APMIS 107: 160-7, 1999 Printed in Denmark . All rights reserved Cop,vrixht 0 APMIS 1999 ISSN 0903-4641 The urokinase plasminogen activator receptor in blood from healthy individuals and patients with cancer Review article NILS BRUNNER,' HANS J0RGEN NIELSEN,2 MARIA HAMERS,' IB JARLE CHRISTENSEN,' OLE THORLACIUS-USSING3 and ROSS W. STEPHENS' Finsen Laboratory, Rigshospitalet, Copenhagen, 2Department of Surgical Gastroenterology, Hvidovre Hospital, Copenhagen, and 3Department of Surgical Gastroenterology, Alborg Hospital, Alborg, Denmark Brunner N, Nielsen HJ, Hamers M, Christensen IJ, Thorlacius-Ussing 0 & Stephens RW. The uroki- nase plasminogen activator receptor in blood from healthy individuals and patients with cancer. APMIS 1999; 107: 160-7. The cell surface plasminogen activation system functions in promoting tumor dissemination, and is facilitated by a glycolipid anchored three domain receptor for urokinase. This receptor can also be found in a soluble form (suPAR) in extracts of tumors, as well as in plasma from both healthy individuals and cancer patients. The suPAR in plasma consists of the intact three domain protein, but neither the precise mechanism of its release from cell surfaces, nor its biological function are understood. Increased levels of plasma suPAR have been found in patients with cancers of the lung, breast, ovary, and colon, and recent data now indicates that the level of the molecule is related to patient prognosis. Key words: uPAR; suPAR; cancer; prognosis. Nils Brunner, Finsen Laboratory, Strandboulevarden 49, DK-2 100 Copenhagen, Denmark. The invasive process by which malignant tumors dis- seminate can now be more accurately described as a process of unregulated tissue remodelling, progress- ively involving normal cells which are recruited into the tumor stroma by the neoplastic cells. In this pro- cess the barriers which normally define the homeo- stasis of differentiated tissue are breached by new sig- nals from the de-differentiating and proliferating neoplastic cells. Recruitment and reorganisation of the normal tissue cells progressively disrupts organ structure, as the neoplastic cells develop and continu- ously remodel a supporting stroma, complete with a new blood capillary network. Tumor tissue remodel- ling is also accompanied by the detachment of tumor cells and their active invasion of blood and lymph vessels, from where they can metastasize to form new tumors at distant sites. The hallmark of invasive tissue remodelling is the degradation of the existing extracellular matrix which normally maintains the integrity of organised 160 tissue structures. In particular, the basement mem- brane is a specialised form of matrix which normally forms a boundary adjacent to the epithelial tissues where carcinomas arise. In order to degrade and tra- verse the first barrier represented by the basement membrane, it is a prerequisite that cancers arising in epithelial tissues either acquire the ability themselves to secrete proteolytic enzymes which can cleave base- ment membrane proteins, or induce the required pro- teases in the normal tissue cells which are progress- ively recruited into the tumor stroma. In addition, proteolysis of the extracellular matrix may in turn stimulate growth of tumors by release and/or acti- vation of growth factors and cytokines and/or by in- ducing neovascularization. Thus, certain proteases are highly expressed in tumor tissue, including mem- bers of both the serine protease family, e.g. uroki- nase-type plasminogen activator and the metallopro- teinase family. Several publications have demonstrated a prognos-

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

APMIS 107: 160-7, 1999 Printed in Denmark . All rights reserved
Cop,vrixht 0 A P M I S 1999
ISSN 0903-4641
The urokinase plasminogen activator receptor in blood from healthy individuals and patients with cancer
Review article
NILS BRUNNER,' HANS J0RGEN NIELSEN,2 MARIA HAMERS,' IB JARLE CHRISTENSEN,' OLE THORLACIUS-USSING3 and ROSS W. STEPHENS'
Finsen Laboratory, Rigshospitalet, Copenhagen, 2Department of Surgical Gastroenterology, Hvidovre Hospital, Copenhagen, and 3Department of Surgical Gastroenterology, Alborg Hospital,
Alborg, Denmark
Brunner N, Nielsen HJ, Hamers M, Christensen IJ, Thorlacius-Ussing 0 & Stephens RW. The uroki- nase plasminogen activator receptor in blood from healthy individuals and patients with cancer. APMIS 1999; 107: 160-7.
The cell surface plasminogen activation system functions in promoting tumor dissemination, and is facilitated by a glycolipid anchored three domain receptor for urokinase. This receptor can also be found in a soluble form (suPAR) in extracts of tumors, as well as in plasma from both healthy individuals and cancer patients. The suPAR in plasma consists of the intact three domain protein, but neither the precise mechanism of its release from cell surfaces, nor its biological function are understood. Increased levels of plasma suPAR have been found in patients with cancers of the lung, breast, ovary, and colon, and recent data now indicates that the level of the molecule is related to patient prognosis.
Key words: uPAR; suPAR; cancer; prognosis.
Nils Brunner, Finsen Laboratory, Strandboulevarden 49, DK-2 100 Copenhagen, Denmark.
The invasive process by which malignant tumors dis- seminate can now be more accurately described as a process of unregulated tissue remodelling, progress- ively involving normal cells which are recruited into the tumor stroma by the neoplastic cells. In this pro- cess the barriers which normally define the homeo- stasis of differentiated tissue are breached by new sig- nals from the de-differentiating and proliferating neoplastic cells. Recruitment and reorganisation of the normal tissue cells progressively disrupts organ structure, as the neoplastic cells develop and continu- ously remodel a supporting stroma, complete with a new blood capillary network. Tumor tissue remodel- ling is also accompanied by the detachment of tumor cells and their active invasion of blood and lymph vessels, from where they can metastasize to form new tumors at distant sites.
The hallmark of invasive tissue remodelling is the degradation of the existing extracellular matrix which normally maintains the integrity of organised
160
tissue structures. In particular, the basement mem- brane is a specialised form of matrix which normally forms a boundary adjacent to the epithelial tissues where carcinomas arise. In order to degrade and tra- verse the first barrier represented by the basement membrane, it is a prerequisite that cancers arising in epithelial tissues either acquire the ability themselves to secrete proteolytic enzymes which can cleave base- ment membrane proteins, or induce the required pro- teases in the normal tissue cells which are progress- ively recruited into the tumor stroma. In addition, proteolysis of the extracellular matrix may in turn stimulate growth of tumors by release and/or acti- vation of growth factors and cytokines and/or by in- ducing neovascularization. Thus, certain proteases are highly expressed in tumor tissue, including mem- bers of both the serine protease family, e.g. uroki- nase-type plasminogen activator and the metallopro- teinase family.
Several publications have demonstrated a prognos-

uPAR IN BLOOD
tic value in cancer patients of molecules involved in the urokinase pathway of plasminogen activation (1- 22). However, almost all these results are based on determination in tumor extracts. The development of more effective diagnostic methods has resulted in earlier detection and thus smaller tumor size at the time of surgery, which is making it increasingly diffi- cult to acquire access to frozen, unfixed tumor samples. Therefore, the conversion of these assays from tests which require tumor extracts to assays that can be performed on plasma or serum would signifi- cantly increase their clinical value. Sample collection would be easier, and the need for careful histological evaluation of the tissue to be analysed as well as the inherent problems of tissue heterogeneity would not apply to blood samples. Even more important, the possibility of obtaining blood samples during clinical follow-up after primary surgery for cancer, would allow sequential measurements of the uPA related an- alytes.
We recently reported that tumor tissue levels of sol- uble uPAR but not total uPAR were associated with prognosis in breast cancer patients (6). At that time we suspected that this form of soluble uPAR (suPAR: full-length receptor and cleavage products, without the glycolipid anchor) after release from the cell sur- face could find its way into the circulation. In order to study plasma and serum levels of suPAR in cancer patients, we developed a kinetic ELISA specifically evaluated for measurements of suPAR in blood (23). Using this ELISA, we have since shown that in fact patients with various forms of cancer including pa- tients with metastatic breast cancer, Dukes D col- orectal cancer and patients with ovarian cancer (22,
23) have elevated levels of suPAR in their blood, and in two recent studies, one of the patients with colorec- tal cancer (24) and another of the patients with ovarian cancer (22) we found a significant association between elevated suPAR blood levels and shorter sur- vival.
BIOCHEMISTRY OF THE UROKINASE PLASMINOGEN ACTIVATION SYSTEM
Fig. 1 shows the urokinase plasminogen activation system. uPA is a 52 kD serine protease which is se- creted as an inactive single chain proenzyme (pro- uPA) (25, 26). This immature form has only 11250 the activity of the mature enzyme (27). Pro-uPA can be activated to uPA by plasmin, cathepsin B and kallikrein (28, 29), which all cleave the single chain pro-uPA within an intrachain loop held closed by a disulfide bridge. Thus the active enzyme consists of two chains (A+B) held together by this disulfide bridge. Active uPA has high specificity for the Arg560-Val561 bond in plasminogen, and cleavage between these residues gives rise to active plasmin. Plasminogen (28) is present at high concentration (1.5-2.0 pM) in plasma and interstitial fluids, and represents a plentiful potential source of plasmin ac- tivity. Unlike uPA, plasmin is a relatively non-specific protease, cleaving many glycoproteins and proteo- glycans of the extracellular matrix, as well as fibrin (30). In addition, it is important to note that plasmin can activate some of the metalloproteases which also degrade tissue matrix (31, 32), and plasmin is known to activate growth factors, e.g. TGF-P (33), which
Fig. 1. The urokinase plas- minogen activation system. uPA: urokinase plasminogen activator; uPAR: plasmino- gen activator receptor; su- PAR: soluble uPAR; PLG: plasminogen; PL: plasmin; PAI: plasminogen activator inhibitor.
161

BRUNNER era / .
may further modulate stromal interactions in the ex- pression of enzymes and tumor neo-angiogenesis.
Pro-uPA and uPA bind to a specific cell-surface re- ceptor (uPAR) (34, 35). This receptor consists of a glycolipid-anchored three-domain 60 kD glyco- protein whose N-terminal domain l contains the high affinity binding site for a growth factor domain within the A-chain of uPA. Since uPAR also has an- other important interaction with vitronectin (36), these two interactions explain the observed localiz- ation of pro-uPA and thus potential uPA activity at the focal contacts and junctions formed between cells and with the extracellular matrix (37). Indeed uPA binding to uPAR promotes binding of uPAR to the matrix protein vitronectin (36), and thus adhesion of cells to a vitronectin substratum.
Low affinity, high capacity binding of plasminogen to cell-surface proteins through the lysine binding sites of plasminogen kringles (38) enhances consider- ably the rate of plasminogen activation by uPA (29, 39). This is probably due to the proximity of bound plasminogen to the laterally mobile uPAR (40) on the cell-surface. Furthermore, plasmin formed on the cell surface is able to catalyse the activation of uPAR- bound pro-uPA (29), while remaining protected from plasma proteinase inhibitors, especially a-2-antiplas- min. Cell-surface bound plasmin can then mediate the non-specific matrix proteolysis which facilitates migration of tumor cells through restraining tissue structures and activates or releases growth factors in the matrix.
Plasminogen activation by uPA is regulated by two important inhibitors, PAI-I and PAI-2 (37, 41). PAI- 1, a 52 kD single chain protein, is secreted in an ac- tive but conformationally unstable form, whose in- hibitory activity is stabilized and prolonged by bind- ing to vitronectin (42, 43). PAI-1 is the most abun- dant fast-acting inhibitor of uPA in tumor tissues, and it is also able to inactivate uPA which is bound to uPAR. Formation of PAI-1 or PAI-2 complexes with uPA on the cell-surface leads to internalization of the uPAR/uPA/PAI complex (44) and probably drives uPAR cycling through the endosomal com- partment back to the cell surface (45). uPA activity may also be lost from the cell-surface by plasmin cleavage at Lys135-Lys136 in the A-chain of uPA, leaving only the so-called amino terminal fragment (ATF) bound on the cell. The general plasma protein- ase inhibitor, a2-macroglobulin, inactivates soluble uPA relatively slowly, but uPA bound to uPAR is ap- parently protected by a steric effect (46), thus allowing plasminogen activation to occur on the sur- face of cells even in the presence of serum (29).
The interaction between PAI-1 and the cell-surface uPA system is now also appreciated as a modulator of cell adhesion and migration (47). The basis of this effect is competition by active PAI-1 for uPAR bind-
162
ing sites on matrix vitronectin. Thus a high local con- centration of PAL1 can overcome uPAR-mediated adhesion and promote cell detachment, increased motility and migration. This may be most important for transformed epithelial cells which have reduced adhesion interactions with other matrix proteins, e.g. fibronectin (48).
SOLUBLE uPAR
As mentioned above uPAR consists of a three-do- main highly glycosylated protein which in patients with cancer may be expressed on the surface of acti- vated macrophages infiltrating the tumor stroma, and/or on the tumor cells themselves (35). uPAR is anchored to the cell membrane by a glycosylphos- phatidylinositol linkage at its carboxy-terminus (34), and this linkage can be cleaved by e.g. phospholipase C resulting in release of a hydrophilic full-length su- PAR molecule. Between domains 1 and 2, there is a “hinge” region of relatively unstructured backbone, containing cleavage sites for the serine proteases uPA, plasmin, trypsin, chymotrypsin, and probably several other proteases (49). Thus domain 1 may be cleaved off the receptor by proteases either prior to, or after cleavage by a phospholipase. The main forms of uPAR one could expect to find in blood are therefore full length, three-domain soluble uPAR (suPAR), two-domain suPAR consisting of domains (2+3) or free domain 1 (Fig. 2). The two former have both been found in extracts of human tumors xenografted on nude mice (50). However, suPAR is also present in patient tumor extracts (37) as well as in plasma from these patients (33), and one could imagine this consisting of full-length, three domain suPAR, as well as the two domain (2+3) and free domain 1. Free domain 1 is relatively small, comparable in MW to
UROKINASE RECEPTOR CLEAVAGE
PROTEOLYTIC CLEAVAGE
PHOSPHOLIPASE
uPAR IN BLOOD
microglobulin, which is known to be lost from blood through the kidneys. Free domain 1 has not been found in either tumor extracts or plasma. All the evi- dence currently available indicates that suPAR in plasma, both from healthy donors and cancer pa- tients, consists of the full-length, three domain pro- tein (22, 23, 51).
MEASUREMENT OF suPAR IN PLASMA AND SERUM
Various techniques including receptor/ligand assays, immunohisto- (cyto)chemistry, ELISA and Western blot, have been employed to determine uPAR and su- PAR in tumor tissues and blood in order to assess their clinical relevance. For this purpose, a variety of monoclonal antibodies to uPAR have been developed and using these antibodies one can distinguish be- tween different molecular forms of uPAR (e.g. full- length receptor, domains 1, 2, 3 as well as free versus complexed forms) (52).
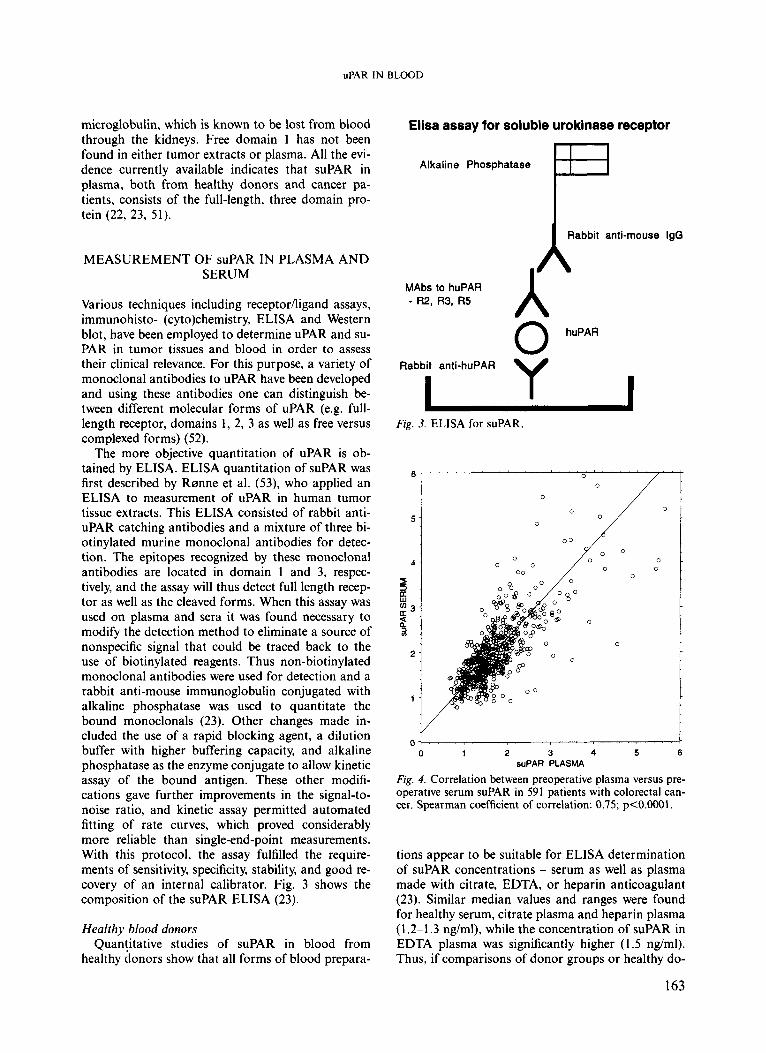
The more objective quantitation of uPAR is ob- tained by ELISA. ELISA quantitation of suPAR was first described by Rsnne et al. (53), who applied an ELISA to measurement of uPAR in human tumor tissue extracts. This ELISA consisted of rabbit anti- uPAR catching antibodies and a mixture of three bi- otinylated murine monoclonal antibodies for detec- tion. The epitopes recognized by these monoclonal antibodies are located in domain 1 and 3, respec- tively, and the assay will thus detect full length recep- tor as well as the cleaved forms. When this assay was used on plasma and sera it was found necessary to modify the detection method to eliminate a source of nonspecific signal that could be traced back to the use of biotinylated reagents. Thus non-biotinylated monoclonal antibodies were used for detection and a rabbit anti-mouse immunoglobulin conjugated with alkaline phosphatase was used to quantitate the bound monoclonals (23). Other changes made in- cluded the use of a rapid blocking agent, a dilution buffer with higher buffering capacity, and alkaline phosphatase as the enzyme conjugate to allow kinetic assay of the bound antigen. These other modifi- cations gave further improvements in the signal-to- noise ratio, and kinetic assay permitted automated fitting of rate curves, which proved considerably more reliable than single-end-point measurements. With this protocol, the assay fulfilled the require- ments of sensitivity, specificity, stability, and good re- covery of an internal calibrator. Fig. 3 shows the composition of the suPAR ELISA (23).
Healthy blood donors Quan!itative studies of suPAR in blood from
healthy donors show that all forms of blood prepara-
Elisa assay for soluble urokinase receptor
B Alkaline Phosphatase
Rabbit anti-mouse IgG
I A A MAbs to huPAR - R2, R3, R5
huPAR 0 Rabbit anti-huPAR
Fig. 3. ELISA for suPAR.
6 0
0
0 ! 1 0 0 7
0 0 0
00
0 1 2 3 4 5 6 suPAR PLASMA
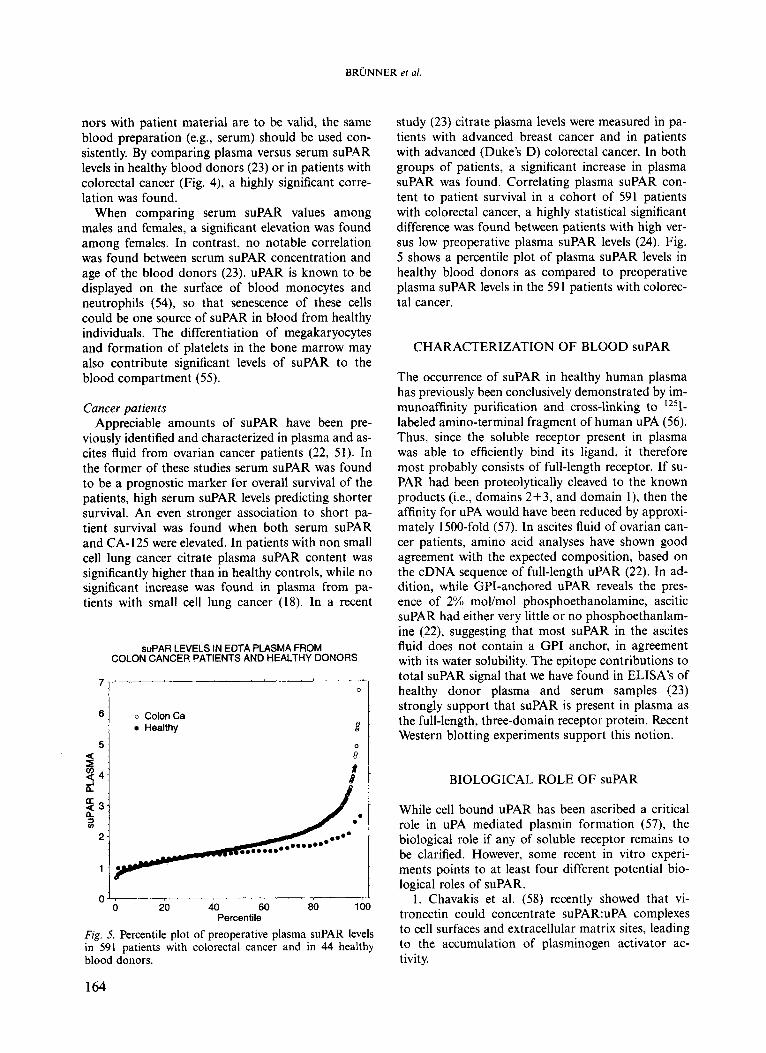
Fig. 4. Correlation between preoperative plasma versus pre- operative serum suPAR in 591 patients with colorectal can- cer. Spearman coefficient of correlation: 0.75; p<O.OOOl.
0
tions appear to be suitable for ELISA determination of suPAR concentrations - serum as well as plasma made with citrate, EDTA, or heparin anticoagulant (23). Similar median values and ranges were found for healthy serum, citrate plasma and heparin plasma (1.2-1.3 ng/ml), while the concentration of suPAR in EDTA plasma was significantly higher (1.5 ng/ml). Thus, if comparisons of donor groups or healthy do-
163
BRUNNER e r a / .
nors with patient material are to be valid, the same blood preparation (e.g., serum) should be used con- sistently. By comparing plasma versus serum suPAR levels in healthy blood donors (23) or in patients with colorectal cancer (Fig. 4), a highly significant corre- lation was found.
When comparing serum suPAR values among males and females, a significant elevation was found among females. In contrast, no notable correlation was found between serum suPAR concentration and age of the blood donors (23). uPAR is known to be displayed on the surface of blood monocytes and neutrophils (54), so that senescence of these cells could be one source of suPAR in blood from healthy individuals. The differentiation of megakaryocytes and formation of platelets in the bone marrow may also contribute significant levels of suPAR to the blood compartment (55).
Cancer patients Appreciable amounts of suPAR have been pre-
viously identified and characterized in plasma and as- cites fluid from ovarian cancer patients (22, 51). In the former of these studies serum suPAR was found to be a prognostic marker for overall survival of the patients, high serum suPAR levels predicting shorter survival. An even stronger association to short pa- tient survival was found when both serum suPAR and CA-125 were elevated. In patients with non small cell lung cancer citrate plasma suPAR content was significantly higher than in healthy controls, while no significant increase was found in plasma from pa- tients with small cell lung cancer (18). In a recent
suPAA LEVELS IN EDTA PLASMA FROM 90LON CANCER PATIENTS AND HEALTHY DONORS
7
6
5
f 3 4 n
3 3 3
2
1
0
o Colon Ca Healthy 1
0
8
P
100
Fig. 5. Percentile plot of preoperative plasma suPAR levels in 591 patients with colorectal cancer and in 44 healthy blood donors.
0 0 20 40 60 80
Percentile
study (23) citrate plasma levels were measured in pa- tients with advanced breast cancer and in patients with advanced (Duke’s D) colorectal cancer. In both groups of patients, a significant increase in plasma suPAR was found. Correlating plasma suPAR con- tent to patient survival in a cohort of 591 patients with colorectal cancer, a highly statistical significant difference was found between patients with high ver- sus low preoperative plasma suPAR levels (24). Fig. 5 shows a percentile plot of plasma suPAR levels in healthy blood donors as compared to preoperative plasma suPAR levels in the 591 patients with colorec- tal cancer.
CHARACTERIZATION OF BLOOD suPAR
The occurrence of suPAR in healthy human plasma has previously been conclusively demonstrated by im- munoaffinity purification and cross-linking to Iz5I- labeled amino-terminal fragment of human uPA (56) . Thus, since the soluble receptor present in plasma was able to efficiently bind its ligand, it therefore most probably consists of full-length receptor. If su- PAR had been proteolytically cleaved to the known products (i.e., domains 2+3, and domain l), then the affinity for uPA would have been reduced by approxi- mately 1500-fold (57). In ascites fluid of ovarian can- cer patients, amino acid analyses have shown good agreement with the expected composition, based on the cDNA sequence of full-length uPAR (22). In ad- dition, while GPI-anchored uPAR reveals the pres- ence of 2% mol/mol phosphoethanolamine, ascitic suPAR had either very little or no phosphoethanlam- ine (22), suggesting that most suPAR in the ascites fluid does not contain a GPI anchor, in agreement with its water solubility. The epitope contributions to total suPAR signal that we have found in ELISA’s of healthy donor plasma and serum samples (23) strongly support that suPAR is present in plasma as the full-length, three-domain receptor protein. Recent Western blotting experiments support this notion.
BIOLOGICAL ROLE OF suPAR
While cell bound uPAR has been ascribed a critical role in uPA mediated plasmin formation (57), the biological role if any of soluble receptor remains to be clarified. However, some recent in vitro experi- ments points to at least four different potential bio- logical roles of suPAR.
1. Chavakis et al. (58) recently showed that vi- tronectin could concentrate suPAR:uPA complexes to cell surfaces and extracellular matrix sites, leading to the accumulation of plasminogen activator ac- tivity.
164

uPAR IN BLOOD
2. Resnati et al. (59) found that suPAR was capable of inducing chemotaxis in cells. However, this activity required proteolytic cleavage of suPAR between do- main 1 and domain 2.
3. Through its interaction with uPAR binding to vitronectin (36, 47), PAI-1 can facilitate cell mi- gration. It is tempting to speculate that suPAR might in a similar way interact with cell surface uPAR:vi- tronectin binding and thereby increase cell migration. It should be mentioned that only full-length and not the cleaved forms of suPAR bind vitronectin (60).
4. In an in vitro study, Wilhelm et al. (61) showed that suPAR can serve as a scavenger for uPA thereby inhibiting proliferation and in vitro invasion of hu- man cancer cells.
CONCLUSION
Significant levels of suPAR are present in plasma and serum from healthy blood donors and cancer patients appear to have increased concentrations of suPAR in their blood, which is in agreement with the potentiat- ing of plasmin formation by binding of uPA to cell surface anchored uPAR and concomitant cell surface binding of plasminogen. Recent studies suggest that full-length suPAR without the glycolipid anchor is the predominant form of suPAR in blood. By com- paring plasma or serum suPAR levels with patient survival, high suPAR levels have been implicated as a parameter of shorter survival.
Neither the exact mechanism(s) of suPAR release from cell surfaces nor its biological role if any is yet clearly defined. Future studies should aim at clarify- ing these questions.
1.
2.
3.
4.
5.
REFERENCES
Duggan C, Maguire T, McDermott E, O’Higgins N, Fennelly JJ, Duffy MJ. Urokinase plasminogen acti- vator and urokinase plasminogen activator receptor in breast cancer. Int J Cancer 1995;61:597-600. Fernti M, Bendahl P-0, Borg A, Brundell J, Hirschberg L, Olsson H, Killander D. Urokinase plasminogen acti- vator, a strong independent prognostic factor in breast cancer, analysed in steroid receptor cytosols with a lum- inometric immunoassay. Eur J Cancer 1996;32A:793- 801. Foekens JA, Schmitt M, van Putten WL, Peters HA, Kramer MD, Janicke F, Klijn JG. Plasminogen activator inhibitor-1 and prognosis in primary breast cancer, J Clin Oncol 1994; 12: 1648-58. Foekens JA, Schmitt M, van PW, Peters HA, Bontenbal M, Janicke F, Klijn JG. Prognostic value of urokinase- type plasminogen activator in 671 primary breast cancer patients. Cancer Res 1992;52:6101-5. Grsndahl-Hansen J, Christensen IJ, Rosenquist C, Brilnner N, Mouridsen HT, Dan0 K, Blichert-Toft M.
High levels of urokinase-type plasminogen activator and its inhibitor PAI-1 in cytosolic extracts of breast carcinomas are associated with poor prognosis. Cancer
6. Grsndahl-Hansen J, Peters HA, van Putten WL, Look MP, Pappot H, Rsnne E, et al. Prognostic significance of the receptor for urokinase plasminogen activator in breast cancer. Clinical Cancer Res 1995; 1: 1079-87.
7. Grsndahl-Hansen J, Christensen IJ, Briand P, Pappot H, Mouridsen HT, Blichert-Toft M, Dan0 K, Brilnner N. Plasminogen activator inhibitor type 1 in cytosolic tumor extracts predicts prognosis in low-risk breast can- cer patients. Clinical Cancer Res 1997;3:233-9.
8. Heiss MM, Allgayer H, Gruetzner KU, Funke I, Babic R, Jauch RW, Schildberg FW. Individual development and uPA-receptor expression of disseminated tumour cells in bone marrow: a reference to early systemic dis- ease in solid cancer. Nat Med 1995; 1 : 1035-9.
9. Hofmann R, Lehmer A, Buresch M, Hartung R, Ulm K. Clinical relevance of urokinase plasminogen acti- vator, its receptor, and its inhibitor in patients with re- nal cell carcinoma. Cancer 1996;78:487-92.
10. Janicke F, Pache L, Schmitt M, Ulm K, Thomssen C, Prechtl A, Graeff H. Both the cytosols and detergent extracts of breast cancer tissues are suited to evaluate the prognostic impact of the urokinase-type plasmino- gen activator and its inhibitor, plasminogen activator inhibitor type 1. Cancer Res 1994;54:2527-30.
11. Janicke F, Schmitt M, Pache L, Ulm K, Harbeck N, Holler H, Graeff H. Urokinase (uPA) and its inhibitor PAI-1 are strong and independent prognostic factors in node-negative breast cancer. Breast Cancer Res Treat
12. Janicke F, Schmitt M, Ulrn K, Gossner W, Graeff H. Urokinase-type plasminogen activator antigen and early relapse in breast cancer [letter]. Lancet 1989;2: 1049.
13. Janicke F, Schmitt M, Graeff H. Clinical relevance of the urokinase-type and tissue-type plasminogen acti- vators and of their type 1 inhibitor in breast cancer. Semin Thromb Hemostasis 1991;17:303-12.
14. Jlnicke F, Thomssen C. Pache L. Schmitt M. Graeff
Res 1993;53:2513-21.
1993;24: 195-208.
15.
16.
H. Urokinase (uPA) and PAI-1 as.selection criteria for adjuvant chemotherapy in axillary node-negative breast cancer patients. In: Scmitt M, et al., editors. Prospects in diagnosis and treatment of breast cancer. Elsevier Science B.V.: 1994. p.207-18. Kobayashi H, Fujishiro S, Terao T. Impact of uroki- nase-type plasminogen activator and its inhibitor type 1 on prognosis in cervical cancer of the uterus. Cancer Res 1994;54:653948. Kuhn W, Pache L, Schmalfeldt B, Dettmar P, Schmitt M, Janicke F, Graeff H. Urokinase (uPA) and PAI-1 predict survival in advanced ovarian cancer patients (FIG0 111) after radical surgery and platinum-based chemotherapy. Gyn Oncol 1994; 401-9.
17. Pappot H,-Gaardsvoll H, Rsmer J, Pedersen AN, Grsndahl-Hansen J, Pyke C, Briinner N. Plasminogen activator inhibitor type 1 in cancer: therapeutic and prognostic implications. Biol Chem Hoppe Seyler 1995;376:259-67.
18. Pappot H, Hsyer-Hansen G, Rsnne E, Hansen HH, Briinner N, Dan0 K, Grmdahl-Hansen J. Elevated plasma levels of urokinase plasminogen activator recep- tor in non-small cell lung cancer patients. Eur J Cancer
19. Pedersen H, Briinner N, Francis D, Osterlind K, Rsnne 1997;33:867-72.
165

BRUNNER el a[.
E, Hansen HH, et al. Prognostic impact of urokinase, urokinase receptor, and type 1 plasminogen activator inhibitor in squamous and large cell lung cancer tissue. Cancer Res 1994;54:4671-5.
20. Pedersen H, Grsndahl-Hansen J, Francis D, Osterlind K, Hansen HH, Dan0 K, Brunner N. Urokinase and plasminogen activator inhibitor type 1 in pulmonary adenocarcinoma. Cancer Res 1994;54: 120-3.
21. Plebani M, Herszhyi L, Carraro P, Paoli MD, Roveroni G, Cardin R, et al. Urokinase-type plasminogen acti- vator receptor in gastric cancer: tissue expression and prognostic role. Clin Exp Metastasis 1997;15:418-25.
22. Sier CFM, Stephens RW, Bizik J, Mariani A, Bassan M, Pedersen N, Frigerio L, Ferrari A, Dan0 K, Brunner N, Blasi E The level of urokinase-type plasminogen acti- vator receptor is increased in serum of ovarian cancer patients. Cancer Res 1998;58:1843-9.
23. Stephens RW, Pedersen NA, Nielsen HJ, Hamers MJAG, Hsyer-Hansen G, Rsnne E, et al. ELISA deter- mination of soluble urokinase receptor in blood from healthy donors and cancer patients. Clin Chem
24. Stephens RW, Nielsen HJ, Christensen IJ, Thorlacius- Ussing 0, Dans K, Brunner N. Soluble urokinase re- ceptor in preoperative plasma is an independent prog- nostic variable in patients with colorectal cancer. Proc Am Ass Cancer Res 1998;39:# 325.
25. Nielsen LS, Hansen JG, Skriver L, Wilson EL, Kaltoft K, Zeuthen J, Dan0 K. Purification of zymogen to plas- minogen activator from human glioblastoma cells by af- finity chromatography with monoclonal antibody. Bio- chemistry 1982;2 1 :6410-15.
26. Skriver L, Nielsen LS, Stephens R, Dan0 K. Plasmino- gen activator released as inactive proenzyme from mu- rine cells transformed by sarcoma virus. Eur J Biochem
27. Petersen LC, Lund LR, Nielsen LS, Dan0 K, Skriver L. One-chain urokinase-type plasminogen activator from human sarcoma cells is a proenzyme with little or no intrinsic activity. J Biol Chem 1988;263: 11 189-95.
28. Dans K, Andreasen PA, Grmdahl-Hansen J, Kristens- en P, Nielsen LS, Skriver L. Plasminogen activators, tissue degradation and cancer. Adv Cancer Res
29. Stephens RW, Pollanen J, Tapiovaara H, Leung KC, Sim PS, Salonen EM, et al. Activation of pro-urokinase and plasminogen on human sarcoma cells: a proteolytic system with surface-bound reactants. J Cell Biol
30. Liotta LA, Goldfarb RH, Brudage R, Siegal GP, Ter- ranova V, Garbisa S. Effect of plasminogen activator (urokinase), plasmin, and thrombin on glycoprotein and collagenous components of basement membrane. Cancer Res 1981;41:4629-36.
31. Werb Z, Mainardi CL, Vater CA, Harris-ED J. Endo- genous activiation of latent collagenase by rheumatoid synovial cells. Evidence for a role of plasminogen acti- vator. N Engl J Med 1977;296:1017-23.
32. DeClerck YA, Laug WE. Cooperation between matrix metalloproteinases and the plasminogen activator-plas- min system in tumor progression. Enzyme Protein
33. Lyons RM, Keski-Oja J, Moses HL. Proteolytic acti- vation of latent transforming growth factor-p from fibroblast-conditioned medium. J Cell Biol 1988;
1997;43: 1868-76.
1982; 124:409-14.
1985;44: 139-266.
l989;108: 1987-95.
1996;49:72-84.
106:1659-65.
34. Ploug M, Rsnne E, Behrendt N, Jensen AL, Blasi F, Dan0 K. Cellular Receptor for Urokinase Plasminogen Activator. Carboxyl-terminal processing and membrane anchoring by glycosyl-phosphatidylinositol. J Biol Chem 1991;266: l92G33.
35. Behrendt N, Ploug M, Patthy L, Houen G, Blasi F, Dan0 K. The ligand-binding domain of the cell surface receptor for urokinase-type plasminogen activator. J Biol Chem 1991;266:7842-7.
36. Wei Y, Waltz DA, Rao N, Drummond RJ, Rosenberg S, Chapman HA. Identification of the urokinase recep- tor as an adhesion receptor for vitronectin. J Biol Chem 1994;269:32380-8.
37. Pollanen J, Stephens RW, Vaheri A. Directed plasmino- gen activation at the surface of normal and malignant cells. Adv Cancer Res 1991;57:273-328.
38. Miles LA, Plow EF. Ubiquitous sites for cellular regula- tion of fibrinolysis. Fibrinolysis 1988; 61-71.
39. Ellis V, Scully MF, Kakkar VV. Plasminogen activation initiated by single-chain urokinase-type plasminogen activator. Potentiation by U937 monocytes. J Biol Chem 1989;264:2185-8.
40. Myohanen HT, Stephens RW, Hedman K, Tapiovaara H, Rsnne E, Hsyer-Hansen G, et al. Distribution and lateral mobility of the urokinase-receptor complex at the cell surface. J Histochem Cytochem 1993;41:1291- 1301.
41. Andreasen PA, Kjeller L, Christensen L, Duffy MJ. The urokinase-type plasminogen activator system in cancer metastasis: a review. Int J Cancer 1997;72:1-22.
42. Lindahl TL, Sigurdardottir 0, Wiman B. Stability of plasminogen activator inhibitor 1 (PAI-I). Thromb Haemost 1989;62:748-51.
43. Salonen EM, Vaheri A, Pollanen J, Stephens R, Andre- asen P, Mayer M, et al. Interaction of plasminogen acti- vator inhibitor (PAI-I) with vitronectin. J Biol Chem 1989;264:633943.
44. Cubellis MV, Wun TC, Blasi F. Receptor-mediated inter- nalization and degradation of urokinase is caused by its specific inhibitor PAI-I. EMBO J 1990;9:1079-85.
45. Nykjaer A, Conese M, Christensen EI, Olson D, Cre- mona 0, Gliemann J, Blasi E Recycling of the urokinase receptor upon internalization of the uPA:serpin com- plexes. EMBO J 1997;16:2610-20.
46. Stephens RW, Tapiovaara H, Reisberg T, Bizik J, Vaheri A. Alpha 2-macroglobulin restricts plasminogen acti- vation to the surface of RC2A leukemia cells. Cell Regul
47. Deng G, Curriden SA, Wang SJ, Rosenberg S, Losku- toff DJ. Is plasminogen activator inhibitor-I the mol- ecular switch that governs urokinase receptor-mediated cell adhesion and release. J Cell Biol 1996; 134: 1563-7 1.
48. Alitalo K, Vaheri A. Pericellular matrix in malignant transformation. Adv Cancer Res 1982;37:111-58.
49. Hsyer-Hansen G, Rsnne E, Solberg H, Behrendt N, Ploug M, Lund LR, et al. Urokinase plasminogen acti- vator cleaves its cell surface receptor releasing the ligand-binding domain. J Biol Chem 1992;267: 18224-9.
50. Solberg H, Rsmer J, Brunner N, Holm A, Sidenius N, Dan0 K, H~lyer-Hansen G. A cleaved form of the recep- tor for urokinase-type plasminogen activator in invasive transplanted human and murine tumors. Int J Cancer
51. Pedersen N, Schmitt M, Rsnne E, Nicoletti MI, Hsyer- Hansen G, Conese M, Giavazzi R, et al. A ligand-free, soluble urokinase receptor is present in the ascitic fluid
I99 1;2: 1057-65.
1994;58:877-81.
166

uPAR IN BLOOD
from patients with ovarian cancer. J Clin Invest 1993;92:2160-7.
52. Luther T, Magdolen V, Albrecht S, Kasper M, Riemer C, Kessler H, et al. Epitope-mapped monoclnal anti- bodies as tools for functional and morphological analy- ses of the human urokinase receptor in tumor tissue. Am J Pathol 1997;150:12314.
53. Rsnne E, Hayer-Hansen G, Brtinner N, Pedersen H, Rank F, Osborne CK, et al. Urokinase receptor in breast cancer tissue extracts. Enzyme-linked immuno- sorbent assay with a combination of mono- and poly- clonal antibodies. Breast Cancer Res Treat 1995; 199- 207.
54. Plesner T, Behrendt N, Ploug M. Structure, function and expression on blood and bone marrow cells of the urokinase-type plasminogen activator receptor, uPAR. Stem Cells 1997;15:398-408.
55. Wohn KD, Kanse SM, Deutsch V, Schmidt T, Eldor A, Preissner KT. The urokinase-receptor (CD87) is ex- pressed in cells of the megakaryoblastic lineage. Thromb Haemost 1997;77:540-7.
56. Ranne E, Pappot H, Grsndahl-Hansen J, Hsyer-Hans- en G, Plesner T, Hansen NE, Dan0 K. The receptor for urokinase plasminogen activator is present in plasma from healthy donors and elevated in patients with par- oxysmal nocturnal haemoglobinuria. Brit J Haematol 1995;89:57&81.
57. Dana K, Behrendt N, Brunner N, Ellis V, Plough M, Pyke C. The urokinase receptor. Protein structure and role in plasminogen activation and cancer invasion. Fibronolysis 1994;s: 189-213.
58. Chavakis T, Kanse SM, Yutzy B, Lijnen HR, Preissner KT. Vitronectin concentrates proteolytic activity on the cell surface and extracellular matrix by trapping soluble urokinase receptor-urokinase complexes. Blood 1998; 9 1 :2305-12.
59. Resnati M, Guttinger M, Valcamonica S, Sidenius N, Blasi F, Fazioli E Proteolytic cleavage of the urokinase receptor subsitutes for the agonist-induced chemotactic effect. EMBO J 1996;15:1572-82.
60. Hsyer-Hansen G, Behrendt N, Ploug M, Dane K, Pre- issner KT. The intact urokinase receptor is required for efficient vitronectin binding: receptor cleavage prevents ligand interaction. FEBS Letters 1997;420:79-85.
61. Wilhelm 0, Weidle U, Hohl S, Rettenberger P, Schmitt M, Graeff H. Recombinant soluble urokinase receptor as a scavenger for urokinase-type plasminogen activator (uPA). Inhibition of proliferation and invasion of hu- man ovarian cancer cells. FEBS Lett 1994;337:1314.
62. Luther T, Magdolen V, Albrecht S, Kasper M, Riemer C, Kessler H, et al. Epitope-mapped monoclonal anti- bodies as tools for functional and morphological analy- ses of the human urokinase receptor in tumor tissue. Am J Pathol 1997;150:123144.
167
Related Documents











