık, 200, Ankara. Bauer, C., Boy, M., Faber, K., Felfer, U., Voss, H. 2001. Activation of Mandelate Racemase via Immobilization in Lyotropic Liquid Crystals for Biocatalysis in Organic Solvents: Application and Modeling. Journal of Molecular Catalysis B: Enzymatic, 16; 91-100. Blanch H.W and Clark D.S. 1996. Biochemical engineering. New York: Marcel Dekker Inc. Braeutigam S., Dennewald, D. Schürmann, M., Lutje-Spelberg, J., Pitner W-R, Weuster-Botz D. 2009. Whole-cell biocatalysis: Evaluation of new hydrophobic ionic liquids for efficient asymmetric reduction of prochiral ketones, Enzyme and Microbial Technology 45:310–316. Buque, E.M., Chin-Joe, I., Straathof, A.J.J., Jongejan, J.A., Heijnen, J.J., “Immobilization affects the rate and enantioselectivity of 3-oxo ester reduction by baker’s yeast”, Enzym. Microb. Technol., 31,656-664 (2002). Carnell, A. J., Cardus, G. J., Trauthwein, H., Riermeir, T. 2004. Microbial Deracemisation of N-(1-hydroxy-1-phenylethyl)benzamide. Tetrahedron: Asymmetry, 15; 239-243. Chadha, A., Baskar, B. 2002. Biocatalytic Deracemisation of α-hydroxy Esters:High Yield Preparation of (S)-ethyl 2-hydroxy-4-phenylbutanoate from the Racemate. Tetrahedron: Asymmetry, 13; 1461-1464. Chadha, A., Padhi, S. K., Pandian, N. G. 2004. Microbial Deracemisation of Aromatic β-Hydroxy Acid Esters. Journal of Molecular Catalysis B: Enzymatic, 29; 25-29. Chadha, A., Padhi, S. K. 2005. Deracemisation of Aromatic β-hydroxy Esters Using Immobilised Whole Cells of Candila Parapsilosis ATCC 7330 and Determination of Absolute Configuration by H NMR. Tetrahedron: Asymmetry, 16; 2790-2798. Choi, W.J., Lee, K.Y., Kang, S.H., Lee, S.B. 2007. Biocatalytic Enantioconvergent Separation of Racemic Mandelic Acid. Separation and Purification Technology, 53; 178-182. Çelebi N., Yıldız, N., Demir, A.S., Çalımlı, A. 2006. Enzymatic Synthesis of Benzoin in Supercritical Carbon dioxide. Journal of Supercritical Fluids, 41;386-390. Demir, A.S., Hamamci, H., Sesenoglu, Ö., Neslihanoglu, R., Asikoglu, B., Capanoglu D. 2002.Fungal deracemization of benzoin, Tetrahedron Letters, 43:6447–6449. Dilsiz, Nihat., Moleküler Biyoloji, Palme Yayıncılık Ankara 2004. Gröger, H., Hummel, W., Rollmann, C., Chamouleau, F., Hüsken, H., Werner, H., Wunderlich, C., Abokitse, K., Drauz K.,Buchholz S. 2004. Preparative asymmetric reduction of ketones in a biphasicmedium with an (S)-alcohol dehydrogenase under in situ-cofactor-recycling with a formate dehydrogenase, Tetrahedron 60:633-640. Faber, K., Strauss, U. T., Felfer, U. 1999a. Biocatalytic Transformation of Racemates into Chiral Building Blocks in 100% Chemical Yield and 100% Enantiomeric Excess. Tetrahedron: Asymmetry, 10; 107-117. Faber, K., Strauss, U.T. 1999b. Deracemization of (±)-Mandelic Acid Using a Lipase-Mandelate Racemase Two-Enzyme System. Tetrahedron: Asymmetry, 10; 4079-4081. Faber, K. 2000. Biotransformations in Organic Chemistry. 4th Edition, Springer-Verlag Berlin Heidelberg New York. Faber, K., Schnell, B. L., Glueck, S. M., Kroutil, W. 2006. Enantio-Complementary Deracemization of (±)-2-hydroxy-4-phenylbutanoic Acid and (±)-3-Phenyllactic Acid Using Lipase-Catalyzed Kinetic Resolution Combined with Biocatalytic Racemization. Tetrahedron, 62; 2912-2916. Ghanem, A. and Aboul-Enein, H.Y. 2004. Lipase-Mediated Chiral Resolution of Racemates in Organic Solvents.Tetrahedron: Asymmetry Report Number 78, 15, 3331-3351. Goswami A., Mirfakhrae, K. D., Patel R.N. 1999. Deracemization of Racemic 1,2-diol by Biocatalytic

Stereoinversion. Tetrahedron: Asymmetry ,10; 4239-4244. Gröger, H., Chamouleau, F., Orologas, N., Rollmann, C., Drauz, K., Hummel, W., Weckbecker, A., ve Oliver May, O., Enantioselective Reduction of Ketones with “Designer Cells” at High Substrate Concentrations: Highly Efficient Access to Functionalized Optically Active Alcohols, Angew. Chem. Int. Ed. 2006, 45, 5677 –5681 http://reference.md/files/D010/mD010626.html Hummel, W., Abokitse, K., Drauz, K., Rollmann, C. and Gröger, H. 2003. Towards a Large-Scale Asymmetric Reduction Process with Isolated Enzymes: Expression of an (S)-Alcohol Dehydrogenase in E. coli and Studies on the Synthetic Potential of this Biocatalyst. Adv. Synth. Catal. 345:153-159. Inoue, K., Makino Y. and Itoh, N. 2005. Purification and Characterization of a Novel Alcohol Dehydrogenase from Leifsonia sp. Strain S749: a Promising Biocatalyst for an Asymmetric Hydrogen Transfer Bioreduction. Applied and Environmental Microbiology, 71: 3633–3641. Kosjek, B., Stampfer, W., Pogorevc, M., Goessler, W., Faber, K. and Kroutil, W. 2004. Purification and Characterization of a Chemotolerant Alcohol Dehydrogenase Applicable to Coupled Redox Reactions” Biotechnology and Bioengineering, 86:55-62. Kragl, U., Kaftzik, N., Kroutil, W., Faber, K. 2004. Mandelate Racemase Activity in Ionic Liquids: Scopes and Limitations. Journal of Molecular Catalysis A: Chemical, 214; 107-112. Kroutil, W., Schnell, B., Faber, K. 2003. Enzymatic Racemisation and its Application to Synthetic Biotransformations. Adv. Synth. Catal., 345; 653-666. Kroutil, W., Voss, C. V., Cruber, C. C. 2007. A Biocatalytic One-Pot Oxidation/Reduction Sequence for the Deracemisation of a sec-alcohol. Tetrahedron: Asymmetry, 18; 276-281. Li, G-Y., Huang, K-L., Jiang, Y-R., Ding, P., Production of (R)-mandelic acid by immobilized cells ofSaccharomyces cerevisiae on chitosan carrier,Process Biochemistry, 42 (2007) 1465–1469 Majewska, P., Kafarski, P., Lejczak, B. 2006. Simple and Effective Method for the Deracemization of ethyl 1-hydroxyphosphinate Using Biocatalysts with Lipolytic Activity. Tetrahedron: Asymmetry, 17; 2870-2875. Olceroglu, A.H., Calık, P., Yılmaz, L. 2006. Chiral Separation of Racemic Benzoin via Enzyme Enhanced Ultrafiltration. Desalination, 200;464-465. Patel, R.N., Goswami, A., Chu, L., Donovan, M.J., Nanduri, V., Goldberg, S., Johnston, R., Siva, P.J., Nielsen, B., Fan, J., He, W., Shi, Z., Wang, K.Y., Eiring, R., Cazzulino, D., Singh A. and Mueller R., Enantioselective microbial reduction of substituted Acetophenones, Tetrahedron: Asymmetry 15, 2004, 1247–1258. Shuler M.L. and Kargi F. 1992. Bioprocess Engineering Basic Concepts. Prentice-Hall, Inc. New Jersey. Suye, S., Kamiya, K., Kawamoto, T. and Tanaka, A. 2002. Efficient Repeated Use of Alcohol Dehydrogenase with NAD+ Regeneration in an Aqueous–organic Two-phase System. Biocatalysis and Biotransformation, 20 (1):23–28. Xiao M., Huang Y., Meng C. And Guo Y. 2006. Kinetics of Asymmetric Reduction of Phenylglyoxylic Acid to R-( -)-Mandelic Acid by Saccharomyces Cerevisiae FDllb, Chinese J. Chem. Eng., 14(1) 73-80.Inoue K., Makino Y., and Itoh N., 2005. Purification and Characterization of a Novel Alcohol Dehydrogenase from Leifsonia sp. Strain S749: a Promising Biocatalyst for an Asymmetric Hydrogen Transfer Bioreduction. Applied and Environmental Microbiology, 71: 3633–3641.
Kurbanoglu, E.B., Zilbeyaz, K., Kurbanoglu, N.I. ve Taskin M. 2007.,”Highly enantioselective reduction of acetophenone by locally isolated Alternaria alternata using ram horn peptone”,Tetrahedron: Asymmetry 18 (2007) 1529–1532. Kurbanoglu, E.B., Zilbeyaz, K., Kurbanoglu, N.I. ve Kilic H. 2007.,” Enantioselective reduction of substituted acetophenones by Aspergillus niger”, Tetrahedron: Asymmetry 18 (2007) 1159–1162. Mertens R., Greiner L., Van den Ban E. C.D., Haaker H. B.C.M. and Liese A. 2003. Practical applications of hydrogenase I from Pyrococcus furiosus for NADPH generation and regenaration.

Journal of Moleculer Catalysis B: Enzymatic 27-25: 39-52. Stampfer, W., Kosjek, B., Faber, K., Kroutil, W. 2003. Biocatalytic Asymmetric Hydrogen Transfer Employing Rhodococcus ruber DSM 44541. J.Org. Chem. 68:402-406. Suan CL, Sarmidi MR. 2004. Immobilised lipase-catalysed resolution of (R,S)-1-phenylethanol in recirculated packed bed reactor. Journal of Molecular Catalysis B: Enzymatic 28:111–119. Telefoncu, A., Zihnioğlu, F., Kılınç, A. ve Salnikow, J. 2000. Biyokimyada Temel ve Modern Teknikler, Biyokimya Lisansüstü Yaz Okulu, Bornova, İzmir Temino D.M., Hartmeier W., Ansorge-Schumacher M.B., 2005. Entrapment of the alcohol dehydrogenase from Lactobacillus kefir in polyvinyl alcohol for the synthesis of chiral hydrophobic alcohols in organic solvents, Enzyme Microbial Technology 36(1): 3-9 Tüzün, C. 1994. Stereokimya. Ankara
Tüzün, C. 2005. Biyokimya. Palme Yayıncılık, Ankara.
Villela Filho M, Stillger T, Müller M, Liese A, Wandrey C. 2003. Is the polarity a convenient criterion to guide the choice of solvents for biphasic enzymatic reactions? Angew Chem Int Ed 42:2993–2996. Weckbecker A. ve Hummel W. 2006. Cloning, expression, and characterization of an (R)-specific alcoholdehydrogenase from Lactobacillus kefir, Biocatalysis and Biotransformation; 24(5): 380-389. Wang, W. Y., Hetter, D.J. 1982. Cell Immobilization in k-carragenenan with Tricalcium Phosphate, Biotechnology and Bioengineering 24: 1827-1838. Ward, O.P., Singh, A. 2000. Enzymatic Asymmetric Synthesis by Decarboxylases. Current Opinion in Biotechnology, 11;520-526.
Zhu, S., Wu, Y., Yu, Z. 2006. An extended model for biochemical kinetic resolution of enantiomers. Process Biochemistry, Elsevier Ltd.

VII. Ekler
a) Mali Bilanço ve Açıklamaları
SIRA NO
EKONOMİK SINIFLANDIRMA
MİKTARI ÖLÇÜ BİRİMİ Durum
Makine Teçhizat Adı (03.7+06.1+06.3)
1 Döner Buharlaştırıcı ve Vakum Sstemi
1 Adet Alındı
2 Çalkalamalı inkübatör 1 Adet Alındı
Ara Toplam Sarf Malzemesi Adı
(03.2)
1 Cam malzemeler 56 Adet Alındı 2 Kimyasal ve enzim 57 Adet Alındı 3 HPLC kolon + guard
kolonu (OB kolon ) 1 Adet Alındı
4 HPLC kolon + guard kolonu (OD kolon )
1 Adet Alındı
5 Milipore Progard kartuş
2 Adet Alındı
6 Milipore Tank filtresi 2 Adet Alındı 7 Milipore Q-Gard
kartuş 1 Adet Alındı
Ara Toplam Hizmet Alımı Adı
(03.5+03.8+06.6)
1 Dolgulu kolon biyoreaktör yapımı
2 Adet Alındı
2 3 4 Ara Toplam Seyahat (03.3) 1 2 3 TOPLAM
b) Makine ve Teçhizatın Konumu ve İlerideki Kullanımına Dair Açıklamalar
Projeden temin edilen “Döner Buharlaştırıcı ve Vakum Sistemi “ çalışır durumda ve ileriki çalışmalarda işlev görebilecek durumdadır.
Projeden temin edilen “Çalkalamalı inkübatör “ çalışır durumda ve ileriki çalışmalarda işlev görebilecek durumdadır.

c) Teknik ve Bilimsel Ayrıntılar (varsa Kesim III'de yer almayan analiz ayrıntıları)
Asetofenon’ a ait kalibrasyon grafiği
y = 4E+06x
R2 = 0,9975
0
20000000
40000000
60000000
80000000
100000000
120000000
0 5 10 15 20 25 30
C, mM
Ala
n
Şekil 41. Asetofenon’ a ait kalibrasyon grafiği, 254 nm
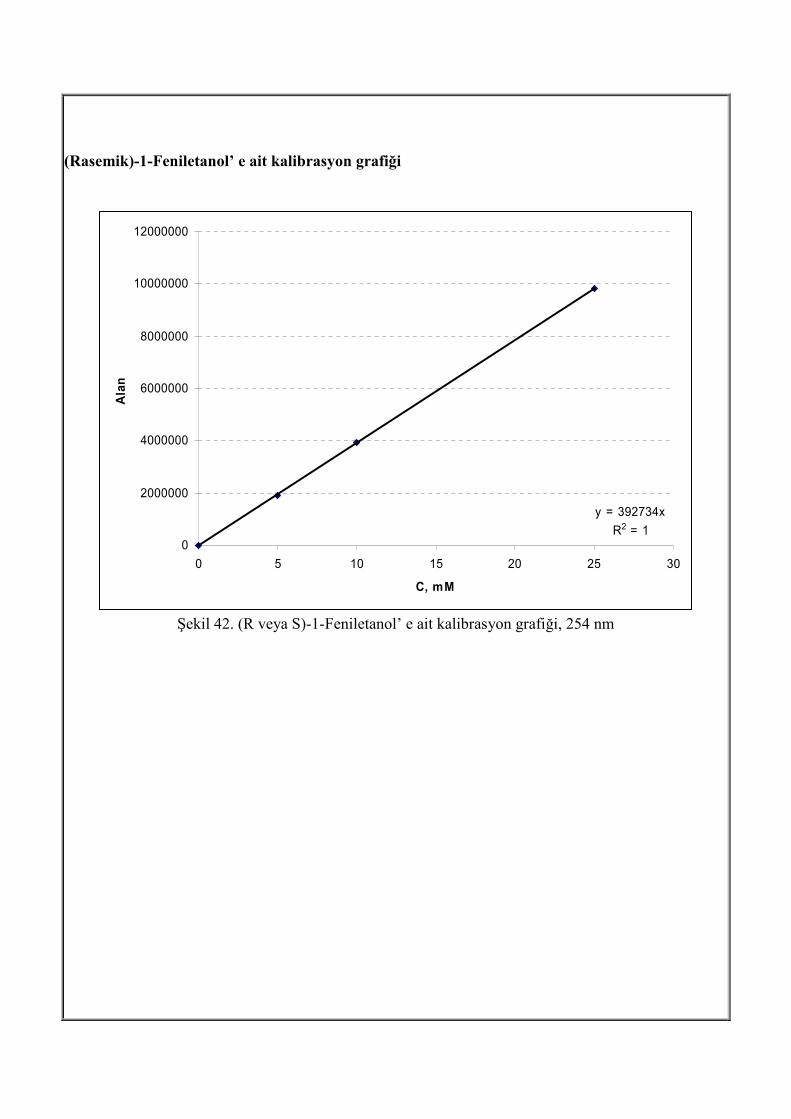
(Rasemik)-1-Feniletanol’ e ait kalibrasyon grafiği
y = 392734x
R2 = 10
2000000
4000000
6000000
8000000
10000000
12000000
0 5 10 15 20 25 30
C, mM
Ala
n
Şekil 42. (R veya S)-1-Feniletanol’ e ait kalibrasyon grafiği, 254 nm

Asetofenona ait fiziksel özellikler
Diğer adı Metil fenil keton
Kapalı formülü CH3COC6H5
CAS no 98-86-2
Molekül ağırlığı 120.15 g/mol
Yoğunluk 1.03 g/mL (25 °C da)
Çözünürlük 5.5 g/L (25 oC da)
Kaynama noktası 202 oC
Erime noktası 19-20 oC
(R,S)-1-Feniletanol’ e ait fiziksel özellikler
Diğer adı Metil fenil karbinol, stiren alkol, alfa-metilbenzil alkol
Kapalı formülü C6H5CH(OH)CH3
CAS no 98-85-1
Molekül ağırlığı 122.16 g/mol
Yoğunluk 1.012 g/mL (25 °C da)
Kaynama noktası 204 oC
Erime noktası 19-20 oC

Lactobacillus kefir NRRL B-1839’ a ait hücre büyüme eğrisi (600 nm)
0
1
2
3
4
5
6
0 5 10 15 20 25 30
t, h
Ab
so
rba
ns
(6
00
nm
)
Şekil 43. (Lactobacillus kefir NRRL B-1839’ a ait hücre büyüme eğrisi
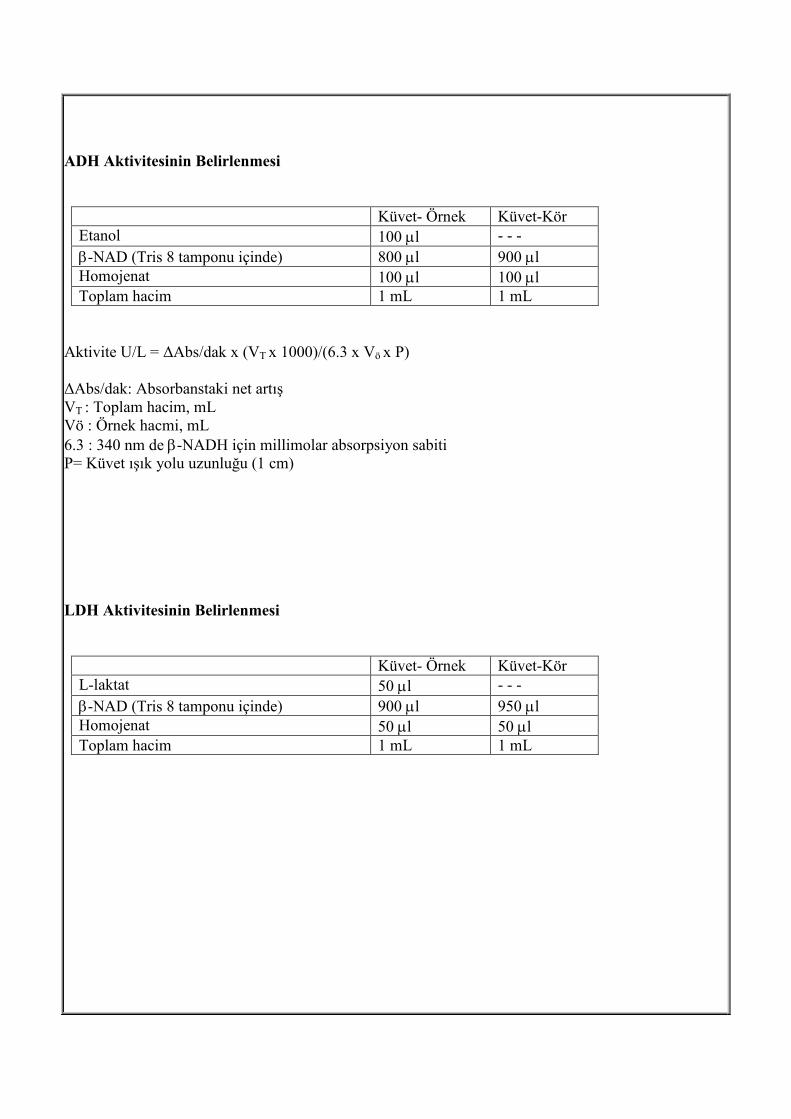
ADH Aktivitesinin Belirlenmesi
Küvet- Örnek Küvet-Kör Etanol 100 µl - - -
β-NAD (Tris 8 tamponu içinde) 800 µl 900 µl Homojenat 100 µl 100 µl Toplam hacim 1 mL 1 mL
Aktivite U/L = ∆Abs/dak x (VT x 1000)/(6.3 x Vö x P) ∆Abs/dak: Absorbanstaki net artış VT : Toplam hacim, mL Vö : Örnek hacmi, mL 6.3 : 340 nm de β-NADH için millimolar absorpsiyon sabiti P= Küvet ışık yolu uzunluğu (1 cm) LDH Aktivitesinin Belirlenmesi
Küvet- Örnek Küvet-Kör L-laktat 50 µl - - -
β-NAD (Tris 8 tamponu içinde) 900 µl 950 µl Homojenat 50 µl 50 µl Toplam hacim 1 mL 1 mL

Asetofenon, R-1-Feniletanol ve S-1-Feniletanol’ e ait HPLC kromatogramı
Minutes
0 5 10 15 20 25 30 35 40
mAU
0
1000
2000
3000
mAU
0
1000
2000
3000
8.288
11.087
13.227
35.013
UV-254nm
Retention Time
Şekil 44. Asetofenon, R-1-Feniletanol ve S-1-Feniletanol’ e ait HPLC kromatogramı (Kolonda kalma süreleri: S-1-Feniletanol: 8. dakika, R-1-Feniletanol: 11. dakika, Asetofenon: 13. dakika)

Farklı kalma sürelerinde, % verim ve % enantiyomerik aşırılık değerlerinin zamanla değişimi Çizelge1 Dolgulu kolon biyoreaktörde zamanla % dönüşüm, % enantiyomerik aşırılık değerleri (τ= 9 h, Cglukoz= 0.02 g/ml (derişim 110mM), Cso=15 mM, T=35 oC)
Zaman, h % Dönüşüm % ee 1 26 99 2 36 99 3 49 99 4 62 99 5 64 99 6 68 99 7 81 99 8 86 99 9 86 99 10 86 99
Çizelge2 Dolgulu kolon biyoreaktörde zamanla % dönüşüm, % enantiyomerik aşırılık değerleri (τ= 4.5 h, Cglukoz=0.02 g/ml (derişim 110mM), Cso=15 mM, T=35 oC)
Zaman, h % Dönüşüm % ee 1 16 99 2 21 99 3 49 99 4 55 99 5 60 99 6 65 99 7 79 99 8 79 99 9 79 99 10 79 99

Çizelge 3Dolgulu kolon biyoreaktörde zamanla % dönüşüm, % enantiyomerik aşırılık değerleri (τ= 3 h, Cglukoz= 0.02 g/ml (derişim 110mM), Cso=15 mM, T=35 oC)
Zaman, h % Dönüşüm % ee 1 29 99 2 36 99 3 60 99 4 60 99 5 62 99 6 65 99 7 68 99 8 68 99 9 68 99 10 68 99
Çizelge 4Dolgulu kolon biyoreaktörde zamanla % dönüşüm, % enantiyomerik aşırılık değerleri (τ= 2.25 h, Cglukoz= 0.02 g/ml (derişim 110mM), Cso=15 mM, T=35 oC)
zaman dönüşüm %ee 1 3 99 2 8 99 3 26 99 4 29 99 5 49 99 6 52 99 7 52 99 8 55 99 9 55 99 10 55 99
Kullanılan Tamponlar 50X TAE (Tris-Asetat-EDTA) Elektroforez Tamponu
Bileşen 1 litre için gereken miktarlar
Son 1X derişim
1 Tris base
242 g 40 mM
2 Dumanlı asedik asit
57,1 ml 20 mM
3 0,5 M EDTA (pH 8,0)
100 ml 1 mM
4 H2O
Gereken miktarda
10X TBE (Tris-Borat-EDTA) Elektroforez Tamponu
Bileşen 1 litre için gereken miktarlar
Son 1X derişim
1 Tris base
108 g 89 mM
2 Borik asit
55 ml 89 mM
3 0,5 M EDTA (pH 8,0)
40 ml 2 mM
4 H2O
Gereken miktarda
TE (Tris-EDTA) Tamponu, pH 7,4, 7,6 veya 8,0
Bileşen 1 litre için gereken miktarlar
Son 1X derişim
1 1 M Tris, pH 7,4, 7,6, 8,0 10 ml 10 mM 2 0,5 M EDTA (pH 8,0) 2 ml 1 mM 3 H2O
Gereken miktarda 2 mM

Şekil 45.Lambda DNA/Pst I Digest marker’ a ait % 1.5 agaroz jel elektroforezi

d) Sunumlar (bildiriler ve teknik raporlar) (Altyapı Projeleri için uygulanmaz)
Uluslararası Tebliğler
Ö. Aydoğan, E. Bayraktar, Ü. Mehmetoğlu, “Enantioselective reduction of ketones catalyzed by Rhyzopus oryzae and Lactobacillus kefiri whole cells”, 19th International Congress of Chemical and Process Engineering CHISA 2010 and the 7th European Congress of Chemical Engineering ECCE-7, 28 August - 1 September 2010, Prague, Czech Republic. Ö. Aydoğan, E. Bayraktar, Ü. Mehmetoğlu, “Enantioselective R-1-Phenlyethanol Production using Free and Immobilized Lactobacillus kefir”, 6th Chemical Engineering Conference for Collaborative Research in Eastern Mediterranean Countries (EMCC6), 7-12.03.2010, Belek, Antalya, Turkey.
Ulusal Tebliğler
Ö. Aydoğan, E. Bayraktar, Ü. Mehmetoğlu, “Lactobacillus Kefir Biyokatalizörlüğünde Asimetrik Biyoindirgemeyle R-1-Feniletanol Üretim Kinetiğinin İncelenmesi”, 9. Ulusal Kimya Mühendisliği Kongresi (UKMK 9) 22-25 Haziran 2010, Gazi Üniversitesi, Ankara.
e) Yayınlar (hakemli bilimsel dergiler) ve tezler (Altyapı Projeleri için uygulanmaz)
Yayınlar
Ö. Aydoğan, E. Bayraktar, Ü. Mehmetoğlu, ”Determination of Effective Diffusion Coefficient of Acetophenone in κ-carrageenan and Asymmetric Bioreduction in Packed Bed Reactor” Journal of Molecular Catalysis B: Enzymatic, Mart 2011 Sunuldu. (SCI-Core)





























