



Genetic diversity and safety aspects of enterococci fromslightly fermented sausages
B. Martin1, M. Garriga1, M. Hugas2 and T. Aymerich1
1IRTA, Meat Technology Centre, Granja Camps i Armet, Girona, Spain, and 2European Food Safety Authority (EFSA), Bruxelles,
Belgium
2004/1134: received 28 September 2004, revised 16 November 2004 and accepted 23 November 2004
ABSTRACT
B. MARTIN, M. GARRIGA, M. HUGAS AND T. AYMERICH. 2005.
Aims: To determine the biodiversity of enterococci from slightly fermented sausages (chorizo and fuet) at species
and strain level by molecular typing, while considering their safety aspects.
Methods and Results: Species-specific PCR and partial sequencing of 16S rRNA and sodA genes were used to
identify enterococcal population. Enterococcus faecium was the most frequently isolated species followed by E.faecalis, E. hirae and E. durans. Randomly amplified polymorphic DNA (RAPD)-PCR revealed species-specific
clusters and allowed strain typing. Sixty strains of 106 isolates exhibited different RAPD profiles indicating a high
genetic variability. All the E. faecalis strains carried virulence genes (efaAfs, esp, agg and gelE) and all E. faeciumisolates carried efaAfm gene. Enterococcus faecalis showed higher antibiotic resistance than the other species. Only
one E. faecium strain showed vanA genotype (high-level resistance to glycopeptides) and E. gallinarum and E.casseliflavus/flavescens isolates showed vanC1 and vanC2/C3 genotypes (low-level resistance only to vancomycin)
respectively.
Conclusions: E. faecalis has been mainly associated with virulence factors and antimicrobial multi-resistance and,
although potential risk for human health is low, the presence of this species in slightly fermented sausages should be
avoided to obtain high quality products.
Significance and Impact of the Study: The enterococcal population of slightly fermented sausages has been
thoroughly characterized. Several relevant safety aspects have been revealed.
Keywords: antibiotic resistance, enterococci, fermented sausages, RAPD-PCR, virulence genes.
INTRODUCTION
Enterococci are ubiquitous micro-organisms that inhabit the
gastrointestinal tract of humans and animals. They are
frequently isolated from fermented meat products with
counts up to 105 CFU g)1 (Teuber et al. 1999; Aymerich
et al. 2003) because of their tolerance to sodium chloride and
nitrite allowing them to survive, and even to multiply, during
fermentation (Giraffa 2002). Their presence in foods is highly
controversial; while some authors consider them undesirable,
indicators of faecal contamination and responsible for the
spoilage of meat products (Franz et al. 1999), others reporttheir important role in flavour development of cheeses,
bioprotection in dairy and meat products and benefits as
probiotics (Coppola et al. 1988; Centeno et al. 1996; Ayme-
rich et al. 2000). In recent decades, although food-borne
enterococci have not yet been clearly proved to be the direct
cause of clinical infections (Adams 1999), enterococci have
unfortunately acquired clinical relevance. They have become
the third cause of nosocomial diseases causing urinary tract
infections, bacteraemia and endocarditis (CDCNNIS System
1998) Enterococcus faecalis has been implicated in 80% of the
cases and E. faecium in 15–20% (Morrison et al. 1997).Virulence mechanisms of enterococci are not com-
pletely known but enterococcal virulence factors includingCorrespondence to: Teresa Aymerich, IRTA, Meat Technology Centre, Granja
Camps i Armet, 17121 Monells, Spain (e-mail: [email protected]).
ª 2005 The Society for Applied Microbiology
Journal of Applied Microbiology 2005, 98, 1177–1190 doi:10.1111/j.1365-2672.2005.02555.x

adherence to host tissue, invasion and abscess formation,
resistance to and modulation of host defence mechanisms,
secretion of cytolysins and production of plasmid-encoded
pheromones have been reported (Jett et al. 1994; Dunny
et al. 1995; Lowe et al. 1995; Singh et al. 1998; Haas and
Gilmore 1999; Shankar et al. 1999).The resistance of enterococci to a wide variety of anti-
microbials contributes to enterococci pathogenicity and
impedes medical treatment of enterococcal infections (Mur-
ray 1990; Mundy et al. 2000). Their ability of gene exchangeby conjugation (Clewell 1990; Franz et al. 1999) may spread
antibiotic resistance and virulence factors between enterococci
or some other pathogenic bacteria (Leclercq et al. 1989;Noble
et al. 1992; Cocconcelli et al. 2003).Vancomycin-resistant
enterococci (VRE) are the main cause of concern, as
enterococci are resistant to many antibiotics and vancomycin
is one of the last options for antimicrobial therapy in some
infections by Gram-positive bacteria (Wegener et al. 1999).The discovery of genes for vancomycin and other antibiotic
resistances in plasmids and transposons (Murray 1990;
Leclercq 1997) increased the concern. Virulence factors have
been found in food strains (Eaton and Gasson 2001; Franz
et al. 2001; Semedo et al. 2003) and antibiotic-resistant
enterococci seem to be widespread in raw food (Giraffa
2002) and have also been isolated from dairy products, ready-
to-eat foods and meat products (Quednau et al. 1998; Teuberet al. 1999; Giraffa et al. 2000; Baumgartner et al. 2001).Accurate species identification and strain typing is
important to evaluate the genetic diversity among entero-
cocci populations and to select nonpathogenic bacteria for
further use in food technology and probiotics (Franz et al.1999; Giraffa 2002). Species identification in routine and
clinical laboratories uses mainly phenotypic methods (Dev-
riese et al. 1996). Nevertheless these methods are often
unreliable and can take several days (Cheng et al. 1997).Several molecular approaches have been developed as an
alternative to facilitate reliable identification of enterococcal
species, such as SDS-PAGE of whole cell proteins (Des-
cheemaeker et al. 1997; Devriese et al. 2002), PCR-basedmethods (Dutka-Malen et al. 1995) and sequenciation of
sodA and 16S rRNA genes (Poyart et al. 2000; Angeletti
et al. 2001).Genetic typing techniques, such as plasmid profiling
(Farber et al. 1996), pulsed-field gel electrophoresis of DNA
macro-restriction patterns (Descheemaeker et al. 1997;
Vancanneyt et al. 2002), randomly amplified polymorphic
DNA (RAPD)-PCR (Cocconcelli et al. 1995; Descheemae-
ker et al. 1997) and amplified fragment length polymor-
phisms (Antonishyn et al. 2000; Vancanneyt et al. 2002)
have been widely used to characterize clinical and dairy
isolates of enterococci.
The aim of this study was to analyse the biodiversity of
enterococci present in slightly fermented sausages at species
and strain level, the incidence of several virulence traits and
their antibiotic susceptibility.
MATERIALS AND METHODS
Bacterial strains and culture conditions
Reference strains listed in Table 1 were obtained from
Spanish Type Culture Collection (CECT) and from our
own collection (CTC Meat Technology Center). All strains
were grown in tryptic soya broth (Difco Laboratories,
Detroit, MI, USA) with 0Æ6% yeast extract (TSBYE) at
37�C for 24 h under anaerobic conditions (Oxoid jars with
AnaeroGen; Oxoid, Basingstoke, UK).
Enterococcal strains isolation
Twelve samples of commercial slightly fermented sausages
(pH 5Æ3–6Æ2), six fuets (cold ripened fermented sausages
with black pepper) and six chorizos (cold ripened fermented
sausages with paprika and garlic) from different producers
were purchased at local butchers/supermarkets.
After removal of the casing, 10 g of each sample were
homogenized in 90 ml of 0Æ1% peptone (Difco) and
0Æ85% NaCl (Merck, Darsmstadt, Germany), pH 7Æ0, in a
Stomacher Lab-Blender (model 400; Cooke Laboratories,
Table 1 Reference strains used in this study
Reference strain
Virulence
determinants Origin and/or comment
Enterococcus
faecium CECT410
efaAfm+ Equivalent to ATCC 19434.
Type strain
Enterococcus
faecium CTC492
efaAfm+ Fermented sausages
Enterococcus
faecium CTC496
efaAfm+ Fermented sausages
Enterococcus
faecalis CECT481
efaAfs+
gelE+ esp+Equivalent to ATCC 19433
Enterococcus
faecalis CECT184
efaAfs+
gelE+ agg+Equivalent to ATCC 27285.
Isolated from cheese
Enterococcus
faecalis CECT795
– Equivalent to ATCC 29212.
Control strain for antibiotic
testing
Lactobacillus
sakei CTC494
– Fermented sausages. Used as
negative control
Lactobacillus
curvatus CTC371
– Fermented sausages. Used as
negative control
Lactobacillus
plantarum CTC305
– Fermented sausages. Used as
negative control
efaAfm, efaAfs: cell wall adhesin EfaA for Enterococcus faecium and E.
faecalis respectively; gelE: gene encoding gelatinase; esp: gene encoding
surface protein Esp; agg: gene encoding aggregation protein.
1178 B. MARTIN ET AL.
ª 2005 The Society for Applied Microbiology, Journal of Applied Microbiology, 98, 1177–1190, doi:10.1111/j.1365-2672.2005.02555.x

Alexandria, VA, USA), pour-plated in kanamycin-esculin-
azide agar (Oxoid) and incubated for 24 h at 37�C.Twelve colonies showing typical morphology from each
sample were randomly picked and grown in plate counting
agar for 24 h at 37�C. In samples with low counts of
enterococci (<10 colonies per plate) all colonies were
picked. Isolates able to agglutinate with latex (Slidex
Strepto B. BioMerieux, Marcy L’Etoile, France) were
grown overnight in TSBYE and stored at )80�C after
addition of 20% of glycerol.
Enterococcal DNA extraction
Genomic DNA from all strains was extracted from
overnight TSBYE cultures using the DNeasy tissue kit
(Qiagen, Hilden, Germany) following manufacturer’s rec-
ommendations. DNA was quantified using the GeneQuant
RNA/DNA Calculator (Amersham Biosciences, Uppsala,
Sweden), adjusted at 0Æ1 mg ml)1 and stored at )20�C.
Identification of recovered isolates
Determination of enterococci at genus level was assessed by
PCR amplification of tuf gene (elongation factor EF-Tu) as
previously described (Ke et al. 1999).Identification of E. faecium and E. faecalis species was
carried out by ddl multiplex-PCR amplification as reported
by Dutka-Malen et al. (1995). A standard PCR was
performed in a final volume of 25 ll containing 20 mmol l)1
Tris–HCl (pH8Æ0), 50 mmol l)1 KCl, 1Æ5 mmol l)1 MgCl2,
0Æ4 mmol l)1 of each dNTP (Promega, Madison, WI, USA),
0Æ2 lmol l)1 of each primer (Table 2) and 1 U of Taqpolymerase (Roche Molecular Biochemicals, Indianapolis,
IN, USA). Thermal cycling was carried out in the
Table 2 List of primers used in this study
Primer Sequence (5¢–3¢) Target gene Reference
Ent1 TACTGACAAACCATTCATGATG tuf Ke et al. (1999)
Ent2 AACTTCGTCACCAACGCGAAC
EFM1 GCAAGGCTTCTTAGAGA E. faecium ddl Dutka-Malen et al. (1995)
EFM2 CATCGTGTAAGCTAACTTC
EFK1 ATCAAGTACAGTTAGTCTT E. faecalis ddl Dutka-Malen et al. (1995)
EFK2 ACGATTCAAAGCTAACTG
BSF8 AGAGTTTGATCATGGCTCAG 16S rRNA
BSF 343 TACGGGAGGCAGCAG
BSF1541 AAGGAGGTGATCCAGCCGCA
sodA1 CCITAYICITAYGAYGCIYTIGARCC sodA Poyart et al. (2000)
sodA2 ARRTARTAIGCRTGYTCCCAIACRTC
vanA1 GGGAAAACGACAATTGC vanA Dutka-Malen et al. (1995)
vanA2 GTACAATGCGGCCGTTA
vanB1 ATGGGAAGCCGATAGTC vanB Dutka-Malen et al. (1995)
vanB2 GATTTCGTTCCTCGACC
vanC1-F GGTATCAAGGAAACCTC vanC1 Dutka-Malen et al. (1995)
vanC1-R CTTCCGCCATCATAGCT
vanC2/C3-F CTCCTACGATTCTCTTG vanC2/C3 Dutka-Malen et al. (1995)
vanC2/C3-R CGAGCAAGACCTTTAAG
esp1 TTGCTAATGCTAGTCCACGACC esp Eaton and Gasson (2001)
esp2 GCGTCAACACTTGCATTGCCGAA
agg1 AAGAAAAAGAAGTAGACCAAC agg Eaton and Gasson (2001)
agg2 AAACGGCAAGACAAGTAAATA
gelE1 ACCCCGTATCATTGGTTT gelE Eaton and Gasson (2001)
gelE2 ACGCATTGCTTTTCCATC
efaAfs1 GACAGACCCTCACGAATA efaAfs Eaton and Gasson (2001)
efaAfs2 AGTTCATCATGCTGTAGTA
efaAfm1 AACAGATCCGCATGAATA efaAfm Eaton and Gasson (2001)
efaAfm2 CATTTCATCATCTGATAGTA
R1 GGTGCGGGAA Random primer
R2 GTTTCGCTCC Random primer
R5 AACGCGCAAC Random primer
M13R2 GGAAACAGCTATGACCATGA Random primer
BIODIVERSITY OF ENTEROCOCCI FROM FERMENTED SAUSAGES 1179
ª 2005 The Society for Applied Microbiology, Journal of Applied Microbiology, 98, 1177–1190, doi:10.1111/j.1365-2672.2005.02555.x

GeneAmp PCR System 2700 (Applied Biosystems, Foster
City, CA, USA) using the following programme: initial
denaturation at 94�C for 2 min; 30 cycles of denaturation at
94�C for 1 min, annealing at 54�C for 1 min and extension
at 74�C for 1 min; ending with a final extension at 74�C for
5 min.
The other enterococcal species were identified by partial
sequenciation of 16S rRNA gene and sodA gene. PCR
amplification of V1–V3 regions of 16S rRNA gene was
assessed by oligonucleotides BSF8 and BSF1541 (Table 2).
The standard PCR reaction was used except for an annealing
temperature of 50�C and an elongation step of 1Æ5 min DNA
was purified with GeneClean kit II (Bio 101, La Jolla, CA,
USA). DNA was sequenced by using the BigDye Termi-
nator v3Æ1 cycle sequencing kit, the oligonucleotides BSF8,
BSF343 (Table 2) and BSR1541 and the sequencing device
ABI PRISM 310 (Applied Biosystems). The sodA gene was
partially sequenced as previously described by Poyart et al.(2000). The BLAST-W2 and ClustalW software from the
European Bioinformatics Institute (Wellcome Trust Gen-
ome Campus, Hinxton, UK. Web site: http://www.ebi.
ac.uk) was used for the analysis of sequences.
PCR amplification of virulence genes
Primers used for the amplification of genes esp (surface
protein gene), agg (aggregation substance gene), gelE (coding
for gelatinase), efaAfs and afaAfm (genes coding for cell wall
adhesins in E. faecalis and E. faecium respectively) were
those described by Eaton and Gasson (2001) (Table 2). PCR
conditions were the same as described in the standard PCR
except for annealing temperature (50�C for efaAfm, 55�C for
efaAfs, 56�C for agg, 60�C for gelE and 61�C for esp). AllPCR performances included positive and negative control
strains (Table 1).
Antibiotic susceptibility testing
Susceptibility testing was based on the agar overlay disc
diffusion test described by Charteris et al. (1998) with some
modifications. Enterococci were grown overnight in TSBYE
at 37�C under anaerobic conditions (Oxoid jars with
AnaeroGen; Oxoid). Eight millilitres of Mueller–Hinton
Agar (MHA, Oxoid) kept at 50�C were inoculated with
0Æ2 ml of the grown culture. Petri dishes containing 15 ml of
MHA were overlaid with 7Æ2 ml of the inoculated MHA and
allowed to solidify at room temperature. Antibiotic discs
(Oxoid) were placed onto the overlaid plates and all plates
were incubated for 20–24 h at 37�C under anaerobic
conditions. All enterococcal recovered isolates were screened
for their susceptibility to ampicillin (10 lg), chloramphen-
icol (30 lg), ciprofloxacin (5 lg), gentamicin (120 lg),erythromycin (15 lg), linezolid (30 lg), nitrofurantoin
(300 lg), penicillin G (10 U), quinupristin/dalfopristin
(15 lg), rifampicin (5 lg), teicoplanin (30 lg), tetracycline(30 lg) and vancomycin (30 lg). The results were inter-
preted following the recommendations of the National
Committee for Clinical Laboratory Standards (NCCLS
2002). Enterococcus faecalis CECT795 (equivalent to
ATCC29212) was used as the control strain for monitoring
the performance of the study conditions.
PCR amplification of van genes
In addition to the disc diffusion test, high and low-level
resistance to glycopeptides was confirmed using the PCR
protocol described by Dutka-Malen et al. (1995) with the
specific primers for the genes vanA, vanB, vanC1 and
vanC2/C3 (Table 2). Two separate PCR amplifications were
carried out, one to detect vanA and vanB genotypes and
another for vanC1 and vanC2/C3 in order to avoid
interferences due to the high number of primers. PCR
mixtures and amplification programmes were the same as
described in the standard PCR except for annealing
temperature (57�C for vanA-vanB and 55�C for vanC1-vanC2/C3) and number of cycles (25).
RAPD-PCR
Four random primers (R1, R2, R5 and M13R2; Table 2)
were tested for their ability to discriminate between strains
of enterococci and for their reproducibility. Primers R5 and
M13R2 showed higher discrimination capacity and they
were selected for RAPD-PCR analysis. Each 25-ll PCR mix
contained 20 mmol l)1 Tris–HCl (pH 8Æ0), 50 mmol l)1
KCl, 1Æ5 mmol l)1 MgCl2 (2Æ5 mmol l)1 when primers R1,
R2 and R5 were used), 0Æ2 mmol l)1 of each dNTP,
0Æ8 lmol l)1 of primer, 2 U of Taq polymerase and 100 ng
of extracted DNA. When primers R1, R2 and R5 were used
the amplification process consisted of 5 min of initial
denaturation at 94�C and 40 cycles consisting of denatur-
ation at 94�C for 1 min, and 40 cycles of denaturation at
94�C for 1 min, annealing at 29�C for 1Æ5 min and
elongation at 72�C for 2 min, followed by a final extension
of 5 min at 74�C. With primer M13R2, 35 cycles were
performed, each consisting of 1 min of denaturation at
94�C, 1 min of annealing at 38�C and elongation at 72�C for
1 min.
After amplification, 10 ll of PCR product were mixed
with 1 ll of loading buffer (40% sucrose, 0Æ25% bromo-
phenol blue) and subjected to agarose electrophoresis for
100 min at 80 mA. Gels were stained with 0Æ1 lg ml)1
ethidium bromide (Sigma Chemical Comp., MO, USA).
Each gel contained two lines of 1 kb DNA ladder
(Invitrogen, Merelbeke, Belgium) as molecular weight
and normalization gel standards. The banding profiles
1180 B. MARTIN ET AL.
ª 2005 The Society for Applied Microbiology, Journal of Applied Microbiology, 98, 1177–1190, doi:10.1111/j.1365-2672.2005.02555.x

were visualized under u.v. light and digitalized by the
Gelprinter photodocumentation equipment (TDI, Barce-
lona, Spain).
Clustering analysis
Relevant safety traits (antibiotic resistance and presence of
virulence factors) and RAPD-PCR electrophoretic profiles
were analysed using the software Fingerprinting II Infor-
matix (Bio-Rad Laboratories, Hercules, CA, USA). Safety
traits were recorded as �1� or �0� if positive or negative
respectively. The similarity matrix was defined by the
Simple Matching coefficient and cluster analysis were
carried out by the unweighted pair group method with
arithmetic averages (UPMGA).
For RAPD-PCR electrophoretic profiles, conversion,
normalization and analysis were performed by the software
package Fingerprinting II Informatix (Bio-Rad). RAPD
profiles of both primers were combined and compared using
the Dice coefficient; correlation coefficients were calculated
by the UPMGA.
RESULTS
Identification of recovered isolates
A total of 115 strains were isolated from fermented sausages.
Among them, 106 strains were assigned to the genus
Enterococcus on the basis of genus-specific PCR identifica-
tion. Nine strains were identified as Pediococcus pentosaceus(five strains) and P. acidilactici (four strains) by 16S rRNA
gene sequencing (data not shown). The enterococcal isolates,
48 from chorizo and 58 from fuet, were subjected to species
identification by species-specific multiplex-PCR, partial
sequencing of 16S rRNA gene and sodA sequencing.
Multiplex-PCR allowed the identification of all E. faecium(51Æ9% of the isolates) and E. faecalis strains (14Æ2% of the
isolates). Partial 16S rRNA gene and sodA gene sequencing
identified the remaining strains. About 13Æ2% of isolates
were assigned to E. hirae and E. durans each, 5Æ7% to
E. casseliflavus/flavescens, 0Æ94% to E. mundtii and 0Æ94% to
E. gallinarum.Comparing the two types of sausages studied, E. faecium,
with 69% of the isolates, was dominant in fuet while E. hiraeand E. durans represented 17Æ2 and 10Æ3% respectively. Only
3Æ4% were allotted to E. faecalis. Chorizo showed more
enterococcal species diversity. Enterococcus faecium and
E. faecalis were balanced representing 31Æ3 and 27Æ1% of
isolates respectively. Enterococcus durans, E. casseliflavus/flavescens and E. hirae comprised 16Æ7, 12Æ5 and 8Æ3% of the
chorizo isolates. Only one strain from chorizo was identified
as E. mundtii (2Æ1%) and another one as E. gallinarum(2Æ1%).
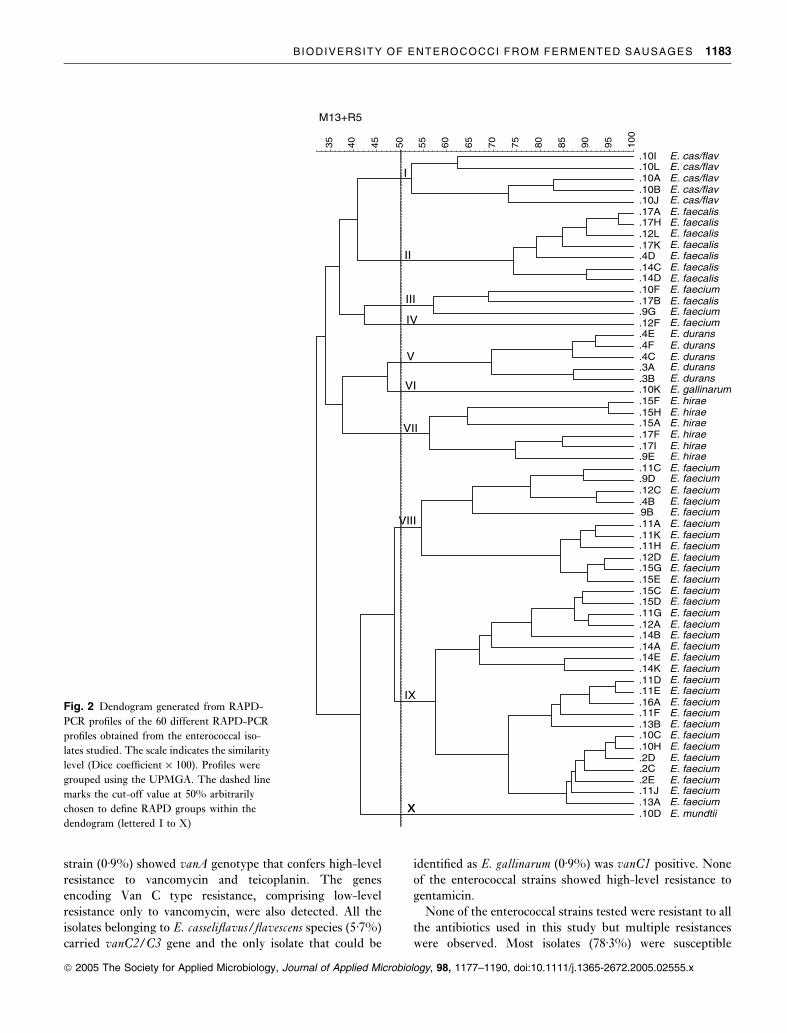
RAPD-PCR typing
To study intraspecies diversity of enterococci isolated from
fermented sausages all strains were subjected to RAPD-PCR
analysis with two different primers, M13R2 and R5 (Fig. 1).
The reproducibility of RAPD-PCR assay and running
conditions estimated by analysis of repeated DNA extracts
of several type strains was >92% (results not shown).
After numerical analyses of the combined RAPD-PCR
profiles of the two primers used, 60 different patterns were
obtained (Fig. 2). When the similarity value of 92% was
considered, 56 strains of 106 could be differentiated. Ten
RAPD clusters were defined at a similarity level of 50% and
eight species-specific clusters were found. Cluster I grouped
all E. casseliflavus/flavescens strains (5Æ7% of isolates)
although they were distributed in two different subgroups.
Cluster II contained all but one E. faecalis strains (13Æ2% of
isolates). Cluster V grouped all E. durans strains (13Æ2% of
isolates). cluster VI included only one strain assigned to E.gallinarum species and cluster VII grouped all E. hiraestrains (13Æ2% of isolates). All E. faecium isolates, except for
three single strains, were grouped in clusters VIII and IX.
Cluster VIII was composed of 11 different profiles distri-
buted in two subgroups defined at a similarity level of 60%.
This cluster included 17 E. faecium strains (16% of isolates).
Cluster IX was the major RAPD group as it included 35 E.faecium isolates (33% of isolates). This cluster was composed
of 20 profiles also distributed in two subgroups defined at a
similarity level of 60%. Cluster X comprised only the isolate
assigned to the species E. mundtii. The remaining clusters
(III and IV) contained the strains that did not group in their
corresponding species cluster. Cluster III contained two
isolates of E. faecium (including the vanA+ E. faecium) andone E. faecalis isolate. Cluster IV comprised one single
isolate of E. faecium.The isolates from fuet and chorizo did not form separated
RAPD clusters. Genotypic grouping was compared with
potentially pathogenic traits and antibiotic resistance of E.faecium strains. The three E. faecium isolates containing all
virulence genes studied were clustered in the same subgroup
of cluster VII. Multi-resistant strains were distributed in
several RAPD groups.
Incidence and distribution of virulencedeterminants
A molecular screening of the genes encoding virulence
factors revealed distinct trends in the occurrence of
virulence between species. Enterococcus faecium and E.faecalis strains carried the virulent-associated genes (efa,esp, agg, gelE) while the other species were clear of them.
All E. faecalis and all E. faecium carried the gene encoding
for efaAfs and efaAfm respectively. All E. faecalis strains
BIODIVERSITY OF ENTEROCOCCI FROM FERMENTED SAUSAGES 1181
ª 2005 The Society for Applied Microbiology, Journal of Applied Microbiology, 98, 1177–1190, doi:10.1111/j.1365-2672.2005.02555.x
carried the gelE gene (coding for gelatinase) and agg gene
while only 5Æ5% of E. faecium isolates harboured them.
About 99Æ3% of E. faecalis strains and 5Æ5% of E. faeciumisolates carried the esp gene.
A higher incidence of virulence determinants was observed
in enterococci from chorizo, due to its larger proportion of E.faecalis strains. The gelE, esp and agg genes were detected in
33Æ3%of isolates and efaAfm and efaAfs geneswere detected in31Æ3 and 27Æ1% of the chorizo strains respectively. In general,
enterococci from fuet presented fewer incidences of virulence
genes, as only 3Æ4% carried gelE, agg and efaAfs genes and1Æ7% carried esp gene. However, the gene encoding efaAfmadhesin was detected in 69Æ0% of isolates from fuet, due to the
large proportion of E. faecium strains in this product.
Antibiotic susceptibility
All enterococcal isolates were subjected to antibiotic
susceptibility testing towards 13 antibiotics using a modified
disc diffusion technique. Vancomycin resistance was con-
firmed by PCR. The prevalence of antibiotic resistance
among enterococcal species is shown in Table 3. A high
resistance to rifampicin was observed in all the enterococcal
species. About 100% of E. faecalis and E. durans isolates,92Æ9% of E. hirae, 83Æ3% of E. casseliflavus/flavescens and69Æ1% of E. faecium isolates presented resistance to
rifampicin. A high incidence of E. faecalis-resistant strainsto chloramphenicol (93Æ3%), erythromycin (93Æ3%) and
tetracycline (86Æ7%) was recorded. Enterococcus hirae strainsshowed an elevated incidence of resistance to ciprofloxacin
(71Æ4%) and nitrofurantoin (85Æ7%).
The disc diffusion test showed poor reliability for
vancomycin resistance as previously reported (Swenson
et al. 1989; Temmerman et al. 2003), four false negative and34 false positive were detected for vancomycin resistance
among enterococci from slightly fermented sausages. A very
low incidence of resistance to glycopeptides was found. The
presumptively vancomycin- and teicoplanin-resistant iso-
lates, obtained by the disc diffusion assay, were confirmed
by PCR amplification of van genes. Only one E. faecium
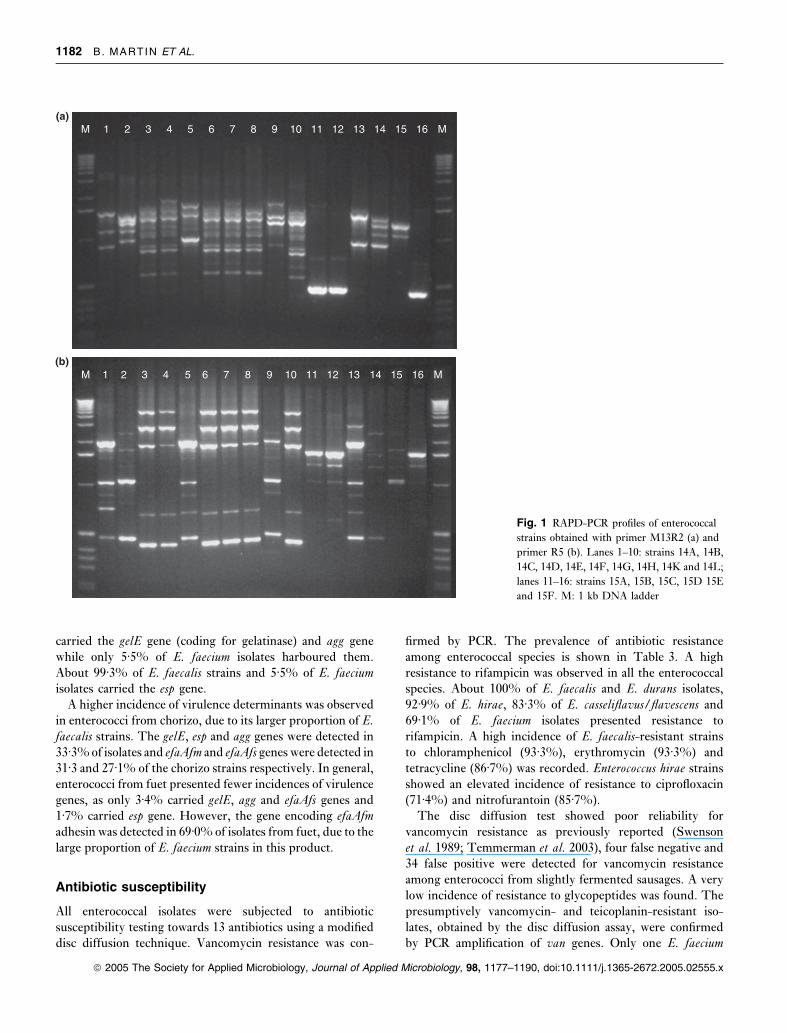
Fig. 1 RAPD-PCR profiles of enterococcal
strains obtained with primer M13R2 (a) and
primer R5 (b). Lanes 1–10: strains 14A, 14B,
14C, 14D, 14E, 14F, 14G, 14H, 14K and 14L;
lanes 11–16: strains 15A, 15B, 15C, 15D 15E
and 15F. M: 1 kb DNA ladder
1182 B. MARTIN ET AL.
ª 2005 The Society for Applied Microbiology, Journal of Applied Microbiology, 98, 1177–1190, doi:10.1111/j.1365-2672.2005.02555.x
strain (0Æ9%) showed vanA genotype that confers high-level
resistance to vancomycin and teicoplanin. The genes
encoding Van C type resistance, comprising low-level
resistance only to vancomycin, were also detected. All the
isolates belonging to E. casseliflavus/flavescens species (5Æ7%)
carried vanC2/C3 gene and the only isolate that could be
identified as E. gallinarum (0Æ9%) was vanC1 positive. None
of the enterococcal strains showed high-level resistance to
gentamicin.
None of the enterococcal strains tested were resistant to all
the antibiotics used in this study but multiple resistances
were observed. Most isolates (78Æ3%) were susceptible
35
M13+R5
40
45
50
55
60
65
70
75
80
85
90
95
10
0
E. mundtii
I
II
III
IV
V
VIII
IX
X
VI
VII
.10I
.10L
.10A
.10B
.10J
.17A
.17H
.12L
.17K
.4D
.14C
.14D
.10F
.17B
.9G
.12F
.4E
.4F
.4C
.3A
.3B
.10K
.15F
.15H
.15A
.17F
.17I
.9E
.11C
.9D
.12C
.4B
.
.11A
.11K
.11H
.12D
.15G
.15E
.15C
.15D
.11G
.12A
.14B
.14A
.14E
.14K
.11D
.11E
.16A
.11F
.13B
.10C
.10H
.2D
.2C
.2E
.11J
.13A
.10D
9B
E. cas/flavE. cas/flavE. cas/flavE. cas/flavE. cas/flavE. faecalisE. faecalisE. faecalisE. faecalisE. faecalisE. faecalisE. faecalis
E. faecalisE. faecium
E. faecium
E. faeciumE. faeciumE. faeciumE. faeciumE. faeciumE. faeciumE. faeciumE. faeciumE. faeciumE. faeciumE. faeciumE. faeciumE. faeciumE. faeciumE. faeciumE. faeciumE. faeciumE. faeciumE. faeciumE. faeciumE. faeciumE. faeciumE. faeciumE. faeciumE. faeciumE. faeciumE. faeciumE. faeciumE. faeciumE. faeciumE. faecium
E. faeciumE. duransE. duransE. duransE. duransE. duransE. gallinarumE. hiraeE. hiraeE. hiraeE. hiraeE. hiraeE. hirae
X
Fig. 2 Dendogram generated from RAPD-
PCR profiles of the 60 different RAPD-PCR
profiles obtained from the enterococcal iso-
lates studied. The scale indicates the similarity
level (Dice coefficient · 100). Profiles were
grouped using the UPMGA. The dashed line
marks the cut-off value at 50% arbitrarily
chosen to define RAPD groups within the
dendogram (lettered I to X)
BIODIVERSITY OF ENTEROCOCCI FROM FERMENTED SAUSAGES 1183
ª 2005 The Society for Applied Microbiology, Journal of Applied Microbiology, 98, 1177–1190, doi:10.1111/j.1365-2672.2005.02555.x

towards six to 10 of the antibiotics and 33% were susceptible
to nine or more antibiotics.
Clustering analysis of relevant safety traits
To study diversity of enterococcal isolates considering their
safety aspects, antibiotic susceptibility and presence or
absence of virulence genes were analysed using the Infor-
matix software from Bio-Rad. By combining all data, we
could differentiate 78 different patterns (Fig. 3) although
some of them showed similarity levels of 95% (differing only
in one trait). The isolates were grouped in four main groups
(A–D). Group A contained E. faecium and E. faecalis isolatespresenting genes associated to virulence traits. These strains
represented 16% of isolates and were distributed in 10
different patterns. The isolates were resistant towards six to
11 antibiotics and all of them showed resistance to
chloramphenicol, erythromycin and rifampicin. Group B
contained 57Æ5% of isolates distributed in 44 different
profiles. These strains were resistant towards one to six
antibiotics (one isolate was susceptible to all antibiotics
tested) and did not carry virulence genes, except for efaAfmin E. faecium isolates. This group included E. gallinarum and
E. casseliflavus/flavescens isolates that carried vanC1 and
vanC2/C3 genes respectively. Group C contained 16% of
isolates and included the vanA+ strain. All of them were
resistant towards six to 12 antibiotics and did not carry
virulence genes except for efaAfm in E. faecium strains. All
isolates were resistant to ampicillin, penicillin and rifampi-
cin. Group D contained 8Æ5% of enterococcal isolates, all
showing resistance towards four to seven antibiotics and
carrying efaAfm gene. All isolates were resistant to eryth-
romycin, penicillin and tetracycline. Two isolates were not
included in any of the four groups, one E. faecalis that didnot carry esp gene and was resistant to only three antibiotics
and one E. faecium that showed an intermediate profile
among groups B, C and D.
DISCUSSION
Identification of recovered isolates
As classical identification of enterococci by phenotypic
methods often gives ambiguous results, several molecular
methods have been developed as alternative tools to reliable
identification of Enterococcus species (Domig et al. 2003). Inthe present study, the identification of enterococci was
carried out by PCR amplification (Dutka-Malen et al. 1995),partial sequence analysis of 16S rRNA and sodA genes as
previously described (Poyart et al. 2000). PCR amplification
of ddl gene allowed the identification of all E. faecium and
E. faecalis strains. The gene sodA constituted a more
discriminative target sequence than 16S rRNA gene and
allowed differentiation of all enterococci except between
E. casseliflavus and E. flavescens which it was not possible to
discriminate. Partial 16S rRNA gene sequencing did not
complete the identification due to the high homology
between closely related enterococcal species. As reported
by Devriese and Pot (1995) and Descheemaeker et al.(1997), E. casseliflavus/flavescens could not even be separated
by SDS-PAGE of whole cell. It has been suggested that
E. flavescens should be considered as an E. casseliflavusbiovar rather than a separate species (Descheemaeker et al.1997).
Table 3 Prevalence of antibiotic-resistant enterococci isolated from slightly fermented sausages to selected antibiotics using disc diffusion test,
expressed as percentage
Antibiotic
Total strains
(n ¼ 106)
E. faecium
(n ¼ 55)
E. faecalis
(n ¼ 5)
E. hirae
(n ¼ 14)
E. durans
(n ¼ 14)
E. casseliflavus/
flavescens (n ¼ 6)
Ampicillin (10 lg) 27Æ4 30Æ9 13Æ3 14Æ3 57Æ1 0
Chloramphenicol (30 lg) 26Æ4 20 93Æ3 14Æ3 7Æ1 0
Ciprofloxacin (5 lg) 52Æ8 54Æ5 46Æ7 71Æ4 50 16Æ7Gentamicin (120 lg) 0 0 0 0 0 0
Erythromycin (15 lg) 50Æ9 56Æ4 93Æ3 35Æ7 8Æ6 0
Linezolid (30 lg) 32Æ1 21Æ8 40 28Æ6 57Æ1 50
Nitrofurantoin (300 lg) 50Æ9 52Æ7 33Æ3 85Æ7 42Æ9 33Æ3Penicillin G (10 U) 42Æ5 58Æ2 6Æ7 28Æ6 57Æ1 0
Quinupristin/dalfopristin (15 lg) 37Æ7 27Æ3 –* 14Æ3 57Æ1 0
Rifampicin (5 lg) 80Æ2 69Æ1 100 92Æ9 100 83Æ3Teicoplanin (30 lg)� (10Æ4) 0Æ9 (9Æ1) 1Æ8 (33Æ3) 0 (7Æ1) 0 (0) 0 (0) 0
Tetracycline (30 lg) 47Æ2 29Æ1 86Æ7 28Æ6 35Æ7 0
Vancomycin (30 lg)� (27Æ4) 7Æ5 (12Æ7) 1Æ8 (86Æ7) 0 (14Æ3) 0 (35Æ7) 0 (33Æ3) 100
*Intrinsic resistance.
�Vancomycin and teicoplanin results using disc diffusion test are expressed in parentheses followed by final results after PCR screening of van genes.
1184 B. MARTIN ET AL.
ª 2005 The Society for Applied Microbiology, Journal of Applied Microbiology, 98, 1177–1190, doi:10.1111/j.1365-2672.2005.02555.x

Fig. 3 Dendogram generated from safety traits data of the 78 different profiles obtained from the enterococcal isolates studied. The scale indicates
the similarity level (Simple Matching coefficient · 100). Profiles were grouped using the UPMGA method. Main groups are lettered A–D. AMP,
ampicillin; C, chloramphenicol; CIP, ciprofloxacin; CN, gentamicin; E, erythromycin; LZD, linezolid; F, nitrofurantoin; P, penicillin G; QD,
quinupristin/dalfopristin; RD, rifampicin; TEC, teicoplanin; TE, tetracycline, VA, vancomycin
BIODIVERSITY OF ENTEROCOCCI FROM FERMENTED SAUSAGES 1185
ª 2005 The Society for Applied Microbiology, Journal of Applied Microbiology, 98, 1177–1190, doi:10.1111/j.1365-2672.2005.02555.x

In this study, the most common enterococcal species found
in the slightly fermented sausages was E. faecium (51Æ9%).
This is in agreement with Reuter (1995) but in contrast with
recent studies that indicate E. faecalis as being more frequent
in food from animal origin thanE. faecium (Peters et al. 2003).Klein (2003) reported difficulties of E. faecium isolation and
enumeration in selective media and a consequent underesti-
mation of this species in meat and meat products when
compared with E. faecalis. In our study, E. faecalis, E. hiraeand E. durans isolates were very balanced in fermented
sausages when compared with the results of Devriese et al.(1995), who identified E. faecium, E. faecalis and, less
frequently, E. hirae/E. durans in meat and fermented meat.
In food from animal origin, Peters et al. (2003) found a higherproportion ofE. faecalis (72%) and a lower proportion (6%) of
E. durans/E. hirae compared with our results. Enterococcuscasseliflavus/flavescens, E. gallinarum and E. mundtii were
minority species (between 0Æ9 and 5Æ7%). Peters et al. (2003)found a similar proportion of E. casseliflavus and E. gallinarumbut they did not detect any E. mundtii strain.The natural contamination of meat from the gastrointes-
tinal content of the slaughtered animals and occasionally from
human origin may explain the species distribution in this kind
of product. Enterococcus faecium and E. faecalis are usually thespecies most isolated from the intestinal tract of domestic
animals and humans while E. durans/E. hirae, E. gallinarumand E. avium are, in general, less frequent (Devriese and Pot
1995; Klein 2003). In pig excrement, E. faecium is dominant
over E. faecalis, and E. hirae is a common enterococcal species
(Devriese et al. 1987; Devriese and Pot 1995). Enterococcusdurans is a frequent inhabitant in preruminant calves and in
chicken (Devriese and Pot 1995) and its presence in fermented
sausages might suggest a cross-contamination from meat of
different animal species in the manufacturing process.
Enterococcus casseliflavus, E. mundtii and E. gallinarum are
rare in animals and humans. Enterococcus mundtii has beenisolated from plants and soils (Collins et al. 1986; Niemi et al.1993) and occasionally from meat and meat products (Dev-
riese et al. 1995; Klein et al. 1998; Peters et al. 2003).The heterogeneity among enterococcal species found in
chorizo when compared with fuet could be explained by
the differences in the composition of both products. Fuet
contains black pepper whereas chorizo is made with
paprika and garlic. Also the initial content of enterococci
in the meat butter, human cross-contamination, the
temperature, the pH and the relative humidity during
processing can affect the final enterococcal population
from product to product.
RAPD-PCR typing
The RAPD-PCR profiles have proved to be a sensitive and
efficient molecular method for the characterization of inter-
strains variations. In this study, the use of two primers
(M13R2 and R5) and two different PCR conditions enabled
the elucidation of the genetic diversity among isolates and
within enterococcal species. Eight species-specific clusters
were obtained at a similarity level of 50%, for which RAPD-
PCR was useful not only for strain typing, but also for
species identification of isolates. All but three E. faeciumisolates were distributed into two separated clusters that
were each further subdivided in two main subgroups, but no
correlation with type of product, pathogenicity or antibiotic
resistance could be established. Vancanneyt et al. (2002) alsofound two genomic groups in E. faecium from various
sources although no phenotypic features could clearly
differentiate one group from the other. These results may
suggest the presence of exchange genetic material in the
enterococcal population.
Clustering analysis of genotypic typing and safety traits
showed 60 different RAPD profiles and 78 different patterns
respectively. Nevertheless, if considering the reproducibility
value of 92% for RAPD analysis and a similarity value of
95% to differentiate among safety traits patterns, 54 and 55
isolates were characterized respectively. These results
suggest that phenotypic and genotypic traits are comple-
mentary to characterize individual strains.
Strains with identical RAPD profiles were found in the
same sample and in products from different origins. This
could be explained by the predominance of a particular
strain among the enterococci population in a product or, by
the fact that different sausage producers can obtain meat
from the same slaughterhouse, representing a common meat
origin.
Incidence and distribution of virulencedeterminants
The incidence of virulence genes in the enterococci isolated
from fermented sausages was found to be lower in E. faeciumstrains than in E. faecalis, in accordance with the results of
several authors on food and clinical isolates (Eaton andGasson
2001; Franz et al. 2001; Dupre et al. 2003; Semedo et al.2003). The most widely spread virulence determinants were
cell-wall adhesins. All E. faecium and E. faecalis strains
harboured efaAfm and efaAfs genes respectively. Eaton and
Gasson (2001) and Semedo et al. (2003) also reported a high
incidence of efaAfs and efaAfm among enterococci from food
origin although lower than in clinical strains. Mannu et al.(2003) reported efaA as the only virulence trait present in E.faecium from dairy origin but in a lower percentage. The high
proportion of different adhesins among enterococcal isolates
may constitute an important advantage to the survival of
enterococci in all environments (Semedo et al. 2003).The genes agg and gelE were detected in all E. faecalis
strains and 5Æ5% of E. faecium strains and the gene esp in
1186 B. MARTIN ET AL.
ª 2005 The Society for Applied Microbiology, Journal of Applied Microbiology, 98, 1177–1190, doi:10.1111/j.1365-2672.2005.02555.x

93Æ3% of E. faecalis and 5Æ5% of E. faecium strains. Semedo
et al. (2003) also detected these genes in most E. faecalisisolates from food and clinical origin. Eaton and Gasson
(2001) and Franz et al. (2001) showed lower incidence
among E. faecalis strains and none of the E. faecium strains
harboured these genes. Franz et al. (2001) explained the
high incidence of gelE among food enterococci by their
origin from a protein-rich source; the production of protease
may be a selection mechanism for enterococci growing as it
may enable them to utilize proteins as a source of amino
acids. The enterococcal surface protein (Esp) plays a role in
adhesion and is also involved in immune evasion (Shankar
et al. 1999). Thus, enterococcal strains harbouring this gene
should be clearly undesirable for use as starter cultures in
food (Franz et al. 2001). The aggregation substance has been
described as characteristic of E. faecalis pheromone response
plasmids (Dunny 1990) and in fact, several authors (Franz
et al. 1999; Eaton and Gasson 2001; Dupre et al. 2003)
found only agg+ genotype among isolates of this species. We
detected the agg gene in three E. faecium strains isolated
from a sample with a high proportion of E. faecalis strainssuggesting that they may have acquired this virulence factor
by a natural conjugation gene transfer process. In fact, all E.faecium harbouring agg, esp and gelE genes belonged to the
same sample with a high presence of E. faecalis. Eaton and
Gasson (2001) showed the possibility of virulence determi-
nants transfer from a strain of E. faecalis with a sex
pheromone plasmid into E. faecalis starter strains; althoughthe transfer into E. faecium strains was not achieved by these
authors, sex pheromone cross talk between E. faecium and E.faecalis has been established (Heaton et al. 1996). The
existence of E. faecium isolates of food origin containing agg,esp and gelE genes is in contrast to previous works (Eaton
and Gasson 2001; Franz et al. 2001). Enterococcus faeciumstrains containing these virulence genes may be involved in
the evolution of pathogenic E. faecium strains (Eaton and
Gasson 2001) and related with E. faecium-derived infections.
Another sample with high prevalence of E. faecalis strainsharbouring the virulence genes studied was characterized. In
this sample the remaining isolates were identified as E. hiraeand they all were clear of virulence traits.
Antibiotic susceptibility
It is difficult to assess the role of the food chain as a possible
source of antibiotic-resistant enterococci, but strains resist-
ant to glycopeptides and other antibiotics has been isolated
from foods (Knudtson and Hartman 1993; Teuber et al.1999; Franz et al. 2001).Our results showed that vancomycin-resistant entero-
cocci, one of the major concerns from the clinical point of
view, are not common in slightly fermented sausages. Only
one E. faecium isolate of 106 enterococci presented vanA
genes associated with a high level of resistance to vancomy-
cin and teicoplanin (Arthur et al. 1996). Six motile entero-
cocci carried the vanC gene, associated with a low-level
resistance to vancomycin and intrinsic to the motile
enterococcal species (Leclercq 1997). Teuber et al. (1999)in a study of cheeses from Europe, Quednau et al. (1998) inenterococci from meat and Franz et al. (2001) in dairy
enterococci also reported a low incidence of VRE. Robredo
et al. (2000) found VRE in 27Æ2% of chicken products but
no VRE were detected in cooked pork or turkey products
from Spain. Peters et al. (2003) did not find any VRE in
foods of animal origin.
The E. faecium strains isolated from this study showed a
higher incidence of penicillin resistance when compared
with E. faecalis as previously reported by Murray (1990) and
Franz et al. (2001). Enterococcus faecium showed a higher
incidence of ampicillin resistance than those reported by
Quednau et al. (1998), Franz et al. (2001) and Peters et al.(2003). This may be a cause of concern due to monotherapy
with penicillin or ampicillin has been used for decades as a
general treatment for enterococci infections. However, none
of the strains presented high-level resistance to gentamicin,
an aminoglycoside that is generally combined with penicillin
or ampicillin to treat serious enterococcal infections, espe-
cially endocarditis. Franz et al. (2001) also reported a low
incidence of gentamicin resistance among E. faecium isolates
but a relatively high incidence among E. faecalis strains
(25Æ5%) and Peters et al. (2003) only found one E. faecalisstrain with a high-level resistance to gentamicin.
From our results, E. faecalis isolates showed a higher
incidence of antibiotic resistance than E. faecium and the
other enterococcal species except for ampicillin, ciprofloxa-
cin, nitrofurantoin and penicillin. Enterococcus faecalisshowed high prevalence of resistance towards chloramphen-
icol, erythromycin, quinupristin/dalfopristin, rifampicin
and tetracycline. Teuber et al. (1999) and Franz et al.(2001) reported lower resistance to tetracycline, erythromy-
cin and chloramphenicol among E. faecalis strains.Although no multi-resistant enterococcal strains could be
grouped in a single cluster group by RAPD typing, three
samples concentrated all these multi-resistant strains. This
suggests a high selective pressure exerted by the use of
antibiotics in food animals that may be an activation of
efficient gene transfer under stress conditions among the
enterococcal community in fermented sausages. In E.faecalis, plasmids encoding antibiotic resistances can be
mobilized within the enterococcal food community during
fermentation at high frequency, even without selective
pressure; the conjugal transfer seems more efficient in the
fermented sausage model than on agar plate or in cheese
(Cocconcelli et al. 2003).Although some enterococcal strains isolated from fer-
mented sausages were resistant to a high number of
BIODIVERSITY OF ENTEROCOCCI FROM FERMENTED SAUSAGES 1187
ª 2005 The Society for Applied Microbiology, Journal of Applied Microbiology, 98, 1177–1190, doi:10.1111/j.1365-2672.2005.02555.x

antibiotics, most of them were susceptible to clinically
relevant antibiotics, such as gentamicin and vancomycin, in
accordance with results obtained in other food enterococci
(Franz et al. 2001; Peters et al. 2003). Nevertheless, there
was a relatively high incidence towards ampicillin and
penicillin among E. faecium and E. durans strains not
previously reported in enterococci from food.
The enterococcal population from slightly fermented
sausages showed a considerable genetic diversity at species
and strain level, which was even higher in chorizo than in
fuet. The enterococci population can be quantitatively
important in slightly fermented sausages achieving counts
of 104–105 CFU g)1, and their use as bioprotective starter
cultures against Listeria monocytogenes and probiotics has
been reported (Coppola et al. 1988; Centeno et al. 1996;
Aymerich et al. 2003). In this study, the presence of
virulence factors and the higher incidence of antibiotic
resistance have been mainly associated with E. faecalisspecies, which are, moreover, recorded as responsible for
most nosocomial infections. Therefore, it is recommended
to use E. faecium instead of E. faecalis as starter culture in
fermented sausages but safety aspects of selected strains
must be considered. Typing of E. faecium isolates will ensure
the selection of safety strains as starter and bioprotective
cultures in fermented sausages. Although the potential risk
for human health associated to the presence of enterococci in
slightly fermented sausages is very low, the production of
these products without any virulent or antimicrobial resist-
ant enterococci is possible, therefore these characteristics
could be considered as a quality trait in this kind of
products.
ACKNOWLEDGEMENTS
This research was funded by the Spanish Inter-Ministerial
Commission of Science and Technology (CICYT ALI99-
0308 and AGL2004-05431/ALI). We thank Y. Beltran, A.
Claret and D. Tibau for technical assistance and the
Ministry of Science and Technology for Belen Martin’s
scholarship.
REFERENCES
Adams, M.R. (1999) Safety of industrial lactic acid bacteria. Journal of
Biotechnology 68, 171–178.
Angeletti, S., Lorino, G., Gherardi, G., Battistoni, F., De Cesaris, M.
and Dicuonzo, G.R. (2001) Routine molecular identification of
enterococci by gene-specific PCR and 16S ribosomal DNA sequen-
cing. Journal of Clinical Microbiology 39, 794–797.
Antonishyn, N.A., McDonald, R.R., Chan, E.L., Horsman, G.,
Woodmansee, C.E., Falk, P.C. and Mayhall, C.G. (2000) Evaluation
of fluorescence-based amplified fragment length polymorphism
analysis for molecular typing in hospital epidemiology: comparison
with pulsed-field gel electrophoresis for typing strains of vancomy-
cin-resistant Enterococcus faecium. Journal of Clinical Microbiology 38,
4058–4065.
Arthur, M., Reynolds, P. and Courvalin, P (1996) Glycopeptide
resistance in enterococci. Trends in Microbiology 4, 401–407.
Aymerich, M.T., Garriga, M., Ylla, J., Vallier, J., Monfort, J.M. and
Hugas, M. (2000) Application of enterocins as biopreservatives
against Listeria innocua in meat products. Journal of Food Protection
63, 721–726.
Aymerich, M.T., Martın, B., Garriga, M. and Hugas, M. (2003)
Microbial quality and direct PCR identification of lactic acid bacteria
and nonpathogenic staphylococci from artisanal low-acid sausages.
Applied and Environmental Microbiology 69, 4583–4594.
Baumgartner, A., Kueffer, M. and Rohner, P. (2001) Occurrence and
antibiotic resistance of enterococci in various ready-to-eat foods.
Archiv fur Lebensmittel Hygiene 52, 16–19.
CDC NNIS System (1998) National Nosocomial Infections Surveil-
lance (NNIS) report, data summary from October 1986–April 1998,
issued June 1998. American Journal of Infection Control 26, 522–533.
Centeno, J.A., Menendez, S. and Rodrıguez-Otero, J.L. (1996) Main
microbial flora present as natural starters in Cebreiro raw cow’s-milk
cheese (Northwest Spain). International Journal of Food Microbiology
33, 307–313.
Charteris, W.P., Kelly, P.M., Morelli, L. and Collins, J.K. (1998)
Antibiotic susceptibility of potentially probiotic Lactobacillus species.
Journal of Food Protection 61, 1636–1643.
Cheng, S., McCleskey, F.K., Gress, M.J., Petroziello, J.M., Liu, R.,
Namdari, H., Beninga, K., Salmen, A. et al. (1997) A PCR assay for
identification of Enterococcus faecium. Journal of Clinical Microbiology
35, 1248–1250.
Clewell, D.B. (1990) Movable genetic elements and antibiotic resist-
ance in enterococci. European Journal of Clinical Microbiology and
Infectious Diseases 9, 90–102.
Cocconcelli, P.S., Porro, D., Galandini, S. and Senini, L. (1995)
Development of RAPD protocol for typing of strains of lactic acid
bacteria and enterococci. Letters in Applied Microbiology 21, 376–379.
Cocconcelli, P.S., Cattivelli, D. and Gazzola, S. (2003) Gene transfer of
vancomycin and tetracycline resistances among Enterococcus faecalis
during cheese and sausage fermentations. International Journal of
Food Microbiology 88, 315–323.
Collins, M.D., Farrow, J.A.E. and Jones, D. (1986) Enterococcus
mundtii sp. Nov. International Journal of Systematic Bacteriology 36,
8–12.
Coppola, S., Parente, E., Dumontet, S. and La Peccerella, A. (1988)
The microflora of natural whey cultures utilized as starters in the
manufacture of Mozzarella cheese from water-buffalo milk. Lait 68,
295–310.
Descheemaeker, P., Lammens, C., Pot, B., Vandamme, P. and
Goossens, H. (1997) Evaluation of arbitrarily primed PCR analysis
and pulsed-field gel electrophoresis of large genomic DNA frag-
ments for identification of enterococci important in human medicine.
International Journal of Systematic Bacteriology 47, 555–561.
Devriese, L.A. and Pot, B. (1995) The genus Enterococcus. In The
Lactic Acid Bacteria ed. Wood, B.J.B. and Holzapfel, W.H. pp. 328–
367. London: Blackie Academic.
Devriese, L.A., Van De Kerckhove, A., Kilpper-Balz, R. and Schleifer,
K.H. (1987) Characterization and identification of Enterococcus
1188 B. MARTIN ET AL.
ª 2005 The Society for Applied Microbiology, Journal of Applied Microbiology, 98, 1177–1190, doi:10.1111/j.1365-2672.2005.02555.x

species isolated from animals. International Journal of Systematic
Bacteriology 37, 257–259.
Devriese, L.A., Pot, B., Van Damme, L., Kersters, K. and Hae-
sebrouck, F. (1995) Identification of Enterococcus species isolated
from foods of animal origin. International Journal of Food Microbio-
logy 26, 187–197.
Devriese, L.A., Pot, B., Kersters, K., Lauwers, S. and Haesebrouck, F.
(1996) Acidification of methyl-alpha-D-glucopyranoside: a useful
test to differentiate Enterococcus casseliflavus and Enterococcus gallin-
arum from Enterococcus faecium species group and from Enterococcus
faecalis. Journal of Clinical Microbiology 34, 2607–2608.
Devriese, L.A., Vancanneyt, M., Descheemaeker, P., Baele, M., Van
Landuyt, H.W., Gordts, B., Butaye, P., Swings, J. et al. (2002)
Differentiation and identification of Enterococcus durans, E. hirae and
E. villorum. Journal of Applied Microbiology 92, 821–827.
Domig, K.J., Mayer, H.K. and Kneifel, W. (2003) Methods used for
the isolation, enumeration, characterisation and identification of
Enterococcus spp. 2. Pheno- and genotypic criteria. International
Journal of Food Microbiology 88, 165–188.
Dunny, G.M. (1990) Genetic functions and cell-cell interactions in the
pheromone-inducible plasmid transfer system of Enterococcus fae-
calis. Molecular Microbiology 4, 689–696.
Dunny, G.M., Leonard, B.A. and Hedberg, P.J. (1995) Pheromone
inducible conjugation in Enterococcus faecalis: interbacterial and host-
parasitechemicalcommunication.JournalofBacteriology177,871–876.
Dupre, I., Zanetti, S., Schito, A.M., Fadda, S. and Sechi, L.A. (2003)
Incidence of virulence determinants in clinical Enterococcus faecium
and Enterococcus faecalis isolates collected in Sardinia (Italy). Journal
of Medical Microbiology 52, 491–498.
Dutka-Malen, S., Evers, S. and Courvalin, P. (1995) Detection of
glycopeptide resistance genotypes and identification to the species
level of clinically relevant enterococci by PCR. Journal of Clinical
Microbiology 33, 24–27.
Eaton, T.J. and Gasson, M.J. (2001) Molecular screening of Entero-
coccus virulence determinants and potential for genetic exchange
between food and medical isolates. Applied and Environmental
Microbiology 67, 1628–1635.
Farber, J.M., Cai, Y. and Ross, W.H. (1996) Predictive modeling of the
growth of Listeria monocytogenes in CO2 environments. International
Journal of Food Microbiology 32, 133–144.
Franz, C.M., Holzapfel, W. and Stiles, M.E. (1999) Enterococci at the
crossroads of food safety?. International Journal of Food Microbiology
47, 1–24.
Franz, C.M., Muscholl-Silberhorn, A.B., Yousif, N.M., Vancanneyt,
M., Swings, J. and Holzapfel, W.H. (2001) Incidence of virulence
factors and antibiotic resistance among enterococci isolated from
food. Applied and Environmental Microbiology 67, 4385–4389.
Giraffa, G. (2002) Enterococci from foods. FEMS Microbiology
Reviews 26, 163–171.
Giraffa, G., Olivari, A.M. and Neviani, E. (2000) Isolation of
vancomycin-resistant Enterococcus faecium from Italian cheeses. Food
Microbiology 39, 2354–2355.
Haas, W. and Gilmore, M.S. (1999) Molecular nature of a novel
bacterial toxin: the cytolysin of Enterococcus faecalis. Medical
Microbiology and Immunology 187, 183–190.
Heaton, M.P., Discotto, L.F., Pucci, M.J. and Handwerger, S. (1996)
Mobilization of vancomycin resistance by transposon-mediated
fusion of a VanA plasmid with an Enterococcus faecium sex
pheromone-response plasmid. Gene 171, 9–17.
Jett, B., Huycke, M.M. and Gilmore, M.S. (1994) Virulence of
enterococci. Clinical Microbiology Reviews 7, 462–478.
Ke, D., Picard, F.J., Martineau, F., Menard, C., Roy, P.H., Ouellette,
M. and Bergeron, M.G. (1999) Development of a PCR assay for
rapid detection of enterococci. Journal of Clinical Microbiology 37,
3497–3503.
Klein, G. (2003) Taxonomy, ecology and antibiotic resistance of
enterococci from food and the gastro-intestinal tract. International
Journal of Food Microbiology 88, 123–131.
Klein, G., Pack, A. and Reuter, G. (1998) Antibiotic resistance patterns
of enterococci and occurrence of vancomycin-resistant enterococci in
raw minced beef and pork in Germany. Applied and Environmental
Microbiology 64, 1825–1830.
Knudtson, L.M. and Hartman, P.A. (1993) Antibiotic resistance
among enterococcal isolates from environmental and clinical sources.
Journal of Food Protection 56, 486–492.
Leclercq, R. (1997) Enterococci acquire new kinds of resistance.
Clinical Infectious Diseases 24, S80–S84.
Leclercq, R., Derlot, M., Weber, J., Duval, J. and Courvalin, P. (1989)
Transferable vancomycin and teicoplanin resistance in Enterococcus
faecium. Antimicrobial Agents and Chemotherapy 33, 10–15.
Lowe, A.M., Lambert, P.A. and Smith, A.W. (1995) Cloning of an
Enterococcus faecalis endocarditis antigen: homology with adhesins
from some oral streptococci. Infection and Immunity 63, 703–
706.
Mannu, L., Paba, A., Daga, E., Comunian, R., Zanetti, S., Dupre, I.
and Sechi, L.A. (2003) Comparison of the incidence of virulence
determinants and antibiotic resistance between Enterococcus faecium
strains of dairy, animal and clinical origin. International Journal of
Food Microbiology 88, 291–304.
Morrison, D., Woodford, N. and Cookson, B. (1997) Enterococci as
emerging pathogens of humans. Journal of Applied Microbiology.
Symposium Supplement 83, 89S–99S.
Mundy, L.M., Sahm, D.F. and Gilmore, M.S. (2000) Relationship
between enterococcal virulence and antimicrobial resistance. Clinical
Microbiology Reviews 13, 513–522.
Murray, B.E. (1990) The life and times of the Enterococcus. Clinical
Microbiology Reviews 3, 46–65.
NCCLS (2002) Performance Standards for Antimicrobial Susceptibility
Testing; Twelfth Informational Supplement. NCCLS document
M100-S12.940. Wayne, PA: National Committee for Clinical
Laboratory Standards.
Niemi, R.M., Niemela, S.I., Bamfort, D.H., Hantula, J., Hyvarinen,
T., Forsten, T. and Raateland, A. (1993) Presumptive fecal
streptococci in environmental samples characterized by one-dimen-
sional sodium dodecyl sulphate-polyacrylamide gel electrophoresis.
Applied and Environmental Microbiology 59, 2190–2196.
Noble, W.C., Virani, Z. and Cree, R.G. (1992) Co-transfer of
vancomycin and other resistance genes from Enterococcus faecalis
NCTC 12201 to Staphylococcus aureus. FEMS Microbiology Letters
72, 195–198.
Peters, J., Mac, K., Wichmann-Schauer, H., Klein, G. and Ellerbroek,
L. (2003) Species distribution and antibiotic resistance patterns of
enterococci isolated from food of animal origin in Germany.
International Journal of Food Microbiology 88, 311–314.
BIODIVERSITY OF ENTEROCOCCI FROM FERMENTED SAUSAGES 1189
ª 2005 The Society for Applied Microbiology, Journal of Applied Microbiology, 98, 1177–1190, doi:10.1111/j.1365-2672.2005.02555.x

Poyart, C., Quesnes, G. and Trieu-Cuot, P. (2000) Sequencing the
gene encoding manganese-dependent superoxide dismutase for rapid
species identification of enterococci. Journal of Clinical Microbiology
38, 415–418.
Quednau, M., Ahrne, S., Petersson, A.C. and Molin, G. (1998)
Antibiotic-resistant strains of Enterococcus isolated from Swedish and
Danish retailed chicken and pork. Journal of Applied Microbiology 84,
1163–1170.
Reuter, G. (1995) Culture media for enterococci and group D-
streptococci. In Culture Media for Food Microbiology Progress in
Industrial Microbiology ed. Corry, J.E.L., Curtis, G.D.W. and Baird,
R.M. pp. 51–61. Philadelphia, PA: Elsevier Health Sciences.
Robredo, B., Singh, K.V., Baquero, F., Murray, B.E. and Torres, C.
(2000) Vancomycin-resistant enterococci isolated from animals and
food. International Journal of Food Microbiology 54, 197–204.
Semedo, T., Almeida, M., Silva Lopes, M.F., Figueiredo Marques,
J.J., Barreto Crespo, M.T. and Tenreiro, R. (2003) Virulence factors
in food, clinical and reference enterococci: a common trait in the
genus? Systematic and Applied Microbiology 26, 13–22.
Shankar, V., Baghdayan, A.S., Huycke, M.M., Kindahl, G. and
Gilmore, M.S. (1999) Infection-derived Enterococcus faecalis strains
are enriched in esp, a gene encoding a novel surface protein. Infection
and Immunity 67, 193–200.
Singh, K.V., Coque, T.M., Weinstock, G.M. and Murray, B.E. (1998)
In vivo testing of an Enterococcus faecalis efaA mutant and use of efaA
homologs for species identification. FEMS Immunology and Medical
Microbiology 21, 323–331.
Swenson, J.M., Bertha, C.H. and Thornsberry, C. (1989) Problems
with the disk diffusion test for detection of vancomycin resistance in
enterococci. Journal of Clinical Microbiology 27, 2140–2142.
Temmerman, R., Pot, B., Huys, G. and Swings, J. (2003) Identification
and antibiotic susceptibility of bacterial isolates from probiotic
products. International Journal of Food Microbiology 81, 1–10.
Teuber, M., Meile, L. and Schwarz, F. (1999) Acquired antibiotic
resistance in lactic acid bacteria from food. Antonie van Leeuwenhoek
76, 115–137.
Vancanneyt, M., Lombardi, A., Andrighetto, C., Knijff, E., Torriani,
S., Bjorkroth, K.J., Franz, C.M.A.P., Moreno, M.R.F. et al. (2002)
Intraspecies genomic groups in Enterococcus faecium and their
correlation with origin and pathogenicity. Applied and Environmental
Microbiology 68, 1381–1391.
Wegener, H.C., Aarestrup, F.M., Jensen, L.B., Hammerum, A.M.
and Bager, F. (1999) Use of antimicrobial growth promoters in
food animals and Enterococcus faecium resistance to therapeutic
antimicrobial drugs in Europe. Emerging Infectious Diseases 5, 329–
335.
1190 B. MARTIN ET AL.
ª 2005 The Society for Applied Microbiology, Journal of Applied Microbiology, 98, 1177–1190, doi:10.1111/j.1365-2672.2005.02555.x






























