UNIVERSIDADE FEDERAL FLUMINENSE INSTITUTO DE SAÚDE DE NOVA FRIBURGO BIOMEDICINA MILLA BONECINI STUMBO POSSÍVEL ALTERAÇÃO NA FORMAÇÃO DA MICROBIOTA ANFIBIÔNTICA A PARTIR DE PRIVAÇÕES E/OU CUIDADOS EXCESSIVOS DURANTE A INFÂNCIA NOVA FRIBURGO 2018

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

UNIVERSIDADE FEDERAL FLUMINENSE
INSTITUTO DE SAÚDE DE NOVA FRIBURGO BIOMEDICINA
MILLA BONECINI STUMBO
POSSÍVEL ALTERAÇÃO NA FORMAÇÃO DA MICROBIOTA ANFIBIÔNTICA A PARTIR DE PRIVAÇÕES E/OU CUIDADOS
EXCESSIVOS DURANTE A INFÂNCIA
NOVA FRIBURGO 2018

MILLA BONECINI STUMBO
POSSÍVEL ALTERAÇÃO NA FORMAÇÃO DA MICROBIOTA ANFIBIÔNTICA A PARTIR DE PRIVAÇÕES E/OU CUIDADOS
EXCESSIVOS DURANTE A INFÂNCIA Trabalho de Conclusão de Curso
apresentado ao Instituto de Saúde de
Nova Friburgo, da Universidade Federal
Fluminense, como requisito à obtenção de
título de Graduação em Biomedicina.
Orientador: Profª. Drª Laís dos Santos
Falcão.
Co-orientador: Profº. Dr. Renato
Guimarães Varges.
NOVA FRIBURGO 2018

S929p Stumbo, Milla Bonecini
Possível Alteração na Formação da Microbiota Anfibiôntica a partir de Privações e/ou Cuidados Excessivos
Durante a Infância : / Milla Bonecini Stumbo ; Laís dos Santos
Falcão, orientadora ; Renato Guimarães Varges, coorientador.
Nova Friburgo, 2018.
61 f. : il.
Trabalho de Conclusão de Curso (Graduação em Biomedicina)-
Universidade Federal Fluminense, Instituto de Saúde de Nova
Friburgo, Nova Friburgo, 2018.
1. A Microbiota Anfibiôntica e sua Colonização Inicial. 2. Funções da Microbiota Anfibiôntica. 3. Privações e Cuidados Excessivos como interferentes na Microbiota. 4. A
Teoria da Higiene e a Possível Interferência no Sistema
Imune. 5. Produção intelectual. I. Falcão, Laís dos Santos,
orientadora. II. Varges, Renato Guimarães,
coorientador. III. Universidade Federal Fluminense. Instituto de
Saúde de Nova Friburgo. IV. Título.
CDD -
Ficha catalográfica automática - SDC/BNF Gerada com informações fornecidas pelo autor
Bibliotecária responsável: Natália Ribeiro de Rezende - CRB7/6422

MILLA BONECINI STUMBO
POSSÍVEL ALTERAÇÃO NA FORMAÇÃO DA MICROBIOTA ANFIBIÔNTICA A
PARTIR DE PRIVAÇÕES E/OU CUIDADOS EXCESSIVOS DURANTE A INFÂNCIA
Trabalho de Conclusão de Curso
apresentado ao Instituto de Saúde de
Nova Friburgo, da Universidade Federal
Fluminense, como requisito à obtenção de
título de Graduação em Biomedicina.
Aprovada em: ______/______/______
BANCA EXAMINADORA
Prof. Dr.: _____________________________________________________________
Instituição: ____________________________ Assinatura:______________________
Prof. Dr.: _____________________________________________________________
Instituição: ____________________________ Assinatura:______________________
Prof. Dr.: _____________________________________________________________
Instituição: ____________________________ Assinatura:______________________
NOVA FRIBURGO 2018

À Deus, pelas oportunidades permitidas. Aos familiares, pela
base, apoio e estímulo em sempre buscar por àquilo que me proporcionasse
realização pessoal e satisfação profissional. Aos amigos, pelos conselhos, apoio,
companheirismo e gargalhadas, as quais tornaram essa caminhada muito mais leve
e divertida.

AGRADECIMENTO
À todos os professores que tive a honra de conhecer nesse Campus, em
especial a Profª. Drª Laís Falcão, Prof. Dr. Renato Varges e Profª. Drª Caroline
Botino, por compartilharem conhecimento, tempo, dedicação e entusiasmo em cada
momento, tornando esses anos muito mais fáceis e prazerosos.

“O período de maior ganho em conhecimento e experiência
é o período mais difícil da vida de alguém”. (Dalai Lama)

RESUMO
Os estudos das interações microbiota-hospedeiro têm levado à compreensão, não
somente sobre a modulação e estabelecimento da microbiota anfibiôntica fundamental para a
manutenção da homeostase humana, como, também, sobre quais fatores impediriam a
composição desse ecossistema microbiano e de que maneira essas alterações se relacionam a
quadros de doença. Até o momento foi possível relacionar fatores como parto cesáreo e a
ausência do aleitamento materno com a desregulação dos padrões de colonização intestinal do
bebê, e, juntamente a esses fatores, os hábitos de vida e alimentares, higiene excessiva, perda
ou diminuição do contato interpessoal ou com animais de estimação como interferentes que
impedem o contato com micro-organismos que seriam essenciais para a composição inicial
adequada da microbiota anfibiôntica da criança, a qual estaria relacionada à modulação do
sistema imunológico e a diminuição da suscetibilidade no desenvolvimento de doenças, como
obesidade, diabetes Mellitus tipo 1, doenças imunomediadas e doença de Crohn.
Palavras-chave: formação da microbiota. Microbioma intestinal. Desenvolvimento da
microbiota. Microbiota e o sistema imune. Funções da microbiota. Microbiota e diabetes.
Interações microbiota-hospedeiro. Hipótese da higiene.
ABSTRACT
Studies of microbial-host interactions have led to the understanding not only of the
modulation and establishment of the amphibionic microbiota fundamental to the maintenance
of human homeostasis, but also of what factors would impede the composition of this
microbial ecosystem and how these diseases. To date, it has been possible to relate factors
such as cesarean delivery and the absence of breastfeeding with the deregulation of the infants
intestinal colonization patterns, and along with these factors, living and eating habits,
excessive hygiene, loss or decrease of interpersonal contact or with pets as interferents that
prevent contact with microorganisms that would be essential for the appropriate initial
composition of the child's amphibitic microbiota, which would be related to the modulation of
the immune system and the decrease of the susceptibility in the development of diseases such
as obesity, type 1 diabetes mellitus, immune-mediated diseases, and Crohn's disease.
Key words: Formation of the microbiota. Gut microbiome. Microbiota development.
Microbiota and the immune system. Functions of the microbiota. Microbiota and diabetes.
Microbe-Human Interactions. Hygiene hypothesis.

SUMÁRIO
1 INTRODUÇÃO ............................................................................................ 10
1.1 Objetivos ............................................................................................... 14
1.2 Metodologia ........................................................................................... 14
2 DESENVOLVIMENTO ................................................................................ 15
2.1 A Microbiota Anfibiôntica e sua Colonização Inicial .............................. 15
2.2 Funções da Microbiota Anfibiôntica ....................................................... 19
2.3 Privações .............................................................................................. 21
2.3.1 Vias de Parto .................................................................................. 21
2.3.2 Aleitamento Materno ....................................................................... 24
2.3.3 Nutricional ....................................................................................... 26
2.4 Cuidados Excessivos ............................................................................ 29
2.4.1 Higiene excessiva ........................................................................... 29
2.4.2 Contato interpessoal ....................................................................... 30
2.4.3 Contato com Animais ...................................................................... 31
2.4.4 Privações Ambientais ..................................................................... 32
2.4.5 Uso de Antimicrobianos .................................................................. 33
2.5 A Teoria da Higiene e a possível interferência no Sistema Imune ........ 34
3 DISCUSSÃO ................................................................................................. 37
4 CONCLUSÃO .............................................................................................. 48
BIBLIOGRAFIA ............................................................................................. 49

10
1 INTRODUÇÃO
Durante muitos séculos a presença de seres microscópicos não fazia parte do
entendimento, nem das descobertas científicas propostas à época. Apenas em 1676 Antoni
Van Leeuwenhoek, após construir um rústico microscópio, pôde observar seres não visíveis a
olho nu, em água e infusões, aos quais se referiu como animálculos. Em 1683, Leeuwenhoek
escreveu uma carta ao “Royal Society of London”, descrevendo quantitativamente e
qualitativamente os tipos de organismos observados, retirados de sua saliva e dentes, e
comparou-os com os de outros indivíduos, dando os primeiros indícios do que se tratava ser a
microbiota humana. Tornou-se, então, o primeiro bacteriologista da história (Porter, 1976;
Bakhtiar et al., 2013).
Mesmo após 40 anos de estudos, feitos por Leeuwenhoek, apenas no final do século
XIX esses micro-organismos despertaram interesse científico, primeiro por Louis Pasteur e,
posteriormente, por Joseph Lister, Robert Koch e Alexander Fleming. A princípio, a presença
de qualquer ser microscópico foi condicionada à causa de patologias, não distinguindo o
mutualismo, praticado por bactérias da microbiota anfibiôntica, do parasitismo, ou
amensalismo, realizado por organismos patogênicos. Pasteur refutou a teoria da geração
espontânea e, juntamente com Koch, defenderam e estabeleceram firmemente a teoria dos
germes da doença, em que organismos seriam os vetores de tais enfermidades, tornando tal
conceito o pilar sobre o qual a microbiologia médica foi construída. (American Society for
Microbiology, 2018).
Por muito tempo esse conjunto de germes foi denominado como flora, por estarem
compreendidos no mesmo Reino das plantas, o vegetal. Em 1956, Copeland dividiu o Reino
Protista, criado por Haeckel, em Reino Mychota, que compreendia organismos procariontes, e
Reino Protoctista, que incluía todos os eucariontes. Treze anos mais tarde, graças ao avanço
da ciência e dos estudos genéticos, os cinco Reinos foram reconhecidos por Whittaker, o qual
levou em consideração as especificidades e características morfofuncionais de cada
organismo. Com isso, o Reino Monera passou a englobar bactérias, o Reino Fungi, os fungos
e o Reino Protista, protozoários (Whittaker, 1969; Margulis, 1971; Whittaker & Margulis,
1978).
Por essa razão, hoje classificamos como microbiota o ecossistema formado pelo
conjunto de micro-organismos que se adaptaram e passaram a conviver em simbiose com o

11
hospedeiro, humano e não-humano, fornecendo benefícios mútuos. (Lozupone et al., 2012;
Gonçalves, 2014; Candido, Tunon e Carneiro, 2009).
Trata-se de uma enorme variabilidade de organismos, com mais de 1000 espécies e
subespécies já catalogadas. (Guarner, 2007; Barbosa et al., 2010). Ainda assim, de acordo
com Trabulsi & Sampaio (2000), cerca de 40% das espécies ainda não puderam ser isoladas e
estudadas. O que deve ser um desafio a ser alcançado, pois, segundo Preidis e Versalovic
(2009), o estudo detalhado sobre a composição da microbiota humana, juntamente com os
produtos metabólicos formados por esses micro-organismos, permitiria estabelecer quais
deles são responsáveis por manter a homeostase e quais seriam responsáveis pelo
desencadeamento de patologias.
No corpo humano, esses micro-organismos não estão distribuídos aleatoriamente, mas
sim, de acordo com a sua adaptabilidade em relação à temperatura, umidade, valor de pH,
concentração de nutrientes e aos diferentes tipos de receptores celulares, dentre os órgãos e
tecidos, internos e externos. (Fiocchi e Pereira de Sousa, 2012; Gonçalves, 2014).
Por muito tempo acreditava-se que o feto se mantinha estéril, ou seja, livre de qualquer
contato com micro-organismos durante a gestação (Pelczar, 1996; Talaro, 2008). Porém,
graças ao avanço de técnicas, como o sequenciamento genético, foi possível identificar
bactérias Gram positivas e negativas na placa basal placentária, além de outros locais
intrauterinos, e que o tempo correto de gestação é primordial para que ocorra uma colonização
adequada. Contudo, trata-se de um estudo continuado para excluir a hipótese de contaminação
dessas amostras por organismos presentes no ambiente (Rautava et al., 2012; Stout et al.,
2013; Aagaard, 2016).
Segundo Stout (2013), após o parto, o bebê adquire novos organismos, os quais irão
complementar sua microbiota. No parto normal, são adquiridos organismos importantes,
provenientes do canal vaginal, da pele e fezes maternas, que porventura são expelidas,
involuntariamente, e do ambiente externo, que irão colonizar a pele, orofaringe, seguida do
trato gastrointestinal (TGI) e trato respiratório superior do recém-nato. Porém, no parto
cesárea, essa complementação se torna comprometida, uma vez que o mesmo só entrará em
contato com micro-organismos da pele materna e do ambiente extra-uterino ao qual foi
inserido (Nicoli e Vieira, 2004; Talaro, 2008; Candido, Tunon e Carneiro, 2009).
A partir da amamentação essa colonização se intensifica (Gronlund et al., 2007;
Martin et al., 2012), sofrendo alterações durante a inserção de alimentos na dieta, no contato

12
interpessoal e com o ambiente, até se estabelecer, entre 1 e 3 anos de idade (Palmer et al.,
2007). A partir daí, a microbiota se mantém quase inalterada até a fase adulta (McFarland,
2000; Penders et al., 2006; Biasucci et al., 2008; Talaro, 2008).
Segundo Knol et al. (2005) e Palmer et al. (2007), o aleitamento materno introduz
novas colônias microbianas no TGI do recém-nato, o que garante o estímulo da maturação da
microbiota intestinal neonatal. O uso de fórmulas, em substituição ao leite materno, prejudica
o desenvolvimento adequado do sistema imunológico do bebê (Innis, 2007), além de alterar o
metabolismo ao longo de sua vida (Gale et al., 2012).
Como a maior parte das comunidades microbianas se encontra no TGI –
aproximadamente 70% -, torna-se o local onde se concentram as principais funções benéficas
oferecidas, majoritariamente pelas bactérias (Salonen e Vos, 2014).
Dentre as principais funções da microbiota intestinal, estão as
metabólicas/nutricionais, através das atividades bioquímicas dos micro-organismos, que inclui
favorecimento na digestão, produção de vitaminas K e B12, além de auxiliarem na absorção de
cálcio e ferro no cólon; função de prevenção à invasão de agentes infecciosos, impedindo o
estabelecimento de organismos patogênicos ou crescimento exacerbado de organismos
oportunistas; e função auxiliar na diferenciação e proliferação do epitélio intestinal, além do
desenvolvimento e maturação do sistema imune (Salonen e Vos, 2014).
De acordo com Wall et al. (2009) e Andrade (2010), aproximadamente 80% de todas
as células imunológicas ativas do corpo humano estão localizadas no TGI.
Os diferentes locais em que albergam micro-organismos, pertencentes à microbiota
anfibiôntica, podem sofrer alterações caso haja algum interferente externo exercendo tal
pressão seletiva. Como, por exemplo, higiene excessiva, o uso de medicações ou o déficit
nutricional (Bernstein e Shanahan, 2008; Sommer e Bäckhed, 2013).
A Hipótese da Higiene foi proposta há cerca de três décadas, porém, ainda é muito
estudada quanto a sua interferência na formação da microbiota anfibiôntica e,
consequentemente, na formação da imunidade (Strachan, 1989).
Strachan propôs, em 1989, a hipótese da higiene de doença alérgica após observar
respostas infecciosas menores em crianças que possuíam irmãos mais velhos, argumentando o
fato de que teriam maior probabilidade de exposição aos micro-organismos na primeira

13
infância, principalmente por ações anti-higiênicas, e que esse contato poderia protegê-los
contra o desenvolvimento de alergias, doenças atópicas e asma.
Segundo Isolauri (2017) e Stiemsma et al. (2015), a privação do contato com germes e
parasitas na primeira infância impede o imput necessário ao desenvolvimento correto do
sistema imune, o que leva a respostas alérgicas exacerbadas, doenças autoimunes e
inflamações crônicas, podendo aumentar a probabilidade ao desenvolvimento de obesidade e,
de acordo com Musso et al. (2010), diabetes Mellitus tipo 2. Essa ausência de micro-
organismos pode, inclusive, agravar os sintomas causados pela Doença de Crohn, ou, até
mesmo, aumentar o número de casos da doença (Zhang et al., 2017; Shen et al., 2017).
Já em relação aos antimicrobianos, se usados de forma indiscriminada durante a
infância, podem afetar o crescimento e o metabolismo da criança ao longo de sua vida. Por
estar em fase de estabelecimento e maturação, a microbiota se encontra particularmente
vulnerável à ação de antibióticos e sua depleção pode levar ao ganho excessivo de peso, ou
atraso no desenvolvimento infantil (Cox e Blaser, 2015). A mesma interferência também é
observada com o uso de medicamentos perinatais (Rautava et al., 2012).
Por outro lado, ao contrário do que muitos imaginam, o contato com animais de
estimação pode auxiliar no desenvolvimento da imunidade durante a primeira infância. De
acordo com Pelucchi et al. (2013), o contato com cães, logo após o nascimento, diminui
significativamente a propensão ao desenvolvimento de Dermatite Atópica e, segundo Lodge
et al. (2012), esse contato diminui, ou não interfere no desenvolvimento de alergias, mesmo
em crianças de alto risco (que possuam predisposição ao desenvolvimento de doenças
alérgicas).

14
1.1 OBJETIVOS
Investigar na literatura se há alteração na formação da microbiota anfibiôntica a partir
da privação do contato com micro-organismos, patogênicos ou não, seja a partir de cuidados
higiênicos excessivos, aumento do número de partos cesariana sobre partos normais, privação
do aleitamento materno, perda do contato com animais, diminuição da diversidade de
ambientes ou escassez nutricional, e, caso ocorra, se essa alteração acarreta prejuízos quanto a
recorrência e gravidade de infecções.
Elucidar tutores quanto à ligação entre o contato com micro-organismos e a formação
da defesa contra patógenos, desde a fase de recém-nato até a primeira infância.
Avaliar a variabilidade da microbiota anfibiôntica em relação a fatores predisponentes
(como gênero e predisposição gênica) e ambientais, além de esclarecer sobre a real
importância dos micro-organismos na vida humana.
1.2 METODOLOGIA
Para a elaboração deste Trabalho de Conclusão de Curso foram utilizadas diversas
ferramentas de pesquisa de artigos científicos, como PubMed, SciELO, Medline, BIREME,
LILACS e Google Schoolar, além de informações obtidas a partir da leitura de livros.
As palavras chave para a localização dos artigos utilizados no trabalho foram
“formation of the microbiota”, “gut microbiome”, “microbiota development”, “microbiota
and the immune system”, “functions of the microbiota”, “microbiota and diabetes”, “Microbe-
Human Interactions” e “hygiene hypothesis”.
Os artigos selecionados compreendem datas entre 1933 e 2018.

15
2 DESENVOLVIMENTO
2.1 A MICROBIOTA ANFIBIÔNTICA E SUA COLONIZAÇÃO INICIAL
A microbiota anfibiôntica é formada por micro-organismos simbióticos (mutualistas),
comensais e oportunistas. Os simbióticos são os que fornecem condições favoráveis ao
desenvolvimento e manutenção da homeostase humana. Os comensais são os inertes, ou seja,
não apresentam benefícios, nem malefícios ao homem. Já os oportunistas são organismos que
causam patologias a partir de uma desordem da microbiota, a partir do uso de
antimicrobianos, ou do sistema imune do hospedeiro, seja por imunodeficiências,
normalmente causadas pelo vírus da imunodeficiência humana, por quimioterápicos ou
terapia imunossupressora em transplantados e doenças autoimunes, extensas queimaduras ou
exposição de mucosas (Cândido, Tunon e Carneiro, 2009).
Diversos autores consideram a microbiota como um microbioma, ou seja, o conjunto
de genes que constituem o genoma do ecossistema formado pelos organismos simbióticos
(Ley et al., 2008). Em 1977, quando Lederberg criou o termo “microbioma”, argumentou que
os micro-organismos que habitam o corpo humano deveriam ser incluídos como parte de seu
genoma, por interferirem diretamente na fisiologia humana (Lerderberg e Maccray, 1977).
Outros consideram ser um “órgão independente”, o qual fornece condições basais para
uma vida saudável, tendo, portanto, funções primordiais na vida humana. Para tal, levam em
consideração o recente sucesso do transplante de microbiota fecal, especialmente no contexto
da infecção por Clostridium difficile (van Nood et al., 2013).
Em 2007 foi desenvolvido o Projeto Microbioma Humano (HMP), que tinha como
objetivo identificar espécies encontradas em diversos locais do corpo humano, como narinas,
pele, cavidade oral e tratos gastrointestinal e urogenital, analisando a interferência desses
micro-organismos na saúde humana, além da forma como estariam alterados em quadros
patológicos. Após a coleta de amostras do corpo feminino e masculino, foi possível identificar
mais de 800 genomas, com aproximadamente 5 milhões de genes, distribuídos em cerca de 10
mil espécies diferentes que compõem uma microbiota saudável (Consortium THMP, 2012;
Ribeiro et. al., 2014).
Com o Projeto, foi possível estimar entre 81-99% dos gêneros, enzimas e análises de
comportamento das comunidades formadas pelo microbioma saudável. Estes resultados
traçam a gama de configurações estruturais e funcionais normais nos ecossistemas
microbianos de uma população saudável, permitindo a futura caracterização das aplicações

16
epidemiológicas, ecológicas e translacionais do microbioma humano (Consortium THMP,
2012).
De acordo com Cândido, Tunon e Carneiro (2009), só foi possível comprovar a
influência substancial exercida pelos micro-organismos em seu hospedeiro após estudos com
animais axênicos, ou seja, germ-free. Segundo eles, esses animais apresentavam diversos
prejuízos metabólico-funcionais ao longo de suas vidas.
Estes animais não são anatômica e fisiologicamente
normais, já que apresentam tecido linfático pouco
desenvolvido, parede do epitélio intestinal fina, ceco, quando
presente, estendido, e baixo título de anticorpos. Além disso,
devem receber suplemento alimentar com vitamina K e
complexo B. Tais animais também apresentam baixa taxa de
metabolismo e baixo batimento cardíaco.
A partir daí, foi possível iniciar a correlação entre a função, que gera o benefício, e as
espécies que exercem esse mutualismo. Dentre os principais benefícios promovidos pelos
micro-organismos, podemos citar a proteção contra infecções por organismos
patogênicos/oportunistas, atividades metabólicas de síntese, degradação e controles
energéticos, além da modulação do sistema imune (Cândido, Tunon e Carneiro, 2009;
Salonen e Vos, 2014).
Porém, a privação do contato com germes, ainda na infância, pode acarretar
disfunções nessa simbiose ao longo da vida, pois, a diversidade de organismos dentro de um
determinado habitat corporal, definida entre volume e variabilidade entre espécies, tem sido
associada ao surgimento de algumas patologias. Como, por exemplo, a baixa diversidade de
micro-organismos do TGI relacionada à obesidade e doença inflamatória intestinal
(Turnbaugh et al., 2009; Qin et al., 2010), enquanto que a elevada variabilidade na vagina é
relacionada à vaginose bacteriana (Fredricks et al., 2005). No primeiro caso, há a falta, ou
insuficiência, de organismos com atividades metabólico-funcionais, no segundo, ausência, ou
depleção, de espécies responsáveis pela proteção contra patógenos oportunistas.
A Hipótese da Higiene foi proposta por Strachan, em 1989, para que a correlação entre
a ausência do contato com micro-organismos na infância e sua interferência na formação da
microbiota anfibiôntica fosse melhor estudada. Para isso, vários interferentes foram
analisados, como ausência do contato interpessoal, principalmente com irmãos mais velhos,
dieta, exposição a antibióticos e higiene excessiva e comparados com o aumento de casos de
doenças imunológicas (Strachan, 1989).

17
Germes podem ser encontrados no ambiente, fômites e superfícies. Porém, o fato de
que as espécies mais predominantes, como o Bifidobacterium spp., presentes na microbiota
anfibiôntica de bebês, não serem encontrados facilmente nesse ambiente extra-uterino, sugere
que essas crianças adquiram micro-organismos simbióticos a partir de outro ser humano,
sendo essa carga mais significativa vinda da mãe (Rautava et al., 2012).
No início do século XX, o pediatra francês Henry Tissier afirmou que o feto se
desenvolve em um ambiente estéril, adquirindo seu inóculo bacteriano inicial a partir do
contato com a microbiota materna, no momento parto (Tissier, 1900). Após aproximadamente
um século essa questão ainda é muito discutida, devido à hipótese de que qualquer presença
bacteriana no útero seria prejudicial ao feto. Isso ocorreu devido ao fato da maioria das
bactérias detectadas em infecções intra-uterinas estar comumente presente no canal vaginal
materno (Goldenberg et al., 2000), sendo relacionadas ao risco de parto prematuro
significantemente aumentado em gestantes diagnosticadas com vaginose bacteriana (Fiscella,
1996).
Contudo, a partir do avanço de métodos moleculares de sequenciamento de genes, foi
possível identificar micro-organismos não cultiváveis, ou seja, organismos que apresentam
exigências nutricionais ou ambientais não reproduzíveis em técnicas convencionais de cultivo
(Ribeiro et al., 2014).
Pesquisas recentes sobre o potencial de transmissão microbiana através da barreira
placentária identificaram micro-organismos na placa basal placentária (Satokari et al., 2009;
Fardini et al., 2010; Rautava et al., 2012; Stout et al., 2013; Aagaard et al., 2014), no líquido
amniótico (Wang et al., 2013), sangue do cordão umbilical (Jimenez et al., 2005) e
membranas fetais (Steel et al., 2005). Contudo, trata-se de um estudo continuado para excluir
a hipótese de contaminação dessas amostras por organismos presentes no ambiente (Rautava
et al., 2012; Stout et al., 2013; Aagaard, 2016).
O tempo correto de gestação é primordial para que o recém-nascido possa estar
preparado para receber novas espécies durante o parto. Bebês prematuros, principalmente os
de alto risco (peso ≤ 1.500g), são muito vulneráveis a perturbações em sua microbiota, por
não serem totalmente desenvolvidos fisiologicamente (Björkström et al., 2009). Embora nem
todos os estudos subsequentes corroborem esse achado, parece que o intestino prematuro
ainda não está totalmente desenvolvido para a colonização ideal por Bifidoacterium spp.
(Butel et al., 2007). Um estudo feito por Chang et al. (2011) utilizou novas técnicas de

18
sequenciamento para avaliar a microbiota fecal em amostras de 10 prematuros de muito baixo
peso e revelou que a colonização intestinal nesses prematuros é um processo dinâmico, que
envolve um grande número de espécies bacterianas, das quais, aproximadamente, 20% são
atualmente não classificadas.
A amamentação exerce uma grande influência nos padrões de colonização intestinal
(Harmsen et al., 2000) e oral (Biagi et al., 2018) no início da infância. Os componentes
presentes no leite fornecem tanto nutrientes para o bebê, quanto para os micro-organismos
presentes na microbiota intestinal, além de atuarem na defesa contra colonização de espécies
oportunistas/patogênicas, a partir de novas espécies que foram introduzidas através do contato
com a pele da mãe e dos micro-organismos presentes no leite materno (DeWeerdt; Biagi et
al., 2018).
A microbiota presente na cavidade oral se inicia no nascimento, a partir de micro-
organismos presentes nas fezes que são expelidas pela mãe a partir do parto vaginal (Stout et
al., 2013), sofrendo influência do tipo de amamentação (materna ou de terceiros) e
alimentação (fórmulas infantis e início da dieta) (Palmer et al., 2007). Em partos cesárea essa
interação inicial não ocorre, e irá depender apenas da introdução da amamentação. (Biagi et
al., 2017, 2018).
Outro fator igualmente importante para o acesso e transferência de micro-organismos
são os componentes familiares, principalmente irmãos mais velhos, os quais, segundo
Strachan, propiciariam um contato anti-higiênico, aumentando a carga de germes que a
criança teria acesso ainda na primeira infância (Strachan, 1989).
Com um ano de idade, acredita-se que a microbiota infantil comece a convergir para
um perfil que se assemelha ao dos adultos, sendo estabelecido por volta dos 2 a 3 anos de
idade (Palmer et al., 2007; Koenig et al., 2011). No entanto, de acordo com Rautava et al.
(2012), a compreensão da dinâmica da composição da microbiota intestinal ainda é
fragmentada, tornando evidente que mudanças essenciais nos processos fisiológico,
exposições dietéticas e estilo de vida, que afetam a composição da microbiota intestinal,
ocorrem além dessa idade.
Esses micro-organismos não se estabelecem, nem se distribuem aleatoriamente pelo
TGI, mas a partir de diversos fatores, como estado nutricional ou tipo de alimentação, idade,
utilização de antimicrobianos, probióticos e prebióticos, ambiente, microbiota materna (a
partir da via do parto e aleitamento), interações microbianas e entre microbiota-hospedeiro,

19
além da presença de certos genes e regiões de complementaridade com seus receptores
celulares específicos (Mackie et al., 1999; Savage, 1999; Bourlioux et al., 2003).
Fatores que permitem que um organismo colonize órgãos e superfícies humanas não
estão totalmente explicados, mas há a convergência sobre o fato de gerar benefícios mútuos,
tanto para o hospedeiro, quanto para bactérias, fungos e parasitos (Bakhtiar et al., 2013).
Novas teorias estão surgindo para explicar essa associação, como a Teoria do Hologenoma,
que considera que o holobionte – um organismo e todos os seus micro-organismos
simbióticos associados, incluindo parasitos, mutualistas, sinergistas e amensalistas (ou seja,
todo o microbioma) – evoluiu como resultado da simbiopoiese, ou co-desenvolvimento do
hospedeiro e simbiontes (Gilbert et al., 2010; Gordon et al., 2013; Carrier e Reitzel, 2017).
Segundo a Teoria, com a utilização de técnicas atuais de sequenciamento foi possível
caracterizar microbiotas pertencentes a indivíduos saudáveis e composições atípicas em casos
de patologias. Contudo, homem e micro-organismos devem ser estudados e compreendidos
juntos, para que processos de saúde e doença sejam melhores esclarecidos. (van de Guchte et
al., 2018).
2.2 FUNÇÕES DA MICROBIOTA ANFIBIÔNTICA
As principais áreas do corpo humano que abrigam micro-organismos simbióticos são:
cavidade oral, trato gastrointestinal e pele em maiores números, tanto qualitativa quanto
quantitativamente; genitália, sendo este, necessariamente, em menor variabilidade de
espécies; e vias aéreas superiores e inferiores, onde, aproximadamente, 20% da população
possui espécies patogênicas presentes nas narinas, como o Staphylococcus aureus incluído em
sua microbiota anfibiôntica, o qual se relaciona diretamente com local ao qual o indivíduo
está, ou esteve, inserido (Riva et al., 2016; van de Guchte et al., 2018).
A microbiota presente na cavidade oral do lactente compreende, em sua maioria, os
gêneros Pseudomonas, Staphylococcus e Streptococcus (Biagi et al., 2018). Trata-se de um
local com diversos microambientes que, até a fase adulta, irá moldar uma vasta variabilidade
de espécies pertencentes ao seu microbioma simbiótico, chegando a alcançar um quantitativo
de, aproximadamente, 1.000 espécies (Aas et a., 2005).
Essas espécies produzem metabólitos que auxiliam na defesa contra agentes externos e
também auxiliam na digestão/meetabolização de alguns alimentos. Porém, alterações como

20
mudança de hábitos alimentares, ou imunossupressão levam a um quadro de disbiose, onde as
bactérias podem passar a produzir fatores de virulência e gerar doenças (Lamont et al., 2018).
A cavidade oral se comunica e compartilha com diversas espécies da microbiota
comensal dos tratos respiratórios superiores e inferiores (Fonkou et al., 2018). A microbiota
presente nas vias aéreas superiores terá a principal função de proteção contra patógenos
presentes em suspensão. Já a inferior, além da proteção contra micro-organismos oportunistas,
irá sensibilizar as células T, presentes na barreira epitelial, auxiliando na modulação imune
(Gollwitzer et al., 2014).
Estudos recentes sobre a composição da microbiota intestinal humana revelaram uma
série de características notáveis deste ecossistema microbiano, as quais acredita ter um papel
extremamente importante, ou mesmo decisivo, na saúde. Os filos mais dominantes, descritos
até o momento são, principalmente, Bacteroidetes (com predominância dos gêneros
Bacteroides e Prevotella) e Firmicutes. Em menor grau, Proteobacteria e Actinobacteria
(CTHMP, 2012).
Dentre as principais funções da microbiota intestinal, estão as
metabólicas/nutricionais, através das atividades bioquímicas dos micro-organismos, que inclui
favorecimento na digestão, produção de vitaminas K e B12, além de auxiliarem na absorção de
cálcio, magnésio e ferro no ceco e cólon; função de prevenção à invasão de agentes
infecciosos, impedindo o estabelecimento de organismos patogênicos ou crescimento
exacerbado de organismos oportunistas; e função auxiliar na diferenciação e proliferação do
epitélio intestinal (Guarner e Malagelada Jr., 2003; Cândido, Tunon e Carneiro, 2009;
Barbosa et al., 2010; Fiocci e Pereira de Souza, 2012; Salonen e Vos, 2014).
Algumas espécies favorecem a digestão por serem responsáveis em metabolizar
componentes alimentares que são indigeríveis pelas enzimas dos mamíferos, como os
oligossacarídeos do leite humano e a fibra alimentar (Marcoal et al., 2010; Atarashi et al.,
2011). A fermentação de carboidratos não digeríveis pelo hospedeiro ocorre principalmente
no ceco e no cólon ascendente, sendo uma importante fonte de energia para a proliferação de
bactérias, além de produzir ácidos graxos de cadeia curta, que são fontes essenciais de energia
para muitos tecidos do hospedeiro e importantes moduladores imunes (Smith et al., 2013;
Park et al., 2015).

21
Isso se traduz em recuperação de energia da dieta, favorecimento da absorção de íons,
produção de vitaminas (K, B12, biotina, ácido fólico e pantotênico) e a síntese de
aminoácidos a partir de amônia ou ureia (Guarner, 2007).
Já a microbiota anfibiôntica vaginal possui baixa variabilidade, pois apresenta
dominância do gênero Lactobacillus, o qual impede que organismos oportunistas colonizem a
região, provocando vaginoses bacterianas, podendo ser extremamente perigoso para gestantes
(Aagaard et al., 2012).
O processo de montagem da microbiota da pele começa durante o parto e procede
principalmente de acordo com o local do corpo ao longo de várias semanas (Chu et al.,
2017). Esse ecossistema se desloca notavelmente durante a puberdade, com aumento da
predominância de Corynebacterium e Cutibacterium (anteriormente denominado como
Propionibacterium ) e diminuição da abundância de Firmicutes (incluindo espécies
de Staphylococcus e Streptococcus ). Na idade adulta, apesar da exposição contínua da pele
ao ambiente, a composição microbiana permanece surpreendentemente estável ao longo do
tempo (Oh et al., 2016).
A pele, por ser o maior órgão do corpo humano, abriga um numeroso ecossistema
microbiano em toda a sua extensão, importantes para a manutenção de sua fisiologia e
imunidade (Chen et al., 2018). A microbiota da pele modula a expressão de vários fatores
inatos, incluindo a interleucina 1α (IL-1α), componentes do complemento e peptídeos
antimicrobianos, que são produzidos por queratinócitos e células sebáceas (Naik et al., 2012).
A Corynebacterium spp., gênero mais abundante na pele, possui diversas funções,
como a de se ligar a receptores celulares, a partir de estruturas presentes em sua parede
bacteriana, e deflagrar respostas inflamatórias ou anti-inflamatórias, dependendo do contexto
em que estão inseridas; demonstrou recentemente inibir o crescimento de Streptococcus
pneumoniae; outras espécies de Corynebacterium suprimem genes de virulência de
Staphylococcus aureus e estimulam genes relacionados ao comensalismo (Chen et al., 2018).
2.3 PRIVAÇÕES
2.3.1 Vias de Parto
Durante a gravidez, a gestante apresenta diversas alterações em seu corpo, incluindo
modificações em sua microbiota vaginal (Aagaard et al., 2012; Romero et al., 2014) e

22
intestinal (Koren et al., 2012), que serão essenciais para a transmissão vertical microbiana ao
recém-nato, através do parto vaginal (Mueller et al., 2015).
Em um estudo feito por Aagaard et al., descobriu-se que as grávidas possuíam menor
variabilidade bacteriana em sua microbiota, quando comparadas à mulheres não grávidas,
com dominância de Lactobacillus, Clostridium, Bacteroides e Actinomyces spp. A presença
primordial de lactobacilos na vagina é necessária para que o pH da região se mantenha ácido,
impedindo que organismos oportunistas colonizem o canal vaginal, útero e,
consequentemente, o líquido amniótico, placenta e feto (Goldemberg et al., 2000; van de
Wijgert et al., 2014).
Um exemplo dessa proteção está nos produtos gerados pelo metabolismo dos
lactobacilos, como o peróxido de hidrogênio e bacteriocinas (Corr et al., 2007), ou em
algumas espécies ( L. jensenii e L. crispatus ) que possuem epítopos específicos em sua
superfície, os quais impedem a colonização de patógenos, como a Neisseria gonorrhoeae
(Spurbeck e Arvidson, 2010), além do surgimento de outras vaginoses bacterianas
(Rosenstein et al., 1997).
Além disso, há evidências de que a microbiota intestinal também sofra modificações,
diminuindo sua diversidade bacteriana na medida em que ocorre a progressão da gravidez
(Koren et al., 2012), ocorrendo diminuição significativa de Streptococcus spp. e bactérias da
família Enterobacteriaceae, já no primeiro trimestre, enquanto há aumento de
Faecalibacterium spp. até o terceiro trimestre da gestação (Kuczynski et al., 2011).
Segundo Koren et al., (2012), as implicações dessas alterações para a saúde da mãe,
feto e, posteriormente, da criança ainda não foram totalmente esclarecidas, contudo, essas
mudanças adaptativas sugerem que essas bactérias evoluíram com o homem, permitindo ao
seu hospedeiro obter maior ganho energético durante a gestação, garantindo a nutrição da
mãe, favorecendo, assim, o desenvolvimento do feto.
Já de acordo com Mueller et al. (2015), essas mudanças provavelmente fariam parte de
uma resposta adaptativa para proteger e promover a saúde da criança, fornecendo ao recém-
nascido um inóculo bacteriano específico, antes de sua exposição aos micro-organismos
ambientais.
A transferência da microbiota intestinal materna para o recém-nascido, durante o parto
vaginal, garante o acesso imediato a micro-organismos responsáveis pelo suprimento
energético, logo nas primeiras horas de vida do bebê (Koren et al., 2012; Thum et al., 2012).

23
Estudos feitos por Makino et al. (2011) mostrou que, nas fezes de bebês nascidos por parto
normal, havia a presença de quantidade significativa de Bifidobacterium spp., proveniente das
fezes maternas, enquanto que em recém-nascidos por via cesariana, não apresentavam.
Já em relação à prematuridade, bebês nascidos antes de 32 semanas são especialmente
vulneráveis a perturbações na microbiota. Muitos nascem por cesariana e, portanto, não
entram em contato com os micro-organismos do canal vaginal. Alguns necessitam de terapias
antimicrobianas de amplo espectro e, além disso, são mantidos em incubadoras estéreis, tendo
contato mínimo com organismos da pele humana e do ambiente extrauterino (Palmer et al.,
2007; Koenig et al., 2011).
Com isso, a microbiota intestinal de bebês prematuros se torna marcadamente
diferente da encontrada em bebês nascidos a termo (a partir de 39 semanas). Tendem,
portanto, a ter uma menor proporção de micro-organismos que são benéficos para a saúde
intestinal, como Bifidobacterium spp. e Lactobacillus spp., bem como uma menor diversidade
de bactérias em geral, ocasionando aumento do número de bactérias patogênicas (DeWeerdt
2018).
De acordo com DeWeerdt (2018), a comunidade bacteriana é muitas vezes caótica,
com consideráveis mudanças em sua composição, em questão de dias. Acredita-se que a
microbiota intestinal anormal de bebês prematuros tenha um papel em sua vulnerabilidade à
enterocolite necrosante, uma forma grave de inflamação intestinal que ocorre repentinamente
nas primeiras semanas de vida e pode causar danos permanentes ao intestino. Embora bebês
nascidos a termo possam desenvolver a doença, pelo menos 75% dos casos ocorrem em bebês
nascidos prematuramente.
O parto cesáreo foi associado a padrões precoces de colonização bacteriana intestinal,
segundo Stokholm et al (2016). Com apenas uma semana de vida, os bebês já apresentavam
espécies como Citrobacter freundii, Clostridium spp., Enterobacter cloaceae, Enterococcus
faecalis, Klebsiella oxytoca e K. pneumoniae, além de Staphylococcus aureus, enquanto que,
em bebês nascidos por parto normal, apresentavam apenas Escherichia coli fora dos padrões
normais.
Em relação à microbiota presente nas vias aéreas, o parto cesáreo está relacionado à
colonização por Streptococcus pneumoniae, em bebês de apenas um mês de vida, e por E.
faecalis aos 3 meses, enquanto que em partos normais essas alterações não são observadas
(Stokholm et al., 2016).

24
Huh et al. (2012), em seus estudos feitos com crianças obesas e não obesas, puderam
estabelecer a relação entre jovens obesos e parto cesárea. Na análise feita identificaram que,
aproximadamente, 16% das crianças que haviam nascido através de parto cesariana
desenvolveram obesidade, em comparação a apenas 7,5% em crianças nascidas por parto
normal. Bervoets et al, (2013) atribuíram à essa interferência o fato de que crianças nascidas
por parto normal possuem uma relação menor entre os filos Bacteroidetes (com predomínio
de Bacteroides spp.) e Firmicutes, enquanto aqueles nascidos por parto cesáreo apresentam
maior relação entre esses filos, tendo Firmicutes em quantidades duas vezes maiores,
indicando a variabilidade microbiana como causalidade da obesidade.
Existem, ainda, diferenças entre os tipos de cesariana, as quais podem ocorrer de
maneira eletiva ou emergencial, sendo ambas relacionadas à prática comum de
antibioticoterapia durante o procedimento, o que torna o parto cesáreo fator de risco para
diversas doenças imunomediadas em crianças (Hakansson et al., 2008) e disbiose de longo
prazo (Azad et al., 2015).
Stokholm et al (2016) puderam concluir com o estudo que, em partos cesáreos, onde
havia uso de antibióticos intraparto, o número de E. faecalis e S. aureus aumentava
consideravelmente, enquanto que em bebês nascidos por parto normal, com o mesmo período
de tempo de terapia medicamentosa, era associado à colonização por K. oxytoca e
Staphylococcus hemolyticus. Além de estabelecerem a hipótese de que tais alterações
ocorreriam devido à interrupção da transmissão vertical dos micro-organismos comensais da
mãe para o recém-nascido, através do parto vaginal.
Embora o parto cesáreo e o nascimento prematuro possam predispor crianças ao
desenvolvimento de asma e doenças atópicas, existem dados insuficientes e conflitantes sobre
o efeito dos fatores perinatais no desenvolvimento da alergia alimentar (Renz-Polster et al.,
2005; Liem et al., 2007; Kvenshagen et al., 2009). Koplin et al. (2008) sugeriram, portanto,
que o parto cesáreo aumenta o risco de sensibilização a alérgenos alimentares, mediada por
imunoglobulina E (IgE), devido à colonização tardia e alterada da microbiota gastrointestinal.
2.3.2 Aleitamento Materno
Diversos estudos mostraram que bebês amamentados com leite materno possuem
menores chances de desenvolver enterocolite necrosante, quando comparados àqueles
alimentados por fórmulas. É considerado um alimento rico por, além de suprir

25
energeticamente o recém-nato, contém ingredientes que promovem o estabelecimento de uma
microbiota intestinal saudável, como, por exemplo, cadeias curtas de moléculas de açúcar, ou
oligossacarídeos de leite humano. Esses açúcares são a segunda fonte de carboidratos mais
abundantes no leite humano após a lactose. Porém, não são para a nutrição dos bebês, mas
sim, atuam como prebióticos, ou seja, fornecem alimento para micro-organismos comensais,
como o Bifidobacterium spp. Eles também revestem o lúmen intestinal e se ligam a bactérias
patogênicas, tornando mais difícil a invasão de micróbios causadores de doenças (DeWeerdt,
2018).
Outro componente do leite materno, a proteína lactoferrina, tem várias propriedades
antimicrobianas. Ele suprime o crescimento de bactérias e pode, até mesmo, desencadear a
morte de certos micro-organismos nocivos, ligando-se ao lipopolissacarídeo presente na
membrana externa das bactérias (DeWeerdt, 2018).
A amamentação exerce uma grande influência nos padrões de colonização intestinal
no início da infância. Estudos vêm relatando que a microbiota intestinal de bebês
amamentados exclusivamente por leite materno é dominada por bifidobactérias, enquanto
bebês alimentados com fórmulas infantis abrigam um microbioma mais diversificado
(Harmsen et al., 2000).
Por outro lado, Penders et al. (2005) examinaram a composição da microbiota
intestinal em 50 crianças amamentadas por leite materno e 50 amamentadas por fórmulas
infantis e descobriram que, em nível de espécie, todas as crianças foram colonizadas por
Bifidobacterium spp., sem diferenças quantitativas ou de prevalência. Os mesmos
investigadores analisaram, posteriormente, amostras fecais obtidas de mais de 1.000 crianças,
todas com um mês de idade, não sendo detectadas diferenças entre a colonização por
bifidobactérias entre lactentes alimentados com leite materno e alimentados com fórmula.
Porém, bebês alimentados exclusivamente com fórmula foram mais frequentemente
colonizados com Escherichia coli, Clostridium difficile, Bacteroides e Lactobacillus spp
(Penders et al., 2006).
Os estudos concluíram que bebês nascidos parto vaginal, com tempo de gestação
normal e em período de amamentação exclusiva, eram mais propensos a ter o maior número
de bifidobactérias intestinais (Penders et al., 2006).
Em estudos feitor por Roger et al., de 2010, sete bebês amamentados com leite
materno e sete amamentados com fórmula foram acompanhados até a idade de 18 meses. Ao

26
longo desse tempo, aproximadamente 20 amostras fecais foram obtidas e analisadas. Durante
a amamentação exclusiva, as bifidobactérias dominaram a microbiota intestinal com uma
participação em mais de 50% do total de colônias bacterianas, enquanto que, no mesmo
período de tempo, em bebês alimentados com fórmula apenas, aproximadamente, 25% do
total de micro-organismos intestinais eram bifidobactérias.
Os bebês amamentados no estudo receberam leite materno até a idade de 6
a 15 meses e abrigaram uma composição mais complexa de Bifidobacterium spp.
Além disso, as diferenças na composição fecal de Bifidobacterium spp. entre
lactentes amamentados por leite materno e alimentados com fórmula pareciam
permanecer após a interrupção da amamentação (Roger et al., 2010).
O que define a formação da microbiota é desconhecido e o desenvolvimento inicial de
enterótipos em lactentes ainda não está claro (Arumugam et al., 2011), mas acredita-se que a
microbiota intestinal individual estável se desenvolva de forma gradual, sofrendo
modificações por fatores dietéticos e ambientais durante a infância, até culminar em um perfil
geral mais semelhante ao adulto. O primeiro passo compreende a colonização precoce,
principalmente pelo filo Firmicutes, incluindo bactérias aeróbias ou anaeróbias facultativas
como, por exemplo, Staphylococcus e Streptococcus spp., e organismos pertencentes à família
Enterobacteriaceae. Esses primeiros colonizadores são, então, seguidos por bactérias do filo
Actinobacteria, assim como por mais micro-organismos anaeróbios, incluindo
Bifidobacterium spp., especialmente em bebês amamentados por leite materno (Hamsen et al.,
2000; Roger et al., 2010).
A próxima etapa de sucessão é o aumento do filo Bacteroidetes, com dominância dos
gêneros Bacteroides e Prevotella, e outros anaeróbios, como Clostridium spp. e Eubacterium
spp. (Palmer et al., 2007; Koenig et al., 2011).
2.3.3 Nutricional
A grande biodiversidade de espécies que habitam o ecossistema intestinal faz com
que esse local, além de acumular diversas funções benéficas para a saúde humana, seja de
extrema importância para a manutenção do estado nutricional do hospedeiro (Guarner, 2007).
Durante a formação da microbiota humana, da gestação até o parto, o bebê passa a
adquirir micro-organismos essenciais, que serão responsáveis por sua homeostase (Stout et
al., 2013). A partir da introdução da dieta, novos organismos serão incorporados nesse

27
ecossistema, gerando modificações ao longo de sua vida, que poderão auxiliar ou deprimir
funções benéficas exercidas por esses organismos (Harmsen et al., 2000).
Fatores ambientais/geográficos, genéticos e hábitos alimentares durante o
desenvolvimento da criança serão primordiais para definir o tipo e o grau de variabilidade da
colonização em cada indivíduo (Yatsunenku et al, 2012).
A amamentação e o desmame anormais, juntamente com ingestão de água e alimentos
contaminados são, geralmente, considerados como fatores de risco para uma formação
inadequada da microbiota, além de serem os desencadeantes da desnutrição (Motarjemi et al.,
1993; ). Essa disbiose, somada a precariedade na ingestão de alimentos ricos em amido,
ausência de carne (ou outra fonte de proteína), frutas e vegetais, que são fontes importantes de
nutrientes, principalmente vitaminas e íons, leva ao quadro de desnutrição aguda grave
(DAG) e desenvolvimento de kwashiorkor em crianças menores de 5 anos (WHO, 2007).
A DAG é caracterizada quando a medida da circunferência do braço de uma criança
tem uma média inferior à 115mm, entre as idades de 6 meses à, aproximadamente, 5 anos.
Tem sido associada a altos índices de mortalidade, além do risco de desenvolver quadros
diarreicos, pneumonias e infecções sistêmicas, causadas por Staphylococcus aureus,
Escherichia coli, além dos gêneros Streptococcus, Salmonella e Klebsiella (WHO, 2009).
Segundo Williams (1933), kwashiorkor é uma doença nutricional infantil, detectada
principalmente em recém-nascidos. Pode desenvolver-se a partir de uma alimentação ou
amamentação anormal: mães ausentes, levando a criança a ser amamentada por terceiros ou
fórmulas infantis; mães que engravidam novamente, ainda no resguardo; mães doentes ou,
também, desnutridas. Mas, em todos os casos, onde a única suplementação é feita a partir de
alimentos derivados do milho. Sugerindo, portanto, que se trata de uma desnutrição
qualitativa, não quantitativa, devido ao fato do milho não possuir nutrientes como caroteno,
triptofano, lisina e glicina.
Pesquisas feitas por Yatsunenku et al. (2012) analisaram diversas amostras fecais, de
bebês e adultos, em 3 localidades: famílias indígenas, na Venezuela e em Malawi, e famílias
de áreas metropolitanas dos Estados Unidos. A partir da análise, verificaram que bebês
americanos possuíam predominância de Bifidobacterium spp., além de quantidades menores
dos gêneros Bacteroides e Prevotella. Porém, amostras fecais de crianças indígenas possuíam
predominância de bactérias da família Enterobacteriaceae. Já nos adultos, houve predomínio
entre as amostras dos gêneros Prevotella e Bacteroides. Isso induz à probabilidade de que o

28
estilo de vida afete sistematicamente os componentes bacterianos presentes na microbiota
intestinal, mesmo levando em consideração a elevada variabilidade existente entre bebês e
crianças dentro da mesma população.
De acordo com Wu et al. (2011), enterótipos humanos têm sido relacionados a hábitos
alimentares de longo prazo, onde o gênero Prevotella parece estar associado,
preferencialmente, a uma dieta rica em fibras, frutas e vegetais, ao passo que o enterótipo
Bacteroides está ligado à um maior consumo de gordura animal e proteínas. Sendo
relativamente estáveis e resistentes a mudanças de curto prazo.
Diferenças significativas entre as espécies formadoras do microbioma intestinal
adequado interferem na saúde humana, pois, essa alteração se relaciona com a falta da função,
exercida por determinadas espécies chave para a homeostase do metabolismo humano,
acarretando prejuízos na síntese, metabolização e/ou absorção de nutrientes (Million et al.,
2017).
Em condições normais, acredita-se que a microbiota se estabeleça em torno dos 3 anos
de idade e ocorram alterações pouco significativas até a fase adulta (Penders et al., 2006;
Palmer et al., 2007). Um exemplo dessas mudanças comuns relacionadas à idade envolve o
metabolismo das vitaminas B12 (cobalamina) e Ácido Fólico (folato). Em contraste com o
folato, que é sintetizado por micro-organismos e plantas, a cobalamina é produzida
principalmente por germes. O microbioma intestinal de bebês é rico em genes envolvidos na
biossíntese do folato, enquanto a dos adultos tem uma representação significativamente maior
de genes que metabolizam o folato, proveniente da dieta, em sua forma reduzida, o
tetraidrofolato (Krautler, 2005).
Segundo Krautler (2005), ao contrário dos componentes da via biossintética de folato,
que diminuem com a idade, a representação proporcional dos genes que codificam a maioria
das enzimas envolvidas na biossíntese da cobalamina aumenta com a idade.
Indo além dessa correlação homem-microbiota, experimentos feitos com
camundongos estabeleceram um papel causal para a microbiota intestinal na obesidade, a
partir da transferência de um fenótipo obeso, retirados de animais obesos, para camundongos
axênicos não-obesos, por transplante fecal (Backhed et al., 2004; Turnbaugh et al., 2006).
Resultados semelhantes já foram relatados em relação à doença inflamatória intestinal
(Schaubeck et al., 2016) e a propensão a desenvolver doenças hepáticas em camundongos (Le

29
Roy et al., 2013; Llopis et al., 2016), ou estado depressivo em ratos (Kelly Jr et al., 2016).
Nos dois últimos casos, após transplante fecal de doadores humanos.
Indivíduos obesos, com baixa diversidade bacteriana intestinal, são caracterizados pela
maior adiposidade global, resistência à insulina e dislipidemia, e um fenótipo inflamatório
mais pronunciado, quando comparado com indivíduos com microbiota mais diversificada.
Essa baixa variabilidade pode, inclusive, aumentar a propensão ao desenvolvimento de
comorbidades cardiometabólicas relacionadas à inflamação (Le Chatelier et al., 2013).
Para bebês nascidos prematuramente, estratégias nutricionais precoces, que visam
limitar a perda de peso corporal e promover o crescimento, logo nas primeiras horas de vida,
também podem impactar positivamente o neurodesenvolvimento da criança (Schneider e
Garcia-Rodenas, 2017).
2.4 CUIDADOS EXCESSIVOS
2.4.1 Higiene excessiva
A partir da década de 1980 foi possível analisar, em países desenvolvidos, o
aumento progressivo na prevalência de doenças atópicas, como asma, rinite alérgica, alergia
alimentar e dermatite atópica, além de doenças imunomediadas, incluindo diabetes Mellitus
tipo 1 e doença inflamatória intestinal (ISAAC, 1998; Okada et al., 2010).
O Curto período de tempo em que esse aumento pôde ser observado diminui a
probabilidade de que haja um gene mutável como responsável pelo aumento expressivo nos
casos de doenças imunológicas. Com isso, fatores ambientais passaram a ser analisados como
potenciais indutores dessas patogenias (Brooks et al., 2013).
Além disso, ainda segundo Brooks et al, o aumento nos casos desses distúrbios
imunológicos é inversamente proporcional ao número de casos de doenças infecciosas, no
mesmo período de tempo, os quais podem ser atribuídos ao aumento de saneamento básico e
ao avanço da área médica, com o acesso a vacinas e antibióticos. Porém, um estudo
aprofundado sobre os efeitos desses fatores ambientais sugere que a falta de exposição a
agentes infecciosos pode ser o culpado pelo aumento da prevalência de doenças
imunomediadas.
Uma das primeiras observações relacionando agentes infecciosos e desregulação
imune ocorreu no oeste da Nigéria, onde Greenwood observou uma diminuição em novos

30
casos de artrite reumatóide e deduziu que essa baixa incidência podia ser atribuída a
distúrbios imunológicos resultantes da exposição frequente à malária (Greenwood et al.,
1969).
No final dos anos 1970, uma discrepância entre ambientes urbanizados e rurais
emergiu quando Gerrard et al. observaram uma menor prevalência de alergia em populações
indígenas no norte do Canadá, em comparação com populações caucasianas urbanas (Gerrard
et al., 1976).
Em 1989, quando Strachan propôs a hipótese da higiene da doença alérgica,
argumentou que crianças que crescem em famílias maiores têm maior probabilidade de
exposição a micro-organismos, ainda na primeira infância, devido a contatos anti-higiênicos
com irmãos mais velhos, além da exposição pré-natal da mãe, através de sua colonização por
contato familiar similar (Strachan, 1989).
Segundo Strachan (1989), esse aumento da exposição microbiana no início da
vida poderia proteger as crianças do desenvolvimento de hipersensibilidades mais tarde na
vida.
Estudos epidemiológicos adicionais, que apoiam a hipótese da higiene, associam uma
redução na sensibilização a alérgenos com a exposição de animais de estimação, frequência
em creches e um aumento no número de irmãos (Ownby et al., 2002; Benn et al., 2004).
2.4.2 Contato interpessoal
De forma cultural, os bebês humanos nascem, se envolvem e recebem cuidados de
vários indivíduos. A díade mãe-bebê frequentemente existe dentro de redes intersociais,
compostas de indivíduos que ocupam diferentes papéis geracionais, familiares e não
familiares (Meehan et al., 2018). Estudos que quantificam a contribuição de cuidadores
diretos demonstram que esses oferecem uma quantidade considerável de micro-organismos
aos bebês (Meehan et al., 2016).
Em pesquisas a respeito do microbioma do leite humano, Meehan et al. (2018)
analisaram que, as mães que possuíam maior contato interpessoal e que seus bebês recebiam
maior contato direto com terceiros, possuíam maior uniformidade bacteriana no leite materno.
Esses padrões sugerem que diferentes dimensões do ambiente social impactam diretamente na
diversidade microbiana, tanto da mãe quanto do bebê. Pois, segundo Meehan et al., o contato

31
físico com recém-natos faz com que este carreie germes através da cavidade oral até as
glândulas mamárias da mãe, alterando a composição da microbiota presente no leite.
Contudo, Strachan já defende, desde 1989, a hipótese de que o contato anti-higiênico
com irmãos mais velhos propiciaria ao recém-nascido um contato mais facilitado com
bactérias e parasitos importantes para a modulação da colonização inicial da sua microbiota.
2.4.3 Contato com Animais
Segundo pesquisas realizadas por Lappalainen et al (2010), é comum bebês
conviverem com animais de estimação, ainda intraútero. De acordo com eles, das 228 mães
finlandesas, 38% haviam sido sensibilizadas por cães, 19% pela presença de gatos, 11% por
cavalos, 8% por vacas, 4% por coelhos e um total de 1% para porco, ovelha, cabras e aves.
Incluiu, ainda, que o cuidado direto com esses animais era expressivamente maior entre as
famílias rurais, quando comparadas às urbanas.
Diversos estudos sobre a interação com animais sugerem que essa exposição favoreça
o contato com compostos microbianos específicos, os quais se relacionam com o
desenvolvimento imune (Gern et al., 2004; Bufford et al., 2008). Além disso, o tempo de
contato com animais durante os primeiros anos de vida parece desempenhar um papel crucial
(Riedler et al., 2001). Acredita-se também que a exposição materna a ambientes próprios de
animais, além de um número crescente de diferentes animais de criação, já durante a gravidez
, afetam a expressão dos receptores na imunidade inata da criança (Ege et al., 2006).
O efeito das exposições precoces a animais de estimação no desenvolvimento de
alergias e do sistema imunológico é de especial interesse, não apenas por ser comum a
presença de pets junto aos humanos, mas, também, devido a achados conflitantes sugerindo
que animais de estimação podem promover (Melen et al., 2001) ou impedir o
desenvolvimento de doenças alérgicas (Ownby et al; Anyo et al., 2002; Waser et al., 2005).
De acordo com um estudo feito com gatos, por Hesselmar et al (2003), apesar de não
ser clara quais vias imunológicas são ativadas após a exposição a esse animal, foi
demonstrado que a exposição à seus alérgenos na infância pode resultar em produção elevada
de IgG, sem o risco de asma ou sensibilização (resposta hiperestimulada de Th2).
Lappalainen et al puderam concluir em 2010 que a presença de um cão no ambiente
familiar, durante a gravidez, foi significativamente associada a uma diminuição da capacidade
de produzir TNF-α, no nascimento e com 1 ano de idade. Além da hipótese de que a

32
exposição a cavalos, durante a gestação, ser associada à diminuição das respostas do TNF-α
no cordão umbilical.
2.4.4 Privações Ambientais
De acordo com Barbiero (2014), em 2009 o número de habitantes em centros
urbanos chegou a corresponder à metade da população total mundial. Estima-se que até 2050
essa proporção chegue a, aproximadamente, 70%.
No Brasil, o êxodo rural foi marcante entre os anos de 1940 e 1970, onde o
número de habitantes em grandes cidades avançou de 68,8 para 84,3% (Telo e David, 2012).
Por um lado, um maior número de indivíduos puderam ter acesso facilitado a serviços de
saúde, programas vacinais e de saneamento básico, levando ao aumento da expectativa de
vida e diminuição da mortalidade e morbidade causadas por doenças infecciosas (Barreto et
al., 2011).
Por outro lado, diversos programas de atendimento básico à população não
acompanharam esse aumento expressivo da densidade demográfica urbana. A falta de ações
assertivas acarreta, consequentemente, maior dano ambiental, aumento de indivíduos vivendo
em áreas de risco, sob condições precárias de saúde e segurança, além de prejuízos
nutricionais (Barbiero, 2014).
De acordo com Vlahov et al. (2005), essa tendência demográfica, com a crescente
expansão da urbanização, muitas vezes avança sobre áreas rurais que abrigam habitat para
muitas espécies animais, gerando reemergência de doenças causadas por vetores, como
dengue e febre amarela, por exemplo.
Todos esses fatores se relacionam, tanto com a hipótese da higiene da doença
infecciosa, proposta por Strachan em 1989, a qual se baseia no fato de que o acesso a
programas e serviços de saúde leva a diminuição de doenças infecciosas, que seriam comuns
na infância, prejudicando, assim, a modulação do sistema imune, quanto com o que é
proposto por Isolauri (2017) e Stiemsma et al. (2015), que defendem o fato de que a privação
do contato com germes e parasitas na primeira infância impede deflagração do correto
desenvolvimento do sistema imune, o que leva a respostas alérgicas exacerbadas, doenças
autoimunes e inflamações crônicas.

33
Mais recentemente, dois estudos europeus foram reunidos, o primeiro
utilizando análises do polimorfismo de conformação das fitas simples de DNA (Single-strand
conformation polymorphism – SSCP) e, no segundo, técnicas de cultura foram usadas para
identificar exposições microbianas entre crianças europeias. Com isso, os pesquisadores
descobriram que as crianças que cresceram em fazendas na Europa Central encontraram uma
gama mais ampla de exposições microbianas e tiveram uma menor prevalência de asma e
atopia, em comparação ao grupo de referência, crescidos em área urbana (Ege et al., 2011).
O ambiente urbano ao qual o indivíduo está exposto também pode acarretar diferenças
na composição de sua microbiota e, consequentemente, prejuízo em sua função metabólica ou
funcional. Um exemplo pode ser observado em pacientes hospitalizados, os quais apresentam
números reduzidos na diversidade de espécies de alguns gêneros anaeróbicos, como
Bacteroides e Bifidobacterium, e um aumento concomitante de enterobactérias (Bartosch et
al., 2004).
As privações ambientais, portanto, se relacionam com a redução ou impedimento do
contato com micro-organismos que seriam mais facilmente encontrados em áreas rurais, bem
como com manobras relacionadas à saúde, que, muitas vezes, levam a exclusão do contato
com germes ambientais que seriam primordiais para a correta colonização da microbiota
anfibiôntica.
2.4.5 Uso de Antimicrobianos
A antibioticoterapia pode afetar não apenas o patógeno alvo, mas também os
habitantes comensais do hospedeiro humano, levando a um desequilíbrio na microbiota
anfibiôntica intestinal e perda da homeostase, gerando quadros como diarreia associada a
antibióticos, por exemplo (McFarland, 1998).
Foi relatado que os bebês abrigam uma microbiota heterogênea que é propensa a
mudanças de composição causadas por distúrbios ambientais, como o uso de antibióticos
(Palmer et al., 2007; Koenig et al., 2011). Os bebês desenvolvem uma microbiota distinta que
é mais individual e aberta à influência de interferentes ao longo da vida do que a dos adultos
(Arumugam et al., 2011).
A perturbação gerada pelo uso de antimicrobianos manifesta-se principalmente como
diminuição da resistência à colonização de membros da microbiota comensal, o que leva a

34
vários estados de doença (com o uso a curto prazo) (Sullivan et al., 2001), bem como a
emergência de cepas resistentes a antibióticos (uso a longo prazo) (Lindgren et al., 2009).
Se usados de forma indiscriminada durante a infância, podem afetar o crescimento e o
metabolismo da criança ao longo de sua vida. Por estar em fase de estabelecimento e
maturação, a microbiota se encontra particularmente vulnerável à ação de antibióticos e sua
depleção pode levar ao ganho excessivo de peso, ou atraso no desenvolvimento infantil (Cox
e Blaser, 2015).
A mesma interferência também é observada com o uso de medicamentos perinatais,
que podem alterar a microbiota intestinal da mãe (Rautava et al., 2012) e, consequentemente,
vir a alterar a conformação da microbiota do leite materno (Meehan et al., 2018)
2.5 A TEORIA DA HIGIENE E A POSSÍVEL INTERFERÊNCIA NO SISTEMA IMUNE
Nos últimos 150 anos os casos de alergia aumentaram de forma expressiva e sua
prevalência ainda está crescendo. De acordo com Lambrecht e Hammad (2017), “Atualmente,
mais de 30% das crianças são alérgicas, até 10% das crianças sofrem de asma e rinite
alérgica e 5% a 7% das crianças desenvolveram alergia alimentar”.
Isso ocorreu devido à diminuição dos casos de doenças infecciosas, graças ao aumento
de medidas de higiene, uso de medicamentos e acesso a vacinas, diminuição do contato com
animais domésticos e, até mesmo, diminuição de integrantes em uma família. Todos esses
fatores reduziram o contato com bactérias, fungos, vírus e parasitos importantes durante a
primeira infância, como é estudado pela teoria da higiene (Strachan, 1989; Haspeslagh et al.,
2018).
Segundo Isolauri (2017) e Stiemsma et al. (2015), a privação do contato com germes e
parasitas na primeira infância impede o imput necessário ao desenvolvimento correto do
sistema imune, o que leva a respostas exacerbadas – hipersensibilidade –, doenças autoimunes
e inflamações crônicas.
Há evidências de que alguns parasitos e micro-organismos co-evoluíram com o
sistema imunológico humano e que a presença desses organismos são vitais para promover
seu desenvolvimento normal (Stiemsma et al., 2015).
Infecções infantis também foram associadas à diminuição da atopia em crianças. Um
estudo feito por Matricardi et al. (2000) mostrou que pacientes atópicos apresentaram menor

35
prevalência de Toxoplasma gondii , Helicobacter pylori e hepatite A, quando comparados a
controles não-atópicos.
É preciso que haja um controle rígido do sistema imunológico para que o organismo
esteja protegido contra sinais estimuladores exagerados, desencadeados por antígenos
inofensivos, como antígenos próprios e substâncias ambientais. Dependendo da natureza do
antígeno, um desequilíbrio nos mecanismos regulatórios do sistema imune pode levar a
distúrbios autoimunes ou doenças alérgicas em indivíduos geneticamente predispostos
(Calzada et al., 2018).
Hipersensibilidade é um termo utilizado para caracterizar respostas imunes que
ocorrem de forma exagerada ou inapropriada, quando há presença de um antígeno, levando a
reações inflamatórias e/ou dano tecidual (Abbas et al., 2015).
De acordo com Abbas et al (2015), a hipersensibilidade pode ser dividida em quatro
tipos, I, II, III e IV, sendo os três primeiros mediados por anticorpos e, o último, mediado por
células T e macrófagos.
Os distúrbios alérgicos originam-se principalmente das respostas das células Th2, e
um equilíbrio entre as células Th1 e Th2 foi considerado fundamental para a homeostase
imune. A premissa original, portanto, era que altos padrões de higiene resultaram em
diminuição da exposição a microrganismos, que impulsionam a atividade das células Th1,
que, por sua vez, deveriam regular de forma cruzada a resposta das células Th2. Esse
desequilíbrio imunorregulatório, no entanto, não pode explicar o aumento simultâneo da
incidência de vários outros distúrbios imunológicos, como a doença de Crohn, o diabetes tipo
1 e a esclerose múltipla, que são todos principalmente causados por células efetoras Th1.
Notavelmente, a incidência de distúrbios alérgicos (Th2) e diabetes tipo 1 (Th1) correlaciona-
se intimamente dentro e fora da Europa (Stene e Nafstad, 2001).
A incidência de diabetes tipo 1 aumentou dramaticamente, a partir de uma prevalência
muito baixa, na virada do século XIX (Gale, 2002). Na Grã-Bretanha, a prevalência de
diabetes tipo 1 entre crianças menores de 11 anos aumentou de 0,1 para 0,6 por 1.000
crianças, entre 1946 e 1958, e então para 1,3 por 1.000 crianças em 1970, demonstrado em
três estudos feitos por Gale.
A incidência de esclerose múltipla também mostrou um aumento acentuado: na
Alemanha, a incidência de esclerose múltipla dobrou em um período de 15 anos (Poser et al.,
1989). A incidência de doença inflamatória intestinal (DII) na infância, nas Ilhas Britânicas, é
agora quase duas vezes maior do que em 1983 (Sawczenko et al., 2001).

36
Um alto nível de higiene foi repetidamente identificado como um fator de risco para o
desenvolvimento de alergias (Stene e Nafstad, 2001), DII (Gent et al., 1994), diabetes tipo 1
(Marshall et al., 2004) e esclerose múltipla (Ponsonby et al., 2005).
As células linfóides intestinais estão localizadas em três compartimentos: tecido
linfoide associado ao trato gastrointestinal (GALT), a lâmina própria e o epitélio superficial.
A indução e a regulação da imunidade da mucosa ocorrem, principalmente, no GALT e nos
gânglios linfáticos mesentéricos. A diferenciação dos plasmócitos em células B efetoras (de
memória), as quais produzem IgA e IgM, ocorre na lâmina própria, a partir de sinais
secundários fornecidos por células dendríticas, ou células apresentadoras de antígeno (APCs).
O papel da microbiota anfibiôntica é importante neste contexto, pois, as APCs se utilizam
desses micro-organismos para sensibilizar plasmócitos (Brandtzaeg e Johansen, 2005).
Como mostrado pelo fato de que camundongos axênicos só iniciam sua produção de
IgA no intestino após a exposição aos micróbios intestinais convencionais (Crabbé et al.,
1970).
A resposta imune sistêmica, com potencial para dano tecidual e inflamação, ou uma
resposta tolerogênica parece estar diretamente relacionada ao impacto microbiano nas APCs e
células T. As células Th virgens CD4+ são ativadas por APCs (principalmente células
dendríticas) que fornecem sinais coestimulatórios apropriados para diferenciação em células
Th1 ou Th2, com a resultante secreção de citocinas. Sob circunstâncias ainda pouco elusivas,
as APCs também podem induzir vários subconjuntos de células T reguladoras (Treg). Ao
secretar citocinas interleucina (IL) -10 e fator transformador de crescimento (TGF) -β, ou por
interações celulares diretas, as células Tregs podem suprimir as respostas de Th1 e Th2, bem
como a atividade imunológica inata (van Eden et al., 2005).
Esse equilíbrio do sistema imunológico parece depender da relação entre imunidade
inata e adaptativa no início do período neonatal. As APCs também podem, subsequentemente,
ser condicionadas a induzir células Treg por fatores ambientais, como lipídeos da parede
celular de algum parasito (van der Kleij et al., 2002), ou proteínas microbianas de choque
térmico, que têm uma capacidade intrínseca de desencadear eventos imunorregulatórios (van
Eden et al., 2005).
As células da barreira epitelial das vias aéreas inferiores respondem a alérgenos
através de seus receptores celulares específicos. Essa ligação irá estimular a produção de
citocinas pró-inflamatórias, dentre elas a IL-33, que, irá atrair células dendríticas, as quais irão
apresentar esses antígenos aos linfócitos T, induzindo-os na diferenciação em Th2, nos

37
linfonodos. A exposição à endotoxina, poeira ou a colonização microbiana diminuem a
resposta epitelial, aumentando a expressão de reguladores negativos. Essa liberação epitelial
de IL-33 também é inibida por produtos secretados derivados de helmintos (Gollwitzer et al.,
2014).
3 DISCUSSÃO
O corpo humano é colonizado por trilhões de micro-organismos, os quais habitam
diversas regiões anatômicas e contribuem para a nossa saúde e bem-estar. Variações
consideráveis na composição da microbiota têm sido descritas entre os indivíduos, por
exemplo, pelo Projeto de Microbioma Humano, nos EUA (HMP), e usadas para auxiliar na
identificação de padrões de composição que norteiam estados de saúde e doença, dentro das
populações (Huttenhower et al., 2012).
A vantagem da classificação baseada em padrões de composição é a potencialização
de diagnósticos baseados em microbiota, terapias ou prevenção de doenças, com implicações
para o tratamento personalizado, por meio de intervenções nutricionais, microbianas e
farmacêuticas (Costea et al., 2018).
O ecossistema microbiano apresenta uma sucessão de diferentes estágios da
microbiota: a composição da comunidade muda rapidamente na primeira infância, se
estabiliza em adultos e se deteriora na velhice (Falony et al., 2016).
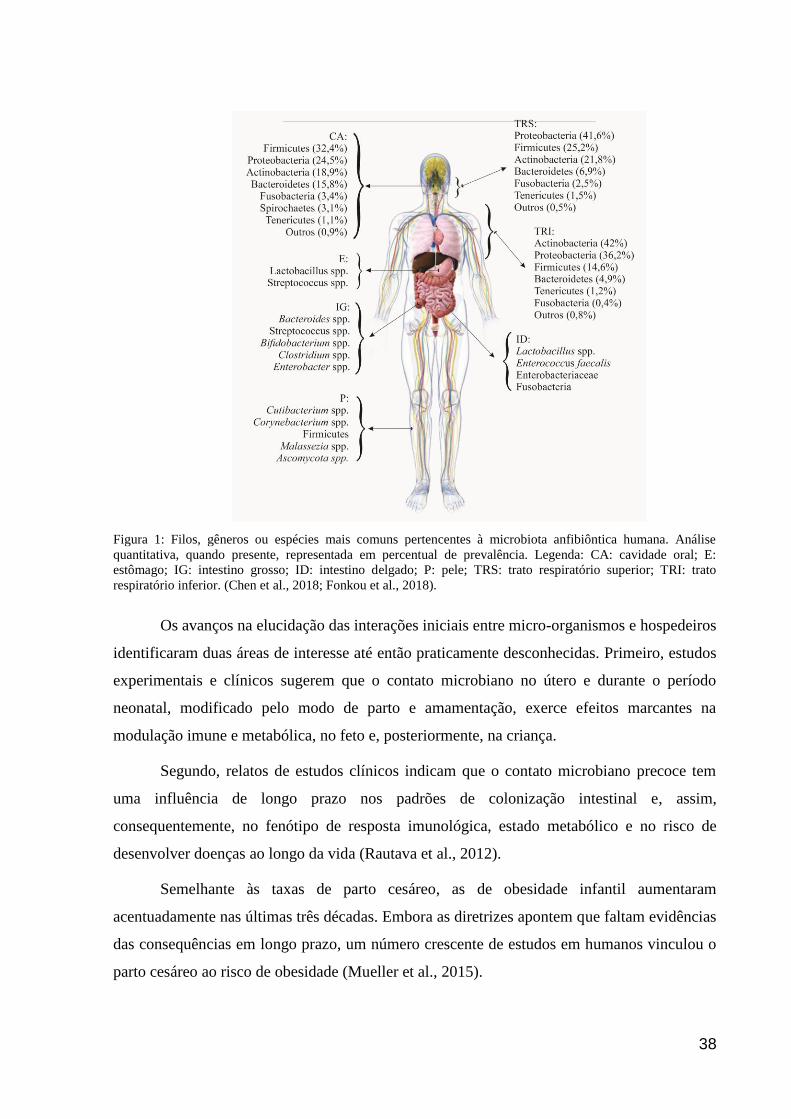
38
Figura 1: Filos, gêneros ou espécies mais comuns pertencentes à microbiota anfibiôntica humana. Análise
quantitativa, quando presente, representada em percentual de prevalência. Legenda: CA: cavidade oral; E:
estômago; IG: intestino grosso; ID: intestino delgado; P: pele; TRS: trato respiratório superior; TRI: trato
respiratório inferior. (Chen et al., 2018; Fonkou et al., 2018).
Os avanços na elucidação das interações iniciais entre micro-organismos e hospedeiros
identificaram duas áreas de interesse até então praticamente desconhecidas. Primeiro, estudos
experimentais e clínicos sugerem que o contato microbiano no útero e durante o período
neonatal, modificado pelo modo de parto e amamentação, exerce efeitos marcantes na
modulação imune e metabólica, no feto e, posteriormente, na criança.
Segundo, relatos de estudos clínicos indicam que o contato microbiano precoce tem
uma influência de longo prazo nos padrões de colonização intestinal e, assim,
consequentemente, no fenótipo de resposta imunológica, estado metabólico e no risco de
desenvolver doenças ao longo da vida (Rautava et al., 2012).
Semelhante às taxas de parto cesáreo, as de obesidade infantil aumentaram
acentuadamente nas últimas três décadas. Embora as diretrizes apontem que faltam evidências
das consequências em longo prazo, um número crescente de estudos em humanos vinculou o
parto cesáreo ao risco de obesidade (Mueller et al., 2015).

39
Composições atípicas da microbiota estão cada vez mais associadas a patologias e, no
caso da obesidade, a correlação é tão intensa que o microbioma intestinal pode ser utilizado
como ferramenta diagnóstica, superando biomarcadores humanos da doença (Le Chatelier et
al., 2013). Para a doença de Crohn e a cirrose, foram propostos índices de disbiose da
microbiota que se correlacionam com a progressão da doença (Bajaj et al., 2014; Gevers et al.,
2014). Para outras doenças, além da obesidade, como a resistência à insulina, doença
inflamatória intestinal e depressão, a associação foi além dessa correlação, tendo efeitos
causais estabelecidos em modelos animais (van de Guchte et al., 2018).
Não é necessário haver grandes alterações para que ocorram perdas de função.
Mudanças discretas também estão ligadas a diferentes estados da saúde humana, sejam por
diferentes doenças manifestadas ou pelo conjunto de parâmetros fisiológicos e imunológicos
que possam indicar uma propensão ao desenvolvimento da doença, como o alerta para um
“estado pré-doença”. Como, por exemplo, a predominância do gênero Bacteroides na
microbiota intestinal, associada ao aumento de linfócitos e níveis de proteína C-reativa
(Costea et al., 2018).
Ainda que se pense que a microbiota apresenta diferentes composições, através das
inúmeras fases da vida, desempenhando, igualmente, diferentes funções em cada uma delas,
não se relaciona ao fato das disbioses estarem associadas a doenças inflamatórias crônicas ou
a um estado pré-doença. Alterações relacionadas a patologias são casos atípicos e não fazem
parte da sucessão habitual das composições do microbioma humano, da infância até a idade
adulta (Kundu et al., 2017).
Fato que leva a crer, portanto, que as diferenças na função exercida pelos micro-
organismos podem indicar alterações na simbiose microbiota-hospedeiro, após a idade de
estabelecimento e maturação do microbioma anfibiôntico – 1 a 3 anos – e em situações em
que haja pressão exercida por fatores perturbadores, como antibioticoterapia, dieta a longo
prazo, predisposição genética, doenças crônicas e fatores ambientais (Le Chatelier et al.,
2013; Mueller et al., 2015).
Quando um sistema é levado ao seu limite de saturação, por movimentos estocásticos,
perturbações, mudanças de condições ou uma combinação desses fatores, pode alcançar um
“ponto de inflexão”, de onde pode facilmente avançar a um estado diferente. Essa transição de
estado também é chamada de “transição crítica”, o que significa que restaurar as condições
para aquelas que existiam antes da mudança não é suficiente para fazer o sistema retornar ao

40
seu estado original. O estado original e o novo são, portanto, estados estáveis alternativos,
diferentes, que podem coexistir sob as mesmas condições externas (Scheffer et al., 2001).
Figura 2: Alterações na microbiota, promovendo a ruptura da simbiose entre microbiota-hospedeiro, imposta por
fatores como hábitos alimentares, uso de antimicrobianos e doenças inflamatórias. A constante perturbação leva
o estado simbiótico ao estado crítico de transição, até a perda do mutualismo, com baixa diversidade entre as
espécies da microbiota, estresse oxidativo, permeabilidade aumentada e inflamações crônicas. (Adaptado de van
de Guchte et al., 2018).
A colonização tardia e alterada da microbiota gastrointestinal, seja por parto cesáreo
ou fatores descritos pela hipótese da higiene, tem sido sugerida como um possível fator
etiológico de diversas patologias, entre elas asma, doenças atópicas e alergias alimentares
(Thavagnanam et al., 2008; Bager et al., 2008).
De acordo com Schlinzig et al., (2017), isso ocorre principalmente em bebês que são
privados do contato com a microflora vaginal da mãe, no momento do parto, pois nascem
através de cesariana, levando ao atraso ou alteração na modulação do sistema imune da
criança.
Em estudos feitos por Van Nimwegen et al. (2011) foi possível descobrir que o parto
cesáreo está fortemente associado a colonização por Clostridium difficile, que aumenta o risco
de desenvolvimento de asma, eczema e sensibilização a alérgenos alimentares na infância.
Diversas alterações na composição da microbiota, a partir da diferença entre tipos de
parto, já puderam ser observadas através de estudos utilizando análise fecal. Como, por
exemplo, em um estudo feito por Stokholm et al., (2016), onde a partir da análise de amostras
de 700 crianças, nascidas por parto normal e cesárea, puderam diferenciar o padrão de
colonização intestinal entre elas, chegando a conclusão preliminar de que partos cesariana

41
geravam um padrão precoce de colonização no TGI dos bebês (gráficos 1 e 2), que, inclusive,
poderiam ser associados a fatores de risco para diversas doenças imunomediadas na criança.
Gráfico 1: Microbiota de bebês de 1 mês de vida, nascidos por cesárea, sem amamentação exclusiva,
apresentando padrões desordenados de colonização. (Stoholm et al., 2016).
Gráfico 2: Microbiota intestinal de crianças com 1 ano de idade, nascidas a partir de parto cesáreo, sem
amamentação exclusiva, mostrando a transição quantitativa de colonização intestinal, porém não
significativamente qualitativa (Stokholm et al., 2016).
0
50
100
150
200
250
300
1 mês
0
100
200
300
400
500
600
1 ano

42
Além de pesquisas realizadas com o intuito de avaliar o impacto das vias de parto para
a criança, já foi possível identificar proporções semelhantes para a influência exercida pelo
leite materno sobre a colonização e proteção intestinal da criança (Harmsen et al., 2000; Knol
et al., 2005; Palmer et al., 2007; Roger et al., 2010; DeWeerdt, 2018). Porém, essas alterações,
mediadas pelo tipo de amamentação/alimentação à qual a criança está sujeita, devem ser
aprofundadas para se excluir interferentes, como o uso de antimicrobianos (durante a gestação
ou perinatais), tipo de microbiota presente na pele da mãe, bem como seus hábitos de vida e
alimentação (Biagi et al., 2017, 2018).
Bebês com prematuridade moderada a grave (abaixo de 34 semanas), não conseguem
amamentar por não possuírem os movimentos de sucção, deglutição e respiração
coordenados, por se tratar de um desenvolvimento que ocorre, intrautero, após esse período,
levando-os à alimentação artificial (Mizuno e Ueda, 2003).
Esse fator fisiológico faz com que o bebê necessite de métodos alternativos e
sintéticos de alimentação, interferindo ou modificando os padrões de colonização intestinal da
criança, como pode ser observado na tabela 1.
Micro-organismo Bebês alimentados
com leite materno
Bebês alimentados
por fórmulas
Bifidobacterium spp. x
Lactobacillus spp. x x
Bacteroides spp. x
Staphylococcys spp. x
Streptococcus spp. x x
Clostridium difficile x
Enterobacteriaceae x
Tabela 1: Micro-organismos mais prevalentes na microbiota de bebês alimentados exclusivamente por
leite materno e alimentados somente por fórmulas infantis, mostrando a variação na colonização e o déficit de
espécies benéficas à saúde da criança (Penders et al., 2006; Roger et al., 2010).
Além disso, a hipótese da higiene, proposta por Strachan, em 1989, estipula que a
composição da microbiota intestinal influencia a tolerância oral e a resposta imune a
alérgenos em crianças (Strachan, 1989). A partir dessa hipótese, foi possível identificar uma

43
relação entre a diminuição de doenças infecciosas (gráficos 3-5) e o aumento de doenças
imunológicas, que, segundo Strachan, estaria relacionada à diminuição ou perda do contato
com micro-organismos importantes durante a infância.
Acredita-se que o rápido aumento da asma e de outros distúrbios atópicos no mundo
industrializado tenha começado entre os anos 1960 e 1970, com aumentos progressivos
durante os anos 80 e 90 (Isolauri et al., 2004). A prevalência de asma, por exemplo, aumentou
cerca de 1% ao ano em torno de 1980 (Wamer, 1999) (Figura 3). Estudos recentes do Reino
Unido confirmam que a asma atópica representa um aumento preponderante em relação à
asma não-atópica, além da comparação feita entre dois espaços de tempo distintos, com
crianças nascidas em 1958 e 1970, em que foi possível constatar aumentos de 3,1 a 6,4% para
o eczema e de 12 a 23,3% para a rinite alérgica (febre do feno), aos 16 anos de idade
(Bloomfeld et al., 2006).
Gráfico 3: Incidência de três doenças infecciosas infantis (DII) no Brasil, de 1978 à 2016: Coqueluche, Rubéola
e Poliomielite, para corroborar a hipótese da higiene de doença infecciosa proposta por Strachan (1989). Trata-se
de um quantitativo percentual em relação à incidência da doença a cada 100 mil habitantes (Centers for Desease
Control and Prevention (CDC).
0
5
10
15
20
1978 1980 1993 1995 1998 2000 2002 2004 2006 2008 2010 2012 2014 2016
Doenças infecciosas infantis
Coqueluche Rubéola Poliomielite
Co
ef. i
nci
dên
cia/
10
0.00
0 h
ab

44
0
0.2
0.4
0.6
0.8
20
01
20
02
20
03
20
10
20
11
20
12
20
13
DII
Meningitepor HIB
0
0.1
0.2
0.3
0.4
0.5
DII
Difteria
Gráficos 4 e 5: Incidências de Difteria e Meningite por HIB (Haemophilus influenzae tipo b) no Brasil,
respectivamente, de 1990 a 2016, para corroborar a hipótese da higiene de doença infecciosa proposta por
Strachan (1989). (DII: Doença Infecciosa Infantil). (Centers for Desease Control and Prevention (CDC).
Figura 3: Representação do aumento de casos de asma no mundo, onde o Brasil ocupa o 8º lugar, com 20% de
crianças doentes. (The International Study of Asthma and Allergies in Childhood – ISAAC).
Porém, um exame mais detalhado dos mecanismos imunológicos por trás da hipótese
de higiene de Strachan para doenças alérgicas pode aumentar a conexão entre exposições
infecciosas no início da vida e o desenvolvimento de tolerância imunológica (Bloomfeld et
al., 2006).
Os mecanismos imunológicos subjacentes para explicar como as exposições
microbianas podem influenciar o desenvolvimento de funções imunológicas e distúrbios
alérgicos ainda são pouco compreendidos, mas inicialmente foi descrito que a diminuição da
exposição a micróbios e seus componentes no início da vida poderia favorecer a manutenção
de respostas pós-natal do tipo Th2, por falha na diferenciação de Th1 (Martinez e Holt, 1999).

45
Indo além da hipótese da higiene, segundo Lappalainen et al. (2010), elucidar
interações entre exposições ambientais e a constituição genética da criança facilitariam a
avaliação da real influência que um determinado estímulo ambiental exerce sobre a mesma,
pois exposição iguais podem levar a respostas distintas, como aumentar ou diminuir (ou não
influenciar) o risco de asma ou sensibilização alérgica na criança, dependendo de sua
suscetibilidade genética.
Por outro lado, a exposição intensiva a longo prazo à endotoxina presente no poeira
doméstica parece estar associada a respostas reguladas por citocinas entre crianças expostas.
(Lappalainen et al., 2010).
Em estudos anteriores, Lappalainen et al (2010) detectaram uma associação entre a
endotoxina do pó doméstico e a capacidade de produção elevada de IFN- γ, desde o
nascimento até os 3 meses de idade. Porém, bebês nascidos em área rural, má limpeza
doméstica ou a presença de um cão ou gato também parecia aumentar, igualmente, os níveis
de IFN- γ, não sendo possível assegurar qual dessas variáveis de exposição foi melhor
relacionada a produção de IFN-γ.
Levando em consideração as respostas imunológicas deflagradas pela criança,
Stiemsma et al. (2015) puderam concluir que não apenas as bactérias seriam importantes para
imput da modulação do sistema imune, mas como, também, a presença de parasitos ainda na
primeira infância.
Embora os helmintos sejam indutores prototípicos da imunidade do tipo II, eles têm
sido correlacionados com a redução da reatividade do teste cutâneo de alérgenos e, até certo
ponto, com a proteção da asma (Cruz et al., 2017). Uma explicação geral para essa associação
é a de que os helmintos induzem a chamada resposta Th2 modificada, com células
imunorreguladoras, como Tregs, complementando a atividade de Th2 e regulando a resposta a
antígenos ambientais, como aeroalérgenos. Com isso, vários grupos tentaram encontrar
produtos derivados de helmintos com propriedades imunomoduladoras que poderiam ser
usados para suprimir a imunidade mediada por Th2. (Navarro et al., 2017).
O impacto de muitas covariáveis na composição e estabelecimento do microbioma
infantil tem sido amplamente explorado, indicando que a idade gestacional, o tipo de parto e o
tipo de nutrição – leite materno, leite de doador ou fórmulas infantis – estar entre as variáveis
de maior impacto (Chu et al., 2017).

46
Recentemente, evidências mostrando a importância do ambiente social e físico em
torno da díade mãe-bebê (por exemplo, quantas pessoas cuidam do bebê e com que
frequência) na determinação da composição da microbiota do leite foram demonstradas
(Meehan et al., 2018). Nesses casos, a microbiota intestinal está sendo cada vez mais
reconhecida como um fator importante que conecta os genes, o meio ambiente e o sistema
imunológico infantil (Musso et al., 2010).
Essas covariáveis podem relacionar, inclusive, o aumento do número de partos cesárea
com o desenvolvimento da obesidade (gráfico 6), apesar de tratar-se de um processo
complexo, envolvendo suscetibilidade genética e fatores ambientais, os quais permanecem
apenas parcialmente compreendidos. Segundo Huh et al. (2012), após estudos feitos com
crianças obesas e não-obesas, puderam identificar diferenças no padrão de colonização
intestinal nas que haviam nascido por parto cesárea, as quais se enquadravam como fatores
causais da obesidade. Em conjunto, esses achados sugerem a existência de uma complexa – e
ainda parcialmente inexplorada – conexão entre os microbiomas materno, infantil e ambiental
no estabelecimento de microbiomas infantis que minimizem a suscetibilidade ao
desenvolvimento de doenças (Biagi et al., 2018).
Em estudos feitos por Musso et al. (2010), foi possível descrever que animais
submetidos à obesidade induzida por alimentação, apresentavam quantitativo
consideravelmente maior do filo Firmicutes em relação ao filo Bacteroidetes. Porém,
retirando a interferência da dieta, os padrões microbianos voltavam à sua normalidade,
demonstrando mais assertivamente que a composição alterada da microbiota intestinal é uma
causa, não uma consequência da obesidade, ou hábitos alimentares alterados.
Além disso, outros mecanismos que ligam a microbiota intestinal à obesidade foram
posteriormente ligados à endotoxinemia crônica de baixo grau, a qual pode induzir o
desenvolvimento de diabetes Mellitus tipo 2. Esse fato foi elucidado a partir de experimentos
animais, onde ratos alimentados com dieta hiperlipídica gerou ativação de macrófagos no
fígado, que atacam células de Kupffer, induzindo a resistência à insulina e a intolerância à
glicose (Neyrinck et al., 2009)
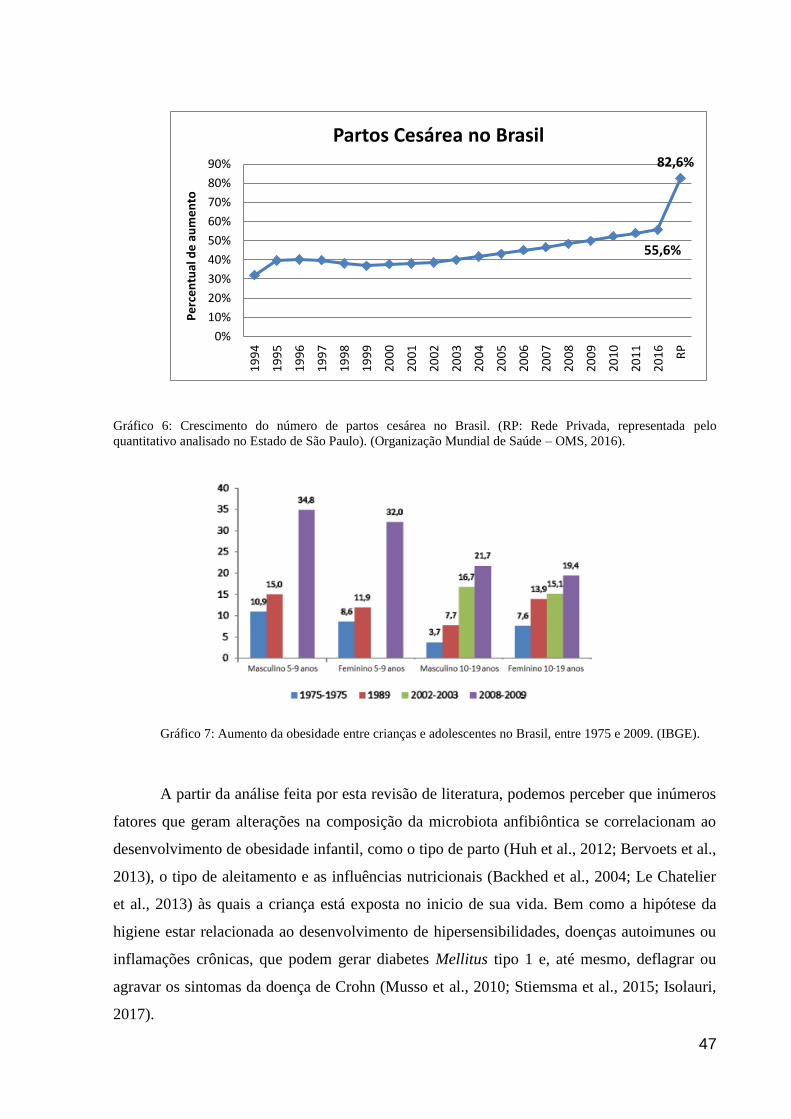
47
Gráfico 6: Crescimento do número de partos cesárea no Brasil. (RP: Rede Privada, representada pelo
quantitativo analisado no Estado de São Paulo). (Organização Mundial de Saúde – OMS, 2016).
Gráfico 7: Aumento da obesidade entre crianças e adolescentes no Brasil, entre 1975 e 2009. (IBGE).
A partir da análise feita por esta revisão de literatura, podemos perceber que inúmeros
fatores que geram alterações na composição da microbiota anfibiôntica se correlacionam ao
desenvolvimento de obesidade infantil, como o tipo de parto (Huh et al., 2012; Bervoets et al.,
2013), o tipo de aleitamento e as influências nutricionais (Backhed et al., 2004; Le Chatelier
et al., 2013) às quais a criança está exposta no inicio de sua vida. Bem como a hipótese da
higiene estar relacionada ao desenvolvimento de hipersensibilidades, doenças autoimunes ou
inflamações crônicas, que podem gerar diabetes Mellitus tipo 1 e, até mesmo, deflagrar ou
agravar os sintomas da doença de Crohn (Musso et al., 2010; Stiemsma et al., 2015; Isolauri,
2017).
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
19
94
19
95
19
96
19
97
19
98
19
99
20
00
20
01
20
02
20
03
20
04
20
05
20
06
20
07
20
08
20
09
20
10
20
11
20
16
RP
Pe
rce
ntu
al d
e a
um
en
to
Partos Cesárea no Brasil82,6%
55,6%

48
4 CONCLUSÃO
A partir da análise feita por esse estudo, podemos concluir, portanto, que alterações
precoces na progressão de eventos que levam à montagem de um microbioma intestinal
saudável podem predispor a várias doenças não transmissíveis na infância, como enterocolite
necrosante, asma, doença atópica, hipersensibilidades, doenças autoimunes, obesidade,
diabetes Mellitus, doença de Crohn, além de distúrbios no neurodesenvolvimento da criança.
Diversos fatores foram enquadrados como agentes perturbadores, assim como vários
mecanismos foram elucidados como sendo de extrema importância para a manutenção e
progressão da saúde infantil.
Embora com efeitos até agora pouco compreendidos sobre a imunologia infantil e a
composição microbiana do leite materno, o parto vaginal parece fornecer não apenas uma
exposição bacteriana substancial, importante nas primeiras horas de vida do bebê, mas,
também, estimular interações saudáveis entre microbiota-hospedeiro e, consequentemente,
modular o desenvolvimento imunológico da criança.
Vimos que o estabelecimento da microbiota anfibiôntica, tanto no período perinatal,
quanto na primeira infância, é tão importante que pode, até, ser considerado a última fase da
organogênese, pois afeta diretamente a fisiologia gastrointestinal e sistêmica do recém-
nascido.
Da mesma forma, a amamentação pode ser vista como uma extensão dos padrões de
colonização materno-infantil, que proporciona condições favoráveis à estabilização das
interações microbiota-criança, necessárias durante o início da vida. Já foi possível, por
exemplo, relacionar componentes presentes no leite materno à interações complexas com
micro-organismos simbióticos, os quais levam ao desenvolvimento da tolerância oral aos
antígenos alimentares.
Trata-se, portanto, de um estudo que deve ser contínuo, para que leve à elucidação real
dos fatores perturbadores da formação adequada da microbiota anfibiôntica, moldando a
compreensão da importância do contato microbiano durante os períodos frágeis da vida fetal,
do parto e da infância, para que uma programação metabólica e imunológica saudável seja
estabelecida, criando novas oportunidades para melhorar a saúde infantil e reduzir o risco de
doenças ao longo da vida adulta.

49
BIBLIOGRAFIA
A
Aagaard, K. et al. The placenta harbors a unique microbiome. Sci. Transl. Med. 2014, vol. 6,
pp. 237-65.
Aas, J. A. et al. Defining the normal bacterial flora of the oral cavity. J. Clin.
Microbiol. 2005, vol. 43, pp. 5721–5732.
Abbas, A. K., Lichtman, A. H., Pillai, S. Imunologia Celular e Molecular. Tradução da 8º
Edição. Elsevier. 2015.
Alpan, O. Oral tolerance and gut-oriented immune response to dietary proteins. Curr Allergy
Asthma Rep. 2001, vol. 1, pp. 572–577.
American Society for Microbiology: Pasteur-Koch: distinctive ways of thinking about
infectious diseases. Disponível em:
<https://www.asm.org/index.php/component/content/article/114-unknown/unknown/ 4469-
pasteur-koch-distinctive-ways-of-thinking-about-infectious-diseases>. Acesso em 30 set.
2018.
Andrade, A. Microflora intestinal: uma barreira imunológica desconhecida. Dissertação de
Mestrado Integrado em Medicina - Instituto de Ciências Biomédicas Abel Salazar na
Universidade do Porto, Porto, 2009/2010.
Anyo, G. et al. Early, current and past pet ownership: associations with sensitization,
bronchial responsiveness and allergic symptoms in school children. Clin Exp Allergy 2002,
vol. 32, pp. 361–6.
Arumugam, M. et al. Enterotypes of the human gut microbiome. Nature 2011, vol. 473, pp.
174–180.
Atarashi, K. et al. Induction of colonic regulatory T cells by
indigenous Clostridium species. Science. 2011, vol. 331(6015), pp. 337–341.
Azad, M. et al. Impact of maternal intrapartum antibiotics, method of birth and breastfeeding
on gut microbiota during the first year of life: a prospective cohort study. BJOG. 2015.
B
Backhed, F. et al. The gut microbiota as an environmental factor that regulates fat
storage. Proc Natl Acad Sci USA. 2004, vol. 101(44), pp. 15718–15723.
Bager, P.; Wohlfahrt, J.; Westergaard, T. Caesarean delivery and risk of atopy and allergic
disease: meta-analyses. Clin Exp Allergy. 2008, vol. 38, pp. 634–642.
Bajaj, J. S. et al. Altered profile of human gut microbiome is associated with cirrhosis and its
complications. J Hepatol. 2014, vol. 60(5), pp. 940–947.
Bakhtiar, S. M. et al. Implications of the human microbiome in inflammatory bowel diseases.
FEMS Microbiology Letters. 2013, vol. 342, pp. 7-10.

50
Barbiero, V. K. Urban health: it's time to get moving!. Glob Health Sci Pract. 2014, vol. 2,
n.2, pp.139-44.
Barbosa, F. et al. Microbiota indígena do trato gastrintestinal. Revista de Biologia e Ciência
da Terra, Aracaju. 2010, vol. 10, n. 1, p. 78-93.
Barreto, M. L. et al. Successes and failures in the control of infectious diseases in Brazil:
social and environmental context, policies, interventions, and research needs. Lancet. 2011,
vol. 377, pp. 1877-86.
Bartosch, S. et al. Characterization of bacterial communities in feces from healthy elderly
volunteers and hospitalized elderly patients by using real-time PCR and effects of antibiotic
treatment on the fecal microbiota. Appl Environ Microbiol. 2004, vol. 70, pp. 3575–3581.
Benn, C. S. et al. Cohort study of sibling effect, infectious diseases, and risk of atopic
dermatitis during first 18 months of life. BMJ. 2004, vol. 328(7450):1223.
Bernstein, C. N.; Shanahan, F. Disorders of a modern lifestyle-reconciling the epidemiology
of inflammatory bowel diseases. Gut. 2008, vol. 57, pp. 1185-1191.
Bervoets, L. et al. Differences in gut microbiota composition between obese and lean
children: a cross-sectional study. Gut Pathog. 2013, vol. 5:10.
Biasucci, G. et al. Cesarean delivery may affect hearly biodiversity of intestinal bactéria. The
Journal of Nutrition. 2008, vol. 138, pp. 1796S-1800S.
Björkström, M. V. et al. Intestinal flora in very low-birth weight infants. Acta Paediatr. 2009,
vol. 98, pp.1762–1767.
Bourlioux, P. et al. The intestine and its microflora are partners for the protection of the host:
report on the Danone Symposium “The Intelligent Intestine”, Am. J. Clin. Nutr. 2003, vol. 78,
pp. 675-683.
Brandtzaeg, P. and Johansen, F-E. Mucosal B cells: phenotypic characteristics, transcriptional
regulation, and homing properties. Immunol Rev. 2005, vol. 206, pp. 32–63
Brooks, C.; Pearce, N.; Douwes, J. The hygiene hypothesis in allergy and asthma: an
update. Curr Opin Allergy Clin Immunol. 2013, vol. 13(1), pp. 70–77.
Bufford, J. D. et al. Effects of dog ownership in early childhood on immune development and
atopic diseases. Clin Exp Allergy 2008, vol. 38, pp. 1635–43.
Butel, M. J. et al. Conditions of bifidobacterial colonization in preterm infants: a prospective
analysis. J. Pediatr. Gastroenterol. Nutr. 2007, vol. 44, pp. 577–582.
C
Calzada, D. et al. Immunological Mechanisms in Allergic Diseases and Allergen Tolerance:
The Role of Treg Cells. Journal of Immunology Research. 2018, vol. 2018, Article ID
6012053, 10 pages.

51
Cândido, A. L.; Tunon, I. L.; Carneiro, M. R. P. Microbiologia Geral. São Cristovão:
Universidade Federal de Sergipe, CESAD, 2009. Disponível em:
<https://www.passeidireto.com/arquivo/31130324/microbiologia-geral-aula-4-cesad-ufs>.
Acesso em: 3 out 2018.
Carrier, T. J.; Reitzel, A. M. The hologenome across environments and the implications of a
host-associated microbial repertoire. Front Microbiol. 2017, vol. 8:802.
Chang, J. Y. et al. Pyrosequencing-based molecular monitoring of the intestinal bacterial
colonization in preterm infants. J. Pediatr. Gastroenterol. Nutr. 2011, vol. 53, pp. 512–519.
Chu, D. M. et al. Maturation of the infant microbiome community structure and function
across multiple body sites and in relation to mode of delivery. Nat Med. 2017, vol. 23, pp.
314–326.
Consortium THMP. Structure, function and diversity of the healthy human
microbiome. Nature. 2012, vol. 486(7402), pp. 207–214.
Corr, S. C. et al. Bacteriocin production as a mechanism for the antiinfective activity
of Lactobacillus salivarius UCC118. Proc Natl Acad Sci U S A. 2007, vol. 104, pp. 7617–
7621.
Costea, P. I. et al. Enterotypes in the landscape of gut microbial community composition. Nat
Microbiol. 2018, vol. 3(1), pp. 8–16.
Cox, L. M.; Blaser, M. J. Antibiotics in early life and obesity. Nat. Rev. Endocrinol. 2015,
vol. 11(3), pp. 182-190.
Crabbé, P. A. et al. Immunohistochemical observations on lymphoid tissues from
conventional and germ-free mice. Lab Invest. 1970, vol. 22, pp. 448–457.
Cruz, A. A. Global issues in allergy and immunology: parasitic infections and allergy. J
Allergy Clin Immunol. 2017, vol. 140, pp. 1217-1228.
D
DeWeerdt, S. How baby’s first microbes could be crucial to future health. Nature. 2018, vol.
555(7695), pp. S18–S19.
E
Ege, M. J. et al. Prenatal farm exposure is related to the expression of receptors of the innate
immunity and to atopic sensitization in school-age children. J Allergy Clin Immunol 2006,
vol. 117, pp. 817–23.
Ege M. J. et al. Exposure to environmental microorganisms and childhood asthma. N Engl J
Med. 2011, vol. 364(8), pp. 701–709.

52
F
Falony, G. et al. Population-level analysis of gut microbiome variation. Science. 2016, vol.
352(80), pp. 560–564.
Fardini, Y. et al. Transmission of diverse oral bacteria to murine placenta: evidence for the
oral microbiome as a potential source of intrauterine infection. Infect Immun. 2010, vol. 78,
pp. 1789–1796.
Fiocchi, C. e Pereira de Sousa, H.S. Microbiota Intestinal - Sua importância e função. Jornal
Brasileiro de Medicina. 2012, vol. 100, pp. 30-38.
Fonkou, M. D. et al. Repertoire of bacterial species cultured from the human oral cavity and
respiratory tract. Future Microbiology. 2018.
Fredricks, D. N.; Fiedler, T. L.; Marrazzo, J. M. Molecular identification of bacteria
associated with bacterial vaginosis. The New England journal of medicine. 2005, vol. 353, pp.
1899–1911.
G
Gale, C. et al. Effect of breastfeeding compared to formula feeding on infant body
composition: a systematic review and meta-analysis. Am J Clin Nutr. 2012, vol. 95, pp. 656-
669.
Gale, E. A. M. The rise of childhood type I diabetes in the 20th century. Diabetes. 2002, vol.
51, pp. 3353–3361.
Gent, A. E. et al. Inflammatory bowel disease and domestic hygiene in infancy. Lancet. 1994,
vol. 343, pp. 766–767.
Gern, J. E. et al. Effects of dog ownership and genotype on immune development and atopy in
infancy. J Allergy Clin Immunol 2004, vol. 113, pp. 307–14.
Gerrard, J. W. et al. Serum IgE levels in white and metis communities in Saskatchewan. Ann
Allergy. 1976, vol. 37(2), pp. 91–100.
Gevers, D. et al. The treatment-naive microbiome in new-onset Crohn’s disease. Cell Host
Microbe. 2014, vol. 15(3), pp. 382–392.
Goldenberg RL, et al. Intrauterine infection and preterm delivery. N Engl J Med. 2000, vol.
342, pp. 1500–1507.
Goldenberg, R. L. et al. Epidemiology and causes of preterm birth. Lancet. 2008, vol. 371, pp.
75–84.
Gollwitzer, E. S. et al. Lung microbiota promotes tolerance to allergens in neonates via PD-
L1. Nat Med, 2014, vol. 20, pp. 642-647.
Gonçalves, M. A. P. Microbiota – implicações na imunidade e no metabolismo. Dissertação
de Mestrado em Ciências Farmacêuticas. Universidade Fernando Pessoa, Porto, Portugal,

53
2014, 53p. Disponível em: <https://bdigital.ufp.pt/bitstream/10284/4516/1/PPG_21951.pdf>.
Acesso em: 30 set 2018.
Gordon, J. et al. Superorganisms and holobionts. Microbe. 2013, vol. 8(4), pp. 152–153.
Greenwood, B. M. Polyarthritis in Western Nigeria. I. Rheumatoid arthritis. Ann Rheum Dis.
1969, vol. 28(5), pp. 488–496.
Gronlund, M. M. et al. Maternal breast-milk and intestinal bifidobacteria guide the
compositional development of the Bifidobacterium microbiota in infants at risk of allergic
disease. Clin Exp Allergy. 2007, vol. 37, pp. 1764–1772.
Guarner, F. Papel de la flora intestinal en la salud y em la enfermedad. Nutrición Hospitalaria,
Madrid, 2007, vol. 22, n. 2, pp. 14-19.
Guarner, F.; Malagelada Jr, R. Gut flora in health and disease. Lancet, London. 2003, vol. 8,
n. 361, pp. 512- 519.
H
Håkansson, S. et al. Group B streptococcal carriage in Sweden: a national study on risk
factors for mother and infant colonization. Acta Obstet Gynecol Scand. 2008, vol. 87, pp. 50–
58
Harmsen, H. J. et al. Analysis of intestinal flora development in breast-fed and formula-fed
infants by using molecular identification and detection methods. J. Pediatr. Gastroenterol.
Nutr. 2000, vol.30, pp.61–67.
Haspeslagh, E. et al. The hygiene hypothesis: immunological mechanisms of airway
tolerance. Current Opinion in Immunology. 2018, vol. 54, pp. 102-108.
Hesselmar, B. et al. High-dose exposure to cat is associated with clinical tolerance – a
modified Th2 immune response? Clin Exp Allergy. 2003, vol. 33, pp. 1681–5.
Huh, S. Y. et al. Delivery by caesarean section and risk of obesity in preschool age children: a
prospective cohort study. Arch Dis Child. 2012, vol. 97(7), pp. 610-6.
Huttenhower, C. et al. Structure, function and diversity of the healthy human
microbiome. Nature. 2012, vol. 486, pp. 207–214.
I
Innis, S. M. Human milk: lipids from maternal diet and child development. Proc Nutr
Soe. 2007, vol. 66, pp. 397-404.
Isolauri, E. Microbiota and Obesity. Nestlé Nutrition Institute Workshop Series. 2017, pp. 95–
105.
J
Jimenez E, et al. Isolation of commensal bacteria from umbilical cord blood of healthy
neonates born by Cesarean section. Curr Microbiol. 2005, vol. 51, pp. 270–274.

54
K
Kelly, Jr. et al. Transferring the blues: depression-associated gut microbiota induces
neurobehavioural changes in the rat. J Psychiatr Res. 2016, vol. 82, pp. 109–118.
Knol J, et al. Colonic microflora in infants fed formula with galacto- and fructo-
oligosaccharides: more like breastfed infants. J Pedi and Gastroenterol Nutr. 2005, vol. 40,
pp. 36–42.
Koenig, J. E. et al. Succession of microbial consortia in the developing infant gut
microbiome. Proc. Natl Acad. Sci. USA 108 (Suppl. 1), 2011, pp. 4578–4585.
Koplin, J. et al. Is caesarean delivery associated with sensitization to food allergens and IgE-
mediated food allergy: a systematic review.Pediatr Allergy Immunol. 2008, vol. 19, pp. 682–
687.
Koren O, et al. Host remodeling of the gut microbiome and metabolic changes during
pregnancy. Cell. 2012, vol. 150, pp. 470–480.
Krautler, B. Vitamin B12: chemistry and biochemistry. Biochem Soc Trans. 2005, vol. 33, pp.
806–810.
Kuczynski J, et al. Experimental and analytical tools for studying the human microbiome. Nat
Rev Genet. 2012, vol. 13, pp. 47–58.
Kvenshagen, B.; Halvorsen, R.; Jacobsen, M. Is there an increased frequency of food allergy
in children delivered by caesarean section compared to those delivered vaginally? Acta
Paediatr. 2009, vol. 98, pp. 324–327.
L
Lambrecht, B. N. & Hammad, H.. The immunology of the allergy epidemic and the hygiene
hypothesis. Nature Immunology. 2017, vol. 18(10), pp. 1076–1083.
Lamont, R. J.; Koo, H.; Hajshengallis, G. The oral microbiota: dynamic communities and host
interactions. Nature Reviews Microbiology. 2018, vol. 16, pp. 745–759.
Lappalainen, M. H. J. et al. Exposure to dogs is associated with a decreased tumour necrosis
factor-α-producing capacity in early life. Clinical & Experimental Allergy. 2010, vol. 40(10),
pp. 1498–1506.
Le Chatelier, E. et al. Richness of human gut microbiome correlates with metabolic
markers. Nature. 2013, vol. 500(7464), pp. 541–546.
Le Roy, T. et al. Intestinal microbiota determines development of non-alcoholic fatty liver
disease in mice. Gut. 2013, vol. 62(12), pp. 1787–1794.
Lederberg, B. J.; Mccray, A. T. ‘Ome Sweet’ Omics – A Genealogical Treasury of Words.
1977.
Liem, J..J. The risk of developing food allergy in premature or low-birth-weight children. J
Allergy Clin Immunol. 2007, vol. 119, pp. 1203–1209.

55
Lindgren, M. et al. Prolonged impact of a one-week course of clindamycin on
Enterococcus spp. in human normal microbiota. Scand J Infect Dis. 2009, vol. 41, pp. 215–
219.
Llopis, M. et al. Intestinal microbiota contributes to individual susceptibility to alcoholic liver
disease. Gut. 2016, vol. 65(5), pp. 830–839.
Lodge, C. J. et al. Pets at birth do not increase allergic disease in at-risk children. 2012, vol.
42(9), pp. 1377-85.
Lozupone, C. A. et al. Diversity, stability and resilience of the human gut microbiota. Nature.
2012, vol. 489, pp. 220-229.
M
Mackie, R.; Sghir, A.; Gaskins, H. R. Developmental microbial ecology of the neonatal
gastrointestinal tract. Am. J. Clin. Nutr. 1999, vol. 69, pp. 1035-1045.
Makino, H. et al. Transmission of intestinal Bifidobacterium longum subsp. longum strains
from mother to infant, determined by multilocus sequencing typing and amplified fragment
length polymporphism. Appl Environ Microbiol. 2011, vol. 77, pp. 6788–6793.
Marcobal, A. et al. Consumption of human milk oligosaccharides by gut-related microbes. J
Agric Food Chem. 2010, vol. 58(9), pp. 5334–5340.
Margulis, L. The Origin of Plant and Animal Cells: The serial symbiosis view of the origin of
higher cells suggests that the customary division of living things into two kingdoms should be
reconsidered. American Scientist. 1971, vol. 59, n. 2, pp 230-235.
Marshall, A. L. et al. Type 1 diabetes mellitus in childhood: a matched case control study in
Lancashire and Cumbria, UK. Diabet Med. 2004, vol. 21, pp. 1035–1040.
Martin, V. et al. Sharing of bacterial strains between breast milk and infant feces. J Hum Lact.
2012, vol. 28, pp. 36–44.
Matricardi, P. M,. et al. Exposure to foodborne and orofecal microbes versus airborne viruses
in relation to atopy and allergic asthma: epidemiological study. BMJ. 2000, vol. 320(7232),
pp. 412–417.
McFarland, L. V. Epidemiology, risk factors and treatments for antibiotic-associated
diarrhea. Dig Dis. 1998, vol. 16, pp. 292–307.
McFarland, L.V. Normal flora: diversity and functions. Microb. Ecol. Health Dis. 2000, vol.
12, pp. 193-207.
Meehan, C.; Helfrecht, C.; Malcom, C. Implications of lengthy development and maternal life
history: Allomaternal investment, peer relationships, and social networks. In C. L. Meehan &
A. Crittenden (Eds.), Childhood: Origin, evolution, and implications. 2016, pp. 199–220.
Meehan, C. L. et al. Social networks, cooperative breeding, and the human milk microbiome.
American Journal of Human Biology. 2018, vol. 30(4), e23131.

56
Melen, E. et al. Influence of early and current environmental exposure factors on sensitization
and outcome of asthma in pre-school children. Allergy 2001, vol. 56, pp. 646–52.
Million, M.; Diallo, A.; Raoult, D. Gut microbiota and malnutrition. Microbial Pathogenesis.
2017, vol. 106, pp. 127–138.
Mizuno, K. and Ueda, A. The maturation and coordination of sucking, swallowing, and
respiration in preterm infants. J. Pediatr. 2003, vol. 142, pp. 36–40.
Motarjemi, Y. et al. Contaminated weaning food: a major risk factor for diarrhoea and
associated malnutrition. Bull World Health Organ 1993, vol. 71(1), pp. 79-92.
Musso, G; Gambino, R; Cassader, M. Obesity, Diabetes, and Gut Microbiota: The hygiene
hypothesis expanded? Diabetes Care. 2010, vol. 33(10), pp. 2277–2284.
N
Navarro, S. et al. Hookworm recombinant protein promotes regulatory T cell responses that
suppress experimental asthma. Sci Transl Med. 2016, vol. 8, pp. 362ra.
Naik, S. et al. Compartmentalized control of skin immunity by resident commensals.
Science. 2012, vol. 337, pp. 1115–1119.
Neyrinck, A. M. et al. Critical role of Kupffer cells in the management of diet-induced
diabetes and obesity. Biochem Biophys Res Commun. 2009, vol. 385, pp. 351–356.
Nicoli, J. R. Normal gastrointestinal microbiota in domestic animals and human beings.
Enferm. Infec. Microbiol. 1995, vol. 15, pp. 183-190.
Nicoli, J. R.; Vieira, L. Q. Microbiota gastrointestinal normal na doença e na saúde.
Gastroenterologia. Rio de Janeiro: Médica e Científica Ltda. 2004, pp. 1037-1047.
O
Oh, J. et al. Temporal stability of the human skin microbiome. Cell. 2016, vol. 165, pp. 854–
866.
Organização Mundial de Saúde (OMS). Disponível em:
<http://apps.who.int/iris/bitstream/handle/10665/161442/WHO_RHR_15.02_por.pdf;jsessioni
d=AEEB2ECCC706FC9A20AB86820251FB37?sequence=3>. Acesso em 14 nov. 2018.
Okada, H. The ‘hygiene hypothesis’ for autoimmune and allergic diseases: an update. Clin
Exp Immunol. 2010, vol. 160(1), pp. 1–9.
Ownby, D. R.; Johnson, C. C.; Peterson, E. L. Exposure to dogs and cats in the first year of
life and risk of allergic sensitization at 6 to 7 years of age. JAMA. 2002, vol. 288(8), pp.
963–972.
P
Palmer, C. et al. Development of the human infant intestinal microbiota. PLOS Biol. 2007,
vol. 5:e177.

57
Park, J. et al. Short-chain fatty acids induce both effector and regulatory T cells by
suppression of histone deacetylases and regulation of the mTOR-S6K pathway. Mucosal
Immunol. 2015, vol. 8(1), pp. 80–93.
Pelucchi, C. et al. Pet exposure and risk of atopic dermatitis at the pediatric age: A meta-
analysis of birth cohort studies. The Journal of Allergy and Clinical Immunology. 2013, vol.
132, Issue 3, pp. 616–622.e7.
Penders, J. et al. Quantification of Bifidobacterium spp., Escherichia coli and Clostridium
difficile in faecal samples of breastfed and formula-fed infants by real-time PCR. FEMS
Microbiol. Lett. 2005, vol. 243, pp. 141–147.
Penders, J. et al. Factors influencing the composition of the intestinal microbiota in early
infancy. Pedriatics. 2006, vol. 118, pp. 511-521.
Ponsonby, A. L. et al. Exposure to infant siblings during early life and risk of multiple
sclerosis. JAMA. 2005, vol. 293, pp. 463–469.
PORTER, J. R. ANTONY VAN LEEUWENHOEK: TERCENTENARY OF HIS
DISCOVERY OF BACTERIA. 1976, VOL. 40, N. 2, PP. 260-269.
Poser, S. et al. Increasing incidence of multiple sclerosis in South Lower Saxony, Germany.
Neuroepidemiology. 1989, vol. 8, pp. 207–213.
Preidis, G. A. e Versalovic, J.. Targeting the human microbiome with antibiotics, probiotics,
and prebiotics: Gastroenterology enters the metagenomics Era. Official Journal of the Aga
Institute. 2009, vol. 136, pp. 2015-2031.
Q
Qin, J. et al. A human gut microbial gene catalogue established by metagenomic
sequencing. Nature. 2010, vol. 464, pp. 59–65.
R
Rautava, S. et al. Microbial contact during pregnancy, intestinal colonization and human
disease. Nature Reviews Gastroenterology & Hepatology. 2012.
Rede Simbiótica de Biologia e Conservação. Disponível em:
<http://simbiotica.org/reinos.htm>. Acesso em: 9 out 2018.
Renz-Polster, H. et al. Caesarean section delivery and the risk of allergic disorders in
childhood. Clin Exp Allergy. 2005, vol. 35, pp. 1466–1472.
Ribeiro, A. A. et al. Human microbiome: A mostly positive interaction? Revista Uningá.
2014, vol. 19, n. 1, pp 38-43.
Riva, G. et al. Nasal and Tracheal Microial Colonization in Laryngectomized Patients. Ann
Otol Rhinol Laryngol. 2016, vol. 125(4), pp. 336-41.
Riedler, J. et al. Exposure to farming in early life and development of asthma and allergy: a
crosssectional survey. Lancet. 2001, vol. 358, pp. 1129–33.

58
Rodriguez, J. M. et al. The composition of the gut microbiota throughout life, with an
emphasis on early life. Microb. Ecol. Health Dis. 2015, vol. 26, 26050.
Roger, L. C. et al. Examination of faecal Bifidobacterium populations in breast- and formula-
fed infants during the first 18 months of life. Microbiology 2010, vol. 156, pp. 3329–3341.
Roger, L. C. & McCartney, A. L. Longitudinal investigation of the faecal microbiota of
healthy full-term infants using fluorescence in situ hybridization and denaturing gradient gel
electrophoresis. Microbiology 2010, vol. 156, pp. 3317–3328.
Romero R, et al. The composition and stability of the vaginal microbiota of normal pregnant
women is different from that of non-pregnant women. Microbiome. 2014, vol. 2:4.
Rosenstein, I. J. et al. Relationship between hydrogen peroxide-producing strains of
lactobacilli and vaginosis-associated bacterial species in pregnant women. Eur J Clin
Microbiol Infect Dis. 1997, vol. 16, pp. 517–522.
S
Salonen, A.; Vos, W. M. Impact of Diet on Human Intestinal Microbiota and Health. The
Annual Review of Food Science and Technology. 2014, vol. 5, pp. 239–262.
Satokari, R. et al. Bifidobacterium and Lactobacillus DNA in the human placenta. Lett Appl
Microbiol. 2009, vol. 48, pp. 8–12.
Savage, D. C. et al. Mucosal microbiota. Mucosal Immunology. San Diego: Academic Press.
1999, pp. 19-30.
Sawczenko, A. et al. Prospective survey of childhood inflammatory bowel disease in the
British Isles. Lancet 2001, vol. 357, pp. 1093–1094.
Schaubeck, M. et al. Dysbiotic gut microbiota causes transmissible Crohn’s disease-like ileitis
independent of failure in antimicrobial defence. Gut. 2016; vol. 65(2), pp. 225–237.
Schlinzig, T. et al. Surge of immune cell formation at birth differs by mode of delivery and
infant characteristics — a population-based cohort study. PLoS One. 2017, 12: e0184748.
Schneider, N.; Garcia-Rodenas, C. L. Early nutritional interventions for brain and cognitive
development in preterm infants: a review of the literature. Nutrients. 2017, vol.9:E187.
Shen, Z. et. al. Update on intestinal microbiota in Crohn's disease 2017: Mechanisms, clinical
application, adverse reactions, and Outlook. Journal of Gastroenterology and Hepatology.
2017, vol. 32, issue 11, pp. 1804-1812.
Smith, P. M. et al. The microbial metabolites, short-chain fatty acids, regulate colonic Treg
cell homeostasis. Science. 2013, vol. 341(6145), pp. 569–573.
Sommer, F.; Bäckhed, F. The gut microbiota - masters of host development and physiology.
Nature Reviews Microbiology. 2013, vol. 11, pp. 227-238.

59
Spurbeck, R. R. Arvidson CG. Lactobacillus jensenii surface-associated proteins
inhibit Neisseria gonorrhoeae adherence to epithelial cells. Infect Immun. 2010, vol. 78, pp.
3103–3111.
Steel, J. H. et al. Bacteria and inflammatory cells in fetal membranes do not always cause
preterm labor. Pediatr Res. 2005, vol. 57, pp. 404–411.
Stene, L. C. and Nafstad, P. Relation between occurrence of type 1 diabetes and asthma.
Lancet. 2001, vol. 357, pp. 607–608.
Stiemsma, L. T. et al. The hygiene hypothesis: current perspectives and future therapies.
Immuno Targets and Therapy. 2015, vol. 2015:4, pp. 143-157.
Stokholm, J. et al. Cesarean section changes neonatal gut colonization. The Journal of Allergy
and Clinical Immunology. 2016, vol. 138; Issue 3; pp. 881-889
Stout M. J. et al. Identification of intracellular bacteria in the basal plate of the human
placenta in term and preterm gestations. Am J Obstet Gynecol. 2013, vol. 208, pp. 226.e1–
226.e7.
Strachan, D. P. Hay fever, hygiene, and household size. BMJ. 1989, vol. 299(6710), pp.
1259–1260.
Sullivan, A.; Edlund, C.; Nord, C. E. Effect of antimicrobial agents on the ecological balance
of human microflora. Lancet Infect Dis. 2001, vol. 1, pp. 101–114.
T
Talaro, K. P. Microbe-Human Interactions. Foundations in Microbiology, 6ª Edição.
Dubuque, Iowa, McGraw-Hill Higher Internacional Education. 2008, pp. 384-387.
Telo, F.; David, C. O rural depois do êxodo: as implicações do despovoamento dos campos
no distrito de Arroio do Só, município de Santa Maria/RS, Brasil. Mundo Agr. 2012, vol.13,
n.25.
Thavagnanam, S. et al. A meta-analysis of the association between Caesarean section and
childhood asthma. Clin Exp Allergy. 2008, vol. 38, pp. 629–633.
The International Study of Asthma and Allergies in Childhood (ISAAC): Worldwide
variation in prevalence of symptoms of asthma, allergic rhinoconjunctivitis, and atopic
eczema Steering Committee. Lancet. 1998, vol. 351(9111), pp. 1225–1232.
Thum, C. et al. Can nutritional modulation of maternal intestinal microbiota influence the
development of the infant gastrointestinal tract? J Nutr. 2012, vol. 142, pp. 1921–1928.
Tissier, H. Recherches sur la flore intestinale des nourrissons (état normal et
pathologique). Paris: G. Carre e C. Naud. 1900.
Trabulsi, L. R.; Sampaio, M. M. S. C. A composição e papel da microflora intestinal na saúde
e proteção do organismo. Os Probióticos e a Saúde Infantil. Nestlé Ltda. Brasil. 2000, vol. 1,
pp. 3-11.

60
Turnbaugh, P. J. et al. An obesity-associated gut microbiome with increased capacity for
energy harvest. Nature. 2006, vol. 444(7122), pp. 1027–1031.
Turnbaugh, P. J. et al. A core gut microbiome in obese and lean twins. Nature. 2009, vol. 457,
pp. 480–484.
V
van de Guchte, M.; Blottière, H. M.; Doré, J. Humans as holobionts: implications for
prevention and therapy. Microbiome. 2018, vol. 6:81.
van de Wijgert JH, et al. The vaginal microbiota: what have we learned after a decade of
molecular characterization? PLoS ONE. 2014, vol. 9:e105998.
van der Kleij, D. et al. A novel host– parasite lipid cross-talk. Schistosomal
lysophosphatidylserine activates Toll-like receptor 2 and affects immune polarization. J Biol
Chem. 2002, vol. 277, pp. 48122–48129.
van Eden, W. et al. Heat-shock proteins in T-cell regulation of chronic inflammation. Nature
Rev Immunol. 2005, vol. 5, pp. 318–330.
van Nimwegen, F. A. et al. Mode and place of delivery, gastrointestinal microbiota, and their
influence on asthma and atopy. Journal of Allergy and Clinical Immunology. 2011, vol. 128,
issue 5, pp. 948-955.e3.
van Nood, E. et al. Duodenal infusion of donor feces for recurrent Clostridium difficile. N
Engl J Med. 2013, vol. 368(5), pp. 407-15.
Vlahov, D. et al. Toward an urban health advantage. J Public Health Manag Pract, 2005,
vol.11, pp.256-8.
W
Wall, R. et al. Role of gut microbiota in early infant development. Clinical Medicine:
Pediatrics, Auckland. 2009, vol. 3, pp. 45-54.
Wang, X. et al. Comparative microbial analysis of paired amniotic fluid and cord blood from
pregnancies complicated by preterm birth and early-onset neonatal sepsis. PLoS ONE. 2013,
vol. 8:e56131.
Waser, M. et al. Exposure to pets, and the association with hay fever, asthma, and atopic
sensitization in rural children. Allergy 2005, vol. 60, pp. 177–84.
Whittaker, R. H. New Concepts of Kingdoms of Organisms. Science, New series. 1969, vol.
163, n. 3863, pp 150-160.
Whittaker, R. H.; Margulis, L. Protist classification and the kingdoms of organisms. Elsevier.
1978, vol. 10, n. 1-2, pp 3-18.
WHO. Child growth standards and the identification of severe acute malnutrition in infants
and children, a joint statement by the World Health Organization and the United Nations
Children's Fund. 2009.

61
WHO. Community-based management of severe acute malnutrition. A joint statement by the
world health organization, the World Food Programme, the United Nations System Standing
Committee on Nutrition and the United Nations Children's Fund. 2007
Williams, C. D. A nutritional disease of childhood associated with a maize diet. Arch Dis
Child 1933, vol. 8(48), pp. 423-33.
Wu G. D. et al. Linking long-term dietary patterns with gut microbial
enterotypes. Science. 2011, vol. 334(6052), pp. 105–108.
Z
Zhang, J; Chen, S. L; Li, L. B. Correlation between intestinal flora and serum inflammatory
factors in patients with Crohn’s disease. European Review for Medical and Pharmacological
Sciences. 2017, vol. 21, pp. 4913-4917.
Related Documents











