The effect of UV-A (352 nm) stress on chlorophyll fluorescence, chlorophyll a content, thickness of upper cortex and determinate DNA damage in Physcia semipinnata Dilek Unal * , _ Inci Tuney, Aylin Esiz-Dereboylu, Atakan Sukatar University of Ege, Faculty of Science, Department of Biology, 35100 Bornova-Izmir, Turkey article info Article history: Received 16 June 2008 Received in revised form 17 September 2008 Accepted 17 October 2008 Available online xxxx Keywords: Lichen Ultraviolet-A irradiation Photosynthesis RAPD-PCR abstract In the present study, Physcia semipinnata samples were exposed to UV-A (352 nm) in 20 J m 2 for 24 h, 48 h and 72 h to seek the alterations in the PSII photosynthetic quantum yield, thallus anatomy and DNA mutation rate in response to radiation. The Fv/Fm ratio decreased in P. semipinnata following expo- sure to UV-A for 24 h, 48 h and 72 h. The data of the present study reported that Chla degradation occurred by exposition of UV-A for 24 h, 48 h and 72 h. Photobiont size and thickness of upper cortex layer also showed a gradually decrease in P. semipinnata thallus sections during 24 h, 48 h and 72 h. The thickness of the upper cortex layer, exposed to UV-A for 72 h, of P. semipinnata thallus reduced 64%, compared with control. The random amplified polymorphic DNA (RAPD) technique was used to detect DNA damage. The main changes observed in the RAPD profiles resulted in both appearance and disappearance of different bands and variation of their intensity. According to RAPD assay, the genetic distance between the control group and 24 h, 48 h and 72 h UV-A exposed, groups was found 56%, 78% and 84%, respectively. Ó 2008 Elsevier B.V. All rights reserved. 1. Introduction Photosynthesis is the main target of UV radiation [1]. UV radia- tion affects photosystem II by two ways. Firstly, it may cause a di- rect damage on key components such as D1 protein of photosystem II, so that the photosynthetic pigments are lost [2,3] and the activation of rubisco is inhibited [4]. In addition to these effects, it was documented that UV-A induced oxidative stress, oxi- dative damage, decrease in photosynthetic efficiency and inhibi- tion of growth. It has been already shown that UV-A light has effect on induction of lipid peroxidation of biological membranes, polyunsaturated fatty acids and phospholipid liposomes [5]. De- spite of its importance related to the inhibition of photosynthesis, our knowledge about the effects of UV-A is eventually limited, compared with visible and UV-B effects. Besides, the harmful ef- fects of UV-A radiation on PS II function were manifested as inhibi- tion of electron transport [6], and degradation of D1 and D2 reaction center proteins. Nonetheless, the mechanistic details of these effects have not been clarified yet [6]. Similar to ionizing radiation, UV-A radiation generates oxygen and hydroxyl radicals which react with DNA to form monomeric photoproducts such as cytosine and thymine photohydrates as well as strand breaks and DNA-protein crosslinks [7]. In addition, of all the DNA lesions, strand breaks are perhaps the most dangerous since they disrupt the linear structure of the DNA and prevent replication and gene expression. The measurement of modulated chlorophyll (Chl)a fluorescence in lichen is one of the several methodologies used for the assess- ment of degree of many environmental stresses including temper- ature and osmotic stress [8], heavy metals [9], or air pollution [10]. Similarly, Chlorophyll a/b and Chlorophyll a degradation are useful ecological parameters for determining physiological conditions of plants, lichens and photobionts subjected to air pollution [11,12], heavy metals [13], high temperature [14], light and osmotic stress [15]. Over the past decade, several molecular techniques have been developed to provide information on diversity, genotoxicology, ge- netic relationships etc. One of these methods that are used for these aims is RAPD-PCR (Random Amplified Polymorphic DNA- Polymerase Chain Reaction). The RAPD procedures were first developed in 1990 [16,17] using PCR to amplify anonymous seg- ments of nuclear DNA with identical primers which are 10 bp in length. Genetic variation and divergence within and between the taxa of interest are assessed by the presence or absence of each product. The RAPD primers are commercially available and do 1011-1344/$ - see front matter Ó 2008 Elsevier B.V. All rights reserved. doi:10.1016/j.jphotobiol.2008.10.004 * Corresponding author. E-mail addresses: [email protected], [email protected] (D. Unal). Journal of Photochemistry and Photobiology B: Biology 94 (2009) 71–76 Contents lists available at ScienceDirect Journal of Photochemistry and Photobiology B: Biology journal homepage: www.elsevier.com/locate/jphotobiol ARTICLE IN PRESS

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Journal of Photochemistry and Photobiology B: Biology 94 (2009) 71–76
ARTICLE IN PRESS
Contents lists available at ScienceDirect
Journal of Photochemistry and Photobiology B: Biology
journal homepage: www.elsevier .com/locate/ jphotobiol
The effect of UV-A (352 nm) stress on chlorophyll fluorescence,chlorophyll a content, thickness of upper cortex and determinateDNA damage in Physcia semipinnata
Dilek Unal *, _Inci Tuney, Aylin Esiz-Dereboylu, Atakan SukatarUniversity of Ege, Faculty of Science, Department of Biology, 35100 Bornova-Izmir, Turkey
a r t i c l e i n f o
Article history:Received 16 June 2008Received in revised form 17 September2008Accepted 17 October 2008Available online xxxx
Keywords:LichenUltraviolet-A irradiationPhotosynthesisRAPD-PCR
1011-1344/$ - see front matter � 2008 Elsevier B.V. Adoi:10.1016/j.jphotobiol.2008.10.004
* Corresponding author.E-mail addresses: [email protected], duna
a b s t r a c t
In the present study, Physcia semipinnata samples were exposed to UV-A (352 nm) in 20 J m�2 for 24 h,48 h and 72 h to seek the alterations in the PSII photosynthetic quantum yield, thallus anatomy andDNA mutation rate in response to radiation. The Fv/Fm ratio decreased in P. semipinnata following expo-sure to UV-A for 24 h, 48 h and 72 h. The data of the present study reported that Chla degradationoccurred by exposition of UV-A for 24 h, 48 h and 72 h. Photobiont size and thickness of upper cortexlayer also showed a gradually decrease in P. semipinnata thallus sections during 24 h, 48 h and 72 h.The thickness of the upper cortex layer, exposed to UV-A for 72 h, of P. semipinnata thallus reduced64%, compared with control. The random amplified polymorphic DNA (RAPD) technique was used todetect DNA damage. The main changes observed in the RAPD profiles resulted in both appearance anddisappearance of different bands and variation of their intensity. According to RAPD assay, the geneticdistance between the control group and 24 h, 48 h and 72 h UV-A exposed, groups was found 56%, 78%and 84%, respectively.
� 2008 Elsevier B.V. All rights reserved.
1. Introduction
Photosynthesis is the main target of UV radiation [1]. UV radia-tion affects photosystem II by two ways. Firstly, it may cause a di-rect damage on key components such as D1 protein ofphotosystem II, so that the photosynthetic pigments are lost [2,3]and the activation of rubisco is inhibited [4]. In addition to theseeffects, it was documented that UV-A induced oxidative stress, oxi-dative damage, decrease in photosynthetic efficiency and inhibi-tion of growth. It has been already shown that UV-A light haseffect on induction of lipid peroxidation of biological membranes,polyunsaturated fatty acids and phospholipid liposomes [5]. De-spite of its importance related to the inhibition of photosynthesis,our knowledge about the effects of UV-A is eventually limited,compared with visible and UV-B effects. Besides, the harmful ef-fects of UV-A radiation on PS II function were manifested as inhibi-tion of electron transport [6], and degradation of D1 and D2reaction center proteins. Nonetheless, the mechanistic details ofthese effects have not been clarified yet [6]. Similar to ionizingradiation, UV-A radiation generates oxygen and hydroxyl radicals
ll rights reserved.
[email protected] (D. Unal).
which react with DNA to form monomeric photoproducts such ascytosine and thymine photohydrates as well as strand breaks andDNA-protein crosslinks [7]. In addition, of all the DNA lesions,strand breaks are perhaps the most dangerous since they disruptthe linear structure of the DNA and prevent replication and geneexpression.
The measurement of modulated chlorophyll (Chl)a fluorescencein lichen is one of the several methodologies used for the assess-ment of degree of many environmental stresses including temper-ature and osmotic stress [8], heavy metals [9], or air pollution [10].Similarly, Chlorophyll a/b and Chlorophyll a degradation are usefulecological parameters for determining physiological conditions ofplants, lichens and photobionts subjected to air pollution [11,12],heavy metals [13], high temperature [14], light and osmotic stress[15].
Over the past decade, several molecular techniques have beendeveloped to provide information on diversity, genotoxicology, ge-netic relationships etc. One of these methods that are used forthese aims is RAPD-PCR (Random Amplified Polymorphic DNA-Polymerase Chain Reaction). The RAPD procedures were firstdeveloped in 1990 [16,17] using PCR to amplify anonymous seg-ments of nuclear DNA with identical primers which are 10 bp inlength. Genetic variation and divergence within and between thetaxa of interest are assessed by the presence or absence of eachproduct. The RAPD primers are commercially available and do

Table 1RAPD primers used for PCR amplification.
RAPD Primers
0PA9 (Primer 1) 5-GGG TAA CGC C-30PB14 (Primer 2) 5-TCC GCT CTG G-3OPB5 (Primer 3) 5-TGC GCC CTT C-3OPB6 (Primer 4) 5-TGC TCT GCC C-3OPB7 (Primer 5) 5-GGT GAC GCA G-3OPB8 (Primer 6) 5-GTC CAC ACG G-3OPB10 (Primer 7) 5-CTG CTG GGA C-3
72 D. Unal et al. / Journal of Photochemistry and Photobiology B: Biology 94 (2009) 71–76
ARTICLE IN PRESS
not require prior knowledge of the target DNA sequence. RAPDmarkers have been used for species identification, analysis of pop-ulation structure, analysis of genetic impact of environmentalstress and analysis of genetic diversity.
Although the number of reports regarding the effects of en-hanced UV irradiation on lichens has considerably increased in re-cent years, our knowledge about avoidance mechanisms onenhanced UV-B radiation and its effects on photosynthesis in li-chens is mainly restricted to UV-absorbing substances. In addition,there is little known about other avoidance mechanisms in lichensand there are few experiments on the effect(s) of UV-A irradiation.The aim of the present study was to evaluate time-and irradiance-relationships of UV-A damages on photosynthesis, chlorophyll acontent and thickness of upper cortex and algal size in Physciasemipinnata thallus. UV-A which induced DNA damage was ana-lyzed with the RAPD-PCR technique.
2. Material and method
2.1. Material
Physcia semipinnata samples were collected from tree branchesin Karagöl, Izmir-Turkey (38�330 N 27�130 E, 840 m) in November2007. Lichen samples were transferred to laboratory, were rinsedthree times (10 s each) to minimize dust contamination and thendivided into several groups. Experiments were conducted within3–4 days after collection. P. semipinnata samples were exposed toUV-A in 20 Jm�2 (352 nm, 50 Hz, 0.60 Amps, model; XX-20BLB)in Petri dishes for 24 h, 48 h and 72 h at 25 �C.
2.2. Chlorophyll a fluorescence
Chlorophyll a fluorescence of samples was measured with aplant efficiency analyzer (Handy PEA, Hansatech). Before measure-ment of chlorophyll fluorescence, thalli were dark-adapted for15 min. The Fv/Fm parameters were calculated with the instru-ment from fluorescence induction curves of 5 s duration recordedat an irradiance of 1800 lmol m�2s�1 from light emitting diodes.The Fv/Fm parameter (maximum quantum yield efficiency of PSII)was used as a stress indicator. Each treatment comprised of tenreplicates.
2.3. Determination of chla content
About 20 mg thalli were extracted in the dark for 1 h at 65 �C in3 ml dimethyl sulfoxide (DMSO) in the presence of polyvinylpo-lyrrolidone to minimize chlorophyll degradation. To assess chloro-phylls, absorbance of the extracts was then read at 665.1 nm,649.1 nm and 435 nm [18]. Each treatment comprised of threereplicates.
2.4. Size of algal cells in lichen thallus and thickness of upper fungaland algal layer
For anatomical studies, thallus sections about in 15 lm thick-ness were taken and examined under a light microscope (OlympusCH40 microscope made in Japanese) Anatomical features weremeasured in 25 different areas of each section that was taken fromP. semipinnata thallus.
2.5. Isolation of DNA
After UV treatment, DNA was isolated from P. semipinnata sam-ples by CTAB method [19].
2.6. RAPD-PCR
Amplification of genomic DNA was performed in a 50 ll reac-tion mixture containing 18.5 Mili Q water, 1.75 ll of 10� Taq poly-merase buffer, 1.2 ll of MgCl2, 1.5 ll of dNTPs, 0.6 ll of eachprimer and 0.6 ll of Taq DNA polymerase before addition of DNA.
RAPD-PCR was performed with 7 different RAPD primers (Table1), following control of DNAs by agarose gel electrophoresis. PCRbands were checked again by agarose gel electrophoresis followingPCR analysis. PCR analysis was performed three times for control.
Amplification was performed in ABI Prisim 9700 Thermal Cyclerwith 5 cycles for denaturation at 94 �C for 1 min, annealing at 40 �Cfor 30 s and extension at 72 �C for 1 min, and then 40 cycles fordenaturation at 94 �C for 45 s, annealing at 60 �C for 1 min andextension at 72 �C for 45 s, final extension at 72 �C for 11 min.Amplified products were kept in 4 �C.
9 ll of each PCR product was electrophoresed on agarose gel(%1.2) in TAE buffer (40 mM Tris acetate, 1 mM EDTA, pH 8.0) at80 V for 1 h. The gel was stained with ethidium bromide andphotographed.
2.7. Estimation of genomic template stability
Each change that was observed in RAPD profiles was given as +1arbitrary score. The average was then calculated for each experi-mental group exposed to UV irradiance. Template genomic stabil-ity (%) was calculated as ‘‘100�(100a/n)” where ‘‘a” is the averagenumber of changes in DNA profiles and ‘‘n” the number of bandsthat were selected in control DNA profiles.
2.8. Statistical analysis
Statistical analyses were performed using SPSS for Windows,including one-way analysis of variance (ANOVA) and Tuckey‘spairwise comparisons.
3. Results
3.1. Fv/Fm values
In this research, photoinhibition was measured with a perma-nent reduction in maximal PSII efficiency (Fv/Fm). These resultsshowed a gradually decrease in the Fv/Fm values during 24 h,48 h and 72 h UV-treatment conditions. The 0.5, 0.2 and 0.1 Fv/Fm, approximate values indicated that there was a photodestruc-tive effect on photosystem II (Table 2. 0.5, 0.2 and 0.1 Fv/Fm valuesas indicated by ANOVA).
3.2. Chla content
The data of present work showed that chla content was signif-icantly (p < 0.05) decreased by exposition to UV-A for 24 h, 48 hand 72 h (chla content values of samples were approx. 23.98,8.89 and 6.65 mg/g dw, respectively, as seen in Fig. 1). However,

Table 2Photosythetic quantum yield (Fv/Fm) of P. semipinnata exposed to UV-A light for 24 h,48 h and 72 h and non-exposed to UV-A light (as a control).
n Control 24 h 48 h 72 h
Fv/Fm 10 0.717 ± 0.02 0.548 ± 0.03 0.226 ± 0.04 0.152 ± 0.04AnovaF ratio 77.61F probability 0.005
Chla content
05
101520253035404550
control 24 h 48 h 72 h
mg/
g dw
Fig. 1. Content of Chla of P. semipinnata. exposed to UV-A for 24 h, 48 h and 72 hand non-treated (as a control). Means with common letters are significantlydifferent at P 6 0.05, according to Tuckey’s test.
D. Unal et al. / Journal of Photochemistry and Photobiology B: Biology 94 (2009) 71–76 73
ARTICLE IN PRESS
samples, non-treated with UV-A, showed that chla content washigher than other groups (chla content values were approx.38.9 mg/g dw as seen in Fig. 1).
3.3. Size of algal cells in lichen thallus and thickness of upper fungaland algal layer
Projected area of algal cells in the sections of thallus segmentswas used as a parameter for estimation of the algal cell size. Itwas measured in P. semipinnata after full-hydrated thallus sectionsfrom non-treated UV-A (control group condition) or the 24 h, 48 hand 72 h UV-A treated groups. Photobiont size gradually decreasedin P. semipinnata thallus during 24 h, 48 h and 72 h UV-treatmentconditions (Table 3). In the control group, the areas of the photobi-ont cells in P. semipinnata thallus were significantly greater(p < 0.05) compared with the 72 h UV-A exposed group (Table 3).
Our results showed a significant (p < 0.05) decrease in the thick-ness of upper cortex layer during 24 h, 48 h and 72 h UV-treatmentconditions (Table 3). The thickness of upper cortex layer in P. semi-pinnata thallus exposed to UV-A for 72 h reduced about 64 % com-pared with control.
3.4. UV-A stress on DNA
Total seven 10-mer priming oligo-nucleotides were used toanalyze the results; however four 10-mer priming oligo-nucleo-tides gave reproducible bands in all samples. In these samples,RAPD patterns generated by UV-exposed P. semipinnata were dif-
Table 3Size of algal cells in lichen thallus and thickness of upper fungal and algal layerexposed to UV-A light for 24 h, 48 h and 72 h and non-exposed to UV-A light (as acontrol).
n Algal layer(lm)
Upper fungallayer (lm)
Algal cellsize (lm)
Control 25 51. 880 ± 0. 846cd 189. 200 ± 0.860d 11.700 ± 0.387d
24 h UV-A 25 45.820 ± 0.930 165.600 ± 0.680d 11.250 ± 0.38148 h UV-A 25 44.120 ± 1.101a 177.200 ± 0.706d 10.900 ± 0.44272 h UV-A 25 39.900 ± 0.504a 125.600 ± 0.793abc 9.950 ± 0.357a
ferent from those obtained from control DNA. The principal eventsobserved in the UV exposed groups were variation in the bandintensity and the disappearance or appearance new bands (Figs.2a and 2b). The differentiation in the band intensity and appear-ance or disappearance of some bands may correlate with level ofphotoproducts in DNA template and structural changes in DNAsuch as deletions, insertions or breaks after UV treatment. Tables4 and 5 present a summary of all RAPD profile modifications. Thedecrease in band intensity was obvious for all UV-A exposed sam-ples. The highest increased band intensity occurred in the 72 h UV-exposed group. The number of disappeared RAPD bands which oc-curred for all UV treatments was higher at 48 h UV exposition time.Bands with a molecular size bigger than 1Boca Ratonkb disap-peared in all the UV-A treated groups. Finally, some extra bandsappeared in the three irradiance exposition periods; however thenumber of appeared RAPD bands was higher in the 72 h UV expo-sition time.
4. Discussion
Although it was shown that UV-A radiation caused damage onPSII in higher plants, lichens and animals [6,20], UV-B radiationhas been generally studied in lichens. Sass and Vass [2] foundout remarkably high tolerance in photosystem II against short termand high intensity of UV-B radiation in Cladonia convolute and Pelti-gera aphthosa lichens. The mycobiont of Xanthoria elegans has alsobeen found to have high resistance to extreme UV irradiances [21].Measurement of chlorophyll fluorescence and thermolumines-cense indicated that lichens are remarkably tolerant to extremeradiation, which kills vascular plants within a day [22]. Maximalphootosytem II efficiency (fluorescence ratio Fv/Fm) has been fre-quently used to monitor in photosynthetic organism. In lichens,decrease of this parameter has been utilized to assess environmen-tal stress on photobiont. Under non-stressed conditions, lichentypically possesses Fv/Fm in the range of 0.45–0.65 [23]. In this re-search, our results showed a gradual decrease in the Fv/Fm valuesunder the 24 h, 48 h and 72 h UV-treatment conditions. Mean Fv/Fm values of about 0.54, determined in this study for P. semipinnatathalli, are similar to values reported for healthy lichens. However,the obtained approx. values of Fv/Fm, 0.2 and 0.1, indicated thatthere was a photodestructive effect on photosystem II (Table 2. ap-prox. 0.5, 0.2 and 0.1 Fv/Fm values indicated by ANOVA). Chloro-phyll degradation is also used as a parameter of being inlichenological studies focused on air pollution and heavy metalsfrequently [10,24,25]. In lichens, to exposure to heavy metalscaused chlorophyll a concentrations to decrease [13,24]. This de-crease has been attributed to inhibition of the reductive steps ofbiosynthesis [24]. In the present study, the chlorophyll degradationalso gradually occurred in all experimental groups except in thecontrol group during UV-A exposure. Similarly results were ob-tained in a study by Gauslaa and Solhaug [26] were exposed tohigh light levels for prolonged periods, showing higher sensitivityto photoinhibition and increased chlorophyll degradation.
Functions of the upper cortex include protection of the Photobi-ont against excessive light, since various light-absorbing com-pounds are located in the upper cortical tissue of the vegetativeand generative parts of the thallus [27]. In a fully sun-exposed hab-itat, the thickness of the cortex was reduced [27,28]. In the presentstudy, thallus sections showed that thickness of upper fungal layerwas significantly influenced by exposition to UV-A radiationdepending on exposition time. Photobiont size gradually decreasedin P. semipinnata thallus under the 24 h, 48 h and 72 h UV-treat-ment conditions (Table 3). In the control samples of P. semipinnatathallus, the layers of the photobiont cells were about 86% greaterthan those in the 72 h UV-A exposed group (Table 3).
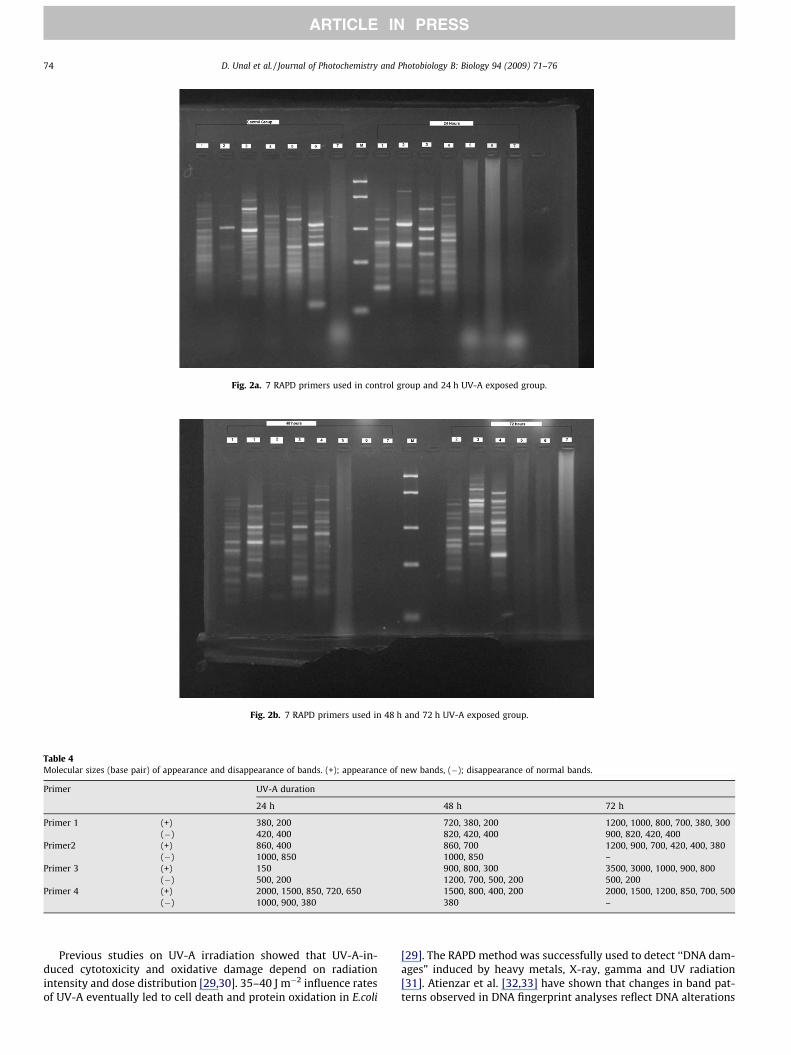
Fig. 2a. 7 RAPD primers used in control group and 24 h UV-A exposed group.
Fig. 2b. 7 RAPD primers used in 48 h and 72 h UV-A exposed group.
Table 4Molecular sizes (base pair) of appearance and disappearance of bands. (+); appearance of new bands, (�); disappearance of normal bands.
Primer UV-A duration
24 h 48 h 72 h
Primer 1 (+) 380, 200 720, 380, 200 1200, 1000, 800, 700, 380, 300(�) 420, 400 820, 420, 400 900, 820, 420, 400
Primer2 (+) 860, 400 860, 700 1200, 900, 700, 420, 400, 380(�) 1000, 850 1000, 850 –
Primer 3 (+) 150 900, 800, 300 3500, 3000, 1000, 900, 800(�) 500, 200 1200, 700, 500, 200 500, 200
Primer 4 (+) 2000, 1500, 850, 720, 650 1500, 800, 400, 200 2000, 1500, 1200, 850, 700, 500(�) 1000, 900, 380 380 –
74 D. Unal et al. / Journal of Photochemistry and Photobiology B: Biology 94 (2009) 71–76
ARTICLE IN PRESS
Previous studies on UV-A irradiation showed that UV-A-in-duced cytotoxicity and oxidative damage depend on radiationintensity and dose distribution [29,30]. 35–40 J m�2 influence ratesof UV-A eventually led to cell death and protein oxidation in E.coli
[29]. The RAPD method was successfully used to detect ‘‘DNA dam-ages” induced by heavy metals, X-ray, gamma and UV radiation[31]. Atienzar et al. [32,33] have shown that changes in band pat-terns observed in DNA fingerprint analyses reflect DNA alterations
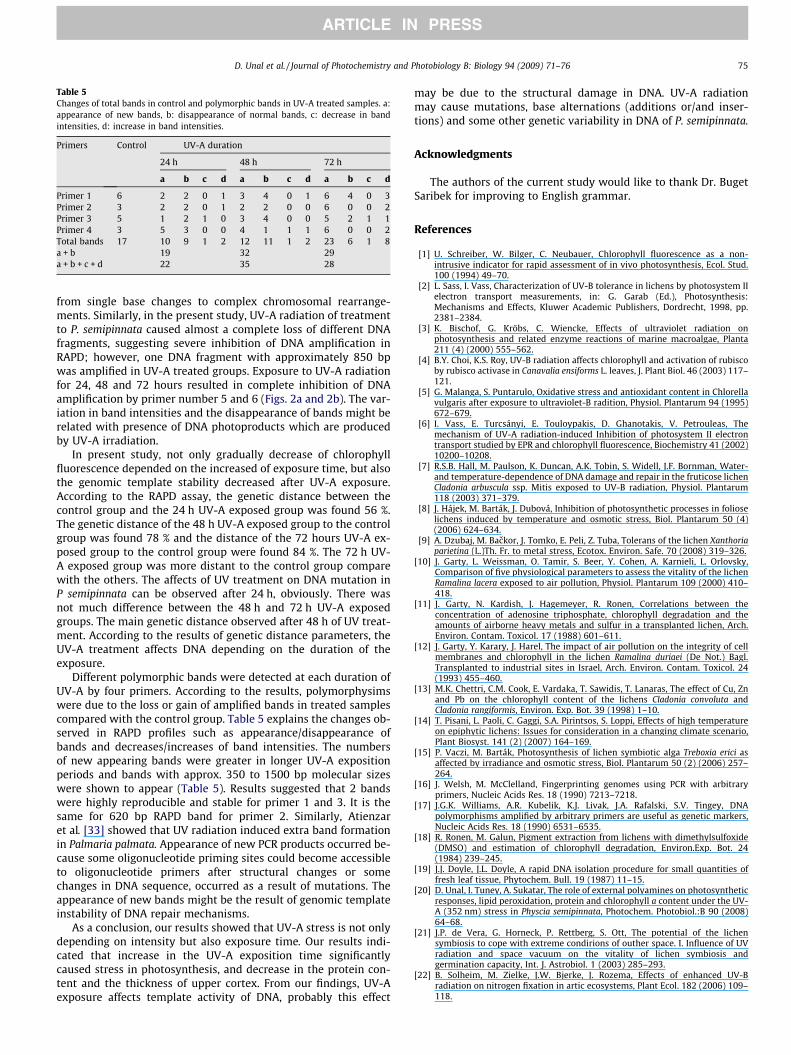
Table 5Changes of total bands in control and polymorphic bands in UV-A treated samples. a:appearance of new bands, b: disappearance of normal bands, c: decrease in bandintensities, d: increase in band intensities.
Primers Control UV-A duration
24 h 48 h 72 h
a b c d a b c d a b c d
Primer 1 6 2 2 0 1 3 4 0 1 6 4 0 3Primer 2 3 2 2 0 1 2 2 0 0 6 0 0 2Primer 3 5 1 2 1 0 3 4 0 0 5 2 1 1Primer 4 3 5 3 0 0 4 1 1 1 6 0 0 2Total bands 17 10 9 1 2 12 11 1 2 23 6 1 8a + b 19 32 29a + b + c + d 22 35 28
D. Unal et al. / Journal of Photochemistry and Photobiology B: Biology 94 (2009) 71–76 75
ARTICLE IN PRESS
from single base changes to complex chromosomal rearrange-ments. Similarly, in the present study, UV-A radiation of treatmentto P. semipinnata caused almost a complete loss of different DNAfragments, suggesting severe inhibition of DNA amplification inRAPD; however, one DNA fragment with approximately 850 bpwas amplified in UV-A treated groups. Exposure to UV-A radiationfor 24, 48 and 72 hours resulted in complete inhibition of DNAamplification by primer number 5 and 6 (Figs. 2a and 2b). The var-iation in band intensities and the disappearance of bands might berelated with presence of DNA photoproducts which are producedby UV-A irradiation.
In present study, not only gradually decrease of chlorophyllfluorescence depended on the increased of exposure time, but alsothe genomic template stability decreased after UV-A exposure.According to the RAPD assay, the genetic distance between thecontrol group and the 24 h UV-A exposed group was found 56 %.The genetic distance of the 48 h UV-A exposed group to the controlgroup was found 78 % and the distance of the 72 hours UV-A ex-posed group to the control group were found 84 %. The 72 h UV-A exposed group was more distant to the control group comparewith the others. The affects of UV treatment on DNA mutation inP semipinnata can be observed after 24 h, obviously. There wasnot much difference between the 48 h and 72 h UV-A exposedgroups. The main genetic distance observed after 48 h of UV treat-ment. According to the results of genetic distance parameters, theUV-A treatment affects DNA depending on the duration of theexposure.
Different polymorphic bands were detected at each duration ofUV-A by four primers. According to the results, polymorphysimswere due to the loss or gain of amplified bands in treated samplescompared with the control group. Table 5 explains the changes ob-served in RAPD profiles such as appearance/disappearance ofbands and decreases/increases of band intensities. The numbersof new appearing bands were greater in longer UV-A expositionperiods and bands with approx. 350 to 1500 bp molecular sizeswere shown to appear (Table 5). Results suggested that 2 bandswere highly reproducible and stable for primer 1 and 3. It is thesame for 620 bp RAPD band for primer 2. Similarly, Atienzaret al. [33] showed that UV radiation induced extra band formationin Palmaria palmata. Appearance of new PCR products occurred be-cause some oligonucleotide priming sites could become accessibleto oligonucleotide primers after structural changes or somechanges in DNA sequence, occurred as a result of mutations. Theappearance of new bands might be the result of genomic templateinstability of DNA repair mechanisms.
As a conclusion, our results showed that UV-A stress is not onlydepending on intensity but also exposure time. Our results indi-cated that increase in the UV-A exposition time significantlycaused stress in photosynthesis, and decrease in the protein con-tent and the thickness of upper cortex. From our findings, UV-Aexposure affects template activity of DNA, probably this effect
may be due to the structural damage in DNA. UV-A radiationmay cause mutations, base alternations (additions or/and inser-tions) and some other genetic variability in DNA of P. semipinnata.
Acknowledgments
The authors of the current study would like to thank Dr. BugetSaribek for improving to English grammar.
References
[1] U. Schreiber, W. Bilger, C. Neubauer, Chlorophyll fluorescence as a non-intrusive indicator for rapid assessment of in vivo photosynthesis, Ecol. Stud.100 (1994) 49–70.
[2] L. Sass, I. Vass, Characterization of UV-B tolerance in lichens by photosystem IIelectron transport measurements, in: G. Garab (Ed.), Photosynthesis:Mechanisms and Effects, Kluwer Academic Publishers, Dordrecht, 1998, pp.2381–2384.
[3] K. Bischof, G. Kröbs, C. Wiencke, Effects of ultraviolet radiation onphotosynthesis and related enzyme reactions of marine macroalgae, Planta211 (4) (2000) 555–562.
[4] B.Y. Choi, K.S. Roy, UV-B radiation affects chlorophyll and activation of rubiscoby rubisco activase in Canavalia ensiforms L. leaves, J. Plant Biol. 46 (2003) 117–121.
[5] G. Malanga, S. Puntarulo, Oxidative stress and antioxidant content in Chlorellavulgaris after exposure to ultraviolet-B radition, Physiol. Plantarum 94 (1995)672–679.
[6] I. Vass, E. Turcsányi, E. Touloypakis, D. Ghanotakis, V. Petrouleas, Themechanism of UV-A radiation-induced Inhibition of photosystem II electrontransport studied by EPR and chlorophyll fluorescence, Biochemistry 41 (2002)10200–10208.
[7] R.S.B. Hall, M. Paulson, K. Duncan, A.K. Tobin, S. Widell, J.F. Bornman, Water-and temperature-dependence of DNA damage and repair in the fruticose lichenCladonia arbuscula ssp. Mitis exposed to UV-B radiation, Physiol. Plantarum118 (2003) 371–379.
[8] J. Hájek, M. Barták, J. Dubová, Inhibition of photosynthetic processes in folioselichens induced by temperature and osmotic stress, Biol. Plantarum 50 (4)(2006) 624–634.
[9] A. Dzubaj, M. Backor, J. Tomko, E. Peli, Z. Tuba, Tolerans of the lichen Xanthoriaparietina (L.)Th. Fr. to metal stress, Ecotox. Environ. Safe. 70 (2008) 319–326.
[10] J. Garty, L. Weissman, O. Tamir, S. Beer, Y. Cohen, A. Karnieli, L. Orlovsky,Comparison of five physiological parameters to assess the vitality of the lichenRamalina lacera exposed to air pollution, Physiol. Plantarum 109 (2000) 410–418.
[11] J. Garty, N. Kardish, J. Hagemeyer, R. Ronen, Correlations between theconcentration of adenosine triphosphate, chlorophyll degradation and theamounts of airborne heavy metals and sulfur in a transplanted lichen, Arch.Environ. Contam. Toxicol. 17 (1988) 601–611.
[12] J. Garty, Y. Karary, J. Harel, The impact of air pollution on the integrity of cellmembranes and chlorophyll in the lichen Ramalina duriaei (De Not.) Bagl.Transplanted to industrial sites in Israel, Arch. Environ. Contam. Toxicol. 24(1993) 455–460.
[13] M.K. Chettri, C.M. Cook, E. Vardaka, T. Sawidis, T. Lanaras, The effect of Cu, Znand Pb on the chlorophyll content of the lichens Cladonia convoluta andCladonia rangiformis, Environ. Exp. Bot. 39 (1998) 1–10.
[14] T. Pisani, L. Paoli, C. Gaggi, S.A. Pirintsos, S. Loppi, Effects of high temperatureon epiphytic lichens: Issues for consideration in a changing climate scenario,Plant Biosyst. 141 (2) (2007) 164–169.
[15] P. Vaczi, M. Barták, Photosynthesis of lichen symbiotic alga Treboxia erici asaffected by irradiance and osmotic stress, Biol. Plantarum 50 (2) (2006) 257–264.
[16] J. Welsh, M. McClelland, Fingerprinting genomes using PCR with arbitraryprimers, Nucleic Acids Res. 18 (1990) 7213–7218.
[17] J.G.K. Williams, A.R. Kubelik, K.J. Livak, J.A. Rafalski, S.V. Tingey, DNApolymorphisms amplified by arbitrary primers are useful as genetic markers,Nucleic Acids Res. 18 (1990) 6531–6535.
[18] R. Ronen, M. Galun, Pigment extraction from lichens with dimethylsulfoxide(DMSO) and estimation of chlorophyll degradation, Environ.Exp. Bot. 24(1984) 239–245.
[19] J.J. Doyle, J.L. Doyle, A rapid DNA isolation procedure for small quantities offresh leaf tissue, Phytochem. Bull. 19 (1987) 11–15.
[20] D. Unal, I. Tuney, A. Sukatar, The role of external polyamines on photosyntheticresponses, lipid peroxidation, protein and chlorophyll a content under the UV-A (352 nm) stress in Physcia semipinnata, Photochem. Photobiol.:B 90 (2008)64–68.
[21] J.P. de Vera, G. Horneck, P. Rettberg, S. Ott, The potential of the lichensymbiosis to cope with extreme condirions of outher space. I. Influence of UVradiation and space vacuum on the vitality of lichen symbiosis andgermination capacity, Int. J. Astrobiol. 1 (2003) 285–293.
[22] B. Solheim, M. Zielke, J.W. Bjerke, J. Rozema, Effects of enhanced UV-Bradiation on nitrogen fixation in artic ecosystems, Plant Ecol. 182 (2006) 109–118.

76 D. Unal et al. / Journal of Photochemistry and Photobiology B: Biology 94 (2009) 71–76
ARTICLE IN PRESS
[23] L. Kappen, B. Schroeter, T.G.A. Green, R.D. Seppelt, Chlorophyll a fluorescenceand CO2 exchange of Umbilicaria aprina under extreme light stress in the cold,Oecologia 113 (1998) 325–331.
[24] M. Backor, D. Fahselt, R. Davidson, C.T. Wu, Effects of copper on wild andtolerant strains of the lichen photobont Trebouxia erici and possible tolerancemechanisms, Arch. Environ. Contam. Toxicol. 45 (2003) 159–167.
[25] J. Garty, Y. Karary, J. Harel, Effect of low pH, heavy metals and anions onchlorophyll degradation in the lichen Ramalina duriaei (De Not.) Bagl, Environ.Exp. Bot. 32 (1992) 229–241.
[26] Y. Gauslaa, K.A. Solhaug, High-light damage in air-dry thalli of the old forestlichen Lobaria pulmonaria-interactions of irradiance, exposure duration andhigh temperature, J. Exp. Bot. 50 (1999) 697–705.
[27] T.H. Nash III, Lichen Biology, Campridge University Press, New York, 1996. pp.171.
[28] L. Kappen, Ecophysiological relationships in different climatic regions. in: M.Galun (Ed.), Handbook of Lichenology, vol. 1. Boca Raton, 1988, pp. 37–99.
[29] J.D. Hoerter, A.A. Arnold, D.A. Kuczynska, A. Shibuya, C.S. Ward, M.G. Sauer, A.Gizachew, T.M. Hotchkiss, T.J. Fleming, S. Johnson, Effects of Sublethal UV-A
irradiation on activity levels of oxidative defense enzymes and proteinoxidation in Escherichia coli, J. Photochem. Photobiol: B 81 (2005) 171–180.
[30] H. Merwald, G. Klosner, C. Kokesch, M. Der-Petrossian, H. Honigsmann,F. Trautinger, UV-A-induced oxidative damage and cytotocity dependon the mode of exposure, J. Photochem. Photobiol.:B 79 (2005) 197–207.
[31] S. Kuroda, H. Yano, Y. Koga-Ban, Y. Tabei, F. Takaiwa, T. Kayano, H. Tanaka,Identification of DNA polymorphism induced by X-ray and UV irradiation inplant cells, Jpn. Agric. Res. Quart. 33 (1999) 223–226.
[32] A.F. Atienzar, M. Conradi, A. Evenden, A. Jha, M. Depledge, Qualitativeassessment of genotoxicity using RAPD: comparison of genomic templatestability with key fitness parameters in Daphnia magna exposed tobenzo(a)pyrene, Environ. Toxicol. Chem. 18 (1999) 2275–2282.
[33] A.F. Atienzar, B. Cordi, E.M. Donkin, J.A. Evenden, N.A. Jha, H. Depledge,Comparison of ultraviolet-induced genotoxicity detected by random amplifiedpolyamorphic DNA with chlorophyll fluorescence and growth in a marinemacroalgae, Palmaria palmate, Aquatic. Toxicol. 50 (2000) 1–12.
Related Documents











