



PRIMARY RESEARCH PAPER
Ecological diatom guilds are useful but not sensitive enoughas indicators of extremely changing water regimes
Viktoria B-Beres • Peter Torok •
Zsuzsanna Kokai • Enik}o T. Krasznai •
Bela Tothmeresz • Istvan Bacsi
Received: 12 January 2014 / Revised: 25 May 2014 / Accepted: 26 May 2014
� Springer International Publishing Switzerland 2014
Abstract Classification of taxa into ecological
guilds is based on the relation of respective taxa to
nutrient enrichment and their resistance to physical
disturbance. We hypothesized that ecological guild’s
proportion and their taxa composition were strongly
effected both by extremely changing water regime and
nutrient contents. Diatom composition, guild dynam-
ics and the diatom-based ecological status assessment
index were studied in the Sebes-Koros River (South-
East Hungary) in a year with extremely changing
water regimes. There were highly pronounced changes
in species composition during the whole vegetation
period including the formation of running and stand-
ing water segments in autumn. While the proportions
of ecological guilds showed no significant correlations
with the studied environmental parameters, they were
more balanced in high water discharge period than in
the low water discharge period. Taxa compositions of
segments were mainly determined by the preferences
and strategies of a respective species and/or genera,
regardless to their guild affiliation. These results point
out that ecological guild characterisation should be
refined using ecological knowledge at the subgenus
level. We suggest to establish several subdivisions
within the guilds to consider the differences in life
strategies (CSR model) and life forms, and to imple-
ment the accumulated knowledge of nutrient prefer-
ences/indication of a respective taxa.
Keywords Diatom species composition � Diversity �Ecological guilds �Medium-sized river �Water quality
Introduction
Nowadays, diatoms are routinely used in environmen-
tal status assessment, because of their importance in
food webs, biochemical linkages and due to their
sensitivity for physical, chemical and biological
disturbances (Stenger-Kovacs et al., 2007; Bolla
et al., 2010; Chetelat et al., 2010; Kireta et al.,
2012). Diatom-based environmental status assessment
requires high taxonomical knowledge. Due to the
expansion of molecular methods and electron
Handling editor: Judit Padisak
V. B-Beres (&) � Z. Kokai � E. T. Krasznai
Environmental Protection and Nature Conservation
Authority, Trans-Tisza Region, Hatvan u. 16,
Debrecen 4025, Hungary
e-mail: [email protected]
P. Torok
MTA-DE Biodiversity and Ecosystem Services Research
Group, Egyetem ter 1, Debrecen 4032, Hungary
B. Tothmeresz
Department of Ecology, University of Debrecen, Egyetem
ter 1, Debrecen 4032, Hungary
I. Bacsi
Department of Hydrobiology, University of Debrecen,
Egyetem ter 1, Debrecen 4032, Hungary
123
Hydrobiologia
DOI 10.1007/s10750-014-1929-y

microscopy in diatom research, several species com-
plexes were divided into several new species (Potap-
ova & Hamilton, 2007; Novais et al., 2009). This
resulted that exact identification to species by light
microscopy techniques (1,000–1,600 fold magnifica-
tion, which is widely used in practice) are not feasable
for each taxa (e.g. Gomphonema parvulum complex,
Achnanthidium minutissimum complex and Encyon-
opsis microcephala complex—Morales et al., 2001;
Rimet & Bouchez, 2012a). Rimet & Bouchez (2012b)
argue that there is only a little loss of ecological
information when the accuracy decreased from spe-
cies to higher taxonomical group. Thus, classifying
taxa into respective ecological functional groups (i.e.
guilds) became more popular in biomonitoring appli-
cations and also in ecological studies (Padisak et al.,
2009, Rimet & Bouchez, 2012a, b). Passy (2007)
ranked the diatom taxa into three guilds (low-profile,
high-profile and motile guilds), which was modified
by Rimet & Bouchez (2012b) as planktic guild.
Changes in flow conditions were found to induce
responses in both composition and physical structure
of benthic diatom communities (Lamb & Lowe, 1987;
Poff et al., 1990; Molloy, 1992; Biggs et al., 1998;
Passy, 2002). Classification of taxa into ecological
guilds is based on the relation of taxa to nutrient
enrichment and on taxa’s resistance to physical
disturbance (e.g. in form of different flow conditions;
Passy, 2007). Studying the relationship between
diatom guilds and environmental factors began in the
last few years (e.g. Berthon et al., 2011; Rimet &
Bouchez, 2012a; Stenger-Kovacs et al., 2013).
Because of the easy use of guilds, it might provide a
great opportunity to apply them in the environmental
status assessment. However, it is still not entirely
clear, how reliable are the guild level ecological
analyses of benthic diatoms in the identification of
relatively rapid changes in the environmental condi-
tions and flow conditions among them as physical
disturbance.
In the year of 2012, the summer and autumn could
be considered as extremely dry in Hungary based on
the data of the Hungarian Meteorological Service and
the General Directorate of Water Management.
Because of the dry weather, the water discharge of
some lowland rivers—like the Sebes-Koros river—
decreased drastically (max. 90.25 m3 s-1 in May;
min. 1.22 m3 s-1 in September). Due to very low
water discharge, segments characterized by different
water flow conditions were formed in Autumn. These
segments were continuously connected with each
other, and the main differences between them occurred
in the flow conditions: an extremely slowed down
water flow was typical in the so-called standing water
segment, which were formed between the riverbank
and the main stream of the riverbed because of its
morphology.
In this work, the diatom species and guild dynamics
and its effects on the changes of diatom-based
ecological status assessment index (in the followings
abbreviated as IPSITI) were studied from April to
November (during the whole vegetation period). We
hypothesized the followings:
(i) Relative abundance of low-profile guild will be
high in the Spring and early Summer (period
with high water discharge), while the decreasing
water discharge from July will decrease the
proportion of this guild.
(ii) The proportion of high-profile guild will be low
in Spring and early Summer (period with high
water discharge), while the decreasing water
discharge from July will favour this guild.
(iii) The proportion of motile guild will be the lowest
in months with the highest water discharge.
(iv) We assumed that running and standing water
segments can be characterized by different guild
ratios: low-profile guild will be present in higher
relative abundance in running water segments,
while high-profile guild and motile guild will be
present in higher relative abundance in the
standing water segments.
Materials and methods
Sampling setup and environmental parameters
Samples were collected at Korosszakal, on the river-
bed of Sebes-Koros (South- Eastern Hungary; N:
47�00041.9500 E: 21�36025.6800; medium-sized river).
For monthly analyses, benthic diatom samples were
collected from April to November, 2012. Conductiv-
ity, pH, dissolved oxygen and water temperature were
measured in situ in the field with a portable-multipa-
rameter digital metre (Multi 350i-WTW, Germany).
Water samples were kept at 4 �C in cooler bag during
transportation to the laboratory for total nitrogen (TN)
Hydrobiologia
123

and total phosphorus (TP) measurements. TN was
calculated by summarising the amount of different
nitrogen forms. Free ammonium concentration and
ammonium concentration disengaged from organic
nitrogen compounds were measured by a manual
spectrophotometric method based on the MSZ ISO
7150-1:1992 (1992) Hungarian Standard. This stan-
dard is totally equivalent in technical content and fully
corresponds in presentation to the International Stan-
dard ISO 7150-1:1984. Measurement of nitrate con-
centration was based on colorimetry by titration of
salicylic acid (MSZ 1484-13:2009, 2009). Colorime-
try was also used to measure nitrite concentration
applying sulphonic acid and aminonaphthalene
reagents (MSZ 1484-13:2009, 2009). Total phospho-
rous was determined by the acid molybdate method
(MSZ EN ISO 6878:2004, 2004). The same physical
and chemical parameters were measured by the same
methods for analyses of the formed segments from
September to November (see below). Water discharge
data were kindly provided by the National Water
Authority, while rainfall data were provided by the
Hungarian Meteorological Service.
The amount of rainfall in 2012 (427 mm) was
25% lower than the hundred-year average (569 mm),
which can be considered as extremely dry according
to the Hungarian Meteorological Service. This
amount is also lower, than in 2011 (441 mm), 2010
(845 mm) and 2009 (485 mm). What was more
important is the distribution of annual rainfall which
was unequial: the quantity of rainfall in the second
part of vegetation period was less than 36% of the
2012’ year amount. This dry weather caused a very
low water discharge from September to November,
resulting that two segments were formed in connec-
tion with each other, but with different water flow
conditions (called ‘running water’ and ‘standing
water’ segment in the followings), because of the
riverbed morphology. This phenomenon was not
observed in the previous years in the Sebes-Koros
at Korosszakal by the colleagues of the Environmen-
tal Protection and the Nature Conservation Authority.
There were no differences in the measured physical
and chemical parameters, only in the water flow
conditions between the formed segments.
Benthic diatom samples were also collected from
these formed segments from September to November.
Sampling was done in these segments in triplicate.
There were no differences in the substrate and depth of
sampling between the standing and running water
segments.
Sample collection and preparation
Five cobbles were used as sampled substrates all
times. Diatoms were removed from the cobbles by
tooth brushing. The sampling area of cobbles was
approximately 10 cm2. Sampling was done in tripli-
cate in all cases. Samples were fixed in Lugol’s
solution on the field. Diatom valves were prepared by
the hot hydrogen-peroxide method. Naphrax synthetic
resin was used for embedding (MSZ EN 13946:2003,
2003). Leica DMRB research microscope and
1,000–1,600-fold magnification were used for identi-
fication of diatom taxa. At least 400 valves were
counted (MSZ EN 14407:2004, 2004); for diatom
identification, Krammer & Lange-Bertalot (1997a, b),
Krammer & Lange-Bertalot (2004a, b), and Potapova
& Hamilton (2007) were used. Identification of
Achnanthidium minutissimum (Kutzing) Czarnecki
and A. eutrophilum (Lange-Bertalot) Lange-Bertalot
belonging to A. minutissimum species complex was
based on Potapova & Hamilton (2007).
Data processing and analyses
Classifying the diatom taxa into guilds was imple-
mented according to Passy (2007) and Berthon et al.
(2011): low-profile, high-profile and motile guild.
According to Rimet & Bouchez (2012b), planktic
guild was also considered. Multimetric diatom index
developed for streams, and small- and medium-sized
rivers (IPSITI) was calculated to characterize the
water quality of the river. The IPSITI index is
calculated by averaging of IPS, SI and TI indices.
The IPS, SI and TI indices were calculated by the
OMNIDIA version 5.2 software package (Lecointe
et al., 1993). For details about IPS, SI and TI, see
Lecointe et al. (1993); for further data about IPSITI,
see Varbıro et al. (2012). To verify main tendencies
between environmental factors and species composi-
tion and to select crucial factors, principal component
analysis (PCA) was calculated (Leps & Smilauer,
2003; Stenger-Kovacs et al., 2013). Those environ-
mental factors were included in the PCA, which
showed higher differences between months than 10%
from the average calculated for the whole study
period. To analyse diatom assemblages in relation of
Hydrobiologia
123
verified environmental factors, a canonical correspon-
dence analysis (CCA) was calculated (Ter Braak &
Smilauer, 2002). Kruskal–Wallis correlations were
calculated for significance of guilds’ proportion (Zar,
1999). Relations of species composition of diatom
assemblages and water flowing conditions were dis-
played by a detrended correspondence analysis (DCA)
based on relative species abundances.
Results
Changes in environmental variables
Altogether, seven environmental factors (TN, TP,
water discharge, conductivity, time, temperature and
dissolved oxygen- DO, sample N-1) were included to
the Principal Component Analysis. The first three axes
of PCA explained 85% of variance. The PCA revealed
that the factors with the highest correlation were with
the first axis Time (0.6432) and TN (0.5954), with the
second axis Water discharge (-0.5774), and with the
third axis TP (-0.6136) and Conductivity (0.5921).
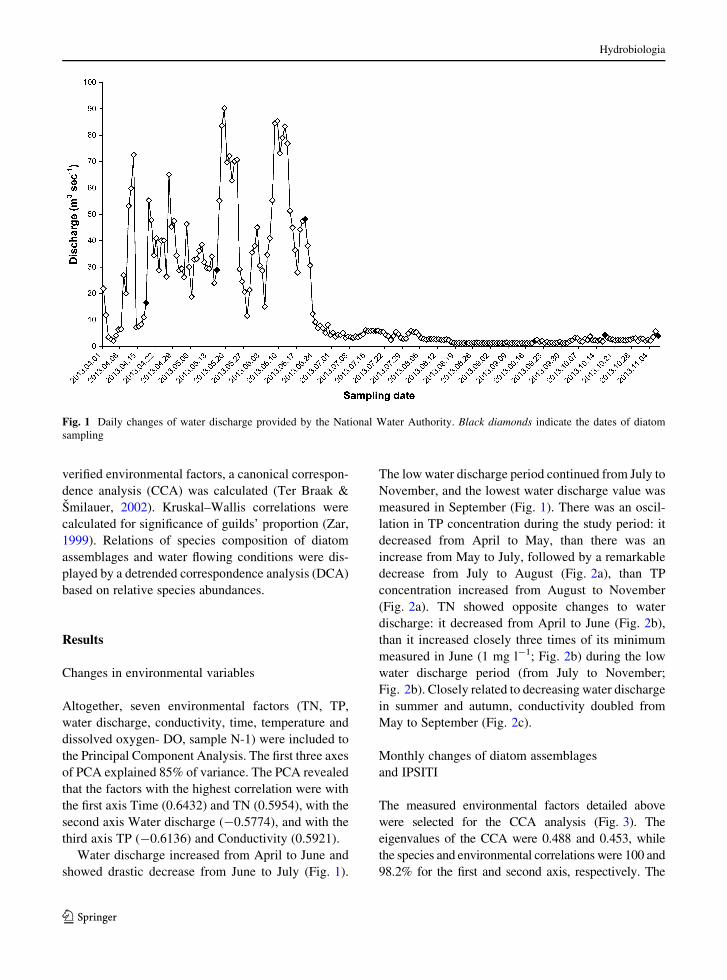
Water discharge increased from April to June and
showed drastic decrease from June to July (Fig. 1).
The low water discharge period continued from July to
November, and the lowest water discharge value was
measured in September (Fig. 1). There was an oscil-
lation in TP concentration during the study period: it
decreased from April to May, than there was an
increase from May to July, followed by a remarkable
decrease from July to August (Fig. 2a), than TP
concentration increased from August to November
(Fig. 2a). TN showed opposite changes to water
discharge: it decreased from April to June (Fig. 2b),
than it increased closely three times of its minimum
measured in June (1 mg l-1; Fig. 2b) during the low
water discharge period (from July to November;
Fig. 2b). Closely related to decreasing water discharge
in summer and autumn, conductivity doubled from
May to September (Fig. 2c).
Monthly changes of diatom assemblages
and IPSITI
The measured environmental factors detailed above
were selected for the CCA analysis (Fig. 3). The
eigenvalues of the CCA were 0.488 and 0.453, while
the species and environmental correlations were 100 and
98.2% for the first and second axis, respectively. The
Fig. 1 Daily changes of water discharge provided by the National Water Authority. Black diamonds indicate the dates of diatom
sampling
Hydrobiologia
123
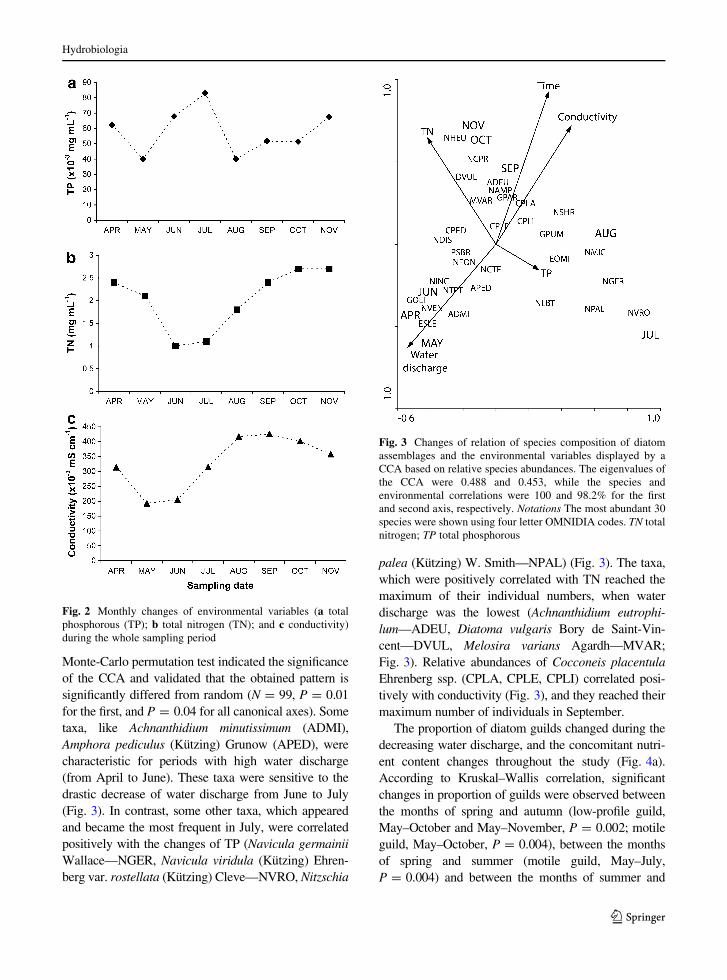
Monte-Carlo permutation test indicated the significance
of the CCA and validated that the obtained pattern is
significantly differed from random (N = 99, P = 0.01
for the first, and P = 0.04 for all canonical axes). Some
taxa, like Achnanthidium minutissimum (ADMI),
Amphora pediculus (Kutzing) Grunow (APED), were
characteristic for periods with high water discharge
(from April to June). These taxa were sensitive to the
drastic decrease of water discharge from June to July
(Fig. 3). In contrast, some other taxa, which appeared
and became the most frequent in July, were correlated
positively with the changes of TP (Navicula germainii
Wallace—NGER, Navicula viridula (Kutzing) Ehren-
berg var. rostellata (Kutzing) Cleve—NVRO, Nitzschia
palea (Kutzing) W. Smith—NPAL) (Fig. 3). The taxa,
which were positively correlated with TN reached the
maximum of their individual numbers, when water
discharge was the lowest (Achnanthidium eutrophi-
lum—ADEU, Diatoma vulgaris Bory de Saint-Vin-
cent—DVUL, Melosira varians Agardh—MVAR;
Fig. 3). Relative abundances of Cocconeis placentula
Ehrenberg ssp. (CPLA, CPLE, CPLI) correlated posi-
tively with conductivity (Fig. 3), and they reached their
maximum number of individuals in September.
The proportion of diatom guilds changed during the
decreasing water discharge, and the concomitant nutri-
ent content changes throughout the study (Fig. 4a).
According to Kruskal–Wallis correlation, significant
changes in proportion of guilds were observed between
the months of spring and autumn (low-profile guild,
May–October and May–November, P = 0.002; motile
guild, May–October, P = 0.004), between the months
of spring and summer (motile guild, May–July,
P = 0.004) and between the months of summer and
Fig. 2 Monthly changes of environmental variables (a total
phosphorous (TP); b total nitrogen (TN); and c conductivity)
during the whole sampling period
Fig. 3 Changes of relation of species composition of diatom
assemblages and the environmental variables displayed by a
CCA based on relative species abundances. The eigenvalues of
the CCA were 0.488 and 0.453, while the species and
environmental correlations were 100 and 98.2% for the first
and second axis, respectively. Notations The most abundant 30
species were shown using four letter OMNIDIA codes. TN total
nitrogen; TP total phosphorous
Hydrobiologia
123
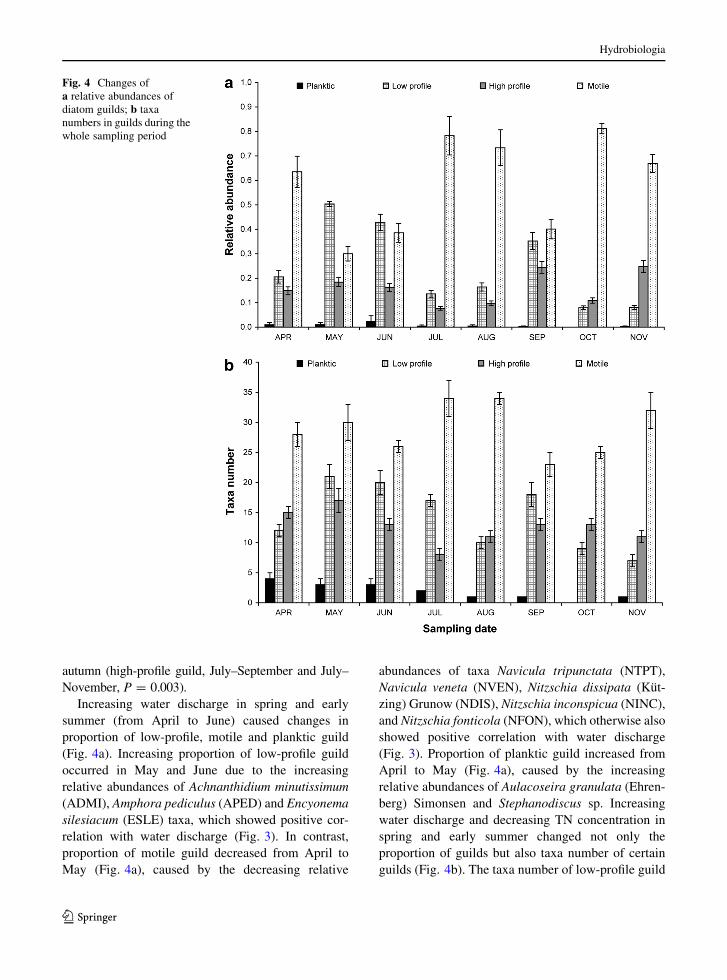
autumn (high-profile guild, July–September and July–
November, P = 0.003).
Increasing water discharge in spring and early
summer (from April to June) caused changes in
proportion of low-profile, motile and planktic guild
(Fig. 4a). Increasing proportion of low-profile guild
occurred in May and June due to the increasing
relative abundances of Achnanthidium minutissimum
(ADMI), Amphora pediculus (APED) and Encyonema
silesiacum (ESLE) taxa, which showed positive cor-
relation with water discharge (Fig. 3). In contrast,
proportion of motile guild decreased from April to
May (Fig. 4a), caused by the decreasing relative
abundances of taxa Navicula tripunctata (NTPT),
Navicula veneta (NVEN), Nitzschia dissipata (Kut-
zing) Grunow (NDIS), Nitzschia inconspicua (NINC),
and Nitzschia fonticola (NFON), which otherwise also
showed positive correlation with water discharge
(Fig. 3). Proportion of planktic guild increased from
April to May (Fig. 4a), caused by the increasing
relative abundances of Aulacoseira granulata (Ehren-
berg) Simonsen and Stephanodiscus sp. Increasing
water discharge and decreasing TN concentration in
spring and early summer changed not only the
proportion of guilds but also taxa number of certain
guilds (Fig. 4b). The taxa number of low-profile guild
Fig. 4 Changes of
a relative abundances of
diatom guilds; b taxa
numbers in guilds during the
whole sampling period
Hydrobiologia
123

changed the most conspicuous; it increased with 75%
from April to May, and rare species like Reimeria
uniseriata Sala, Guerrero & Ferrario, Rhoicosphenia
abbreviata (Agardh) Lange-Bertalot and Encyonopsis
microcephala (Grunow) Krammer appeared. Increase
of the proportion of planktic guild was due to the
increase of the already present species, not due to the
increase of taxa number (Fig. 4b).
Drastic decrease in water discharge (from a mean of
51.3 m3 s-1 to a minimum of 7.7 m3s-1; Fig. 1) and
the increase in total nitrogen and total phosphorus
(Fig. 2a, b) favoured taxa showing positive correlation
with TN (Fig. 3, see above), some of them (NAMP,
NGER, NVIR, NVRO, NPAL) belonging to motile
guild. Not only the number of taxa increased in motile
guild (Fig. 4b), but also some of the motile guild taxa
became characteristic due to their increasing individ-
ual numbers. The mean proportion and the taxa
number of low-profile guild decreased from May to
July (Fig. 4). However, this decrease of taxa number
continued in August as well (Fig 4b), but the relative
abundances of those low-profile guild taxa, which
significantly correlated with TN and conductivity
(ADEU, Cocconeis placentula —CPLA ssp., Fig. 3),
increased multiple in August (2.6- and 2.5-fold for
ADEU and CPLA spp., respectively), causing increas-
ing proportion of low-profile guild in these months
(Fig. 4a).
Water discharge was very low (Fig. 1), and TN
concentration reached almost its maximum (Fig. 2b)
in September. Proportions of both low-profile and
high-profile guilds were higher than in August
(Fig. 4a). On one hand, it was due to the increase of
some species’ relative abundances (2.4-fold for Cocc-
oneis species and 1.8-fold for Achnanthidium eutro-
philum in low-profile guild; 7.6-fold for Gomphonema
parvulum and 17.3-fold for Diatoma vulgaris in high-
profile guild) and on the other hand, due to the increase
of taxa number (Fig 4b). Meanwhile, proportion of
motile guild decreased from 0.73 to 0.40 (Fig. 4a) to
September, because of the decrease of relative abun-
dances of some motile guild taxa (Eolimna minima
(Grunow) Lange-Bertalot—EOMI, Nitzschia micro-
cephala Grunow—NMIC, Navicula schroeteri Mei-
ster—NSHR). Proportion of high-profile guild
decreased from September to October (Fig. 4a), due
to the decreasing relative abundances of Gomphonema
parvulum and Fragilaria species. Drastic decrease of
proportion of low-profile guild occurred due to
decreasing relative abundances of Cocconeis species
and the drastic decrease of taxa number (Fig. 4). In
contrast to decreasing proportion of high and low-
profile guilds, proportion of motile guild increased
from September to October due to increasing taxa
number in motile guild (Fig. 4) and increasing relative
abundance of Navicula capitatoradiata Germain. The
proportion of high-profile guild increased again from
October to November (Fig. 4a), mainly because of the
increasing relative abundance of Melosira varians and
Diatoma vulgaris and the increasing taxa number
(28%) (Fig. 4b).
The tendency changes of both the Berger–Parker
diversity and the number of taxa were the same in the
high water period (May and June; Fig. 5a). There were
many rare taxa detected (e. g. Reimeria uniseriata,
Rhoicosphenia abbreviata, Encyonopsis microcepha-
la, Diatoma ehrenbergii Kutzing, Pseudostaurosira
parasitica (W. Smith) Morales, Pseudostaurosira
parasitica var. subconstricta (Grunow) Morales,
Navicula radiosa Kutzing, Nitzschia dissipata, etc.),
which appeared only in these months. Later, there
were no significant correlation between taxa number
and BP diversity (Fig. 5). This is probably due to the
drastic changes in physical–chemical parameters
(from June to July/August; Figs. 1, 2), which led to
the reorganization of assemblages: some Navicula and
Nitzschia species became dominant with extremely
high abundance (e.g. Navicula viridula var. rostellata
31.5%; Nitzschia microcephala 20.5%) in July and/or
in August.
The index IPSITI reached its maximum in May (the
month with high water discharge and low TN
concentration—Fig. 5b.). Some taxa, which corre-
lated positively with water discharge (Achnanthidium
minutissimum and Encyonema silesiacum; Fig. 3),
were most abundant in this period. In July and August,
both the value of IPSITI and water discharge
decreased in correlation with increasing TN concen-
tration. In this period, taxa such as Nitzschia micro-
cephala, Diadesmis confervacea Kutzing, Navicula
viridula var. rostellata and Eolimna minima were
dominant.
Composition of diatom assemblages in running
and standing water segments
The species composition in the standing water
segments in September was clearly separated from
Hydrobiologia
123
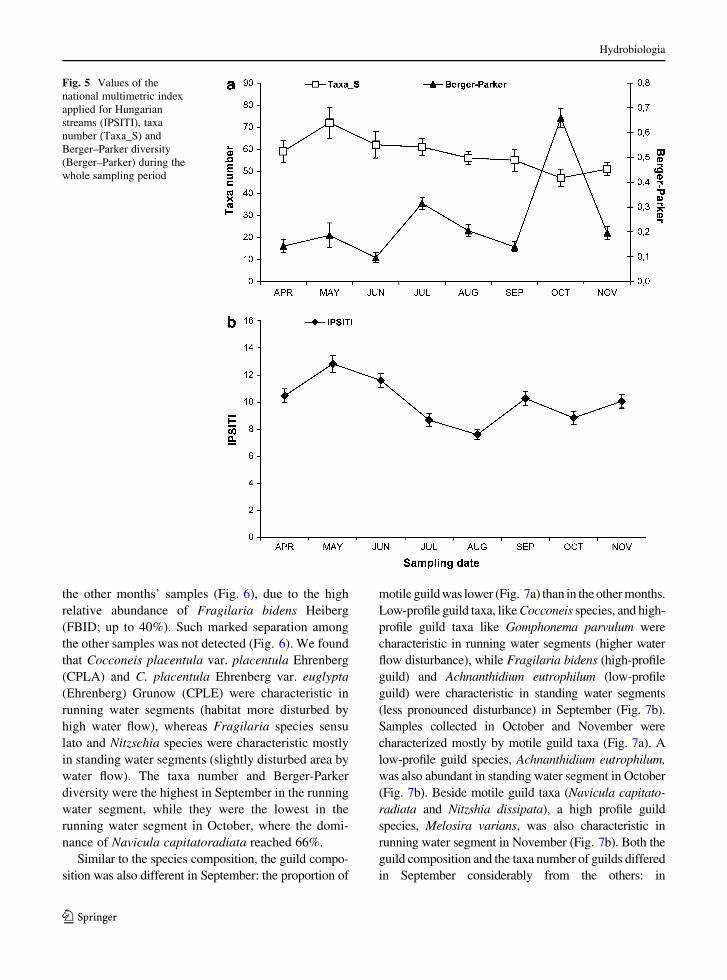
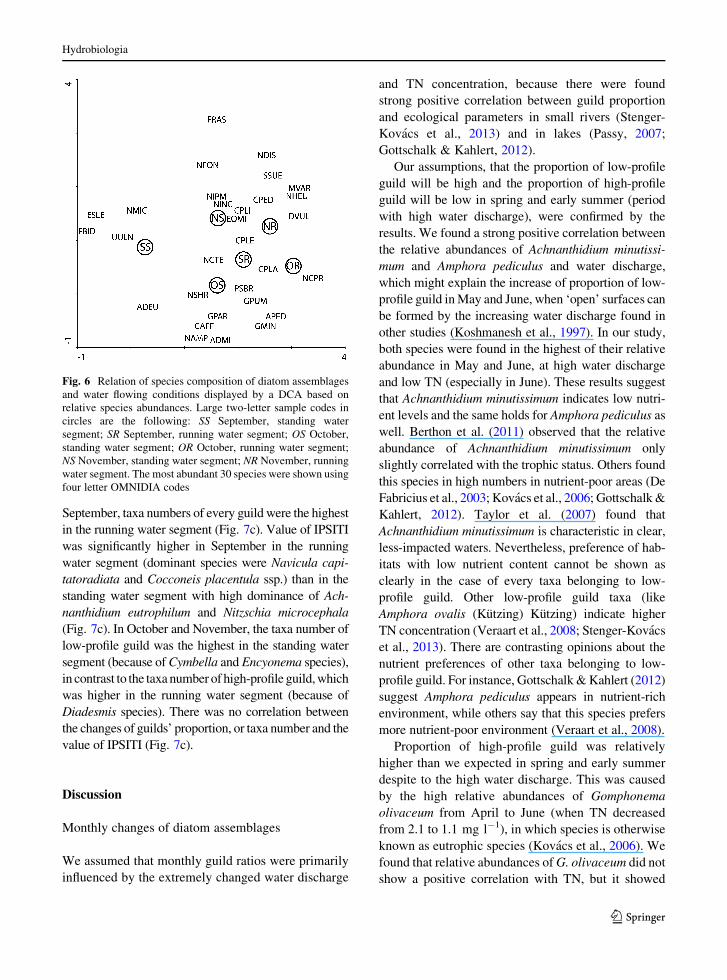
the other months’ samples (Fig. 6), due to the high
relative abundance of Fragilaria bidens Heiberg
(FBID; up to 40%). Such marked separation among
the other samples was not detected (Fig. 6). We found
that Cocconeis placentula var. placentula Ehrenberg
(CPLA) and C. placentula Ehrenberg var. euglypta
(Ehrenberg) Grunow (CPLE) were characteristic in
running water segments (habitat more disturbed by
high water flow), whereas Fragilaria species sensu
lato and Nitzschia species were characteristic mostly
in standing water segments (slightly disturbed area by
water flow). The taxa number and Berger-Parker
diversity were the highest in September in the running
water segment, while they were the lowest in the
running water segment in October, where the domi-
nance of Navicula capitatoradiata reached 66%.
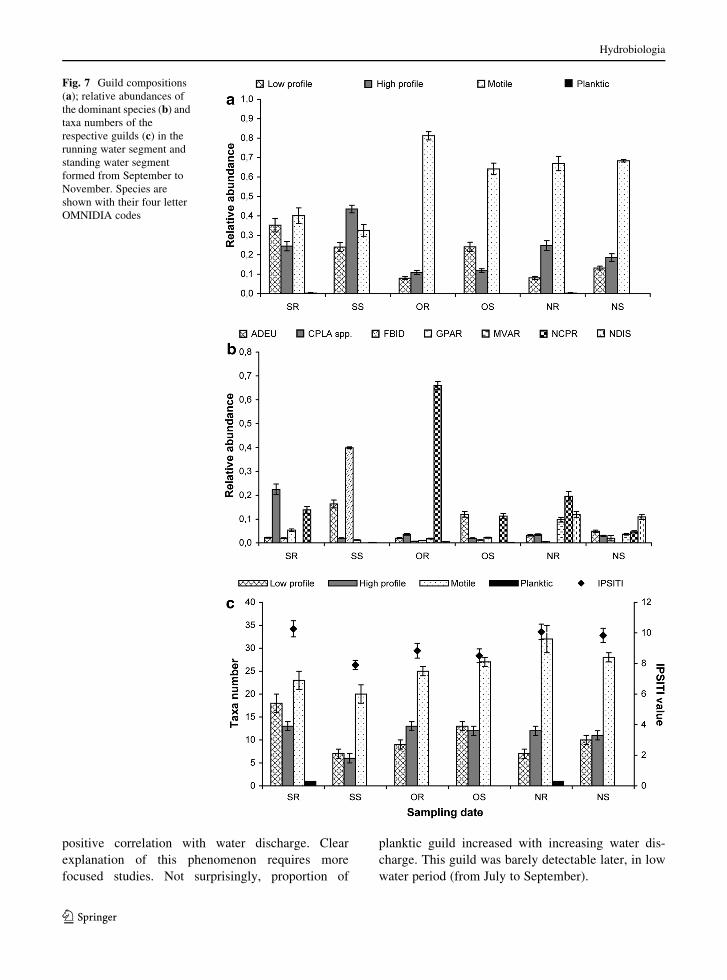
Similar to the species composition, the guild compo-
sition was also different in September: the proportion of
motile guild was lower (Fig. 7a) than in the other months.
Low-profile guild taxa, like Cocconeis species, and high-
profile guild taxa like Gomphonema parvulum were
characteristic in running water segments (higher water
flow disturbance), while Fragilaria bidens (high-profile
guild) and Achnanthidium eutrophilum (low-profile
guild) were characteristic in standing water segments
(less pronounced disturbance) in September (Fig. 7b).
Samples collected in October and November were
characterized mostly by motile guild taxa (Fig. 7a). A
low-profile guild species, Achnanthidium eutrophilum,
was also abundant in standing water segment in October
(Fig. 7b). Beside motile guild taxa (Navicula capitato-
radiata and Nitzshia dissipata), a high profile guild
species, Melosira varians, was also characteristic in
running water segment in November (Fig. 7b). Both the
guild composition and the taxa number of guilds differed
in September considerably from the others: in
Fig. 5 Values of the
national multimetric index
applied for Hungarian
streams (IPSITI), taxa
number (Taxa_S) and
Berger–Parker diversity
(Berger–Parker) during the
whole sampling period
Hydrobiologia
123
September, taxa numbers of every guild were the highest
in the running water segment (Fig. 7c). Value of IPSITI
was significantly higher in September in the running
water segment (dominant species were Navicula capi-
tatoradiata and Cocconeis placentula ssp.) than in the
standing water segment with high dominance of Ach-
nanthidium eutrophilum and Nitzschia microcephala
(Fig. 7c). In October and November, the taxa number of
low-profile guild was the highest in the standing water
segment (because of Cymbella and Encyonema species),
in contrast to the taxa number of high-profile guild, which
was higher in the running water segment (because of
Diadesmis species). There was no correlation between
the changes of guilds’ proportion, or taxa number and the
value of IPSITI (Fig. 7c).
Discussion
Monthly changes of diatom assemblages
We assumed that monthly guild ratios were primarily
influenced by the extremely changed water discharge
and TN concentration, because there were found
strong positive correlation between guild proportion
and ecological parameters in small rivers (Stenger-
Kovacs et al., 2013) and in lakes (Passy, 2007;
Gottschalk & Kahlert, 2012).
Our assumptions, that the proportion of low-profile
guild will be high and the proportion of high-profile
guild will be low in spring and early summer (period
with high water discharge), were confirmed by the
results. We found a strong positive correlation between
the relative abundances of Achnanthidium minutissi-
mum and Amphora pediculus and water discharge,
which might explain the increase of proportion of low-
profile guild in May and June, when ‘open’ surfaces can
be formed by the increasing water discharge found in
other studies (Koshmanesh et al., 1997). In our study,
both species were found in the highest of their relative
abundance in May and June, at high water discharge
and low TN (especially in June). These results suggest
that Achnanthidium minutissimum indicates low nutri-
ent levels and the same holds for Amphora pediculus as
well. Berthon et al. (2011) observed that the relative
abundance of Achnanthidium minutissimum only
slightly correlated with the trophic status. Others found
this species in high numbers in nutrient-poor areas (De
Fabricius et al., 2003; Kovacs et al., 2006; Gottschalk &
Kahlert, 2012). Taylor et al. (2007) found that
Achnanthidium minutissimum is characteristic in clear,
less-impacted waters. Nevertheless, preference of hab-
itats with low nutrient content cannot be shown as
clearly in the case of every taxa belonging to low-
profile guild. Other low-profile guild taxa (like
Amphora ovalis (Kutzing) Kutzing) indicate higher
TN concentration (Veraart et al., 2008; Stenger-Kovacs
et al., 2013). There are contrasting opinions about the
nutrient preferences of other taxa belonging to low-
profile guild. For instance, Gottschalk & Kahlert (2012)
suggest Amphora pediculus appears in nutrient-rich
environment, while others say that this species prefers
more nutrient-poor environment (Veraart et al., 2008).
Proportion of high-profile guild was relatively
higher than we expected in spring and early summer
despite to the high water discharge. This was caused
by the high relative abundances of Gomphonema
olivaceum from April to June (when TN decreased
from 2.1 to 1.1 mg l-1), in which species is otherwise
known as eutrophic species (Kovacs et al., 2006). We
found that relative abundances of G. olivaceum did not
show a positive correlation with TN, but it showed
Fig. 6 Relation of species composition of diatom assemblages
and water flowing conditions displayed by a DCA based on
relative species abundances. Large two-letter sample codes in
circles are the following: SS September, standing water
segment; SR September, running water segment; OS October,
standing water segment; OR October, running water segment;
NS November, standing water segment; NR November, running
water segment. The most abundant 30 species were shown using
four letter OMNIDIA codes
Hydrobiologia
123
positive correlation with water discharge. Clear
explanation of this phenomenon requires more
focused studies. Not surprisingly, proportion of
planktic guild increased with increasing water dis-
charge. This guild was barely detectable later, in low
water period (from July to September).
Fig. 7 Guild compositions
(a); relative abundances of
the dominant species (b) and
taxa numbers of the
respective guilds (c) in the
running water segment and
standing water segment
formed from September to
November. Species are
shown with their four letter
OMNIDIA codes
Hydrobiologia
123

We hypothesized that decreasing water discharge and
increasing nutrient (mainly TN) concentration from July
to September will increase the proportion of high-profile
guild, and decrease that of low-profile guild. We detected
that increasing nutrient concentration from July to
September really caused an increase in the proportion
of high-profile guild, but this change was not accompa-
nied with the decrease in that of low-profile guild.
However, the proportion of low-profile guild was high
under high TN level in September as well. The relative
abundance of several low-profile guild taxa (Achnanthi-
dium minutissimum, Amphora pediculus) showed a
strong negative correlation with TN. A stronger positive
effect of TN was detected only on the relative abundance
of Achnanthidium eutrophilum from low-profile guild.
These observations suggest that Achnanthidium minu-
tissimum and Achnanthidium eutrophilum, both belong-
ing to the Achnanthidium minutissimum species complex
and to the low-profile guild containing disturbance-
tolerant R-strategist species, have quite different ecolog-
ical demands. We observed that the relative abundance of
Achnanthidium minutissimum (mean width 1.5–3.3 lm,
linear lanceolate shape) increased with the decreasing
TN values, while relative abundance of this taxa had a
decrease with one order of magnitude, when the TN
levels increased. In contrast, the relative abundance of
Achnanthidium eutrophilum (mean width 2.1–4.4 lm,
rhombic shape) increased with the increasing TN. Acs
et al. (2006) also showed in their study for river Danube
that relative abundance of Achnanthidium minutissimum
was high under those circumstances which was charac-
terized by high disturbance due to high water discharge
and low TN concentration. De Fabricius et al. (2003)
found also higher relative abundance of Achnanthidium
minutissimum in river segments with low nutrient
contents (low nitrate and nitrite) than in segments with
high nutrient contents, studying the river Cuarto. Acs
et al. (2006) draw attention to the presence of several
variant of Achnanthidium minutissimum taxa with
different ecological demands. Results of Gottschalk &
Kahlert (2012) also suggest that there could be different
ecological demands for species with different morpho-
logical characteristics (i.e. cell size). Studying 73
Swedish lakes, they found that the relative abundance
of Achnanthidium minutissimum with a mean width
between 2.2 and 2.8 lm was higher in lakes with low
nutrient contents. The relative abundance of Achnanthi-
dium minutissimum wider than 2.8 lm was almost the
same in both nutrient-rich and nutrient-poor lakes.
High-profile guild taxa attached by stalks (Gom-
phonema species) prefer to uptake nutrients dissolved
in water than nutrients absorbed in substratum (Prin-
gle, 1990; Berthon et al., 2011). High-profile guild
taxa were dominant from September to November (at
high TN concentration and low water discharge). We
found that the relative abundance of G. parvulum
showed a positive correlation with TN. This species
was dominant in September (at high TN level). Similar
to our results, Gomphonema species were found in
nutrient-rich water earlier (Kovacs et al., 2006;
Gottschalk and Kahlert, 2012); however, in our case,
Gomphonema olivaceum showed different character-
istics (see above).
Motile guild taxa are present in high numbers in
nutrient-rich habitats as competitors (Fairchild et al.,
1985; Van der Grinten et al., 2004), having the ability
to leave the inappropriate habitats (Johnson et al.,
1997). The relative abundance of motile guild taxa
was high in April (when water discharge was
relatively low, and TN was relatively high), and then
(after a temporary decrease in May and June) it was
almost the highest, when water discharge decreased
nearly by 85% from June to July. This decrease in
water discharge was accompanied with a 64% increase
in TN simultaneously. The resulting circumstances
(mainly the higher nutrient content) were not advan-
tageous any more for low-profile guild taxa, but for
motile guild taxa. They are able to produce extracel-
lular enzymes (Pringle, 1990), so presumably they
have higher nutrient uptake rates than low-profile
guild. Further decline of water discharge and increas-
ing TN favoured to increase the individual number of
high-profile guild taxa, which can explain the decrease
of taxa number and proportion of motile guild in
September. Relative abundance of motile guild
increased again in October and November, probably
due to competitive skills of the species.
Composition of diatom assemblages in running
and standing water segments
We assumed that the species composition is different
in running and standing water segments in a given
month. Fragilaria bidens was the most characteristic
to standing water segments in September. Although,
such clear separation was not revealed in the case of
the other samples based on taxa composition, Cocc-
oneis placentula var. placentula, C. placentula var.
Hydrobiologia
123

euglypta and Gomphonema species were characteris-
tic mainly to running water segments. Cocconeis
species able to adapt easily to disturbed circumstances
(Passy, 2007; Stenger-Kovacs et al., 2013), which
explains their presence in running water segments.
Since Gomphonema parvulum and Gomphonema
pumilum (Grunow) Reichardt & Lange-Bertalot are
known as nutrient-rich water preferring taxa (Kovacs
et al., 2006; Gottschalk & Kahlert, 2012), it explains
the high relative abundance of these species in
September (month with high TN concentration). TN
concentrations were the highest in October and
November. Certain taxa usually appear in nutrient-
rich water (Passy, 2007; Berthon et al., 2011; Gotts-
chalk & Kahlert, 2012; Stenger-Kovacs et al., 2013),
which explains the dominance of Diadesmis conferv-
acea in October and Melosira varians and Diatoma
vulgaris in November in running water segment.
Melosira varians and Diatoma vulgaris, similarly to
Cocconeis species, are able to adapt to high distur-
bances (R-strategy—Passy, 2007). These disturbances
were caused by high flow in the running water
segments. Moreover, we observed an interesting
phenomenon in the case of A. minutissimum and A.
eutrophilum: the proportion of A. minutissimum and A.
eutrophilum differed in samples collected from stand-
ing and running water segments in autumn: relative
abundance of A. minutissimum was higher in running
water segments. Conversely, the relative abundance of
A. eutrophilum was higher in standing water segments.
Because of the riverbed morphology and low water
discharge, fast water flow appeared in running water
segments. This was favourable for Achnanthidium
minutissimum, which are tolerant to physical distur-
bance (Passy, 2007; Berthon et al., 2011; Passy &
Larson, 2011). In contrast, we supposed that appear-
ance of Achnanthidium eutrophilum was defined
primarily by nutrients and not physical disturbances.
Potapova & Hamilton (2007) also reported Achnan-
thidium eutrophilum with higher relative abundance at
nutrient-rich habitats. These observations also confirm
the possible necessity of revising their current classi-
fication into guilds.
We assumed that running and standing water
segments can be characterized with different guild
ratios. These expectations were not confirmed by the
results. Taxa did not appear in the different segments
according to their guild affiliation. Rather, it was
observed that each genus—regardless of their
belonging to guilds—preferred more one or the other
segment. Within low-profile guild Cocconeis species,
which are known as R-strategist (see above), appeared
in running water segment with higher abundance.
Cymbella and Encyonema species—also belonging to
low-profile guild, which can form colonies and tubes
or can be stalked (Berthon et al., 2011)—preferred the
standing water segment. Similar contradictions
occurred in the high-profile guild as well: Fragilaria
species—except F. capucina var. vaucheriae (Kut-
zing) Lange-Bertalot—were found in standing water
segment with high abundance, in contrast Diatoma
vulgaris and Melosira varians were abundant in
running water segment. Each of these taxa are known
as colonial and/or stalked species (Berthon et al.,
2011). Overall, it seems that preferences and life forms
of given species or genera defined the taxa composi-
tion of segments, regardless to their current guild
affiliation.
Conclusions
Our results confirmed that the proportion of low-
profile guild increased with decreasing nutrient avail-
ability (increasing water discharge), and the propor-
tion of high-profile guild was lower in periods with
high water discharge and low nutrient availability.
Changes of relative abundance of motile guild
confirmed current knowledge that motile guild pre-
ferred nutrient-rich habitats and relatively sensitive to
increased water discharge. Our results suggest that
differences in seasonal patterns and compositional
changes are well reflected by the ecological guilds.
The expectation that running and standing water
segments can be characterized with different guild
ratios was not supported by the results. Our findings
for the relatively short time at the presence of
segments (from September to November) suggested
that further refinement of guild concept was necessary
based on life strategies (CSR model), life forms, and
more current knowledge about nutrient preferences of
taxa.
Our findings clearly indicate that there is a great
potential in the use of diatom guilds for environmental
status assessment. However, our results also suggest
that subdivisions within the current guilds are neces-
sary to make them more feasible for environmental
status assessment. Based on the results presented here
Hydrobiologia
123

and in literature data, the creation of subdivisions
within the low-profile guild could be based on the
nutrient preference of certain taxa. In light of this, a
subgroup in low-profile guild should include first
colonizers/pioneer species, which prefer low nutrient
concentration (e.g. Achnanthidium minutissiumum,
Encyonema and Cymbella species), another subgroup
should contain taxa, which are tolerant to disturbance
and tolerate or require higher nutrient contents (e.g.
Cocconeis placentula varietas, A. eutrophilum). Sub-
groups of high-profile guild may be based on the
disturbance tolerance and life forms of taxa. A
subgroup in high-profile guild should contain taxa
which can be characterized with high nutrient require-
ments and high disturbance tolerance (e.g. Diatoma
species, Melosira varians), an other subgroup in high-
profile guild should include taxa with low disturbance
tolerance (e.g. Fragilaria species). Based on our data,
similar subdivisions cannot be proposed for motile and
planktic guilds, but future research may indicate the
necessity of splitting also these guilds to several
subdivisions. It is pertinent to emphasize that the
confirmation of these suggestions definitely requires
further studies. More precise definitions of life forms,
life strategies and ecological needs of certain taxa are
strictly necessary for this kind of work, to which the
data presented in this work may be contributed.
Acknowledgments The authors are thankful for the support of
the Bolyai Janos Research Scholarship of the Hungarian Academy
of Sciences (Bacsi I.), Internal Research Project of the University of
Debrecen (Bacsi I.) and Hungarian Scientific Research Found PD
100192 (Torok P.) during manuscript preparation. The work was
supported by TAMOP 4.2.1./B-09/1/KONV-2010-0007, TAMOP-
4.2.2/B-10/1-2010-0024, TAMOP 4.2.2.C-11/1/KONV-2012-0010
and TAMOP 4.2.4.A/2-11-1-2012-0001 ‘National Excellence
Program—Elaborating and operating an inland student and
researcher personal support system’, projects. The TAMOP
projects are implemented through the New Hungary Development
Plan, co-financed by the European Social Fund and the European
Regional Development Fund. The authors are thankful for
measuring the physical and chemical parameters to the
Environmental Protection and Nature Conservation Authority,
Trans-Tisza Region, for water discharge data and rainfall data to the
National Water Authority and to the Hungarian Meteorological
Service.
References
Acs, E., K. Szabo, A. K. Kiss, B. Toth, G. Zaray & K. T. Kiss,
2006. Investigation of epilithic algae on the River Danube
from Germany to Hungary and the effect of a very dry year
on the algae of the River Danube. Archive fur Hydrobiol-
ogie 158: 389–417.
Berthon, V., A. Bouchez & F. Rimet, 2011. Using diatom life-
forms and ecological guilds to assess organic pollution and
trophic level in rivers: a case study of rivers in south eastern
France. Hydrobiologia 673: 259–271.
Biggs, B. J. F., D. G. Goring & V. I. Nikora, 1998. Subsidy and
stress responses of stream periphyton to gradients in water
velocity as a function of community growth form. Journal
of Phycology 34: 598–607.
Bolla, B., G. Borics, K. T. Kiss, M. N. Reskone, G. Varbıro & E.
Acs, 2010. Recommendations for ecological status
assessment of lake Balaton (largest shallow lake of central
Europe), based on benthic diatom communities. Vie Milieu
60: 1–12.
Chetelat, J., L. Cloutier & M. Amyot, 2010. Carbon sources for
lake food webs in the Canadian High Arctic and other
regions of Arctic North America. Polar Biology 33:
1111–1123.
De Fabricius, A. L. M., N. Maidana, N. Gomez & S. Sabater,
2003. Distribution patterns of benthic diatoms in a Pam-
pean river exposed to seasonal floods: the Cuarto River
(Argentina). Biodiversity and Conservation 12:
2443–2454.
Fairchild, G. W., R. L. Lowe & W. B. Richardson, 1985. Algal
periphyton growth on nutrient-diffusing substrates: an
in situ bioassay. Ecology 66: 465–472.
General Directorate of Water Management, homepage. http://
www.vizugy.hu.
Gottschalk, S. & M. Kahlert, 2012. Shifts in taxonomical and
guild composition of littoral diatom assemblages along
environmental gradients. Hydrobiologia 694: 41–56.
Hungarian Meteorological Service, homepage. http://www.met.
hu.
Johnson, R. E., N. C. Tuchman & C. G. Peterson, 1997. Changes
in the vertical microdistribution of diatoms within a
developing periphyton mat. Journal of the North American
Benthological Society 16: 503–519.
Kireta, A. R., E. D. Reavie, G. V. Sgro, T. R. Angradi, D.
W. Bolgrien, B. H. Hill & T. M. Jicha, 2012. Planktonic
and periphytic diatoms as indicators of stress on great
rivers of the United States: testing water quality and dis-
turbance models. Ecological Indicators 13: 222–231.
Koshmanesh, A., F. Lawson & I. G. Prince, 1997. Cell surface
area as a major parameter in the uptake of cadmium by
unicellular green microalgae. Chemical Engineering
Journal 65: 13–19.
Kovacs, Cs, M. Kahlert & J. Padisak, 2006. Benthic diatom
communities along pH and TP gradients in Hungarian and
Swedish streams. Journal of Applied Phycology 18:
105–117.
Krammer, K. & H. Lange-Bertalot, 1977a. Bacillariophyceae 1.
Teil: Naviculaceae. In Ettl, H., J. Gerloff, H. Heynig & D.
Mollenhauer (eds), Susswasserflora von Mitteleuropa.
Elsevier, Heidelberg.
Krammer, K. & H. Lange-Bertalot, 1977b. Bacillariophyceae 2.
Teil: Bacillariaceae, Epithemiaceae, Surirellaceae. In Ettl,
H., J. Gerloff, H. Heynig & D. Mollenhauer (eds), Sus-
swasserflora von Mitteleuropa. Elsevier, Heidelberg.
Krammer, K. & H. Lange-Bertalot, 2004a. Bacillariophyceae 3.
Teil: Centrales Fragilariaceae, Eunotiaceae. In Ettl, H., J.
Hydrobiologia
123

Gerloff, H. Heynig & D. Mollenhauer (eds), Die Sus-
swasserflora von Mitteleuropa. Spektrum Akademischer
Verlag, Heidelberg.
Krammer, K. & H. Lange-Bertalot, 2004b. Bacillariophyceae 4.
Teil: Achnanthaceae. Kritische Erganzungen zu Achnan-
thes s. l., Navicula s. str., Gomphonema. Gesamtliter-
aturverzeichnis Teil 1–4. In Ettl, H., J. Gerloff, H. Heynig
& D. Mollenhauer (eds), Susswasserflora von Mitteleuro-
pa. Spektrum Akademischer Verlag, Heidelberg.
Lamb, M. A. & R. L. Lowe, 1987. Effects of current velocity of
the physical structuring of diatom (Bacillariophyceae)
communities. Ohio Journal of Science 87: 72–78.
Lecointe, C., M. Coste & J. Prygiel, 1993. ‘‘Omnidia’’: software
for taxonomy, calculation of diatom indices and invento-
ries management. Hydrobiologia 269(270): 509–513.
Leps, J. & P. Smilauer, 2003. Multivariate analysis of ecological
data using CANOCO. Cambridge University Press,
Cambridge.
Molloy, J. M., 1992. Diatom communities along stream longi-
tudinal gradients. Freshwater Biology 28: 59–69.
Morales, E. A., P. A. Siver & F. R. Trainor, 2001. Identification
of diatoms (Bacillariophyceae) during ecological assess-
ments: comparison between light microscopy and scanning
electron microscopy techniques. Proceedings of the
Academy of Natural Sciences of Philadelphia 151: 95–103.
MSZ 1484-13:2009, 2009. Water quality. Part 13: Determina-
tion of nitrate and nitrite content by spectrophotometric
method.
MSZ EN 13946:2003, 2003. Water quality. Guidance standard
for the routine sampling and pretreatment of benthic dia-
toms from rivers.
MSZ EN 14407:2004, 2004. Water quality. Guidance standard
for the identification, enumeration and interpretation of
benthic diatom samples from running waters.
MSZ EN ISO 6878:2004, 2004. Water quality. Determination of
phosphorus. Ammonium molybdate spectrometric method
(ISO 6878:2004).
MSZ ISO 7150-1:1992, 1992. Determination of ammonium in
water. Manual spectrophotometric method.
Novais, M. H., S. Blanco, D. Hlubikova, E. Falsco, J. Goma, C.
Delgado, P. Ivanov, E. Acs, M. Morais, L. Hoffmann & L.
Ector, 2009. Morphological examination and biogeogra-
phy of the Gomphonema rosenstockianum and G. tergest-
inum species complex (Bacillariophyceae). Fottea 9(2):
257–274.
Padisak, J., L. O. Crossetti & L. Naselli-Flores, 2009. Use and
misuse in the application of the phytoplankton functional
classification: a critical review with updates. Hydrobiolo-
gia 621: 1–19.
Passy, S. I., 2002. Environmental randomness underlies mor-
phological complexity of colonial diatoms. Functional
Ecology 16: 690–695.
Passy, S. I., 2007. Diatom ecological guilds display distinct and
predictable behavior along nutrient and disturbance gra-
dients in running waters. Aquatic Botany 86: 171–178.
Passy, S. I. & C. A. Larson, 2011. Succession in stream biofilms
is an environmentally driven gradient of stress tolerance.
Microbial Ecology 62: 414–424.
Poff, N. L., N. J. Voelz & J. V. Ward, 1990. Algal colonization
under four experimentally-controlled current regimes in a
high mountain stream. Journal of the North American
Benthological Society 9: 303–318.
Potapova, M. & P. B. Hamilton, 2007. Morphological and
ecological variation within the Achnanthidium minutissi-
mum (Bacillariophyceae) species complex. Journal of
Phycology 43: 561–575.
Pringle, C. M., 1990. Nutrient spatial heterogeneity: effects on
community structure, physiognomy, and diversity of
stream algae. Ecology 71: 905–920.
Rimet, F. & A. Bouchez, 2012a. Biomonitoring river diatoms:
implications of taxonomic resolution. Ecological Indica-
tors 15: 92–99.
Rimet, F. & A. Bouchez, 2012b. Life-forms, cell-sizes and
ecological guilds of diatoms in European rivers. Knowl-
edge and Management of Aquatic Ecosystems 406: 01.
Stenger-Kovacs, Cs, K. Buczko, E. Hajnal & J. Padisak, 2007.
Epiphytic, littoral diatoms as bioindicators of shallow lake
trophic status: trophic diatom index for lakes (TDIL)
developed in Hungary. Hydrobiologia 589: 141–154.
Stenger-Kovacs, C., E. Lengyel, L. O. Crossetti, V. Uveges & J.
Padisak, 2013. Diatom ecological guilds as indicators of
temporally changing stressors and disturbances in the small
Torna-stream, Hungary. Ecological Indicators 24:
138–147.
Taylor, J. C., J. Prygiel, A. Vosloo, P. A. de la Rey & L. van
Rensburg, 2007. Can diatom-based pollution indices be
used for biomonitoring in South Africa? A case study of the
Crocodile West and Marico water management area.
Hydrobiologia 592: 455–464.
Ter Braak, C. J. F. & P. Smilauer, 2002. CANOCO reference
manual and CanoDraw for Windows user’s guide: software
for canonical community ordination (version 4.5). Ithaca,
NY: Microcomputer Power. http://www.canoco.com.
Accessed 2013.
Van der Grinten, E., M. Janssen, S. G. H. Simis, C. Barranguet &
W. Admiraal, 2004. Phosphate regime structures species
composition in cultured phototrophic biofilms. Freshwater
Biology 49: 369–381.
Varbıro, G., G. Borics, B. Csanyi, G. Feher, I. Grigorszky, K.
T. Kiss, A. Toth & E. Acs, 2012. Improvement of the
ecological water qualification system of rivers based on
first results of the Hungarian phytobenthos surveillance
monitoring. Hydrobiologia 695: 125–135.
Veraart, A. J., A. M. Romani, E. Tornes & S. Sabater, 2008.
Algal response to nutrient enrichment in forested oligo-
trophic stream. Journal of Phycology 44: 564–572.
Zar, J. H., 1999. Biostatistical analysis. Prentice Hall Interna-
tional, London.
Hydrobiologia
123






























