The allogeneic effect revisited: exogenous help for endogenous, tumor-specific T cells Heather J. Symons 1,2 , Moshe Y. Levy 1,3 , Jie Wang 1 , Xiaotao Zhou 1 , Gang Zhou 1 , Sarah E. Cohen 4 , Leo Luznik 3 , Hyam I. Levitsky 1,3 , and Ephraim J. Fuchs 1,3 1Division of Cancer Immunology, Sidney Kimmel Comprehensive Cancer Center at Johns Hopkins, Baltimore, MD 2Division of Pediatric Oncology, Sidney Kimmel Comprehensive Cancer Center at Johns Hopkins, Baltimore, MD 3Division of Hematologic Malignancies, Sidney Kimmel Comprehensive Cancer Center at Johns Hopkins, Baltimore, MD 4Thomas Jefferson Medical College, Philadelphia, PA Abstract The “allogeneic effect” refers to the induction of host B cell antibody synthesis or host T cell cytotoxicity, including tumoricidal activity, by an infusion of allogeneic lymphocytes. We show that treatment of mice with cyclophosphamide (Cy) followed by CD8 + T cell-depleted allogeneic donor lymphocyte infusion (Cy + CD8 − DLI) induces regression of established tumors with minimal toxicity in models of both hematologic and solid cancers, even though the donor cells are eventually rejected by the host immune system. The optimal anti-tumor effect of Cy + CD8 − DLI required the presence of donor CD4 + T cells, host CD8 + T cells, and alloantigen expression by normal host but not tumor tissue. The results support a model in which a donor CD4+ T cell-mediated graft-versus- host reaction effectively awakens anti-tumor immunity among Cy-resistant host CD8 + T cells. These events provide the cellular mechanism of the “allogeneic effect” in anti-tumor immunity. Cy + CD8 − DLI may be an effective and minimally toxic strategy for awakening the host immune response to advanced cancers. Introduction Allogeneic blood or marrow transplantation (alloBMT) is a well-established therapy for hematologic malignancies. The graft-versus-leukemia, or GVL, effect of alloBMT is perhaps the best evidence that T cells can mediate the destruction of advanced, metastatic human malignancies via direct killing of tumor cells by donor T cells reactive to host histocompatibility antigens [1–8]. Despite the curative potential of allogeneic T cells, alloBMT as a procedure is severely hampered by three factors: (a) toxicity, (b) lack of efficacy, especially in solid tumors, and (c) limited availability of human leukocyte antigen (HLA)-identical donors. The conditioning regimen, infection, and GVHD all contribute to toxicity, and lack of efficacy manifests itself as progression of the underlying malignancy after transplantation [9, 10]. Corresponding Author: Ephraim Joseph Fuchs, M.D., 488 Blunting-Blaustein Cancer Research Building, 1650 Orleans St, Baltimore, MD 21231, E-mail:[email protected], Phone: 410-955-8143, Fax: 410-502-7163. Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customers we are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resulting proof before it is published in its final citable form. Please note that during the production process errors may be discovered which could affect the content, and all legal disclaimers that apply to the journal pertain. NIH Public Access Author Manuscript Biol Blood Marrow Transplant. Author manuscript; available in PMC 2009 May 1. Published in final edited form as: Biol Blood Marrow Transplant. 2008 May ; 14(5): 499–509. NIH-PA Author Manuscript NIH-PA Author Manuscript NIH-PA Author Manuscript

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

The allogeneic effect revisited: exogenous help for endogenous,tumor-specific T cells
Heather J. Symons1,2, Moshe Y. Levy1,3, Jie Wang1, Xiaotao Zhou1, Gang Zhou1, Sarah E.Cohen4, Leo Luznik3, Hyam I. Levitsky1,3, and Ephraim J. Fuchs1,3
1Division of Cancer Immunology, Sidney Kimmel Comprehensive Cancer Center at Johns Hopkins,Baltimore, MD
2Division of Pediatric Oncology, Sidney Kimmel Comprehensive Cancer Center at Johns Hopkins, Baltimore,MD
3Division of Hematologic Malignancies, Sidney Kimmel Comprehensive Cancer Center at Johns Hopkins,Baltimore, MD
4Thomas Jefferson Medical College, Philadelphia, PA
AbstractThe “allogeneic effect” refers to the induction of host B cell antibody synthesis or host T cellcytotoxicity, including tumoricidal activity, by an infusion of allogeneic lymphocytes. We show thattreatment of mice with cyclophosphamide (Cy) followed by CD8+ T cell-depleted allogeneic donorlymphocyte infusion (Cy + CD8− DLI) induces regression of established tumors with minimaltoxicity in models of both hematologic and solid cancers, even though the donor cells are eventuallyrejected by the host immune system. The optimal anti-tumor effect of Cy + CD8− DLI required thepresence of donor CD4+ T cells, host CD8+ T cells, and alloantigen expression by normal host butnot tumor tissue. The results support a model in which a donor CD4+ T cell-mediated graft-versus-host reaction effectively awakens anti-tumor immunity among Cy-resistant host CD8+ T cells. Theseevents provide the cellular mechanism of the “allogeneic effect” in anti-tumor immunity. Cy +CD8− DLI may be an effective and minimally toxic strategy for awakening the host immune responseto advanced cancers.
IntroductionAllogeneic blood or marrow transplantation (alloBMT) is a well-established therapy forhematologic malignancies. The graft-versus-leukemia, or GVL, effect of alloBMT is perhapsthe best evidence that T cells can mediate the destruction of advanced, metastatic humanmalignancies via direct killing of tumor cells by donor T cells reactive to hosthistocompatibility antigens [1–8]. Despite the curative potential of allogeneic T cells, alloBMTas a procedure is severely hampered by three factors: (a) toxicity, (b) lack of efficacy, especiallyin solid tumors, and (c) limited availability of human leukocyte antigen (HLA)-identicaldonors. The conditioning regimen, infection, and GVHD all contribute to toxicity, and lack ofefficacy manifests itself as progression of the underlying malignancy after transplantation [9,10].
Corresponding Author: Ephraim Joseph Fuchs, M.D., 488 Blunting-Blaustein Cancer Research Building, 1650 Orleans St, Baltimore,MD 21231, E-mail:[email protected], Phone: 410-955-8143, Fax: 410-502-7163.Publisher's Disclaimer: This is a PDF file of an unedited manuscript that has been accepted for publication. As a service to our customerswe are providing this early version of the manuscript. The manuscript will undergo copyediting, typesetting, and review of the resultingproof before it is published in its final citable form. Please note that during the production process errors may be discovered which couldaffect the content, and all legal disclaimers that apply to the journal pertain.
NIH Public AccessAuthor ManuscriptBiol Blood Marrow Transplant. Author manuscript; available in PMC 2009 May 1.
Published in final edited form as:Biol Blood Marrow Transplant. 2008 May ; 14(5): 499–509.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Recently, there have been reports of disease responses in patients with hematologic [11–13]orsolid [14] malignancies despite rejection of the donor leukocytes. Earlier studies had shownthat anti-tumor effects can be mediated by donor lymphocyte infusions, or DLI, in patientsreceiving no or minimal conditioning. Interestingly, while alloBMT has not been conclusivelyshown to be effective in the treatment of solid tumors, allogeneic or even xenogeneiclymphocyte infusions have induced objective responses in as many as 20–30% of patients withadvanced solid malignancies [15]. These observations raise the question of whether transientlyengrafting lymphocytes mediate clinical responses via direct killing of tumor cells. Alexanderand colleagues were the first to speculate that cells of the host mediate the anti-tumor effect ofallogeneic lymphocytes [16]. Katz, Ellman, and colleagues subsequently found that allogeneicDLI prolonged the survival of guinea pigs subsequently challenged with a host strain leukemia,even when the challenge occurred after the allogeneic lymphocytes had been rejected by thehost immune system. This was dubbed the “allogeneic effect” of anti-tumor immunity [17,18], yet the underlying cellular mechanisms of this effect have been poorly defined.
Here, we have developed a mouse model to investigate the cellular interactions involved in theanti-tumor effect of transiently engrafting lymphocytes. These studies reveal a potentialcollaboration in which donor CD4+ T cells cooperate with, and effectively awaken, hostCD8+ T cells to induce tumor regression in both hematologic and solid malignancies.Therefore, infusion of allogeneic lymphocytes after minimal conditioning can mediate an anti-tumor effect against a wide spectrum of malignancies with reduced toxicity compared withstandard allogeneic bone marrow transplantation.
Materials and MethodsAnimals
C57BL/6 (B6; H-2b), BALB/c (H-2d, Thy1.2+/+), BALB/c × B6 (CB6) F1 (H-2d/b), and B6 ×C3H (B6C3) F1(H-2b/k) mice were all obtained from the National Cancer Institute (Frederick,MD). Mice were maintained in microisolator cages and were fed ad libitum with autoclavedlaboratory chow and acidified water. All mice were approximately 6 to 12 weeks of age at thetime of treatment. All manipulations were performed in a laminar flow hood. The Animal Careand Use Committee of the Johns Hopkins University approved all procedures on animals inaccordance with guidelines established by the Association for Assessment and Accreditationof Laboratory Animal Care (AAALAC). In specified GVHD experiments, mice were weighedsemiweekly beginning at the time of adoptive transfer of lymphocytes.
Tumor Cell LinesA20 is a spontaneous B cell leukemia/lymphoma of BALB/c origin [19]. Cells were obtainedoriginally from American Type Culture Collection (ATCC; Manassas, VA) and weremaintained in culture and administered intravenously (IV) at the lethal dose of 1 × 106 aspreviously described [20]. Cells were cultured in vitro in Eagle Hanks Amino Acid (EHAA)medium (Biofluids, Rockville, MD), 10% fetal calf serum (FCS; GIBCO BRL, Gaithersburg,MD), 5 × 10−5 M 2-mercaptoethanol (2-ME), glutamine, and antibiotics (complete medium;CM). RENCA, a murine carcinogen-induced renal cell carcinoma of BALB/c origin and B16,a murine melanoma cell line of C57Bl/6 origin, were used in specified experiments andmaintained in vitro in RPMI 1640 (Life Technologies, Grand Island, NY), supplemented with10% heat-inactivated FCS (HyClone, Logan, UT), 5 × 10−5 M 2-ME, 20 mmol/L HEPES, 30Ag/mL gentamicin (Schering Corporation, Kenilworth, NJ), and 0.2% sodium bicarbonate.
Cell preparationsDonor spleens were removed aseptically and pressed through a nylon mesh to obtain singlecell suspensions. In some experiments, T cells were depleted from donor splenocytes by
Symons et al. Page 2
Biol Blood Marrow Transplant. Author manuscript; available in PMC 2009 May 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

incubation with antibodies to CD4 (RL172.4; gift of Dr Albert Bendelac, U. Chicago, Chicago,Ill.) and/or CD8 (3.155; ATCC, Manassas, VA), and guinea pig complement (GIBCO BRL),as previously described [21]. Cell suspensions were vigorously pipetted, counted, and washedin sterile phosphate buffered saline (PBS) prior to injection. The purity of depleted cells wasgreater than 99.4% in all experiments.
Adoptive Transfer of T cellsCyclophosphamide (Cy; Bristol-Myers, Evansville, IN) was dissolved in PBS to aconcentration of 20 mg/mL and administered at a dose of 200 mg/kg intraperitoneally (IP)before or after tumor inoculation as specified in each experiment. Twenty-five millionsplenocytes, prepared as above, were injected IV into the lateral tail vein unless otherwisespecified.
In vivo depletion of T cellsMice were depleted of CD8+ T cells in vivo by giving 1.4 mg 2.43 (rat anti-mouse CD8monoclonal antibody) [22,23] IP on days 9 and 30 after tumor inoculation. The 2.43 antibodywas produced and purified from the supernatant of a bioreactor cell culture at the National CellCulture Center (Division of Biovest International, Worcester, MA) and was diluted in sterilePBS prior to injection. Control mice were injected with sterile PBS only.
Antibodies and flow cytometryAntibodies for flow cytometry were anti-CD4 (allophycocyanin, peridinin chlorophyll protein[PerCP], and phycoerythrin-Cy5 [Cyc]), Thy1.1 (FITC, PerCP and phycoerythrin [PE]),Thy1.2-allophycocyanin and –fluorescein isothiocyanate (FITC), H-2Kb FITC, CD-45 PerCP,CD8-PE and -FITC, and B220-FITC (eBioscience, San Diego, Ca). All antibodies werepurchased from BD Biosciences (Mountain View, CA) unless otherwise specified. Gatedevents were collected on a FACSCalibur (Becton Dickinson, San Jose, CA) and analyzed usingCellQuest software (Becton Dickinson).
Analysis of leukocyte chimerismAt designated times after DLI, blood was obtained from the lateral tail vein, or animals werekilled and suspensions of spleen and/or bone marrow were prepared. Erythrocytes from theperipheral blood were lysed by using ammonium chloride buffer before initiation of staining.For determination of lineage-specific chimerism in BALB/c chimeras, 1 million cells werestained with fluorescein (FITC)–conjugated antibody to H-2Kb or H-2Kd-FITC with or withoutphycoerythrin (PE)–conjugated antibodies to CD4 or CD8 (all from Becton Dickinson, SanJose, CA). In each experiment, samples of peripheral blood from at least 3 host strain mice notreceiving transplants were stained for donor H-2 or Thy antigens. The mean + (3 × SEM) ofthe percentage of FITC cells in hosts not receiving transplants was calculated (and was < 0.5%for every experiment), and any transplant recipient containing a greater percentage of FITCcells than this value was considered to have donor cell engraftment.
Lymphocyte quantificationTruCOUNT™ Tubes (BD Biosciences, San Jose, CA) were used for determining absolutecounts of leucocytes in blood. The tubes contain a known number of lyophilized 4.2-mmfluorescent beads. In specified experiments, peripheral blood was obtained from the lateral tailvein at specified times after adoptive lymphocyte transfer. Trucount™ was performedaccording to the manufacturer’s recommendations. Briefly, 20 µL containing CD45-PerCPantibody was distributed to each Trucount™ tube. Then 50 µl of peripheral blood from eachmouse was added, gently mixed, and incubated for 15 min at room temperature in the dark.The lysis and fixation were obtained by adding 450 µL of FACS Lysing solution™ (BD
Symons et al. Page 3
Biol Blood Marrow Transplant. Author manuscript; available in PMC 2009 May 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Biosciences). Samples were analyzed within 1 hour on a FACSCalibur (Becton Dickinson,San Jose, CA). Samples were mixed gently immediately prior to analysis. Absolute cell countwas calculated using the following formula:
where * can be found on the TruCOUNT Absolute Count Tube foil pouch label.
Statistical analysisAll survival data were analyzed by the nonparametric rank sum test of Wilcoxon. P < .05 wasconsidered statistically significant. In mice receiving both tumor and donor lymphocytes, themechanism of death (tumor versus GVHD) was verified by necropsy of representative animals(data not shown).
Medians and SEMs were calculated for lymphocyte and chimerism data and analyzed usingSigma Plot, v9.0 (Systat Software, San Jose, CA). All experiments were repeated at least once.
ResultsCyclophosphamide plus CD8+ T cell-depleted DLI is effective against early and established,metastatic hematologic or solid cancers
In order to model the anti-tumor effects of transiently engrafting allogeneic lymphocytes, micewere conditioned for transplantation with cyclophosphamide (Cy) only, a drug that isminimally toxic to host hematopoietic stem cells. In the first set of experiments, Cy wasadministered the day before tumor challenge, with or without concomitant lymphocytetransfusion, so that any anti-tumor effects observed could be unequivocally attributed to thelymphocytes and not the drug. In a tumor survival experiment employing BALB/c recipientsand fully MHC-incompatible B6 donors, administration of Cy followed by 50 million wholesplenocytes led to severe acute graft-versus-host disease (GVHD), marked by hunched posture,ruffled fur, weight loss, diarrhea, and ultimately death (Figure 1A). In contrast, animalsreceiving the same number of splenocytes depleted of CD8+ cells (CD8− DLI) had no clinicalevidence of GVHD and survived longer than animals receiving Cy plus splenocytes depletedof both CD4+ and CD8+ cells (median survival, 60 versus 30 days; p = .04). Recipients ofCD4+ cell-depleted splenocytes did not survive significantly longer than recipients of T cell-depleted splenocytes (median survival, 56 versus 30 days; p = .30). Therefore, donor CD4+ Tcells are required for the optimal anti-tumor effect of Cy + CD8-depleted DLI.
In a second tumor survival experiment employing BALB/c recipients and fully MHC-incompatible B6 donors (Figure 1B), animals that received CD8− DLI survived significantlylonger than mice receiving Cy alone (median survival, 73 versus 19.5 days; p < 0.0001).Additionally, recipients of CD8− DLI survived significantly longer than mice receivingCD4− CD8− DLI (median survival, 73 versus 22 days; p=0.0002). Furthermore, there was nodifference between mice receiving no DLI (Cy only) and CD4−CD8− DLI (median survival,19.5 versus 22 days; p= .17). This experiment demonstrates that CD4−CD8− DLI does notprovide any anti-tumor benefit over no DLI. Deaths in mice in Figures 1A and 1B that receivedA20 alone, or A20 with Cy conditioning +/−, CD8− DLI or CD4−CD8− DLI were secondaryto tumor, as confirmed by necropsy or visualization of protuberant abdomens and/or hindlegparalysis, whereas deaths in mice receiving whole spleen DLI were solely from GVHD.Recipients of CD4− DLI died either early from GVHD or later from tumor.
To investigate the therapeutic potential of Cy + CD8− DLI, disseminated tumor was establishedby intravenous (IV) injection fourteen days prior to any treatment. Compared to animals
Symons et al. Page 4
Biol Blood Marrow Transplant. Author manuscript; available in PMC 2009 May 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

receiving no treatment, A20- or RENCA (renal cell carcinoma)-bearing BALB/c mice treatedwith Cy alone survived a median of 34 (Figure 1C) or 6 days (Figure 1D) longer (p<.0001 andp = .01) respectively. Addition of CD8− DLI from B6 donors further prolonged survival by amedian of 10 (Figure 1C, p = .02) and 7 (Figure 1D, p = .001) days, respectively. These datademonstrate that transiently engrafting allogeneic lymphocytes achieve modest but significantprolongations of survival in mice with disseminated hematologic or solid malignancies.
CD8− DLI engrafts transiently and does not cause GVHD or leukopeniaThe two most common complications of transfusing viable allogeneic lymphocytes intounconditioned human recipients are severe acute GVHD and bone marrow aplasia, both ofwhich require the sustained engraftment of the donor cells [6,24]. To characterize the toxicitiesof Cy + DLI, tumor-free BALB/c mice were conditioned with Cy, transfused with 50 millionB6 splenocytes, and were monitored serially for survival, and leukocyte count in the peripheralblood. Additionally, A20-bearing BALB/c mice were conditioned with Cy, transfused with 50million B6 splenocytes, and were monitored for donor CD4+, CD8+, and B cell chimerism.Recipients of either whole or CD4+ T cell-depleted splenocytes in non-tumor bearing mice hadGVHD-associated mortality (Figure 2C), and leukopenia prior to death (Figure 2D). Incontrast, splenocytes depleted of CD8+ cells, alone or together with CD4+ cells, did not induceeither GVHD-associated mortality or sustained leukopenia in non-tumor bearing mice. In A20-bearing mice, recipients of either whole or CD4+ T cell-depleted splenocytes had sustainedengraftment of donor CD4+ (Figure 2A), CD8+(data not shown), and B cells (Figure 2B),whereas recipients of splenocytes depleted of CD8+ cells, alone or together with CD4+ cellsengrafted only transiently. The effect of donor T cell subset depletion on donor chimerism wasthe same in tumor-free versus tumor-bearing recipients of donor lymphocyte infusions (datanot shown). Therefore, in immunocompetent recipients, the combination of Cy + CD8−allogeneic DLI induces anti-tumor effects without DLI-associated toxicities.
A direct graft-versus-tumor reaction is not required for prolongation of survival by Cy +CD8− DLI
A variety of mechanisms have been to proposed to account for the anti-tumor effect oftransiently engrafting lymphocytes, including a direct graft-versus-tumor effect [25,26], or thestimulation of endogenous anti-tumor immunity [27–29]. In some instances, it has been shownthat a host-versus-graft reaction [30–32] can augment anti-tumor immunity. We wished tocharacterize the anti-tumor effect of a graft-versus-host (GVH) reaction in the absence of anyT cell-mediated host-versus-graft or graft-versus-tumor (GVT) effects. To do so, CB6 F1 micebearing the B16 melanoma, of B6 origin, were treated with Cy followed by nothing or DLIfrom syngeneic CB6 F1 or parental strain B6, or BALB/c donors. Whereas BALB/c donorswould induce both GVH reactions and GVT effects, and CB6 F1 donors would provide neither,the B6 donors are syngeneic to the tumor and would induce only a GVH reaction. Recipientsof DLI from either parental strain prolonged survival compared to recipients of either no DLI(Figure 3; p =.04 and p=.06 for B6 and BALB/c donors, respectively) or DLI that was syngeneicto the recipient (p=.04 and .03 for B6 and BALB/c donors, respectively). The results with B6DLI demonstrate that a GVH reaction is sufficient to prolong survival in the absence of a directGVT effect. The failure of CB6 F1 DLI to prolong survival demonstrates that host expressionof alloantigens is required for the induction of anti-tumor immunity.
Host CD8+ T cells participate in the anti-tumor effect of Cy + CD8− DLICD8+ T cells have been shown to be capable of mediating anti-tumor immunity in both mousemodels [33,34] and in human clinical trials [35] Since CD4+ T cells deliver help to CD8+ Tcells by “licensing” antigen-presenting cells [36–38], we postulated that a donor CD4+ T cell-mediated GVH reaction effectively licenses host APCs to provide help to host-derived, anti-
Symons et al. Page 5
Biol Blood Marrow Transplant. Author manuscript; available in PMC 2009 May 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

tumor CD8+ T cells. To characterize the role of host CD8+ T cells in the anti-tumor effect ofCy + CD8− DLI, the A20 therapy experiment of Figure 1c was repeated but with the additionof separate groups of mice treated with the in vivo CD8-depleting monoclonal antibody 2.43on days 9 and 30 after tumor inoculation. Table I shows the effect of antibody treatment onperipheral blood lymphocyte populations in recipients of Cy + CD8− DLI, and Figure 4 showsthe effect of antibody treatment on the anti-tumor effect of Cy + CD8− DLI. Several conclusionsemerge from these data. First, depletion of host CD8+T cells was associated with a higher peakand longer duration of donor CD4+ T cell chimerism (Table I), implicating a role for hostCD8+ T cells in determining the kinetics of donor graft rejection. Second, the survival ofanimals treated with Cy alone was longer in antibody-untreated than in CD8-depleted mice(Figure 4A; median survival, 99 versus 51 days, p =.005). This result suggests that hostCD8+ T cells contribute to the anti-tumor effect of Cy. Third, CD8-depleted recipients of Cy+ CD8− DLI survived longer than CD8-depleted recipients of Cy alone (Figure 4A; mediansurvival >130 versus 51 days, p=.005), suggesting that allogeneic donor CD4+ T cells mediatean anti-tumor effect that does not require host CD8+ T cells. Fourth, Cy + CD8− DLI wasmarginally more effective in antibody-untreated than in antibody-treated recipients (Figure4A; median survival >130 days for both groups; p = .08). This result raises the possibility thathost CD8+ T cells participate in the anti-tumor effect of Cy + CD8- DLI and demonstrates thatprolonged survival of donor cells does not guarantee a more potent anti-tumor effect.
In the A20 model system described above, a clear role of host CD8+ T cells in the anti-tumoreffects of CD8− DLI was difficult to discern because donor CD4+ T cells could kill tumor cellsdirectly and because the 2.43 antibody prolonged donor cell survival in addition to depletinghost CD8+ T cells. To eliminate the possibility of a direct GVT effect of DLI, we examinedthe effect of host CD8+ cell depletion in B16-bearing CB6 F1 recipients of Cy +/− B6 CD8−DLI, which is syngeneic to the tumor (Figure 4B). When host CD8+ T cells were not depleted,recipients of Cy + CD8− DLI survived significantly longer than recipients of Cy alone (Figure4B; median survival 56.5 versus 70 days, p=.02). In contrast, when host CD8+ T cells weredepleted, the recipients of Cy + CD8− DLI did not survive significantly longer than recipientsof Cy alone (Figure 4B; median survival 52.5 versus 53.5 days, p=.41). This result demonstratesthat host CD8+ cells are required for the anti-tumor effect of CD8− DLI in the treatment ofmetastatic B16 melanoma.
Additionally, because the anti-tumor effect of B6 CD8− DLI is abrogated completely bydepleting host CD8+ T cells (Figure 4C), it rules out any direct GVT effects from the donorB6 cells, otherwise this effect would have been seen even in the absence of host CD8+ T cells.
DiscussionThe results presented here demonstrate that transiently engrafting allogeneic lymphocytes canmediate the regression of established solid or hematologic malignancies with minimalassociated toxicity. Treatment of tumor-bearing animals with Cy was required to unmask theanti-tumor activity of CD8-depleted DLI. This anti-tumor activity involves at least two distinctmechanisms: (a) a direct GVT effect that requires CD4+ T cells in the DLI and alloantigenexpression by the tumor itself, and (b) an indirect anti-tumor effect mediated by host CD8+ Tcells and requiring a GVH reaction against non-malignant host tissue.
Although it is possible that the relative contribution of CD4 and CD8 cells in GVHD and GVTis model specific, we have found that the combination of Cy followed by CD8− DLI is effectiveat inducing tumor regression without producing GVHD in two fully MHC-mismatched andone MHC-haploidentical strain combination (data not shown). We do find, however, that whenthe recipients’ tumor is MHC Class II+, there are both direct and indirect anti-tumor effects ofdonor CD4+ T cells, whereas the anti-tumor effect against MHC Class II− tumors is indirect
Symons et al. Page 6
Biol Blood Marrow Transplant. Author manuscript; available in PMC 2009 May 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

only, i.e. mediated by host CD8+ T cells. We specifically chose to study a variety of tumormodels including both hematologic and solid tumors of BALB/c background (A20 andRENCA, respectively), as well as a solid tumor of C57BL/6 background (i.e. B16) to makeour conclusions more generalizable and applicable to a variety of cancer types.
Survival is not always a proper measure of a GVT effect. However, we performed paralleltransfers of allogeneic cells into tumor-free mice, and found that the only groups with mortalitywere those that received DLI containing CD8+ T cells. These deaths were GVHD related, asmice exhibited ruffled fur, hunched posture, diarrhea, and weight loss (data not shown). Tumor-free mice that received Cy with or without CD8− DLI appeared healthy and survived long-term. From this, we conclude that CD8+ T cells are required for GVHD-associated mortality,and that deaths among tumor-bearing mice not receiving CD8+ T cells were attributable tocomplications of progressive tumor.
Previous studies have demonstrated an anti-tumor effect of allogeneic or even xenogeneiclymphocyte infusions given to unconditioned or minimally conditioned humans [13,39–48] orexperimental animals [49–53]. A variety of mechanisms have been invoked to explain tumorregression induced by transiently engrafting DLI, including transient graft-versus-tumorresponses mediated by donor T cells or NK cells [12,54–57], conditioning effects on hostimmunity [58,59], or even abrogation of host tolerance by cytokines liberated during therejection reaction [60,61]. Moreover, immunosuppressive conditioning has been shown toaugment the capacity of adoptively transferred allogeneic cells to induce GVHD [62]or GVTeffects [63], and recent studies have shown that lymphopenia-induced proliferation per se issufficient to augment anti-tumor immunity by adoptively transferred T cells [64,65]. We arecurrently investigating why Cy pre-treatment is required to unmask the anti-tumor activity ofCD8− DLI in animals with established tumors. Preliminary experiments demonstrate that Cymitigates the inhibitory influence of tumor-induced, Foxp3+ regulatory T cells (H.J.S. andE.J.F., unpublished data), but other explanations remain possible, such as the enhancement oftumor-and allo-antigen presentation resulting from the Cy-induced apoptosis of tumor and hostcells. Any or all of these mechanisms may be contributing to the anti-tumor effect of transientlyengrafting DLI in our models. Indeed, results in the A20 model demonstrate that at least someof the anti-tumor effect of CD8-depleted DLI is independent of host CD8+ T cells (Figure 4),and thus may be produced directly by donor lymphocytes. However, when the donorlymphocytes were syngeneic to the tumor, an anti-tumor effect of allogeneic lymphocyteinfusion that is dependent upon the presence of host CD8+ T cells became evident, suggestingthat donor CD4+ T cells and host CD8+ T cells cooperate to induce tumor regression. Thisimplies that the reason that CD8-depleted recipients of Cy + CD8− DLI survived longer thanCD8-depleted recipients of Cy alone in the A20 model (Figure 4A), but not in the B16melanoma model (Figures 4B–C), may be explained by direct tumor recognition of MHC classII on A20 tumor cells, an event that would not occur with B16 because it does not expressMHC Class II molecules. Taken together, the results of Figure 4 suggest that CD8− DLImediates anti-tumor effects through two distinct mechanisms- a direct GVT effect that requiresdirect tumor expression of alloantigens, and an indirect anti-tumor effect that is mediated byhost CD8+ T cells and does not require tumor expression of alloantigens. Therefore, CD8+ Tcells of host origin may play a critical role in producing tumor regression in the context of agraft-versus-host reaction, as described here, or in a host-versus-graft reaction, as describedpreviously by Sykes and colleagues [32].
The idea that a graft-versus-host reaction could awaken a dormant anti-tumor response fromthe host was first proposed by Alexander and colleagues [66]. The term “allogeneic effect”was originally coined to describe how a graft-versus-host reaction could substitute for cognateT cell help in the secondary antibody response to a hapten [67], but was extended to describehow an allogeneic lymphocyte infusion could augment host resistance to a subsequent tumor
Symons et al. Page 7
Biol Blood Marrow Transplant. Author manuscript; available in PMC 2009 May 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

challenge, even at a time when the donor cells had been rejected. Due to limitations in existingtechnology, these early studies could not define the precise cellular interactions involved inthe anti-tumor effect of transiently engrafting lymphocytes [52,68,69]. The current studydescribes a novel cooperation between donor CD4+ and host CD8+ T cells in mediating ananti-tumor effect against a subsequent tumor challenge as well as against an established burdenof either a solid or hematologic malignancy. Since CD4+ T cells provide “help” to CD8+
cytotoxic T cells by activating APCs [70–72], it seems reasonable to propose that alloreactivedonor CD4+ T cells recognize and activate host APCs, which in turn can activate tumor-specifichost CD8+ T cells even after the donor CD4+ T cells have been rejected. This model is consistentwith the finding that alloantigen expression by non-malignant host tissue is sufficient for ananti-tumor effect of parental DLI given to an F1 host (Figure 3A).
Allogeneic BMT has an established role in the treatment of a variety of hematologicmalignancies through the induction of a graft-versus-tumor effect. While there has been nodearth of attempts to apply alloBMT to the treatment of advanced solid tumors, results havebeen mixed at best. A putative GVT effect of alloBMT has been demonstrated against a varietyof solid tumors [73] including renal cell carcinoma [74], breast cancer [75,76], colon cancer[77], and ovarian carcinoma [78], but the anti-tumor effect usually occurs in the context ofGVHD, and no conclusive benefits in overall or event-free survival have been reported [79].In contrast, Cy + CD8− DLI induced anti-tumor effects against systemically disseminated solidtumors with minimal toxicity. While encouraging, these results need to be confirmed in moreclinically relevant models, such as the treatment of endogenous rather than transplanted tumors.We postulate that distinct populations of T cells mediate anti-tumor effects after alloBMTversus Cy + CD8− DLI. The anti-tumor effectors after myeloablative alloBMT are thought tobe donor T cells reactive to host minor histocompatibility antigens. While these T cells cancure hematologic malignancies through a lymphohematopoietic GVH reaction that destroysboth normal and malignant blood cells, there is no a priori reason for them to be able todistinguish between normal and malignant solid tissue. In contrast, at least some of the anti-tumor effectors of Cy + CD8− DLI are host CD8+ T cells, which are presumably tolerant tonormal peripheral self tissues. It is possible that the systemic activation of host APCs byalloreactive donor CD4+ T cells could also unmask the activity of autoreactive T and/or B cellclones. While we have not rigorously evaluated the mice for autoimmune phenomena, nonewas clinically evident (data not shown).
The results described herein demonstrate that chemotherapy, with or without adoptive cellularimmunotherapy, works best in the presence of an intact host CD8+ T cell compartment. Indeed,the participation of host CD8+ T cells in the anti-tumor response to chemotherapy (Figure 4A)may explain why the pre-treatment absolute lymphocyte count correlates with the response tochemotherapy of lymphoma [80] and solid tumors [81]. By the same token, it is also possiblethat chemotherapy-induced lymphopenia underlies the inexorable decline in responsiveness ofmetastatic solid tumors to cytotoxic drugs. If, as our results suggest, the optimal response toimmunotherapy requires an intact host immune system, then the sequence of tryingimmunotherapy after the failure of standard chemotherapy would appear to becounterproductive. Likewise, the strategy of intensive, non-selective lymphodepletion prior toadoptive immunotherapy [82–84] makes sense only when donor cells are the sole mediatorsof anti-tumor immunity.
Finally, our results support a strategy of awakening dormant anti-tumor immunity againstadvanced cancer by providing exogenous CD4+ T cell help for endogenous tumor-specificCD8+ T cells [85]. The capacity of a graft-versus-host reaction to provide this help suggeststhat allogeneic lymphocytes are a readily available tool for breaking functional tolerance toadvanced cancers in the clinic.
Symons et al. Page 8
Biol Blood Marrow Transplant. Author manuscript; available in PMC 2009 May 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Acknowledgments
We thank Dr. Drew Pardoll for his assistance in reviewing and editing this manuscript. This work was supported bythe National Institutes of Health grants PO1 CA-15396 (Richard J. Jones, M.D., PI) and R01 CA105148 (EJF). EJFis a Leukemia and Lymphoma Society Scholar in Clinical Research
REFERENCES1. Weiden PL, Flournoy N, Thomas ED, et al. Antileukemic effect of graft-versus-host disease in human
recipients of allogeneic-marrow grafts. N Engl J Med 1979;300:1068–1073. [PubMed: 34792]2. Weiden PL, Flournoy N, Sanders JE, Sullivan KM, Thomas ED. Antileukemic effect of graft-versus-
host disease contributes to improved survival after allogeneic marrow transplantation. Transplant Proc1981;13(Pt 1):248–251. [PubMed: 7022833]
3. Maraninchi D, Gluckman E, Blaise D, et al. Impact of T-cell depletion on outcome of allogeneic bone-marrow transplantation for standard-risk leukaemias. Lancet 1987;2:175–178. [PubMed: 2885638]
4. Horowitz MM, Gale RP, Sondel PM, et al. Graft-versus-leukemia reactions after bone marrowtransplantation. Blood 1990;75:555–562. [PubMed: 2297567]
5. Kolb HJ. Graft-versus-leukemia effects of donor lymphocyte transfusions in marrow grafted patients.Blood 1995;86:2041–2050. [PubMed: 7655033]
6. Collins RH Jr, Shpilberg O, Drobyski WR, et al. Donor leukocyte infusions in 140 patients with relapsedmalignancy after allogeneic bone marrow transplantation [see comments]. J Clin Oncol 1997;15:433–444. [PubMed: 9053463]
7. Tsukada N, Kobata T, Aizawa Y, Yagita H, Okumura K. Graft-versus-leukemia effect and graft-versus-host disease can be differentiated by cytotoxic mechanisms in a murine model of allogeneic bonemarrow transplantation. Blood 1999;93:2738–2747. [PubMed: 10194454]
8. Schmaltz C, Alpdogan O, Horndasch KJ, et al. Differential use of Fas ligand and perforin cytotoxicpathways by donor T cells in graft-versus-host disease and graft-versus-leukemia effect. Blood2001;97:2886–2895. [PubMed: 11313285]
9. Szydlo R, Goldman JM, Klein JP, et al. Results of allogeneic bone marrow transplants for leukemiausing donors other than HLA-identical siblings. J Clin Oncol 1997;15:1767–1777. [PubMed: 9164184]
10. Drobyski WR, Klein J, Flomenberg N, et al. Superior survival associated with transplantation ofmatched unrelated versus one-antigen-mismatched unrelated or highly human leukocyte antigen-disparate haploidentical family donor marrow grafts for the treatment of hematologic malignancies:establishing a treatment algorithm for recipients of alternative donor grafts. Blood 2002;99:806–814.[PubMed: 11806980]
11. O'Donnell PV, Luznik L, Jones RJ, et al. Nonmyeloablative bone marrow transplantation frompartially HLA-mismatched related donors using posttransplantation cyclophosphamide. Biol BloodMarrow Transplant 2002;8:377–386. [PubMed: 12171484]
12. Dey BR, McAfee S, Colby C, et al. Anti-tumour response despite loss of donor chimaerism in patientstreated with non-myeloablative conditioning and allogeneic stem cell transplantation. British Journalof Haematology 2005;128:351–359. [PubMed: 15667537]
13. Colvin GA, Lambert J-F, Lum LG, et al. Effective universal outpatient immunotherapeutic approachfor refractory acute myelocytic leukemia: HLA-haploidentical transplants in 100 cGy-conditionedhosts [abstract]. Blood 2002;100:363a. [PubMed: 12096715]
14. Strair RK, Schaar D, Medina D, et al. Antineoplastic Effects of Partially HLA-Matched IrradiatedBlood Mononuclear Cells in Patients With Renal Cell Carcinoma. J Clin Onc 2003;21:3785–3791.
15. Kondo M, McCarty MF. Rationale for a novel immunotherapy of cancer with allogeneic lymphocyteinfusion. Med Hypotheses 1984;15:241–277. [PubMed: 6394972]
16. Alexander P, Delorme EJ, Hall JG. The effect of lymphoid cells from the lymph of specificallyimmunized sheep on the growth of primary sarcomata in rats. Lancet 1966 May 28;:1186–1189.
17. Ellman L, Katz DH, Green I, Paul WE, Benacerraf B. Mechanisms involved in the antileukemic effectof immunocompetent allogeneic lymphoid cell transfer. Cancer Res 1972;32:141–148. [PubMed:4400015]
Symons et al. Page 9
Biol Blood Marrow Transplant. Author manuscript; available in PMC 2009 May 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

18. Katz DH, Ellman L, Paul WE, Green I, Benacerraf B. Resistance of guinea pigs to leukemia followingtransfer of immunocompetent allogeneic lymphoid cells. Cancer Res 1972;32:133–140. [PubMed:4400014]
19. Kim KJ, Kanellopoulos-Langevin C, Merwin RM, Sachs DH, Asofsky R. Establishment andcharacterization of BALB/c lymphoma lines with B cell properties. J Immunol 1979;122:549–554.[PubMed: 310843]
20. Levitsky HI, Montgomery J, Ahmadzadeh M, et al. Immunization with granulocyte-macrophagecolony-stimulating factor-transduced, but not B7-1-transduced, lymphoma cells primes idiotype-specific T cells and generates potent systemic antitumor immunity. J Immunol 1996;156:3858–3865.[PubMed: 8621924]
21. Fuchs EJ, Matzinger P. B cells turn off virgin but not memory T cells. Science 1992;258:1156–1159.[PubMed: 1439825]
22. Zhou G, Lu Z, McCadden JD, Levitsky HI, Marson AL. Reciprocal Changes in Tumor Antigenicityand Antigen-specific T Cell Function during Tumor Progression. J Exp Med 2004;200:1581–1592.[PubMed: 15596524]
23. Zhao Y, Swenson K, Wekerle T, Rodriguez-Barbosa JI, Arn JS, Sykes M. The critical role of mouseCD4+ cells in the rejection of highly disparate xenogeneic pig thymus grafts. Xenotransplantation2000;7:129–137. [PubMed: 10961297]
24. Schroeder ML. Transfusion associated graft-versus-host-disease. Brit J Haematol 2002:275–287.[PubMed: 11972509]
25. Boranic M, Tonkovic I. Time pattern of the antileukemic effect of graft-versus-host reaction in mice.Cancer Res 1971;31:1140–1147. [PubMed: 4398523]
26. Boranic M. Transient graft-versus-host reaction in the treatment of leukemia in mice. J Natl CancerInst 1968;41:421–437. [PubMed: 4876445]
27. Alexander P, Delorme EJ, Hall JG. The effect of lymphoid cells from the lymph of specificallyimmunized sheep on the growth of primary sarcomata in rats. Lancet 1966 May 28;:1186–1189.
28. Ellman L, Katz DH, Green I, Paul WE, Benacerraf B. Mechanisms involved in the antileukemic effectof immunocompetent allogeneic lymphoid cell transfer. Cancer Res 1972;32:141–148. [PubMed:4400015]
29. Katz DH, Ellman L, Paul WE, Green I, Benacerraf B. Resistance of guinea pigs to leukemia followingtransfer of immunocompetent allogeneic lymphoid cells. Cancer Res 1972;32:133–140. [PubMed:4400014]
30. Rubio MT, Kim YM, Sachs T, Mapara M, Zhao G, Sykes M. Antitumor effect of donor marrow graftrejection induced by recipient leukocyte infusions in mixed chimeras prepared withnonmyeloablative conditioning: critical role for recipient-derived IFN-{gamma}. Blood2003;102:2300–2307. [PubMed: 12791660]
31. Zhou G, Drake CG, Levitsky HI. Amplification of tumor-specific regulatory T cells followingtherapeutic cancer vaccines. Blood 2006;107:628–636. [PubMed: 16179369]
32. Rubio MT, Saito TI, Kattleman K, Zhao G, Buchli J, Sykes M. Mechanisms of the AntitumorResponses and Host-versus-Graft Reactions Induced by Recipient Leukocyte Infusions in MixedChimeras Prepared with Nonmyeloablative Conditioning: A Critical Role for Recipient CD4+ TCells and Recipient Leukocyte Infusion-Derived IFN-{gamma}-Producing CD8+ T Cells. J Immunol2005;175:665–676. [PubMed: 16002661]
33. Hanson HL, Donermeyer DL, Ikeda H, et al. Eradication of established tumors by CD8+ T celladoptive immunotherapy. Immunity 2000;13:265–276. [PubMed: 10981969]
34. Overwijk WW, Theoret MR, Finkelstein SE, et al. Tumor regression and autoimmunity after reversalof a functionally tolerant state of self-reactive CD8+ T cells. J Exp Med 2003;198:569–580. [PubMed:12925674]
35. Dudley ME, Wunderlich JR, Robbins PF, et al. Cancer Regression and Autoimmunity in PatientsAfter Clonal Repopulation with Antitumor Lymphocytes. Science. 2002
36. Ridge JP, Di Rosa F, Matzinger P. A conditioned dendritic cell can be a temporal bridge between aCD4+ T- helper and a T-killer cell. Nature 1998;393:474–478. [PubMed: 9624003]
Symons et al. Page 10
Biol Blood Marrow Transplant. Author manuscript; available in PMC 2009 May 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

37. Schoenberger SP, Toes RE, van der Voort EI, Offringa R, Melief CJ. T-cell help for cytotoxic Tlymphocytes is mediated by CD40-CD40L interactions. Nature 1998;393:480–483. [PubMed:9624005]
38. Bennett SR, Carbone FR, Karamalis F, Flavell RA, Miller JF, Heath WR. Help for cytotoxic-T-cellresponses is mediated by CD40 signalling. Nature 1998;393:478–480. [PubMed: 9624004]
39. Strair RK, Schaar D, Medina D, et al. Antineoplastic Effects of Partially HLA-Matched IrradiatedBlood Mononuclear Cells in Patients With Renal Cell Carcinoma. J Clin Oncology 2003;21:3785–3791.
40. Kondo M, McCarty MF. Rationale for a novel immunotherapy of cancer with allogeneic lymphocyteinfusion. Med Hypotheses 1984;15:241–277. [PubMed: 6394972]
41. Nadler SH, Moore GE. Immunotherapy of malignant disease. Arch Surg 1969;99:376–381. [PubMed:5798432]
42. Sumner WC, Foraker AG. Spontaneous regression of human melanoma: clinical and experimentalstudies. Cancer 1960;13:79–81. [PubMed: 13835693]
43. Teimourian B, McCune WS. Surgical management of malignant melanoma. Am Surg 1963;29:515–519. [PubMed: 14046056]
44. Woodruff MF, Nolan B. Preliminary obervations on treatment of advanced cancer by injection ofallogeneic spleen cells. Lancet 1963;13:426–429. [PubMed: 14044314]
45. Schwarzenberg L, Mathe G, Schneider M, Amiel JL, Cattan A, Schlumberger JR. Attempted adoptiveimmunotherapy of acute leukaemia by leucocyte transfusions. Lancet 1966;2:365–368. [PubMed:4161720]
46. Porter DL, Connors JM, Van Deerlin VM, et al. Graft-versus-tumor induction with donor leukocyteinfusions as primary therapy for patients with malignancies. J Clin Oncol 1999;17:1234. [PubMed:10561184]
47. Ballen KK, Becker PS, Emmons RVB, et al. Low-dose total body irradiation followed by allogeneiclymphocyte infusion may induce remission in patients with refractory hematologic malignancy.Blood 2002;100:442. [PubMed: 12091334]
48. Yonemoto RH. Adoptive immunotherapy utilizing thoracic duct lymphocytes. Ann N Y Acad Sci1976;277:7–19. [PubMed: 11733]
49. Alexander P, Delorme EJ, Hall JG. The effect of lymphoid cells from the lymph of specificallyimmunized sheep on the growth of primary sarcomata in rats. Lancet 1966 May 28;:1186–1189.
50. Katz DH, Ellman L, Paul WE, Green I, Benacerraf B. Resistance of guinea pigs to leukemia followingtransfer of immunocompetent allogeneic lymphoid cells. Cancer Res 1972;32:133–140. [PubMed:4400014]
51. Boranic M. Transient graft-versus-host reaction in the treatment of leukemia in mice. J Natl CancerInst 1968;41:421–437. [PubMed: 4876445]
52. Medzihradsky J. Modification of tumor homograft immunity during the graft-versus-host reaction.Neoplasma 1966;13:223–226. [PubMed: 5911878]
53. Fefer A. Treatment of a Moloney lymphoma with cyclophosphamide and H-2-incompatible spleencells. Cancer Res 1973;33:641–644. [PubMed: 4696464]
54. Boranic M. Transient graft-versus-host reaction in the treatment of leukemia in mice. J Natl CancerInst 1968;41:421–437. [PubMed: 4876445]
55. Fefer A. Treatment of a Moloney lymphoma with cyclophosphamide and H-2-incompatible spleencells. Cancer Res 1973;33:641–644. [PubMed: 4696464]
56. Ruggeri L, Capanni M, Casucci M, et al. Role of natural killer cell alloreactivity in HLA-mismatchedhematopoietic stem cell transplantation. Blood 1999;94:333–339. [PubMed: 10381530]
57. Dey BR, McAfee SL, Sackstein R, et al. Achievement of sustained remissions despite loss of donorchimerism (DC) patients with chemotherapy-refractory non-Hodgkin's lymphoma (NHL) treatedwith nonmyeloablative conditioning and allogeneic stem cell transplantation (SCT). [abstract]. Blood2001;98:1753.
58. Mokyr MB, Hengst JC, Dray S. Role of antitumor immunity in cyclophosphamide-induced rejectionof subcutaneous nonpalpable MOPC-315 tumors. Cancer Res 1982;42:974–979. [PubMed: 6977410]
Symons et al. Page 11
Biol Blood Marrow Transplant. Author manuscript; available in PMC 2009 May 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

59. North RJ. Gamma-irradiation facilitates the expression of adoptive immunity against establishedtumors by eliminating suppressor T cells. Cancer Immunol Immunother 1984;16:175–181. [PubMed:6231095]
60. Rubio MT, Kim YM, Sachs T, Mapara M, Zhao G, Sykes M. Antitumor effect of donor marrow graftrejection induced by recipient leukocyte infusions in mixed chimeras prepared withnonmyeloablative conditioning: critical role for recipient-derived IFN-{gamma}. Blood2003;102:2300–2307. [PubMed: 12791660]
61. Rubio MT, Saito TI, Kattleman K, Zhao G, Buchli J, Sykes M. Mechanisms of the AntitumorResponses and Host-versus-Graft Reactions Induced by Recipient Leukocyte Infusions in MixedChimeras Prepared with Nonmyeloablative Conditioning: A Critical Role for Recipient CD4+ TCells and Recipient Leukocyte Infusion-Derived IFN-{gamma}-Producing CD8+ T Cells. J Immunol2005;175:665–676. [PubMed: 16002661]
62. Lehnert S, Rybka WB. Amplification of the graft-versus-host reaction by cyclophosphamide:dependence on timing of drug administration. Bone Marrow Transplant 1994;13:473–477. [PubMed:8019473]
63. Fefer A. Treatment of a Moloney lymphoma with cyclophosphamide and H-2-incompatible spleencells. Cancer Res 1973;33:641–644. [PubMed: 4696464]
64. Dummer W, Niethammer AG, Baccala R, et al. T cell homeostatic proliferation elicits effectiveantitumor autoimmunity. J Clin Invest 2002;110:185–192. [PubMed: 12122110]
65. Brown IE, Blank C, Kline J, Kacha AK, Gajewski TF. Homeostatic Proliferation as an IsolatedVariable Reverses CD8+ T Cell Anergy and Promotes Tumor Rejection. J Immunol 2006;177:4521–4529. [PubMed: 16982889]
66. Alexander P, Delorme EJ, Hall JG. The effect of lymphoid cells from the lymph of specificallyimmunized sheep on the growth of primary sarcomata in rats. Lancet 1966 May 28;:1186–1189.
67. Katz DH, Davie JM, Paul WE, Benacerraf B. Carrier function in anti-hapten antibody responses. IV.Experimental conditions for the induction of hapten-specific tolerance or for the stimulation of anti-hapten anamnestic responses by "nonimmunogenic" hapten-polypeptide conjugates. J Exp Med1971;134:201–223. [PubMed: 4397662]
68. Ellman L, Katz DH, Green I, Paul WE, Benacerraf B. Mechanisms involved in the antileukemic effectof immunocompetent allogeneic lymphoid cell transfer. Cancer Res 1972;32:141–148. [PubMed:4400015]
69. Katz DH, Ellman L, Paul WE, Green I, Benacerraf B. Resistance of guinea pigs to leukemia followingtransfer of immunocompetent allogeneic lymphoid cells. Cancer Res 1972;32:133–140. [PubMed:4400014]
70. Bennett SR, Carbone FR, Karamalis F, Flavell RA, Miller JF, Heath WR. Help for cytotoxic-T-cellresponses is mediated by CD40 signalling. Nature 1998;393:478–480. [PubMed: 9624004]
71. Ridge JP, Di Rosa F, Matzinger P. A conditioned dendritic cell can be a temporal bridge between aCD4+ T- helper and a T-killer cell. Nature 1998;393:474–478. [PubMed: 9624003]
72. Schoenberger SP, Toes RE, van der Voort EI, Offringa R, Melief CJ. T-cell help for cytotoxic Tlymphocytes is mediated by CD40-CD40L interactions. Nature 1998;393:480–483. [PubMed:9624005]
73. Blaise D, Bay JO, Faucher C, et al. Reduced-intensity preparative regimen and allogeneic stem celltransplantation for advanced solid tumors. Blood 2004;103:435–441. [PubMed: 12946991]
74. Childs R, Chernoff A, Contentin N, et al. Regression of metastatic renal-cell carcinoma afternonmyeloablative allogeneic peripheral-blood stem-cell transplantation. N Engl J Med2000;343:750–758. [PubMed: 10984562]
75. Carella AM, Beltrami G, Corsetti MT, et al. Reduced intensity conditioning for allograft aftercytoreductive autograft in metastatic breast cancer. Lancet 366:318–320. [PubMed: 16039336]
76. Ueno NT, Cheng YC, Rondon G, et al. Rapid induction of complete donor chimerism by the use ofa reduced-intensity conditioning regimen composed of fludarabine and melphalan in allogeneic stemcell transplantation for metastatic solid tumors. Blood 2003;102:3829–3836. [PubMed: 12881308]
77. Carnevale-Schianca F, Cignetti A, Capaldi A, et al. Allogeneic nonmyeloablative hematopoietic celltransplantation in metastatic colon cancer: tumor-specific T cells directed to a tumor-associatedantigen are generated in vivo during GVHD. Blood 2006;107:3795–3803. [PubMed: 16403911]
Symons et al. Page 12
Biol Blood Marrow Transplant. Author manuscript; available in PMC 2009 May 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

78. Silverman LR, Demakos EP, Peterson BL, et al. Randomized controlled trial of azacitidine in patientswith the myelodysplastic syndrome: a study of the cancer and leukemia group B. J Clin Oncol2002;20:2429–2440. [PubMed: 12011120]
79. Bregni M, Ueno NT, Childs R. The Second International Meeting on Allogeneic Transplantation inSolid Tumors. Bone Marrow Transplant 2006;38:527–537. [PubMed: 16953213]
80. Siddiqui M, Ristow K, Markovic SN, et al. Absolute lymphocyte count predicts overall survival infollicular lymphomas. British Journal of Haematology 2006;134:596–601. [PubMed: 16889618]
81. Lissoni P, Brivio F, Fumagalli L, et al. Efficacy of cancer chemotherapy in relation to the pretreatmentnumber of lymphocytes in patients with metastatic solid tumors. Int J Biol Markers 2004;19:135–140. [PubMed: 15255546]
82. Dudley ME, Wunderlich JR, Robbins PF, et al. Cancer Regression and Autoimmunity in PatientsAfter Clonal Repopulation with Antitumor Lymphocytes. Science. 2002
83. Gattinoni L, Finkelstein SE, Klebanoff CA, et al. Removal of homeostatic cytokine sinks bylymphodepletion enhances the efficacy of adoptively transferred tumor-specific CD8+ T cells. J ExpMed 2005;202:907–912. [PubMed: 16203864]
84. Wrzesinski C, Restifo NP. Less is more: Lymphodepletion followed by hematopoietic stem celltransplant augments adoptive T-cell-based anti-tumor immunotherapy. Curr Opin Immunol2005;17:195–201. [PubMed: 15766681]
85. Antony PA, Piccirillo CA, Akpinarli A, et al. CD8+ T Cell Immunity Against a Tumor/Self-AntigenIs Augmented by CD4+ T Helper Cells and Hindered by Naturally Occurring T Regulatory Cells. JImmunol 2005;174:2591–2601. [PubMed: 15728465]
Symons et al. Page 13
Biol Blood Marrow Transplant. Author manuscript; available in PMC 2009 May 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Figure 1. Cy + Non-engrafting CD8-depleted DLI abrogates risk of GVHD, induces anti-tumorimmunity, and prolongs survival of animals with established, metastatic hematologic and solidtumor malignancies(a,b) CD8 depletion of DLI abrogates risk of GVHD without compromising anti-tumorimmunity. BALB/c mice (H-2d; n=10/group) either received 106 A20 lymphoma cells IV onDay 0 alone (△) or were conditioned with Cy 200 mg/kg IP on day −1 and received 106 A20lymphoma cells IV on Day 0 (○). Additional mice were then treated with 5 × 107 whole spleencells from fully MHC-mismatched C57BL/6 (H-2b) donors, either undepleted (■) or depletedof CD4+ T cells (●), CD8+ T cells (▲), or both (□). Results for 1a: (▲) v (□) p =.04, (●) v (□)p=.03. Results for 1b: (▲) v (○) v (○) p<.0001, (▲) v (□) v p=.0002, (○) v ((□) p=.17. (c) Cy+ CD8-depleted allogeneic DLI prolongs survival of animals with established, metastaticlymphoma. BALB/c mice (H-2d; n=10/group) received 106 A20 cells IV on Day 0 either alone(■), or followed by treatment with Cy 200 mg/kg IP on day 14 (●), or Cy 200 mg/kg IP on day14 and 5 × 107 spleen cells from fully MHC-mismatched C57BL/6 (H-2b) donors, depleted ofCD8+ T cells on Day 15 (▲). Results show: (●) v (■) p<.0001, (●) v (▲) p=.02. (d) Cy +CD8-depleted allogeneic DLI prolongs survival of animals with established, metastatic renalcell carcinoma. BALB/c mice (H-2d; n=10/group) received 106 RENCA cells IV alone on Day0 either alone (■), or followed by treatment with Cy 200 mg/kg IP on day 14 (●), or Cy 200mg/kg IP on day 14 and 5 × 107 spleen cells from fully MHC-mismatched C57BL/6 (H-2b)donors, depleted of CD8+ T cells on Day 15 (▲). Results show: (●) v (■) p=.01, (●) v (▲)p=.001. All experiments were repeated at least once.
Symons et al. Page 14
Biol Blood Marrow Transplant. Author manuscript; available in PMC 2009 May 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Figure 2. Cy + CD8 depletion of DLI abrogates sustained engraftment of donor cells, GVHD andGVHD-associated aplasia(a,b) CD8 depletion of DLI abrogates sustained engraftment of donor cells administered afterCy. BALB/c mice (H-2d; n=10/group) were conditioned with Cy 200 mg/kg IP on day −1 andreceived 106 A20 lymphoma cells IV on Day 0. Mice received 5 × 107 spleen cells from fullyMHC-mismatched C57BL/6 (H-2b) donors on Day 0, either undepleted (■) or depleted ofCD4+ T cells (●), CD8+ T cells (▲), or both (□). Donor CD4+ T cell (a) and B220 (b) chimerismin peripheral blood was measured on days 3, 7, 14, and 21 after DLI via staining for H-2Kb
and H-2Kd antibody. (c) CD8 depletion abrogates DLI-induced GVHD. BALB/c mice(H-2d; n=10/group) were conditioned with Cy 200 mg/kg IP on day −1. Mice received 5 ×107 spleen cells from fully MHC-mismatched C57BL/6 (H-2b) donors on Day 0, eitherundepleted (■) or depleted of CD4+ T cells (●), CD8+ T cells (▲), or both (□). (d) CD8depletion of DLI does not induce GVHD-associated aplasia. BALB/c mice (H-2d; n=10/group)were conditioned with Cy 200 mg/kg IP on day −1. Mice received 5 × 107 spleen cells fromfully MHC-mismatched C57BL/6 (H-2b) donors on Day 0, either undepleted (■) or depletedof CD4+ T cells (●), CD8+ T cells (▲), or 5 × 107 spleen cells from syngeneic BALB/c mice(H-2d) (○). ▲ indicates the WBC in a naïve BLAB/c mouse. Peripheral white blood cell count
Symons et al. Page 15
Biol Blood Marrow Transplant. Author manuscript; available in PMC 2009 May 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

was monitored via TruCOUNT™ on days 3, 7, 14, and 21 after DLI. All experiments wererepeated at least once.
Symons et al. Page 16
Biol Blood Marrow Transplant. Author manuscript; available in PMC 2009 May 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

Figure 3. Tumor expression of alloantigens is not required for the beneficial effect of Cy plus CD8-depleted DLICB6 F1 mice (H-2dxb; n=10/group) received 3×104 B16 melanoma cells IV on Day 0 followedby treatment with Cy 200 mg/kg IP on day 14 (■), or Cy 200 mg/kg IP on Day 14 and 5 ×107 spleen cells on Day 15 from syngeneic (CB6F1) (□), one haplotype matched (C57Bl/6)(●), or fully MHC mismatched (BALB/c) (○) donors, depleted of CD8+ T cells. The tableincluded describes the GVT and GVH relationships between the donor and recipient strainsgiven that B16 melanoma is of B6 background. Results show: (●) v (■) p=.04, (○) v (●) p=.06, (●) v (□) p= .04, (○) v (□) p=.03. All experiments were repeated at least once.
Symons et al. Page 17
Biol Blood Marrow Transplant. Author manuscript; available in PMC 2009 May 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
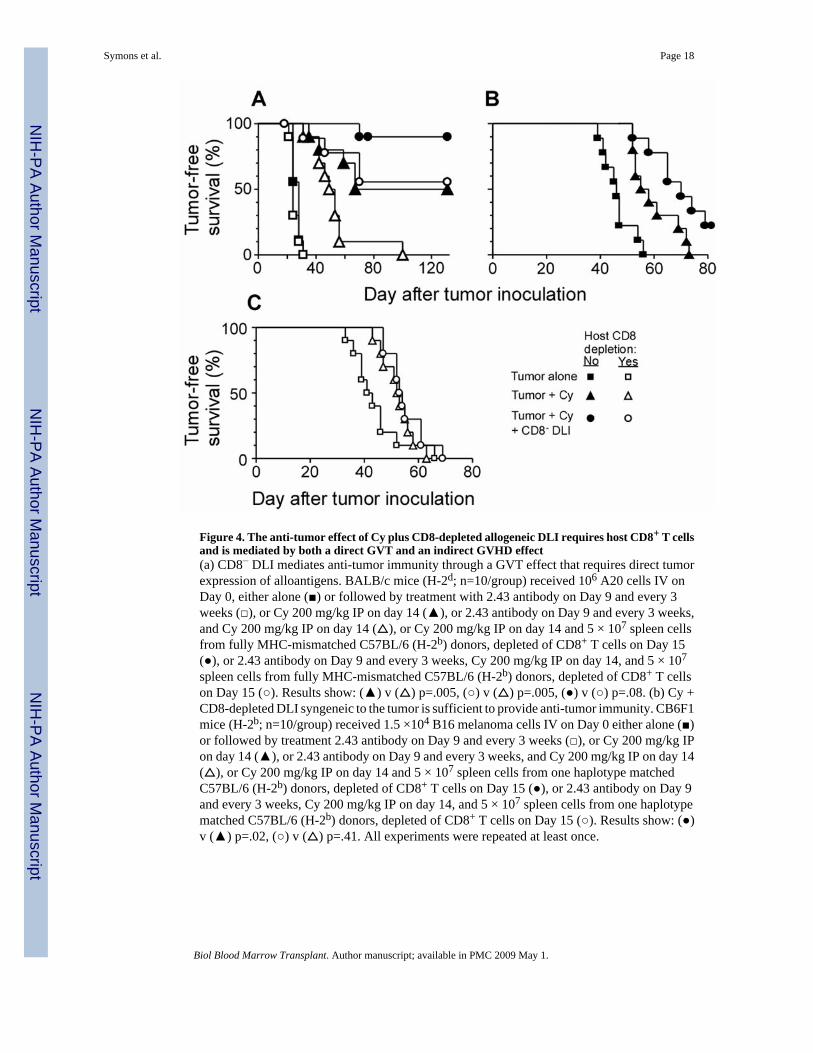
Figure 4. The anti-tumor effect of Cy plus CD8-depleted allogeneic DLI requires host CD8+ T cellsand is mediated by both a direct GVT and an indirect GVHD effect(a) CD8− DLI mediates anti-tumor immunity through a GVT effect that requires direct tumorexpression of alloantigens. BALB/c mice (H-2d; n=10/group) received 106 A20 cells IV onDay 0, either alone (■) or followed by treatment with 2.43 antibody on Day 9 and every 3weeks (□), or Cy 200 mg/kg IP on day 14 (▲), or 2.43 antibody on Day 9 and every 3 weeks,and Cy 200 mg/kg IP on day 14 (△), or Cy 200 mg/kg IP on day 14 and 5 × 107 spleen cellsfrom fully MHC-mismatched C57BL/6 (H-2b) donors, depleted of CD8+ T cells on Day 15(●), or 2.43 antibody on Day 9 and every 3 weeks, Cy 200 mg/kg IP on day 14, and 5 × 107
spleen cells from fully MHC-mismatched C57BL/6 (H-2b) donors, depleted of CD8+ T cellson Day 15 (○). Results show: (▲) v (△) p=.005, (○) v (△) p=.005, (●) v (○) p=.08. (b) Cy +CD8-depleted DLI syngeneic to the tumor is sufficient to provide anti-tumor immunity. CB6F1mice (H-2b; n=10/group) received 1.5 ×104 B16 melanoma cells IV on Day 0 either alone (■)or followed by treatment 2.43 antibody on Day 9 and every 3 weeks (□), or Cy 200 mg/kg IPon day 14 (▲), or 2.43 antibody on Day 9 and every 3 weeks, and Cy 200 mg/kg IP on day 14(△), or Cy 200 mg/kg IP on day 14 and 5 × 107 spleen cells from one haplotype matchedC57BL/6 (H-2b) donors, depleted of CD8+ T cells on Day 15 (●), or 2.43 antibody on Day 9and every 3 weeks, Cy 200 mg/kg IP on day 14, and 5 × 107 spleen cells from one haplotypematched C57BL/6 (H-2b) donors, depleted of CD8+ T cells on Day 15 (○). Results show: (●)v (▲) p=.02, (○) v (△) p=.41. All experiments were repeated at least once.
Symons et al. Page 18
Biol Blood Marrow Transplant. Author manuscript; available in PMC 2009 May 1.
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript

NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
NIH
-PA Author Manuscript
Symons et al. Page 19Ta
ble
ILy
mph
ocyt
e su
bset
s afte
r Cy
+ C
D8−
DLI
: Eff
ect o
f in
vivo
CD
8 de
plet
ion
Med
ian
% C
D8+ c
ells
in b
lood
(SE
M)
Med
ian
% d
onor
CD
4+ chi
mer
ism
(SE
M)
Day
afte
r C
D8−
DL
IN
o de
plet
ion
CD
8+ dep
lete
dp
No
depl
etio
nC
D8+ d
eple
ted
p7
2.9
(.76)
.12
(.03)
.000
67.
9 (4
.0)
26.9
(3.2
).0
114
5.3
(3.2
).3
0 (.4
3).0
3.5
4 (.7
5)31
.7 (5
.6)
.000
321
ND
*.2
4 (.1
3)-
0 (0
)0.
32 (1
.0)
.02
28N
D2.
0 (.6
9)-
0 (0
)0.
63 (.
50)
.02
BA
LB/c
mic
e (H
-2d ;
n=1
0/gr
oup)
eith
er d
eple
ted
in v
ivo
of C
D8+
T c
ells
with
2.4
3 an
tibod
y on
Day
9 a
nd e
very
3 w
eeks
, or u
ndep
lete
d, re
ceiv
ed C
y 20
0 m
g/kg
IP o
n D
ay 1
4, a
nd 5
× 1
07 sp
leen
cel
ls
from
fully
MH
C-m
ism
atch
ed C
57B
L/6
(H-2
b ) d
onor
s, de
plet
ed o
f CD
8+ T
cel
ls o
n D
ay 1
5. D
onor
CD
4+ c
ell c
him
eris
m (%
of t
otal
CD
4+ c
ells
) was
mea
sure
d w
eekl
y, fr
om d
ays 7
–28,
by
stai
ning
tail
bloo
d w
ith fl
uoro
chro
me-
conj
ugat
ed a
ntib
odie
s aga
inst
CD
4 an
d H
-2K
b (d
onor
). Pe
rcen
tage
of h
ost C
D8+
T c
ells
in ta
il bl
ood
was
als
o m
easu
red.
* ND
, not
det
erm
ined
Biol Blood Marrow Transplant. Author manuscript; available in PMC 2009 May 1.
Related Documents











