Jordi Serangeli Verbreitung der großen Jagdfauna in Mittel- und Westeuropa im oberen Jungpleistozän Ein kritischer Beitrag

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Jordi Serangeli
Verbreitungder großen Jagdfauna
in Mittel- und Westeuropaim oberen Jungpleistozän
Ein kritischer Beitrag

Tübinger Arbeiten zurUrgeschichte
Herausgegeben von
Nicholas J. ConardHans-Peter UerpmannHarald FlossMiriam N. HaidleMichael Bolus
TAU 3

Jordi Serangeli
Verbreitungder großen Jagdfauna
in Mittel- und Westeuropaim oberen Jungpleistozän
Ein kritischer Beitrag
Verlag Marie Leidorf GmbH . Rahden/Westf.
2006

Bibliografische Information der Deutschen Bibliothek
Serangeli, Jordi:
Verbreitung der großen Jagdfauna in Mittel- und Westeuropa im oberenJungpleistozän ; ein kritischer Beitrag / von Jordi Serangeli.Rahden/Westf.: Leidorf, 2006
(Tübinger Arbeiten zur Urgeschichte ; Bd. 3)Zugl.: Tübingen, Univ. ; Diss. ; 2004ISBN 3-89646-853-7
Die Deutsche Bibliothek verzeichnet diese Publikation in der Deutschen Nationalbibliografie.Detaillierte bibliografische Daten sind im Internet über http://dnb.ddb.de abrufbar.
Alle Rechte vorbehalten© 2006
Verlag Marie Leidorf GmbHGeschäftsführer: Dr. Bert Wiegel
Stellerloh 65 . D-32369 Rahden/Westf.
Tel: +49/(0)5771/ 9510-74Fax: +49/(0)5771/ 9510-75
E-Mail: [email protected]: http://www.vml.de
ISBN 3-89646-853-7ISSN 1438-8618
Kein Teil des Buches darf in irgendeiner Form (Druck, Fotokopie, CD-ROM, DVD, Internet oder einemanderen Verfahren) ohne schriftliche Genehmigung des Verlages Marie Leidorf GmbH reproduziert werden
oder unter Verwendung elektronischer Systeme verarbeitet, vervielfältigt oder verbreitet werden.
Umschlagentwurf: Harald Floss, TübingenTitelvignette – Umzeichnung: Jordi Serangeli, Tübingen
Scans, Satz und Layout: Jordi Serangeli, TübingenRedaktion: Maria Ronniger, Jörg Götze, Christoph Kümmel,
Harald Floss, Michael Bolus und Miriam N. Haidle, Tübingen
Kontaktadresse der Herausgeber:Eberhard Karls Universität Tübingen
Institut für Ur- und Frühgeschichte und Archäologie des Mittelalters,Abteilung Ältere Urgeschichte und Quartärökologie, Schloss, Burgsteige 11, D-72070 Tübingen
Internet: www.uni-tuebingen.de/uni/afa/index.html
Druck und Produktion: DSC-Heinz J. Bevermann KG, Fleethweg 1, D-49196 Bad Laer
XII, 255 Seiten mit 89 Abbildungen und 131 Tabellen

Vorwort der Herausgeber
Tierdarstellungen in der steinzeitlichen Kunst sind seitlangem ein Gegenstand der archäologischenForschung. Meist ging es dabei jedoch weniger um diedargestellten Tiere sondern mehr um den darstel-lenden Menschen und seine jeweiligen künstlerischenFähigkeiten oder um die zeitliche Einordnung dieserbesonderen Fundgattung. Daneben werden urge-schichtliche Tierdarstellungen auch von zoologischerSeite benutzt, beispielsweise um die Fellfärbung voneiszeitlichen Tieren zu bestimmen, die sich ausKnochenfunden nicht erschließen lässt. In den meistenFällen stehen einzelne Darstellungen oder bestimmteKunstformen im Zentrum solcher Betrachtungen. Dievorliegende Arbeit von Jordi Serangeli betritt inverschiedener Hinsicht thematisches und methodi-sches Neuland. Hier wird die paläolithische Kunst alshistorische Quelle in die Erschließung zoologischerErkenntnisse über die jungpleistozäne Jagdfaunaeingebunden – eine Vorgehensweise, die in vielenBereichen über das hinausgeht, was sich aus demVorkommen von Knochenfunden erschließen lässt.Denn der bildliche Bericht eines eiszeitlichenKünstlers, aus dem hervorgeht, dass er die darge-stellte Tierart kennt, hat einen anderen Stellenwert alsein mehr oder minder zufällig im Boden überlieferterKnochenrest. In der Kombination mit einerErschließung der Umweltverhältnisse und auf derBasis naturwissenschaftlicher Erkenntnisse über dieehemalige Verbreitung der Tierarten, die für die
paläolithischen Jäger wichtig waren, kommt JordiSerangeli zu Ergebnissen, die sowohl für Paläonto-logen und Archäozoologen wie für Archäologen undKunsthistoriker von Bedeutung sind. In manchenFällen erschließt die paläolithische Kunst Teile desVerbreitungsgebiets von Tieren, in denen entspre-chende Knochenfunde fehlen. In anderen Fällen fehleneher die Darstellungen, und man muss sich die Fragenach fehlender Überlieferung und ihren Gründenstellen. In der erfolgreichen Kombination einer kultur-wissenschaftlichen Quellengattung mit einer naturwis-senschaftlichen Fragestellung entsteht ein Ergebnis,das sowohl aus kulturgeschichtlicher wie aus zoologi-scher Sicht neue Erkenntnismöglichkeiten schafft.Dadurch wird einmal mehr der fließende Übergangzwischen Natur- und Kulturwissenschaft sichtbar, derfür zeitgemäße archäologische Forschung charakte-ristisch ist.
Tübingen 15.12.2005
H.-P. Uerpmann N.J.Conard
M. Bolus H. Floss M. N. Haidle


Vorwort
Die Einführung von Ackerbau und Viehzucht mit dendamit verbundenen Folgen (u. a. Sesshaftigkeit,Entwaldung, Einführung von fremden Pflanzen- undTierarten, lokales und regionales Aussterben voneinheimischen Tier- und Pflanzenarten) gilt als dererste große Eingriff des Menschen in die Natur. Davoragierten die Menschen in ihrem Ökosystem alsSammler und Jäger. Die Bevölkerungsdichte war,verglichen mit der heutigen, sehr gering. Der mensch-liche Einfluss auf die Natur war begrenzt und unterUmständen vergleichbar mit dem anderer Raubtiere.
In dieser Arbeit wird von dem Standpunkt ausge-gangen, dass es kaum möglich ist, die Lebensweisebzw. die Kultur im weitesten Sinne der Menschen imEuropa der letzten Eiszeit zu verstehen, ohne derenUmwelt gut zu kennen. Für dieses Verständnis spieltdie große Jagdfauna aus drei Gründen die zentraleRolle. Erstens sind archäozoologische Reste vongroßen Säugetieren nach den Steinartefakten diegrößte archäologische Fundgruppe. Zweitens sind inder paläolithischen Kunst Tausende von Tieren darge-stellt, die nach einer kritischen Betrachtung als Quellefür eine Umweltrekonstruktion zur Verfügung stehen.Drittens erlaubt uns die Kartierung von archäozoologi-schen Resten und bildlichen Darstellungen der großenJagdfauna, gekoppelt mit der Analyse der Bedürfnissederselben Fauna, Vergleiche zwischen der heutigenUmwelt und der Umwelt der letzten Eiszeit anzu-stellen.
Die Angaben über die Fauna und die Abbildungender Tierwelt werden in dieser Arbeit als zwei vonein-ander getrennte, unabhängige und gleichwertigeQuellen behandelt. Da die Datenbasis aus der Literaturstammt, können in dieser Hinsicht Fehlerübernommen worden sein. Eine eigene Aufnahme vonDaten oder eine Überprüfung der publizierten Datenam „Material“ war jedoch weder vorgesehen, nochhätte sie angesichts der großen Zahl an Funden undBefunden stattfinden können. Der Autor ist zudemArchäologe und nicht Paläontologe oder Archäo-zoologe.
Ich sehe diese Arbeit als einen Beitrag zumVerständnis der Lebensweise der Menschen imJungpaläolithikum Europas. Sie basiert auf Daten ausden einzelnen Fundstellen und Regionen, will aberweiter gehen, als die Besonderheit der einzelnenFundstellen und Regionen zu unterstreichen.
Es sind viele, denen ich hier gerne danken möchte.Daher geht mein erstes Dankeschön an alle dieje-nigen, die ich hier nicht namentlich erwähnen konnte.
Besonders möchte ich mich jedoch bei Prof. Hans-Peter Uerpmann bedanken, der sowohl als Dozent alsauch als Doktorvater an mich und an diese Arbeitimmer stark geglaubt und dieser Arbeit Zeit undInteresse gewidmet hat, und bei Prof. Nicholas JohnConard, der als Abteilungsdirektor diese Arbeitebenfalls immer unterstützt hat.
Ganz herzlich möchte ich mich für das Korrektur-lesen und die ausdauernde Diskussionsbereitschaft beiMaria Ronniger, Jörg Götze, Christoph Kümmel,Sascha Schmidt, Priv.-Doz. Dr. Michael Bolus, Priv.-Doz. Dr. Harald Floss, Dr. Miriam Noël Haidle, Prof.em. Hansjürgen Müller-Beck sowie bei Dr. ReinhardZiegler bedanken.
Durch ein Forschungsstipendium im Rahmen deseuropäischen Projektes „Migration and diffusion ofhominids and anatomically modern humans in theMediterranean basin in early prehistory: palaeoenvi-ronments, routes, settlements, subsistence“ konnte ichan der Universität Rovira i Virgili in Tarragona inSpanien forschen. Für dieses Stipendium möchte ichmich hier bei Prof. Eudald Carbonell (Tarragona,Spanien) herzlich bedanken. Im Laufe der Doktor-arbeit habe ich zudem folgende Institutionen besucht:die Universität von Girona (Prof. Narcis Soler), dasDeutsche Archäologische Institut in Madrid (Prof.Michael Kunst), die Universität Autonoma in Madrid(Prof. Arturo Morales), die Universität La Sapienza diRoma (Frau Prof. Margherita Mussi), die Universitätin Ferrara (Prof. Carlo Peretto) und das Museum fürdie Archäologie des Eiszeitalters auf SchlossMonrepos (Prof. Gerhard Bosinski). In allen diesenInstitutionen und von allen Professoren bin ich immersehr herzlich empfangen und unterstützt worden.Ihnen allen gebührt mein herzlicher Dank.
Durch das Landesgraduiertenförderungsprogrammdes Landes Baden-Württemberg konnte ich zwei Jahrelang konzentriert an der Dissertation arbeiten. DieserInstitution und deren Entscheidungsträgern giltebenfalls mein herzlicher Dank.
Schließlich möchte ich mich bei meiner Familiebedanken. Bei meinem Sohn Pierre, der geduldigakzeptiert hat, dass ich wegen dieser Arbeit oft spätnach Hause kam und nicht allzu viel Zeit gehabt habe,um mit ihm zu spielen. Bei meiner Frau Céline, weiles diese Arbeit ohne sie gar nicht gegeben hätte.
Jordi Serangeli


Inhaltsverzeichnis
Vorwort der Herausgeber ....................................................................................................................... V
Vorwort ..................................................................................................................................................... VII
Definition des Themas .......................................................................................................................... 1
Forschungsziel ................................................................................................................................... 1
Zeitlicher und geographischer Rahmen ............................................................................................. 1
Vorgehensweise ................................................................................................................................ 1
Abgrenzung des Themas ................................................................................................................... 2
Vergleichbarkeit der Daten ............................................................................................................... 3
Die Fauna . ................................................................................................................................... 3
Die Kunst ..................................................................................................................................... 3
Forschungsgeschichte in Mittel- und Westeuropa (Schwerpunkt Jungpaläolithikum) ................. 5
Frankreich ............................................................................................................................................ 7
Spanien ................................................................................................................................................. 10
Portugal ................................................................................................................................................ 12
Italien .................................................................................................................................................... 12
Deutschland .......................................................................................................................................... 14
Schweiz ................................................................................................................................................ 16
Niederlande, Belgien und Luxemburg ................................................................................................. 17
Großbritannien ..................................................................................................................................... 18
Skandinavien ........................................................................................................................................ 19
Forschungsimpulse aus dem Rest Europas .......................................................................................... 20
Forschungsimpulse aus den Vereinigten Staaten von Amerika ........................................................... 21
Überregionaler Vergleich .................................................................................................................... 21
Europa am Ende der letzten Eiszeit ....................................................................................................... 25
Topographie Europas ........................................................................................................................... 25
Klimaübersicht ..................................................................................................................................... 32
Jäger und Beute in Europa am Ende der letzten Eiszeit ...................................................................... 39
Der Mensch .......................................................................................................................................... 39
Die Raubtiere ....................................................................................................................................... 41
Raubtiere in der paläolithischen Kunst ................................................................................................ 45

Die große Jagdfauna der letzten Eiszeit in Europa .............................................................................. 49
Unpaarhufer (Perissodactyla) ............................................................................................................... 49
Pferd (Equus ferus) ......................................................................................................................... 49
Hydruntinus (Equus hydruntinus) .................................................................................................. 55
Wollnashorn (Coelodonta antiquitatis) .......................................................................................... 61
Waldnashorn (Stephanorhinus kirchbergensis) ............................................................................. 67
Steppennashorn (Stephanorhinus hemitoechus) ............................................................................. 67
Rüsseltiere (Proboscidea) ..................................................................................................................... 69
Mammut (Mammuthus primigenius) .............................................................................................. 69
Waldelefant (Elephas antiquus) ..................................................................................................... 81
Paarhufer (Artiodactyla) ....................................................................................................................... 83
Wildschwein (Sus scrofa) ............................................................................................................... 83
Rothirsch (Cervus elaphus) ............................................................................................................ 89
Damhirsch (Dama dama) ............................................................................................................... 93
Reh (Capreolus capreolus) ............................................................................................................. 97
Elch (Alces alces) ........................................................................................................................... 101
Rentier (Rangifer tarandus) ........................................................................................................... 105
Riesenhirsch (Megaloceros giganteus) .......................................................................................... 109
Steppenbison (Bison priscus) ......................................................................................................... 115
Auerochse (Bos primigenius) ......................................................................................................... 119
Moschusochse (Ovibos moschatus) ................................................................................................ 123
Steinbock (Capra ibex und Capra pyrenaica) ............................................................................... 129
Gämse (Rupicapra rupicapra) ....................................................................................................... 135
Saiga-Antilope (Saiga tatarica) ..................................................................................................... 141
Die große Jagdfauna an den Küsten Europas ....................................................................................... 147
Seehunde (Phocidae) .......................................................................................................................... 147
Waltiere (Cetacea) ............................................................................................................................... 159
Riesenalk (Pinguinus impennis) ........................................................................................................... 163
Mögliche Interpretation der Daten ........................................................................................................ 167
Paläolithische Kunst und ihre Deutung ................................................................................................ 167
Die Wandkunst ..................................................................................................................................... 168
Die Kleinkunst ..................................................................................................................................... 170
Die Verbreitung der Knochenreste und der Darstellungen der großen Jagdfauna ............................... 171
... und die Neandertaler? ...................................................................................................................... 175

Zusammenfassung ................................................................................................................................... 177
Abstract ..................................................................................................................................................... 178
Résumé ...................................................................................................................................................... 179
Riassunto ................................................................................................................................................... 180
Resumen .................................................................................................................................................... 181
Resum ........................................................................................................................................................ 182
Bibliographie ............................................................................................................................................ 185
Katalog der Faunenlisten ........................................................................................................................ 211
Fundstellen aus Großbritannien ..................................................................................................... 215
Fundstellen aus Belgien .................................................................................................................. 216
Fundstellen aus Frankreich ............................................................................................................. 218
Fundstellen aus Spanien ................................................................................................................. 232
Fundstellen aus Portugal ................................................................................................................. 242
Fundstellen aus Deutschland .......................................................................................................... 243
Fundstellen aus der Schweiz .......................................................................................................... 246
Fundstellen aus Italien .................................................................................................................... 247
Fundstellen aus Slowenien ............................................................................................................. 255


Definition des Themas
Forschungsziel
Primäres Ziel dieser Arbeit ist es, die Knochenresteund Darstellungen der großen Jagdfauna aus einemökologischen Blickwinkel zu untersuchen, umgenauere Aussagen über ihre Verbreitung zu erhalten.Dabei wird der Anwesenheit und Abwesenheit dereinzelnen Tierarten, deren starkem oder schwachemVorkommen sowie der Zusammensetzung der Faunen-listen besondere Aufmerksamkeit gewidmet. Währendder Arbeit hat sich weiterhin herausgestellt, dass die„seltenen“ Tierarten besonders relevante Ergebnisseliefern können.
Auf eine Analyse der kulturellen Komponente derjungpaläolithischen Kunst sowie der Jagdstrategienwurde bewusst verzichtet, da diese andere Methodensowie eine detailliertere Kenntnis der Funde vorausge-setzt hätte. Die Umwelt wird hier als Fundament ver-standen, auf dem die kulturelle Komponente aufbaut.Diese Arbeit will keineswegs die Bedeutung der kultu-rellen Komponente verneinen, die hinter den mensch-lichen Handlungen (Kunst, Jagdstrategie, Glaubens-vorstellungen usw.) steckt oder stecken könnte, jedochwird sie sich damit nicht beschäftigen.
Aufgrund des großen zeitlichen und geographi-schen Rahmens, der zahlreichen berücksichtigtenPublikationssprachen und wegen der weit zerstreutenPublikationsorgane erhebt diese Arbeit keineswegsAnspruch auf Vollständigkeit.
Zeitlicher und geographischer Rahmen
Der zeitliche Rahmen dieser Arbeit umfasst das oberePleistozän, das heißt die Zeit zwischen 40.000 und10.000 BP. Schwerpunkte sind durch die Anzahl, dieQualität und den Forschungsstand der Fundstellenvorgegeben. Ältere Fundstellen werden nur vereinzeltangesprochen, u. a. weil die Radiokarbon-Datierungs-methode jenseits von ca. 40.000 BP zu ungenau wird.
Der geographische Rahmen dieser Arbeit ist daswestliche Mitteleuropa und Westeuropa, das Gebietvon Deutschland bis Portugal und nach Südosten bisItalien (Abb.1). Einzelne Daten aus England undDänemark werden ebenfalls einbezogen. Derwestliche Teil von Österreich liegt zwar im unter-suchten Gebiet, lag aber während der Eiszeit unterGletschern begraben.
Aufgrund des großen Forschungsgebietes kannman die Kenntnis der Forschungsgeschichte in denjeweiligen Ländern nicht voraussetzen, diese spieltaber eine wesentliche Rolle für das Verständnis des
Forschungsstandes. Daher wurden in einem eigenenKapitel, vor der Behandlung des eigentlichen Themasder Arbeit, die wesentlichen Persönlichkeiten, Theo-rien, Ereignisse und Publikationen behandelt, die dieurgeschichtliche Forschung geprägt haben. Ein klarerSchwerpunkt wurde auf die Forschungsgeschichte desJungpaläolithikums gelegt.
Vorgehensweise
Die Angaben aus der Literatur über die große jungpa-läolithische Jagdfauna, nämlich bildliche Darstel-lungen und Faunenlisten, bilden die Datenbasis dieserArbeit. Alle Daten sind trotz Berücksichtigung der Artihrer Gewinnung für diese Arbeit relevant. Es werdendaher sowohl sichere als auch mögliche und in einigenFällen sogar fragliche Darstellungen ange-sprochen.Teilweise wird gerade dieser Vergleich es dem Leserermöglichen, sich seine eigene Meinung zu bilden.Große, in einigen Fällen polychrome Malereien habenin diesem Sinn den gleichen „Wert“ wie kleine,unscheinbare Gravierungen. Aspekte wie Größe,Farbe und Lage in der Fundstelle, Art des Trägersusw. spielen hier kaum eine Rolle. Auf solche Infor-mationen wurde demzufolge bei den Abbildungenverzichtet. Die Literaturangaben sollen dabei weiterhelfen.
Bei den Knochenfunden werden gleichermaßenAngaben über Anwesenheit oder Abwesenheit derTierarten, Mindestindividuenzahlen (MIZ) und die
Abb. 1 Geographischer Rahmen der Arbeit

2 Verbreitung der großen Jagdfauna in Mittel und Westeuropa im oberen Jungpleistozän
Zahl der bestimmten Knochen oder die Gewichtsan-gabe berücksichtigt. Welche Methode zur Gewinnungder Daten benutzt wurde, wird immer angegeben.
Es ist hier zu betonen, dass beide Datenkate-gorien, Fauna und Kunst, zwei voneinander getrennteund unabhängige Quellen sind.
Zur Auswertung werden die Daten in Pro-zentzahlen angegeben. Da in manchen Fundstellen derProzentanteil von bestimmten Tieren verschwindendgering ist, werden die Prozentzahlen zum Teil mit biszu zwei Dezimalstellen angegeben. Da in der vorlie-genden Arbeit aber eher Tendenzen thematisiert wer-den, sollen die Prozentzahlen nicht überbewertetwerden. Bei zahlreichen Tierarten wird insbesondereihre Vergesellschaftung berücksichtigt.
Bei der Darstellung der Daten wird auf ihre Aussa-gekraft geachtet. Gut datierte Fundstellen mit einemaußerordentlich reichen Vorkommen einer Tierartwerden eine wichtige Rolle spielen. Gleichzeitigwerden aber auch seltene Tiere besonders starkberücksichtigt. So sind z. B. Reste oder Darstellungenvon Rentieren und Mammuten in Südeuropa viel inte-ressanter als die viel zahlreicheren Reste und Darstel-lungen aus Frankreich und Deutschland. Daherwerden die wenigen Hinweise auf diese Arten ausSpanien und Italien ausführlicher präsentiert unddiskutiert als diejenigen aus Mitteleuropa.
Um eine korrekte Deutung der Daten zu ermögli-chen, werden, dem aktualistischen Prinzip entspre-chend, die Umweltansprüche rezent vorkommenderHerbivoren (u. a. Steinbock, Saiga-Antilope, Mo-schusochse, Rentier) und vom Wildschwein unter-sucht und dargestellt. Dabei werden bewusst nicht nurdie Lebensansprüche der in Europa lebenden Arten(siehe z. B. Griggo 1995), sondern auch die aus demasiatischen Teil der ehemaligen Sowjetunion zumVergleich herangezogen (Heptner et al. 1966). Weiter-hin sollen, soweit möglich, auch ausgestorbene Arten(u. a. Mammut und Riesenhirsch) auf ihre möglichenUmweltansprüche untersucht werden. Um das Bild derVerbreitung der großen Jagdfauna in der letzten Eis-zeit in Mittel- und Westeuropa zu ergänzen, werdenebenfalls Meeressäuger (Cetacea und Phocidae) so-wie der Riesenalk (Pinguinus impennis) berücksich-tigt, da sie die Beziehung des Menschen zurWasserwelt gut veranschaulichen.
Daten über die Verbreitung von Tierarten werdenanhand von Verbreitungskarten dargestellt. Hierknüpft diese Arbeit sowohl an Arbeiten über die Ver-breitung heutiger Tierarten (z. B. Heptner et al. 1966;
Niethammer und Krapp 1986; Griggo 1995; Mitchell-Jones et al. 1999) als auch an solche über die Verbrei-tung der Tiere während der letzten Eiszeit in Europa(z. B. Villeneuve und Boule 1906–1910; Kahlke 1994)oder in der Vorgeschichte des Vorderen Orients(Uerpmann 1987) an.
Ein Katalog mit den Daten aus den in dieser Arbeitberücksichtigten Fundstellen wird am Ende der Arbeitden Vergleich der Daten erleichtern. Die stets vorhan-dene Angabe über die Quellen will hier nicht nur alsbibliographischer Hinweis verstanden werden, son-dern als Bitte, die primäre Literatur, wo die Fundstel-len ausführlich behandelt worden sind, ergänzend zulesen.
Abgrenzung des Themas
Unter „großer Jagdfauna“ werden in dieser ArbeitHerbivoren ab der Größe von Saiga tatarica (Saiga-Antilope) bis zum Mammuthus primigenius (Mam-mut), sowie Sus scrofa (Wildschwein) verstanden.Raubtiere, wie z. B. Canis lupus (Wolf) oder Pantheraleo (Löwe), sowie kleinere Säuger wie Oryctolaguscuniculus (Wildkaninchen) und Erinaceus europaeus(Igel), wie auch Kleinsäuger werden nicht berück-sichtigt. Zum einen, weil es bekannt ist, dassRaubtiere, Hasenartige und Kleinsäuger, anders als diegroßen Herbivoren, selbst Höhlen nutzten. Daherkönnen ihre Knochen in Höhlen auf natürliche Weisein großer Zahl vorkommen. Zum anderen, weil dieJagd auf sie kaum die gleichen Ziele oder die gleicheTechnik und Rolle wie die Jagd bei der „großenJagdfauna“ gehabt haben kann. Man kann sich z. B.nicht auf die Jagd auf Raubtiere zum Zweck derErnährung verlassen, da es schnell zur Überjagungkommen würde. Auch können bei der Jagd aufRaubtiere andere Faktoren wie Prestige oder Bedarf anFellen eine größere Rolle gespielt haben (u. a. Charles1997). Die Jagd auf Hasen und Wildkaninchen1
andererseits, die in Südeuropa und insbesondere aufder Iberischen Halbinsel mit Sicherheit eine großeRolle gespielt hat (u. a. Hockett und Bicho 2000), hatwahrscheinlich unter Verwendung anderer Waffen alsdie Jagd auf die „große Jagdfauna“ stattgefunden.Reste von Vögeln, Amphibien, Fischen, Reptilien undMollusken werden ebenfalls in dieser Arbeit nichtberücksichtigt.
Als Darstellungen von Tieren sind sowohl Gravie-rungen als auch Malereien, Halbreliefs oder Vollpla-stiken gemeint.
1 Darstellungen von Leporidae sind auch in der Kunst nicht so exzeptionell, wie man zuerst glauben würde: u. a.Acosta und Molinero 2003; Airvaux 2001, 120–121; Altuna 1996b, 156; Gaussen 1993; vielleicht Lejeune 1987,39; Leonardi 1988, 158, Abb. 41; Pales und de Saint Pereuse 1989; Radimilli 1974.

Definition des Themas 3
Vergleichbarkeit der Daten
Die Fauna
Faunenreste sind von Natur aus genormte Körper. DasMaterial ist bekannt, nämlich Knochen, Zahn,Elfenbein oder Geweih. Die anatomische Position desKnochens ist, einmal bestimmt, vorgegeben. Selbst dieFunktion des Knochens ist bekannt.
Das verhindert jedoch nicht, dass Schwierigkeitenbeim Vergleich der unterschiedlichen Faunenlistenauftreten (siehe dazu Münzel 1988; Uerpmann 1997).Der Grund dafür besteht darin, dass die Archäozoo-logen sich nicht auf einheitliche Aufnahmesystemegeeinigt haben oder sich nicht einigen konnten.Manche Autoren (z. B. Delpech 1983, 362; Sala 1985)zählen bei ihren Faunenlisten Herbivoren ab derGröße von Saiga tatarica (Saiga-Antilope) mit,andere, insbesondere in Südeuropa, nehmen auchOryctolagus cuniculus (Wildkaninchen) undErinaceus europaeus (Igel), Fleischfresser (Nadal etal. 1997) und Geweihfragmente in ihre Prozentzahlenauf (Cassoli und Tagliacozzo 1994). Andere rechnenzusätzlich noch Vögel und Fische dazu (z. B. Soffer1985, 123–135).
Verwirrung entsteht auch dadurch, dass mancheAutoren nur auf Artniveau bestimmte Tiere in ihrenListen aufnehmen, während andere auch die Knochenaufführen, die lediglich einer Familie zugeordnet wer-den können. Manche Autoren publizieren, wie vieleKnochen nicht bestimmt werden konnten, anderenicht.
Auch die Methoden, die Daten zu präsentieren,unterscheiden sich wesentlich voneinander. MancheAutoren rechnen nach Bestimmung der Knochen nurdie Mindestindividuenzahl aus (MIZ; englisch: Mini-mal Number of Individuals, MNI). Andere errechnendiese nicht und geben nur die Zahl der bestimmtenKnochen an (n; englisch: Number of Identified Speci-men, NISP). Wieder andere halten das Gewicht derKnochen für relevanter (Münzel 1988). In manchenFaunenlisten ist nicht einmal erwähnt, welcheMethode verwendet wurde.
Zusätzlich besteht oft bei älteren Faunenlisten derVerdacht, dass nur leicht bestimmbare Knochen, ins-besondere Zähne und vollständige Stücke, gezähltwurden, während man die anderen „entsorgte“.
Als letztes Hindernis sei hier auf die fehlerhafteWiedergabe der Daten insbesondere in der Sekundärli-teratur hingewiesen. Hier erhält man nach demZusammenzählen der einzelnen Prozentzahlen manch-mal deutlich weniger oder deutlich mehr als die zuerwartenden 100 Prozent (siehe hierzu die Einführungim Katalog der Faunenlisten, S. 213–214).
Das alles wird erwähnt, um darauf hinzuweisen, dassein überregionaler Vergleich sehr schwierig ist undman daher als Ergebnis nicht mehr als Tendenzenerwarten kann. Beschränkt man sich auf Tendenzen,was bei einer so weiträumig angelegten Arbeit sinn-voll erscheint, spielt die Methode, die zur Gewinnungder publizierten Daten verwendet wurde, eher einezweitrangige Rolle.
Die Kunst
In dieser Arbeit soll, wie schon erwähnt, der Kunsteine besondere Rolle beigemessen werden. Es stehtaußer Frage, dass die Themen der Kunst eine großekulturelle Komponente haben2, sie wird jedoch viel zuoft als rein „kulturelle“ Erscheinung betrachtet,wodurch ihre Bedeutung für die Verbreitung derTierarten abgestritten oder vernachlässigt wird (siehedas Kapitel „Mögliche Interpretation der Daten undErgebnisse“, S. 167–175). Die mehrfach nachge-wiesene Tatsache, dass die Anzahl der dargestelltenTiere oft diametral gegenüber der Anzahl der durchKnochenfunde belegten gejagten und verspeistenFauna stehen kann (u. a. Delporte 1984b; Altuna 1994,1996b), zeigt nur, dass diese zwei Gruppen nichtkongruent sind. Der überregionale Vergleich der
Abb. 2 Oben: „Mammut von Altamira“ (Jordá1983, 267). Mitte: Eine Boa, welche einen Elefantverschlungen hat. Unten: Erklärung des mittlerenBildes (Saint-Exupéry 1946).
2 U. a. siehe Serangeli 2004.

4 Verbreitung der großen Jagdfauna in Mittel und Westeuropa im oberen Jungpleistozän
Themen sowie deren Verbreitung zeigen, dass dieKunst wertvolle zusätzliche Informationen liefernkann (u. a. Serangeli 2003). Dabei ist in manchenFällen das schwierigste Problem die sichereZuordnung der von Menschen verursachten Striche,Linien und Bemalungen zu einer konkreten Figur. Da,wo manche Autoren ein bestimmtes Tier zu erkennenglauben, können andere, mit genau so gutenArgumenten, etwas anderes oder sogar nichtserkennen. Damit steht man oft vor dem Dilemma, obdie eine oder die andere Meinung richtig ist.
Das wiederholte Lesen eines Klassikers, „Le PetitPrince“ (Der Kleine Prinz) von Saint-Exupery, zeigtemir, dass manchmal die eine und die andere Interpre-tation richtig sein kann (Abb. 2), es kommt lediglichauf den Betrachter an. Daher habe ich michentschieden, auch fragliche Interpretationen demLeser zu präsentieren, um eine subjektive Vorauswahlzu vermeiden. Die Ansprache einer Darstellung alseine bildliche Wiedergabe eines bestimmten Tieresbasiert auf der Interpretation der zitierten Autoren.Eine tiefere und genauere Diskussion über diesesThema hätte verlangt, sich die Originale vor Ortanzuschauen, was im Rahmen dieser Arbeitunmöglich zu erreichen gewesen wäre.
Einige weitere Vorbemerkungen sind an dieserStelle notwendig.
Zuerst muss betont werden, dass das, was man die„Kunst des Jungpaläolithikums“ nennt, sich über eineZeit von über 25.000 Jahren erstreckt und eine Regionabdeckt, welche von Südspanien bis zum Ural reicht.Jean-Pierre Bocquet-Appel und Pierre-Yves Demarsgehen für das Gebiet von Frankreich bis Deutschlandund Großbritannien von einer Population von 5.400bis 8.900 Individuen für die Zeit vom Aurignacien biszum zweiten Kältemaximum und von mehr als 40.000für das Magdalénien aus (Bocquet-Appel und Demars2000, 568).
Weiterhin sollte man bedenken, dass das, was anKunst überliefert ist, durch das engmaschige Sieb derZeit und der Einflüsse von Geologie und Chemie desBodens gefiltert wurde, wobei viele andere Zerstö-rungsfaktoren beteiligt waren. Wir werden wahr-scheinlich kaum etwas über die Malereien außerhalbder Höhlen oder über die bildlichen Darstellungen aufHolz oder Leder erfahren (Holdermann 2001; Müller-Beck 2001). Unzählige Beispiele aus der Ethnologielassen keinen Zweifel an der ehemaligen Existenzderartiger Kunstwerke aufkommen. Daher basierenalle Studien und Theorien über die paläolithischeKunst, so auch in dieser Arbeit, nicht auf der gesamten
von den Menschen des Jungpaläolithikums produ-zierten Kunst, sondern auf der kleinen Anzahl anDarstellungen, die uns überliefert ist.
Zugleich darf man nicht vergessen, dass die Men-schen des Jungpaläolithikums, anders als wir, nichtgleichzeitig die Höhlen von Chauvet, Cosquer, Las-caux, Altamira, Nerja, Paglicci, Kapova oder aus Süd-westdeutschland kannten. Sie kannten noch nichteinmal eine einzige Höhle so wie wir sie heute ken-nen. Dank der elektrischen Beleuchtung und der sorg-fältigen und zeitlich aufwändigen Aufnahme derWände können wir heute, dort wo die Wände erhaltensind, auch die feinsten Gravierungen wahrnehmen.Dank der Ausgrabungen sehen wir nebeneinanderObjekte, die teilweise im Abstand von 100, 1000 odersogar 10.000 Jahren in den Boden kamen. Daherreicht es nicht aus, wenn man sich mit paläolithischerKunst beschäftigt, einen detaillierten und überregio-nalen Überblick des Themas zu haben, sondern manmuss ebenfalls in der Lage sein, davon Abstand zugewinnen, um die bildlichen Darstellungen in einembegrenzten Zeitfenster von nur 40 Jahren (eine opti-mistische Einschätzung der damaligen Lebens-erwartung), in ihrem lokalen Kontext mit denbeschränkten, damals existierenden Beleuchtungs-möglichkeiten zu sehen.
Als ob das Ganze noch nicht kompliziert genugwäre, kamen in den letzten 15 Jahren, dank derEntwicklung der 14C-AMS-Methode, direkte Datie-rungen der Höhlenmalereien hinzu. Man bedenke nurdie Diskussion über die Grotte Chauvet und derenDatierung. Einige Autoren sind der Ansicht, dass die14C-Datierungen um 30.000 BP aus der Höhle vonChauvet zeigen, dass die Malereien aus demAurignacien stammen müssen (u. a. Clottes 2001;Valladas und Clottes 2003). Damit wäre die bis dahinübliche stilistische Datierung nach den Kriterieninsbesondere von André Leroi-Gourhan nicht mehranwendbar. Andere (insbesondere Züchner 1999 und2001) meinen, dass die Höhle von Chauvet sich stili-stisch sehr wohl mit dem Magdalénien vergleichenlässt. Dies würde implizieren, dass die zahlreichenDatierungen aus dieser Höhle alle falsch oder proble-matisch seien. Es ist möglich, dass aufgrund systema-tischer Verunreinigungen der Proben durchWandkarbonate die Datierung fraglich ist (Pettitt undBahn 2003). Diese Diskussion mit ihren Implika-tionen scheint noch weit von einer Lösung und einemEnde entfernt zu sein.
Schließlich muss man noch erwähnen, dass dergrößte Teil der Wandkunst, nämlich die Gravie-rungen, nicht direkt zu datieren ist.

Forschungsgeschichte in Mittel- und Westeuropa (Schwerpunkt Jungpaläolithikum)
Vorwiegend verwendete Literatur: Barton 1997; Bosinski 1982; Brizzi 1977; Campbell 1977; Canal undCarbonell 1989; Daniel 1990; Estévez und Vila 1999; Groenen 1994; Kristiansen 1985; Le Tensorer 1998;Lorblanchet 1995; Müller-Karpe 1966; Otte 1979; Palma di Cesnola 1993; Reitz und Wing 1999; Straus 1992;Zilhão 1997.
„Auch die Wissenschaft ist Zeitströmungen und sogarModen unterworfen“ (Hediger 1980, 13–14). Dies giltnaturgemäß besonders bei einer solchen Disziplin wieder Urgeschichte, die als Ziel Erkenntnisse über diemenschliche Vergangenheit hat. Daher muss zumVerständnis des Forschungstandes am Beginn dieserArbeit ein Überblick über die Entwicklung dieserDisziplin in den verschiedenen Ländern stehen.Manche Schlussfolgerungen werden direktangesprochen, andere kann man aus den hier gesam-melten und wiedergegebenen Daten entnehmen. DieForschungsgeschichte sagt uns nicht, wohin die Reisegeht, aber sie sagt uns, woher wir und viele unsererGedanken kommen.
Grundsätzlich kann man vier Perioden unter-scheiden, welche sich in allen europäischen Ländernungefähr gleich wiederholen (u. a. Müller-Karpe 1966,1–18; Straus 1992, 24–37).
Eine erste Phase, nach der Zeit der Kuriosi-tätenkabinette (Daniel 1990, 48–54), erstreckte sichvon den Anfängen des 19. bis zum Beginn des 20.Jahrhunderts. Das Interesse an der Urgeschichte kamaus Disziplinen wie Geologie, Paläontologie, Anthro-pologie, Ethnologie, Kunstgeschichte. Es gingeinerseits um grundlegende Fragen, beispielsweise, obund wie viele Eiszeiten es gegeben hatte, welche Tiereund welche Menschen in Europa während derUrgeschichte gelebt hatten, inwieweit diese Menschen„entwickelt“ oder „primitiv“, wie ihre geistigen undkünstlerischen Fähigkeiten ausgeprägt waren. Ande-rerseits ging es darum, möglichst viele und „schöne“Stücke zu sammeln, um diese dann mit Objekten ausanderen archäologischen oder ethnologischen Samm-lungen zu vergleichen. Der Zusammenhang der Fundespielte in dieser ersten Periode meistens eine unter-geordnete Rolle3. Klare Gesetze für den Schutz vonarchäologischen Fundstellen gab es kaum (Cleyet-Merle et al. 1990, 56–57). Aus dieser Zeit stammenviele der Privatsammlungen, die heute als Bestand-teile von Museums- und Universitätssammlungen inder ganzen Welt verstreut sind. Darunter sei hier auf
die reiche Sammlung des „Museums für Vor- undFrühgeschichte“ in Berlin hingewiesen, das sogar dieEntfernung des Reliefs eines Lachses aus dem Abri duPoisson à Gorges-d’Enfer (Dordogne) in Auftraggegeben hatte (Peyrony 1932, 246; Cleyet-Merle et al.1990, 72–75). Manche Privatsammlungen entstandensogar später durch fragwürdigen Erwerb (Bayer 1925).
Die Kunst spielte in den damaligen Diskussioneneine besondere Rolle. Die Herstellung eines nichtzweckgebundenen Gegenstandes ist vielleicht daseindeutigste Merkmal, welches Mensch und Tiervoneinander trennen lässt. Die Entdeckung einerMammutgravierung im Abri von La Madeleine imJahr 1864 lieferte einen nicht anzuzweifelnden Beweisfür die Gleichzeitigkeit des Menschen und ausge-storbener Tierarten (Müller-Karpe 1966, 3). Diebesondere Kunstfertigkeit und Feinfühligkeit derpaläolithischen Künstler für die Formen, die Verzie-rungen und oft auch für das Detail führte in dieserPhase der Forschung zu der Theorie der „l’art pourl’art“ (Lorblanchet 1995, 89–90). Die bildlichenDarstellungen wurden insbesondere als ästhetischeAusdrücke betrachtet und die Vorliebe für Verzie-rungen wurde als der Funke, der Motor, der Grund fürdie Kunst gesehen. Die Objekte der Kleinkunst – dieWandkunst war zwar schon entdeckt, aber von denmeisten Forschern nicht anerkannt worden – wurdenals Sammlerstücke betrachtet (Groenen 1994, 305–350).
Die Tierknochen wurden damals insbesondere imHinblick auf die Frage untersucht, welche ausgestor-benen Tiere gleichzeitig mit dem Menschen in derUrgeschichte gelebt haben. Gerade die ausgestor-benen Tierarten waren der Beleg für das hohe Altervieler Fundstellen. Etwas später kam den Tierknocheneine feinere chronologische Bedeutung zu (Reitz undWing 1999, 16). So schlug Edouard Lartet eine Unter-teilung des Paläolithikums in eine erste Periode desHöhlenbären, eine zweite Periode des Wollmammutsund des Wollnashorns, eine dritte Periode des Rentiersund eine vierte Periode des Auerochsen und des
3 Man denke an die Höhlen im Neandertal, aus denen bis zu den Nachgrabungen von 1997 kaum mehr als dieKnochen des Neandertalers bekannt war (Schmitz und Thissen 2000).

6 Verbreitung der großen Jagdfauna in Mittel und Westeuropa im oberen Jungpleistozän
Steppenbisons vor. Diesen setzte Joseph Garrigou eineZeitstufe des Elefanten warmer Klimate und derFlusspferde voraus (Daniel 1990, 75).
Eine zweite Phase, grob gerechnet vom Ende des 19.Jahrhunderts bis in die 1930er Jahre, begann mit derAnerkennung der paläolithischen Wandkunst. Diese,zusammen mit der immer größer werdenden Anzahlan Objekten aus der mobilen Kunst, erforderte einInterpretationsmodell. Zahlreiche Gelehrte, insbe-sondere katholische Geistliche wie Abbé Henri Breuil,welche häufig ebenfalls an der Ethnologie interessiertwaren, setzten sich mit der paläolithischen Kunstauseinander. Der religiöse und magische Charakter derpaläolithischen Darstellungen wurde unterstrichen. Esentstand ein Modell, in dem die Jagd, das Leben undder Tod der Tiere die zentrale Rolle spielten(Lorblanchet 1995, 90).
Der Zweite Weltkrieg, während dem naturgemässkeine internationalen Kongresse stattfanden4, bildeteeinen Wendepunkt nicht nur in der GeschichteEuropas, sondern auch in der paläolithischenForschung. Über Themen wie physische Anthropo-logie sowie über ethnologische Vergleiche mit „primi-tiven“ Völkern wurde stark nachgedacht. Damit fingeine dritte Phase an, die bis zum Ende der 1960erJahre andauerte. Die von Libby 1949 bekanntgegebene Entdeckung der Radiokarbondatierungrevolutionierte die Archäologie wie keine andere. DiePalynologie, die seit 1836 bekannt war, nahm stark anBedeutung zu. Man versuchte, mit Hilfe von Natur-wissenschaften zusätzliche Daten zu gewinnen (siehe:Daniel 1990, 215; Filip 1969, 1067). Eine strengeTypologie der Artefakte, begleitet durch dieEntwicklung statistischer Methoden, schien fürOrdnung unter den Technokomplexen zu sorgen (u. a.Sonneville-Bordes und Perrot 1954). André Leroi-Gourhan war die dominierende Figur im Bereich derpaläolithischen Kunst in dieser Zeit. Er setzte die stati-stische Analyse der bildlichen Darstellungen, sowohlin thematischer Hinsicht als auch in Hinblick auf dieVerteilung der Themen in der Höhlen, durch.
Weiterhin entwickelte er die Hypothese einer fastlinearen stilistischen Entwicklung der Kunst währenddes Jungpaläolithikums in Westeuropa. Diese sollteeine zeitliche Einordnung der Kunst, insbesondere derWandkunst, anhand von Vergleichen ermöglichen(Leroi-Gourhan 1965).
Die letzte Periode, die mit den 1970er Jahren begannund bis heute andauert, ist durch eine starke Ausbrei-tung naturwissenschaftlicher Methoden gekennzeich-net. Um die Archäologie entstanden neue Methodenund andere entwickelten sich zu eigenständigenForschungsrichtungen, wie die Archäozoologie, diePollenanalyse, die radiometrische Datierung und dieGebrauchsspurenanalyse. In der Untersuchung vonSteinartefakten nahmen Fragen nach der Herkunft derRohmaterialien, den Beschaffungsstrategien und demtechnischen Abbau an Bedeutung zu. Man wollte undwill nicht nur den Menschen als kulturelle Erschei-nung untersuchen, sondern man will auch seine Weltund seine Umwelt verstehen. Im spezifischen Bereichder paläolithischen Kunst sind ebenfalls wichtige Ent-wicklungen in dieser Richtung zu vermerken. Heutekann man Malereien, welche einen hohen Anteil anorganischem Material haben, beispielsweise Kohle,direkt datieren. Chemische Untersuchungen erlaubenzu bestimmen, welche Mischung benutzt wurde, umeine Farbe herzustellen. Jedoch hat diese Entwicklungeine Kehrseite. Manche Forscher lehnen schon alleindie Idee des Versuches einer Interpretation der paläoli-thischen Kunst ab oder halten die Beschäftigung mitFelsbildern für unseriös5.
Erst in dieser letzten Periode entwickelt sich dieUntersuchung der zoologischen Funde aus archäolo-gischen Grabungen als eigene Forschungsrichtung.Vorher war dieser Bereich entweder den Paläon-tologen überlassen worden oder wurde von nicht dafürausgebildeten Archäologen nebenbei geführt. Daherist es nicht verwunderlich, dass so manche alteBestimmung heutzutage als fehlerhaft oder falschbezeichnet werden muss6, andere sind in ihrerAussage zwar für paläontologische Erkenntnisse nütz-lich, für eine archäozoologische Auswertung jedoch
4 Der vorgesehene dritte Kongress des CISPP (Congrès international des Sciences préhistoriques et protohisto-riques), Vorgänger der UISPP (Union internationale des Sciences préhistoriques et protohistoriques), der in Buda-pest 1940 hätte stattfinden sollen, fand erst 1950 in Zürich statt (Nenquin 1994).5 In diesem Kontext sei die Position von drei der bedeutendsten Experten für prähistorische Kunst durch zweiZitate geschildert. „Allzu häufig sind die Prähistoriker – die sich nur dafür interessieren, was sie für „Fakten“ hal-ten – der Ansicht, dass die Erklärungsversuche nicht mehr als Spekulationen seien, die manchmal zwar ganzannehmbar klingen (aber Hypothesen sind), in den meisten Fällen jedoch wie wildgewordene Phantasien auseinem Fieberwahn anmuten. Die Mehrzahl der Fachleute für paläolithische Kunst befaßt sich mit den Höhlen,ohne auf die Frage der Interpretation überhaupt einzugehen“ (Clottes und Lewis-Williams 1997, 78–79). „Geradein Deutschland wird die Beschäftigung mit Felsbildern gewöhnlich von der Fachforschung als mehr oder wenigerexotisch bis unseriös abgelehnt, ohne dass man sich darüber im Klaren wäre, welche Erkenntnismöglichkeitendiese Quellengattung für prähistorische Kulturen und Kulturzusammenhänge bietet“ (Züchner 2000).6 Hier sei nur als Beispiel auf die fehlerhafte Ansprache von Knochen, welche zu den Elephantidae gehören, alsWale hingewiesen (Poplin 1983).

Forschungsgeschichte in Mittel- und Westeuropa (Schwerpunkt Jungpaläolithikum) 7
viel zu ungenau7. Noch heutzutage ist es fast immerdie Regel, dass der Archäozoologe kaum eine Chancehat, auf die Bergung der Funde und Befunde Einflusszu nehmen, so dass damit manche Informationenverloren gehen (Altuna 1972, 426).
Da dieser Forschungszweig relativ jung ist, scheintseine Rolle für die Archäologie noch nicht ganzgeklärt zu sein. Während man im kontinentalenEuropa eher von Archäozoologie spricht, wird imenglischsprachigen Raum vor allem die Bezeichnungzooarchaeology verwendet (Reitz und Wing 1999).Obwohl im Grunde die gleiche Datenbasis und auchdie gleichen Arbeitsmethoden verwendet werden, istdie Archäozoologie im engeren Sinne also einearchäologische Disziplin mit allen archäologischenPflichten, die speziell zoologische Methoden anwen-det und die Entwicklung der Tiere anhand der Datenaus archäologischen Fundstellen untersucht, währendsich die zooarchaeology bemüht, die kulturelle Rolleder Tiere im archäologischen Zusammenhang zurekonstruieren. Andere zum Teil nur leicht abwei-chende Forschungsrichtungen sind die Palethnozoolo-gie8, die Anthropozoologie9 und Osteoarchäologie10
(u. a. Reitz und Wing 1999, 2ff.). Um keine unnötigeVerwirrung zu verursachen, wird in dieser Arbeit aus-schließlich der Begriff Archäozoologie verwendet,womit auch der eine oder andere abweichende Begriffgemeint ist.
Frankreich
Vorwiegend verwendete Literatur: Groenen 1994;Cleyet-Merle et al. 1990.
Von 1837 bis 1847 führte Jaques Boucher de Perthesin Abbeville, Département Somme, eine Ausgrabungdurch, so, wie bis dahin viele stattgefunden hatten. Erbemerkte aber, dass Knochen ausgestorbener Säuge-tiere (u. a. Elephas antiquus und Stephanorhinuskirchbergensis) und Steinartefakte (hauptsächlichFaustkeile) nebeneinander in ungestörten, damals alsvorsintflutlich bezeichneten Schichten lagen. Obwohl
er schon 1839 seine Schlussfolgerung in der PariserAkademie vortrug, dauerte es noch 20 Jahre, bis diesedamals revolutionäre Ansicht allgemein akzeptiertwurde (Groenen 1994, 62ff.). Dazu führten vielekleine Schritte, darunter die Publikation der „Anti-quités celtiques et antédiluviennes“ durch JacquesBoucher de Perthes11 in drei Bänden (1847–1864), dieAusgrabungen von Marcel-Jérôme Rigollot (1854) inder Kiesgrube von Saint Acheul12 und 1859 dieBegutachtung des Fundplatzes von Abbeville durchden englischen Geologieprofessor Charles Lyell. Nochim Herbst desselben Jahres wurde die Existenz vonSteinartefakten in „vorsintflutlichen“ Schichten vonden wissenschaftlichen Gesellschaften in London undvon der Pariser Akademie anerkannt (Groenen 1994,66). Die Bedeutung dieser Anerkennung fällt in dasgleiche Jahr wie die Publikation „On the origin of thespecies by means of natural selection“ von CharlesDarwin.
In den Jahren 1860 bis 1864 führten der FranzoseÉdouard Lartet und der Brite Henri Christy zahlreicheAusgrabungen in Frankreich, darunter in Aurignac,Laugerie-Basse und Laugerie-Haute, La Madeleineund Le Moustier durch. Mit den neugewonnenenDaten und durch den Vergleich der Steinartefakte imSinne einer Typologie konnte das erste chronologi-sche System für das Paläolithikum entwickelt wer-den13. Ihre Publikation „Reliquiae Aquitanicae“, diein London in der Zeit von 1865 bis 1875 publiziertwurde, gilt als Meilenstein in der damaligen urge-schichtlichen Diskussion. Gabriel de Mortillet grün-dete 1864 die „Materiaux pour l’histoire positive etphilosophique de l’homme“, die als die erste derurgeschichtlichen Forschung gewidmete Zeitschriftgilt. Die Entdeckung zahlreicher paläolithischerFunde, sowie Menschenreste auch außerhalb Frank-reichs, führte ihn zu der Überzeugung, dass es an derZeit wäre, einen internationalen Kongress einzu-berufen. So wurde der „Congrès PaléoethnologiqueInternational“ 1865 in La Spezia (Italien) beschlos-sen, der 1866 in Neuchâtel (Schweiz) stattfand. Diesergilt als der erste internationale urgeschichtliche Kon-
7 Bei vielen altgegrabenen Fundstellen ist die Zugehörigkeit der Knochenfunde zu bestimmten Schichten unklar.Auch ist das fast vollständige Fehlen von Daten über Knochen von Vögeln oder Kleinsäugern charakteristisch.Dazu kommt, dass bei alten Grabungen von Archäologen oft nur leicht bestimmbare Knochen aufgesammelt wur-den, Bruchstücke wurden schon bei der Grabung ausgelesen. 8 Teil der Ethnologie, welcher sich mit der Rolle des Tieres in alten Kulturen beschäftigt. 9 Die Anthropozoologie hat als Ziel, die Beziehung zwischen Mensch und Tier zu untersuchen. Ein Zweig, dienaturhistorische Athropozoologie, beschäftigt sich mit dem Menschen als Tier, die kulturelle Anthropozoologiebeschäftigt sich mit der Geschichte der Integration der Tiere in die Gesellschaft. Dabei deckt sie sich größtenteilsmit den Zielen der Palethnozoologie.10 Im engeren Sinne ist damit die Anwendung von osteologischen und osteometrischen Methoden an Knochenund Zähnen aus archäologischen Fundstellen gemeint.11 Hier trennte er als Erster zwischen dem Neolithikum, das er als keltisch anspricht, und dem Paläolithikum, daser als vorsintflutlich anspricht.12 In Saint Acheul wurden massenweise Faustkeile, etwa 800 jährlich, zutage gefördert.13 Wie schon erwähnt, die Perioden des großen Höhlenbären, des Mammuts, des Rentiers und des Auerochsen.

8 Verbreitung der großen Jagdfauna in Mittel und Westeuropa im oberen Jungpleistozän
gress. Zum zweiten Mal fand er 1867 in Paris statt(gleichzeitig mit der Weltausstellung); der Namewurde in „Congrès International d’Anthropologie etArchéologie Préhistoriques“ umgewandelt. DieseKongresse, deren Nachfolger der Kongress der „UnionInternationale des Sciences Préhistoriques et Protohis-toriques“ ist, führten zu einer weiten Verbreitung derUrgeschichte als wissenschaft (Nenquin 1994). DieZahl an Publikationen und das Interesse für diese neueDisziplin sowie die Anzahl an Ausgrabungen undAusstellungen nahmen stark zu. Gabriel de Mortilletselbst wurde 1867 zum Direktor des Museums in SaintGermain bei Paris und Professor an der „Écoled’Anthropologie“ ernannt, wo er bis zu seinem Tod1898 tätig war (Groenen 1994, 459). Ihm ist weiterhinder Versuch einer Chronologie anhand von archäo-logischen Leittypen zu verdanken14. Émile Cartailhacfolgte 1867 Gabriel de Mortillet als Herausgeber derZeitschrift „Materiaux pour l’histoire positive etphilosophique de l’homme“ und blieb bis 1889 in die-ser Funktion. Er reiste und forschte in Nordafrika,Griechenland, Italien (Grimaldi-Höhlen), Russlandund Spanien. Im Jahr 1882 wurde er Professor in Tou-louse (Groenen 1994, 425).
Die Entdeckung der Gravuren der Höhle vonChabot durch Léopold Chiron im Jahr 1878 und zweiJahre später die Publikation der Entdeckungen inAltamira bei Santander in Spanien durch Marcelino deSautuola erregten bei den Gelehrten kaum Aufmerk-samkeit. Gabriel de Mortillet und Émile Cartailhacsprachen sich entschieden gegen eine Anerkennungvon solchen Darstellungen aus. Eine Diskussion fandkaum statt. Erst nach der Entdeckung vonWandbildern in den Höhlen von La Mouthe (1895),von Marsoulas (1897), in der Höhle von Pair-non-Pair1896 durch François Daleau, und zwar an Stellen, dievorher vollkommen von den Kulturschichten desoberen Aurignacien bedeckt waren, sowie in LesCombarelles (1901) und Font-de-Gaume (1901) gabÉmile Cartailhac seine ablehnende Haltung gegenüberder Wandkunst mit dem selbstkritischen Artikel „Lescavernes ornées de dessins. La grotte d’Altamira,Espagne. ‚Mea culpa’ d’un sceptique“ auf. Ab 1894begannen Louis Capitan und Denis Peyrony damit,zahlreiche Fundstellen der Dordogne (u. a. Laugerie-Haute, La Ferrassie und La Madeleine) auszugrabenund zu publizieren (u. a. Capitan und Peyrony 1912).Louis Capitan folgte 1898 Gabriel de Mortillet alsProfessor an der „École d’Anthropologie“ in Paris. Ab1901 arbeitete mit ihnen auch der Abbé Henri Breuil.Zusammen entdeckten sie die Wandkunst von LesCombarelles (1901), von Font-de-Gaume (1901),Bernifal (1902) und Teyjat (1903). Breuil wurde in
wenigen Jahren der bedeutendste Urgeschichtlerseiner Zeit. Er revidierte 1906 die chronologischeAbfolge von Gabriel de Mortillet und stellte dasAurignacien vor das Solutréen (Breuil 1906). Ab 1910lehrte er „ethnographie préhistorique“ am „Institut dePaléontologie Humaine“ in Paris. In den folgendenJahrzehnten gab es kaum eine Entdeckung, insbe-sondere bei der Wandkunst, an der Breuil nichtbeteiligt gewesen wäre. Sein Werk besteht aus mehrals 500 Publikationen. Um so wichtiger erscheintheute seine Arbeit, wenn man bedenkt, dass für vieleFundstellen seine Zeichnungen, trotz aller Kritik(siehe Bahn und Vertut 1988, 43; Bégouën und Clottes1987), immer noch die einzigen Dokumente sind.
Während des Zweiten Weltkrieges musste der„conservateur adjoint“ des Museums Guimet vonParis, Philippe Stern, seinen Platz verlassen, um derVerfolgung durch die Nationalsozialisten zuentkommen. An seiner Stelle wurde der jungeEthnologe André Leroi-Gourhan berufen, der aufdiesem Posten bis 1943 tätig blieb. Ab 1946 arbeiteteer für das „Musée de l’Homme“ als stellvertretenderDirektor (Gaucher 1987). Aufgrund seiner breitenKenntnisse und zahlreichen Publikationen in Anthro-pologie, Ethnologie und Urgeschichte, stieg seineAnerkennung. Er wurde Mitglied oder Direktor inzahlreichen Forschungszentren, Kongressen undZeitschriften. Mit der Unterstützung von Henri Breuilwidmete er sich der Erforschung der paläolithischenKunst. 1965 erschien die Monographie „Préhistoire del’Art occidentale“, in der er eine Unterteilung derjungpaläolithischen Kunst in vier Stile mit chronologi-scher Abfolge vorschlug. Weiterhin entwickelte er dieTheorie eines starken Dualismus, in dem einesexuelle, teilweise versteckte Komponente insbe-sondere hinter den Darstellungen von Steppenbisonund Pferd anzunehmen sei. Diese Theorie wurdejedoch sehr stark kritisiert (Laming-Emperaire 1972;Lorblanchet 1995, 92). 1964 fand unter seiner Leitungdie erste Grabungskampagne in der FundstellePincevent bei Paris statt, in der er die flächigeGrabungstechnik mit hervorragenden Ergebnissenanwenden konnte (Leroi-Gourhan und Brezillon1983).
Eine prominente Figur der französischenurgeschichtlichen Forschung war ohne Zweifel auchFrançois Bordes. Als ausgebildeter Geologe, wurde er1956 Professor am „Institut de Préhistoire“ inBordeaux“ (ab 1969 „Institut du Quaternaire“), wo erdie Verbindung der „klassischen“ Urgeschichte mitnaturwissenschaftlichen Fächern wie Sedimentologie,Paläontologe und Palynologie konsequent verfolgte.Sein Hauptinteresse galt der Erforschung des Alt- und
14 Er unterschied 1872 die Technokomplexe Chélléen, Acheuléen, Moustérien, Solutréen, Magdalénien. DieIndustrie von Aurignac, welche er 1869 als eigenen Technokomplex anerkannt hatte, erschien in seiner Abfolgenun nicht mehr (Mortillet 1872).

Forschungsgeschichte in Mittel- und Westeuropa (Schwerpunkt Jungpaläolithikum) 9
Mittelpaläolithikums. Neben zahlreichen Ausgra-bungen und Publikationen ist seine typologischeEinordnung des Alt- und Mittelpaläolithikums zuerwähnen, die einen Meilenstein für die statistischeTypologie darstellt (Jelinek 1982). Der Versuch, eineähnlich geordnete Typologie auch für das Jungpaläoli-thikum zu erstellen, wurde von Denise de Sonneville-Bordes und Jean Perrot realisiert (u. a. Sonneville-Bordes und Perrot 1954).
Seit den 1970er Jahren, besonders in den 1980er und1990er Jahren, nahm in Frankreich wie auch anderswoin Europa die Spezialisierung verstärkt zu. Es wurdenauf dem Gebiet der 14C-Datierung, der Pollenanalyse,der Archäobotanik und der Archäozoologie vieleSpezialisten ausgebildet. Hier sei auf Forscher wieFrançoise Delpech und François Poplin verwiesen. ImRahmen der „klassischen“ Urgeschichte nahm imLaufe der Jahre die Erforschung der technischenVorgänge, die „Chaîne opératoire“, durch die Arbeitenvon André Leroi-Gourhan und noch mehr von JacquesTixier, ständig an Bedeutung zu.
Noch wichtiger als einzelne Forscher ist in Frank-reich in dieser Zeit, neben den zahlreichen Institutenan den Universitäten, die Gründung von reinen For-schungszentren, wie des CNRS, „Centre National deRecherche Scientifique“, oder der UMR, „Unité Mixtede Recherche“ zu erwähnen.
Nach zahlreichen Diskussionen mit französischenFachkollegen und nach der Durchforstung der Listender Mitarbeiter der unterschiedlichen Institutionen inFrankreich scheint die Zahl von derzeit etwa 150 bis250 Planstellen im Bereich der älteren Urgeschichteplausibel. Ein Teil der Stellen beim CNRS sind zwarkeine offiziellen Planstellen, es ist jedoch für einenWissenschaftler, der eine solche Stelle bekommen hatund den Erwartungen entspricht, sehr schwierig, siewieder zu verlieren.
Weiterhin haben die lange Forschungstradition, diehäufigen Entdeckungen von wichtigen und zum Teilspektakulären Fundstellen, die zahlreichen Publi-kationsorgane, Ausstellungen und Kongresse und dieTatsache, dass Französisch als internationale Spracheim Bereich der älteren Urgeschichte und insbesonderedes Jungpaläolithikums eine breite Akzeptanz ge-nießt, dazu geführt, dass die französische Forschungeine mehr oder weniger zentrale Rolle im Rahmen derpaläolithischen Forschung einnimmt. In vielen Län-dern, wie z. B. Italien, Spanien, Deutschland, richtetsich der Blick der Forscher sehr aufmerksam auf dieEntwicklungen in Frankreich. Dem stehen jedoch einhäufig eher mäßiges Interesse oder mäßige Kenntnis-se der französischen Wissenschaftler für das, was au-ßerhalb ihres Landes passiert, gegenüber. Hier, um dieProblematik schon jetzt zu verdeutlichen, ein Zitat vonYves Coppens aus dem Vorwort zur sehr interessan-ten Arbeit von Jean-Pierre Duhard „Réalisme de
l’image masculine paléolithique“ (1996, 5): „ ... lecorpus des représentations masculines embrasse ... unespace grand comme l’Europe, ... de l’Atlantique àl’Oural“ („die Sammlung der männlichen Darstellun-gen erstreckt sich .. über eine Region so groß wie Eu-ropa, ... vom Atlantik bis zum Ural“; eigeneÜbersetzung). Tatsächlich werden jedoch nur diefranzösischen Darstellungen behandelt. Das gleichegilt für den Artikel desselben Autors „Les humains it-hyphalliques dans l’art paléolithique“ (1992). Die seitlangem bekannten, eindeutigen Darstellungen vonAltamira (Spanien) oder Addaura (Italien) werdennicht einmal erwähnt. Dieses Problem ist keineswegsneu. Schon 1877 sprach Paolo Mantegazza bei der Re-zension einer anthropologischen Arbeit von Quatrefa-ges dieses Thema mit deutlichen Worten an: „Imgesamten Buch sticht ein Grundfehler hervor, und die-ses ist die Missachtung für alles, was nicht in Frank-reich gemacht wurde. ... Und hat man in Italienwirklich nichts im Bereich der Kraniologie und Ethno-logie gemacht? Wenn das Schweigen von Quatrefa-ges Verachtung ist – so erinnern wir ihn hier daran,dass Wissenschaft etwas kosmopolitisches ist. Ist esIgnoranz, so sagen wir ihm, dass dies, bei einem soschnellen Fluss von Gedanken von Volk zu Volk, heu-te unverzeihlich ist“ (eigene Übersetzung aus Mante-gazza 1877, 27f).
Unter den vielen forschenden Institutionen Frank-reichs seien hier in Kürze das „Laboratoire d’Ethnolo-gie Préhistorique“ der Universität Paris I, Panthéon–Sorbonne, das „Laboratoire de Préhistoire et Techno-logie“ der Universität Paris X–Nanterre, das „Institutdu Quaternaire“ in Bordeaux und das „Centre euro-péen des recherches préhistoriques“ in Tautavel (Tal-teüll) erwähnt.
Unter den zahlreichen Zeitschriften mit paläolithi-schem Schwerpunkt sind „L’Anthropologie“ (gegrün-det 1890, herausgegeben vom „Institut de Paléon-tologie Humaine“, Paris), das „Bullettin de la SociétéPréhistorique Française“ (gegründet 1904, Paris),„Gallia Préhistoire“ (gegründet 1958, herausgegebenvom CNRS, Nanterre), „Bulletin de la Société Préhi-storique de l'Ariège“ (gegründet 1946) und „Paléo“(gegründet 1989, herausgegeben von der „Société desAmis du Musée National de Préhistoire et de laRecherche Archéologique, Les Eyzies“) unbedingt zunennen. Als französischsprachige Zeitschrift ist in die-ser Gruppe ebenfalls das „Bulletin du Muséed’Anthropologie Préhistorique de Monaco“ (gegrün-det 1954) zu erwähnen.
Weiterhin genießt das Paläolithikum in Frankreicheine große Anerkennung in der Öffentlichkeit. Diebemalten Höhlen beispielsweise bilden die wichtigstetouristische Attraktion in der Region des Périgord. Alldies ist aber auch das Ergebnis von vielen populärwis-senschaftlichen Publikationen, unter deren Autorenviele renommierte Professoren zu zählen sind.

10 Verbreitung der großen Jagdfauna in Mittel und Westeuropa im oberen Jungpleistozän
Im Ausland ist Frankreich sowohl in Osteuropa alsauch in Afrika präsent. Im Bezug auf Osteuropa musshier erwähnt werden, dass dort bis zum Zusammen-bruch der Sowjetunion die allgemeine Akzeptanz vonFranzösisch als wissenschaftlicher Fremdsprachedeutlich höher war als die von Englisch. In Afrika hatnatürlich Frankreich traditionell enge Verbindungenzu seinen ehemaligen Kolonien, insbesondere zuTunesien, Algerien und Marokko.
Spanien
Vorwiegend verwendete Literatur: Altuna 1993; Canalund Carbonell 1989; Estévez und Vila 1999; Straus1992.
Gerade die Geschichte der Entdeckung von Altamira,der bekanntesten paläolithischen Höhle Spaniens, istbeispielhaft für die Anfänge der Forschungsge-schichte auf der Iberischen Halbinsel. 1868 durchZufall von einem Jäger entdeckt (Cartailhac undBreuil 1906, 2), wurde sie ab 1875 wiederholt vonMarcelino Sanz de Sautuola untersucht. Dabei fielenihm einige schwarze Zeichnungen auf (Madariaga1976, 90–91). Aber erst nach dem Besuch derWeltausstellung in Paris 1878, wo unterschiedlichevorgeschichtliche Sammlungen vorgestellt wurden,schien ihm klar zu sein, dass auch um SantanderSpuren aus der Urgeschichte vorhanden sein könnten(Madariaga 1976, 24). Im folgenden Jahr entdeckte eroder genauer seine Tochter die polychromen Steppen-bisondarstellungen und er publizierte kurz danachseine „Breves apuntes sobre algunos objetos prehi-stóricos de la provincia de Santander“ (1880). Obwohler durch seine Publikation eine offene Diskussionanstrebte, stieß er auf die Ablehnung der damaligenFachwelt, personifiziert durch Gabriel de Mortilletund Émile Cartailhac. Letzterer bekannte, wie schonerwähnt, seinen Fehler öffentlich erst 1902 (Cartailhac1902).
Unter den ältesten urgeschichtlichen Ausgra-bungen ist ferner auf der Mittelmeerseite die Grabungvon 1871 von Pere Alsius i Torrent in der Höhle derBora Grand (bei Girona, Katalonien) zu erwähnen.Hier wurden Harpunen, Steinwerkzeuge und Kno-chenreste ausgegraben (Canal und Carbonell 1989,38). In den folgenden Jahren wurden in diesem Gebietweitere Ausgrabungen durchgeführt und Ent-deckungen gemacht. Darunter ist der Fund des Unter-kiefers von Banyoles 1887 zu erwähnen, derheutzutage um ca. 45.000 Jahren BP datiert wird (Juliáund Bischoff 1993) und damals große Aufmerk-samkeit im Bereich der Paläoanthropologie erweckte.
Am Anfang des 20. Jahrhunderts wird die Höhlen-kunst zum Schwerpunkt der Forschung. In Kantabrienfanden planmäßige Untersuchungen statt, dabeiwurden zahlreiche bemalte Höhlen entdeckt, u. a. El
Castillo, Hornos de la Peña, Covalanas, El Pendo undLa Loja. In Südspanien wurden bedeutende Malereienin der Höhle von Los Ardales entdeckt. Die ersteErforschung all dieser Höhlen ist vor allem HermilioAlcalde del Rio, Émile Cartailhac, Henri Breuil undHugo Obermaier zu verdanken. Besondere Erwähnungverdienen hier auch die Ausgrabungen der Höhle vonEl Castillo in den Jahren 1910–1914, wo man eine fürdas Mittel- und Jungpaläolithikum besonders reicheSchichtenabfolge aufdecken konnte (Acheuléen,Moustérien, Aurignacien, Gravettien, Solutréen,frühes Magdalénien, spätes Magdalénien und Azilien).
Zwischen 1912 und 1922 ist weiterhin alswichtiger Forscher der Conde de la Vega del Sella zuerwähnen, welcher in den Höhlen von La Riera, Cuetode la Mina und Morín ausgrub.
In den 1920er und 1930er Jahren nahm imBaskenland insbesondere die Arbeit des Priesters JoséMiguel Barandiarán zu, der bis zum Beginn desBürgerkriegs (1936) zusammen mit Telesforo deAranzadi und Enrique de Eguren und später nach derZeit seiner Verbannung (1936–1953) allein dieForschung in Kantabrien stark prägte. Unter denvielen seiner wissenschaftlichen Ausgrabungen seienhier Santimamiñe, Bolinkoba, Urtiaga, Ermittia undLezetxiki sowie die Arbeiten über die Malereien derHöhlen Altxerri und Ekain erwähnt (Altuna 1990).Am Mittelmeer ist die Ausgrabung der Höhle vonParpalló (1929-1930) durch Luis Pericot García mitihren unzähligen gravierten Steinplättchen sicher eineder wichtigsten Fundstellen mit Kleinkunst überhaupt(Villaverde 1994).
Die Zeit des Bürgerkrieges und die Jahre danachwaren in Spanien nicht nur für die Bevölkerung,sondern auch für die urgeschichtliche Forschung eintrauriges Kapitel (siehe das Kapitel 3 „La Epocaoscura 1936/9–1960“ von Estévez und Vila 1999, 61–110). Manche Forscher starben während des Krieges,andere, wie José Miguel Barandiarán und Pere BoschGimpera, mussten ins Exil. Nur diejenigen, die dienationale und katholische Ideologie uneingeschränktvertraten, konnten weiter als Archäologen in leitendenPositionen arbeiten. Spanien wurde in dieser Zeitkaum von ausländischen Forschern besucht. Erst abder Mitte der 1950er Jahre fand, durch die Rückkehrvon José Miguel Barandiarán, die Arbeiten vonJoaquín González Echegaray in El Juyo, La Chora, ElOtero und von Francisco Jordá Cerdá über dasSolutréen Spaniens und über die Ausgrabungen in LaLloseta, eine Wiederbelebung der urgeschichtlichenForschung statt. 1961 wurde das „Departamento dePrehistoria de la Sociedad de Ciencias Aranzadi“gegründet. Ausländische Forscher arbeiteten wieder inKantabrien. Darunter muss die Person von Leslie G.Freeman, Professor an der „University of Chicago“,hervorgehoben werden, da ein großer Teil seiner

Forschungsgeschichte in Mittel- und Westeuropa (Schwerpunkt Jungpaläolithikum) 11
Forschung jahrzehntelang mit Spanien verbunden war.In Südspanien verdient die Entdeckung der bemaltenHöhle von Nerja 1959 erwähnt zu werden.
Die 1960er und 1970er Jahre bis zum Ende derDiktatur sind in der Politik durch langsame,vorsichtige Öffnung nach Europa charakterisiert.Auch in der Archäologie spiegelt sich dieses Bildwider.
Die Situation änderte sich ab dem Ende der 1970erJahre. Durch die wiedergewonnene Demokratie, dieÖffnung zum Tourismus und den Beitritt zur Europäi-schen Gemeinschaft verbesserte sich der Lebens-standard in wenigen Jahren enorm. Gleichzeitig stiegauch die Zahl der Wissenschaftler und der wissen-schaftlichen Institutionen erheblich. Geländearbeitwurde nun auch in bis dahin fast fundleeren Regionendurchgeführt und zwar mit sehr positiven Ergeb-nissen. Ausländische Forscher interessierten sichvermehrt für dieses Land. Darunter ist Lawrence GuyStraus von der „University of New Mexico ofAlbuquerque“, USA, zu erwähnen, der zusammen mitspanischen Kollegen die Bearbeitung zahlreicherpaläolithischer Fundstellen insbesondere in Kanta-brien vorangetrieben hat.
Die Entfaltung von lokaler Autonomie, die allge-meine Stärkung der Regionen und die damitverbundene Dezentralisierung führte dazu, dass dieserProzess bis heute noch nicht abgeschlossen ist. Aberschon jetzt gibt es in Spanien, obwohl dieses Landökonomisch noch immer unter dem Durchschnitt derEuropäischen Union liegt, etwa 80 bis 100 Stellen imBereich der älteren Urgeschichte. Viele der Profes-soren sind zwar Experten für die ältere Steinzeit, siesind aber gleichzeitig in der Lehre und Forschungauch für jüngere Epochen zuständig.
Es wäre schwer hier alle in Spanien forschendenUniversitäten mit einem Schwerpunkt in der älterenUrgeschichte zu erwähnen, allein in Katalonien wärenes z. B. vier (Girona, „Autonoma“ und „Central“ inBarcelona, Tarragona). Ferner ist die Struktur derInstitutionen noch nicht gefestigt, so dass fast jedesJahr neue Institutionen entstehen oder alte umstruktu-riert werden. Daher werde ich hier nur einige derWichtigsten erwähnen können: Die „Universidad deCantabria“ in Santander (von der unter der Leitungvon Alfonso Moure Romanillo viele Ausgrabungenund Forschungsprojekte im kantabrischen Raumorganisiert worden sind), die Universitäten vonAlcalá, Oviedo, Salamanca, Saragoza sowie die„Universidad a Distancia“ in Madrid (Universität fürFernstudien) und die „Universidad de Valencia“.
Für die Datenbasis der vorliegenden Arbeit ist diegeographische Lage der verschiedenen Universitätenin Spanien besonders relevant, da die meisten jungpa-läolithischen Fundstellen auf der Iberischen Halbinsellange Zeit in der Nähe der Universitäten oder dergrößeren Städte zu liegen schienen, während in den
ländlichen Regionen kaum Fundstellen bekannt waren(Jordá 1986, 51, 56, 63). Dieses Bild ändert sich abden 1990er Jahren, als u.a. die Entdeckungen vonzahlreichen Felsbildern in Siega Verde und DomingoGarcia in Spanien und Foz do Côa in Portugal bekanntwurden (Ripoll und Municio 1999, 17–18). Diesewenigen Fundstellen zeigen, welches große Potentialfür die Erforschung des Jungpaläolithikums in denländlichen Regionen Spaniens noch steckt.
Die Archäozoologie ist im Bereich der älterenUrgeschichte Spaniens sehr eng mit der Arbeit vonJesus Altuna verbunden. Seit dem Beginn der 1970erJahre haben er und seine Mitarbeiter die archäozoolo-gische Bestimmung der Faunen aus fast allenwichtigen kantabrischen Fundstellen durchgeführt.Weiterhin ist in diesem Bereich Arturo Morales zuerwähnen, Direktor des archäozoologischen Laborsder „Universidad Autonoma de Madrid“ und Heraus-geber der Zeitschrift „Archeofauna“. Zusammen mitseinen Mitarbeitern hat er zahlreiche Fundstellen ausganz Spanien untersucht und publiziert (u. a. aus Nerjaund Amalda).
Unter den bedeutenden spanischen Zeitschriftenmit einem Schwerpunkt in der älteren Urgeschichtesind „Munibe“ (gegründet 1971, herausgegeben vonder „Sociedad de Ciencias Aranzadi“, Donostia bzw.San Sebastian), „Trabajos de Prehistoria“ (gegründet1960, herausgegeben vom „Departamento de Prehi-storia des CSIC, „Consejo Superior de Investiga-ciones Científicas“, Madrid) und „Zephyrus“(gegründet 1950, herausgegeben von der Universitätvon Salamanca) zu erwähnen. Eine für die ältereUrgeschichte bedeutende Reihe an Monographienwird vom „Centro de Investigación y Museo deAltamira“ in Santander herausgegeben.
Wie in Frankreich hat das Paläolithikum auch inSpanien und insbesondere in Kantabrien eine langeForschungsgeschichte sowie sensationelle und zahl-reiche Fundstellen aufzuweisen. Es ist jedoch vonAnfang an und noch verstärkt während der Diktaturein Mangel an wissenschaftlicher Anerkennung insbe-sondere im Ausland zu vermerken. Die klareEntwicklung seit den 1980er Jahren in den Wissen-schaften allgemein und in der älteren Urgeschichteinsbesondere, sowie die zahlreichen Projekte, beidenen durch Museen, Ausstellungen und Publika-tionen die Aufmerksamkeit der breiten Öffentlichkeitgesucht wird, lassen eine weitere Entwicklung undFestigung der Forschung erwarten.
Ein Schwerpunkt für die spanische Forschung imAusland ist aus geschichtlichen und sprachlichenGründen Mittel- und Südamerika gewesen. Auchdieser Bereich befindet sich im Aufschwung.

12 Verbreitung der großen Jagdfauna in Mittel und Westeuropa im oberen Jungpleistozän
Portugal
Vorwiegend verwendete Literatur: Zilhão 1997.
Obwohl die erste urgeschichtliche Ausgrabung inPortugal in Casa da Moura schon 1865 durch NeryDelgado für die „Serviços Geológicos de Portugal“erfolgte, folgte mit wenigen Ausnahmen eine langeforschungsleere Zeit (Zilhão 1997, Vol. II, 11).
Erst in den 1930er und 1940er Jahren fandenzahlreiche Ausgrabungen auf Freilandstationen durchManuel Heleno, Direktor des „Museu Nacional deArqueologia e Etnologia“, statt. Leider publizierte erseine Befunde kaum. Ihm reichten oft die Einzelob-jekte allein, was einen beträchtlichen Verlust für diejungpaläolithische Forschung in Portugal bedeutete.Seiner Meinung nach waren die charakteristischenWerkzeugtypen der verschiedenen Technokomplexe(z. B. Kielkratzer, Gravettespitzen) als Leitfossilien zubetrachten und dem entsprechenden Technokomplexgleichzusetzen. So ordnete er Funde, die aus einerFundeinheit stammten, unterschiedlichen Technokom-plexen und Epochen zu.
Von Bedeutung ist weiterhin der Portugal-aufenthalt von Henri Breuil (1944), der, zusammenmit Georges Zbyszewski, bei der Untersuchung der im19. Jahrhundert ausgegrabenen Funde die Präsenz vonWerkzeugen aus dem Aurignacien, Gravettien,Solutréen und Magdalénien erkannte. Sie verwen-deten dabei konsequent die gleiche Abfolge anTechnokomplexen, wie sie im Périgord anhanddeutlich zahlreicherer Funde und Fundzusammen-hänge bekannt geworden war, für das gesamte Jungpa-läolithikum Portugals.
Von 1950 bis 1980 wirkte Abbé Juan Roche, dereine Kontinuität in der urgeschichtlichen Forschung inPortugal durch zahlreiche Artikel (oft in Zusammen-arbeit mit Forschern der Serviços Geológicos)sicherte. Auch er versuchte, einzelne Fundstellenanhand der Typologie an die bekannten Technokom-plexe anzuschließen.
Seit dem Ende der Diktatur in Portugal (1974)erlebte die Forschung wie in Spanien einen deutlichenWandel. Alte Strukturen und Institutionen wurdenerneuert, neue Institutionen wurden gegründet, und dieZahl der Planstellen für das Paläolithikum nahmschnell zu. In diesem Entwicklungskontext spielenu. a. das G.E.A.P. („Grupo de Estudios Arqueo-lógicos de Porto“) und das G.E.P.P. („Grupo para oEstudio do Paleolítico Português“) eine bedeutendeRolle, da sie das Interesse einer heranwachsendenneuen Generation an Forschern bündeln. Eineführende Rolle für die Erforschung des Jungpaläoli-thikums ist João Zilhão zuzuerkennen. Er untersuchteund revidierte die alten Sammlungen, führte zahl-reiche Ausgrabungen, oft in Zusammenarbeit mitausländischen Forschern (u. a. Anthony E. Marks von
der „Southern Methodist University of Dallas“, USAund Lawrence Guy Straus von der „University of NewMexico“, Albuquerque, USA), durch und – was nochwichtiger ist – er weckte das internationale Interessefür die Urgeschichte Portugals, das bis dahin de factonicht existent war. Zu diesem letzten Punkt sind die14C-Datierungen der Fundstellen mit Knochenrestenvom Neandertaler und die Gravierungen von Mazoucound Val do Côa hervorzuheben. Diese sind interna-tional besonders relevant und lassen ein starkesForschungspotential auch für die Zukunft erhoffen.
Unter den in Portugal forschenden Institutionenmit einer Tradition auch in der älteren Urgeschichtewurde schon auf die enge Verbindung mit derGeologie hingewiesen. Ferner sind das „MuseoNacional de Arqueologia e Etnologia“ (Herausgeberder Zeitschrift „O Arqueólogo Português, N. S.“,gegründet 1951, Nachfolger der Zeitschrift „OArqueólogo Português“ erschienen von 1895 bis1938) und das „Instituto Português de Arqueologia“(Herausgeber der Zeitschrift „Revista Portuguesa deArqueologia“, gegründet 1998) zu erwähnen, die imRahmen ihrer Tätigkeiten auch die Forschung imBereich der älteren Urgeschichte vorantreiben. Diemeisten älteren Publikationen sind in den Reihen derServiços Geológicos de Portugal erschienen.
Italien
Vorwiegend verwendete Literatur: Brizzi 1977; Palmadi Cesnola 1993.
Schon in der zweiten Hälfte des 16. Jahrhundertserkannte Michele Mercati (1544–1593), Archivar beiden Vatikanischen Museen, dass die Steinartefakte ausEuropa, die damals als „keraunia“ oder „ceraunias“bezeichnet (vom griechischen keraunós = Blitz, inDeutschland bekannt als „Donnerkeile“) und fürNaturprodukte gehalten wurden, eindeutig mit denSteinartefakten und Werkzeugen aus der „NeuenWelt“ vergleichbar waren. Er schloss daraus, dassauch die europäischen Funde genau so von Menschenproduziert worden sein mussten wie die anderen, dieaus bekannten, ethnologischen Zusammenhängenentnommen waren. Seine Arbeit wurde aber erst 1717,lange nach seinem Tod, unter dem Titel „Metallo-theca vaticana“ publiziert (Vigliardi 1979, 4). Dadurchgilt in Italien Mercati als Vorgänger der urgeschicht-lichen Forschung, und auch im Ausland gab sein Werkden frühesten Forschern wichtige Impulse (Cleyet-Merle 1990, 7).
Italien wurde jedoch von Künstlern und Literatenseit der Renaissance nicht wegen der prähistorischen,sondern wegen der klassischen archäologischen Fundeimmer wieder besucht. Es galt, die Reste des altenRom sowie die römischen Repliken griechischerStatuen zu bewundern. Unter dem prachtvollen

Forschungsgeschichte in Mittel- und Westeuropa (Schwerpunkt Jungpaläolithikum) 13
Schatten der klassischen Archäologie waren und sinddie Bedingungen für die Entwicklung einer urge-schichtlichen Archäologie damit grundlegend andersals beispielsweise in Frankreich oder in Mitteleuropa.
Als die erste Publikation über paläolithische Fundein Italien ist wahrscheinlich der Artikel von GiuseppeScarabelli, „Intorno alle armi di pietra dura che sonoraccolte nell’Imolese“ (Annuali di Scienze naturali diBologna, serie III, t. II, 1850) zu sehen (Brizzi 1977,39). Diesem folgten mehrere Artikel hauptsächlichsammlerischen Charakters, darunter auch der ersteArtikel von Luigi Pigorini (1867) über die vorge-schichtlichen Funde im noch nicht vereinigten Italien.Im Jahr 1871 fanden in Bologna gleichzeitig derfünfte internationale Kongress für prähistorischeAnthropologie und Archäologie und die ersteurgeschichtliche Ausstellung Italiens („Esposizioneitaliana di archeologia ed antropologia preistorica“)statt, in denen das junge Fach Urgeschichte sich Gehörverschaffen konnte. Im Jahr 1875 wurde der ersteLehrstuhl für Paläoethnologie in Rom eingerichtet,den Luigi Pigorini für etwa 40 Jahre besetzte, und eswurde das Prähistorische und EthnographischeMuseum in Rom (heute „Museo Luigi Pigorini“)eröffnet. Im gleichen Jahr wurde weiterhin vonGaetano Chierici, Luigi Pigorini und PellegrinoStrobel die Zeitschrift „Bullettino di PaletnologiaItaliana“ gegründet.
Um Luigi Pigorini und das EthnographischeMuseum bildete sich eine historisch-ethnographischeSchule, die sogenannte „scuola romana“. Diese sah dieprähistorische Forschung als einen Teil der Ethno-logie (Palma di Cesnola 1993, 11). Daher waren dieEthnologie und ihre Methoden die wichtigste Quellefür die heranwachsende neue Wissenschaft. DieseSchule dominierte das Ende des 19. Jahrhunderts.Aldobrandino Mochi (Professor für Anthropologie inFlorenz) und Gian Alberto Blanc setzten dagegen denAkzent eher auf die Naturwissenschaften, Umwelt-rekonstruktion und Sedimentanalysen. Im Jahr 1912gründen sie den „Comitato per le ricerche di Paleon-tologia Umana in Italia“, der im Jahr 1927 zu einemrichtigen Institut wurde, dem „Istituto Italiano diPaleontologia Umana“. Um diese Forschungsrichtungentstand die sogenannte „scuola fiorentina“. Bezeich-nend für diese Zeit ist die Diskussion über das italie-nische Jungpaläolithikum. Noch 1904 sprach sichLuigi Pigorini trotz der Ergebnisse der ersten Untersu-chungen in der Höhle von Romanelli in Apulien durchPaolo Emilio Stasi und Ettore Regalia gegen dieExistenz eines Jungpaläolithikums in Italien aus, dadieses seinem historisch-ethnographischen Modellwidersprach. Erst 1913 kann man durch den Vortragvon Aldobrandino Mochi „La fine del dibattitosull’esistenza in Italia del paleolitico superiore“ („DasEnde der Diskussion über die Existenz des Jungpaläo-lithikums in Italien“) dieses Kapitel als abgeschlossen
betrachten (Brizzi 1977, 43). Signifikant in diesemKontext ist weiterhin die Erforschung der Grimaldi-Höhlen in Ligurien. Die erste Grabung fand bereits1846 durch Prinz Florestan I von Monaco statt. Ersandte eine Kiste Knochen nach Paris zur Begut-achtung. Ob er bei diesen Grabungen die Steinarte-fakte als solche erkannte, ist ungewiss. Es folgtenzahlreiche Ausgrabungen, insbesondere in den 1870erund 1880er Jahren, bei denen, so wie damals überallüblich, eher die Objekte an sich eine Rolle spielten,selten die Zusammenhänge. So konnte beispielsweiseGabriel de Mortillet jahrelang die Meinung vertretenund verbreiten, dass die Bestattungen, die in denGrimaldi-Höhlen entdeckt wurden, nicht paläoli-thisch, sondern neolithisch wären. 1892 interessiertesich Prinz Albert I von Monaco für die Grimaldi-Höhlen und verpflichtete als Grabungsleiter MarcellinBoule, Émile Cartailhac und Louis de Villeneuve von1895 bis 1902. Erst 1928–29 und dann 1938 grub dasIstituto Italiano di Paleontologia Umana (unter derLeitung von Alberto Carlo Blanc, Luigi Cardini, PaoloGraziosi und Aldobrandino Mochi) in den Grimaldi-Höhlen.
Auch die erste wissenschaftliche Ausgrabung aneiner anderen bedeutenden Fundstelle in Süditalien,der Höhle von Romanelli (namensgebend für dasRomanelliano, ein Technokomplex des Endpaläolithi-kums Italiens), fand erst 1914 unter der Leitung vonGian Alberto Blanc statt.
Nach der Mitte der 1940er Jahre steigt die Zahl derpaläolithischen Forscher, der Publikationsorganesowie der Ausgrabungen so stark, dass es unmöglichist, einen umfassenden Überblick zu bieten. Daherkönnen hier nur einige der Bekanntesten erwähntwerden.
Paolo Graziosi gründete 1946 (und führte bis1988) die „Rivista di Scienze Preistoriche“, die späterdas offizielle Publikationsorgan des im Jahr 1954gegründeten „Istituto Italiano di Preistoria e Proto-storia“ mit Sitz in Florenz wurde. Ein Forschungs-schwerpunkt von Paolo Graziosi war die paläo-lithische Wandkunst. In diesem Bereich untersuchte erinsbesondere die Höhlen im Süden Italiens undversuchte durch zahlreiche Publikationen zu erreichen,dass auch das Ausland davon Kenntnis nahm. AlbertoCarlo Blanc grub 1936–1953 am Monte Circeo. Ihmsind ferner die Erforschung der Schädel von Homoerectus aus Saccopastore sowie die Ausgrabungen vonTorre in Pietra (1954–55) zu verdanken. AntonioMario Radmilli führte die erste Ausgrabung in derGrotta Polesini bei Rom im Jahr 1953 sowiezahlreiche Untersuchungen in den Abruzzen durch.Arturo Palma di Cesnola untersuchte in den Jahren1961 und 1963 bis 1966 die Fundstelle der Grotta delCavallo in Apulien, wo er das Uluzziano charakteri-sieren konnte. Später übernahm er die Grabungen inder Höhle von Paglicci. Neben Apulien lag bei ihm

14 Verbreitung der großen Jagdfauna in Mittel und Westeuropa im oberen Jungpleistozän
ein weiterer Schwerpunkt in der Toskana, wo er an derUniversität von Siena seit 1966 Professor ist. Nordost-italien ist traditionell das Forschungsgebiet derUniversität von Ferrara. Hier konnte Alberto Brogliodie wichtigen Fundstellen von Riparo Tagliente undGrotta Fumane ausgraben und untersuchen. DieseGrabungen sind heute noch nicht abgeschlossen. Ander gleichen Universität ist auch der Paläontologe undArchäozoologe Benedetto Sala tätig.
Für die italienische Urgeschichte sind GeorgesLaplace und sein Aufnahmesystem besonders wichtig,da es die meisten Forscher in Italien verwendet haben(Gioia 1988, 71; Palma di Cesnola 1993, 21).
Zwei Meilensteine in der ForschungsgeschichteItaliens, die es verdienen erwähnt zu werden, sind diezwei Kongresse des UISPP („Union Internationale desSciences Préhistoriques et Protohistoriques“), die inden Jahren 1962 in Rom (Präsidenten Alberto CarloBlanc und Massimo Pallottino) und 1996 in Forlí(Präsident Antonio Mario Radimilli, GeneralsekretärCarlo Peretto) stattfanden. Beide führten zu einerbedeutenden Steigerung des Interesses, der wissen-schaftlichen Diskussion und der Erforschung desEiszeitalters in Italien.
Auch in Italien beginnt sich die Archäozoologieals eigenes Fach mit eigenen Schwerpunkten zuentwickeln. Der erste „Convegno Nazionale diArcheozoologia“ fand im Jahr 1993 in Rovigo statt(Padusa Quaderni n°1). Unter den in Italienforschenden Archäozoologen seien hier BenedettoSala (Universität von Ferrara) und Antonio Taglia-cozzo (“Museo Luigi Pigorini”, Roma) stellvertretendfür die vielen anderen erwähnt.
Neben dem „Bullettino di Paletnologia Italiana”(gegründet 1875, herausgegeben vom „Museo LuigiPigorini“, Rom) und der „Rivista di Scienze Preisto-riche” (gegründet 1946, herausgegeben vom „IstitutoItaliano di Preistoria e Protostoria”, Florenz), sind alsfür die ältere Urgeschichte besonders relevanteZeitschriften noch Quaternaria (1954–1981) undQuaternaria Nova (gegründet 1991, herausgegebenvom „Istituto di Paleontologia Umana”, Rom) sowie„Origini” (gegründet 1954, herausgegeben vom„Museo delle Origini”, „Universitá degli Studi LaSapienza”, Rom) hervorzuheben.
Neben dem „Istituto Italiano di Preistoria e Proto-storia“ in Florenz und dem „Istituto di PaleontologiaUmana“ in Rom sind als Institutionen mit einerTradition in der Erforschung der älteren Urgeschichtedie Universitäten von Bari, Ferrara, Florenz, Pisa,Rom („La Sapienza“) und Siena zu nennen. Für ganzItalien kann man die Zahl der festen Stellen imBereich der älteren Urgeschichte mit ca. 50 bis 80angeben.
Wie schon erwähnt, darf man nicht vergessen, dass inItalien die paläolithische Archäologie im Schatten derKlassischen Archäologie stand und zum Teil nochimmer steht. Das betrifft nicht nur die Mittel, die fürdie Forschung in Italien zur Verfügung stehen,sondern auch das öffentliche und leider auch das inter-nationale wissenschaftliche Interesse. Während Italienin der Klassischen Archäologie eine zentrale Bedeu-tung hat, wird es im Bereich der älteren Urgeschichteinsbesondere von Wissenschaftlern aus Frankreich,England und Mitteleuropa oft als Randbereichbetrachtet. Dafür ist wahrscheinlich neben einergewissen Sprachbarriere auch die Tatsache verant-wortlich, dass das Jungpaläolithikum Italiens, mit demweitgehenden Fehlen an Technokomplexen desMagdaléniens und des Solutréens und der Präsenzeigener Technokomplexe mit eigener Problematik,wie des Uluzziano und des Epigravettiano, nicht ohneWeiteres mit Technokomplexen aus anderen TeilenEuropas verglichen werden kann. Weiterhin muss mandaran erinnern, dass Luigi Pigorini und vieleAnhänger der „scuola romana“ bis zum Beginn des20. Jahrhunderts sich entschieden gegen das wissen-schaftliche Interesse aus dem Ausland aussprachenund eine fast vollständige Autonomie in derEntwicklung der „italienischen“ Vorgeschichteverlangten (Palma di Cesnola 1993, 12). Weiterhinerschwert die Methode von Laplace den Vergleich mitdem typologischen Aufnahmensystem, das im RestEuropas angewendet wird (siehe Gioia 1988, 71). Dasalles ist dennoch keine Entschuldigung für vieleVeröffentlichungen, in denen italienische Fundstellenund Forscher fast vollständig ignoriert werden (z. B.Groenen 1994).
Deutschland
Vorwiegend verwendete Literatur: Bosinski 1982;Müller-Karpe 1966.
Wenn man ein Datum wählen sollte, um den Beginnder paläolithischen Forschung in Deutschland zufixieren, würden die meisten wahrscheinlich das Jahr1856 wählen. In diesem Jahr, als eine kleine Höhle imNeandertal bei Düsseldorf ausgeräumt wurde, kamendie Reste eines Menschen bis dahin unbekannter oderunerkannter Art ans Tageslicht. Es war dieEntdeckung des Neandertalers durch Johann CarlFuhlrott. Die Existenz dieser Art wurde lange Zeitbestritten, insbesondere durch den berühmtenAnthropologen Rudolf Virchow, der die Knocheneinem „rachitischen Greis“ zuordnete.
Als die erste geplante altsteinzeitliche GrabungMitteleuropas gilt jedoch die Erforschung der Schus-senquelle bei Bad Schussenried durch den Geologenund Paläontologen Oscar Fraas im Jahre 1866. Dortwurden zum ersten Mal in Deutschland Steinwerk-

Forschungsgeschichte in Mittel- und Westeuropa (Schwerpunkt Jungpaläolithikum) 15
zeuge und eine gravierte Rentierdarstellung alsArtefakte erkannt. In Folge dieser Entdeckungkonnten Steinartefakte im gleichen Jahr auch imHohlenstein im Lonetal erkannt werden. Aufgrundseiner Stellung als Konservator und später alsVorsteher des Naturalienkabinetts (heute „StaatlichesMuseum für Naturkunde“) in Stuttgart konnte OscarFraas in den folgenden Jahren konsequent weitereGrabungen in Baden-Württemberg leiten, u. a. imHohle Fels bei Schelklingen (1870–71) und in derOfnethöhle im Nördlinger Ries (1875–76). SeinSchwerpunkt war allerdings die Paläontologie.
Im Rheinland fanden die ersten archäologischenGrabungen 1874 durch August von Cohausen in derWildscheuer und im Wildhaus bei Steeden an derLahn statt. Für diese Region ist ferner die Arbeit vonHermann Schaaffhausen zu erwähnen, der u. a. dieAusgrabung des Magdalénienfundplatzes am Hangdes Martinsbergs in Andernach (1883) leitete, einerStation, welche unter einer 4 m mächtigen Bimschichtlag.
Zahlreiche mehr oder weniger systematischeGrabungen fanden bis zum Ende des 19. Jahrhundertsstatt. Viele davon wurden leider nicht dokumentiert,bei anderen lag der Schwerpunkt eindeutig auf derPaläontologie, so dass die Funde nicht oder nurschwer einzelnen Schichten zuzuordnen sind. Einewesentliche qualitative Veränderung fand ab demBeginn des 20. Jahrhunderts statt. Die Ur- undFrühgeschichte setzte sich auch in Deutschland durchdie Einrichtung von Lehrstühlen als eigenständigeDisziplin durch. Robert Rudolf Schmidt promovierteals erster Urgeschichtler im Jahr 1907 an der Univer-sität Tübingen, wo er sich 1912 habilitierte. Ihm sindneben zahlreichen Ausgrabungen im Blautal beiSchelklingen und Blaubeuren (u. a. Schmiechenfels,Gansersfels, Sirgenstein) auch die Gründung desUrgeschichtlichen Forschungsinstituts an der Univer-sität Tübingen, dessen Direktor er von 1921 bis 1929war, zu verdanken. Zur gleichen Generation gehörtHugo Obermaier, der im Komplex der Klausenhöhlennoch vor 1914 tätig wurde, aber sonst kaum inDeutschland forschte. Er promovierte 1904 und habili-tierte sich 1908 in Wien. Sein Hauptinteresse widmeteer aber der Urgeschichte Frankreichs und Spaniens.Ab 1922 war er Professor für „Urgeschichte desMenschen“ an der „Universidad Central de Madrid“,ab 1924 besaß er die spanische Staatsbürgerschaft.
In den 1930er Jahren ist die Urgeschichte als Lehrfachetabliert, die Zahl der Forscher nahm stark zu. Hierkönnen nur einige unter den bekanntesten erwähntwerden. Gustav Riek, der ab 1935 den Lehrstuhl ander Universität Tübingen übernahm, grub zahlreicheFundstellen in Süddeutschland aus (u. a. Vogelherd,Große Grotte, Brillenhöhle). Alfred Rust, der durchdie Ausgrabungen von Meiendorf (1933 und 1934) bei
Hamburg, Stellmoor (1934–36) und Poggenwisch(1946–50) bei Ahrensburg die Kenntnisse über dieAhrensburger und Hamburger Kultur festigte, seiebenfalls genannt.
Durch die Teilung Deutschlands nach demZweiten Weltkrieg entstand auch eine Grenze in derForschung.
In den 50er und 60er Jahren wurde in der Bundes-republik Deutschland die Grabungstätigkeit gesetzlichgeregelt. Die Landesdenkmalämter wurden neugegründet oder umstrukturiert. Der Denkmalschutzerhielt ein viel größeres Gewicht. Die ältereUrgeschichte etablierte sich hauptsächlich an dreiUniversitäten: Erlangen (jetziger LehrstuhlinhaberLudwig Reisch) und Tübingen (jetziger Lehrstuhl-inhaber Nicholas John Conard), später auch Köln(jetzige urgeschichtliche Professur Jürgen Richter).Mit mehreren Ausnahmen kann man jedoch dieseInstitute über Jahre hinweg mit einer Forschungs-region oder mit einem Projekt identifizieren. So gräbtdie Universität Erlangen hauptsächlich in Bayern mitSchwerpunkt im Altmühltal und der Fränkischen Alb.Die Universität Köln gräbt hauptsächlich imMittelrheingebiet, wobei hier die Ausgrabungen vonGerhard Bosinski ab dem Ende der 1960er Jahre inGönnersdorf und deren Bearbeitung unbedingterwähnt werden müssen. Die Abteilung für ÄltereUrgeschichte und Quartärökologie des Instituts für Ur-und Frühgeschichte und Archäologie des Mittelaltersin Tübingen gräbt seit den 1970er Jahren auf derSchwäbischen Alb insbesondere in den HöhlenGeißenklösterle und Hohle Fels (Leitung von JoachimHahn bis 1996, seit 1997 von Nicholas John Conardund Hans-Peter Uerpmann). An diesem Institut istauch die einzige Professur für Archäobiologie(Professor Hans-Peter Uerpmann) in Deutschlandangesiedelt.
Im Jahr 2003 wurde wieder eine Professur mitSchwerpunkt ältere Urgeschichte an der UniversitätMainz errichtet und mit der Leitung des Forschungs-bereichs Altsteinzeit des Römisch-GermanischenZentralmuseums in Neuwied gekoppelt (jetzigeurgeschichtliche Professur Sabine Gaudzinski).
In der Deutschen Demokratischen Republiketablierte sich die urgeschichtliche Forschung eher inund um einige Museen (Halle, Potsdam, Schwerin,Weimar). Ferner waren die Mittel für die Forschung,verglichen mit der BRD, um ein Vielfaches geringer.Das führte dazu, dass nur besonders relevanteGrabungsprojekte fachmännisch untersucht wurdenund bis zu einer angemessenen Publikationsformkommen konnten. Darunter sind für das Mesoli-thikum die Grabungen von Ewald Schuldt (MuseumSchwerin) in Hohen Viecheln (1953–55) und vonBernhard Gramsch (Museum Potsdam) in Friesack (ab1977) hervorzuheben. Die Travertinfundstelle vonBilzingsleben (Ausgrabungen von Dietrich Mania seit

16 Verbreitung der großen Jagdfauna in Mittel und Westeuropa im oberen Jungpleistozän
1969) gehört aufgrund einiger Menschenreste sowiezahlreicher Befunde zu den wichtigsten altpaläolithi-schen Fundstellen Mitteleuropas. Viele andereFundstellen wurden von Laien, Lehrern, Schülern undzahlreichen ehrenamtlichen Mitarbeitern ausgegrabenund weiter gepflegt. Die „Tradition“, breite Schichtender Gesellschaft an die Archäologie zu binden,entsprach u. a. auch sozialistischen Idealen und wirdseit der Wiedervereinigung viel weniger gepflegt, sodass die Zahl der im Bereich der Archäologie tätigenehrenamtlichen Mitarbeiter stark gesunken ist.
Ältere Urgeschichte wurde an der Humboldt-Universität in Ost-Berlin (hier in engem Zusam-menhang mit der Anthropologie) und an der Martin-Luther-Universität Halle-Wittenberg in Halle gelehrt.
Nach der Wiedervereinigung wurde auch in denneuen Bundesländern das System der Landesdenkmal-ämter eingeführt. Die Grabungen im Stadtbereich unddamit die Mittelalterarchäologie erhielten aufgrundgroßflächiger Sanierungsaktionen oberste Priorität.Die ältere Urgeschichte wurde nur selten von dieserEntwicklung betroffen. Ein für die Urgeschichteaußerordentlich interessanter Bereich, insbesondere inOstdeutschland, ist dagegen der Braunkohletagebau.Da hier u. a. Schichten aus dem Quartär auf einerFläche von mehreren Quadratkilometern entferntwerden, ist die Chance relativ hoch, paläolithischeFunde zu entdecken.
In Jena wurde 2002 eine neue Professur (jetzigeurgeschichtliche Professur Clemens Pasda) mitSchwerpunkt Paläolithikum eingerichtet.
Die Präsenz im Ausland hat in Deutschland einelange Tradition und hat zu zahlreichen Forschungspro-jekten geführt. Darunter sei hier nur auf die berühmtenGrabungen in der Mumba-Höhle in Tansania durchMargit Kohl-Larsen (1938) und in Jabrud in Syriendurch Alfred Rust 1930–33 hingewiesen, denenzahlreiche andere im Bereich der älteren Urgeschichteund der jägerischen Archäologie folgten, wie z. B. inSpanien, Frankreich, Österreich, in der Türkei, Syrien,den Vereinigten Arabischen Emiraten, in Choukotka(Nordostsibirien), Ägypten, Namibia und Südafrika,um nur einige der Projekte allein aus der Abteilung fürÄltere Urgeschichte und Quartärökologie in Tübingenzu erwähnen. In manchen dieser Länder war dieUnterstützung des Deutschen ArchäologischenInstituts (DAI) besonders ausgeprägt, obwohl derInteressenschwerpunkt dieser Institution auf derKlassischen Archäologie liegt.
Schließlich soll hier noch die hohe Qualität sowohlder Gewinnung der Daten (z. B. genaue bis geradezupenible Grabungsmethode, sorgfältige Grabungs-dokumentation) als auch der Publikationen im Bereichder älteren Urgeschichte in Deutschland erwähnt wer-den. Diese hohe Qualität wird im Ausland durchausanerkannt und hoch geschätzt.
Unter den Publikationen, die in der älteren Ur-geschichte einen Schwerpunkt und einen überregio-nalen Blickwinkel haben, sind „Quartär“ (gegründet1938, herausgegeben von der „Hugo Obermaier-Gesellschaft“, Erlangen) und „Eiszeitalter undGegenwart“ (gegründet 1951, herausgegeben von der„Deutschen Quartärvereinigung e. V. Hannover“) zuerwähnen. In einigen anderen Zeitschriften, wie z. B.„Archäologisches Korrespondenzblatt“ (gegründet1971), „Jahrbuch des Römisch-Germanischen Zentral-museums Mainz“ (gegründet 1954, beide Organe des„Römisch-Germanischen Zentralmuseums Mainz“)sowie „Germania“ (gegründet 1917, herausgegebenvon der „Römisch-Germanischen Kommission desDeutschen Archäologischen Instituts“, Frankfurt amMain), nehmen paläo- und mesolithische Themen nureinen kleinen Anteil ein.
Schweiz
Vorwiegend verwendete Literatur: Le Tensorer 1998.
In der Schweiz ist die Erforschung der älterenUrgeschichte noch mehr als anderswo die Suche nachden Spuren der Eiszeit. Da der größte Teil derSchweiz im späten Pleistozän unter dem alpinenEisschild lag, waren es insbesondere Geologen, diezuerst die natürlichen Vorgänge der Vergletscherungverstehen wollten. Bernhard Friedrich Kühn warderjenige, der als erster (1787) die Existenz alterMoränen erkannte und diese in Zusammenhang miteiner viel breiteren Ausdehnung der Gletscher in derVergangenheit brachte (Hantke 1978, 28). Die weitereUntersuchung der Mechanismen der Entstehung undVorstöße der Gletscher sowie die Idee von mehrerenEiszeiten ist drei Schweizer Forschern zu verdanken,Jean de Charpentier, Ignaz Venetz und Louis Agassiz.Diese konnten anhand von erratischen Steinen undMoränen zwischen 1825 und 1840 die Ausdehnungder eiszeitlichen Gletscher aufnehmen und damit dieBasis für die Diskussion über die klimatischenSchwankungen während des Quartärs liefern.
Nach der Entdeckung von Pfahlbauten undzahlreichen prähistorischen Siedlungen in der zweitenHälfte des 19. Jahrhunderts am Zürich- und amBodensee konzentrierte sich fast das gesamte archäo-logische Interesse auf dieses Gebiet. Diese Art vonBefunden wurde auch im Ausland bald so bekannt,dass es kein Zufall ist, dass der erste „Congrès Inter-national de Paléoethnologie“ im Jahr 1866 inNeuchâtel stattfand. Im Schatten der Seeufersied-lungen wurde die Erforschung von Höhlen eher denPaläontologen überlassen, und so wurde auch dieAufsammlung von paläolithischen Steinwerkzeugen inder Freilandstation von Moosbühl ab 1860 von derFachwelt kaum zur Kenntnis genommen (Le Tensorer1998, 23). Als die erste paläolithische Ausgrabung in

Forschungsgeschichte in Mittel- und Westeuropa (Schwerpunkt Jungpaläolithikum) 17
der Schweiz galt daher lange Zeit die Entdeckung1868 des Abris du Scé (Villeneuve, Kanton vonVaud), in dem eine Schicht aus dem spätenMagdalénien durch Louis Taillefer und Henri deSaussure erkannt wurde.
In das Jahr 1873 fällt die Entdeckung der HöhleKesslerloch durch den Botaniker Konrad Merk, derein Jahr später die Entdeckung der Höhle vonFreudenthal und ferner vom Schweizersbild folgten.
Mit dem Beginn des 20. Jahrhunderts erscheinen inganz Europa die ersten großen Synthesen. Für dasPaläolithikum der Schweiz sind die zusammenfas-senden Arbeiten von Jakob Heierli (1901) undAlexander Schenk (1910) zu erwähnen. Im Jahr 1907wird die „Société Suisse de Préhistoire“ gegründet.
Eine Sensation war weiterhin die Entdeckung derWildkirchli-Höhle auf einer Höhe von 1500 m durchEmil Bächler im Jahr 1904. Durch die Entdeckungvon weiteren Höhlen bis ins Hochgebirge in derSchweiz und in Österreich (u. a. Drachenloch in2455 m Höhe) entwickelte sich das Konzept einesalpinen Paläolithikums, welches, charakterisiert durchdie Höhlenbärenjagd, bis in die Mitte des letztenJahrhunderts diskutiert wurde.
Durch einige Gesetze aus den 1960er Jahrenwurden so wie auch in Deutschland die archäo-logischen Tätigkeiten reglementiert. Dabei wurde einklarer Schwerpunkt auf den Denkmalschutz gesetzt,um einerseits die bekannten Fundstellen zu schützenund um andererseits die vorhandenen Mittel aufAusgrabungen im Bereich großer Bauvorhaben zukonzentrieren. In diesem Zusammenhang sind die1983 und 1989 entdeckten Magdalénien-FundstellenChampréveyres und Monruz zu erwähnen.
Die Universitäten, die in der älteren Urgeschichteeinen Schwerpunkt haben oder gehabt haben, sindBern und Basel.
Otto Tschumi, der schon seit 1911 am BernischenHistorischen Museum tätig war, wurde 1924 zumProfessor für Urgeschichte, römische Zeit und Mittel-alter an der Universität Bern ernannt. Da er aber auchfür weitere Aufgaben zuständig war, konnte er seineFunktion als Dozent de facto kaum wahrnehmen. Erstab 1950, mit der Berufung von Hans-Georg Bandi alsProfessor, fängt das Seminar für Urgeschichte an, sichrichtig zu entwickeln. Die ältere Urgeschichte nahmaber nach seiner Emeritierung in der Lehre und in dervon der Universität Bern ausgehenden Forschung anBedeutung deutlich ab.
Der einzige Professor für ältere Urgeschichte inder Schweiz ist derzeit Jean-Marie Le Tensorer, derseit 1981 am Institut de Préhistoire in Basel lehrt undforscht.
In der Schweiz gibt es keine auf die ältereUrgeschichte spezialisierte Zeitschrift, daher werdendie Beiträge oft in lokalen Organen publiziert. Unter
den überregionalen Zeitschriften ist das „Jahrbuch derSchweizerischen Gesellschaft für Ur- und Frühge-schichte“ aus Basel (gegründet 1908) zu erwähnen.
Niederlande, Belgien und Luxemburg
Vorwiegend verwendete Literatur: Groenen 1994;Otte 1979.
Als der „Vater der älteren Urgeschichte“ in dieserRegion gilt Philippe-Charles Schmerling, der ausHolland stammte und in Liège studierte. In derUmgebung von Liège untersuchte er zahlreicheHöhlen: u. a. Fonds-de-Forêt, Engis, Engihoul,Ramioul und Moha. Schon 1833 versuchte er, dieSilex-Artefakte nicht nur als von Menschen gemachteWerkzeuge, sondern auch in ihrer Funktion zuinterpretieren. Seine Ideen wurden in seiner Zeit aberkaum wahrgenommen und gerieten bald in Verges-senheit.
Erst im Jahr 1864, nach der Entdeckung von paläo-lithischen Fundstellen in Frankreich, beginnt mit derUnterstützung des Staates ein ganzes Programm anUntersuchungen an den Höhlen Belgiens. Mit dieserAufgabe wird der Geologe Edouard Dupont beauf-tragt. In den Jahren 1885–1886 graben Marcel dePuydt und Max Lohest in der Höhle von Spy. Dankdieser Ausgrabungen konnte zum Beginn des 20.Jahrhunderts Henri Breuil die Abfolge von Spy inseinem zeitlichen Gerüst für das Jungpaläolithikumverwenden. An dem zeitlichen Gerüst arbeitete in derselben Zeit auch der Geologe und Prähistoriker Aimé-Louis Rutot, der die Chronologie von Dupont weiterverfolgt und insbesondere mit den Daten Frankreichsverglich. In den 1920er und 1930er Jahren wurdenaltbekannte Höhlen (u. a. de la Princesse à Marche-les-Dames, Fonds de Forêt und Spy) weiter unter-sucht. Die Frage nach der Korrelation der belgischenund der französischen Abfolge konnte jedoch, trotzder Beteiligung von vielen Forschern insbesondere ausFrankreich, nicht geklärt werden. Dazu seien hier dieArbeiten von Denis Peyrony (1948), Henri Angelroth(1953), Henri Delporte (1956) und Denise de Sonne-ville-Bordes (1961) erwähnt.
Seit den 1970er Jahren spielt die Universität vonLiège („Service de Préhistoire“) und insbesondereProfessor Marcel Otte in der belgischen Urgeschichts-forschung eine herausragende Rolle. Ihm ist fürBelgien die Überarbeitung zahlreicher Fundstellen imInland und Ausland zu verdanken. Auf internationalerEbene sind neben zahlreichen Ausgrabungen imAusland auch wichtige Reihen an Publikationen zuerwähnen: die Zeitschrift „Préhistoire Européenne“15
(gegründet im Jahr 1992 und herausgegeben von derUniversität Liège) und die Publikationsreihe„ERAUL“ („Études et Recherches Archéologiques del’Université de Liège“, gegründet 1973). In diesem

18 Verbreitung der großen Jagdfauna in Mittel und Westeuropa im oberen Jungpleistozän
Zusammenhang ist es wichtig zu erwähnen, dass inbeiden Publikationsorganen zahlreiche Ausgrabungenund Berichte aus dem östlichen Mitteleuropa undOsteuropa publiziert wurden. Diese Publikationendienen explizit als wichtige wissenschaftliche Brückezwischen West- und Osteuropa.
In den Niederlanden lag und liegt der Schwer-punkt der urgeschichtlichen Forschung auf denjüngeren Perioden. Das Jungpaläolithikum istaufgrund der starken Sedimentation des Rheines, deshohen Grundwasserspiegels, der Seltenheit an Höhlenund der überwiegend niedrigen Topographie16 sehrselten. Es sei hier auf einige Fundstellen der Stiel-spitzen-Gruppen hingewiesen (Taute 1968, 150–154).Dem entsprechend existieren in den Niederlandenkeine Zeitschriften mit älterer Urgeschichte alsSchwerpunkt. Nur an der Universität Leiden, durchdie Arbeit von Wil Roebroeks und Elco Rensink, undan der Universität Groningen, durch Dick Stapert,wird ein Schwerpunkt auf die ältere Urgeschichtegesetzt.
Luxemburg hat zwar nie eine zentrale Rolle in derErforschung der älteren Urgeschichte gespielt, dieArbeiten und das Engagement insbesondere von PierreZiesaire seit den 1980er Jahren haben jedoch dieluxemburgischen Fundstellen weit über die Grenzendieses kleinen Landes bekannt gemacht. Weiterhin isthier Foni Le Brun-Ricalens zu erwähnen, Kustos am„Musée National d’Histoire et d’Art du Luxem-bourg“, der an zahlreichen Forschungsprojekten inFrankreich beteiligt ist.
Das „Bulletin de la Société Préhistorique Luxem-bourgeoise“ (erste Ausgabe im Jahr 1979), in dem fastdie Hälfte der Artikel sich auf die ältere Urgeschichtebeziehen, hat inzwischen einen deutlichen überregio-nalen Charakter erreicht (Ziesaire 2000).
Großbritannien
Vorwiegend verwendete Literatur: Barton 1997;Campbell 1977.
Der überwiegende Teil der britischen Inseln lagwährend der Eiszeit unter Gletschern, nur Südenglandund Südirland waren davon nicht bedeckt. Daher ist esverständlich, dass die Zahl der paläolithischen Fundein Großbritannien deutlich geringer ist als in anderenRegionen (Barton 1997). Das führte vielleicht auchdazu, dass, während man sich „auf dem Kontinent“stark auf die lokalen Besonderheiten der einzelnenTechnokomplexe fixierte, die Urgeschichte Europasaus dem Blickwinkel von Großbritannien als Teil der
globalen Urgeschichte erschien. Diese Perspektivekann man in der ganzen paläolithischen Forschung inGroßbritannien erkennen.
Der Begriff „Paläolithikum“ ist dem EngländerJohn Lubbock zu verdanken, der 1865 die bis dahinals Einheit betrachtete Periode der Steinzeit in einePhase der geschlagen (Paläolithikum) und eine Phaseder geschliffenen Steinwerkzeuge (Neolithikum)unterteilte. Außer ihm sind aus der Zeit der „Pioniere“der Urgeschichte die Geologen Charles Lyell undJoseph Prestwich und der Prähistoriker John Evans zuerwähnen, die der Interpretation des FranzosenJacques Boucher de Perthes und der Akzeptanz des„fossilen Menschen“ in den Jahren 1858–1860 zumDurchbruch verhalfen. Auch Charles Darwin verhalfdurch sein Werk „On the origin of the species bymeans of natural selection“ (1859) dazu, denMenschen als Teil der Natur und die Entwicklungaller Lebewesen als Evolution zu sehen, Konzepte, dieauch in der beginnenden Urgeschichtsforschung vonfundamentaler Bedeutung waren. Auch die Arbeit vonEdouard Lartet wäre ohne die Zusammenarbeit undUnterstützung des Bankiers Henry Christy nicht soerfolgreich gewesen.
Obwohl schon William Buckland (1823) in derPaviland Cave und John McEnery in der Kent’s Cave(1825–29) Ausgrabungen mit zahlreichen Fundendurchgeführt hatten, glaubten sie offensichtlich nichtan das damals unvorstellbare Alter der Funde, das diestratigraphischen Zusammenhänge eigentlich anzeig-ten. Daher kann man erst mit der Grabung 1859–1863im Wookey Hole durch William Boyd Dawkins undder unmittelbaren Publikation der dort geborgenenFunde (Stein- und Knochenartefakte sowie archäozoo-logische Reste) von einer wissenschaftliche Ausgra-bung im Bereich der älteren Urgeschichte sprechen.Von 1865 bis 1880 wurden die Grabungen in derKent’s Cave von William Pengelly geleitet, der schondamals mit Hilfe eines Gittersystems arbeitete und diePosition aller Knochen und Artefakte dreidimensionalaufnahm. Ferner ist für die Zeit der „Pioniere“Reverend John Magens Mello zu erwähnen, der durchseine Ausgrabungen an den Höhlen des Creswell Crag(1884–1879) den Technokomplex des Creswellienbeschreiben konnte.
Während in anderen Ländern das Paläolithikum zudiesem Zeitpunkt immer mehr an Bedeutung zunahm,gab es aus Großbritannien nur vereinzelte Ent-deckungen und Ausgrabungen, darunter 1903 dieEntdeckung eines menschlichen Skelettes in derGough’s Cave, das ins Spätpaläolithikum/Frühmesoli-thikum datiert wurde (Campbell 1977, 4). 1924 grubLeslie Armstrong im vorderen Bereich der schon von
15 Seit Dezember 2002 erscheint diese Zeitschrift als jeweils 5. Heft eines Jahrganges von „L’Anthropologie“,Paris. 16 Große Teile der Niederlande liegen unter dem Meeresspiegel, und Höhen von über 50 m sind sehr selten.

Forschungsgeschichte in Mittel- und Westeuropa (Schwerpunkt Jungpaläolithikum) 19
William Boyd Dawkins ausgegrabenen MotherGrundy’s Parlour. Ferner grub Leslie Armstrong auchim Pin Hole, R.F. Parry von 1927 bis 1931 in derGough’s Cave. Von Bedeutung ist in dieser Zeit auchdie „University of Bristol Spelaeological Society“, diein den Jahren 1919–21 im Aveline’s Hole, von 1925bis 1927 in der King Arthur’s Cave und von 1926 bis1928 im Sun Hole Ausgrabungen durchführte.
Durch den Zweiten Weltkrieg wurden viele Fundebei Bombenangriffen zerstört, so dass in den 1950erJahren, bei der Wiederbelebung der paläolithischenForschung in Großbritannien, viele der alten Fund-stellen neu untersucht wurden.
Eine Besonderheit der Forschung in Großbritan-nien sind die unterschiedlichen, oft lokal angelegten„Societies“, die als Vereine sehr aktiv sind.
In Großbritannien gibt es keine Zeitschrift, die sichschwerpunktmäßig mit der älteren Urgeschichte aus-einandersetzt. Artikel über Themen aus dem Paläoli-thikum kommen nur gelegentlich in den verschie-denen englischen internationalen Zeitschriften vor.Andererseits muss man ebenfalls erwähnen, dass Zeit-schriften wie Antiquity (gegründet 1927) oder Procee-dings of the Prehistoric Society (erste Ausgabe 1911),eine weltweite Verbreitung haben, was bei vielenPublikationsorganen, die die ältere Urgeschichte alsSchwerpunkt haben, nicht der Fall ist.
Skandinavien
Vorwiegend verwendete Literatur: Kristiansen 1985.
Da Skandinavien abgesehen vom westlichen TeilDänemarks während der letzten Eiszeit unter denGletschern lag, fehlen hier die „klassischen“jungpaläolithischen Technokomplexe vollkommen(z. B. Aurignacien, Gravettien, Solutréen, Magda-lénien). Erst ab dem Spätpaläolithikum mit derHamburger Kultur scheint diese Region durchgehendbesiedelt gewesen zu sein. Weiterhin dauerte dasMesolithikum hier länger als in anderen Regionen unddie Technokomplexe Skandinaviens sind nur schwermit anderen und insbesondere mit westeuropäischenRegionen zu vergleichen. Daher hat Skandinavienzum Beginn der Forschungsgeschichte im Bereich derälteren Urgeschichte nur eine zweitrangige Rollegespielt; im Rahmen der jüngeren Urgeschichte undFrühgeschichte dagegen war die skandinavischeForschung von Anfang an sehr bedeutend17. Man darfauch nicht vergessen, dass der Beginn der Erfor-schung der älteren Urgeschichte in vielen Regionensich fast ausschließlich auf Höhlen konzentrierte, undHöhlen wie auch Höhlenfundstellen sind in Skandi-
navien außerordentlich selten. Erst als das „Gerüst“der älteren Urgeschichte, insbesondere in zeitlicherHinsicht, etwas fester wurde, bekam diese Regiondeutlich mehr Aufmerksamkeit. So wurde beispiels-weise trotz weiterer Entdeckungen zwischen 1920 und1940 und wichtiger neuer Erkenntnisse durch GustavSchwantes die zeitliche Einordnung der Gruppe derStielspitzen erst 1946 durch Erik Westerby anhand derDaten aus der 1944 ausgegrabenen FundstelleBromme endgültig geklärt (Kristiansen 1985, 83),obwohl die erste Fundstelle dieser Gruppe inDänemark (Fjellerup Mark) schon 1890 entdecktwurde. Ähnlich lange dauerte die Diskussion um dieMuschelhaufen der Ertebølle Kultur. Während dieFundstelle Ertebølle schon am Ende des 19. Jahrhun-derts von Sophus Müller und anderen ausgegrabenworden war, erfolgte ihre richtige Einstufung an dasEnde des Mesolithikums erst 1927 durch ErikWesterby.
Die extreme Bodenfeuchtigkeit sowie die zahlreichenMoore haben jedoch dazu geführt, dass sich gerade inSkandinavien andere Schwerpunkte bilden konnten,darunter sei hier auf die hervorragenden Erhaltungsbe-dingungen für organisches Material hingewiesen.
So sind aus Dänemark, Südschweden und FinnlandBoote (u. a. Tybrind Vig, Dänemark), Fischnetze,zahlreiche Bögen und Pfeile (u. a. Holmegaard,Dänemark) sowie andere Artefakte mit vollständigerhaltener Schäftung bekannt. Das sind Funde undBefunde, die im übrigen Europa, außer inNorddeutschland, Polen, den Baltischen Staaten undRussland, bis jetzt weitgehend fehlen.
Die besonderen Bodenbedingungen haben schonfrüh zur Entwicklung unterschiedlicher Analysever-fahren an organischem Material geführt, darunter seihier auf die Pollenanalyse und auf die Analyse vonsonstigen Pflanzenresten hingewiesen. Es ist eben keinZufall, dass schon 1901, durch Hartz und Milthers, dieAlleröd-Schwankung erkannt und nach dem dänischenDorf Allerød benannt wurde, und das Bölling-Inter-stadial bekam seinen Namen von dem dänischen SeeBølling.
Ferner förderte die geringe Tiefe des Meeres in derOstsee die organische Erhaltung und steigerte so dieZahl der Untersuchungsmöglichkeiten. Durch eusta-tische Veränderung befinden sich weite Teile derehemaligen Küstenregion heute einige Meter unterdem Meeresniveau. Der Begriff Unterwasser-Archäo-logie steht hier längst nicht mehr nur für die Bergungversunkener Schiffe, sondern für die genaueAusgrabung von sehr unterschiedlichen Befunden amMeeresboden.
17 Man braucht hier nur an den Dänen Christian Jürgensen Thomsen (1788–1865), Direktor des KöniglichenMuseums für Nordische Altertümer und Begründer des Drei-Perioden-Systems, und an den Schweden OscarMontelius (1843–1921), den Entwickler der „Typologische Methode“, zu erinnern.

20 Verbreitung der großen Jagdfauna in Mittel und Westeuropa im oberen Jungpleistozän
Unter den für die Erforschung der Eiszeit relevantenZeitschriften aus Skandinavien ist „BOREAS“(gegründet 1972, herausgegeben vom „The NationalCouncils for Scientific Research in Denmark, Finland,Norway and Sweden“, Oslo) zu erwähnen.
Forschungsimpulse aus dem Rest Europas
Vorwiegend verwendete Literatur: Groenen 1994;Neugebauer-Maresch 1999.
Es würde zu weit führen, hier ausführlich auf dieForschungsgeschichte der Länder des östlichen Mittel-europa, Osteuropas sowie des ehemaligen Jugosla-wiens und Griechenlands einzugehen, da dies imvorliegenden Zusammenhang nur von geringerBedeutung wäre. Weiterhin war die politische TeilungEuropas von der Mitte der 40er Jahre bis zum Endeder 80er Jahre des letzten Jahrhunderts auch in derErforschung der älteren Urgeschichte zu spüren. Eswar in dieser Zeit kaum möglich, gemeinsameProjekte mit Ländern östlich des Eisernen Vorhangsdurchzuführen. Die Teilnahme an internationalenFachkongressen im „westlichen“ Ausland war für dieWissenschaftler in den Ländern des „Ostblocks“ allesandere als selbstverständlich und der hohe Preis derPublikationen aus dem „Westen“ war oft für dieBibliotheken in Osteuropa unerschwinglich.
Sprachbarrieren spielten und spielen ebenfalls einegroße Rolle. Während nach dem Zweiten Weltkrieg inWesteuropa und im westlichen Mitteleuropa Englischund Französisch internationale Fachsprachen wurden,gebrauchte man in Osteuropa und im östlichen Mittel-europa Russisch. Jedoch muss man hier auch differen-zieren. Während in den meisten Ländern derSowjetunion fast nur russisch publiziert wurde,wurden im östlichen Mitteleuropa (insbesondere inden Baltischen Staaten, Polen, in der Tschecho-slowakei und Ungarn) teilweise Französisch, Deutschund Englisch ebenfalls als internationale Sprachenakzeptiert. In diesem Zusammenhang sind hierfolgende Zeitschriften zu erwähnen: „Anthropologie“(gegründet 1962, herausgegeben vom „MoravianMuseum Brno“, in der Tschechische Republik), „ActaArchaeologica Hungarica“ (gegründet 1951, herausge-geben von den „Academiae Scientiarum Hunga-ricae“) sowie „Starunia“ (gegründet 1934, letzteAusgabe 1953, herausgegeben von der PolnischeAkademie der Künste und der Wissenschaften), diespäter den Name „Folia Quaternaria“ übernahm(herausgegeben von 1960 bis 1992 von der PolnischeAkademie der Wissenschaften in Krakau und seit1993 von der Kommission der Paläogeographie desQuartärs der Polnischen Akademie der Künste und derWissenschaften).
Während viele Wissenschaftler im östlichen Mittel-europa und teilweise in Osteuropa sich mit Franzö-sisch, Deutsch und Englisch auseinander setzten, gabund gibt es nur wenige Wissenschaftler im westlichenMitteleuropa und in Westeuropa, die russisch odereine andere slawische Sprache gut lesen können.Daraus folgt, dass Artikel oder ganze Zeitschriften mitSchwerpunkt auf der älteren Urgeschichte, die aufTschechisch, Bulgarisch oder auf Russisch publiziertwurden und werden, kaum im westlichen Mittel-europa, in Westeuropa und den USA wahrgenommenwerden. Seit der politischen Abnahme des Einflussesder Sowjetunion bzw. der Russischen Föderationpublizieren viele Zeitschriften aus Osteuropa nunimmer häufiger Artikel auf Englisch. Durch dieseneue Situation entstand in den letzten Jahren ein bisdahin unbekannter Austausch an Informationen undZusammenarbeit in den Bereichen Quartärgeologie,Ökologie und Archäologie.
Unter den Entdeckungen, die auch in den Westenwichtige Impulse weitergaben, sei hier auf die erstenflächigen Ausgrabungen in den 1920er Jahren durchSerguéï Nicolaïevitch Zamiatnine in Gagarino (Rus-sland) und Piotr Petrovitch Efimenko in Kostienki I(Russland), sowie auf die Ausgrabungen von 1955 bis1980 durch Aleksandr Černyš in Molodova (Ukraine)hingewiesen. Wegweisend waren auch die Arbeiten inder Sowjetunion in den 1930er Jahren von SerguéïAristarkhovitch Sémionov über die Gebrauchsspuren-analyse (Groenen 1994, 105–109). Unter den Wissen-schaftlern, die nach dem Zweiten Weltkrieg einebedeutende Rolle im östlichen Mitteleuropa gespielthaben und auch außerhalb des „Ostblocks“ bekanntwaren, sind hier Janusz Krzysztof Kozłowski (Polen),Bohuslav Klima, Martin Oliva, Jiři Svoboda und KarelValoch (Tschechische Republik), Ladislav Bánesz(Slowakische Republik), Viola Dobosi (Ungarn) undMarín Cârciumaru (Rumänien), Andrei Velichko(Russland) und Pavel Iosifovich Boriskovskii(Ukraine) zu erwähnen.
Als forschende Institutionen sind hier die natio-nalen Akademien der Wissenschaften zu nennen, die,manchmal in Zusammenarbeit mit spezialisiertenMuseen, in vielen Ländern des östlichen Mitteleu-ropas und Osteuropas den Hauptteil der Forschungs-arbeit durchgeführt haben.
Österreich gehört geographisch gesehen größten-teils zum östlichen Mitteleuropa, sprachlich gehört esjedoch zum deutschsprachigen wissenschaftlichenRaum. Aus der ersten Hälfte des 20. Jahrhunderts isthier unbedingt die ethnologische „Wiener Schule“(u. a. Wilhelm Schmidt, Fritz Graebner und WilhelmKoppers) zu erwähnen. Diese entwickelte als Alter-native zur Theorie der einseitigen Entwicklung derKultur die Hypothese, dass nach der gemeinsamenGenese der primitiven Kulturen eine parallele, unhab-hängige Entwicklung stattfand. Diese Konzepte, die

Forschungsgeschichte in Mittel- und Westeuropa (Schwerpunkt Jungpaläolithikum) 21
man als historisch-kulturelle Entwicklung bezeichnet,haben sehr stark die urgeschichtliche Forschungbeeinflusst und wurden insbesondere durch die Werkevon Josef Bayer, Oswald Menghin und HugoObermaier verbreitet (Broglio und Kozłowski 1987,19). In dieser Zeit wurden die ersten jungpaläo-lithischen Fundstellen in Österreich entdeckt undausgegraben (u. a. in Willendorf und Krems-Hunds-steig). Im Jahre 1924 (letzte Ausgabe 1930) gründeteJosef Bayer die Zeitschrift „Die Eiszeit“ (ab 1928„Die Eiszeit und Urgeschichte“). Nach dem Tod vonJosef Bayer 1931 erlebte die paläolithische Forschungin Österreich eine Phase der Stagnation, die erst amAnfang der 1950er Jahre dank Fritz Felgenhauer,Friedrich Brandtner und der Paläontologin MariaMottl endete (Neugebauer-Maresch 1999, 9). Unterden wichtigsten Arbeiten der letzten Jahrzehnte sindhier die Ausgrabungen von Christine Neugebauer-Maresch in Stratzing und von Friedrich Brandtner inGrubgraben zu erwähnen.
Forschungsimpulse aus den Vereinigten Staaten von Amerika
Schon bei der Ansprache der ForschungsgeschichteGroßbritanniens habe ich auf die besondere Rolle derbritischen Archäologie hingewiesen, die gegenüberKontinentaleuropa einen ganz anderen Blickwinkelhat. Noch stärker gilt das für die Forschungsge-schichte in den USA. Aus US-amerikanischer Sicht istEuropa in mancher Hinsicht auch für die Forschungrecht klein und weit entfernt. Ethnologische Ansätzesowie überregionale Vergleiche sind nicht nur selbst-verständlich, sondern üblich. Themen aus der europäi-schen älteren Urgeschichte werden häufig Erkennt-nissen oder Theorien aus Beobachtungen vonamerikanischen Ureinwohnern gegenübergestellt. Dasbeste Beispiel sind die kritischen Ansätze von LewisRoberts Binford (1962, 1982 sowie Binford undBinford 1969), der im Rahmen der „New Archaeo-logy“ für wichtige Impulse in Mittel- und Westeuropagesorgt hat.
Unter den bekanntesten Zeitschriften können hiernur einige erwähnt werden: „Current Anthropology“(gegründet 1960, herausgegeben von der „Universityof Chicago“), „Journal of AnthropologicalArchaeology“ (gegründet 1982), „Journal of Archaeo-logical Research“ (gegründet 1992, herausgegebenvon der „University of Wisconsin“), „Journal ofArchaeological Science“ (gegründet 1974, heraus-gegeben von der „Society for ArchaeologicalSciences“), „Journal of World Prehistory„ (gegründet1987, herausgegeben von der „University of
Washington“), „Journal of Field Archaeology“(gegründet 1974, herausgegeben von der „BostonUniversity“) und „American Antiquity“ (gegründet1935, herausgegeben von der „Society for AmericanArchaeology“). Viele US-amerikanische Zeitschriftensind außerdem über das Internet erhältlich, was ihreVerbreitung noch stärker begünstigt.
Zu erwähnen ist hier ferner die langjährigeTeilnahme mancher Professoren oder Universitäten inden Vereinigten Staaten an Forschungsprojekten inEuropa. Darunter sind Leslie Freeman („University ofChicago“), der seit Jahrzehnten in Spanien tätig ist,Lawrence Guy Straus („University of New Mexico,Albuquerque“), der in Belgien und Spanien tätig ist,Olga Soffer („University of Illinois, Urbana“), die aufOsteuropa einen Schwerpunkt ihrer Forschung gelegthat und Anta Montet-White („University of Kansas“),die seit Jahrzehnten in Bosnien und Österreich tätigist.
Die weltweite Akzeptanz von Englisch als derinternationalen Wissenschaftssprache hat außerdemviel zur Verbreitung von Theorien und der weltweitenBekanntheit von Fundstellen beigetragen. Demge-genüber muss man aber auch die fast sprichwörtlicheGleichgültigkeit vieler Kollegen – und nicht mal zusehr solche, die aus englischsprachigen Ländernstammen – für alles, was nicht auf Englisch publiziertwurde, erwähnen. Manche davon lesen oft nichteinmal die Publikationen in ihrer eigenen Mutter-sprache. Wenn man manche Publikationen mithochgesetztem Anspruch über Themen liest, die ganzEuropa oder große Teile davon behandeln, und manweiß, dass die Autoren zum Beispiel gar kein Deutschlesen oder besser gesagt lesen können, wird die Kritikvon Mantegazza aus dem Jahr 1877 (siehe S. 9) füreine bestimmte Haltung in der Wissenschaft leiderwieder sehr aktuell.
Überregionaler Vergleich
Die ältere Urgeschichte hat in den verschiedenenLändern nicht nur eine unterschiedliche Forschungs-geschichte, sondern auch einen deutlich unterschied-lichen Stellenwert und einen anderen Forschungs-stand. Dies spiegelt sich natürlich auch in den Datenwieder, die in dieser Arbeit untersucht werden.
Als wichtigster Aspekt muss unterstrichen wer-den, dass man trotz der länderspezifischen Entwick-lung, der unterschiedlichen Forschungsorgane18 und-schwerpunkte, in Mittel- und Westeuropa mittler-weile über qualitativ einheitliche Daten verfügt. Injedem Land stehen Funde und Daten aus älteren, zumTeil ungenau durchgeführten Ausgrabungen solchenaus modernen Untersuchungen gegenüber. Zuverläs-
18 In jedem Land existiert z. B. eine unterschiedliche Organisation des Denkmalschutzes, der Universitäten, derwissenschaftlichen Forschung und der Publikationsorgane.
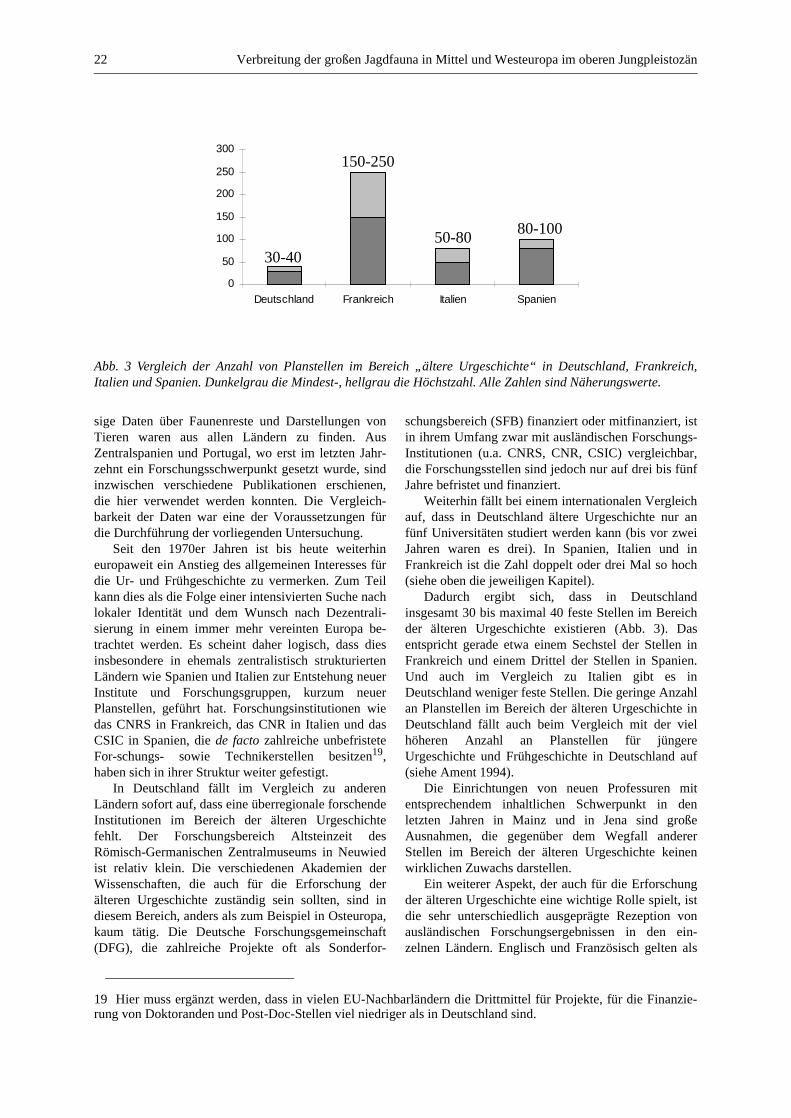
22 Verbreitung der großen Jagdfauna in Mittel und Westeuropa im oberen Jungpleistozän
sige Daten über Faunenreste und Darstellungen vonTieren waren aus allen Ländern zu finden. AusZentralspanien und Portugal, wo erst im letzten Jahr-zehnt ein Forschungsschwerpunkt gesetzt wurde, sindinzwischen verschiedene Publikationen erschienen,die hier verwendet werden konnten. Die Vergleich-barkeit der Daten war eine der Voraussetzungen fürdie Durchführung der vorliegenden Untersuchung.
Seit den 1970er Jahren ist bis heute weiterhineuropaweit ein Anstieg des allgemeinen Interesses fürdie Ur- und Frühgeschichte zu vermerken. Zum Teilkann dies als die Folge einer intensivierten Suche nachlokaler Identität und dem Wunsch nach Dezentrali-sierung in einem immer mehr vereinten Europa be-trachtet werden. Es scheint daher logisch, dass diesinsbesondere in ehemals zentralistisch strukturiertenLändern wie Spanien und Italien zur Entstehung neuerInstitute und Forschungsgruppen, kurzum neuerPlanstellen, geführt hat. Forschungsinstitutionen wiedas CNRS in Frankreich, das CNR in Italien und dasCSIC in Spanien, die de facto zahlreiche unbefristeteFor-schungs- sowie Technikerstellen besitzen19,haben sich in ihrer Struktur weiter gefestigt.
In Deutschland fällt im Vergleich zu anderenLändern sofort auf, dass eine überregionale forschendeInstitutionen im Bereich der älteren Urgeschichtefehlt. Der Forschungsbereich Altsteinzeit desRömisch-Germanischen Zentralmuseums in Neuwiedist relativ klein. Die verschiedenen Akademien derWissenschaften, die auch für die Erforschung derälteren Urgeschichte zuständig sein sollten, sind indiesem Bereich, anders als zum Beispiel in Osteuropa,kaum tätig. Die Deutsche Forschungsgemeinschaft(DFG), die zahlreiche Projekte oft als Sonderfor-
schungsbereich (SFB) finanziert oder mitfinanziert, istin ihrem Umfang zwar mit ausländischen Forschungs-Institutionen (u.a. CNRS, CNR, CSIC) vergleichbar,die Forschungsstellen sind jedoch nur auf drei bis fünfJahre befristet und finanziert.
Weiterhin fällt bei einem internationalen Vergleichauf, dass in Deutschland ältere Urgeschichte nur anfünf Universitäten studiert werden kann (bis vor zweiJahren waren es drei). In Spanien, Italien und inFrankreich ist die Zahl doppelt oder drei Mal so hoch(siehe oben die jeweiligen Kapitel).
Dadurch ergibt sich, dass in Deutschlandinsgesamt 30 bis maximal 40 feste Stellen im Bereichder älteren Urgeschichte existieren (Abb. 3). Dasentspricht gerade etwa einem Sechstel der Stellen inFrankreich und einem Drittel der Stellen in Spanien.Und auch im Vergleich zu Italien gibt es inDeutschland weniger feste Stellen. Die geringe Anzahlan Planstellen im Bereich der älteren Urgeschichte inDeutschland fällt auch beim Vergleich mit der vielhöheren Anzahl an Planstellen für jüngereUrgeschichte und Frühgeschichte in Deutschland auf(siehe Ament 1994).
Die Einrichtungen von neuen Professuren mitentsprechendem inhaltlichen Schwerpunkt in denletzten Jahren in Mainz und in Jena sind großeAusnahmen, die gegenüber dem Wegfall andererStellen im Bereich der älteren Urgeschichte keinenwirklichen Zuwachs darstellen.
Ein weiterer Aspekt, der auch für die Erforschungder älteren Urgeschichte eine wichtige Rolle spielt, istdie sehr unterschiedlich ausgeprägte Rezeption vonausländischen Forschungsergebnissen in den ein-zelnen Ländern. Englisch und Französisch gelten als
Abb. 3 Vergleich der Anzahl von Planstellen im Bereich „ältere Urgeschichte“ in Deutschland, Frankreich,Italien und Spanien. Dunkelgrau die Mindest-, hellgrau die Höchstzahl. Alle Zahlen sind Näherungswerte.
0
50
100
150
200
250
300
Deutschland Frankreich Italien Spanien
30-40 50-80
80-100
150-250
19 Hier muss ergänzt werden, dass in vielen EU-Nachbarländern die Drittmittel für Projekte, für die Finanzie-rung von Doktoranden und Post-Doc-Stellen viel niedriger als in Deutschland sind.

Forschungsgeschichte in Mittel- und Westeuropa (Schwerpunkt Jungpaläolithikum) 23
internationale Sprachen. Es ist daher verständlich,dass die Ergebnisse aus diesen Ländern sofort interna-tional rezipiert werden.
Bei der Hugo-Obermaier Tagung in Tübingen imJahr 2000 ist allgemein festgestellt worden, dass diedeutschsprachige Forschung im Bereich der älterenUrgeschichte im Ausland kaum wahrgenommen wird.Dies gilt im internationalen Vergleich jedoch nochstärker für die slawische und auch skandinavischeLiteratur. Dadurch ergibt sich automatisch, dass esheutzutage unerlässlich ist, die wissenschaftlichenErgebnisse auch in einer internationalen Sprache zupublizieren. Andererseits wird aber auch mehrEngagement in Richtung Öffentlichkeitsarbeit erwar-tet, um das gestiegene Interesse der anspruchsvollenÖffentlichkeit besser zu befriedigen. Dies kann man
nur in der jeweiligen Muttersprache erreichen. ImBaskenland und in Katalonien wurden beispielsweisein den letzten 20 Jahren wissenschaftliche Publika-tionen vermehrt auch in den offiziellen, lokalenSprachen publiziert, Baskisch und Katalanisch (siehedie Zeitschriften Munibe und Cypsela). Dies steht dortaber keineswegs im Gegensatz zur parallelenVeröffentlichung in internationalen Publikations-organen auf Englisch oder Französisch.
Solange jedoch die Erforschung der älteren Urge-schichte in Deutschland nur auf eine geringe Anzahlan fest angestellten Wissenschaftlern zurückgreifenkann, wird sie dem gestiegenen Anspruch an die wis-senschaftliche Arbeit und die damit verbundene Ver-breitung der Forschungsergebnisse nicht gerechtwerden können.


Europa am Ende der letzten Eiszeit
Topographie Europas
Die Geographie Europas am Ende des Pleistozänswurde im wesentlichen vom Ausdehnen und Zurück-ziehen der Gletscher und dem damit verbundenenAuf- und Absteigen des Meeresspiegels bestimmt. Dadie Kontinentalplatten sich in der Größenordnung vonnur wenigen Zentimetern pro Jahr bewegen, spieltdieser Faktor für diese geologisch „kurze“ Zeitspannekaum eine Rolle. So haben auch Vulkane,Landhebung oder -absenkung meistens nur in einemregionalen Rahmen und in Verbindung mit anderenEreignissen (z. B. Rückzug von Gletschern, lokaleTektonik) eine Rolle gespielt.
Die Abbildung 4 zeigt das mögliche AussehenEuropas zwischen 23.000 und 18.000 BP, als dieGletscher in Europa ihre maximale Ausdehnungerreichten und der Meeresspiegel weltweit um ca.100 m unter dem heutigen Niveau lag. Die einzelnenNummern werden im Text erklärt und aufgegriffen.Die geographischen Namen und die Berghöhenstammen aus „Knaurs Großem Weltatlas“ (Bartho-lomew et al. 1980).
1. Das Packeis auf dem Atlantischen Ozean erreichteim Winter während des zweiten Kältemaximums nachOdette Conchon, Marton Pécsi, Andrej Velichko undanderen Wissenschaftlern (Velichko et al. 1991, 49,51 und 59) Nordspanien. Der Golf von Biscaya war,nach dieser Rekonstruktion, im Winter zugefroren.Nach der Rekonstruktion von Burkhard Frenzel(Velichko et al. 1991, 39, 43 und 45) soll die Packeis-grenze nur bis auf die Höhe der Bretagne nach Südenvorgestoßen sein. Nach William Ruddimann undAndrew McIntyre (Ruddimann und McIntyre 1981,177, Abb. 19) soll die Polarfront im Atlantik zu dieserZeit bis Lissabon gereicht haben. Da solche Grenzenals Orientierung bei den Schwankungen innerhalbmehrerer Jahrtausende zu verstehen sind, kann manannehmen, dass im Winter das Meer bis zur Bretagne,häufig aber auch deutlich südlicher zugefroren war.
2. Im Sommer schmolz das Eis auf dem Ozeanwährend des zweiten Kältemaximums nach BurkhardFrenzel (Velichko et al. 1991, 41, 43, 45 und 47) biszu einer hypothetischen Linie von Südirland bis fastnach Island. Nach Odette Conchon u. a. (Velichko etal. 1991, 49, 51 und 59) war der Bereich zwischendem nordeuropäischen Eisschild und dem grönlän-
dischen Eisschild nicht das ganze Jahr über vereist.Auch hier lag wahrscheinlich die Grenze des ewigenEises auf dem Meer meistens zwischen diesen zweiRekonstruktionsvorschlägen. Andererseits ist dieseFrage für diese Arbeit nicht besonders relevant, daman davon ausgehen kann, dass während des zweitenKältemaximums keine Menschen in diesem Bereichlebten.
3. Die nordeuropäische Vergletscherung bedeckte dieskandinavische Halbinsel, den östlichen Teil vonDänemark und Norddeutschland (bis Hamburg undBerlin), Nordpolen, die baltischen Staaten, dennördlichen Teil von Weißrussland (bis südlich vonVilnius und nördlich von Minsk) und den nordwest-lichen Teil von Russland (ca. 100 km nördlich vonMoskau) bis zur Kanin Halbinsel (Woldstedt 1958).Auf der Kanin Halbinsel (Woldstedt 1958, 120) undauf dem Weißen Meer (Velichko et al. 1991, 59)gingen in dieser Zeit die nordostsibirischen (mitZentrum auf der Insel Novaya Zemlya) und dienordeuropäischen Vergletscherungen ineinander über.
Dem skandinavischen Eisschild wird eineMächtigkeit von 3000 m im nördlichen Teil derheutigen Ostsee20, von ca. 2000 m auf der Linie vonOslo, Karlskrona, Gotland, Tallinn, St. Petersburg,und von noch 1000 m auf der Linie Ålborg, Szczecin,Gdansk, Tikhivin, Arkhangel‘sk zugerechnet (sieheWoldstedt 1958, 5, Abb. 1). Das heißt, dass währendder letzten Vereisung dieser Schild ein 3000 m hohesund von Nordosten nach Südwesten mehrere hundertKilometer langes Hindernis für die Luftströmung mitüberregionalen Folgen darstellte, insbesondere was dieNiederschläge betrifft (man denke an die Bedeutungz. B. der Alpen für das Klima in Europa).
Das skandinavische Gebirge ist im Durchschnittetwa 1500 m hoch und überschreitet heute nur inEinzelfällen 2000 m (u. a. im Südwesten Glittertind,2470 m, und Snøhetta, 2286 m, im Norden Kebne-kaise, 2111 m, und Sarektjåkkå, 2090 m). Es liegtdamit ca. 1500 m tiefer als der Eisschild während desKältemaximums.
4. Es ist umstritten, inwieweit und ob der fennoskan-dische und der britische Gletscher überhaupt inein-ander übergingen (Woldstedt 1958, 4–14; Velichko etal. 1991, 43, 59; Lang 1994, 90, Abb. 4.2–1). Es wirdaber generell angenommen, dass dieser Bereich dasganze Jahr lang vereist war (Velichko et al. 1991),
20 Paul Woldstedt (1958, 121, Abb. 39) präsentiert auch eine Karte von S. A. Jakowlew von 1956, in der eineHöhe von bis zu 3500 m für denkbar gehalten wird.

26 Verbreitung der großen Jagdfauna in Mittel und Westeuropa im oberen Jungpleistozän
daher gab es hier eine Eiswüste, die genauso lebens-feindlich wie der fennoskandische und der britischeGletscher war. Diese Eisfront und die Tatsache, dassder größte Teil der Nordsee über dem damaligenMeeresspiegel lag, verhinderte das Entwässern derFlüsse nach Norden. Heutzutage liegt die Meerestiefe
zwischen Schottland und Norwegen meistens bei 50bis 100 m, zwischen England und Dänemark,Deutschland, den Niederlanden und Belgien bei 25 bis50 m, wobei hier die Doggerbank zu erwähnen ist, diezum Teil noch höher liegt.
Abb. 4 Europa während des zweiten Kältemaximums (ca. 23.000–18.000 BP). Die Küstenlinie liegt 100 munter dem heutigen Normalnull. Die Gletscher wurden anhand von Angaben aus folgenden Publikationengezeichnet: Andersen und Borns 1997, 44, 55; Bartholomew et al. 1980; Blanc 1938; Cremaschi 1990;Duphorn et al. 1970; Frenzel 1968; Lang 1994, 90; Schmidt-Thomé 1973; Woldstedt 1958, 168; InternationaleQuartär-Karte von Europa 1:2.500.000.
-100m / -50m -50m / 0m
0m / 300m 300m / 900m
> 900mGletscher
Packeis (Winter)Packeis (Sommer)
Eisstausee
3 2
4
5
6
8
7 9
10 1 11
13 12
14 15 20
22 21 17 16
18
23 24
25 19

Europa am Ende der letzten Eiszeit 27
5. Die Britischen Inseln lagen wie schon erwähnt untereinem Eisschild. Dieser reichte im Westen bis zurKüstenlinie von Irland und der Äußeren Hebriden, imSüden bis zur Linie Limerick, Dublin, Bristol,Birmingham und Leeds. Im Osten ging der Gletschermöglicherweise ab der Höhenlinie von Middlesbo-rough und Hull in die heutige Nordsee über, imNorden wurde er vom Packeis begrenzt.
6. Im Bereich der Nordsee entstand ein Eisstausee,welcher insbesondere im Sommer als Sammelbeckenfür die Schmelzwässer aus den umgebendenGletschern diente. Die Elbe, welche zum Teil auch dasSchmelzwasser vom fennoskandischen Gletschertransportierte, mündete direkt in diesen See(Woldstedt 1958, 13, Abb. 5). Da im Norden derEntwässerungsweg durch Gletscher gesperrt war,musste dieser See im Südwesten über den Ärmelkanalentwässern. Aufgrund der geringen Tiefe der heutigenGebiete der Nordsee zwischen England undDänemark, Deutschland, den Niederlanden undBelgien kann man annehmen, dass dieser See eineTiefe von wenigen Metern kaum überschritten hat undeher wie ein riesiges Sumpfgebiet ausgesehen hat. ImWinter war dieses Gebiet zugefroren.
7. Die Themse war, ebenso wie der Rhein und dieSeine, ein Nebenfluss des Urstroms, der aus demEisstausee stammte und durch den Ärmelkanal floss.Der Urstrom, welcher nach dem Aussehen der Höhen-linien beim Ärmelkanal wahrscheinlich ab und zulanggestreckte Seen gebildet hatte, mündete in denAtlantik auf der Höhe von Cornwall im Norden undder Bretagne im Süden. Dieser Fluss, sowie alleandere Flüsse Nordeuropas, konnte aber nur imFrühling und im Sommer fließen, da er im Winterselbst zugefroren war oder das Meer vor ihm. Daherwaren die Britischen Inseln in dieser Zeit vom RestEuropas nicht durch ein Meer getrennt, sondern durcheinen gewaltigen Fluss, der in wenigen Monaten imJahr das Schmelz- und Regenwasser aus einemriesigen Gebiet entwässerte (Williams et al. 1998, 151,Abb. 8.5).
8. Aufgrund der Absenkung des Meeresspiegels lagenBelgien, die Niederlande, Norddeutschland und derTeil von Dänemark, der nicht unter der nordeuropäi-schen Vergletscherung lag, über dem Meeresspiegel.Da die Mündung des Urstroms (Woldstedt undDuphorn 1974, 3; Andersen und Borns 1997, 61)mehrere hundert Kilometer entfernt war, war dasGefälle andererseits so niedrig, dass im Frühling,wenn die Flüsse wieder fließen konnten, und imSommer, wenn die Temperaturen teilweise höher alsdie heutigen lagen (Frenzel 1980, 20), hier weitrei-chende Überschwemmungen stattfanden. Dies hat
während der Eiszeit eine besonders wichtige Rollegespielt, da das Klima generell von starkerTrockenheit geprägt war (Bosinski 1987, 4).
Die deutschen Mittelgebirge (u. a. Harz, ThüringerWald, Westerwald, Eifel, Hunsrück, Taunus, Oden-wald) waren einerseits zu weit südlich, um vomnordeuropäischen Gletscher bedeckt zu sein, anderer-seits, da diese Gebirgsketten gerade 800 bis 900 m mitihren höchsten Gipfeln erreichen, zu niedrig, um großeeigene Gletscher zu bilden. Der einzige Berg, der inder kältesten Phase zum Teil einen Gletscher bildenkonnte, ist der Brocken im Harz mit 1142 m.
9. Gerade an der heutigen Grenze zwischen Tsche-chien und der Slowakei zu Deutschland und Polenbefinden sich etwas höhere Gebirgsketten. Hier sind,insbesondere bei den höchsten Bergen, einige lokaleGletscher zu vermuten (Bayrischer Wald mit demGroßen Rachel, 1452 m und dem Großen Arber,1457 m; Erzgebirge mit dem Klinovec/Keilberg,1244 m; Sudety/Sudeten mit dem Snìzka/Schnee-koppe, 1603 m und dem Pradìd, 1490 m; Tatry/HoheTatra mit dem Gerachovsky/Gerlsdorfer Spitze,2663 m und dem Bystrá, 2250 m).
Eine wichtige Rolle spielen hier die Flusstäler derLabe/Elbe, Odřa/Oder, Morava/Moldau und derWisła/Weichsel. Diese, mit einer Höhe von wenigerals 500 m über Normalnull, stellten während derganzen Eiszeit die wichtigste Verbindung fürMenschen und Tiere zwischen der norddeutschen undnordpolnischen Ebene auf der einen und demDonautal und der ungarischen Ebene auf der anderenSeite dar. Weitere niedrige Pässe (etwa 500–600 m)lagen im Osten zwischen den Flusstälern der Oravaund Raba, Hornad und Poprad. Die ostbayerischenMittelgebirge werden ihrerseits zwar nicht vonFlüssen geschnitten, ihre Überquerung ist aber anmehreren Stellen nicht besonders schwer, da mancheTäler knapp oberhalb einer Höhe von 500 m liegen(z. B. vom Naabtal zum Tal der Eger/Ohře, vomRegen- und vom Chamtal zum Tal der Radbuza).
10. Das Donautal schneidet auf einer Länge vonmehreren tausend Kilometern Europa von Westennach Osten. Gerade in einer Zeit, in der Gebirge nochstärker als heute als eine Grenze wirkten, ist dieDonau während der Eiszeit als wichtige Ost-West-Verbindung anzusehen.
11. Zwischen den Vogesen (höchster Berg ist derGrand Ballon/Großer Belchen, 1424 m) und demSchwarzwald (Feldberg, 1493 m, Belchen, 1414 mund Kandel, 1241 m) befindet sich der Rheintal-graben. Hier fließt der Rhein, der, zusammen mit denFlüssen Saône und Rhône, die wichtigste Nord-Süd-Verbindung in Westeuropa darstellt (u. a. m Floss2002).

28 Verbreitung der großen Jagdfauna in Mittel und Westeuropa im oberen Jungpleistozän
12. Die atlantische Küste Frankreichs lag im Südenziemlich nahe der heutigen Küstenlinie. Auf der Höhevon Bordeaux war sie mehr als 20 km, auf der Höhevon Nantes ca. 50 km von der heutigen Küste entfernt.
13. Die Alpen, die noch heute nur über sehr hochgelegene Pässe, meistens in ca. 2000 m Höhe zuüberqueren sind, waren während der Eiszeit voneinem riesigen Gletscher bedeckt, aus dem nur diehöchsten Gipfel herausragten. Der alpine Eisschildwar abschnittsweise über 100 km breit und erstrecktesich von Ligurien am Mittelmeer bis in die Steiermarkin Österreich über eine Länge von mehr als 1000 km.Eine Überquerung dieser Barriere durch Menschenoder Tiere (Zugvögel ausgenommen) ist für diegesamte Eiszeit auszuschließen. Die Alpen bildetenschon immer aufgrund ihrer Höhe und der Ost-West-Ausrichtung ein Hindernis für warme Luftströmungenvom Mittelmeer nach Norden und für kalte Luft ausSkandinavien nach Süden. Durch die Präsenz desalpinen Eisschildes war dieser Einfluss noch verstärkt.
Diese riesige weiße Eisfläche, die schon durch denAlbedoeffekt die umgebende Region abkühlte, hatwohl auch einen starken „Kühlschrankeffekt“ produ-ziert. Da warme Luft nach oben steigt, kalte Luft nachunten sinkt, kam aus dem alpinen Eisschild ständigkalte Luft in die umgebenden, tiefer liegendenRegionen (siehe dazu das Kapitel „Klimaübersicht“,S. 32–37).
14. Das Massif Central war an mehreren Stellenvergletschert. Seine höchsten Gipfel sind der Puy deSancy, 1886 m, der Plomb du Cantal, 1858 m, der PuyMary, 1787 m, der Mont Mézenc, 1754 m, Finiels,1702 m und Pierre sur Haute, 1640 m. Diese Regionliegt größtenteils über 1000 m hoch. Nur dasFlusssystem von Saône und Rhône, welches abChalon-sur-Saône fast geradlinig von Norden nachSüden zieht, trennt mit seinem zum Teil recht engenTal das Massif Central im Osten von den Alpen. NachWesten und Nordwesten nimmt das Massif Centralsanft an Höhe ab. Nach Südosten erreichen nur verein-zelte Gebirgsgruppen die 1000 m, die meisten liegenzwischen 500 und 1000 m, bis dann die Pyrenäenanfangen. Das Tal zwischen Toulouse und Carcas-sonne, wo heute der Canal du Midi fließt, liegt unter200 m Höhe und schneidet von Osten nach Westendiese Hügellandschaft.
15. Die Po-Ebene war um die Zeit des zweiten Kälte-maximums fast 300 km länger als heutzutage undverband damit das heutige Dalmatien mit der Apenni-nenhalbinsel bis südlich von Ancona. Durch dieseverlängerte Po-Ebene flossen alle nach Südenentwässernden Wasserläufe aus den Alpen und allenach Norden entwässernden Wasserläufe aus denNordapenninen. Das Wasser trug sehr viel Sediment
und das Gefälle war sehr gering (ca. 1 m Höhenunter-schied auf 3000 m Distanz). Dies führte zu einer sehrstarken Sedimentation in der gesamten Po-Ebene.Damit liegen die jungpaläolithischen Fundstellenunerreichbar unter mehreren Metern Flusssedimentund lassen bei Kartierungen die Po-Ebene so gut wiefundleer erscheinen (u. a. Mussi 2001, 289). Nuraußerordentlich glückliche Zufälle zeigen die Präsenzdes Menschen in dieser Region (Gallini und Sala2000).
16. Der östliche Teil der adriatischen Küste, der heutedie stark zerklüftete kroatische Küste bildet, steigtschnell auf über 1000 m Höhe an (Snežnik, 1798 m,Rajinac, 1699 m, Vaganjski, 1758 m, Dinara, 1831 mund Troglav, 1913 m). Manche Gebirgszüge weiter imInneren und weiter im Süden erreichen sogar 2000 mHöhe (Klekovaèa, 1961 m, Èvrsnica, 2228 m, Žabljak,2522 m, Komovi, 2484 m und Korabi, 2764 m) undwaren während der letzten Eiszeit vergletschert. Nurin der Nähe des heutigen Triest gab es natürlichePässe, die niedrig genug waren (ca. 500 m), um diePassage bestimmter Tierarten und Menschen zwischendem Balkan und der Apenninenhalbinsel zu ermög-lichen. Die Ausrichtung von Nordwesten nachSüdosten der meisten Gebirgsketten dieser Region istals weiteres Hindernis für West-Ost-Kontakte zusehen.
17. Die Apenninen bildeten entlang der ganzenApenninenhalbinsel durch ihre Höhe ein Hindernisvon Nordwesten nach Südosten für Menschen undTiere. Sie erreichen an verschiedenen Stellen entlangihrer ganzen Länge eine Höhe von 2000 m (Cimone,2165 m, Gran Sasso, 2914 m, Petroso, 2247 m, SerraDolcedorme, 2271 m und Aspromonte, 1956 m).Durch ihre Ausläufer, die zum Teil bis ans Meerreichen, gliedern sie die Landschaft in viele kleineRegionen. Der höchste Berg Italiens außerhalb derAlpen ist jedoch der Vulkan Ätna auf Sizilien, derheute 3323 m erreicht.
Grundsätzlich weist die Adriatische Küste inMittel- und Süditalien während der letzten Eiszeit eintrockeneres Klima als die Tyrrenische Küste auf. Sogibt es östlich der Apenninen viele Hinweise auf einesteppenartige Vegetation, während es westlich davonzahlreiche Hinweise auf Wälder gibt.
18. Aufgrund des niedrigen Meeresniveaus war dieInsel Elba mit dem Kontinent verbunden. Zwischender Apenninenhalbinsel und Korsika lag ein nur ca.20 km breiter Kanal. Die Straße von Bonifacio lagtrocken, damit war Sardinien mit Korsika verbunden.
19. Während sich die Nord- und die Ostküste vonSizilien trotz des niedrigen Meeresspiegels kaum inder Gestalt verändert haben, sahen die Süd- und

Europa am Ende der letzten Eiszeit 29
Westküste Siziliens ganz anders aus. Hier entstandeine ungefähr 20 km breite Ebene, die als Streifen vonSyrakus bis Trapani die Küste entlang reichte. Maltaund Gozo bildeten eine einzige Insel, die sich nochetwa 60 km in Richtung Osten ausdehnte. Dazwischen Malta und Sizilien und zwischen Sizilienund Pantelleria an mehrere Stellen das Meerheutzutage weniger als 100 m tief ist, kann man hierfür diese Zeit mit zum Teil sehr breiten Inseln rechnen(ohne den hier sehr aktiven Einfluss der Tektonik zuberücksichtigen). In diesem Fall wäre Sizilien nichtmehr 150, sondern nur 60 km von Afrika entfernt.Heutzutage beträgt die Distanz zwischen Sizilien undItalien gerade einmal 3 km und das Meer ist zwischen100 und 200 m tief. Ob Sizilien vor 20.000 Jahren mitItalien verbunden war, kann nicht mit Gewissheitbeantwortet werden. Tatsache ist, dass die Tektonik ander Meerenge von Messina sehr aktiv und die Meeres-strömung stark ist. Eine Verbindung mit langen Unter-brechungen scheint jedoch aufgrund der Faunenunter-schiede wahrscheinlich (Zampetti et al. 2000).
20. Die galizische und kantabrische Küste sind sehrsteil, daher lag die eiszeitliche Küste von der heutigenKüste meistens nur wenige 100 m bis einigeKilometer entfernt. Nur an wenigen Stellen erreichtediese Distanz bis zu 10 km.
21. Die Pyrenäen sind eine Gebirgskette, die sich über400 km hauptsächlich von Westen nach Ostenerstreckt. Sie erreichen an verschiedenen StellenHöhen von 3000 m (Vignemale, 3303 m, MontePerdido, 3355 m, Aneto, 3404 m und Pic d’Estats,3115 m). Noch heute sind sie nur an wenigen Stellenüber hohe Pässe zu überqueren. In der Zeit des Kälte-maximums waren sie zu einem wesentlichen Teil vonGletschern bedeckt und daher kaum zu passieren.Durch ihre Lage trennen sie die iberische Halbinselvom übrigen Europa.
22. Die Cordillera Cantabrica steht den Pyrenäen nurwenig nach, was die Höhe und ihre Rolle alsHindernis betrifft. Auch sie ist von Westen nach Ostenausgerichtet. Ihre Gipfel erreichen ca. 2500 m (PeñaUbina, 2417 m, Picos de Europa, 2648 m, Peña Prieta,2536 m).
23. Das Zentrum der iberischen Halbinsel liegtgrößtenteils über 500 m über dem Meeresspiegel undwird von fünf großen Flusssystemen entwässert. DerEbro fließt im Norden von der Cordillera Cantabricanach Südwesten bis ins Mittelmeer, in das auch dieanderen Flüsse aus dem Süden der Pyrenäen münden.Der Duero (Douro auf Portugiesisch), der Tajo (Tejoauf Portugiesisch), der Guadiana und der Guadal-quivir fließen dagegen von Osten nach Westen undmünden in den Atlantik. Zwischen diesen Flüssen
liegen zum Teil hohe Gebirge (Sierra de la Demandamit dem Gipfel San Lorenzo, 2262 m, Sierra deGuadarrama mit dem Gipfel Peñalara, 2430 m, Sierrade Gredos mit dem Pico Almanzor, 2592 m, Sierra deSegura mit dem Gipfel Sagra, 2381 m und die SierraNevada mit dem Gipfel Mulhacen, 3478 m), diezumeist von Westen nach Osten ausgerichtet sind.
24. Die Mittelmeerküste vor der Ebromündung (Golfode San Jorge und Golfo de Valencia) sowie vor derMündung der Rhône (Golf du Lion) ist aufgrund derSedimente dieser zwei wasserreichen Flüsse ziemlichflach. Während der letzten Eiszeit war daher die Küstemehr als 60 km vorgelagert. Die Balearen Mallorcaund Menorca bildeten eine einzige größere Insel sowieauch Ibiza und Formentera. Sie waren allerdingswährend der letzten Eiszeit nicht mit Spanienverbunden, was aus dem Verbreitungsbild der erst inhistorischer Zeit ausgestorbenen endemischen Antilo-penart Myotragus balearicus hervorgeht (Martin undKlein 1984, 79).
25. Die Südküste Spaniens am Mittelmeer ist sehrsteil. Gipfel von über 1000 m Höhe liegen nur 5 bis10 km vom Meer entfernt. Die eiszeitliche Küste lagnur wenige hundert Meter oder wenige Kilometer vonder heutigen entfernt. Die Straße von Gibraltar,welche heute 15 km breit ist, war wahrscheinlich nurgeringfügig enger.
Betrachtet man die weitere Entwicklung derTopographie Europas auf der Abbildung 5, kann manfür die Zeit von 13.000 bis 11.000 BP folgendewesentliche Änderungen feststellen.
26. William Ruddimann und Andrew McIntyre konn-ten anhand von Untersuchungen an Foraminiferenzeigen, dass in der Zeit zwischen 13.000 und 11.000BP (ca. Bölling bis Alleröd) die Polarfront im Atlantiksehr stark zurückgedrängt und das Meer zwischenSchottland und Island in dieser Zeit eisfrei war(Ruddimann und McIntyre 1981, 180, Abb. 21).Zwischen 11.000 und 10.000 BP (ca. Jüngere Dryas)war dagegen die Polarfront wieder bis Nordspanienvorgerückt (Ruddimann und McIntyre 1981, 181,Abb. 22; Andersen und Borns 1997, 81, Abb. 2–53).
27. Die Rückbildung des Eisschildes in Skandinavienist seit langem sehr gut untersucht worden (Aaris-Sørensen 1992; Woldstedt 1958). Eine der Folgen desRückzugs des Eises ist eine Entlastung der Erdkrustevon dessen gewaltigem Gewicht. Das Gebiet um dennördlichen Teil der Ostsee hebt sich noch heute, wasdazu führt, dass sich andere Gebiete an der Peripheriedes ehemaligen Eisschildes wie Dänemark oderMecklenburg-Vorpommern absenken (Lampe 2002).Für die ältere Urgeschichte spielt das eine wichtige

30 Verbreitung der großen Jagdfauna in Mittel und Westeuropa im oberen Jungpleistozän
Rolle, da dadurch ganze Fundstellen unter Wasserverschwunden sind und damit häufig organischesMaterial erhalten geblieben ist.
28. Während der fennoskandische Eisschild sich nurlangsam zurückzog, ging dieser Prozess auf den Briti-schen Inseln viel schneller. Im Alleröd gab es
anscheinend keine Gletscher mehr in Irland, Walesund England. Nur noch in Schottland waren größereGletscher vorhanden.
29 Beim Rückzug des fennoskandischen Eisschildesbildete sich im Ostseeraum ein Eisstausee (BaltischerEissee).
Abb. 5 Europa am Ende der letzten Eiszeit (ca. 13.000–11.000 BP). Die Küstenlinie entspricht 50 m bis 25 munter dem heutigen Normalnull (Bartholomew et al. 1980). Die Gletscher wurden für die Zeit von ca. 13.000 bis11.000 BP dargestellt. Für Südeuropa und die Alpen schematisiert hauptsächlich anhand der Höhenlinien (fürNordeuropa nach den Daten von Aaris-Sørensen 1992, 144; Lang 1994, 268; Lowe und Walker 1984; Woldstedt1958; Woldstedt und Duphorn 1974).
26 27
28 29
30 31 32 33
34 35
-50m / -25m -25m / 0m
0m / 300m 300m / 900m
> 900mGletscher
Eisstausee

Europa am Ende der letzten Eiszeit 31
30. Die Nordsee war in dieser Zeit südlich einer Linievon der schottisch-englischen Grenze über dieDoggerbank bis nördlich von Dänemark trocken.Durch den Rückzug des Gletscher- und Packeiseskonnte das kontinentale Wasser aus dem nach Südenentwässernden fennoskandischen Eisschild sowie ausden nach Norden entwässernden mitteleuropäischenFlusssystemen über das Gebiet der heutigen Nordseenach Norden gelangen. Aufgrund des geringen bisfehlenden Gefälles der ganzen Region und wegen derstarken Sedimentation stauten sich die Flüssewahrscheinlich in sich ständig verändernden Süßwas-serseen (siehe Andersen und Borns 1997, 74, Abb. 2–46 und 81, Abb. 2–53).
31. Die westlich des Rheins fließenden Flüsse entwäs-serten weiter über den Ärmelkanal in den Atlantik. Dadie Nordsee sehr flach ist, könnte der Rhein selbersowohl nach Westen als auch nach Norden entwässerthaben. Hier ist jedoch zu erwähnen, dass es beimHöhepunkt der Jüngeren Dryaszeit, als die Polarfrontwieder bis Nordspanien vorgerückt war, Permafrostbis im Süden Frankreichs sowie zugefrorene Flüsse imWinter und starke Überschwemmungen in Frühlingund Sommer gegeben haben muss.
32. Der Meeresspiegel lag zu dieser Zeit ca. 50 munter dem heutigen Normalnull. Die kontinentalefranzösische Atlantikküste lag dadurch mehrereKilometer von der heutigen Küste entfernt. Dies istsowohl in West- als auch in Nordfrankreich der Fall.Das Gebiet des heutigen Ärmelkanals lag über demMeeresspiegel. Nur die Pyrenäen sowie Teile derBretagne und der Normandie, wo es noch heute einesteile Küste gibt, hatten damals wohl recht ähnlicheKüstenverläufe wie heute.
33. Die Moränen in den Alpen zeigen verschiedenePhasen des Rückzugs der Gletscher, die zum Teilschwierig zu korrelieren sind. In der Abbildung 5 sollnur ein Überblick angegeben werden.
34. Die meisten Gletscher Spaniens waren schon imAlleröd verschwunden oder auf ein minimalesAusmaß reduziert. Nur auf den Pyrenäen lagen nochgrößere Gebiete das ganze Jahr lang unter dem Eis.
35. Die Mittelmeerküste in Westeuropa sah mögli-cherweise zu dieser Zeit kaum anders aus als heute,nur die Po-Ebene war mehr als 100 km länger alsheute. Der Golf du Lion sowie der Golfo de Valenciawaren ebenfalls noch zu einem wesentlichen Teiltrocken, deren Küstenlinien lagen aber wesentlichnäher zu den heutigen als im Fall der Po-Ebene.

32 Verbreitung der großen Jagdfauna in Mittel und Westeuropa im oberen Jungpleistozän
Klimaübersicht
Um Europa und die Verbreitung der Tiere währendder letzten Eiszeit besser verstehen und untersuchenzu können, ist eine Übersicht über das Klima in derbehandelten Zeit unerlässlich. Dabei soll hier jedochder Akzent weniger auf die üblichen Eisbohrkerne ausGrönland, Tiefseesedimente vom Atlantik, Pollen-profile und Pollenstufen, Böden und Gletschervor-
stöße sowie errechnete Temperaturmittelwerte gesetztwerden. Diese sind in vielen Publikationen in Formvon Tabellen und Kurven behandelt (u. a. Jöris undWeninger 1996; Weißmüller 1997). Es sollen hiervielmehr die Tag- und Nacht-, die Sommer- undWinter- sowie regionale Unterschiede betont werden,die u. a. von den Meeresströmungen und Windverhält-nissen verursacht wurden.
Als Erstes möchte ich hier die Sonneneinstrahlungansprechen, deren Ausprägung immer als bekanntvorausgesetzt und daher selten diskutiert wird.
Die Sonneneinstrahlung ist die Energie, die vonder Sonne abgegeben und von der Erde aufgenommenwird. Auf der Energie der Sonne beruht das ganzeLeben auf der Erde, daher haben Differenzen in ihrerVerteilung eine wichtige Bedeutung in Bezug auf dieVerbreitung der Flora und der Fauna. Trotz leichter
Änderungen der Erdachsenneigung und der Sonnen-einstrahlung kann man für die Eiszeit von einerähnlichen zeitlichen Verteilung der Sonnenein-strahlung wie der heutigen ausgehen. Zwischen dergeographischen Breite von Gibraltar und Malta (ca.37° nördliche Breite) und der von Bremen (ca. 53°nördliche Breite) liegen deutliche Unterschiede in derSonneneinstrahlung vor. Während das Tageslicht inMitteleuropa zur Sonnenwende im Sommer 16 bis 17
Abb. 6 A Verbreitung der entdeckten Weichteile von Mammuten (nach Joger und Koch 1994, 56–57, Abb. 35 undnach Guthrie 1990). 1–2 Mammut, Adams (Russland); 3–7 Mammutfriedhof, Berelekh (Russland); 8–9 Mammut,Khatanga (Russland); 10 Mammut, Beresowka (Russland); 11 Mammut, Kirigilyak (Russland); 12 Mammut,Juribei (Russland); 13 Mammut, Jamal (Russland); 14 Mammut, Elephant (USA); 15 Mammut, Fairbanks (USA);16 Mammut, Starunia (Ukraine).

Europa am Ende der letzten Eiszeit 33
Stunden andauert, ist der Tag in Südeuropa nur 14 bis15 Stunden lang. Dies gleicht sich dann im Winteraus, wenn in Südeuropa die Tage länger als in Mittel-europa sind (zur Wintersonnenwende circa 9 bis 10Stunden gegenüber nur 7 bis 8 in Mitteleuropa).Zusätzlich steht die Sonne, je weiter man nach Nordengeht, um so tiefer am Himmel und um so schwächersind die Sonnenstrahlen, die die Erde erreichen. DieUnterschiede zwischen dem südlichen Südeuropa unddem nördlichen Mitteleuropa sind zwar bedeutsam, siesind jedoch im Vergleich zur Sonneneinstrahlung amPolarkreis (ab ca. 66’30° nördliche Breite) ehergering. Hier gibt es im Sommer bei einem niedrigenSonnenstand bis zu 24 Stunden Sonneneinstrahlungam Tag. Die winterliche Polarnacht dauert ebenfallsbis zu 24 Stunden. Damit ist es klar, dass man Funde
aus dem Polarkreisgebiet mit Funden aus Mittel- undWesteuropa nur mit Vorsicht vergleichen sollte, wasnicht immer geschieht.
Als Beispiel sei hier auf die gefrorenen Mammutsaus der Tundra Nordsibiriens hingewiesen (Abb. 6Aund 6B), die häufig unkritisch mit Mammut-darstellungen aus Höhlen Südfrankreichs verglichenwerden.
Aufgrund der geographischen Lage sind amehesten die Überreste aus Starunia (früher in der öster-reichischen Provinz Galizien, dann Polen, heuteUkraine) mit den Faunenresten aus Mittel- undWesteuropa vergleichbar21.
Die beste und kompakteste Beschreibung desKlimas der Eiszeit für Mitteleuropa befindet sich beiGerhard Bosinski, daher wird diese hier wiederge-geben (Bosinski 1987, 5): „Obwohl damals wie heuteWestwind vorherrschte, brachte der Wind keine
Abb. 6 B Verbreitung der entdeckten Weichteile von Wollnashörnern, Pferden und Steppenbisons (nach Jogerund Koch 1994, 56–57, Abb. 35; Guthrie 1990). 1–2 Pferd, Selerikan (Russland); 3 Pferd, Dukarskoje(Russland); 4 Nashorn, Vilyui (Russland); 5 Nashorn, Churapcha (Russland); 6 Nashorn, Starunia (Ukraine);7 Steppenbison, Mylakhchyn (Russland); 8 Pferd und Steppenbison, Fairbanks (USA).

34 Verbreitung der großen Jagdfauna in Mittel und Westeuropa im oberen Jungpleistozän
Wolken und Niederschläge, da der Ozean kalt war undkaum verdunstete. Der blaue Himmel ermöglichte eineungehinderte Sonneneinstrahlung. Im Juli und Augustwar es teilweise wärmer als heute.
Doch auch die Kälte wurde nicht durchBewölkung abgehalten. So gab es starke Temperatur-unterschiede zwischen Tag und Nacht und besondersnatürlich zwischen Sommer und bitterkaltem Winter“.
Für Mitteleuropa kann man von einer offenenGraslandschaft ausgehen. „Das weitgehende Fehlenvon Bäumen und Wald wurde nicht durch die Kälte,sondern durch die Trockenheit verursacht. Die Bäumewuchsen nur in den Flusstälern“ (Bosinski 1987, 4).
Mitteleuropa war während der Eiszeit von einerLösssteppe bedeckt. Lössschichten sind auf einembreiten, sich von Westfrankreich bis zum SchwarzenMeer über den Ural bis nach Sibirien ziehenden
Streifen (Bosinski 1987, 5, Abb. 2) zu finden22. Indieser Lösssteppe kamen sowohl Pflanzen und Tiereaus der heutigen Subarktis als auch aus den Trocken-gebieten Zentralasiens vor. Heutzutage sind diese zweiÖkosysteme durch einen zum Teil mehrere tausendKilometer breiten Waldstreifen, die Taiga, getrennt.
Ein Biotop wie die eiszeitliche Lösssteppe kommtheute nirgends mehr vor. Der einfache Vergleich mitder heutigen Subarktis wäre, insbesondere wegen desflachen Sonnenstandes sowie der langen Winternacht,sehr problematisch (Poplin 1976, 66). Die damaligeLösssteppe vereinte Charakteristika, Pflanzen undTiere aus der Subarktis (z. B. Moschusochse undRentier), aus der Trockensteppe (Saiga-Antilope undPferd) und aus den baumlosen Hochgebirgen(Steinbock und Gämse).
Während des Kältemaximums der letzten Eiszeit(ca. 18.000 BP) kam die Polarfront im Atlantik vielweiter nach Süden als es heute der Fall ist. Sieerreichte den 42. Breitengrad, also die Höhe vonNordportugal. Bis hierher gab es im Winter Packeis.Der Golfstrom, der heute warmes Wasser bis vorSkandinavien führt, war „gekippt“, und lief nachSüden statt nach Norden (Abb. 7). Relevant istweiterhin, dass, während heute die gemäßigteStromzone an der europäischen Küste ca. 2500 kmbreit ist (Sarnthein 1980, Abb.11), diese während desKältemaximums auf nur ca. 500 bis 600 kmgeschrumpft war. Das heißt, dass im Atlantikzwischen der Polarströmung, die sich heute zwischenIsland und Grönland befindet, und den subtropischenWassermassen nur knapp 1000 km lagen.
Auch die Wasserverhältnisse im Mittelmeer sahenwährend der letzten Eiszeit deutlich anders aus (Mussi2001, 219–222). Die Verengung des Kanals vonSizilien von den heutigen 150 auf nur 60 km führte zueiner noch deutlicheren Trennung zwischen demwestlichen und dem östlichen Mittelmeerbecken. ImWesten, auch durch die Zufuhr von kaltem Wasser ausdem Atlantik, erreichten die Temperaturen (Abb. 8)im Winter 7°C bis 13°C (gegenüber den heutigen13°C bis 15°C) und im Sommer 13°C bis 19°C(gegenüber den heutigen 22°C bis 24°C). Es mussjedoch betont werden, dass es sich hier keineswegs umsehr kalte oder „polare“ Temperaturen gehandelt hatte.Die Nord- und die Ostsee weisen gegenwärtig Winter-und Sommertemperaturen auf, die teilweise nochdarunter liegen.
21 Dort wurden Weichteile von einem Mammut und einigen Wollnashörnern in einer Erdwachsgrube aus Ozoke-rit zu Beginn des 20. Jahrhunderts entdeckt (Nowak et al. 1930). Nach dem Zweiten Weltkrieg befand sich Staru-nia jedoch an der Grenze der Sowjetunion und war daher für Wissenschaftler lange unzugänglich. Erst in denletzten Jahren haben polnische und ukrainische Paläontologen die Erforschung dieser Fundstelle wieder auf-nehmen können (Kowalski 2000).
Abb. 7 Oberflächenströmungen im nordöstlichenAtlantik während des Kältemaximums um 18.000 BP.Nach der Auswertung von Plankton-Foraminiferen imSediment (nach Ruddiman und McIntyre 1976 undSarnthein 1980).
22 Löss scheint auch in einigen Regionen Italiens vorzukommen (Guidi und Piperno 1992, 25, Abb. 8), wobeinicht immer klar ist, ob es sich hier um ein und dasselbe handelt.
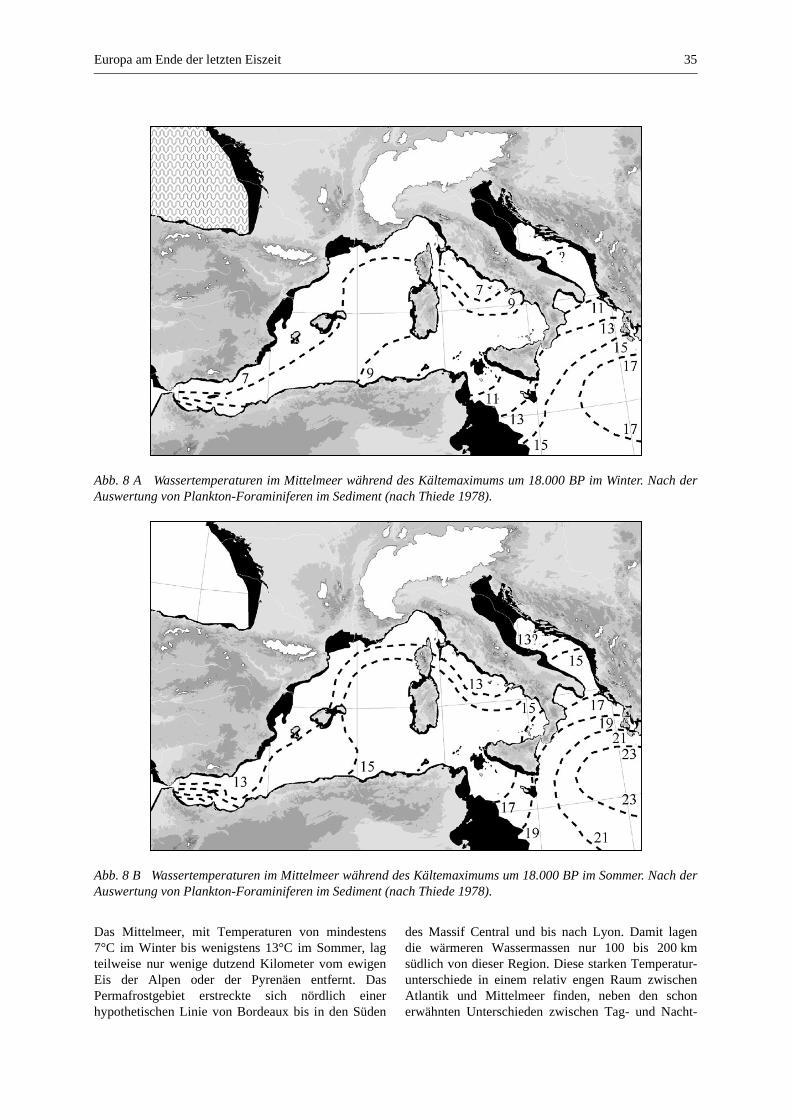
Europa am Ende der letzten Eiszeit 35
Das Mittelmeer, mit Temperaturen von mindestens7°C im Winter bis wenigstens 13°C im Sommer, lagteilweise nur wenige dutzend Kilometer vom ewigenEis der Alpen oder der Pyrenäen entfernt. DasPermafrostgebiet erstreckte sich nördlich einerhypothetischen Linie von Bordeaux bis in den Süden
des Massif Central und bis nach Lyon. Damit lagendie wärmeren Wassermassen nur 100 bis 200 kmsüdlich von dieser Region. Diese starken Temperatur-unterschiede in einem relativ engen Raum zwischenAtlantik und Mittelmeer finden, neben den schonerwähnten Unterschieden zwischen Tag- und Nacht-
Abb. 8 A Wassertemperaturen im Mittelmeer während des Kältemaximums um 18.000 BP im Winter. Nach derAuswertung von Plankton-Foraminiferen im Sediment (nach Thiede 1978).
Abb. 8 B Wassertemperaturen im Mittelmeer während des Kältemaximums um 18.000 BP im Sommer. Nach derAuswertung von Plankton-Foraminiferen im Sediment (nach Thiede 1978).

36 Verbreitung der großen Jagdfauna in Mittel und Westeuropa im oberen Jungpleistozän
temperaturen sowie zwischen Sommer- und Winter-temperaturen, keine Parallele in den heutigenKlimaverhältnissen. Nichtsdestotrotz kann mananhand der Lössvorkommen und durch Analogiensowie aus Modellberechnungen einige Aussagentreffen.
Thermisch bedingte Luftströmungen waren entlangder Küste und der Berge stärker ausgeprägt als heute.Aus den Gletschern kam auch im Sommer ständigkalte, schwerere Luft auf Bodenhöhe in dieumgebenden Regionen. „Über dem skandinavischenEisschild sowie über Grönland befanden sich flacheHochdruckgebiete“ (Meyer und Kottmeier 1989, 17).Da Hochdruckgebiete in der Nordhemisphäre sichimmer im Uhrzeigersinn drehen (Roth 1995, 172–177), ist anzunehmen, dass die Winde entlang desSüdrandes des skandinavischen Eisschildes vonNordosten und Osten nach Westen bliesen. Da aberauch während der Eiszeit in Mitteleuropa die üblicheWest-Ost-Windrichtung vorherrschte, gingen dieOstwinde in Westwinde über. Zum Vergleich werdenhäufig die aktuellen Verhältnisse am Rande Grönlandsund der Antarktis herangezogen (Meyer und
Kottmeier 1989, 16). Die Lössvorkommen in dieserRegion (Abb. 9A) scheinen dieses Bild zu bestätigen,und ähnliche Verhältnisse werden ebenfalls für denlaurentischen Eisschild formuliert (Williams et al.1998, Abb. 12.11).
Eine weitere für diese Arbeit relevante Hypothesebezieht sich auf die Verlagerung der üblichen Bahnder Tiefdruckgebiete und ist eng verbunden mit derHauptwindrichtung in Europa (Abb. 9B). DieAusdehnung des Packeises im Winter während desKältemaximums bis zum Norden der iberischenHalbinsel führte dazu, dass sich die Tiefdruckgebiete,die die hauptsächlichen Niederschläge bringen,meistens südlich dieser Linie bewegten (Meyer undKottmeier 1989, 16, Abb. 14; Florineth 1998, 35, Abb.7). Das bedeutet, dass kaum bis gar keine Nieder-schläge auf dem heutigen Weg durch den Nord-atlantik und Nordfrankreich bis nach Mitteleuropavom Westen her kommen konnten. Die Tiefdruck-gebiete scheinen eher südlich der iberischen Halbinselverlaufen zu sein, um dann Südfrankreich, denSüdrand des alpinen Eisschildes und den Balkan zuerreichen (Florineth und Schlüchter 2000).
Abb. 9 A Paläowindkarte von Mitteleuropa anhand der aus den Lössvorkommen abgeleiteten Lösswinde (nachMeyer und Kottmeier 1989, 12, Abb. 1 und 13, Abb. 2). Die Strich-Punkt-Linie stellt die Grenze des Permafrostesdar.

Europa am Ende der letzten Eiszeit 37
Diese Hypothese lässt sich gut mit globalen Zirkulati-onsmodellen sowie mit der Gletscherentwicklung inEuropa korrelieren. Die Gletscher in den Vogesen, imMassif Central und den Pyrenäen scheinen zumindestteilweise von dieser Winterzirkulation abgeschnittengewesen zu sein. Sie erreichen während des letztenKältemaximums nicht die Ausdehnung, die siewährend der Sauerstoff-Isotopenstufe 4 erreichthatten. Der Südrand des alpinen Eisschildes dagegen,welcher während des letzten Kältemaximums seinemaximale Ausdehnung erreichte, scheint im Wintervon ausreichenden Niederschlägen genährt worden zusein (Pons et al. 1989, 53, Abb. 3; Florineth undSchlüchter 2000, 304, Abb. 5). In Ungarn ist diePräsenz von Wäldern, die dem Südteil der heutigenTaiga entsprechen, zu dieser Zeit (Willis et al. 2000,211) nur mit ausreichenden Niederschlägen zuerklären. Schließlich kommen auch die pollenanaly-tischen Untersuchungen zum Ergebnis, dass die
Mittelmeerregion weniger von der Trockenheit, diewährend des letzten Kältemaximums herrschte,betroffen war als West- und Mitteleuropa (Peyron etal. 1998, 192, Abb. 5d).
Im Sommer, am Rand des Meereseises, dürftensich auf der Höhe von Island aufgrund der Tempera-turunterschiede zwischen Wasser und Eis Tiefdruck-gebiete gebildet haben, die aber auf ihrer östlichenZugbahn vor dem britischen und skandinavischenEisschild nach Nordosten abgedrängt wurden (Meyerund Kottmeier 1989, 17).
Aus der Beschreibung der wahrscheinlichen klimati-schen Zustände während des Kältemaximums inEuropa lässt sich jetzt vielleicht besser verstehen,weshalb das Gebiet Deutschlands in dieser Zeit sehrdünn besiedelt oder teilweise sogar menschenleer war(Street und Terberger 2000).
Abb. 9 B Wahrscheinlich typischer Verlauf der Tiefdruckgebiete über dem Mittelmeer und Europa im Winterwährend des letzten Kältemaximums (nach Florineth und Schlüchter 2000, 306, Abb. 6). Die Strich-Punkt-Liniestellt die Grenze des Permafrostes dar.


Jäger und Beute in Europa am Ende der letzten Eiszeit
Der Mensch
Obwohl der Mensch während der Eiszeit als Teil derNatur anzusehen ist, hatte er schon damals das obereEnde der Nahrungskette erreicht (Abb. 10 und 11). Erwar der einzige Räuber, der für erwachsene Elefanten,Mammuts und Nashörner eine Gefahr darstellte.Zusätzlich konnte er auch für erwachsene Raubtiereeine Bedrohung darstellen.
Die Entdeckung der hölzernen Wurfspeere vonSchöningen (Thieme 1997) zeigte, dass der Homoerectus schon vor mindestens 400.000 Jahren überhervorragende Waffen verfügte. Experimente habengezeigt (Steguweit 1999), dass damit bei einer Distanzvon 30 m eine große Treffgenauigkeit erreichbar war,wobei die übliche Wurfdistanz wahrscheinlichdarunter lag. Holz bleibt aber nur unter außeror-dentlich günstigen Verhältnissen erhalten, so dass dieurgeschichtliche Archäologie sich fast ausschließlichmit der technologischen Entwicklung der Stein- undKnochenspitzen befassen kann. Diese zeigen, dass abdem Jungpaläolithikum eine Erweiterung der Durch-schlagskraft und Treffdistanz der Geschosse durchSpeerschleuder und Pfeil und Bogen stattfand (Abb.10). Dadurch baute der Homo sapiens durch dieTechnologie seine Position noch mehr aus. Zu dieserhervorgehobenen Stellung führten aber nicht nur dietechnischen, sondern auch – und insbesondere – diegeistigen Errungenschaften. Der Homo sapiensbrachte viele Neuerungen nach Europa, manche vonüberwiegend technischer Art (u. a. gezielte Klingen-technologie, intensive Knochenbearbeitung), anderevon überwiegend kultureller Art (vielfältige Schmuck-herstellung, Kunst und komplexe Bestattungssitten).Diese Letzteren sind ein Beweis dafür, dass der Homosapiens nicht nur seine Umwelt erlebt, sondern dass erversucht, sie auch zu verstehen und unter anderemdurch Kunst anderen verständlich zu machen. SeineUmwelt zu verstehen bedeutete damals, logischeRückschlüsse über Wanderungsrouten potentiellerBeutetiere zu ziehen, die Bedeutung von Schluchten,Flüssen und offenen Landschaften für die Jagd richtigzu bewerten. Die Erfolgsgeschichte des Menschen alsJäger – in Europa Homo erectus, Homo neandertha-lensis und Homo sapiens – bedeutete gleichzeitigeinen größeren Selektionsdruck für Tiere, insbe-sondere für andere Räuber. Da der Mensch lokal die„besten“ Stellen für die Jagd und wiederholt Höhlenfür sich allein beanspruchte, kam er bestimmt immerwieder mit Raubtieren in Konflikt. Diese Begeg-nungen sind aber meist von zufälligem Charakter
gewesen. Eine aktive Jagd auf große Raubtiere wird,außer vielleicht auf den Bären (Morel 1998; Münzel etal. 2001), sehr begrenzt gewesen sein (Altuna 1972,424). Bildliche Darstellungen von Raubtieren mit„Pfeilen“ oder Linien kennt man aus verschiedenenFundstellen, und diese deuten an, dass es eine aktiveJagd auf große Raubtiere gab. Darstellungen vonRaubkatzen mit „Pfeilen“ sind u. a. aus Les TroisFrères (Bégouën und Breuil 1958, 10) und Lascaux(Nougier und Robert 1965, 23) bekannt. Darstel-lungen dieser Art von Höhlenbären stammen aus LesTrois Frères (Bégouën und Breuil 1958, 48, Abb. 4)und Les Combarelles (Breuil et al. 1956, 63). InFreilandfundstellen sind Raubtierknochen beinahenicht vertreten. In Kulturschichten von Höhlen sindsie, mit Ausnahme von Bär und Hyäne, immerseltener als Ungulaten. Da alle Fleischfresser natürlichin Höhlen vorkommen können, beispielsweise zumÜberwintern oder zum Setzen, ist die Herkunft dieserKnochen außerdem nur selten unzweideutig mit demMenschen in Verbindung zu bringen; noch seltenerkann man von sicherer Jagd sprechen.
Interessant ist in diesem Zusammenhang zu zeigen,dass eigentlich viele Tiere in der archäologischenLiteratur als „besonders gefährlich“ und damit alsmindestens „ungünstige“ Beute interpretiert wurden:
– Joachim Hahn betonte die Gefährlichkeit von Tierenwie Löwe, Bär, Mammut und Wollnashorn. Aus derTatsache, dass diese starken und aggressiven Tiere imAurignacien Südwestdeutschlands häufig auftreten,sowie wegen der aggressiven Haltung der Pferdedar-stellung vom Vogelherd entwickelte er die Theorieeiner Botschaft von Kraft und Aggression aus dieserZeit für die Kunst der Schwäbischen Alb (Hahn 1986).Martin Porr (2001) scheint diese Meinung zu teilen. – André Leroi-Gourhan betrachtete Löwe, Bär undNashorn als gefährliche Tiere (1965, 82).– Claude Guérin und Martine Faure vertreten dieThese, dass es unwahrscheinlich ist, dass dieMenschen im Paläolithikum Nashörner gejagt habenkönnen (Guérin und Faure 1983, 35), da diese als „zukraftvoll, zu aggressiv, zu gut gepanzert“ („troppuissants, trop agressifs, trop bien cuirassés“) definiertwerden. – Lawrence Guy Straus erklärt den Mangel an Restenvon Sus scrofa im älteren Jungpaläolithikum Spaniensdamit, dass Wildschweine von den Jägern gemiedenwurden (Straus 1992, 84) und dass sie gefährlichgewesen seien (Straus 1995, 352; siehe auch Cardoso1996, 239).

40 Verbreitung der großen Jagdfauna in Mittel und Westeuropa im oberen Jungpleistozän
Abb. 10 Der Mensch und seine bisher nachgewiesenen Waffen während der letzten Eiszeit (verändert nach Stodiek und Paulsen 1996, 14 und 15, Abb. 6 und 7).
– Anne Bridault interpretiert die Tatsache, dass dieAuerochsen in den Fundstellen aus dem Endpaläolithi-kum und Mesolithikum des französischen Jura seltensind, so, dass die Auerochsen gefährlicher als andereTiere gewesen sein sollen (Bridault 1998, 76). – Vladimir Georgievich Heptner, welcher zahlreichewichtige Arbeiten über die Ökologie der Sowjetuniongeschrieben hat, zitiert bezüglich der Jagd aufPrzewalski-Pferde in Sibirien, dass diese bis zumAussterben in freier Wildbahn als besondersgefährlich betrachtet wurden. Hengste werden alsaggressive Tiere beschrieben, die in der Lage waren,Reiter anzugreifen, um sie aus dem Sattel zu werfen(Heptner et al. 1966, 868)
Daraus kann man nur schließen, dass in den Augenverschiedener Forscher viele Tiere für den Menschenals Jäger gefährlich gewesen sein können. UnsereEinstufung entspricht aber möglicherweise nicht denKriterien der prähistorischen Menschen.Die von Straus vermutete besondere Gefährlichkeitvon Wildschweinen passt nicht mit den Ergebnisse auszahlreichen spätpaläolithischen und mesolithischen
Fundstellen zusammen, in denen Wildschweinedurchaus intensiv gejagt werden23. Auch wenn hierdie verbreitete Benutzung von Pfeil und Bogen einewichtige Rolle spielt (Straus 1995, 352), kann mansich nicht erklären, wieso Wildschweine gefährlicherals beispielsweise Steppenbisons gewesen sein sollen.
Schließlich gibt es von fast jeder Tierart bildlicheDarstellungen, in denen möglicherweise „Pfeile“ zuerkennen sind (Abb. 12). Es sei hier auf den Steinbockund die Steppenbisons von Niaux (Clottes 1995, 51,Abb. 40, 105, Abb. 119, und 123, Abb. 150;Lorblanchet 1995, 110), auf die Robben, dieHirschkuh und das unbestimmte Tier aus der HöhleCosquer (Clottes und Courtin 1994, 93, Abb. 64, 122,Abb. 112, und 131 Abb. 123), auf die Steppenbisonsund Rentiere von Les Trois Frères (Bégouën undBreuil 1958, 35, Abb. 39, und 66–67, Abb. 70–71),auf die Steppenbisons und Pferde von Lascaux (Leroi-Gourhan 1965, 354–355), auf das Wollnashorn von LaColombière (Graziosi 1956, Taf. 21; Leroi-Gourhan
Abb. 10 Der Mensch und seine bisher nachgewiesenen Waffen während der letzten Eiszeit (verändert nach Stodiek und Paulsen 1996, 14 und 15, Abb. 6 und 7).
23 U. a. in Spanien Zatoya IIb, Straus 1992, E3; in Italien Paglicci, Sala 1985; in Frankreich Mannlefelsen I Sund I Q, Ruffey-sur-Seille R4, Rochedane A3, Choisey C2, Bridault 1998).

Jäger und Beute in Europa am Ende der letzten Eiszeit 41
1965, 376) und auf das Pferd von der Grotta Pagliccihingewiesen24 (Vigliardi 1979, 234; Palma di Cesnola1993, 465; Lorblanchet 1995, 15).
Weiterhin sind fast alle Tiere, außer Raubtiere,auch auf Speerschleudern dargestellt (Tab. 1). Daszeigt, dass fast alle Tiere von Menschen im Paläoli-thikum als geeignet empfunden wurden, um imZusammenhang mit der Jagd und daher aufJagdwaffen abgebildet zu werden.
Die Raubtiere
Der Mensch war nicht der einzige Jäger der Eiszeit.Seine Aktivitäten fanden in einer Umwelt statt, in derviele andere „Nahrungskonkurrenten“ ihre eigeneökologische Nische hatten. Je nach Größe undJagdstrategie waren die Raubtiere auf bestimmteTierarten mehr oder weniger spezialisiert (Abb. 11und 13).
Die kleinen Fleischfresser wie Wildkatze (Felis silve-stris; 3 bis 8 kg), Luchs (Lynx lynx; 8 bis 38 kg),Fuchs (Vulpes vulpes, 3 bis 14 kg), Eisfuchs (Alopexlagopus; 2,5 bis 4,5 kg) und Vielfraß (Gulo gulo; 7 bis32 kg) konnten aufgrund ihrer Größe und ihrerJagdstrategie als Einzelgänger in der Regel nur fürkleine Beute oder junge und geschwächte Tieregefährlich werden (bis ca. 20–25 kg). Sie sind allebesonders scheu und meistens nachtaktiv. Sie sinddaher vom Menschen im Paläolithikum kaum alsKonkurrenz betrachtet worden.
Unter den großen Fleischfressern gab es in Europawährend der letzten Eiszeit den Löwen, den Bären, dieHyäne und den Wolf. Obwohl die Hauptnahrung auchbei größeren Räubern häufig aus kleiner Beute sowieaus jungen und geschwächten Tieren besteht, könnendiese auch größere Beute jagen. Dabei wenden sieunterschiedliche Jagdstrategien an.
Der heutige Löwe (Panthera leo), der 180 kg Körper-gewicht bei den Weibchen und 250 kg bei denMännchen erreichen kann, kann dank seiner Größeschon allein eine Beute bis zu einem Gewicht von ca.300 kg jagen. Höhlenlöwen dürften 5 bis 10% größergewesen sein (Koenigswald 2002, 114). Zwei Phasen
sind in ihrer Jagdstrategie wichtig: das Anschleichenund das Sprinten. In dieser letzten Phase wird kaumeine Distanz von 200 m überschritten (Weniger 1982,111). Auch bei der Jagd in der Gruppe spielen dasAnschleichen und Sprinten eine wichtige Rolle, wobeifast nur eine perfekte und koordinierte Aktion zumErfolg führen kann.
Da der Höhlenlöwe (Panthera leo spelaea) amEnde der Eiszeit ausgestorben ist, kann sein Verhaltennur vom Verhalten der heutzutage in Afrika und derals Restpopulation in einem Naturschutzgebiet inIndien im Freien unter natürlichen Verhältnissenlebenden Tiere abgeleitet werden25. Die dortigenklimatischen Verhältnisse sind aber von denen derEiszeit in Europa weit entfernt. In diesem Zusam-menhang wäre von Interesse zu wissen, bis zuwelchem Grad der wahrscheinlich bis in die erstenJahrhunderte nach Christus (Becker 1986, 169–173;Heptner und Sludskij 1980, 82) in Südosteuropalebende Löwe mit dem eiszeitlichen Höhlenlöwenverwandt war.
Die jungpaläolithischen Darstellungen von Löwenzeigen kein einziges Tier mit einer solch ausgeprägtenMähne, wie sie für männliche afrikanische Löwencharakteristisch ist. Die Mähne ist ein klarer Ausdruckfür Kraft, sie ist jedoch auffällig und behindertdeswegen das Anschleichen (Guthrie 1990, 103). Inder afrikanischen Savanne, wo eine hohe Dichte anBeute im Allgemeinen und an Zebras im Besonderenvorhanden ist, brauchen sich erwachsene männlicheLöwen, die ein großes Rudel führen, kaum an der Jagdzu beteiligen. Diese wird von weiblichen Individuendurchgeführt. Die wichtigste Aufgabe für männlicheTiere ist es, ihr Territorium zu verteidigen. Unterdiesen Voraussetzungen macht eine ausgeprägteMähne einen Sinn. In Regionen, wo die Dichte derpotentiellen Beute niedrig ist, sind die Rudel kleinerund die männlichen Tiere müssen sich an der Jagdbeteiligen. In diesem Fall ist die Mähne deutlichweniger auffällig. Das fast vollständige Fehlen26 vonUnterscheidungsmerkmalen zwischen männlichen undweiblichen Höhlenlöwen in der paläolithischen Kunstweist wahrscheinlich auf sehr kleine Rudel hin, woIndividuen beider Geschlechter zusammen jagten(Guthrie 1990, 103).
24 Nicht aus der Höhle Polesini, wie nach d’Errico und Ucelli Gnesutta 1999, 149, Abb. 24,4.25 Der Höhlenlöwe wird von den meisten Autoren als naher Verwandter des heutigen Löwen betrachtet (Guérinund Patou-Mathis 1996, 206). Trotzdem sei hier darauf hingewiesen, dass nach anderen Autoren der Höhlenlöwefür einen Verwandten des Tigers gehalten wird (u. a. Groiss 1996. Zum Thema siehe auch Koenigswald 2002,113–117 und Ziegler 1994, 51).26 Henri Lhote (1988) weist zum Beispiel darauf hin, dass die so genannte „Lionne des Trois-Frères“ kein weib-liches, sondern ein männliches Individuum darstellt. Einziges Unterscheidungsmerkmal sind zwei kleine Halb-kreise unter dem Schwanz, welche er als Hoden interpretiert.
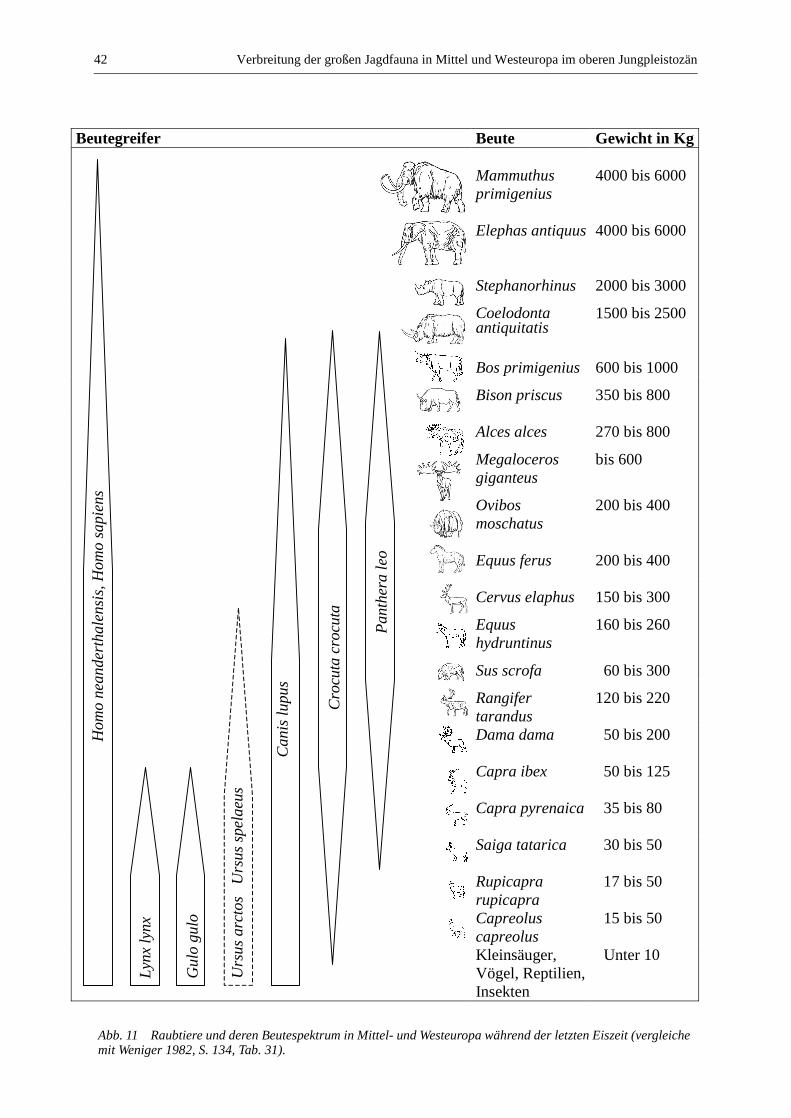
42 Verbreitung der großen Jagdfauna in Mittel und Westeuropa im oberen Jungpleistozän
B
eutegreifer Beute Gewicht in Kg
Mammuthus primigenius
4000 bis 6000
Elephas antiquus
4000 bis 6000
Stephanorhinus
2000 bis 3000
Coelodonta antiquitatis
1500 bis 2500
Bos primigenius
600 bis 1000
Bison priscus
350 bis 800
Alces alces
270 bis 800
Megaloceros giganteus
bis 600
Ovibos moschatus
200 bis 400
Equus ferus
200 bis 400
Cervus elaphus
150 bis 300
Equus hydruntinus
160 bis 260
Sus scrofa
60 bis 300
Rangifer tarandus
120 bis 220
Dama dama
50 bis 200
Capra ibex
50 bis 125
Capra pyrenaica
35 bis 80
Saiga tatarica
30 bis 50
Rupicapra rupicapra
17 bis 50
Capreolus capreolus
15 bis 50
Kleinsäuger, Vögel, Reptilien, Insekten
Unter 10
Hom
o ne
ande
rtha
lens
is, H
omo
sapi
ens
Cro
cuta
cro
cuta
Pan
ther
a le
o
Urs
us a
rcto
s U
rsus
spe
laeu
s
Gul
o gu
lo
Lyn
x ly
nx
Can
is lu
pus
Abb. 11 Raubtiere und deren Beutespektrum in Mittel- und Westeuropa während der letzten Eiszeit (vergleiche mit Weniger 1982, S. 134, Tab. 31).

Jäger und Beute in Europa am Ende der letzten Eiszeit 43
Inwieweit Höhlenlöwen in Europa Tiere, die in ihremFluchtverhalten bestens die Gegebenheit des Geländesausnutzen können, wie Elch oder Steinbock, erfolg-reich jagen konnten, ist unklar (Abb. 13, Mitte oben),da die Verbreitung dieser Tiere sich heute nicht mitder Verbreitung von Löwen überschneidet.
Der Höhlenbär (Ursus spelaeus) ist ausgestorben.Dank genauer Untersuchungen der Knochenrestesowie der Sedimente der Höhlen, die er besucht hat,kann man aber heute seine Lebensweise ziemlichgenau rekonstruieren (Rabeder et al. 2000). Er wardeutlich größer als die heutigen europäischen Braun-bären (Ursus arctos). Diese erreichen in der Regel170 cm bis 220 cm Länge und wiegen zwischen 100und 340 kg. Anhand von Vergleichen der Knochen derin Nordamerika noch heute lebenden Grizzly- undKodiakbären und der Höhlenbären kommt man auf einmaximales Gewicht für den Höhlenbären von1500 kg. Seine maximale Länge betrug 3 ½ m, dieWiderristhöhe 175 cm (Rabeder et al. 2000, 38).
Über die Ernährung des Höhlenbären gibt esverschiedene Ansichten. Es wird zwar immer wiederbehauptet, dass dieses Tier sich fast ausschließlich vonPflanzen ernährte und wahrscheinlich nicht mehr inder Lage war, sich räuberisch zu ernähren (Rabeder etal. 2000, 31), andererseits aber bleibt die Frage offen,
wie ein so riesiges Tier ausreichende Kalorien imSommer speichern konnte, um dann den Winterhindurch den Winterschlaf oder genauer dieWinterruhe zu überleben. Weiterhin ist das Gebiss desHöhlenbären das Gebiss eines Fleischfressers und füreinen Vegetarier, der auf Almenkräuter spezialisiertsein soll, ungeeignet. Der Höhlenbär war daherwahrscheinlich eher ein Allesfresser so wie derheutige Braunbär. Bei günstigen Gelegenheiten wirder in der Lage gewesen sein, unerfahrene oder krankegrößere Säugetiere zu töten und zu fressen. Weiterhinwird er dort, wo es Fische gab, wahrscheinlich so wiedie meisten Bären diese Nahrungsquelle ebenfallsgenutzt haben. Denkbar ist aber auch, dass er sichteilweise von Aas ernährt haben könnte, das er durchseine Größe, seine Kraft, sein Gebiss und seineKrallen jedem anderen Konkurrenten streitig machenkonnte. Und auch wenn er mit Sicherheit langsamerals andere Tiere gewesen ist, war er bestimmtschneller als ein Mensch.
Der Eisbär (Ursus maritimus) ernährt sich fastausschließlich von Robben. Obwohl man ihn berech-tigterweise an der atlantischen Küste Europas währendder letzten Eiszeit vermuten kann, sind bis heute keineReste von ihm im frankokantabrischen Bereichbekannt (Rouzaud 2002, 212). Die einzigen eiszeit-
Abb. 12 Bildliche Darstellungen von Tieren mit wahrscheinlichen Einschüssen. Ausgewählte Beispiele. 1 Les Trois Frères (Bégouën und Breuil 1958, 66, Abb. 70); 2 La Colombière (Graziosi 1956, Taf. 21); 3 Coug-nac (nach Clottes 1995, 123, Abb. 150); 4 Les Trois Frères (Bégouën und Breuil 1958, 35, Abb. 39); 5 Paglicci(Vigliardi 1979, 234).
2 3 1 4 5
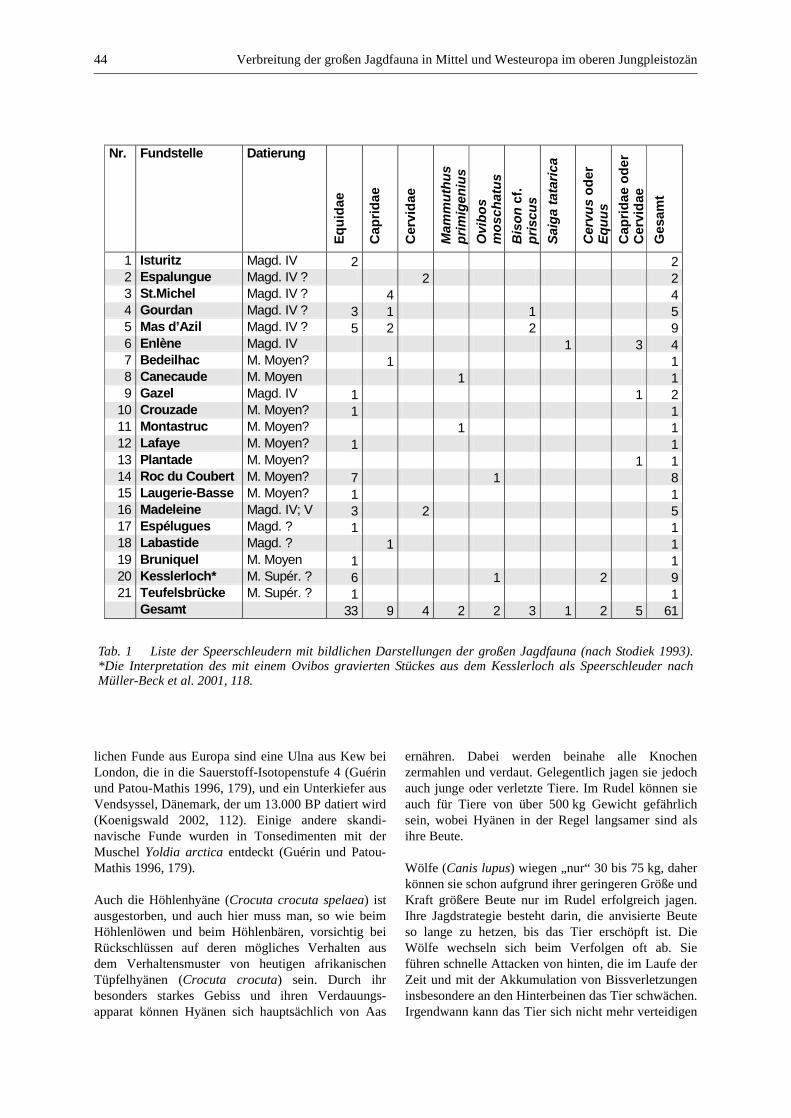
44 Verbreitung der großen Jagdfauna in Mittel und Westeuropa im oberen Jungpleistozän
lichen Funde aus Europa sind eine Ulna aus Kew beiLondon, die in die Sauerstoff-Isotopenstufe 4 (Guérinund Patou-Mathis 1996, 179), und ein Unterkiefer ausVendsyssel, Dänemark, der um 13.000 BP datiert wird(Koenigswald 2002, 112). Einige andere skandi-navische Funde wurden in Tonsedimenten mit derMuschel Yoldia arctica entdeckt (Guérin und Patou-Mathis 1996, 179).
Auch die Höhlenhyäne (Crocuta crocuta spelaea) istausgestorben, und auch hier muss man, so wie beimHöhlenlöwen und beim Höhlenbären, vorsichtig beiRückschlüssen auf deren mögliches Verhalten ausdem Verhaltensmuster von heutigen afrikanischenTüpfelhyänen (Crocuta crocuta) sein. Durch ihrbesonders starkes Gebiss und ihren Verdauungs-apparat können Hyänen sich hauptsächlich von Aas
ernähren. Dabei werden beinahe alle Knochenzermahlen und verdaut. Gelegentlich jagen sie jedochauch junge oder verletzte Tiere. Im Rudel können sieauch für Tiere von über 500 kg Gewicht gefährlichsein, wobei Hyänen in der Regel langsamer sind alsihre Beute.
Wölfe (Canis lupus) wiegen „nur“ 30 bis 75 kg, daherkönnen sie schon aufgrund ihrer geringeren Größe undKraft größere Beute nur im Rudel erfolgreich jagen.Ihre Jagdstrategie besteht darin, die anvisierte Beuteso lange zu hetzen, bis das Tier erschöpft ist. DieWölfe wechseln sich beim Verfolgen oft ab. Sieführen schnelle Attacken von hinten, die im Laufe derZeit und mit der Akkumulation von Bissverletzungeninsbesondere an den Hinterbeinen das Tier schwächen.Irgendwann kann das Tier sich nicht mehr verteidigen
Tab. 1 Liste der Speerschleudern mit bildlichen Darstellungen der großen Jagdfauna (nach Stodiek 1993).*Die Interpretation des mit einem Ovibos gravierten Stückes aus dem Kesslerloch als Speerschleuder nachMüller-Beck et al. 2001, 118.
Nr. Fundstelle Datierung
Eq
uid
ae
Cap
rid
ae
Cer
vid
ae
Mam
mu
thu
s p
rim
igen
ius
Ovi
bo
s m
osc
hat
us
Bis
on
cf.
p
risc
us
Sai
ga
tata
rica
Cer
vus
od
er
Eq
uu
s
Cap
rid
ae o
der
C
ervi
dae
Ges
amt
1 Isturitz Magd. IV 2 2 2 Espalungue Magd. IV ? 2 2 3 St.Michel Magd. IV ? 4 4 4 Gourdan Magd. IV ? 3 1 1 5 5 Mas d’Azil Magd. IV ? 5 2 2 9 6 Enlène Magd. IV 1 3 4 7 Bedeilhac M. Moyen? 1 1 8 Canecaude M. Moyen 1 1 9 Gazel Magd. IV 1 1 2
10 Crouzade M. Moyen? 1 1 11 Montastruc M. Moyen? 1 1 12 Lafaye M. Moyen? 1 1 13 Plantade M. Moyen? 1 1 14 Roc du Coubert M. Moyen? 7 1 8 15 Laugerie-Basse M. Moyen? 1 1 16 Madeleine Magd. IV; V 3 2 5 17 Espélugues Magd. ? 1 1 18 Labastide Magd. ? 1 1 19 Bruniquel M. Moyen 1 1 20 Kesslerloch* M. Supér. ? 6 1 2 9 21 Teufelsbrücke M. Supér. ? 1 1
Gesamt 33 9 4 2 2 3 1 2 5 61

Jäger und Beute in Europa am Ende der letzten Eiszeit 45
(Guthrie 1990, 287–289). Diese Jagdtechnik ist soerfolgreich, dass in Nordamerika, Europa und imnördlichen Asien kein Tier sich vor Wölfen sicherfühlen kann. Einzige Verteidigung bleibt dabeientweder in der Gruppe zu bleiben oder die Flucht.Gegen eine kompakte Herde von Bisons oderRentieren haben Wölfe keine Chance. Wenig erfolg-reich sind sie auch gegen Tiere wie z. B. Elch,Steinbock oder Gämse (Abb. 13 Mitte unten), welchein ihrem Fluchtverhalten bestens die Gegebenheitendes Geländes ausnutzen können.
Eine solche Jagdstrategie, die auf das Hetzen derBeute abzielt, kann vielleicht auch bei der gezieltenJagd auf junge Elefanten, Mammuts und Nashörnererfolgreich gewesen sein (Abb. 13 rechts unten) undeventuell einen Beitrag zu deren Aussterben am Endedes Pleistozäns und im Holozän geleistet haben. DaElefanten und Nashörner heute kaum in den gleichenGebieten wie Wölfe verbreitet sind, ist dieseÜberlegung rein spekulativ. Schließlich muss manerwähnen, dass Wölfe besonders im Winter als Jägererfolgreich sind. Das Verhältnis von ihrem Gewichtzur Breite der Pfoten fällt so aus, dass sie bei dickerSchneedecke nicht einsinken. In dieser Situationhaben Wölfe einen Vorteil gegenüber den meistenihrer Beutetiere.
Als weitere große Raubkatzen sollte man hier noch dieLeoparden (Panthera pardus) erwähnen. Diese Tieresind deutlich kleiner als die Löwen (ihr Gewichtbeträgt maximal 90 kg), und waren bis zum Beginndes 20. Jahrhunderts noch sehr weit verbreitet, u. a. imKaukasus, Vorderasien und Nordafrika (Heptner undSludskij 1980, 187). In den Fundstellen Europas istder Leopard zwar sehr selten, wurde aber immerwieder nachgewiesen (u. a. Dubois und Stehlin 1933,91ff.). Eine Darstellung aus der Höhle von Chauvet(Chauvet et al. 1996, 34) wurde als die erste sichereDarstellung eines Leoparden interpretiert.
Raubtiere in der paläolithischen Kunst
Raubtiere wurden nicht besonders häufig dargestellt,aber ihre Bilder sind zahlreicher als man denkt.Insgesamt kommt man auf ungefähr 120 Darstel-lungen von Raubkatzen, wobei nur wenige anhand derDetails (u. a. Schwanzspitze mit Quaste) mitSicherheit als Löwe interpretiert werden können27.Bei manchen Darstellungen von Raubkatzen, z. B. inChauvet oder in La Vache, kommt, da mehrereIndividuen dargestellt wurden, ebenfalls nur der Löweals Rudeltier für die letzte Eiszeit in Europa in Frage.
Abb. 13 Verhalten von verschiedenen Beutetieren, wenn sie von Löwen (oben) oder von einem Rudel von Wöl-fen (unten) angegriffen werden (verändert nach Guthrie 1990, 288, Abb. 10.12).Einige Verhaltensweisen können nur als Hypothese betrachtet werden, da diese Tiere heute nicht mehr in dengleichen Regionen leben. Links: Löwe und Steppenbison, Wolf und Steppenbison; Mitte: Löwe und Elch (hypo-thetisch), Wolf und Elch; Rechts: Löwe und Nashorn, Wolf und Nashorn mit Nachwuchs (hypothetisch).
27 Schon 1967 konnte Michel Rousseau (Seite 173) 81 von ihm als sicher angenommene Darstellungen von Feli-den zählen. Diesen sind weitere Funde zuzurechnen, darunter die 36 Darstellungen aus der Höhle von Chauvet(Chauvet et al. 1996, 105).

46 Verbreitung der großen Jagdfauna in Mittel und Westeuropa im oberen Jungpleistozän
Obwohl die bekanntesten Darstellungen diejenigensind, die besonders „wild“ aussehen, wie z. B. in LaMarche (Abb. 14, 2; Pales und de Saint Pereuse 1969),sind die meisten Raubkatzen als ruhige Tiere darge-stellt. Sie trippeln über die Wände von Höhlen oderauf Steinen und Knochen genau so wie andere Tiere(Serangeli 2004). Manchmal sind zwar Raubkatzenfrontal dargestellt, wie z. B. in Les Trois Frères(Bégouën und Breuil 1958, 10–11, Pl. VIII und IX),dies ist aber an sich keine „bedrohliche“ Haltung. Inder Höhle von Les Trois Frères sind ebenfalls auchEulen, eine menschlich wirkende Maske, Bären undder „Dieu Cornu“ frontal dargestellt worden (Bégouënund Breuil 1958, 17, Pl. XI, 35, 64, Pl. XX). BeiLebewesen wie Raubtieren, Raubvögeln undMenschen, die die Augen vorne und nicht seitlichhaben, ist eine frontale Darstellung schon aus techni-scher Sicht her eher etwas Natürliches.
Es ist weiterhin interessant zu vermerken, dassKonzepte wie „Töten“, „Aggression“, „Fleisch-fresser“, die man in Zusammenhang mit Raubtierenbestens u. a. von der korinthischen Vasenmalerei, von
den römischen Arenaspielen oder von der mittelalter-lichen christlichen Tradition kennt, sich in denbildlichen Darstellungen des Jungpaläolithikums nichtwiederfinden lassen. Auch die Tatsache, dass inSüdwestdeutschland Raubtiere die größere Gruppeunter den dargestellten Tieren sind, muss nichtzwangsläufig mit „Aggression“ zu tun haben. Auchdas Mischwesen aus Mensch und Raubkatze aus demHohlenstein-Stadel, der so genannte „Adorant“ (Porr2001, 100) aus dem Geißenklösterle und der neuer-liche Fund eines kleinen „Löwenmenschen“ aus demHohle Fels (Conard et al. 2003, 26), sind qualitativnicht anders dargestellt als andere Mischwesen vonMensch und Rind, Mensch und Rentier oder Menschund Steinbock (Serangeli 2004). Es sind, bis jetzt,auch keine Darstellungen bekannt, in denenRaubkatzen beim Angriff auf ein anderes Tier oder beider Gefährdung eines Menschen zu sehen sind28.Dadurch scheint die Theorie einer u. a. vonRaubkatzen ausgehenden Botschaft von „Kraft undAggression“ für das Aurignacien Südwest-deutschlands, so wie sie Joachim Hahn 1986 präsen-tiert hat29, nicht mehr haltbar. Interessant ist
Abb. 14 Ausgewählte Löwendarstellungen. 1 Elfenbeinfigur vom Vogelherd (Hahn 1986, 207); 2 Gravierungauf Steinplatte von La Marche (Pales und de Saint Pereuse 1969, Pl. 10); 3 Gravierung auf Steinplatte ausRiparo Tagliente (Bosinski 1994, 64); 4 Löwengravierung auf Knochen aus La Vache (Bosinski 1994, 60);5 Löwenmensch aus dem Hohlenstein-Stadel (Hahn 1986, 247, Taf. 17).
1 2 3 4 5
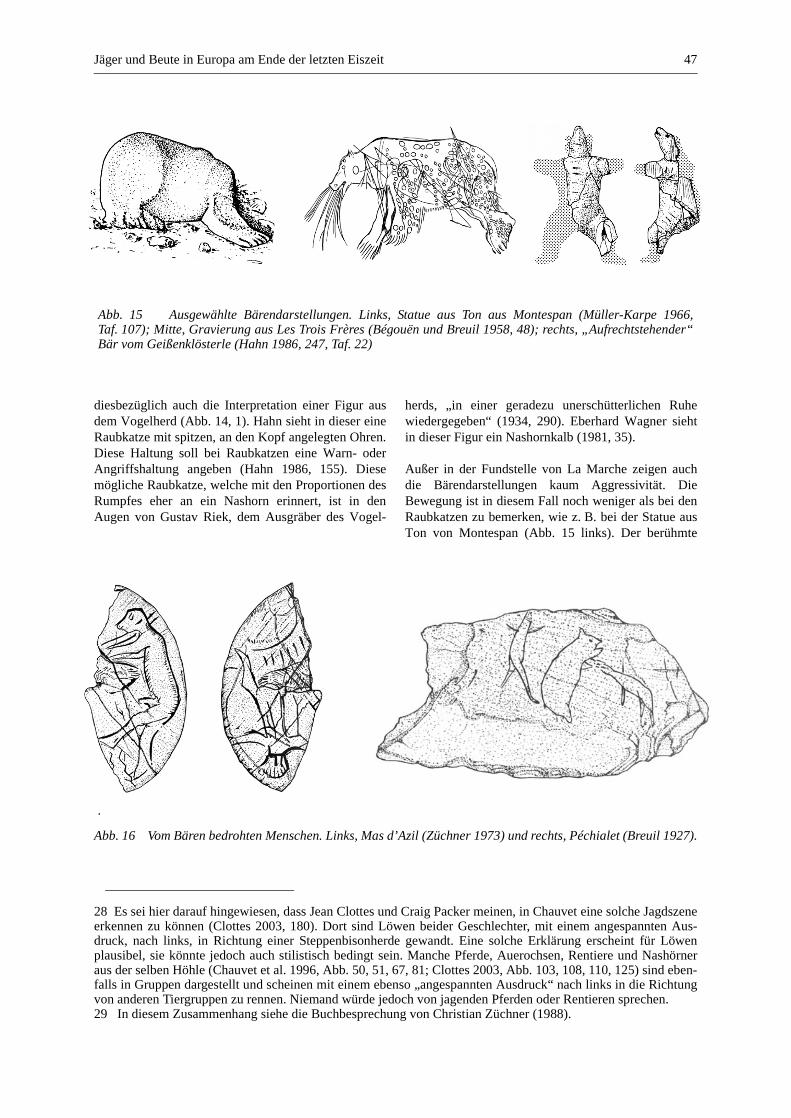
Jäger und Beute in Europa am Ende der letzten Eiszeit 47
diesbezüglich auch die Interpretation einer Figur ausdem Vogelherd (Abb. 14, 1). Hahn sieht in dieser eineRaubkatze mit spitzen, an den Kopf angelegten Ohren.Diese Haltung soll bei Raubkatzen eine Warn- oderAngriffshaltung angeben (Hahn 1986, 155). Diesemögliche Raubkatze, welche mit den Proportionen desRumpfes eher an ein Nashorn erinnert, ist in denAugen von Gustav Riek, dem Ausgräber des Vogel-
herds, „in einer geradezu unerschütterlichen Ruhewiedergegeben“ (1934, 290). Eberhard Wagner siehtin dieser Figur ein Nashornkalb (1981, 35).
Außer in der Fundstelle von La Marche zeigen auchdie Bärendarstellungen kaum Aggressivität. DieBewegung ist in diesem Fall noch weniger als bei denRaubkatzen zu bemerken, wie z. B. bei der Statue ausTon von Montespan (Abb. 15 links). Der berühmte
28 Es sei hier darauf hingewiesen, dass Jean Clottes und Craig Packer meinen, in Chauvet eine solche Jagdszeneerkennen zu können (Clottes 2003, 180). Dort sind Löwen beider Geschlechter, mit einem angespannten Aus-druck, nach links, in Richtung einer Steppenbisonherde gewandt. Eine solche Erklärung erscheint für Löwenplausibel, sie könnte jedoch auch stilistisch bedingt sein. Manche Pferde, Auerochsen, Rentiere und Nashörneraus der selben Höhle (Chauvet et al. 1996, Abb. 50, 51, 67, 81; Clottes 2003, Abb. 103, 108, 110, 125) sind eben-falls in Gruppen dargestellt und scheinen mit einem ebenso „angespannten Ausdruck“ nach links in die Richtungvon anderen Tiergruppen zu rennen. Niemand würde jedoch von jagenden Pferden oder Rentieren sprechen. 29 In diesem Zusammenhang siehe die Buchbesprechung von Christian Züchner (1988).
Abb. 15 Ausgewählte Bärendarstellungen. Links, Statue aus Ton aus Montespan (Müller-Karpe 1966,Taf. 107); Mitte, Gravierung aus Les Trois Frères (Bégouën und Breuil 1958, 48); rechts, „Aufrechtstehender“Bär vom Geißenklösterle (Hahn 1986, 247, Taf. 22)
.
Abb. 16 Vom Bären bedrohten Menschen. Links, Mas d’Azil (Züchner 1973) und rechts, Péchialet (Breuil 1927).

48 Verbreitung der großen Jagdfauna in Mittel und Westeuropa im oberen Jungpleistozän
gravierte Bär von Les Trois Frères, der sogar Blutspuckt (Abb. 15 Mitte), ist ein sterbendes, keinbedrohliches Tier. Einzig dem aufgerichteten Bär vomGeißenklösterle (Abb. 15 rechts) könnte eine bedroh-liche Haltung zugeschrieben werden (Hahn 1986,191), wobei betont werden muss, dass Bären meistensauf den hinteren Beinen stehen, um Ausschau zuhalten oder um besser die Herkunft eines Geruches zuorten, was die gehobene Kopfhaltung zu suggerierenscheint. Als weitere denkbare Möglichkeit sei hier aufdas Besteigen eines Baumes hingewiesen. Zu einersolchen Aktion würde sowohl die Haltung des Kopfesals auch der Pranken passen. Nur die unvollständigeDarstellung von Mas d’Azil (Abb. 16, links; Züchner1973) und die Gravierung von Péchialet (Abb. 16rechts; Breuil 1927) zeigen einen angreifenden Bär. Esmuss jedoch erwähnt werden, dass die Szene ausPéchialet zur Zeit der Publikation des Fundes im Jahr1927 schon seit ca. 100 Jahren bekannt war und kaumInformationen über die Lage oder die Bergung derSchieferplatte zu finden sind.
Zusammenfassend kann man sagen, dass Raubtierestilistisch kaum anders behandelt wurden als andereTiere. Unter den Raubtieren ist nur der Bär in zweiDarstellungen beim Angriff auf einen Menschendargestellt. Das ist um so interessanter, wenn manbedenkt, dass Rinder häufiger als Gefährdung odersogar beim Töten von Menschen dargestellt wurden,so wie bei den Darstellungen in Lascaux, Roc de Sers,Villars und Font-de-Gaume.
Wenn man weiterhin annimmt, dass für die Erzeugerder bildlichen Darstellungen des Jungpaläolithikumsdiese eine religiöse, magische oder schamanischeFunktion mit Auswirkungen auf die Realität hatten,sollte es nicht verwundern, dass man keine „Aggressi-vität“, sondern möglicherweise eher eine besondereArt der „Harmonie“ mit den Bildern darstellen wollte(siehe dazu Serangeli 2004).

Die große Jagdfauna der letzten Eiszeit in Europa
Unpaarhufer (Perissodactyla)
Die Unpaarhufer zeichnen sich nach den Worten vonVolker Storch und Ulrich Welsch (1993, 446) durchfolgende Eigenschaften aus: „Die mittleren Digitidominieren, die anderen bilden sich verschiedenartigzurück. Gebiß vollständig, bisweilen fehlen die
Eckzähne. Prämolaren und Molaren von ungefähr dergleichen Größe und Form. Große Pflanzenfresser.Mehrzahl von Ausrottung bedroht. Tapirus, Rhino-ceros, Dicerorhinus, Ceratotherium, Diceros, Equus.“
Pferd (Equus ferus)
Englisch – Horse; Französisch – Cheval; Spanisch – Caballo; Katalanisch – Cavall; Italienisch – Cavallo; Portu-giesisch – Cavalo. Im Deutschen findet man neben der Bezeichnung (Wild-) Pferd auch gelegentlich, insbesondere in Bezug aufrezente Wildpferde, die Bezeichnung Tarpan oder Przewalskipferd.Ordnung: Perissodactyla Owen 1848; Familie: Equidae Gray 1821; Gattung: Equus Linnaeus 1758; Art: Equusferus Boddaert 1785.Unterarten: In historischer Zeit hat man hauptsächlich drei Gruppen unterschieden, nämlich das Przewalski-Pferd (E. f. przewalskii Poljakov 1881, letzte Beobachtung in freier Wildbahn 1967), das Südrussische Steppen-Wildpferd (E. f. gmelini Antonius 1912, ausgestorben 1918) und den Waldtarpan (E. f. silvaticus Vetulani 1928,ausgestorben 1814). Mit dem Namen Equus ferus werden hier, entsprechend der Arbeit von Birgit Cramer(2002, 141), auch die zahlreichen abweichenden Bezeichnungen für das eiszeitliche Wildpferd (u. a. E. arceliniGuadelli 1986, E. antunensi Cardoso und Eisenmann 1989) miteingeschlossen. Vorwiegend verwendete Literatur: Boyd und Houpt 1994; Girard 1995; Guthrie 1990; Heptner et al. 1966, 858–874; Meadow und Uerpmann 1991; Uerpmann 1987; Volf 1996.
Habitat: E. f. przewalskii und E. f. gmelini lebten inder offenen Steppe, Waldsteppe und Halbwüste. E. f.silvaticus, das manchmal als unterdrückte Form desSteppen-Wildpferdes angesehen wird, lebte dagegenin Wäldern (Heptner et al. 1966, 865).
Verbreitung: Hinweise auf Wildpferde gibt es vonder Antike bis ins 19. Jahrhundert von der IberischenHalbinsel, aus Frankreich, Deutschland, Polen und derehemaligen Sowjetunion bis zur mongolischen Steppe(Heptner et al. 1966) (Abb. 17). Die Einführung vonHaustieren sowie die Zerstörung von Wald und Steppefür Viehzucht und Ackerbau führte schon früh zurAusrottung dieser Art in Westeuropa, wobei mancheAutoren erwogen haben, dass Equus ferus schon vorder Einführung von domestizierten Pferden auf derIberischen Halbinsel ausgestorben sein könnte (Schule1969, 92). Im frühen Mittelalter, aber möglicherweisenoch früher, lebte E. f. silvaticus nicht mehr imwestlichen Mitteleuropa. Die letzten Bestände lebtenim Urwaldmassiv Belowesher (Białowieża) in Polenund in Weißrussland bis zum Ende des 18. Jahrhun-derts. Die letzten Exemplare wurden 1814 im Gebietvon Königsberg erlegt.
E. f. gmelini kam in historischer Zeit nur im europäi-schen Teil der ehemaligen Sowjetunion, im Gebietzwischen den Flüssen Prut im Westen und dem Uralim Osten, vielleicht bis zur Wolga vor. Möglicher-weise lebte es auch in Rumänien. Es ist unklar,inwieweit sein Verbreitungsgebiet sich über Mittel-und Westeuropa erstreckte. Wahrscheinlich ist aber,dass diese Tiere im Westen wenig verbreitet warenund dort hauptsächlich der Waldtarpan (E. f. silva-ticus) lebte.
E. f. przewalskii wurde bis zuletzt stark bejagt, seies wegen des Fells und Fleisches, sei es als „Sport“(Heptner et al. 1966, 869f.). Bis zum 18. Jahrhundertkam es jedoch noch in den Steppen zwischen demFluss Ural und der Mongolei und Nordchina vor. Im19. Jahrhundert ging durch die massive Veränderungder Steppe (planmäßig geführte Trockenlegung vonSumpfgebieten, Verbreitung von Ackerbau undViehzucht) die Zahl dieser Tiere rasch zurück. Dieletzte Beobachtung von Przewalski-Pferden in derNatur durch einen Zoologen fand am 27. Mai 1967 imWesten der Mongolei an der Grenze zu China statt(Girard 1995, 35).

50 Verbreitung der großen Jagdfauna in Mittel und Westeuropa im oberen Jungpleistozän
In der Steppenregion zwischen den Flüssen Ural undWolga, wo sowohl E. f. gmelini als auch E. f.przewalskii vorkamen, kam es zu einer Übergangs-zone, in der sich die zwei Unterarten vermutlich starkvermischt haben.
In der Eiszeit kam das Wildpferd (Equus ferus) inganz Eurasien von der portugiesischen Atlantikküstebis zur Küste Asiens am Stillen Ozean vor. Auch inAlaska belegen mehrere datierte Reste die Präsenzdieser Art während des ganzen Jungpaläolithikums(Guthrie 1990, 243).
Körpermaße von E. f. przewalskii: Körperlänge220–280 cm; Widerristhöhe 120–146 cm; Gewicht200–300 kg oder sogar 450 kg (Girard 1995, 25). E. f.gmelini (Südrussisches Steppen-Wildpferd) soll etwaskleiner und E. f. silvaticus (Waldtarpan) noch kleinergewesen sein.
Körperbeschreibung: Schweif mit langen Haarenbewachsen, der Aalstrich30 ist meistens vorhanden,die Mähne ist kurz und aufrechtstehend. Die Ohrensind kurz, aufrecht stehend und spitz. Das gesamteErscheinungsbild ist gedrungen und stämmig. DerKopf ist groß und schwer, die Schnauze ist kurz. DerHals ist kurz und dick, die Brust ist breit, die Rippensind steil und der Rücken ist gerade. Kräftige Beine
mit hohen Hufen. E. f. przewalskii ist im gesamtenKörperbau schwerer und kräftiger als E. f. gmelini undE. f. silvaticus.
Fell: Sehr kurz im Sommer, recht lang bis welligim Winter. Bei den südrussischen Steppenwildpfer-den scheint es besonders lang gewesen zu sein, so dasses eher wie ein Pelz aussah. Die Gesamtfärbung ist beiE. f. gmelini und E. f. silvaticus gelb oder fuchsrötlichbis gelb, bei E. f. przewalskii brauner. Der Bauchscheint stets heller gewesen zu sein als das Rücken-fell. Die Beine sind schwarz oder schwarzbraun biszum Knie, in diesem Bereich können zebraartigeStreifen auftreten (Girard 1995, 29, Abb. 4). Bei man-chen Individuen jedoch war die dunkle Verfärbungnur an den Hufen vorhanden. Die Schnauze war mei-stens heller, die Mähnen, der Aalstrich und derSchwanz auch schwarz bis braun. Bei manchen Indivi-duen war der Aalstrich nicht oder nicht während desganzen Jahres zu erkennen.
Das Fell scheint im Winter bei E. f. gmelini undE. f. silvaticus meistens eine etwas trübere Färbunggehabt zu haben als im Sommer, bei E. f. przewalskiischeint eine etwas hellere Farbe im Winter die Regelgewesen zu sein.
Druck auf den Boden: Besonders hoher Druck, ca.760 g/cm². Bei einer hohen Schneedecke oder Eis wardas gegenüber dem des Wolfs (140–160 g/cm²) eingroßer Nachteil.
Paarungszeit: Juni. Tragzeit: 11 Monate. Setzzeit:Mai.
Natürliche Lebenserwartung: bis 30 Jahre. Nahrung: Gräser und Kräuter.Verhalten: Im Freien lebte Equus ferus meistens in
Gruppen von 10 bis 15 Individuen unter der Führungeines Leithengstes. Dieser beschützte die Gruppesowohl vor Gefahren als auch vor anderen Hengsten.Equus ferus war besonders schnell und von beson-derer Ausdauer (Heptner et al. 1966, 870). EineVermischung zwischen Wild- und Hauspferden fandin der russischen Steppe häufig statt. In Europa ist diewiederholte Verwilderung von Hauspferden bekannt.
Bei schlechten klimatischen Bedingungen konntenWildpferde weite Gebiete in kurzer Zeit durchziehen.Im Winter gruben sie mit den Hufen im Schnee, um anFutter zu kommen. Man nimmt an, dass diese Tiere inGebieten mit einer maximalen Schneetiefe von 60 bis70 cm leben konnten (Girard 1995, 39). In besondersschneereichen und frostigen Wintern sowie bei Eiskamen diese Tiere in großer Zahl um. Besonderswichtig war im Sommer das Trinken. Die Trocken-legung vieler Sumpfgebiete in der Steppe sowie dievon Viehzüchtern übernommene Kontrolle derübrigen Wasserquellen führten zum Rückgang dieserArt.
Abb. 17 Verbreitung von Equus ferus im oberenPleistozän (dunkelgrau) (nach Uerpmann 1987, 36)und letztes bewohntes Gebiet von Equus ferus prze-walskii im 20. Jahrhundert (nach Volf 1996, 31).
30 Über die Mitte des Rückens verlaufender, von der Mähne bis zur Schwanzwurzel durchgehender Strich.

Pferd (Equus ferus) 51
Da es überall gejagt und verfolgt wurde, war Equusferus besonders scheu. Menschen zu Fuß konnten bisca. 200 m herankommen, wobei die Regel eher 500bis 600 m gewesen sein soll (Girard 1995, 38). EinReiter wurde schon auf einigen Kilometer Distanz alsGefahr und als Anlass zur Flucht angesehen.Erwachsene Wildpferde waren kaum zu zähmen,daher wurden sie oft getötet.
Natürliche Feinde: In den letzten Jahrhunderten istder Mensch sicher der größte Feind gewesen.Inwieweit Wölfe in der Lage waren, gesunde,erwachsene Wildpferde zu jagen, ist unklar. Durchihre Geschwindigkeit, Ausdauer und die Gefähr-lichkeit ihrer Tritte kann man davon ausgehen, dasssie für Wölfe in einer offenen Landschaft keineeinfache Beute gewesen sein können. Der Vergleichmit der afrikanischen Savanne, wo Zebras zur regel-mäßigen Beute von Löwen gehören, lässt dagegen alswahrscheinlich erscheinen, dass Pferde zu denüblichen Beutetieren der eiszeitlichen Großkatzengehört haben.
Klimatische Aussage: Das Auftreten von verein-zelten Hinweisen auf Pferde lässt kaum eine klimati-sche oder ökologische Aussage zu. Pferde kommenzwar zahlreich in der offenen Steppe vor, da aber E. f.silvaticus im Wald lebte, ist nicht auszuschließen, dasswährend der Eiszeit Equus ferus teilweise ebenfalls imWald und in Galeriewäldern entlang der Flüsse anzu-treffen war. Die Tatsache, dass Kadaver in den Dauer-frostböden von Jakutien und Alaska entdeckt wurden,die in die Eiszeit datieren (z. B. Dukarskoje, 26.000Jahre), und dass sie noch im Neolithikum in Südspa-nien vorkamen (Uerpmann 1976), zeigt, wie anpas-sungsfähig diese Art war. Erst das Auftreten vonzahlreichen Pferderesten weist auf eine offene Land-schaft hin. Manche Fundstellen, wie beispielsweisedie Höhle Paglicci (Sala 1985), belegen durch die sta-tistisch relevante Abnahme von Equus ferus zugun-sten von Equus hydruntinus eine trockener werdendeLandschaft. Während des zweiten Kältemaximumsscheint auch Equus ferus in dem Gebiet zwischen demskandinavischen und dem alpinen Eisschild seltener
zu sein (Koenigswald 2002, 63f.), wobei das Pferd umca. 19.000 BP in Wiesbaden-Igstadt nachgewiesen ist(Street und Terberger 2000).
Darstellungen aus dem Jungpaläolithikum
Es wird immer wieder betont, dass das Pferd das amhäufigsten dargestellte Tier sei und dass es eine„tiefere“ Bedeutung haben soll (siehe Delporte 1990,225ff.). Dabei muss man bedenken, dass das Pferd,von manchen Raubtieren abgesehen, das einzigegrößere Säugetier ist, das fast in ganz Europa undwährend des ganzen Jungpaläolithikums vorkam(Abb. 19). Das Wildpferd wird mit unterschiedlichenTieren dargestellt. In Südspanien und Italien wird esneben Gravierungen und Malereien von Auerochsenund Rothirschen präsentiert, in Frankreich undDeutschland dagegen kommt es in Kontexten vor, indenen ausschließlich kälteliebende Fauna dargestelltist, z. B. in Gönnersdorf, im Vogelherd und inRouffignac. Die Darstellungen von Pferden sindweiterhin auch in Regionen vorhanden, in denenKnochen kaum oder gar nicht erhalten sind, so wie inMittelspanien (Ripoll und Municio 1999) oder in derFundstelle Döbritz (Hanitzsch 1972).
In diesem Zusammenhang sei hier auf manchehervorragende Darstellungen (Abb. 18) insbesondereaus dem Magdalénien hingewiesen. Die Details,insbesondere die des Felles, sind manchmal so präzisedargestellt worden, dass einige Autoren meinen, unter-schiedliche Rassen erkennen zu können (u. a. Capitanet al. 1924). Als sicher gilt, dass manche Darstel-lungen so detailliert sind, dass man das Winter- oderSommerfell des Tieres erkennen kann (Schmid 1984;Crémadès 1997).
Reste von Pferden aus dem Jungpaläolithikum
Wie schon oben erwähnt, kam das Pferd in fast ganzEuropa und während des ganzen Jungpaläolithikumsvor. Es war daher mit allen größeren Säugetierenvergesellschaftet. Obwohl es in den Grimaldi-Höhlen
Abb. 18 Ausgewählte Pferdedarstellungen. 1 Lascaux; 2 Niaux; 3 Ekain; 4 Le Portel (Schmid 1984, 156 und158).
1 2 3 4

52 Verbreitung der großen Jagdfauna in Mittel und Westeuropa im oberen Jungpleistozän
in allen Schichten vorkommt, sind hier jeweils nurwenige Reste vorhanden (Villeneuve und Boule 1910,178). In Nerja, La Vache, Les Eglises, Rhodes II,Arene Candide, Riparo Tagliente, Grotta di Fumaneund in den jungpaläolithischen slowenischenFundstellen (siehe die Faunenlisten, S. 215–255) ist esebenfalls stark unterrepräsentiert oder es fehlt sogarganz. In fast allen diesen Fundstellen ist dagegen derSteinbock sehr gut vertreten, oft sogar das häufigsteTier.
Diskussion
Obwohl Knochenreste von Wildpferden in fast ganzEuropa zu finden sind, gibt es Regionen, in denendiese stärker vertreten und andere, wo sie stark unter-repräsentiert sind oder sogar fehlen. Dafür scheinenunterschiedliche Gründe verantwortlich zu sein.– Die Topographie. Das Przewalski-Pferd miedGebirge (Heptner et al. 1966, 863). Man kann daherdavon ausgehen, dass Equus ferus während der Eiszeitebenfalls lieber in Ebenen oder auf Plateaus gelebt hat.Hier kamen seine ausgeprägten Fähigkeiten beimschnellen und ausdauernden Laufen am besten zurGeltung. Weiterhin haben Ebenen gegenüber engenSchluchten, steilen Hängen und felsigen Küsten denVorteil, dass sie einen weiten Blick ermöglichen unddamit den Tieren erlauben, eine ausreichende Flucht-distanz zwischen sich und potentiellen Gefahreneinzuhalten. In Landschaften, wo sowohl felsigerUntergrund als auch breite, offene Ebenen vorhandensind, sind Steinbock und Pferd gleichzeitig undteilweise gleich stark repräsentiert. Beispiele dafürsind die Schicht 19b von Paglicci, Italien (Sala 1985)und die Schicht 8 von Parpalló, Spanien (Davidson1989, 66, Tabla 7.9). In Nerja, einer Höhle, die sich ineiner steilen Küstenlandschaft in Südspanien befindet,ist die Zahl der Pferde, insbesondere am Ende desJungpaläolithikums, auffällig gering (Jordá et al. 2003,86). Auch hier ist die Wirkung der Topographiedeutlich. – Das Klima. Wie beim Przewalski-Pferd und beimasiatischen Wildesel (Heptner et al. 1966, 870 und856) kann man auch für Equus ferus annehmen, dassraue Winter eine große Gefahr für dessen Bestandwaren. Eis und eine hohe Schneedecke haben eszumindest im Winter aus den Gebirgsregionenverdrängt. Die eisige, von den Gletschern herabsin-kende Luft kann während der Eiszeit bis weit in die
Ebenen das Klima beeinflusst haben. Im Sommerjedoch, bei jahreszeitlichen Wanderungen, kam Equusferus auch in höher gelegenen Regionen vor, so z. B.im Massif Central (Fontana 1998) oder im Tal desFucino in den Apenninen (Palma di Cesnola 1993,222; Radimilli 1956). In der Fundstelle von Solutré isteine solche Wanderung besonders gut dokumentiert(Turner 2002). In Zentralspanien sind kaum jungpa-läolithische Fundstellen mit Fauna publiziert31. Daaber Pferde zahlreich in den Höhlen und an den Felsendieser Region dargestellt wurden, ist anzunehmen,dass auch hier Pferde regelmäßig, wenn nicht sogarzahlreich vorhanden waren.– Die Rückzugsgebiete und die „Konkurrenz“. DieEbenen, in die sich die Pferde zumindest im Winterzurückzogen, mussten offen sein. Ein dichter Wald, indem z. B. Rehe, Rothirsche und Wildschweine ingroßer Anzahl leben konnten, bot kaum Lebensraumfür diese Tiere. Das gleiche gilt für einen lichten Waldmit sumpfigen Gebieten, in denen Rinder, Rothirscheund Elche, jedoch kaum Pferde leben können32.
Sieht man diese drei Aspekte zusammen, ver-wundert es nicht, dass nördlich der Apenninen so gutwie keine Pferdeknochen aus Fundstellen des Epigra-vettien (Bridault und Chaix 2002, 55; Palma diCesnola 1993; Sala 1985; Masseti et al. 1995, 90) undnur ein einziges Pferd für das Aurignacien in RiparoFumane publiziert sind (Cassoli und Tagliacozzo1994; Gurioli 2002). Aufgrund der Alpengletscher undder im Vergleich mit anderen Regionen Europasreicheren Niederschläge (Florineth und Schlüchter2000), waren die Flüsse in der Po-Ebene im Sommerimmer reich an Wasser33. Wegen des geringen oderkaum vorhandenen Gefälles der Ebene (wie schonerwähnt, betrug das Gefälle heute wie während derEiszeit ca. 1 m Höhenunterschied auf 3 km Distanz)gab es ständig Überschwemmungen, mit allentypischen Folgen wie beispielsweise entwurzelteBäume, Sümpfe, Altwasserarme usw. Im Winterfroren die stillen Wasserflächen zu. Im Frühling undHerbst kann man von häufigem, starkem Nachtfrostausgehen. Das sind Voraussetzungen, die in denEbenen für Elche, Rinder und Rothirsche, in denBergen für Steinböcke und Gämsen, jedoch nicht fürPferde günstig sind. Dabei darf man jedoch, wie schonerwähnt, nicht vergessen, dass gerade infolge derstarken Sedimentation in der Po-Ebene die jungpaläo-lithischen Fundstellen mehrere zehn Meter unter derheutigen Oberfläche liegen (u. a. Mussi 2001, 289;
31 Die Höhle von Jarama VI ist eine der wenigen Ausnahmen. Hier konnten nur wenige Knochen, darunter einPferd, bestimmt werden (Arribas und Jordá 1999).32 Manche Pferderassen leben zwar heute in Sumpfgebieten, wie z. B. in der Camargue in Frankreich, das Klimaist aber heute ganz anders als das Klima der Eiszeit.33 In diesem Zusammenhang darf nicht unerwähnt bleiben, dass noch heute die Region um die Po-Ebene (Alpen,Voralpen und Appennino settentrionale) eine der niederschlagsreichsten Regionen Europas mit Spitzenwerten inden Alpen von über 2000 mm pro Jahr ist.

Pferd (Equus ferus) 53
Palma di Cesnola 1993). Nur unter besonderenUmständen finden sich Knochen aus dieser Zeit indieser Region. Die Fundstelle Settepolesini di Bodeno,in der Nähe von Ferrara in der Po-Ebene, ist einesolche Ausnahme (Gallini und Sala 2000 und 2001;Sala und Gallini 2002). Hier ist zwar das Pferd belegt,dessen Verhältnis zu den Faunenresten aus dem
oberen Jungpleistozän ist jedoch unklar (mündlicheMitteilung von Marco Marchetti, Universitá diFerrara). Das zeigt aber, dass neue Fundstellen denMangel an Pferden in Norditalien korrigieren könnten.Eine topographische und klimatische Situation, wie inden Alpen und in der Po-Ebene während der letztenEiszeit, scheint in Europa einmalig gewesen zu sein.
Abb. 19 Verbreitung von Equus ferus in Europa. Dunkel sind die Gebiete hervorgehoben, in denen das Pferdvorhanden, jedoch nie stark repräsentiert ist (oder wo es aufgrund der Topographie angenommen werden kann),hell sind die Gebiete hervorgehoben, in denen das Pferd zum Teil stark repräsentiert ist, frei sind die Gebiete indenen das Pferd nicht oder durch einzelne Knochen für das Jungpaläolithikum belegt ist. Die in diesem Kapitel erwähnten paläolithischen Fundstellen mit Resten (Dreieck) oder Darstellungen (Kreis)von Pferden dienen hier nur zur Orientierung und sind keineswegs repräsentativ, da sonst Hunderte Fundstellenmit Fauna (u. a. Vogelherd) und Darstellungen (u. a. Gönnersdorf) erwähnt werden müssten, insbesondere inKantabrien, den Pyrenäen und im Périgord.1 La Peña del Candamo; 2 La Paloma; 3 Altamira; 4 Ekain; 5 La Vache; 6 Arbreda; 7 Mazouco; 8 Siega Verde;9 Domingo Garcia; 10 La Griega; 11 Los Casares; 12 João Ramos; 13 El Niño; 14 Parpalló; 15 La Pileta;16 Nerja; 17 Isturitz; 18 Laugerie-Haute und Laugerie-Basse; 19 Gannat; 20 Marolles-sur-Seine; 21 Solutré;22 Cornille; 23 Chauvet; 24 Champréveyres; 25 Grimaldi; 26 Grotta dei Fanciulli; 27 Settepolesini di Bodeno;28 Fumane; 29 Riparo Tagliente; 30 Ponte di pietra, 31 Polesini; 32 Maurizio Tronci; 33 Paglicci; 34 Castelci-vita; 35 Gönnersdorf; 36 Andernach; 37 Brillenhöhle; 38 Vogelherd; 39 Nebra; 40 Döbritz; 41 Oelknitz;42 Trou de Chaleux; 43 Ossom’s Cave.

54 Verbreitung der großen Jagdfauna in Mittel und Westeuropa im oberen Jungpleistozän
Interessant ist auch die bisherige Abwesenheit vonKnochen von Equus ferus in Sizilien. Die IsolationSiziliens durch das Meer kann nicht der einzige Grunddafür sein, da Equus hydruntinus am Ende der Eiszeitnach Sizilien eindringt. Möglicherweise hat dersüdliche Teil Kalabriens mit seinen hohen Gebirgenund steilen Küsten eine wichtige Rolle als Filtergespielt (Zampetti et al. 2000, 26).
Als weiterer Aspekt ist hier anzumerken, dass alleFundstellen, in denen das Pferd stark dominant ist(u. a. Gönnersdorf, Andernach, Teufelsbrücke,Oelknitz, Champréveyres, Solutré, Rond-du-Barry,Arbreda und Paglicci), in hügeligen Landschaften
liegen, durch die möglicherweise die jahreszeitlichenWanderungen dieser Tiere verliefen. Aufgrund derLebensweise der Pferde in Grassteppen und wegenihrer hohen Fluchtdistanz ist eine Jagd in der offenenLandschaft kaum anzunehmen.
Die Vorstellung einer Pferdejagd aus demHinterhalt und bei gezielter Ausnutzung und Kenntnisder regionalen Topographie scheint hiermit in ganzEuropa denkbar, wenn nicht sogar wahrscheinlich. DieJagd in felsigen Regionen und insbesondere entlangsteiler Küsten scheint eher eine gelegentliche Formvon Jagd wiederzuspiegeln.

Hydruntinus (Equus hydruntinus)
Für diese ausgestorbene Tierart wird meistens den Begriff Hydruntinus verwendet.Ordnung: Perissodactyla Owen 1848; Familie: Equidae Gray 1821; Gattung: Equus Linnaeus 1758; Art: Equushydruntinus Regalia 1904. Vorwiegend verwendete Literatur: Burke et al. 2003; Cleyet-Merle und Madelaine 1991; Denzau und Denzau1999; Heptner et al. 1966, 834–858; Stehlin und Graziosi 1935; Uerpmann 1976; Uerpmann 1987.
Ausgestorben am Ende der Eiszeit, unterscheidet sichEquus hydruntinus deutlich von Equus hemionusPallas 1775, insbesondere durch Zähne und Extremi-täten (Stehlin und Graziosi 1935). Die Beziehungzwischen diesen Arten ist unklar (Uerpmann 1976).Die mögliche Präsenz von Equus hemionus in Europawährend der letzten Eiszeit wird immer wiedererwähnt. Die Bezeichnung Equus hemionus kommt inmanchen Publikationen über paläolithische Fund-stellen anstelle Equus hydruntinus vor (u. a. Bégouënund Breuil 1958). Bis heute konnten jedoch keineKnochen aus Fundstellen des europäischen Jungpaläo-lithikums zweifelsfrei Equus hemionus zugerechnetwerden (Aujoulat 1993, 108). Weiterhin ist zuerwähnen, dass in der Literatur zahlreiche weitereNamen für kleine Equiden in Europa während derletzten Eiszeit zu finden sind (u. a. Guérin und Patou-Mathis 1996, 126–129). Manche dieser Arten, z. B.Equus cazurroi Cabrera 1919, konnten durch Ver-gleiche der Art Equus hydruntinus zugeordnet werden(Wernert 1956). Bei den anderen kleinen Equidenwird trotz der Unterschiede inzwischen dieBezeichnung Equus hydruntinus als Überbegriffakzeptiert und angewendet (mündliche Mitteilung vonH.-P. Uerpmann). Neue Untersuchungen, die Equushydruntinus sowohl von einem paläontologischen alsauch ethologischen Standpunkt her Equus hemionusnahe stellen (Burke et al. 2003), zeigen, dass diesesThema noch offen bleibt.
Der Onager und der Kulan, Equus hemionus,liegen ökologisch Equus hydruntinus näher als Equusafricanus, zu dem der Nubische und der Somali-Wildesel gehören. Die ökologischen und klimatischenAnsprüche von Equus africanus (Denzau und Denzau1999, 51–54), der während des oberen Pleistozäns inweiten Teilen Nordafrikas lebte und noch heute imOsten Afrikas zwischen 10° und 25° nördlicher Breitevorkommt (Abb. 20), können kaum zum Vergleich mitder eiszeitlichen europäischen Umwelt herangezogenwer-den. Daher wird hier Equus hemionusbeschrieben, während auf eine nähere Betrachtung vonEquus africanus verzichtet wird.
Equus hemionus (E. – Hemione, Half-Ass, Onager;F. – Hémion; I. – Asino selvatico) hat zahlreicheUnterarten: Mongolischer Kulan (E. h. hemionusPallas 1775); Kasachischer Kulan (E. h. finschi
Matschie 1911); Turkmenischer Halbesel oderTurkmenischer Kulan (E. h. onager Boddaert 1785);Indischer Kulan (E. h. khur Less 1827); Arabischeroder Syrischer Kulan (E. h. hemippus, Geoffr 1855);Kiang oder Tibetischer Kulan (E. h. kiang Moorcroft1841).
Habitat: Steppenzonen, teilweise Waldsteppe,Halbwüste und Wüste. Hier können Temperaturunter-schiede eine wichtige Rolle spielen. Während in denSteppen Indiens und Pakistans, wo E. h. khur lebt, dieTemperaturen auch im Winter kaum unter denGefrierpunkt fallen, erreichen diese in der Mongoleiund in der tibetischen Region, in der E. h. hemionusund E. h. kiang leben, jeweils bis 30°C und 40°C unterNull in der Nacht. Die Sommertemperaturen liegenzwischen Werten von ca. 25°C für E. h. kiang und vonfast 50°C für E. h. khur. In allen Regionen, wo Equushemionus lebt, ist das Wasser knapp. Die Nieder-schläge bewegen sich im Bereich von 100 bis
Abb. 20 Verbreitung von Equus hemionus (hell-grau), Equus africanus (schwarz) und Equus hydrun-tinus (dunkelgrau) während des oberen Pleistozäns(nach Uerpmann 1987, 36 mit Ergänzungen nachmündlicher Mitteilung von Hans-Peter Uerpmann).

56 Verbreitung der großen Jagdfauna in Mittel und Westeuropa im oberen Jungpleistozän
300 mm, selten bis 400 mm pro Jahr. Ab einerSchneedecke von mehr als 10–15 cm Dicke sowie beihartem Schnee bekommen die Tiere Probleme bei derFutterbeschaffung. Eine Schneehöhe von mehr als40 cm kann ein Massensterben verursachen.
Verbreitung: Ursprünglich kam Equus hemionus inSteppen- und Wüsten- oder Halbwüstengebieten imSüden der Ukraine und Russlands, in Kasachstan,Zentralasien, Tibet, Vorderasien und in Nordwest-indien vor. Im größten Teil dieses Areals ist die Artausgerottet und erst im letzten Jahrhundert anmanchen Stellen wieder angesiedelt worden. DieUnterart E. h. hemippus ist ausgestorben.
Körpermaße: Länge 200–220 cm; Widerristhöhe110–137 cm; Gewicht 160–260 kg.
Körperbeschreibung: Der Schwanz ist meistensvon kurzen Haaren bedeckt, die am Ende eine Quastebilden. Der Aalstrich ist dunkel und immer deutlich zuerkennen, er verläuft bis zum Widerrist. Hier beginntdie aufrechtstehende, kurze Mähne, die bis in denBereich zwischen den Ohren reicht. Diese sind bis zu21 cm lang und länger als bei Pferden. Der Kopf wirktverhältnismäßig groß und schwer.
Fell: Kurz im Sommer, länger im Winter. Inmanchen Gegenden wächst im Winter ein richtigerPelz (Denzau und Denzau 1999, 132 und 140).Sommerfell: Rücken bis zum Kopf, Schulterpartie,Flanken, Außenfläche der Schenkel, Kopf undSchnauze intensiv gefärbt. Die Farbe variiert vonhellsandgelb (insbesondere bei E. h. kuhr) bis rötlich-braun (bei E. h. kiang). Der untere Teil des Halses, derBauch und die innere Fläche der Beine sind deutlichheller bis ganz weiß. Bei E. h. khur gibt es Tiere, diefast ganz weiß sind. Das Winterfell ist in der Farbeetwas trüber und dunkler.
Paarungszeit: Meistens von Juni bis August.Unterschiede finden sich auch in ein und demselbenAreal. Tragzeit: 11 Monate. Setzzeit: In den wärmstenMonaten.
Nahrung: Kraut- und grasartige Pflanzen, Seggenund Stauden.
Verhalten: Im Freien leben sie in Gruppen von 10bis 20 Individuen oder als Einzelgänger. Inbestimmten Jahreszeiten, und insbesondere in Abhän-gigkeit von verfügbarem Wasser, können die Gruppenaus mehreren hundert Individuen bestehen. Wande-rungen betragen oft 500 bis 600 km. In einzelnenJahren kann die Reichweite einer Wanderung nochgrößer sein. In einigen Regionen sind die Tiere
Standwild. Bei Wanderungen vom TurkmenischenKulan hängt die Zahl der Individuen einer Herde vonder Größe des Gesamtbestandes der Art in dem Gebietab. Ein Umfang von maximal 200 Tieren scheint dieRegel zu sein. Nur in manchen Regionen wurdenHerden von über 1000 Kulanen gesehen.
Natürliche Feinde: Der einzige Feind neben demMenschen ist der Wolf. Dieser greift im Rudel dieKulane (Equus hemionus) an und verfolgt einzelneTiere bis zur Erschöpfung. Eine leichtere Beute sindnatürlich Fohlen und kranke Tiere. In manchenGebieten ist es jedoch so trocken, dass sogar Wölfedort nicht leben können.
Klimatische Aussage: Über Equus hemionus kannkeine Aussage hinsichtlich der Temperaturverhält-nisse gemacht werden, da er Temperaturen von -40°Cbis +50°C ertragen kann. Nur seine besondereAnpassung an Trockensteppen ist sehr charakteri-stisch und wird allgemein auch für Equus hydruntinusangenommen (Stehlin und Graziosi 1935, 56;Uerpmann 1976, 214). In archäologischen Fundstellenist daher die Präsenz oder Abwesenheit sowie Ab- undZunahme des Anteils von Equus hydruntinus in Bezugauf die Reste von Equus ferus relevant.
Darstellungen aus dem Jungpaläolithikum
Sichere Darstellungen vom Hydruntinus sind imJungpaläolithikum äußerst selten, so dass inAbbildung 21 annäherend alle bekannten Bilder, diedieser Art zugerechnet wurden34, dargestellt sind.Dazu gehören jedoch wahrscheinlich auch einige derRückenlinien und unvollständigen oder ungenauenDarstellungen, die zu „Equiden“ gerechnet wurden.Manchmal könnten sogar einige der zu Hirschkühengerechneten Zeichnungen doch Hydruntini darstellen.Umgekehrt könnte eine Darstellung aus Montespan(Abb. 21, 5) sowohl eine Hirschkuh als auch einenHydruntinus darstellen. Eine Gravierung aus Isturitz(Saint-Périer und Saint-Périer 1936, 98) kann sowohlals Hydruntinus als auch als junges Pferd interpretiertwerden. Unter den wahrscheinlicheren Darstellungenvon Hydruntinus sind die Gravierungen aus der HöhleLes Trois Frères zu erwähnen (Abb. 21, 1 und 2). Hiersind es die langen, spitz zulaufenden und aufrechtstehenden Ohren sowie die gesamte grazile Prägungder Figuren, die eine solche Interpretation begründen.Auch bei den Darstellungen von Bernifal und Gabillou(Abb. 21, 3 und 4) sind es die langen Ohren, die eine
34 Zwar weisen Jan Jelinek (1972, 342–345), Philippe Novel (1986, 90), Norbert Aujoulat (1993, 108) sowieJean-Jacques Cleyet-Merle und Stephane Madelaine (1991, 127) auf einen sicheren „Esel“ in der Höhle von Las-caux hin, leider bilden sie ihn nicht ab und ihre Hinweise reichen nicht, um festzustellen, welche Darstellunggemeint ist. André Leroi-Gourhan (1965) so wie Arlette Leroi-Gourhan und Jacques Allain (1979) nennen dieseDarstellung nicht. Weiterhin sollten zwei Tierdarstellungen von Schweizersbild, die auf einem Kalkplättchen ein-geritzt und von Jakob Nüesch 1892 als „Wildesel“ bzw. „Steppenesel“ interpretiert wurden, eher als Hirschegedeutet werden (dazu Höneisen und Peyer 1994, 108–111).

Hydruntinus (Equus hydruntinus) 57
Abb. 21 Als Hydruntinus, Hemionus oder Esel publizierte Darstellungen. 1–2 Les Trois Frères (Bégouën undBreuil 1958, 13, Abb. 5, und 61, Abb. 65); 3 Bernifal (Breuil 1952, 290, Abb. 331); 4 Gabillou (Gaussen 1964,pl. 27,4); 5 Montespan (Trombe und Dubuc 1947, Abb. 77 und 43); 6 Les Combarelles (Novel 1987, 92);7–8 Levanzo (Graziosi 1962, tav. 19 und 20); 9 La Marche (Pales und de Saint Pereuse 1981, Abb. 47); 10 Pole-sini (Radimilli 1974, 97); 11 Domingo Garcia (Ripoll und Municio 1999, 65); 12 Los Casares (Acosta und Moli-nero 2003, 107); 13 La Peña del Candamo (Ucko 1987, 61); 14 Putois II (Cleyet-Merle und Madelaine 1991,126). Zwei weitere Hydruntinus-Köpfe soll es in der Höhle von Llonin geben (Acosta und Molinero 2003, 107).
1 2 3 4 5 6 7 8 9 10 11 12 13 14

58 Verbreitung der großen Jagdfauna in Mittel und Westeuropa im oberen Jungpleistozän
Interpretation als Hydruntinus nahe legen. Bei denGravuren von Levanzo (Abb. 21, 7 und 8) sind dieOhren kaum zu erkennen oder sie wurden stilisiertdargestellt. Hier ist eine Ansprache als Hydruntinusnur als eine der Möglichkeiten zu betrachten. Diesewird durch die Präsenz von Equus hydruntinus undden Mangel an Equus ferus in den archäologischenSchichten unterstützt (Graziosi 1962, 58). Zweimögliche Darstellungen von Hydruntini sind ausZentralspanien bekannt, die eine aus Cueva de losCasares (Abb. 21, 12), die andere aus Domingo Garcia(Abb. 21, 11). Darstellungen von sicheren oderzumindest wahrscheinlichen Hydruntini sind wederaus Südspanien noch aus Kantabrien bekannt.
Reste aus dem Jungpaläolithikum
Equus hydruntinus kommt bevorzugt in Südeuropavor (Abb. 22). In Süditalien ist er nur in Paglicci,Apulien, am Ende der Eiszeit besonders häufig, unddies sowohl in der Dryas II wie auch im Bölling undAlleröd. In Paglicci zeigt sich Equus hydruntinusmeistens umgekehrt proportional zum Pferd (Equusferus).
In anderen Fundstellen Mittel- und Süditaliens,wie in der Grotta delle Veneri in Apulien, und in denFundstellen von Grotta Polesini und Palidoro im Lazioist er immer wieder vorhanden. In Norditalien ist ernur in Riparo Tagliente mit einem einzigen Knochenvertreten, sonst fehlt er vollkommen. Hier gilt mögli-cherweise die gleiche Argumentation wie beim Pferd(siehe S. 49–54). In Frankreich kommt er immer wie-der vereinzelt in der Gironde, Dordogne (Delpech1983) und im Languedoc (Sacchi 1986) vor. In Süd-ostfrankreich ist er am Ende der Eiszeit in Cornilleund Valorgues gut vertreten (Escalon de Fonton et al.1979). Equus hydruntinus fehlt in Deutschland undGroßbritannien während des gesamten Jungpaläolithi-kums. Die Reste dieses Tieres, die in Deutschlandgefunden wurden, sind entweder nicht datiert, odersind älter als das Jungpaläolithikum (Stehlin und Gra-ziosi 1935). Manche gehören zum Eem-Interglazial(Musil 1978). Die publizierten Reste von Zähnen vonEquus hydruntinus aus der Magdalénien-FundstelleSchweizersbild in der Schweiz sind allgemein nichtakzeptiert (Höneisen und Peyer 1994, 150).
Im Höhlenkomplex bei Serinya (Arbreda, BoraGran d'en Carreres, Reclau Viver, Cau de les Gogesund Mollet III) ist Equus hydruntinus während desganzen Jungpaläolithikums mit einzelnen Knochenvertreten. Die Jagdfauna ist in diesen Höhlen jedochvon Equus ferus und Cervus elaphus dominiert.
Davidson konnte eine kleine und eine größereEquiden-Form in Parpalló, Malletes und Volcánerkennen. Er äußerte sich bewusst nicht über ihreZugehörigkeit (Davidson 1989, 67), andererseits kann
es sich bei der kleineren Form nur um Equus hydrun-tinus handeln. In Cueto de la Mina in Asturien solleneinige Reste von Equus zu Equus hydruntinus gehören(Wernert 1956). In Portugal konnte er noch nicht mitSicherheit identifiziert werden, wobei einige Fundeaus Pedreira das Salemas, Algar de Cascais undFigueira brava wahrscheinlich zu dieser Art zurechnen sind (Cardoso 1993, 252). Ob er inSüdspanien verbreitet war, was wahrscheinlich ist,konnte anhand der Literatur nicht festgestellt werden.In Nerja scheint er zu fehlen.
Diskussion
Es ist interessant zu vermerken, dass Equus hydrun-tinus in Spanien zumeist selten ist. In Südspanienkonnte er weder anhand von Darstellungen nochanhand von Knochen belegt werden. DieseVerbreitung ist um so auffälliger, wenn man bedenkt,dass Spanien eigentlich aufgrund des kontinentalenKlimas im Inneren und der verstärkten Trockenheitzumindest während des zweiten Kältemaximumsideale Lebensbedingungen für eine an die Trockenheitangepasste Tierart geboten haben sollte. Gleichzeitigist Equus hydruntinus in Frankreich, in der Dordogneund in der Gironde immer wieder belegt. Equushydruntinus wird zusätzlich in Paglicci in wärmerenPhasen zahlreicher, z. B. in der Schicht 18b9, die demLaugerie-Interstadial zugeordnet wird, sowie vomBölling bis zum Alleröd. In allen diesen Schichtenverschwindet gleichzeitig Equus ferus. Weiterhinkommt Equus hydruntinus immer mit Cervus elaphus,häufig sogar mit Sus scrofa vor. Natürlich liegt es zumTeil daran, dass Equus hydruntinus an wärmereKlimate angepasst ist, jedoch wird auch deutlich, dassdie einfache Gleichsetzung Equus hydruntinus =„mehr Trockenheit“ nicht stimmt. Eine wichtigeVariable in der Verbreitung von Equus hydruntinusscheint mit dem Vorkommen des Pferdes korreliert zusein. Das Pferd ist größer (ca. 200–300 kg bis zueinem Maximum von 450 kg beim Przewalski-Pferdgegenüber 160–260 kg bei Hydruntinus), schnellerund aggressiver als der Hydruntinus, dadurch konntees, dort wo es gute Lebensbedingungen fand, denHydruntinus als Konkurrenten unter Druck setzen undzum Ausweichen auf Randregionen gezwungenhaben. Erst das Verschwinden des Wildpferdes auseiner Region, aus welchem Grund auch immer, wiez. B. in Süditalien, ermöglichte dem Hydruntinus, dieökologische Nische des Pferdes zu erobern und sichstark zu vermehren. Andererseits jedoch darf mannicht übersehen, dass in Sizilien, wo es keine Pferdeim Jungpaläolithikum gab, der Hydruntinus weiterhinselten ist (Zampetti et al. 2000).

Hydruntinus (Equus hydruntinus) 59
In der Höhle von Polesini, in der Nähe von Rom,kommen Pferd und Hydruntinus in allen Schichten desEpigravettiens vor. Ihr Anteil an den Resten ist stabilzwischen 1,9% und 4,8% beim Pferd und 3,6% und7,9% beim Hydruntinus gegenüber einem Anteil an
Rothirsch zwischen 64,6% und 74,0%. Auffällig istaußerdem, dass es hier keine wesentlichen Unter-schiede zwischen den Schichten gibt, die denStadialen (Dryas II und Dryas III) oder den Intersta-dialen (Bölling und Alleröd) zugeordnet werden.
Abb. 22 Mögliche maximale Verbreitung von Equus hydruntinus in Mittel- und Westeuropa während des Jung-paläolithikums (u. a. anhand der Kartierung von Uerpmann 1987, 36) sowie für das Thema relevante Fundstel-len mit Resten (Dreieck) oder Darstellungen (Kreis) von Hydruntinus. Die Pfeile verweisen auf möglicheWanderungsrouten außerhalb ihres annehmbaren, üblichen Verbreitungsgebietes.1 Pedreira das Salemas undAlgar de Cascais; 2 Figueira Brava; 3 Malletes, Parpalló, Volcán; 4 Cueto de la Mina; 5 Domingo Garcia;6 Los Casares; 7 Arbreda , Bora Gran d’en Carreres Reclau Viver, Cau de les Goges, Mollet III; 8 Montespanund Putois II; 9 Les Trois Frères; 10 La Marche; 11 Gabillou; 12 Abri Morin, Saint Germain de la Rivière;13 Pont-d‘Ambon; 14 Bernifal; 15 La Ferrassie, Combe Saunière; Le Flageolet, Caminade Est; 16 Abri desBattuts; 17 Valorgues; 18 Grimaldi; 19 Riparo Tagliente; 20 Palidoro; 21 Polesini; 22 Paglicci; 23 Grottadell’Erica; 24 Grotta delle Veneri; 25 San Teodoro; 26 Levanzo.


Wollnashorn (Coelodonta antiquitatis)
D. – Wollnashorn, Wollhaariges Nashorn oder Fellnashorn; E. – Woolly rhinoceros; F – Rhinocéros laineux;S. – Rinoceronte lanudo; Kat. – Rinoceront llanut; P. – Rinoceronte lanígero; I. – Rinoceronte lanoso. Ausgestorben: am Ende der letzten Eiszeit.Ordnung: Perissodactyla Owen 1848; Familie: Rhinocerotidae Gray 1821; Gattung: Coelodonta Bronn 1831;Art: Coelodonta antiquitatis (Blumenbach 1799). Vorwiegend verwendete Literatur: Kahlke 1994; Guérin und Patou-Mathis 1996.
Die wissenschaftliche Entdeckung und Untersuchungdieser Art fand in verschiedenen Ländern gleichzeitigstatt, so dass viele Namen parallel entstanden, diemindestens zum Teil als Synonyme zu sehen sind.Darunter Rhinoceros tichorhinus, Tichorhinus antiqui-tatis und Rhinoceros antiquitatis. Die BezeichnungCoelodonta (nach Bronn 1831) für das Genus undantiquitatis (nach Blumenbach 1799) für die Speziesist jedoch der einzige wissenschaftlich gültige Name(Arsuaga und Aguirre 1979, 24).
Nashörner leben zur Zeit nur in sehr warmenRegionen, so dass ein Vergleich mit dem ausgestor-benen Wollnashorn außerordentlich schwierig ist. Dieheutigen Nashörner leben in Afrika, so das Breitmaul-oder Weiße Nashorn (Ceratotherium simum Burchell1817) und das Spitzmaul- oder Schwarze Nashorn(Diceros bicornis Linnaeus 1758), in Indien, dasIndische Panzernashorn (Rhinoceros unicornisLinnaeus 1758), auf Java, das Javanashorn (Rhino-ceros sondaicus Desmarest 1822) und auf Sumatra,das Sumatranashorn (Dicerorhinus sumatrensisFischer 1814). Die afrikanischen Nashörner leben inder Savanne, die asiatischen im Tropenwald. Dasgrößte ist das Breitmaulnashorn (Kopf-Rumpf-Länge360 bis 375 cm; Schulterhöhe 175 bis 190 cm;Gewicht 1,8 bis 2,3 t), das auch die längste Tragzeit(490 Tage) und die längste Lebenserwartung (45Jahre) hat. Das kleinste ist das Sumatranashorn (Kopf-Rumpf-Länge 210 cm; Schulterhöhe 135 cm; Gewicht0,8 t), das auch die kürzeste Tragzeit (400 Tage) unddie kürzeste Lebenserwartung (35 Jahre) unter denNashörnern hat.
Sehr wichtige Informationen über Wollnashörnerkonnten anhand mehrerer mumifizierten Kadaver ins-besondere aus Starunia in der Ukaine, ehemaligesPolen (Nowak et al. 1930) und aus Vilyui undChurapcha in Sibirien gewonnen werden (Guthrie1990; Joger und Koch 1994).
Habitat: Tundra und Steppe.Verbreitung: Nach Kahlke (1994, Abb. 15)
bewohnte Coelodonta antiquitatis in seiner maximalenVerbreitung ein Gebiet von Kantabrien bis Korea. Im
Norden kam es bis zur Grenze der Gletscher, nachSüden bis Süditalien, Griechenland, in den Kaukasusund bis zur chinesischen Küste am Pazifischen Ozeanvor.
Größe eines weiblichen Individuums von Starunia:Kopf-Rumpf-Länge 360 cm; Schulterhöhe: 160 cm.
Hörner: Das vordere Horn erreichte 130 cm, dashintere 90 cm Länge. Die abgeschliffene Vorderseitedes Hornes zeigt, dass das Wollnashorn sein Horn fürdie Suche nach Pflanzen unter dem Schnee einsetzte.
Fortpflanzung: Wenn man annimmt, dass diegünstigste Jahreszeit zum Setzen für Wollnashörner sowie für Mammuts April–Mai gewesen ist und dass dieTragzeit des Wollnashorns in der Nähe der Tragzeitvon Breitmaulnashörnern (fast 17 Monate) lag undeventuell sogar etwas länger gedauert haben könnte,kommt man auf eine Paarungszeit am Ende desSommers und Beginn des Herbstes.
Verhalten: Narben auf der Haut eines derIndividuen von Starunia (Königswald 2002, 59) sowieSpuren von Traumata am Schädel verschiedenerweiterer Nashörner (Garutt 1999, 175) lassen heftigeinnerartliche Rangeleien vermuten. Norbert Aujoulatund Claude Guérin (Clottes 2003, 174–176) meinen,dass viele ökologische und ethologische Affinitätenzwischen dem Wollnashorn (Coelodonta antiquitatis)und dem noch heute lebenden Weißen Nashorn(Ceratotherium simum) zu erkennen seien. DieGruppen von Weißen Nashörnern bestehen häufig ausmehr als zehn Tieren, zu denen ein dominanter Bulle,mehrere Weibchen und junge Tiere beiderGeschlechter gehören. Paläontologische Fundstellenlegen nahe, dass beim Wollnashorn eine ähnlicheStruktur der Familienverbände anzunehmen sei.Weiterhin soll das Wollnashorn so wie das WeißeNashorn ein ausgeprägtes Revierverhalten gehabthaben und Standwild gewesen sein (Clottes 2003,176). Sein vermutetes Revierverhalten, seine Ortsge-bundenheit und die geringe Größe der Gruppenkönnten die Gründe dafür sein, dass das Wollnashorn,anders als Rentiere, Pferde, Steppenbisons, Saiga-Antilopen und Mammuts, Nordamerika nie erreichte.

62 Verbreitung der großen Jagdfauna in Mittel und Westeuropa im oberen Jungpleistozän
Nahrung: Anhand der Untersuchungen von Magen-inhalt der im Permafrost erhaltenen Wollnashornka-daver weiß man, dass sie Gräser, Seggen, niederePflanzen und gelegentlich Blätter gegessen haben.
Klimatische Aussage: Das Wollnashorn ist inEurasien ein ständiger Begleiter von Mammuthusprimigenius. Sein gesenkter Kopf zeigt, dass esSchwierigkeiten mit einer zu hohen Schneedeckegehabt haben muss. Bestens ausgerüstet gegentrockene Kälte war sein Fell andererseits nicht wasser-dicht. Daher war Coelodonta antiquitatis, so wieMammuthus primigenius und Ovibos moschatus, sehrempfindlich gegen Regen im Winter.
Darstellungen aus dem Jungpaläolithikum
Bis vor einigen Jahren galt das Wollnashorn als selten;Louis-René Nougier und Romain Robert zählten amEnde der 1950er Jahre 28 Darstellungen in der Wand-und 15 in der mobilen Kunst (1957, 48). 1993 zählteBarrière nur ca. 20 Darstellungen in der Wandkunst(Barrière 1993c, 158). Die Höhle von Rouffignac mitneun (Nougier und Robert 1957, 46), zwölf (Barrière1982, 150) oder elf Darstellungen (Plassard 1999, 62)war in diesem Kontext die bei weitem reichste Höhlean Darstellungen von Wollnashörnern.
Louis-René Nougier und Romain Robert betontenin Bezug auf Rouffignac, dass das Besondere an denDarstellungen dieser Höhle ist, dass dort nicht nureinzelne Individuen, sondern auch kleine Gruppendargestellt seien (Nougier und Robert 1957, 38). DieEntdeckung der Höhle von Chauvet, in der bis jetzt 65Nashörner gezählt wurden (Clottes 2003, 174),änderte dieses Bild natürlich stark, sowohl wegen derAnzahl, wie auch wegen der Qualität der Darstel-lungen. Die Form der Nashörner von Chauvetentspricht vollkommen der Form der Nashörner vonFont-de-Gaume (Abb. 23, 8), von Les Trois Frères(Abb. 23, 9), von Rouffignac (Abb. 23, 10), von LesCombarelles II (Abb. 23, 11) oder von Gönnersdorf(Abb. 23, 12). Der Kopf hängt tiefer als die Schultern.Diese wirken dadurch noch mächtiger. In Rouffignacscheinen die Nashörner beim Gehen oder beimWeiden dargestellt zu sein, in Chauvet dagegen sinddie Tiere unruhig (Abb. 23, 5, 6 und 7) abgebildet. Inmanchen Fällen sind sie kämpfend dargestellt. Einerder Kontrahenten blutet eindeutig sehr stark (Clottes2003, 143). Geht man davon aus, dass diese Kämpfeim Zusammenhang mit der Paarungszeit stehen, washier als reine Hypothese geäußert wird, könnte maneine Zusatzinformation über die Jahreszeit der darge-stellten Szene haben, nämlich Spätsommer, bzw.Anfang Herbst.
Der auffällige Streifen in der Mitte des Rumpfes derNashörner aus der Höhle Chauvet ist einmalig in derpaläolithischen Kunst und erscheint zunächstunerklärlich (Clottes 2003, 174). Er könnte aber dasAuftreten des neuen Winterfelles symbolisieren,welches bei Wollnashörnern aus sehr langen Haarenbestand, so wie man es aus den Darstellungen vonFont-de-Gaume, Rouffignac, Les Combarelles undGönnersdorf kennt (Abb. 23, 8, 10–12). In der Höhlevon Chauvet sind Farbunterschiede in Form vondunklerer Farbgebung oder hellen, frei geschabtenOberflächen als stilistische Konvention für das Fellvon Tieren verwendet worden, man denke an dieRentiere, die Riesenhirsche, die Moschusochsen, dieSteppenbisons, die Pferde oder die Löwen (Chauvet etal. 1996; Clottes 2003). Weiterhin sind bei manchenDarstellungen von Nashörnern die Bauchpartie unddie Wange durch Farbunterschiede betont worden(Clottes 2003, 132–135 und 176). Der Bauch und dieFlanken sind die sensibelsten Stellen eines Tieres, wowahrscheinlich der Winterhaarwuchs als erstes einge-setzt hat. Die Wange ist ebenfalls ein Körperbereich,in dem der Haarwuchs stärker ist als im Augen- oderNasenbereich, wie es bei den Nashorn- und Mammut-darstellungen von Rouffignac zu sehen ist (Barrière1982).
Die Tatsache, dass manche Tiere keinen Streifenaufweisen, während andere einen deutlichen Streifenzeigen und andere schon fast vollständig dunkel sind(Clottes 2003, 133), könnte daher unterschiedlicheBehaarungszustände zeigen.
In Gönnersdorf existieren weiterhin mindestenszehn Gravierungen dieses Tieres (Bosinski 1982, 41).Leider sind die meisten davon kaum publiziert.Insgesamt kommt man in Mittel- und Westeuropa aufca. 120 Darstellungen von Nashörnern, von denen diemeisten aus Frankreich stammen. Damit ist dasNashorn zahlenmäßig nördlich der Pyrenäen und derAlpen nicht weiter als seltene Tierart zu betrachten.Äußerst selten bleibt das Nashorn in den Darstel-lungen von Spanien, in Italien fehlt es (Abb. 24). Dieeinzigen sicheren Darstellungen in Spanien stammenaus Las Caldas (Abb. 23, 1–2). Die Gravierungen vonEkain könnten zwar Nashörner sein, aber sie könntengenau so gut andere Tiere darstellen (Abb. 23, 3–4).Die immer wieder zitierte Darstellung eines Wollnas-hornes aus der Höhle von Los Casares35 ist, nach dergenaueren Aufnahme durch Rodrigo de BalbínBehrmann und José Javier Alcolea González sowiedurch das Foto derselben Figur (1992, 422, Abb. 27und Abb. 28) nicht mehr haltbar. Ich nehme an, dasses sich um ein Wildschwein handelt, dementspre-chend wird diese Figur im Kapitel über dasWildschwein (S. 83–88) besprochen und abgebildet
35 Breuil 1952, 389; Arsuaga und Aguirre 1979, 26; Corchón 1992, 57; Álvarez und Jöris 1998, 70; Acosta undMolinero 2003, 129–131; Alcolea und Balbín 2003, 236).
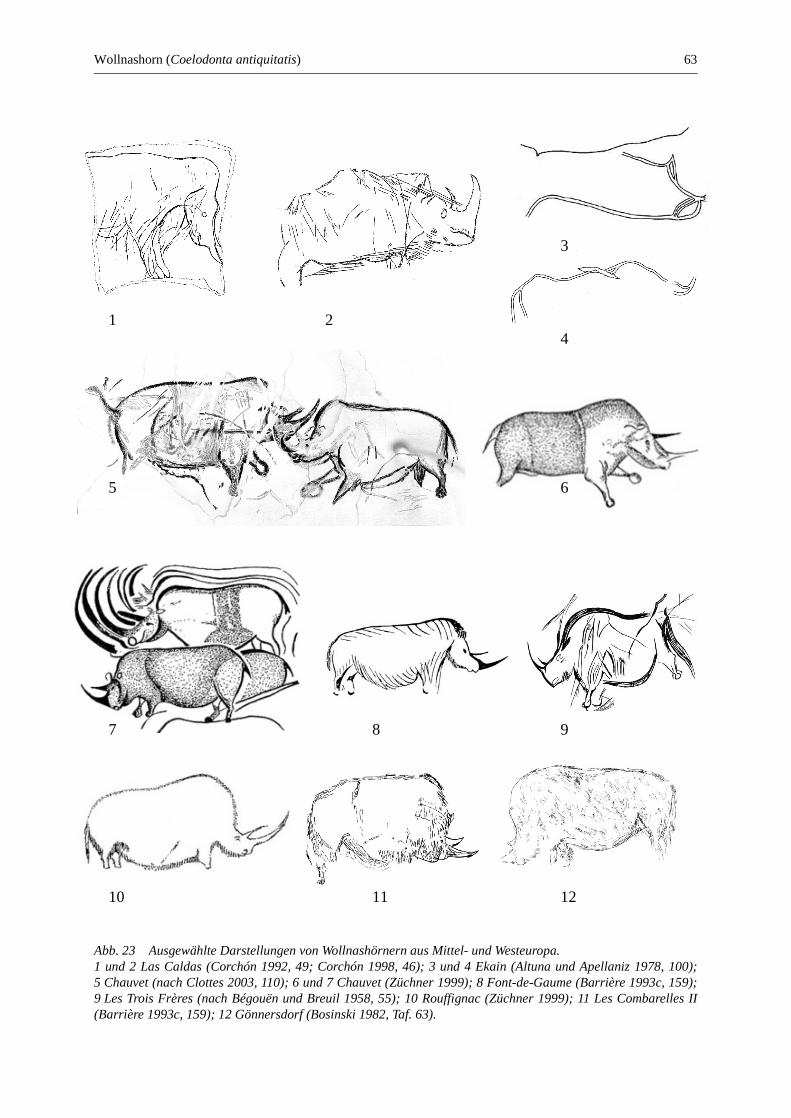
Wollnashorn (Coelodonta antiquitatis) 63
Abb. 23 Ausgewählte Darstellungen von Wollnashörnern aus Mittel- und Westeuropa.1 und 2 Las Caldas (Corchón 1992, 49; Corchón 1998, 46); 3 und 4 Ekain (Altuna und Apellaniz 1978, 100);5 Chauvet (nach Clottes 2003, 110); 6 und 7 Chauvet (Züchner 1999); 8 Font-de-Gaume (Barrière 1993c, 159);9 Les Trois Frères (nach Bégouën und Breuil 1958, 55); 10 Rouffignac (Züchner 1999); 11 Les Combarelles II(Barrière 1993c, 159); 12 Gönnersdorf (Bosinski 1982, Taf. 63).
3 1 2 4 5 6
7 8 9 10 11 12

64 Verbreitung der großen Jagdfauna in Mittel und Westeuropa im oberen Jungpleistozän
(Abb. 34). Die angebliche Darstellung eines Wollnas-hornes aus Siega Verde (Balbín und Alcolea 1995)scheint anhand des Fotos (Alcolea und Balbín 2003,237) als Naturprodukt zu interpretieren zu sein undwird daher hier nicht besprochen. Schließlich sollteman auch die rötliche Malerei in serpentiformerTechnik aus der Höhle von La Pileta in der Provinzvon Granada erwähnen. Diese Darstellung wurde vonBreuil sowohl als sichere (Breuil et al. 1915, 18–19)als auch als fragliche (Breuil et al. 1915, Pl. V)Darstellung eines Nashornes angesprochen. Sie weistjedoch keine Charakteristika auf, die für ein Nashornsprechen würden. Daher wird hier auch dieseDarstellung als Nashorn abgelehnt und nicht weiterbehandelt.
Reste aus dem Jungpaläolithikum
Das Wollnashorn ist ein typischer Vertreter derFaunenkomplexe des älteren und mittleren Jungpaläo-lithikums in Mitteleuropa. Es kommt immer wieder imAurignacien und Gravettien Südwestdeutschlands(u. a. Bockstein, Hohlenstein-Stadel, Geißenklösterleund Vogelherd), Frankreichs (u. a. Roc de Combe,Aurignac, Gargas und Gatzarria) und Spaniens(Lezetxiki) vor; etwas seltener sind sie imMagdalénien (u. a. Oelknitz, Kesslerloch, Spitzbuben-höhle in Deutschland und Orciers in Frankreich).
In El Castillo wurden Splitter eines Nashorn-zahnes entdeckt, die von Altuna als Wollnashorninterpretiert wurden, da die Gattung Dicerorhinus(oder Stephanorhinus) aus Kantabrien und Europa vordem Würm III/IV verschwunden sei (Altuna 1995,93). Diese Aussage widerspricht vollkommen der
Zuordnung von anderen Nashornresten aus demAurignacien und Gravettien der gleichen Höhle durchdenselben Autor (siehe das Kapitel über dasWaldnashorn, S. 67). Daher muss dieser Fund alsCoelodonta antiquitatis unter Vorbehalt betrachtetwerden.
Interessant zu erwähnen sind hier die Reste aus derHöhle La Carigüela in der Nähe von Granada und ausLa Unquera in der Nähe von Santander. In beidenFundstellen kamen die Reste von Coelodonta antiqui-tatis zusammen mit Steinartefakten des Moustérienvor (Arsuaga und Aguirre 1979). Der Fund aus LaCarigüela, einer Höhle in Südspanien, ist dersüdlichste Fund eines Wollnashornes in Spanien.Zusammen mit den Resten aus der Grotta diCardamone in Apulien (Abb. 24) zeigt er, dass dasWollnashorn im Mittelpaläolithikum bis in diesüdlichsten Regionen Südeuropas vorgedrungen war.
Diskussion
Es ist auffällig, dass Reste von Wollnashörnern ausjungpaläolithischen Fundstellen immer aus einzelnen,bestenfalls einigen wenigen Individuen bestehen. Esgab möglicherweise nie eine spezialisierte Jagd aufdiese Tiere. Man darf natürlich nicht vergessen, dasswie beim Mammut die Erlegung eines Nashornes vielmehr Fleischausbeute brachte als die Erlegung einesRentieres, Rothirsches oder Pferdes. Tatsache bleibtjedoch, dass es in Osteuropa Fundstellen gibt, wo einespezialisierte Jagd auf Mammuts bewiesen ist (u. a.Pavlov, Dolni Vestonice). Mir ist aber keineFundstelle bekannt, wo man von einer spezialisiertenJagd auf Wollnashörner sprechen könnte.

Wollnashorn (Coelodonta antiquitatis) 65
Abb. 24 Verbreitung von Resten und Darstellungen von Coelodonta antiquitatis. Fundstellen mit Darstellungenaus Spanien und eine Auswahl aus anderen Gebieten Europas sind mit einem Kreis, Faunenreste aus Spanien miteinem Dreieck und mittelpaläolithische oder paläontologische Fundstellen aus Spanien und Italien mit einemKreuz markiert. 1 Las Caldas; 2 El Castillo; 3 Labeko-Koba; 4 Lezetxiki; 5 fragliche Darstellungen in Ekain;6 La Unquera; 7 Udias; 8 Cueva de las Cascaras; 9 Cueva de Coscobillo; 10 Cueva de Toll; 11 Cueva Arenys deMar; 12 Arroyo del Culebro; 13 La Carigüela; 14 Polesine; 15 Settepolesini di Bodeno; 16 Monte Circeo;17 Grotta dei Pipistrelli; 18 Grotta di Cardamone; 19 Gönnersdorf; 20–22 Font-de-Gaume, Rouffignac und LesCombarelles II; 23 Chauvet; 24 Les Trois Frères.


Waldnashorn (Stephanorhinus kirchbergensis)
D. – Waldnashorn, Mercksches Nashorn; E. – Merck’s Rhinoceros; F. – Rhinocéros de Merck; S. – Rinocerontede Merck; K. – Rinoceront de Merck; I. – Rinoceronte di Merck. Sehr häufig wird jedoch nur der lateinischeName verwendet.Ordnung: Perissodactyla Owen 1884; Familie: Rhinocerotidae Gray 1821; Gattung: Stephanorhinus Kretzoi1942; Art: Stephanorhinus kirchbergensis (Jaeger 1839). Synonym ist die Bezeichnung Stephanorhinus mercki(Kaup 1841). Vorwiegend verwendete Literatur: Altuna 1972; Guérin und Patou-Mathis 1996.
Dieses Nashorn war im Eem-Interglazial weitverbreitet. Funde stammen aus England, Spanien,Italien ebenso wie aus Gebieten des mittleren Orients.Am Beginn der Würm-Eiszeit verschwand es ausMitteleuropa sowie aus Italien (Tagliacozzo 1992, 78;Sala 1980, 237). Anscheinend überlebte es bis zumBeginn des Jungpaläolithikums nur auf der IberischenHalbinsel (Abb. 26). Funde stammen aus Schicht IVa,Aurignacien Antiguo, und aus der Schicht IIIa, dem
oberen Aurignacien, von Lezetxiki sowie aus denSchichten 18, Aurignacien, 16, Gravettien, und 13,gravettienzeitlich, von El Castillo (Straus 1992, 262).In manchen Publikationen wird das Aussterben vonStephanorhinus kirchbergensis sowie das Aussterbenvon Stephanorhinus hemitoechus irrtümlicherweiseauf das Ende des Eem-Interglazials datiert(Klostermann 1999, 167).
Steppennashorn (Stephanorhinus hemitoechus)
E. – Narrow-nosed Rhinoceros; F. – Rhinocéros de prairie; S. – Rinoceronte de estepa; K. – Rinoceront depraderia; I. – Rinoceronte di prateria. Sehr häufig wird jedoch nur der lateinische Name verwendet.Ordnung: Perissodactyla Owen 1848; Familie: Rhinocerotidae Gray 1821; Gattung: Stephanorhinus Kretzoi1942; Art: Stephanorhinus hemitoechus (Falconer 1868).Vorwiegend verwendete Literatur: Cardoso 1993; Guérin und Patou-Mathis 1996.
Dieses Nashorn wird oft mit dem Waldnashornverwechselt, obwohl es etwas kleiner ist. Es trägt denKopf auf mittlerer Höhe oder tief. Die Größeentspricht der eines heutigen Schwarzen Nashornes. InItalien soll es zusammen mit dem Waldnashorn amBeginn des Würms verschwunden sein (Tagliacozzo1992, 78; Sala 1980, 237). Ein Fußknochen einesStephanorhinus cf. hemitoechus stammt aus einerSchicht des späten Moustériens der Grotta di Castel-civita (Abb. 26), die auf ungefähr 40.000 BP datiertwurde (Masini und Abbazzi 1997, 34). Es könnte sichaber auch um Coelodonta antiquitatis handeln. Leidererlaubt dieser Fußknochen keine sichere Bestimmungder Art. Aus der Schicht 14 (Uluzziano, 29.120 ± 300BP) der Grotta della Cala stammt ebenfalls einKnochen, der als Stephanorhinus cf. hemitoechusbestimmt wurde (Benini et al. 1997, 69–70). InPortugal überlebte das Steppennashorn bis ins Jungpa-läolithikum. Verschiedene Reste stammen aus der Zeitzwischen 30.000 und 22.000 BP (Cardoso 1993, 273).In Spanien ist Stephanorhinus hemitoechus imAurignacien von Cueva del Otero (Madariaga 1966,
71), vielleicht in der Höhle von El Castillo (Altuna1972, 395–398; Madariaga 1966, 71) und vielleichtauch in Lezetxiki (Altuna 1972, 409) vorhanden.
Claude Guérin meint, dass diese Art in der „scènedu puits“ der Höhle von Lascaux zu erkennen sei(Guérin und Faure 1983, 31; Guérin und Patou-Mathis1996, 119; Clottes 2003, 174) (Abb. 25). Im Bereichdes Unterkiefers bis zum Beginn des Oberschenkelssind jedoch mehrere schwache Striche als Behaarungzu deuten. Diese gehören zu der Zeichnung, die vordem Auftragen der schwarzen Farbe durch Blasenangefertigt worden war (Nougier und Robert 1957,32). Die üblichen Konturen sind sehr kräftig undwirken verschwommen. Diese Figur hebt sich nichtvon anderen Nashorndarstellungen ab, die alle der ArtCoelodonta antiquitatis zugerechnet werden, daher isteine Interpretation dieser Darstellung als Stephanor-hinus hemitoechus wenig überzeugend. Von anderenAutoren wird diese mögliche Zuordnung nichtübernommen (u. a. Barrière 1993c; Leroi-Gourhan etal. 1984). Louis-René Nougier und Romain Robertbetonen in Bezug auf eine mögliche Zuordnung dieser

68 Verbreitung der großen Jagdfauna in Mittel und Westeuropa im oberen Jungpleistozän
Darstellung als „rhinocéros Mercki“, dass es sich umein Rhinocéros tichorhinus (oder Coelodonta antiqui-tatis) handelt (Nougier und Robert 1957, 32). Bedenktman, dass das Steppennashorn im Jungpaläolithikumnur noch in Portugal für die Zeit zwischen 30.000 und20.000 BP als sicher nachgewiesen gilt (Guérin undPatou-Mathis 1996, 119; Cardoso 1993, 273), ist dieDarstellung eines Steppennashornes in Lascaux wennauch nicht unmöglich, so doch zumindest unwahr-scheinlich.
Es sei ergänzend erwähnt, dass die Fundstellen inPortugal sowie in Zentral- und Südspanien meistenseine eher schlechte Knochenerhaltung aufweisen.Dadurch sind die Hinweise auf die letzten Wald- undSteppennashörner noch seltener und schwieriger zubestimmen.
Abb. 25 Darstellung aus Lascaux, die von ClaudeGuérin als Stephanorhinus hemitoechus interpretiertwurde (Guérin und Faure 1983, 31).
Abb. 26 Fundstellen in Spanien im oberen Jungpleistozän mit Resten von Stephanorhinus kirchbergensis(1 El Castillo; 3 Lezetxiki), Stephanorhinus hemitoechus (1 El Castillo (?), 2 El Otero, 3 Lezetxiki (?); 5 GrutaNova da Columbeira; 6 Lapa da Raínha; 7 Pedreira das Salemas; 8 Figueira Brava; 9 Castelcivita (?)) und Ste-phanorhinus sp. (4 Aitzbitarte).

Rüsseltiere (Proboscidea)
Nach Volker Storch und Ulrich Welsch (1993, 446):„Größte landlebende Säugetiere. Die fünf Zehentragen nagelähnliche Hufe. Säulenartige Beine. Naseund Oberlippe bilden einen langen Rüssel. Zwei obereSchneidezähne sind immerwachsende Stoßzähne,derentwegen v. a. der afrikanische Elefant verfolgtwird und von Ausrottung bedroht ist. Pflanzenfressermit sehr großen Backenzähnen, die aus mehrerentransversalen Lamellen zusammengesetzt sind.Elephas (indischer Elefant), Loxodonta (afrikanischerElefant).“
Aus Fundstellen des Jungpaläolithikums Europas sindnur zwei Arten bekannt, Elephas antiquus (Wald-elefant) und Mammuthus primigenius (Mammut). Fer-
ner sind von den Mittelmeerinseln Reste von Zwerg-elefanten aus paläontologischen Fundstellen bekannt,deren Stammesgeschichte jedoch nicht vollständiggeklärt worden ist (Caloi et al. 1996, 234–239). Aufder griechischen Insel Tilos sollen Zwergelefanten biszum späten Pleistozän gelebt haben (Bachmayer et al.1976). Claude Guérin und Marylène Patou-Mathis(1996, 151) halten es für wahrscheinlich, dass derenAussterben in Verbindung mit der Ankunft desMenschen am Ende des Paläolithikums oder im Neoli-thikum zu sehen sei. Da es keine Funde dieser Tierartaus archäologischen Fundstellen gibt, wird sie indieser Arbeit nicht behandelt.
Mammut (Mammuthus primigenius)
E. – Mammoth; F. – Mammouth; S. – Mamut; Kat. – Mamut; P. – Mamute; I. – Mammut.Ordnung: Proboscidea Illiger 1811; Familie: Elephantidae Gray 1821; Gattung: Mammuthus Brookes 1828; Art:Mammuthus primigenius (Blumenbach 1799).Vorwiegend verwendete Literatur: Cavarretta et al. 2001; Guthrie 1990, 2001; Haynes et al. 1999; Joger undKoch 1994; Lister und Bahn 1997; Mol und van Essen 1992.
Heutige Verwandte: Wie beim Wollnashorn lebenauch beim Mammut alle heutigen Verwandten in sehrwarmen Regionen, nämlich der afrikanische Elefant(Loxodonta africana) in Savannen, Tropenwäldernund Halbwüsten Afrikas und der asiatische Elefant(Elephas maximus) in Savannen und TropenwäldernIndiens und Südostasiens. Der afrikanische Elefant(Kopf-Rumpf-Länge 600 bis 750 cm; Schulterhöhe220 bis 370 cm, selten bis 400 cm; Gewicht bis 7,5 t)lebt in Herden. Diese erreichten in der Vergangenheitdurchaus hundert Individuen. Er ist tag- und nacht-aktiv. Die Tragzeit dieser Art beträgt 22 Monate, dieLebenserwartung 50 bis 70 Jahre. Ihre Nahrungs-quellen sind Gras, Blätter, Zweige, Früchte, Knospenund Wurzeln.
Der asiatische Elefant (Kopf-Rumpf-Länge 550 bis640 cm; Schulterhöhe 240 bis 290 cm; Gewicht bis4,7 t) lebt in Gruppen aus mehreren Familien von 5 bis120 Tieren. Er ist sowohl tag- als auch nachtaktiv. DieTragzeit beträgt ebenfalls ca. 22 Monate, die Lebens-erwartung 40 bis 60 Jahre. Die wichtigste Nahrung istGras; in den Wäldern kommen Laubbäume undPalmen dazu.
Habitat: Mammutsteppe. Die letzten Mammuts lebtenin der heutigen Tundra.
Körpermaße: Schulterhöhe bis 3,5 m; Gewicht 4bis 6 t. Auf der Wrangel-Insel gab es kleinere Formen.
Stoßzähne: Der weltweit größte Stoßzahn stammtvon einem Exemplar vom Kolyma-Fluss (Ostsi-birien). Er ist 490 cm lang, gemessen entlang deräußeren Krümmung, und wiegt 84 kg.
Verhalten: Man nimmt an, dass eine Mammut-herde im Durchschnitt aus 10 bis 30 Individuenbestand. Ihr Nahrungs- und Wasserbedarf sowie dasNahrungsangebot der eiszeitlichen Tundra setzenausgedehnte Wanderungen voraus (Velichko undZelikson 2001).
Paarungszeit: Juli bis August (Guthrie 2001).Tragzeit: Guthrie nimmt ca. 22 Monate als Tragzeit inAnalogie zu den heute lebenden Elefanten alswahrscheinlich an (Guthrie 2001, 276). Setzzeit: Aprilbis Mai.
Nahrung: Anhand von Mageninhaltsanalysen aneinigen Mammutkadavern konnte festgestellt werden,dass sie hauptsächlich Gras, Zweige und Moosegefressen haben. Ihr Nahrungsbedarf wurde auf ca.

70 Verbreitung der großen Jagdfauna in Mittel und Westeuropa im oberen Jungpleistozän
175 bis 200 kg pflanzlichen Materials und über 150Liter an Wasser pro Tag geschätzt (Velichko undZelikson 2001).
Natürliche Lebenserwartung: Für das Mammutwird eine ähnlich lange Lebenserwartung wie beimAfrikanischen Elefanten geschätzt, also über 55 Jahre.
Klimatische Aussage: Das Mammut ist derbekannteste Vertreter des Mammuthus-Coelodonta-Faunenkomplexes (Kahlke 1994) bzw. der Mammut-steppe (u. a. Guthrie 1990). Sein Auftreten stehtimmer mit einem sehr kalten und extrem trockenenKlima in Verbindung. Unter den Anpassungen an einsolches Klima sei hier auf seine „Wolle“ und seine füreinen Elefanten kleinen Ohren (Guthrie 1990, 6)hingewiesen. Gerade die Wolle, welche aus einerkurzen (2,5 bis 8 cm) und sehr dichten Unterwollesowie aus langen deckenden Haaren (bis zu 90 cm)bestand, weist auf seine klimatischen Grenzen hin.Bestens ausgerüstet gegen trockene Kälte war dasMammut bei Regen im Winter ungeschützt. Sein Fellwar nicht wasserdicht, daher hätte das Wasser denKörper erreicht und das Tier hätte sich erkältet (Jogerund Koch 1994, 126). Bei der Annahme, dass währendder letzten Eiszeit die Temperaturen im Sommerteilweise höher als die heutigen lagen (Frenzel 1980,20), sind Niederschläge während dieser Jahreszeit inForm von Regen vorauszusetzen. Damit ist eindichtes, wolliges Fell bei Mammuts im Sommerunwahrscheinlich. Weiterhin könnte ein Tier von vierbis sechs Tonnen Gewicht unmöglich in einerbaumarmen Tundra bei starker Hitze einen solchenPelz tragen, so wie man es von den sibirischen Fundenkennt. Ein Kreislaufkollaps wäre vorprogrammiertgewesen.
Logischerweise müssen Mammuts, so wie andereTiere des Mammuthus-Coelodonta-Faunenkomplexes(Kahlke 1994, 69–71), ein Sommer- und einWinterfell gehabt haben. Wahrscheinlich gab es auch
im Laufe der Jahrzehntausende Änderungen undlokale Anpassungen, die nur auf der Grundlage derKnochen nicht zu erkennen sind. So ist es zumindestunwahrscheinlich, dass Mammuts aus dem Mittelpa-läolithikum Süditaliens und Südspaniens das gleichedicke Fell unserer „klassischen“, sibirischenVorstellung vom Mammut gehabt haben. Daher darfdas Auftreten von Mammuten im Mittelpaläolithikumin Süditalien und Südspanien nicht sofort mitextremen, „sibirischen“ Temperaturen in Verbindunggebracht werden.
Darstellungen aus dem Jungpaläolithikum
Darstellungen von Mammuten sind in Mitteleuropaund Frankreich üblich. Da hier die Präsenz vonMammuthus primigenius sowie das Verschwinden vonElephas antiquus schon ab dem frühen Würmallgemein akzeptiert ist, werden uns Darstellungen ausMitteleuropa kaum weiter beschäftigen. Es sei hier nurals Beispiel auf eine Gravierung aus Gönnersdorf undeine schwarzgraue Wandmalerei aus der Höhle vonChauvet hingewiesen.
In Gönnersdorf werden die Schichten auf ca.12.500 BP datiert (Bosinski 1992, 86). Hier wurdenElfenbein und Mammutknochen sowie Mammutdar-stellungen entdeckt, die um mindestens 500 Jahrejünger als die letzten datierten Mammutknochen ausSüdschweden sind (Vereshchagin und Baryshnikov1984, 495). In der abgebildeten Gravierung vonGönnersdorf (Abb. 27, links) sind zahlreiche Details,darunter die Behaarung, die Ohren und die Spitze desRüssels zu sehen, so dass man kaum von einer überGenerationen überlieferten Erinnerung, sondern vontatsächlich gesehenen Tieren sprechen muss.
Die Darstellungen aus der Höhle Chauvet zeigenMammuts mit zahlreichen Details, darunter die Ohren,die Spitze des Rüssels und die Stoßzähne (Abb. 27,
Abb. 27 Links, Gravierung auf einer Schieferplatte aus Gönnersdorf (Bosinski und Fischer 1980, Taf. 13);rechts, schwarzgraue Wandmalerei aus der Höhle von Chauvet (verändert nach Clottes 2003, 136).

Mammut (Mammuthus primigenius) 71
rechts). Diese Tiere sind „nackt“ dargestellt, und beieiner solchen Vorliebe für Details ist die immerwieder erwähnte künstlerische Freiheit kaumanzunehmen.
Besonders relevant für diese Arbeit sind jedocheher die Darstellungen aus Südeuropa, da sie aus einerRegion stammen, in der die Mammuts ihre südlichsteAusdehnungsgrenze hatten.
In Italien sind keine Mammutdarstellungenbekannt, andererseits muss man aber betonen, dassMammuts für das Jungpaläolithikum in Italien nur imAurignacien und nur in Norditalien belegt sind.Norditalien jedoch ist an sich eine an paläolithischenTierdarstellungen arme Region. Es fehlt also die stati-stische Basis, um überhaupt anhand der Präsenz, desMangels oder der Abwesenheit von Darstellungendarüber diskutieren zu können.
Anders sieht es in Spanien aus. Manche Autorenbehaupten, dass in Spanien nur vier Mammutdarstel-lungen aus drei Fundplätzen bekannt seien (Bellier etal. 1999) und zählen darunter eine Darstellung auf,welche zwar Henri Breuil und Hugo Obermaier (1935)als Mammut ansprechen, die jedoch heute allgemeinals Steppenbisondarstellung anerkannt ist (Freeman etal. 1987, 87–93). Damit entsteht ein Bild, nach demMammuts in Spanien die ganz große Ausnahme seien.Tatsache ist aber, dass 13 Elefanten- bzw. Mammut-darstellungen aus neun Fundstellen publiziert sind.Tatsache ist aber auch, dass viele davon umstrittensind. Bei den Gravierungen aus Las Caldas (Abb. 28,10–12) sind Stoßzähne, Augen und Behaarungdeutlich zu erkennen. Damit ist eine Ansprache alsMammut sicher. Die Darstellung von Los Casares(Abb. 28, 9) bildet ein Unikat in seiner Art. FrontaleDarstellungen von Herbivoren sind sehr selten, unddiese ist die einzige bekannte dieser Art einesMammuts bzw. einer Mammutmaske. Die Form desKopfes, die vermuteten Stoßzähne, der möglicheRüssel und die Behaarung lassen eine Interpretationals Mammut bzw. als Mammutmaske zu. Links vondieser Darstellung wurde eine zweite Gravur ebenfallsals Mammut interpretiert (Jordá 1983). Da dieseGravierungen die südlichste Darstellung einesMammuts sein könnten, wären eine neue, moderneUmzeichnung sowie detaillierte Fotos der Wandwünschenswert; leider erfüllt auch die neueste Publi-kation über Los Casares diesen dringenden Wunschnicht (Acosta und Molinero 2003, 79–81). Diesebeiden möglichen Mammutdarstellungen aus LosCasares werden aber von José Javier AlcoleaGonzález und Rodrigo de Balbín Behrmann bei derKartierung der Mammutdarstellungen im InnerenSpaniens gar nicht erwähnt (Alcolea und Balbín 2003,229). Man muss daraus folgern, dass die Autorendiese Gravierung nicht für Darstellungen vonMammuten halten.
Die Bestimmung der Darstellung aus La Palomera beiOjo Guareña (Balbín und Alcolea 1992, 406, Abb. 7)ist in der Literatur ebenfalls umstritten (Corchón et al.1996, 54) und scheint mir keine Elefantendarstellungzu sein (Abb. 28, 6). Das gleiche gilt für die vorkurzem publizierte Entdeckung von vier weiterenElephantidae-Darstellungen in der Höhle von ElPindal (Balbín et al. 1999). Drei davon werden vonJavier Fortea Pérez (2000) als Fiktion, eine (Abb. 28,3) als Möglichkeit betrachtet. Die Darstellungen vonEl Reguerillo (Abb. 28, 7) werden ihrerseits vonRodrigo de Balbín Behrmann und José Javier AlcoleaGonzález als hypothetisch und zweifelhaft bezeich-net (1992, 414; 1995, 106) und werden ebenfalls beider Kartierung der Mammutdarstellungen im InnerenSpaniens nicht berücksichtigt (Alcolea und Balbín2003, 229). Eine letzte Darstellung sollte aus derCueva de la Lloseta stammen (Balbín et al. 2003,127). Die Deutung als Mammut ist jedoch fraglich.
Die Darstellung aus El Castillo (Abb. 28, 1) zeigtkeine Details der Behaarung und wurde von Leonardiund Graziosi als mögliche Darstellung eines jungenMammuts, nach Eduardo Ripoll Perelló jedoch alsElephas antiquus angesprochen (Pericot und Ripoll1964, 93–96). Die Darstellung an sich erlaubt wedereine sichere Zuordnung als Elephas antiquus, noch alsMammuthus primigenius. Die weit verbreiteteMeinung, dass es sich um ein Mammut handeln müsseund dass eine Interpretation als Elephas antiquus einFehler sei (Barrière 1993b, 155), beruht auf dermindestens zum Teil falschen Annahme, dass auf derIberischen Halbinsel nur noch Mammuthus primi-genius als Vertreter der Rüsseltiere im Jungpaläoli-thikum existierte (siehe unten). Die Darstellung aus ElPindal (Abb. 28, 2) zeigt ebenfalls keine Behaarung.
Schließlich muss man erwähnen, dass zahlreichejungpaläolithische Darstellungen von Mammutenkeine Stoßzähne aufweisen (u. a. in El Pindal undGönnersdorf), während alle modernen Rekonstruk-tionen ausgewachsene Mammuts stets mit gewaltigenStoßzähnen darstellen (u. a. Lister und Bahn 1997).Auch hier sieht man eine zu einfache Übertragung desBildes des Mammuts, so wie man es von sibirischenFunden kennt, auf Europa. Es ist bekannt, dass dieElefantenkühe von vielen asiatischen Populationenkeine oder nur rudimentäre Stoßzähne besitzen(Gröning und Saller 1998, 466). Das könnte ebenfallsin manchen Mammutpopulationen der Fall gewesensein (Lister und Bahn 1997, 99). Lange Stoßzähnesind eine aufwendige energetische Investition, die sichgewissermaßen auch „lohnen“ muss. Stoßzähnespielen noch heute bei männlichen Elefanten eineRolle bei der Einschüchterung von Rivalen in derPaarungszeit und werden von beiden Geschlechternbei Rangeleien insbesondere um Nahrung eingesetzt.In einer Landschaft, wie in Mittel- und Westeuropawährend der letzten Eiszeit, in der im Winter kaum

72 Verbreitung der großen Jagdfauna in Mittel und Westeuropa im oberen Jungpleistozän
Abb. 28 Als Mammut publizierte jungpaläolithische Darstellungen aus Spanien. 1 El Castillo (Jordá 1983, 267, Abb. 2); 2 Pindal (Balbín et al. 1999, 86); 3 Pindal (Fortea 2000, 40); 4 Cuevadel Arco B (González und San Miguel 1996, 13); 5 La Peña del Candamo (Jordá 1983, 269, Abb. 4); 6 Ojo Gua-reña (Balbín und Alcolea 1992, 406, Abb. 7); 7 El Reguerillo (Lucas 2000, 134, Abb. 4); 8 Altamira (Jordá
1 2 3 4 5 6 7 8 9
10 11 12
1983, 267, Abb. 3); 9 Los Casares (Jordá 1983, 270); 10–12 Las Caldas (Corchón 1992, 48, Abb. 5, und 49,Abb. 6).

Mammut (Mammuthus primigenius) 73
Schnee fiel und es ausreichend Pflanzennahrung vomFrühling bis zum Sommer gab (Joger und Koch 1994,70ff.), könnten Stoßzähne für Weibchen überflüssiggewesen sein. Auch als Verteidigungswaffe darf manStoßzähne nicht überschätzen. Schon die körperlicheMasse von Mammuten und ihr ausgeprägtes Sozial-verhalten (wie man es insbesondere unter Elefanten-kühen und -kälbern kennt) waren ein ausreichenderSchutz. Der Mangel an vollständigen Schädeln vonMammutkühen aus Mittel- und Westeuropa lässt dieseFrage, ob es in Europa Mammutpopulationen ohneStoßzähne gab, jedoch bis auf Weiteres offen.
Knochenreste von Mammuten in Südeuropa
Es ist auffallend, dass sowohl in Spanien wie auch inItalien die maximale Verbreitung dieser Tiere bis weitnach Süden reicht (Abb. 30). Die Funde sind jedochälter als das Jungpaläolithikum. Die Reste von zweiIndividuen von Mammuthus primigenius aus derFundstelle von El Padul in der Provinz Granadagehören zum mittleren Pleistozän. Die Tiere sindwahrscheinlich im Sumpf versunken (Carrasco 1999;Made und Mazo 2001). Die Annahme, dass dieMammutreste aus Cardamone in Apulien, Süditalien,in das zweite Kältemaximum der letzten Eiszeit bzw.zwischen 22.000 und 18.000 BP zu datieren seinkönnten (Rustioni et al. 2003), scheint sich nur auf dieFaunenvergesellschaftung zu stützen und ist daher alsspekulativ zu betrachten. Antonio Tagliacozzo stelltdieselben Funde an den Beginn des Würms (Taglia-cozzo 1992, 79). Dies wird von den meisten Autorenebenso gesehen (u. a. Mussi 2001, 127; CapassoBarbato et al. 1991). Andererseits darf man nichtvergessen, dass die adriatische Küste während derletzten Eiszeit ein strengeres Klima als die thyrre-nische hatte (Mussi 2001, 503). Daher ist die Präsenzdes Mammuts während der letzten Eiszeit inSüditalien nicht a priori auszuschließen, sie ist jedochnicht belegt und erscheint nach dem Vergleich mit dergut datierten Fauna aus dieser Region als unwahr-scheinlich
In Portugal sind die angeblichen Reste vonMammuthus primigenius (Cardoso 1993, 290–293)umstritten (Zilhão 1997, 65), so dass diese hier alsnicht gesichert gelten müssen. Die meistens radiome-trisch oder archäologisch datierten jungpaläolithi-schen Fundstellen liegen jedoch viel weiter nördlich(Kantabrien, Arene Candide bei Savona und Settepo-lesini bei Ferrara).
Die einzige archäologische Fundstelle nördlich derApenninen mit Resten von Mammuthus primigenius,die mit modernen Methoden gegraben wurde, istRiparo Tagliente bei Verona. Die Schicht 37–36gehört aber zum Mittelpaläolithikum. Mittelpaläoli-thische Funde begleiteten auch einige andereMammutfunde aus der Po-Ebene (u. a. Asolo) und aus
Mittelitalien (u. a. Grotta della Iena). Auch in derGrotta del Principe in Ligurien wurde ein Stoßzahneines potentiellen Mammuts zusammen mit Levallois-Abschlägen in der Nähe der Feuerstelle A gefunden(Mussi 1992, 233). Nur in der Schicht P7 von AreneCandide, die ein 14C-Datum von 19.400 ± 230 BPerbracht hat (Bietti und Molari 1994, 24), kommenMammutfunde (fünf Reste von mindestens zweiIndividuen; Cassoli und Tagliacozzo 1994, 181–185;Alhaique 1994, 265) in einem jungpaläolithischenKontext vor. Zahlreiche weitere Mammutfunde ausItalien wurden entweder außerhalb eines stratigraphi-schen Zusammenhangs aufgesammelt (z. B. Lese-funde nach Überschwemmungen insbesondere in derPo-Ebene) oder wurden bei Grabungen, oft paläonto-logischer Natur, geborgen. Zu solchen Fundstellengehört auch Settepolesini di Bodeno (Gallini und Sala2000, 2001; Sala und Gallini 2002). Hier liegen dieFunde ca. 20 m unter der Oberfläche und sind für eineübliche Ausgrabung unerreichbar. Sie werden dahermit Hilfe einer „Saugpumpe“ herausgesaugt. DieDatierung einzelner Knochen erfolgte anhand vondirekten 14C-Datierungen. Bei unterschiedlichenKnochen von Mammuthus primigenius hat man Datenzwischen ca. 39.000 und 33.000 Jahren BP gewonnen.Diese Fundstelle und die weiteren Funde von Coelo-donta antiquitatis und Megaloceros giganteuserlauben die Annahme, dass es eine Mammutsteppe inder älteren Phase des Jungpaläolithikums in der Po-Ebene gegeben hat.
In Spanien scheint das Bild etwas anders zu sein.Auch hier sind viele paläontologische Fundstellen garnicht oder nur grob datiert worden. Datierungen wie„Würm-Eiszeit“ oder „Oberes Pleistozän“, diedurchaus für Paläontologen ausreichend sein können,erweisen sich für Archäologen als völlig unzurei-chend. In Nordspanien sind Mammutreste (Elfenbeinausgenommen) häufiger als allgemein angenommen.Sie verteilen sich über das ganze Jungpaläolithikum.So findet man sie im Châtelperronien und Aurignacienin Labeko-Koba, in der Übergangsschicht zwischenMittelpaläolithikum und Aurignacien sowie im Gra-vettien und Solutréen der Arbreda, im Magdaléniender Bora-Gran d’en Carreres, im Aurignacien von ElCierro, im Solutréen von Las Caldas, Cau de lesGoges und Cueto de la Mina. Dabei handelt es sichhäufig um Dentinreste von Molaren, so dass sich hierdie Frage stellt, ob solche Molaren nicht als Rohma-terial in die Höhle gebracht wurden.
Verbreitung von Elfenbein
Die Verbreitung von Elfenbein muss hier als eineeigenständige Quelle außerhalb der Verbreitung vonMammutresten angesprochen werden. Die Proble-matik, die sich dahinter verbirgt, zwingt weiterhin,diese soweit wie möglich wertungsneutral zu präsen-

74 Verbreitung der großen Jagdfauna in Mittel und Westeuropa im oberen Jungpleistozän
tieren, weil hier der Import von Elfenbein als Rohma-terial, als Fertigprodukt oder als fossiles Rohmaterialin Frage kommt. Die Präsenz von Elfenbein in einerarchäologischen Fundstelle setzt damit nicht voraus,dass Mammuts zur selben Zeit und in derselbenRegion gelebt haben müssen. Die 14C-Daten aus denMammutknochen aus Gönnersdorf sind deutlich älterals die Fundschicht, daher ist wahrscheinlich, dass sieschon als Fossilien aufgesammelt wurden (Hedges etal. 1998, 231). Zusätzlich könnten die Elfenbeinfundevon anderen Tieren als vom Mammut stammen. DieTatsache, dass Elephas antiquus in Kantabrien minde-stens bis zum Aurignacien in El Castillo inNordspanien und bis zum Ende des Mittelpaläoli-thikums in Foz do Enxarrique in Portugal überlebte(siehe das Kapitel über den Waldelefanten), zeigt, dassElfenbein auf der Iberischen Halbinsel (Abb. 29)mindestens bis 30.000 BP nicht zweifelsfrei dem
Mammuthus primigenius zugeordnet werden kann.Schließlich kann Elfenbein, wenn auch nur in wohläußerst seltenen Fällen, auch von Walross- undPottwahlzähnen stammen (Poplin 1983).
In Italien findet man Elfenbein in jungpaläolithischemKontext lediglich in den Grimaldi-Höhlen, nämlicheinige Venusfiguren vermutlich aus dem Gravettien,aus den Höhlen Barma Grande und Grotta del Principe(Bolduc et al. 1996) und als Anhänger bei einerGravettien-Bestattung in der Höhle von AreneCandide (Bolduc et al. 1996, 44). Deutlich zahlreichersind die Funde aus Spanien, die von Esteban ÁlvarezFernandez (2001; 2002) wie folgt aufgelistet wurden:
– Aus dem Châtelperronien: Elfenbeinreste ausLabeko-Koba.
Abb. 29 Fundstellen mit Elfenbein in Spanien und Italien. 1 Las Caldas; 2 Viña; 3 Cueto de la Mina; 4 LaRiera; 5 Llonin; 6 Altamira; 7 El Castillo; 8 El Pendo; 9 Labeko-Koba; 10 Jarama II; 11 Bora Gran d’en Car-reres; 12 Arbreda; 13 Grimaldi-Höhlen; 14 Arene Candide.

Mammut (Mammuthus primigenius) 75
– Aus dem Aurignacien: eine Geschossspitze aus ElCastillo, ein Pfriem oder Geschossspitze aus La Viña,Elfenbeinreste aus Labeko-Koba, ein wahrschein-licher Elfenbeinkern und Abfallprodukte aus Arbreda(Maroto 1994, 225-231).
– Aus dem Aurignacien bzw. Gravettien: Zweimögliche Geschossspitzen aus Cueto de la Mina.
– Aus dem Gravettien: Zwei Geschossspitzen in LaViña.
– Aus dem Solutréen: Ein Pfriem und Dentinfrag-mente in Cueto de la Mina, der mediale Teil einerGeschossspitze in El Castillo, eine Geschossspitze vonLlonin, drei Geschossspitzen aus Altamira, eineGeschossspitze in El Pendo, zwei unvollständigerhaltene, mit Kerben versehene Plättchen, von deneneines gelocht war, sowie unbearbeitete Elfenbeinresteaus der Höhle von Las Caldas, zwei Anhänger und einSchaft in La Riera, ein kleines, bearbeitetes Bruch-stück aus der Höhle von El Pendo.
– Aus dem Magdalénien: Eine Geschossspitze inAltamira, eine Geschossspitze und ein Schaft inLlonin, unbearbeitete Elfenbeinreste aus der Höhlevon Las Caldas, eine Elfenbeinfigur aus Jarama II,eine mögliche Geschossspitze aus der Bora Gran d’enCarreres (Pallarès und Wernert 1915–1920, 431;Canal und Carbonell 1989, 343).
– Möglicherweise aus dem Azilien: Ein Pfriem aus LaRiera.
Elfenbein war also in der nördlichen Hälfte Spanienswährend des ganzen Jungpaläolithikums und vielleichtsogar noch im Azilien (wenn diese Zuordnung stimmt,siehe Álvarez und Sánchez 2002, 169) als Materialbekannt.
Die jüngsten Fundplätze
Anhand der publizierten Daten lässt sich der langsameRückzug des Mammuthus primigenius in Europa gutverfolgen (Tab. 2), folgende Bemerkungen sindjedoch notwendig: Erstens stammen die Daten ausunterschiedlichen Laboratorien, und jeder hat, trotzinternationaler Standards, eigene Verfahren. Weiterhinsind die Oxford-Daten (OxA) AMS-Daten, währenddie anderen Daten meistens konventionelle 14C-Datensind. Drittens basieren unsere Informationen über dieletzten Mammuts auf viel zu wenigen Funden. Eineletzte entscheidende Bemerkung muss man über dieNatur der Daten äußern. Sie beweisen, wenn man dieMöglichkeit eines Imports ausschließen kann, die
Präsenz von Mammuten in einer bestimmten Regionbis zu einem bestimmten Datum. Sie beweisen aberkaum, ab wann diese Tierart ausgestorben ist oder abwann sie nicht mehr gejagt wurde. Wenn man diegeringe Zahl an ausgegrabenen, datierten Fundstellenmit den Tausenden von Tieren, die es im Laufe derJahrhunderte und Jahrtausende gegeben haben muss,in Relation stellt, kann man nur von einer statistischkaum relevanten Stichprobe sprechen.
Die jüngsten Knochenreste aus Italien stammenaus den Fundstellen von Settepolesini di Bodeno (ca.34.000 BP) und Arene Candide (ca. 19.500 BP). Fürdie Po-Ebene vermutet man eine Einwanderung sowieeinen Rückzug von Mammuten aus bzw. nachNordosten, während die Fundstellen in Ligurienwahrscheinlich von Frankreich aus entlang der Küsteerreicht wurden. Die Apenninen werden von manchenArchäozoologen als eine wichtige Grenze betrachtet(Cassoli und Tagliacozzo 1994, 199).
In Spanien datieren die jüngsten Mammutreste ausCueto de la Mina in Asturien 19.110±205 und17.545±205 BP, aus der Arbreda in Katalonien17.320±290 und 17.720±290 BP. Die sehr detail-reichen Gravierungen aus Las Caldas zeigen jedoch,dass Mammuts noch um ca. 13.370±110 BP denMenschen bekannt waren.
Ungefähr aus der gleichen Zeit (13.390±300 BP)stammen die Knochenreste aus der Fundstelle von LaColombière, im Osten Frankreichs. Die deutlichjüngeren Daten aus Praz Rodet (ca. 12.270±210 BP)scheinen jedoch zu zeigen, dass Mammuts in dieserRegion vielleicht als Restpopulation noch ca. 1000Jahre lang präsent waren. Älter sind die jüngstenDaten aus Österreich (ca. 15.560±200 BP) und Süd-(ca. 14.610±90 BP) und Ostdeutschland (ca.14.100±100 BP). Die Mammutreste aus Gönnersdorfsind um ca. 14.500 BP datiert, während die archäolo-gische Schicht der Fundstelle um 12.900–12.500 BPdatiert. Wie schon erwähnt, scheint damit die vonPoplin geäußerte Vermutung bestätigt, dass dieGönnersdorfer Mammutknochen aufgesammeltwurden (Poplin 1976, 50). Andererseits darf man nichtvergessen, dass bei so wenigen Daten wie inGönnersdorf, die außerdem von zwei unterschied-lichen Labors durchgeführt wurden, es eineEntscheidung der Archäologen ist, die Mammutresteabzusondern. In Osteuropa sind Freilandstationen wieKostienki über Jahrtausende immer wieder besuchtworden.
Zu einem „kontinuierlichen“ Rückzug derMammuts nach Osten und Norden passen ebenfallsdie Daten aus Schweden (ca. 13.090 BP) und dasjüngste Datum aus Dänemark (ca. 13.240 BP), dasheute jedoch umstritten ist36. Die Daten von Etiollesin der Nähe von Paris (ca. 12.000 BP), und von
36 Siehe Anmerkung ** zu Tabelle 2.

76 Verbreitung der großen Jagdfauna in Mittel und Westeuropa im oberen Jungpleistozän
Condover in England (ca. 12.300 BP) passen jedochnicht gut zu einem „kontinuierlichen“ Rückzug derMammuts nach Osten und Norden. Diese 14C-Datenwürden für eine Restpopulation zwischen Nordfrank-reich und Südengland in einer Zeit sprechen, in der bisjetzt keine Mammuts auf der Achse Schweden-Däne-mark-Deutschland-Österreich nachgewiesen wurden.
Die zahlreichen Mammutdarstellungen auf lokalenSchieferplatten aus Gönnersdorf und die Datierungder Fundstelle könnten ebenfalls für eine Restpopu-lation sprechen37. Obwohl eine solche Hypotheseanhand der uns heute zur Verfügung stehenden Datenvertretbar ist, basiert sie, wie schon erwähnt, auf zupunktuellen, oft sehr zufälligen Funden. Weiter nachOsten sind die Daten aus Kostienki, Russland (ca.11.000 BP) und aus Kunda, Estland (ca. 9.780 BP) zuerwähnen, die deutlich jünger als alle andere Datie-rungen aus Europa sind.
Die vor wenigen Jahren erfolgte Datierung derMammuts von der Wrangel-Insel (Vartanyan et al.1995, 4) auf 3.730±40 BP erschien erstaunlich. Hierhat eindeutig eine kleinere Form von Mammuten38 inIsolation lange überlebt, nachdem anderswo ihreArtgenossen schon längst ausgestorben waren. Eineganze Serie an Daten, von denen das jüngste jetzt bei3.685±60 BP liegt, lässt keinen Zweifel über dieDaten und zeigt, dass neue Ergebnisse und Datenunsere Vorstellungen über das Aussterben derMammuts noch ändern können.
Diskussion
Die Seltenheit von Mammuthus primigenius in denFaunenlisten Südeuropas zeigt eindeutig, dass dortseine klimatische Grenze erreicht war. Norditalien warnach dem heutigen Forschungsstand39 nur zu Beginndes Jungpaläolithikums kalt und trocken genug, um
die Lebensbedürfnisse dieser Tiere zu erfüllen.Danach scheint das Mammut aus Italien zuverschwinden. Reste von Mammuten kommen dannnur in Arene Candide in Ligurien und nur imSolutréen um die Zeit des zweiten Kältemaximumsvor. Während des Jungpaläolithikums sind dieHinweise auf Elefantidae, sei es als Knochen, Dentinoder Elfenbein, auf der Iberischen Halbinsel imVergleich zu Italien deutlich häufiger. Es ist zwar einSchwerpunkt im Solutréen zu erkennen, die Hinweisedecken jedoch das gesamte Jungpaläolithikum vomChâtelperronien über Aurignacien, Gravettien,Solutréen bis ins Magdalénien ab40. Eine feinereGliederung, die sich an den gesicherten klimatischenSchwankungen während des Jungpaläolithikumsaufhängen ließe und die die Präsenz von Mammutennur in „ganz spezifischen Momenten“41 auf der Iberi-schen Halbinsel nahe legen würde, ist zwar sehr inter-essant, jedoch zur Zeit nicht möglich. Die Reste oderHinweise auf Mammut, aber auch auf Wollnashorn,Rentier oder Saiga-Antilope, sind zahlenmäßig nichtausreichend und bestehen immer aus einzelnen oderwenigen Bruchstücken. Erfahrungsgemäß ist derUnterschied zwischen „so gut wie gar nichts“ und „garnichts“ immer sehr gering und häufig durch Zufällebestimmt. Daher dürften diese Knochenreste stati-stisch nicht überbewertet werden42.
Der Mangel an Hinweisen auf eiszeitliche Faunaauf der Iberischen Halbinsel für eine bestimmteZeitspanne bedeutet nicht automatisch, dass dieseseltenen Tiere aus Spanien verschwunden waren, undihr Auftreten mit ein paar Bruchstücken im Befundmuss nicht heißen, dass sie wieder eingewandert sind.Die Hypothese eines „Hiatus“ des Mammuthus primi-genius auf der Iberischen Halbinsel zwischen 17.500und 15.200 BC-kalibriert43 (erst Vorschlag beiÁlvarez und Jöris 1998, 81; schon Tatsache bei
37 Man nimmt an, dass die Menschen in Gönnersdorf Kontakte nach Norden hatten bzw. regelmäßig dorthingewandert sind. Das ortsfremde Rohmaterial lässt eine solche Annahme zu (Floss 1994, 195, Abb. 115b, und 220,Abb. 135b). Andererseits aber, wenn man Mammuts zwischen England, Frankreich und nördlich von Belgien undden Niederlanden vermutet, muss man auch die Möglichkeit anerkennen, dass Mammuts einmalig, gelegentlichoder sogar regelmäßig nach Süden gewandert sein könnten, ohne dass sie als Knochenmaterial in die uns bekann-ten Fundstellen dieser Zeit gelangen konnten. 38 Diese werden oft als Zwergmammuts angesprochen, da ihre Schulterhöhe nur 1,8 m erreichte (Lister undBahn 1997, 34). Die Zwergelefanten aus Sizilien waren jedoch noch kleiner. Ihre Schulterhöhe erreichte geradeeinmal 90 cm (Koenigswald 2002, 53).39 Wie schon mehrmals erwähnt, muss man berücksichtigen, dass in der Po-Ebene wegen der Mächtigkeit derSedimentation kaum paläolithische Fundstellen untersucht werden konnten. 40 Eine Spitze, die aus einer Azilien-Schicht stammt, soll, entsprechend der schlüssigen Argumentation vonEsteban Álvarez Fernández und Begoña Sanchez Chillon (2002, 169), entweder auf eine Vermischung zurückzu-führen sein, oder ihr Rohmaterial ist fossiles Elfenbein. Eine direkte Datierung des Objektes wäre wünschens-wert.41 „Periodos temporales especificos“, Álvarez und Sánchez 2002, 168, aber s. a. Álvarez und Jöris 1998, 67.42 Siehe diesbezüglich auch die Diskussion im Kapitel über die Saiga-Antilope.43 Die Autoren geben leider diese Zeitspanne als kalibrierte 14C-Daten (BC cal) an, dadurch ist jedoch jede Ver-gleichbarkeit mit älteren 14C-Daten (14C-BP) schwierig. Man kann nur schätzen, dass es sich um eine Zeitspannevon ca. 16.700 bis 14.500 BP handelt.
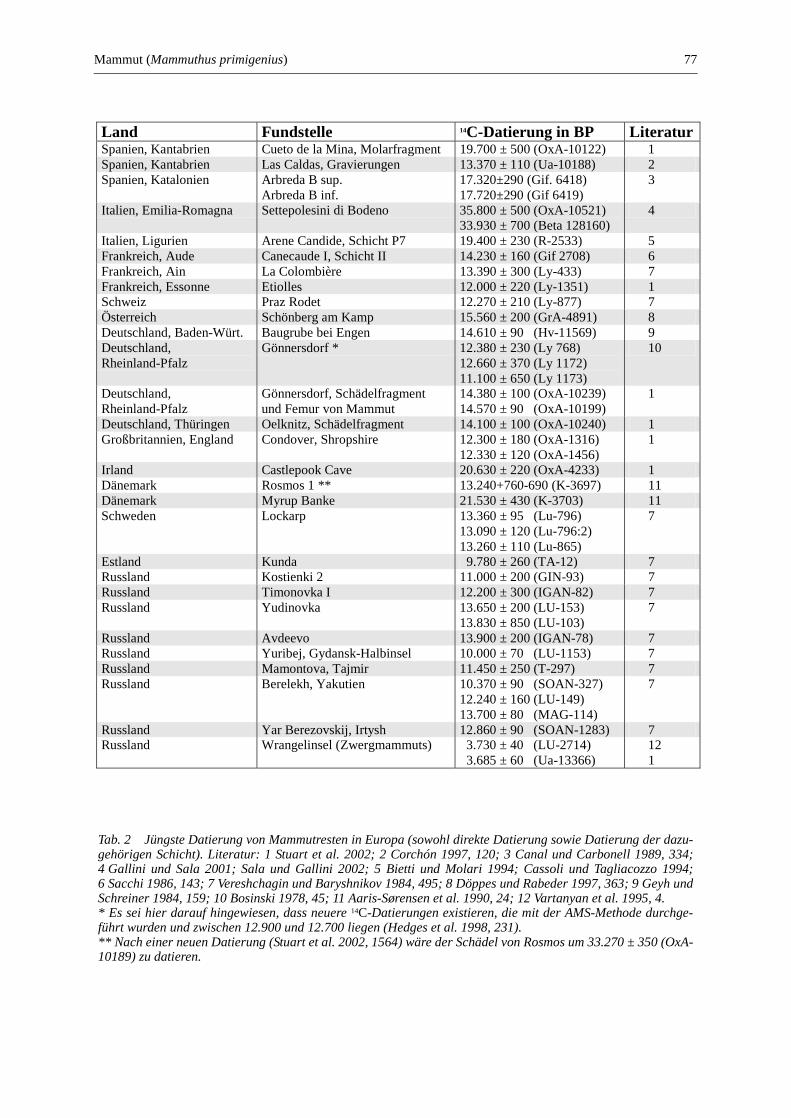
Mammut (Mammuthus primigenius) 77
Tab. 2 Jüngste Datierung von Mammutresten in Europa (sowohl direkte Datierung sowie Datierung der dazu-gehörigen Schicht). Literatur: 1 Stuart et al. 2002; 2 Corchón 1997, 120; 3 Canal und Carbonell 1989, 334;4 Gallini und Sala 2001; Sala und Gallini 2002; 5 Bietti und Molari 1994; Cassoli und Tagliacozzo 1994;6 Sacchi 1986, 143; 7 Vereshchagin und Baryshnikov 1984, 495; 8 Döppes und Rabeder 1997, 363; 9 Geyh undSchreiner 1984, 159; 10 Bosinski 1978, 45; 11 Aaris-Sørensen et al. 1990, 24; 12 Vartanyan et al. 1995, 4.* Es sei hier darauf hingewiesen, dass neuere 14C-Datierungen existieren, die mit der AMS-Methode durchge-führt wurden und zwischen 12.900 und 12.700 liegen (Hedges et al. 1998, 231).** Nach einer neuen Datierung (Stuart et al. 2002, 1564) wäre der Schädel von Rosmos um 33.270 ± 350 (OxA-10189) zu datieren.
Land Fundstelle 14C-Datierung in BP Literatur Spanien, Kantabrien Cueto de la Mina, Molarfragment 19.700 ± 500 (OxA-10122) 1 Spanien, Kantabrien Las Caldas, Gravierungen 13.370 ± 110 (Ua-10188) 2 Spanien, Katalonien Arbreda B sup.
Arbreda B inf. 17.320±290 (Gif. 6418) 17.720±290 (Gif 6419)
3
Italien, Emilia-Romagna Settepolesini di Bodeno 35.800 ± 500 (OxA-10521) 33.930 ± 700 (Beta 128160)
4
Italien, Ligurien Arene Candide, Schicht P7 19.400 ± 230 (R-2533) 5 Frankreich, Aude Canecaude I, Schicht II 14.230 ± 160 (Gif 2708) 6 Frankreich, Ain La Colombière 13.390 ± 300 (Ly-433) 7 Frankreich, Essonne Etiolles 12.000 ± 220 (Ly-1351) 1 Schweiz Praz Rodet 12.270 ± 210 (Ly-877) 7 Österreich Schönberg am Kamp 15.560 ± 200 (GrA-4891) 8 Deutschland, Baden-Würt. Baugrube bei Engen 14.610 ± 90 (Hv-11569) 9 Deutschland, Rheinland-Pfalz
Gönnersdorf * 12.380 ± 230 (Ly 768) 12.660 ± 370 (Ly 1172) 11.100 ± 650 (Ly 1173)
10
Deutschland, Rheinland-Pfalz
Gönnersdorf, Schädelfragment und Femur von Mammut
14.380 ± 100 (OxA-10239) 14.570 ± 90 (OxA-10199)
1
Deutschland, Thüringen Oelknitz, Schädelfragment 14.100 ± 100 (OxA-10240) 1 Großbritannien, England Condover, Shropshire 12.300 ± 180 (OxA-1316)
12.330 ± 120 (OxA-1456) 1
Irland Castlepook Cave 20.630 ± 220 (OxA-4233) 1 Dänemark Rosmos 1 ** 13.240+760-690 (K-3697) 11 Dänemark Myrup Banke 21.530 ± 430 (K-3703) 11 Schweden Lockarp 13.360 ± 95 (Lu-796)
13.090 ± 120 (Lu-796:2) 13.260 ± 110 (Lu-865)
7
Estland Kunda 9.780 ± 260 (TA-12) 7 Russland Kostienki 2 11.000 ± 200 (GIN-93) 7 Russland Timonovka I 12.200 ± 300 (IGAN-82) 7 Russland Yudinovka 13.650 ± 200 (LU-153)
13.830 ± 850 (LU-103) 7
Russland Avdeevo 13.900 ± 200 (IGAN-78) 7 Russland Yuribej, Gydansk-Halbinsel 10.000 ± 70 (LU-1153) 7 Russland Mamontova, Tajmir 11.450 ± 250 (T-297) 7 Russland Berelekh, Yakutien 10.370 ± 90 (SOAN-327)
12.240 ± 160 (LU-149) 13.700 ± 80 (MAG-114)
7
Russland Yar Berezovskij, Irtysh 12.860 ± 90 (SOAN-1283) 7 Russland Wrangelinsel (Zwergmammuts) 3.730 ± 40 (LU-2714)
3.685 ± 60 (Ua-13366) 12 1

78 Verbreitung der großen Jagdfauna in Mittel und Westeuropa im oberen Jungpleistozän
Álvarez und Sánchez 2002, 168) basiert ebenfalls aufeiner unzureichenden Datenbasis. In Spanien, dessenTopographie und dessen Forschungsgeschichte schonin anderen Kapiteln behandelt wurden, könnten dieseTiere in Refugien jahrhunderte-, wenn nicht sogarjahrtausendelang überlebt haben, ohne dass es dafürBeweise in Form von Knochen in den uns bekanntenFundstellen geben müsste. In Zentralspanien gibt es
weiterhin kaum Fundstellen mit erhaltener jungpaläo-lithischer Fauna, daher kann man nicht annehmen,dass diese Tiere dort nicht gelebt bzw. in bestimmtenPhasen überlebt haben, sondern man muss feststellen,dass aus dieser Region keine ausreichenden Datenexistieren. Die Präsenz von Elfenbein sowie diemöglichen (aber auch zweifelhaften) Mammutdarstel-lungen von Los Casares, El Reguerillo und Ojo
Abb. 30 Lage der als Mammuthus primigenius publizierten jungpaläolithischen Darstellungen (Kreis mitPunkt) und der Faunenreste vom Mammut aus Spanien und Italien aus der Zeit zwischen 40.000 und 10.000 BP(Dreiecke). Mit Fragezeichen wurden die Fundstellen von Elephas sp. ohne weitere Artzuordnung gekennzeich-net. Mit einem Stern wurde die Elephas antiquus-Fundstelle von Foz do Exanrique und mit einem Kreuz wurdendie paläontologischen und mittelpaläolithischen Fundstellen markiert. 1 Figueira Brava; 2 João Ramos; 3 Foz do exanrique; 4 La Peña de Candamo; 5 Las Caldas; 6 Cueto de laMina; 7 Pindal; 8 Altamira; 9 El Castillo; 10 Morin; 11 Cueva del Arco; 12 Labeko-Koba; 13 Bora Gran d’enCarreres; 14 Cau de les Goges; 15 Ojo Guareña; 16 El Reguerillo; 17 Los Casares; 18 Cantera del Buján;19 Arroyo del culebro; 20 La Olha I; 21 Clot del Llop; 22 Pedralves; 23 El Padul; 24 Arene Candide; 25 Sette-polesini di Bodeno; 26 Lambro; 27 Pizzighettone; 28 Borgo San Giacomo; 29 Breccia Serbaro; 30 Riparo Tagli-ente; 31 Asolo; 32 Vidor; 33 Buca della Iena; 34 Valdarno; 35 Cava Gorini; 36 Borgo Montello37; Canale delleAcque alte; 38 Cardamone.

Mammut (Mammuthus primigenius) 79
Guareña sind zumindest Hinweise dafür, dassElephantidae im Jungpaläolithikum in Zentralspaniengelebt haben könnten. Weiterhin muss der Mangel an größeren Knochen-stücken von Mammuten in den spanischen Fundstellenebenfalls durch einige Überlegungen ergänzt werden.Die Megafauna bietet so viel Fleisch (ein Mammutwiegt so viel wie ungefähr 20 Pferde, 25 Rothirscheoder 10 Steppenbisons), dass es häufig sinnvoller war,das Lager zu dem erlegten Tier zu versetzen, als denTierkadaver zu dem Lager zu bringen, wenn manannimmt, dass Mammuts und Wollnashörner auch inSpanien gejagt wurden. Weiterhin sind die meistenReste von Mammuten Elfenbein- und Dentinbruch-stücke. Es liegt auf der Hand, dass sie als Rohmaterial
in die Fundstellen, meistens Höhlen, gelangten. DasMitnehmen von Mammutknochen ist stark von ihrergezielten Anwendung abhängig (z. B. Münzel 2001),und da Holz in Spanien nie so selten war wiebeispielsweise in Mittel- und Osteuropa, kommt hierauch die Verwendung von Mammutknochen alsBrennmaterial oder als Baumaterial, wie z. B. inMezin, nicht in Frage (dazu Perlès 1977, 46–51). Dasheißt, dass man sich über die Abwesenheit bzw. dieSeltenheit von Mammutknochen in archäologischenFundstellen in Südeuropa nicht wundern darf. Inmanche Lager könnte sogar das knochenfreie Fleischvon Elephas sp. und Wollnashorn häufig eingebrachtworden sein, ohne dass man dafür Belege im archäolo-gischen Befund hätte.


Waldelefant (Elephas antiquus)
E. – Straight-tusked elephant, forest elephant; F. – Éléphant antique; S. – Elefante antiguo; Kat. – Elefant antic;P. – Elefante antigo; I. – Elefante antico.Ordnung: Proboscidea Illiger 1811; Familie: Elephantidae Gray 1821; Gattung: Elephas Linnaeus 1758; Art:Elephas antiquus Falconer und Cautley 1847. Der Waldelefant ist näher verwandt mit dem indischen (Elephas maximus) als mit dem afrikanischen Elefanten(Loxodonta africana), daher wird meistens der Name Elephas antiquus verwendet. Die Bezeichnung Palaeolo-xodon antiquus wird gelegentlich als Untergattung von Elephas verwendet (Koenigswald 2002, 51). Von Ele-phas antiquus stammen die Zwergelefanten auf den Inseln Sizilien und Tilos (Elephas falconeri Busk 1855) ab. Vorwiegend verwendete Literatur: Cardoso 1993; Cavarretta et al. 2001; Guérin und Patou-Mathis 1996.
Diese ausgestorbene Art kann hier nur am Randeangesprochen werden, da sie durch nur wenige sichereFunde in der Zeit nach 40.000 BP vertreten ist. DieInformationen über die Lebensweise von heutigenElefanten können für das Verständnis von Elephasantiquus zum Vergleich gezogen werden (siehe dasKapitel über das Mammut, S. 69–79).
Elephas antiquus ist eine an wärmeres undmilderes Klima angepasste Tierart. Daher wird er inMitteleuropa und Frankreich immer mit Interglazialenin Verbindung gebracht. Diese Art verschwindet ausMitteleuropa mit dem Beginn der letzten Eiszeit. AusItalien sind keine Reste von Elephas antiquus nach derSauerstoff-Isotopenstufe 4 bekannt (Bolduc et al.1996, 45). Auch in Sizilien ist die Zwergform Palaeo-loxodon falconeri im Jungpaläolithikum nicht mehrpräsent (Zampetti et al. 2000).
Auf der Iberischen Halbinsel hat Elephas antiquusbis zum Ende des Mittelpaläolithikums bzw. zumBeginn des Jungpaläolithikums überlebt (Abb. 31).Aus der Terrasse von Foz do Enxarrique stammenReste des Waldelefanten aus einer Schicht, die auf33.600±500 BP datiert wird (Cardoso 1993, 68; 1996,241). Möglicherweise gehören ebenfalls die Funde ausJoão Ramos und Figueira Brava dazu (Zilhão 1997,65). Diese Funde, die von Cardoso als mögliche Restevon Mammuthus primigenius interpretiert wurden(Cardoso 1993 und 1996, 241), sind eine distaleZahnlamelle aus Figueira Brava, aus einer Schicht, dieauf 30.930±700 BP datiert wird und ein Bruchstückeines Femurs aus der ein bis zwei Meter dicken
Schicht von João Ramos44. Ebenfalls aus dem Frühau-rignacien stammen fünf Reste von Elephas antiquusaus El Castillo in Spanien (Straus 1992, 70; Pike-Tayet al. 1999).
Eine sichere Trennung zwischen Elephas antiquusund Mammuthus primigenius scheint fast nur anhandder Zähne möglich zu sein. Weiterhin, wie schonerwähnt, sind postkraniale Reste von Elephas sp. inarchäologischen Fundstellen aus verschiedenen Grün-den selten. So konnte auch der untere Teil einesFemurs aus dem Gravettien des Reclau Viver (Canalund Carbonell 1989, 339) nur zu Elephas sp. ohneeine genauere Ansprache zugeordnet werden. Ausdem Gravettien der Arbreda, Schicht D, scheinthingegen der einzige Rest von Elephas sp. eher zuElephas antiquus als zu Mammuthus primigenius zugehören (Canal und Carbonell 1989, 327). Dadurchsteht fest, dass Elfenbeinreste aus Spanien nichtunbedingt Mammuthus primigenius zuzuordnen sind(Abb. 30). Im Bezug auf Italien ist die mittelpaläoli-thische45 Fundstelle Canale delle Acque Alte inte-ressant (Blanc et. al 1957). Hier scheinen die letztenExemplare von Elephas antiquus neben dem vonNorden eingewanderten Mammuthus primigeniusgelebt zu haben (Caloi et al. 1989, 81; Mussi undZampetti 1987, 8).
Es existieren keine eindeutigen Darstellungen vomWaldelefanten. Lediglich einige Mammutdarstel-lungen in Spanien könnten eventuell als Waldele-fanten interpretiert werden (Abb. 28).
44 Die Schicht ist zwar auf 14.400±330 BP datiert, die Mächtigkeit derselben lässt aber offen, wie alt die Fundetatsächlich sind. 45 Zwar ist eine Schicht (E2–E3) als „older than 55.000 years“ datiert worden (Blanc et al. 1957, 90–91). Da aberhier die 14C-Methode angewendet wurde, sollte dieses Datum als Tendenz verstanden werden.

82 Verbreitung der großen Jagdfauna in Mittel und Westeuropa im oberen Jungpleistozän
Abb. 31 Verbreitung von Resten und Darstellungen von Elephas antiquus in der Zeit des Jungpaläolithikums.Mit Dreiecken sind die sicheren (4 und 11), die wahrscheinlichen (6) und die möglichen (9 und 10) Reste vonWaldelefanten sowie die Elfenbeinfigur aus Jarama II (8) markiert. Mit einem Kreis sind die Darstellungenangegeben, die eventuell als Waldelefant in Frage kommen könnten. 1 La Peña del Candamo; 2 Pin-dal; 3 Altamira; 4 El Castillo; 5 Cueva del Arco; 6 Arbreda; 7 El Reguerillo; 8 Jarama II; 9 Figueira Brava; 10João Ramos; 11 Foz do Exanrique.

Paarhufer (Artiodactyla)
Nach Storch und Welsch (1993, 446): „Dritte undvierte Zehe dominieren, die anderen sind mehr oderweniger rudimentär. Die Prämolaren sind von den
Molaren schärfer unterschieden als bei Unpaarhufern.Obere Schneidezähne und Eckzähne oft reduziert.Pflanzenfresser.“
Wildschwein (Sus scrofa)
E. – Wild boar; F. – Sanglier; S. – Jabali; Kat. – Senglar; P. – Javali; I. – Cinghiale.Ordnung: Artiodactyla Owen 1848; Familie: Suidae Gray 1821; Gattung: Sus Linnaeus 1758; Art: Sus scrofaLinnaeus 1758; Unterarten (nach Herre 1986a, 42–43): Sus scrofa scrofa Linnaeus 1758, von Frankreich bisWeißrussland; Sus scrofa meridionalis Forsyth Major 1882, Sardinien und Corsica; Sus scrofa castilianusThomas 1912, in der Iberischen Halbinsel; Sus scrofa majori de Beaux und Festa 1927, in Italien. Vorwiegend verwendete Literatur: Herre 1986a; Heptner et al. 1966, 32–83.
Habitat: Das Wildschwein lebt in dichten Laub- undMischwäldern mit feuchten Gebieten und Sümpfen;sowohl in Bergen wie auch in Ebenen und Tälern,teilweise aber auch in Nadelwäldern. In den Karpatensoll das Wildschwein im Sommer in die Almenregionbis 1900 m, im Kaukasus bis 2600 m und in denBergen Mittelasiens bis 4000 m vordringen.
Verbreitung: Lokal ausgerottet (in England,Dänemark und Schweden im 17., in Mitteleuropa im18. und 19. Jahrhundert), wurde das Wildschweinvielerorts wieder eingebürgert oder ist wieder einge-wandert. Derzeit ist es in ganz Europa bis nachSüdschweden und Südfinnland vorhanden (Abb. 32).Es fehlt in Dänemark, auf den Britischen Inseln und ingroßen Teilen Siziliens. Es fehlt auch in manchen vomMenschen stark bewirtschafteten und besiedeltenRegionen wie z. B. in Teilen der Po-Ebene, derNiederlande und Belgiens.
Außerhalb von Europa (Abb. 33) ist es inNordafrika sowie in fast ganz Asien südlich des 60.Breitengrades außer in Teilen Zentralchinas und aufder Arabischen Halbinsel verbreitet. Es kommt auchauf Sumatra, Java und Neuguinea vor; auf Borneo lebteine andere Art, Sus barbatus. In Ägypten und imSudan ist es am Ende des 19. bzw. am Anfang des 20.Jahrhunderts ausgerottet worden. Seine derzeitigeVerbreitung und der Erfolg der Einbürgerungspro-gramme stehen in einem klaren Zusammenhang mitder Ausrottung des Wolfes, seines wichtigsten natür-lichen Feindes.
Nach Augusto Azzaroli (1971, 90) sind dieWildschweine auf Korsika und Sardinien erst vomMenschen im Neolithikum eingeführt worden.
Maximale Schneehöhe: 30 bis 40 cm.Körperbeschreibung: Das Wildschwein ist ein
massives, wuchtig wirkendes Tier. Der Hals ist sehrkurz, so dass der Kopf direkt in den Rumpf überzu-gehen scheint. Der vordere Rumpf ist kräftig, derhintere Teil etwas abfallend. Der Kopf ist gestrecktund keilartig. Die Beine sind kurz und dünn. DerSchwanz ist dünn. Bei erwachsenen Keilern ragen dieoberen und unteren Eckzähne aus dem Maul. DasGeruchsvermögen ist ausgezeichnet. Es soll eineGefahr über mehrere 100 m gegen den Wind witternkönnen (Heptner et al. 1966, 70). Sehr gut ist auch dasGehör. Die Augen dagegen sind klein, und dasSehvermögen ist beschränkt.
Abb. 32 Heutige Verbreitung von Sus scrofa inWest- und Mitteleuropa (nach Mitchell-Jones et al.1999, 381).

84 Verbreitung der großen Jagdfauna in Mittel und Westeuropa im oberen Jungpleistozän
Körpermaße: Die Körpermaße sind sehr unter-schiedlich, je nach Unterart und je nach Region. EineGrößenzunahme zeichnet sich von Südwesteuropanach Ost- und Nordosteuropa und Fernost ab. DieJagd, die Ausrottung der natürlichen Feinde an vielenOrten sowie die Vermischung mit Hausschweinenscheinen weiterhin zu einer deutlichen Reduktion derKörpermaße bei Wildschweinen geführt zu haben.Daher sollen die folgenden Angaben nur als richtungs-gebend verstanden werden. Die Körperlänge beiKeilern beträgt bis 170 cm, in Einzelfällen kann eineLänge von 200 cm erreicht werden. Berichte, nachdenen Keiler 230 bis 240 cm groß gewesen seinsollen, sind entweder zweifelhaft oder beziehen sichauf außergewöhnliche Individuen (Heptner et al. 1966,36). Bachen sind meistens kleiner, bis 150 cm lang.Die Schulterhöhe erreicht bei Keilern 90 bis 100 cm,bei Bachen 80 bis 90 cm. Kleine Keiler können etwa60 kg schwer sein (Bachen ca. 50 kg), besondersgroße Wildschweine des Fernen Ostens erreichenjedoch 300 bis 320 kg. Während der Brunstzeit und imWinter erleiden Keiler einen starken Gewichtsverlust,der bis zur Hälfte des Herbstgewichtes betragen kann.
Fell: Es besteht aus langen, steifen Borsten,darunter wächst eine kurze Unterwolle aus dünnen,welligen Wollhaaren. Die Borsten sind in der Mittedes vorderen Teiles des Rückens am längsten und siesträuben sich bei Erregung. Borsten sind beiWildschweinen oft proximal dunkler, distal heller. DieFarbe ist sehr unterschiedlich, je nach Region undBiotop. Sie kann dunkelbraun bis schwarz sein, z. B.in Weißrussland, aber es sind auch Populationenbekannt, bei denen die Farbe eher hellbraun bisstrohgelb oder sandfarbig ist, z. B. im Balchasch-Gebiet (Heptner et al. 1966, 34).
Druck auf den Boden: Die mittleren Zehen tragengewöhnlich die Körperlast (500 g/cm²), bei weicheremBoden stützen Wildschweine sich aber auf alle vierPhalangen (320–390 g/cm²).
Paarungszeit: November. Tragzeit: 4 bis 5 Monate.Setzzeit: März bis April, in der Regel bis sechs,ausnahmsweise bis zwölf Jungtiere. Unter natürlichenVerhältnissen erreicht nur jedes zehnte Tier das dritteLebensjahr.
Natürliche Lebenserwartung: Gewöhnlich 10 bis15, kann aber auch 25 Jahre erreichen.
Nahrung: Das Nahrungsspektrum des Wild-schweins ist sehr breit. Je nach Region und Jahreszeitgehören dazu u. a. Wurzeln, Knollen, Zwiebeln,Früchte, Beeren, Samen, Gras, Rinde, Zweige, Vogel-eier, Schlangen, Aas. Den größten Teil seiner Nahrungfindet es im Boden (unterirdische Pflanzenteile undTiere) dank seines stark entwickelten Geruchssinns.
Verhalten: Wildschweine sind vorwiegend nacht-aktiv. Keiler sind Einzelgänger, alle übrigen Tiereleben in kleinen Herden. Wenn sie bedrängt werdenbzw. wenn die Fluchtdistanz unterschritten ist oder dieTiere sich einfach gereizt fühlen, greifen Keiler oderBachen mit Frischlingen häufig an. In solch einerSituation bleibt dem Jäger nur übrig, die Flucht zuergreifen46.
Noch erstaunlicher ist das Verhalten einzelnerTiere gegenüber dem Meer. Nach Augusto Azzaroli(1971, 92–93) könnte eine bestimmte Art vonWildschweinen (Potamochoerus) übers Meer die InselMadagaskar von Afrika aus erreicht haben. Beobach-tungen von Wildschweinen, die mehrere Kilometervon der Küste entfernt im offenen Meer entdecktwurden, sind nicht allzu selten (Vigne 1988, 9). Dasspielt eine Rolle bei der Besiedlungsgeschichteeinzelner Inseln, insbesondere für die Zeit, in der dasMeeresniveau sehr niedrig lag und mancheMeeresarme deutlich schmaler waren als heute.
Wildschweine führen in ebenen Gebieten meistenskeine jahreszeitliche Wanderung durch. Es findet nurein gewisser Konzentrationswechsel zu Gunsten dernahrungsreichsten Plätze statt. In Gebirgen sindHöhenwanderungen die Regel. Bei Mangel anNahrung können jedoch auch in ebenen RegionenDistanzen von 100 bis 200 km überwunden werden.Dabei können die Wildschweine Strecken von bis zu40 km in vier Stunden zurücklegen (Heptner et al.1966, 67).
Abb. 33 Ursprüngliches Verbreitungsgebiet von Susscrofa (nach Heptner et al. 1966, 31).
46 Hansjürgen Müller-Beck berichtet von einem Fall aus dem Gebiet von Bad-Nauheim/Friedberg (Hessen), indem ein Keiler eine Lokomotive von vorne „angegriffen“ hat. Vom Keiler blieb kaum etwas übrig (mündlicheMitteilung).

Wildschwein (Sus scrofa) 85
Natürliche Feinde: Der wichtigste Feind ist der Wolf,der insbesondere in schneereichen Wintern diePopulation der Wildschweine in einem Gebiet starkdezimieren kann. Ein erwachsener Wolf kann 50 bis80 Wildschweine unterschiedlichen Alters im Laufeeines Jahres töten (Heptner et al. 1966, 76). KleineWildschweine können auch von Luchsen gejagtwerden.
Klimatische Aussage: Oft wird das Wildschweinals Anzeiger für dichten Wald angesehen. Aufgrunddes Hinweises, dass Wildschweine im Sommer auchin die Almregion vordringen (Heptner et al. 1966),sowie wegen des wiederholten Auftretens meistenseinzelner Funde in von Rentieren dominierten Faunen-komplexen ist bei Einzelfunden Vorsicht geboten. DerKörperbau, der eine Bevorzugung weicher Bödenanzeigt, sowie vor allem die Gewohnheit, Nahrungdurch Wühlen in der Erde aufzunehmen, beschränkensein Vorkommen wohl auf Regionen ohne Permafrost-Böden. Weiterhin ist es schon ab einer Schneetiefevon 30 cm in seinen Bewegungsmöglichkeiten starkeingeschränkt, und bei zusätzlichem Fehlen der natür-lichen Deckung im Wald sind Frischlinge und Bachenfür Wölfe eine leichte Beute.
Darstellungen aus dem Jungpaläolithikum
Nach den Zeichnungen und der Interpretation vonHenri Breuil und Hugo Obermaier (1935) gäbe es inAltamira zwei Darstellungen von Wildschweinen.Dieser Interpretation folgt auch André Leroi-Gourhan(1971, 372–374), der später (1984, 84) hinzufügt, dassdas Wildschwein in der paläolithischen Wandkunstnur durch diese Darstellung repräsentiert sei. Erstdurch eine neue Aufnahme und eine kritische Untersu-chung dieser Bilder (Freeman et al. 1987, 81–87;Sacchi 1993b, 163) konnte nachgewiesen werden,dass keine der beiden Darstellungen als sicheresWildschwein, sondern viel eher als Steppenbisonsanzusehen sind. Die Annahme, dass mindestens eineder zwei Darstellungen ein Wildschwein ist, lebt
jedoch in der Literatur weiter, indem André Leroi-Gourhan und Henri Breuil immer wieder zitiertwerden (u. a. Lorblanchet 1995, 125).
Eine wahrscheinliche Darstellung eines Wild-schweins ist das so genannte Wollnashorn von LosCasares. Während in der älteren Umzeichnungen vonCabré (Breuil 1952, 389; Corchón 1992, 58; Balbínund Alcolea 1992, 422, Abb. 26) die Interpretation als„Nashorn“ noch wahrscheinlich erscheint, muss diesenach der Aufnahme und der Beschreibung vonRodrigo de Balbín Behrmann und José Javier AlcoleaGonzález (1992, 417 und 422, Abb. 27) zurückge-wiesen werden. Das Horn des Nashorns existiert nicht,es soll sich um eine natürliche Spalte in der Felswandhandeln (Balbín und Alcolea 1992, 417), wie das Fotoebenfalls nahe legt (Balbín und Alcolea 1992, 423,Abb. 28).
Betrachtet man die Umzeichnung genau (Abb. 34),so sieht man, dass die Nase spitz zuläuft, und dasAuge liegt, in Vergleich mit den Nashorndarstel-lungen (Abb. 23), viel zu hoch in einem Bereich, derbei den Wollnashörnern schon im Schulterbereichliegt. Der Hals und der Kopf des Tieres gehen sanft indie Rückenlinie über, wo der höchste Punkt die Schul-terpartie ist. Das entspricht der Form einesWildschweins und nicht der eines Wollnashorns, beidem der Kopf gesenkt gehalten wird und derÜbergang zu den Schultern meistens abrupt ist (Abb.23). Der ventrale Bereich ist weiterhin nur leichtgewölbt; der tiefste Punkt liegt am Ende des Brust-korbes. Bei fast allen Wollnashorndarstellungen ist derBauch dagegen stark gewölbt und „hängt“ (Abb. 23).Daher ist meiner Ansicht nach diese Darstellung, trotzder Möglichkeit einer ausgeprägten „künstlerischenFreiheit“ eher als Wildschwein (Abb. 34) und nicht alsWollnashorn47 zu interpretieren.
Erst in der nacheiszeitlichen Felskunst der Spani-schen Levante kommt das Wildschwein häufiger vor(Abb. 35). Es bleibt jedoch immer ein Tier mit sekun-därer Bedeutung (Bandi und Maringer 1952). DieseDarstellungen zeigen jedoch, obwohl sie aus einem
Abb. 34 Links, wahrscheinliches Sus scrofa (nach der Umzeichnung von Balbín und Alcolea 1992, 422) ausLos Casares. Rechts, wissenschaftliche Darstellung von Sus scrofa (Sacchi 1993b, 161).

86 Verbreitung der großen Jagdfauna in Mittel und Westeuropa im oberen Jungpleistozän
anderen kulturellen Komplex stammen, deutlicheÄhnlichkeiten mit der Gravur von Los Casares (Abb.34) und bestärken mich in der Überzeugung, dass essich dort um eine Wildschweindarstellung handelt.
Darstellungen von Wildschweinen im Jungpaläoli-thikum bleiben dennoch außerordentlich selten, wobeimanche dieser Zuordnungen auch noch fraglicherscheinen (Abb. 36).
Reste aus dem Jungpaläolithikum
Sus scrofa fehlt im Moustérien der IberischenHalbinsel, im älteren Jungpaläolithikum ist es nurdurch einzelne Individuen vertreten und erst ab demSolutréen scheint es stärker vertreten zu sein (Straus1992, 56 und 84), erreicht aber vor dem Azilien niemehr als 10% der faunistischen Reste. Auf derApenninenhalbinsel sind Wildschweine nicht nurwährend des ganzen Jungpaläolithikums präsent,sondern der Anteil an ihren Resten liegt in manchenFundstellen im Süden im Bölling und Alleröd schonüber 30% (u. a. Grotta Paglicci und Grotta dell’Erica).In Südfrankreich ist das Wildschwein immer wiederdurch einzelne Funde repräsentiert. Darunter fällt dieFundstelle von La Ferrassie auf, wo unter 155Knochen 13,64% zum Wildschwein gehören (Delpech
1983, Tableau 6). In Mitteleuropa ist das Wildschweindurch einzelne Funde aus dem Aurignacien vomVogelherd (Baden-Württemberg, Deutschland) undvon der Grotte Princesse Pauline und Trou du Renard(Belgien) bekannt. Von einer WiederbesiedlungMitteleuropas kann man aber erst ab dem spätenMagdalénien sprechen (in Deutschland: Petersfels,Niederbieber und Stellmoor).
In Großbritannien fehlt es bis zum Ende derEiszeit. Die ältesten Funde stammen aus den frühme-solithischen Fundstellen von Star Carr und Thatcham(Koenigswald 1986).
Diskussion
Das weitgehende Fehlen von Sus scrofa nördlich desPérigords in Frankreich und in Mitteleuropa lässt sichleicht erklären. Hier fehlte ihm wegen der Trockenheitund des andauernden Frostes im Winter die Nahrungs-grundlage, denn den größten Teil davon findenWildschweine im Boden. Weiterhin wären sie in einerfreien Landschaft ohne Versteckmöglichkeiten eineleichte Beute für Löwen, Hyänen und Wölfe gewesen.Nur einzelne Individuen können es unter günstigenUmständen geschafft haben, weit in den Nordenvorzudringen.
Die Tatsache, dass Sus scrofa im älteren Jungpa-läolithikum Spaniens sehr selten ist, erklärt LawrenceGuy Straus damit, dass es gemieden wurde (Straus1992, 84). Cardoso fügt hinzu, dass es daran liegenkönnte, dass Wildschweine zu schwer zu jagen waren(Cardoso 1996, 239). Diese beiden Behauptungenerscheinen aus heutiger Sicht, wegen des schon obenerwähnten starken Selbstbewusstseins und derAngriffslust dieser Art, zunächst plausibel. StarkeZweifel treten aber bei der genaueren Betrachtung derEthologie, der Geschichte und der archäologischenDaten auf. Man darf nicht vergessen, dass dasWildschwein seinen heutigen besonders schlechtenRuf bekommen hat, da es aufgrund seiner anpassungs-fähigen Ernährungsweise seit dem Beginn desAckerbaus einer der größten Schädlinge für dieBauern ist. Ein Rudel kann in kürzester Zeit ganzeFelder verwüsten. Aus einem solchen bäuerlichenUmfeld stammt auch die altgriechische mytholo-gische Erzählung über den Fang des ungeheueren„Erymanthischen Ebers“, der erst vom Halbgott
47 Es sei hier ergänzend erwähnt, dass Rodrigo de Balbín Behrmann und José Javier Alcolea González in einemArtikel von 2003 unerklärlicherweise eine deutlich andere Umzeichnung dieses Tieres präsentieren (Alcolea undBalbín 2003, 236), wo doch ein Horn auf der Nase dargestellt wird und damit die Interpretation dieses Tieres alsNashorn untermauert werden soll. Diese Umzeichnung stimmt jedoch nicht mit der Evidenz des von denselbenAutoren 1992 publizierten Fotos überein (Balbín und Alcolea 1992, 423). Weiterhin widersprechen sie damit dia-metral ihrer eigenen Aussage, dass das Wollnashorn bis zu ihren Artikeln von 1992 und 1995 leichtfertig interpre-tiert worden sei und dass es gar kein Horn gäbe (Balbín und Alcolea 1992, 417). Die Umzeichnung von 2003stellt daher keine wissenschaftliche Umzeichnung, sondern vielmehr die spätere Interpretation von Alcolea undBalbín dar. Daher wird sie in dieser Arbeit nicht berücksichtigt.
Abb. 35 Wildschwein-Jagdszene aus der Höhle vonder Cueva Remigia, Wandmalerei in Grauschwarz,Arte Levantino (Bandi und Maringer 1952, 126,Abb. 160).
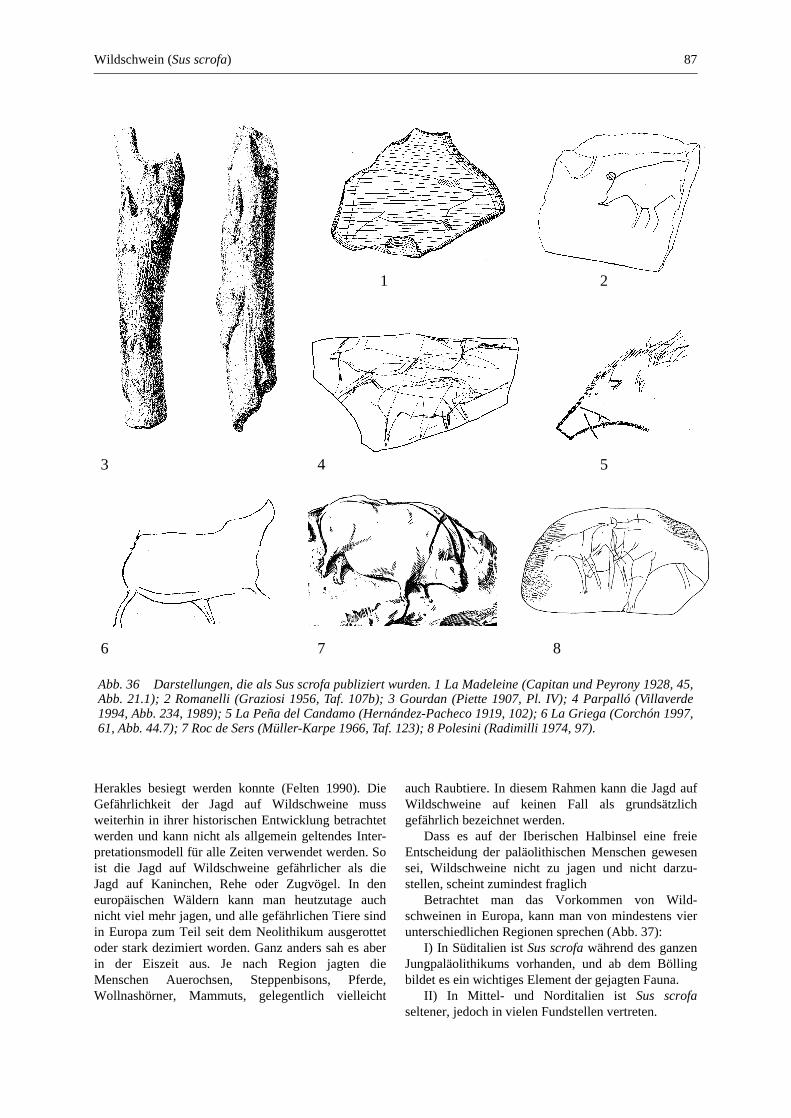
Wildschwein (Sus scrofa) 87
Herakles besiegt werden konnte (Felten 1990). DieGefährlichkeit der Jagd auf Wildschweine mussweiterhin in ihrer historischen Entwicklung betrachtetwerden und kann nicht als allgemein geltendes Inter-pretationsmodell für alle Zeiten verwendet werden. Soist die Jagd auf Wildschweine gefährlicher als dieJagd auf Kaninchen, Rehe oder Zugvögel. In deneuropäischen Wäldern kann man heutzutage auchnicht viel mehr jagen, und alle gefährlichen Tiere sindin Europa zum Teil seit dem Neolithikum ausgerottetoder stark dezimiert worden. Ganz anders sah es aberin der Eiszeit aus. Je nach Region jagten dieMenschen Auerochsen, Steppenbisons, Pferde,Wollnashörner, Mammuts, gelegentlich vielleicht
auch Raubtiere. In diesem Rahmen kann die Jagd aufWildschweine auf keinen Fall als grundsätzlichgefährlich bezeichnet werden.
Dass es auf der Iberischen Halbinsel eine freieEntscheidung der paläolithischen Menschen gewesensei, Wildschweine nicht zu jagen und nicht darzu-stellen, scheint zumindest fraglich
Betrachtet man das Vorkommen von Wild-schweinen in Europa, kann man von mindestens vierunterschiedlichen Regionen sprechen (Abb. 37):
I) In Süditalien ist Sus scrofa während des ganzenJungpaläolithikums vorhanden, und ab dem Böllingbildet es ein wichtiges Element der gejagten Fauna.
II) In Mittel- und Norditalien ist Sus scrofaseltener, jedoch in vielen Fundstellen vertreten.
Abb. 36 Darstellungen, die als Sus scrofa publiziert wurden. 1 La Madeleine (Capitan und Peyrony 1928, 45,Abb. 21.1); 2 Romanelli (Graziosi 1956, Taf. 107b); 3 Gourdan (Piette 1907, Pl. IV); 4 Parpalló (Villaverde1994, Abb. 234, 1989); 5 La Peña del Candamo (Hernández-Pacheco 1919, 102); 6 La Griega (Corchón 1997,61, Abb. 44.7); 7 Roc de Sers (Müller-Karpe 1966, Taf. 123); 8 Polesini (Radimilli 1974, 97).
1 2 3 4 5 6 7 8

88 Verbreitung der großen Jagdfauna in Mittel und Westeuropa im oberen Jungpleistozän
III) In Spanien und an der Mittelmeerküste Frank-reichs sind Wildschweine eher selten anzutreffen; erstab dem Alleröd, wenn nicht sogar ab dem Präborealsind sie deutlich häufiger.
IV) In Südwest- und im Zentrum Frankreichs sindWildschweine eine Ausnahme bis zum Alleröd.
In Nordfrankreich und Mitteleuropa sind Wild-schweine bis zum Ende des Jungpaläolithikums nurdurch Einzelfunde bekannt.
Die Lage der Iberischen Halbinsel als „Sackgasse“Europas kann dabei nur eine geringe Rolle gespielthaben. Süditalien ist gewiss durch die verlängerte Po-Ebene viel leichter als Spanien für Einwanderungenaus dem Osten erreichbar gewesen, aber aufgrund derhohen Geburtsraten und der hervorragenden Fähigkeitder Wildschweine zur Adaptation hätten diese auch in
Südspanien in kürzester Zeit bei gleichen Bedin-gungen genauso erfolgreich bzw. zahlreich seinmüssen und annäherend genauso oft auf der „Speise-karte“ der paläolithischen Menschen vertreten seinsollen wie in Süditalien. Da die Technokomplexe desJungpaläolithikums in Italien mit denen von Spanienin groben Zügen vergleichbar sind, kann man schlechtmit kulturellen oder technologischen Unterschiedenargumentieren. Vielmehr kann man annehmen, dassWildschweine, so wie sie in weiten Teilen Europaswährend des Jungpaläolithikums aus ökologischenbzw. klimatischen Gründen gefehlt haben, ausdenselben Gründen in Spanien und Portugal nicht sozahlreich wie in Italien gewesen sind. Motive dafürkönnten niedrigere Wintertemperaturen mit längerandauerndem Frost, höhere Schneeniederschläge odergeringerer Schutz vor Räubern (hauptsächlich Wölfen)durch weniger Unterholz im Wald gewesen sein.
Abb. 37 Mögliche Verbreitung von Sus scrofa in Europa während der letzten Eiszeit mit den wichtigsten im Texterwähnten Fundstellen. I bis IV: Unterschiedliche Verbreitungsregionen (Erklärung im Text). 1 Stellmoor; 2 Troudu Renard; 3 Princesse Pauline; 4 Niederbieber; 5 Ölknitz; 6 Vogelherd; 7 Roc de Sers; 8 La Madeleine;9 Gourdan; 10 La Peña del Candamo; 11 La Griega; 12 Los Casares; 13 Parpalló; 14 Polesini; 15 Romanelli.

Rothirsch (Cervus elaphus)
E. – Red deer (in Großbritannien) und elk (in Nordamerika); F. – Cerf élaphe, rouge ou noble; S. – Ciervocomun; Kat. – Cervol; P. – Veado; I. – Cervo elafo.Ordnung: Artiodactyla Owen 1848; Familie: Cervidae Goldfuss 1820; Gattung: Cervus Linnaeus 1758; Art:Cervus elaphus Linnaeus 1758.Unterarten: C. e. elaphus Linnaeus 1758, in Südschweden; C. e. hippelaphus Erxleben 1777, von den Pyrenäenbis Weißrussland und von Dänemark bis zur Apenninen- und Balkanhalbinsel; C. e. corsicanus Erxleben 1777,Korsika und Sardinien; C. e. atlanticus Lönnberg 1906, Norwegen; C. e. scoticus Lönnberg 1906, BritischeInseln; C. e. hispanicus Hilzheimer 1909, Iberische Halbinsel.Vorwiegend verwendete Literatur: Bützler 1977; Bützler 1986.
Habitat: Er lebt meistens im Wald. Im Sommer ist erauch in offenen Flächen anzutreffen. In Schottlandleben die Rothirsche das ganze Jahr in der offenenLandschaft.
Verbreitung: Nördliches Eurasien und Amerika(Abb. 38 und 39).
Körpermaße: Kopf-Rumpf-Länge 140 bis 250 cm;Schulterhöhe 100 bis 150 cm. Nach Osten werden siegrößer und schwerer. Gewicht 90 bis 350 kg.Hirschkühe sind um ein Viertel bis ein Drittel leichterals die Hirsche.
Verhalten: Sie sind vorwiegend dämmerungsaktiv.Alte Hirsche sind oft Einzelgänger. Der typischeBrunftschrei der Hirsche ist ein lautes Röhren. Beiausreichendem Nahrungsangebot und nicht allzuhoher Schneedecke überwintert der Rothirsch in dergleichen Region, in der er auch im Sommer gelebt hat.In Gebirgsgegenden mit starken Höhenunterschiedenunternimmt er Wanderungen von 50 bis 140 kmLänge, wobei 1000 bis 1500 m Höhenmeterüberwunden werden können (Heptner et al. 1966,192).
Fluchtdistanz: Es ist schwer, über eine allgemeineFluchtdistanz für diese Tiere zu sprechen. In manchenNaturschutzgebieten, insbesondere dort, wo dieseTiere nicht gejagt werden und relativ oft mitMenschen zu tun haben, kann die Fluchtdistanzweniger als 10 m betragen (Bützler 1977, 145).
Paarungszeit: September bis Oktober. Tragzeit: 8Monate. Setzzeit: Mai und Juni, in der Regel einJungtier, selten zwei.
Natürliche Lebenserwartung: bis 20 Jahre.Nahrung: Von der Jahreszeit abhängig äsen sie
Gräser, Kräuter, Triebe, Blätter, Flechten, Pilze,Beeren, Früchte, Eicheln, Kastanien, Bucheckern,Rinde und Feldfrüchte.
Geweih: Eine Geweihstange kann bis über 1 mLänge, 7 kg Gewicht und bis zu zwölf Endenerreichen. Der Rothirsch verliert das Geweih vonFebruar bis März, wobei dieses sofort wieder anfängtnachzuwachsen.
Klimatische Aussage: Eine starke Präsenz vonRothirsch ist ein klarer Hinweis auf mildes Klima undWald, er kann aber auch in Regionen mit Durch-schnittstemperaturen im Januar von bis zu -35 °Cvorkommen.
Darstellungen aus dem Jungpaläolithikum
Es ist interessant zu vermerken, dass bei Rothirschdar-stellungen fast immer zwischen männlichen Hirschenund Hirschkühen unterschieden wird (u. a. Sauvet1993, 84). Diese Praxis basiert hauptsächlich auf derAnwesenheit oder Abwesenheit von Geweihen; andereFaktoren spielen eine viel geringere Rolle (Crémadès1993 und 1997). Männliche Hirsche sind weiterhinfast immer mit voll ausgebildetem Geweih dargestellt(Abb. 40). Es ist jedoch schwierig, darin eine jahres-zeitliche Information zu lesen, da die paläolithischeKunst eine symbolische Komponente hat, die jenseitsdes einfachen Bezuges auf das reelle Leben liegt(Crémadès 1997, 80).
Abb. 38 Heutige Verbreitung von Cervus elaphus inEuropa (nach Mitchell-Jones et al. 1999, 389).

90 Verbreitung der großen Jagdfauna in Mittel und Westeuropa im oberen Jungpleistozän
Darstellungen von Rothirschen sind besonders häufigin Südeuropa, wo sie immer auch in der Fauna belegtsind. Nördlich der Pyrenäen sind sie gut vertreten(siehe das Kapitel „Mögliche Interpretation der Datenund Ergebnisse“, S. 167–175). Ihre Darstellungen sindin Frankreich jedoch nicht gleichmäßig verbreitet.Dort, wo die Darstellungen von Vertretern desMammuthus-Coelodonta-Faunenkomplexes zahlreichsind, sind Darstellungen von Rothirschen selten. InChauvet, wo 66 Mammut-, 65 Wollnashorn- und 12Rentierdarstellungen gezählt werden konnten, wurdennur 2 Bilder von Rothirschen entdeckt (Clottes 2003).In Rouffignac, wo es 158 Mammut- und 11 Wollnas-horndarstellungen gibt, konnte keine einzigeRothirschdarstellung entdeckt werden (Plassard 1999,62). In Font de Gaume, wo 29 Mammuts, 17 Rentierebzw. Cerviden und 2 Nashörner erkannt wurden(Leroi-Gourhan et al. 1984, 131), konnte nur eineDarstellung mit Sicherheit dem Rothirsch zugeordnetwerden (Capitan et al. 1910, 182). Umgekehrt, dortwo in Frankreich mehrere Rothirsche abgebildet sind,sind wenige oder gar keine kälteliebenden Tieredargestellt. Dies ist der Fall in Cosquer, wo sechsRothirsche und keine Mammuts, Wollnashörner oderRentiere dargestellt sind. In Lascaux wurden 88Rothirsche (Leroi-Gourhan und Allain 1979, 344)gegenüber einem fraglichen Rentier (Leroi-Gourhan etal. 1984, 200) und einem einzigen Nashorn, das einWollnashorn, aber auch ein Steppennashorn seinkönnte (siehe das Kapitel über das Steppennashorn, S.
67–68), gezählt. Man muss also daraus schließen, dassRothirsche und kälteliebende Fauna sich in der Regelausschließen. Dies erscheint zwar logisch, ist abernicht selbstverständlich. Es sei hier erwähnt, dassAndré Leroi-Gourhan die Komposition Steppenbison-Mammut-Rothirsch in der Liste der möglichenKompositionen aufführt, diese aber gleichzeitig inkeiner Höhle als ein Hauptthema in der Wandkunsterwähnt (Leroi-Gourhan 1995, 584–585).
Reste aus dem Jungpaläolithikum
Der Rothirsch ist in Südeuropa unter der großenJagdfauna die wichtigste Beute gewesen. In Deutsch-land und Frankreich kommt er während des ganzenJungpaläolithikums, meistens in geringer Zahl, invielen Fundstellen vor (siehe Fundstellenkatalog). Esist interessant zu vermerken, dass dort, wo Cervuselaphus über 70% der Fauna darstellt, er fast immervon relativ hohen Anteilen an Capreolus capreolusoder Sus scrofa begleitet wird (z. B. Le Flageolet I,Schicht VII; La Riera; Riparo Tagliente; GrottaPolesini; Grotta della Cala). In der Regel ist dieAnzahl an Resten von Cervus elaphus umgekehrtproportional zu der Anzahl an Resten von Capra ibexbzw. Capra pyrenaica. Sehr deutlich ist dies in RiparoTagliente und Grotta Paglicci zu sehen. In Italien, dortwo Cervus elaphus überwiegt, ist Capra ibex kaumvertreten, z. B. in Grotta Polesini, Palidoro, Grottadella Cala, Grotta della Calanca. In Spanien und inPortugal sind Steinbock und Rothirsch dagegen häufigdie zwei wichtigsten Spezies, z. B. in Parpalló,Malletes, Cendres, Cova Beneito, Tossal de la Roca,Nerja, Ekain, Tito Bustillo, La Riera, Rascaño,Fontainhas, Cabeco do Morto Caldeirao und Colum-beira.
Diskussion
Cervus elaphus ist der einzige unter den in wärmerenRegionen lebenden europäischen Hirschen, derzahlreich sowohl in der Kunst als auch in der Faunavertreten ist. Dama dama und Capreolus capreolussind unter den Faunenresten sehr selten und in derpaläolithischen Kunst nur in einzelnen Darstellungenvorhanden.
Das Verhältnis zwischen der Anzahl an Resten vonCervus elaphus und an Resten von Capra ibex bzw.Capra pyrenaica scheint in hügligen und bergigenRegionen das Verhältnis zwischen Wald und baum-loser Landschaft widerzuspiegeln.
Da Rothirsche während des Jungpaläolithikums inganz Europa nachgewiesen sind, wird hier auf eineVerbreitungskarte verzichtet.
Abb. 39 Verbreitung von Cervus elaphus in histori-scher Zeit.
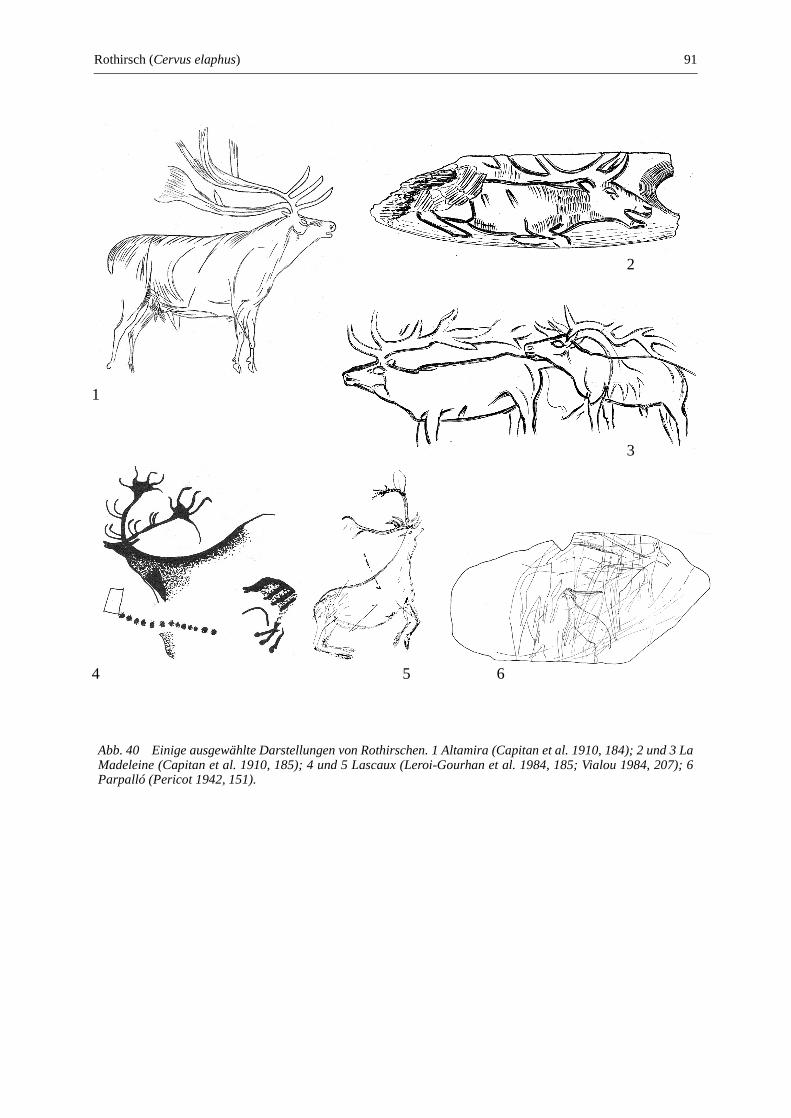
Rothirsch (Cervus elaphus) 91
Abb. 40 Einige ausgewählte Darstellungen von Rothirschen. 1 Altamira (Capitan et al. 1910, 184); 2 und 3 LaMadeleine (Capitan et al. 1910, 185); 4 und 5 Lascaux (Leroi-Gourhan et al. 1984, 185; Vialou 1984, 207); 6Parpalló (Pericot 1942, 151).
2
1 3 4 5 6


Damhirsch (Dama dama)
E. – Fallow deer; F. – Daim; S. – Gamo; Kat. – Daina; P. – Gamo; I. – Daino. Ordnung: Artiodactyla Owen 1848; Familie: Cervidae Goldfuss 1820; Gattung: Dama Frisch 1775; Art: Damadama (Linnaeus 1758). Unterarten: Europäischer Damhirsch (D. d. dama Linnaeus 1758) und Mesopotamischer Damhirsch (D. d.mesopotamica Brook 1875).Vorwiegend verwendete Literatur: Heidemann 1986.
Habitat: Hauptsächlich lichte Laub- und Misch-wälder. Nach Donald und Norma Chapman (1980)leben von allen Damhirschen auf der Welt etwa 31%in Laubwäldern, 31% in Mischwäldern, 14% in reinenNadelwäldern, 6% in Buschgebieten, 6% in Savannenund 1% in Tundren oder alpiner Vegetation.
Heutige Verbreitung: Das natürliche nacheiszeit-liche Vorkommen ist unklar, da diese Art in Europa inihrem gesamten Verbreitungsgebiet anscheinendausgerottet und später vielerorts weitgehend wiedereingebürgert wurde. Während einige Autoren dasnatürliche Verbreitungsgebiet auf die Türkei undSüdosteuropa beschränkt sehen (u. a. Niethammer undKrapp 1986, 144), rechnen andere mit fast demgesamten Mittelmeerraum für die Unterart D. d. damaund Syrien, Palästina, Mesopotamien, West- undSüdwestiran für die Unterart D. d. mesopotamica(Heptner et al. 1966, 218, Abb. 65). Entsprechendunklar ist, inwieweit die Römer, die diese Art inseuropäische Mittelmeergebiet eingeführt haben, diesenatürliche Verbreitung beeinflusst bzw. veränderthaben können.
Ab dem 8. Jahrhundert n. Chr. ist diese Art auch inMitteleuropa präsent (Niethammer und Krapp 1986,144). In Europa ist D. d. dama heute bis nachSüdskandinavien und zu den Britischen Inselnverbreitet (Abb. 41).
Körpermaße: Kopf-Rumpf-Länge 130 bis 230 cm;Schulterhöhe 80 bis 110 cm; Gewicht 35 bis 200 kg.Hirsche sind größer und schwerer als Hirschkühe. DasGeweih hat eine Länge von bis zu 80 cm.
Paarungszeit: Ende Oktober bis November.Tragzeit: circa 7 Monate. Setzzeit: April und Mai,meist ein, selten bis drei Jungtiere.
Natürliche Lebenserwartung: bis 20 Jahre.Nahrung: Gräser, Kräuter, Kastanien, Eicheln,
Bucheckern, Triebe und Rinde.Verhalten: Der Damhirsch ist vorwiegend dämme-
rungsaktiv und bildet nahezu während des ganzenJahres Gruppen, die man als Mutterfamilie,Weibchengruppe, Hirschgruppe, Jugendgruppe odergemischter Verband beschreiben kann. Gelegentlichfindet man auch Einzeltiere. Damhirsche lebenmeistens in ein und demselben Revier während des
ganzen Jahres und gelten dadurch als eindeutigesStandwild. Es sind jedoch auch Wanderungen von biszu 90 km bekannt (Heidemann 1986, 154).
Klimatische Aussage: Der Damhirsch gilt alsVertreter von warmem, eher trockenem Klima. NachWighart von Koenigswald (1986, 151) verschwindetder Damhirsch schon am Beginn der letzten Eiszeitaus Mitteleuropa und später auch aus den nördlichenMittelmeerregionen. Da Donald und Norma Chapman(1980) behaupten, dass der Damhirsch durchausvereinzelt in Tundren- bzw. in hochalpinen Regionenvorkommen kann, dürfen einzelne Reste odermögliche Abbildungen dieser Art nicht als Anzeigervon wärmeren Klimaverhältnissen bzw. von Waldüberbewertet werden.
Darstellungen aus dem Jungpaläolithikum
Nach Eduardo Ripoll Perelló (1984, 270) soll dieGravierung eines Cerviden aus der Höhle von Parpalló(Pericot 1942, 224, Abb. 488) eine unsichereDarstellung eines Damhirsches sein. Die Darstellungdes Geweihes ist jedoch so unvollständig, dass einesolche Annahme sehr spekulativ ist. Zu den als
Abb. 41 Heutige Verbreitung von Dama dama (nachMitchell-Jones et al. 1999, 387).

94 Verbreitung der großen Jagdfauna in Mittel und Westeuropa im oberen Jungpleistozän
Damhirsch publizierten Darstellungen sind hier dievon Addaura, El Buxu und Les Combarelles(Crémadès 1993, 146–147) zu zählen (Abb. 42, 1–3).Diesbezüglich muss man jedoch die hohe Variabilitätder oberen Geweihverzweigung bei Cervus elaphusbetonen (Bützler 1986, 112), die zu geradezu abson-derlichen Geweihformen führen kann, so wie man esoft in Jagdmuseen zu sehen bekommt. Daher mussman damit rechnen, dass alle „Damhirschdarstel-lungen“ Rothirsche sein könnten.
Reste aus dem Jungpaläolithikum
Aufgrund der wenigen als Dama dama bestimmtenReste scheint es, dass diese Art bis zum Beginn desJungpaläolithikums in Südeuropa überlebt hat(Abb. 43). In Saint-Jean de Verges, Frankreich, sindReste von mindestens zwei Individuen in derAurignacien-Schicht bestimmt worden (Bahn 1983,Tab. 24, 26). Im westlichen Languedoc undRoussillon wurde der Damhirsch im mittlerenMagdalénien der Grotte Gazel und der GrotteCanecaude I und im oberen Magdalénien der Grottede Belvis nachgewiesen (Sacchi 1986, 128, 143 und179). In Portugal ist er nach 30.000 BP in derFundstelle Gruta Nova da Columbeira, Algar de JoãoRamos sowie in Pedreira das Salemas bestimmtworden (Cardoso 1993, 169; Cardoso 1996). Damadama ist in Italien in verschiedenen Schichten derGrotta dei Fanciulli gefunden worden (Palma diCesnola 1993, 194), außerdem ist er im Aurignaciender Grotta della Fabbrica (Mussi 2001, 180), imAurignacien der Grotta del Fossellone (Mussi 1992,360), im Aurignacien der Grotta Barbara (Mussi 1992,362), im Uluzziano und Aurignacien der Grotta diCastelcivita (Masini und Abbazzi 1997, 39) und imAurignacien des Abri Perriere Sottano auf Sizilien(Mussi 1992, 352) präsent. In der Grotta della Cala,einer Station Süditaliens, erreicht er mit 152 Resten in
der Schicht des Uluzziano 43,8% der Faunenreste undbleibt auch in den darüber liegenden Schichten desAurignacien gut vertreten (Benini et al.1997, 70).
Diskussion
Es fällt auf, dass Dama dama nur in ganz wenigenFundstellen, meist durch wenige Funde vertreten ist.Eine einleuchtende Erklärung dafür gibt João LuisCardoso. Er schreibt, „dass es schwierig ist, Zahn- undKnochenreste von Dama dama und Cervus elaphus zutrennen; da es sich um Cerviden von mittlerer Größeund mit einer erheblichen Variabilität in den Dimen-sionen handelt, sind die auf Größenunterschiedenbasierenden Argumente nicht sehr zuverlässig. Nurdas Geweih ist das einzige, sofortige und sichereUnterscheidungsmerkmal. [...] So entsteht gewisser-maßen ein Teufelskreis: einerseits rechtfertigt derscheinbare Mangel an Dama dama in den pleisto-zänen Fundstellen keine systematische Studie überdessen Trennung von Cervus elaphus; andererseitserlaubt es der Mangel an solchen Studien nicht, diesezwei Tierarten auseinander zu halten“ (eigeneÜbersetzung aus Cardoso 1993, 169). Dies kann auchdie Erklärung dafür sein, dass in den aufgelistetenFundstellen des Jungpaläolithikums Spaniens Damasp. nur in El Castillo, in einer Schicht des Aurignacien(Straus 1992, 262), bestimmt wurde, obwohl er inPortugal und Südfrankreich gut belegt ist. Neben denSchwierigkeiten in der sicheren Bestimmung vonDama dama sei noch auf die zum Teil sehr schlechtenErhaltungsbedingungen für Knochen auf der Iberi-schen und auf der Apenninenhalbinsel hingewiesen.
Trotzdem scheint es so, als ob der Damhirsch mitdem zweiten Kältemaximum in Italien ausstirbt, umdann in römischer Zeit wieder angesiedelt zu werden(Mussi 2001, 304), wobei manche Autoren meinen,dass der Damhirsch in Süditalien bis zum Ende desPleistozäns überlebt haben könnte (Masseti und
Abb. 42 Als Damhirsch publizierte Darstellungen des Jungpaläolithikums. 1 Addaura (Brizzi 1977, 209); 2 El Buxu (Ripoll 1984, 271); 3 Les Combarelles (Novel 1987, 92).
1 2 3

Damhirsch (Dama dama) 95
Rustioni 1988; Masseti et al. 1995, 92). Im Languedococcidental und im Roussillon, wo er immer durchwenige oder einzelne Funde vertreten ist, scheint er imMagdalénien überlebt zu haben (Sacchi 1986, 128,143 und 179). Nimmt man an, dass diese Bestim-mungen korrekt sind48, kann es sich um kleineRückzugsgebiete handeln. Dominique Sacchi betont,dass in Belvis die Hinweise auf mesophile undthermophile Pflanzen beweisen, dass dort einRefugium auf mittlerer Höhe (960 m) existierte, dasdurch seine Ausrichtung nach Süden begünstigt war.Aus einem ähnlichen refugialen Kontext stammt
eventuell der „Cervus dama somonensis“ (dieBestimmung ist jedoch unsicher) aus der Feuerstelle Eder Grotta dei Fanciulli, die dem Epigravettianoevoluto zugeordnet wird (Mussi 2001, 538).
Um das Bild der Fundstellen mit Resten von Damhir-schen zu ergänzen, sei hier darauf hingewiesen, dassDama dama ebenfalls in Slowenien am Ende derEiszeit (Pohar 1997) und vielleicht noch im Neoli-thikum in Süditalien präsent war (Bökönyi 1982, 346).
Abb. 43 Im Text erwähnte Fundstellen mit Resten (Dreiecke) oder möglichen Darstellungen (Kreise mit Frage-zeichen) von Dama dama. Die Verbreitungskartierung ist als Hypothese zu betrachten, da für eine sichere Aus-sage die Daten nicht ausreichend sind. 1 Les Combarelles; 2 El Buxu; 3 Addaura; 4 Saint-Jean de Verges,5 Gazel; 6 Canecaude; 7 Belvis; 8 El Castillo (Dama sp.); 9 Algar de João Ramos; 10 Gruta Nova da Colum-beira; 11 Grotta dei Fanciulli; 12 Grotta La fabbrica; 13 Grotta del Fossellone und Grotta Barbara; 14 Grottadi Castelcivita; 15 Grotta della Cala; 16 Perriere Sottano.
48 Manche dieser Bestimmungen sind ziemlich alt und es gibt keinen Hinweis auf die Art der bestimmten Reste.

96 Verbreitung der großen Jagdfauna in Mittel und Westeuropa im oberen Jungpleistozän
Die vorgeschlagene Verbreitung der Damhirsche imJungpaläolithikum ist daher als Hypothese zubetrachten, da die Daten für eine sichere Aussagenicht ausreichend sind. Vielleicht waren Damhirscheausschließlich entlang der Mittelmeerküste verbreitet.Dort sicherte das nicht allzu kalte Meer (Abb. 8)mildere Temperaturen. Im Inland waren sie wahr-scheinlich von den Rothirschen verdrängt worden.Rothirsche sind am Mittelmeer etwas kleiner (u. a.Cardoso 1993, 167–168; Bützler 1986, 121) undkönnten daher im Jungpaläolithikum die ökologischeNische der Damhirsche besetzt haben.
Es sei hier schließlich darauf hingewiesen, dass es inder älteren Literatur manche Autoren davon ausgehen,dass keine sicheren Reste von Damhirschen an derwestlichen und zentralen Mittelmeerküste aus demoberen Pleistozän und frühen Holozän gibt (siehe dazuUerpmann 1987, 57). Eine systematische Studie überdie europäischen Reste von Damhirschen aus demoberen Pleistozän wäre wünschenswert.

Reh (Capreolus capreolus)
E. – Roe deer; F. – Chevreuil; S. – Corzo; Kat. – Cabirol; P. – Corço; I. – Capriolo.Ordnung: Artiodactyla Owen 1848; Familie: Cervidae Goldfuss 1820; Gattung: Capreolus Gray 1821; Art:Capreolus capreolus (Linnaeus 1758).Vorwiegend verwendete Literatur: Lehmann und Sägesser 1986; Heptner et al. 1966.
Habitat: Rehe verbergen sich tagsüber in dichtemWald und weiden in der Dämmerung in offenemGelände.
Verbreitung: Sie kommen im nördlichen Eurasien(Abb. 44), nicht aber im hohen Norden vor. Siebesiedeln Gebiete mit unterschiedlichem Klima,nämlich vom feuchten See- bis zum trockenen Konti-nentalklima (Heptner et al. 1966, 262).
Körpermaße: Kopf-Rumpf-Länge 95 bis 135 cm;Schulterhöhe 60 bis 80 cm; Gewicht 15 bis 35 kg. InSibirien bis 50 kg.
Verhalten: Rehböcke leben oft allein, sonstpaarweise oder in kleinen Gruppen. In Europa sindRehe Standwild, in Sibirien dagegen führen sie jahres-zeitliche Wanderungen von über 300 km durch. BeiWanderungen überwinden sie kaum mehr als 800Höhenmeter (Heptner et al. 1966, 268).
Paarungszeit: Juli–August, aber auch November–Dezember. Tragzeit: 7 bis 11 Monate (die Einnistungdes Keims wird bei der Sommerpaarung um bis zuvier Monate verzögert). Setzzeit: Mai bis Juni.
Natürliche Lebenserwartung: bis 15 Jahre.Nahrung: Blätter, Triebe, Kräuter, Gras, Beeren,
Eicheln und Pilze.Geweih: Sie besitzen bis zu sechs Enden (jede
Seite 3). Die Geweihe erreichen eine Länge von 20–25 cm. Sie werden von November bis Dezemberabgeworfen und wachsen während des Winters nach.Ihr Wachstum ist vom Nahrungszugang abhängig.
Klimatische Aussage: Das Vorkommen vonCapreolus capreolus ist ein klarer Hinweis für diePräsenz von dichtem Wald. Durch seine Anpassung anTemperaturen von bis zu -40°C Durchschnittstempera-turen im Januar und von 10°C bis 30°C Durch-schnittstemperaturen im Juli (Hokr 1951) ist es jedochsowohl in wärmeren wie auch in extrem kaltenRegionen verbreitet.
Darstellungen aus dem Jungpaläolithikum
Die sichere Zuordnung zu dieser Art ist schwierig, dasie kaum Merkmale aufweist, die eine zuverlässigeZuordnung erlauben. In der Wandkunst ist bis heutekeine Darstellung dieses Tieres bekannt. In der Klein-kunst sind hier eine Darstellung von Lortet (Abb. 45,links) auf Knochen (Chollot 1964, 150–152) und eineGravierung von La Marche (Abb. 45, rechts) auf
einem kleinen Plättchen (Pales und de Saint Pereuse1989) zu erwähnen. Aber auch hier ist die Interpre-tation als Reh nicht sicher, da Hirsche nicht immer eingroßes Geweih tragen. Wenn das Geweih bei Rothir-schen nachwächst, erreicht es immer eine Phase, inder es ebenfalls aus nur wenigen Sprossen besteht.Männliche Rothirsche im Alter von zwei bis dreiJahren besitzen ebenfalls nur kurze Geweihe mitwenigen Enden, wie sie bei der möglichen Gravierungeines Rehes von Lortet zu sehen sind.
Reste aus dem Jungpaläolithikum
Die Reste von Capreolus capreolus sind immer sehrselten. Das Reh ist nie die dominierende Tierart, undauch in den Fundstellen, in denen es häufigervorkommt, erreicht es einen Anteil an der Fauna vongerade 10 bis 20%. Unter den Fundstellen, in denendas Reh etwas besser vertreten ist, sind hier dieGravettienschichten von La Ferrassie, RiparoTagliente, Grotta Polesini, Grotta della Cala, Grottadella Calanca und Grotta dell’Erica (siehe Fundstel-lenkatalog) zu nennen (Abb. 46).
Abb. 44 Heutige Verbreitung von Capreolus capre-olus in Europa (nach Mitchell-Jones et al. 1999,401).

98 Verbreitung der großen Jagdfauna in Mittel und Westeuropa im oberen Jungpleistozän
In Südeuropa kommt das Reh während des gesamtenJungpaläolithikums vor. In Italien scheint es etwaszahlreicher gewesen zu sein. In Südfrankreich ist dasReh dagegen, mit der erwähnten Ausnahme von LaFerrassie, nur punktuell und fast immer durch nurwenige Knochen vertreten (meistens weniger als 10Knochen und Prozentzahlen zwischen 0,01 und 2%).In Kantabrien ist es wiederum in zahlreichenSchichten zu finden (Straus 1992), obwohl ebenfallsin geringen Zahlen, so wie in Portugal. Grundsätzlichwird es häufiger am Ende des Magdaléniens und inden Schichten des Aziliens. In Mitteleuropa ist es imMagdalénien der Teufelsbrücke, im Magdalénien desHohle Fels (Münzel 2004) und, wie schon dasWildschwein, in Fundstellen der Federmessergruppen(Andernach) und der Stielspitzen-Gruppen (HohlerStein bei Kallenhardt) erwähnt. Es kommt fast immermit Cervus elaphus, häufig auch mit demWildschwein vor, dagegen nimmt es ab, sobald Tiereder offenen Landschaft wie Pferd oder Steinbockbedeutender werden.
Diskussion
Die Tatsache, dass Rehe in den meisten Fundstellennur durch einzelne oder wenige Funde vertreten sind,zeigt wie beim Wildschwein, dass diese Tiere nurselten zur Nahrung der Menschen im Paläolithikumgehörten. Diese scheuen Tiere sind im Wald wegen
ihres tarnenden Felles und ihrer geringen Größe ohneJagdhunde kaum auszumachen. Die gleiche Argumen-tation gilt für die Seltenheit der Darstellungen diesesTieres. In Italien sind Fundstellen mit Resten vonRehen etwas häufiger als in anderen Regionen. Diesscheint auf eine stärkere Präsenz dieser Art in dieserRegion hinzuweisen und passt gut zu dem für dieEiszeit vermuteten milden Klima.
Die punktuelle Präsenz von Capreolus capreolusmit Spezies der offenen Mammuthus-Coelodonta-Steppenfauna in Frankreich und Mitteleuropa kannverschiedenen Gründe haben, die von Fall zu Fallüberprüft werden sollten. Es kann sich natürlich umheterogene Schichten handeln, so wie es wahrschein-lich in der Grotte de l’Œil (Sacchi 1986, 177) der Fallist oder auch für manche Fundstellen der Stielspitzen-Gruppen (Baales 1996, 220–221) für möglich gehaltenwird. Andererseits, so wie es auch für dasWildschwein erwähnt wurde (siehe das Kapitel überdas Wildschwein, S. 83–88), darf man nichtvergessen, dass Rehe in Sibirien sehr weite jahreszeit-liche Wanderungen unternehmen (Heptner et al. 1966,268). Dadurch scheint es möglich, dass Rehe imSommer nach Norden wanderten, in die Regionen, indenen die übliche Jagdfauna aus Tieren derMammuthus-Coelodonta-Steppenfauna bestand. Dieweitverbreitete Präsenz von Galeriewäldern entlangder Flüsse könnte ihnen dort eine passende ökolo-gische Nische geboten haben.
Abb. 45 Mögliche Darstellungen von Rehen. Links, Lortet (Capitan et al. 1910, 186); rechts, La Marche (Cré-madès 1997, 68).

Reh (Capreolus capreolus) 99
Abb. 46 Im Text erwähnte Fundstellen mit Resten aus Deutschland oder mit zahlreichen Resten aus Frankreichund Italien (Dreiecke) und Darstellungen (Kreise) von Rehen im Jungpaläolithikum. Die graue Fläche zeigt diemögliche Hauptverbreitung des Rehes im Jungpaläolithikum. Es wurden keine der zahlreichen Fundstellen Spa-niens und Portugals kartiert, da hier die Rehe immer nur durch wenige Knochen vertreten sind. Die Pfeile ver-weisen auf mögliche Wanderungsrouten außerhalb ihres annehmbaren, üblichen Verbreitungsgebietes. 1 Lortet; 2 La Marche; 3 Hohler Stein bei Kallenhardt; 4 Andernach; 5 Teufelsbrücke; 6 Hohle Fels; 7 La Fer-rassie; 8 Riparo Tagliente; 9 Grotta Polesini; 10 Grotta della Cala; 11 Grotta della Calanca; 12 Grotta diCastelcivita.


Elch (Alces alces)
E. – Elk (in Großbritannien) und Moose (in Nordamerika); F. – Elan (in Eurasien) und Orignal (in Nord-amerika); S. – Alce europeo; Kat. – Ant; P. – Alce; I. – Alce.Ordnung: Artiodactyla Owen 1848; Familie: Cervidae Goldfuss 1820; Gattung: Alces Gray 1821; Art: Alcesalces (Linnaeus 1758).Vorwiegend verwendete Literatur: Gaussen und Merlaud 1987; Nygrén 1986.
Habitat: Wald, Steppe, Taiga.Verbreitung: Elche leben im nördlichen Eurasien
und Nordamerika (Abb. 47). Obwohl sie heute nichtmehr im alpinen Raum vorkommen, sind sie durcheinen Fund aus dem Elchlöchli (1630 ± 50 BP bzw.328–554 n.Chr.) weit in der römischen Kaiserzeit inder Schweiz belegt (Blant et al. 2002).
Körpermaße: Kopf-Rumpf-Länge 200 bis 290 cm,manchmal bis 310 cm; Schulterhöhe 150 bis 225 cm;Gewicht 270 bis 600 kg. Bullen sind größer undschwerer als Weibchen.
Verhalten: Sie sind meistens Einzelgänger undkönnen im Winter kleine bis größere Gruppen bilden.In der Brunftzeit sind sie auch paarweise anzutreffen.
Paarungszeit: September und Oktober. Tragzeit: 8Monate. Setzzeit: Mai und Juni, ein bis drei Jungtiere,meistens jedoch 2.
Natürliche Lebenserwartung: bis 25 Jahre.Nahrung: Triebe, Blätter, Laubholzrinde, Sumpf-
und Wasserpflanzen. Klimatische Aussage: Elche können in Regionen
leben, in denen die Wintertemperatur bis -70°C reicht.Im Juli ertragen sie kaum Mittelwerttemperaturen vonüber 20°C. Sie können in Regionen mit erheblichenNiederschlägen leben. Die langen Beine mit dembreiten Fuß erlauben es ihnen, sich bestens in schnee-reichen sowie sumpfigen Gebieten zu bewegen.
Darstellungen aus dem Jungpaläolithikum
Bilder von Elchen sind außerordentlich selten. Alssichere Darstellung gelten nur zwei Gravierungen(Abb. 48, 4 und 5) von Gargas (Crémadès 1993, 147).Eine Gravierung auf einem bearbeiteten Bruchstückeines Rentiergeweihes (Chollot 1964, 104–105) ausGourdan (Abb. 48, 1–2) scheint ebenfalls sehrwahrscheinlich. Zu diesen kommt eine sehr schöneGravierung auf einer Schieferplatte (Abb. 48, 3), die,obwohl deren Herkunft nicht bestimmt werden konnte,wahrscheinlich aus einer Magdalénien-Fundstelle desPérigords stammt (Gaussen und Merlaud 1987). Beidiesen Darstellungen ist es entweder das charakte-ristische Geweih (Gourdan) oder die besonderen
Proportionen des Körpers (Gargas), die zu einerBestimmung als Elch führen. Die detailreicheGravierung aus einer nicht näher bestimmbarenFundstelle des Périgord ist ohne diese Charakteristikanur als wahrscheinliche Darstellung eines weiblichenElchs anzusprechen (Gaussen und Merlaud 1987,169). Etwas Ähnliches gilt auch für eine sehr schöneGravierung aus Gönnersdorf (Abb. 73, 3), die trotz derQualität und der Details nicht mit Sicherheit dem Elchoder der Saiga-Antilope zugeordnet werden kann(Bosinski 1981, 100; Koenigswald und Hahn 1981,89; Dubourg 1997, Abb. 37)49.
Reste aus dem Jungpaläolithikum
Elche sind unter der jungpaläolithischen Fauna außer-ordentlich selten und spielen nie eine relevante Rolle(Abb. 49). In Südwestfrankreich sind sie erst ab demMagdalénien nachgewiesen (Delpech 1983, 177–178).
Abb. 47 Heutige Verbreitung von Alces alces (nachHeptner et al. 1966, 281).
49 Die Saiga-Antilope ist mit Sicherheit, der Elch wahrscheinlich in der Fauna von Gönnersdorf vertreten (Street1994, 548).

102 Verbreitung der großen Jagdfauna in Mittel und Westeuropa im oberen Jungpleistozän
Unter den wenigen Fundstellen kann man Gourdan(Bahn 1983, Table 14), wo der Elch schon imSolutréen anwesend zu sein scheint und die Fauna vonRentier und Rothirsch dominiert ist, und Rochereil(Astre 1950) erwähnen.
Die nacheiszeitlichen Fundstellen sind etwaszahlreicher (Delpech 1983, 177). Dem oberenMagdalénien gehören auch die Funde von LesDouattes, Veyrier und Pierre-Châtel (Desbrosse undPrat 1974). In der Schicht IIa von Pierre-Châtel sindvon 45 Resten 22 als Elch bestimmt worden(Desbrosse und Prat 1974, 153).
Auch in Norditalien scheint der Elch erst ab demEpigravettiano nachgewiesen zu sein. In RiparoSoman erreicht er bei der Berechnung der Mindestin-dividuenzahl (MIZ) der Schicht der Dryas III 8% derFauna (was jedoch lediglich 2% der Knochenrestedarstellt), in der Schicht 11–12 von Riparo Tagliente,am Übergang zwischen Dryas I und Bölling, beträgtsein Anteil in der Fauna 29,7% (30 von 101 Faunen-resten), in der Schicht 13–15, die in die Dryas I datiertwird, beträgt sein Anteil 11,2%. Seine Präsenz inNorditalien ist ebenfalls in der Höhle Paina und inArene Candide nachgewiesen. Hier ist zu vermerken,dass der Elch zusammen mit dem Riesenhirschvorkommt. Die Trennung zwischen diesen beidenArten ist jedoch schwierig, so dass mehrere Reste hier
in die Kategorie Alces/Megaloceros fallen. Das Bildüber die Verbreitung des Elches in Norditalien wirddurch seine wiederholte Präsenz in Slowenien ergänzt.In Deutschland kommt der Elch an der Schussen-quelle, im Hohlenstein-Stadel und im Hohlen Stein beiKallenhardt vor, wo immer das Rentier vorherrscht,sowie in der Federmesserschicht von Andernach, woder Rothirsch durch mehrere Individuen ebenfallsvertreten ist. Der Elch scheint in Kantabrien (Altuna1972, 347) sowie im Rest Spaniens zu fehlen.
Diskussion
In der untersuchten Region wird nur in Arene Candidevon der gleichzeitigen Präsenz von Elch und Riesen-hirsch berichtet. In Slowenien kommen beideTierarten in zwei weiteren Fundstellen vor: Županovspodmol und Lukenjska jama. Dafür lassen sich zweiErklärungen finden. Zum einen hat es mit denUmweltansprüchen dieser beiden Tierarten zu tun, diezwar beide ein großes Geweih tragen und sich in derGröße nahe stehen, aber sonst unterschiedlich gebautsind. Der Elch mit seinem kurzen Hals und seinenlangen Beinen zeugt von einer Anpassung anfeuchtere Regionen unabhängig von den Temperatur-verhältnissen. Vielleicht konnte er deswegen erst amEnde der Eiszeit nach Südwestfrankreich vordringen.
Abb. 48 Mögliche Darstellungen von Elchen. 1–2 Gourdan (Capitan et al. 1910, Abb. 177); 3 unbekannteFundstelle aus dem Périgord (Gaussen und Merlaud 1987); 4–5 Gargas (Barrière 1976, 222 und 235).
2 3 1 4 5

Elch (Alces alces) 103
Der Elch konnte außerdem auch in etwas dichterenWäldern als der Riesenhirsch vorkommen. DerRiesenhirsch mit seinen im Verhältnis kürzerenBeinen brauchte ein trockeneres Klima. Der größereTeil seiner Reste aus Südwestfrankreich stammt ausdem älteren oder mittleren Jungpaläolithikum.Weiterhin darf man nicht vergessen, dass diese beidenTierarten, deren Verbreitungsgebiete sehr ähnlicherscheinen (Abb. 49 und 55), immer nur durch ganz
wenige wenn nicht sogar einzelne Knochen vertretensind, daher ist die Wahrscheinlichkeit, sie beide inderselben Schicht zu finden (und auseinander haltenzu können), gering.
Die Verbreitung des Elchs um die Alpen passtübrigens sehr gut mit der erhöhten Feuchtigkeit dieserRegion auch während der letzten Eiszeit zusammen(siehe S. 35–36).
Abb. 49 Im Text erwähnte Fundstellen mit publizierten Resten (Dreiecke) oder Darstellungen (Kreise) vonElchen und die mögliche, maximale Verbreitung von Elchen im Jungpaläolithikum. Die Pfeile verweisen aufmögliche Wanderungsrouten außerhalb ihres annehmbaren, üblichen Verbreitungsgebietes. 1 Eine nicht näher zu definierende Fundstelle im Périgord (siehe Abb. 48, 3); 2 Gargas; 3 Gourdan (Kunst)sowie Gourdan und Rochereil (Fauna); 4 Gönnersdorf; 5 Riparo Soman; 6 Riparo Tagliente; 7 Grotta Paina; 8Arene Candide; 9 Les Douattes und Veyrier; 10 Pierre-Châtel; 11 Rochereil; 12 Hohler Stein bei Kallenhardt;13 Andernach; 14 Hohlenstein-Stadel; 15 Hohle Fels und Brillenhöhle; 16 Schussenquelle.


Rentier (Rangifer tarandus)
E. – Reindeer (in Großbritannien) und Cariboo (in Nordamerika); F. – Renne; S. – Reno; Kat. – Ren; P. – Rena;I. – Renna.Ordnung: Artiodactyla Owen 1848; Familie: Cervidae Goldfuss 1820; Gattung: Rangifer Frisch 1775; Art:Rangifer tarandus (Linnaeus 1758).Vorwiegend verwendete Literatur: Altuna 1996a; Corchón 1992; Delpech 1983, 144–174; Herre 1986b;Weinstock 2000.
Habitat: Tundra, Taiga, Wald/Tundra, hohe Gebirge. Verbreitung: Rentiere leben in den nördlichsten
Regionen Eurasiens und Amerikas (Abb. 50).Verhalten: Die Rentiere in der Tundra leben in
großen, die Rentiere der Taiga in kleinen Herden. Siesind Wandertiere und überwiegend tagaktiv.
Körpermaße: Kopf-Rumpf-Länge 170 bis 220 cm;Schulterhöhe bis 140 cm; Gewicht 120 bis 220 kg.Renhirsche sind größer und schwerer als Weibchen.
Sowohl Hirsche als auch Weibchen tragenGeweihe, wobei das Geweih der Hirsche größer ist.
Paarungszeit: Ende September und Oktober.Tragzeit: 7 bis 8 Monate. Setzzeit: Mai bis Juni.
Natürliche Lebenserwartung: 15 Jahre.Nahrung: Flechten, Kräuter, Gräser, Zweige,
Triebe, Blätter, Pilze, an der Küste auch Algen.Klimatische Aussage: Rentiere leben sowohl in der
Taiga, im Wald als auch in der baumlosen Tundra. Siekönnen im Winter extrem niedrige Temperaturenertragen, in den Regionen, in denen sie heute leben,erreichen die Sommer kaum den Mittelwert von 17°C.
Darstellungen aus dem Jungpaläolithikum
Bildliche Darstellungen von Rentieren sind, wennauch nicht sehr häufig, in Frankreich und Deutschlandgewöhnlich. Man denke hier an die Gravierung aufeinem Lochstab vom Kesslerloch und auf einemLochstab im Petersfels (Müller-Beck und Albrecht1987, 88 und 104–105), an die Malereien und Gravie-rungen in der Chauvet-Höhle (Clottes 2003) und andie Gravierungen in Les Trois Frères (Bégouën undBreuil 1958). Daher werden uns diese hier nicht weiterbeschäftigen.
In Spanien sind Darstellungen von Rentierendagegen selten. Sie befinden sich außer in Kantabrienauch in der spanischen Meseta, nämlich in der Cuevadel Reno (Abb. 51, 13), La Hoz (Abb. 51, 15) undSiega Verde50 (Abb. 51, 16). Die deutlichsten Darstel-lungen stammen aus Las Caldas (Abb. 51, 3–4) undTito Bustillo (Abb. 51, 11–12). Einige andere
Darstellungen, die von María Soledad CorchónRodríguez abgebildet und als mögliche Darstellungenangesprochen werden (1992, 60–62), erscheinen sehrfraglich, daher werden sie hier nicht abgebildet.
Reste aus dem Jungpaläolithikum
Das Rentier ist während des gesamten Jungpaläoli-thikums die Hauptbeute in vielen Regionen in Mittel-und Westeuropa. Fundstellen mit über 80% Rangifertarandus sind häufig. Darunter seien hier Petersfels,Kesslerloch, Munzingen, Schussenquelle (Deutsch-land), Roque de Combe, Le Flageolet, Laugerie-Hauteund La Madeleine (Frankreich) erwähnt. In allendiesen Fundstellen ist das Pferd die zweithäufigsteBeute, wodurch eine offene Landschaft belegt ist. InSpanien (Abb. 52), wo das Rentier immer selten ist,
50 In der Fundstelle von Los Casares wird zwar eine mögliche Rentierdarstellung erwähnt (Acosta und Molinero 2003, 41), da diese aber nicht abgebildet ist, kann man dies nicht überprüfen.
Abb. 50 Heutige Verbreitung von Rangifer tarandus(nach Heptner et al. 1966, 385).
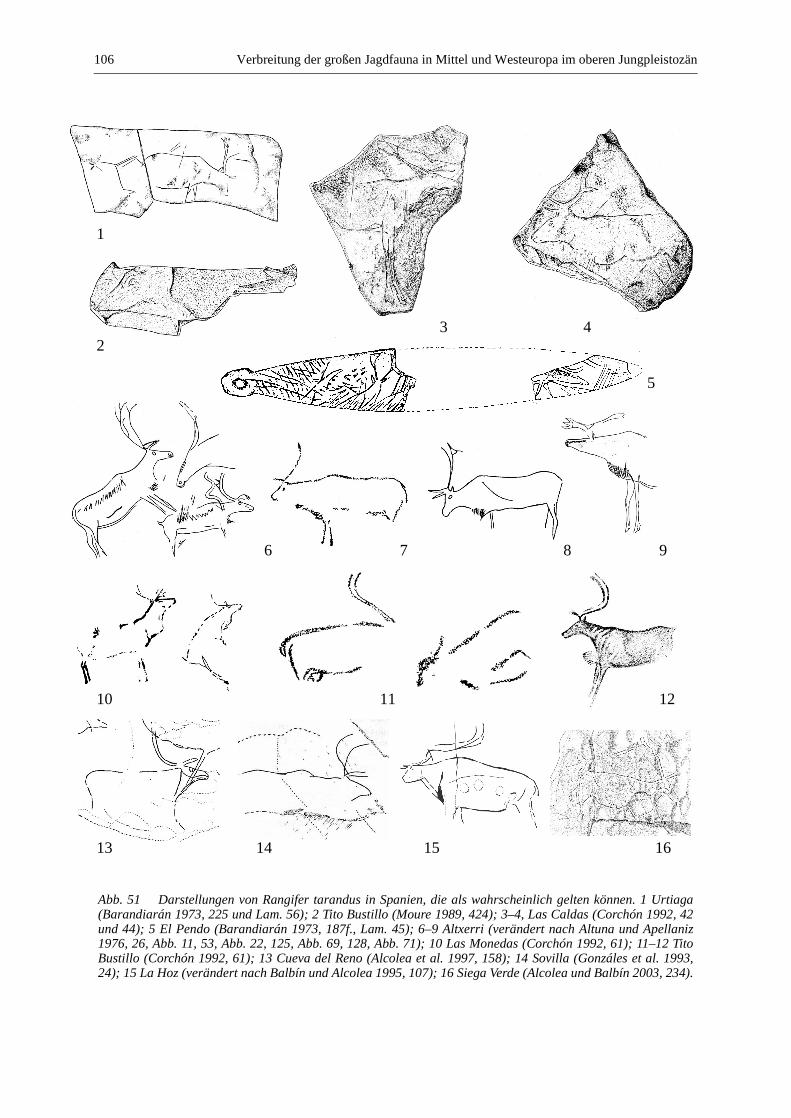
106 Verbreitung der großen Jagdfauna in Mittel und Westeuropa im oberen Jungpleistozän
Abb. 51 Darstellungen von Rangifer tarandus in Spanien, die als wahrscheinlich gelten können. 1 Urtiaga(Barandiarán 1973, 225 und Lam. 56); 2 Tito Bustillo (Moure 1989, 424); 3–4, Las Caldas (Corchón 1992, 42und 44); 5 El Pendo (Barandiarán 1973, 187f., Lam. 45); 6–9 Altxerri (verändert nach Altuna und Apellaniz1976, 26, Abb. 11, 53, Abb. 22, 125, Abb. 69, 128, Abb. 71); 10 Las Monedas (Corchón 1992, 61); 11–12 TitoBustillo (Corchón 1992, 61); 13 Cueva del Reno (Alcolea et al. 1997, 158); 14 Sovilla (Gonzáles et al. 1993,24); 15 La Hoz (verändert nach Balbín und Alcolea 1995, 107); 16 Siega Verde (Alcolea und Balbín 2003, 234).
1 3 4 2 5 6 7 8 9 10 11 12 13 14 15 16

Rentier (Rangifer tarandus) 107
kommt es in den Schichten aller jungpaläolithischer„Technokomplexe“ häufig mit Cervus elaphus undteilweise sogar mit Sus scrofa vor, so im Magdalénienvon Bora Gran d’en Carreres, im Solutréen vonReclau Viver, in der Schicht des oberen Aurignacienvon Lezetxiki, im Solutréen von Ermittia, im Azilienund Magdalénien von Aitzbitarte, in der Magdalénien-Schicht von Urtiaga, im Châtelperronien und Aurigna-cien von Labeko-Koba, im Magdalénien von La Riera,in der Schicht des oberen Magdalénien sowie in dervermischten Magdalénien-Solutréen-Schicht vonSantimamiñe, in der Gravettien- und der Solutréen-Schicht der Höhle von Amalda, im Aurignacien vonEl Castillo, im Solutréen von Altamira, im Solutréenund Magdalénien von Ermittia, im Solutréen vonCueto de la Mina und im Magdalénien von Morin. Andieser etwas ausführlicheren Auflistung sieht man,dass es eine kontinuierliche Präsenz von Rentieren inKantabrien und im Baskenland während des gesamtenJungpaläolithikums gab.
In Slowenien ist das Rentier gut vertreten, darunterin den Fundstellen von Županov spodmol, Zakajenispodmol, Pološiška cerkev, Babja jama undLukennjska jama (Pohar 1997). In Italien dagegen istes fast völlig abwesend. Im nordöstlichen Teil sindReste von Rangifer tarandus nur aus zwei Fundstellenbekannt: Ca’ Negra und Pocala. Ca’ Negra ist ein„Pozzo ossifero“, also eine Knochenansammlung.Nahe der Lokalität Pocala wurde ein Teil einesRentiergeweihes mit fluviatilen Spuren aus einerSchicht von „Terra rossa“ entdeckt, diewahrscheinlich zur Würm-Eiszeit gehört. Aus dergleichen Schicht stammen Steinartefakte, die demMoustérien zugeordnet wurden (Bartolomei et al.1982, 7). Aus dem nordwestlichen Teil Italiensstammen verschiedene Reste von Rentieren aus denGrimaldi-Höhlen, darunter Reste aus den jungpaläoli-thischen Feuerstellen A und B der Grotta del Principe(Blanc 1938, 17; Boule 1910, 213), ein (Mussi 1992,540) oder zwei (Boule 1910, 213) Rentiergeweihe aus
der Grotta dei Fanciulli und ein Unterkiefer aus derBarma Grande (Boule 1910, 212). Aus der weiteröstlich gelegenen Fundstelle von Arene Candide, diedurchaus zahlreiche faunistische Reste aufweist undsorgfältiger als die Grimaldi-Höhlen ausgegrabenwurde, ist kein einziger Rest eines Rentieres bekannt.Es scheint also, dass die Verbreitung von Rangifertarandus an den Grenzen Italiens, am Karst imöstlichen Teil des Friuli-Venezia-Giulia und imwestlichen Teil Liguriens, aufhörte. In der paläontolo-gischen Fundstelle von Settepolesini di Bodeno in derPo-Ebene sind, bis jetzt, keine jungpaläolithischenReste von Rentieren entdeckt worden (Gallini undSala 2000 und 2001).
Diskussion
Die Koexistenz von Rentier und Rothirsch und nochmehr die Koexistenz von Rentier und Wildschwein inSpanien lassen vermuten, dass es hier eine Art vonTaiga-Rentier gab. Natürlich ist hier zu berücksich-tigen, so wie Jesús Altuna und Koro Mariezkurrenazurecht erwähnen, dass manche Schichten aus diesenFundstellen sehr mächtig sind. Die Schicht IX vonLabeko-Koba ist z. B. einen Meter dick und dieOrographie ist um diese Fundstelle „sehr komplex undlabyrinthartig“ (Altuna und Mariezkurrena 2000, 112),so dass es sicher verschiedene Lebensräume neben-einander gegeben hat. Andererseits aber zeigt dieEntdeckung von Rentierresten aus dem Präboreal unddem Boreal Deutschlands, nämlich einige Knochen inRottenburg-Siebenlinden I (Hahn et al. 1993, 46–47)und zahlreiche Reste in Bettenroder Berg IX undSphinx II (Staesche 1994), dass um ca. 9.000 BPneben den Rentieren der Tundra schon Waldformenexistieren. Diese ebenfalls an der südlichsten Grenzeder Verbreitung der Rentiere und am Übergang zumVerbreitungsareal vom Rothirsch zu vermuten, scheintplausibel (dazu Delpech 1983, 144–174).

108 Verbreitung der großen Jagdfauna in Mittel und Westeuropa im oberen Jungpleistozän
Abb. 52 Fundstellen in Südeuropa (hier Spanien und Italien) mit publizierten Resten (JungpaläolithischeFundstellen = Dreiecke; Knochenansammlung und ältere Fundstellen = Kreuze) oder Darstellungen (Kreise)vom Rentier und dessen mögliche, maximale südliche Verbreitung im Jungpaläolithikum. Hier sind auch Dar-stellungen angegeben, die mir fraglich erscheinen. 1 A Valiña; 2 La Paloma; 3 Las Caldas; 4 Puebla de Lillo; 5 La Viña; 6 La Riera und Cueto de la Mina; 7 TitoBustillo; 8 El Castillo und Altamira; 9/10 Las Monedas und El Pendo; 11 La Haza und Covalanas; 12 Atxurri,Santimamiñe, Lumentxa, Armotxe und Armiña; 13 Ekain, Astigarraga, Eralla, Urtiaga, Urtiagako Leizea,Ermittia und Amalda; 14 Aitzbitarte III und IV, Torre; 15 Morín und Rascaño; 16 Axlor; 17 Lezetxiki undLabeko Koba; 18 Abauntz; 19 Zatoya; 20 Siega Verde; 21 Cueva del Reno; 22 La Hoz; 23 Bora Gran d’en Car-reras und Arbreda; 24 Grotta del Principe; 25 Ca’ Negra; 26 Grotta Pocala.

Riesenhirsch (Megaloceros giganteus)
E. – Giant deer, irish elk; F. – Mégacéros; S. – Megaceros, alcé irlandés oder ciervo gigante; Kat. – Megaceros,cérvol gegant; P. – Alce cigante o alce irlandês; I. – Cervo megacero oder nur Megacero.Ordnung: Artiodactyla Owen 1848; Familie: Cervidae Goldfuss 1820; Gattung: Megaloceros Brookes 1828; Art:Megaloceros giganteus (Blumenbach 1799). Vorwiegend verwendete Literatur: Gould 1974; Koenigswald 2002, 71–75.
Da der Riesenhirsch am Ende der Eiszeit bzw. zuBeginn des Holozäns ausgestorben ist51, kann mandessen Umweltansprüche nur mit Hilfe von Rück-schlüssen untersuchen. Als wichtigster Hinweis giltsein Geweih, das bis 3,5 (Joger und Koch 1994, 64)oder sogar 5 m breit sein konnte (Yalden 1999, 40).Die Größe des Geweihs lag in direktem Verhältnis zurKörpergröße, so dass ein Riesenhirsch mit einem 5 mbreiten Geweih bis zu 2 m Schulterhöhe und einGewicht von bis zu 600 kg erreichen konnte. Miteinem solchen Geweih ist es klar, dass der Riesen-hirsch in freiem Gelände leben musste, da es für ihnunmöglich gewesen wäre, sich im Wald zu bewegen.Das Geweih hatte weiterhin ein beträchtlichesGewicht (bis über 45 kg) und musste jedes Jahrerneuert werden. Damit war der Riesenhirsch auf einean Mineralien und Kalk reichhaltige Nahrungangewiesen. Das Vorrücken des Waldes und dieweiteren Veränderungen der Umwelt am Ende derEiszeit führten unweigerlich zu seinem Aussterben.Die jüngsten Funde aus Dänemark und Irland sind um11.000 BP (Aaris-Sørensen 1992; Yalden 1999)datiert. Der bis jetzt jüngste Fund stammt jedoch vonder Insel Man und wurde um 9.200 BP datiert(González et al. 2000).
Klimatische Aussage: Der Riesenhirsch ist inerster Linie ein Indikator für eine weitgehend offeneLandschaft, eventuell für sehr lichten Wald. Er konntesehr niedrige Temperaturen im Winter ertragen,bevorzugte aber ein mildes oder leicht kühles Klima(Guérin und Patou-Mathis 1996, 62).
Darstellungen aus dem Jungpaläolithikum
Die Kennzeichen des Riesenhirsches sind der Rücken-höcker bei beiden Geschlechtern und das riesigeGeweih bei den männlichen Tieren. Dadurch lassensich einige Darstellungen dieser Art zuordnen.
Aus Spanien stammen nur unsichere Darstel-lungen von Riesenhirschen. Die erste wurde in derCueva del Reno entdeckt (Abb. 53, 1). Sie wurde mitder Pick-Technik erstellt. Das Foto derselbenDarstellung (Alcolea und Balbín 2003, 235) ist so„trüb“, dass alle Zweifel bestehen bleiben müssen.
Eine Gravierung von Siega Verde in Spanien, dieebenfalls einen Riesenhirsch darstellen soll (Balbínund Alcolea 1995, 122–123), zeigt zwar einen Cer-viden mit stark ausgeprägtem Geweih, was an einenRiesenhirsch denken lassen könnte, aber die sehr gutdargestellte Rückenpartie zeigt keinen Rückenhöcker.Dadurch muss man auch diese Darstellung als Riesen-hirsch zumindest als unsicher betrachten52.
Eine dritte Darstellung, diesmal eine Malerei,wurde in der Höhle von La Lloseta entdeckt (Abb. 53,3). Ihre Bestimmung als Riesenhirsch scheint eben-falls sehr fraglich.
Als sicher gelten dagegen einige Darstellungen ausFrankreich. Es sind sieben Individuen in der Höhlevon Chauvet (Clottes 2003, 192), von denen aber nurzwei publiziert wurden (Abb. 54, 1–2), eine Malereiund eine Gravierung in der Höhle von Cosquer (Abb.54, 3–4), eine Gravierung aus der Höhle von Gargas(Abb. 54, 5), die Malerei von drei Individuen ohneGeweih sowie die Finger-Gravierung eines männ-lichen Individuums in der Höhle von Pech-Merle53
(Abb. 54, 6–7), drei gemalte Individuen aus der Höhlevon Cougnac (Abb. 54, 8), eine Gravierung aus derHöhle von La Grèze (Abb. 54, 9), eine Gravierung aus
51 Die Annahme, dass der Riesenhirsch in Irland erst im Mittelalter ausgestorben wäre (Mitchell und Parkes1949), wird heute gar nicht mehr diskutiert und damit als falsch betrachtet (Yalden 1999).52 Es sei hier ergänzend erwähnt, dass Rodrigo de Balbín Behrmann und José Javier Alcolea González in einemArtikel von 2003 eine wesentlich andere Umzeichnung dieses Tieres präsentieren (Alcolea und Balbín 2003,232). Hier sind die Schaufeln im Vergleich mit der anderen Umzeichnung (Abb. 53) ergänzt und hervorgehoben;auf der Rückenlinie ist ein Rückenhöcker zu erkennen. 53 Die drei gemalten Tiere werden von Jan Jelinek als „phantasievoll gestaltete Huftiere“ angesprochen (Jelinek1972, 371), André Leroi-Gourhan meint, dass es unmöglich sei, diese Figuren einer bestimmten Tierart zuzuord-nen (Leroi-Gourhan 1965, 263).

110 Verbreitung der großen Jagdfauna in Mittel und Westeuropa im oberen Jungpleistozän
der Höhle von Pair-non-Pair (Abb. 54, 10) undzahlreiche Gravierungen aus der Höhle vonRoucadour (Abb. 54, 11–15). Über diese letzte Höhle,in der es über 80 Darstellungen geben soll, schreibtMichel Lorblanchet, dass 30% der ZeichnungenCerviden, die meisten davon weibliche Riesenhirschesein sollen (Lorblanchet 1985, 512). Leider ist dieseHöhle kaum publiziert. Als wahrscheinlich gilt nocheine Gravur aus Gargas, die ein Individuum ohneGeweih darstellen soll (Abb. 54, 5).
Zu den Fundstellen mit Darstellungen von Riesen-hirschen wird häufig auch die Höhle von Roc-de-Sersgerechnet, weil hier auf einem Block die Gravierungeines mächtigen Geweihs dargestellt ist. Ob dieseallein eine Zuordnung zum Riesenhirsch, wie AndréLeroi-Gourhan es vorschlägt (1965, 419), erlaubt, istfraglich. Jean Airvaux und Henri Delporte meinen, eshandle sich um die Darstellung eines Rentiergeweihs(Airvaux 2001, 67; Delporte 1984a, 580–581).
Wie man sieht, ist der Riesenhirsch in der paläoli-thischen Kunst nur in wenigen Fundstellen abgebildetworden, die sich alle in Frankreich befinden.
Es fällt weiterhin auf, dass gegenüber den ca. 30Wandbildern aus Frankreich54 keine einzigeDarstellung von Riesenhirschen auf einer tragbarenUnterlage bekannt ist. Ob dahinter mehr als nur einZufall zu sehen ist, kann hier nicht geklärt werden.
Reste aus dem Jungpaläolithikum
Der Riesenhirsch ist durch zahlreiche Reste insbe-sondere aus den Mooren Irlands bekannt, wo ganzeSkelette geborgen werden konnten. Die Bestimmungvon einzelnen postkranialen Resten von Riesenhir-schen ist jedoch zumindest schwierig, da sie haupt-sächlich auf metrischen Merkmalen beruht. Dadurchentstehen oft Überschneidungen mit anderen großenCerviden (Koenigswald et al. 1974, 96–97). In Europa
kommt er während des ganzen Jungpaläolithikumsvor. In Frankreich und Kantabrien ist ein Schwer-punkt im älteren und mittleren Jungpaläolithikum zuerkennen. In Italien sind nur drei Fundstellen mitResten von Riesenhirschen publiziert; er kommt imUluzziano von Castelcivita in Süditalien sowie imEpigravettiano von Fumane und Arene Candide inNorditalien vor. Die Funde aus dem Ende desJungpaläolithikums Sloweniens (Županov spodmol,Poljšiška und Lukenjska jama) unterstreichen seinePräsenz in Südeuropa.
In Südwestdeutschland ist das Vorkommen desRiesenhirsches für das Mittelpaläolithikum bewiesen.Ein Fersenbein eines Cerviden aus der Aurigna-cienschicht AH II und ein Mittelfußknochen aus demaurignacienzeitlichen Horizont GH 17 der HöhleGeißenklösterle gehören aufgrund ihrer Größewahrscheinlich zum Riesenhirsch (Münzel 2004;Münzel und Conard 2004, 228).
Diskussion
Betrachtet man die Kartierung der Reste vom Megalo-ceros giganteus (Abb. 55), fällt auf, dass die meistenaus Regionen mit einer vielseitigen Geländemor-phologie stammen. Schaut man weiterhin die Faunen-listen dieser Fundstellen an, kann man einigeGemeinsamkeiten feststellen. Erstens ist der Riesen-hirsch fast immer durch nur ganz wenige odereinzelne Knochen vertreten und tritt hier und dort nurin wenigen oder einzelnen Schichten auf. EineAusnahme sind hier die Grotta delle Arene Candideund die Grotta di Fumane. In der letzteren ist derRiesenhirsch im Epigravettiano durchgehend mit biszu vier Individuen vertreten und erreicht bei dergesamten Aufzählung mit 13 Individuen 7,6% derMIZ der großen Jagdfauna aus den Schichten desEpigravettiano dieser Höhle. In anderen Fundstellen
Abb. 53 Mögliche Darstellungen von Riesenhirschen aus Spanien: Links, Cueva del Reno (nach: Alcolea undBalbín 2003, 235); Mitte, Siega Verde (Balbín und Alcolea 1995, 122); rechts, La Lloseta (nach Balbín et al.2003, 132).
54 Wie schon erwähnt, weiß man nicht, wie viele Riesenhirschdarstellungen es in der Höhle Roucadour gibt.

Riesenhirsch (Megaloceros giganteus) 111
Abb. 54 Als Riesenhirsch publizierte Darstellungen. 1–2 Chauvet (Züchner 1999); 3–4 Cosquer (Züchner1999; Lorblanchet 1995, 255); 5 Gargas (Barrière 1976); 6–7 Pech-Merle (Züchner 1999; Crémadès 1993,150); 8 Cougnac (Züchner 1999); 9 La Grèze (Crémadès 1993); 10 Pair-non-Pair (Cheynier und Breuil 1963,199); 11–15 Roucadour (verändert nach Lorblanchet 1984, 512; Lorblanchet 1995, 255).
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15

112 Verbreitung der großen Jagdfauna in Mittel und Westeuropa im oberen Jungpleistozän
ist sein Anteil oft nur 0,1%. Weiterhin fällt auf, dass infast allen Fundstellen, in denen Megaloceros gigan-teus bestimmt wurde, die Faunenvergesellschaftungaus auffällig zahlreichen Pflanzenfressern besteht,nämlich sieben bis elf Arten55. Diese große Vielsei-tigkeit der Begleitfauna der Reste von Megalocerosgiganteus zeigt, dass er in Gebieten vorkam, in denenzahlreiche verschiedene Nischen existierten. Dort, woder Riesenhirsch nachgewiesen ist, sind auch immerandere Hirscharten vorhanden. In La Ferrassie, LeFlageolet, Gargas, Lezetxiki, Labeko Koba undAmalda kommen sogar Rothirsch, Reh und Rentierzusammen vor. In Italien, wo es kaum Rentiere gab,tritt er immer mit Rothirsch und Reh, in AreneCandide auch mit dem Elch zusammen auf.
Obwohl die meisten Fundstellen, in denen derRiesenhirsch vorhanden ist, eine ausgewogeneFaunenliste aufweisen, ist der Riesenhirsch auch inSchichten mit einer mehr oder weniger „speziali-sierten“ Jagd nachgewiesen: in La Ferrassie (F1–F4fr), wo der Rothirsch 65,7% erreicht, in Le Flageolet Isowohl in der Schicht IX, wo das Rentier mit 68,9%vorkommt als auch in der Schicht VII, in der derRothirsch mit 69,2% dominiert, in Amalda (IV), wodie Gämse 40% der Individuenzahl und 62,9% der
Reste stellt, in Arene Candide (P3), wo der Steinbockmit 74,4% vertreten ist, und in Castelcivita (rsa UA),wo das Pferd 76,2% der Fauna bildet.
Es ist schließlich auffällig, dass die Verbreitung desRiesenhirsches sich am Übergangsbereich zwischender Verbreitung des Rentieres und des Rothirschesbefindet. In derselben Region ist ebenfalls der Elch zufinden (Abb. 49), der noch heute am Übergangzwischen der Verbreitung des Rentieres und desRothirsches lebt. Der Riesenhirsch scheint gegenüberdem Elch auch in bergigen und trockeneren Regionenwie Kantabrien vorzukommen. Die Apenninenscheinen durch die Funde von Castelcivita ebenfallskein Hindernis für die Verbreitung des Riesenhirschesgewesen zu sein. Die enge Überschneidung desVerbreitungsgebietes des Riesenhirsches und desElches sowie dessen Lage zwischen den Arealen vonRentier und Rothirsch wird auch durch ihre fastgleichzeitige Ankunft im Süden Skandinaviens um ca.11.500 BP, ca. 1.000 Jahre nach den Rentieren und ca.1.800 Jahre vor den Rothirschen deutlich (Aaris-Sørensen 1992, 145–146).
Möglicherweise hat die Einwanderung des Elchesam Ende der Eiszeit nach Westeuropa einen zusätz-lichen ökologischen Druck auf die Riesenhirschpopu-lation ausgeübt.
55 In den wenigen Fundstellen, in denen nur fünf Spezies der großen Jagdfauna vertreten sind, beruht diesesErgebnis auf der geringen Anzahl der bestimmten Reste oder der MIZ.

Riesenhirsch (Megaloceros giganteus) 113
Abb. 55 Fundstellen mit publizierten Resten (Dreiecke), deutliche (Kreise mit punkt) und fragliche (Kreise mitFragezeichen) Darstellungen von Riesenhirschen und die mögliche Verbreitung des Riesenhirsches im Jungpa-läolithikum. 1 Pair-non-Pair; 2 La Grèze; 3 Cougnac; 4 Roucadour; 5 Pech-Merle; 6 Chauvet; 7 Cosquer;8 Gargas; 9 Lloseta; 10 Siega Verde; 11 Cueva del Reno; 12 Hohle Fels; 13 Pierre-Châtel; 14 La Ferrassie undLe Flageolet; 15 Gatzarria; 16 Valorgues; 17 El Pendo; 18 Urtiaga und Amalda; 19 Lezetxiki und LabekoKoba; 20 Arene Candide; 21 Fumane; 22 Polesini di Bodeno; 23 Castelcivita.


Steppenbison (Bison priscus)
E. – Steppe Wisent; F. – Bison des steppes; S. – Bisonte; Kat. – Bisó; P. – Bisão europeu; I. – Bisonte.Ausgestorben: In Europa starb der Bison priscus am Ende der Eiszeit aus. In der Angara-Ebene könnte er jedochbis vor ca. 1000 Jahren noch gelebt haben (Vereshagin und Baryshnikov 1984). Der heutige europäische Wisent(Bison bonasus) ist eng mit dem amerikanischen Bison (Bison bison) und dem eiszeitlichen Steppenbison (Bisonpriscus) verwandt. Der Grad der Verwandtschaft wird jedoch unterschiedlich beurteilt (Pucek 1986, 302–303). Ordnung: Artiodactyla Owen 1848; Familie: Bovidae Gray 1821; Gattung: Bison Smith 1827; Eiszeitliche Art:Bison priscus Bojanus 1838; Heutige Art: Bison bonasus (Linnaeus 1758). Unterarten: Europäischer Wisent, B. bonasus bonasus (Linnaeus 1758); Kaukasischer Wisent, B. bonasus cauca-sicus Grevé 1906, ausgestorben 1926; Möglicherweise gab es auch in Ungarn und den Karpaten eine eigeneUnterart, B. bonasus hungarorum Kretzoi 1947, die im 18. Jahrhundert ausgestorben sein soll und viel kleinerals der B. bonasus bonasus war.Vorwiegend verwendete Literatur: Brugal et al. 1999; Heptner et al. 1966, 488–522; Pucek 1986.
Da der Steppenbison ausgestorben ist, wird hier stell-vertretend der europäische Wisent beschrieben.
Habitat: B. b. bonasus lebt meist in Laubwäldernim Flachland, in der Waldsteppe und sogar in reinerSteppe. Die dichten, feuchten Wälder des Urwaldge-biets Belowesher (Białovieża) an der Grenze zwischenPolen und der Ukraine, in denen die letzten Individuendes Wisents im Freien leben, sind als Rückzugsgebietanzusehen (Abb. 56). B. b. caucasicus lebte imKaukasus, im Gürtelgebirge von 900–2100 m.
Verbreitung: In historischer Zeit war B. b. bonasusim Westen bis Frankreich und bis in die Schweiz, imOsten bis zu den Flüssen Don und Wolga verbreitet.B. b. caucasicus war im Kaukasus beheimatet.
Während der Eiszeit reichte die Verbreitung vonBison priscus von der atlantischen Küste Frankreichsund Spaniens bis zum Stillen Ozean und nach Nordsi-birien und von Alaska bis zu den Prärien Nordame-rikas.
Körpermaße: B. b. bonasus, Körperlänge 215–310cm; Schulterhöhe der Stiere 185 cm; Gewicht Kühe320–540 kg, Stiere 530–920 kg. Im Jahr 1555 wurdein Preußen ein Tier getötet, das eine Länge von 4 m,eine Schulterhöhe von 2,6 m und ein Gewicht von952 kg gehabt haben soll (Kempf 1990, 55). B. b.caucasicus war etwas kleiner; Schulterhöhe bis160 cm, Gewicht bis 700 kg, selten 800 kg.
Fell: Bei B. b. bonasus sind die Haare auf demganzen Körper fast gerade oder schwach gekräuselt,an den Seiten des Beckens länger. Die Haare an derVorderseite sind ebenfalls etwas länger. Stierebesitzen einen 37 bis 40 cm langen Bart, unter demHals sind die Haare 18 bis 25 cm lang. Bei B. b.caucasicus waren die Haare gekräuselt – am Körperfein, an Kopf und Hals etwas gröber. Die Haare desBartes waren bis 25 cm, die des Halses bis 15 cm lang.
Paarungszeit: August und September. .
Tragzeit: 40–41 Wochen. Setzzeit: Mai bis Juni; ein,selten zwei Junge. Natürliche Lebenserwartung: 30Jahre.
Nahrung: Laub von Bäumen und Sträuchern,kleine Zweige, Gräser, Baumrinde.
Verhalten: Der heutige Wisent ist tag- und nacht-aktiv. Er lebt in Herden von sechs bis 40 Individuen.Nur alte Bullen sind Einzelgänger. Alle modernenBeobachtungsaussagen über die Herden der Wisentebeziehen sich auf Zeiten, in denen diese Tiere überallschon stark dezimiert waren. Die Herden des Ameri-kanischen Bisons (Bison bison) konnten aus mehrerentausend Tieren bestehen. Eine solche Größenordnung
Abb. 56 Letztes bewohntes Gebiet von Bison bona-sus im 20. Jahrhundert (nach Pucek 1986, 292).

116 Verbreitung der großen Jagdfauna in Mittel und Westeuropa im oberen Jungpleistozän
kann, zumindest in bestimmten Jahreszeiten, für deneiszeitlichen Steppenbison (Bison priscus) nichtausgeschlossen werden.
Klimatische Aussage: Der Wisent scheint sowohlin Steppen- als auch in Waldarealen im Flachland(B. b. bonasus) und in Bergregionen (B. b.hungarorum und B. b. caucasicus) beheimatetgewesen zu sein. Es gibt zwei Unterarten des Ameri-kanischen Bisons (Bison bison): die Nominalform,Bison bison bison, welche in der Prärie Nordamerikasgelebt hat, und Bison bison athabascae, den Amerika-nischen Waldbison. Es lässt sich daher vermuten, dassauch der Steppenbison (Bison priscus), trotz seinesNamens, sowohl in der offenen Landschaft desMammuthus-Coelodonta-Faunenkomplexes als auchin Waldregionen beheimatet war.
Es ist weiterhin interessant zu bemerken, dass dassüdliche Verbreitungsareal des Steppenbisons und dasnördliche Verbreitungsareal des Auerochsen sich invielen Regionen, sowohl in historischer als auch inprähistorischer Zeit stark überlappten. Hier istaußerdem eine Trennung der zwei Arten anhand derKnochenreste schwierig, leichter erscheint sie anhandder Kunst.
Darstellungen aus dem Jungpaläolithikum
In Kantabrien und in den Pyrenäen sind Darstel-lungen von Steppenbisons üblich, wenn nicht sogarhäufig. In manchen Höhlen spielen sie eine zentraleRolle, wie z. B. in Altamira, wo eine ganze Herdedargestellt wurde. Südlich von Kantabrien jedoch sindsolche Darstellungen absolut selten oder sehr fraglich.Keine der von Rodrigo de Balbín Behrmann und JoséJavier Alcolea González (2003, 229) erwähntenSteppenbisondarstellungen in den Höhlen La Hoz, LosCasares und Siega Verde kann als sicher in dieseArbeit eingehen (Alcolea und Balbín 2003, 229). DieGravierung des so genannten „Gran bisonte“ aus derHöhle von Los Casares stellt anhand der Fotos(Acosta und Molinero 2003, 116–117) wahrscheinlichkeinen Steppenbison dar. Die anderen zwei Steppen-bisondarstellungen (aus La Hoz und Siega Verde)konnten nicht überprüft werden. Schließlich soll hier
eine schon immer als unsicher behandelte Darstellungeines Steppenbisons in roter Farbe der Cueva del Reno(Alcolea et al. 1997, 151) erwähnt werden, die imArtikel der gleichen Autoren (Alcolea und Balbín2003, 229) einige Jahre später nicht mehr erwähntwird.
Die vermutete Darstellung eines Steppenbisons ausder Cueva de la Pileta in Südspanien (Breuil et al.1915, Taf. VII), bei der der Kopf fehlt, kann heutenicht mehr als solche akzeptiert werden.
Aus Italien sind drei Darstellungen bekannt: Einevollständige Gravierung auf einer Steinplatte und eineauf einem Knochen von Riparo Tagliente (Abb. 57,links und Mitte) sowie eine Gravierung auf einemGeröll aus Lustigiano (Abb. 57, rechts) in derToskana.
Reste aus dem Jungpaläolithikum
Die Unterscheidung der Knochen von Bison priscusund Bos primigenius ist sehr schwierig (Altuna 1971,349–368; Delpech 1983, 185–191). In den meistenFundstellen in Mitteleuropa wurde nur der Steppen-bison mit Sicherheit bestimmt. Der Auerochse wurdenur während milder Phasen und insbesondere am Endeder Eiszeit nachgewiesen. In Deutschland ist derSteppenbison jedoch nie häufig und sein Anteil in derFauna bleibt meistens viel geringer als der Anteil vonRentier und Pferd. Im Aurignacien Südwestdeutsch-lands ist er nur durch einzelne Reste im Bockstein undVogelherd im Lonetal vertreten. Große Bovidenscheinen im Aurignacien des Achtals zu fehlen.
In Frankreich wird, neben Fundstellen, wo nur derSteppenbison nachgewiesen wurde (u. a. Solutré;Turner 2002), meistens die Kategorie Bovinae oderBos/Bison verwendet. Françoise Delpech meint, dassdie Präsenz von Bos in Südwestfrankreich nur für denBeginn und das Ende des Jungpaläolithikums paläon-tologisch bewiesen sei. Die Bilder von Lascaux so wievon zahlreichen anderen Fundstellen zeigen jedoch,dass der Auerochse nicht verschwunden, sondern nurwährend kälterer Phasen viel seltener als der Steppen-bison war (Delpech 1983, 189; Wüst 1999).
Abb. 57 Ausgewählte Darstellungen von eiszeitlichen Steppenbisons aus Italien. Links und Mitte, Riparo Tagli-ente (Leonardi 1988, 150); rechts, Lustigiano, Lesefund (Leonardi 1988, 166).

Steppenbison (Bison priscus) 117
In Kantabrien ist der Steppenbison immer nochhäufiger als der Auerochse (Altuna 1974) und unter-streicht damit weiter die Verbindung dieser Region zuFrankreich. Südlich der Cordillera Cantabrica und derPyrenäen wird das Bild schlagartig anders. DerSteppenbison fehlt bis jetzt in Portugal (Cardoso 1993,208). In Katalonien ist er viel seltener als derAuerochse (Estévez 1979). In Mittelspanien fehlensichere Informationen (Abb. 58).
In Norditalien werden die Reste von großenBoviden meistens zu der Gruppe Bos/Bison gerechnet.In Mittel- und Süditalien dagegen werden diese fastnur Bos primigenius zugeordnet. Unter den hier unter-suchten Fundstellen wurde nur in der Grotta di Castel-civita die Präsenz des Steppenbisons vom Mittelpaläo-lithikum bis ins Aurignacien nachgewiesen. Die Restevon Bison priscus aus Cava Nuzzo bei Malpignano inApulien (Bologna und Petronio 1994; Bologna et al.1994) sind älter als das Jungpaläolithikum undgehören möglicherweise zu der Isotopenstufe 5c–5a(Bologna et al. 1994, 273).
Diskussion
Der Steppenbison war schon während des Mittel-paläolithikums in der Fauna einiger Fundstellendominierend, z. B. über 90% in Mauran (Delpech1999, 69; Gaudzinski 1996, 23), über 70% in zweiSchichten von Wallertheim (Conard 1999, 308;Gaudzinski 1996, 24). Dies zeigt, dass schon dieNeandertaler in der Lage waren, eine gezielte Jagd aufSteppenbisons durchzuführen. Der Steppenbison stelltjedoch in der Regel nur einen Teil der gejagten Fauna.In den meisten Fundstellen des Mittelpaläolithikumsist sein Anteil an der Fauna sehr gering, oder er fehltganz (Delpech 1999, 69). Im Jungpaläolithikum isteine sehr spezialisierte Jagd auf Steppenbisons nur fürdie Magdalénien-Horizonte von Fongaban bewiesen,wo der Steppenbison die Fauna mit einem Anteil bis
98% dominiert (Delpech 1983, Tableau 25). Trotzeiniger weiterer Fundstellen wie Marcamps undDuruthy mit einem bedeutenden Anteil an Steppen-bisons ist dieses Tier in Frankreich im Allgemeinennicht sehr häufig (Delpech 1999, 74). FrançoiseDelpech konnte für Südwestfrankreich sehr gutzeigen, dass die stärkere oder niedrigere Präsenz vonSteppenbisons direkt mit den klimatischenÄnderungen zu korrelieren sei. Die Spezialisierung inmanchen Fundstellen wäre daher vielmehr Ergebniseiner klimatisch bedingten großen Anzahl an Steppen-bisons als auf ein kulturelles Verhaltensmusterzurückzuführen (Delpech 1999, 82). Ihre Arbeit führtweiterhin die Theorie an, dass das Vorkommen derSteppenbisons in Südwestfrankreich von derAusbreitung der Prärie abhängig gewesen sei. In „zu“milden Perioden verschwanden die Prärien wegen derAusbreitung der Wälder; in „zu“ kalten Periodenverschwanden sie dagegen wegen der zu geringenGrasproduktion (Delpech 1999, 82).
Schließlich ist noch zu erwähnen, dass man, wie esschon für das Mammut und das Wollnashorn der Fallwar, auch vom Steppenbison mittelpaläolithischeReste in Apulien in Süditalien findet. Er überlebt ander tyrrhenischen Küste in Castelcivita (Masini undAbbazzi 1997) mindestens bis zum Aurignacien. Eine14C-Datierung hat ein Datum für diese Schicht von ca.32.000 BP ergeben. Da diese Schicht leider auch diejüngste der stratigraphischen Abfolge dieser Höhle ist,ist die weitere Entwicklung in dieser Region unklar.Ich habe in keiner anderen Höhle Süditaliens einesichere Bestimmung eines Steppenbisons gefunden.Federico Masini und Laura Abbazzi schreiben diesbe-züglich jedoch, „dass man nicht ausschliessen kann,dass die Daten über den Steppenbison in Italien inAllgemeinen und in Süditalien im Besonderen unter-schätzt worden sind“ (eigene Übersetzung aus Masiniund Abbazzi 1997, 42).

118 Verbreitung der großen Jagdfauna in Mittel und Westeuropa im oberen Jungpleistozän
Abb. 58 Mögliche maximale Verbreitung von Bison priscus in Mittel- und Westeuropa während des Jungpaläo-lithikums sowie für das Thema relevante Fundstellen mit Resten (Jungpaläolithische Fundstellen = Dreiecke;Knochenansammlung und ältere Fundstellen = Kreuze) oder Darstellungen (Kreise) von Steppenbisons. DerPfeil weist auf die mögliche Ausbreitung außerhalb des üblichen Verbreitungsgebietes von Bison priscus. 1 Las Caldas; 2 Altamira; 3 Jarama; 4 Riparo Tagliente; 5 Polesini di Bodeno; 6 Lustigiano; 7 Castelcivita;8 Melpignano; 9 San Teodoro; 10 Grotta dei Puntali.

Auerochse (Bos primigenius)
D. Auerochse oder Ur; E. – Aurochs; F. – Aurochs; S. – Uro; Kat. – Ur; P. – Auroque; I. – UroAusgestorben im Jahr 1627 in Jakterowo, Polen, etwa 65 km südlich von Warschau. Der „Rückzucht-Auerochse“ entspricht nur im Äußeren seinem ausgestorbenen Vorbild (Uerpmann 1999). Ordnung: Artiodactyla Owen 1848; Familie: Bovidae Gray 1821; Gattung: Bos Linnaeus 1758; Art: Bos primi-genius Bojanus 1827. Vorwiegend verwendete Literatur: Heptner et al. 1966, 474–482; Weniger 1999a; Wüst 1999.
Habitat: Im Sommer waren Auerochsen oft aufoffenen Wiesen anzutreffen, im Winter suchten sieden Schutz des Waldes, wo sie sich teilweise vonLaub ernährten. In den Pyrenäen lebten sie auch aufden alpinen Wiesen. In manchen Regionen kamen sieaber auch in lichten Wäldern, Waldsteppen undSteppenräumen mit schwach entwickelter Vegetationvor.
Die Gebiete in Polen und Litauen, wo die letztenIndividuen erlegt wurden, sind eher als Zufluchts-areale anzusehen (Abb. 59). Der Auerochse vermiedRegionen mit einer Schneehöhe von über 50 cm.
Historische Verbreitung: Auerochsen waren vonSkandinavien bis Nordafrika und Mesopotamienbeheimatet. Fossile Reste des Auerochsen sind auchaus China und bis zum Pazifik bekannt.
Körpermaße: Körperlänge bei Stieren 260–310 cm; Schulterhöhe 170–180 cm; Gewicht 800–1000 kg. Kühe waren etwas kleiner und leichter.
Fell: In Sommer trug der Auerochse kurzes Haar,im Winter etwas längeres.
Hörner sind sowohl bei Stieren als auch bei Kühenvorhanden. Sie sind hoch aufgesetzt. Die Basen liegenweit voneinander entfernt und am seitlichen Teil desSchädels. Die Hörner sind zunächst auf die Seitegerichtet, dann aber nach vorne gebogen. Sie konnteneine Länge von 40 bis 80 cm erreichen.
Paarungszeit: September. Tragzeit: circa 8 Monate.Setzzeit: Frühjahr.
Nahrung: Hauptsächlich Gräser und Blätter.Verhalten: Besonders angriffslustig, so dass es in
vielen Regionen als Zeichen von Tapferkeit galt, einenAuerochsen erlegt zu haben. Sie lebten meistens inkleinen Gruppen, wobei sie im Sommer eher einzeln,in Winter eher in größeren Herden zu finden waren.
Klimatische Aussage: Bos primigenius ist, beimassivem Auftreten, ein guter Indikator für warmes,feuchtes Klima. Sein Areal ist, im Vergleich zumAreal des Steppenbisons, nach Süden versetzt.Françoise Delpech konnte zeigen, dass eine Korre-lation zwischen seinem Auftreten in Südwestfrank-reich und milderen Klimaverhältnissen existiert(Delpech 1999, 82).
Darstellungen aus dem Jungpaläolithikum
Darstellungen von Auerochsen sind in ganzSüdeuropa und in Südfrankreich häufig (Wüst 1999;Weniger 1999b). In Deutschland scheinen sie, so wiees schon beim Rothirsch der Fall war, zu fehlen. Dieeinzige Ausnahme ist Gönnersdorf (Abb. 60, 9). InBelgien ist eine sehr schöne Gravierung aus demAzilien des Trou de Chaleux zu vermerken (Abb. 60,11).
Die Art der Darstellungen ist zum Teil sehr unter-schiedlich, so dass es, bevor man verschiedeneFundstellen vergleicht, interessant ist, diese Diversitätinnerhalb einer einzigen Fundstelle, Lascaux, zuwürdigen (Abb. 60, 1–8). Lascaux ist mit über 50Bildern die Fundstelle mit den meisten Darstellungenan Auerochsen in Frankreich (Wüst 1999). Hier sindAuerochsen mit einem massiven Kopf und einem sehrkurzen, massiven Hals (Abb. 60, 4 und 5) nebenanderen, die einen verhältnismäßig kleinen Kopf undeinen längeren und feinen Hals besitzen (Abb. 60, 2, 3
Abb. 59 Letztes Vorkommen des Auerochsen in denverschiedenen Ländern des Arbeitsgebietes, die Zif-fern bezeichnen das Jahrhundert n.Chr. (nach Guin-tard 1999, 8).
XI
XII XVII
XIII
X V
XII
XI

120 Verbreitung der großen Jagdfauna in Mittel und Westeuropa im oberen Jungpleistozän
Abb. 60 Ausgewählte Darstellungen von Bos primigenius. 1–8 Lascaux (Leroi-Gourhan et al. 1984, 182–185);9 Gönnersdorf (Bosinski 1981, 100); 10 Teyjat (Leroi-Gourhan et al. 1984, 235); 11 Troux du Chaleux (Lejeune1987, 27); 12 Levanzo; 13 Font-de-Gaume; 14 La Pileta; 15 Pair-non-Pair (12–15 Barriére 1993a, 112).
1 2 3 4 5 6 7 8 9 10 11 12 13 14 15

Auerochse (Bos primigenius) 121
und 6) zu finden. Auch die Hörner sind sehrunterschiedlich dargestellt. Sie sind mal massiv (Abb.60, 5), mal so fein, dass sie mit den Hörnern einerGämse übereinstimmen würden (Abb. 60, 3 und zumVergleich Abb. 70). Auch die Pigmentierung desKörpers betont unterschiedliche Bereiche, nämlich dieSchnauze, den Kopf, den Hals und die Beine. MancheTiere weisen eindeutige Flecken auf. Manche sindrötlicher, andere schwarz abgebildet.
Auffällig ist, dass die Hörner des Auerochsenimmer die Aufmerksamkeit der paläolithischenKünstler erweckt haben. Sie wurden oft nach vorne,seltener nach oben gerichtet dargestellt. Bei manchenDarstellungen wurden die Tiere mit erhobenem Kopfin einer “imponierenden“ oder „sichernden“ Haltungdargestellt (z. B. in Font-de-Gaume und Pair-non-Pair;Abb. 60, 13 und 15). Bei den Darstellungen vonLevanzo (Abb. 60, 12) scheint ein Angriff unmit-telbar bevorzustehen.
Reste aus dem Jungpaläolithikum
In Südfrankreich ist der Auerochse selten. Es sei hierauf das Châtelperronien der Höhle von Belvis, dasAurignacien der Höhle von Canecaude, das Gravettiender Grotte de la Crouzade, das mittlere Magdaléniender Grotte Gazel und das obere Magdalénien derGrotte de la Crouzade für das Languedoc occidentalund das Roussillon (Sacchi 1986, 39, 48, 50, 57, 124,161) sowie auf viele andere Fundstellen insbesonderein Südwestfrankreich hingewiesen (Delpech 1999). Imwestlichen Mitteleuropa dagegen scheint er nie vordem Ende des Jungpaläolithikums (z. B. in Ander-nach, Federmessergruppen) mit Sicherheit nachge-wiesen worden zu sein.
Diskussion
Die Grenze der Verbreitung des Auerochsen imJungpaläolithikum verlief durch die Alpen, durchZentralfrankreich und durch das Massif Central
(Abb. 61). Dafür sprechen, mehr als die zum Teilschwer zu bestimmenden Knochenreste, insbesonderedie Darstellungen. Ohne ausschließen zu können, dassdiese Tiere in besonders günstigen Jahren auchnördlich dieser „Verbreitung“ insbesondere entlangder Flusstäler gewandert sein könnten, ist der Belegbis jetzt nur für das Ende des Jungpaläolithikumserbracht.
Neben einer bestimmten Freiheit, die dem Künstleroder den Künstlern zugebilligt werden muss, insbe-sondere in Bezug auf die Körpermaßen, kann man dieEigenart der Darstellung des Felles bei den Rindernvon Lascaux, bei denen jedes Bild fast die Darstellungeines bestimmten Individuums zu sein scheint, nichtübersehen. Die Tatsache, dass domestizierte Rinderebensolche charakteristischen Farben (schwarz, weißund braun) und Merkmale aufweisen, zeigt, dassmöglicherweise diese Art der Darstellung mindestenszum Teil vom tatsächlichen Aussehen dieser Tiereinspiriert wurde. Es ist bekannt, dass es für Herden-tiere wie Zebras, Gnus, Bisons, Saiga-Antilopen oderRentiere von Vorteil ist, gleich auszusehen. Dadurchkönnen Räuber kein Einzeltier in der Herdeausmachen. Ein „anders“ aussehendes Tier könnte imGegenteil leicht identifiziert und gezielt verfolgtwerden. Dadurch entsteht eine natürliche Selektion,die gegen die „Abweichler“ gerichtet ist. DieAbbildung in Lascaux von im Aussehen stark vonein-ander abweichenden Rindern könnte zeigen, dass dieerwachsenen Tiere keine natürlichen Feinde zufürchten hatten, so dass eine Abweichung imAussehen für sie kein Nachteil war.
Wichtig ist schließlich, dass in Portugal, Zentral-und Südspanien sowie in Italien paläolithischeMalereien grundsätzlich viel seltener und die Art derAusführung der Gravuren nicht so detailreich wie inKantabrien und Frankreich sind (Weniger 1999b,139). Daher verwundert es nicht, wenn die schönstenDarstellungen von Auerochsen in Lascaux an derPeripherie des Verbreitungsgebietes dieser Artwährend der letzten Eiszeit liegen.

122 Verbreitung der großen Jagdfauna in Mittel und Westeuropa im oberen Jungpleistozän
Abb. 61 Im Text erwähnte Fundstellen mit publizierten Resten (Dreiecke) oder Darstellungen (Kreise) vonAuerochsen und deren mögliche Verbreitung im Jungpaläolithikum. Die Pfeile verweisen auf mögliche Wande-rungsrouten außerhalb ihres annehmbaren, üblichen Verbreitungsgebiets. 1 Trou du Chaleux; 2 Gönnersdorf; 3 Lascaux; 4 Font-de-Gaume; 5 Pair-non-Pair; 6 Teyjat; 7 Chauvet;8 La Pileta; 9 Levanzo; 10 Andernach; 11 Pont d’Ambon; 12 Morin; 13 La Ferrassie; 14 Duruthy; 15 Belvis;16 Canecaude und Gazel; 17 Crouzade; 18 Valorgues; 19 Cornille.

Moschusochse (Ovibos moschatus)
E. – Musk-ox; F. – Boeuf musqué; S. – Buey almizclado; Kat. – Bou mesquer; P. – Boi-almiscarado; I. – Buemuschiato.Ordnung: Artiodactyla Owen 1848; Familie: Bovidae Gray 1821; Gattung: Ovibos de Blainville 1816; Art:Ovibos moschatus (Zimmermann 1780).Ausgestorben in Europa, hat in Kanada und Grönland überlebt und wurde 1772 an der Hudson Bay wiederent-deckt. Von Kanada und Grönland wurde er wieder nach Europa eingeführt. Vorwiegend verwendete Literatur: Bohlken 1986; Crégut-Bonnoure 1984; Gessain 1981; Rathgeber 1994;Raufuss und Koenigswald 1999; Soergel 1942.
Habitat: Der Moschusochse ist regen- und feuchtig-keitsempfindlich, daher ist seine Verbreitung auf dieoffene, trockene Tundra beschränkt. Sein Fell schütztihn vor kalten Temperaturen und Wind.
Verbreitung: Zur Zeit kommt er in der arktischenTundra Grönlands und Kanadas vor (Abb. 62). Erwurde in Norwegen ebenfalls in sehr trockenenRegionen erfolgreich eingeführt.
Körpermaße: Länge 190 bis 250 cm; Schulterhöhe110 bis 150 cm; Gewicht: 190 bis über 400 kg.
Körperbeschreibung: Stämmig mit kurzen Beinenund langem, im Winter bis zum Boden reichendenFell. Damit gewinnt man den Eindruck einer geschlos-senen Körperform. Die Hörner sind charakteristisch.Sie sind in den ersten drei Vierteln nach unten undnach außen gerichtet, im letzten Viertel richten siesich jedoch nach vorne und nach oben. Die Männchenriechen während der Brunst stark nach Moschus. Indieser Jahreszeit ist ihr Fleisch ungenießbar.
Fell: Die Oberhaare sind grob und 45 bis 62 cmlang; Die Unterwolle ist weich, dicht und bis 7 cmlang. Das Fell der Moschusochsen ist das längste unterden lebenden Säugetieren.
Paarungszeit: Vom Klima abhängig, zweite Juli-hälfte bis September und Oktober. Tragzeit: 8 bis 8½Monate. Setzzeit: Ende April bis Mitte Juni.
Natürliche Lebenserwartung im Freiland 20 Jahre.Nahrung: Gräser, Kräuter und Zwergsträucher.
Verhalten: Moschusochsen leben gesellig in Herdenvon 4 bis 65 Individuen. In manchen älteren Berichtenwird von Gruppen von 100 Individuen gesprochen.Die Sommerverbände sind kleiner. Bullen könnenEinzelgänger sein. Bei Bedrohung fliehen sie meistensnicht, sondern drücken sich eng aneinander. Sie bildeneinen Kreis und bieten dem Feind die Stirn. DiesesVerhalten erwies sich jedoch gegenüber demMenschen als kontraproduktiv. Wenn ein einzelnerBulle zur Verteidigung gezwungen ist, kann erplötzlich angreifen. Dabei haben seine Hörner fürWölfe oft fatale Folgen.
Auch bei starkem Wind oder extremer Kälte drückensie sich eng aneinander, dabei werden junge Tierebesonders geschützt. Sie sind die kälteresistenteste Artunter den Pflanzenfressern und ertragen ohneProbleme Temperaturen von -50°C.
Moschusochsen sind ortstreu. Sie wandern inner-halb derselben Region auf der Suche nach Essbarem,ohne eine feste Route zu haben. Es ist beobachtetworden, dass Moschusochsen im Winter auf Hügelsteigen, da hier der Wind den Schnee wegfegt. ImSommer sind sie auf der sonnigeren Seite von Hügelnund Gebirgen sowie entlang von Flüssen und Seen zufinden (Gessain 1981, 59). Sie können leicht sehrsteile Hänge ersteigen.
Abb. 62 Gebiete, in denen Ovibos moschatus bisheute überlebt hat (nach Pedersen 1958, 11).

124 Verbreitung der großen Jagdfauna in Mittel und Westeuropa im oberen Jungpleistozän
Klimatische Aussage: Wegen seines Felles ist derMoschusochse ein deutlicher Hinweis auf mitextremer Trockenheit verbundener Kälte. Wegen derkurzen Beine ist er schon bei einer leicht höherenSchneedecke in der Bewegung behindert. Sieerkranken leicht bei feuchten Verhältnissen anLungenentzündung.
Darstellungen aus dem Jungpaläolithikum
Man findet nur wenige Darstellungen von Moschus-ochsen. Manche davon sind fraglich. Die Darstellungauf dem Lochstab von Arudy-Espalungue (Abb. 63, 4)wurde schon früh in Frage gestellt, da auf der anderenSeite der Kopf eines Steinbockes vorhanden ist, undweil das Horn eine Rinne aufweist, die auf die Falteder Hörner bei Steinböcken hinweisen könnte (Chollot1964, 191). Die Biegung des Hornes wäre also von derForm des Lochstabes diktiert. Bei der Gravierung vonPech-Merle (Abb. 63, 8) wird sogar deren Existenzangezweifelt (Barrière 1993a, 122). Die möglichenGravierungen von La Mouthe (Abb. 63, 10) undLascaux (Abb. 63, 11) sind unsicher (Barrière 1993a,122). Die Gravierung auf einem Geröll (Abb. 63, 3)von La Colombière (Leroi-Gourhan 1966, 426) wirdsowohl als möglicher Moschusochse als auch alsmöglicher Steppenbison angesprochen, der Block vonRoc-de-Sers (Abb. 63, 9) wird ebenfalls aufgrund derProportionen, Größe und der ikonographischenVergleiche mit anderen Mensch-Tier-Darstellungen(siehe Abb. 63, 9) eher als ein Steppenbison als einMoschusochse angesprochen (Leroi-Gourhan 1966,419), obwohl andere Autoren es als sichereDarstellung annehmen (Barrière 1993a, 122; Airvaux2001, 68). Nur die Darstellungen von Laugerie-Haute(Abb. 63, 1), zwei Darstellungen aus Enlène, die eineauf einer Steinplatte, die andere auf Rentiergeweih(leider kann hier kein Bild davon wiedergegebenwerden), das Speerschleuderbruchstück aus demKesslerloch (Abb. 63, 5), die Speerschleuder (Abb.63, 2) von Bruniquel (Stodiek 1993, 445–446) und diezwei Malereien aus der Höhle von Chauvet (Abb. 63,
7–8) gelten in der Literatur als sicher. Dieseeindeutigen Darstellungen reichen jedoch aus, um beider allzu schnellen Ablehnung der unsicherenVorsicht zu verlangen56.
Reste aus dem Jungpaläolithikum
Die Arbeiten von Françoise Delpech und Emile Heintz(1976) hinsichtlich Frankreich sowie von WolfgangSoergel (1942), Ingo Raufuss und Wighart vonKoenigswald (1999) geben eine Liste der Funde ausMittel- und Westeuropa wieder57.
Die Reste von Ovibos moschatus aus dem spätenPleistozän stammen meistens aus magdalénienzeit-lichen Schichten. Dies ist im Kesslerloch (Schweiz),in der Burghöhle Dietfurt (Deutschland) und im Abridu Poisson in Les Eyzies (Frankreich) der Fall.Einzelne Funde stammen jedoch auch aus demSolutréen von Laugerie-Haute (Frankreich), aus derÜbergangsphase vom Gravettien zum Solutréen derFundstelle von Arbreda (Spanien), aus einer Schichtder Potočka Höhle (Slowenien), die zwischen 27.000und 35.750 BP datiert wird, und aus der FundstelleBeckford (Großbritannien), die auf ca. 27.650 BPdatiert wird. In Mitteleuropa fehlen anscheinend Restevon Moschusochsen aus dem Aurignacien undGravettien (Raufuss und Koenigswald 1999, 388). Diemeisten Fundstellen haben nur Reste von einemIndividuum erbracht (u. a. in Baden-Württemberg:Rathgeber 1994, 9).
Wie zu erwarten, tritt Ovibos moschatus immerzusammen mit anderen Vertretern des Mammuthus-Coelodonta-Faunenkomplexes auf. Das ist z. B. imKesslerloch (Schweiz), wo er zusammen mit Rentier(ca. 85%), Wollnashorn und Mammut auftritt, und inder Burghöhle Dietfurt (Deutschland) der Fall, wo erzusammen mit Rentierknochen gefunden wurde. Inder Schicht C der Höhle von Arbreda kam ein Zahnvon Moschusochse aus einer Schicht mit zahlreichenResten von Equus ferus und Cervus elaphus vor.
56 André Leroi-Gourhan schrieb in Bezug auf die mögliche Moschusochsendarstellung von La Colombière1965: „Je ne connais pourtant aucune représentation de cet animal qui ne puisse aussi bien s’appliquer au bison“(1965, 426). Diese Aussage wurde ebenfalls in der deutschen Übersetzung 1971 wiedergegeben: „Obwohl dieseAnnahme ... sehr wahrscheinlich klingt, gibt es meines Wissens keine Darstellung dieser Spezies, die nichtgenauso als Bison gedeutet werden könnte“ (1971, 544). In der neueren erweiterten Auflage von Brigitte und Gil-les Delluc (Leroi-Gourhan 1995, 422) bleibt der Text unverändert.57 Es sei hier darauf hingewiesen, dass die zwei Reste aus dem Vogelherd (Rathgeber 1994, 13; Raufuss undKoenigswald 1999, 391) bei einer nachträglichen Untersuchung durch François Poplin die Bestimmung einesRentiers und eines Boviden ergeben haben sollen (Weniger 1982, 88). Die Angabe von Gerd Christian Weniger,dass es Reste von Moschusochsen aus dem Aurignacien der Höhle Bockstein IV gäbe (Weniger 1982, 119),konnte bei einer Überprüfung der Fauna aus dem gesamten Bocksteinkomplex durch Petra Krönneck im Jahr2003 nicht bestätigt werden. Aus der Bocksteinschmiede konnten dafür neben dem bekannten distalen Ende einesFemurs (Lehmann 1969, Taf. 166.6; Rathgeber 1994, 14) ein weiteres Bruchstück eines Femurs sowie ein Meta-carpus dem Moschusochsen zugeordnet werden (mündliche Mitteilung Petra Krönneck). Alle diese Stücke stam-men aus Schichten, die dem Micoquien zugeordnet werden (Rathgeber 1994, 14–15; Hahn et al. 1985, 64).

Moschusochse (Ovibos moschatus) 125
Diskussion
Die letzten Moschusochsen in Eurasien starben vor ca.4.000 Jahren auf der Halbinsel Tajmyr in Sibirien,also fast gleichzeitig mit den letzten Mammuts auf derWrangel-Insel aus. Auch das Fell und die klimatischeAnpassung des Moschusochsen entsprechen der desWollnashorns und des Mammuts. Die Tatsache, dassReste von Coelodonta antiquitatis und Mammuthusprimigenius bis Andalusien bekannt sind, währendOvibos moschatus in Italien und in Spanien (mitAusnahme der Höhle von Arbreda) fehlt58, wird vonIngo Raufuss und Wighart von Koenigswald (1999,386) als Indiz verstanden, dass diese Art sehr trockeneGebiete bevorzugt und jegliche Feuchtigkeitvermeidet, fast noch mehr als Mammut undWollnashorn. Dieser Vergleich muss jedoch relati-viert werden: Einerseits, da alle Reste von Coelo-donta antiquitatis und Mammuthus primigenius ausSüdspanien und Süditalien älter als 40.000 Jahre sind(siehe S. 64 und 73) und man folglich nicht sicher seinkann, ob die Tiere, zu denen diese Knochen gehörten,die gleichen Klimaverhältnisse brauchten wie dieWollnashörner und Mammuts, die 30.000 Jahre späterlebten. Andererseits darf man nicht vergessen, dass dieKnochen der Megafauna immer wegen ihrer Größeauffallen, während bei anderen Tieren dieBestimmung der postkranialen Knochen deutlichschwieriger ist und sie meistens nur anhand der Zähnebestimmt werden.
Die Tatsache, dass nur wenige Reste von Ovibosmoschatus sowie nur zwei Moschusochsendarstel-lungen aus der Zeit des frühen Jungpaläolithikumsstammen, hat mit Sicherheit mit der klimatischenAnpassung dieser Art zu tun. Dies wird von derstärkeren Verbreitung dieser Art in Mitteleuropagegenüber der in Westeuropa verdeutlicht (Crégut-Bonnoure 1984, 138; Raufuss und Koenigswald 1999,387) (Abb. 64). Andererseits haben sicherlich andereAspekte auch eine Rolle gespielt; sonst wäre es kaumzu erklären, dass Reste von Moschusochsen häufigerin einer Zeit auftreten, in der gleichzeitig die Restevon Wollnashörnern und Mammuts seltener werden.Heutzutage ist, abgesehen vom Menschen, der einzigeFeind des Moschusochsen der Wolf. In der Eiszeit gabes jedoch in Europa auch Höhlenhyänen und Löwen.Diese scheinen am Ende der Eiszeit (Solutréen undMagdalénien) deutlich seltener zu werden, und sieverschwinden aus manchen Regionen vollständig.Gleichzeitig scheint die Zahl der Moschusochsenrestezu steigen. Nun wissen wir nichts über dasVerteidigungsverhalten von Moschusochsen gegen-
über einem Rudel von Löwen oder gegenüber einemeinzelnen 250 kg schweren Löwen. Man kann jedochannehmen, dass die Bildung eines Kreises und dieDarbietung der Stirn und der Hörner gegenüber einemRudel von Löwen weniger erfolgreich war, alsgegenüber einem Rudel von Wölfen, bei dem jedeseinzelne Tier maximal 75 kg wiegt. Ein Zusam-menhang zwischen der Zunahme an Moschusochsenin Europa bei der gleichzeitigen Abnahme von Löwenund Hyänen scheint mindestens möglich zu sein.Wenn man dann in die Argumentation die Rolle desMenschen einbringt, wird klar, dass Moschusochsenmit der Bildung eines Kreises und der Darbietung derStirn und der Hörner, so wie sie es heute tun,gegenüber dem Menschen und seinen Fernwaffen garkeine Chance hatten.
Von den wenigen Darstellungen von Moschus-ochsen stammen zwei von Speerschleudern (Grotte deBruniquel und Kesslerloch; Abb. 63, 2 und 5). Eineweitere Darstellung (Roc-de-Sers; Abb. 63, 9) zeigtden Angriff eines möglichen Moschusochsen aufeinen Menschen. Dies zeigt, dass der MenschMoschusochsen eventuell häufiger gejagt hat, als esdie Knochenfunde uns zeigen.
Noch heute leben Moschusochsen nur in extremenRegionen, in denen es sogar für die Feinde undKonkurrenten zu schwierig ist zu überleben. Gerade inDeutschland war das Klima während des zweitenKältemaximums der letzten Eiszeit für die Menschenund viele Tiere so hart, dass mehrere jahrhunderte-oder sogar jahrtausendelang keine Menschen hierleben konnten. In einem solchen Raum kann man sichgut vorstellen, dass Moschusochsen eine stabilePopulation entwickelten, die auch den ökologischenDruck einer geringen menschlichen Populationsdichteim Magdalénien ertragen konnte. Und tatsächlichstammen die meisten Reste von Moschusochsen ausDeutschland (u. a. Rathgeber 1994). Ganz anders siehtes für Frankreich aus. Hier war der Mensch ohneUnterbrechungen ansässig, und zwar mit einer imVergleich zu Deutschland höheren Populationsdichte(u. a. Bocquet-Appel und Demars 2000; Demars 1996und 2002). Schließlich zeigt die Tatsache, dass Restevon Moschusochsen nie in großer Anzahl in paläoli-thischen Fundstellen zu finden und meistens nur aufein einzelnes Individuum zurückzuführen sind, dass,auch wenn diese Tiere vorhanden waren, sie niehäufig erbeutet wurden. Dies könnte gut mit demHinweis zusammenpassen, dass die IndianerNordamerikas das Fleisch von Rentieren dem Fleischvon Moschusochsen vorzogen (Gessain 1981, 32) .
58 In diesem Zusammenhang sei auf die Präsenz von Praeovibos priscus in Andalusien, nahe Granada, hinge-wiesen. Dieser Fund stammt möglicherweise aus dem Mittelpleistozän (Moya-Sola et. al. 1981).
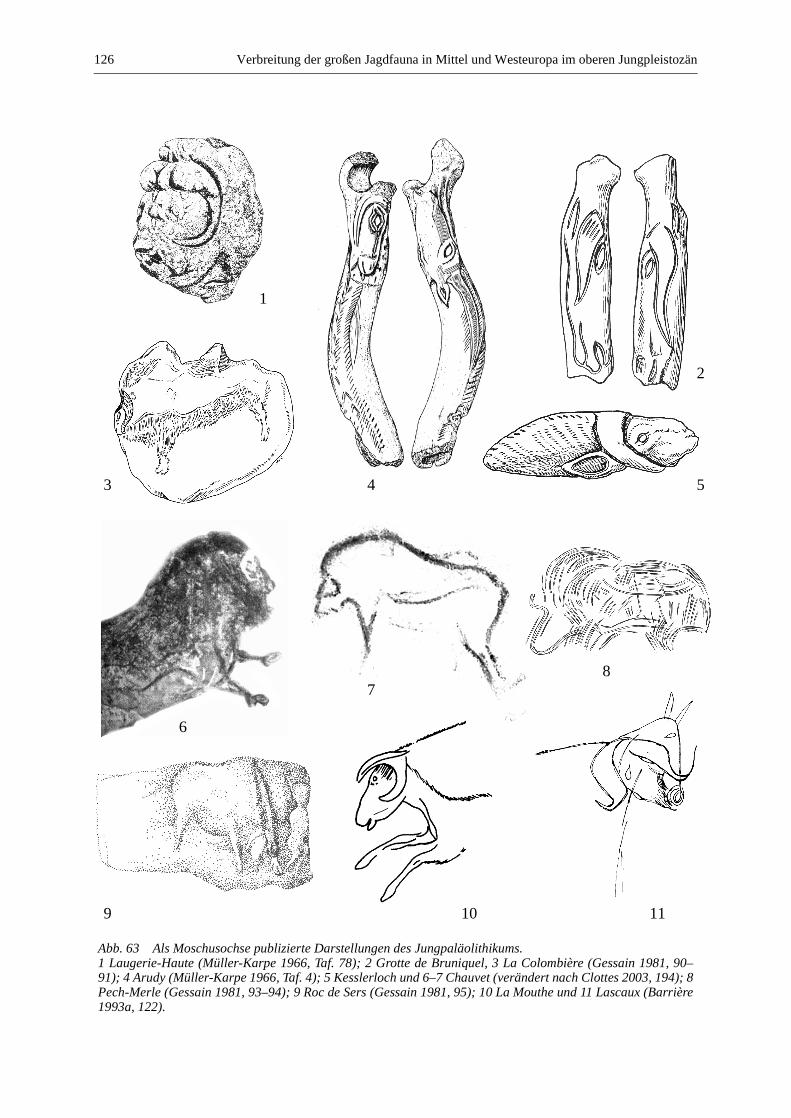
126 Verbreitung der großen Jagdfauna in Mittel und Westeuropa im oberen Jungpleistozän
Abb. 63 Als Moschusochse publizierte Darstellungen des Jungpaläolithikums.1 Laugerie-Haute (Müller-Karpe 1966, Taf. 78); 2 Grotte de Bruniquel, 3 La Colombière (Gessain 1981, 90–91); 4 Arudy (Müller-Karpe 1966, Taf. 4); 5 Kesslerloch und 6–7 Chauvet (verändert nach Clottes 2003, 194); 8Pech-Merle (Gessain 1981, 93–94); 9 Roc de Sers (Gessain 1981, 95); 10 La Mouthe und 11 Lascaux (Barrière1993a, 122).
1 2 3 4 5 8 7 6 9 10 11

Moschusochse (Ovibos moschatus) 127
Abb. 64 Verbreitung von Moschusochsen im Jungpaläolithikum sowie in diesem Kapitel erwähnte paläolithi-sche Fundstellen mit Resten (Dreieck) oder Darstellungen (Kreis) dieser Tiere. Die Daten über die Fauna(sowohl paläontologische wie auch archäologische Fundstellen) nach Raufuss und Koenigswald 1999. 1 Lauge-rie-Haute; 2 Bruniquel; 3 La Colombière; 4 Arudy; 5 Roc-de-Sers; 6 Chauvet; 7 Pech-Merle; 8 Beckford;9 Maidenhead/Thames; 10 Trou de Chaleux; 11 Furfooz (Trou de Reuviau); 12 Trou de Goyet; 13 Unkelstein;14 Dömnitz; 15 Siersleben; 16 Ranis; 17 Regensburg; 18 Vogelherd (zweifelhaft, siehe Fußnote 57); 19 HohleFels; 20 Burghöhle-Dietfurt; 21 St. Jakob; 22 Kesslerloch; 23 Abri du Poisson; 24 La Madeleine; 25 Arbreda.


Steinbock (Capra ibex und Capra pyrenaica)
E. – Alpine Ibex, Spanish Ibex; F. – Bouquetin des Alpes, Bouquetin des Pyrénées; S. – Ibice de los Alpes,Cabra montes iberica; Kat. Cabra dels Alps (Ibex dels Alps), Cabra pirenenca; P. – Cabra montez, cabra doGerês; I. – Stambecco, Stambecco iberico.Ordnung: Artiodactyla Owen 1848; Familie: Bovidae Gray 1821; Gattung: Capra Linnaeus 1758; Art: Capraibex Linnaeus 1758. Art: Capra pyrenaica Schinz 1838; Unterarten: C. p. lusitanica França 1909; C. p.pyrenaica Schinz 1838; C. p. victoriae Cabrera 1911; C. p. hispanica Schimper 1848.Vorwiegend verwendete Literatur: Engländer 1986; Griggo 1991; Nievergelt und Zingg 1986; Sacchi 1993a.
Habitat: Der Lebensraum des Steinbockes bestehtheute aus hohen Gebirgen und felsigem Untergrund.Im Sommer oberhalb der Waldgrenze (in den Alpen2500 bis 3000 m), im Winter in tieferen Regionen.
Verbreitung: Im 19. Jahrhundert war Capra ibexmit Ausnahme des italienischen NationalparksGranparadiso aus ganz Europa verschwunden(Niethammer und Krapp 1986, 392). Mit Hilfe ausdieser Restpopulation stammender Tiere wurde dieArt in den ganzen Alpen wieder eingebürgert. Caprapyrenaica lebt auf der Iberischen Halbinsel. C. p.lusitanica ist im Jahre 1892 ausgestorben(Niethammer und Krapp 1986, 418), sie lebte in denGebirgen von Galizien und Nordportugal, C. p.pyrenaica hat nur in den zentralen Pyrenäen überlebt,C. p. victoriae lebt in der Serra de Gredos, in Zentral-spanien, C. p. hispanica im Süden und SüdwestenSpaniens (Abb. 65).
Körpermaße von Capra ibex: Kopf-Rumpf-Länge100 bis 150 cm; Schulterhöhe 70 bis 100 cm; GewichtBöcke 80 bis 125 kg, Weibchen ca. 50 kg.
Körpermaße von Capra pyrenaica: Kopf-Rumpf-Länge 90 bis 140 cm; Schulterhöhe bis 75 cm;Gewicht: Böcke 35 bis 80 kg. Weibchen 35 bis 40 kg.Größe und Gewicht sind in Südspanien noch geringer.
Die Hörner von Capra ibex drehen sich bogen-förmig nach hinten. Bei Böcken sind sie in der Regelbis 75 cm lang, in Ausnahmefällen bis 140 cm.Weibchen besitzen viel kleinere Hörner. Die Hörnerbilden ein bis drei Knoten pro Jahr. Die Hörner vonCapra pyrenaica sind leierförmig. Bei Böcken sind siein der Regel bis 90 cm lang, selten darüber. Weibchenbesitzen viel kleinere Hörner (ca. 20 cm). Die Hörnerbilden jährlich ein bis drei kleine Wülste und sind umihre Achse gedreht, so dass die Seite, die an der Basisnach vorne gerichtet ist, in der Mitte nach außen undan der Spitze nach hinten zeigt. Dabei muss jedochbetont werden, dass unter den Iberischen Steinböckendie Hörner eine große Variabilität haben, so dass inSüdspanien manche Böcke Hörner tragen, die denendes Alpensteinbocks stark ähneln (Engländer 1986,414).Paarungszeit von Capra ibex: Dezember und Januar.Setzzeit von Capra ibex: Mai/Juni.
Paarungszeit von Capra pyrenaica: Oktober/November. Setzzeit von Capra pyrenaica: April/Mai,in Südspanien auch im März. Tragzeit: 5½ Monate.
Natürliche Lebenserwartung: 15 bis 20 Jahre.Nahrung: Kräuter, Zwergsträucher, Flechten.Verhalten: Steinböcke sind Tiere, die auf das
Leben auf Felsen spezialisiert sind. In einer solchenLandschaft sind sie für Landraubtiere schwererreichbar. Männliche Tiere bleiben oft auf Felsenstehen. Folglich wurden sie zu einer leichten Beute fürFernwaffen (Lenardi 1990, 106). Sie sind tagaktiv.Alte Böcke sind oft Einzelgänger, sie leben sonst inVerbänden von bis zu 30 Tieren.
Klimatische Aussage: Obwohl Steinböcke imAllgemeinen mit extremen Klimazonen in Verbindunggebracht werden, zeigen gerade die Funde aus demJungpaläolithikum, dass diese Tiere früher inSüditalien bis zum Meer vorkamen (wie in Grottadella Cala und Grotta Paglicci; Sala 1985). Erst durchdas Aufkommen des Waldes, die starke Bejagung, dieVerbreitung von Landwirtschaft und Viehzucht wurde
Abb. 65 Heutige Verbreitung von Capra ibex undCapra pyrenaica (nach Mitchell-Jones et al. 1999,412–415).
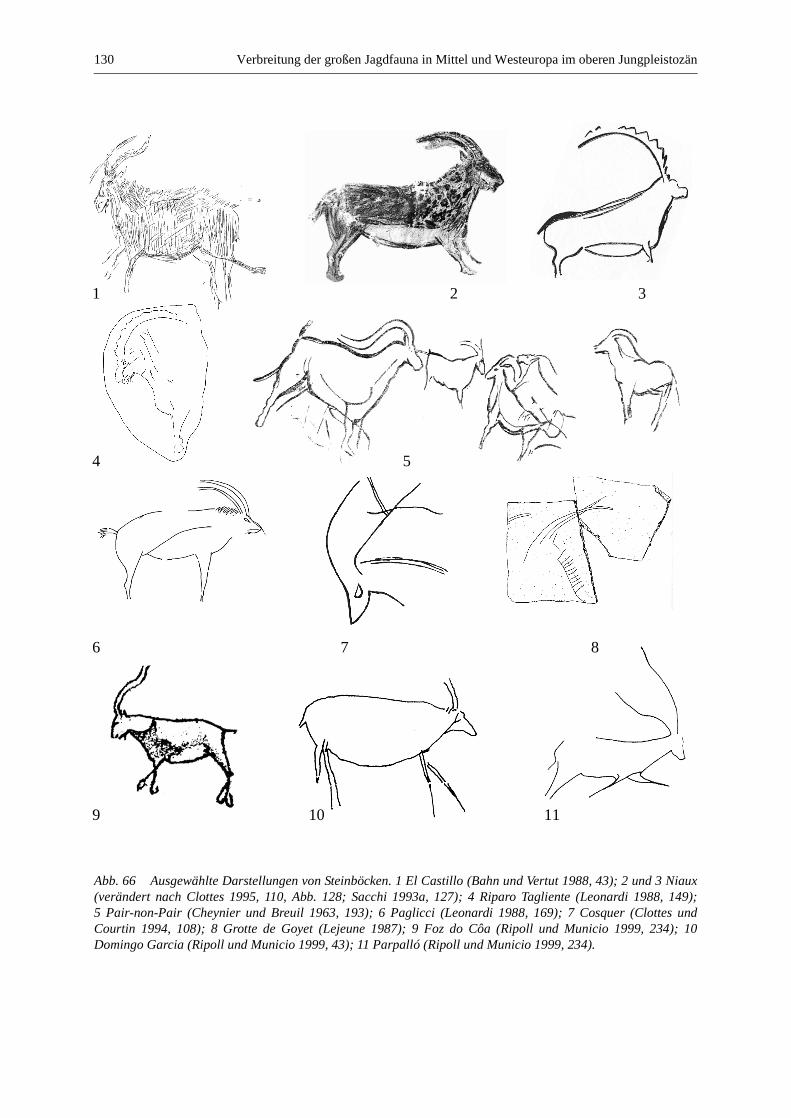
130 Verbreitung der großen Jagdfauna in Mittel und Westeuropa im oberen Jungpleistozän
Abb. 66 Ausgewählte Darstellungen von Steinböcken. 1 El Castillo (Bahn und Vertut 1988, 43); 2 und 3 Niaux(verändert nach Clottes 1995, 110, Abb. 128; Sacchi 1993a, 127); 4 Riparo Tagliente (Leonardi 1988, 149);5 Pair-non-Pair (Cheynier und Breuil 1963, 193); 6 Paglicci (Leonardi 1988, 169); 7 Cosquer (Clottes undCourtin 1994, 108); 8 Grotte de Goyet (Lejeune 1987); 9 Foz do Côa (Ripoll und Municio 1999, 234); 10Domingo Garcia (Ripoll und Municio 1999, 43); 11 Parpalló (Ripoll und Municio 1999, 234).
1 2 3 4 5 6 7 8 9 10 11

Steinbock (Capra ibex und Capra pyrenaica) 131
ihr Lebensraum auf hohe Gebirge begrenzt. Sie lebenheute in Regionen, wo die Niederschläge im Jahr 300bis 700 mm, die durchschnittliche Temperatur imJanuar -20 bis 0°C und im Juli 15 bis 25°C beträgt.
Darstellungen aus dem Jungpaläolithikum
Steinböcke gehören zu den am häufigsten darge-stellten Tieren. Darstellungen sind aus Südspanien(Nerja), aus Süditalien (Paglicci) und vielleicht ausGroßbritannien (Church Cave) bekannt. DieVerbreitung der Darstellungen konzentriert sicheindeutig dort, wo die Landschaft eher den Lebensbe-dürfnissen dieser Tiere entspricht.
Es ist interessant zu vermerken, dass die heutigeTrennung zwischen Alpen- und Pyrenäensteinbockschon im Jungpaläolithikum nachvollziehbar ist. InItalien und fast ganz Frankreich kommt immer einSteinbock vor, der Knoten und stark gebogene Hörneraufweist. Die Darstellungen von Steinböcken inSpanien, sowie in Südfrankreich um den Pyrenäenzeigen dagegen häufig leierförmige, glatte Hörner.Diese Unterschiede sind um so stärker, wenn manDarstellungen aus Kantabrien, wie beispielsweise ausEl Castillo (Abb. 66, 1), mit der aus Riparo Taglienteauf der Südseite der Alpen vergleicht (Abb. 66, 4).
Die Darstellung eines Steinbockes mit deutlichenKnoten und nach hinten gebogenen Hörnern in Niaux(Abb. 66, 3) erinnert stark an einen Alpensteinbockund scheint aufgrund der geographischen Lagezunächst überraschend. Diese Darstellung kann aberauch durch die große Variabilität der Hörner bei denIberischen Böcken erklärt werden. Auch in der Höhlevon Nerja in Südspanien entsprechen nur die Hörnervon zwei Individuen der leierförmigen, geschwun-genen Gestalt (Dams 1987, 214). Daraus folgt, dassnur die Darstellung von leierförmigen Hörnern in derKunst als Kriterium für die Bestimmung der Artgelten kann. Gebogene Hörner können überall darge-stellt sein. In diesem Zusammenhang ist eine Serievon Steinböcken (Abb. 66, 5) aus der Höhle von Pair-non-Pair (Gironde, Frankreich) sehr aufschlussreich,da diese die nördlichsten Darstellungen vomIberischen Steinbock bietet59. Aus dem Périgordwurden drei Darstellungen von Iberischen Stein-böcken publiziert (Delpech 1983, 202) . Diese müssenjedoch zumindest als fraglich eingestuft werden undwurden daher in dieser Arbeit nicht als iberische
Steinböcke berücksichtigt. Die Gravierung des Stein-bockes vom Abri Delluc, die Abbé Glory publizierthat (1949), wurde von Alain Roussot und NorbertAujoulat als unlesbar und die zeitliche Zuordnung alssehr fraglich bezeichnet (Leroi-Gourhan et. al 1984,74). Die Identifizierung der Gravierung in der GrotteNancy, die durch die Umzeichnung von Henri Breuileindeutig erscheint, wird von Alain Roussot alsextrem schwierig bezeichnet, so dass er nicht in derLage war, den Kopf zu finden (Leroi-Gourhan et. al1984, 149). Die Umzeichnung von Henri Breuil voneiner 26 cm langen Gravierung eines Steinbockes ausder Grotte de Comarque (Capitan et al. 1915, 511;Delpech 1983, 195; Cardoso 1993, 236) sieht bei derUmzeichnung von Brigitte und Gilles Delluc (Leroi-Gourhan et. al 1984, 121) ganz anders aus.
Schließlich ist noch die Darstellung in der HöhleCosquer, wo die Hörner aus einzelnen Linienbestehen, zu erwähnen (Abb. 70, 14). Hier ist eineTrennung zwischen Steinbock und Gämse nichtimmer leicht.
Da die Hörner bei erwachsenen, männlichen Stein-böcken länger als bei weiblichen und jungen Tierensind, kann man annehmen, dass meistens männlicheTiere dargestellt sind.
Die Malerei aus der levantinischen Kunst derCueva Remigia in Spanien (Abb. 67) zeigt, wie dieseTiere nach der Eiszeit in Spanien dargestellt wurden.
59 Es sei ergänzend erwähnt, dass im Abri Roc-aux-Sorciers in Angles-sur-l’Anglin eine Darstellung einesmännlichen Steinbockes Hörner besitzt, die die für den Steinbock der Pyrenäen typische Biegung aufzuweisenscheinen (Airvaux 2001, 166–167). Ob der erste Eindruck stimmt – Jean Airvaux äußert sich sehr vorsichtig(2001, 151) – bedarf einer Überprüfung. Daher wurde diese einzelne Darstellung in die Untersuchung über dieVerbreitung von C. ibex und C. pyrenaica nicht einbezogen.
Abb. 67 Cueva Remigia, Levantine-Kunst (Beltrán1979, 360).

132 Verbreitung der großen Jagdfauna in Mittel und Westeuropa im oberen Jungpleistozän
Reste aus dem Jungpaläolithikum
Der Steinbock ist während der letzten Eiszeit in ganzEuropa zu finden (Griggo 1991). In Großbritannien ister durch einzelne Funde belegt (Robin Hood’s Cave)(Abb. 68). Um so häufiger ist er dort, wo felsigerUntergrund existiert. In manchen Fundstellen ist er alsbevorzugte Jagdbeute zu betrachten. Steinbock istunter den Knochenresten mit ca. 90% in Rascaño undNerja (Spanien), mit über 95% in Les Eglises und LaVache (Frankreich), mit über 75% in Riparo Taglienteund Paglicci sowie ebenfalls mit einem hohen Anteilin Arene Candide und Grotta del Mezzogiorno(Italien) vertreten. Durch seine weite Verbreitungkommt der Steinbock sowohl in Regionen vor, woCervus elaphus, als auch dort, wo Equus ferus, diegroßen Boviden oder Rangifer tarandus starkvertreten sind. Die Untersuchung der Hornbasis vonmehreren Schädeln insbesondere in Frankreich undSpanien konnte zeigen, dass eine Trennung zwischenCapra ibex und Capra pyrenaica schon im Jungpaläo-lithikum vorhanden war (Griggo 1991; Altuna 1972,409).
Diskussion
Die Tatsache, dass männliche Tiere häufiger alsWeibchen und junge Tiere dargestellt sind, kannunterschiedlich interpretiert werden. Einerseits könntees damit zusammenhängen, dass Steinböcke geradedurch ihre Hörner zu charakterisieren sind. Daherkönnten, um sicher zu sein, dass der Inhalt des Bildesverstanden wird, zu der Darstellung eines Steinbockeslange Hörner gehört haben. Eine sexuelle Informationder Art „das ist ein männlicher Steinbock“ wäre, indiesem Sinne, von sekundärer oder sogar von garkeiner Bedeutung. Andererseits könnten die Hörnergerade als Unterscheidungsmerkmal zwischenmännlichen und weiblichen Tieren verwendet wordensein. Die Information „das ist ein männlicherSteinbock“ wäre von primärer Bedeutung. Das ist mitSicherheit der Fall bei Darstellungen, in denen sichzwei männliche Steinböcke gegenüberstehen, wie aufder Speerschleuder von Les Trois Frères und demHochrelief vn Roc-de-Sers (Airvaux 2001, 70). Diesewerden als Kampfszenen in der Paarungszeit interpre-tiert. Hörner sind weiterhin Symbole von Stärke undhaben ohne Zweifel die Aufmerksamkeit derMenschen auf sich gezogen. Schließlich darf nichtunerwähnt bleiben, dass weibliche und junge Stein-böcke schneller zur Flucht übergehen als männlicheSteinböcke (Rauer-Gross 1992), die dadurch leichterder Jagd zum Opfer fallen.

Steinbock (Capra ibex und Capra pyrenaica) 133
Abb. 68 Verbreitung von Capra ibex (dunkelgrau) und Capra pyrenaica (hellgrau) sowie in diesem Kapitelerwähnte paläolithische Fundstellen mit Resten (Dreieck) oder Darstellungen (Kreis) dieser Tiere (siehe dazuGriggo 1991). 1 Grotte de Goyet (Kunst, Magdalénien) und Grotte de Goyet (Fauna Aurignacien), Grotte du Prince und Grottede la Princesse; 2 Trou du Renard und Trou Magrite; 3 Robin Hood’s Cave; 4 Church Cave; 5 Limeuil, LaMouthe, Les Combarelles, Rouffignac; 6 Cougnac; 7 Chauvet; 8 Cosquer; 9 Arene Candide; 10 RiparoTagliente; 11 Grotta del Mezzogiorno; 12 Paglicci; 13 El Castillo; 14 Rascaño; 15 Domingo Garcia; 16 Foz doCôa; 17 Parpalló; 18 Nerja; 19 Les Eglises; 20 Niaux; 21 Isturitz; 22 Pair-non-Pair; 23 Roc-de-Sers.


Gämse (Rupicapra rupicapra)
E. – Chamois; F. – Chamois (für die Alpen), Isard (für die Pyrenäen); S. – Rebeco, Sarrio oder Gamuza;Kat. – Isard; P. – Camurça; I. – Camoscio.Ordnung: Artiodactyla Owen 1848; Familie: Bovidae Gray 1821; Gattung: Rupicapra Blainville 1816; Art:Rupicapra rupicapra (Linnaeus 1758).Unterarten: Rupicapra rupicapra rupicapra Linnaeus 1758, R. r. pyrenaica Bonaparte 1845, R. r. parva Cabrera1911, R. r. cartusiana Couturier 1938, R. r. ornata Neumann 1899. Vorwiegend verwendete Literatur: Sacchi 1993a; Sägesser und Krapp 1986.
Habitat: Gämsen leben im Hochgebirge bis 3500 m.Sie kommen meistens oberhalb der Waldgrenze, imWinter auch in geringeren Höhen (bis 800 m) vor.
Verbreitung: Sehr hohe, felsige Gebirgszüge inMittel- und Südeuropa sind ihre Heimat; R. r.rupicapra in den Alpen, R. r. pyrenaica in denPyrenäen, R. r. parva in den Picos d’Europa inKantabrien, R. r. ornata in den Abruzzen in Italien,R. r. cartusiana im Massif de la Chartreuse in Frank-reich (Abb. 69).
Körpermaße: Kopf-Rumpf-Länge 110 bis 140 cm;Schulterhöhe 70 bis 85 cm; Gewicht: 17 bis 50 kg.Weibchen ca. 10% leichter als Böcke. Die spanischenGämsen, R. r. parva und R. r. pyrenaica, sind kleinerund erreichen nur 35 kg.
Hörner: Bis ca. 30 cm lang. Bei Böcken sind dieHörner manchmal hakenförmiger und etwas weitervoneinander entfernt als bei den Weibchen. R. r.ornata zeigt feinere Hörner.
Paarungszeit: Oktober bis Dezember. Tragzeit: 6Monate. Setzzeit: Mai bis Juni, ein Jungtier, seltenerzwei.
Natürliche Lebenserwartung: 15 bis 20 Jahre. Nahrung: Gräser, Kräuter, Blätter, Triebe, Moose,
Flechten. Verhalten: Tagaktiv, Rudel im Herbst und Winter
mit bis zu über 100 Tieren.Klimatische Aussage: So wie es bei Capra ibex
und Capra pyrenaica erwähnt wurde, spielt eher derfelsige Untergrund als das Klima eine Rolle. Beiklimatischen Änderungen haben sie ihren Lebensraumauf höhere oder niedrigere Areale verschoben, so wiesie es in begrenztem Ausmaß jedes Jahr bei jahreszeit-lichen klimatischen Änderungen tun.
Darstellungen aus dem Jungpaläolithikum Es gibt nur wenige sichere Darstellungen vonGämsen, darunter sind sicher der Lochstab vonGourdan, auf dem sieben Gämsen abgebildet sind(Abb. 70, 1a und 1b) und die 19 „contours découpés“aus der Höhle Labastide (Abb. 70, 6; Gämsen nachThiault und Roy 1996, 245–246; Steinböcke nachGraziosi 1956 und Leroi-Gourhan 1965) hervorzu-
heben. Die Schwierigkeit der Bestimmung vonGämsendarstellungen liegt wahrscheinlich darin, dasssie nur durch die hakenförmigen Hörner zweifelsfreizu bestimmen sind. Bei einer Gravur aus Isturitz (Abb.70, 17), wo vom Tier nur die Hörner erhalten sind,reichen diese aus, um eine sichere Bestimmung zuermöglichen. In vielen Fällen jedoch, wo lediglicheine Bestimmung der Unterfamilie Caprinae möglichist, tendiert man zu einer Zuordnung der Darstellungzu Gunsten der Steinböcke. In manchen Fällen istsogar unklar, ob es sich um die Darstellung einesSteinbockes, einer Gämse oder einer Saiga-Antilopehandelt.
In der Wandkunst ist eine einzige gemalteDarstellung bekannt, und zwar in El Castillo inSpanien (Abb. 70, 9), alle anderen sind Gravierungen.Darunter seien hier die vier stilisierten Gravuren ausder Höhle Cosquer erwähnt (Abb. 70, 13–14), wo dieHörner zwar deutlich zu lang, jedoch in ihrer Formund Ausrichtung korrekt dargestellt sind.
Abb. 69 Heutige Verbreitung von Rupicapra rupica-pra in Europa (nach Mitchell-Jones et al. 1999, 406–409).

136 Verbreitung der großen Jagdfauna in Mittel und Westeuropa im oberen Jungpleistozän
1a 1b 2 3 4
5 6 7 8 9 10 11 12
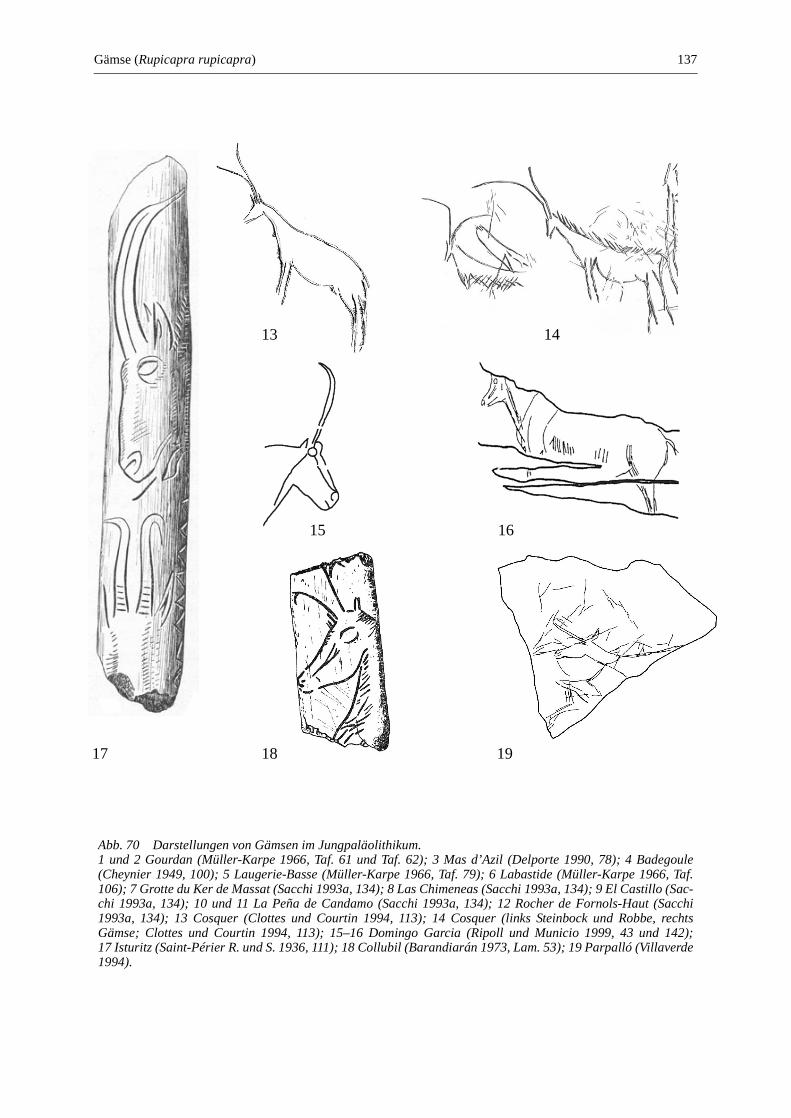
Gämse (Rupicapra rupicapra) 137
Abb. 70 Darstellungen von Gämsen im Jungpaläolithikum. 1 und 2 Gourdan (Müller-Karpe 1966, Taf. 61 und Taf. 62); 3 Mas d’Azil (Delporte 1990, 78); 4 Badegoule(Cheynier 1949, 100); 5 Laugerie-Basse (Müller-Karpe 1966, Taf. 79); 6 Labastide (Müller-Karpe 1966, Taf.106); 7 Grotte du Ker de Massat (Sacchi 1993a, 134); 8 Las Chimeneas (Sacchi 1993a, 134); 9 El Castillo (Sac-chi 1993a, 134); 10 und 11 La Peña de Candamo (Sacchi 1993a, 134); 12 Rocher de Fornols-Haut (Sacchi1993a, 134); 13 Cosquer (Clottes und Courtin 1994, 113); 14 Cosquer (links Steinbock und Robbe, rechtsGämse; Clottes und Courtin 1994, 113); 15–16 Domingo Garcia (Ripoll und Municio 1999, 43 und 142);17 Isturitz (Saint-Périer R. und S. 1936, 111); 18 Collubil (Barandiarán 1973, Lam. 53); 19 Parpalló (Villaverde1994).
13 14
15 16
17 18 19

138 Verbreitung der großen Jagdfauna in Mittel und Westeuropa im oberen Jungpleistozän
Reste aus dem Jungpaläolithikum
Die Gämse ist zwar in zahlreichen Fundstellenwährend des ganzen Jungpaläolithikums vertreten, sieist aber in der Regel kaum mehr als eine Rander-scheinung (Abb. 70), so wie beispielsweise inAndernach und in der Brillenhöhle in Deutschland, inLe Flageolet (Delpech et al. 2000) und in Laugerie-Haute (Delpech 1983) in der Dordogne in Frankreich,in Arbreda (Canal und Carbonell 1989) im NordostenSpaniens, in der Grotta Polesini (Sala 1985) undGrotta della Cala in Italien (Benini et al. 1997). Nurdort, wo die Landschaft den Bedürfnissen dieser Artbesser entspricht, steigt die Zahl der Gämsen. Das istder Fall in der Magdalénien-Schicht von Abauntz imspanischen Baskenland (ca. 31%), in Caldeirao inPortugal (ca. 28%), in der Magdalénien-Schicht ausder Grabung 1958 von La Vache (ca. 7,5%) in Frank-reich und in der Epigravettien-Schicht in RiparoTagliente (ca. 14,9%) und Grotta di Fumane (ca. 31%)sowie in den Gravettienschichten der Höhle Paglicci(ca. 16% und 33%) in Italien. Besonders erwäh-nenswert sind jedoch die Höhlen von Amalda, ElBuxú und Ekain. In Amalda erreicht die Gämse 63,6%der MIZ im Solutréen und 69,4% der MIZ imGravettien. In El Buxú sind es 50% der MIZ imSolutréen. In Ekain konnten 61,5% der gezähltenIndividuen in der Schicht 9b den Gämsen zugerechnetwerden. Interessant ist weiterhin zu erwähnen, dassauch andere baskische und kantabrische Fundstelleneinen Anteil an Gämsen zwischen 40% und 50%aufweisen, u. a. Lezetxiki (Aurignacien undGravettien) und Piélago II, 1 (Azilien).
Die Gämse stellt die relative Mehrheit derKnochenreste oder der MIZ auch in Fundstellen miteinem hohen Anteil an Equus ferus (u. a. in Abauntzund in der Schicht 22a von Paglicci), an Cervuselaphus (Riparo Soman, Dryas II und Alleröd) oder anCapra ibex (Riparo Soman, Dryas III). Die typischenBegleiter von Rupicapra rupicapra sind einerseits
Capra ibex aufgrund der ähnlichen Umweltan-sprüche, andererseits Cervus elaphus, der inSüdeuropa annährend in jeder Schicht vorhanden ist.
Anhand der Distanz zwischen den Hörnern konnteJesús Altuna zeigen, dass die Gämsen in den Pyrenäender Unterart Rupicapra rupicapra pyrenaica ange-hörten (Altuna 1972, 408).
Diskussion
Obwohl die Gämse in fast ganz Europa verbreitet war,ist sowohl bei den Knochenresten als auch bei denDarstellungen ein Schwerpunkt in den Pyrenäen undin Kantabrien zu sehen (Abb. 71). Dies ist ein gutesBeispiel dafür, dass, obwohl bei einzelnen Fundstellenmehrfach nachgewiesen wurde, dass die Anzahl derdargestellten Tiere oft diametral zur Anzahl derKnochenreste steht (u. a. Delporte 1984b; Altuna1994, 1996), im überregionalen Vergleich eine Korre-lation zwischen dargestellter und gejagter Faunadennoch möglich ist.
Ihre Seltenheit liegt zumindest teilweise daran,dass die Verbreitung der Gämsen auf sehr hohen,felsigen Bergen jenseits der üblichen Jagdgebiete derMenschen lag. Schließlich ist es interessant zuerwähnen, dass Rupicapra rupicapra weder in Nerja,noch in Malletes oder Parpalló unter der Faunaerwähnt ist. In Nerja, wo Darstellungen von Stein-böcken bekannt sind, erscheint sie auch nicht unterden Bildern. In Parpalló scheint eine Gämse auf einereinzigen Gravierung dargestellt zu sein. Es sieht soaus, als ob die Gämse in Südspanien nicht gejagtwurde oder nicht einmal vorhanden war. Vergleichtman die heutige Verbreitung von Rupicapra rupicapramit der heutigen Verbreitung von Capra pyrenaica,sieht man, dass Rupicapra rupicapra auch heute inSpanien nicht so weit südlich wie Capra pyrenaicavorkommt. Möglicherweise besetzt in Südspanien dieCapra pyrenaica, die in dieser Region etwas kleinerist, die ökologische Nische, die anderswo von derGämse belegt ist.

Gämse (Rupicapra rupicapra) 139
Abb. 71 Verbreitung von Rupicapra rupicapra sowie in diesem Kapitel erwähnte paläolithische Fundstellenmit Resten (Dreieck) oder Darstellungen (Kreis) dieser Tiere. 1 Badegoule; 2 Laugerie-Haute und Laugerie-Basse; 3 Cosquer; 4 Isturitz; 5 Gourdan; 6 Labastide; 7 Grotte du Ker de Massat; 8 Mas d’Azil; 9 La Vache;10 Rocher de Fornols-Haut; 11 La Peña del Candamo; 12 El Buxú; 13 Collubil; 14 El Castillo und Las Chime-neas; 15 El Pielago; 16 Amalda; 17 Lezetxiki; 18 Ekain; 19 Abauntz; 20 Domingo Garcia; 21 Arbreda;22 Caldeirao; 23 Gruta das Salemas und Pego do Diabo; 24 Parpalló; 25 Grotta di Fumane; 26 Riparo Tagli-ente; 27 Polesini; 28 Paglicci; 29 La Cala; 30 Brillenhöhle; 31 Andernach.


Saiga-Antilope (Saiga tatarica)
E. – Saiga; F. – Saiga; S. – Saiga; Kat. – Saiga; P. – Antílope saiga I. – Antilope saiga Ordnung: Artiodactyla Owen 1848; Familie: Bovidae Gray 1821; Gattung: Saiga Gray 1843; Art: Saiga tatarica(Linnaeus 1766).Unterarten: S. t. tatarica (Linnaeus 1766) hauptsächlich in den Steppen im Süden der ehemaligen UdSSR,S. t. mongolica (Bannikov 1946) in den Senken der großen Seen in der Nordwestmongolei. Die Reste aus denwesteuropäischen Fundstellen der letzten Eiszeit werden der Unterart Saiga tatarica borealis (Čserskij 1876)zugerechnet.Vorwiegend verwendete Literatur: Bannikov 1963; Dubourg et al. 1994; Heptner et al. 1966, 570–602; Kahlke1992; Nougier und Robert 1958.
Habitat: Die Saiga-Antilope lebt in Salz-, Sand- undLehmsteppen bis 1600 m Höhe. Sie meidetentschieden alle unebenen Regionen, nicht nurGebirge, sondern auch einigermaßen zerfurchtesGelände. Sie sucht im Winter während der Schnee-stürme Schutz in Dickichten an Fluss- und Seeufern.Die Saiga-Antilopen meiden weiterhin auch Eis undKrustenschnee, da sie sonst kein Futter erreichenkönnen und sich noch dazu auf solchen FlächenVerletzungen zufügen könnten.
Verbreitung: Zum Beginn der 1930er Jahreschätzte man den Bestand der Saiga-Antilopen aufkaum mehr als 1000 Individuen. Ihr Areal war aufeinzelne begrenzte Steppengebiete beschränkt. Durchden konsequenten Schutz dieser Art stieg ihre Zahl1958 wieder auf ca. 2.000.000 Individuen (Heptner etal. 1966, 579–580); ihr Areal erstreckte sich von derUkraine bis in die Mongolei (Abb. 72). Bis zum Jahr2001 sind die Bestände jedoch wegen Wilderei wiederauf wenige zehntausend Individuen zurückgegangen(Reading 2001).
Fossile Reste von Saiga-Antilopen zeigen, dass ihrVerbreitungsgebiet in prähistorischen Zeiten deutlichgrößer war. Funde stammen nicht nur aus Westeuropa,sondern auch aus Nordgriechenland (Kahlke 1975),Nordsibirien (Mündung des Flusses Lena und InselBolschoi Ljachow; Bannikov 1963, 55f.) sowie ausAlaska und Kanada (Kahlke 1975).
Körpermaße: bei S. t. tatarica Körperlänge beiBöcken 123 bis 146 cm, Geißen 108–125 cm; Schul-terhöhe bei Böcken 60–73 cm, Geißen 57–69 cm;Gewicht S. t. tatarica, Böcke 32–51 kg, Geißen 21,5–41 kg. S. t. mongolica unterscheidet sich von der S. t.tatarica durch kleinere Gesamtmaße.
Kopfbeschreibung: Ein kurzer, beweglicher Rüsselcharakterisiert den Kopf dieses Tieres und beweisteine besondere Anpassung an seine Umwelt. Diegroßen Nasenhöhlen erwärmen die Luft im Winter,bevor diese in die Lungen gelangt, im Sommerdagegen kühlen und befeuchten sie die Luft.
Weiterhin filtert dieser Atmungsapparat den in aridenund semiariden Regionen reichlich vorhandenenStaub.
Fell: Die Haardecke ist von gleichmäßiger Länge(bis 3 cm im Sommer, bis 7 cm im Winter). Im Winterbildet sich am Hals eine kurze Mähne (bis 15 cmLänge). An der Schnauze und unter den Augen bildetsich der so genannte „Backenbart“.
Die Hörner bestehen aus vielen deutlichen Ringenund sind bis 30 cm lang. Weibliche Tiere besitzenmeistens keine Hörner.
Druck auf den Boden: ca. 480 g/cm²Paarungszeit: November/Dezember. Tragzeit: 4 bis
5 Monate. Setzzeit: April, ein bis drei Jungtiere,meistens zwei.
Abb. 72 Historisches Verbreitungsgebiet von Saigatatarica (nach: Bannikov 1963, 54).

142 Verbreitung der großen Jagdfauna in Mittel und Westeuropa im oberen Jungpleistozän
Abb. 73 Als Saiga-Antilope publizierte Darstellungen. 1 Bruniquel (Bétirac 1952, 226); 2 Bize-Minervois(Dubourg et al. 1994, 258); 3 Gönnersdorf (Dubourg et al. 1994, 258); 4 La Vache (Nougier und Robert 1958,15); 5 Peyrat (Dubourg et al. 1994, 258); 6 Gourdan (Nougier und Robert 1958, 23); 7 Enlène (Bégouën et al.1987, 14); 8 Mas d’Azil (Nougier und Robert 1958, 23); 9 Abri de la Souquette (Dubourg et al. 1994, 256); 10Altxerri (Sacchi 1993a, 136); 11 Les Combarelles (Nougier und Robert 1958); 12 Rouffignac (Sacchi 1993a,136).
1 2 3 4 5 6 7 8 9 10 11 12

Saiga-Antilope (Saiga tatarica) 143
Natürliche Lebenserwartung: Bestenfalls 11 Jahre. InGebieten mit zahlreichen Wölfen erreichen die Böckenicht mehr als 5 Jahre.
Nahrung: Wenn möglich sind sie sehr wählerisch,haben aber sonst ein sehr breites Spektrum anNahrungspflanzen. Insgesamt gehören bis zu 120unterschiedliche Pflanzen dazu, wobei in jeder Regionnur eine kleinere Anzahl überhaupt zur Auswahl steht,und diese auch nicht während des ganzen Jahres. ImSommer werden Pflanzen mit hohem Wassergehaltbevorzugt.
Verhalten: Sowohl tag- als auch nachtaktiv. Beistarker Hitze im Sommer ist sie nur dämmerungs- undnachtaktiv. Die Saiga tatarica tatarica ist ständig inBewegung. Ihre bevorzugte Gangart ist der Passgang,in dem sie gleichzeitig beide Beine einer Seite hebt.Dies erlaubt für kurze Zeit Geschwindigkeiten von biszu 80 km pro Stunde, erschwert jedoch das Springen,so dass alle auch nur unbedeutenden Grubenvermieden werden (Bannikov 1963, 99). Es wurdenTagesstrecken bis 120 km beobachtet (Heptner et al.1966, 586). Einzelne Wanderungen betragen jedochmaximal 350 km (Bannikov 1963, 68). Ihr Wandernhängt eher mit dem Nahrungs- und Wasserangebot alsmit klaren Wanderungsrouten zusammen. So sprichtman von saisonalen, aber auch von aperiodischenWanderungen (Bannikov 1963, 65–76). In Kasachstanwandern die Herden im Frühling nach Norden, imHerbst nach Süden. In schneearmen Wintern undfeuchten Sommern bleiben die Saiga-Antilopen odermindestens ein Teil der Population jedoch im gleichenAreal. Bei Wanderungen betragen die Ansammlungenvon Saiga-Antilopen bei extremen klimatischenBedingungen bis zu 200.000 Individuen, dabeiwimmelt es von Saiga-Antilopen soweit das Augereicht. Die Tiere der Unterart S. t. mongolica in derMongolei wandern nicht.
Saiga-Antilopen sind gute Schwimmer, beiWanderungen werden Ströme wie die Wolga oderFlussarme von mehreren Dutzend Metern Breiteüberquert.
Ihr Sehvermögen ist sehr gut; Gefahren können bisauf 1 km Distanz bemerkt werden. Gut ist weiterhinauch ihr Geruchsvermögen, während das Gehörschlecht zu sein scheint (Heptner et al. 1966, 587).
Erhebliche Probleme beim Laufen sowie bei derNahrungssuche bekommen die Saiga-Antilopen abeiner Schneetiefe von 25 cm, u. a. weil ihr Trittsiegelklein ist und eine hohe Gewichtsbelastung ertragenmuss (viel höher als z. B. beim Wolf, ihrem Haupt-feind).
Aufgrund des unentwegten Umherziehens verbrau-chen Saiga-Antilopen ständig Energie, andererseitsaber sind ihre Speicherkapazitäten aufgrund dergeringen Körpergröße begrenzt. Sommerliche Hitze-wellen und anhaltender Schneefall oder plötzlicherEisregen im Winter können die Herden dezimieren.
Klimatische Aussage: Saiga-Antilopen weisen ohneZweifel auf extrem trockene Klimaverhältnisse hin,andererseits jedoch darf man nicht vergessen, dass dieAnwesenheit oder Abwesenheit ihrer Feinde ebenfallseinen entscheidenden Einfluss auf ihre Populationausübt.
Darstellungen aus dem Jungpaläolithikum
Die Wandbilder aus dem Abri de la Souquette sowieaus den Höhlen von Altxerri, Les Combarelles undRouffignac sind Gravierungen (Abb. 73, 9–12), zumTeil von sehr grober Art. Dadurch sind sie nur alsmögliche oder wahrscheinliche, jedoch nicht alssichere Darstellungen von Saiga-Antilopen zubetrachten. Ganz anders sieht es bei den feinenGravierungen der Kleinkunst aus. Die Darstellungenvon Bize-Minervois, La Vache und Gourdan (Abb. 73,2, 4 und 6) zählen zu den schönsten Beispielen derMagdalénien-Kunst. Die Gravierung eines Hornes ausMas d’Azil (Abb. 73, 8) ist so typisch, dass sie,obwohl das ganze Tier fehlt, ausreicht, um eineBestimmung als Saiga-Antilope zu begründen.Umgekehrt muss mit den Gravierungen vonGönnersdorf (Abb. 73, 3) argumentiert werden. Hierist zwar das ganze Gesicht mit dem für Saiga-Antilopen üblichen Profil abgebildet, da aber dieHörner nicht dargestellt sind, könnte es sich ebenfallsum die Darstellung eines Elchs handeln, was verschie-denen Autoren bereits aufgefallen ist (Bosinski 1981;Koenigswald und Hahn 1981, 89; Dubourg 1997,Abb. 37).
Reste aus dem Jungpaläolithikum
Aus Spanien ist die Saiga tatarica nur aus Abauntzbekannt (Abb. 74). Der ältere Fund von El Pendo, dervon Arlette Leroi-Gourhan 1959 bestimmt und vonJesús Altuna zitiert wurde (1972, 59), wird imjüngeren Artikel von Jesús Altuna und Koro Mariez-kurrena (1996) nicht mehr erwähnt. Damit wird impli-ziert, dass der Fund von El Pendo von den beidenAutoren als nicht vertretbar eingestuft worden ist. InItalien gibt es keine Reste dieser Art. In Frankreich istihre Verbreitung sehr detailliert untersucht worden, sodass man anhand der 14C-Datierungen die Präsenz derSaiga-Antilope in Westeuropa als Ergebnis einerEinwanderung um ca. 18.500 BP charakterisierenkann. Die Saiga-Antilopen sind in Südwestfrankreichbis zum Beginn des Bölling-Interstadials (ca. 13.300BP) vorhanden60 (Delpech 1989). In den französi-schen Pyrenäen sind sie bis ca. 12.500 BP nachge-wiesen (La Vache und vielleicht Grotte de Bize;Álvarez und Jöris 1998, 76). In Südostfrankreich sindsie im Abri Cornille in der Dryas II nachgewiesen(Escalon de Fonton et al. 1979, 278).

144 Verbreitung der großen Jagdfauna in Mittel und Westeuropa im oberen Jungpleistozän
In Deutschland ist die Saiga-Antilope schon imGravettien der Brillenhöhle, obwohl nur durch eineneinzelnen Rest, nachgewiesen. Sie ist weiterhin inmehreren Schichten des Magdalénien der Teufels-brücke sowie im Magdalénien der Kniegrotte und ausGönnersdorf nachgewiesen.
In Belgien ist sie sowohl in Fonds-de-Forêt alsauch im Trou du Renard (Otte 1979, 101) bestimmtworden. Diese letzte Fundstelle ist auf ca. 24.000Jahre BP datiert (Otte 1979, 103).
Dort, wo die Saiga-Antilope dominiert, wie z. B. inMoulin-Neuf und Saint-Germain de la Rivière, wirdsie von Tierarten der offenen Landschaft begleitet, wieRentier, Steppenbison oder Pferd. Rothirsche undandere Waldtiere sind dort, wo die Saiga-Antilopeauftritt, immer selten, außer in Abauntz.
Diskussion
Da Saiga-Antilopen Gebirge strikt meiden, sind diewenigen Reste aus Abauntz, einer Fundstelle in denspanischen Pyrenäen, wenige Kilometer von derGrenze zwischen Spanien und Frankreich entfernt, mitgroßer Wahrscheinlichkeit als Import aus den flachenEbenen Frankreichs zu betrachten. Diese Reste sindaußer mit dem Pferd (25,1%) auch mit einem hohenAnteil an Gämse (34,4%), Steinbock (15,6%) undRothirsch (17,0%) vergesellschaftet, was für Saiga-Antilopen nicht üblich ist. Die Fundstelle Abauntzkann daher nicht als Hinweis darauf betrachtet
werden, dass die Saiga-Antilopen je die IberischeHalbinsel erreicht hätten61. Inwieweit weiterhin das„Saiga-Phänomen“ als einheitlich betrachtet werdenkann, ist unklar. Aufgrund der besonderen Geschwin-digkeit, mit der diese Art (wohl mit Hilfe desMenschen) ihren alten Lebensraum in kürzester Zeitwieder erobert hat (von 1930 bis 1958 war ihre Zahlvon 1.000 auf über 2.000.000 Individuen gestiegen),aufgrund ihres kurzen Generationswechsels(Weibchen sind mit 8 Monaten geschlechtsreif) undihres maximalen Ausbreitungsareals während derletzten Eiszeit (von der Atlantikküste Europas überganz Eurasien bis nach Alaska und Nordkanada;Harington 1980, 215, Abb. 11), kann man nichtausschließen, dass es unter Umständen mehrereEinwanderungswellen dieser Art gab. Die wiederholtePräsenz dieser Art in Mitteleuropa könnte dafürsprechen. Leider sind jedoch viele der Fundstellen inMittel- und Osteuropa paläontologischer Natur(Kahlke 1975), so dass ihre grobe Datierung vieleFragen offen lässt.
Der anscheinende Widerspruch zwischenFundstellen, wo die Saiga-Antilope über 50% derFauna darstellt (Moulin-Neuf und Saint-Germain de laRivière), und die Seltenheit der Darstellungen dieserArt könnte ein Hinweis dafür sein, dass Saiga-Antilopen sowohl regional als auch zeitlich in ihremBestand sehr großen Schwankungen unterworfenwaren.
60 Paul Bahn, der sich auf eine Publikation von 1922 von René de Saint-Périer bezieht, zitiert zwei Fragmentevon Saiga-Antilopen für die Aurignacien/Gravettien-Schichten der Rideaux-Höhle im Komplex der Grottes deLespugue (Bahn 1983, Tab. 27). Es würde sich mit Sicherheit lohnen, diese Reste direkt zu datieren.61 Anderer Meinung sind Esteban Álvarez Fernández und Olaf Jöris, die eine lange Präsenz von Saiga-Antilopenauf der Iberischen Halbinsel annehmen (Álvarez und Jöris 1998).

Saiga-Antilope (Saiga tatarica) 145
Abb. 74 Verbreitung von Saiga tatarica sowie in diesem Kapitel erwähnte paläolithische Fundstellen mitResten (Dreieck) oder Darstellungen (Kreis) dieser Tiere. 1 Soldier’s Hole und Gough’s Cave; 2 Sun Hole;3 Trou du Renard; 4 Fonds-de-Forêt; 5 Gönnersdorf; 6 Kniegrotte und Teufelsbrücke; 7 Brillenhöhle; 8 Durif àEnval; 9 Coude; 10 Combe Saunière, Le Flageolet, Laugerie-Haute; 11 Combe Cullier; 12 Saint-Germain de laRivière und Fongaban; 13 Moulin Neuf; 14 Dufaure; 15 Isturitz; 16 Abauntz; 17 Cornille; 18 Peyrat und Abride la Souquette; 19 Les Combarelles und Rouffignac; 20 Altxerri; 21 Gourdan; 22 Mas d’Azil; 23 Enlène undLa Vache; 24 Bruniquel; 25 Bize.


Die große Fauna an den Küsten Europas
Mit den Robben wird die erste von drei Tiergruppenangesprochen (Seehunde und Robben, Riesenalke undWaltiere), die eng mit dem Meer verbunden sind. DieAnzahl an Fundstellen, in denen Reste oder Darstel-lungen dieser Tierarten entdeckt wurden, ist sehrbegrenzt. Man darf aber nicht vergessen, dass dasMeer während der letzten Eiszeit zeitweilig bis zu140 m tiefer als heute lag (Aksu et al. 1995, 96–97,Abb. 13; Sacchi 1986, Abb. 199; Verhart 1995, 291;Zilhão 1997, 53). Daraus folgt, dass der eiszeitlicheKüstenlandstrich sich aufgrund seiner heutigen Lageunter dem Meeresspiegel der direkten archäologi-schen Forschung entzieht. Die Untersuchung derwenige Reste und der Darstellungen solcher
Tiergruppen wie Seehunde und Robben, Riesenalkeund Waltiere, die ohne Zweifel bevorzugt an derKüste gesehen, gejagt und verwertet werden konnten,ist als stellvertretend für eine deutlich größere Anzahlan solchen Funden und solchen Stationen zu sehenund können uns wichtige Hinweise über diesenBereich liefern.
Die Höhle von Cosquer, deren Eingang 60 m unterWasser liegt, gilt hier als das beste Beispiel. Durchihre acht gravierten Robbendarstellungen (Abb. 82,21) und drei gemalten Riesenalke (Abb. 87, 1) zeigtsie, wie viel unter dem Meeresspiegel verlorengegangen ist.
Seehunde (Phocidae)
Vorwiegend verwendete Literatur: Cleyet-Merle 1990; Duguy und Robineau 1992; Serangeli 2003; Sonneville-Bordes und Laurent 1983; Wandrey 1997.
Körperbeschreibung: Seehunde und Robben gehörenzur Familie der Wasserraubtiere. Ihre Gestalt istspindelförmig, die Gliedmaßen sind flach, Finger undZehen wachsen zusammen in Form von Flossen(daher der Name der Pinnipedia, vom Lateinischenpinna = Flosse und pes = Fuß). Die hinteren Glied-maßen stehen in einer Achse mit dem Rest desKörpers und sind das Hauptantriebssystem im Wasser.Die vorderen Extremitäten werden beim Schwimmenmeistens am Körper angelegt und dienen eher zurRichtungsänderung als zur Fortbewegung. Da dieBewegung des Fußgelenkes sehr stark eingeschränktist, können Seehunde (Phocidae) auf dem Land fastnur die Vorderextremitäten benutzen. Ganz anderssieht es bei den verwandten Ohrenrobben (Otariidae)aus dem Pazifischen Ozean aus, die man aus Zoos undZirkussen kennt. Diese können die hinteren Flossennach vorne drehen und sich mit allen vier Gliedmaßenan Land geschickt fortbewegen.
Der Körper ist von zwei Arten von Haarenbedeckt: Deckhaaren und feineren Wollhaaren. DieHaare sind durch ein Sekret aus Talgdrüsen einge-fettet und damit wasserundurchlässig.
An den Seiten der Schnauze wachsen Vibrissen,die sich in Anzahl und Länge von Art zu Art unter-scheiden. Einige Tasthaare befinden sich auch überden Augen.
Die Anpassung ans Wasser ist so weit fortgeschritten,dass Seehunde keine Ohrmuscheln besitzen.Obwohl der größte Teil dieser Tiere im Meer lebt, gibtes Populationen von Ringelrobben in Europa (Phocahispida saimensis und Phoca hispida ladogensis)sowie Seehunde in Nordamerika (Phoca vitulina) unddie Baikalrobbe in Asien (Phoca sibirica), die dasganze Jahr in Süßwasserseen leben.
Vereinzelte Seehunde (Phoca vitulina), Kegel-robben (Halichoerus grypus), sowie andere nichtnäher bestimmbare Robben sind in FrankreichsFlüssen im Sommer gelegentlich beobachtet worden,bis zu 100 km vom Meer entfernt (Sonneville-Bordesund Laurent 1983, 72). Ein trächtiges Individuum derArt Phoca groenlandica wurde im März 1856 in derElbe nahe Dessau (500 km von der Elbmündungentfernt) gefangen (Kapel 1992, 204). Gruppen vonKaspischen Robben (Phoca caspica) wandern regel-mäßig vom Kaspischen Meer in den Fluss Ural oderentlang der Wolga, manchmal bis Wolgograd ca.300 km landeinwärts (Krylov 1992, 166). Auch dieSeehunde vom Baikalsee sind wahrscheinlich imQuartär vom mehr als 1500 km entfernten Nordpo-larmeer entlang des Jenissei-Angara-Flusssystems inden Baikalsee eingewandert (Zemsky 1992, 188).
Es zeigt sich deutlich, dass das Eindringen vonRobben in Flusssysteme, das oft mit dem Begriff„Verirren“ umschrieben wird, aufgrund der Regel-

148 Verbreitung der großen Jagdfauna in Mittel und Westeuropa im oberen Jungpleistozän
mäßigkeit solcher Einwanderungsphänomene nichtweiter als „Fehler“ interpretiert werden darf. EinSeehund ist in erster Linie ein Säugetier, daher ist esnicht besonders relevant für seine Atemfunktionen, ober im Süß- oder Salzwasser lebt. Die in Süßwasserseenlebenden Unterarten von Phoca hispida (Phocahispida saimensis und Phoca hispida ladogensis) gabes in der Eiszeit sicher noch nicht, da die Seen, indenen sie heute leben, in der Eiszeit nicht existiertenund deren Gebiet unter mehreren hundert Metern Eislag. Es sind jedoch Reste von Robben aus der ganzenGeschichte der Ostsee bekannt, auch aus der Zeit desAncylus-Sees, ca. 9.400 bis 8.200 BP (Clark 1946,39), als die Ostsee ein Süßwassersee war. Weiterhinist der Seehund ein außerordentlich intelligenter undneugieriger Fleischfresser; unter diesem Aspekt stehter dem Menschen besonders nah. Die Wanderungenvon einzelnen Individuen sind möglicherweise sogarsehr wichtig für die Überlebenschancen ihrer Gruppe,weil dadurch neue Lebensräume erschlossen werdenkönnen. Wenn eine Robbe auf der Suche nachNahrung in den im Vergleich zu heute deutlichbreiteren Flussbetten des Rheins, der Elbe oder derDordogne hochgeschwommen ist, hat sie sich kaumverirrt, sondern ist nur Wanderfischen gefolgt.
Am Ufer sind Seehunde und Robben, aufgrundihres Körperbaus, für den Menschen eine leichte, bis300 kg wiegende Beute. Sie liefern noch heute nichtnur Fleisch, sondern auch Haut und Fett. Die Haut istwasserdicht und eignet sich hervorragend für dieHerstellung sowohl von Kleidung als auch von Lederfür Zelte, Taschen, Schnüre und Decken (Bogoras1909, insbesondere 234–263). Der Anteil an Fett istbei Robben um ein Vielfaches größer als bei anderenTieren und es kann, je nach Jahreszeit und Alter desTieres, bei Ringelrobben bis zur Hälfte des Körper-gewichts ausmachen.
Aufgrund der geringen Zahl an Funden und dergroßen Ähnlichkeiten sowohl im Körperbau wie auchim Verhalten der Robben werden diese hier als eineGruppe betrachtet, und nur die spezifischen Merkmaleder unterschiedlichen Tierarten werden vereinzeltangesprochen.
So wird auch die Behandlung der Darstellungensowie der Knochenreste am Ende des Kapitels statt-finden.
Mittelmeer-Mönchsrobbe (Monachus monachus)
E. – Monk seal; F. – Phoque moine; S. – Foca monje,Foca fraile; Kat. – Foca mediterranea, Vedell marí;P. – Lobo-marinho; I. – Foca monaca.
Ordnung: Carnivora Bowdich 1821; Familie:Phocidae Gray 1821; Gattung: Monachus Flemming1822; Art: Monachus monachus (Hermann 1779).
Habitat: Mönchsrobben leben zur Zeit fast nur anfelsigen Küsten. Früher waren sie auch an Sand-stränden vorhanden (Kolonien von bis zu 5000Individuen).
Heutige Verbreitung: Monachus monachus kommtvereinzelt im Mittelmeer, im Schwarzen Meer und ander nordafrikanischen Atlantikküste vor (Abb. 75).
Körpermaße: Länge Männchen bis 380 cm,Weibchen 290 cm; Gewicht bis ca. 320 kg.
Verhalten: Die Vorderflossen der Mönchsrobbensind mit Krallen versehen. Damit können diese Tiere,anders als z. B. die Seehunde, auch an klippenreichenFelsküsten das Wasser verlassen. Sie leben oft inHöhlen. Aufgrund der geringen Zahl der Tiere ist eswahrscheinlich, dass ihr heutiges Verhalten in vielenAspekten anders als ihr ursprüngliches ist.
Paarungszeit: Hauptsächlich Sommer und Herbst.Tragzeit: 11 Monate. Setzzeit: Sommer und Herbst.
Natürliche Lebenserwartung: 30 bis 40 Jahre.Nahrung: Fische und andere Meerestiere.
Abb. 75 Heutige Verbreitung von Monachus mona-chus (nach Wandrey 1997, 220).

Seehunde (Phocidae) 149
Seehund (Phoca vitulina)
E. – Harbour Seal, Sand Seal; F. – Phoque veau-marin; S. – Foca comun, Lobo marino; P. – Focapeixe-homem, Foca-vitulina; I – Foca vitulina.Ordnung: Carnivora, Bowdich 1821; Familie:Phocidae Gray 1821; Gattung: Phoca Linnaeus 1758;Art: Phoca vitulina Linnaeus 1758.
Habitat: Seehunde leben an Meeresküsten undFlussmündungen mit Sandbänken und seichtemWasser. Selten kommen sie an Steilküsten vor.
Verbreitung: Ihr Lebensraum ist die KüsteNordamerikas und die atlantische Küste Europas. Inder Hudson Bay bewohnen sie Süßwasserseen dasganze Jahr über (Abb. 76).
Körpermaße: Länge Männchen 130 bis 195 cm,Weibchen 120 bis 169 cm; Gewicht Männchen100 kg (max. 200 kg), Weibchen 45 bis 100 kg.
Verhalten: Phoca vitulina ist meist ortstreu. Paarungszeit: Mai bis Juni. Tragzeit: 10 ½ – 11
Monate (Verzögerung der Entwicklung des Embryosca. 2½ Monate). Setzzeit: Mai – Juni.
Natürliche Lebenserwartung: 38 Jahre beimännlichen, 25 bei weiblichen Individuen.
Nahrung: Fische, Tintenfische, junge Krabben.
Ringelrobbe (Phoca hispida)
E. – Ringed Seal; F. – Phoque marbré; S. – Focaocelada; I. – Foca dagli anelli. Ordnung: Carnivora Bowdich 1821; Familie: PhocidaeGray 1821; Gattung: Phoca Linnaeus 1758; Art:Phoca hispida Schreber 1775.
Habitat: Phoca hispida lebt im Küsteneis, an Küstenund Fjorden. Zwei Unterarten leben in Süßwasserseen(P. h. saimensis und P. h. ladogensis).
Verbreitungsgebiet: Vorwiegend arktisch. Eserstreckt sich allerdings bis in die Nord- und Ostsee.Eine Unterart lebt in Finnland im Saimasee, eineandere in Russland im Ladogasee (Abb. 77).
Verhalten: Ringelrobben sind besonders ortstreu.Sie leben am Rande der Eisgrenze. Im Winter müssensie sich im Eis Atemlöcher frei halten. Im Wintergraben sie sich weiterhin eine bis zu 10 m lange Höhleim Schnee, wo die Jungen zur Welt kommen.
Körpermaße: Länge 110–157 cm; Gewicht: 36 bis110 kg.
Weitere Merkmale: Sie besitzen unter der Hauteine im Vergleich zu Seehunden viel dickereFettschicht, die es ihnen erlaubt, tief ins Eismeervorzudringen. Fett kann bis zur Hälfte ihres Körperge-wichtes ausmachen.
Paarungszeit: Frühjahr. Tragzeit: 10½–11 Monate(die Entwicklung des Embryos ist um 3 ½ Monateverzögert). Setzzeit: Frühjahr.
Natürliche Lebenserwartung: bis 45 Jahre, nur 15–16% erreichen das 10., nur 6–8% das 20. Lebensjahr.
Nahrung: Fische, Tintenfische und Wirbellose.
Abb. 76 Heutige Verbreitung von Phoca vitulina(nach Wandrey 1997, 238).
Abb. 77 Heutige Verbreitung von Phoca hispida(nach Wandrey 1997, 240).

150 Verbreitung der großen Jagdfauna in Mittel und Westeuropa im oberen Jungpleistozän
Sattelrobbe (Phoca groenlandica)
E. – Harp Seal, Groenland Seal, Saddle-Back;F. – Phoque du Groenland; S. – Foca pía; I. – Focadella Groenlandia.
Ordnung: Carnivora Bowdich 1821; Familie:Phocidae Gray 1821; Gattung: Phoca Linnaeus 1758;Art: Phoca groenlandica Erxleben 1777.
Habitat: Die Sattelrobbe lebt im Treibeis.Verbreitung: Arktisch (Abb. 78). Körpermaße: Länge 155–200 cm, im Durchschnitt
170 cm sowohl für Weibchen als auch für Männchen;Gewicht 115 bis 190 kg.
Verhalten: Sie wandert im Oktober zu denWurfplätzen und im Mai nordwärts zurück. Sie lebtviel mehr im offenen Meer als die anderen Robben-arten.
Paarungszeit: März–April. Tragzeit: ca. 11–11 ½Monate (mit einer Verzögerung der Entwicklung desEmbryos von 3 ½–4 Monaten). Setzzeit: EndeFebruar–März.
Natürliche Lebenserwartung: 30–40 JahreNahrung: Fische, insbesondere Lodden, Heringe
und Dorsche, Krebse und andere Wirbellose.
.
Kegelrobbe (Halichoerus grypus)
E. – Grey seal; F. – Phoque gris; S. – Foca gris;I. – Foca grigia; P. – Foca-cinzenta.
Ordnung: Carnivora Bowdich 1821; Familie:Phocidae Gray 1821; Gattung: Halichoerus Nillson1820; Art: Halichoerus grypus (Fabricius 1791).
Habitat: Die Kegelrobbe lebt in Küstengewässern undbevorzugt felsige Ufer.
Verbreitung: Eine Population kommt im Westat-lantik, eine in der Ostsee und eine zwischen derBretagne, den Britischen Inseln, Island und Norwegenvor (Abb. 79).
Körpermaße: Länge bis 220 cm; Gewicht bis300 kg bei Männchen und bis 150 kg bei Weibchen.
Verhalten: Wenn sie in die Enge getrieben werden,greifen insbesondere männliche Tiere an und benutzendabei ihr kräftiges Gebiss.
Paarungszeit: Frühjahr. Tragzeit: 11½ Monate (dieEntwicklung des Embryos ist um 3½ Monateverzögert). Setzzeit: Frühjahr.
Natürliche Lebenserwartung: bis 40 Jahre.Nahrung: Fische, vor allem Lachse in Küstennähe.
Abb. 78 Heutige Verbreitung von Phoca groenlan-dica (nach Wandrey 1997, 245).
Abb. 79 Heutige Verbreitung von Halichoerus gry-pus (nach Wandrey 1997, 248).

Seehunde (Phocidae) 151
Klappmütze (Cystophora cristata)
E. – Hooded seal; F. – Phoque à crête, Phoque àcapuchon; S. – Foca de casco; P. – Foca de crista;I. – Cistofora crestata.
Ordnung: Carnivora Bowdich 1821; Familie:Phocidae Gray 1821; Gattung: Cystophora Nilsson1820; Art: Cystophora cristata (Erxleben 1777).
Habitat: Die Klappmütze lebt im offenen Meer, imTreibeis und in Küstennähe.
Verbreitungsgebiet: Arktis und Subarktis. Haupt-sächlich zwischen Island, Grönland und Kanada (Abb.80).
Körpermaße: Länge 250 cm (max. 283 cm) beiMännchen, 220 cm bei Weibchen; Gewicht 400 kg(max. 435 kg).
Paarungszeit: März bis Anfang April. Tragzeit:11½ Monate (mit einer Verzögerung der Entwicklungdes Embryos von 4 Monaten). Setzzeit: März bisAnfang April.
Natürliche Lebenserwartung: 30–35 Jahre.Nahrung: Fische in tiefen Gewässern.
Bartrobbe (Erignathus barbatus)
E. – Bearded seal; F.– Phoque à barbe, Phoque barbu;S. – Foca barbuda; P. – Foca-barbuda; I. – Focabarbuta.
Ordnung: Carnivora Bowdich 1821; Familie:Phocidae Gray 1821; Gattung: Erignathus Gill 1866;Art: Erignathus barbatus (Erxleben 1777).
Habitat: Die Bartrobbe lebt in der Arktis undbevorzugt flache Gewässer.
Verbreitungsgebiet: Nordpazifik, Nordatlantik,Polarmeer sowie die Nordküsten Amerikas, Asiensund Europas (Abb. 81).
Verhalten: Vorwiegend einzelgängerisch, eswerden jedoch auch lose Verbände in der Zeit vonApril bis Juni gebildet. Sie halten sich häufig imBereich des Treibeises auf. Sie gehen auch an Küstenan Land und schwimmen in Flüsse.
Körpermaße: Länge 225 cm; Gewicht ca. 230 kg.Paarungszeit: Mitte März bis Anfang April.
Tragzeit: 10½–11 Monate (mit einer Verzögerung derEntwicklung des Embryos von 2½–3 Monaten).Setzzeit: Februar und März.
Natürliche Lebenserwartung: 25 Jahre.Nahrung: Krebse, Weichtiere, Meeresringel-
würmer, Fische.
Abb. 80 Heutige Verbreitung von Cystophora cri-stata (nach Wandrey 1997, 236).
Abb. 81 Heutige Verbreitung von Erignathus barba-tus (nach Wandrey 1997, 234).
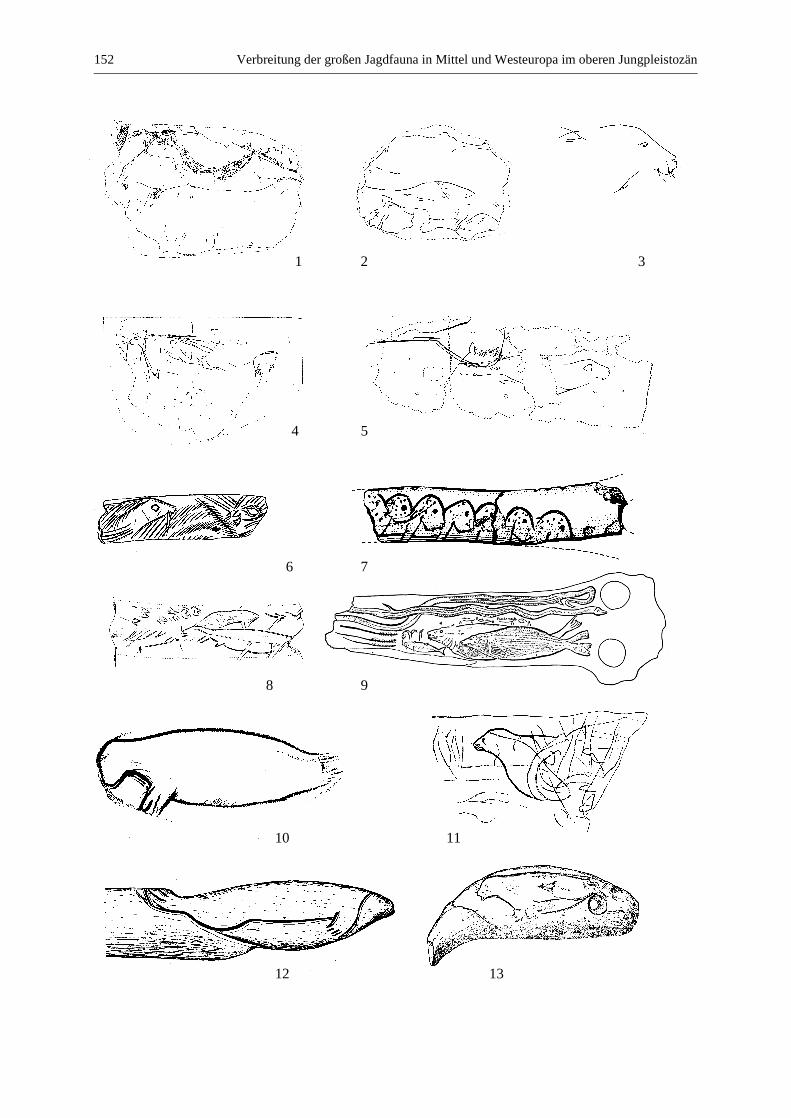
152 Verbreitung der großen Jagdfauna in Mittel und Westeuropa im oberen Jungpleistozän
1 2 3 4 5 6 7 8 9 10 11 12 13
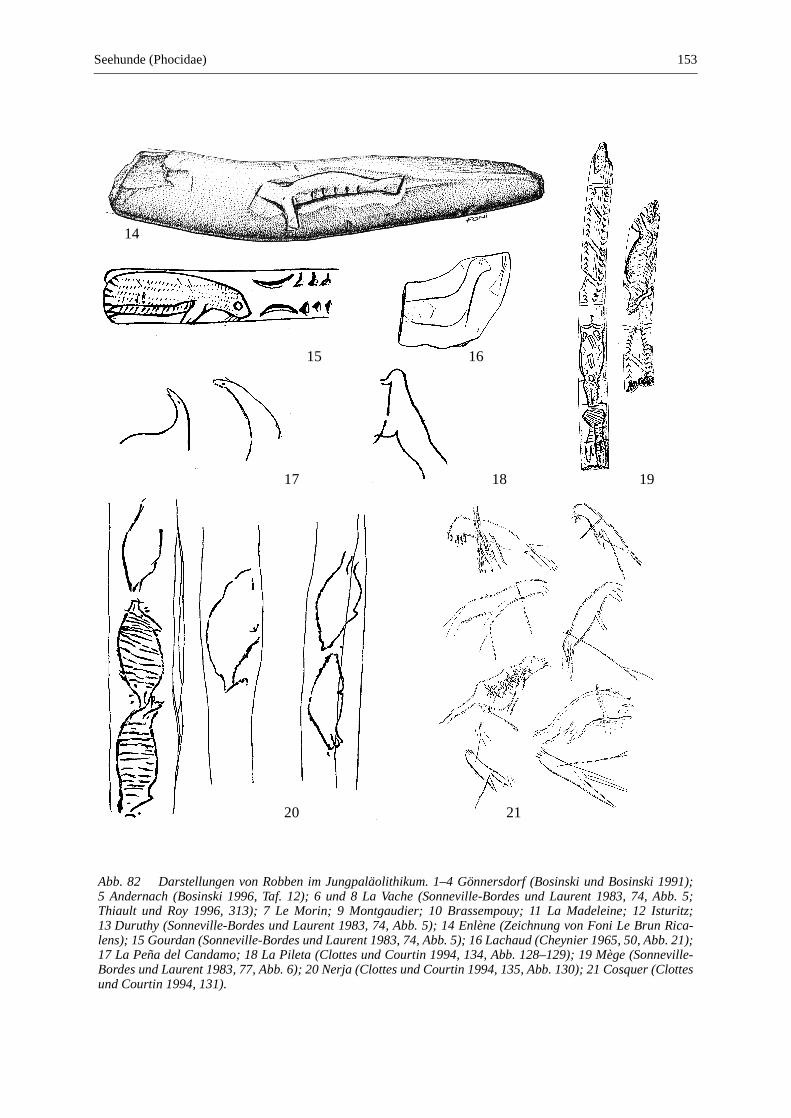
Seehunde (Phocidae) 153
Abb. 82 Darstellungen von Robben im Jungpaläolithikum. 1–4 Gönnersdorf (Bosinski und Bosinski 1991);5 Andernach (Bosinski 1996, Taf. 12); 6 und 8 La Vache (Sonneville-Bordes und Laurent 1983, 74, Abb. 5;Thiault und Roy 1996, 313); 7 Le Morin; 9 Montgaudier; 10 Brassempouy; 11 La Madeleine; 12 Isturitz;13 Duruthy (Sonneville-Bordes und Laurent 1983, 74, Abb. 5); 14 Enlène (Zeichnung von Foni Le Brun Rica-lens); 15 Gourdan (Sonneville-Bordes und Laurent 1983, 74, Abb. 5); 16 Lachaud (Cheynier 1965, 50, Abb. 21);17 La Peña del Candamo; 18 La Pileta (Clottes und Courtin 1994, 134, Abb. 128–129); 19 Mège (Sonneville-Bordes und Laurent 1983, 77, Abb. 6); 20 Nerja (Clottes und Courtin 1994, 135, Abb. 130); 21 Cosquer (Clottesund Courtin 1994, 131).
14 15 16 17 18 19
20 21

154 Verbreitung der großen Jagdfauna in Mittel und Westeuropa im oberen Jungpleistozän
Darstellungen aus dem Jungpaläolithikum
Die Zahl der sicheren und der wahrscheinlichenDarstellungen von Robben ist relativ gering, so dasshier der Versuch unternommen wurde, alle publi-zierten Daten zu behandeln.
Bis vor einigen Jahren war ein Dutzend Darstel-lungen von Robben bekannt und wurde immer wiederzitiert (Sonneville-Bordes und Laurent 1983; Dams1987; Cleyet-Merle 1990; Bosinski und Bosinski1991; Cleyet-Merle und Madelaine 1995). DieDarstellungen von Nerja62 (Abb. 82, 20) waren zwarbekannt, es gab jedoch keine Klarheit über ihre Deu-tung. Die Darstellung von Mas d’Azil war zwar publi-ziert worden (Chollot 1964, 307), aber sie wurdeübersehen. Es gibt weiterhin einige als „Phantome“und „Anthropomorphe“ angesprochene oder nichtnäher bestimmte Darstellungen, die durchaus Bildervon Robben sein könnten. Da ich noch nicht die Gele-genheit hatte, diese Darstellungen im Original zusehen, kann hier nicht näher auf diese Stücke einge-gangen werden.
Robbendarstellungen sind auf sehr unter-schiedlichen Unterlagen angebracht worden: kleinenSteinplatten wie in Gönnersdorf, Andernach, LaMadeleine oder Abri Lachaud (Abb. 82, 1–5, 11 und16), auf Lochstäben wie in Montgaudier und Le Morin(Abb. 82, 9 und 7), auf Zähnen wie in Duruthy (Abb.82, 13) oder auf Knochen wie in Brassempouy (Abb.82, 10) und Mas d’Azil63. In der Wandkunst treten sieals Malereien und Gravierungen auf, an Wänden wiein Cosquer und an Stalaktiten wie in Nerja (Abb. 82,21 und 20).
Fast alle Gravierungen der mobilen Kunststammen aus Schichten des Magdalénien. Die ältesteDarstellung in dieser Gruppe könnte die vom AbriLachaud aus dem Proto-Magdalénien sein.
Für die Wandkunst gibt es keine Datierung, da essich in Nerja um rote Malereien, in den anderenHöhlen um Gravierungen handelt. Falls die Gravie-rungen aus der Höhle von Cosquer genau so alt wärenwie die benachbarten Malereien, wäre eine zeitlicheZuordnung dieser Darstellungen um ca. 17.000 BPmöglich. Hier ist weiterhin zu vermerken, dass dieacht Gravierungen einen oder mehrere Stricheaufweisen, die wahrscheinlich als Projektile, etwa alsWurfspeere oder Pfeile, zu charakterisieren sind.
Reste aus dem Jungpaläolithikum
Robben sind meistens nur durch einzelne Fundevertreten, oft durch Zähne. Auch wenn Zähne wider-standsfähiger sind als andere Knochen und daherbessere Erhaltungschancen haben, kann dies nicht füralle Fundstellen als Hauptgrund für ihre bevorzugteAuffindung angenommen werden. Dem Archäologenkönnte das häufigere Auftreten von Zähnen gegenüberanderen Körperteilen (u. a. Castanet, Grottes deSaulges, Isturitz, La Marche, Raymonden, Altamira,Devil’s Tower, Arene Candide) zuerst auch alsErgebnis von sammlerischer Tätigkeit erscheinen,deren Ziel vielleicht die weitere Verwendung derZähne als Anhänger gewesen sein könnte. Aber außerden Funden aus La Marche und aus den Grottes deSaulges scheinen alle anderen keine gezielteBearbeitung aufzuweisen. Denise de Sonneville-Bordes und Pierre Laurent (1983, 72) erwägen für dieUnterkiefer von Castanet und Raymonden die Inter-pretation als Trophäen im weitesten Sinne.Gelegentlich kann es sein, dass ganze Robbenunter-kiefer oder einzelne Zähne gesammelt und als „etwasBesonderes“ betrachtet wurden, aber die auffälligeÜberrepräsentation der Zähne kann nicht dadurchallein erklärt werden. Eine wichtige Rolle spieltwahrscheinlich die Tatsache, dass Seehunde nicht zuder „üblichen” eiszeitlichen Fauna gehören. Daher,und insbesondere bei älteren Grabungen, ist dieBestimmung von Robben und deren Arten meistensnur anhand von Zähnen möglich gewesen, währenddie postkranialen Knochen meistens in der Kategorie„unbestimmt” landeten.
Aufgrund der geringen Zahl an Fundstellen mitResten von Robben kann ich hier alle mir bekanntenpublizierten erwähnen: In Spanien Tito Bustillo, LaRiera, Altamira, Nerja und Les Cendres; in PortugalFigueira Brava; auf Gibraltar Gorhams Cave undDewil’s Tower; in Italien Grimaldi, Arene Candide,Romanelli und Sant’Agostino. Alle diese Fundstellenbefinden sich direkt an der Küste oder nur wenigeKilometer von dieser entfernt. Dazu kommen dieFundstellen aus Frankreich: Isturitz, Castanet, Lartet,Raymonden und Grottes des Saulges. DieseFundstellen befinden sich, außer Isturitz, weit imInneren Frankreichs (Abb. 83).
62 Hier ist jedoch zu erwähnen, dass Michel Lorblanchet (1993, 186) sich entschieden gegen eine Interpretationdieser drei Figuren als Robben ausspricht. Er erkennt in diesen Darstellungen drei nicht näher zu bestimmendeFische.63 Von dieser Gravierung (Chollot 1964, 307) existiert keine Umzeichnung, sie wird daher hier nicht abgebildet.

Seehunde (Phocidae) 155
Diskussion
Die Fundstellen mit Darstellungen, La Peña delCandamo, La Pileta und Nerja in Spanien, Cosquer inSüdfrankreich, sowie die Fundstellen mit Resten, TitoBustillo, La Riera, Altamira und Nerja in Spanien,Gorhams Cave auf Gibraltar, Grimaldi und Romanelliin Italien, liegen alle an felsigen Küsten. Die heutigeKüste liegt nicht sehr weit von der eiszeitlichen Küsteentfernt. Hier ist ein Zusammenhang mit der direktenBeobachtung und Jagd auf Robben allgemein akzep-tiert. Die Linien auf den Darstellungen der HöhleCosquer könnten sogar Wurfwaffen darstellen. Ob essich bei den Darstellungen von La Peña del Candamound La Pileta tatsächlich um Robben handelt, istzumindest unsicher.
Für die Fundplätze im Inland stellt sich die Fragenach der Herkunft der auf Robben hinweisendenFunde (Bosinski und Bosinski 1991, 86; Sonneville-Bordes und Laurent 1983, 78–79).
Die Interpretationsmöglichkeiten lassen sich auf viereingrenzen:
1) Die Menschen des Paläolithikums haben dieseTiere an Flüssen oder Seen im Inland gesehen und siedort dargestellt.2) Die Menschen des Paläolithikums haben dieseTiere am Meer gesehen und haben dann diese Tiere imInland gezeichnet. In diesem Fall wäre das abstrakteKonzept des Tieres mit dem Künstler ins Inlandgereist.3) Die Menschen des Paläolithikums haben dieseTiere am Meer gesehen und dargestellt, und habenspäter deren Bilder oder Reste ins Inland mitgebracht.In diesem Fall wären es die konkreten Funde, diegereist sind.4) Die Funde im Inland können durch Tausch in denBesitz von Menschen gekommen sein, die weit vomMeer entfernt gelebt haben. In diesem Fall ist es nichtzwingend, dass Robben von den Menschen im Inlandje gesehen, gejagt oder dargestellt wurden.
Die Gravierungen von Robben aus Gönnersdorfund Andernach (Bosinski und Bosinski 1991;Terberger 1997) sind auf lokalen Schieferplattenangebracht worden, genau wie Hunderte andereGravierungen aus diesen Fundstellen. In diesem Fallkann man ausschließen, dass es sich um Stückehandelt, die anderswo graviert und dann importiertwurden. Nur die Vorkenntnisse, die den Künstler oderdie Künstler in die Lage versetzten, dieseDarstellungen von Robben anzufertigen, könnten vonanderswo stammen. Woher kannten die Künstler dieseTiere, um solche zum Teil detailreichen Darstel-lungen zu vollbringen? Der atlantische Ozean befandsich damals etwa 800 km entfernt im Westen, dieNordsee 450 km im Norden. Wenn man sich den
Rhein breiter, reicher an Wasser und an Fischenvorstellt, scheint doch wahrscheinlich, dass Robbendiesen Fluss bis ins Neuwieder Becken, wo sich dieFundstellen von Gönnersdorf und Andernachbefinden, hoch geschwommen sein könnten. Anderer-seits müssen diese Tiere nicht zwangsläufig vom weitentfernten Meer gekommen sein. Eine Population vonRobben könnte während des ganzen Jahres im Bereichder heutigen Nordsee gelebt haben. Es istanzunehmen, dass hier wegen des geringen Gefällesund der starken Sedimentation, zahlreiche flacheSüßwasserseen entstanden sind, wie man sie heute ausFinnland kennt und wo eben noch heute die Unter-arten Phoca hispida saimensis und Phoca hispidaladogensis leben. Weiterhin stammen mehr als 6 kgdes lithischen Rohmaterials aus Gönnersdorf und fast1 kg aus Andernach (Floss 1994, 195 und 220) aus derRegion 100 km nördlich des Neuwieder Beckens.Angesichts dieser Tatsache scheint es wahrscheinlich,dass die Menschen von Gönnersdorf und AndernachRobben eher im Rhein oder in einer ihnen vertrautenRegion 100 km nördlich von ihren Lagerplätzen alsam weit entfernten atlantischen Ozean gesehen haben.Ferner erinnern einige schlangenförmige Linien aufeiner der Gravierungen von Gönnersdorf mit ihrenKurven an einen Fluss (Abb. 82. 1), welcher der Rheinsein könnte.
Obwohl einige Darstellungen in diesem wie inanderen Fällen äußerliche Details der dargestelltenTiere zeigen, so dass eine Bestimmung der Artmöglich erscheint (Bosinski und Bosinski 1991, 85;Sonneville-Bordes und Laurent 1983), muss manvorsichtig bleiben. Die Freiheit des Künstlers sowiedie Ähnlichkeit zwischen den unterschiedlichen Artensind so groß, dass die Zuordnung meistens alsHypothese, selten als sicher betrachtet werden kann.
Bei den französischen Gravierungen von Isturitz,Duruthy, Brassempouy, Gourdan, Enlène, Mas d‘Azilund La Vache in den Pyrenäen, sowie bei den Kno-chenfunden von Castanet und Raymonden und bei denGravierungen von Montgaudier, Le Morin, Mège undLa Madeleine stellt sich die gleiche Frage. Hier istgegenüber Gönnersdorf das Meer viel näher gewesen(ca. 200–300 km). Da einzelne Seehunde oder kleineGruppen noch heutzutage gelegentlich die französi-schen Flüsse hochschwimmen, ist es wohlwahrscheinlich, dass sie es auch und sogar häufigerwährend der Eiszeit getan haben. Da es sich immerum einzelne Funde handelt, ist andererseits die Mög-lichkeit eines „Importes“ von der Küste, wo Robbenbestimmt häufiger waren, nicht auszuschließen.
Importe über Distanzen von hunderten von Kilo-metern sind während des Jungpaläolithikums fürMuscheln und lithisches Rohmaterial bekannt (Floss1994, 336–341) und für die Region der französischenPyrenäen und Vorpyrenäen sehr gut belegt (Bahn1982). In Mas d’Azil sind ein Zahn eines Pottwals und

156 Verbreitung der großen Jagdfauna in Mittel und Westeuropa im oberen Jungpleistozän
eine Muschel der Art Pecten maximus (Poplin 1983;Chollot 1964, 319; Thiault, Roy 1996, 274, 284) indiesem Zusammenhang hervorzuheben. Bei der„mobilen Kunst” stellen Knochen, Zähne oder Geweihdie am häufigsten verwendete Grundlage. DieseObjekte könnten von überall her stammen. Nur bei dermöglichen Robbendarstellung auf der Plakette vomAbri Lachaud kann man, wie schon für Gönnersdorfund Andernach, von lokalem Rohmaterial sprechen(Cheynier 1965, 29).
Die Darstellung von Montgaudier (Abb. 82, 9)wird als zwei hinter einem Fisch schwimmendeRobben interpretiert (Sonneville-Bordes und Laurent1983, 76). Ob es sich bei dem Fisch um einen Lachs,eine Forelle, eine Makrele oder eine andere Fischarthandelt, ist umstritten (Sonneville-Bordes und Laurent1983). Wäre dieser Fisch tatsächlich eine Forelle oderein männlicher Lachs mit gebogener Schnauze(Marshak 1970, 325), würde diese Darstellung eineSzene in einem Fluss, wahrscheinlich im Landesin-neren, darstellen. Drei Linien vor dieser Gruppekönnten als Lanzen oder Pfeile verstanden werden;dann hätte diese Gruppe einen jägerischen Charakter.Auch auf der Baguette-Demironde vom Abri Mègefindet man in der Nähe einer der Robben-darstellungen (Sonneville-Bordes und Laurent 1983,77) das Bild eines Lachses. Auch hier könnte mandieses als Hinweis deuten, dass sich diese Szene imFluss oder am Ufer eines Flusses abspielte. Fernerwird die Art der Darstellung der vermuteten Robbenvon Sonneville-Bordes und Laurent als eine Erläu-terung zur Schlachtung von Robben verstanden, damitwäre auch hier ein jägerischer Charakter enthalten.
In Enlène, Gourdan, Montgaudier und La Vachesind außer den Darstellungen von Robben auchDarstellungen von Lachsen mit gebogener Schnauzebekannt, die auf die Zeit hinweisen, in der die Lachsezur Fortpflanzung die Flüsse hoch schwimmen(Thiault und Roy 1996, 319). Dieser saisonal bedingteReichtum an Nahrung kann ein weiteres Motiv für dieRobben gewesen sein, auch Hunderte von Kilometernflussaufwärts zu schwimmen.
Bei der Kartierung (Abb. 83) der publiziertenDarstellungen und Reste von Robben fällt auf, dassalle Fundstellen weit im Inneren nördlich derPyrenäen und der Alpen liegen. Die Fundstellen inKüstennähe liegen dagegen alle südlich der Pyrenäen,der Alpen und an der französischen Mittelmeerküste.Zwei Interpretationsmodelle lassen sich erstellen.
Das erste Modell betrachtet dieses Muster als Ergebnisvon natürlichen Bedingungen. Ein Schwerpunkt in derVerteilung von paläolithischen Fundstellen in Frank-reich liegt in und um die Region des Périgord undentlang der Pyrenäen; entlang der heutigen Küste sindHöhlen und paläolithische Fundstellen seltener. InSüdeuropa und insbesondere in Spanien gibt es
umgekehrt an der Küste viel mehr Fundstellen als imInneren. Es ist daher nicht erstaunlich, dass diemeisten Nachweise von Robben aus fundreichenRegionen stammen. Ferner kann man auch eine Kom-ponente dieses Ergebnisses im Verhalten von Robbenfinden. Es ist sehr wahrscheinlich, dass atlantische,subarktische und arktische Robben während derEiszeit in den flachen, breiten, wasserreichen Flüssennördlich der Pyrenäen und der Alpen regelmäßighochgeschwommen sind. Möglicherweise spielte diesaisonale Wanderung von Lachsen dabei eine wichtigeRolle. Robben können in den Flüssen von Menschengesehen und eventuell auch gejagt und dargestelltworden sein. Südlich der Pyrenäen und der Alpen sinddagegen die meisten Flüsse deutlich schmaler undsteiler. Im Sommer führen sie heutzutage kaumWasser. Robben können durch diese nicht weit insInnere Spaniens, Frankreichs oder Italiens gelangen.Weiterhin ist die einzige gesicherte Robbenart ausdem Mittelmeer die Mönchsrobbe, welche heuteextrem selten ist. Mir ist kein Bericht bekannt, nachdem diese Art je in einem Fluss gesichtet worden ist.Im Mittelmeer gibt es außerdem auch keine Lachse,hinter denen diese Robben her gewesen sein könnten.Die Jagd auf Robben konnte daher südlich derPyrenäen und der Alpen nur an der Küste stattfinden.
Das zweite Modell stellt den anthropogenen Faktor inden Vordergrund. In den meistens durch die Tundrageprägten Landschaften Frankreichs und Deutsch-lands gab es keine natürlichen Hindernisse fürKontakte oder Wanderungen, die über Hunderte vonKilometern laufen konnten. Es fand ein regerAustausch im weitesten Sinne statt. Mehr als dieRobben an sich sind es das Konzept dieses Tieres unddessen Darstellungen oder Reste, die transportiertworden sind. Am Mittelmeer dagegen, wo dieLandschaft sehr stark durch Berge in kleinereRegionen unterteilt ist und wo es viel mehr Wald alsim Norden gegeben hat, blieben Austausch oderWanderung über größere Distanzen weit seltener. DieLebensweise könnte weiterhin aufgrund der Nutzungvon solchen „sicheren“ Nahrungsquellen aus demMeer durch eine geringere Mobilität zu charakteri-sieren sein (Pérez und Raga 1998, 259).
Unter den Befürwortern für eine saisonale Wanderungder Menschen und den folgenden Import von Objektenauch im Zusammenhang mit den Resten von Robbenaus der Küstenregion ist hier Paul Bahn und seinArtikel “Seasonal migration in South-West Franceduring the Late Glacial Period” (1977) zu zitieren.Denise de Sonneville-Bordes und Pierre Laurent(1983, 79) machen jedoch zu Recht auch auf diewahrscheinlich wiederholte Präsenz von Robben inFlüssen aufmerksam.

Seehunde (Phocidae) 157
Wahrscheinlich sind manche Darstellungen oderReste von Robben auf Importe, jahreszeitliche Wande-rungen und auf Kontakte zurückzuführen, währendandere Darstellungen an Ort und Stelle hergestellt und
Tiere dort erlegt wurden, wo die Knochen gefundenwurden. Sicherheit darüber, welche Darstellung woentstand und welcher Knochen woher stammt, wirdwohl nie bestehen.
Abb. 83 Fundstellen mit publizierten Resten (Dreiecke) oder Darstellungen (Kreise) von Robben. 1 Gönners-dorf; 2 Andernach; 3 La Marche; 4 Mège; 5 Raymonden; 6 La Madeleine; 7 Lachaud; 8 Castanet; 9 Lartet;10 Le Morin; 11 Montgaudier; 12 Isturitz; 13 Duruthy; 14 Brassempouy; 15 Gourdan; 16 Enlène; 17 Masd’Azil; 18 La Vache; 19 Cosquer; 20 La Peña del Candamo; 21 Tito Bustillo; 22 La Riera; 23 Altamira; 24 LesCendres; 25 La Pileta; 26 Nerja; 27 Gorhams Cave; 28 Devil’s Tower; 29 Figueira Brava; 30 Grimaldi;31 Arene Candide; 32 Grotta di Sant’Agostino; 33 Romanelli; 34 Grottes de Saulges.


Waltiere (Cetacea)
Die Waltiere zeichnen sich nach den Worten von Volker Storch und Ulrich Welsch (1993, 447) durch folgendeEigenschaften aus: „Rein wasserlebend und dadurch stark umgewandelt. Körper fischförmig, Haarkleidgewöhnlich vollkommen rückgebildet. Hochentwickeltes Gehirn. Vorderextremitäten flossenartig. Hinter-extremitäten äußerlich völlig fehlend, ihr Skelett und der Beckengürtel fast völlig rückgebildet. Schwanz zueiner breiten, horizontalen Flosse umgebildet“Vorwiegend verwendete Literatur: Feruglio und Robinau 2001; Pérez und Raga 1998; Wandrey 1997.
Zur Ordnung der Waltiere (Cetacea) gehören ca. 80Walarten. Diese unterscheiden sich in zwei Unterord-nungen: Bartenwale (Mysticeti), mit den FamilienGlattwale (Balaenidae), Grauwale (Eschrichitiidae)und Furchenwale (Balaenopteridae), sowie Zahnwale(Odontoceti), mit den Familien der Pottwale(Physeteridae), Gründelwale (Monodontidae), Schna-belwale (Ziphiidae), Schwert- und Grindwale (Globi-cephalidae), Delphine (Delphinidae), Schweinswale(Phocoenidae) und Flussdelphinartige (Platani-stoidae). Die üblichen vereinfachten BezeichnungenWal oder Delphin beschreiben lediglich grobeGrößenklassen. So steht z. B. der Schwert- oderKillerwal in der Verwandtschaft den Delphinen näherals den Walen.
Waltiere sind in ihren Körpermaßen, Nahrungs-gewohnheiten und Verhalten sehr unterschiedlich. Dasie aber in jungpaläolithischen Fundstellen nur durchsehr wenige Knochenreste und Darstellungen vertretensind, werden sie hier zusammen behandelt.
Habitat: Waltiere leben meistens im offenen Meer.Eine Art der Familie Delphinidae (Sotalia fluviatilis)sowie einige Arten der Familie Platanistoidea (Plata-nista gangetica; Platanista minor; Inia geoffrensis;Lipotes vexillifer) leben in den großen Flusssystemendes Amazonas, Ganges, Indus, Mekong und Yang-Tse-Kiang. Es gibt verschiede Hinweise darauf, „daßWale durchaus in der Lage sind, größere Strömehinaufzuschwimmen, sofern sie sich in dasMündungsgebiet verirrten. Bekannt ist ja, dass vieleArten bevorzugt Küstengewässer und enge Buchtenaufsuchen“ (Jux und Rosenbauer 1959, 113).
Verbreitung: Bis zur Erfindung der Harpunen-kanone im Jahr 1868 war das Verhältnis zwischenWalfängern und Walen einigermaßen ausgewogen.Durch die Industrialisierung, den steigenden Bedarf anÖl sowie die Entwicklung immer schnellerer Schiffekam es rasch zu eine Überjagung der Wale, dievielerorts zu deren Ausrottung geführt hat. Delphineund Wale kommen auch am Mittelmeer vor, sie sindaber hier seltener als im Atlantik.
Länge und Gewicht: Das größte Waltier ist derBlauwal, mit einer Länge von 30 m und einemGewicht von über 130 t (soviel wie ca. 30 Elefanten).
Tragzeit: bei Walen 11–16 Monate. Die natürliche Lebenserwartung ist sehr unter-
schiedlich. Größere Wale wie der Blauwal, der Seiwaloder der Pottwal haben eine natürliche Lebenser-wartung von über 60 Jahren. Die Lebenserwartungliegt beim Großen Tümmler bei 37 Jahren, beimBeluga bei 25–35 Jahren. Beim Schweinswal wirdeine durchschnittliche Lebenserwartung von 9 Jahrenangegeben; als Höchstalter wurden 14 Jahre registriert(Wandrey 1997).
Klimatische Aussage: Waltiere leben in allenMeeren, oft mit einem sehr breiten Lebensraum. Allebis heute entdeckten ins Jungpaläolithikum datie-renden Funde wurden auf Arten zurückgeführt, dienoch heute an der atlantischen Küste Europas oder amMittelmeer vorkommen.
Nahrung: Bartenwale ernähren sich von Plankton,das in einer Tiefe von bis zu 100 m anzutreffen ist.Dadurch brauchen sie nicht so tief wie Pottwale zutauchen, die sich bis zu 80% von Kalmaren undseltener sogar von Riesenkalmaren ernähren. ZumNahrungsspektrum der Schwertwale gehören nebenFischen und Weichtieren auch andere Wale, Robben,Seekühe und Meeresvögel. Delphine ernähren sichmeistens von Fischen, die sie in den oberen Wasser-schichten jagen.
Verhalten: Manche Wale sollen bis in 3000 mTiefe tauchen können (Wandrey 1997, 21). DerNormaltauchgang eines Pottwales dauert bis zu 40Minuten. Dabei erreicht er bis zu 500 m Tiefe.Während Delphine ausgesprochen gesellig sind, sodass manche Arten Gruppen von über 1000 Tierenbilden können, scheinen Blauwale eher als Einzel-gänger oder in Mutter-Kind-Gruppen zu leben.
Ein weiterer für diese Arbeit relevanter Aspektihres Verhaltens ist das Stranden zahlreicher Tiere.Die Ursache dieses Phänomens, das schon für dasMittelalter schriftlich belegt ist (Cleyet-Merle 1990,39), ist noch nicht ganz klar, möglicherweise ist beiden verschiedenen Arten der Ausschlag gebendeGrund auch sehr unterschiedlich. Da diese Tierebestens an das Leben im Meer angepasst sind, mussman davon ausgehen, dass sie kaum aktiv auf demMeer gejagt wurden. Man wird aber mit Sicherheit diegestrandeten Tiere zu verwerten gewusst haben.

160 Verbreitung der großen Jagdfauna in Mittel und Westeuropa im oberen Jungpleistozän
Darstellungen aus dem Jungpaläolithikum
Bis vor einigen Jahren waren kaum Darstellungen vonWaltieren allgemein bekannt, so dass auch in Fachbü-chern (Wandrey 1997, 27) die Felsbilder aus derJungsteinzeit Norwegens oder generell in Nordeuropaals die ältesten Darstellungen angesprochen werden.Drei sichere und vier wahrscheinliche Darstellungenvon Waltieren aus dem Magdalénien Spaniens undFrankreichs erlauben heute, diese Sicht zu revidieren.
Sichere Darstellungen von Waltieren in dermobilen Kunst sind ein Anhänger aus Las Caldas, dermöglicherweise aus einem Zahn eines Pottwalesgefertigt ist (Corchón 1994, 249), ein Anhänger ausArancou und eine Baguette-Demironde aus La Vache,auf der ein Pottwal dargestellt ist (Abb. 84, 1–3). Dreisehr kleine Gravierungen auf dem Lochstab vonMontgaudier (Robineau 1984), auf dem ebenfalls zweiRobben dargestellt wurden (Abb. 84, 4 und Abb. 82,9), können als wahrscheinliche Darstellungen vonWalen im Meer interpretiert werden.
Die einzige mögliche Darstellung eines Waltieresin der Wandkunst ist eine schwarze Malerei aus derHöhle von Tito Bustillo in Spanien (Abb. 84, 5).
Resten aus dem Jungpaläolithikum
Jungpaläolithische Reste von Walen und Delphinensind äußerst selten. François Poplin überprüfte die bis1983 bekannten und publizierten Reste von Cetaceen.Dabei erwiesen sich einige Bestimmungen alsFehlbestimmungen, während andere Reste verschwun-den zu sein scheinen. Eine sehr schöne Darstellung
eines Steinbockes von Mas d’Azil auf einem Träger,der bis 1983 als Mammut-Elfenbein interpretiertwurde, erwies sich bei einer näheren Untersuchung alsein Pottwal-Zahn64 (Poplin 1983). Dies zeigt, dass esmöglich ist, dass sich hinter manchen Bestimmungenals Mammut ebenfalls Reste von größeren Waltierenverbergen können. Insbesondere bei kleinen, starkbearbeiteten Elfenbeinobjekten ist eine genaueBestimmung nicht mehr möglich.
Als sichere Reste von Pottwalen (Physetercatodon) gelten ein Zahn aus El Castillo und der schonerwähnte Zahn aus Mas d’Azil (Poplin 1983, 90).Ebenfalls zu einem Pottwal könnte der gravierte Zahnaus Las Caldas gehören (Corchón 1994).
In der Schicht des Magdalénien IV von Duruthywurden drei einzelne Zähne bestimmt. Zwei davon(nach Delpech 1970, 23) könnten zu dem GroßenTümmler (Tursiops truncatus) oder (nach Poplin1983, 91) dem Gewöhnlichen Grindwal (Globice-phala melanaea) und dem Beluga (Delphinapterus)gehören. Der dritte Zahn konnte nicht näher bestimmtwerden.
In den Magdalénien-Horizonten der Höhle vonNerja (Pérez und Raga 1998, 254) wurden siebenReste von größeren Delphinen entdeckt, wahr-scheinlich vom Großen Tümmler (Tursiops truncatus)und vom Rundkopf-Delphin (Grampus griseus).Weiterhin wurden 30 Reste vom Gemeinen Delphin(Delphinus delphis) entdeckt. In den jungpaläolithi-schen Schichten B und D der Gorhams Cave inGibraltar wurden 12 Reste von Waltieren bestimmt(Waechter 1964, 214).
Abb. 84 Darstellungen von Walen. 1 Las Caldas (Corchón 1994, 250, Abb. 15); 2 Arancou (Crémadès 1997,79); 3 La Vache; 4 Montgaudier (Robineau 1984, 664); 5 Tito Bustillo (Balbín 1989, 446).
1 2 3
4 5
64 Pottwalzähne sind kegelförmig und können einzeln über ein Kilogramm Gewicht erreichen (Wandrey 1997, 133).

Waltiere (Cetacea) 161
Diskussion
Von allen Fundstellen mit Resten von Waltieren istdie Höhle von Mas d’Azil diejenige, die sich amweitesten im Landesinneren befindet. Sie liegt ca.150 km vom Mittelmeer und 230 km vom Atlantikentfernt. Wegen seiner Größe ist es unmöglich, dassder Pottwal die Flüsse aufwärts geschwommen ist. Esfolgt daraus, dass dieser Zahn, der wie schon erwähntvon den Wissenschaftlern jahrzehntelang alsMammut-Elfenbein betrachtet wurde, auch in derEiszeit wahrscheinlich nur als Rohmaterial gehandeltund in die Höhle importiert wurde. Bestenfalls galt dieauffällige Form als etwas Besonderes, aber dieBearbeitung dieses Stückes zu einem Hochrelief einesSteinbockes erfolgte so wie beim Mammut-Elfenbein.
Es fällt auf, dass die Fundstellen mit Resten oderDarstellungen von Waltieren in Frankreich in einemgewissen Abstand zum Meer liegen (Abb. 85),während sich solche Fundstellen südlich der Pyrenäenin der Nähe des Meeres befinden. In Italien sind keinejungpaläolithischen Fundstellen mit Resten vonWaltieren entdeckt worden. Die Reste von Waltierenaus der Grotta dell’Uzzo sind mesolithisch (Mussi1992, 692). Die angeblichen Reste von Pottwal undDelphin aus den Grimaldi-Höhlen sind verschwundenund werden von François Poplin als fraglichbezeichnet (Poplin 1983, 91). Schließlich ist noch zuvermerken, dass bis heute keine Reste und keineDarstellungen des doch so charakteristischenNarwales aus paläolithischen Fundstellen bekanntsind. Die heutige Verbreitung dieses Tieres lässtjedoch vermuten, dass es in der Eiszeit ebenfalls vorder atlantischen Küste Europas vorgekommen ist.
Abb. 85 Fundstellen mit Resten (Dreiecke) oder Darstellungen (Kreise) von Walen im Jungpaläolithikum. 1 Las Caldas; 2 Tito Bustillo; 3 El Castillo; 4 Nerja; 5 Gorhams Cave; 6 Arancou; 7 Duruthy; 8 Mas d’Azil;9 La Vache; 10 Montgaudier.


Riesenalk (Pinguinus impennis)
E. – Great Auk; F. – Pinguin geant; S. – Alca Gigante; Kat. – Gavot gegant; P. – alca-gigante o grande pinguim;I. – Alca gigante.Ordnung: Charadriiformes; Familie: Alcidae; Gattung: Pinguinus Bonnaterre 1790; Art: Pinguinus impennis,(Linnaeus 1758).Ausgestorben: Die letzten beiden Individuen wurden am 3. Juli 1844 vor Island getötet.Vorwiegend verwendete Literatur: Clottes und Courtin 1994; Errico 1994; Fuller 2002.
Aussehen bei Erwachsenen im Sommer: Anhand derzahlreichen modernen Zeichnungen und der ausge-stopften Tiere in den verschiedenen Museen der Welt(Fuller 2002) weiß man, dass der Kopf, der Rückenund die Flügel schwarz, manchmal mit Tendenz zumDunkelbraunen waren. Die Kehle war schwarz oderdunkelbraun. Zwischen Auge und Schnabel lag einweißer ovaler Fleck. Der dorsale Teil der Flügel warebenfalls schwarz mit einem weißen Rand amOberflügel, die palmare Seite des Flügels war weiß.Weiß war ebenfalls der gesamte ventrale Bereich desTieres sowie die Brust und der ventrale Teil desSchwanzes. Die Füße und der Schnabel warenschwarz. Der Schnabel weist weiße Einkerbungen auf(Abb. 86).
Verbreitung: Während der Eiszeit ist er an dereuropäischen Küste im westlichen Teil des Mittel-meers nachgewiesen worden. Der östlichste Fundstammt aus der Höhle von Romanelli in Apulien.Fossile Reste stammen von den Kanarischen Inselnund aus Florida.
Obwohl er mehrmals als typischer Vertreter fürsubarktische und arktische klimatische Verhältnissebezeichnet worden ist (u. a. Capasso et al. 1991, 239;Brizzi 1977, 98), scheint diese Annahme nicht zustimmen. Betrachtet man die Wassertemperaturen amMittelmeer während der Eiszeit (Abb. 9), weisen diesekeineswegs auf extrem kalte Bedingungen hin. Fernerlebte der Alk noch bis mindestens ins 4. Jahrhundertnach Christus auf der Iberischen Halbinsel (Hernández1995, 259).
Die Populationen, die im 17. und 19. Jahrhundertvollständig ausgerottet wurden, waren schon damalsRestpopulationen. Noch im 15. Jahrhundert umfassteihre Verbreitung in Europa die gesamte atlantischekontinentale Küste nördlich der Bretagne bis in denNorden Norwegens sowie die Britischen Inseln(Cassoli et al. 1979).
Verhalten: Ständig im Wasser, bildete derRiesenalk Kolonien mit bis zu Tausenden vonIndividuen. Paarungszeit, Tragzeit der Eier, Brüt- undSchlüpfzeit sind nie wissenschaftlich untersuchtworden und mögen im Norden Norwegens andersgewesen sein als in Frankreich. Es scheint jedoch,dass die Riesenalke nur sehr kurz an Land blieben. Sie
legten ihre Eier wahrscheinlich Ende Mai, dasSchlüpfen erfolgte um die Sommersonnenwende undsie verließen das Land mit den Jungen schon in derersten Julihälfte (Fuller 2002, 41).
Körpermaße: Kopf-Rumpf-Länge 75 bis 80 cm;Gewicht 5 kg.
Natürliche Lebenserwartung: 20 bis 25 Jahre.Nahrung: Fische. Klimatische Aussage: Betrachtet man die natür-
liche Verbreitung dieser Art während der letztenEiszeit sowie ihre Ausbreitung bis zum 15.Jahrhundert, so zeigt der Riesenalk, dass dasMittelmeer kühler war, ohne jedoch arktische Tempe-raturen zu erreichen.
Abb. 86 Riesenalk im Sommer (links) und im Winter(rechts) (Naumann 1903, Taf. 17).

164 Verbreitung der großen Jagdfauna in Mittel und Westeuropa im oberen Jungpleistozän
Darstellungen aus dem Jungpaläolithikum
Nur in drei Fundstellen wurden Darstellungen ent-deckt, die als mögliche Riesenalke angesprochenworden sind. Die drei Tiere aus der Höhle vonCosquer (Frankreich) sind wohl die eindeutigsten.Hier sind die Vögel in Bewegung dargestellt. Mansieht nicht nur die Silhouette der Tiere, in der dieGröße des Körpers gegenüber Kopf und Extremitätenbetont ist, sondern man sieht, dass es sich um flugun-fähige Vögel handelt, die sich an Land nur mit Mühefortbewegen konnten (Abb. 87, links). Ganz anders istes bei den Gravierungen von El Pendo in Spanien undGrotta Paglicci in Italien (Abb. 87, Mitte und rechts).Hier sind neben der Silhouette nur wenige Details desTieres zu erkennen. Die Flügel sind am Körpergehalten, so dass sie für eine Argumentation nicht zuVerfügung stehen. Die Haltung und die Form desKörpers, die Größe und die Form des Kopfes, desSchnabels und die Lage des Auges lassen in beidenFällen die Ansprache als „mögliche Darstellung“65 zu.
Reste aus dem Mittel- und Jungpaläolithikum
Alle paläolithischen Fundstellen, außer der Mousté-rien-Fundstelle von Cotte de Saint Brélade (Jersey),befinden sich in Südeuropa (Abb. 88). Das ist zwareinerseits, sowie bei Robben und Waltieren, mit demdamals tieferen Meeresspiegel und der verändertenKüstenlinie in Verbindung zu setzen, andererseits darfman nicht vergessen, dass der Vorstoß des Packeisesnach Süden, das während des zweiten Kältema-
ximums die Höhe von Nordportugal im Wintererreichte, impliziert, dass im Winter die Riesenalkesüdlich dieser Grenze leben mussten.
Der Mangel an Resten von Riesenalken rührteinerseits von der damals niedrigeren Meeres-spiegelhöhe her, andererseits liegen konkrete Hinder-nisse für die sichere Bestimmung der Knochen diesesVogels vor. Im Jahr 1928, also mehr als 75 Jahrenachdem Pinguinus impennis ausgestorben war,kannte Gian Alberto Blanc (1928, 10) nur zwei präpa-rierte Skelette, eines in London, das andere in Paris.Dazu kommen in einigen Museen Skelette, die aus denAufsammlungen insbesondere von Funk Islandstammen. Diese wurden teilweise aus den Knochenunterschiedlicher Individuen zusammengestellt (Fuller2002, 342ff.).
Wegen des Mangels an gutem, zugänglichemVergleichsmaterial ist die Bestimmung der Restebeeinträchtigt und führt zu einer geringeren Zahl anbestimmten Knochen. Dass dies nicht nur eineHypothese ist, zeigt die Art der bestimmten Knochenvon Pinguinus impennis. Fast immer handelt es sichum Sternum, Femur oder Humerus. Diese sind dieaussagekräftigsten Knochen für eine Bestimmung alsPinguinus impennis.
Diskussion
Alle bis heute bekannten Reste von Riesenalkenstammen aus Küstenregionen. Die Tatsache, dassdieser Vogel weder fliegen, noch sich am Bodenwehren konnte, lässt ausschließen, dass er weit insInland eindringen konnte.
Abb. 87 Als Riesenalk publizierte Darstellungen des Jungpaläolithikums. Links, Cosquer (verändert nachClottes und Courtin 1994, 127); Mitte, El Pendo; rechts, Paglicci (Clottes und Courtin 1994, 129).
65 Francesco d‘Errico hält nur die Gravierung von der Höhle El Pendo, neben den Malereien von der HöhleCosuer, für mögliche Darstellungen von Riesenalken. Er schließt die Gravierung von Paglicci aus, in der er ehereinen Greifvogel (Familie der Accipitriformes) zu erkennen meint (Errico 1994, 44–45). Die Ähnlichkeiten unterden Darstellungen von Cosquer, El Pendo und Paglicci und zwischen Letzterer und der Art Pinguinus impennissind meiner Meinung nach jedoch bedeutender als die Unterschiede.

Riesenalk (Pinguinus impennis) 165
Bis zuletzt wurden diese Tiere kaum gejagt, sonderneher gesammelt, und ihr Fleisch und ihre außerge-wöhnlich großen Eier sollen ähnlich wie die vonGänsen geschmeckt haben (Fuller 2002, 60–77).Daher kann man davon ausgehen, dass, falls sie anStränden an Land kamen, zu denen die Menschenirgendwie Zugang finden konnten, sie als sichereNahrungsquelle betrachtet werden konnten.
Obwohl von dieser Tierart nur drei sichere und zweiweitere wahrscheinliche Darstellungen bekannt sind,sollte man bedenken, dass Vogeldarstellungen an sichals selten gelten. Weiterhin, wie schon am Beginn desKapitels auf S. 147 erwähnt, zeigt die Höhle vonCosquer mit ihrem Eingang unter Wasser, dass Belegefür diese Tiere eher in der heute unter dem Meeres-niveau verschwundenen Küstenregion als im Binnen-land zu erwarten sind.
Abb. 88 Fundstellen mit Resten (Dreiecke) oder Darstellungen (Kreise) von Riesenalken im Mittel- und Jung-paläolithikum. 1 El Pendo; 2 Nerja; 3 Gorhams Cave; 4 Devil’s Tower; 5 Figueira Brava; 6 Cosquer; 7 Arene Candide;8 Paglicci; 9 Romanelli; 10 Archi; 11 Cotte de Saint Brélade.


Mögliche Interpretation der Daten
Paläolithische Kunst und ihre Deutung
Bis heute sind zahlreiche Theorien um diepaläolithische Kunst entstanden66. Unter denwichtigsten sei hier als älteste die Theorie der „L’artpour l’art“ erwähnt, der dann die Vorstellung einerJagdmagie durch Henri Breuil, einer Strukturierungder Wandkunst um ein männliches und ein weiblichesPrinzip durch André Leroi-Gourhan oder einesSchamanismus durch Jean Clottes und Lewis-Williams folgten. Alle diese Theorien haben ihrenmehr oder weniger deutlich geäußerten Anspruch aufeine Erklärung der gesamten paläolithischen Kunstgemeinsam. Kurz nach ihrer Entstehung67 haben siemeistens eine weite Zustimmung bekommen. Mankann fast sagen, dass sie die Erforschung der paläo-lithischen Kunst in ihrer Zeit geprägt haben, um danneinige Jahre später unerbittliche Gegner zu finden(dazu u. a. Laming-Emperaire 1972 und Paul Bahn2003). Neben diesen Theorien mit hohem Anspruchseien hier einige etwas bescheidenere Interpretations-modelle erwähnt, die sich als Ziel gesetzt haben, einenBeitrag zur Interpretation der Kunstdarstellungen vonbestimmten Fundstellen oder Regionen zu leisten.Darunter seien hier die Theorie von Joachim Hahn zueiner Botschaft von Kraft und Aggression für dasAurignacien in Südwestdeutschland (Hahn 1986), dieIdee von Barbara Olins Alpert zu einem spielerischenSinn („sens ludique“) hinter einigen der jungpaläoli-thischen Darstellungen (Olins Alpert 1992), sowie dieInterpretation von Derek Hodgson zu einigenjungpaläolithischen Darstellungen in Höhlen alsgezielte 3D-Bilder (Hodgson 2003) aufgeführt.
Wie man sieht, haben sich bis heute viele Forschermit der Interpretation der paläolithischen Kunstbeschäftigt. Man hat Vorstellungen wie die eines„geheimen“ Interpretationsschlüssels (u. a. AndréLeroi-Gourhan), einer geheimen Botschaft (Hahn1986) oder einer „Grammatik“ (Sauvet und Wlodar-czyk 1995) verfolgt, die als „kulturelle“, bewußteEntscheidung zu bewerten sein solle (dazu sieheSauvet und Wlodarczyk 2001, 223). Bis jetzt hat sichjedoch niemand systematisch mit dem Spektrum derdargestellten Tiere und deren effektiver Verbreitungauseinandergesetzt.
Die Verbreitung der Tierdarstellungen verglichen mitder tatsächlichen Verbreitung derselben Tierartenanhand der Knochenreste (S. 49–165) hat gezeigt, dasseine Korrelation zwischen gejagter und dargestellterFauna zumindest auf einer regionalen und überregio-nalen Skala zu erkennen ist. Diese Korrelation beruhtauf der Anwesenheit und Abwesenheit von Tierenunter den Faunenresten und in der Kunst. Ausnahmensind außerordentlich selten.
Dieses Ergebnis zeigt, dass ein Filter vor jeder kultu-rellen Entscheidung existiert: die Umwelt. Die kultu-relle Komponente spielt mit Sicherheit eine wichtigeRolle, sowohl bei der Auswahl der Anzahl an dar-gestellten Tieren, die selten den Zahlenverhältnissender Faunenreste in den zeitgleichen Fundstellen ent-spricht als auch bei der Auswahl der dargestelltenTierarten. Allerdings bewegen sich diese Entschei-dungsmöglichkeiten wohl nur in dem Rahmen, den dieUmwelt vorgibt. Die Tatsache, dass im Jungpaläoli-thikum Kontakte über hunderte von Kilometernanhand von Schnecken, lithischem Rohmaterial oderKnochen nachgewiesen sind, darf nicht überbewertetwerden. Importe sind gegenüber lokalen Schnecken,Silex oder Knochenresten die Ausnahme (u. a. Bahn1982; Floss 1994; Féblot-Augustins 1997; Djindjian etal. 1999, 79–84). Im Bereich der paläolithischenKunst bedeutet dies, dass das Spektrum der bekanntenTierarten und der Arten, die dargestellt werdenkonnten, in erster Linie von der regionalen Umweltbeeinflusst oder sogar determiniert war. Beginnt manbei der Analyse der paläolithischen Kunst nicht beider Umwelt, sondern bei der „kulturellen“ Auswahlder dargestellten Tiere, zäumt man das Pferd vonhinten auf und übersieht zentrale Aspekte derdamaligen menschlichen Existenz.
Dieses Ergebnis, das vielleicht vordergründig banalwirkt, ist nicht nur relevant, sondern sogar wesentlich,da fast alle Forscher, die sich mit der paläolithischenKunst beschäftigen oder beschäftigt haben, es einfachvergessen oder nur am Rande erwähnen.
66 Für eine Vertiefung des Themas siehe: Floss 2004; Delporte 1990, 188–247; Lorblanchet 1995; Clottes undLewis-Williams 1997, 61–79.67 Im Fall der Theorie des Schamanismus sollte man eher von Wiederentdeckung sprechen, da schon anderefrüer von schamanistischen Elementen in der paläolithischen Kunst gesprochen hatten (u. a. Narr 1966).

168 Verbreitung der großen Jagdfauna in Mittel und Westeuropa im oberen Jungpleistozän
Die Wandkunst
Für die Analyse der jungpaläolithischen Wandkunsthabe ich mich entschieden, die publizierten Daten vonGeorges Sauvet und André Wlodarczyk zu ver-wenden. Auf der Basis der Arbeit von André Leroi-Gourhan pflegen sie seit dem Ende der 1970er Jahreeine Datenbank mit allen entdeckten Tierdarstel-lungen in der Wandkunst (Sauvet 1988; Sauvet undWlodarczyk 1995; Sauvet und Wlodarczyk 2001).Eine eigene Datenbank anhand der Literatur aufzu-bauen, wäre nicht möglich gewesen, da sie die Kennt-nis aller dieser Bilder im Original vorausgesetzt hätte.
Die Gruppierung von jungpaläolithischen Wanddar-stellungen von großen Jagdtieren nach Regionen (Tab.3) zeigt, dass ihre Anzahl sehr unterschiedlich ist.Während im Périgord 991 (27,8% von allen Darstel-lungen), in Kantabrien 994 (27,9%) und in denPyrenäen 775 (21,7%) einzelne Figuren erkanntworden sind, beträgt ihre Anzahl in Nordfrankreichnur 51 (1,4% von allen Darstellungen) und in Zentral-und Südspanien lediglich 203 (5,7%). Daher kann maneinen überregionalen Vergleich der Daten nur mitäußerster Vorsicht durchführen.
Es fällt weiterhin auf, dass folgende Tiere fehlen:Hydruntinus, Steppennashorn, Waldelefant, Gämse,Saiga-Antilope, Wildschwein, Damhirsch, Reh, Elch,Riesenhirsch, Moschusochse sowie die Meerestiere.Das liegt daran, dass alle diese Tiere in der paläoli-thischen Wandkunst selten und daher statistisch kaumrelevant sind.
Beim Hydruntinus wurde schon erwähnt, dass ernur als Hydruntinus erkannt wird, wenn das BildMerkmale eines Esels aufweist, insbesondere dieOhren und die Proportionen des Körpers. Alles wasnur als Equus zu erkennen ist, wird automatisch demEquus ferus und nie dem Equus hydruntinuszugeordnet. Dies gilt auch für das Steppennashorngegenüber dem Wollnashorn, für den Waldelefantengegenüber dem Mammut, für Damhirsch und Rehgegenüber dem Rothirsch.
Die gewöhnliche Seltenheit der Gämse liegtwahrscheinlich auch darin begründet, dass sie auf sehrhohen felsigen Bergen jenseits der üblicherweise vonMenschen bewohnten Gebiete lebt. Weiterhin ist sieviel scheuer als der Steinbock, der dagegen relativhäufig dargestellt wurde (ca. 11,1% aller Wandbilderder jungpaläolithischen Jagdfauna).
Die Saiga-Antilope, die in der Kleinkunst durchsehr schöne Darstellungen bekannt ist, ist in derWandkunst nur durch wenige grobe, unsichere Gravie-rungen vertreten. Die fast irrelevante Anzahl anmöglichen Darstellungen von Saiga-Antilopen in derWandkunst hat wahrscheinlich u. a. mit ihrer zeitlichsehr begrenzten Präsenz in Westeuropa zu tun.
Der Elch ist zwar nur zweimal in Gargas dargestelltworden, er ist aber in Frankreich erst ab demMagdalénien nachgewiesen. Der Riesenhirsch wurdeca. 30 mal in den Höhlen Frankreichs abgebildet,wobei er nur in der Höhle von Roucadour, einerHöhle, die leider kaum publiziert ist, zahlreich seinsoll.
Die Tatsache, dass nur ein paar mögliche Bildervon Damhirschen und keine einzige Darstellung vonRehen (die zwei wahrscheinlichen Gravuren sindKleinkunst) bekannt sind, hat neben ihrer Seltenheitauch damit zu tun, dass diese Tiere hauptsächlich imWald leben, kleine Gruppen bilden, teilweise sehrscheu sind und daher kaum von Menschenwahrgenommen werden konnten. Diese Anmerkunggilt natürlich auch für das Wildschwein, das ebenfallsäußerst selten dargestellt wurde. Zusätzlich darf mannicht vergessen, dass im Wald andere Materialien wiebeispielsweise Rinde oder Holz vorliegen, die demKünstler gute Unterlagen lieferten. Davon ist abernichts erhalten geblieben.
Nicht erhalten geblieben ist ebenfalls die jungpa-läolithische Küstenregion (Serangeli 2003). Diewenigen Darstellungen von Tieren aus dieser Regionmüssen daher als Indizien für eine im Meerverschwundene größere Anzahl an Darstellungeninterpretiert werden. Robben mit ihren 17 möglichenDarstellungen würden gerade 0,7% der Bilder derjungpaläolithischen Wandkunst ausmachen.
Setzt man sich mit der Anzahl der dargestelltenTierarten auseinander, fällt auf, dass manche Tierehäufiger repräsentiert sind als andere. Darunter ist dasPferd das am häufigsten dargestellte Tier (30,8% vonallen Tieren), während das Rentier (3,9%) viel seltenerdargestellt wurde. Rentiere zählen jedoch zu den ammeisten gejagten Tieren (Delporte 1990, 212–215).Diese Diskordanz hat dazu geführt, dass viele Autorendie kulturelle Komponente bei der Deutung der paläo-lithischen Darstellungen betont und der damaligenUmwelt sehr wenig Aufmerksamkeit geschenkt haben.Betrachtet man aber außer der Anzahl der Rentiere,die in manchen Höhlen in Frankreich sehr seltendargestellt sind, und der Anzahl der Pferde, die überallvertreten sind, auch die anderen Tierarten, könnenklare Tendenzen erkannt werden.
Auerochsen sind selten in den Pyrenäen (1,5%),sie sind im Périgord, Quercy, Languedoc-Rhône-Provence und in Kantabrien gut (5,3% bis 9,3%) undin Zentral- und Südspanien (12,8%) noch etwas besservertreten. In Süditalien kommen sie noch etwashäufiger vor (u. a. Leonardi 1988). In Nordfrankreichzeigen Auerochsen mit 7,8% eine sehr hohe Präsenz,andererseits wurden sie nur in der Höhle von Gouyentdeckt. Diese Höhle wird zum spätestenMagdalénien gerechnet (Leroi-Gourhan 1995, 517).
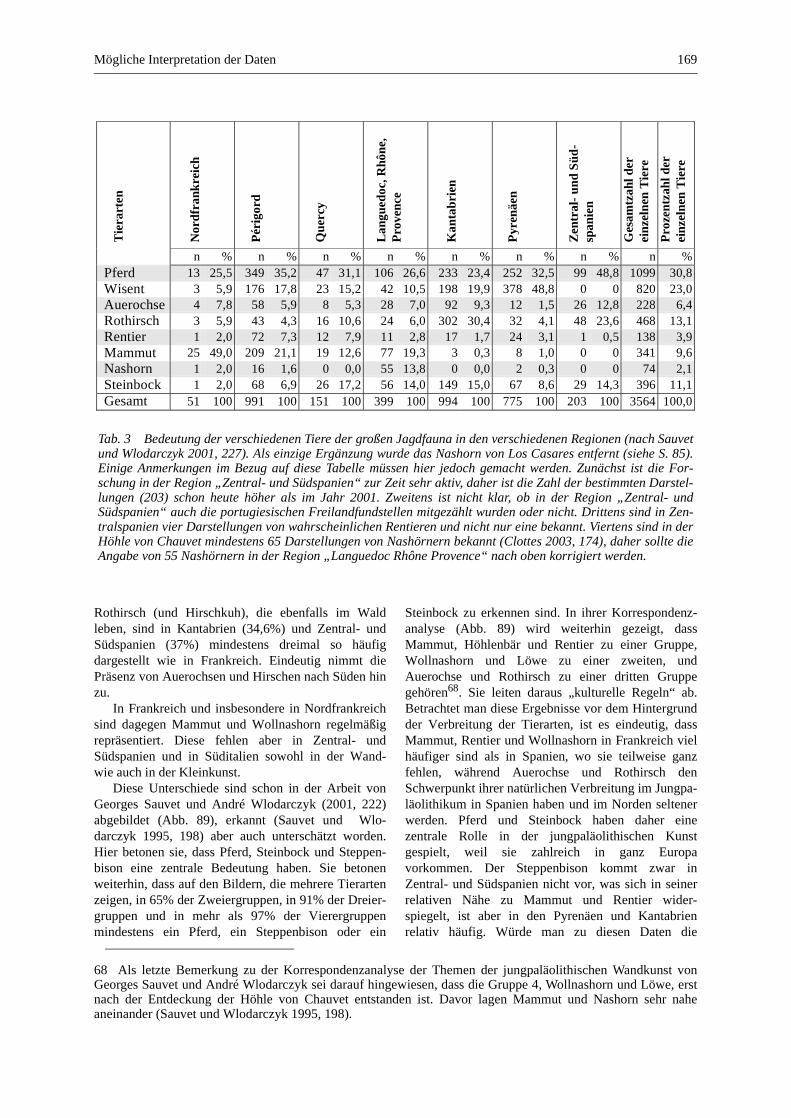
Mögliche Interpretation der Daten 169
Rothirsch (und Hirschkuh), die ebenfalls im Waldleben, sind in Kantabrien (34,6%) und Zentral- undSüdspanien (37%) mindestens dreimal so häufigdargestellt wie in Frankreich. Eindeutig nimmt diePräsenz von Auerochsen und Hirschen nach Süden hinzu.
In Frankreich und insbesondere in Nordfrankreichsind dagegen Mammut und Wollnashorn regelmäßigrepräsentiert. Diese fehlen aber in Zentral- undSüdspanien und in Süditalien sowohl in der Wand-wie auch in der Kleinkunst.
Diese Unterschiede sind schon in der Arbeit vonGeorges Sauvet und André Wlodarczyk (2001, 222)abgebildet (Abb. 89), erkannt (Sauvet und Wlo-darczyk 1995, 198) aber auch unterschätzt worden.Hier betonen sie, dass Pferd, Steinbock und Steppen-bison eine zentrale Bedeutung haben. Sie betonenweiterhin, dass auf den Bildern, die mehrere Tierartenzeigen, in 65% der Zweiergruppen, in 91% der Dreier-gruppen und in mehr als 97% der Vierergruppenmindestens ein Pferd, ein Steppenbison oder ein
Steinbock zu erkennen sind. In ihrer Korrespondenz-analyse (Abb. 89) wird weiterhin gezeigt, dassMammut, Höhlenbär und Rentier zu einer Gruppe,Wollnashorn und Löwe zu einer zweiten, undAuerochse und Rothirsch zu einer dritten Gruppegehören68. Sie leiten daraus „kulturelle Regeln“ ab.Betrachtet man diese Ergebnisse vor dem Hintergrundder Verbreitung der Tierarten, ist es eindeutig, dassMammut, Rentier und Wollnashorn in Frankreich vielhäufiger sind als in Spanien, wo sie teilweise ganzfehlen, während Auerochse und Rothirsch denSchwerpunkt ihrer natürlichen Verbreitung im Jungpa-läolithikum in Spanien haben und im Norden seltenerwerden. Pferd und Steinbock haben daher einezentrale Rolle in der jungpaläolithischen Kunstgespielt, weil sie zahlreich in ganz Europavorkommen. Der Steppenbison kommt zwar inZentral- und Südspanien nicht vor, was sich in seinerrelativen Nähe zu Mammut und Rentier wider-spiegelt, ist aber in den Pyrenäen und Kantabrienrelativ häufig. Würde man zu diesen Daten die
Tab. 3 Bedeutung der verschiedenen Tiere der großen Jagdfauna in den verschiedenen Regionen (nach Sauvetund Wlodarczyk 2001, 227). Als einzige Ergänzung wurde das Nashorn von Los Casares entfernt (siehe S. 85).Einige Anmerkungen im Bezug auf diese Tabelle müssen hier jedoch gemacht werden. Zunächst ist die For-schung in der Region „Zentral- und Südspanien“ zur Zeit sehr aktiv, daher ist die Zahl der bestimmten Darstel-lungen (203) schon heute höher als im Jahr 2001. Zweitens ist nicht klar, ob in der Region „Zentral- undSüdspanien“ auch die portugiesischen Freilandfundstellen mitgezählt wurden oder nicht. Drittens sind in Zen-tralspanien vier Darstellungen von wahrscheinlichen Rentieren und nicht nur eine bekannt. Viertens sind in derHöhle von Chauvet mindestens 65 Darstellungen von Nashörnern bekannt (Clottes 2003, 174), daher sollte dieAngabe von 55 Nashörnern in der Region „Languedoc Rhône Provence“ nach oben korrigiert werden.
n % n % n % n % n % n % n % n % 13 25,5 349 35,2 47 31,1 106 26,6 233 23,4 252 32,5 99 48,8 1099 30,8 3 5,9 176 17,8 23 15,2 42 10,5 198 19,9 378 48,8 0 0 820 23,0 4 7,8 58 5,9 8 5,3 28 7,0 92 9,3 12 1,5 26 12,8 228 6,4 3 5,9 43 4,3 16 10,6 24 6,0 302 30,4 32 4,1 48 23,6 468 13,1 1 2,0 72 7,3 12 7,9 11 2,8 17 1,7 24 3,1 1 0,5 138 3,9 25 49,0 209 21,1 19 12,6 77 19,3 3 0,3 8 1,0 0 0 341 9,6 1 2,0 16 1,6 0 0,0 55 13,8 0 0,0 2 0,3 0 0 74 2,1 1 2,0 68 6,9 26 17,2 56 14,0 149 15,0 67 8,6 29 14,3 396 11,1
51 100 991 100 151 100 399 100 994 100 775 100 203 100 3564 100,0
Pferd Wisent Auerochse Rothirsch Rentier Mammut Nashorn Steinbock Gesamt
Tie
rart
en
Nor
dfra
nkre
ich
P
érig
ord
Q
uerc
y
Lan
gued
oc, R
hône
, P
rove
nce
K
anta
brie
n
Pyr
enäe
n
Zen
tral
- un
d Sü
d-sp
anie
n
Ges
amtz
ahl d
er
einz
elne
n T
iere
P
roze
ntza
hl d
er
einz
elne
n T
iere
68 Als letzte Bemerkung zu der Korrespondenzanalyse der Themen der jungpaläolithischen Wandkunst vonGeorges Sauvet und André Wlodarczyk sei darauf hingewiesen, dass die Gruppe 4, Wollnashorn und Löwe, erstnach der Entdeckung der Höhle von Chauvet entstanden ist. Davor lagen Mammut und Nashorn sehr naheaneinander (Sauvet und Wlodarczyk 1995, 198).

170 Verbreitung der großen Jagdfauna in Mittel und Westeuropa im oberen Jungpleistozän
Darstellungen aus Italien hinzuzählen, würden dieseErgebnisse noch stärker hervorgehoben werden, da inItalien keine Mammuts, Wollnashörner, Steppenbi-sons oder Rentiere in der Wandkunst abgebildet sind.
Vergleicht man statt der Arten die Familien unter-einander, ergibt sich, dass die Equiden (Equus ferusund Equus hydruntinus) 30,8%, die Boviden (Bisonpriscus und Bos primigenius) 29,4% und die Cerviden(Cervus elaphus und Rangifer tarandus) 15% derabgebildeten Tiere umfassen. Die Unterschiede sindnicht mehr so stark. Sieht man die Verbreitungs-unterschiede von Darstellungen und Resten vonSteppenbison und Auerochse, sowie von Rentier undRothirsch, so lassen sich diese Paarungen als unter-schiedliche Anpassungen der Familie der Boviden undCerviden begreifen. Es fällt auf, dass das Pferddagegen im Hydruntinus keinen vergleichbarenGegenpol unter den Equiden gehabt hat. Die sicherenund möglichen Darstellungen des Hydruntinus in derWandkunst entsprechen nicht einmal 0,1% allerDarstellungen. Dies spiegelt sich auch in den Faunen-resten wider. Das Pferd hatte im Jungpaläolithikum inMittel- und Westeuropa, anders als Boviden undCerviden, keinen ernsthaften Konkurrenten in seinerökologischen Nische.
Eine wichtige Anmerkung muss man noch zurAuswahl der dargestellten Tiere machen. Die amhäufigsten abgebildeten Tiere sind solche, die in
größeren Herden im Freien leben, nämlich Pferd,Steinbock und Steppenbison. Zusammen stellen sie ca.64,8% aller Jagdtiere in der paläolithischenWandkunst dar. Wie schon für Damhirsch, Reh undWildschwein erwähnt, darf man jedoch nichtvergessen, dass für den Künstler, der Tiere im Waldoder in der Nähe eines Waldes beobachtet hat, auchdie typischen Materialien aus dem Wald in Fragekommen. Davon ist gar nichts erhalten geblieben.Daher könnte es auch sein, dass Hirsche etwas seltenerin der Wandkunst sind, aber auch, dass sie vielleichthäufiger auf Bäume graviert oder gemalt wurden.Natürlich ist dies eine reine Hypothese, die nichtdiskutiert, jedoch erwähnt werden soll.
Die Kleinkunst
Leider existiert keine Arbeit, in der die Darstellungender Kleinkunst europaweit systematisch behandeltoder zumindest aufgezählt werden. In der Arbeit vonAndré Leroi-Gourhan (1995, 578) sind die Funde auseinigen Fundstellen aufgezählt, die jedoch wesentlichandere Prozentzahlen ergeben, als die Fundstellen, dievon Henri Delporte (1990, 209) gezählt wurden.
Die Schwierigkeit einer überregionalen Arbeitüber die Kleinkunst liegt zum Teil darin, dass dieFundstellen sich in einem sehr unterschiedlichenForschungsstand befinden. Die Informationen über dieim 20. Jahrhundert aufgesammelten Funde, die oft zuPrivatsammlungen gehören, sind lückenhaft dokumen-tiert in Bezug auf die Herkunft und erst recht auf dieFundlage. Die Fundstelle von Gönnersdorf, um einjüngeres Beispiel zu nennen, ist am Ende der 1960erund während der 1970er Jahren ausgegraben worden,und ein Teil der Schieferplatten wartet immer nochdarauf, ausführlich untersucht zu werden. Weiterhinsollte mam auch bedenken, dass, während Objekte derKleinkunst in Museen oder Forschungsinstitutionenaufbewahrt und untersucht werden, sich Darstel-lungen der Wandkunst seit ihrer Entstehung immer ander gleichen Stelle befinden, was ihre Untersuchungdeutlich erleichtert. Schließlich muss man nochbedenken, dass die Kleinkunst durch die Art derUnterlage, meistens Knochen oder dünne Stein-platten, einer Zerteilung und Zerstörung unterzogenworden ist, die die Bestimmung der Tierartenwesentlich erschwert.
Mit einer solchen Datenbasis ist es schwierig,überregionale Vergleiche anzustellen. Die Durchsichtder publizierten Kleinkunstdarstellungen der wich-tigsten Fundstellen in Mittel- und Westeuropa,darunter Gönnersdorf (u. a. Bosinski 1981; Bosinskiund Fischer 1980; Bosinski und Bosinski 1991),Parpalló (Pericot 1942; Villaverde 1994) und LaMarche (Pales und de Saint Pereuse 1969, 1981 und1989), der Publikationen über die Sammlung Piette
Abb. 89 Korrespondenzanalyse der Themen derjungpaläolithischen Wandkunst (nach Sauvet undWlodarczyk 2001, 222). An = Mensch; Bi = Hirschkuh; Bn = Steppenbison;Bq = Steinbock; Ce = Rothisch; Ch = Pferd; Div =Verschiedene Tiere; Li = Löwe; Ma = Mammut; Po =Fisch; Re = Rentier; Rh = Nashorn; Ur = Bär.

Mögliche Interpretation der Daten 171
(Chollot 1964), über die Funde aus den Pyrenäen(Thiault und Roy 1996) sowie über die jungpaläoli-thische Kunst in Italien (Leonardi 1988) erlaubt mirjedoch zu unterstreichen, dass die direkte Verbindungzwischen natürlicher Fauna und dargestellter Faunaebenfalls bestätigt wird. Es sind keine wesentlichenUnterschiede in der Verbreitung der Darstellungen inder Kleinkunst, in der Wandkunst und in der Fauna zubemerken. In den Kapiteln „Die große Jagdfauna derletzten Eiszeit“ und „Die große Fauna an den KüstenEuropas“ wurden von Tierart zu Tierart immer wiederkonkrete Funde zur Diskussion herangezogen undabgebildet.
Als ein gutes Beispiel, bei dem die Kenntnis über dieVerbreitung der Tierarten das Verständnis der paläoli-thischen Kunst unterstützen kann, sei hier auf dieDarstellungen von Mischwesen aus dem Aurignacienvon Südwestdeutschland, nämlich dem Löwen-menschen vom Hohlenstein-Stadel und dem kleinerenLöwenmenschen vom Hohle Fels, hingewiesen(Wehrberger 1994; Conard 2003; Conard et al. 2003).Außerhalb dieser Region und dieser Zeit sind bisheute keinen anderen Löwenmenschendarstellungenim Jungpaläolithikum Europas bekannt. Natürlichfragt man sich: Warum? Schaut man nach Frankreich,dann sieht man, dass dort mehrere Mischwesen darge-stellt sind, meistens haben sie den Körper einesMenschen und den Kopf eines Rindes (u. a. Clottes2003, 168). Warum ist es im Aurignacien in Südwest-deutschland anders? Selbstverständlich steckt eine„kulturelle“ Komponente dahinter, die maßgeblich zuder Entstehung dieser Darstellungen geführt hat, aberes gibt auch eine wichtige Umweltkomponente. Restevon Rindern sind in der Aurignacien-Fauna Südwest-deutschlands sehr selten und fehlen in den Schichtendes Aurignacien des Hohle Fels (Bolus et al. 2000, 47)und Hohlenstein-Stadels (Gamble 1979, 40; Weniger1982, 119). In dieser letzteren Fundstelle sind 29Reste des Höhlenlöwen bestimmt worden. Es liegtfolglich nahe, dass, trotz der bekannten Darstellungeneines Steppenbisons aus dem Vogelherd und eines ausdem Geißenklösterle (Müller-Beck et al. 2001, Taf. 2und 17), Rinder in der Welt der Menschen desAurignacien in Südwestdeutschland eine andere Rollespielten als Rinder in Frankreich, wo sie während desganzen Jungpaläolithikums fast immer zahlreichwaren. Daher scheint es mir zumindest wichtig zubetonen, dass die auffällige Seltenheit der Rinder imAurignacien in Südwestdeutschland bei der Wahl derKünstler, die bewusst zwei Löwenmenschfigurenhergestellt haben, sicher eine wichtige Rolle gespielthat.
Die Verbreitung der Knochenreste und derDarstellungen der großen Jagdfauna
In dieser Arbeit wurden zum ersten Mal die Knochen-reste der großen Jagdfauna von zahlreichen jungpaläo-lithischen Fundstellen aus Mittel- und Westeuropastatistisch so analysiert, dass man sie vergleichen kann(siehe S. 3). Dadurch konnte Tierart für Tierart derenmögliche maximale Verbreitung, sowie in manchenFällen deren Verschwinden dokumentiert und disku-tiert werden. Der Vergleich mit den Tierdarstellungenhat, wie schon erwähnt, wesentliche Ergänzungengebracht.
Beim Pferd konnte gezeigt werden, dass es sowohlsteile bergige (u. a. Alpen, Apenninen, Kantabrien,Pyrenäen und die gebirgige Region in Südspanien) alsauch sumpfige Regionen (Po-Ebene) so weit wiemöglich mied, während es dort zahlreich war, wo esweite offene Flächen gab (u. a. Apulien, fast überall inFrankreich und Deutschland). In der Wandkunst istdas Pferd das am meisten abgebildete Tier.Ausnahmen sind Nordfrankreich, wo jedoch die Zahlvon nur 51 dargestellten Tieren ihre statistischeAussagekraft begrenzt, Kantabrien und die Pyrenäen.Diese letzten beiden Regionen sind bergig. DieTatsache, dass viele der Fundstellen, wo Pferde außer-ordentlich zahlreich sind, in hügligen Landschaftenliegen, könnte weiterhin ein Indiz dafür sein, dass diePferde an Übergangsplätzen während jahreszeitlicherWanderungen erlegt wurden. Jahreszeitliche Wande-rungen dürfen jedoch nicht immer nur als Wande-rungen im Frühling in kühlere und im Spätsommerund Herbst in wärmere Regionen verstanden werden.Zugang zu Trinkwasser sowie zu Futter können zubedeutenden Wanderungen auch in anderen Jahres-zeiten und in unterschiedliche Richtungen geführthaben. Man denke zum Beispiel daran, dass im Winteran exponierten, höheren und kälteren Stellen, dort, woder Wind stärker den Schnee wegwehen konnte, mehrFutter zu Verfügung stand als in geschützten, niedri-geren und leicht wärmeren Regionen. Das giltnatürlich nicht nur für das Pferd, sondern auch fürandere Tierarten.
Die Verbreitung des Hydruntinus, der in der Regelim Jungpaläolithikum viel seltener als das Pferd undfast nur in südlichen Regionen verbreitet war, warwahrscheinlich stark von der Verbreitung des Pferdesabhängig. Seine anhand der Funde annehmbarestärkere Präsenz in Süditalien gegenüber Spaniensowie seine Präsenz auf Sizilien zeigen, dass derHydruntinus sich in steilen Landschaften (Sizilienkonnte er nur erreichen, indem er Kalabrien durch-

172 Verbreitung der großen Jagdfauna in Mittel und Westeuropa im oberen Jungpleistozän
querte), aber auch in wärmeren Perioden (z. B. imAlleröd und Bölling in Paglicci) sich regionalgegenüber dem Pferd durchsetzen konnte.
Beim Wollnashorn zeigten die Faunenreste, dass esim Jungpaläolithikum Spaniens fehlt und nur nördlichder kantabrischen Gebirge nachgewiesen ist. Diewürmeiszeitlichen Reste u. a. aus Cardamone inSüditalien und aus der Cueva de la Cariguela inSüdspanien zeigen jedoch, dass diese Art im Würmvor dem Jungpaläolithikum viel weiter nach Südengekommen war. Die Datierung dieser Fundstellen istmeist schwierig und ungenau. Die Präsenz einigerArtefakte aus dem Moustérien hilft auch kaum beieiner genaueren Datierung. Man kann jedochannehmen, dass diese Funde in die Sauerstoff-Isoto-penstufe 4, also ins erste Kältemaximum der letztenEiszeit zu datieren sind. Die Tatsache, dass die anmildere Klimate angepassten Nashörner, Stephanorhi-nus kirchbergensis und Stephanorhinus hemitoe-chus, bis weit in die Sauerstoff-Isotopenstufe 3nachgewiesen sind (u. a. in El Castillo, Lezetxiki, inPortugal sowie vielleicht in Süditalien), zeigt, dassdiese kältere Phase nicht so extrem gewesen ist, da siesie überlebt hatten.
So wie beim Wollnashorn kann man auch für viele imSüden vorkommende Mammutreste aus Italien undSpanien annehmen, dass sie ins erste Kältemaximumder letzten Eiszeit zu datieren sind. Die Verbreitungdes Mammuts im Jungpaläolithikum ist auf der Iberi-schen Halbinsel nur für Kantabrien und Nord-katalonien, auf der Apenninenhalbinsel nur fürNorditalien nachgewiesen. Der an mildere Klimateangepasste Waldelefant, Elephas antiquus, scheintebenfalls wie die an mildere Klimate angepasstenNashörner auf der Iberischen Halbinsel bis weit in dieSauerstoff-Isotopenstufe 3 (mindestens ca. 33.000 BP)überlebt zu haben. Auch diese Daten unterstützen dieHypothese, dass das erste Kältemaximum der letztenEiszeit nicht so extrem gewesen ist. Die Präsenz desElephas antiquus nach 40.000 BP auf der IberischenHalbinsel erschwert weiterhin die sichere Bestimmungvon Resten von Elephantidae im Jungpaläolithikumals Mammut, so dass einige Reste aus Portugal alsElephantidae ohne Spezifikation der Art geltenmüssen.
Das Wildschwein, eine Art, die im Jungpaläolithikumimmer als selten gilt, zeigte nach einer näherenBetrachtung, dass sie in Süditalien fast immer in derFauna vertreten ist. In Norditalien, Südostfrankreichund im größten Teil der Iberischen Halbinsel ist esregelmäßig vorhanden, während es in Kantabrien und
Südwestfrankreich selten ist. In Belgien und Deutsch-land wird das Wildschwein häufig zur nacheiszeit-lichen Fauna gerechnet. Obwohl in mehrerenFundstellen Knochen von Wildschweinen durchSchnittspuren von Metallmessern tatsächlich aufVermischungen hingewiesen haben, darf man nichtvergessen, dass diese Tiere in der freien Wildbahnsehr weit wandern und auch in baumlosen Regionenvorkommen können. Daher sollten einzelne Reste vonWildschweinen nicht allzu schnell als Indikatoren fürdichte Wälder interpretiert werden. Seine stärkerePräsenz in Süditalien gegenüber anderen Regionenwie z. B. Südspanien hat wahrscheinlich mit derPräsenz eines ausreichenden Schutzes im Unterholzaufgrund höherer Niederschläge in Süditalien zu tun.Dieses Ergebnis scheint mit der Theorie von DuriFlorineth und Christian Schlüchter (2000) übereinzu-stimmen, die für eine Luftzirkulation der Tiefdruckge-biete im Winter über dem Mittelmeer argumentieren(siehe Seite 40, Abb. 9 B).
Der Rothirsch ist während des Jungpaläolithikumseines der wichtigsten Jagdtiere in Südeuropa gewesen.Seine Verbreitung nach Norden ist zum Teil von derVerbreitung des Waldes und des Rentiers bestimmt.Wenn man bedenkt, dass Rothirsche Wanderungenvon bis zu 140 km unternehmen und dass mancheUnterarten noch heute in waldlosen Regionen leben,ist jedoch eine Interpretation von wenigen Resten vonRothirschen als Indikator für Wald nicht möglich. Erstseine starke Präsenz in der Fauna und in der Kunst inmanchen Fundstellen in Frankreich kann als klarerHinweis auf mildere Klimaverhältnisse betrachtetwerden. In Deutschland ist der Rothirsch fast immerselten und häufig sogar abwesend. In Belgien, wo er infast allen Fundstellen nachgewiesen ist, ist er dennochzahlenmäßig selten.
Die Verbreitung des Damhirsches erscheint nacheiner intensiven Analyse der Literatur unklar. InItalien ist der Damhirsch im Uluzziano undAurignacien häufig. Nach dem zweiten Kältema-ximum ist diese Art so gut wie nie nachgewiesenworden, so dass vermutet wurde, dass sie ausge-storben sei. Die Darstellung eines wahrscheinlichenDamhirsches in Sizilien, die nach typologischenMerkmalen ans Ende des Jungpaläolithikums zustellen ist, sowie die Hinweise aus dem Neolithikumin Süditalien und aus dem Epigravettien in Slowenienlassen jedoch vermuten, dass diese Art auch imJungpaläolithikum noch existierte. In Spanien ist derDamhirsch so gut wie nie im Jungpaläolithikumnachgewiesen worden. Dies ist umso merkwürdiger,als er in Portugal und in Südfrankreich (hier sogar imMagdalénien) bestimmt wurde. Die Seltenheit und das

Mögliche Interpretation der Daten 173
lokale Verschwinden dieser Art könnten zum Teil mitder Tatsache erklärt werden, dass der Rothirsch inSüdeuropa, wo er kleiner ist, zum Teil die ökolo-gische Nische des Damhirsches übernommen unddamit diese Art verdrängt haben könnte.
Das Reh ist sowohl in der Kunst als auch in der Faunaaußerordentlich selten. Nur in Italien und insbe-sondere im Süden scheint es etwas häufiger zu sein.Die Tatsache, dass in Sibirien Rehe Wanderungen vonbis zu 300 km unternehmen, zeigt wie beimWildschwein und beim Hirsch, dass einzelne Restevon Rehen nicht zu schnell als Indikatoren für dichteWälder interpretiert werden dürfen. Seine vereinzeltePräsenz in Mitteleuropa scheint das Bild von Galerie-wäldern entlang der Flüsse zu unterstreichen.
Der Elch, der durch einzelne Funde im Gravettien derBrillenhöhle sowie im Solutréen von Gourdan nachge-wiesen ist, scheint erst ab dem Magdalénien undEpigravettien in Europa verstärkt präsent zu sein. SeinKörperbau sowie die Kenntnisse über sein Verhaltenweisen darauf hin, dass er in der Ebene und hügeligenLandschaften lebte und steile Gebirge mied. Vielleichterreichte er deswegen nie die Iberische Halbinsel, undwahrscheinlich konnte er ebenfalls die Apenninennicht überqueren. Die Verbreitung der Reste und derDarstellungen vom Elch zeigt, dass er in der Regionam Übergang zwischen der Verbreitung des Rothir-sches und des Rentieres lebte. In diesen Breiten lebteebenfalls der Riesenhirsch. Die Verbreitung der Resteund der Darstellungen vom Riesenhirsch zeigen, dasser im gesamten Jungpaläolithikum in Ebenen (u. a. inder Po-Ebene), aber insbesondere in zerklüftetenLandschaften mit zahlreichen unterschiedlichenÖkosystemen (u.a Kantabrien und Pyrenäen) vorkam.Seine Präsenz in Süditalien jenseits der Apenninenzeigt, dass diese für ihn kein Hindernis waren. Ausdem Vergleich der Verbreitung dieser beidenTierarten kommt man zu dem Schluss, dass dieTrockenheit und die Gebirgsketten in Südeuropa denRiesenhirsch vor dem ökologischen Druck des Elchswährend der letzten Eiszeit bewahrt haben. Die klima-tischen Veränderungen am Ende der letzten Eiszeitzwangen sowohl den Elch als auch den Riesenhirschnach Norden. Hier war der Riesenhirsch, aufgrund dererhöhten Feuchtigkeit und des Fehlens von steilenGebirgsketten mit einem nicht allzu extremen Klima,dem vollen Konkurrenzdruck des Elches ausgesetzt.Vielleicht spielte der Konkurrenzdruck des Elcheseine größere Rolle beim Aussterben dieser Art, als dieimmer wieder erwähnte Hypothese, dass sein Geweihunverhältnismäßig groß war.
Das Rentier ist das am meisten gejagte Tier desJungpaläolithikums in Deutschland und Frankreich.Fundstellen mit einem sehr hohen Anteil an Rentierensind in diesen Ländern häufig. Dem gegenüber stehtseine absolute Seltenheit in Italien sowie seine vielgeringere Bedeutung in Kantabrien. Aus Zentral-spanien sind zwar keine Reste vom Rentier bekannt,wobei dort auch überhaupt kaum Fundstellen mitfaunistischen Resten bekannt sind. Drei überzeugendeDarstellungen aus Zentralspanien können als Indizinterpretiert werden, dass auf der Meseta mindestenszuweilen Rentiere gelebt haben könnten.
Auerochse und Steppenbison sind in ihremKnochenbau so ähnlich, dass eine sichere Trennunghäufig schwierig, bisweilen unmöglich ist. Nur inRegionen, in denen man annimmt, dass nur eine derbeiden Arten vorhanden war, kann eine Artbe-stimmung ohne größere Bedenken durchgeführtwerden. Auf der Iberischen Halbinsel ist der Steppen-bison in Kantabrien zahlreich und in Nordkataloniendurch einige Reste nachgewiesen. Im Rest Spaniensund in Portugal scheint er dagegen zu fehlen. DiesesBild wird von der Verbreitung der Darstellungenbestätigt. In Süditalien ist der Steppenbison imUluzziano nachgewiesen, danach scheint er zuverschwinden. Dies könnte, zusammen mit derPräsenz vom Riesenhirsch in denselben Schichteneinen weiteren Hinweis darauf geben, dass die Feuch-tigkeit auf der Apenninenhalbinsel am Ende derSauerstoff-Isotopenstufe 3 und während der Sauer-stoff-Isotopenstufe 2, d.h. während des zweiten Kälte-maximums der letzten Eiszeit, zunahm. Dadurchverbesserten sich die Lebensbedingungen für denAuerochsen und den Rothirsch und verschlechtertensich gleichzeitig für Steppenbison und Riesenhirsch.Die Verbreitung von Steppenbison und Auerochsewären folglich stark voneinander abhängig. InSüdfrankreich lebten Auerochse und Steppenbisonwährend des gesamten Jungpaläolithikums nebenein-ander. In wärmeren Perioden überwog der Auerochse,in kälteren Phasen der Steppenbison.
Der Moschusochse, die einzige der „wolligen“ Tier-arten des Mammuthus-Coelodonta-Faunenkomplexes,die bis in unsere Zeit überlebt hat, erlaubt uns einenEinblick in die sehr fragilen klimatischen Verhält-nisse während der letzten Eiszeit. Sein Vorkommen istein Hinweis auf extrem trockene klimatischeBedingungen und fehlt dementsprechend in Italien.Die verschiedenen Darstellungen dieser Art lassenvermuten, dass der Moschusochse den Menschenbesser bekannt war, als die aufgefundenen Knochen-reste es vermuten ließen.

174 Verbreitung der großen Jagdfauna in Mittel und Westeuropa im oberen Jungpleistozän
Beim Steinbock konnte anhand der Kunst dieTrennung zwischen dem Alpensteinbock (Capra ibex)und dem Iberischen Steinbock (Capra pyrenaica), dieschon in anderen Arbeiten behandelt wurde (u. a.Griggo 1995), dargestellt und diskutiert werden. SeinePräsenz in der Wandkunst bewegt sich zwischenbescheidenen 6,9% im Périgord und 17,2% imQuercy. Da er aber in fast ganz Mittel- undWesteuropa dargestellt wurde, erreicht er mit ca.11,1% die vierte Stelle unter den am meisten darge-stellten Tieren (nach Pferd, Steppenbison undRothirsch). Die Tatsache, dass meistens Steinböckemit stark ausgeprägten Hörnern dargestellt wurden,könnte sowohl rein kulturell bedingt sein als auch vonder Tatsache bestimmt sein, dass Böcke nicht soschnell wie Weibchen und junge Tiere fliehen unddadurch eine leichtere Beute waren.
Gämsen haben eine größere Fluchtdistanz als Stein-böcke und leben oft in sehr unwegsamen Gebieten.Dies könnte ein Grund dafür sein, dass Gämsenseltener gejagt und noch seltener dargestellt wurden.Das anscheinende Fehlen von Gämsen während desJungpaläolithikums in Südspanien, wo der Steinbockbestens vertreten ist, sowie deren Seltenheit könntejedoch auch ein Hinweis darauf sein, dass Steinbockund Gämse teilweise Konkurrenten waren und dassder Steinbock, der von beiden die größere und aggres-sivere Tierart ist und die stärkeren Hörner aufweist,hier die Gämse teilweise verdrängt haben könnte.
Die Saiga-Antilope, eine Steppenart in Osteuropa undSibirien, erreichte während der letzten Eiszeit Mittel-und Westeuropa. Während sie im Südwesten Frank-reichs nur in der Zeit zwischen 18.500 und 12.500Jahren vor heute nachgewiesen ist, erscheint ihreVerbreitung in anderen Regionen komplexer. In dieseRichtung weisen eine Fundstelle aus Südwest-deutschland, zwei Fundstellen aus Belgien und zweiaus den französischen Pyrenäen. Die Seltenheit derjungpaläolithischen Darstellungen von Saiga-Antilo-pen gegenüber Fundstellen, wo diese Art bisweilenden überwiegenden Teil der gejagten Fauna darstellt,weist weiterhin darauf hin, dass diese Art regional undzeitlich sehr große Unterschiede in ihrem Bestandhatte.
Die Verbreitung der Reste und Darstellungen vongroßen Tieren aus der Küstenregion zeigt, dass dieHinweise auf den Küstenlandstrich relativ selten sind,nur weil das Meeresniveau seit der letzten Eiszeit ummehr als 100 m gestiegen ist und damit den überwie-genden Teil der Funde überschwemmt hat. Wenn mandie Verbreitung der Reste von Robben aus dem
Mesolithikum und Neolithikum, aber auch aus ethno-logischen Beispielen von Sammlern, Jägern undFischern aus Europa, Asien oder Amerika, als dasMeeresniveau nicht wesentlich anders als das heutigewar, zum Vergleich nimmt (u. a. Clark 1946; Fisher1995), muss man feststellen, dass alle Fundstellenentlang der Küste bis zu einer Höhe von weniger als100 m liegen. Es liegt in der Natur der Sache, dass dieHinweise auf die Nutzung der Ressourcen des Meeresin der Nähe des Meeres liegen. Wenn man darübernachdenkt, dass bei einem rein hypothetischenMeeresniveau von 100 m über dem heutigen Meeres-spiegel so gut wie keine einzige mesolithische oderneolithische Fundstelle mit Hinweisen auf die großenTiere aus der Küstenregion übrig bliebe, kann mansich die Masse der im Meer versunkenen jungpaläoli-thischen Fundstellen mit Resten und Darstellungenvon Robben, Riesenalken und Waltieren vorstellen.Die auf uns gekommenen Fundstellen dieser Art sindals die Spitze des „Eisbergs“ zu betrachten, demgegenüber man sich das Vielfache an Fundstellenunter Wasser vorstellen muss. Das Verhältnis istwahrscheinlich 1:100, aber auch wenn man von 1:10ausgeht, sind die 34 Fundstellen mit Resten undDarstellungen von Robben stellvertretend für 3400oder auch nur 340, die man im Küstenlandstrich zuerwarten hätte. Dieses Bild ändert wesentlich unsereKonzeption von den Menschen der Eiszeit underlaubt, über die Küstenregion nachzudenken. DerBericht der „Jesup North Pacific Expedition“, die amBeginn des 20. Jahrhunderts die materielle Kultur, dieReligion und die soziale Organisation der Tschukt-schen in Nordostsibirien und der Inuit in Alaska unter-suchte, bietet eine weitere Hilfe auf dem Weg zumVerständnis der Lebensweise der Menschen imJungpaläolithikum Europas. „Even before the arrivalof the Russians, a lively traffic was carried on betweenthe Reindeer Chukchee camps and the maritimevillages (both Chukchee and Eskimo), and alsobetween Asia and America across Bering Strait. TheReindeer people were in need of blubber, thongs, andseal-skins, and would offer in exchange reindeer-skinsand ready-made garments, just as is done at thepresent day“ (Bogoras 1909, 53).
Die Verbreitung der Reste und Darstellungen vongroßen Tieren aus der Küstenregion zeigt weiterhinam deutlichsten, dass die „Trennungslinie“, die vonKantabrien über die Pyrenäen und die Alpen Mittel-und Westeuropa in eine südliche und eine nördlicheRegion teilt, von der Topographie wesentlich geprägtist. Die „nördliche“ Region ist meistens durch Hügel-landschaften, weite Ebenen und wasserreiche Flüssegekennzeichnet. Erst dies ermöglicht es den Robben,flussaufwärts zu schwimmen, und den Menschen,Kontakte über hunderte von Kilometern zu pflegen.Die „südliche“ Region ist dagegen stark zerklüftet, diezum Teil sehr hohen Gebirge reichen oft bis ans Meer,

Mögliche Interpretation der Daten 175
schließen kleinere Regionen ein und können kaumüberquert werden (siehe dazu Djindjian 1994). DieFlüsse am Mittelmeer, außer Ebro, Rhône und Po,führen nur wenig Wasser und bilden erst an ihrenUnterläufen kleinere Ebenen. Bis heute sind keineReste von großen Tieren aus der Küstenregion imInneren Spaniens oder Italiens bekannt, was dagegenspricht, dass diese Tiere in Südeuropa Flüsse aufwärtsgeschwommen sind oder dass es enge Kontaktezwischen Küstenregion und Inland gab.
Es sei mir hier eine letzte Anmerkung im Bezug aufdie Bedeutung der Festlegung der Verbreitung dergroßen Jagdfauna im Jungpaläolithikum erlaubt. DieVerbreitung von Rentier und Mammut ist logischer-weise auch die Verbreitung von Rentiergeweih undElfenbein. Da viele „Technokomplexe“ u. a. durch diePräsenz von organischem Material – sei es alsGeschossspitze, als Schmuck oder als Unterlage fürKunstobjekte – definiert oder mitdefiniert sind,darunter das Aurignacien und das Magdalénien (sieheDjindjian et al. 1999), ist es einleuchtend, dass dasFehlen oder die Knappheit von organischem Rohma-terial entscheidend für die Ansprache eines Techno-komplexes sind.
In dieser Arbeit konnte nur ein Überblick über dieVerbreitung der Hinweise (Faunenreste und Darstel-lungen) auf Tierarten während des Jungpaläoli-thikums in Mittel- und Westeuropa gegeben werden.Die Unerlässlichkeit der Analyse der Umweltbedin-gungen im Allgemeinen und der Verbreitung derTierarten im Speziellen vor jeder Untersuchung derkulturellen Bedeutung der menschlichen Hinterlassen-schaften wird durch diese Arbeit untermauert.
... und die Neandertaler?
Betrachtet man die jüngsten Reste von Elephasantiquus, Stephanorhinus hemitoechus, Stephano-rhinus kirchbergensis und Dama dama, alles typischeVertreter eines gemäßigten Klimas und Relikte ausdem letzten Interglazial, stellt man fest, dass diese inSpanien (El Castillo, Lezetxiki), Portugal (Gruta Novada Columbeira, Lapa da Raínha, Pedreira das Salemas,Figueira Brava, Foz do Exanrique) und Süditalien(Grotta della Cala) bis weit ins Isotopen-Stadium 3überlebt haben. Sie sind erst mit dem Auftreten vonHomo sapiens und des ihm zugeordneten Technokom-plexes des Aurignaciens ausgestorben (endgültigElephas antiquus, Stephanorhinus hemitoechus undStephanorhinus kirchbergensis, regional Dama dama).In denselben Regionen hat ebenfalls bis weit in dasIsotopen-Stadium 3 auch eine andere Art gelebt, diedes Homo neanderthalensis. Die jüngsten Knochen-funde dieser Art, aber noch mehr die Technokom-
plexe, die ihr zugeordnet werden und daher auf ihrePräsenz hinweisen (spätes Moustérien in Portugal undSüdspanien, Châtelperronien in Nordspanien undUluzziano in Italien), decken sich erstaunlich gut mitder Verbreitung der gerade erwähnten Vertreter einesgemäßigten Klimas. Das Aussterben der Neandertalervollzieht sich ebenfalls mit dem Auftreten von Homosapiens und des ihm zugeordneten Technokomplexesdes Aurignaciens. Dies erlaubt die Neandertaler nicht„nur“ als Menschen, sondern auch als Teil ihrerUmwelt zu betrachten.
Die Idee, dass die „Folge“ des Homo sapiens aufden Homo neanderthalensis zu einem bedeutendenTeil vom Klima abhängig gewesen sei, wurde schonfür einige Fundstellen in Israel formuliert, u. a.Qafzeh, Skhul, Amud und Kebara (Gibbons 2003;Bar-Yosef 1989). Hier jedoch ist der Neandertaler derVertreter eines kühleren, der moderne Mensch derVertreter eines gemäßigten bis warmen Klimas. Auchfür Mitteleuropa wird die enge Verbindung zwischender Verbreitung des Neandertalers und von dessenTechnokomplexen mit den Klimaveränderungen,insbesondere mit der Südverschiebung des Nordeuro-päischen Eisschildes als Modell diskutiert (Jöris2002). Es ist jedoch wichtig zu vermerken, dass derHomo sapiens, eine Art, die während mehrererzehntausende von Jahren in warmen und milderenRegionen gelebt hatte, nach Südeuropa von Nordenher kam. In Mitteleuropa, in Norditalien undNordspanien sind die ältesten Fundschichten desAurignaciens um ca. 35.000–40.000 BP datiert(Conard und Bolus 2003; Richter 1996; Bischoff et al.1989). Der älteste datierte Fund der Art Homo sapiensin Europa, Pestera cu Oase in Rumänien, ist zwischen34.000 und 36.000 Jahren datiert (Trinkaus et al.2003a und 2003b). In Spanien jedoch, südlich der sogenannten „Ebro-Grenze“ (Zilhão 1993), sind diejüngsten Funde eines Neanderthalers in der Höhle vonZafarraya um ca. 27.000 BP datiert. In Süditalien, inder Höhle von Castelcivita, konnte der Übergangzwischen den Schichten des Uluzziano und desAurignaziano um ca. 32.000 Jahren datiert werden(Mussi 2001, 172–187). Gleichzeitig ist zu erwähnen,dass die ältesten 14C-Daten des Aurignaciens südlichdes Ebro und in Süditalien zwischen 32.000 und28.000 Jahren liegen und damit deutlich jünger als dieDaten nördlich der „Ebro-Grenze” und in Norditaliensind. Daher kommt der Homo sapiens nach Südeuropaals Vertreter eines kalten Klimas, während die letztenAngehörigen der Art Homo neanderthalensis inSüdeuropa als Vertreter eines gemäßigten Klimas zubetrachten sind. Würde man das Aussterben desNeandertalers und die Ankunft des modernenMenschen als das Verhältnis innerhalb eines Arten-paares betrachten, also zwischen zwei Arten dergleichen Familie, in diesem Fall sogar der gleichenGattung, die teilweise in Konkurrenz zueinander

176 Verbreitung der großen Jagdfauna in Mittel und Westeuropa im oberen Jungpleistozän
stehen, würde dies bedeuten können, dass der Homosapiens, eine Art, die wie schon erwähnt bestens anmildere und wärmere Klimaverhältnisse, sowie aneine entsprechende Fauna angepasst war, den Homoneanderthalensis vielerorts abgelöst hat, so wie es fürviele andere Artenpaare gezeigt werden konnte.Beispielweise liegen die Vorteile des Equus ferusgegenüber dem Equus hydruntinus in seinem größerenKörperbau, in seiner höheren Aggressivität und seinerweit überlegenen Geschwindigkeit. Die Vorteile desHomo sapiens gegenüber dem Homo neandertha-
lensis lagen wahrscheinlich eher in der kulturellenKomponente, in der unterschiedlichen Steinbear-beitung, in den Jagdwaffen, in den Jagdstrategien,vielleicht sogar in einer unterschiedlichen sozialenStruktur. Da jedoch in dieser Arbeit auf eine Analyseder kulturellen Aspekte bewusst verzichtet wurde(siehe das Unterkapitel Forschungsziel, S. 1), konntedas Verhältnis Homo sapiens zu Homo neandertha-lensis hier nicht näher untersucht werden und bleibtein Ausblick für die Zukunft.

Zusammenfassung
Für die Übersetzung und Korrekturen möchte ich mich hier bei Anna Budweg, Emma Dalmau, Prof. PaulGoldberg, Prof. Anthony E. Marks, Luc Moreau, Céline Richard und Prof. Narcis Soler herzlich bedanken.
Durch eine intensive Literaturarbeit und Kontakte mitKollegen im In- und Ausland konnten die Forschungs-geschichte sowie der heutige Forschungsstand imBereich der älteren Urgeschichte in Mittel- undWesteuropa untersucht und hier dargestellt werden.Trotz der unvermeidlichen Vereinfachung war dieserTeil der Arbeit notwendig, um die verwendeten Datenüber die Verbreitung der großen Jagdfauna im Jungpa-läolithikum in Mittel- und Westeuropa unter einemforschungsgeschichtlichen Blickwinkel kritischbetrachten zu können. Der deutliche Unterschied inden errechneten Planstellen zwischen Spanien, Italien,Frankreich und Deutschland ist nicht nur das Ergebnisvon unterschiedlichen Systemen, sondern weist auchauf den unterschiedlichen Stellenwert der älterenUrgeschichte in den verschiedenen Ländern hin underlaubt ebenfalls Schlussfolgerungen für die nächsteZukunft (S. 5–23).
Eine detaillierte Übersicht über die Geographie unddas Klima Europas diente ebenfalls als Voraus-setzung, um die Verbreitung der großen Jagdfauna imJungpaläolithikum in Mittel- und Westeuropa auseiner ökologischen Perspektive kritisch betrachten zukönnen (S. 25–37). Die Küstenregion und derenVeränderungen konnten kartiert und in ihrerBedeutung hervorgehoben werden.
Der Mensch als Jäger konnte sich mindestens seit400.000 Jahren hölzerner Lanzen und Wurfspeere beider Jagd bedienen. Im Jungpaläolithikum kamen dieSpeerschleuder und am Ende Pfeil und Bogen hinzu.Dadurch hatte er das Potential, jedes Tier zu töten. DieTatsache, dass in jungpaläolithischen Darstellungenfast alle Tierarten einmal von Geschossen durchbohrterscheinen und dass fast alle Tiere auf Speer-schleudern abgebildet sind, zeigt, dass dieses Potentialauch in die Tat umgesetzt wurde. Dadurch stand derpaläolithische Mensch am oberen Ende der Nahrungs-kette (S. 39–48).
Zum ersten Mal wurden so gut wie alle publiziertenDarstellungen von „seltenen“ Tierarten unter dergroßen Jagdfauna zusammengetragen: Equus hydrun-tinus, Sus scrofa, Dama dama, Capreolus capreolus,Alces alces, Megaloceros giganteus, Ovibosmoschatus, Rupicapra rupicapra, Saiga tatarica,Phocidae, Cetacea und Pinguinus impennis.
Die Analyse der Verbreitung der großen Jagdfaunaanhand der faunistischen Reste sowie der Darstel-lungen hat Indizien für zahlreiche Hypothesengeliefert (S. 49–145). Die Verbreitung jeder einzelnenTierart wurde im Vergleich zur Verbreitung deranderen Tierarten diskutiert. Dadurch erwies sich diebesondere Bedeutung von Tierarten, die zur gleichenFamilie, teilweise zur gleichen Gattung gehören,jedoch unterschiedliche, in den Überlappungsgebietenteilweise konkurrierende ökologische Anpassungenaufweisen, als ausschlaggebend. Diese sind: Equusferus und Equus hydruntinus, Coelodonta antiquitatisund Stephanorhinus kirchbergensis / Stephanorhinushemitoechus, Mammuthus primigenius und Elephasantiquus, Cervus elaphus und Rangifer tarandus,Alces alces und Megaloceros giganteus, Bison priscusund Bos primigenius, Rupicapra rupicapra und Caprapyrenaica / Capra ibex.
Besonders interessant war es aufzuzeigen, dasswährend der OIS 4 (Sauerstoff-Isotopenstufe 4), demersten Kältemaximum der letzten Eiszeit, dieVerbreitung von Tierarten wie Mammut undWollnashorn bis nach Südspanien und Süditalienreichte, ohne dass jedoch die Waldelefanten, dieWald- und Steppennashörner ausstarben. In denRegionen südlich von Kantabrien wurden aus der Zeitdes Jungpaläolithikums außer einigen sehrwahrscheinlichen Bildern von Rentieren nur zweifel-hafte Bestimmungen von Vertretern des Mammuthus-Coelodonta-Faunenkomplexes entdeckt. In Italien sindkaum jungpaläolithische Reste und keine Darstel-lungen von Mammuthus primigenius, Coelodontaantiquitatis oder Rangifer tarandus entdeckt worden.
Das Aussterben von Waldelefanten sowie Wald-und Steppennashörnern scheint sich erst nach 30.000Jahren vor heute ereignet zu haben. Um ca. 30.000sollen auch Steppenbison und Riesenhirsch ausSüditalien verschwunden sein. Beginnend um dieseZeit und spätestens mit dem zweiten Kältemaximumder letzten Eiszeit (OIS 2) nahm die Zahl derDamhirsche sehr stark ab, vielleicht starb er inSüdwesteuropa sogar aus. Dies ist das Bild einerstarken Verarmung der größeren Jagdfauna inSüdeuropa. Zu den in Südeuropa, am Ende der OIS 3ausgestorbenen Arten könnte man ebenfalls den Homoneanderthalensis hinzurechnen, der jedoch in dieserArbeit nicht untersucht wurde.

178 Verbreitung der großen Jagdfauna in Mittel und Westeuropa im oberen Jungpleistozän
Die Analyse der bescheidenen Hinweise auf größereTiere aus den Küstenlandstrichen zeigt nicht nur, dassdiese Region vom Meer überschwemmt wurde (derMeeresspiegel ist seit dem zweiten Kältemaximum derletzten Eiszeit um mehr als 100 m gestiegen), sondernauch, dass sie in der bisherigen Forschung oft einfachvergessen wurden. Die wenigen Fundstellen mitFunden und Befunden dieser Art sind jedoch als Stell-vertreter für eine viel höhere Anzahl an solchenFundstellen, die heutzutage unter Wasser liegen, zuverstehen (S. 147–165).
Schließlich hat sich bei der Interpretation der Datengezeigt, dass eine direkte Beziehung zwischen demnatürlichen Vorkommen einer Art und ihrer Darstel-lung existiert. In der Wandkunst konnte bewiesen wer-den, dass das Zusammenkommen von Tierarten vielmehr von den Umweltbedingungen als von jeglicherkulturellen Komponente abhängig ist (S. 167–175).
Als Ausblick wurde die Hypothese formuliert, dassdas Schicksal des Artenpaares Homo sapiens undHomo neanderthalensis, so wie das vieler andererArtenpaare, großenteils auf die klimatischen undökologischen Änderungen in Westeuropa in der Zeitzwischen 40.000 und 25.000 BP zurückzuführen ist.
Abstract
The status of current research about the distribution oflarge game during the Upper Paleolithic in Middle andWestern Europe and its history can be only investi-gated and presented through intensive literary researchand contact with colleagues locally and abroad. Inspite of the unavoidable simplification about thehistory of this research, this portion of the work wasnecessary, in order to be able to critically consider thedata. The marked difference in the number ofestablished research positions for the study of thePaleolithic in Spain, Italy, France, and Germany is notonly a consequence of differing research systems andresearch institutions, it reflects the importance givento Paleolithic research in each of the differentcountries and also permits predictions about futurereserches (pp. 5–23).
Likewise, a detailed overview of the topography andthe climate of Europe, as well as the environnmentalchanges that occurred, serve as a basis to criticallyevaluate the distribution of large game during theUpper Paleolithic in Middle and Western Europe froman ecological point of view (pp. 25–37). Theimportance of the coastal region and its sea levelchanges have been mapped and emphasized.
For at least 400 kya, especially after the appearance ofwooden lances and throwing spears used for hunting,human beings have been considered hunters. Duringthe Upper Paleolithic, the spear thrower wasdeveloped and later was followed by the bow andarrow. Thus, there then existed the ability to kill anyanimal. Many drawings and engrawings from theUpper Paleolithic show scenes in which almost allspecies of animals are pierced by projectiles. Inaddition almost all species have been depicted onspear throwers. This place the Upper Palaeolithic manat the top of the ecological pyramid (pp. 39–47).
For the first time, many of the published depictions ofuncommon animal species are brought together:Equus hydruntinus, Sus scrofa, Dama dama,Capreolus capreolus, Alces alces, Megalocerosgiganteus, Ovibos moschatus, Rupicapra rupicapra,Saiga tatarica, Phocidae, Cetacea and Pinguinusimpennis.
The analysis of the distribution of large game inMiddle and Western Europe, on the basis of faunalremains, as well as artistic depictions, provides fornumerous hypotheses (especially 49–145). The distri-bution of specific species, in comparison to that ofother species is discussed, such as the significance ofanimals in overlapping regions that belong to the samefamily and, sometimes, to the same genus. Theseanimals exhibited competition-driven ecologicaladaptations as seen between the Equus ferus andEquus hydruntinus, Coelodonta antiquitatis andStephanorhinus kirchbergensis and Stephanorhinushemitoechus, Mammuthus primigenius and Elephasantiquus, Cervus elaphus and Rangifer tarandus,Alces alces and Megaloceros giganteus, Bison priscusand Bos primigenius, Capra pyrenaica and Capra ibexand Rupicapra rupicapra.
It is especially significant that during OIS4(oxygen isotope stage), the first glacial maximum ofthe last ice age, the distribution of species such asmammoth and woolly rhino extended as for south assouthern Spain and southern Italy, without theextinction of the forest elephant, forest rhino, andsteppe rhino, which ware living to these temperateregions. Aside from a few definitive pictures ofreindeer, researchers have made only a few ambiguousidentifications from the „mammuthus-coelodonta“faunal complex during the time of the Upper Paleo-lithic, south of Cantabria. There are few Upper Paleo-lithic remains that have been discovered in Italy andno representations of Mammuthus primigenius, Coelo-donta antiquitatis or Rangifer tarandus are reported.
The extinction of the forest elephant, as well as theforest and steppe rhino, did not occur until shortlyafter 30 kya. At about the same time, the Europeanbison and the giant deer disappeared from southernItaly. Beginning around this period, and no later than

Zusammenfassung 179
the second glacial maximum (OIS2) of the last iceage, the number of fallow deer declined sharply andeven may have completely died out in southwesternEurope. This sugeste a strong impoverishment of thelarger game in southern Europe. Though not analyzedin this study, one could also add Homo neandertha-lensis to the list of these extinct animals.
The analysis of the few indications of the largeranimals in the coastal tract shows that this region wasnot only later submersed by the sea (sea level has risenmore than 100 m) but also that researchers have oftenforgetten about them. The few sites with elementsfrom the coast, must only represente a small numberof such original sites that currently lie underwater(pp.147–165).
In conclusion, the interpretation of the data shows thata direct relationship exists between the natural appea-rance of a species and its artistic representation.Therefore, both the presence of the species and theirrepresentations are more dependent on environmentalcomponents than from any cultural components (pp.167–175).
Finally, this hypothesis would be formulated asfollows, that the destiny of Homo sapiens and Homoneanderthalensis, as well as other linked species, isstrongly influenced by factors such as climatic andecological changes during the period between 40 and25 kya.
Résumé
Partant d’une recherche intensive au sein de lalittérature completée par les informations fournies pardes collègues en Allemagne et à l’étranger, nous avonspu dresser un tour d’horizon de l’histoire desrecherches d’une part, de l’état actuel de la recherchesur le Paléolithique en Europe centrale et occidentaled’autre part. Bien que ce thème n’ait été traité que defaçon succincte, cette analyse a permis de traiter demanière critique les informations sur la distribution dela macrofaune chassée au cours du Paléolithiquesupérieur à la lumière de l’historique de la recherche.L’importante différence dans le nombre de places aucontrat à durée indéterminée en Espagne, Italie,France et Allemagne n’est pas tant le résultat desystèmes différents, que de la valeur accordée auPaléolithique dans ces différents pays. C’est pourtantlà un élément déterminant de la recherche à courtterme (pp. 5–23).
Une vue d’ensemble préalable sur la géographie et leclimat en Europe d’un point de vue écologique futnécessaire pour pouvoir traiter les informations sur ladistribution de la macrofaune chassée au cours du
Paléolithique supérieur en Europe centrale etoccidentale (pp. 25–37). La région côtière et seschangements sont présentés sous forme de cartes etleur importance a pu être mise en évidence.
L’homme chasseur a été dès 400.000 ans en mesure dese servir de lances en bois pour la chasse. Par la suite,au cours du Paléolithique Supérieur, vinrent s’ajouterle propulseur et, plus récemment, l’arc-à-flèches.Ainsi il a été très tôt en position de force par rapport àl’animal. Le fait que dans les représentations figuréesdu Paléolithique supérieur presque tous les espècesanimales soient représentées au moins une foistouchés par une flèche ou représentés sur des propul-seurs montre que le potentiel a également porté à sesfins. L’homme du Paléolithique était donc sur la mar-che la plus haute de la chaîne alimentaire (pp. 39–47).
Pour la première fois, toutes les représentations desanimaux «rares» parmi la macrofaune chassée ont étéprésentées ensemble : Equus hydruntinus, Sus scrofa,Dama dama, Capreolus capreolus, Alces alces,Megaloceros giganteus, Ovibos moschatus, Rupicaprarupicapra, Saiga tatarica, Phocidae, Cetacea etPinguinus impennis.
L’analyse de la distribution de la macrofaunechassée à partir des restes fauniques et des représenta-tions a livré des éléments pour de nombreuseshypothèses (en particulier pp. 49–145). La distri-bution de chaque espèce animale a été discutée et leurimportance mise en évidence. Elle nous montre, parailleurs, que des espèces d’une même famille, parfoisdu même genre, ayant des habitudes différentes, setrouvent en concurrence là où leur distribution sesuperpose. Celles ci sont: Equus ferus et Equushydruntinus, Coelodonta antiquitatis et Stephano-rhinus kirchbergensis / Stephanorhinus hemitoechus,Mammuthus primigenius et Elephas antiquus, Cervuselaphus et Rangifer tarandus, Alces alces et Megalo-ceros iganteus, Bison priscus et Bos primigenius,Rupicapra rupicapra et Capra pyrenaica / Capraibex.
Il est particulièrement intéressant de constaterqu’au cours du OIS 4 (stade isotopique d’oxygène 4),correspondant au premier maximum glaciaire de ladernière glaciation, le mammouth et le rhinocéroslaineux ont atteint le sud de l’Espagne et de l’Italiesans pour autant provoquer l’extinction de l’éléphantantique, du rhinocéros de Merk et du rhinocéros desprairies. Pour la période du Paléolithique supérieur, ausud de la chaîne cantabrique, seules des représenta-tions douteuses de la faune froide ont été signalées, àl’exception de quelques représentations probables derenne. En Italie, la pauvreté de restes fauniques etl’absence de représentations figurées de Mammuthusprimigenius, Coelodonta antiquitatis ou Rangifertarandus ont été mises en évidence.

180 Verbreitung der großen Jagdfauna in Mittel und Westeuropa im oberen Jungpleistozän
L’extinction de l’éléphant antique, du rhinocéros deMerk et du rhinocéros des prairies semble s’êtreproduite après 30.000 ans BP. Aux alentours de30.000 BP le bison et le mégacéros semblent eux aussiavoir disparu de l’Italie méridionale. Au plus tard avecle deuxième maximum glaciaire (OIS 2) le nombre dedaims se réduisit au point qu’ils disparurent peut-êtred’Europe occidentale. Parmi les espèces qui ont subileur extinction pendant la phase OIS 3, il nous fautencore signaler l’Homo neanderthalensis qui n’acependant pas été traité dans cette œuvre.
L’analyse des rares témoins de la région côtièremontre que cette région a non seulement étésubmergée par la mer (le niveau de la mer aujourd’huise trouve à plus de 100 mètres plus haut qu’aux tempsdu deuxième maximum glaciaire de la dernièreglaciation), mais qu’elle a été quasiment oubliée par larecherche archéologique. Les quelques sites qui ontlivré des restes fauniques ou des représentationsd’animaux marins doivent donc être considéréscomme représentatifs d’un nombre bien plus grand desites de ce type, qui aujourd’hui se trouvent sous lamer (pp.147–165). Enfin, nous avons pu montrer qu’il existe une relationdirecte entre la distribution d’une espèce animale et ladistribution de ses représentations. Ainsi dans l’artpaléolithique la composition des tableaux dépenddavantage d’une composante naturelle que d’unequelconque composante culturelle (pp. 167–175).
Comme perspective nous avons formulél’hypothèse que le sort du couple des espèces Homosapiens et Homo neanderthalensis, comme beaucoupde couples d’animaux par ailleurs, est à mettre enrelation, pour une bonne part, avec les changementsécologiques qui se déroulèrent en Europe occidentaleentre 40.000 et 25.000 BP.
Riassunto
Attraverso un intenso lavoro di ricerca fondato sullaletteratura cosí come grazie a contatti con colleghi inGermania ed all’estero è stato possibile presentare lastoria e l’attuale stato della ricerca nel campo dellapaleoetnologia in Europa centrale ed occidentale.Nonostante l’indispensabile stringatezza, questa partedel lavoro è stata necessaria per poter comprendere idati sulla distribuzione della fauna di grandi dimen-sioni cacciata durante il Paleolitico Superiore inEuropa centrale ed occidentale anche alla luce dellastoria della ricerca scientifica. Le differenze sostan-ziali tra il numero di posti di lavoro a tempo indeter-minato calcolato in modo approssimativo per laSpagna, l’Italia, la Francia e la Germania non è solo ilrisultato di sistemi amministrativi differenti ma
evidenzia il differente valore dato alla Paleoetnologianei differenti paesi e permette di farsi un’idea suglisviluppi futuri (pp. 5–23).
Una sintesi dettagliata della topografia e del clima inEuropa centrale ed occidentale è servita comepremessa per esaminare in modo critico la distribu-zione della grande fauna cacciata durante il Paleo-litico Superiore sotto il punto di vista dell’ecologia(pp. 25–37). La regione costiera ed i suoi cambia-menti sono stati presentati in forma di carte geogra-fiche ed in questo modo è stato possibile sottolinearela loro importanza.
L’uomo quale predatore si è potuto servire di lance dilegno nella caccia da almeno 400.000 anni. Nel Paleo-litico Superiore si sono aggiunti a queste propulsori e,nel periodo finale, anche arco e freccia. Tramitequeste armi l’uomo aveva il potenziale di ucciderequalsiasi animale. Il fatto che quasi ogni specieanimale é stata rappresentata almeno una volta colpitada freccie o lance ed inoltre il fatto che quasi tutte lespecie animali sono state rappresentate sui propulsori,dimostra, che questo potenziale è stato anche tradottoin fatti. È quindi lecito vedere l’uomo del Paleoliticosul piú alto gradino della scala alimentare (pp. 39–47).
Per la prima volta sono state presentate assieme laquasi totalitá delle immagini dei cosiddetti “animalirari”: Equus hydruntinus, Sus scrofa, Dama dama,Capreolus capreolus, Alces alces, Megaloceros gigan-teus, Ovibos moschatus, Rupicapra rupicapra, Saigatatarica, Phocidae, Cetacea e Pinguinus impennis.L’analisi della distribuzione della grande fauna sullabase dei resti di fauna e delle rappresentazioni hamesso in evidenza indizi per numerose ipotesi (vederein particolare pp. 49–145). La distribuzione di ognisingola specie animale è stata discussa comparandolacon la distribuzione delle altre specie. In questo modoè stato possibile sottolineare l’importanza di animaliche appartengono alla stessa famiglia, alle volte addi-rittura allo stesso genere, e che mostrano capacitá diadattamento in parte differenti ma in parte anche con-correnziali nelle zone in cui le loro aree di distribu-zione si sovrappongono. Queste coppie di animalisono: Equus ferus e Equus hydruntinus, Coelodontaantiquitatis e Stephanorhinus kirchbergensis / Stepha-norhinus hemitoechus, Mammuthus primigenius e Ele-phas antiquus, Cervus elaphus e Rangifer tarandus,Alces alces e Megaloceros giganteus, Bison priscus eBos primigenius, Rupicapra rupicapra e Capra pyre-naica / Capra ibex. Si è dimostrato particolarmente interessante poterdimostrare che durante la OIS 4 (stadio isotopicodell’ossigeno 4), il primo massimo glaciale dell’ultimaglaciazione, la distribuzione del mammut e delrinoceronte lanoso abbia raggiunto sia il sud della

Zusammenfassung 181
Spagna che il sud dell’Italia senza comunque portareall’estinzione ne l’elefante antico, ne le due specietemperate di rinoceronte, il rinoceronte delle steppe edil rinoceronte della foresta. Durante il paleoliticosuperiore invece, a sud della cordigliera cantabrica, aparte alcune chiare rappresentazioni di renna, sonostate trovate solo dubbiose immagini di rappresentantidella fauna fredda. In Italia sono stati trovatipochissimi resti e nessuna rappresentazione del Paleo-litico Superiore di mammut, rinoceronte lanoso erenna.
L’estinzione dell’elefante antico cosí come delrinoceronte delle steppe e della foresta sembra esserepiú recente di 30.000 anni BP. Nello stesso periodosembra che anche il bisonte ed il megacero sianoscomparsi dal Suditalia. A partire da ca. 30.000 anniBP ed al piú tardi con il secondo massimo glaciale(OIS 2) pare che anche il daino sia diventato moltoraro o che sia addirittura scomparso nell’Europasudoccidentale. Questo è il panorama di un forteimpoverimento della grande fauna nell’Europa delsud. Nella lista delle specie estintesi nella parte finaledel periodo OIS 3 ci sarebbe anche da aggiungerel’Homo neanderthalensis, che comunque non è statotrattato in questo lavoro.
I modesti indizi riguardo ai grandi animali viventinelle zone costiere mostrano che questa regione nonsolo è stata sommersa dalle acque del mare (il livellodel mare è salito rispetto all’ultimo massimo glacialedi oltre 100 metri), ma anche che questa regione èstata spesso dimenticata dai ricercatori. I pochi siti conreperti di questo tipo sono da considerare anche comevicari del numero estremamente maggiore di sitiscomparsi sotto il mare (pp. 147–165).
Infine è stato possibile dimostrare che esiste unrapporto diretto tra la distribuzione di una specieanimale e la distribuzione delle sue rappresentazioni.Grazie all’analisi fattoriale delle corrispondenze èstato possibile provare, che la correlazione di specieanimali nell’arte parietale dipende molto di piú dafattori ambientali che da fattori culturali (pp. 167–175).
Sulla base dei dati sulla distribuzione ed in partesull’estinzione dei grandi mammiferi del PaleoliticoSuperiore in Europa centrale ed occidentale è statopossibile formulare l’ipotesi tutta da verificare e daportare avanti, che anche la sorte della coppia Homosapiens e Homo neandertalensis sia da ricondurre ingran parte ai cambiamenti ecologici avvenuti inEuropa occidentale tra 40.000 e 25.000 anni BP.
Resumen
Ha sido posible presentar los antecedentes y tambiénel estado actual de la investigación en lo que se refierea la Paleoetnología paleolítica en Europa central yoccidental a través de un intenso trabajo en la consultade textos y contactos con diferentes colegas, tantoalemanes como de otros países. Aunque breve, estaparte del trabajo ha sido necesaria para permitir unamejor comprensión de todo lo que afecta a la distri-bución de los grandes mamíferos cazados durante elPaleolítico Superior en Europa central y occidental taly como resulta a la luz de la investigación científica.Si observamos las diferencias en el estado de lainvestigación y las confrontamos con el número depuestos estables ocupados en el sector del Paleolíticoen España, Italia, Francia y Alemania, a pesar de lafuerte aproximación, estas diferencias nos demuestranque no son sólo el resultado de sistemas administra-tivos distintos, sino también del hecho de que el valorque se da al Paleolítico en los diferentes países esdesigual. Todo ello nos permite hacernos una ideasobre el futuro desarrollo de la investigación (pp. 5–23).
Una síntesis detallada de la topografía y del clima enEuropa central y occidental sirvió de premisa paraexaminar de forma crítica la distribución de la granfauna cazada durante el Paleolítico superior tambiéndesde el punto de vista de la ecología (pp. 25–37). Sehan presentado las regiones costeras y sus cambiosmorfológicos mediante mapas, remarcando así suimportancia.
El hombre cazador se sirve de lanzas de madera paracazar ya desde hace unos 400.000 años. Durante elPaleolítico superior se añadieron a éstas los propul-sores y a finales del mismo periodo el arco y lasflechas. Con estas armas el hombre acrecienta supotencial de muerte sobre cualquier tipo de animal. Laexistencia de representaciones de casi todos los tiposde animales alcanzados con flechas o lanzas y elhecho que casi todos ellos se encuentran figuradoscomo decoración en propulsores demuestra que estepotencial se tradujo en hechos. Es por tanto lícitoconsiderar el hombre del Paleolítico en el más altopeldaño de la cadena alimenticia (pp. 39–47).
Por primera vez se han presentado conjuntamente lacasi totalidad de imágenes de “animales raros”: Equushydruntinus, Sus scrofa, Dama dama, Capreoluscapreolus, Alces alces, Megaloceros giganteus,Ovibos moschatus, Rupicapra rupicapra, Saigatatarica, Phocidae, Cetacea y Pinguinus impennis.

182 Verbreitung der großen Jagdfauna in Mittel und Westeuropa im oberen Jungpleistozän
El análisis de la distribución de los grandes mamíferosen base a los hallazgos y a las representaciones de lamisma ha puesto en evidencia indicios para numerosashipótesis (pp. 49–145). La distribución de cada uno delos animales por sí mismos se discutió comparándolacon la de los otros animales. Así se ha puesto enevidencia la importancia específica de los animalespertenecientes a la misma familia, incluso al mismogénero, y que demuestran capacidades de adaptacióndistintas, pero que entran en competición entre ellosen las zonas donde su distribución se sobrepone. Estasparejas de animales son: Equus ferus y Equus hydrun-tinus, Coelodonta antiquitatis y Stephanorhinus kirch-bergensis / Stephanorhinus hemitoechus, Mammuthusprimigenius y Elephas antiquus, Cervus elaphus yRangifer tarandus, Alces alces y Megalocerosgiganteus, Bison priscus y Bos primigenius, Rupi-capra rupicapra y Capra pyrenaica / Capra ibex.
Se ha demostrado particularmente interesante elponer en evidencia que durante el OIS 4 (estadioisotópico del oxígeno 4), que corresponde al primermáximo glacial de la última glaciación, la distri-bución del mamut y del rinoceronte lanudo llegó hastael sur de España y Italia sin por ello provocar laextinción del elefante antiguo, Elephas antiquus, y delas dos especies de rinoceronte de clima templado:Stephanorhinus kirchbergensis y Stephanorhinushemitoechus. Durante el Paleolítico superior, alcontrario, al sur de la cordillera Cantábrica, apartealgunas claras representaciones de reno, se han encon-trado sólo dudosas imágenes de fauna fría. En Italiahan aparecido poquísimos restos y ninguna represen-tación de mamut, rinoceronte lanudo y reno en elPaleolítico superior.
La extinción de Elephas antiquus, Stephanorhinuskirchbergensis y Stephanorhinus hemitoechus pareceser más reciente de 30.000 años BP. Durante el mismoperiodo desaparecieron también del sur de Italia elBison priscus y el Megaloceros giganteus. A partir de30.000 BP y a lo más tarde durante el OIS 2, que secorresponde con el segundo máximo glacial de laúltima glaciación, Dama dama fue desapareciendohasta su posible extinción en Europa sur occidental.Este es el panorama de un fuerte empobrecimiento dela fauna mayor en Europa meridional. En la lista de lasespecies extintas en la parte final del periodo OIS 3podría añadirse también el Homo neanderthalensis,que sin embargo no es tratado en este trabajo. Losmodestos indicios respecto a los grandes animales quevivían en las zonas costeras demuestran que el pococonocimiento de esta zona se debe no sólo a la subidade la barrera litoral en una cota unos 100 metrosrespecto al máximo glacial, sino también a que losinvestigadores la han tenido muy olvidada. Los pocosyacimientos existentes de este tipo debemos conside-
rarlos como representantes del número extrema-damente mayor de los que desaparecieron bajo la mar(pp. 147–165).
En fin, ha sido posible demostrar que existe un nexoentre la distribución de una especie animal y sus repre-sentaciones. Gracias al „análisis factorial de corres-pondencias“ ha sido posible probar que la correlaciónde especies animales en el arte parietal depende másde factores ambientales que de factores culturales (pp.167–175).
En base a los datos sobre la distribución y en partesobre la extinción de los grandes mamíferos del Paleo-lítico en Europa central y occidental ha sido posibleformular la hipótesis, a verificar y profundizar, que eldestino de la pareja de Homo sapiens y Homoneanderthalensis puede ser en buena parte reconsi-derado de acuerdo con los cambios ecológicos quetuvieron lugar entre hace 40.000 y 25.000 años BP.
Resum
Ens ha estat possible presentar la història i l'estatactual de la investigació de la paleoetnologiapaleolítica d’Europa central i occidental mitjançant untreball intensiu, a través de la investigació de textos idels contactes amb investigadors alemanys i d'altrespaïsos. Malgrat la indispensable brevetat d’aquestcapítol, aquesta part del treball ha estat necessària perpermetre una comprensió crítica de la distribució delsgrans mamífers caçats durant el paleolític superior aEuropa central i occidental, feta també a la llum de lahistòria de la investigació científica. Les diferènciesen aquesta confrontades amb el nombre de placesestables ocupades en el sector del paleolític a Espanya,Itàlia, França i Alemanya, malgrat la forta aproxi-mació, ens demostren que tals diferències no sónnomés el resultat de sistemes administratius diferents,sinó degudes també al valor diferent que es reconeix alpaleolític en cada un dels països. A més tot això enspermet fer-nos una idea del futur desenvolupament dela investigació (pp. 5–23).
Una síntesi detallada de la topografia i del clima aEuropa central i occidental ha servit per examinar demanera crítica la distribució de la fauna de gransdimensions caçada durant el paleolític superior tambédes del punt de vista de l'ecologia (pp. 25–37). Lazona litoral i els seus canvis morfològics han estatpresentats sota forma de mapes, remarcant així la sevaimportància.
L’home caçador es serveix de llances de fusta ja desde fa uns 400.000 anys. Durant el paleolític superiores varen afegir a las llances els propulsors i a finals del

Zusammenfassung 183
mateix període els arcs i fletxes. D’aquesta maneral’home podia caçar qualsevol tipus de animal.L'existència de representacions de quasi tots els tipusd'animals afectats per fletxes o bé llances, i el fet quequasi tots ells es trobin representats com a decora-cions en els propulsors demostra que aquell potenciales va traduir també en fets. Podem per tant situarl’home del paleolític en el graó més alt de la cadenaalimentària (pp. 39–47).
Per primera vegada s'han presentat conjuntament quasila totalitat d’imatges “d’animals rars”: Equus hydrun-tinus, Sus scrofa, Dama dama, Capreolus capreolus,Alces alces, Megaloceros giganteus, Ovibosmoschatus, Rupicapra rupicapra, Saiga tatarica,Phocidae, Cetacea i Pinguinus impennis. L’anàlisi dela distribució dels grands mamífers, prenent com apunt de partida les seves troballes i representacions,ens ha obert un ample ventall d'hipòtesis (vegeu pp.49–145). La distribució de cada un dels animals s'hadiscutit comparant-la amb la d’altres animals. Aixíhem pogut veure la importància específica delsanimals que pertanyen a la mateixa família, fins i tot almateix gènere i que demostren capacitats d’adaptaciódiferents. Això no treu que entrin en competició entresí a les zones on la seva distribució es sobreposa.Aquestes parelles d'animals són: Equus ferus i Equushydruntinus, Coelodonta antiquitatis i Stephano-rhinus kirchbergensis / Stephanorhinus hemitoechus,Mammuthus primigenius i Elephas antiquus, Cervuselaphus i Rangifer tarandus, Alces alces i Megalo-ceros giganteus, Bison priscus i Bos primigenius,Rupicapra rupicapra i Capra pyrenaica / Capra ibex.
Se'ns ha demostrat particularment interessant elnotar que durant el OIS 4 (estadi isotòpic del oxigen4) corresponent al primer màxim glacial de l'últimaglaciació, la distribució del mamut i del rinocerontllanut arriba fins i tot fins al sud d'Itàlia i Espanyasense provocar l'extinció de l'elefant antic i de les duesespècies de rinoceront de clima temperat. Durant elpaleolític superior, al contrari, al sud de la serraladaCantàbrica, a part d'algunes clares representacions deren, s'han trobat només imatges dubtoses de fauna declima fred. A Itàlia s'han trobat poques restes i cap
representació de mamut, rinoceront llanut o ren en elpaleolític superior. L'extinció de l’Elephas antiquus,Stephanorhinus kirchbergensis i Stephanorhinushemitoechus sembla que sigui més propera de 30.000anys BP. Durant el mateix període varen desaparèixertambé del sud d’Itàlia el Bison priscus i el Megalo-ceros giganteus. Fa uns 30.000 anys o com a més tardamb el OIS 2, corresponent al segon màxim glacial del'última glaciació, Dama dama va esdevenir molt rarai fins i tot pot ser que deixés d'existir a Europa sud-occidental. Aquest és el panorama d'un fort descens dela fauna major a Europa del sud. Dins la llista de lesespècies extinguides durant la part final del períodeOIS 3 podríem afegir fins i tot l’Homo neandertha-lensis, encara que no tractarem aquest tema en aquesttreball.
Cal assenyalar que els pocs indicis que tenim sobre elsanimals que vivien a la zona litoral demostren que elpoc coneixement que tenim d'aquesta zona no és degutsolament a la pujada de la barrera litoral de més de100 metres amb referència al màxim glacial sinótambé al fet que els investigadors l'han deixat moltoblidada. Els pocs jaciments existents amb indicisd’animals que vivien a la zona litoral podem consi-derar-los com a testimonis d'un nombre molt més grandels desapareguts sota el mar (pp. 147–165).
També s'ha pogut demostrar que existeix un nexeentre la distribució d'una espècie animal i la sevarepresentació. Gràcies a “l’anàlisi factorial de corres-pondències“ ha estat també possible provar que lacorrelació d'espècies animals en l'art parietal depènmés de factors ambientals que de factors culturals(167–175).
Tenint en compte les dades de la distribució i en partles de l’extinció dels grans mamífers del paleolíticsuperior a Europa Central i Occidental, ens ha estatpossible formular la hipòtesi, que serà necessariampliar i aprofundir, que el destí de la parella d’Homosapiens i Homo neanderthalensis podria ser reconsi-derat a la llum dels canvis ecològics que tingueren llocentre 40.000 i 25.000 anys BP.


Bibliographie
Aaris-Sørensen, K. 1992. Deglaciation chronologyand re-immigration of large mammals. A southscandinavian example from late Weichselian – earlyFlandrian. In: von Koenigswald, W., Werdelin, L.(eds.), Mammalian Migration and Dispersal Events inthe European Quaternary. Courier ForschungsinstitutSenckenberg, 153: 143–149. Frankfurt am Main.
Aaris-Sørensen, K., Strand Petersen, K., Tauber, H.1990. Danish Finds of Mammoth (Mammuthus primi-genius (Blumenbach)). Stratigraphical position, datingand evidence of Late Pleistocene environment.Danmarks Geologiske Undersøgelse, Serie B 14: 1–44. København.
Acosta González, A., Molinero Barroso, J. M. 2003.Los grabados de la cueva de los Casares. Guada-lajara: AACHE.
Airvaux, J. 2001. L'art préhistorique du Poitou-Charentes. Paris: La maison des roches.
Aksu, A.E., Yasar, D., Mudie, P.J. 1995. Paleocli-matic and paleoceanographic conditions leading todevelopment of sapropel layer S1 in the Aegean Sea.Palaeogeography, Palaeoclimatology, Palaeoecology116: 71–101.
Alcalde del Rio, H., Breuil, H., Sierra, L. 1912. Lescavernes de la région Cantabrique. Monaco: Impri-merie Chêne.
Alcolea González, J.J., Balbín Behrmann, R. de,Garcia Valero, M.A., Jiménez Sanz, P.J. 1997.Nouvelles découvertes d’art rupestre paléolithiquedans le centre de la Péninsule Iberique: La cueva delReno (Valdesotos, Guadalajara). L’Anthropologie101: 144–164.
Alcolea González, J.J., Balbín Behrmann, R. de, 2003.El Arte Rupestre Paleolítico del interior peninsular:Nuevos elementos para el estudio de su variabilidadregional. In: Balbín Behrmann, R. de, Bueno Ramirez,P. (eds.), El Arte Prehistórico desde los inicios delsiglo XXI. Primer Symposium Internacional de ArtePrehistorico de Ribadesella: 223–253. Ribadesella:Asociación Cultural Amigos de Ribadesella.
Alhaique, F. 1994. Taphonomic analysis of the faunalremains from the “P” and “M” layers of the AreneCandide (Savon, Italy). Quaternaria Nova 4: 263–295.
Altuna, J. 1971. El reno en el Würm de la PenínsulaIbérica. Munibe 23: 71–90.
Altuna, J. 1972. Fauna de los mamiferos de losyacimientos de Guipuzcoa. Tesi Doctoral 1971.Munibe 24.
Altuna, J. 1974. Hallazgo de un Uro (Bos primigeniusBoj.) en la sierra de Gibijo (Alava). Estudio de suesqueleto y de la fauna asociada al mismo. Munibe 26:27–51.
Altuna, J. 1976. Los mamiferos del yacimiento prehi-storico de Tito Bustillo (Asturias). In: MoureRomanillo, J.A., Cano Herrera, M. Excavaciones en laCueva de „Tito Bustillo“ (Asturias): 149–194.Oviedo: Instituto de estudios asturianos.
Altuna, J. 1981. Restos óseos del yacimiento prehi-stórico del Rascaño. In: González Echegaray, J.,Barandiarán Maestu, I., El Paleolitico Superior de laCueva del Rascaño: 221–269. Centro de Investiga-ciónes Paleontológicas y Prehistóricas, Memorias 3.Santander: Centro de investigacion y museo deAltamira.
Altuna, J. 1986. The Mammalian faunas from theprehistoric site of La Riera Cave: MNI, NR, body partrepresentation, breakage patterns; age, sex andmetrical data by levels. In: Straus, L.G., Clark, G.A.,La Riera Cave. Stone age hunter-gatherer adapta-tions in northern Spain: 237–274. Arizona stateuniversity, Anthropological research papers N° 36.Tempe: Arizona state university.
Altuna, J. 1990. D. José Miguel de Barandiarán: Notasbiográficas. Munibe 42: 7–9.
Altuna, J. 1993. Archaeozoological research in theIberian Peninsula. Archaeozoologia V/2: 93–100.
Altuna, J. 1994. La relación fauna consumida-faunarepresentada en el paleolítico superior cantábrico.Complutum 5: 303–313.
Altuna, J. 1995. Faunas de mamíferos y cambiosambientales durante el tardiglacial cantábrico. In:Moure Romanillo, A., González Sáinz, C. El final delpaleolítico cantábrico: 77–117. Santander: Univer-sidad de Cantabria.

186 Verbreitung der großen Jagdfauna in Mittel und Westeuropa im oberen Jungpleistozän
Altuna, J. 1996a. Faunas de clima frio en la peninsulaiberica durante el pleistoceno superior. In: Ramil-Rego, P., Fernández Rodríguez, C., RodríguezGuitían, M. (coord.), Biogeografia Pleistocena –Holocena de la Península Ibérica: 13–42. Santiago deCompostela: Xunta de Galicia.
Altuna, J. 1996b. Ekain und Altxerri bei SanSebastian. Sigmaringen: Thorbecke.
Altuna, J., Straus, L.G. 1976. The solutrean ofAltamira: the Artifactual and Faunal Evidence.Zephyrus 26: 175–182.
Altuna, J., Apellaniz, J.M. 1978. Las figuras rupestresde la cueva de Ekain (Deva). Munibe 30.
Altuna, J., Mariezkurrena, K. 1996. Primer hallazgode restos óseos de antílope Saiga (Saiga tatarica L) enla Península Ibérica. Munibe 48: 3–6.
Altuna, J., Mariezkurrena, K. 2000. Macromammi-feros del yacimiento de Labeko Koba (Arrasate, PaísVasco). In: Arrizabalaga, A, Altuna, J. 2000. LabekoKoba. Hyenas y Humanos en los albores delPaleolítico superior. Munibe 52: 107–181.
Álvarez Fernández, E. 2001. Ivory as a raw materialduring the Upper Paleolithic of the northern edge ofthe Iberian Peninsula: personal ornaments and otherartifacts. In: Cavarretta, G., Gioia, P., Mussi, M.,Palombo, M.R. (eds.), The World of Elephants.Proceeding of the 1st International Congress, Roma16–20 Ottobre 2001: 464. Roma: CNR.
Álvarez Fernández, E. 2002. The use of ivory duringthe upper palaeolithic at the Northern edge of theiberian peninsula. Journal of Iberian Archaeology 4:7–19.
Álvarez Fernández, E., Jöris, O. 1998. El significadocronológico de algunas especies de fauna fría duranteel tardiglaciar en la peninsula ibérica. Zephyrus 51:61–86.
Álvarez Fernández, E., Sánchez Chillón, B. 2002.Objetos de marfil del paleolítico superior conservadosen el museo nacional de ciencias naturales. Boletín delreal instituto de estudios asturianos 159: 164–176.
Ament, H. 1994. Ergebnisse einer Umfrage zurBeschäftigungslage von Absolventen des Studiumsder Ur- bzw. Vor- und Frühgeschichte sowieverwandter Fächer. Archäologische Informationen 17/1: 119–121.
Andersen, B., Borns, H.W. 1997. The Ice Age World.Oslo: Scandinavian university Press.
Angelroth, H. 1953. Le Périgordien et l’Aurignacien.Essai d’introduction des industries de stations belgesdans les stades périgordien et aurignacien. Bulletin dela Société Royale Belge d’Anthropologie et de Préhi-stoire 64: 163–183.
Antunes, M.T. 1991. O Homen da Gruta da FigueiraBrava (ca. 30.000 BP). Contexto ecológico, alimen-tação, canibalismo. Memorias da academia daciências de Lisboa. Classe de Ciências 31: 487–356.
Arribas Herrera, A., Jordá Pardo, J.F. 1999. Losmamiferos del Cuaternario kárstico de Guadalajara(Castilla-La Mancha, España). In: Aguirre, E.,Rábano, I. (eds.), La Huella del Pasado. Fósiles deCastilla-La Mancha: 327–353. Patrimonio Histórico.Arqueologia Castilla-La Mancha. Madrid.
Arsuaga Ferreras, P., Aguirre Enríquez, E. 1979.Rinocerontes lanudos en la provincia de Madrid(Coelodonta antiquitatis Blumenbach). Bolletin de laReal Sociedad Española de Historia Natural(Geologia) 77: 23–59.
Ascenzi, A., Segre, A.G. 1971. Il giacimento conmandibola neandertaliana di Archi, Reggio Calabria.Rendiconti/Accademia Nazionale dei Lincei, Ser. VIII,Volume 50: 763–771.
Astre, G. 1950. Faune magdalénienne et azilienne deRochereil (Dordogne). Bulletin de la Sociétéd'Histoire naturelle de Toulouse 85: 151–171.
Aujoulat, N. 1993. Les équidés. In: Aujoulat, N. et al.,L'art pariétal paléolithique. Documents préhisto-riques 5: 97–108. Paris: Comité des Travaux Histo-riques et Scientifiques.
Aujoulat, N., Barrière, C., Bégouën, Bouvier, J.M.,Clottes, J., Crémadès, M., Dauvois, M., Féruglio, V.,Gaussen, J., Lorblanchet, M., Martin, Y., Rouzaud, F.,Sacchi, D., Sauvet, G. 1993. L'art pariétal paléo-lithique. Documents préhistoriques 5. Paris: Comitédes Travaux Historiques et Scientifiques.
Avellino, E., Bietti, A., Giacopini, L., Lo Pinto, A.,Vicari, M. 1986. Riparo Salvini: a New Dryas II Sitein Southern Lazio. Thoughts on the Late Epi-Gravettian of Middle and Southern Tyrrhenian Italy.In: Bonsall, Cl. (ed.) The Mesolithic in Europe: 516–532. Edinburgh: John Donald Publishers LTD.
Azzaroli, A. 1971. Il significato delle faune insulariquaternarie. Le Scienze 30: 84–93.

Bibliographie 187
Baales, M. 1996. Umwelt und Jagdökonomie derAhrensburger Rentierjäger im Mittelgebirge.Monographien, Band 38. Römisch-GermanischesZentralmuseum, Mainz. Bonn: Rudolf Habelt.
Bachmayer, F., Simeonidis, N. Seeman, R., Zapfe, H.1976. Die Ausgrabungen in der Zwerg-elephantenhöhle „Charkaido“ auf der Insel Tilos.Annalen des Naturhistorischen Museums Wien 80:113–144.
Bahn, P.G. 1977. Seasonal migration in South-WestFrance during the Late Glacial Period. Journal ofArcheological Science 4: 245–257.
Bahn, P.G. 1982. Inter-site and inter-regional linksduring the Upper Palaeolithic: the Pyrenean evidence.Oxford Journal of Archaeology 1/3: 247–268.
Bahn, P.G. 1983. Pyrenean prehistory. Wilshire: Aris& Phillips.
Bahn, P.G. 2003. Librenme del ultimo trance: Unavaloración del mal uso del chamanismo en losestudios de arte rupestre. In: Balbín Behrmann, R. de,Bueno Ramirez, P. (eds.), El Arte Prehistórico desdelos inicios del siglo XXI. Primer Symposium Interna-cional de Arte Prehistorico de Ribadesella: 53–73.Ribadesella: Asociación Cultural Amigos de Ribade-sella.
Bahn, P.G., Vertut, J. 1988. Images of the ice age.Leicester: Windward.
Balbín Behrmann, R. de, 1989. L'art de la grotte deTito Bustillo (Ribadesella, Espagne). Une vision desynthèse. L'Anthropologie 93: 435–462.
Balbín Behrmann, R. de, Alcolea González, J.J. 1992.La grotte de los Casares et l'art paléolithique de laMeseta espagnole. L'Anthropologie 96: 397–452.
Balbín Behrmann, R. de, Alcolea González, J.J. 1995.Arte paleolítico de la Meseta española. Complutum 5:97–138.
Balbín Behrmann, R. de, Alcolea González, J.J.,González Pereda, M.A. 1999. Une vision nouvelle dela grotte de el Pindal, Pimiango, Ribadedeva, Asturies.L'Anthropologie 103: 51–92.
Balbín Behrmann, R. de, Alcolea González, J.J.,González Pereda, M.A. 2003. El macizo de Ardines,Ribadesella, España. Un lugar mayor del artepaleolítico europeo. In: Balbín Behrmann, R. de,Bueno Ramirez, P. (eds.), El Arte Prehistórico desde
los inicios del siglo XXI. Primer Symposium Interna-cional de Arte Prehistorico de Ribadesella: 91–151.Ribadesella: Asociación Cultural Amigos de Ribade-sella.
Bandi, H.-G., Maringer, J. 1952. Kunst der Eiszeit,Levantekunst, Arktische Kunst. Basel: Holbein.
Bannikov, A.G. 1963. Die Saiga-Antilope. Leipzig:Buchdruckerei Richard Hahn.
Barandiarán Maestu, I. 1973. Arte mueble delpaleolítico Cantabrico. Zaragoza: Librería general.
Barandiarán Maestu, I. 1995. La prehistoria. In:VVAA, Gran atlas historico de Euskal Herria.Bilbao: Lur Argitaletxea.
Barrière, Cl. 1976. L’art pariétal de Gargas. BARSupplementary Series 14. Oxford: Archaeopress.
Barrière, Cl. 1982. L'art pariétal de Rouffignac. Lagrotte aux cent mammouths. Paris: Picard.
Barrière, Cl. 1993a. Les bovidés. In: Aujoulat, N. etal., L'art pariétal paléolithique. Documents préhisto-riques 5: 109–122. Paris: Comité des Travaux Histo-riques et Scientifiques.
Barrière, Cl. 1993b. Les proboscidiens. In: Aujoulat,N. et al., L'art pariétal paléolithique. Documentspréhistoriques 5: 151–156. Paris: Comité des TravauxHistoriques et Scientifiques.
Barrière, Cl. 1993c. Les Rhinocérotidés. In: Aujoulat,N. et al., L'art pariétal paléolithique. Documentspréhistoriques 5: 157–158. Paris: Comité des TravauxHistoriques et Scientifiques.
Bartholomew, J. et al. 1980. Knaurs großer Weltatlas.München: Droemer Knaur Verlag.
Bartoli, G., Galiberti, A., Gorini, P. 1978. Oggettid'arte mobiliare rinvenuti nelle province di Grosseto ePisa. Rivista di Scienze Preistoriche 33 (1977): 193–218.
Bartolomei, G., Broglio, A., Cattani, L., Cremaschi,M., Guerreschi, A., Mantovani, E., Peretto, C., Sala,B. 1982. I depositi würmiani del Riparo Tagliente.Annali dell'universitá di Ferrara N.S., Sez. XV,Paleontologia umana e Paletnologia, Vol. III, N. 4:61–105.
Barton, N. 1997. Stone Age Britain. London:B.T.Batsford/Englisch Heritage.

188 Verbreitung der großen Jagdfauna in Mittel und Westeuropa im oberen Jungpleistozän
Bar-Yosef, O. 1989. Upper Pleistocene cultural strati-graphy in southwest Asia. In: Trinkaus, E. (ed.) Theemergence of modern humans. Biocultural adapta-tions in the later Pleistocene: 154–180. Cambridge:Cambridge University Press.
Bayer, J. 1925. Dr. Otto Hauser, eine Gefahr für dieUrgeschichtsforschung. Die Eiszeit 2: 135.
Becker, C. 1986. Kastanas. Die Tierknochenfunde.Prähistorische Archäologie in Südosteuropa, Band 5.Berlin: Wissenschaftsverlag Volker Spiess.
Bégouën, H., Breuil, H. 1958. Les cavernes du Volp.Paris: Artes et métiers graphiques.
Bégouën, R., Clottes, J., Giraud, J.-P., Rouzaud, F.1986. Le propulseur au saïga d’Enlène. PréhistoireAriégeoise 41: 11–22.
Bégouën, R., Clottes, J. 1987. Les Trois-Frères afterBreuil. Antiquity 61: 180–187.
Bellier, Cl., Bott, S., Cattelain, P., Fritz, C., Jadin, I.1999. La rondelle au mammouth de Chaleux. In:Julien, M. et al., Préhistoire d'os: 97–121. Aix-en-Provence: Publications de l'Université de Provence.
Beltrán, A. 1979. Les animaux de l'art rupestre deschasseurs du Levant espagnol. In: Bandi, H.-G.,Huber, W., Sauter, M.-R., Sitter, B. (eds.), La contri-bution de la zoologie et de l'éthologie à l'interpré-tation de l'art des peuples chasseurs préhistoriques.3e Colloque de la Société suisse de sciences humaines(1979): 353–369. Fribourg: Editions UniversitairesFribourg.
Benini, A., Boscato, P., Gambassini, P. 1997. Grottadella Cala (Salerno): industrie litiche e fauneuluzziane ed aurignaziane. Rivista di Scienze Preisto-riche 48: 37–95.
Bétirac, B. 1952. L'abri Montastruc à Bruniquel (Tarn-et-Garonne). L'Anthropologie 56: 213–231.
Biagi, P. 1976. Strumento litico del Paleoliticosuperiore della cavernetta Büs dei lader (97 LO).Natura Bresciana 13: 117–120.
Bietti, A., Martini, F., Tozzi, C. 1985. L'Épigravettienévolué et final de la zone moyenne et basse tyrrhé-nienne. Rivista di Scienze Preistoriche 38 (1983):319–349.
Bietti, A., Molari, C. 1994. The upper pleistocenedeposit of the Arene Candide Cave (Savona, Italy):General introduction and stratigraphy. Quaternarianova 4: 9–27.
Binford, L.R. 1962. Archaeology as Anthropology.American Antiquity 28: 217–225.
Binford, L.R. 1982. The Archaeology of Place.Journal of Anthropological Archaeology 1: 5–31.
Binford, L.R. und Binford, S.R. 1969. Stone Tools andHuman Behavior. Scientific American 220, 4: 70–84.
Bischoff, J.L., Soler, N., Maroto, J., Julià, R. 1989.Abrupt Mousterian/Aurignacian Boundary at c. 40 kabp: Accelerator 14C dates from l'Arbreda Cave(Catalunya, Spain). Journal of Archaeological Science16: 563–576.
Blanc, G.A. 1928. Sulla presenza di Alca impennisLinn. nella formazione pleistocenica superiore digrotta Romanelli in Terra d'Otranto. Archivio perl'Antropologia e l'Etnografia 58: 1–34.
Blanc, A.C. 1938. Sulla penetrazione e diffusione inEuropa ed in Italia del Paleolitico Superiore infunzione della paleoclimatologia e paleogeografiaglaciali. Quartär 1: 1–26.
Blanc, A.C. Vries, Hl. de, Follieri, M. 1957. A first C14 Date for the Würm I Chronology on the ItalianCoast. Quaternaria 4: 83–91.
Blant, M., Blant, D., Della Toffolo, R., Wahren-berger, F. 2002. Nouvelles découvertes d’élans (Alcesalces) et d’un cerf (Cervus elaphus) holocènes à laSchrattenfluh (Flühli, LU). Cavernes 2/2002: 17–20.
Bocquet-Appel, J.-P., Demars, P.-Y. 2000. PopulationKinetics in the Upper Palaeolithic in Western Europe.Journal of Archaeological Science 27: 551–570.
Bogoras, W. 1909. The Chukchee. Memoirs of theAmerican Museum of Natural History, Vol. XI.,Edited by F.Boas, Reprint from Vol. VII, Part I-III ofthe Jesup North Pacific Expedition. New York,London: Johnson reprint corporation, Johnson reprintcompany.
Bohlken, H. 1986. Ovibos moschatus (Zimmermann,1780) – Moschusochse. In: Niethammer, J. Krapp, F.(Hrsg.), Handbuch der Säugetiere Europas. Band 2/IIPaarhufer: 349–361. Wiesbaden: Aula.

Bibliographie 189
Bökönyi, S. 1982. The Early Neolithic fauna ofRendina: a preliminary report. Origini 9 (1977–1982):345–354.
Bolduc, P., Cinq-Mars, J., Mussi, M. 1996. Lesfigurines des Balzi Rossi (Italie): une collectionperdue et retrouvée. Préhistoire Ariégeoise 51: 15–53.
Bologna, P., Petronio, C. 1994. The first occurrence ofBison priscus Bojanus in Melpignano area (Lecce,Southern Italy). Bollettino della Societá Paleonto-logica Italiana 33: 275–278.
Bologna, P., Di Stefano, G., Manzi, G., Petronio, C.,Sardella, R., Squazzini, E. 1994. Late Pleistocenemammals from the Melpignano (LE) „Ventarole“:preliminary analysis and correlations. Bollettino dellaSocietá Paleontologica Italiana 33: 265–274.
Bolus, M. 1992. Die Siedlungsbefunde des späteiszeit-lichen Fundplatzes Niederbieber (Stadt Neuwied).Augrabungen 1981–1988. Römisch-GermanischesZentralmuseum, Monographien 22. Bonn: RudolfHabelt.
Bolus, M., Conard, N.J., Floss, H. (Hrsg.) 2000.Exkursionsführer. Paläolithische Fundstellen imAchtal, Lonetal und Schmiechtal. Tübingen: Institutfür Ur- und Frühgeschichte und Archäologie desMittelalters.
Bonifay, M.-F. 1978. Le gisement de la Baume deValorgues à Saint-Quentin-La-Poterie (Gard) (Paléo-lithique supérieur final et Épipaléolithique). II Lagrande Faune. Gallia Préhistoire 21: 123–142.
Borzatti von Löwenstern, E., 1970. Prima Campagnadi scavi nella grotta „Mario Bernardini“. Rivista discienze Preistoriche 25: 89–125
Boscato, P., Ronchitelli, A., Wierer, U. 1997. IlGravettiano antico della Grotta della Cala a Marina diCamerota. Paletnologia e ambiente. Rivista di ScienzePreistoriche 48: 97–186.
Bosinski, G. 1981. Gönnersdorf. Eiszeitjäger amMittelrhein. Koblenz: Landesmuseum Koblenz.
Bosinski, G. 1982. Die Kunst der Eiszeit inDeutschland und in der Schweiz. Bonn: RudolfHabelt.
Bosinski, G. 1987. Die große Zeit der Eiszeitjäger.Europa zwischen 40.000 und 10.000 v.Chr. Jahrbuchdes Römisch-Germanischen Zentralmuseums Mainz34: 3–139.
Bosinski, G. 1992. Eiszeitjäger im Neuwieder Becken.Dritte erweiterte und veränderte Auflage. Archäologieam Mittelrhein und Mosel, Band 1.
Bosinski, G. 1994. Menschendarstellungen derAltsteinzeit. In: Ulmer Museum (Hrsg.). Der Löwen-mensch. Tier und Mensch in der Kunst der Eiszeit.Ausstellung im Ulmer Museum vom 11. September bis13. November 1994. Ausstellungskatalog: 77–99.Sigmaringen: Thorbecke.
Bosinski, G. 1996. Die Gravierungen desMagdalénien-Fundplatzes Andernach-Martinsberg.Jahrbuch des Römisch-Germanischen Zentralmu-seums Mainz 41 (1994): 19–58.
Bosinski, G., Fischer, G. 1980. Mammut- und Pferde-darstellungen von Gönnersdorf. Wiesbaden: FranzSteiner Verlag.
Bosinski, G., Bosinski H. 1991. Robbendarstellungenvon Gönnersdorf. In: Festschrift Karl Brunnacker.Sonderveröffentlichung des Geologischen Institutesder Universität Köln 82: 81–87.
Boule, M. 1910. Paléontologie. In: Villeneuve, L.,Boule, M. Les Grottes de Grimaldi (Baussé – Roussé):157–236. Monaco: Imprimerie de Monaco.
Boyd, L., Houpt, K. 1994. Przewalski's horse. Thehistory and biology of an endagered species. NewYork: State university of New York press.
Breuil, H. 1906. Les gisements Présolutréens du typed’Aurignac. Coup d’œil sur le plus ancien âge duRenne. In: Congrès International d’Anthropologie etArchéologie Préhistoriques, 13ème session, Monaco:323–346.
Breuil, H. 1927. Oevres d'art paléolithique inédites duPérigord et art oriental d'Espagne. Revue anthropolo-gique 37: 1–8.
Breuil, H. 1952. Quatre cents siècles d'art pariétal.Montignac: Centre d'études et de documentationpréhistoriques.
Breuil, H., Obermaier, H., Verner, W. 1915. La Piletaa Benaojan. Mónaco: Institut de PaléontologieHumaine.
Breuil, H., Obermaier, H. 1935. The Cave of Altamiraat Santillana del Mar, Spain. Madrid: Tipografia deArchivos.

190 Verbreitung der großen Jagdfauna in Mittel und Westeuropa im oberen Jungpleistozän
Breuil, H., Nougier, L.-R., Robert, R. 1956. Le„Lissoir aux Ours“ de la grotte de la Vache, à Alliat, etl'ours dans l'art franco-cantabrique occidentale.Bulletín de la Société Préhistorique de l'Ariège. Préhi-stoire et Spéléologie Ariégeoises 9: 15–78.
Bridault, A. 1997. Chasseurs, ressources animales etmilieux dans le Nord de la France de la fin du Paléo-lithique à la fin du Mésolithique: Problématique et étatde la recherche. In: Fagnart, J.-P., Thévenin, A. (dir.),Le tardiglaciaire en Europe du Nord-Ouest. Actes du119e congrès national des sociétés historiques et scien-tifiques, Amiens 1994: 165–176. Paris: Éditions duComité des travaux historiques et scientifiques.
Bridault, A. 1998. L’environnement animal et sonexploitation dans le Massif jurassien (13000–6500BP). In: Cupillard, Chr., Richard, A. (éd.), Lesderniers chasseurs-cueilleurs du massif jurassien et deses marges: 73–78. Lons-le-Saunier: Centre Jurassiendu Patrimoine.
Bridault, A., Bemilli, C. 1999. La chasse et letraitement des animaux. In: Julien, M., Rieu, J.-L.(dir.), Occupations du Paléolithique supérieur dans lesud-est du Bassin parisien. Documents d'archéologiefrançaise 78: 50–64.
Bridault, A., Chaix, L., Pion, G., Oberlin, S.,Thiébault, Argant, J. 2000. Position chronologique durenne à la fin du Tardiglaciaire dans les Alpes du Nordfrançaises et le Jura méridional. In: Pion, G. (dir.) LePaléolithique Supérieur récent: nouvelles données surle peuplement et l’environnement. Actes de la TableRonde de Chambéry, 12–13 mars 1999. Société Préhi-storique Française, Mémoires 28: 47–57.
Bridault, A., Chaix, L. 2002. Ruptures et Équilibresdans la grande faune à la fin du Pleistocène et durantl'Holocène ancien en Europe occidentale. In: RichardH., Vignot A. (dir.), Équilibres et ruptures dans lesécosystèmes depuis 20.000 ans en Europe de l'Ouest,Actes du colloque international de Besançon: 53–60.Besançon: Presses Universitaires Franche-Comté.
Brizzi, B. 1977. L'Italia nell'etá della pietra. Roma:Edizioni Quasar.
Broglio, A., Kozłowski, J. 1987. Il Paleolitico. Uomo,ambiente e culture. Milano: Jaca Book.
Brooks, S., Gorissen, M. 1991. Elefantenjagd inAfrika. In: Studenten der Universität Hamburg,Landkreis Verden und Heimatmuseum Verden.Begleitheft zur Ausstellung. Jagdtechniken derSteinzeit: 35–42. Verden: Verdener Heimatbund.
Brugal, J.-Ph., David, F., Enloe, J.G., Jaubert, J. (dir.)1999. Le Bison: gibier et moyen de subsistance deshommes du Paléolithique aux Paléoindiens desGrandes Plaines. Actes du colloque international,Toulouse 1995. Antibes: Éditions APDCA.
Bützler, W. 1977. Rotwild. München: BLV Verlagsge-sellschaft.
Bützler, W. 1986. Cervus elaphus Linnaeus, 1758 –Rothirsch. In: Niethammer, J. Krapp, F. (Hrsg.),Handbuch der Säugetiere Europas. Band 2/IIPaarhufer: 107–139. Wiesbaden: Aula.
Burke, A., Eisenmann, V., Ambler, G. K. 2003. Thesystematic position of Equus hydruntinus, an extinctspecies of Pleistocene equid. Quaternary Research 59:459–469.
Cacho Quesada, C., Ripoll López, S., Jordá Pardo,J.F., Muñoz Ibáñez, Yaravedra Sáinz de los Terreros,J., Maicas Ramos, R. 2003. El registro arqueologicodel Pleistoceno Superior Final en el abrigo de la Peñade Estebanvela (S de la cuenca del Duero, Segovia,España). XI reunión nacional de cuaternario, Oviedo(Asturias) 2–3 y 4 de julio 2003: 191–198. http://info.uned.es/dpto-pha/estebanvela/aequa.pdf
Caloi, L., Coccolini, G., Mussi, M., Palombo, M. R.,Vitagliano, S., Zampetti, D. 1989. Le Moustérien duLatium (Italie Centrale): archéologie, milieu naturel,chronologie. L'Anthropologie 92 (1988): 73–98.
Caloi, L., Kotsakis, T., Palombo, M., Petronio, C.1996. The Pleistocene dwarf elephants of Mediter-ranean islands. In: Shoshani, J., Tassy, P. 1996. TheProboscidea. Evolution and Palaeoecology ofElephants and Their Relatives: 234–239. Oxford, NewYork, Tokyo: Oxford University Press.
Campbell, J.B. 1977. The Upper Palaeolithic ofBritain. Oxford: Claredon press.
Canal Roquet, J., Carbonell Roura, E. 1989.Catalunya paleolitica. Girona: Patronat Eiximenis.
Capasso Barbato, L., Minieri, M.R., Scarano, M.,1991. Paleobiografia dei mammiferi „freddi“ delpleistocene medio-superiore italiano. Atti AccademiaPeleoritana dei Pericolanti. Classe I di ScienzeFisiche, Matematiche e Nataturali. Vol. LXVII (1989)– Supplemento N° 1. Messina.
Capitan, L., Breuil, H., Peyrony, D. 1910. La Cavernede Font-de-Gaume aux Eyzies (Dordogne). Monaco:Imprimerie Chêne.

Bibliographie 191
Capitan, L., Peyrony, D. 1912. Station préhistoriquede La Ferrassie. Commune de Savignac-du-Bugue(Dordogne). Revue anthropologique 22: 76–99.
Capitan, L., Breuil, H., Peyrony, D. 1915. Nouvellesgrottes ornées de la vallée de la Beune. L'Anthropo-logie 26: 505–518.
Capitan, L., Breuil, H., Peyrony D. 1924. Les Comba-relles aux Eyzies. Institut de Paléontologie Humaine.Paris: Masson.
Capitan, L., Peyrony, D. 1928. La Madeleine. Songisement, son industrie, ses oevres d'art. Paris:Librairie Émile Nourry.
Cardoso, J.L. 1993. Contribução para o conheci-mento dos grandes mamíferos do plistocénicosuperior de Portugal. Oeiras: Câmara Municipal deOeiras.
Cardoso, J.L. 1996. Les grands mammifères duPléistocène supérieur du Portugal. Essai de synthèse.GEOBIOS 29: 235–250.
Carrasco Duarte, M. 1999. El Padul. Ayuntamento dePadul.
Cartailhac, E. 1902. Les cavernes ornées de dessins.La grotte d’Altamira, Espagne. „Mea culpa“ d’unsceptique. L’Anthropologie 13: 350–352.
Cartailhac, E., Breuil, H. 1906. La Caverne d'Altamiraà Santillane près Santander (Espagne). Monaco:Imprimerie de Monaco.
Cassoli, P.F. 1992. Avifauna del Pleistocene superioredelle Arene Candide, Praia e Grotta Romanelli (Italia).Quaternaria nova 2: 239–246.
Cassoli, P.F., Segre, A.G., Segre, E., 1979. Evolutionmorphologique et écologique de la côte de Castro(Pouilles) dans le Pléistocène final. In: de Sonneville-Bordes, D. (ed.), La fin de temps glaciaires en Europe.Colloques internationaux du Centre nationale de larecherche scientifique, N° 271, Talence 24–28 Mai1977: 325–332. Paris: C.N.R.S.
Cassoli, P.F., Tagliacozzo, A. 1991. Considerazionipaleontologiche, paleoecologiche e archeozoologichesui macromammiferi e gli uccelli dei livelli delPleistocene superiore del Riparo di Fumane (Vr),(scavi 1988–1991). Bollettino Museo civico di StoriaNaturale di Verona 18: 349–445.
Cassoli, P.F., Tagliacozzo, A. 1992. La Macrofaune.Colloque international: Adaptations au milieumontagnard au paléolithique superieur et aumésolithique. Museo Tridentino di Scienze Naturali,Trento 5–11 Octobre 1992: 28–30. Ferrara: Universitádi Ferrara.
Cassoli, P.F., Tagliacozzo, A. 1994. I macromam-miferi dei livelli tardopleistocenici delle AreneCandide (Savona, Italia): Considerazioni paleontolo-giche e archeozoologiche. Quaternaria nova 4: 101–262.
Castel, J.Ch., Liolios, D., Chadelle, J.-P., Geneste, J.-M. 1998. De l’alimentaire et du technique: la consom-mation du renne dans le Solutréen de la grotte deCombe Saunière. In: Brugal, J.-Ph., Meignen, L.,Patou-Mathis, M.. Économie préhistorique: lescomportements de subsistence au Paléolithique. Actesdes Rencontres, 23-24-25 octobre 1997: 433–450.
Cavarretta, G., Gioia, P., Mussi, M., Palombo, M.R.(eds.) 2001. The World of Elephants. Proceeding ofthe 1st International Congress, Roma 16–20 Ottobre2001. Roma: CNR.
Chapman, D., Chapman, N. 1980. The distribution ofFallow deer: a worldwide review. Mammal Review 10:61–138.
Charles, R. 1997. The exploitation of carnivores andother furbearing mammals during the north-westerneuropean Late Upper Palaeolithic and Mesolithic.Oxford Journal of Archaeology 16 (3): 253–277.
Charles, R. 1998. Late Magdalenian Chronology andFaunal Exploitation in the North-Western Ardennes.BAR International Series 737. Oxford: Archaeopress.
Chauvet, J.-M., Deschamps, E., Hillaire, Ch. 1996.Chauvet Cave. London: Thames and Hudson.
Cheynier, A. 1949. Badegoule, station solutréenne etproto-magdalénienne.Archives de l'Institut de Paléon-tologie Humaine. Memoire 23. Paris: Masson.
Cheynier, A. 1965. L’abri Lachaud à Terrasson,Dordogne. Préhistoire XVI. Paris: Presses universi-taires.
Cheynier, A., Breuil, H. 1963. La caverne de Pair-non-Pair. Gironde. Documents d'Aquitaine (II). Publi-cation de la Société Archeologique de Bordeaux.

192 Verbreitung der großen Jagdfauna in Mittel und Westeuropa im oberen Jungpleistozän
Chilardi, S., Frayer, D.W., Gioia, P., Macchiarelli, R.,Mussi, M. 1996. Fontana Nuova di Ragusa (Sicily,Italy): southernmost Aurignacian site in Europe.Antiquity 70: 553–563.
Chollot, M. 1964. Musée des Antiquités Nationales.Collection Piette. Paris: Musées Nationaux.
Clark, J.G.D. 1946. Seal-hunting in the Stone Age ofnorth-western Europe: a study in economic prehi-story. Proceedings of the Prehistoric Society: 12–48.
Cleyet-Merle, J.-J. 1990. La préhistoire de la peche.Paris: Éditions Errance.
Cleyet-Merle, J.-J. et al. 1990. Lartet, Breuil, Peyronyet les autres. Sarlat: Imprimerie nouvelle.
Cleyet-Merle, J.-J., Madelaine, St. 1991. Lapendeloque magdalénienne gravée d'un „equushydruntinus“ de la grotte du Putois II. Paléo 3: 119–129.
Cleyet-Merle, J.-J., Madelaine, St. 1995. Inlandevidence of human sea coast exploitation in Palaeo-lithic France. In: Fischer, A. (ed.), Man and Sea in theMesolithic: 303–308. Oxford: Oxbow Books.
Clottes, J. 1995. Les Cavernes de Niaux. Paris:Éditions du Seuil.
Clottes, J. (ed.) 2001. La Grotte Chauvet. L'Art desorigines. Paris: Éditions du Seuil.
Clottes, J. (ed.) 2003. Chauvet Cave. The Art ofearliest times. Salt Lake City: The University of UtahPress.
Clottes, J., Courtin, J. 1994. La Grotte Cosquer. Paris:Éditions du Seuil.
Clottes, J., Lewis-Williams, D. 1997. Schamanen.Trance und Magie in der Höhlenkunst der Steinzeit.Sigmaringen: Thorbecke.
Clottes, J., Packer, C. 2001. Les lions. In: Clottes, J.(ed.) La Grotte Chauvet. L'Art des origines: 177–180.Paris: Éditions du Seuil.
Conard, N.J. 1999. Bovid Remains from the 1991–1994 Excavation in Wallertheim and their Bearing onthe Interpretation of the Locality. In: Brugal, J.-Ph.,David, F., Enloe, J.G., Jaubert, J. (dir.). Le Bison:gibier et moyen de subsistance des hommes du Paléo-lithique aux Paléoindiens des Grandes Plaines. Actesdu colloque international, Toulouse 1995: 301–318.Antibes: Éditions APDCA.
Conard, N.J. 2003. Palaeolithic ivory sculptures fromsouthwestern Germany and the origins of figurativeart. Nature 426: 830–832.
Conard, N. J., Bolus, M. 2003. Radiocarbon dating theappearance of modern humans and timing of culturalinnovations in Europe: new results and newchallenges. Journal of Human Evolution 44: 331–371.
Conard, N.J., Langguth, K., Uerpmann, H.-P. 2003.Einmalige Funde aus dem Aurignacien und ersteBelege für ein Mittelpaläolithikum im Hohle Fels beiSchelklingen, Alb-Donau-Kreis. ArchäologischeAusgrabungen in Baden-Württemberg 2002: 21–27.
Corchón Rodríguez, M.S. 1992. Representaciones defauna fría en el arte mueble de la cueva de Caldas(Asturias, España). Significación e implicaciones en elarte parietal. Zephyrus 44–45: 35–64.
Corchón Rodríguez, M.S. 1994. Últimos hallazgos ynuevas interpretaciones del arte mueble paleolítico enel occidente asturiano. Complutum 5: 235–264.
Corchón Rodríguez, M.S. 1997. La cornicheCantabrique entre 15.000 et 13.000 ans BP: Laperspective donnée par l'art mobilier. L'Anthropologie101: 114–143.
Corchón Rodríguez, M.S. 1998. Nuevas representa-ciones de antropomorfos en el magdaleninse mediocantábrico. Zephyrus 51: 35–60.
Corchón Rodríguez, M.S. 1999. Solutrense y magda-leniense del oeste de la cornisa cantábrica: Dataciones14C (calibradas) y marco cronologico. Zephyrus 52: 3–32.
Corchón Rodríguez, M.S, Valladas, H., Bécares, J.,Arnold, M., Tisnerat, N., Cachier, H. 1996. Dataciónde las pinturas y revisión del Arte Paleolítico de cuevapalomera (Ojo Guareña, Burgos, España). Zephyrus54: 37–60.
Costamagno, S. 1999. Stratégies de Chasse etFonction des Sites au Magdalénien dans le Sud de laFrance. Thèse doctoral à l’université Bordeaux I.
Costamagno, S. 2000. Stratégies d'approvisionnementet traitement des carcasses au Magdalénien: L'exemplede Moulin-Neuf (Gironde). Paléo 12: 77–95.
Cramer, B. 2002. Morphometrische Untersuchungenan quartären Pferden in Mitteleuropa. Dissertation ander Geowissenschaftlichen Fakultät der EberhardKarls Universität Tübingen.

Bibliographie 193
Crégut-Bonnoure, E. 1984. The Pleistocene ovibo-vinae of western Europe: Temporo-spatial expantionand paleoecological implications. In: Klein, D.R.,White, R.G., Keller, S. (eds.), Proceedings of the FirstInternational Muskox Symposium. Biological Papersof the University of Alaska, Special Report No. 4:136–144.
Crémadès, M. 1993. Les Cervidés. In: Aujoulat et al.,L'art pariétal paléolithique. Documents préhisto-riques 5: 137–150. Paris: Comité des Travaux Histo-riques et Scientifiques.
Crémadès, M. 1997. La Représentation des variationssaisonnières dans l’art pariétal paléolithique.L'Anthropologie 101: 36–82.
Cremaschi, M. 1990. The Loess in Northern andCentral Italy. Quaderni di geodinamica alpina equaternaria 1.
Cziesla, E., Eickhoff, S., Arts, N., Winter, D. (eds.)1990. The Big Puzzle. Bonn: HOLOS.
Dams, L. 1987. L’Art paléolithique de la Grotte deNerja (Malaga, Espagne). BAR Int. Series 385.Oxford: Archaeopress.
Daniel, G. 1990. Geschichte der Archäologie. Köln:Corvus Verlag.
Darwin, Ch. 1859. On the origin of Species by Meansof Natural Selection, or the Preservation of FavouredRaces in the Struggle for Life. London.
Davidson, I 1975. Saisonality in Spain. Zephyrus 26–27: 167–173.
Davidson, I. 1989. La economía del Final delPaleolítico en la Epaña oriental. Valencia: Diput-ación provincial de Valencia.
Davis, S.J.M. 2002. The mammals and birds from theGruta do Caldeirão, Portugal. Revista Portuguesa deArqueologia 5, 2: 29–98.
Delpech, F. 1970. Faune du Magdalenien IV dugisement de Duruthy, commune de Sorde-l'Abbaye(Landes). Bulletin de l'associacion française pourl'étude du Quaternaire 7: 13–26.
Delpech, F. 1983. Les Faunes du Paléolithiquesupérieur dans le Sud-ouest de la France. Cahiers duQuaternaire N° 6, Éditions du Centre National de laRecherche scientifique, Paris.
Delpech, F., 1989. Le Temps de l'antilope Saiga. In:Mohen, J.-P., Le Temps de la Préhistoire, Tome 2, 48-49 (Dijon).
Delpech, F. 1999. La chasse au Bison dans le sud-ouest de la France au cours du Würm: choix humainou contraintes paléoenvironnementales? In: Brugal, J.-Ph., David, F., Enloe, J.G., Jaubert, J. (dir.). Le Bison:gibier et moyen de subsistance des hommes du Paléo-lithique aux Paléoindiens des Grandes Plaines. Actesdu colloque international, Toulouse 1995: 63–84.Éditions APDCA, Antibes.
Delpech, F., Heintz, E. 1976. Les Artiodactyles:Bovidés. In: de Lumley, H. (direction) La préhistoireFrançaise. Tome I. Les civilisations Paléolithiques etMésolithiques de la France. 386-394.
Delpech, F., Grayson, D. K., Rigaud, J.-Ph. 2000.Biostratigraphie et paléoenvironnements du début duWürm récent d'après les grands mammifères de l'Abridu Flageolet I (Dordogne, France). Paléo 12: 97–126.
Delporte, H. 1956. Note sur le Périgordien belge.Bulletin de la Société Préhistorique Française 53: 11–15.
Delporte, H. 1984a. Abri du Roc-de-Sers. In: Leroi-Gourhan, A. et al. L'art des cavernes, Atlas des grottesornées paléolithiques françaises: 578–582. Paris:Imprimerie Nationale.
Delporte, H. 1984b. L'art mobilier et ses rapports avecla faune paléolithique. In: Bandi, H.-G., Huber, W.,Sauter, M.-R., Sitter, B. (eds.), La contribution de lazoologie et de l'éthologie à l'interprétation de l'art despeuples chasseurs préhistoriques. 3e Colloque de laSociété suisse de sciences humaines (1979): 111–142.Fribourg: Editions Universitaires Fribourg.
Delporte, H. 1990. L’image des Animaux dans l’ArtPréhistorique. Paris: Picard.
Demars, P.-Y. 1996. Démographie et occupation del'espace au Paléolithique Supérieur et au Mésolithiqueen France. Préhistoire Européenne 8: 3–26.
Demars, P.-Y. 2002. L'occupation de l'Europeoccidentale à la fin de la dernière glaciation. In: Actesdu Congrés de Besançon „Equilibres et ruptures dansles écosystèmes durant les 20 derniers millénaires:durabilité et mutation“: 143–151.
Denzau, G., Denzau, H. 1999. Wildesel. Stuttgart:Thorbecke.

194 Verbreitung der großen Jagdfauna in Mittel und Westeuropa im oberen Jungpleistozän
Desbrosse, R. Prat, F. 1974. L’élan magdalénien dePierre-Châtel (Ain). Quartär 25: 143–157.
Djindjian, F. 1994. L’influence des frontièresnaturelles dans les déplacements des chasseur-cueil-leurs au würm récent. Preistoria Alpina 28 (1992): 7–28.
Djindjian, F., Kozłowski, J., Otte, M. 1999. Le paléo-lithique supérieur en Europe. Paris: Armand Colin.Döppes, D., Rabeder, G. (Hrsg.) 1997. Pliozäne undpleistozäne Faunen Österreichs. Wien: Verlag derÖsterreichischen Akademie der Wissenschaften.
Dubois, Auguste, Stehlin, H.G. 1933. La grotte deCotencher, station moustérienne. Mémoires de laSociété Paléontologique Suisse, Volume LII–LIII.Bâle: Emile Birkhauser & Cie.
Dubourg, Chr., Bouvier, J.-M., Castanet, R. 1994. Unbloc gravé de l’abri de la Souquette (Sergeac,Dordogne). Une nouvelle figuration d’antilope Saiga.Paléo 6: 247–259.
Dubourg, Chr. 1997. Les expressions du naturalismedans les arts graphiques du Paléolithique supérieur.Une vision du monde des chasseurs préhistoriques.Thèse doctorale présentée à l'université Bordeaux I.
Duguy, R., Robineau, D. (Hrsg.) 1992. Handbuch derSäugetiere Europas, Band 6: Meeressäuger. Teil II:Robben – Pinnipedia. Wiesbaden: Aula.
Duhard, J.-P. 1992. Les humains ithyphallique dansl'art paléolithique. Préhistoire Ariégeoise 47: 133-159.
Duhard, J.-P. 1996. Réalisme de l'image masculinepaléolithique. Grenoble: Éditions Jérôme Millon.
Duphorn, K., Grahlet, H.-O., Schneider, H. (Wissen-schaftliche Redaktion), 1970. Internationale Quartär-karte von Europa, 1:2500000, Blatt 6 København.Hannover.
Eastham, A. 1968. The avifauna of Gorham's Cave,Gibraltar. Bulletin of the institute of Archaeology 7(1967): 37–42.
Eastham, A. 1986. The birds of the cueva of Nerja. In:Jordá Pardo, J.F., La Prehistoria de la Cueva de Nerja(Malaga): 109–131. Malaga: Imprenta de la univer-sidad de Malaga.
Engländer, H. 1986. Capra pyrenaica Schinz, 1838 –Spanischer Steinbock, Iberiensteinbock. In:Niethammer, J. Krapp, F. (Hrsg.), Handbuch derSäugetiere Europas. Band 2/II Paarhufer: 405–422.Wiesbaden: Aula.
Errico, F. d‘. 1994. Birds of the Grotte Cosquer: theGreat Auk and Paleolithic prehistory. Antiquity 68:39–47.
Errico, F. d‘., Ucelli Gnesutta, P. 1999. L'art mobilierépigravettien de la grotte de Settecannelle (Viterbo,Italie). Contexte archéologique, analyse technique etstylistique. L'Anthropologie 103: 121–160.
Escalon de Fonton, M., Bonifay, M.F., Onoratini, G.1979. Les industries de filiation magdalénienne dansle Sud-Est de la France, leur positions géo-chronolo-giques et les faunes. In: de Sonneville-Bordes, D.(ed.), La fin des temps glaciaires en Europe.Colloques internationaux du CNRS. N° 271: 269–286.Paris: CNRS.
Estévez, J. 1978. Primer hallazgo de buey almizcladoen el Pleistoceno peninsular. Acta geologicaHispanica 13 (2): 59–60.
Estévez, J. 1979. La fauna del Pleistoceno catalàn.Tesis doctoral. Cerdanyola del Vallés, UniversidadAutonoma de Barcelona.
Estévez, J. 1987. La fauna de l'Arbreda (sector Alfa)en el conjunt de faunes del Plistocè Superior. Cypsela6: 73–87.
Estévez, J., Vila, A. 1999. Piedra a piedra. Historiade la construcción del Paleolítico en la PenínsulaIbérica. BAR International Series 805. Oxford:Archaeopress.
Féblot-Augustins, J. 1997. La Circulation desMatières premières au Paléolithique. ERAUL 75.Liège: Université de Liège.
Felten, W. 1990. Herakles and the Erymanthian Boar(Labour III). In: Lexicon iconographicum mytho-logiae (LIMC) V.1: 43–48.
Feruglio, V., Robinau, D. 2001. Sur une gravure decétacé de la grotte de La Vache (Ariège, France).Bulletin de la Société Préhistorique Ariège-Pyrénées56: 63–72.
Filip, J. 1969. Enzyklopädisches Handbuch zur Ur-und Frühgeschichte Europas. Stuttgart: Kohlhammer.

Bibliographie 195
Fischer, A. (ed.) 1995. Man and sea in the Mesolithic.Oxford: Oxbow Brooks.
Florineth, D. 1998. Surface geometry of the LastGlacial Maximum (LGM) in the southeastern SwissAlps (Graubünden) and its paleoclimatological signifi-cance. Eiszeitalter und Gegenwart 48: 23–37.
Florineth, D., Schlüchter, Ch. 2000. Alpine Evidencefor Atmospheric Circulation Patterns in Europe duringthe Last Glacial Maximum. Quaternary Research 54:295–308.
Floss, H. 1994. Rohmaterialversorgung im Paläoli-thikum des Mittelrheingebietes. Römisch-Germani-sches Zentralmuseum, Monographien, Band 21.Mainz. Bonn: Rudolf Habelt.
Floss, H. 2002. La Saône: Lien ou limite del’occupation humaine au Paléolithique? In: Bravard,J.-P., Combier, J., Commerçon, N. (dir.) La Saône.Axe de civilisation: 113–125. Lyon: Presses Universi-taires de Lyon.
Floss, H. 2004. Die Kunst der Eiszeit in Europa. In:Schürle, W. und Conard, N. J. (Hrsg.), ZweiWeltalter. Eiszeitkunst und die Bildwelt Willi Baumei-sters: 8–69, 213–217. Ostfildern: Hatje Cantz Verlag.
Fontana, L. 1998. Mobilité et subsistance auMagdalénien supérieur et final en Auvergne. In:Brugal, J.-Ph., Meignen, L., Patou-Mathis, M., (dir.)Économie préhistorique: les comportements de subsi-stance au Paléolithique. Actes des XIIIe rencontresinternationales d'archéologie et d'histoire d'Antibes.23 – 24 – 25 octobre 1997: 373–385.
Fontana, L. 2000. Stratégies de subsistance auBadegoulien et au Magdalénien en Auvergne:nouvelles données. In: Pion, G. (dir.) Le PaléolithiqueSupérieur récent: nouvelles données sur le peuplementet l’environnement. Actes de la Table Ronde deChambéry, 12–13 mars 1999. Société PréhistoriqueFrançaise, Mémoires 28: 59–65.
Fortea Pérez, J. 1989. Cuevas de La Lluera. Avance alestudio de sus artes parietales. In: González Morales,M.R. (ed.), Cien años después de Sautuola: 189–202.Santander.
Fortea Pérez, J. 2000. El Pindal, vision nouvelle oufiction? Bulletin de la Société Préhistorique Ariège-Pyrénées 55: 35–62.
Fortelius, M., Mazza, P., Sala, B. 1993. Stephano-rhinus (Mammalia: Rhinocerotidae) of the westernEuropean Pleistocene, with a revision of S.etruscus(Falconer, 1868). Palaeontographia Italica 80: 63–155.
Freeman, L.G., González Echegaray, J., Bernaldo deQuirós, F., Ogden, J. 1987. Altamira revisited.Chicago: Institut for Prehistoric Investigations.
Frenzel, B. 1968. Grundzüge der pleistozänen Vegeta-tionsgeschichte Nord-Eurasiens. Wiesbaden: FranzSteiner.
Frenzel, B. 1980. Klima der Letzten Eiszeit und derNacheiszeit in Europa. In: Klimaänderungen, Menschund Lebensraum. Vorträge gehalten auf der Tagungder Joachim Jungius-Gesellschaft der Wissenschaf-ten, Hamburg, am 20. und 21. Oktober 1979: 9–46.
Fuller, E. 2002. The Great Auk. New York: Harry N.Abrams, Inc.
Gallini, V., Sala, B. 2000. Il giacimento a vertebratifossili di Settepolesini di Bodeno (FE). Padusa 9, 1/2:2–4.
Gallini, V., Sala, B. 2001. Settepolesini di Bodeno(Ferrara Eastern Po Valley): the first example ofmammoth steppe in Italy. In: Cavarretta, G., Gioia, P.,Mussi, M., Palombo, M.R. The World of Elephants.Proceeding of the 1st International Congress, Roma16–20 Ottobre 2001: 272–275. Roma: CNR.
Gamble, Cl. 1979. Hunting Strategies in the CentralEuropean Palaeolithic. Proceeding of the PrehistoricSociety 45: 35–52.
Garrod, D.A.E., Buxton, L.H.D., Smith, G.E., Bate,D.M.A. 1928. Excavation of a Mousterian Rock-Shelter at Devil's tower, Gibraltar. Journal of theRoyal Anthropological Institute 58, January–June1928: 33–113.
Garutt, N. 1999. Skull pathologies in Coelodontaantiquitatis: implications about social behaviour andecology. In: Haynes, G., Klimowicz, J., Reumer,J.W.F. Mammoths and the Mammoth Fauna: Studiesof an Extinct Ecosystem. Proceedings of the FirstInternational Mammoth Conference. St. Petersburg,Russia. October 16–21, 1995. Deinsea 6, Annual ofthe natural history museum Rotterdam: 175–185.
Gaucher, G. 1987. André Leroi-Gourhan 1911–1986.Bulletin de la société préhistorique française 84: 302–323.

196 Verbreitung der großen Jagdfauna in Mittel und Westeuropa im oberen Jungpleistozän
Gaudzinski, S. 1996. On Bovid Assemblages and theirConsequences for the Knowledge of SubsistencePatterns in the Middle Paleolithic. Proceedings of thePrehistoric Society 62: 19–39.
Gaussen, J. 1964. La grotte ornée de Gabillou.Bordeaux: Imprimeries Delmas.
Gaussen, J. 1993. Les léporidés. In: Aujoulat, N. et al.,L'art pariétal paléolithique. Documents préhisto-riques 5: 173–180. Paris: Comité des Travaux Histo-riques et Scientifiques.
Gaussen, J., Merlaud, B. 1987. Une identificationdifficile. Préhistoire Ariégeoise 42: 165–169.
Gessain, R. 1981. Ovibos. La grande aventure deshommes et des boeufs musqués. Paris: Éditions RobertLaffont.
Geyh, M.A., Schreiner, A. 1984. 14C-Datierung anKnochen- und Stoßzahn-Fragmenten aus würmeiszeit-lichen Ablagerungen im westlichen Rheingletscher-gebiet (Baden-Württemberg). Eiszeitalter undGegenwart 34: 155–161.
Gibbons, A. 2003. A miss for Moderns andNeandertals. Science 300: 893–894.
Gioia, P. 1988. Problems related to the origins ofitalian Upper Palaeolithic: Uluzzian and Aurignacian.In: Kozłowski, J.K. (ed.), L'Homme de Néandertal vol.8, ERAUL. 35: 71–101.
Girard, I. 1995. Le cheval de Przewalski: sa dispa-rition et les tentatives actuelles de reintroduction.Thèse pour le diplôme d'Etat de Docteur Veterinairepresenté le 27 juin 1995 à l'ecole nationale veterinairede Nantes.
Giraudi, C., Mussi, M. 1999. The Central andSouthern Apennine (Italy) during OIS 3 and 2: thecolonisation of a changing environment. ERAUL 90:118–129.
Glory, A. 1949. Les gravures préhistoriques de l'AbriDelluc. Les Eyzies (Dordogne). Bulletin de la SociétéPréhistorique Française 36: 217–219.
González, S., Kitchener, A., Lister, A.M. 2000.Survival of the Irish elk into the Holocene. Nature405: 753–754.
González Morales, M.R. 1994. Pero ... ¿Hubo algunavez mil bisontes? Los temas del arte parietal Paléo-litico de la región cantábrica. Complutum 5: 291–302.
González Sáinz, C., Montes Barquín, R., MuñózFernández, E. 1993. La cueva de Sovilla (San Felicesde Buelna, Cantabria). Zephyrus 46: 7–36.
González Sáinz, C., San Miguel Llamosas, C. 1996.Les grottes du défilé de Carranza. Nouveauxensembles rupestres paléolithiques dans la régionCantabrique. International Newsletter on Rockart: 12–13.
González Sáinz, C. 2002. Sovilla. In: ACDPS (ed.),Las cuevas con arte paleolitico en Cantabria: 131–134. Santander: Cantabria en imágenes undAsociación Cántabra para la Defensa del PatrimonioSubterráneo (ACDPS).
Gould, St. J. 1974. The origin and function of„bizarre“ structures: antler size and skull size in the„irish elk“, megaloceros giganteus. Evolution 28: 191–220.
Graziosi, P. 1956. Die Kunst der Altsteinzeit. Stuttgart:Kohlhammer.
Graziosi, P. 1962. Levanzo. Pitture e incisioni.Firenze: Sansoni.
Griggo, Ch., 1991. Le bouquetin de Malarnaud(Ariège); implications paléobiogéographiques.Quaternaire 2: 76–82.
Griggo, Ch. 1995. Significations paléoenvironnemen-tales des communautés animales pléistocènesreconnues dans l'abri Suard (Charente) et la grotte deBois-Ragot (Vienne): Essai de quantification devariables climatiques. Thèse présentée à l'universitéde Bordeaux I.
Groenen, M. 1994. Pour une Histoire de la Préhi-stoire. Grenoble: Jérôme Millon.
Gröning, K., Saller, M. 1998. Der Elefant in Naturund Kulturgeschichte. Könemann, Köln.
Groiss, J.Th. 1996. Der Höhlentiger Panthera tigrisspelaea (Goldfuss). Neues Jahrbuch für Geologie undPaläontologie, Monatshefte: 399–414.
Guérin, C., Faure, M. 1983. Les hommes du paléo-lithique européen ont-ils chassé le rhinocéros? In:Poplin, F. (dir.), La faune et l’homme préhistoriques.Dix études en hommage à Jean Bouchud. Mémoiresde la Société préhistorique française, 16: 29–36. Paris:Société Préhistorique française.

Bibliographie 197
Guérin, C., Patou-Mathis, M. 1996. Les grandsmammifères plio-pléistocènes d'Europe. Paris, Milano,Barcelona: Masson.
Guidi, A., Piperno, M. 1992. Italia preistorica.Manuali Laterza, Volume 34. Bari: Laterza.
Guintard, Cl., 1999. On the size of the Ure-Ox orAurochs (Bos primigenius bojanus, 1827). In:Weniger, G.-Ch. (Hrsg.), Archäologie und Biologiedes Auerochsen. Archaeology and Biology of theAurochs. Wissenschaftliche Schriften des NeanderthalMuseums 1: 7–21. Mettmann: Neanderthal Museum.
Gurioli, F. 2002. Analisi dei reperti in materia duraanimale provenienti dal deposito aurignaziano dellaGrotta di Fumane (VR): Strumenti, Oggettiornamentali e Superfici con tracce di interventiantropici. Tesi di laurea in Scienze Naturali pressol'universitá di Ferrara.
Guthrie, R.D. 1990. Frozen fauna of the mammothsteppe. Chicago: The University of Chicago Press.
Guthrie, R.D. 2001. Reconstructions of WoollyMammoth life history. In: Cavarretta, G., Gioia, P.,Mussi, M., Palombo, M.R. The World of Elephants.Proceeding of the 1st International Congress, Roma16–20 Ottobre 2001: 276–279. Roma: CNR.
Hahn, J. 1986. Kraft und Aggression. Die Botschaftder Eiszeitkunst im Aurignacien Süddeutschlands?Tübingen: Archaeologica Venatoria.
Hahn, J., Müller-Beck, H., Taute, W. 1985. Eiszeit-höhlen im Lonetal. Führer zu archäologischenDenkmälern in Baden-Württemberg 3. Stuttgart:Theiss.
Hahn, J., Kind, Cl.-J., Steppan, K. 1993. Mesoli-thische Rentier-Jäger in Südwestdeutschland?Fundberichte aus Baden-Württemberg 18: 29–52.
Hanitzsch, H. 1972. Groitzsch bei Eilenburg. Schlag-und Siedlungsplätze der späten Altsteinzeit. Berlin:VEB. Deutscher Verlag der Wissenschaften.
Hantke, R. 1978. Eiszeitalter. Die jüngste Erdge-schichte der Schweiz und ihrer Nachbargebiete. Band1. Thun: Ott.
Haynes, G., Klimowicz, J., Reumer, J.W.F. 1999.Mammoths and the Mammoth Fauna: Studies of anExtinct Ecosystem. Proceedings of the First Interna-tional Mammoth Conference. St. Petersburg, Russia.
October 16–21, 1995. Deinsea 6, Annual of thenatural history museum Rotterdam. Rotterdam:Natural history museum Rotterdam.
Hedges, R.E.M., Pettitt, P.B., Bronk Ramsey, C., vanLinken, G.J. 1998. Radiocarbon dates from the oxfordAMS system: archaeometry datelist 25. Achaeometry40, 1: 227–239.
Hediger, H. 1980. Tiere verstehen: Erkenntnisse einesTierpsychologen. München: Kindler.
Heidemann, G. 1986. Cervus dama (Linnaeus, 1758) –Damhirsch. In: Niethammer, J. Krapp, F. (Hrsg.),Handbuch der Säugetiere Europas. Band 2/IIPaarhufer: 140–158. Wiesbaden: Aula.
Heptner, V.G., Nasimovic , A.A., Bannikov, A.G.1966. Die Säugetiere der Sowjetunion. Band I.Paarhufer und Unpaarhufer. Jena: VEB GustavFischer.
Heptner, V.G., Sludskij, A.A. 1980. Die Säugetiereder Sowjetunion. Band III. Raubtiere (Feloidea). Jena:VEB Gustav Fischer.
Hernández Carrasquilla, F. 1995. El papel de las avesen la reconstrucción paleoecológica. Su aplicación alcuaternario del sur de la península Ibérica. ThesisDoctoral a la Universidad Autónoma de Madrid,Facultad de ciencias, departamento de biología.
Hernández-Pacheco, E. 1919. La caverna de la Penadel Candamo (Asturias). Comision de investigacionespaleontologicas y prehistóricas, Memoria número 24.Madrid: Museo nacional de ciencias naturales.
Herre, W. 1986a. Sus scrofa Linnaeus, 1758 –Wildschwein. In: Niethammer, J. Krapp, F. (Hrsg.),Handbuch der Säugetiere Europas. Band 2/IIPaarhufer: 36–66. Wiesbaden: Aula.
Herre, W. 1986b. Rangifer tarandus (Linnaeus, 1758)– Ren, Rentier. In: Niethammer, J. Krapp, F. (Hrsg.),Handbuch der Säugetiere Europas. Band 2/IIPaarhufer: 198–216. Wiesbaden: Aula.
Hodgson, D. 2003. Seeing the „Unseen“: FragmentedCues and the Implicit in Palaeolithic Art. CambridgeArchaeological Journal 13, 1: 97–106.
Hockett, B.S., Bicho, N.F. 2000. The Rabbits ofPicareiro Cave: Small Mammal Hunting During theLate Upper Palaeolithic in the Portuguese Estre-madura. Journal of Archaeological Science 27: 715–723.

198 Verbreitung der großen Jagdfauna in Mittel und Westeuropa im oberen Jungpleistozän
Höneisen, M., Peyer, S. (eds.) 1994. Schweizersbild –ein Jägerlager der Späteiszeit. Schaffhauser Archäo-logie 2. Schaffhausen: Stadt Schaffhausen.
Hokr, Z. 1951. A Method of the Quantitative Determi-nation of the Climate in the Quaternary Period byMeans of Mammal Associations. Sbornik of the geolo-gical survey of Czechoslovakia 18: 209–218.
Holdermann, Cl.-St. 2001. Verlorene Kunst. In:Müller-Beck, H., Conard, N.J., Schürle, W. (Hrsg.),Eiszeitkunst im Süddeutsch-Schweizerischen Jura: 30–37. Stuttgart: Theiss.
Jelgersma, S. 1979. Sea-level changes in the NorthSeabasin. In: Oele, E., Schüttenhelm, R.T.E., Wiggers,A.J. (eds.), The Quaternary History of the North Sea.Acta Universitatis Upsaliensis. Annum Quingente-simum Celebrantis 2: 233–248.
Jelinek, J. 1972. Das große Bilderlexikon desMenschen in der Vorzeit. Gütersloh: BertelsmannLexikon-Verlag.
Jelinek, A.J. 1982. François Bordes, 1919-1981.American Antiquity 47: 785–792.
Joger, U., Koch, U. 1994. Mammuts aus Sibirien.Begleitbuch zur Ausstellung im Hessischen Landes-museum Darmstadt von 20.10.1994–19.02.1995.Darmstadt: Hessisches Landesmuseum.
Jordá Cerda, F. 1983. El Mamut en el Arte paleoliticopeninsular y la hierogamia de los Casares. In: RipollPerelló, E. (ed.), Homaje al Prof. Martin AlmagroBasch: 265–272. Madrid: Ministero de Cultura.
Jordá Pardo, J.F. (Coord.) 1986. La Prehistoria de laCueva de Nerja (Malaga). Malaga: Imprenta de launiversidad de Malaga.
Jordá Pardo, J.F., Aura Tortosa, J.E., Rodrigo García,M.J., Pérez Ripoll, M., Badal García, E. 2003. Elregistro paleobiológico cuaternario del yacimientoarqueológico de la Cueva de Nerja (Málaga, España).Boletín de la Real Sociedad Española de HistoriaNatural. Sección Geológica 98 (1–4): 73–89.
Jöris, O. 2002. Die aus der Kälte kamen... Von derKultur Später Neandertaler in Mitteleuropa. Mittei-lungen der Gesellschaft für Urgeschichte 11: 5–32.
Jöris, O., Weninger, B. 1996. Calendric Age-Conversion of glacial Radiocarbon Data at theTransition from the Middle to Upper Palaeolithic inEurope. Bulletin de la Société Préhistorique Luxem-bourgeoise 18: 43–55.
Julià, R., Bischoff, J.L. 1993. Datación radiometricade los depósitos cuaternarios y de la mandíbulahumana del lago de Banyoles. In: Maroto, J. (ed.) Lamandíbula de Banyoles en el context dels fòssilshumans del Pleistocè. Centre d'investigacionsarqueològiques, sèrie monogràfica 13. Girona: PalahíA.G.
Jux, U., Rosenbauer, K. 1959. Zum Vorkommen vonCetaceen-Resten in jungpleistozänen Flußablage-rungen der Niederrheinischen Bucht. Neues Jahrbuchfür Geologie und Paläontologie. Abhandlungen 108:81–126.
Kahlke, H.D., 1975. Der Saiga-Fund von Bottrop/Westfalen. Zur Fossilgeschichte, paläogeographi-schen Verbreitung und Systematik der Gattung SaigaGray, 1843. Quartär 26: 135–146.
Kahlke, R.-D., 1992. Repeated immigration of saigainto Europe. In: Koenigswald, W. von, und Werdelin,L. (eds.), Mammalian Migration and Dispersal Eventsin the European Quaternary. Courier Forschungsin-stitut Senkenberg, 153, 187-195 (Frankfurt am Main).
Kahlke, R.-D. 1994. Die Entstehung-, Entwicklungs-und Verbreitungsgeschichte des oberpleistozänenMammuthus-Coelodonta-Faunenkomplexes in Eura-sien (Graßsäuger). Abhandlungen der Senckenbergi-schen Naturforschenden Gesellschaft 546. Frankfurta.M.
Kapel, J. 1992. Phoca groenlandica Erxleben, 1777 –Sattelrobbe. In: Duguy, R., Robineau, D. (Hrsg.),Handbuch der Säugetiere Europas, Band 6: Meeres-säuger. Teil II: Robben – Pinnipedia: 196–224.Wiesbaden: Aula.
Kempf, Ch. 1990. Los señores del bosque. Barcelona:Lynx edicions.
Klostermann, J. 1999. Das Klima im Eiszeitalter.Stuttgart: Schweizerbart'sche Verlagsbuchhandlung.
Koenigswald, W. v. 1986. Ergänzungen zu Herre, W.Sus scrofa Linnaeus, 1758 – Wildschwein. In:Niethammer, J., Krapp, F. (Hrsg.), Handbuch derSäugetiere Europas, Band 2/II Paarhufer: 36–66.Wiesbaden: Aula.
Koenigswald, W. v. 2002. Lebendige Eiszeit. Klimaund Tierwelt im Wandel. Darmstadt: Wissenschaft-liche Buchgesellschaft.

Bibliographie 199
Koenigswald, W. v., Müller-Beck, H, Pressmar, E.1974. Die Archäologie und Paläontologie in denWeinberghöhlen bei Mauern (Bayern). Grabungen1937–1967. Tübingen: Archaeologica Venatoria.
Koenigswald, W. v., Müller-Beck, H., Pressmar, E.1975. Das Pleistozän der Weinberghöhlen bei Mauern(Bayern). Quartär 26: 107–118.
Koenigswald, W. v., Hahn, J. 1981. Jagdtiere undJäger der Eiszeit. Sonderausstellung im HessischenLandesmuseum Darmstadt, vom 26.11.1981–31.01.1982. Stuttgart: Theiss.
Kowalski, K. 2000. Der Pleistozäne Ölsumpf beiStarunia, Ukraine. In: Meischer, D. EuropäischeFossillagerstätten: 232–236. Berlin: Springer.
Kristiansen, K. 1985. A Short History of DanishArchaeology. An Analytical Perspective. In: Kristi-ansen, K., Archaeological formation process. Therepresentativity of archaeological remains fromDanish Prehistory. Copenhagen: Nationalmuseet.
Krylov, V.I. 1992. Phoca caspica Gmelin, 1788 –Kaspischer Seehund. In: Duguy, R., Robineau, D.(Hrsg.), Handbuch der Säugetiere Europas, Band 6:Meeressäuger. Teil II: Robben – Pinnipedia: 162–181.Wiesbaden: Aula.
Kubiak, H. 2001. The finds of four woolly rhinoceros(Coelodonta antiquitatis) and one mammoth fromStarunia. Krakau: Wydawnictwo Instytutu Syste-matyki i Ewolucji Zwierzat PAN.
Lambeck, K. 1997. Sea-level change along the FrenchAtlantic and Channel coasts since the time of the LastGlacial Maximum. Palaeogeography, Palaeoclima-tology, Palaeoecology 129: 1–22.
Laming-Emperaire, A. 1972. Art Rupestre et organi-sation sociale. In: Almagro Basch, M., García Guinea,M.A. Santander Symposium: 65–79. Santander: Artesgráficas resma.
Lampe, R. 2002. Post-glacial water-level variabilityalong the south Baltic coast – a short overview.Greifswalder Geographische Arbeiten 27: 13–19.
Lang, G. 1994. Quartäre VegetationsgeschichteEuropas. Jena, Stuttgart, New York: Fischer.
Lartet, É., Christy, H. 1865–1875. Reliquiae Aquita-nicae, being Contributions to Archaeology andPalaeontology of Périgord and the adjoining Provinceof Southern France. London.
Lehmann U. 1969. Die Fauna. In: Wetzel, R.,Bosinski, G. Die Bocksteinschmiede im Lonetal(Markung Rammingen, Kr. Ulm). Veröffentlichungendes Staatlichen Amtes für Denkmalpflege Stuttgart,Reihe A, Heft 15: 133–168. Stuttgart: Müller & Gräff.
Lehmann, E. von, Sägesser, H. 1986. Capreoluscapreolus Linnaeus, 1758 – Reh. In: Niethammer, J.Krapp, F. (Hrsg.), Handbuch der Säugetiere Europas.Band 2/II Paarhufer: 233–268. Wiesbaden: Aula.
Lejeune, M. 1987. L'art Mobilier Paléolithique etMésolithique en Belgique. Artefacts 4. Treignes-Viroinval: Editions du C.E.D.A.
Lenardi, F. 1990. Per una cultura faunisticovenatoria. Udine: Arti grafiche friulane. Leonardi, P. 1988. Art paléolithique mobilier etpariétal en Italie. L'Anthropologie 92: 129–202.
Leonardi, P., Broglio, A. 1962. Il Paleolitico superioredei Colli Berici. Arheološki vestnik 13–14: 109–128.Leroi-Gourhan, A. 1965. Préhistoire de l'artoccidental. Paris: Mazenod.
Leroi-Gourhan, A. 1971. Prähistorische Kunst. DieUrsprünge der Kunst in Europa. Freiburg, Basel,Wien: Herder.
Leroi-Gourhan, A. 1995. Préhistoire de l'artoccidental. Nouvelle édition revue et augmenté parBrigitte et Gilles Delluc. Paris: Citadelles & Mazenod.
Leroi-Gourhan, A. et al. 1984. L'art des cavernes.Atlas des grottes ornées Paléolithiques françaises.Paris: Imprimerie nationale.
Leroi-Gourhan, A., Brezillon, M. 1983. Fouilles dePincevent. Essai d'analyse ethnographique d'unhabitat magdalénien (la section 36). Paris: CNRS.
Leroi-Gourhan, Arl., Allain, J. 1979. Lascauxinconnou. Paris: CNRS.
Le Tensorer, J.-M. 1998. Le Paléolithique en Suisse.Collection des origines, Série „Préhistoire d'Europe“N° 5. Grenoble: Millon.
Lhote, H. 1988. À propos de la „Lionne“ des Trois-Frères. L'Anthropologie 92: 371–372.
Lister, A., Bahn, P.G. 1997. Mammuts. Die Riesen derEiszeit. Sigmaringen: Thorbecke.

200 Verbreitung der großen Jagdfauna in Mittel und Westeuropa im oberen Jungpleistozän
Lorblanchet, M. 1984. Grotte de Roucadour. In: Leroi-Gourhan, A. et al. L'art des cavernes, Atlas des grottesornées paléolithiques françaises: 511–513. Paris:Imprimerie Nationale.
Lorblanchet, M. 1993. Les Poissons. In: Aujoulat, N.et al., L'art pariétal paléolithique. Documents préhi-storiques 5: 181–190. Paris: Comité des TravauxHistoriques et Scientifiques. Lorblanchet, M. 1995. Les grottes ornées de la préhi-stoire, nouveaux regards. Paris: Errance.
Lowe, J.J., Walker, M.J.C. 1984. Reconstructingquaternary environments. Singapore: Longman scien-tific & technical.
Lucas Pellicer, M.R. 2000. Arte rupestre paleolítico enMadrid: La cueva del Reguerillo. Boletín de laAsociación Española de Amigos de la Arqueologia39–40: 127–139.
Lumley, H. de, Kahlke, H.-D., Moigne, A.-M.,Moulle, P.-E. 1988: Les faunes de grands mammi-feres de la grotte du Vallonnet. Roquebrune-Cap-Martin, Alpes-Maritimes. L’Anthropologie 92: 465–496.
Madariaga de la Campa, B. 1964. El mar y el hombreprehistórico. Zephyrus 15: 37–45.
Madariaga de la Campa, B. 1966. In: GanzálesEchegaray, P.J., Garciá Guinea, M.A., BeginesRamírez, A. Cueva del Otero. Excavaciones Arqueo-logicas en España: 53: 61–80. Madrid: Langa y Cía.
Madariaga de la Campa, B. 1976. Marcelino Sanz deSautuola, escritos y documentos. Santander: Insti-tucion cultural de Cantabria.
Made, van der, J., Mazo, A.V. 2001. SpanishPleistocene Proboscidean diversity as a function ofclimate. In: Cavarretta, G., Gioia, P., Mussi, M.,Palombo, M.R. (eds.), The World of Elephants.Proceeding of the 1st International Congress, Roma16–20 Ottobre 2001: 214–218. Roma: CNR.
Mania, D. 1999. Nebra – eine jungpaläolithischeFreilandstation im Saale-Unstrut-Gebiet. Veröffentli-chungen des Landesamtes für Archäologie Sachsen-Anhalt – Landesmuseum für Vorgeschichte – Band54. Halle (Saale): Landesamt für Archäologie.
Mantegazza, P. 1877. Rezension von A. de Quatre-fages, L'Espèce humaine. Archivio per l'antropologiae l'etnologia 7: 26–28.
Maroto, J. 1994. El pas del paleolític mitjà alpaleolític superior a Catalunya i la seva interpretaciódins del context geogràfic franco-ibèric. Tesi Doctoralal Departement de Geografia, Història i Història del'Art, Facultat de Lletres, Universitat de Girona.
Maroto, J., Soler, N., Fullola, J.M. 1996. Culturalchange between Middle and Upper Palaeolithic inCatalonia. In: Carbonell, E., Vaquero, M. (eds.), Thelast Neandertals, the first anatomically ModernsHumans: A tale about the human diversity. Culturalchange and human evolution: the crisis at 40 KA BP.Santa Margarida de Montbui: Grafiques Lluc.
Marshack, A. 1970. Le bâton de commandement deMontgaudier (Charente). Réexamen au microscope etinterprétation nouvelle. L'Anthropologie 74: 321–352.Martin, P., Klein R.G. 1984. Quaternary Extinctions.Tucson, Arizona: The university of Arizona Press.
Masini, F., Abbazzi, L. 1997. L'associazione dimammiferi della Grotta di Castelcivita. In:Gambassini, P. (ed.), Il Paleolitico di Castelcivita:cultura e ambiente: 33–59. Napoli: Electa.
Masseti, M., Rustioni, R. 1988. Considerazioni preli-minari sulla diffusione di Dama dama (Linneus, 1758)durante le epoche tardiglaciale e postglaciale nell'Italiamediterranea. Studi per l'ecologia del Quaternario 10:93–119.
Masseti, M., Mazza, P., Rustioni, M., Sala, B. 1995.Large-sized italian ungulates at the late Pleistocene-Holocene transition: an overwiew. Atti del 1°convegno nazionale di archeozoologia. Rovigo 5–7Marzo 1993. Padusa Quaderni 1: 89–96. Rovigo:Centro Polesano di studi storici, archeologici edetnografici di Rovigo.
Meadow, R.H., Uerpmann, H.-P. 1991. Equids in theancient World. Wiesbaden: Ludwig Reichert.
Meyer, H.-H., Kottmaier, Chr. 1989. Die atmosphä-rische Zirkulation in Europa im Hochglazial derWeichsel-Eiszeit – abgeleitet von Paläowind-Indika-toren und Modellsimulationen. Eiszeitalter undGegenwart 39: 10–18.
Ministère de la Culture, Société des Amis du Musée etde la Recherche Archéologique (éds.) 1990. Lartet,Breuil, Peyrony et les autres. Sarlat: Imprimerienouvelle.
Mitchell, G.F., Parkes, H.M. 1949. The giant deer inIrland. Proceeding of the Royal Irish Academy 52(B7): 291–314.

Bibliographie 201
Mitchell-Jones, A.J., Amori, G., Bogdanowicz, W., etal. 1999. The atlas of european Mammals. London:Academic Press.
Mohr, E. 1952. Die Robben der europäischenGewässer. Frankfurt am Main: Paul Schöps.
Mol, D., van Essen, H. 1992. De mammoet. Sporen uitde Ijstijd. Den Haag: Uitgeverij BZZTôH.
Morel, Ph. 1998. La grotte du Bichon (La Chaux-deFonds, canton de Neuchâtel, Suisse): un site archéolo-gique singulier, ou l'histoire d'une chasse à l'ours brunil y a 12.000 ans dans le Jura suisse. In: Cupillard,Chr., Richard, A. (dir.), Les derniers chasseurs-cueil-leurs du massif jurassien et de ses marges: 88–93.Lons-le-Saurier: Centre Jurassien du Patrimoine.
Morel, Ph., Müller, W. 1997. Un campementmagdalénien au bord du lac de Neuchâtel, étudearchéozoologique (secteur 1). Archéologieneuchâteloise 23. Neuchâtel.
Mortillet, G. de, 1872. Classification des diversespériodes de l'Age de la Pierre. In: Congrès Interna-tional d’Anthropologie et Archéologie Préhistoriques,6ème session. Bruxelles: 432–459.
Mourer-Chauviré, C., Antunes, M.T. 1991. Pinguinusimpennis (Aves, Charadriformes) dans le Pléistocènedu Portugal. Géobios 24: 201–205.
Moure Romanillo, A. 1989. La caverne de TitoBustillo (Asturies, Espagne). Le gisement paléo-lithique. L'Anthropologie 93: 407–434.
Moya-Sola, S., Augusti, J., Gilbert, J., Pons-Moya, J.1981. El yacimiento cuaternario de Venta Micena(España) y su importancia entro de las asociacionesfaunisticas del Pleistoceno Medio inferior europeo.Paleontologia i Evolució 16: 39–53.
Müller-Beck, H. 2001. Was bleibt ? Die Rolle desZufalls. In: Müller-Beck, H., Conard, N.J., Schürle,W. (Hrsg.), Eiszeitkunst im Süddeutsch-Schweizeri-schen Jura: 23–30. Stuttgart: Theiss.
Müller-Beck, H., Albrecht, G. (Hrsg.) 1987. DieAnfänge der Kunst vor 30.000 Jahren. Stuttgart:Theiss.
Müller-Beck, H., Conard, N.J., Schürle, W. (Hrsg.)2001. Eiszeitkunst im Süddeutsch-SchweizerischenJura. Stuttgart: Theiss.
Müller-Karpe, H. 1966. Handbuch der VorgeschichteI, Altsteinzeit. München: Beck'sche Verlag.
Münzel, S. 1988. Quantitative analysis and archaeolo-gical site interpretation. ArchaeoZoologia II (1.2).Actes du 5e congrès international d'archéozoologie.Bordeaux – Août 1986: 93–109. Bordeaux.
Münzel, S. 2001. The production of Upper Palaeo-lithic mammoth bone artifacts from southwesternGermany. In: Cavarretta, G., Gioia, P., Mussi, M.,Palombo, M.R. (eds.), La Terra degli Elefanti. Atti del1° Congresso Internazionale, Roma 16–20 Ottobre:448–454. Roma: CNR.
Münzel, S. 2004. Subsistence Patterns in theGravettian of the Ach Valley. A former tributary ofthe Danube in the Swabian Jura. In: Svoboda, J.A.,Sedláčková L. (eds.), The Gravettian along theDanube. Proceeding of the Mikulov conference, 20.–21. November 2002. The Dolní Věstonice Studies,Vol. 11: 71–85. Brno: Institute of Archaeology
Münzel, S., Langguth, K., Conard, N.J., Uerpmann,H.-P. 2001. Höhlenbärenjagd auf der SchwäbischenAlb vor 30.000 Jahren. Archäologisches Korrespon-denzblatt 31: 317–328.
Münzel, S., Conard, N.J. 2004. Change and continuityin subsistence during the Middle and Upper Paleo-lithic in the Ach Valley of Swabia (South-westGermany). International Journal of Osteoar-chaeology 14: 225–243.
Musil, R. 1974. Tiergesellschaft der Kniegrotte. In:Feustel, R. Die Kniegrotte. Veröffentlichungen desMuseums für Ur- und Frühgeschichte Thüringens 9:30–95.
Musil, R. 1978. Die fossilen Equiden-Reste aus denTravertinen von Burgtonna in Thüringen. Quartärpa-läontologie 3: 137–138.
Musil, R. 1985. Die Fauna der Magdalénien-SiedlungOelknitz. Weimarer Monographien zur Ur- undFrühgeschichte 17. Museum für Ur- und Frühge-schichte Thüringens, Weimar. Gera: Druckerei Volks-wacht.
Mussi, M. 1992. Il Paleolitico e il Mesolitico in Italia.Popoli e civiltá dell'Italia antica, Volume 10 (Treviso).Bologna: Stilus BSP.
Mussi, M. 2001. Earliest Italy. An Overview of theItalian Paleolithic and Mesolithic. New York: KluwerAcademic / Plenum Publishers.
Mussi, M., Zampetti, D. 1987. La presenza umananella pianura Pontina durante il paleolitico medio esuperiore. Origini 13, 1984–1987: 7–26.

202 Verbreitung der großen Jagdfauna in Mittel und Westeuropa im oberen Jungpleistozän
Nadal, J., Albert, R.M., Juan, J. 1997. Nuevas aporta-ciones arqueozoológicas y arqueobotánicas delyacimiento magdaleniens de la Bora Gran d'enCarreras (Serinyà, Pla de l'Estany). In: Fullola, J.M.,Soler, N. (dir.), El Món Mediterrani després delpleniglacial (18.000–12.000 BP). Colloqui. Banyoles1995. Centre d'investigacions arqueològiques, Sèriemonografica núm. 17: 365–373. Girona: Palahí A.G.
Narr, K.J. 1966. Religion und Magie in der jüngerenAltsteinzeit. In: Narr K.J. (Hrsg.), Handbuch derUrgeschichte 1. Ältere und mittlere Steinzeit: 298–320. München, Bern: Francke.
Nauman, J. 1903. Naturgeschichte der Vögel Mitteleu-ropas. (Hennicke, C.R. Hrsg.) Band XII. Gera:Köhler.
Nenquin, J. 1994. A short history of the internationalunion of prehistoric and protohistoric sciences, and istorganisation. Bollettino del XIII Congresso U.I.S.P.P.n° 2: 34–38.
Neugebauer-Maresch, Chr. 1999. Le Paléolithique enAutriche. Collection L'hommes des origines, série„Préhistoire d'Europe“ N° 8. Grenoble: Millon.
Niethammer, J. Krapp, F. 1986. Handbuch der Säuge-tiere Europas. Band 2/II Paarhufer. Wiesbaden: Aula.
Nievergelt, B., Zingg, R. 1986. Capra ibex Linnaeus,1758 – Steinbock. In: Niethammer, J. Krapp, F.(Hrsg.), Handbuch der Säugetiere Europas. Band 2/IIPaarhufer: 384–404. Wiesbaden: Aula.
Nougier, L.-R., Robert, R. 1957. Le Rhinocéros dansl'art franco-cantabrique occidental. Bulletín de laSociété Préhistorique de l'Ariège. Préhistoire, Spéléo-logie Ariégeoises 12: 16–52.
Nougier, L.-R., Robert, R. 1958. Le „Lissoir auxSaigas“ de la grotte de la Vache, à Alliat, et l'antilopeSaiga dans l'art franco-cantabrique. Bulletín de laSociété Préhistorique de l'Ariège. Préhistoire, Spéléo-logie Ariégeoises 13: 13–44.
Nougier, L.-R., Robert, R. 1965. Le „Frise des Lions“gravure sur os de la grotte de la Vache, à Alliat, et lesgrands félins dans l'art franco-hispanique. Bulletín dela Société Préhistorique de l'Ariège. Préhistoire,Spéléologie Ariégeoises 20: 17–84.
Novel, Ph. 1986. Les animaux rares dans l'art pariétalaquitain. Préhistoire Ariégeoise 41: 63–93.
Novel, Ph. 1987. Les animaux rares dans l'art pariétalaquitain (suite). Préhistoire Ariégeoise 42: 83–118.
Nowak, J., Pankow, E., Tokarski, J., Szafer, W.L.,Stach, J. 1930. The second woolly rhinoceros (Coelo-donta antiquitatis BLUM.) from Starunia, Poland.Bulletin International de l’Académie Polonaise desSciences et des Lettres: 1–45.
Nygrén, K. 1986. Alces alces (Linnaeus, 1758) – Elch.In: Niethammer, J. Krapp, F. (Hrsg.), Handbuch derSäugetiere Europas. Band 2/II Paarhufer: 173–197.Wiesbaden: Aula.
Olins Alpert, B. 1992. Des preuves de sens ludiquedans l'art au pléistocène supérieur. L'Anthropologie96: 219–244.
Ortega y Gasset, J. 1957. Über die Jagd. Hamburg:Rowohlt.
Otte, M. 1979. Le Paléolithique Superieur ancien enBelgique. Musées Royaux d'art et d'histoire, Monogra-phies d'Archéologie Nationale 5. Bruxelles.
Pales, L., de Saint Pereuse, M. 1969. Les gravures dela Marche. I: Félins et ours. Bordeaux: Delmas.
Pales, L, de Saint Pereuse, M. 1981. Les gravures dela Marche. III: Équidés et Bovidés. Paris: Ophrys.
Pales, L, de Saint Pereuse, M. 1989. Les gravures dela Marche. IV: Cervidés, Mammouths et divers. Paris:Ophrys.
Pallarès, M., Wernert, P. 1915–1920. Paleolithic. Elsolutrià de Sant Julià de Ramis: El Cau de las Goges.Annuari de l’Institut d’Estudis Catalans 6: 425–444.
Palma di Cesnola, A. 1966. Il Paleolitico SuperioreArcaico (facies uluzziana) della Grotta del Cavallo,Lecce (continuazione). Rivista di Scienze Preistoriche21: 3–59.
Palma di Cesnola, A. 1988: Paglicci. RignanoGarganico. Mostra iconografica permanente. Foggia:Regione Puglia.
Palma di Cesnola, A. 1993. Il Paleolitico Superiore inItalia. Introduzione allo studio. Firenze: Garlatti &Razzai.
Passemard, E. 1944. La caverne d'Isturitz en PaysBasque. Préhistoire 9: 7–95.
Pedersen, A. 1958. Der Moschusochs. WittenbergLutherstadt: Ziemsen.

Bibliographie 203
Péquart, M, Péquart, St.-J. 1926. Un kjökkenmöddingmorbihannais: Er Yoh. Premier outillage en osdécouvert dans le Morbihan. Revue Antropologique36: 206–211.
Péquart, M., Péquart, St.-J., Boule, M., Vallois, H.1937. Téviec station-nécropole mésolithique duMorbihan. Archives de l’institut de paléontologiehumaine, Mémoires 18. Paris: Masson et Cie éditeurs.
Pérez Ripoll, M. 1986. Avance al estudio de losmamiferos de la cueva de Nerja. In: Jordá Pardo, J.F.(coord.), La Prehistoria de la Cueva de Nerja(Malaga): 98–106. Malaga: Universidad de Malaga.
Pérez Ripoll, M., Raga, J. A. 1998. Los mamíferosmarinos en la vida y en el arte de la prehistoria de lacueva de Nerja. In: Sanchidrían Torti, J. L., SimónVallejo, M.D. (coord.), Las culturas del pleistocenosuperior en Andalucia: 251–275.
Pericot García, L. 1942. La cueva del Parpalló(Gandia). Madrid: Consejo Superior de Investiga-ciones Científicas, Instituto Diego Velazques.
Pericot García, L., Ripoll Perelló, E. 1964. Prehi-storic art of the western mediterranean and theSahara. Chicago: Aldine.
Perlès, C. 1977. Préhistoire du Feu. Paris: Masson.
Pettitt, P.B. 2003. Discovery, nature and preliminarythoughts Britain’s first cave art. Capra 5.http://www.shef.ac.uk/~capra/5/pettitt.html
Pettitt, P.B., Bahn, P.G. 2003. Current problems indating Palaeolithic cave art: Candamo and Chauvet.Antiquity 77: 134–141.
Peyron, O., Guiot, J., Cheddadi, R., Tarasov, P.,Reille, M., de Beaulieu, J.-L., Bottema, S., Andrieu,V. 1998. Climatic Reconstruction in Europe for18,000 YR B.P. from Pollen Data. QuaternaryResearch 49: 183–196.
Peyrony, D. 1932. Les Abris Lartet et du Poisson àGorges d'Enfer (Dordogne). L’Antropologie 42: 241–268.
Peyrony, H. 1948. Le Périgordien, l’Aurignacien et leSolutréen en Eurasie, d’après les dernières fouilles.Bulletin de la Société Préhistorique Française 45:305–328.
Pieper, H. 1985. The fossil land birds of Madeira andPorto Santo. Bocagiana 88: 1–6.
Piette, E. 1907. L'art pendant l'âge du renne. Paris:Masson.
Pigeaud, R., Colleter, R., Hinguant, St., Molines, N.,Arellano-Moullé, A., Moullé, P.-E., Rodet, J. 2001. Leréveil de la préhistoire en Mayenne. Les Grottes deSaulges. Archéologia 384, décembre 2001: 50–59.
Pike-Tay, A., Cabrera Valdés, V., Bernaldo de Quirós,F. 1999. Seasonal variations of the Middle-UpperPaleolithic transition at El Castillo, Cueva Morín andEl Pendo (Cantabria, Spain). Journal of HumanEvolution 1999: 283–317.
Pitti, C., Tozzi, C. 1971. La grotta del Capriolo e laBuca della Iena pressoMommio (Camaiore, Lucca).Rivista di Scienze Preistoriche 26: 213–258.
Plassard, J. 1999. Rouffignac. Stuttgart: Thorbecke.
Pohar, V. 1997. Late Glacial mammal macrofauna inSlovenia. Quartär 47/48: 149–158.
Pons, A., Campy, M., Guiot, J. 1989. The last climaticcycle in France: the diversity of records. QuaternaryInternational 3/4: 49–56.
Poplin, F. 1976. Les grands vertébrés de Gönnersdorf.Fouilles 1968. Der Magdalénien-Fundplatz Gönners-dorf, Band 2. Wiesbaden: Franz Steiner.
Poplin, F. 1978. Großtierfauna. In: Brunnacker, K.Geowissenschaftliche Untersuchungen in Gönners-dorf. Der Magdalénien-Fundplatz Gönnersdorf, Band4: 98–105. Wiesbaden: Franz Steiner.
Poplin, F. 1983. La dent de cachalot sculptée du Masd'Azil avec remarques sur les autres restes de cétacésde la préhistoire française. La faune et l’homme préhi-storiques. Dix études en hommage à Jean Bouchud.Mémoires de la Société préhistorique française, 16:81–94. Paris: Société Préhistorique française.
Porr, M. 2001. Schamanismus im Aurignacien. EineAnnährung. In: Müller-Beck, H., Conard, N.J.,Schürle, W. (Hrsg.), Eiszeitkunst im Süddeutsch-Schweizerischen Jura: 95–102. Stuttgart: Theiss.
Pucek, Z. 1986. Bison bonasus (Linnaeus, 1758) –Wisent. In: Niethammer, J. Krapp, F. (Hrsg.),Handbuch der Säugetiere Europas. Band 2/IIPaarhufer: 278–315. Wiesbaden: Aula.
Rabeder, G., Nagel, D., Pacher, M. 2000. DerHöhlenbär. Stuttgart: Thorbecke.

204 Verbreitung der großen Jagdfauna in Mittel und Westeuropa im oberen Jungpleistozän
Radimilli, A.M. 1956. Il Paleolitico Superiore dellaGrotta Clemente Tronci presso venere dei Marsi,Territorio del Fucino. Bollettino della SocietáGeologica Italiana 75.2: 94–116.
Radimilli, A.M. 1974. Gli scavi nella Grotta Polesinia Ponte Lucano di Tivoli e la piú antica arte nelLazio. Origines, Istituto Italiano di Preistoria e Proto-storia. Firenze: Sansoni.
Rakovec, I. 1975. Razvoj kvartarne sesalske favneSlovenije (Über quartäre Säugetierfaunen Sloweniens(NW Jugoslawien). Arheološki vestnik 24, 1973: 225–270.
Rathgeber, Th. 1994. Nachweise des pleistozänenMoschusochsen (Ovibos moschatus) in Baden-Württemberg. Stuttgarter Beiträge zur Naturkunde.Serie B (Geologie und Paläontologie), Nr. 214: 1–36.
Rauer-Gross, B. 1992. Sichern und Fluchtverhaltenvon Gemsen Rupicapra rupicapra in einem touristi-schen stark gestörten Gebiet der Ostalpen (Raxalpe).Säugerkundliche Mitteilungen 34: 51–57.
Raufuss, I., Koenigswald, W. v. 1999. New remains ofPleistocene Ovibos moschatus from Germany and istgeographic and stratigraphic occurrence in Europe.Geologie en Mijnbouw 78: 383–394.
Reading, R. P. 2001. Erhaltung der Saiga inKalmykien. ZGAP Mitteilungen: 5–7.
Reitz, E. J., Wing, E. S. 1999. Zooarchaeology.Cambridge: Cambridge university press.
Richter, J. 1996. „Out of Africa II“. Die Theorie überdie Einwanderung des modernen Menschen nachEuropa auf dem archäologischen Prüfstand. Archäolo-gische Informationen 19/1&2: 67–73.
Riek, G. 1934. Die Eiszeitjägerstation am Vogelherdim Lonetal. Tübingen: Akademische Verlagsbuch-handlung Franz F. Heine.
Ripoll López, S. 1986. El Solutrense de la cueva deAmbrosio. Excavaciones arqueologicas en España148. Madrid: Ministero de Cultura
Ripoll López, S., Municio González, L.J. 1999.Domingo García. Arte rupestre paleolítico al airelibre en la Meseta Castellana. Arqueologia en Castillay León 8. Madrid: Universidad Nacional de educacióna distancia.
Ripoll Perelló, E. 1984. Notes sur certaines représen-tations d'animaux dens l'art paléolithique de lapéninsule ibérique. In: Bandi, H.-G., Huber, W.,Sauter, M.-R., Sitter, B. (Ed.) La contribution de lazoologie et de l'éthologie à l'interprétation de l'art despeuples chasseurs préhistoriques. 3e Colloque de laSociété suisse de sciences humaines (1979), 263-282.
Robineau, D. 1984. Sur les mammifères marins dubâton gravé préhistorique de Montgaudier. L’Anthro-pologie 88: 661–664.
Roth, G.D. 1995. Wetterkunde für alle. 7. Auflage.München, Zürich, Berlin: BLV.
Rousseau M. 1967. Les grands félins dans l’art denotre préhistoire. Paris: Picard.
Rouzaud, F., 2002. L'ours dans l'art paléolitique. In:Tillet, Th., Binford, L., L'ours et l'homme. ERAUL100: 201–217.
Ruddiman, W. F., McIntyre, A. 1981. The NorthAtlantic ocean during the last deglaciation. Palaeo-geography, Palaeoclimatology, Palaeoecology 35:145–214.
Ruiz de Gaona, M. 1958. Todavia algo más sobre elyacimiento de Coscobilo (Olazagutía). Principe deViana 72–73: 279–287.
Rustioni, M., Ferretti, M.P., Mazza, P., Pavia, M.,Varola, A. 2003. The vertebrate Fauna fromCardamone (Apulia, southern Italy): an example ofmediterranean mammoth fauna. In: Reumer, J.W.F.,de Vos, J., Mol, D. (eds.), Advances in Mammothresearch. Proceedings of the Second InternationalMammoth Conference, Rotterdam, May 16–20 1999.Deinsea 9: 395–403. Rotterdam: Natural historymuseum Rotterdam.
Sacchi, D. 1986. Le paléolithique supérieur duLanguedoc occidental et du Roussillon. 21.Supplément à Gallia Préhistoire. Paris: CNRS.
Sacchi, D. 1993a. Les Caprinés, Antilopinés, Rupica-prinés. In: Aujoulat et al., L'art pariétal paléolithique.Documents préhistoriques 5: 123–136. Paris: Comitédes Travaux Historiques et Scientifiques.
Sacchi, D. 1993b. Les suidés. In: Aujoulat et al., L'artpariétal paléolithique. Documents préhistoriques 5:161–163. Paris: Comité des Travaux Historiques etScientifiques.

Bibliographie 205
Sägesser, H., Krapp, F. 1986. Rupicapra rupicapra(Linnaeus, 1758) – Gemse, Gams. In: Niethammer, J.Krapp, F. (Hrsg.), Handbuch der Säugetiere Europas.Band 2/II Paarhufer: 316–348. Wiesbaden: Aula.
Saint-Exupéry, A. 1946. Le Petit Prince. Paris:Gallimard.
Saint-Périer, R. und Saint-Périer, S. de 1936. La grotted'Isturitz. II. Le Magdalénien de la grande salle.Archives de l'Institut de Paléontologie Humaine,Mémoires 17. Paris: Masson et Cie.
Sala, B. 1980. Faune a grossi mammiferi nelPleistocene superiore. In: AAVV. I vertebrati fossiliitaliani. Catalogo della mostra di Verona. Vago(Verona): La Grafica.
Sala, B. 1985. Variations climatiques et séquenceschronologiques sur la base des variations des associa-tions fauniques à grands mammifères. Rivista discienze preistoriche 38, 1983: 161–180.
Sala, B., Gallini, V. 2002. La Steppa-Taiga a mammute rinoceronti lanosi di Settepolesini. Studi di geomor-fologia, zoologia e paleontologia nel ferrarese.Supplemento al volume 78° degli Atti dell'Accademiadelle Scienze di Ferrara: 39–45. Ferrara.
Sarnthein, M. 1980. Das Paläoklima Nordafrikas derletzten 25 Millionen Jahre – dokumentiert in Tiefsee-Sedimenten. In: Frenzel, B. et al, Klimaänderungen,Mensch und Lebensraum. Tagung der JoachimJungius-Gesellschaft der Wissenschaften, Hamburg,20.–21. Oktober 1979: 47–76. Göttingen: Vanden-hoeck und Ruprecht.
Sauvet, G. 1988. La communication graphique paléo-lithique (De l'analyse quantitative d'un corpus dedonnées à son interpétation sémiologique). L'Anthro-pologie 92: 3–16.
Sauvet, G. 1993. Le problème de la détermination. In:Aujoulat, N. et al., L'art pariétal paléolithique.Documents préhistoriques 5: 83–86. Paris: Comité desTravaux Historiques et Scientifiques.
Sauvet, G. Wlodarczyk, A. 1995. Éléments d’unegrammaire formelle de l’art pariétal paléolithique.L’Anthropologie 99: 193–211.
Sauvet, G., Wlodarczyk, A. 2001. L’art pariétal,miroir des sociétés paléolithiques. Zephyrus 53–54:217–240.
Schmid, E. 1984. Some anatomical observations onthe palaeolithic depictions of horses. In: Bandi, H.-G.,Huber, W., Sauter, M.-R., Sitter, B. (eds.), La contri-bution de la zoologie et de l'éthologie à l'interpré-tation de l'art des peuples chasseurs préhistoriques.3e Colloque de la Société suisse de sciences humaines(1979): 155–160. Fribourg: Editions UniversitairesFribourg.
Schmidt-Thomé, P. 1973. Neue, niedrig gelegeneZeugen einer würmeiszeitlichen Vergletscherung imNordteil der Iberischen Halbinsel. Eiszeitalter undGegenwart 23/24: 384–389.
Schmitz, R., Thissen, J. 2000. First archaeologicalfinds and new human remains at the rediscovered siteof the Neanderthal type specimen. A preliminaryreport. In: Orschiedt, J., Weniger, G.-Ch. (Hrsg.),Neanderthals and Modern Humans – Discussing theTransition. Central and Eastern Europe from 50.000–30.000 B.P. Wissenschaftliche Schriften 2. Mettmann:Neanderthal Museum.
Scott, K. 1986. Man in Britain in the late devensian:evidence from Ossom's cave. In: Roe, D.A. (éd.)Studies in the Upper Palaeolithic of Britain andNorthwest Europe. BAR International Series 296: 63–87. Oxford: Archaeopress.
Schüle, W. 1969. Glockenbecher und Hauspferde. In:Boessneck, J. (Hrsg.), Archäologie und Biologie.Forschungsberichte 15, Deutsche Forschungsgemein-schaft: 88–93. Wiesbaden: Franz Steiner.
Serangeli, J. 2003. La zone côtière et son rôle dans lescomportements alimentaires des chasseurs-cueilleursdu Paléolithique supérieur. In: Patou-Mathis, M.,Bocherens, H., Le rôle de l’environnement dans lescomportements des chasseurs-cueilleurs préhisto-riques. Actes du XIVe Congres UISPP, Liège, 2–8Septembre 2001. BAR International Series 1105: 67–82. Oxford: Archaeopress.
Serangeli, J. 2004. Kraft und Aggression. Existe-t-ilun message de „force“ et d’„agressivité“ dans l’artpaléolithique ? In: Lejeune M. (dir.), L’art pariétalpaléolithique dans son contexte naturel. Actes ducolloque 8.2, Congrès de l’union internationale dessciences préhistoriques et protohistoriques, Liège, 2–6septembre 2001. ERAUL 107: 115–125.
Soergel, W. 1942. Die Verbreitung des diluvialenMoschusochsen in Mitteleuropa. Beiträge zurGeologie von Thüringen 7: 75–95.
Soffer, O. 1985. The Upper Paleolithic of the CentralRussian Plain. San Diego: Academic Press.

206 Verbreitung der großen Jagdfauna in Mittel und Westeuropa im oberen Jungpleistozän
Sonneville-Bordes, D. de 1961. Le Paléolithiquesupérieur en Belgique. L’Anthropologie 65: 421–443.
Sonneville-Bordes, D. de, Perrot, J. 1954. Lexiquetypologique du Paléolithique supérieur. Outillagelithique: I. Grattoirs. Bulletin de la société préhisto-rique française 51, 1953: 323–333.
Sonneville-Bordes, D. de, Laurent, P. 1983. Lephoque á la fin des temps glaciaires. In: Poplin, F., Lafaune et l’homme préhistoriques. Dix études enhommage à Jean Bouchud. Mémoires de la Sociétépréhistorique française 16: 69–80. Paris: SociétéPréhistorique française.
Staesche, U. 1994. Die Tierreste aus den Buntsand-steinabris im Leinebergland bei Göttingen. In: Grote,K., Die Abris im südlichen Leinebergland beiGöttingen. Archäologische Befunde zum Leben unterFelsschutzdächern in urgeschichtlicher Zeit. Teil II,naturwissenschaftlicher Teil. Oldenburg: Isensee.
Steguweit, L. 1999. Die Recken von Schöningen –400.000 Jahre Jagd mit dem Speer. Mitteilungen derGesellschaft für Urgeschichte 8: 5–14.
Stehlin, H.G., Graziosi, P. 1935. Ricerche sugliAsinidi fossili d’Europa. Mémoires de la SociétéPaléontologique Suisse 56: 1–73.
Stodiek, U. 1993. Zur Technologie der jungpaläolithi-schen Speerschleuder. Eine Studie auf der Basisarchäologischer, ethnologischer und experimentellerErkentnisse. Tübinger Monographien zur Urge-schichte 9. Tübingen: Archäologica Venatoria.
Stodiek, U., Paulsen, H. 1996. „Mit dem Pfeil, demBogen ...“. Technik der steinzeitlichen Jagd.Begleitheft zur Ausstellung des Staatlichen Museumsfür Naturkunde und Vorgeschichte Oldenburg.Archäologische Mitteilungen aus Nordwest-deutschland. Beiheft 16. Oldenburg: Isensee.
Storch, V., Welsch U. 1993. Kükenthals Leitfaden fürdas Zoologische Praktikum. 21, Auflage. Stuttgart,Jena: Fischer
Straus, L.G. 1992. Iberia before the Iberians.Albuquerque: University of New Mexico Press.
Straus, L.G. 1995. A través de la frontera Pleistoceno-Holoceno en Aquitania y en la Península Ibérica:Cambios ambientales y respuestas humanas. In:Moure Romanillo, A., González Sáinz, C. El final delpaleolítico cantábrico: 341–363. Santander: Univer-sidad de Cantabria.
Street, M. 1994. Faunal succession and humans subsi-stence in the northern Rhineland 13,000–9,000 BP. In:Fagnart, J.-P., Thévenin, A. (eds.), Le tardiglaciaireen Europe du Nord-Ouest. 119e Congrès national dessociétés historiques et scientifiques, Amiens 26–30octobre 1994: 545–567. Paris: Éditions du Comité destravaux historiques et scientifiques.
Street, M., Terberger, Th. 2000. The German UpperPalaeolithic 35,000–15,000 bp. New dates and insightswith emphasis on the Rhineland. In: Roebroeks, W.,Mussi, M., Svoboda, J., Fennema, K. Hunters of thegolden age. Analecta Praehistorica Leidensia 31: 165–172. Leiden: Faculty of Archaeology.
Stuart, A.J., Sulerzhitsky, L.D., Orlova, L.A., Kuzmin,Y.V., Lister, A.M. 2002. The latest woolly mammoths(Mammuthus primigenius Blumenbach) in Europe andAsia: a review of the current evidence. QuaternarySciences Reviews 21: 1559–1569.
Tagliacozzo, A. 1992. I mammiferi dei giacimenti pre-e protostorici italiani. Un inquadramento peleonto-logico e archeozoologico. In: Guidi, A., Piperno, M.Italia preistorica. Manuali Laterza, Volume 34: 68–102. Bari: Laterza.
Taute, W. 1968. Die Stielspitzen-Gruppen imnördlichen Mitteleuropa. Ein Beitrag zur Kenntnis derspäten Altsteinzeit. Fundamenta, Reihe A, Band 5.Köln: Böhlau.
Teheux, E. 1997. Approche écologique, économiqueet sociale du Magdalénien de la vallée de la Lesse(Belgique). In: Fagnart, J.-P., Thévenin, A. (dir.), Letardiglaciaire en Europe du Nord-Ouest. Actes du119e congrès national des sociétés historiques et scien-tifiques, Amiens 1994: 367–380. Paris: Éditions duComité des travaux historiques et scientifiques.
Terberger, Th. 1997. Die Siedlungsbefunde desMagdalénien-Fundplatzes Gönnersdorf. Konzentra-tionen III und IV. Stuttgart: Steiner.
Terberger, Th. 1998. Siedlungsspuren zwischen20.000 und 16.000 B.P. am Mittelrhein. Germania 76:403–437.
Thiault, M.-H., Roy, J.-B. 1996. L’art préhistoriquedes Pyrénées. Musée des Antiquités nationales –château de Saint-Germain-en-Laye. 2 April – 8 Juin1996. Paris: Réunion des musées nationaux.
Thiede, J. 1978. A glacial Mediterranean. Nature 276:680–683.

Bibliographie 207
Thieme, H. 1997. Altpaläolithische Wurfspeere ausSchöningen, Nordharzvorland. In: Wagner, G.,Beinhauer, W. (Hrsg.), Homo heidelbergensis vonMauer: 304–312. Heidelberg: Uni-Verlag C. Winter.
Trinkaus, E., Milota, St., Rodrigo, R., Mircea, G.,Moldovan, O., 2003a. Early modern human cranialremain from the Pestera cu Oase, Romania. Journal ofHuman Evolution 45: 245–253.
Trinkaus, E., Moldovan, O., Milota, St., Bilgar, A.,Sarcina, L., Athreya, S., Bailey, S.E., Rodrigo, R.,Mircea, G., Higham, Th., Bronk Ramsey, Ch., van derPlicht, J. 2003b. An early modern human from thePestera cu Oase, Romania. PNAS, vol. 100, no. 20:11231–11236.
Trombe, F., Dubuc, G. 1947. Le centre préhistoriquede Ganties-Montespan (Haute-Garonne). Paris:Masson.
Turner, E. 2002. Solutré. An archeozoologicalanalysis of the magdalenian horizon. Römisch-Germanisches Zentralmuseum, Monographien 46.Bonn: RGZM.
Ucko, P. 1987. Débuts illusoires dans l'étude de latradition artistique. Préhistoire Ariégeoise 42: 15–81.
Uerpmann, H.-P. 1976. Equus (Equus) caballus undEquus (Asinus) hydruntinus im Postpleistozän derIberischen Halbinsel (Perissodactyla, Mammalia).Säugetierkundliche Mitteilungen 24: 206–218.
Uerpmann, H.-P. 1987. The ancient distribution ofUngulate Mammals in the Middle East: fauna andarchaeological sites in Southwest Asia and NortheastAfrica. Wiesbaden: Reichert.
Uerpmann, H.-P. 1991. Die Kartierung urgeschicht-licher Umweltgegebenheiten. Ein methodischesProblem. In: Röllig, W. (Hrsg.), Von der Quelle zurKarte: 93–111. Weinheim.
Uerpmann, H.-P. 1997. Buchbesprechung von: HansReichstein, Die Fauna des Germanischen DorfesFeddersen Wierde. Bonner Jahrbücher 197: 432–435.
Uerpmann, H.-P. 1999. Der Rückzucht-Auerochse undsein ausgestorbenes Vorbild. In: Weniger, G.-Ch.(Hrsg.), Archäologie und Biologie des Auerochsen.Archaeology and Biology of the Aurochs. Wissen-schaftliche Schriften des Neanderthal Museums 1: 93–102. Mettmann: Neanderthal Museum.
Valladas, H., Clottes, J. 2003. Style, Chauvet andradiocarbon. Antiquity 77: 142–145.
Vartanyan, S.L., Arslanov, Kh.H., Tertychnaya, T.V.,Chernov, S.B. 1995. Radiocarbon dating evidence formammoths on Wrangel Island, Artic Ocean, until2000 BC. Radiocarbon 37: 1–6.
Velichko, A.A., Frenzel, B., Pécsi, M. 1991. Atlas ofPaleoclimates and Paleoenvironments of the NorthernHemisphere. Late Pleistocene – Holocene. Stuttgart:Fischer.
Velichko, A.A., Zelikson, E.M. 2001. Landscape,climate and mammoth food resources in the EastEuropean plain during the Late Paleolithic. In: Cavar-retta, G., Gioia, P., Mussi, M., Palombo, M.R. (eds.),The World of Elephants. Proceeding of the 1st Inter-national Congress, Roma 16–20 Ottobre 2001: 724–727. Roma: CNR.
Vereshchagin, N.K., Baryshnikov, G.F. 1984.Quaternary Mammalian Extinctions in NorthernEurasia. In: Martin, P., Klein R.G. (eds.), QuaternaryExtinctions: 483–516. Tucson, Arizona: TheUniversity of Arizona.
Verhart, L.B.M. 1995. Fishing for the Mesolithic. TheNorth Sea: a submerged Mesolithic landscape. In:Fischer, A. (ed.) Man and sea in the Mesolithic: 291–302. Oxford: Oxbow Brooks.
Vialou, D. 1984. Les cervidés de Lascaux. In: Bandi,H.-G., Huber, W., Sauter, M.-R., Sitter, B. (eds.), Lacontribution de la zoologie et de l'éthologie à l'interp-rétation de l'art des peuples chasseurs préhistoriques.3e Colloque de la Société suisse de sciences humaines(1979): 199–216. Fribourg: Editions UniversitairesFribourg.
Vigliardi, A. 1968. L'industria litica della grotta di SanTeodoro in provincia di Messina. Rivista di ScienzePreistoriche 23: 33–144.
Vigliardi 1979. Corso introduttivo di preistoria.Firenze: Elite.
Vigne, J.-D. 1988. Les mammifères post-glaciaires deCorse, étude archéozoologique. XXVIe supplément àGallia Préhistoire. Paris: CNRS.
Villaverde Bonilla, V. 1994. Arte Paleolítico de lacova del Parpalló. Valencia: Diputació de Valencia.
Villaverde Bonilla, V., Martinez-Valle, R. 1992.Economia y aprovechamiento del medio en el Paléo-litico de la región central del Mediterráneo español.In: Moure Romanillo (ed.) Elefantes, ciervos y ovica-

208 Verbreitung der großen Jagdfauna in Mittel und Westeuropa im oberen Jungpleistozän
prinos: economía y aprovechamiento del medio en laprehistoria de España y Portugal: 77–95. Santander:Universidad de Cantabria.
Villaverde Bonilla, V., Martinez-Valle, R. 1995.Características culturales y económicas del final delPaleolítico Superior en el Mediterráneo español. In:Villaverde Bonilla, V. (ed.), Los últimos cazadores.Transformaciones culturales y económicas durante eltardiglaciar y el inicio del Holoceno en el ámbitomediterraneo. Patrimonio, V. 22: 79–118. Alicante:Instituto de cultura Juan Gil-Albert.
Villeneuve, L., Boule, M. 1906–1910. Les Grottes deGrimaldi (Baussé – Roussé). Monaco: Imprimerie deMonaco.
Volf, J. 1996. Das Urwildpferd. Die Neue Brehm-Bücherei Bd. 249. Heidelberg, Berlin, Oxford:Spektrum Akademischer Verlag.
Voges, O. (Wissenschaftliche Redaktion) 1980. Inter-nationale Quartärkarte von Europa, 1:2.500.000,Blatt 9 Madrid.
Waechter, A. 1964. The excavation of Gorham's Cave,Gibraltar, 1951–1954. Bulletin of the Institute ofArchaeology 4: 189–221.
Wagner, E. 1981. Eine Löwenkopfplastik von derVogelherdhöhle. Fundberichte aus Baden-Württemberg 6: 29–58.
Wagner, E. 1995. Cannstatt I. Stuttgart: Theiss.
Wandrey, R. 1997. Die Wale und Robben der Welt.Kosmos Naturführer. Stuttgart: Kosmos.
Wehrberger, K. 1994. Der Löwenmensch. In: UlmerMuseum (Hrsg.), Der Löwenmensch. Tier und Menschin der Kunst der Eiszeit. Ausstellung im UlmerMuseum vom 11. September bis 13. November 1994.Ausstellungskatalog: 29–45. Sigmaringen: Thorbecke.
Weinstock, J. 2000. Late Pleistocene reindeer popula-tions in Middle and Western Europe. An osteome-trical study of Rangifer tarandus. BioArchaeologica 3.Tübingen: Mo Vince Verlag.
Weißmüller, W. 1997. Eine Korrelation der delta-18O-Ereignisse des grönländischen Festlandeis mit denInterstadialen des atlantischen und des kontinentalenEuropa im Zeitraum von 45 bis 14 ka. Quartär 47/48:89–111.
Weniger, G.-Ch. 1982. Wildbeuter und ihre Umwelt.Ein Beitrag zum Magdalénien Südwestdeutschlandsaus ökologischer und ethno-archäologischer Sicht.Tübingen: Archaeologica Venatoria.
Weniger, G.-Ch. (Hrsg.), 1999a. Archäologie undBiologie des Auerochsen. Archaeology and Biology ofthe Aurochs. Wissenschaftliche Schriften desNeanderthal Museums 1. Mettmann: NeanderthalMuseum.
Weniger, G.-Ch. 1999b. Representations of theauerochs in the upper palaeolithic and epipalaeolithicon the iberian peninsula. In: Weniger, G.-Ch. (Hrsg.),Archäologie und Biologie des Auerochsen.Archaeology and Biology of the Aurochs. Wissen-schaftliche Schriften des Neanderthal Museums 1:133–140. Mettmann: Neanderthal Museum.
Wernert, P. 1956. Equus Cazurroi Cabrera 1919, oEquus (Asinus) Hydruntinus Regàlia 1907, en elabrigo de Cueto de la Mina (Asturias). In: Librohomenaje al Conde de la Vega de Sella. Oviedo:Diputacion provincial de Asturias.
Williams, M.A., Dunkerley, D.L., Deckker, de, P.,Kershaw, A.P., Chappell, J. 1998. QuaternaryEnvironments. Second Edition. London, New York,Sydney, Auckland: Edward Arnold.
Willis, K., Rudner, E., Sümegi, P. 2000. The Full-Glacial Forests of Central and Southeastern Europe.Quaternary Research 53: 203–213.
Woldstedt, P. 1958. Das Eiszeitalter, zweiter Band.Stuttgart: Enke.
Woldstedt, P., Duphorn, Kl. 1974. Norddeutschlandund angrenzende Gebiete im Eiszeitalter. Stuttgart:Koehler.
Wüst, K. 1999. Auerochsendarstellungen im französi-schen Jungpaläolithikum. In: Weniger, G.-Ch. (Hrsg.),Archäologie und Biologie des Auerochsen.Archaeology and Biology of the Aurochs. Wissen-schaftliche Schriften des Neanderthal Museums 1:141–150. Mettmann: Neanderthal Museum.
Yalden, D. 1999. The history of british mammals.London: Academic press.
Zampetti, D., Caloi, L., Chilardi, S., Palombo, M. R.2000. Le peuplement de la Sicile pendant lePleistocene: l'homme et les faunes. In: Mashkour, M.,Choyke, A.M., Buitenhuis, H., Poplin, F. (eds.),Archaeozoology of the Near East IV A. Proceeding of

Bibliographie 209
the fourth international symposium on the archaeo-zoology of southwestern Asia and adjacent areas.ARC-Publicatie 32. Groningen.
Zemsky, V.A. 1992. Phoca sibirica Gmelin, 1788 –Sibirische oder Baikal-Ringelrobbe. In: Duguy, R.,Robineau, D. (Hrsg.), Handbuch der SäugetiereEuropas, Band 6: Meeressäuger. Teil II: Robben –Pinnipedia: 182–195. Wiesbaden: Aula.
Ziegler, R 1994. Löwen aus dem EiszeitalterSüddeutschlands. In: Ulmer Museum (Hrsg.), DerLöwenmensch. Tier und Mensch in der Kunst derEiszeit. Ausstellung im Ulmer Museum vom 11.September bis 13. November 1994. Ausstellungska-talog: 47–51. Sigmaringen: Thorbecke.
Ziesaire, P. 2000. Zwanzeg Joer „Bulletin de laSociété Préhistorique Luxembourgeoise“. E statiste-schen Iwwerbléck. Bulletin de la Société Préhisto-rique Luxembourgeoise 22: 7–13.
Zilhão, J. 1993. O paleolitico superior em Portugal.Retrospectiva historica e estado dos conhecimientos.In: A.A.V.V. Quaternario em Portugal, Balanço ePerspectivas: 163–172. Lisboa: Colibri.
Zilhão, J. 1997. O Paleolítico superior da estre-madura portuguesa. Lisboa: Colibri.
Züchner, Ch. 1973. Eine Jungpaläolithische Jagddar-stellung aus Mas-d'azil (Ariège, Frankreich). Archäo-logisches Korrespondenzblatt 3: 387–392.
Züchner, Ch. 1988. Buchbesprechung von: Hahn, J.1986. Kraft und Aggression. Die Botschaft derEiszeitkunst im Aurignacien Süddeutschlands?Archaeologica Venatoria 7, Tübingen. BonnerJahrbücher 188: 536–537.
Züchner, Ch. 1999. Grotte Chauvet ArchaeologicallyDated. Communication. International Rock ArtCongress IRAC '98. Vila Real – Portugal 1998. Http://www.uf.uni-erlangen.de/chauvet/chauvet.html.
Züchner, Ch. 2000. Felsbildkunst und PrähistorischeArchäologie. Überlegungen zu einer verkannten Quel-lengattung. Http://www.uf.uni-erlangen.de/felskunst/felsarch.html, 20. 06. 2000.
Züchner, Ch. 2001. Dating rock art by archaeologicalreasoning – an antiquated method. Http://www.uf.uni-erlangen.de/zuechner/rockart/dating/archdating.html,12. 12. 2001.


Katalog der Faunenlisten


Katalog der Faunenlisten
In diesem Teil der Arbeit werden die verwendetenFaunenlisten vorgelegt. Unter jeder Liste ist einebibliographische Angabe zu finden.
Die Angaben nach Arten entsprechen so weit wiemöglich der in dieser Arbeit verwendeten Ansprache,so dass Unterschiede zur zitierten Literatur entstehenkönnen. Beispielsweise wurden die BezeichnungenEquus caballus (u. a. Canal und Carbonell 1989,Delpech 1983), Equus germanicus (u. a. Musil 1985,28), Equus przewalskii (u. a. Weniger 1982, 119)sowie die regionale Bezeichnung Equus caballusantunesi (Cardoso 1993, 241) durch Equus ferusersetzt. Statt Palaeoloxodon antiquus und Stephano-rhinus mercki wurde die Bezeichnung Elephasantiquus und Stephanorhinus kirchbergensisverwendet.
Die Bezeichnungen „MIZ“ und „n“ weisen daraufhin, ob die angegebenen Daten sich auf die Mindestin-dividuenzahlen oder auf die Zahl der bestimmtenKnochen beziehen.
Manche Faunenlisten stammen aus älterenGrabungen oder unterschiedlichen Publikationen, sodass sich bisweilen leichte Unstimmigkeiten ergebenkönnen. In vereinzelten Fällen sind diese Wider-sprüche nicht zu erklären. So zitiert Jesús Altuna1972, Seite 59, Arlette Leroi-Gourhan, die in El PendoReste von Saiga-Antilopen erkannt haben soll, sprichtjedoch im Jahr 1996 (Altuna und Mariezkurrena 1996)vom ersten Nachweis von Saiga tatarica auf der Iberi-schen Halbinsel, ohne die ältere Angabe zu kommen-tieren. Man kann nur vermuten, dass Altuna nach 24Jahren die ältere Angabe nicht mehr als glaubwürdigbetrachtet.
Die Daten über die Fauna der Cueva de La Rieravon Jesús Altuna 1986 und von Lawrence Guy Straus1992, der Jesús Altuna 1986 zitiert, stimmen nichtüberein. Die MIZ an Bovinae ist in der Schicht 7 nachAltuna (1986, 434, Table B.23) 1, während in derArbeit von Lawrence Guy Straus (1992, 267,Appendix C3) 2 steht. Auch die Zahl der bestimmtenKnochen zeigt gravierende Unterschiede. Während indieser Schicht bei Altuna nur von 677 Knochen vomSteinbock die Rede ist (1986, 434, Tabl. B.23), sind esbei Straus 5677 (1992, 467, Appendix C3)69. SolcheUnstimmigkeiten findet man aber auch weiter in derSchicht 14. Hier zählt Jesús Altuna 2 Bovinae (1986,443, Table B.45), Lawrence Guy Straus nur 1 (1992,
267, Appendix C3). In der Schicht 18 bestimmt JesúsAltuna 8 Steinböcke und 20 Hirsche (1986, 452, Table61), Lawrence Guy Straus zitiert nur 6 Steinböckeund 15 Hirsche (1992, 281, Appendix D6). In derSchicht 19 bestimmt Altuna 7 Steinböcke und 23Hirsche (1986, 452, Table 61), Straus publiziert nur 5Steinböcke und 18 Hirsche (1992, 281, Appendix D6).Da die MIZ bei der jüngeren Publikation (Straus1992) gegenüber der älteren (Altuna 1986) abnimmt,kann man diese nicht mit dem jeweiligen Forschungs-stand verknüpfen. Auch bei der Höhle von Rascaño,Schicht 7, gibt Lawrence Guy Straus (1992, 261,Appendix B6) andere Zahlen zur MIZ an als Altuna(1981, Table 3), den er zitiert. Altuna bestimmtnämlich einen Cervus elaphus, der aber bei LawrenceGuy Straus gar nicht erwähnt wird. Widersprüche gibes manchmal aber auch in ein und derselben Arbeit.So kommt man z. B., wenn man die MIZ dereinzelnen Tierarten in der Schicht 1 der Höhle von LaRiera zählt (Altuna 1986, 426, Tabl. B.7), auf 23einzelne Individuen. Bei der Gesamtzahl sind aber nur22 Individuen aufgezählt. Ähnliches passiert bei denProzentzahlen der Fauna aus der Schicht C der Höhlevon San Teodoro, Italien (Vigliardi 1968, 137). Hierkommt man beim addieren der Prozentzahlen auf eineSumme von 102,6%. Obwohl der Fehler offen-sichtlich ist, kann dieser aufgrund des Fehlens dereffektiven Zahl der bestimmten Knochenreste bei derPublikation nicht aufgehoben werden. Bei der zusam-menfassenden Publikation über das gesamte Paläoli-thikum von Alberto Broglio und Janusz Kozłowski(1987) erreicht man bei den Faunenlisten aus demöstlichen Mitteleuropa und Osteuropa fehlerhafteGesamtzahlen zwischen 84,6% und 114,4%. Nur ineiner der dort angegebenen 19 Fundstellen bzw.Schichten erreicht man genau 100% (Broglio undKozłowski 1987, Tab. XXVII, S. 334-335). Obwohldies eine der ganz wenigen zusammenfassendenTabellen der Fauna aus Osteuropa ist, kann mansolche Daten leider nur als Tendenz betrachten.Schließlich sind hier die Daten aus der Arbeit vonFrançoise Delpech (Delpech 1983; Delpech et al.2000) als ein besonderer Fall anzusprechen. Hier istdas Fortschreiten der Forschung sehr gut dokumen-tiert. Daten über bestimmte Faunenkomplexe auseinigen Höhlen aus der Publikation von 1983 werdenin der neuen Publikation von 2000 „korrigiert“. So ist
69 Da in beiden Publikationen eine MIZ von 14 angegeben wird und da die Zahl der Knochen bei Menschen undTieren um 200 liegt, kann man davon ausgehen, dass die Zahl von 677 bestimmten Knochenfragmenten (imDurchschnitt 48 pro Mindestindividuum) richtig ist, während die von 5677 bestimmten Knochenfragmenten (imDurchschnitt 405 pro Mindestindividuum) unwahrscheinlich erscheint.

214 Verbreitung der großen Jagdfauna in Mittel und Westeuropa im oberen Jungpleistozän
es für Le Flageolet I, La Ferrassie und Roc de Combeder Fall, da diese „n’étaient pas fiables dans leurtotalité“ (Delpech, et al. 2000, 119). Natürlich stelltsich nun diesbezüglich die Frage, wie es mit anderenDaten aus anderen Fundstellen aussieht, die nichtgleichermaßen überprüft worden sind. Der Vergleichzeigt jedoch, dass die Unterschiede sich in für dieseArbeit unproblematischen Grenzen bewegen. Miteinzelnen bzw. sehr wenigen Knochen auftretendeTierarten bleiben weiterhin sehr selten. Die amhäufigsten vertretene Tierart bleibt in ihrer Bedeutungebenfalls unangetastet. Die Faunenkombination ändertsich nicht oder nur geringfügig.
Trotz aller Gelassenheit zeigen jedoch dieseBeispiele, dass es möglich und sogar wahrscheinlichist, dass in manchen Fällen solche „Fehler“ nichtentdeckt werden konnten und in den Faunenlistenunbewusst wiedergegeben wurden. In manchen Fällenkann es auch passieren, dass die ältere an Stelle derneueren und nur die sekundäre an Stelle der primärenLiteratur zitiert und behandelt wurde. Schließlich sindmöglicherweise auch in dieser Arbeit bei derÜbertragung der zahlreichen Daten Fehler entstanden.Wie schon im Kapitel „Definition des Themas“, S. 1,angesprochen, geht es hier jedoch eher um überre-gionale Tendenzen als um Zahlen.
Dem Leser wird mit Nachdruck empfohlen, dieprimäre Literatur ebenfalls zu benutzen.
Die Prozentzahlen beziehen sich immer auf diegesamten Ungulaten. Es entstehen daher Unterschiedezu solchen Publikationen, wo auch andere Tierartenwie z. B. Carnivora einbezogen werden. BeiFundstellen oder Schichten mit sehr wenigen Restenwurde auf die Prozentangabe verzichtet.
Bei Tabellen, in denen einzelne „X“ als Markierungangebracht sind, bedeutet ein „X“ nur, dass diePräsenz einer bestimmten Tierart nachgewiesenwurde.
Bei Tabellen, in denen sowohl einzelne als auchdoppelte und dreifache „x“ vorkommen, weist ihreZahl auf die Häufigkeit einer Tierart hin.
x = selten oder auch nur vorhandenxx = gewöhnlich präsentxxx = häufig ? = unsichere Bestimmung
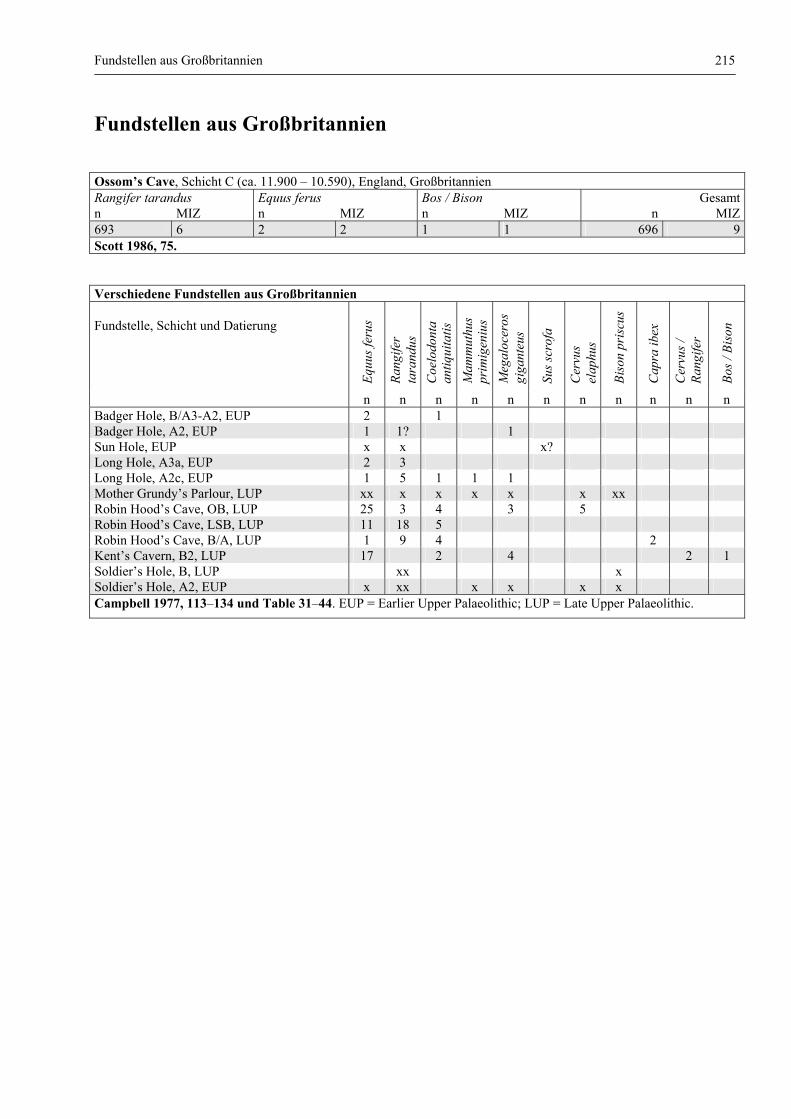
Fundstellen aus Großbritannien 215
Fundstellen aus Großbritannien Ossom’s Cave, Schicht C (ca. 11.900 – 10.590), England, Großbritannien Rangifer tarandus Equus ferus Bos / Bison Gesamtn MIZ n MIZ n MIZ n MIZ693 6 2 2 1 1 696 9Scott 1986, 75. Verschiedene Fundstellen aus Großbritannien Fundstelle, Schicht und Datierung
Equu
s fer
us
Rang
ifer
tara
ndus
Co
elod
onta
an
tiqui
tatis
M
amm
uthu
s pr
imig
eniu
s M
egal
ocer
os
giga
nteu
s
Sus s
crof
a
Cer
vus
elap
hus
Biso
n pr
iscus
Capr
a ib
ex
Cer
vus /
Ra
ngife
r
Bos /
Bis
on
n n n n n n n n n n n Badger Hole, B/A3-A2, EUP 2 1 Badger Hole, A2, EUP 1 1? 1 Sun Hole, EUP x x x? Long Hole, A3a, EUP 2 3 Long Hole, A2c, EUP 1 5 1 1 1 Mother Grundy’s Parlour, LUP xx x x x x x xx Robin Hood’s Cave, OB, LUP 25 3 4 3 5 Robin Hood’s Cave, LSB, LUP 11 18 5 Robin Hood’s Cave, B/A, LUP 1 9 4 2 Kent’s Cavern, B2, LUP 17 2 4 2 1 Soldier’s Hole, B, LUP xx x Soldier’s Hole, A2, EUP x xx x x x x Campbell 1977, 113–134 und Table 31–44. EUP = Earlier Upper Palaeolithic; LUP = Late Upper Palaeolithic.
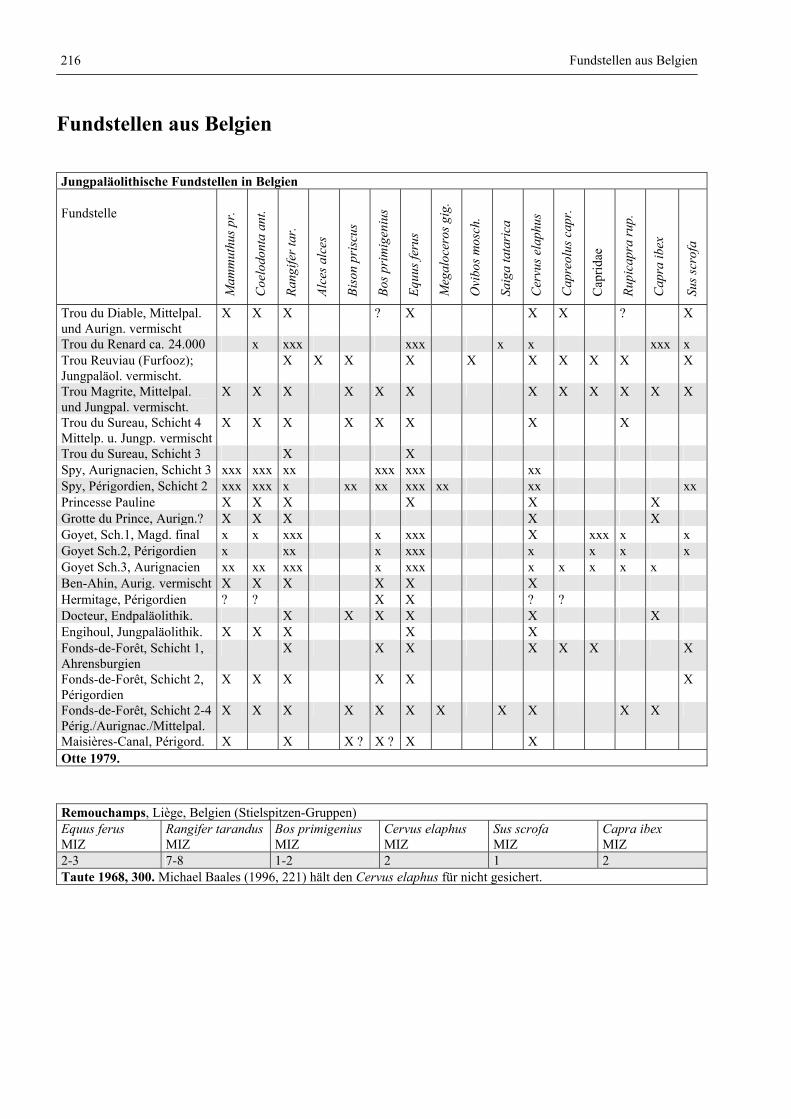
Fundstellen aus Belgien 216
Fundstellen aus Belgien Jungpaläolithische Fundstellen in Belgien Fundstelle
Mam
mut
hus p
r.
Coel
odon
ta a
nt.
Rang
ifer t
ar.
Alce
s alc
es
Biso
n pr
iscus
Bos p
rim
igen
ius
Equu
s fer
us
Meg
aloc
eros
gig
.
Ovi
bos m
osch
.
Saig
a ta
tari
ca
Cerv
us e
laph
us
Capr
eolu
s cap
r.
Cap
ridae
Rupi
capr
a ru
p.
Capr
a ib
ex
Sus s
crof
a
Trou du Diable, Mittelpal. und Aurign. vermischt
X X X ? X X X ? X
Trou du Renard ca. 24.000 x xxx xxx x x xxx x Trou Reuviau (Furfooz); Jungpaläol. vermischt.
X X X X X X X X X X
Trou Magrite, Mittelpal. und Jungpal. vermischt.
X X X X X X X X X X X X
Trou du Sureau, Schicht 4 Mittelp. u. Jungp. vermischt
X X X X X X X X
Trou du Sureau, Schicht 3 X X Spy, Aurignacien, Schicht 3 xxx xxx xx xxx xxx xx Spy, Périgordien, Schicht 2 xxx xxx x xx xx xxx xx xx xx Princesse Pauline X X X X X X Grotte du Prince, Aurign.? X X X X X Goyet, Sch.1, Magd. final x x xxx x xxx X xxx x x Goyet Sch.2, Périgordien x xx x xxx x x x x Goyet Sch.3, Aurignacien xx xx xxx x xxx x x x x x Ben-Ahin, Aurig. vermischt X X X X X X Hermitage, Périgordien ? ? X X ? ? Docteur, Endpaläolithik. X X X X X X Engihoul, Jungpaläolithik. X X X X X Fonds-de-Forêt, Schicht 1, Ahrensburgien
X X X X X X X
Fonds-de-Forêt, Schicht 2, Périgordien
X X X X X X
Fonds-de-Forêt, Schicht 2-4 Périg./Aurignac./Mittelpal.
X X X X X X X X X X X
Maisières-Canal, Périgord. X X X ? X ? X X Otte 1979.
Remouchamps, Liège, Belgien (Stielspitzen-Gruppen) Equus ferus Rangifer tarandus Bos primigenius Cervus elaphus Sus scrofa Capra ibex MIZ MIZ MIZ MIZ MIZ MIZ 2-3 7-8 1-2 2 1 2 Taute 1968, 300. Michael Baales (1996, 221) hält den Cervus elaphus für nicht gesichert.
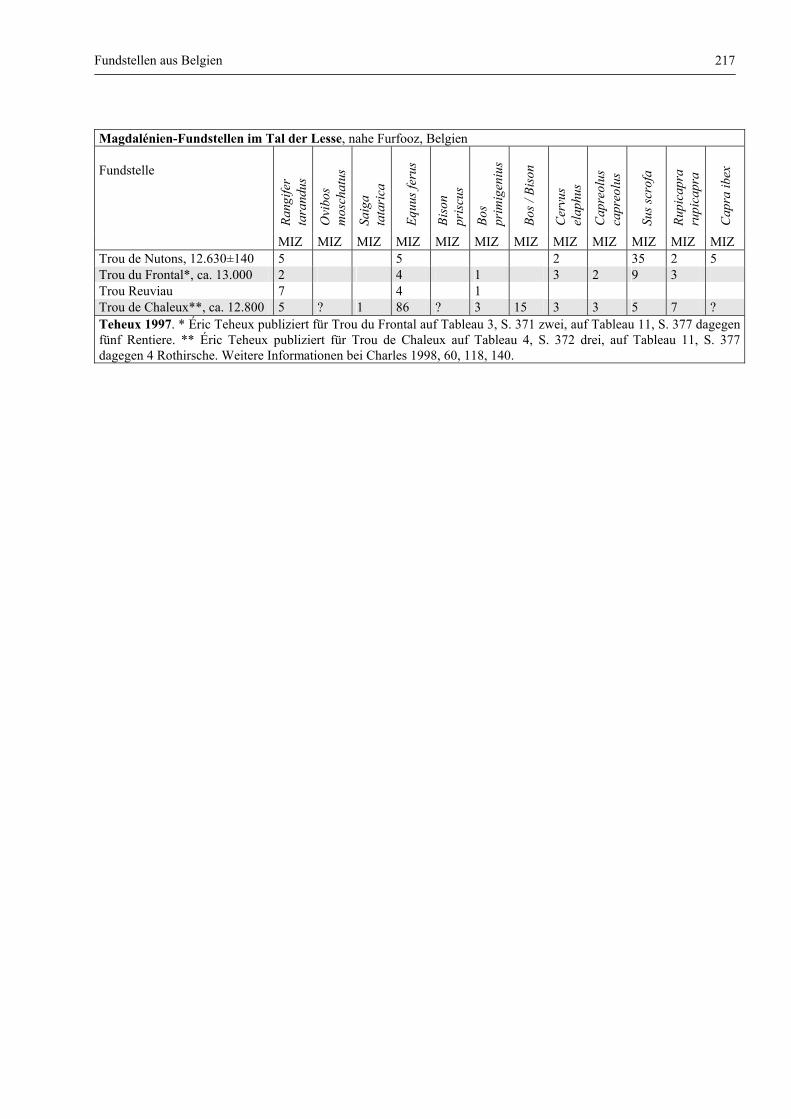
Fundstellen aus Belgien
217
Magdalénien-Fundstellen im Tal der Lesse, nahe Furfooz, Belgien Fundstelle
Rang
ifer
tara
ndus
Ovi
bos
mos
chat
us
Saig
a ta
taric
a
Equu
s fer
us
Biso
n pr
iscus
Bos
prim
igen
ius
Bos /
Bis
on
Cer
vus
elap
hus
Capr
eolu
s ca
preo
lus
Sus s
crof
a
Rupi
capr
a ru
pica
pra
Capr
a ib
ex
MIZ MIZ MIZ MIZ MIZ MIZ MIZ MIZ MIZ MIZ MIZ MIZ Trou de Nutons, 12.630±140 5 5 2 35 2 5 Trou du Frontal*, ca. 13.000 2 4 1 3 2 9 3 Trou Reuviau 7 4 1 Trou de Chaleux**, ca. 12.800 5 ? 1 86 ? 3 15 3 3 5 7 ? Teheux 1997. * Éric Teheux publiziert für Trou du Frontal auf Tableau 3, S. 371 zwei, auf Tableau 11, S. 377 dagegen fünf Rentiere. ** Éric Teheux publiziert für Trou de Chaleux auf Tableau 4, S. 372 drei, auf Tableau 11, S. 377 dagegen 4 Rothirsche. Weitere Informationen bei Charles 1998, 60, 118, 140.
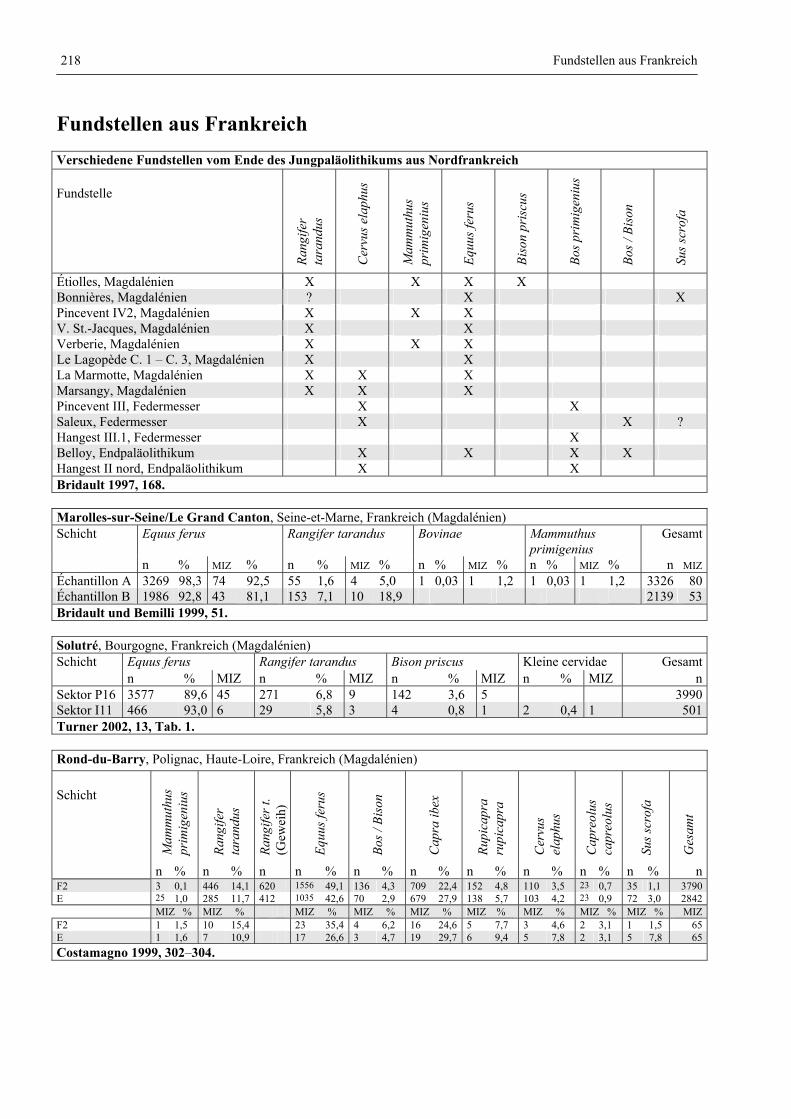
Fundstellen aus Frankreich 218
Fundstellen aus Frankreich Verschiedene Fundstellen vom Ende des Jungpaläolithikums aus Nordfrankreich Fundstelle
Rang
ifer
tara
ndus
Cerv
us e
laph
us
Mam
mut
hus
prim
igen
ius
Equu
s fer
us
Biso
n pr
iscus
Bos p
rim
igen
ius
Bos /
Bis
on
Sus s
crof
a
Étiolles, Magdalénien X X X X Bonnières, Magdalénien ? X X Pincevent IV2, Magdalénien X X X V. St.-Jacques, Magdalénien X X Verberie, Magdalénien X X X Le Lagopède C. 1 – C. 3, Magdalénien X X La Marmotte, Magdalénien X X X Marsangy, Magdalénien X X X Pincevent III, Federmesser X X Saleux, Federmesser X X ? Hangest III.1, Federmesser X Belloy, Endpaläolithikum X X X X Hangest II nord, Endpaläolithikum X X Bridault 1997, 168. Marolles-sur-Seine/Le Grand Canton, Seine-et-Marne, Frankreich (Magdalénien) Schicht Equus ferus Rangifer tarandus Bovinae Mammuthus
primigenius Gesamt
n % MIZ % n % MIZ % n % MIZ % n % MIZ % n MIZÉchantillon A 3269 98,3 74 92,5 55 1,6 4 5,0 1 0,03 1 1,2 1 0,03 1 1,2 3326 80Échantillon B 1986 92,8 43 81,1 153 7,1 10 18,9 2139 53Bridault und Bemilli 1999, 51. Solutré, Bourgogne, Frankreich (Magdalénien) Schicht Equus ferus Rangifer tarandus Bison priscus Kleine cervidae Gesamt
n % MIZ n % MIZ n % MIZ n % MIZ nSektor P16 3577 89,6 45 271 6,8 9 142 3,6 5 3990Sektor I11 466 93,0 6 29 5,8 3 4 0,8 1 2 0,4 1 501Turner 2002, 13, Tab. 1. Rond-du-Barry, Polignac, Haute-Loire, Frankreich (Magdalénien) Schicht
Mam
mut
hus
prim
igen
ius
Rang
ifer
tara
ndus
Rang
ifer t
. (G
ewei
h)
Equu
s fer
us
Bos /
Bis
on
Capr
a ib
ex
Rupi
capr
a ru
pica
pra
Cer
vus
elap
hus
Capr
eolu
s ca
preo
lus
Sus s
crof
a
Ges
amt
n % n % n n % n % n % n % n % n % n % nF2 3 0,1 446 14,1 620 1556 49,1 136 4,3 709 22,4 152 4,8 110 3,5 23 0,7 35 1,1 3790E 25 1,0 285 11,7 412 1035 42,6 70 2,9 679 27,9 138 5,7 103 4,2 23 0,9 72 3,0 2842 MIZ % MIZ % MIZ % MIZ % MIZ % MIZ % MIZ % MIZ % MIZ % MIZF2 1 1,5 10 15,4 23 35,4 4 6,2 16 24,6 5 7,7 3 4,6 2 3,1 1 1,5 65E 1 1,6 7 10,9 17 26,6 3 4,7 19 29,7 6 9,4 5 7,8 2 3,1 5 7,8 65Costamagno 1999, 302–304.
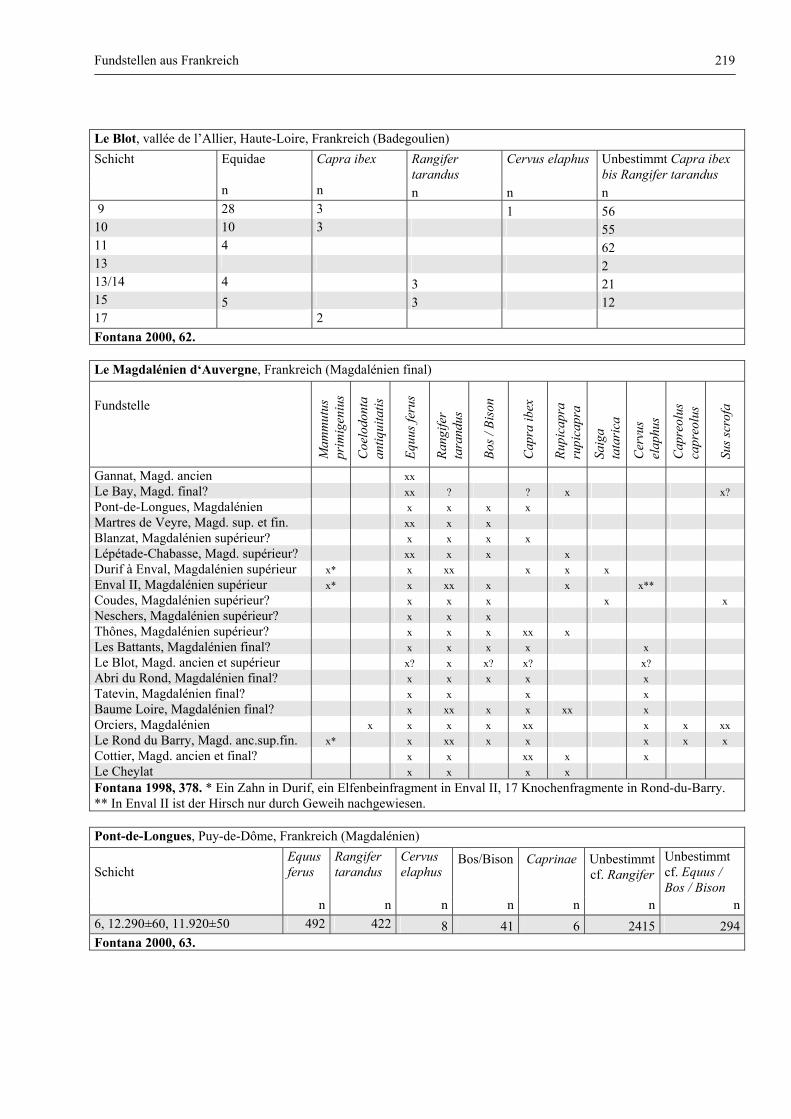
Fundstellen aus Frankreich 219
Le Blot, vallée de l’Allier, Haute-Loire, Frankreich (Badegoulien) Schicht Equidae Capra ibex Rangifer
tarandus Cervus elaphus Unbestimmt Capra ibex
bis Rangifer tarandus n n n n n 9 28 3 1 56 10 10 3 55 11 4 62 13 2 13/14 4 3 21 15 5 3 12 17 2 Fontana 2000, 62. Le Magdalénien d‘Auvergne, Frankreich (Magdalénien final) Fundstelle
Mam
mut
us
prim
igen
ius
Coel
odon
ta
antiq
uita
tis
Equu
s fer
us
Rang
ifer
tara
ndus
Bos /
Bis
on
Capr
a ib
ex
Rupi
capr
a ru
pica
pra
Saig
a ta
taric
a
Cer
vus
elap
hus
Capr
eolu
s ca
preo
lus
Sus s
crof
a
Gannat, Magd. ancien xx Le Bay, Magd. final? xx ? ? x x? Pont-de-Longues, Magdalénien x x x x Martres de Veyre, Magd. sup. et fin. xx x x Blanzat, Magdalénien supérieur? x x x x Lépétade-Chabasse, Magd. supérieur? xx x x x Durif à Enval, Magdalénien supérieur x* x xx x x x Enval II, Magdalénien supérieur x* x xx x x x** Coudes, Magdalénien supérieur? x x x x x Neschers, Magdalénien supérieur? x x x Thônes, Magdalénien supérieur? x x x xx x Les Battants, Magdalénien final? x x x x x Le Blot, Magd. ancien et supérieur x? x x? x? x? Abri du Rond, Magdalénien final? x x x x x Tatevin, Magdalénien final? x x x x Baume Loire, Magdalénien final? x xx x x xx x Orciers, Magdalénien x x x x xx x x xx Le Rond du Barry, Magd. anc.sup.fin. x* x xx x x x x x Cottier, Magd. ancien et final? x x xx x x Le Cheylat x x x x Fontana 1998, 378. * Ein Zahn in Durif, ein Elfenbeinfragment in Enval II, 17 Knochenfragmente in Rond-du-Barry. ** In Enval II ist der Hirsch nur durch Geweih nachgewiesen. Pont-de-Longues, Puy-de-Dôme, Frankreich (Magdalénien) Schicht
Equus ferus
Rangifer tarandus
Cervus elaphus
Bos/Bison Caprinae Unbestimmt cf. Rangifer
Unbestimmt cf. Equus / Bos / Bison
n n n n n n n6, 12.290±60, 11.920±50 492 422 8 41 6 2415 294Fontana 2000, 63.
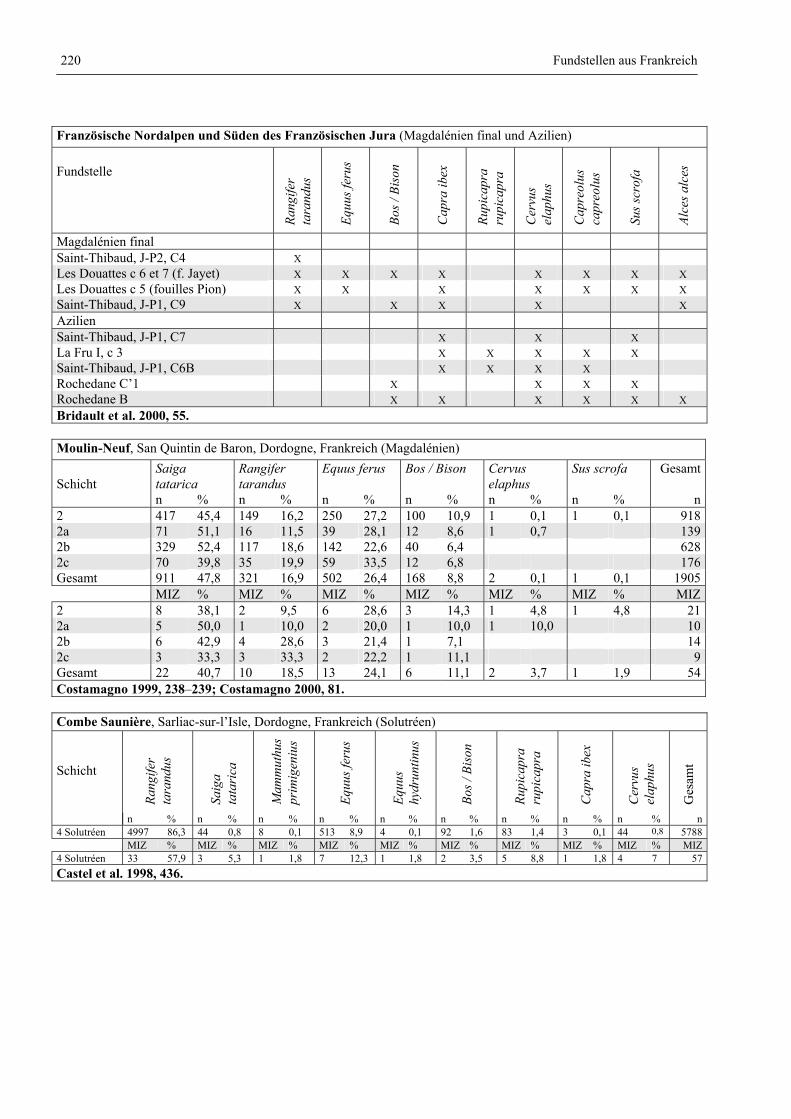
Fundstellen aus Frankreich 220
Französische Nordalpen und Süden des Französischen Jura (Magdalénien final und Azilien) Fundstelle
Rang
ifer
tara
ndus
Equu
s fer
us
Bos /
Bis
on
Capr
a ib
ex
Rupi
capr
a ru
pica
pra
Cer
vus
elap
hus
Capr
eolu
s ca
preo
lus
Sus s
crof
a
Alce
s alc
es
Magdalénien final Saint-Thibaud, J-P2, C4 X Les Douattes c 6 et 7 (f. Jayet) X X X X X X X X Les Douattes c 5 (fouilles Pion) X X X X X X X Saint-Thibaud, J-P1, C9 X X X X X Azilien Saint-Thibaud, J-P1, C7 X X X La Fru I, c 3 X X X X X Saint-Thibaud, J-P1, C6B X X X X Rochedane C’1 X X X X Rochedane B X X X X X X Bridault et al. 2000, 55. Moulin-Neuf, San Quintin de Baron, Dordogne, Frankreich (Magdalénien) Schicht
Saiga tatarica
Rangifer tarandus
Equus ferus Bos / Bison Cervus elaphus
Sus scrofa Gesamt
n % n % n % n % n % n % n2 417 45,4 149 16,2 250 27,2 100 10,9 1 0,1 1 0,1 9182a 71 51,1 16 11,5 39 28,1 12 8,6 1 0,7 1392b 329 52,4 117 18,6 142 22,6 40 6,4 6282c 70 39,8 35 19,9 59 33,5 12 6,8 176Gesamt 911 47,8 321 16,9 502 26,4 168 8,8 2 0,1 1 0,1 1905 MIZ % MIZ % MIZ % MIZ % MIZ % MIZ % MIZ2 8 38,1 2 9,5 6 28,6 3 14,3 1 4,8 1 4,8 212a 5 50,0 1 10,0 2 20,0 1 10,0 1 10,0 102b 6 42,9 4 28,6 3 21,4 1 7,1 142c 3 33,3 3 33,3 2 22,2 1 11,1 9Gesamt 22 40,7 10 18,5 13 24,1 6 11,1 2 3,7 1 1,9 54Costamagno 1999, 238–239; Costamagno 2000, 81. Combe Saunière, Sarliac-sur-l’Isle, Dordogne, Frankreich (Solutréen) Schicht
Rang
ifer
tara
ndus
Saig
a ta
taric
a
Mam
mut
hus
prim
igen
ius
Equu
s fer
us
Equu
s hy
drun
tinus
Bos /
Bis
on
Rupi
capr
a ru
pica
pra
Capr
a ib
ex
Cer
vus
elap
hus
Ges
amt
n % n % n % n % n % n % n % n % n % n4 Solutréen 4997 86,3 44 0,8 8 0,1 513 8,9 4 0,1 92 1,6 83 1,4 3 0,1 44 0,8 5788 MIZ % MIZ % MIZ % MIZ % MIZ % MIZ % MIZ % MIZ % MIZ % MIZ4 Solutréen 33 57,9 3 5,3 1 1,8 7 12,3 1 1,8 2 3,5 5 8,8 1 1,8 4 7 57Castel et al. 1998, 436.
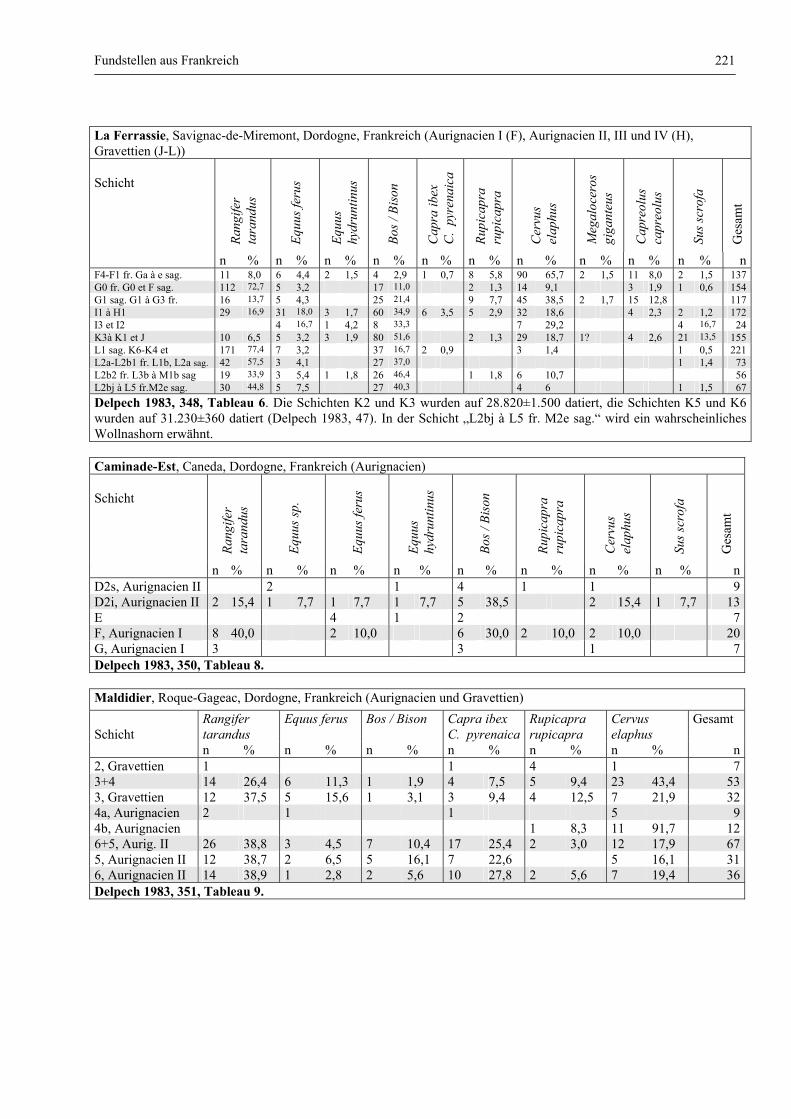
Fundstellen aus Frankreich 221
La Ferrassie, Savignac-de-Miremont, Dordogne, Frankreich (Aurignacien I (F), Aurignacien II, III und IV (H), Gravettien (J-L)) Schicht
Rang
ifer
tara
ndus
Equu
s fer
us
Equu
s hy
drun
tinus
Bos /
Bis
on
Capr
a ib
ex
C. p
yren
aica
Rupi
capr
a ru
pica
pra
Cer
vus
elap
hus
Meg
aloc
eros
gi
gant
eus
Capr
eolu
s ca
preo
lus
Sus s
crof
a
Ges
amt
n % n % n % n % n % n % n % n % n % n % nF4-F1 fr. Ga à e sag. 11 8,0 6 4,4 2 1,5 4 2,9 1 0,7 8 5,8 90 65,7 2 1,5 11 8,0 2 1,5 137G0 fr. G0 et F sag. 112 72,7 5 3,2 17 11,0 2 1,3 14 9,1 3 1,9 1 0,6 154G1 sag. G1 à G3 fr. 16 13,7 5 4,3 25 21,4 9 7,7 45 38,5 2 1,7 15 12,8 117I1 à H1 29 16,9 31 18,0 3 1,7 60 34,9 6 3,5 5 2,9 32 18,6 4 2,3 2 1,2 172I3 et I2 4 16,7 1 4,2 8 33,3 7 29,2 4 16,7 24K3à K1 et J 10 6,5 5 3,2 3 1,9 80 51,6 2 1,3 29 18,7 1? 4 2,6 21 13,5 155L1 sag. K6-K4 et 171 77,4 7 3,2 37 16,7 2 0,9 3 1,4 1 0,5 221L2a-L2b1 fr. L1b, L2a sag. 42 57,5 3 4,1 27 37,0 1 1,4 73L2b2 fr. L3b à M1b sag 19 33,9 3 5,4 1 1,8 26 46,4 1 1,8 6 10,7 56L2bj à L5 fr.M2e sag. 30 44,8 5 7,5 27 40,3 4 6 1 1,5 67Delpech 1983, 348, Tableau 6. Die Schichten K2 und K3 wurden auf 28.820±1.500 datiert, die Schichten K5 und K6 wurden auf 31.230±360 datiert (Delpech 1983, 47). In der Schicht „L2bj à L5 fr. M2e sag.“ wird ein wahrscheinliches Wollnashorn erwähnt. Caminade-Est, Caneda, Dordogne, Frankreich (Aurignacien) Schicht
Rang
ifer
tara
ndus
Equu
s sp.
Equu
s fer
us
Equu
s hy
drun
tinus
Bos /
Bis
on
Rupi
capr
a ru
pica
pra
Cer
vus
elap
hus
Sus s
crof
a
Ges
amt
n % n % n % n % n % n % n % n % nD2s, Aurignacien II 2 1 4 1 1 9D2i, Aurignacien II 2 15,4 1 7,7 1 7,7 1 7,7 5 38,5 2 15,4 1 7,7 13E 4 1 2 7F, Aurignacien I 8 40,0 2 10,0 6 30,0 2 10,0 2 10,0 20G, Aurignacien I 3 3 1 7Delpech 1983, 350, Tableau 8. Maldidier, Roque-Gageac, Dordogne, Frankreich (Aurignacien und Gravettien) Schicht
Rangifer tarandus
Equus ferus Bos / Bison Capra ibex C. pyrenaica
Rupicapra rupicapra
Cervus elaphus
Gesamt
n % n % n % n % n % n % n2, Gravettien 1 1 4 1 73+4 14 26,4 6 11,3 1 1,9 4 7,5 5 9,4 23 43,4 533, Gravettien 12 37,5 5 15,6 1 3,1 3 9,4 4 12,5 7 21,9 324a, Aurignacien 2 1 1 5 94b, Aurignacien 1 8,3 11 91,7 126+5, Aurig. II 26 38,8 3 4,5 7 10,4 17 25,4 2 3,0 12 17,9 675, Aurignacien II 12 38,7 2 6,5 5 16,1 7 22,6 5 16,1 316, Aurignacien II 14 38,9 1 2,8 2 5,6 10 27,8 2 5,6 7 19,4 36Delpech 1983, 351, Tableau 9.
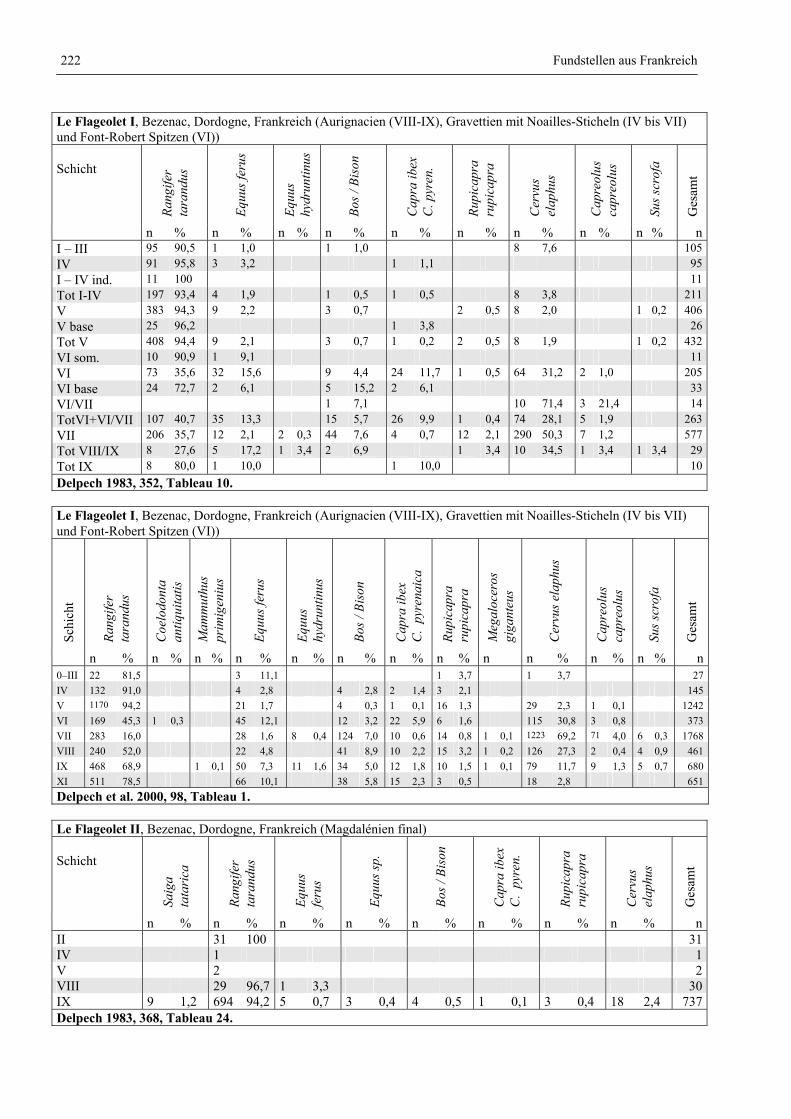
Fundstellen aus Frankreich 222
Le Flageolet I, Bezenac, Dordogne, Frankreich (Aurignacien (VIII-IX), Gravettien mit Noailles-Sticheln (IV bis VII) und Font-Robert Spitzen (VI)) Schicht
Rang
ifer
tara
ndus
Equu
s fer
us
Equu
s hy
drun
tinus
Bos /
Bis
on
Capr
a ib
ex
C. p
yren
.
Rupi
capr
a ru
pica
pra
Cer
vus
elap
hus
Capr
eolu
s ca
preo
lus
Sus s
crof
a
Ges
amt
n % n % n % n % n % n % n % n % n % nI – III 95 90,5 1 1,0 1 1,0 8 7,6 105IV 91 95,8 3 3,2 1 1,1 95I – IV ind. 11 100 11Tot I-IV 197 93,4 4 1,9 1 0,5 1 0,5 8 3,8 211V 383 94,3 9 2,2 3 0,7 2 0,5 8 2,0 1 0,2 406V base 25 96,2 1 3,8 26Tot V 408 94,4 9 2,1 3 0,7 1 0,2 2 0,5 8 1,9 1 0,2 432VI som. 10 90,9 1 9,1 11VI 73 35,6 32 15,6 9 4,4 24 11,7 1 0,5 64 31,2 2 1,0 205VI base 24 72,7 2 6,1 5 15,2 2 6,1 33VI/VII 1 7,1 10 71,4 3 21,4 14TotVI+VI/VII 107 40,7 35 13,3 15 5,7 26 9,9 1 0,4 74 28,1 5 1,9 263VII 206 35,7 12 2,1 2 0,3 44 7,6 4 0,7 12 2,1 290 50,3 7 1,2 577Tot VIII/IX 8 27,6 5 17,2 1 3,4 2 6,9 1 3,4 10 34,5 1 3,4 1 3,4 29Tot IX 8 80,0 1 10,0 1 10,0 10Delpech 1983, 352, Tableau 10. Le Flageolet I, Bezenac, Dordogne, Frankreich (Aurignacien (VIII-IX), Gravettien mit Noailles-Sticheln (IV bis VII) und Font-Robert Spitzen (VI))
Schi
cht
Rang
ifer
tara
ndus
Coel
odon
ta
antiq
uita
tis
Mam
mut
hus
prim
igen
ius
Equu
s fer
us
Equu
s hy
drun
tinus
Bos /
Bis
on
Capr
a ib
ex
C. p
yren
aica
Rupi
capr
a ru
pica
pra
Meg
aloc
eros
gi
gant
eus
Cerv
us e
laph
us
Capr
eolu
s ca
preo
lus
Sus s
crof
a
Ges
amt
n % n % n % n % n % n % n % n % n n % n % n % n0–III 22 81,5 3 11,1 1 3,7 1 3,7 27IV 132 91,0 4 2,8 4 2,8 2 1,4 3 2,1 145V 1170 94,2 21 1,7 4 0,3 1 0,1 16 1,3 29 2,3 1 0,1 1242VI 169 45,3 1 0,3 45 12,1 12 3,2 22 5,9 6 1,6 115 30,8 3 0,8 373VII 283 16,0 28 1,6 8 0,4 124 7,0 10 0,6 14 0,8 1 0,1 1223 69,2 71 4,0 6 0,3 1768VIII 240 52,0 22 4,8 41 8,9 10 2,2 15 3,2 1 0,2 126 27,3 2 0,4 4 0,9 461IX 468 68,9 1 0,1 50 7,3 11 1,6 34 5,0 12 1,8 10 1,5 1 0,1 79 11,7 9 1,3 5 0,7 680XI 511 78,5 66 10,1 38 5,8 15 2,3 3 0,5 18 2,8 651Delpech et al. 2000, 98, Tableau 1. Le Flageolet II, Bezenac, Dordogne, Frankreich (Magdalénien final) Schicht
Saig
a ta
taric
a
Rang
ifer
tara
ndus
Equu
s fe
rus
Equu
s sp.
Bos /
Bis
on
Capr
a ib
ex
C. p
yren
.
Rupi
capr
a ru
pica
pra
Cer
vus
elap
hus
Ges
amt
n % n % n % n % n % n % n % n % nII 31 100 31IV 1 1V 2 2VIII 29 96,7 1 3,3 30IX 9 1,2 694 94,2 5 0,7 3 0,4 4 0,5 1 0,1 3 0,4 18 2,4 737Delpech 1983, 368, Tableau 24.
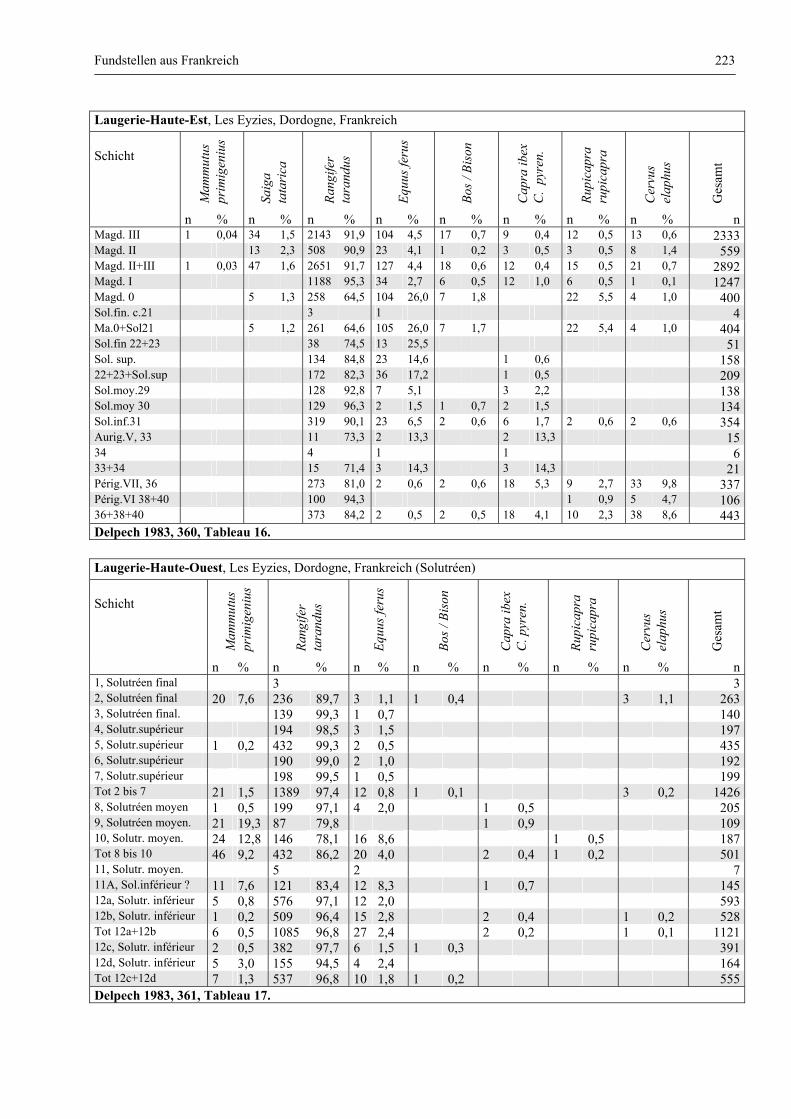
Fundstellen aus Frankreich 223
Laugerie-Haute-Est, Les Eyzies, Dordogne, Frankreich Schicht
Mam
mut
us
prim
igen
ius
Saig
a ta
taric
a
Rang
ifer
tara
ndus
Equu
s fer
us
Bos /
Bis
on
Capr
a ib
ex
C. p
yren
.
Rupi
capr
a ru
pica
pra
Cer
vus
elap
hus
Ges
amt
n % n % n % n % n % n % n % n % nMagd. III 1 0,04 34 1,5 2143 91,9 104 4,5 17 0,7 9 0,4 12 0,5 13 0,6 2333Magd. II 13 2,3 508 90,9 23 4,1 1 0,2 3 0,5 3 0,5 8 1,4 559Magd. II+III 1 0,03 47 1,6 2651 91,7 127 4,4 18 0,6 12 0,4 15 0,5 21 0,7 2892Magd. I 1188 95,3 34 2,7 6 0,5 12 1,0 6 0,5 1 0,1 1247Magd. 0 5 1,3 258 64,5 104 26,0 7 1,8 22 5,5 4 1,0 400Sol.fin. c.21 3 1 4Ma.0+Sol21 5 1,2 261 64,6 105 26,0 7 1,7 22 5,4 4 1,0 404Sol.fin 22+23 38 74,5 13 25,5 51Sol. sup. 134 84,8 23 14,6 1 0,6 15822+23+Sol.sup 172 82,3 36 17,2 1 0,5 209Sol.moy.29 128 92,8 7 5,1 3 2,2 138Sol.moy 30 129 96,3 2 1,5 1 0,7 2 1,5 134Sol.inf.31 319 90,1 23 6,5 2 0,6 6 1,7 2 0,6 2 0,6 354Aurig.V, 33 11 73,3 2 13,3 2 13,3 1534 4 1 1 633+34 15 71,4 3 14,3 3 14,3 21Périg.VII, 36 273 81,0 2 0,6 2 0,6 18 5,3 9 2,7 33 9,8 337Périg.VI 38+40 100 94,3 1 0,9 5 4,7 10636+38+40 373 84,2 2 0,5 2 0,5 18 4,1 10 2,3 38 8,6 443Delpech 1983, 360, Tableau 16. Laugerie-Haute-Ouest, Les Eyzies, Dordogne, Frankreich (Solutréen) Schicht
Mam
mut
us
prim
igen
ius
Rang
ifer
tara
ndus
Equu
s fer
us
Bos /
Bis
on
Capr
a ib
ex
C. p
yren
.
Rupi
capr
a ru
pica
pra
Cer
vus
elap
hus
Ges
amt
n % n % n % n % n % n % n % n1, Solutréen final 3 32, Solutréen final 20 7,6 236 89,7 3 1,1 1 0,4 3 1,1 2633, Solutréen final. 139 99,3 1 0,7 1404, Solutr.supérieur 194 98,5 3 1,5 1975, Solutr.supérieur 1 0,2 432 99,3 2 0,5 4356, Solutr.supérieur 190 99,0 2 1,0 1927, Solutr.supérieur 198 99,5 1 0,5 199Tot 2 bis 7 21 1,5 1389 97,4 12 0,8 1 0,1 3 0,2 14268, Solutréen moyen 1 0,5 199 97,1 4 2,0 1 0,5 2059, Solutréen moyen. 21 19,3 87 79,8 1 0,9 10910, Solutr. moyen. 24 12,8 146 78,1 16 8,6 1 0,5 187Tot 8 bis 10 46 9,2 432 86,2 20 4,0 2 0,4 1 0,2 50111, Solutr. moyen. 5 2 711A, Sol.inférieur ? 11 7,6 121 83,4 12 8,3 1 0,7 14512a, Solutr. inférieur 5 0,8 576 97,1 12 2,0 59312b, Solutr. inférieur 1 0,2 509 96,4 15 2,8 2 0,4 1 0,2 528Tot 12a+12b 6 0,5 1085 96,8 27 2,4 2 0,2 1 0,1 112112c, Solutr. inférieur 2 0,5 382 97,7 6 1,5 1 0,3 39112d, Solutr. inférieur 5 3,0 155 94,5 4 2,4 164Tot 12c+12d 7 1,3 537 96,8 10 1,8 1 0,2 555Delpech 1983, 361, Tableau 17.
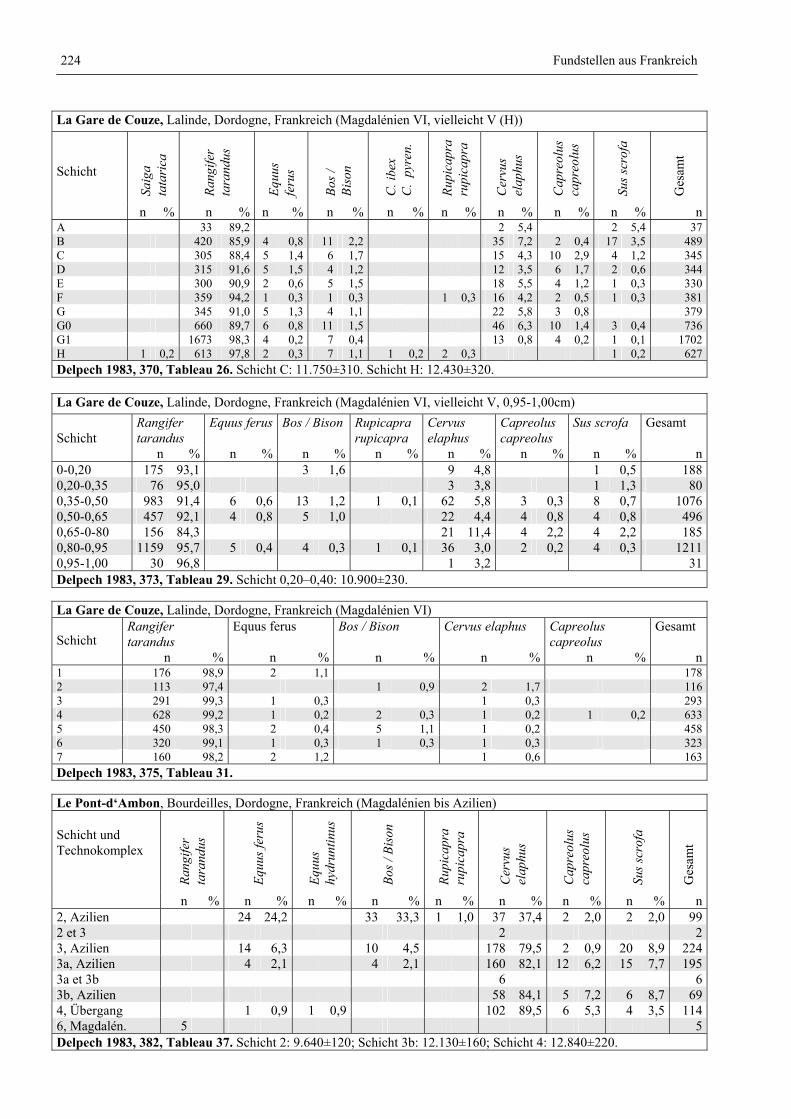
Fundstellen aus Frankreich 224
La Gare de Couze, Lalinde, Dordogne, Frankreich (Magdalénien VI, vielleicht V (H)) Schicht
Saig
a ta
taric
a
Rang
ifer
tara
ndus
Equu
s fe
rus
Bos /
Bi
son
C. i
bex
C. p
yren
.
Rupi
capr
a ru
pica
pra
Cer
vus
elap
hus
Capr
eolu
s ca
preo
lus
Sus s
crof
a
Ges
amt
n % n % n % n % n % n % n % n % n % nA 33 89,2 2 5,4 2 5,4 37B 420 85,9 4 0,8 11 2,2 35 7,2 2 0,4 17 3,5 489C 305 88,4 5 1,4 6 1,7 15 4,3 10 2,9 4 1,2 345D 315 91,6 5 1,5 4 1,2 12 3,5 6 1,7 2 0,6 344E 300 90,9 2 0,6 5 1,5 18 5,5 4 1,2 1 0,3 330F 359 94,2 1 0,3 1 0,3 1 0,3 16 4,2 2 0,5 1 0,3 381G 345 91,0 5 1,3 4 1,1 22 5,8 3 0,8 379G0 660 89,7 6 0,8 11 1,5 46 6,3 10 1,4 3 0,4 736G1 1673 98,3 4 0,2 7 0,4 13 0,8 4 0,2 1 0,1 1702H 1 0,2 613 97,8 2 0,3 7 1,1 1 0,2 2 0,3 1 0,2 627Delpech 1983, 370, Tableau 26. Schicht C: 11.750±310. Schicht H: 12.430±320. La Gare de Couze, Lalinde, Dordogne, Frankreich (Magdalénien VI, vielleicht V, 0,95-1,00cm) Schicht
Rangifer tarandus
Equus ferus Bos / Bison Rupicapra rupicapra
Cervus elaphus
Capreolus capreolus
Sus scrofa Gesamt
n % n % n % n % n % n % n % n0-0,20 175 93,1 3 1,6 9 4,8 1 0,5 1880,20-0,35 76 95,0 3 3,8 1 1,3 800,35-0,50 983 91,4 6 0,6 13 1,2 1 0,1 62 5,8 3 0,3 8 0,7 10760,50-0,65 457 92,1 4 0,8 5 1,0 22 4,4 4 0,8 4 0,8 4960,65-0-80 156 84,3 21 11,4 4 2,2 4 2,2 1850,80-0,95 1159 95,7 5 0,4 4 0,3 1 0,1 36 3,0 2 0,2 4 0,3 12110,95-1,00 30 96,8 1 3,2 31Delpech 1983, 373, Tableau 29. Schicht 0,20–0,40: 10.900±230. La Gare de Couze, Lalinde, Dordogne, Frankreich (Magdalénien VI) Schicht
Rangifer tarandus
Equus ferus Bos / Bison Cervus elaphus Capreolus capreolus
Gesamt
n % n % n % n % n % n1 176 98,9 2 1,1 1782 113 97,4 1 0,9 2 1,7 1163 291 99,3 1 0,3 1 0,3 2934 628 99,2 1 0,2 2 0,3 1 0,2 1 0,2 6335 450 98,3 2 0,4 5 1,1 1 0,2 4586 320 99,1 1 0,3 1 0,3 1 0,3 3237 160 98,2 2 1,2 1 0,6 163Delpech 1983, 375, Tableau 31. Le Pont-d‘Ambon, Bourdeilles, Dordogne, Frankreich (Magdalénien bis Azilien) Schicht und Technokomplex
Rang
ifer
tara
ndus
Equu
s fer
us
Equu
s hy
drun
tinus
Bos /
Bis
on
Rupi
capr
a ru
pica
pra
Cer
vus
elap
hus
Capr
eolu
s ca
preo
lus
Sus s
crof
a
Ges
amt
n % n % n % n % n % n % n % n % n2, Azilien 24 24,2 33 33,3 1 1,0 37 37,4 2 2,0 2 2,0 992 et 3 2 23, Azilien 14 6,3 10 4,5 178 79,5 2 0,9 20 8,9 2243a, Azilien 4 2,1 4 2,1 160 82,1 12 6,2 15 7,7 1953a et 3b 6 63b, Azilien 58 84,1 5 7,2 6 8,7 694, Übergang 1 0,9 1 0,9 102 89,5 6 5,3 4 3,5 1146, Magdalén. 5 5Delpech 1983, 382, Tableau 37. Schicht 2: 9.640±120; Schicht 3b: 12.130±160; Schicht 4: 12.840±220.
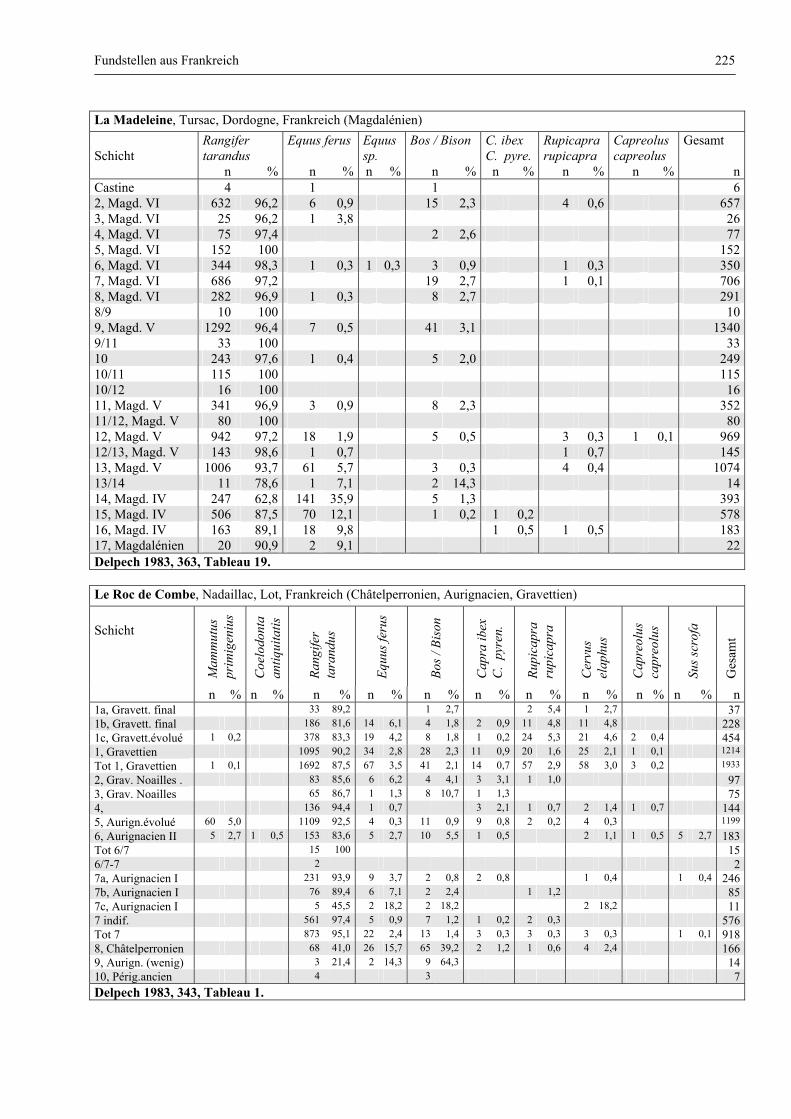
Fundstellen aus Frankreich 225
La Madeleine, Tursac, Dordogne, Frankreich (Magdalénien) Schicht
Rangifer tarandus
Equus ferus Equus sp.
Bos / Bison C. ibex C. pyre.
Rupicapra rupicapra
Capreolus capreolus
Gesamt
n % n % n % n % n % n % n % nCastine 4 1 1 62, Magd. VI 632 96,2 6 0,9 15 2,3 4 0,6 6573, Magd. VI 25 96,2 1 3,8 264, Magd. VI 75 97,4 2 2,6 775, Magd. VI 152 100 1526, Magd. VI 344 98,3 1 0,3 1 0,3 3 0,9 1 0,3 3507, Magd. VI 686 97,2 19 2,7 1 0,1 7068, Magd. VI 282 96,9 1 0,3 8 2,7 2918/9 10 100 109, Magd. V 1292 96,4 7 0,5 41 3,1 13409/11 33 100 3310 243 97,6 1 0,4 5 2,0 24910/11 115 100 11510/12 16 100 1611, Magd. V 341 96,9 3 0,9 8 2,3 35211/12, Magd. V 80 100 8012, Magd. V 942 97,2 18 1,9 5 0,5 3 0,3 1 0,1 96912/13, Magd. V 143 98,6 1 0,7 1 0,7 14513, Magd. V 1006 93,7 61 5,7 3 0,3 4 0,4 107413/14 11 78,6 1 7,1 2 14,3 1414, Magd. IV 247 62,8 141 35,9 5 1,3 39315, Magd. IV 506 87,5 70 12,1 1 0,2 1 0,2 57816, Magd. IV 163 89,1 18 9,8 1 0,5 1 0,5 18317, Magdalénien 20 90,9 2 9,1 22Delpech 1983, 363, Tableau 19. Le Roc de Combe, Nadaillac, Lot, Frankreich (Châtelperronien, Aurignacien, Gravettien) Schicht
Mam
mut
us
prim
igen
ius
Coel
odon
ta
antiq
uita
tis
Rang
ifer
tara
ndus
Equu
s fer
us
Bos /
Bis
on
Capr
a ib
ex
C. p
yren
.
Rupi
capr
a ru
pica
pra
Cer
vus
elap
hus
Capr
eolu
s ca
preo
lus
Sus s
crof
a
Ges
amt
n % n % n % n % n % n % n % n % n % n % n1a, Gravett. final 33 89,2 1 2,7 2 5,4 1 2,7 371b, Gravett. final 186 81,6 14 6,1 4 1,8 2 0,9 11 4,8 11 4,8 2281c, Gravett.évolué 1 0,2 378 83,3 19 4,2 8 1,8 1 0,2 24 5,3 21 4,6 2 0,4 4541, Gravettien 1095 90,2 34 2,8 28 2,3 11 0,9 20 1,6 25 2,1 1 0,1 1214
Tot 1, Gravettien 1 0,1 1692 87,5 67 3,5 41 2,1 14 0,7 57 2,9 58 3,0 3 0,2 1933
2, Grav. Noailles . 83 85,6 6 6,2 4 4,1 3 3,1 1 1,0 973, Grav. Noailles 65 86,7 1 1,3 8 10,7 1 1,3 754, 136 94,4 1 0,7 3 2,1 1 0,7 2 1,4 1 0,7 1445, Aurign.évolué 60 5,0 1109 92,5 4 0,3 11 0,9 9 0,8 2 0,2 4 0,3 1199
6, Aurignacien II 5 2,7 1 0,5 153 83,6 5 2,7 10 5,5 1 0,5 2 1,1 1 0,5 5 2,7 183Tot 6/7 15 100 156/7-7 2 27a, Aurignacien I 231 93,9 9 3,7 2 0,8 2 0,8 1 0,4 1 0,4 2467b, Aurignacien I 76 89,4 6 7,1 2 2,4 1 1,2 857c, Aurignacien I 5 45,5 2 18,2 2 18,2 2 18,2 117 indif. 561 97,4 5 0,9 7 1,2 1 0,2 2 0,3 576Tot 7 873 95,1 22 2,4 13 1,4 3 0,3 3 0,3 3 0,3 1 0,1 9188, Châtelperronien 68 41,0 26 15,7 65 39,2 2 1,2 1 0,6 4 2,4 1669, Aurign. (wenig) 3 21,4 2 14,3 9 64,3 1410, Périg.ancien 4 3 7Delpech 1983, 343, Tableau 1.
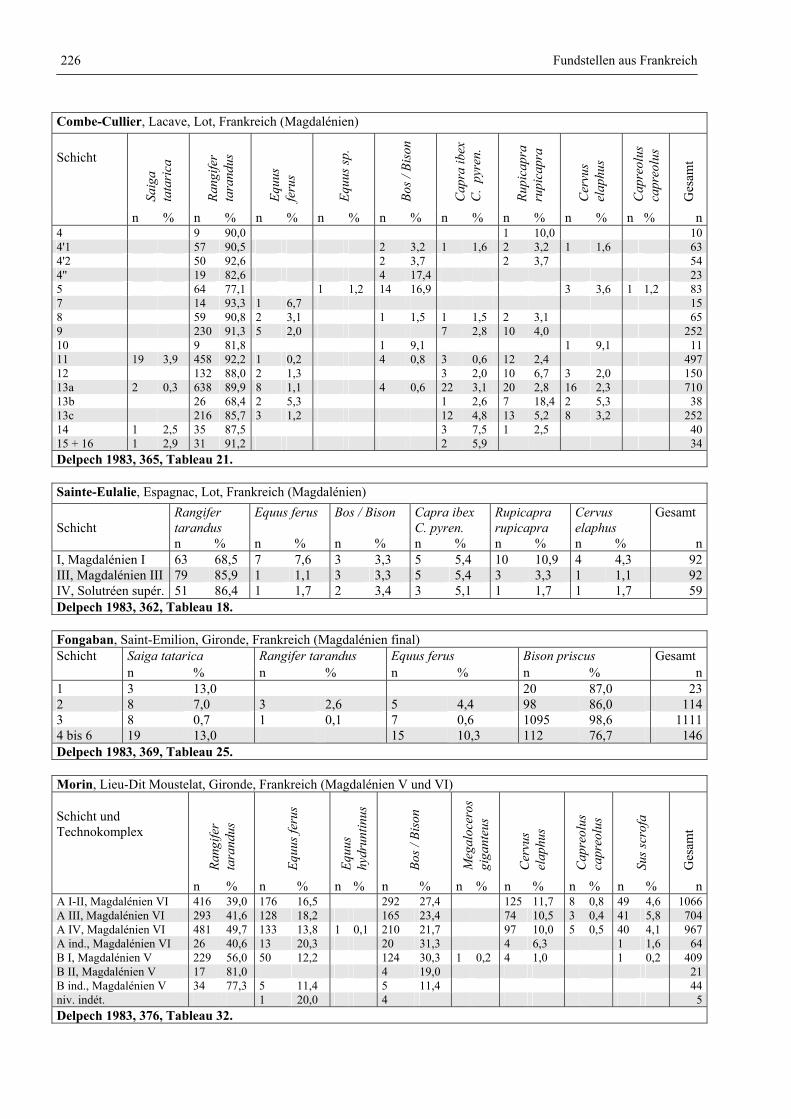
Fundstellen aus Frankreich
226
Combe-Cullier, Lacave, Lot, Frankreich (Magdalénien) Schicht
Saig
a ta
taric
a
Rang
ifer
tara
ndus
Equu
s fe
rus
Equu
s sp.
Bos /
Bis
on
Capr
a ib
ex
C. p
yren
.
Rupi
capr
a ru
pica
pra
Cer
vus
elap
hus
Capr
eolu
s ca
preo
lus
Ges
amt
n % n % n % n % n % n % n % n % n % n4 9 90,0 1 10,0 104'1 57 90,5 2 3,2 1 1,6 2 3,2 1 1,6 634'2 50 92,6 2 3,7 2 3,7 544'' 19 82,6 4 17,4 235 64 77,1 1 1,2 14 16,9 3 3,6 1 1,2 837 14 93,3 1 6,7 158 59 90,8 2 3,1 1 1,5 1 1,5 2 3,1 659 230 91,3 5 2,0 7 2,8 10 4,0 25210 9 81,8 1 9,1 1 9,1 1111 19 3,9 458 92,2 1 0,2 4 0,8 3 0,6 12 2,4 49712 132 88,0 2 1,3 3 2,0 10 6,7 3 2,0 15013a 2 0,3 638 89,9 8 1,1 4 0,6 22 3,1 20 2,8 16 2,3 71013b 26 68,4 2 5,3 1 2,6 7 18,4 2 5,3 3813c 216 85,7 3 1,2 12 4,8 13 5,2 8 3,2 25214 1 2,5 35 87,5 3 7,5 1 2,5 4015 + 16 1 2,9 31 91,2 2 5,9 34Delpech 1983, 365, Tableau 21. Sainte-Eulalie, Espagnac, Lot, Frankreich (Magdalénien) Schicht
Rangifer tarandus
Equus ferus Bos / Bison Capra ibex C. pyren.
Rupicapra rupicapra
Cervus elaphus
Gesamt
n % n % n % n % n % n % nI, Magdalénien I 63 68,5 7 7,6 3 3,3 5 5,4 10 10,9 4 4,3 92III, Magdalénien III 79 85,9 1 1,1 3 3,3 5 5,4 3 3,3 1 1,1 92IV, Solutréen supér. 51 86,4 1 1,7 2 3,4 3 5,1 1 1,7 1 1,7 59Delpech 1983, 362, Tableau 18. Fongaban, Saint-Emilion, Gironde, Frankreich (Magdalénien final) Schicht Saiga tatarica Rangifer tarandus Equus ferus Bison priscus Gesamt
n % n % n % n % n1 3 13,0 20 87,0 232 8 7,0 3 2,6 5 4,4 98 86,0 1143 8 0,7 1 0,1 7 0,6 1095 98,6 11114 bis 6 19 13,0 15 10,3 112 76,7 146Delpech 1983, 369, Tableau 25. Morin, Lieu-Dit Moustelat, Gironde, Frankreich (Magdalénien V und VI) Schicht und Technokomplex
Rang
ifer
tara
ndus
Equu
s fer
us
Equu
s hy
drun
tinus
Bos /
Bis
on
Meg
aloc
eros
gi
gant
eus
Cer
vus
elap
hus
Capr
eolu
s ca
preo
lus
Sus s
crof
a
Ges
amt
n % n % n % n % n % n % n % n % nA I-II, Magdalénien VI 416 39,0 176 16,5 292 27,4 125 11,7 8 0,8 49 4,6 1066A III, Magdalénien VI 293 41,6 128 18,2 165 23,4 74 10,5 3 0,4 41 5,8 704A IV, Magdalénien VI 481 49,7 133 13,8 1 0,1 210 21,7 97 10,0 5 0,5 40 4,1 967A ind., Magdalénien VI 26 40,6 13 20,3 20 31,3 4 6,3 1 1,6 64B I, Magdalénien V 229 56,0 50 12,2 124 30,3 1 0,2 4 1,0 1 0,2 409B II, Magdalénien V 17 81,0 4 19,0 21B ind., Magdalénien V 34 77,3 5 11,4 5 11,4 44niv. indét. 1 20,0 4 5Delpech 1983, 376, Tableau 32.
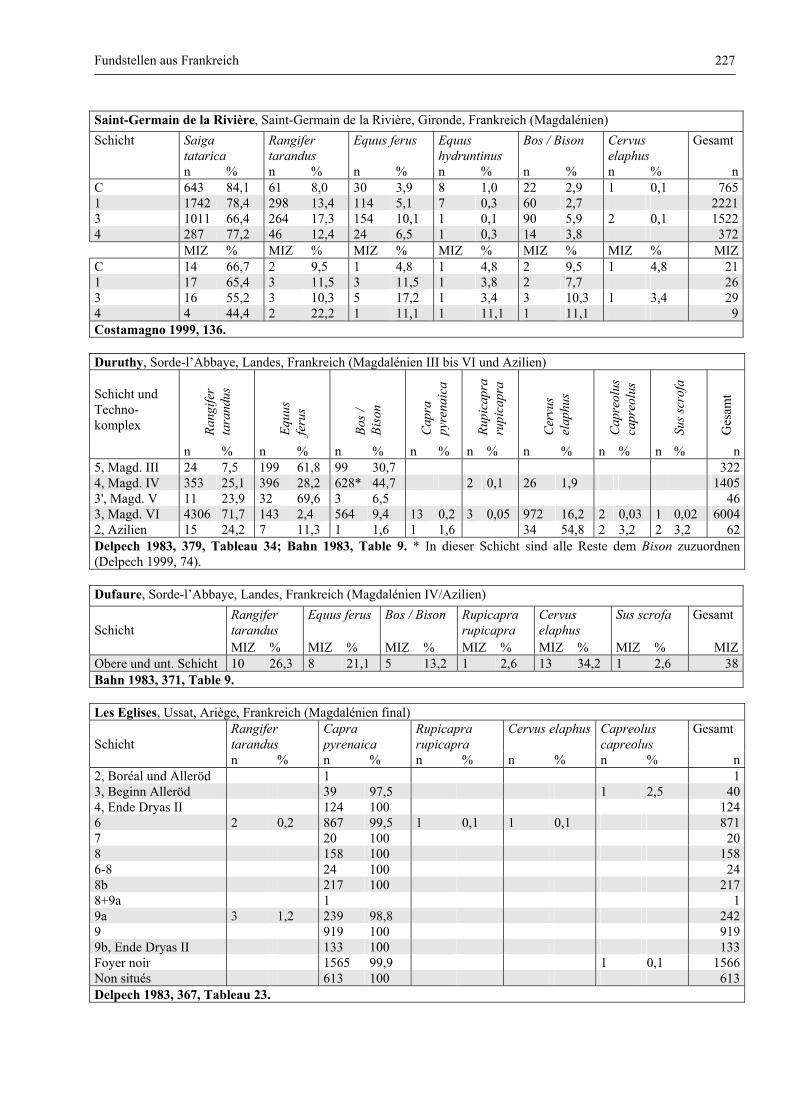
Fundstellen aus Frankreich 227
Saint-Germain de la Rivière, Saint-Germain de la Rivière, Gironde, Frankreich (Magdalénien) Schicht Saiga
tatarica Rangifer tarandus
Equus ferus Equus hydruntinus
Bos / Bison Cervus elaphus
Gesamt
n % n % n % n % n % n % nC 643 84,1 61 8,0 30 3,9 8 1,0 22 2,9 1 0,1 7651 1742 78,4 298 13,4 114 5,1 7 0,3 60 2,7 22213 1011 66,4 264 17,3 154 10,1 1 0,1 90 5,9 2 0,1 15224 287 77,2 46 12,4 24 6,5 1 0,3 14 3,8 372
MIZ % MIZ % MIZ % MIZ % MIZ % MIZ % MIZC 14 66,7 2 9,5 1 4,8 1 4,8 2 9,5 1 4,8 211 17 65,4 3 11,5 3 11,5 1 3,8 2 7,7 263 16 55,2 3 10,3 5 17,2 1 3,4 3 10,3 1 3,4 294 4 44,4 2 22,2 1 11,1 1 11,1 1 11,1 9Costamagno 1999, 136. Duruthy, Sorde-l’Abbaye, Landes, Frankreich (Magdalénien III bis VI und Azilien) Schicht und Techno-komplex Ra
ngife
r ta
rand
us
Equu
s fe
rus
Bos /
Bi
son
Capr
a py
rena
ica
Rupi
capr
a ru
pica
pra
Cer
vus
elap
hus
Capr
eolu
s ca
preo
lus
Sus s
crof
a
Ges
amt
n % n % n % n % n % n % n % n % n5, Magd. III 24 7,5 199 61,8 99 30,7 3224, Magd. IV 353 25,1 396 28,2 628* 44,7 2 0,1 26 1,9 14053', Magd. V 11 23,9 32 69,6 3 6,5 463, Magd. VI 4306 71,7 143 2,4 564 9,4 13 0,2 3 0,05 972 16,2 2 0,03 1 0,02 60042, Azilien 15 24,2 7 11,3 1 1,6 1 1,6 34 54,8 2 3,2 2 3,2 62Delpech 1983, 379, Tableau 34; Bahn 1983, Table 9. * In dieser Schicht sind alle Reste dem Bison zuzuordnen (Delpech 1999, 74). Dufaure, Sorde-l’Abbaye, Landes, Frankreich (Magdalénien IV/Azilien) Schicht
Rangifer tarandus
Equus ferus Bos / Bison Rupicapra rupicapra
Cervus elaphus
Sus scrofa Gesamt
MIZ % MIZ % MIZ % MIZ % MIZ % MIZ % MIZObere und unt. Schicht 10 26,3 8 21,1 5 13,2 1 2,6 13 34,2 1 2,6 38Bahn 1983, 371, Table 9. Les Eglises, Ussat, Ariège, Frankreich (Magdalénien final) Schicht
Rangifer tarandus
Capra pyrenaica
Rupicapra rupicapra
Cervus elaphus Capreolus capreolus
Gesamt
n % n % n % n % n % n2, Boréal und Alleröd 1 13, Beginn Alleröd 39 97,5 1 2,5 404, Ende Dryas II 124 100 1246 2 0,2 867 99,5 1 0,1 1 0,1 8717 20 100 208 158 100 1586-8 24 100 248b 217 100 2178+9a 1 19a 3 1,2 239 98,8 2429 919 100 9199b, Ende Dryas II 133 100 133Foyer noir 1565 99,9 1 0,1 1566Non situés 613 100 613Delpech 1983, 367, Tableau 23.
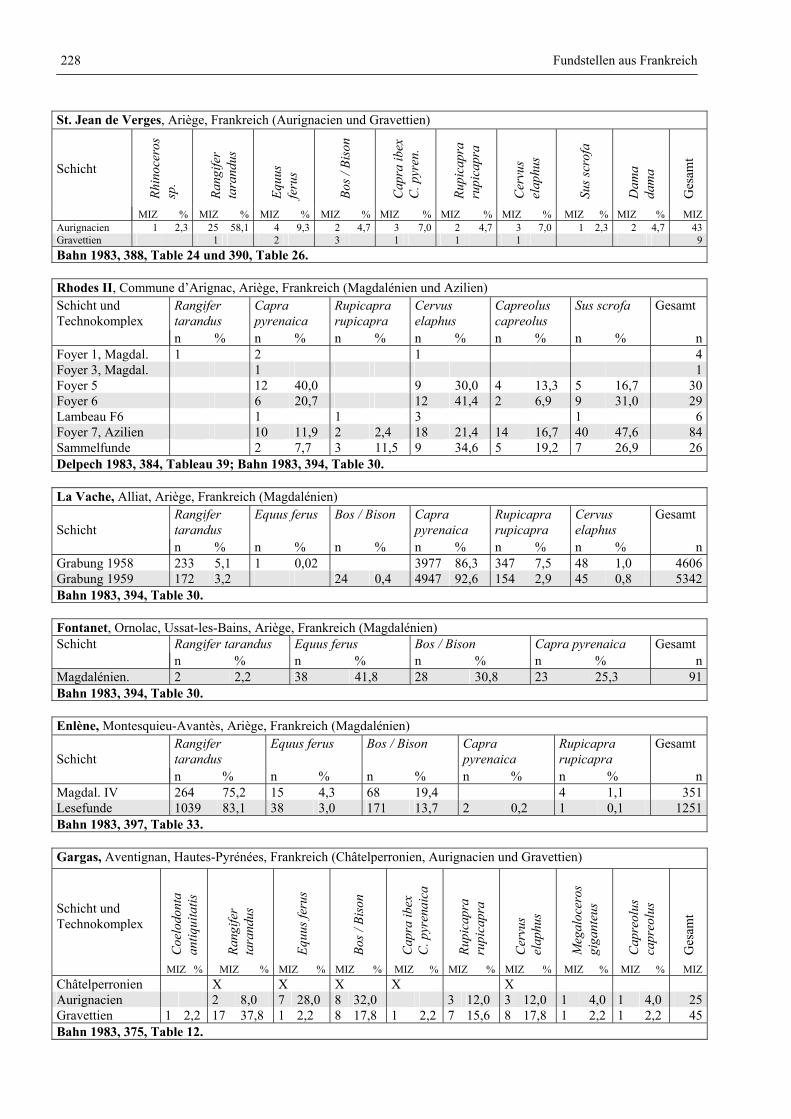
Fundstellen aus Frankreich 228
St. Jean de Verges, Ariège, Frankreich (Aurignacien und Gravettien) Schicht
Rhin
ocer
os
sp.
Rang
ifer
tara
ndus
Equu
s fe
rus
Bos /
Bis
on
Capr
a ib
ex
C. p
yren
.
Rupi
capr
a ru
pica
pra
Cer
vus
elap
hus
Sus s
crof
a
Dam
a da
ma
Ges
amt
MIZ % MIZ % MIZ % MIZ % MIZ % MIZ % MIZ % MIZ % MIZ % MIZAurignacien 1 2,3 25 58,1 4 9,3 2 4,7 3 7,0 2 4,7 3 7,0 1 2,3 2 4,7 43Gravettien 1 2 3 1 1 1 9Bahn 1983, 388, Table 24 und 390, Table 26. Rhodes II, Commune d’Arignac, Ariège, Frankreich (Magdalénien und Azilien) Schicht und Technokomplex
Rangifer tarandus
Capra pyrenaica
Rupicapra rupicapra
Cervus elaphus
Capreolus capreolus
Sus scrofa Gesamt
n % n % n % n % n % n % nFoyer 1, Magdal. 1 2 1 4Foyer 3, Magdal. 1 1Foyer 5 12 40,0 9 30,0 4 13,3 5 16,7 30Foyer 6 6 20,7 12 41,4 2 6,9 9 31,0 29Lambeau F6 1 1 3 1 6Foyer 7, Azilien 10 11,9 2 2,4 18 21,4 14 16,7 40 47,6 84Sammelfunde 2 7,7 3 11,5 9 34,6 5 19,2 7 26,9 26Delpech 1983, 384, Tableau 39; Bahn 1983, 394, Table 30. La Vache, Alliat, Ariège, Frankreich (Magdalénien) Schicht
Rangifer tarandus
Equus ferus Bos / Bison Capra pyrenaica
Rupicapra rupicapra
Cervus elaphus
Gesamt
n % n % n % n % n % n % nGrabung 1958 233 5,1 1 0,02 3977 86,3 347 7,5 48 1,0 4606Grabung 1959 172 3,2 24 0,4 4947 92,6 154 2,9 45 0,8 5342Bahn 1983, 394, Table 30. Fontanet, Ornolac, Ussat-les-Bains, Ariège, Frankreich (Magdalénien) Schicht Rangifer tarandus Equus ferus Bos / Bison Capra pyrenaica Gesamt
n % n % n % n % nMagdalénien. 2 2,2 38 41,8 28 30,8 23 25,3 91Bahn 1983, 394, Table 30. Enlène, Montesquieu-Avantès, Ariège, Frankreich (Magdalénien) Schicht
Rangifer tarandus
Equus ferus Bos / Bison Capra pyrenaica
Rupicapra rupicapra
Gesamt
n % n % n % n % n % nMagdal. IV 264 75,2 15 4,3 68 19,4 4 1,1 351Lesefunde 1039 83,1 38 3,0 171 13,7 2 0,2 1 0,1 1251Bahn 1983, 397, Table 33. Gargas, Aventignan, Hautes-Pyrénées, Frankreich (Châtelperronien, Aurignacien und Gravettien) Schicht und Technokomplex
Coel
odon
ta
antiq
uita
tis
Rang
ifer
tara
ndus
Equu
s fer
us
Bos /
Bis
on
Capr
a ib
ex
C. p
yren
aica
Rupi
capr
a ru
pica
pra
Cer
vus
elap
hus
Meg
aloc
eros
gi
gant
eus
Capr
eolu
s ca
preo
lus
Ges
amt
MIZ % MIZ % MIZ % MIZ % MIZ % MIZ % MIZ % MIZ % MIZ % MIZChâtelperronien X X X X X Aurignacien 2 8,0 7 28,0 8 32,0 3 12,0 3 12,0 1 4,0 1 4,0 25Gravettien 1 2,2 17 37,8 1 2,2 8 17,8 1 2,2 7 15,6 8 17,8 1 2,2 1 2,2 45Bahn 1983, 375, Table 12.
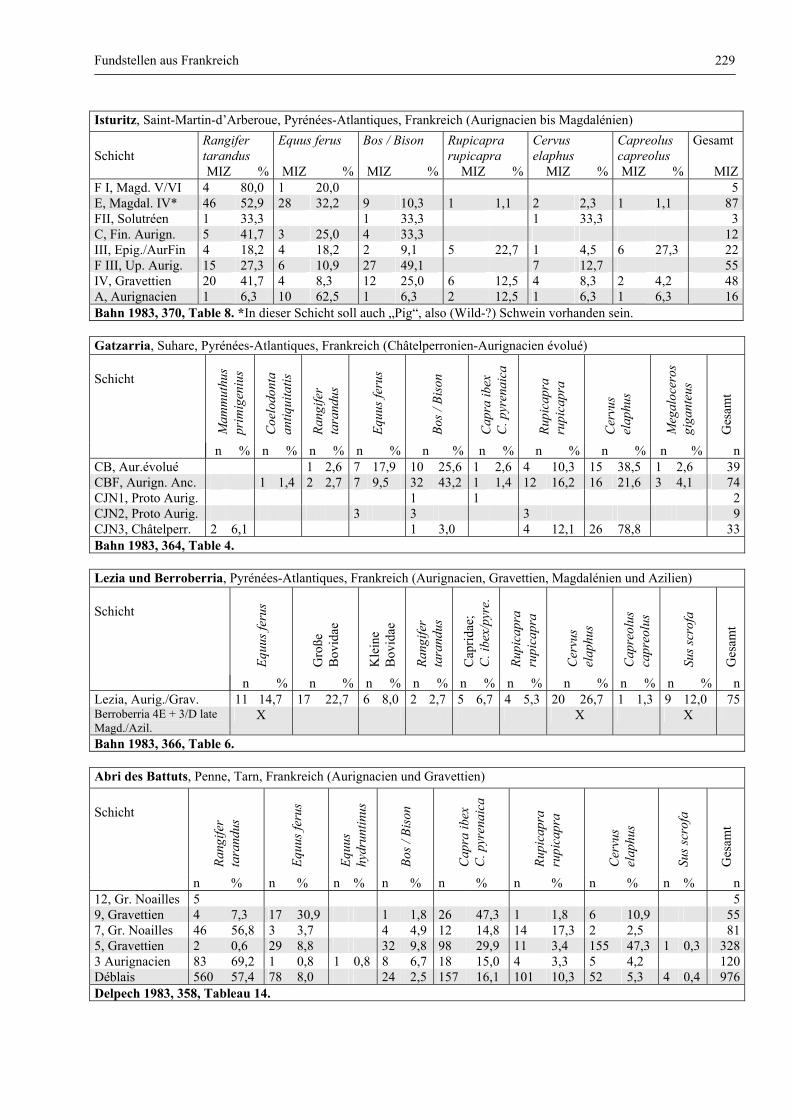
Fundstellen aus Frankreich 229
Isturitz, Saint-Martin-d’Arberoue, Pyrénées-Atlantiques, Frankreich (Aurignacien bis Magdalénien) Schicht
Rangifer tarandus
Equus ferus Bos / Bison Rupicapra rupicapra
Cervus elaphus
Capreolus capreolus
Gesamt
MIZ % MIZ % MIZ % MIZ % MIZ % MIZ % MIZF I, Magd. V/VI 4 80,0 1 20,0 5E, Magdal. IV* 46 52,9 28 32,2 9 10,3 1 1,1 2 2,3 1 1,1 87FII, Solutréen 1 33,3 1 33,3 1 33,3 3C, Fin. Aurign. 5 41,7 3 25,0 4 33,3 12III, Epig./AurFin 4 18,2 4 18,2 2 9,1 5 22,7 1 4,5 6 27,3 22F III, Up. Aurig. 15 27,3 6 10,9 27 49,1 7 12,7 55IV, Gravettien 20 41,7 4 8,3 12 25,0 6 12,5 4 8,3 2 4,2 48A, Aurignacien 1 6,3 10 62,5 1 6,3 2 12,5 1 6,3 1 6,3 16Bahn 1983, 370, Table 8. *In dieser Schicht soll auch „Pig“, also (Wild-?) Schwein vorhanden sein. Gatzarria, Suhare, Pyrénées-Atlantiques, Frankreich (Châtelperronien-Aurignacien évolué) Schicht
Mam
mut
hus
prim
igen
ius
Coel
odon
ta
antiq
uita
tis
Rang
ifer
tara
ndus
Equu
s fer
us
Bos /
Bis
on
Capr
a ib
ex
C. p
yren
aica
Rupi
capr
a ru
pica
pra
Cer
vus
elap
hus
Meg
aloc
eros
gi
gant
eus
Ges
amt
n % n % n % n % n % n % n % n % n % nCB, Aur.évolué 1 2,6 7 17,9 10 25,6 1 2,6 4 10,3 15 38,5 1 2,6 39CBF, Aurign. Anc. 1 1,4 2 2,7 7 9,5 32 43,2 1 1,4 12 16,2 16 21,6 3 4,1 74CJN1, Proto Aurig. 1 1 2CJN2, Proto Aurig. 3 3 3 9CJN3, Châtelperr. 2 6,1 1 3,0 4 12,1 26 78,8 33Bahn 1983, 364, Table 4. Lezia und Berroberria, Pyrénées-Atlantiques, Frankreich (Aurignacien, Gravettien, Magdalénien und Azilien) Schicht
Equu
s fer
us
Gro
ße
Bov
idae
Kle
ine
Bov
idae
Rang
ifer
tara
ndus
Cap
ridae
;
C. i
bex/
pyre
.
Rupi
capr
a ru
pica
pra
Cer
vus
elap
hus
Capr
eolu
s ca
preo
lus
Sus s
crof
a
Ges
amt
n % n % n % n % n % n % n % n % n % nLezia, Aurig./Grav. 11 14,7 17 22,7 6 8,0 2 2,7 5 6,7 4 5,3 20 26,7 1 1,3 9 12,0 75Berroberria 4E + 3/D late Magd./Azil.
X X X
Bahn 1983, 366, Table 6. Abri des Battuts, Penne, Tarn, Frankreich (Aurignacien und Gravettien) Schicht
Rang
ifer
tara
ndus
Equu
s fer
us
Equu
s hy
drun
tinus
Bos /
Bis
on
Capr
a ib
ex
C. p
yren
aica
Rupi
capr
a ru
pica
pra
Cer
vus
elap
hus
Sus s
crof
a
Ges
amt
n % n % n % n % n % n % n % n % n12, Gr. Noailles 5 59, Gravettien 4 7,3 17 30,9 1 1,8 26 47,3 1 1,8 6 10,9 557, Gr. Noailles 46 56,8 3 3,7 4 4,9 12 14,8 14 17,3 2 2,5 815, Gravettien 2 0,6 29 8,8 32 9,8 98 29,9 11 3,4 155 47,3 1 0,3 3283 Aurignacien 83 69,2 1 0,8 1 0,8 8 6,7 18 15,0 4 3,3 5 4,2 120Déblais 560 57,4 78 8,0 24 2,5 157 16,1 101 10,3 52 5,3 4 0,4 976Delpech 1983, 358, Tableau 14.
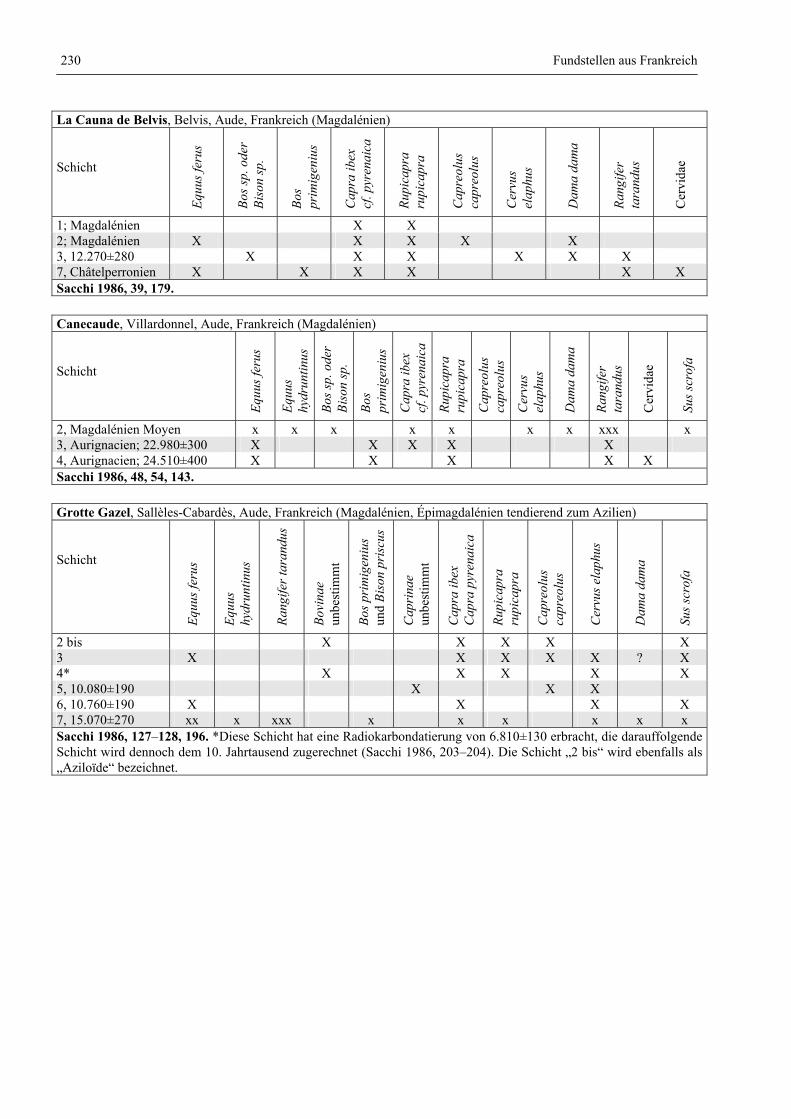
Fundstellen aus Frankreich 230
La Cauna de Belvis, Belvis, Aude, Frankreich (Magdalénien) Schicht
Equu
s fer
us
Bos s
p. o
der
Biso
n sp
.
Bos
prim
igen
ius
Capr
a ib
ex
cf. p
yren
aica
Rupi
capr
a ru
pica
pra
Capr
eolu
s ca
preo
lus
Cer
vus
elap
hus
Dam
a da
ma
Rang
ifer
tara
ndus
Cer
vida
e
1; Magdalénien X X 2; Magdalénien X X X X X 3, 12.270±280 X X X X X X 7, Châtelperronien X X X X X X Sacchi 1986, 39, 179. Canecaude, Villardonnel, Aude, Frankreich (Magdalénien) Schicht
Equu
s fer
us
Equu
s hy
drun
tinus
Bos s
p. o
der
Biso
n sp
.
Bos
prim
igen
ius
Capr
a ib
ex
cf. p
yren
aica
Rupi
capr
a ru
pica
pra
Capr
eolu
s ca
preo
lus
Cer
vus
elap
hus
Dam
a da
ma
Rang
ifer
tara
ndus
Cer
vida
e
Sus s
crof
a
2, Magdalénien Moyen x x x x x x x xxx x 3, Aurignacien; 22.980±300 X X X X X 4, Aurignacien; 24.510±400 X X X X X Sacchi 1986, 48, 54, 143. Grotte Gazel, Sallèles-Cabardès, Aude, Frankreich (Magdalénien, Épimagdalénien tendierend zum Azilien) Schicht
Equu
s fer
us
Equu
s hy
drun
tinus
Rang
ifer t
aran
dus
Bovi
nae
unbe
stim
mt
Bos p
rim
igen
ius
und
Biso
n pr
iscus
Capr
inae
un
best
imm
t
Capr
a ib
ex
Capr
a py
rena
ica
Rupi
capr
a ru
pica
pra
Capr
eolu
s ca
preo
lus
Cerv
us e
laph
us
Dam
a da
ma
Sus s
crof
a
2 bis X X X X X 3 X X X X X ? X 4* X X X X X 5, 10.080±190 X X X 6, 10.760±190 X X X X 7, 15.070±270 xx x xxx x x x x x x Sacchi 1986, 127–128, 196. *Diese Schicht hat eine Radiokarbondatierung von 6.810±130 erbracht, die darauffolgende Schicht wird dennoch dem 10. Jahrtausend zugerechnet (Sacchi 1986, 203–204). Die Schicht „2 bis“ wird ebenfalls als „Aziloïde“ bezeichnet.
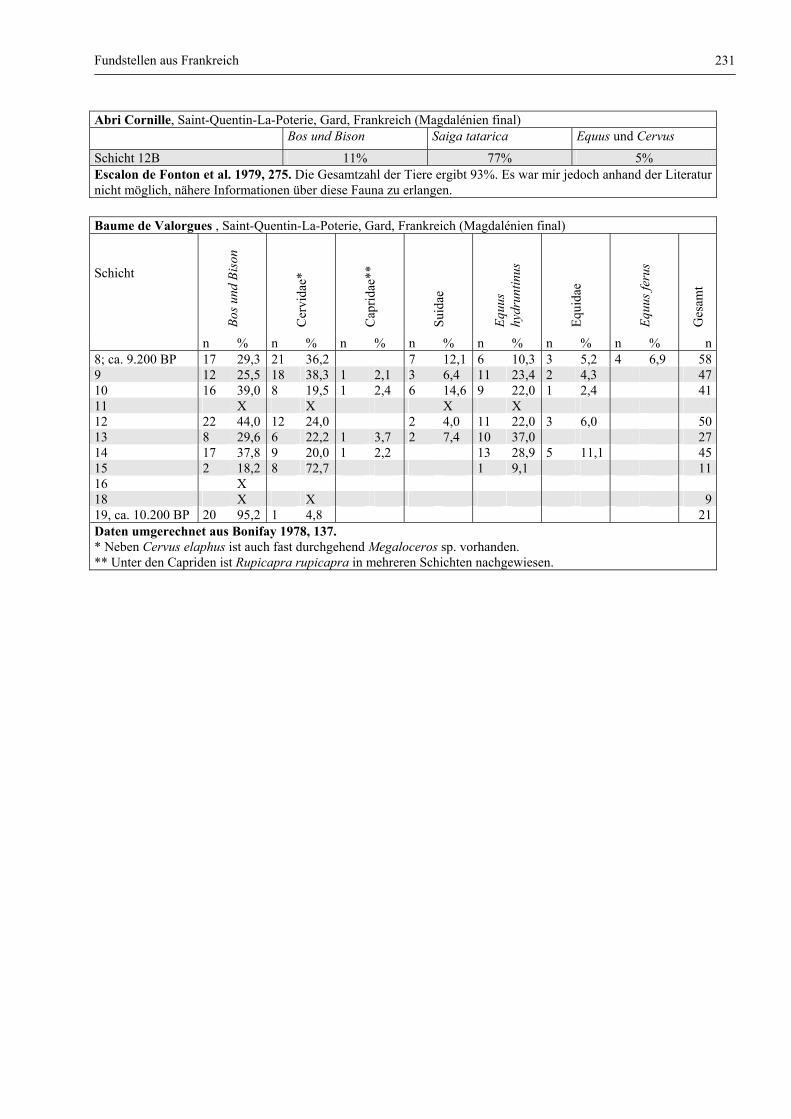
Fundstellen aus Frankreich
231
Abri Cornille, Saint-Quentin-La-Poterie, Gard, Frankreich (Magdalénien final) Bos und Bison Saiga tatarica Equus und Cervus
Schicht 12B 11% 77% 5% Escalon de Fonton et al. 1979, 275. Die Gesamtzahl der Tiere ergibt 93%. Es war mir jedoch anhand der Literatur nicht möglich, nähere Informationen über diese Fauna zu erlangen. Baume de Valorgues , Saint-Quentin-La-Poterie, Gard, Frankreich (Magdalénien final) Schicht
Bos u
nd B
ison
Cer
vida
e*
Cap
ridae
**
Suid
ae
Equu
s hy
drun
tinus
Equi
dae
Equu
s fer
us
Ges
amt
n % n % n % n % n % n % n % n8; ca. 9.200 BP 17 29,3 21 36,2 7 12,1 6 10,3 3 5,2 4 6,9 589 12 25,5 18 38,3 1 2,1 3 6,4 11 23,4 2 4,3 4710 16 39,0 8 19,5 1 2,4 6 14,6 9 22,0 1 2,4 4111 X X X X 12 22 44,0 12 24,0 2 4,0 11 22,0 3 6,0 5013 8 29,6 6 22,2 1 3,7 2 7,4 10 37,0 2714 17 37,8 9 20,0 1 2,2 13 28,9 5 11,1 4515 2 18,2 8 72,7 1 9,1 1116 X 18 X X 919, ca. 10.200 BP 20 95,2 1 4,8 21Daten umgerechnet aus Bonifay 1978, 137. * Neben Cervus elaphus ist auch fast durchgehend Megaloceros sp. vorhanden. ** Unter den Capriden ist Rupicapra rupicapra in mehreren Schichten nachgewiesen.
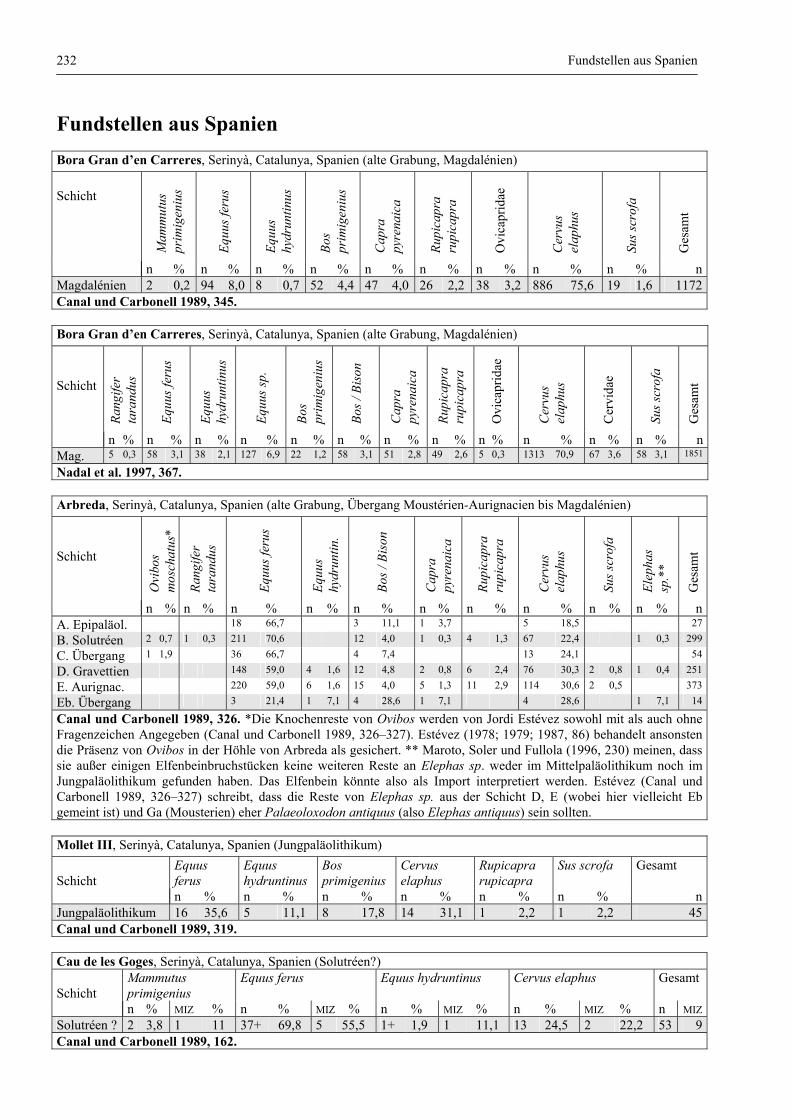
Fundstellen aus Spanien 232
Fundstellen aus Spanien Bora Gran d’en Carreres, Serinyà, Catalunya, Spanien (alte Grabung, Magdalénien) Schicht
Mam
mut
us
prim
igen
ius
Equu
s fer
us
Equu
s hy
drun
tinus
Bos
prim
igen
ius
Capr
a py
rena
ica
Rupi
capr
a ru
pica
pra
Ovi
capr
idae
Cer
vus
elap
hus
Sus s
crof
a
Ges
amt
n % n % n % n % n % n % n % n % n % nMagdalénien 2 0,2 94 8,0 8 0,7 52 4,4 47 4,0 26 2,2 38 3,2 886 75,6 19 1,6 1172Canal und Carbonell 1989, 345. Bora Gran d’en Carreres, Serinyà, Catalunya, Spanien (alte Grabung, Magdalénien) Schicht
Rang
ifer
tara
ndus
Equu
s fer
us
Equu
s hy
drun
tinus
Equu
s sp.
Bos
prim
igen
ius
Bos /
Bis
on
Capr
a py
rena
ica
Rupi
capr
a ru
pica
pra
Ovi
capr
idae
Cer
vus
elap
hus
Cer
vida
e
Sus s
crof
a
Ges
amt
n % n % n % n % n % n % n % n % n % n % n % n % nMag. 5 0,3 58 3,1 38 2,1 127 6,9 22 1,2 58 3,1 51 2,8 49 2,6 5 0,3 1313 70,9 67 3,6 58 3,1 1851
Nadal et al. 1997, 367. Arbreda, Serinyà, Catalunya, Spanien (alte Grabung, Übergang Moustérien-Aurignacien bis Magdalénien) Schicht
Ovi
bos
mos
chat
us*
Rang
ifer
tara
ndus
Equu
s fer
us
Equu
s hy
drun
tin.
Bos /
Bis
on
Capr
a py
rena
ica
Rupi
capr
a ru
pica
pra
Cer
vus
elap
hus
Sus s
crof
a
Elep
has
sp.*
*
Ges
amt
n % n % n % n % n % n % n % n % n % n % nA. Epipaläol. 18 66,7 3 11,1 1 3,7 5 18,5 27
B. Solutréen 2 0,7 1 0,3 211 70,6 12 4,0 1 0,3 4 1,3 67 22,4 1 0,3 299
C. Übergang 1 1,9 36 66,7 4 7,4 13 24,1 54
D. Gravettien 148 59,0 4 1,6 12 4,8 2 0,8 6 2,4 76 30,3 2 0,8 1 0,4 251
E. Aurignac. 220 59,0 6 1,6 15 4,0 5 1,3 11 2,9 114 30,6 2 0,5 373
Eb. Übergang 3 21,4 1 7,1 4 28,6 1 7,1 4 28,6 1 7,1 14
Canal und Carbonell 1989, 326. *Die Knochenreste von Ovibos werden von Jordi Estévez sowohl mit als auch ohne Fragenzeichen Angegeben (Canal und Carbonell 1989, 326–327). Estévez (1978; 1979; 1987, 86) behandelt ansonsten die Präsenz von Ovibos in der Höhle von Arbreda als gesichert. ** Maroto, Soler und Fullola (1996, 230) meinen, dass sie außer einigen Elfenbeinbruchstücken keine weiteren Reste an Elephas sp. weder im Mittelpaläolithikum noch im Jungpaläolithikum gefunden haben. Das Elfenbein könnte also als Import interpretiert werden. Estévez (Canal und Carbonell 1989, 326–327) schreibt, dass die Reste von Elephas sp. aus der Schicht D, E (wobei hier vielleicht Eb gemeint ist) und Ga (Mousterien) eher Palaeoloxodon antiquus (also Elephas antiquus) sein sollten. Mollet III, Serinyà, Catalunya, Spanien (Jungpaläolithikum) Schicht
Equus ferus
Equus hydruntinus
Bos primigenius
Cervus elaphus
Rupicapra rupicapra
Sus scrofa Gesamt
n % n % n % n % n % n % nJungpaläolithikum 16 35,6 5 11,1 8 17,8 14 31,1 1 2,2 1 2,2 45Canal und Carbonell 1989, 319. Cau de les Goges, Serinyà, Catalunya, Spanien (Solutréen?) Schicht
Mammutus primigenius
Equus ferus Equus hydruntinus Cervus elaphus Gesamt
n % MIZ % n % MIZ % n % MIZ % n % MIZ % n MIZSolutréen ? 2 3,8 1 11 37+ 69,8 5 55,5 1+ 1,9 1 11,1 13 24,5 2 22,2 53 9Canal und Carbonell 1989, 162.
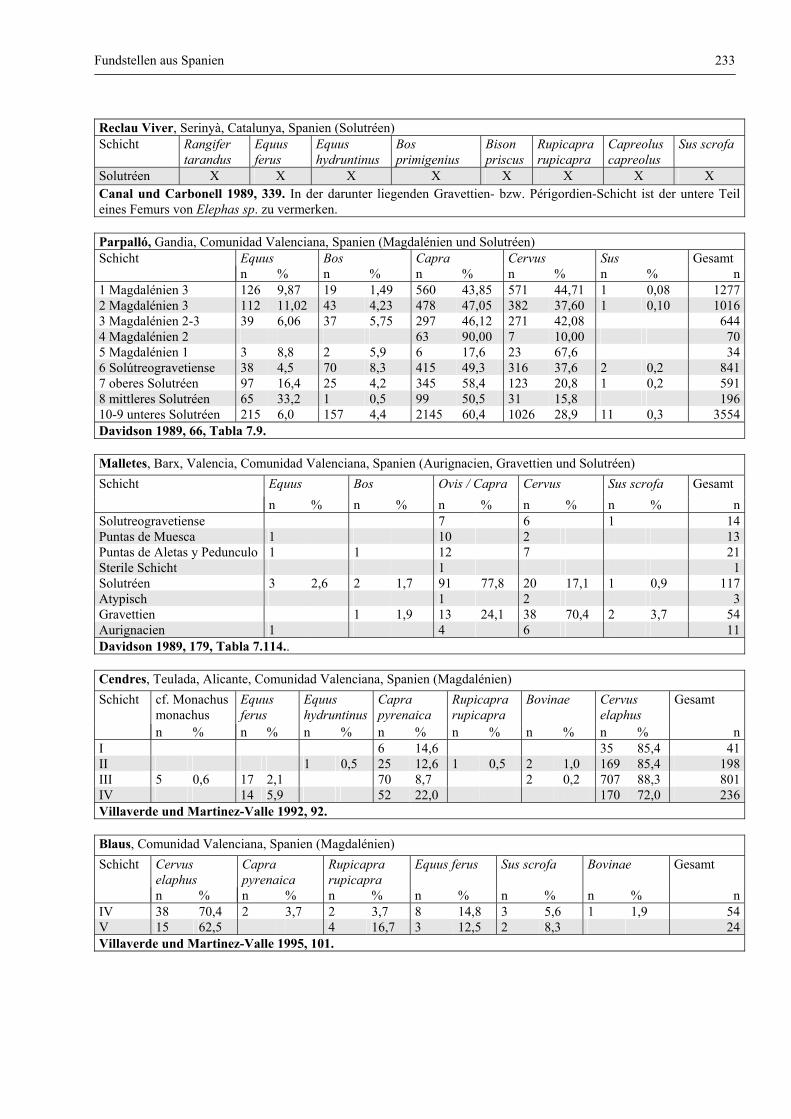
Fundstellen aus Spanien 233
Reclau Viver, Serinyà, Catalunya, Spanien (Solutréen) Schicht Rangifer
tarandus Equus ferus
Equus hydruntinus
Bos primigenius
Bison priscus
Rupicapra rupicapra
Capreolus capreolus
Sus scrofa
Solutréen X X X X X X X X Canal und Carbonell 1989, 339. In der darunter liegenden Gravettien- bzw. Périgordien-Schicht ist der untere Teil eines Femurs von Elephas sp. zu vermerken.
Parpalló, Gandia, Comunidad Valenciana, Spanien (Magdalénien und Solutréen) Schicht Equus Bos Capra Cervus Sus Gesamt
n % n % n % n % n % n1 Magdalénien 3 126 9,87 19 1,49 560 43,85 571 44,71 1 0,08 12772 Magdalénien 3 112 11,02 43 4,23 478 47,05 382 37,60 1 0,10 10163 Magdalénien 2-3 39 6,06 37 5,75 297 46,12 271 42,08 6444 Magdalénien 2 63 90,00 7 10,00 705 Magdalénien 1 3 8,8 2 5,9 6 17,6 23 67,6 346 Solútreogravetiense 38 4,5 70 8,3 415 49,3 316 37,6 2 0,2 8417 oberes Solutréen 97 16,4 25 4,2 345 58,4 123 20,8 1 0,2 5918 mittleres Solutréen 65 33,2 1 0,5 99 50,5 31 15,8 19610-9 unteres Solutréen 215 6,0 157 4,4 2145 60,4 1026 28,9 11 0,3 3554Davidson 1989, 66, Tabla 7.9.
Malletes, Barx, Valencia, Comunidad Valenciana, Spanien (Aurignacien, Gravettien und Solutréen) Schicht Equus Bos Ovis / Capra Cervus Sus scrofa Gesamt
n % n % n % n % n % nSolutreogravetiense 7 6 1 14Puntas de Muesca 1 10 2 13Puntas de Aletas y Pedunculo 1 1 12 7 21Sterile Schicht 1 1Solutréen 3 2,6 2 1,7 91 77,8 20 17,1 1 0,9 117Atypisch 1 2 3Gravettien 1 1,9 13 24,1 38 70,4 2 3,7 54Aurignacien 1 4 6 11Davidson 1989, 179, Tabla 7.114..
Cendres, Teulada, Alicante, Comunidad Valenciana, Spanien (Magdalénien) Schicht cf. Monachus
monachus Equus ferus
Equus hydruntinus
Capra pyrenaica
Rupicapra rupicapra
Bovinae Cervus elaphus
Gesamt
n % n % n % n % n % n % n % nI 6 14,6 35 85,4 41II 1 0,5 25 12,6 1 0,5 2 1,0 169 85,4 198III 5 0,6 17 2,1 70 8,7 2 0,2 707 88,3 801IV 14 5,9 52 22,0 170 72,0 236Villaverde und Martinez-Valle 1992, 92.
Blaus, Comunidad Valenciana, Spanien (Magdalénien) Schicht Cervus
elaphus Capra pyrenaica
Rupicapra rupicapra
Equus ferus Sus scrofa Bovinae Gesamt
n % n % n % n % n % n % nIV 38 70,4 2 3,7 2 3,7 8 14,8 3 5,6 1 1,9 54V 15 62,5 4 16,7 3 12,5 2 8,3 24Villaverde und Martinez-Valle 1995, 101.
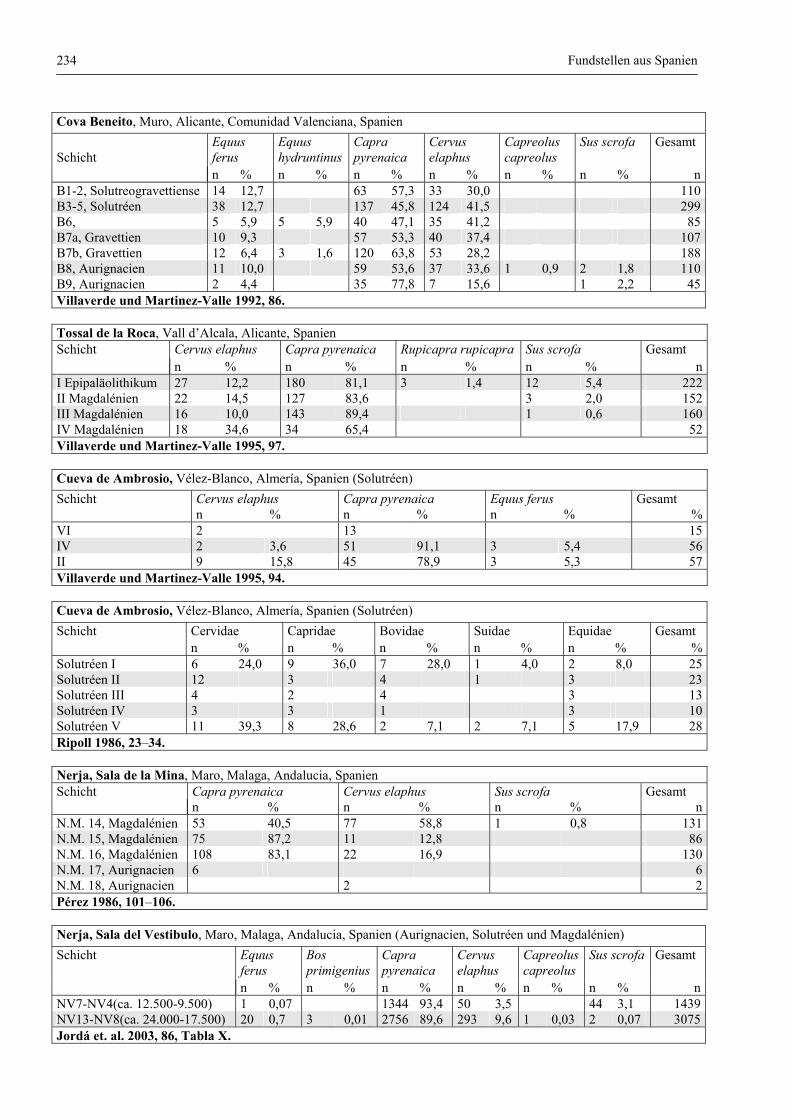
Fundstellen aus Spanien 234
Cova Beneito, Muro, Alicante, Comunidad Valenciana, Spanien Schicht
Equus ferus
Equus hydruntinus
Capra pyrenaica
Cervus elaphus
Capreolus capreolus
Sus scrofa Gesamt
n % n % n % n % n % n % nB1-2, Solutreogravettiense 14 12,7 63 57,3 33 30,0 110B3-5, Solutréen 38 12,7 137 45,8 124 41,5 299B6, 5 5,9 5 5,9 40 47,1 35 41,2 85B7a, Gravettien 10 9,3 57 53,3 40 37,4 107B7b, Gravettien 12 6,4 3 1,6 120 63,8 53 28,2 188B8, Aurignacien 11 10,0 59 53,6 37 33,6 1 0,9 2 1,8 110B9, Aurignacien 2 4,4 35 77,8 7 15,6 1 2,2 45Villaverde und Martinez-Valle 1992, 86. Tossal de la Roca, Vall d’Alcala, Alicante, Spanien Schicht Cervus elaphus Capra pyrenaica Rupicapra rupicapra Sus scrofa Gesamt n % n % n % n % nI Epipaläolithikum 27 12,2 180 81,1 3 1,4 12 5,4 222II Magdalénien 22 14,5 127 83,6 3 2,0 152III Magdalénien 16 10,0 143 89,4 1 0,6 160IV Magdalénien 18 34,6 34 65,4 52Villaverde und Martinez-Valle 1995, 97. Cueva de Ambrosio, Vélez-Blanco, Almería, Spanien (Solutréen) Schicht Cervus elaphus Capra pyrenaica Equus ferus Gesamt n % n % n % %VI 2 13 15IV 2 3,6 51 91,1 3 5,4 56II 9 15,8 45 78,9 3 5,3 57Villaverde und Martinez-Valle 1995, 94. Cueva de Ambrosio, Vélez-Blanco, Almería, Spanien (Solutréen) Schicht Cervidae Capridae Bovidae Suidae Equidae Gesamt n % n % n % n % n % %Solutréen I 6 24,0 9 36,0 7 28,0 1 4,0 2 8,0 25Solutréen II 12 3 4 1 3 23Solutréen III 4 2 4 3 13Solutréen IV 3 3 1 3 10Solutréen V 11 39,3 8 28,6 2 7,1 2 7,1 5 17,9 28Ripoll 1986, 23–34. Nerja, Sala de la Mina, Maro, Malaga, Andalucia, Spanien Schicht Capra pyrenaica Cervus elaphus Sus scrofa Gesamt n % n % n % nN.M. 14, Magdalénien 53 40,5 77 58,8 1 0,8 131N.M. 15, Magdalénien 75 87,2 11 12,8 86N.M. 16, Magdalénien 108 83,1 22 16,9 130N.M. 17, Aurignacien 6 6N.M. 18, Aurignacien 2 2Pérez 1986, 101–106. Nerja, Sala del Vestibulo, Maro, Malaga, Andalucia, Spanien (Aurignacien, Solutréen und Magdalénien) Schicht Equus
ferus Bos primigenius
Capra pyrenaica
Cervus elaphus
Capreolus capreolus
Sus scrofa Gesamt
n % n % n % n % n % n % nNV7-NV4(ca. 12.500-9.500) 1 0,07 1344 93,4 50 3,5 44 3,1 1439NV13-NV8(ca. 24.000-17.500) 20 0,7 3 0,01 2756 89,6 293 9,6 1 0,03 2 0,07 3075Jordá et. al. 2003, 86, Tabla X.
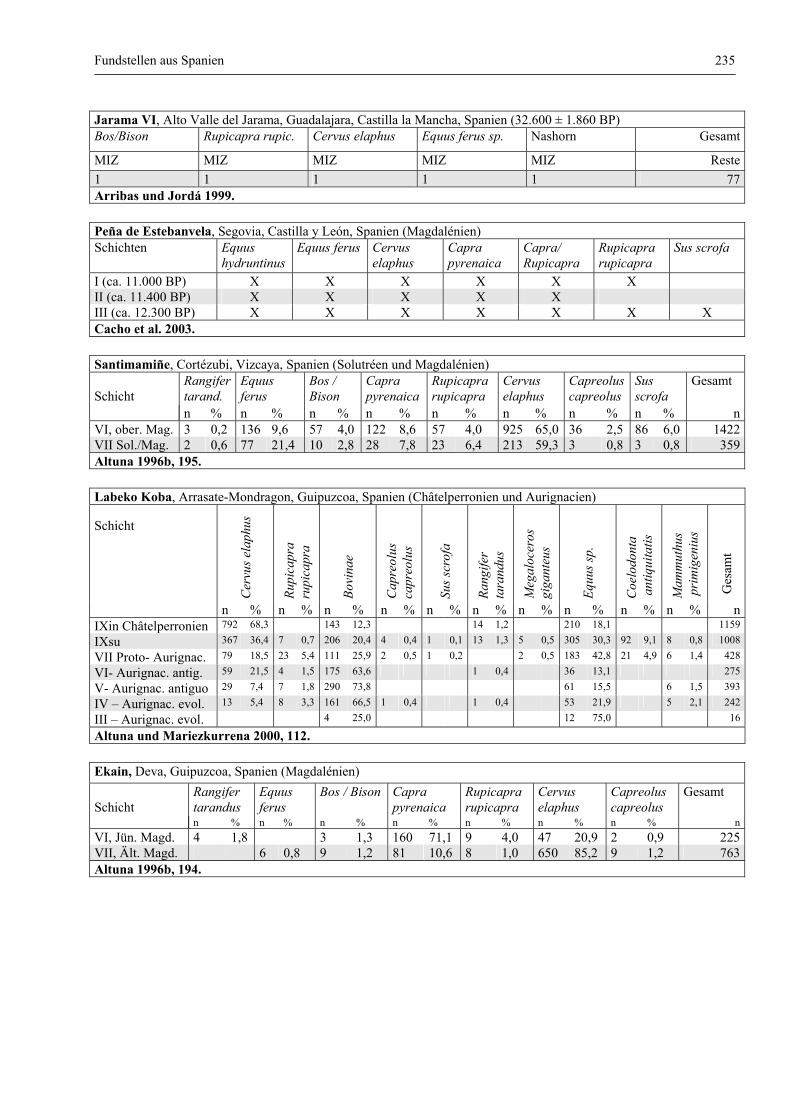
Fundstellen aus Spanien 235
Jarama VI, Alto Valle del Jarama, Guadalajara, Castilla la Mancha, Spanien (32.600 ± 1.860 BP) Bos/Bison Rupicapra rupic. Cervus elaphus Equus ferus sp. Nashorn Gesamt
MIZ MIZ MIZ MIZ MIZ Reste1 1 1 1 1 77Arribas und Jordá 1999. Peña de Estebanvela, Segovia, Castilla y León, Spanien (Magdalénien) Schichten Equus
hydruntinus Equus ferus Cervus
elaphus Capra pyrenaica
Capra/ Rupicapra
Rupicapra rupicapra
Sus scrofa
I (ca. 11.000 BP) X X X X X X II (ca. 11.400 BP) X X X X X III (ca. 12.300 BP) X X X X X X X Cacho et al. 2003. Santimamiñe, Cortézubi, Vizcaya, Spanien (Solutréen und Magdalénien) Schicht
Rangifer tarand.
Equus ferus
Bos / Bison
Capra pyrenaica
Rupicapra rupicapra
Cervus elaphus
Capreolus capreolus
Sus scrofa
Gesamt
n % n % n % n % n % n % n % n % nVI, ober. Mag. 3 0,2 136 9,6 57 4,0 122 8,6 57 4,0 925 65,0 36 2,5 86 6,0 1422VII Sol./Mag. 2 0,6 77 21,4 10 2,8 28 7,8 23 6,4 213 59,3 3 0,8 3 0,8 359Altuna 1996b, 195. Labeko Koba, Arrasate-Mondragon, Guipuzcoa, Spanien (Châtelperronien und Aurignacien) Schicht
Cerv
us e
laph
us
Rupi
capr
a ru
pica
pra
Bovi
nae
Capr
eolu
s ca
preo
lus
Sus s
crof
a
Rang
ifer
tara
ndus
Meg
aloc
eros
gi
gant
eus
Equu
s sp.
Coel
odon
ta
ant
iqui
tatis
Mam
mut
hus
pri
mig
eniu
s
Ges
amt
n % n % n % n % n % n % n % n % n % n % nIXin Châtelperronien 792 68,3 143 12,3 14 1,2 210 18,1 1159
IXsu 367 36,4 7 0,7 206 20,4 4 0,4 1 0,1 13 1,3 5 0,5 305 30,3 92 9,1 8 0,8 1008
VII Proto- Aurignac. 79 18,5 23 5,4 111 25,9 2 0,5 1 0,2 2 0,5 183 42,8 21 4,9 6 1,4 428
VI- Aurignac. antig. 59 21,5 4 1,5 175 63,6 1 0,4 36 13,1 275
V- Aurignac. antiguo 29 7,4 7 1,8 290 73,8 61 15,5 6 1,5 393
IV – Aurignac. evol. 13 5,4 8 3,3 161 66,5 1 0,4 1 0,4 53 21,9 5 2,1 242
III – Aurignac. evol. 4 25,0 12 75,0 16
Altuna und Mariezkurrena 2000, 112. Ekain, Deva, Guipuzcoa, Spanien (Magdalénien) Schicht
Rangifer tarandus
Equus ferus
Bos / Bison Capra pyrenaica
Rupicapra rupicapra
Cervus elaphus
Capreolus capreolus
Gesamt
n % n % n % n % n % n % n % nVI, Jün. Magd. 4 1,8 3 1,3 160 71,1 9 4,0 47 20,9 2 0,9 225VII, Ält. Magd. 6 0,8 9 1,2 81 10,6 8 1,0 650 85,2 9 1,2 763Altuna 1996b, 194.
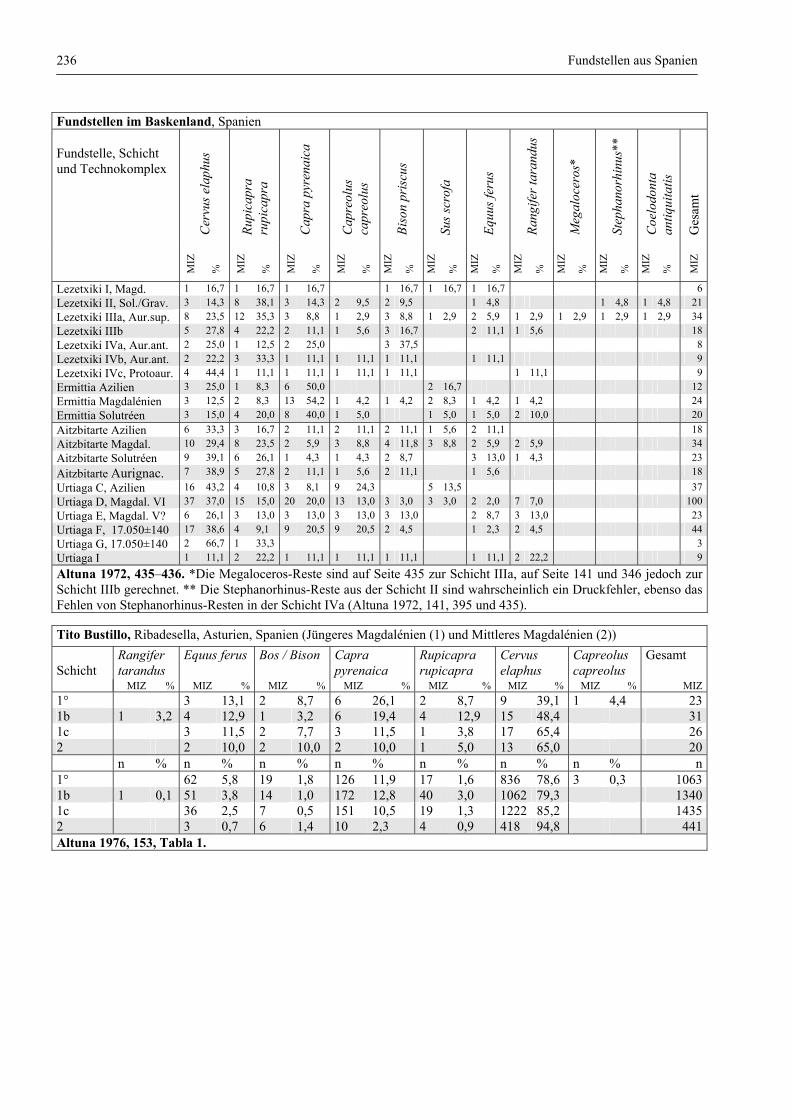
Fundstellen aus Spanien
236
Fundstellen im Baskenland, Spanien Fundstelle, Schicht und Technokomplex
Cerv
us e
laph
us
Rupi
capr
a ru
pica
pra
Capr
a py
rena
ica
Capr
eolu
s ca
preo
lus
Biso
n pr
iscus
Sus s
crof
a
Equu
s fer
us
Rang
ifer t
aran
dus
Meg
aloc
eros
*
Step
hano
rhin
us**
Coel
odon
ta
antiq
uita
tis
Ges
amt
MIZ
%
MIZ
%
MIZ
%
MIZ
%
MIZ
%
MIZ
%
MIZ
%
MIZ
%
MIZ
%
MIZ
%
MIZ
%
MIZ
Lezetxiki I, Magd. 1 16,7 1 16,7 1 16,7 1 16,7 1 16,7 1 16,7 6Lezetxiki II, Sol./Grav. 3 14,3 8 38,1 3 14,3 2 9,5 2 9,5 1 4,8 1 4,8 1 4,8 21Lezetxiki IIIa, Aur.sup. 8 23,5 12 35,3 3 8,8 1 2,9 3 8,8 1 2,9 2 5,9 1 2,9 1 2,9 1 2,9 1 2,9 34Lezetxiki IIIb 5 27,8 4 22,2 2 11,1 1 5,6 3 16,7 2 11,1 1 5,6 18Lezetxiki IVa, Aur.ant. 2 25,0 1 12,5 2 25,0 3 37,5 8Lezetxiki IVb, Aur.ant. 2 22,2 3 33,3 1 11,1 1 11,1 1 11,1 1 11,1 9Lezetxiki IVc, Protoaur. 4 44,4 1 11,1 1 11,1 1 11,1 1 11,1 1 11,1 9Ermittia Azilien 3 25,0 1 8,3 6 50,0 2 16,7 12Ermittia Magdalénien 3 12,5 2 8,3 13 54,2 1 4,2 1 4,2 2 8,3 1 4,2 1 4,2 24Ermittia Solutréen 3 15,0 4 20,0 8 40,0 1 5,0 1 5,0 1 5,0 2 10,0 20Aitzbitarte Azilien 6 33,3 3 16,7 2 11,1 2 11,1 2 11,1 1 5,6 2 11,1 18Aitzbitarte Magdal. 10 29,4 8 23,5 2 5,9 3 8,8 4 11,8 3 8,8 2 5,9 2 5,9 34Aitzbitarte Solutréen 9 39,1 6 26,1 1 4,3 1 4,3 2 8,7 3 13,0 1 4,3 23Aitzbitarte Aurignac. 7 38,9 5 27,8 2 11,1 1 5,6 2 11,1 1 5,6 18Urtiaga C, Azilien 16 43,2 4 10,8 3 8,1 9 24,3 5 13,5 37Urtiaga D, Magdal. VI 37 37,0 15 15,0 20 20,0 13 13,0 3 3,0 3 3,0 2 2,0 7 7,0 100Urtiaga E, Magdal. V? 6 26,1 3 13,0 3 13,0 3 13,0 3 13,0 2 8,7 3 13,0 23Urtiaga F, 17.050±140 17 38,6 4 9,1 9 20,5 9 20,5 2 4,5 1 2,3 2 4,5 44Urtiaga G, 17.050±140 2 66,7 1 33,3 3Urtiaga I 1 11,1 2 22,2 1 11,1 1 11,1 1 11,1 1 11,1 2 22,2 9
Altuna 1972, 435–436. *Die Megaloceros-Reste sind auf Seite 435 zur Schicht IIIa, auf Seite 141 und 346 jedoch zur Schicht IIIb gerechnet. ** Die Stephanorhinus-Reste aus der Schicht II sind wahrscheinlich ein Druckfehler, ebenso das Fehlen von Stephanorhinus-Resten in der Schicht IVa (Altuna 1972, 141, 395 und 435). Tito Bustillo, Ribadesella, Asturien, Spanien (Jüngeres Magdalénien (1) und Mittleres Magdalénien (2)) Schicht
Rangifer tarandus
Equus ferus Bos / Bison Capra pyrenaica
Rupicapra rupicapra
Cervus elaphus
Capreolus capreolus
Gesamt
MIZ % MIZ % MIZ % MIZ % MIZ % MIZ % MIZ % MIZ1° 3 13,1 2 8,7 6 26,1 2 8,7 9 39,1 1 4,4 231b 1 3,2 4 12,9 1 3,2 6 19,4 4 12,9 15 48,4 311c 3 11,5 2 7,7 3 11,5 1 3,8 17 65,4 262 2 10,0 2 10,0 2 10,0 1 5,0 13 65,0 20 n % n % n % n % n % n % n % n1° 62 5,8 19 1,8 126 11,9 17 1,6 836 78,6 3 0,3 10631b 1 0,1 51 3,8 14 1,0 172 12,8 40 3,0 1062 79,3 13401c 36 2,5 7 0,5 151 10,5 19 1,3 1222 85,2 14352 3 0,7 6 1,4 10 2,3 4 0,9 418 94,8 441Altuna 1976, 153, Tabla 1.
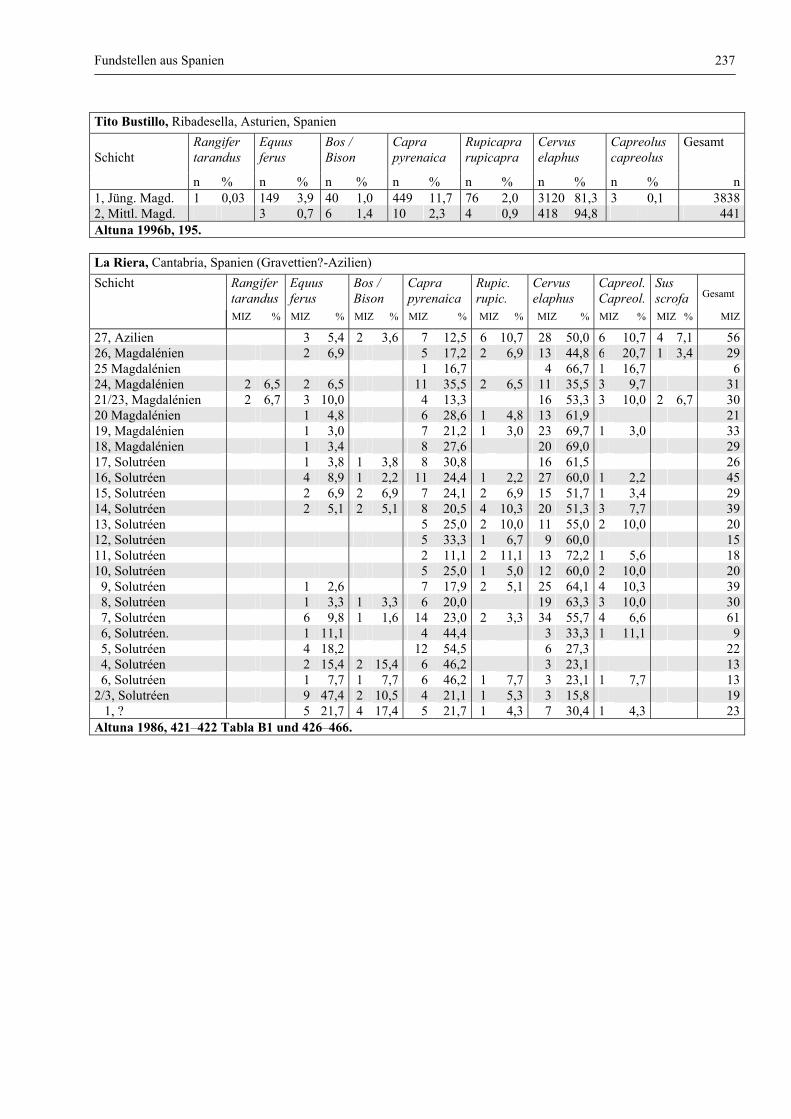
Fundstellen aus Spanien 237
Tito Bustillo, Ribadesella, Asturien, Spanien Rangifer tarandus
Equus ferus
Bos / Bison
Capra pyrenaica
Rupicapra rupicapra
Cervus elaphus
Capreolus capreolus
Gesamt Schicht
n % n % n % n % n % n % n % n1, Jüng. Magd. 1 0,03 149 3,9 40 1,0 449 11,7 76 2,0 3120 81,3 3 0,1 38382, Mittl. Magd. 3 0,7 6 1,4 10 2,3 4 0,9 418 94,8 441Altuna 1996b, 195. La Riera, Cantabria, Spanien (Gravettien?-Azilien) Schicht Rangifer
tarandus Equus ferus
Bos / Bison
Capra pyrenaica
Rupic. rupic.
Cervus elaphus
Capreol. Capreol.
Sus scrofa
Gesamt
MIZ % MIZ % MIZ % MIZ % MIZ % MIZ % MIZ % MIZ % MIZ
27, Azilien 3 5,4 2 3,6 7 12,5 6 10,7 28 50,0 6 10,7 4 7,1 5626, Magdalénien 2 6,9 5 17,2 2 6,9 13 44,8 6 20,7 1 3,4 2925 Magdalénien 1 16,7 4 66,7 1 16,7 624, Magdalénien 2 6,5 2 6,5 11 35,5 2 6,5 11 35,5 3 9,7 3121/23, Magdalénien 2 6,7 3 10,0 4 13,3 16 53,3 3 10,0 2 6,7 3020 Magdalénien 1 4,8 6 28,6 1 4,8 13 61,9 2119, Magdalénien 1 3,0 7 21,2 1 3,0 23 69,7 1 3,0 3318, Magdalénien 1 3,4 8 27,6 20 69,0 2917, Solutréen 1 3,8 1 3,8 8 30,8 16 61,5 2616, Solutréen 4 8,9 1 2,2 11 24,4 1 2,2 27 60,0 1 2,2 4515, Solutréen 2 6,9 2 6,9 7 24,1 2 6,9 15 51,7 1 3,4 2914, Solutréen 2 5,1 2 5,1 8 20,5 4 10,3 20 51,3 3 7,7 3913, Solutréen 5 25,0 2 10,0 11 55,0 2 10,0 2012, Solutréen 5 33,3 1 6,7 9 60,0 1511, Solutréen 2 11,1 2 11,1 13 72,2 1 5,6 1810, Solutréen 5 25,0 1 5,0 12 60,0 2 10,0 20 9, Solutréen 1 2,6 7 17,9 2 5,1 25 64,1 4 10,3 39 8, Solutréen 1 3,3 1 3,3 6 20,0 19 63,3 3 10,0 30 7, Solutréen 6 9,8 1 1,6 14 23,0 2 3,3 34 55,7 4 6,6 61 6, Solutréen. 1 11,1 4 44,4 3 33,3 1 11,1 9 5, Solutréen 4 18,2 12 54,5 6 27,3 22 4, Solutréen 2 15,4 2 15,4 6 46,2 3 23,1 13 6, Solutréen 1 7,7 1 7,7 6 46,2 1 7,7 3 23,1 1 7,7 132/3, Solutréen 9 47,4 2 10,5 4 21,1 1 5,3 3 15,8 19 1, ? 5 21,7 4 17,4 5 21,7 1 4,3 7 30,4 1 4,3 23Altuna 1986, 421–422 Tabla B1 und 426–466.
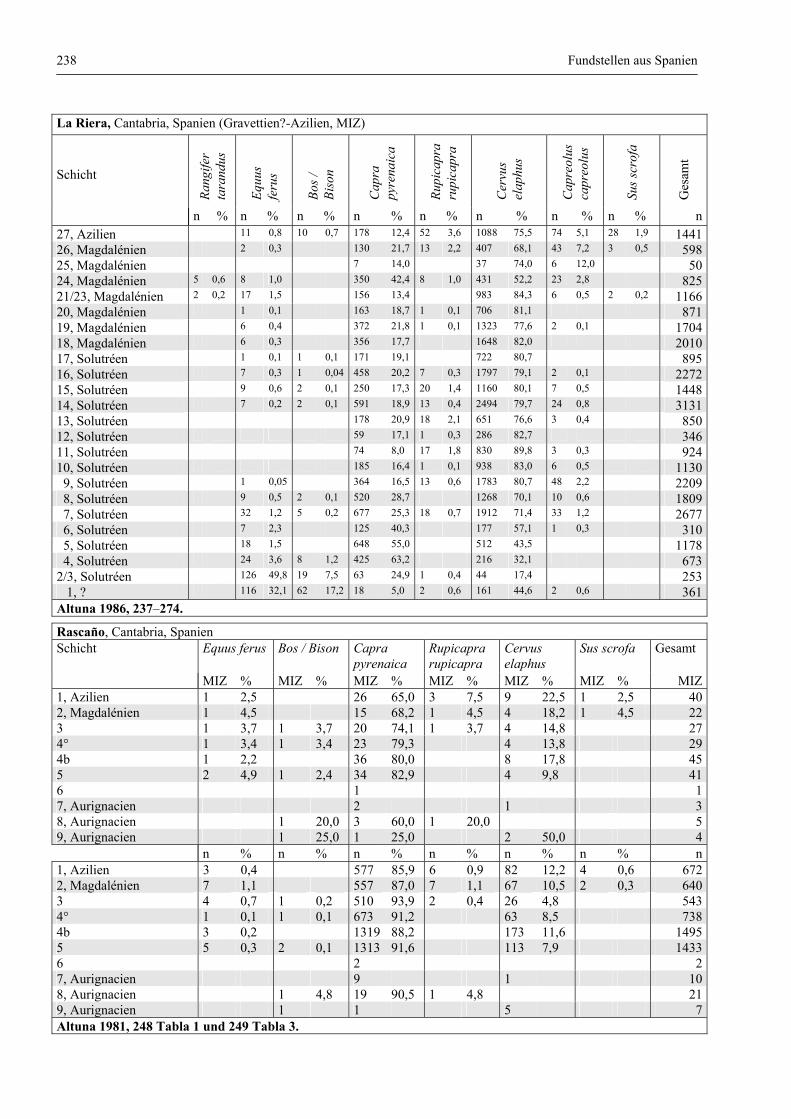
Fundstellen aus Spanien 238
La Riera, Cantabria, Spanien (Gravettien?-Azilien, MIZ) Schicht
Rang
ifer
tara
ndus
Equu
s fe
rus
Bos /
Bi
son
Capr
a py
rena
ica
Rupi
capr
a ru
pica
pra
Cer
vus
elap
hus
Capr
eolu
s ca
preo
lus
Sus s
crof
a
Ges
amt
n % n % n % n % n % n % n % n % n27, Azilien 11 0,8 10 0,7 178 12,4 52 3,6 1088 75,5 74 5,1 28 1,9 144126, Magdalénien 2 0,3 130 21,7 13 2,2 407 68,1 43 7,2 3 0,5 59825, Magdalénien 7 14,0 37 74,0 6 12,0 5024, Magdalénien 5 0,6 8 1,0 350 42,4 8 1,0 431 52,2 23 2,8 82521/23, Magdalénien 2 0,2 17 1,5 156 13,4 983 84,3 6 0,5 2 0,2 116620, Magdalénien 1 0,1 163 18,7 1 0,1 706 81,1 87119, Magdalénien 6 0,4 372 21,8 1 0,1 1323 77,6 2 0,1 170418, Magdalénien 6 0,3 356 17,7 1648 82,0 201017, Solutréen 1 0,1 1 0,1 171 19,1 722 80,7 89516, Solutréen 7 0,3 1 0,04 458 20,2 7 0,3 1797 79,1 2 0,1 227215, Solutréen 9 0,6 2 0,1 250 17,3 20 1,4 1160 80,1 7 0,5 144814, Solutréen 7 0,2 2 0,1 591 18,9 13 0,4 2494 79,7 24 0,8 313113, Solutréen 178 20,9 18 2,1 651 76,6 3 0,4 85012, Solutréen 59 17,1 1 0,3 286 82,7 34611, Solutréen 74 8,0 17 1,8 830 89,8 3 0,3 92410, Solutréen 185 16,4 1 0,1 938 83,0 6 0,5 1130 9, Solutréen 1 0,05 364 16,5 13 0,6 1783 80,7 48 2,2 2209 8, Solutréen 9 0,5 2 0,1 520 28,7 1268 70,1 10 0,6 1809 7, Solutréen 32 1,2 5 0,2 677 25,3 18 0,7 1912 71,4 33 1,2 2677 6, Solutréen 7 2,3 125 40,3 177 57,1 1 0,3 310 5, Solutréen 18 1,5 648 55,0 512 43,5 1178 4, Solutréen 24 3,6 8 1,2 425 63,2 216 32,1 6732/3, Solutréen 126 49,8 19 7,5 63 24,9 1 0,4 44 17,4 253 1, ? 116 32,1 62 17,2 18 5,0 2 0,6 161 44,6 2 0,6 361Altuna 1986, 237–274.
Rascaño, Cantabria, Spanien Schicht Equus ferus Bos / Bison Capra
pyrenaica Rupicapra rupicapra
Cervus elaphus
Sus scrofa Gesamt
MIZ % MIZ % MIZ % MIZ % MIZ % MIZ % MIZ1, Azilien 1 2,5 26 65,0 3 7,5 9 22,5 1 2,5 402, Magdalénien 1 4,5 15 68,2 1 4,5 4 18,2 1 4,5 223 1 3,7 1 3,7 20 74,1 1 3,7 4 14,8 274° 1 3,4 1 3,4 23 79,3 4 13,8 294b 1 2,2 36 80,0 8 17,8 455 2 4,9 1 2,4 34 82,9 4 9,8 416 1 17, Aurignacien 2 1 38, Aurignacien 1 20,0 3 60,0 1 20,0 59, Aurignacien 1 25,0 1 25,0 2 50,0 4
n % n % n % n % n % n % n1, Azilien 3 0,4 577 85,9 6 0,9 82 12,2 4 0,6 6722, Magdalénien 7 1,1 557 87,0 7 1,1 67 10,5 2 0,3 6403 4 0,7 1 0,2 510 93,9 2 0,4 26 4,8 5434° 1 0,1 1 0,1 673 91,2 63 8,5 7384b 3 0,2 1319 88,2 173 11,6 14955 5 0,3 2 0,1 1313 91,6 113 7,9 14336 2 27, Aurignacien 9 1 108, Aurignacien 1 4,8 19 90,5 1 4,8 219, Aurignacien 1 1 5 7Altuna 1981, 248 Tabla 1 und 249 Tabla 3.
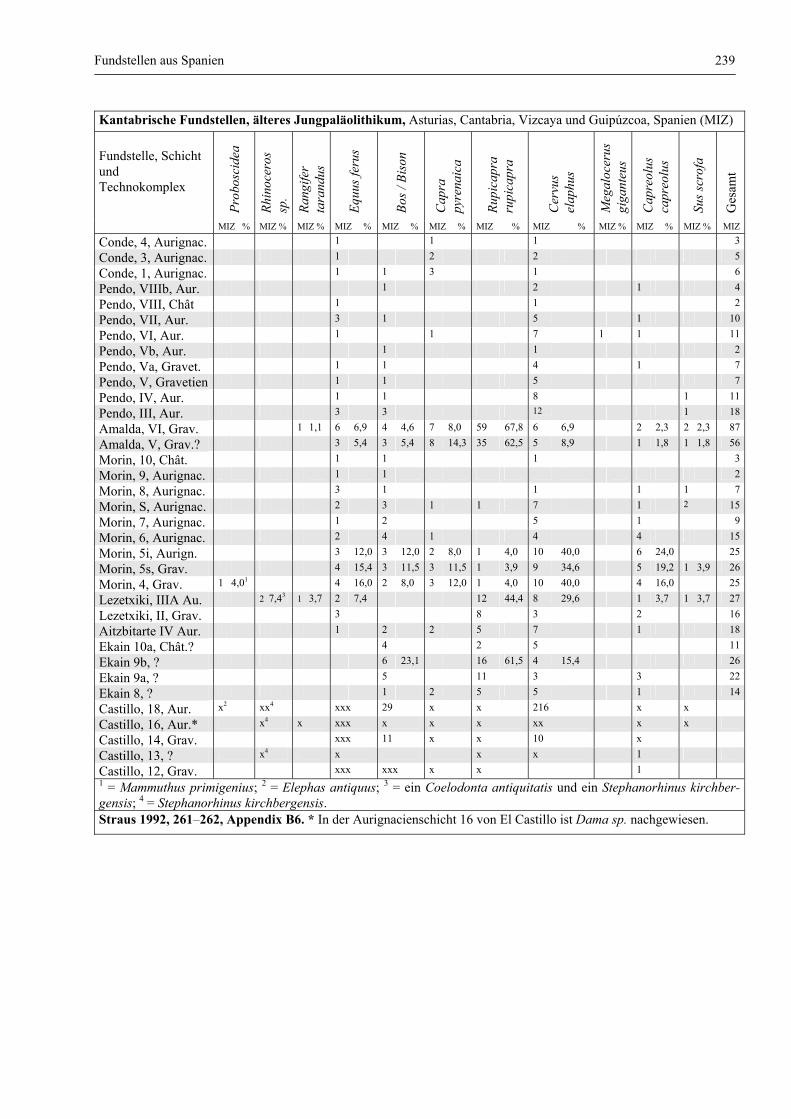
Fundstellen aus Spanien 239
Kantabrische Fundstellen, älteres Jungpaläolithikum, Asturias, Cantabria, Vizcaya und Guipúzcoa, Spanien (MIZ) Fundstelle, Schicht und Technokomplex
Prob
osci
dea
Rhin
ocer
os
sp.
Rang
ifer
tara
ndus
Equu
s fer
us
Bos /
Bis
on
Capr
a py
rena
ica
Rupi
capr
a ru
pica
pra
Cer
vus
elap
hus
Meg
aloc
erus
gi
gant
eus
Capr
eolu
s ca
preo
lus
Sus s
crof
a
Ges
amt
MIZ % MIZ % MIZ % MIZ % MIZ % MIZ % MIZ % MIZ % MIZ % MIZ % MIZ % MIZ
Conde, 4, Aurignac. 1 1 1 3
Conde, 3, Aurignac. 1 2 2 5
Conde, 1, Aurignac. 1 1 3 1 6
Pendo, VIIIb, Aur. 1 2 1 4
Pendo, VIII, Chât 1 1 2
Pendo, VII, Aur. 3 1 5 1 10
Pendo, VI, Aur. 1 1 7 1 1 11
Pendo, Vb, Aur. 1 1 2
Pendo, Va, Gravet. 1 1 4 1 7
Pendo, V, Gravetien 1 1 5 7
Pendo, IV, Aur. 1 1 8 1 11
Pendo, III, Aur. 3 3 12 1 18
Amalda, VI, Grav. 1 1,1 6 6,9 4 4,6 7 8,0 59 67,8 6 6,9 2 2,3 2 2,3 87
Amalda, V, Grav.? 3 5,4 3 5,4 8 14,3 35 62,5 5 8,9 1 1,8 1 1,8 56
Morin, 10, Chât. 1 1 1 3
Morin, 9, Aurignac. 1 1 2
Morin, 8, Aurignac. 3 1 1 1 1 7
Morin, S, Aurignac. 2 3 1 1 7 1 2 15
Morin, 7, Aurignac. 1 2 5 1 9
Morin, 6, Aurignac. 2 4 1 4 4 15
Morin, 5i, Aurign. 3 12,0 3 12,0 2 8,0 1 4,0 10 40,0 6 24,0 25
Morin, 5s, Grav. 4 15,4 3 11,5 3 11,5 1 3,9 9 34,6 5 19,2 1 3,9 26
Morin, 4, Grav. 1 4,01 4 16,0 2 8,0 3 12,0 1 4,0 10 40,0 4 16,0 25
Lezetxiki, IIIA Au. 2 7,43 1 3,7 2 7,4 12 44,4 8 29,6 1 3,7 1 3,7 27
Lezetxiki, II, Grav. 3 8 3 2 16
Aitzbitarte IV Aur. 1 2 2 5 7 1 18
Ekain 10a, Chât.? 4 2 5 11
Ekain 9b, ? 6 23,1 16 61,5 4 15,4 26
Ekain 9a, ? 5 11 3 3 22
Ekain 8, ? 1 2 5 5 1 1 14
Castillo, 18, Aur. x2 xx4 xxx 29 x x 216 x x
Castillo, 16, Aur.* x4 x xxx x x x xx x x
Castillo, 14, Grav. xxx 11 x x 10 x
Castillo, 13, ? x4 x x x 1
Castillo, 12, Grav. xxx xxx x x 1 1 = Mammuthus primigenius; 2 = Elephas antiquus; 3 = ein Coelodonta antiquitatis und ein Stephanorhinus kirchber-gensis; 4 = Stephanorhinus kirchbergensis. Straus 1992, 261–262, Appendix B6. * In der Aurignacienschicht 16 von El Castillo ist Dama sp. nachgewiesen.
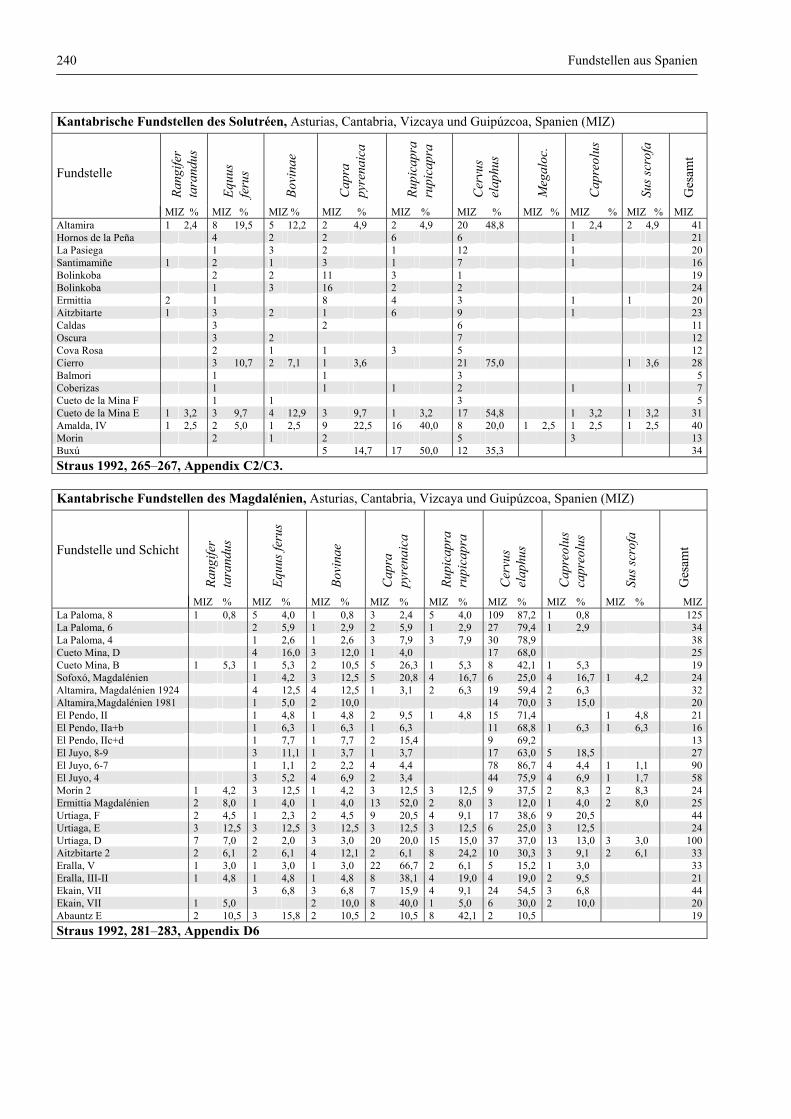
Fundstellen aus Spanien 240
Kantabrische Fundstellen des Solutréen, Asturias, Cantabria, Vizcaya und Guipúzcoa, Spanien (MIZ) Fundstelle
Rang
ifer
tara
ndus
Equu
s fe
rus
Bovi
nae
Capr
a py
rena
ica
Rupi
capr
a ru
pica
pra
Cer
vus
elap
hus
Meg
aloc
.
Capr
eolu
s
Sus s
crof
a
Ges
amt
MIZ % MIZ % MIZ % MIZ % MIZ % MIZ % MIZ % MIZ % MIZ % MIZ Altamira 1 2,4 8 19,5 5 12,2 2 4,9 2 4,9 20 48,8 1 2,4 2 4,9 41Hornos de la Peña 4 2 2 6 6 1 21La Pasiega 1 3 2 1 12 1 20Santimamiñe 1 2 1 3 1 7 1 16Bolinkoba 2 2 11 3 1 19Bolinkoba 1 3 16 2 2 24Ermittia 2 1 8 4 3 1 1 20Aitzbitarte 1 3 2 1 6 9 1 23Caldas 3 2 6 11Oscura 3 2 7 12Cova Rosa 2 1 1 3 5 12Cierro 3 10,7 2 7,1 1 3,6 21 75,0 1 3,6 28Balmori 1 1 3 5Coberizas 1 1 1 2 1 1 7Cueto de la Mina F 1 1 3 5Cueto de la Mina E 1 3,2 3 9,7 4 12,9 3 9,7 1 3,2 17 54,8 1 3,2 1 3,2 31Amalda, IV 1 2,5 2 5,0 1 2,5 9 22,5 16 40,0 8 20,0 1 2,5 1 2,5 1 2,5 40Morin 2 1 2 5 3 13Buxú 5 14,7 17 50,0 12 35,3 34Straus 1992, 265–267, Appendix C2/C3.
Kantabrische Fundstellen des Magdalénien, Asturias, Cantabria, Vizcaya und Guipúzcoa, Spanien (MIZ) Fundstelle und Schicht
Rang
ifer
tara
ndus
Equu
s fer
us
Bovi
nae
Capr
a py
rena
ica
Rupi
capr
a ru
pica
pra
Cer
vus
elap
hus
Capr
eolu
s ca
preo
lus
Sus s
crof
a
Ges
amt
MIZ % MIZ % MIZ % MIZ % MIZ % MIZ % MIZ % MIZ % MIZLa Paloma, 8 1 0,8 5 4,0 1 0,8 3 2,4 5 4,0 109 87,2 1 0,8 125La Paloma, 6 2 5,9 1 2,9 2 5,9 1 2,9 27 79,4 1 2,9 34La Paloma, 4 1 2,6 1 2,6 3 7,9 3 7,9 30 78,9 38Cueto Mina, D 4 16,0 3 12,0 1 4,0 17 68,0 25Cueto Mina, B 1 5,3 1 5,3 2 10,5 5 26,3 1 5,3 8 42,1 1 5,3 19Sofoxó, Magdalénien 1 4,2 3 12,5 5 20,8 4 16,7 6 25,0 4 16,7 1 4,2 24Altamira, Magdalénien 1924 4 12,5 4 12,5 1 3,1 2 6,3 19 59,4 2 6,3 32Altamira,Magdalénien 1981 1 5,0 2 10,0 14 70,0 3 15,0 20El Pendo, II 1 4,8 1 4,8 2 9,5 1 4,8 15 71,4 1 4,8 21El Pendo, IIa+b 1 6,3 1 6,3 1 6,3 11 68,8 1 6,3 1 6,3 16El Pendo, IIc+d 1 7,7 1 7,7 2 15,4 9 69,2 13El Juyo, 8-9 3 11,1 1 3,7 1 3,7 17 63,0 5 18,5 27El Juyo, 6-7 1 1,1 2 2,2 4 4,4 78 86,7 4 4,4 1 1,1 90El Juyo, 4 3 5,2 4 6,9 2 3,4 44 75,9 4 6,9 1 1,7 58Morín 2 1 4,2 3 12,5 1 4,2 3 12,5 3 12,5 9 37,5 2 8,3 2 8,3 24Ermittia Magdalénien 2 8,0 1 4,0 1 4,0 13 52,0 2 8,0 3 12,0 1 4,0 2 8,0 25Urtiaga, F 2 4,5 1 2,3 2 4,5 9 20,5 4 9,1 17 38,6 9 20,5 44Urtiaga, E 3 12,5 3 12,5 3 12,5 3 12,5 3 12,5 6 25,0 3 12,5 24Urtiaga, D 7 7,0 2 2,0 3 3,0 20 20,0 15 15,0 37 37,0 13 13,0 3 3,0 100Aitzbitarte 2 2 6,1 2 6,1 4 12,1 2 6,1 8 24,2 10 30,3 3 9,1 2 6,1 33Eralla, V 1 3,0 1 3,0 1 3,0 22 66,7 2 6,1 5 15,2 1 3,0 33Eralla, III-II 1 4,8 1 4,8 1 4,8 8 38,1 4 19,0 4 19,0 2 9,5 21Ekain, VII 3 6,8 3 6,8 7 15,9 4 9,1 24 54,5 3 6,8 44Ekain, VII 1 5,0 2 10,0 8 40,0 1 5,0 6 30,0 2 10,0 20Abauntz E 2 10,5 3 15,8 2 10,5 2 10,5 8 42,1 2 10,5 19Straus 1992, 281–283, Appendix D6
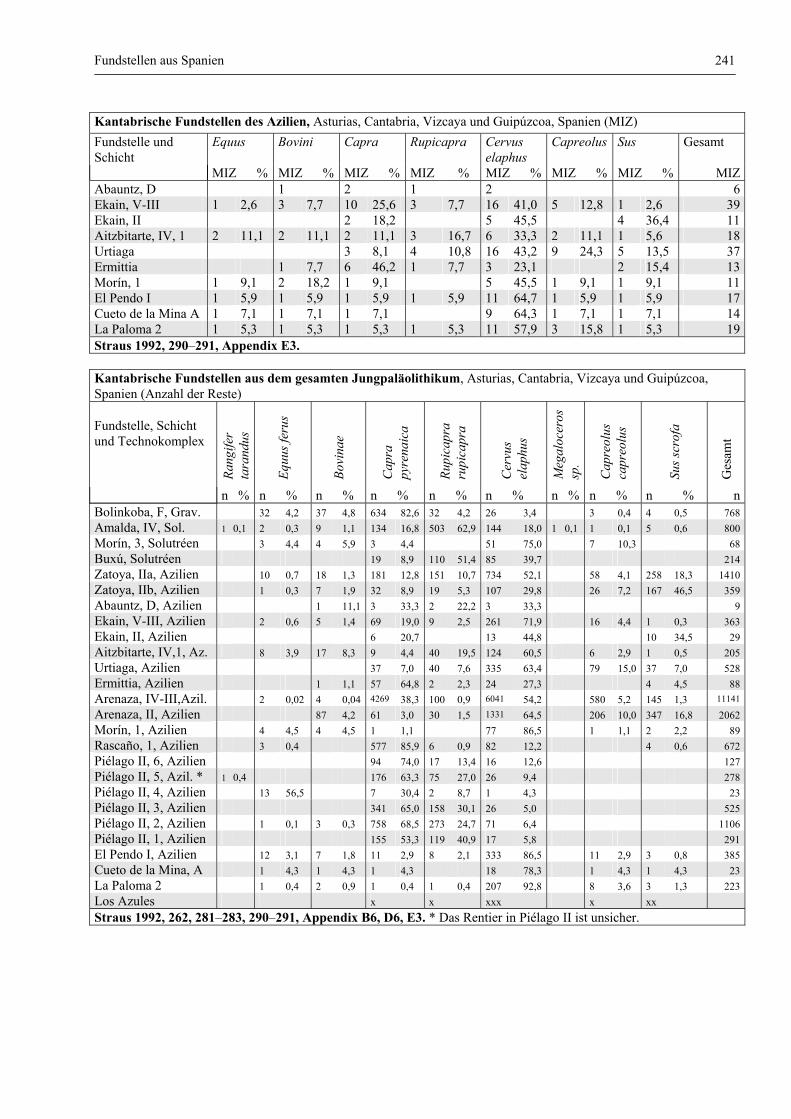
Fundstellen aus Spanien
241
Kantabrische Fundstellen des Azilien, Asturias, Cantabria, Vizcaya und Guipúzcoa, Spanien (MIZ) Fundstelle und Schicht
Equus Bovini Capra Rupicapra Cervus elaphus
Capreolus Sus Gesamt
MIZ % MIZ % MIZ % MIZ % MIZ % MIZ % MIZ % MIZAbauntz, D 1 2 1 2 6Ekain, V-III 1 2,6 3 7,7 10 25,6 3 7,7 16 41,0 5 12,8 1 2,6 39Ekain, II 2 18,2 5 45,5 4 36,4 11Aitzbitarte, IV, 1 2 11,1 2 11,1 2 11,1 3 16,7 6 33,3 2 11,1 1 5,6 18Urtiaga 3 8,1 4 10,8 16 43,2 9 24,3 5 13,5 37Ermittia 1 7,7 6 46,2 1 7,7 3 23,1 2 15,4 13Morín, 1 1 9,1 2 18,2 1 9,1 5 45,5 1 9,1 1 9,1 11El Pendo I 1 5,9 1 5,9 1 5,9 1 5,9 11 64,7 1 5,9 1 5,9 17Cueto de la Mina A 1 7,1 1 7,1 1 7,1 9 64,3 1 7,1 1 7,1 14La Paloma 2 1 5,3 1 5,3 1 5,3 1 5,3 11 57,9 3 15,8 1 5,3 19Straus 1992, 290–291, Appendix E3. Kantabrische Fundstellen aus dem gesamten Jungpaläolithikum, Asturias, Cantabria, Vizcaya und Guipúzcoa, Spanien (Anzahl der Reste) Fundstelle, Schicht und Technokomplex
Rang
ifer
tara
ndus
Equu
s fer
us
Bovi
nae
Capr
a py
rena
ica
Rupi
capr
a ru
pica
pra
Cer
vus
elap
hus
Meg
aloc
eros
sp
.
Capr
eolu
s ca
preo
lus
Sus s
crof
a
Ges
amt
n % n % n % n % n % n % n % n % n % nBolinkoba, F, Grav. 32 4,2 37 4,8 634 82,6 32 4,2 26 3,4 3 0,4 4 0,5 768Amalda, IV, Sol. 1 0,1 2 0,3 9 1,1 134 16,8 503 62,9 144 18,0 1 0,1 1 0,1 5 0,6 800Morín, 3, Solutréen 3 4,4 4 5,9 3 4,4 51 75,0 7 10,3 68Buxú, Solutréen 19 8,9 110 51,4 85 39,7 214Zatoya, IIa, Azilien 10 0,7 18 1,3 181 12,8 151 10,7 734 52,1 58 4,1 258 18,3 1410Zatoya, IIb, Azilien 1 0,3 7 1,9 32 8,9 19 5,3 107 29,8 26 7,2 167 46,5 359Abauntz, D, Azilien 1 11,1 3 33,3 2 22,2 3 33,3 9Ekain, V-III, Azilien 2 0,6 5 1,4 69 19,0 9 2,5 261 71,9 16 4,4 1 0,3 363Ekain, II, Azilien 6 20,7 13 44,8 10 34,5 29Aitzbitarte, IV,1, Az. 8 3,9 17 8,3 9 4,4 40 19,5 124 60,5 6 2,9 1 0,5 205Urtiaga, Azilien 37 7,0 40 7,6 335 63,4 79 15,0 37 7,0 528Ermittia, Azilien 1 1,1 57 64,8 2 2,3 24 27,3 4 4,5 88Arenaza, IV-III,Azil. 2 0,02 4 0,04 4269 38,3 100 0,9 6041 54,2 580 5,2 145 1,3 11141
Arenaza, II, Azilien 87 4,2 61 3,0 30 1,5 1331 64,5 206 10,0 347 16,8 2062Morín, 1, Azilien 4 4,5 4 4,5 1 1,1 77 86,5 1 1,1 2 2,2 89Rascaño, 1, Azilien 3 0,4 577 85,9 6 0,9 82 12,2 4 0,6 672Piélago II, 6, Azilien 94 74,0 17 13,4 16 12,6 127Piélago II, 5, Azil. * 1 0,4 176 63,3 75 27,0 26 9,4 278Piélago II, 4, Azilien 13 56,5 7 30,4 2 8,7 1 4,3 23Piélago II, 3, Azilien 341 65,0 158 30,1 26 5,0 525Piélago II, 2, Azilien 1 0,1 3 0,3 758 68,5 273 24,7 71 6,4 1106Piélago II, 1, Azilien 155 53,3 119 40,9 17 5,8 291El Pendo I, Azilien 12 3,1 7 1,8 11 2,9 8 2,1 333 86,5 11 2,9 3 0,8 385Cueto de la Mina, A 1 4,3 1 4,3 1 4,3 18 78,3 1 4,3 1 4,3 23La Paloma 2 1 0,4 2 0,9 1 0,4 1 0,4 207 92,8 8 3,6 3 1,3 223Los Azules x x xxx x xx Straus 1992, 262, 281–283, 290–291, Appendix B6, D6, E3. * Das Rentier in Piélago II ist unsicher.
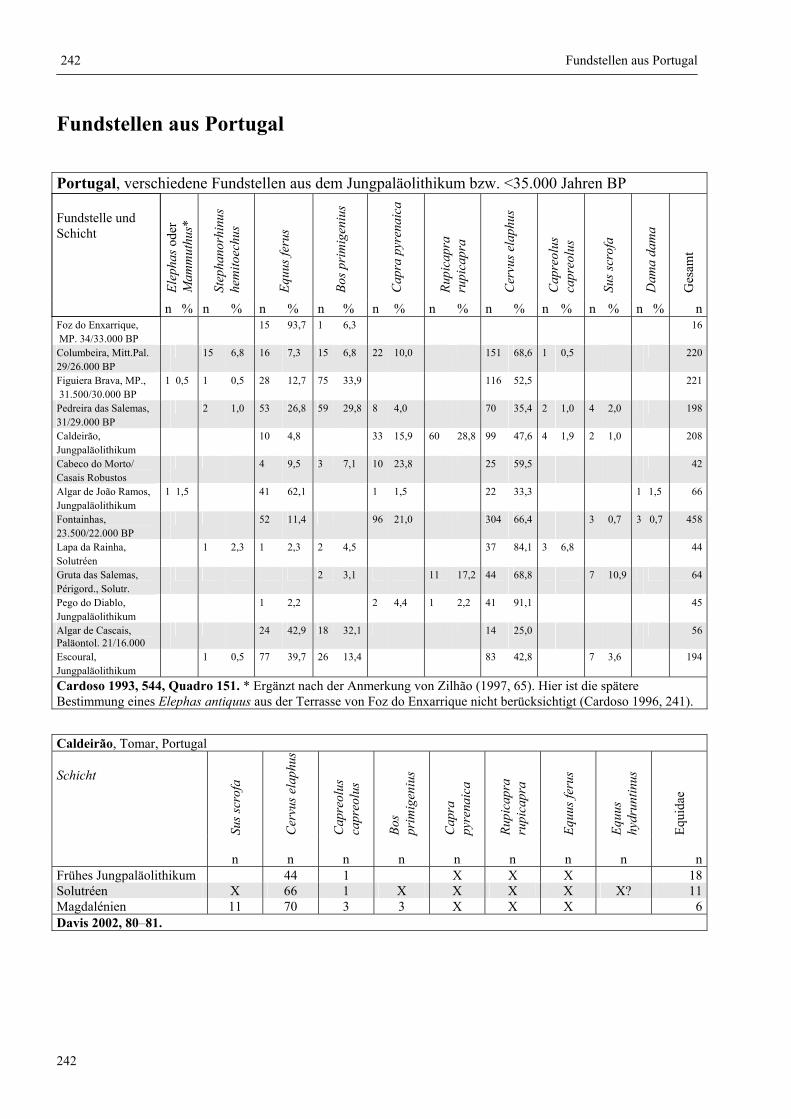
Fundstellen aus Portugal 242
Fundstellen aus Portugal Portugal, verschiedene Fundstellen aus dem Jungpaläolithikum bzw. <35.000 Jahren BP Fundstelle und Schicht
Elep
has o
der
Mam
mut
hus*
Step
hano
rhin
us
hem
itoec
hus
Equu
s fer
us
Bos p
rim
igen
ius
Capr
a py
rena
ica
Rupi
capr
a ru
pica
pra
Cerv
us e
laph
us
Capr
eolu
s ca
preo
lus
Sus s
crof
a
Dam
a da
ma
Ges
amt
n % n % n % n % n % n % n % n % n % n % nFoz do Enxarrique, MP. 34/33.000 BP
15 93,7 1 6,3 16
Columbeira, Mitt.Pal. 29/26.000 BP
15 6,8 16 7,3 15 6,8 22 10,0 151 68,6 1 0,5 220
Figuiera Brava, MP., 31.500/30.000 BP
1 0,5 1 0,5 28 12,7 75 33,9 116 52,5 221
Pedreira das Salemas, 31/29.000 BP
2 1,0 53 26,8 59 29,8 8 4,0 70 35,4 2 1,0 4 2,0 198
Caldeirão, Jungpaläolithikum
10 4,8 33 15,9 60 28,8 99 47,6 4 1,9 2 1,0 208
Cabeco do Morto/ Casais Robustos
4 9,5 3 7,1 10 23,8 25 59,5 42
Algar de João Ramos, Jungpaläolithikum
1 1,5 41 62,1 1 1,5 22 33,3 1 1,5 66
Fontainhas, 23.500/22.000 BP
52 11,4 96 21,0 304 66,4 3 0,7 3 0,7 458
Lapa da Rainha, Solutréen
1 2,3 1 2,3 2 4,5 37 84,1 3 6,8 44
Gruta das Salemas, Périgord., Solutr.
2 3,1 11 17,2 44 68,8 7 10,9 64
Pego do Diablo, Jungpaläolithikum
1 2,2 2 4,4 1 2,2 41 91,1 45
Algar de Cascais, Paläontol. 21/16.000
24 42,9 18 32,1 14 25,0 56
Escoural, Jungpaläolithikum
1 0,5 77 39,7 26 13,4 83 42,8 7 3,6 194
Cardoso 1993, 544, Quadro 151. * Ergänzt nach der Anmerkung von Zilhão (1997, 65). Hier ist die spätere Bestimmung eines Elephas antiquus aus der Terrasse von Foz do Enxarrique nicht berücksichtigt (Cardoso 1996, 241). Caldeirão, Tomar, Portugal Schicht
Sus s
crof
a
Cerv
us e
laph
us
Capr
eolu
s ca
preo
lus
Bos
prim
igen
ius
Capr
a py
rena
ica
Rupi
capr
a ru
pica
pra
Equu
s fer
us
Equu
s hy
drun
tinus
Equi
dae
n
n
n
n
n
n
n
n n
Frühes Jungpaläolithikum 44 1 X X X 18Solutréen X 66 1 X X X X X? 11Magdalénien 11 70 3 3 X X X 6Davis 2002, 80–81.
242
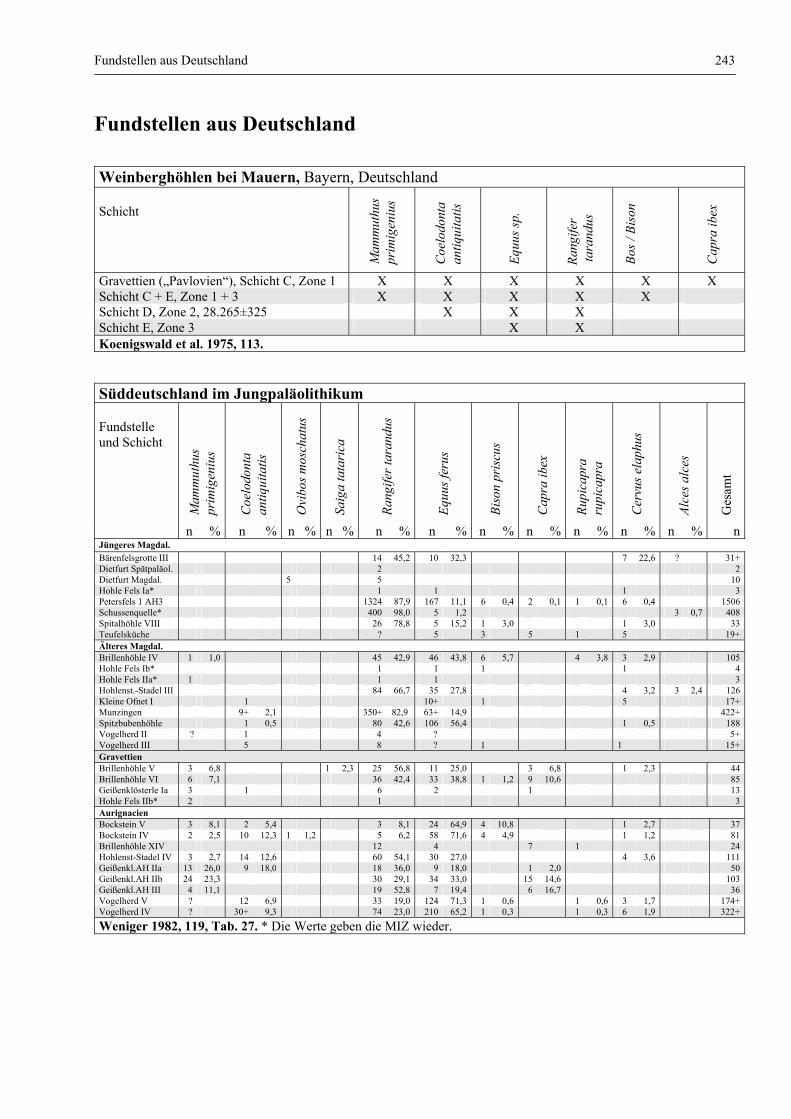
Fundstellen aus Deutschland 243
Fundstellen aus Deutschland Weinberghöhlen bei Mauern, Bayern, Deutschland Schicht
Mam
mut
hus
prim
igen
ius
Coel
odon
ta
antiq
uita
tis
Equu
s sp.
Ran
gife
r
tara
ndus
Bos /
Bis
on
Capr
a ib
ex
Gravettien („Pavlovien“), Schicht C, Zone 1 X X X X X X Schicht C + E, Zone 1 + 3 X X X X X Schicht D, Zone 2, 28.265±325 X X X Schicht E, Zone 3 X X Koenigswald et al. 1975, 113.
Süddeutschland im Jungpaläolithikum Fundstelle und Schicht
Mam
mut
hus
prim
igen
ius
Coel
odon
ta
antiq
uita
tis
Ovi
bos m
osch
atus
Saig
a ta
tari
ca
Rang
ifer t
aran
dus
Equu
s fer
us
Biso
n pr
iscus
Capr
a ib
ex
Rupi
capr
a ru
pica
pra
Cerv
us e
laph
us
Alce
s alc
es
Ges
amt
n % n % n % n % n % n % n % n % n % n % n % n Jüngeres Magdal. Bärenfelsgrotte III 14 45,2 10 32,3 7 22,6 ? 31+Dietfurt Spätpaläol. 2 2Dietfurt Magdal. 5 5 10Hohle Fels Ia* 1 1 1 3Petersfels 1 AH3 1324 87,9 167 11,1 6 0,4 2 0,1 1 0,1 6 0,4 1506Schussenquelle* 400 98,0 5 1,2 3 0,7 408Spitalhöhle VIII 26 78,8 5 15,2 1 3,0 1 3,0 33Teufelsküche ? 5 3 5 1 5 19+Älteres Magdal. Brillenhöhle IV 1 1,0 45 42,9 46 43,8 6 5,7 4 3,8 3 2,9 105Hohle Fels Ib* 1 1 1 1 4Hohle Fels IIa* 1 1 1 3Hohlenst.-Stadel III 84 66,7 35 27,8 4 3,2 3 2,4 126Kleine Ofnet I 1 10+ 1 5 17+Munzingen 9+ 2,1 350+ 82,9 63+ 14,9 422+Spitzbubenhöhle 1 0,5 80 42,6 106 56,4 1 0,5 188Vogelherd II ? 1 4 ? 5+Vogelherd III 5 8 ? 1 1 15+Gravettien Brillenhöhle V 3 6,8 1 2,3 25 56,8 11 25,0 3 6,8 1 2,3 44Brillenhöhle VI 6 7,1 36 42,4 33 38,8 1 1,2 9 10,6 85Geißenklösterle Ia 3 1 6 2 1 13Hohle Fels IIb* 2 1 3Aurignacien Bockstein V 3 8,1 2 5,4 3 8,1 24 64,9 4 10,8 1 2,7 37Bockstein IV 2 2,5 10 12,3 1 1,2 5 6,2 58 71,6 4 4,9 1 1,2 81Brillenhöhle XIV 12 4 7 1 24Hohlenst-Stadel IV 3 2,7 14 12,6 60 54,1 30 27,0 4 3,6 111Geißenkl.AH IIa 13 26,0 9 18,0 18 36,0 9 18,0 1 2,0 50Geißenkl.AH IIb 24 23,3 30 29,1 34 33,0 15 14,6 103Geißenkl.AH III 4 11,1 19 52,8 7 19,4 6 16,7 36Vogelherd V ? 12 6,9 33 19,0 124 71,3 1 0,6 1 0,6 3 1,7 174+Vogelherd IV ? 30+ 9,3 74 23,0 210 65,2 1 0,3 1 0,3 6 1,9 322+
Weniger 1982, 119, Tab. 27. * Die Werte geben die MIZ wieder.
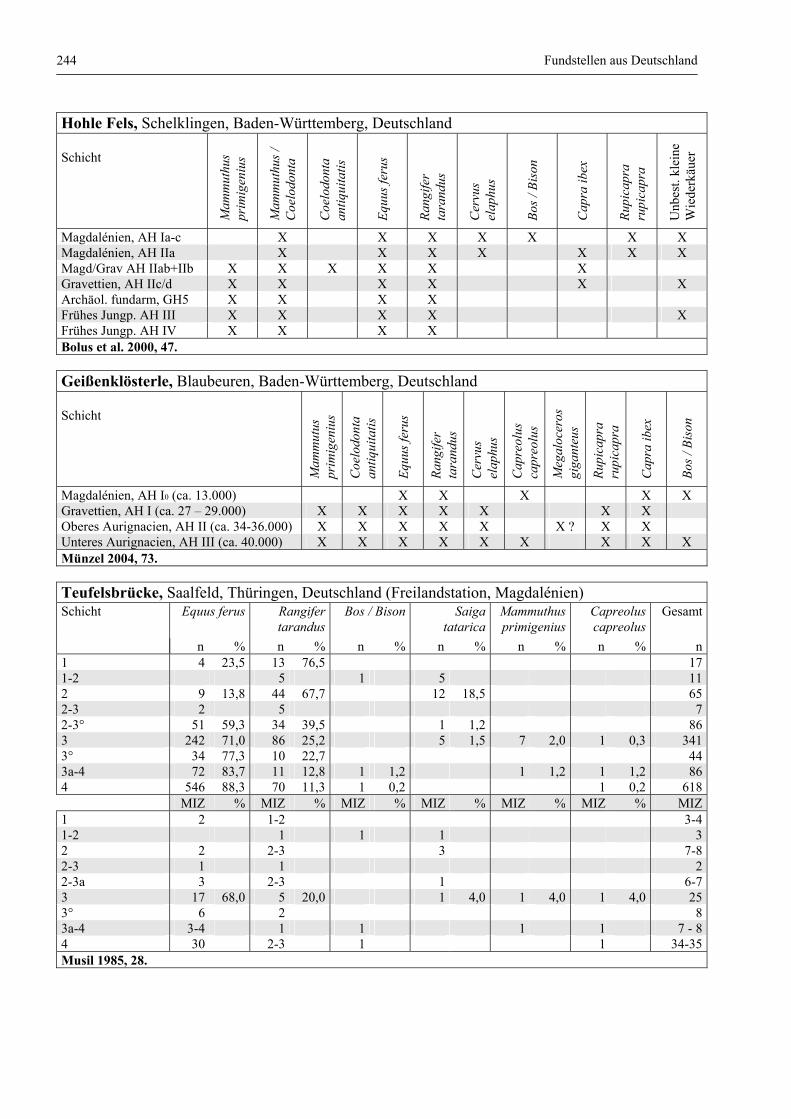
Fundstellen aus Deutschland 244
Hohle Fels, Schelklingen, Baden-Württemberg, Deutschland Schicht
Mam
mut
hus
prim
igen
ius
Mam
mut
hus /
Co
elod
onta
Coel
odon
ta
antiq
uita
tis
Equu
s fer
us
Rang
ifer
tara
ndus
Cer
vus
elap
hus
Bos /
Bis
on
Capr
a ib
ex
Rupi
capr
a ru
pica
pra
Unb
est.
klei
ne
Wie
derk
äuer
Magdalénien, AH Ia-c X X X X X X X Magdalénien, AH IIa X X X X X X X Magd/Grav AH IIab+IIb X X X X X X Gravettien, AH IIc/d X X X X X X Archäol. fundarm, GH5 X X X X Frühes Jungp. AH III X X X X X Frühes Jungp. AH IV X X X X Bolus et al. 2000, 47.
Geißenklösterle, Blaubeuren, Baden-Württemberg, Deutschland Schicht
Mam
mut
us
prim
igen
ius
Coel
odon
ta
antiq
uita
tis
Equu
s fer
us
Rang
ifer
tara
ndus
Cer
vus
elap
hus
Capr
eolu
s ca
preo
lus
Meg
aloc
eros
gi
gant
eus
Rupi
capr
a ru
pica
pra
Capr
a ib
ex
Bos /
Bis
on
Magdalénien, AH I0 (ca. 13.000) X X X X X Gravettien, AH I (ca. 27 – 29.000) X X X X X X X Oberes Aurignacien, AH II (ca. 34-36.000) X X X X X X ? X X Unteres Aurignacien, AH III (ca. 40.000) X X X X X X X X X Münzel 2004, 73.
Teufelsbrücke, Saalfeld, Thüringen, Deutschland (Freilandstation, Magdalénien) Schicht Equus ferus Rangifer
tarandus Bos / Bison Saiga
tataricaMammuthus primigenius
Capreolus capreolus
Gesamt
n % n % n % n % n % n % n1 4 23,5 13 76,5 171-2 5 1 5 112 9 13,8 44 67,7 12 18,5 652-3 2 5 72-3° 51 59,3 34 39,5 1 1,2 863 242 71,0 86 25,2 5 1,5 7 2,0 1 0,3 3413° 34 77,3 10 22,7 443a-4 72 83,7 11 12,8 1 1,2 1 1,2 1 1,2 864 546 88,3 70 11,3 1 0,2 1 0,2 618
MIZ % MIZ % MIZ % MIZ % MIZ % MIZ % MIZ1 2 1-2 3-41-2 1 1 1 32 2 2-3 3 7-82-3 1 1 22-3a 3 2-3 1 6-73 17 68,0 5 20,0 1 4,0 1 4,0 1 4,0 253° 6 2 83a-4 3-4 1 1 1 1 7 - 84 30 2-3 1 1 34-35Musil 1985, 28.
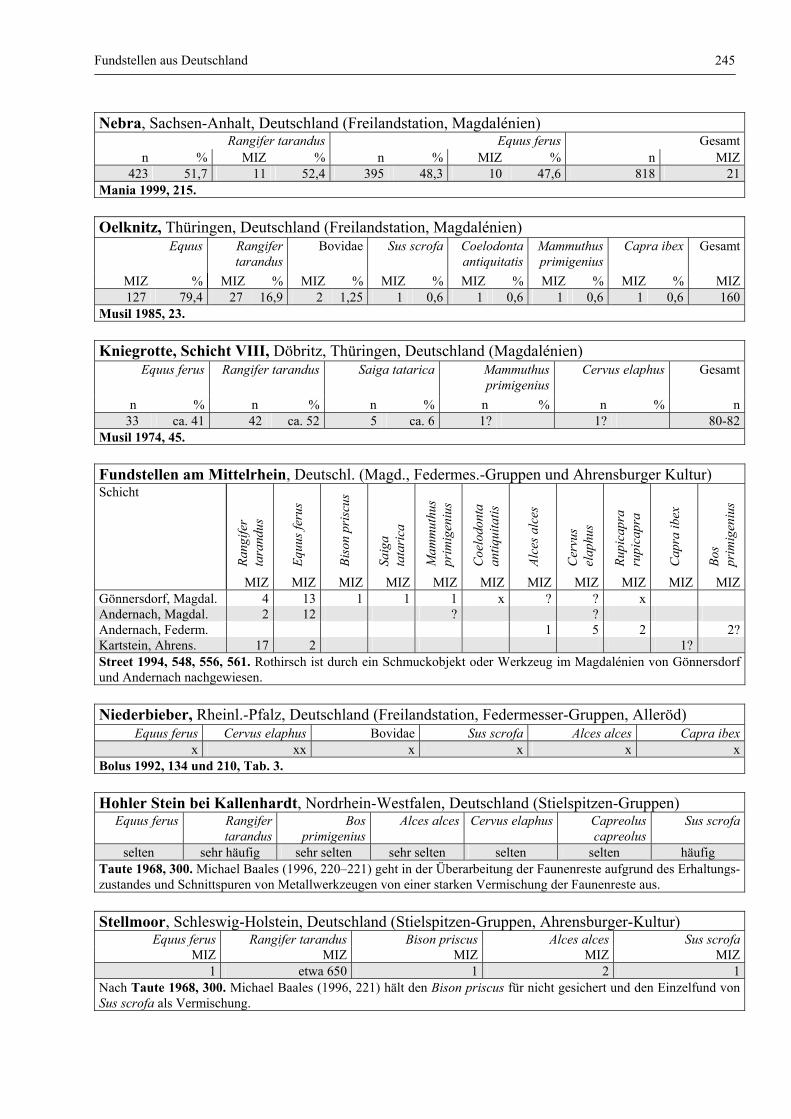
Fundstellen aus Deutschland
245
Nebra, Sachsen-Anhalt, Deutschland (Freilandstation, Magdalénien) Rangifer tarandus Equus ferus Gesamt
n % MIZ % n % MIZ % n MIZ423 51,7 11 52,4 395 48,3 10 47,6 818 21
Mania 1999, 215. Oelknitz, Thüringen, Deutschland (Freilandstation, Magdalénien)
Equus Rangifer tarandus
Bovidae Sus scrofa Coelodonta antiquitatis
Mammuthus primigenius
Capra ibex Gesamt
MIZ % MIZ % MIZ % MIZ % MIZ % MIZ % MIZ % MIZ127 79,4 27 16,9 2 1,25 1 0,6 1 0,6 1 0,6 1 0,6 160
Musil 1985, 23. Kniegrotte, Schicht VIII, Döbritz, Thüringen, Deutschland (Magdalénien)
Equus ferus Rangifer tarandus Saiga tatarica Mammuthus primigenius
Cervus elaphus Gesamt
n % n % n % n % n % n 33 ca. 41 42 ca. 52 5 ca. 6 1? 1? 80-82Musil 1974, 45.
Fundstellen am Mittelrhein, Deutschl. (Magd., Federmes.-Gruppen und Ahrensburger Kultur) Schicht
Rang
ifer
tara
ndus
Equu
s fer
us
Biso
n pr
iscus
Saig
a ta
taric
a
Mam
mut
hus
prim
igen
ius
Coel
odon
ta
antiq
uita
tis
Alce
s alc
es
Cer
vus
elap
hus
Rupi
capr
a ru
pica
pra
Capr
a ib
ex
Bos
prim
igen
ius
MIZ MIZ MIZ MIZ MIZ MIZ MIZ MIZ MIZ MIZ MIZGönnersdorf, Magdal. 4 13 1 1 1 x ? ? x Andernach, Magdal. 2 12 ? ? Andernach, Federm. 1 5 2 2?Kartstein, Ahrens. 17 2 1?Street 1994, 548, 556, 561. Rothirsch ist durch ein Schmuckobjekt oder Werkzeug im Magdalénien von Gönnersdorf und Andernach nachgewiesen.
Niederbieber, Rheinl.-Pfalz, Deutschland (Freilandstation, Federmesser-Gruppen, Alleröd)
Equus ferus Cervus elaphus Bovidae Sus scrofa Alces alces Capra ibex x xx x x x x
Bolus 1992, 134 und 210, Tab. 3. Hohler Stein bei Kallenhardt, Nordrhein-Westfalen, Deutschland (Stielspitzen-Gruppen)
Equus ferus Rangifer tarandus
Bos primigenius
Alces alces Cervus elaphus Capreolus capreolus
Sus scrofa
selten sehr häufig sehr selten sehr selten selten selten häufig Taute 1968, 300. Michael Baales (1996, 220–221) geht in der Überarbeitung der Faunenreste aufgrund des Erhaltungs-zustandes und Schnittspuren von Metallwerkzeugen von einer starken Vermischung der Faunenreste aus.
Stellmoor, Schleswig-Holstein, Deutschland (Stielspitzen-Gruppen, Ahrensburger-Kultur)
Equus ferus Rangifer tarandus Bison priscus Alces alces Sus scrofaMIZ MIZ MIZ MIZ MIZ
1 etwa 650 1 2 1Nach Taute 1968, 300. Michael Baales (1996, 221) hält den Bison priscus für nicht gesichert und den Einzelfund von Sus scrofa als Vermischung.
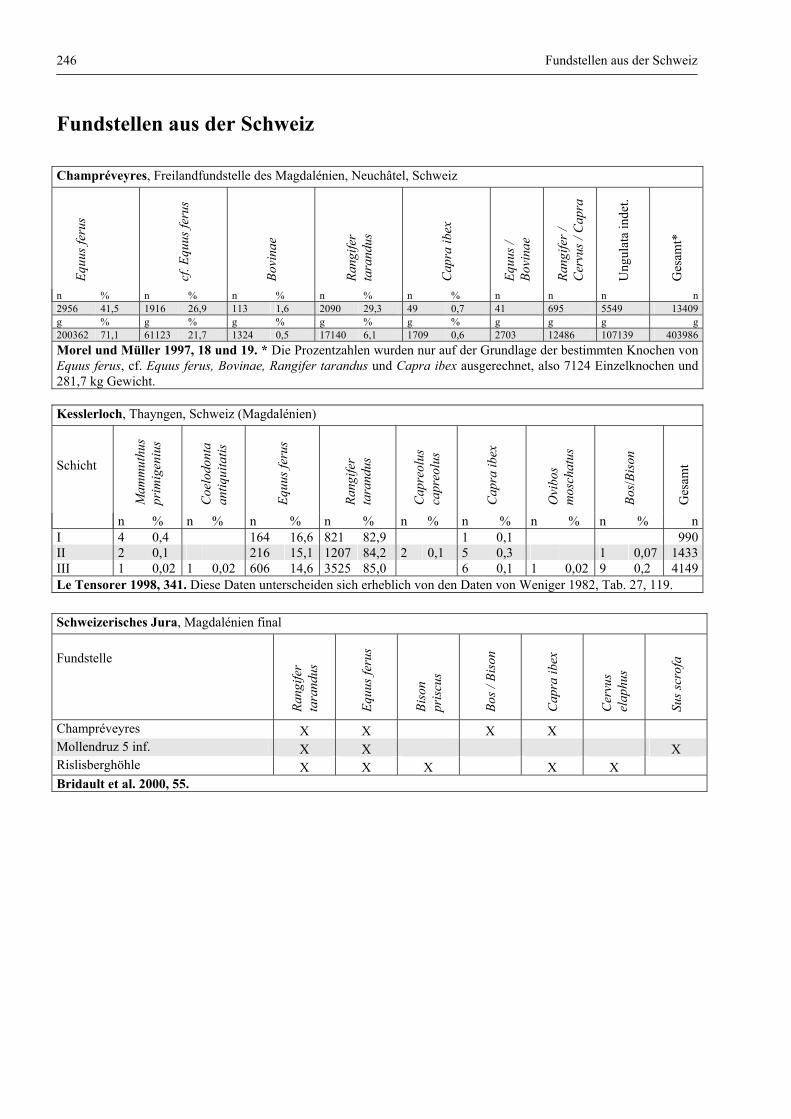
Fundstellen aus der Schweiz 246
Fundstellen aus der Schweiz Champréveyres, Freilandfundstelle des Magdalénien, Neuchâtel, Schweiz
Equu
s fer
us
cf. E
quus
feru
s
Bovi
nae
Rang
ifer
tara
ndus
Capr
a ib
ex
Equu
s /
Bovi
nae
Rang
ifer /
Ce
rvus
/ Ca
pra
Ung
ulat
a in
det.
Ges
amt*
n % n % n % n % n % n n n n2956 41,5 1916 26,9 113 1,6 2090 29,3 49 0,7 41 695 5549 13409g % g % g % g % g % g g g g200362 71,1 61123 21,7 1324 0,5 17140 6,1 1709 0,6 2703 12486 107139 403986Morel und Müller 1997, 18 und 19. * Die Prozentzahlen wurden nur auf der Grundlage der bestimmten Knochen von Equus ferus, cf. Equus ferus, Bovinae, Rangifer tarandus und Capra ibex ausgerechnet, also 7124 Einzelknochen und 281,7 kg Gewicht. Kesslerloch, Thayngen, Schweiz (Magdalénien) Schicht
Mam
mut
hus
prim
igen
ius
Coel
odon
ta
antiq
uita
tis
Equu
s fer
us
Rang
ifer
tara
ndus
Capr
eolu
s ca
preo
lus
Capr
a ib
ex
Ovi
bos
mos
chat
us
Bos/B
ison
Ges
amt
n % n % n % n % n % n % n % n % nI 4 0,4 164 16,6 821 82,9 1 0,1 990II 2 0,1 216 15,1 1207 84,2 2 0,1 5 0,3 1 0,07 1433III 1 0,02 1 0,02 606 14,6 3525 85,0 6 0,1 1 0,02 9 0,2 4149Le Tensorer 1998, 341. Diese Daten unterscheiden sich erheblich von den Daten von Weniger 1982, Tab. 27, 119.
Schweizerisches Jura, Magdalénien final Fundstelle
Rang
ifer
tara
ndus
Equu
s fer
us
Biso
n pr
iscus
Bos /
Bis
on
Capr
a ib
ex
Cer
vus
elap
hus
Sus s
crof
a
Champréveyres X X X X Mollendruz 5 inf. X X X Rislisberghöhle X X X X X Bridault et al. 2000, 55.
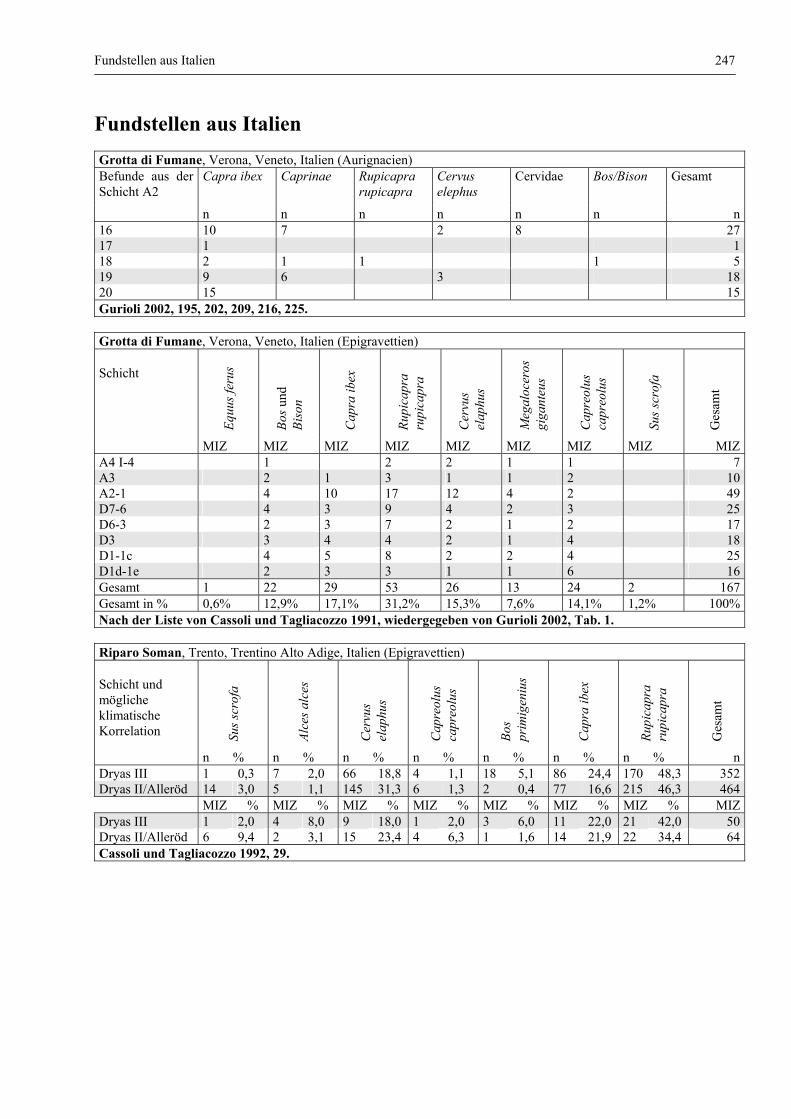
Fundstellen aus Italien 247
Fundstellen aus Italien Grotta di Fumane, Verona, Veneto, Italien (Aurignacien) Befunde aus der Schicht A2
Capra ibex Caprinae Rupicapra rupicapra
Cervus elephus
Cervidae Bos/Bison Gesamt
n n n n n n n16 10 7 2 8 2717 1 118 2 1 1 1 519 9 6 3 1820 15 15Gurioli 2002, 195, 202, 209, 216, 225.
Grotta di Fumane, Verona, Veneto, Italien (Epigravettien) Schicht
Equu
s fer
us
Bos u
nd
Biso
n
Capr
a ib
ex
Rupi
capr
a ru
pica
pra
Cer
vus
elap
hus
Meg
aloc
eros
gi
gant
eus
Capr
eolu
s ca
preo
lus
Sus s
crof
a
Ges
amt
MIZ MIZ MIZ MIZ MIZ MIZ MIZ MIZ MIZA4 I-4 1 2 2 1 1 7A3 2 1 3 1 1 2 10A2-1 4 10 17 12 4 2 49D7-6 4 3 9 4 2 3 25D6-3 2 3 7 2 1 2 17D3 3 4 4 2 1 4 18D1-1c 4 5 8 2 2 4 25D1d-1e 2 3 3 1 1 6 16Gesamt 1 22 29 53 26 13 24 2 167Gesamt in % 0,6% 12,9% 17,1% 31,2% 15,3% 7,6% 14,1% 1,2% 100%Nach der Liste von Cassoli und Tagliacozzo 1991, wiedergegeben von Gurioli 2002, Tab. 1.
Riparo Soman, Trento, Trentino Alto Adige, Italien (Epigravettien) Schicht und mögliche klimatische Korrelation Su
s scr
ofa
Alce
s alc
es
Cer
vus
elap
hus
Capr
eolu
s ca
preo
lus
Bos
prim
igen
ius
Capr
a ib
ex
Rupi
capr
a ru
pica
pra
Ges
amt
n % n % n % n % n % n % n % nDryas III 1 0,3 7 2,0 66 18,8 4 1,1 18 5,1 86 24,4 170 48,3 352Dryas II/Alleröd 14 3,0 5 1,1 145 31,3 6 1,3 2 0,4 77 16,6 215 46,3 464 MIZ % MIZ % MIZ % MIZ % MIZ % MIZ % MIZ % MIZDryas III 1 2,0 4 8,0 9 18,0 1 2,0 3 6,0 11 22,0 21 42,0 50Dryas II/Alleröd 6 9,4 2 3,1 15 23,4 4 6,3 1 1,6 14 21,9 22 34,4 64Cassoli und Tagliacozzo 1992, 29.
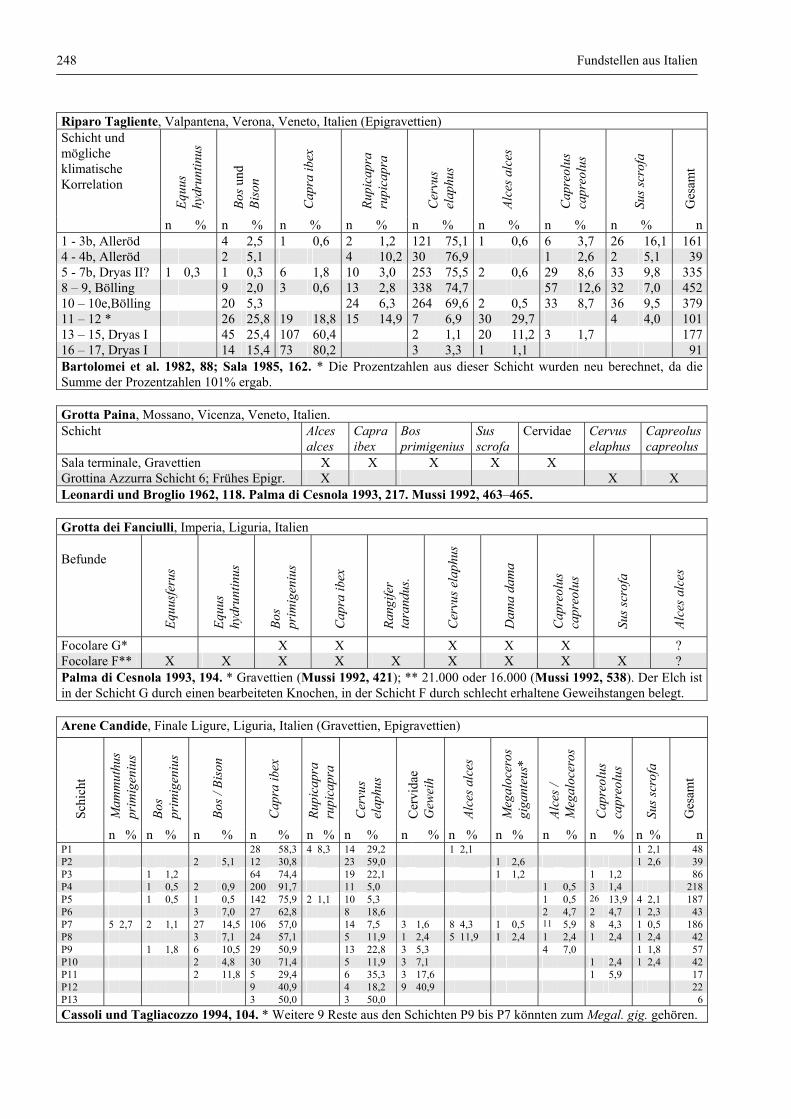
Fundstellen aus Italien 248
Riparo Tagliente, Valpantena, Verona, Veneto, Italien (Epigravettien) Schicht und mögliche klimatische Korrelation
Equu
s hy
drun
tinus
Bos u
nd
Biso
n
Capr
a ib
ex
Rupi
capr
a ru
pica
pra
Cer
vus
elap
hus
Alce
s alc
es
Capr
eolu
s ca
preo
lus
Sus s
crof
a
Ges
amt
n % n % n % n % n % n % n % n % n1 - 3b, Alleröd 4 2,5 1 0,6 2 1,2 121 75,1 1 0,6 6 3,7 26 16,1 1614 - 4b, Alleröd 2 5,1 4 10,2 30 76,9 1 2,6 2 5,1 395 - 7b, Dryas II? 1 0,3 1 0,3 6 1,8 10 3,0 253 75,5 2 0,6 29 8,6 33 9,8 3358 – 9, Bölling 9 2,0 3 0,6 13 2,8 338 74,7 57 12,6 32 7,0 45210 – 10e,Bölling 20 5,3 24 6,3 264 69,6 2 0,5 33 8,7 36 9,5 37911 – 12 * 26 25,8 19 18,8 15 14,9 7 6,9 30 29,7 4 4,0 10113 – 15, Dryas I 45 25,4 107 60,4 2 1,1 20 11,2 3 1,7 17716 – 17, Dryas I 14 15,4 73 80,2 3 3,3 1 1,1 91Bartolomei et al. 1982, 88; Sala 1985, 162. * Die Prozentzahlen aus dieser Schicht wurden neu berechnet, da die Summe der Prozentzahlen 101% ergab.
Grotta Paina, Mossano, Vicenza, Veneto, Italien. Schicht Alces
alces Capra ibex
Bos primigenius
Sus scrofa
Cervidae Cervus elaphus
Capreolus capreolus
Sala terminale, Gravettien X X X X X Grottina Azzurra Schicht 6; Frühes Epigr. X X X Leonardi und Broglio 1962, 118. Palma di Cesnola 1993, 217. Mussi 1992, 463–465.
Grotta dei Fanciulli, Imperia, Liguria, Italien Befunde
Equu
sfer
us
Equu
s hy
drun
tinus
Bos
prim
igen
ius
Capr
a ib
ex
Rang
ifer
tara
ndus
.
Cerv
us e
laph
us
Dam
a da
ma
Capr
eolu
s ca
preo
lus
Sus s
crof
a
Alce
s alc
es
Focolare G* X X X X X ? Focolare F** X X X X X X X X X ? Palma di Cesnola 1993, 194. * Gravettien (Mussi 1992, 421); ** 21.000 oder 16.000 (Mussi 1992, 538). Der Elch ist in der Schicht G durch einen bearbeiteten Knochen, in der Schicht F durch schlecht erhaltene Geweihstangen belegt.
Arene Candide, Finale Ligure, Liguria, Italien (Gravettien, Epigravettien)
Schi
cht
Mam
mut
hus
prim
igen
ius
Bos
prim
igen
ius
Bos /
Bis
on
Capr
a ib
ex
Rupi
capr
a ru
pica
pra
Cer
vus
elap
hus
Cer
vida
e G
ewei
h
Alce
s alc
es
Meg
aloc
eros
gi
gant
eus*
Alce
s /
Meg
aloc
eros
Capr
eolu
s ca
preo
lus
Sus s
crof
a
Ges
amt
n % n % n % n % n % n % n % n % n % n % n % n % nP1 28 58,3 4 8,3 14 29,2 1 2,1 1 2,1 48P2 2 5,1 12 30,8 23 59,0 1 2,6 1 2,6 39P3 1 1,2 64 74,4 19 22,1 1 1,2 1 1,2 86P4 1 0,5 2 0,9 200 91,7 11 5,0 1 0,5 3 1,4 218P5 1 0,5 1 0,5 142 75,9 2 1,1 10 5,3 1 0,5 26 13,9 4 2,1 187P6 3 7,0 27 62,8 8 18,6 2 4,7 2 4,7 1 2,3 43P7 5 2,7 2 1,1 27 14,5 106 57,0 14 7,5 3 1,6 8 4,3 1 0,5 11 5,9 8 4,3 1 0,5 186P8 3 7,1 24 57,1 5 11,9 1 2,4 5 11,9 1 2,4 1 2,4 1 2,4 1 2,4 42P9 1 1,8 6 10,5 29 50,9 13 22,8 3 5,3 4 7,0 1 1,8 57P10 2 4,8 30 71,4 5 11,9 3 7,1 1 2,4 1 2,4 42P11 2 11,8 5 29,4 6 35,3 3 17,6 1 5,9 17P12 9 40,9 4 18,2 9 40,9 22P13 3 50,0 3 50,0 6Cassoli und Tagliacozzo 1994, 104. * Weitere 9 Reste aus den Schichten P9 bis P7 könnten zum Megal. gig. gehören.
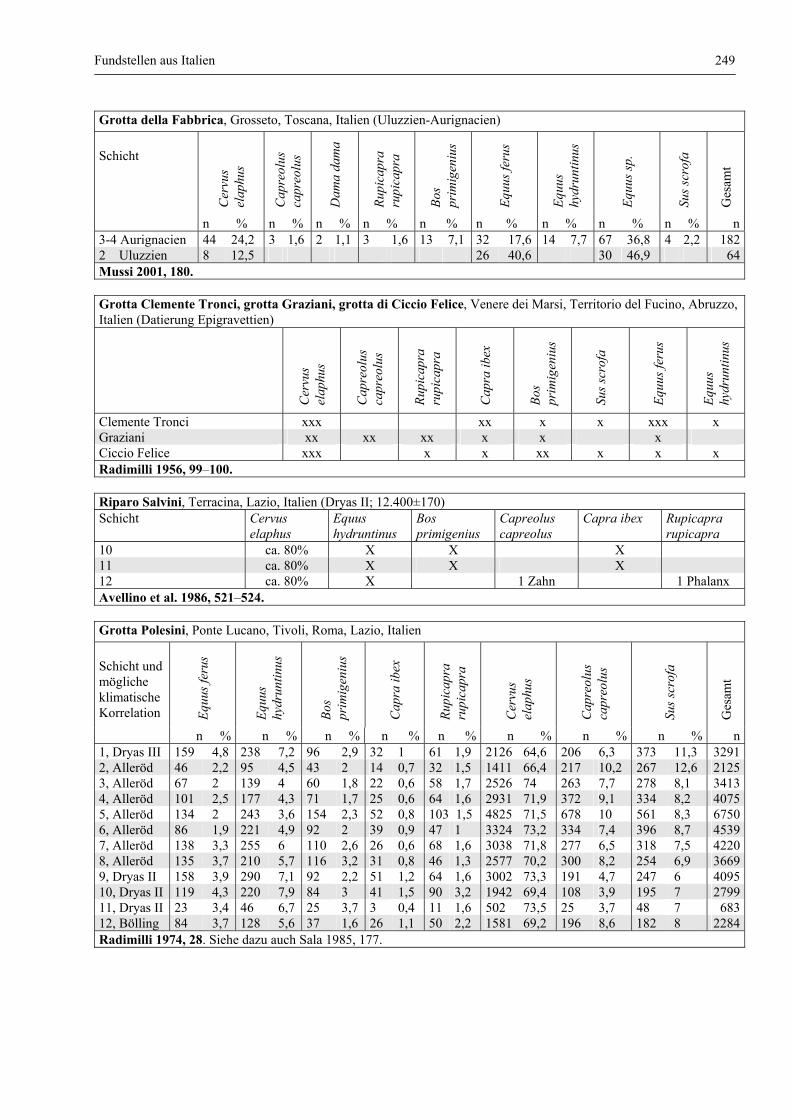
Fundstellen aus Italien 249
Grotta della Fabbrica, Grosseto, Toscana, Italien (Uluzzien-Aurignacien) Schicht
Cer
vus
elap
hus
Capr
eolu
s ca
preo
lus
Dam
a da
ma
Rupi
capr
a ru
pica
pra
Bos
prim
igen
ius
Equu
s fer
us
Equu
s hy
drun
tinus
Equu
s sp.
Sus s
crof
a
Ges
amt
n % n % n % n % n % n % n % n % n % n3-4 Aurignacien 44 24,2 3 1,6 2 1,1 3 1,6 13 7,1 32 17,6 14 7,7 67 36,8 4 2,2 1822 Uluzzien 8 12,5 26 40,6 30 46,9 64Mussi 2001, 180.
Grotta Clemente Tronci, grotta Graziani, grotta di Ciccio Felice, Venere dei Marsi, Territorio del Fucino, Abruzzo, Italien (Datierung Epigravettien)
Cer
vus
elap
hus
Capr
eolu
s ca
preo
lus
Rupi
capr
a ru
pica
pra
Capr
a ib
ex
Bos
prim
igen
ius
Sus s
crof
a
Equu
s fer
us
Equu
s hy
drun
tinus
Clemente Tronci xxx xx x x xxx x Graziani xx xx xx x x x Ciccio Felice xxx x x xx x x x Radimilli 1956, 99–100.
Riparo Salvini, Terracina, Lazio, Italien (Dryas II; 12.400±170) Schicht Cervus
elaphus Equus hydruntinus
Bos primigenius
Capreolus capreolus
Capra ibex Rupicapra rupicapra
10 ca. 80% X X X 11 ca. 80% X X X 12 ca. 80% X 1 Zahn 1 Phalanx Avellino et al. 1986, 521–524.
Grotta Polesini, Ponte Lucano, Tivoli, Roma, Lazio, Italien Schicht und mögliche klimatische Korrelation Eq
uus f
erus
Equu
s hy
drun
tinus
Bos
prim
igen
ius
Capr
a ib
ex
Rupi
capr
a ru
pica
pra
Cer
vus
elap
hus
Capr
eolu
s ca
preo
lus
Sus s
crof
a
Ges
amt
n % n % n % n % n % n % n % n % n1, Dryas III 159 4,8 238 7,2 96 2,9 32 1 61 1,9 2126 64,6 206 6,3 373 11,3 32912, Alleröd 46 2,2 95 4,5 43 2 14 0,7 32 1,5 1411 66,4 217 10,2 267 12,6 21253, Alleröd 67 2 139 4 60 1,8 22 0,6 58 1,7 2526 74 263 7,7 278 8,1 34134, Alleröd 101 2,5 177 4,3 71 1,7 25 0,6 64 1,6 2931 71,9 372 9,1 334 8,2 40755, Alleröd 134 2 243 3,6 154 2,3 52 0,8 103 1,5 4825 71,5 678 10 561 8,3 67506, Alleröd 86 1,9 221 4,9 92 2 39 0,9 47 1 3324 73,2 334 7,4 396 8,7 45397, Alleröd 138 3,3 255 6 110 2,6 26 0,6 68 1,6 3038 71,8 277 6,5 318 7,5 42208, Alleröd 135 3,7 210 5,7 116 3,2 31 0,8 46 1,3 2577 70,2 300 8,2 254 6,9 36699, Dryas II 158 3,9 290 7,1 92 2,2 51 1,2 64 1,6 3002 73,3 191 4,7 247 6 409510, Dryas II 119 4,3 220 7,9 84 3 41 1,5 90 3,2 1942 69,4 108 3,9 195 7 279911, Dryas II 23 3,4 46 6,7 25 3,7 3 0,4 11 1,6 502 73,5 25 3,7 48 7 68312, Bölling 84 3,7 128 5,6 37 1,6 26 1,1 50 2,2 1581 69,2 196 8,6 182 8 2284Radimilli 1974, 28. Siehe dazu auch Sala 1985, 177.
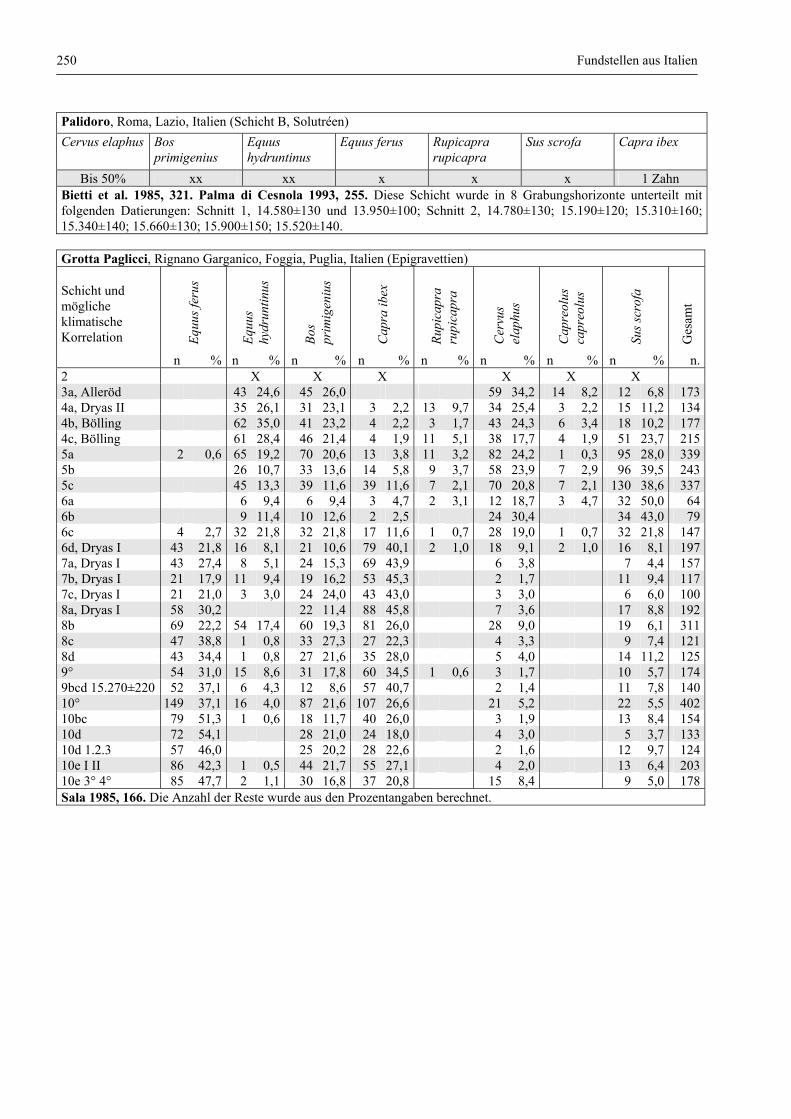
Fundstellen aus Italien 250
Palidoro, Roma, Lazio, Italien (Schicht B, Solutréen) Cervus elaphus Bos
primigenius Equus hydruntinus
Equus ferus Rupicapra rupicapra
Sus scrofa Capra ibex
Bis 50% xx xx x x x 1 Zahn Bietti et al. 1985, 321. Palma di Cesnola 1993, 255. Diese Schicht wurde in 8 Grabungshorizonte unterteilt mit folgenden Datierungen: Schnitt 1, 14.580±130 und 13.950±100; Schnitt 2, 14.780±130; 15.190±120; 15.310±160; 15.340±140; 15.660±130; 15.900±150; 15.520±140.
Grotta Paglicci, Rignano Garganico, Foggia, Puglia, Italien (Epigravettien) Schicht und mögliche klimatische Korrelation Eq
uus f
erus
Equu
s hy
drun
tinus
Bos
prim
igen
ius
Capr
a ib
ex
Rupi
capr
a ru
pica
pra
Cer
vus
elap
hus
Capr
eolu
s ca
preo
lus
Sus s
crof
a
Ges
amt
n % n % n % n % n % n % n % n % n.2 X X X X X X 3a, Alleröd 43 24,6 45 26,0 59 34,2 14 8,2 12 6,8 1734a, Dryas II 35 26,1 31 23,1 3 2,2 13 9,7 34 25,4 3 2,2 15 11,2 1344b, Bölling 62 35,0 41 23,2 4 2,2 3 1,7 43 24,3 6 3,4 18 10,2 1774c, Bölling 61 28,4 46 21,4 4 1,9 11 5,1 38 17,7 4 1,9 51 23,7 2155a 2 0,6 65 19,2 70 20,6 13 3,8 11 3,2 82 24,2 1 0,3 95 28,0 3395b 26 10,7 33 13,6 14 5,8 9 3,7 58 23,9 7 2,9 96 39,5 2435c 45 13,3 39 11,6 39 11,6 7 2,1 70 20,8 7 2,1 130 38,6 3376a 6 9,4 6 9,4 3 4,7 2 3,1 12 18,7 3 4,7 32 50,0 646b 9 11,4 10 12,6 2 2,5 24 30,4 34 43,0 796c 4 2,7 32 21,8 32 21,8 17 11,6 1 0,7 28 19,0 1 0,7 32 21,8 1476d, Dryas I 43 21,8 16 8,1 21 10,6 79 40,1 2 1,0 18 9,1 2 1,0 16 8,1 1977a, Dryas I 43 27,4 8 5,1 24 15,3 69 43,9 6 3,8 7 4,4 1577b, Dryas I 21 17,9 11 9,4 19 16,2 53 45,3 2 1,7 11 9,4 1177c, Dryas I 21 21,0 3 3,0 24 24,0 43 43,0 3 3,0 6 6,0 1008a, Dryas I 58 30,2 22 11,4 88 45,8 7 3,6 17 8,8 1928b 69 22,2 54 17,4 60 19,3 81 26,0 28 9,0 19 6,1 3118c 47 38,8 1 0,8 33 27,3 27 22,3 4 3,3 9 7,4 1218d 43 34,4 1 0,8 27 21,6 35 28,0 5 4,0 14 11,2 1259° 54 31,0 15 8,6 31 17,8 60 34,5 1 0,6 3 1,7 10 5,7 1749bcd 15.270±220 52 37,1 6 4,3 12 8,6 57 40,7 2 1,4 11 7,8 14010° 149 37,1 16 4,0 87 21,6 107 26,6 21 5,2 22 5,5 40210bc 79 51,3 1 0,6 18 11,7 40 26,0 3 1,9 13 8,4 15410d 72 54,1 28 21,0 24 18,0 4 3,0 5 3,7 13310d 1.2.3 57 46,0 25 20,2 28 22,6 2 1,6 12 9,7 12410e I II 86 42,3 1 0,5 44 21,7 55 27,1 4 2,0 13 6,4 20310e 3° 4° 85 47,7 2 1,1 30 16,8 37 20,8 15 8,4 9 5,0 178Sala 1985, 166. Die Anzahl der Reste wurde aus den Prozentangaben berechnet.
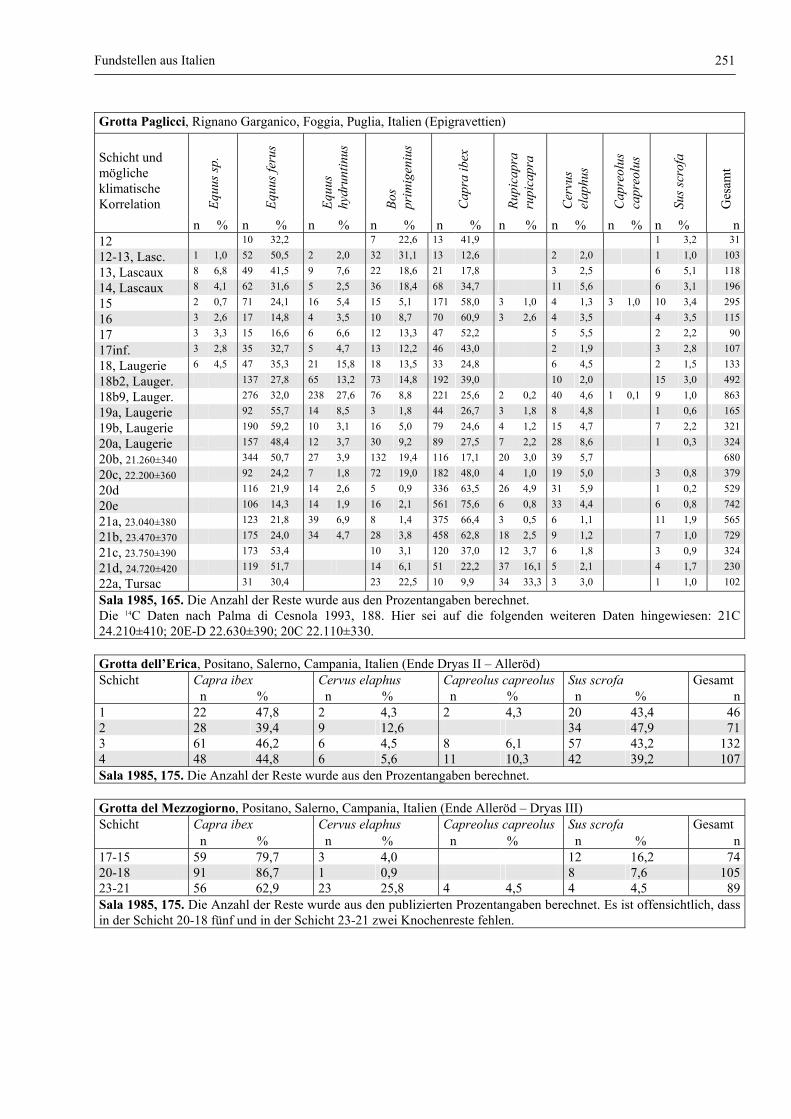
Fundstellen aus Italien 251
Grotta Paglicci, Rignano Garganico, Foggia, Puglia, Italien (Epigravettien) Schicht und mögliche klimatische Korrelation Eq
uus s
p.
Equu
s fer
us
Equu
s hy
drun
tinus
Bos
prim
igen
ius
Capr
a ib
ex
Rupi
capr
a ru
pica
pra
Cer
vus
elap
hus
Capr
eolu
s ca
preo
lus
Sus s
crof
a
Ges
amt
n % n % n % n % n % n % n % n % n % n12 10 32,2 7 22,6 13 41,9 1 3,2 31
12-13, Lasc. 1 1,0 52 50,5 2 2,0 32 31,1 13 12,6 2 2,0 1 1,0 103
13, Lascaux 8 6,8 49 41,5 9 7,6 22 18,6 21 17,8 3 2,5 6 5,1 118
14, Lascaux 8 4,1 62 31,6 5 2,5 36 18,4 68 34,7 11 5,6 6 3,1 196
15 2 0,7 71 24,1 16 5,4 15 5,1 171 58,0 3 1,0 4 1,3 3 1,0 10 3,4 295
16 3 2,6 17 14,8 4 3,5 10 8,7 70 60,9 3 2,6 4 3,5 4 3,5 115
17 3 3,3 15 16,6 6 6,6 12 13,3 47 52,2 5 5,5 2 2,2 90
17inf. 3 2,8 35 32,7 5 4,7 13 12,2 46 43,0 2 1,9 3 2,8 107
18, Laugerie 6 4,5 47 35,3 21 15,8 18 13,5 33 24,8 6 4,5 2 1,5 133
18b2, Lauger. 137 27,8 65 13,2 73 14,8 192 39,0 10 2,0 15 3,0 492
18b9, Lauger. 276 32,0 238 27,6 76 8,8 221 25,6 2 0,2 40 4,6 1 0,1 9 1,0 863
19a, Laugerie 92 55,7 14 8,5 3 1,8 44 26,7 3 1,8 8 4,8 1 0,6 165
19b, Laugerie 190 59,2 10 3,1 16 5,0 79 24,6 4 1,2 15 4,7 7 2,2 321
20a, Laugerie 157 48,4 12 3,7 30 9,2 89 27,5 7 2,2 28 8,6 1 0,3 324
20b, 21.260±340 344 50,7 27 3,9 132 19,4 116 17,1 20 3,0 39 5,7 680
20c, 22.200±360 92 24,2 7 1,8 72 19,0 182 48,0 4 1,0 19 5,0 3 0,8 379
20d 116 21,9 14 2,6 5 0,9 336 63,5 26 4,9 31 5,9 1 0,2 529
20e 106 14,3 14 1,9 16 2,1 561 75,6 6 0,8 33 4,4 6 0,8 742
21a, 23.040±380 123 21,8 39 6,9 8 1,4 375 66,4 3 0,5 6 1,1 11 1,9 565
21b, 23.470±370 175 24,0 34 4,7 28 3,8 458 62,8 18 2,5 9 1,2 7 1,0 729
21c, 23.750±390 173 53,4 10 3,1 120 37,0 12 3,7 6 1,8 3 0,9 324
21d, 24.720±420 119 51,7 14 6,1 51 22,2 37 16,1 5 2,1 4 1,7 230
22a, Tursac 31 30,4 23 22,5 10 9,9 34 33,3 3 3,0 1 1,0 102
Sala 1985, 165. Die Anzahl der Reste wurde aus den Prozentangaben berechnet. Die 14C Daten nach Palma di Cesnola 1993, 188. Hier sei auf die folgenden weiteren Daten hingewiesen: 21C 24.210±410; 20E-D 22.630±390; 20C 22.110±330.
Grotta dell’Erica, Positano, Salerno, Campania, Italien (Ende Dryas II – Alleröd) Schicht Capra ibex Cervus elaphus Capreolus capreolus Sus scrofa Gesamt n % n % n % n % n1 22 47,8 2 4,3 2 4,3 20 43,4 462 28 39,4 9 12,6 34 47,9 713 61 46,2 6 4,5 8 6,1 57 43,2 1324 48 44,8 6 5,6 11 10,3 42 39,2 Sala 1985, 175. Die Anzahl der Reste wurde aus den Prozentangaben berechnet.
107
Grotta del Mezzogiorno, Positano, Salerno, Campania, Italien (Ende Alleröd – Dryas III) Schicht Capra ibex Cervus elaphus Capreolus capreolus Sus scrofa Gesamt n % n % n % n % n17-15 59 79,7 3 4,0 12 16,2 7420-18 91 86,7 1 0,9 8 7,6 10523-21 56 62,9 23 25,8 4 4,5 4 4,5 89Sala 1985, 175. Die Anzahl der Reste wurde aus den publizierten Prozentangaben berechnet. Es ist offensichtlich, dass in der Schicht 20-18 fünf und in der Schicht 23-21 zwei Knochenreste fehlen.
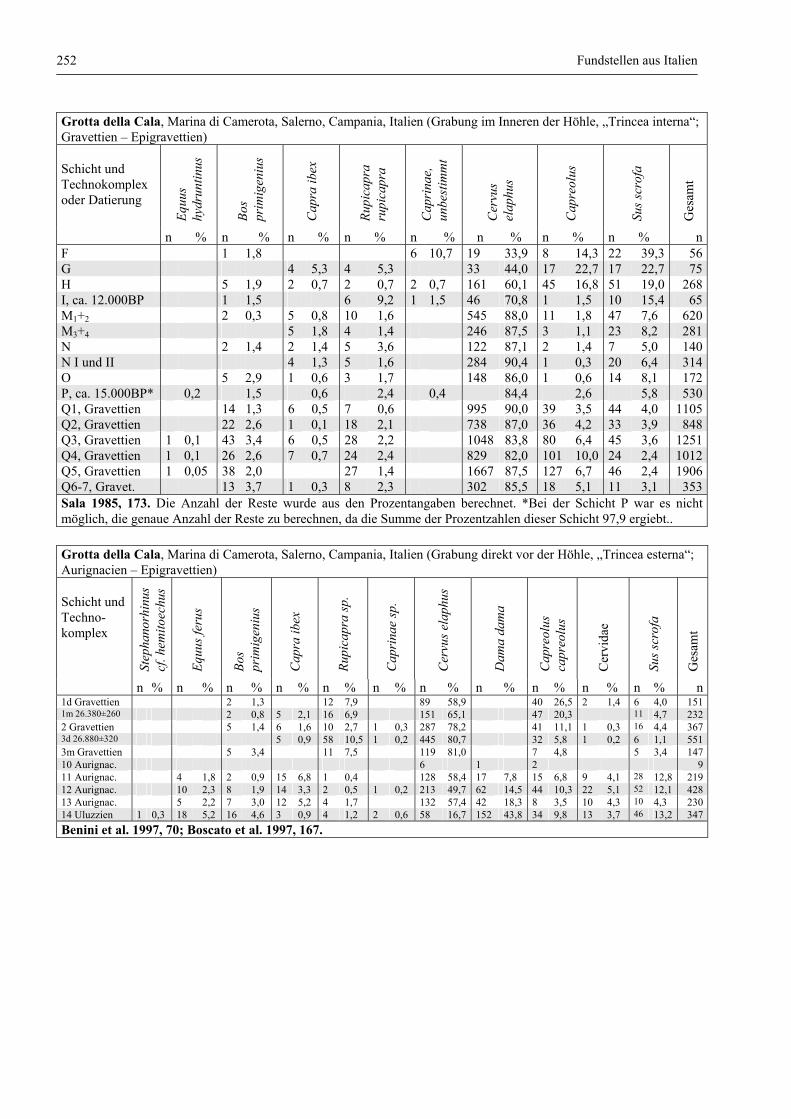
Fundstellen aus Italien 252
Grotta della Cala, Marina di Camerota, Salerno, Campania, Italien (Grabung im Inneren der Höhle, „Trincea interna“; Gravettien – Epigravettien) Schicht und Technokomplex oder Datierung Eq
uus
hydr
untin
us
Bos
prim
igen
ius
Capr
a ib
ex
Rupi
capr
a ru
pica
pra
Capr
inae
, un
best
imm
t
Cer
vus
elap
hus
Capr
eolu
s
Sus s
crof
a
Ges
amt
n % n % n % n % n % n % n % n % n F 1 1,8 6 10,7 19 33,9 8 14,3 22 39,3 56G 4 5,3 4 5,3 33 44,0 17 22,7 17 22,7 75H 5 1,9 2 0,7 2 0,7 2 0,7 161 60,1 45 16,8 51 19,0 268I, ca. 12.000BP 1 1,5 6 9,2 1 1,5 46 70,8 1 1,5 10 15,4 65M1+2 2 0,3 5 0,8 10 1,6 545 88,0 11 1,8 47 7,6 620M3+4 5 1,8 4 1,4 246 87,5 3 1,1 23 8,2 281N 2 1,4 2 1,4 5 3,6 122 87,1 2 1,4 7 5,0 140N I und II 4 1,3 5 1,6 284 90,4 1 0,3 20 6,4 314O 5 2,9 1 0,6 3 1,7 148 86,0 1 0,6 14 8,1 172P, ca. 15.000BP* 0,2 1,5 0,6 2,4 0,4 84,4 2,6 5,8 530Q1, Gravettien 14 1,3 6 0,5 7 0,6 995 90,0 39 3,5 44 4,0 1105Q2, Gravettien 22 2,6 1 0,1 18 2,1 738 87,0 36 4,2 33 3,9 848Q3, Gravettien 1 0,1 43 3,4 6 0,5 28 2,2 1048 83,8 80 6,4 45 3,6 1251Q4, Gravettien 1 0,1 26 2,6 7 0,7 24 2,4 829 82,0 101 10,0 24 2,4 1012Q5, Gravettien 1 0,05 38 2,0 27 1,4 1667 87,5 127 6,7 46 2,4 1906Q6-7, Gravet. 13 3,7 1 0,3 8 2,3 302 85,5 18 5,1 11 3,1 353Sala 1985, 173. Die Anzahl der Reste wurde aus den Prozentangaben berechnet. *Bei der Schicht P war es nicht möglich, die genaue Anzahl der Reste zu berechnen, da die Summe der Prozentzahlen dieser Schicht 97,9 ergiebt..
Grotta della Cala, Marina di Camerota, Salerno, Campania, Italien (Grabung direkt vor der Höhle, „Trincea esterna“; Aurignacien – Epigravettien) Schicht und Techno-komplex
Step
hano
rhin
us
cf. h
emito
echu
s
Equu
s fer
us
Bos
prim
igen
ius
Capr
a ib
ex
Rupi
capr
a sp
.
Capr
inae
sp.
Cerv
us e
laph
us
Dam
a da
ma
Capr
eolu
s ca
preo
lus
Cer
vida
e
Sus s
crof
a
Ges
amt
n % n % n % n % n % n % n % n % n % n % n % n1d Gravettien 2 1,3 12 7,9 89 58,9 40 26,5 2 1,4 6 4,0 1511m 26.380±260 2 0,8 5 2,1 16 6,9 151 65,1 47 20,3 11 4,7 2322 Gravettien 5 1,4 6 1,6 10 2,7 1 0,3 287 78,2 41 11,1 1 0,3 16 4,4 3673d 26.880±320 5 0,9 58 10,5 1 0,2 445 80,7 32 5,8 1 0,2 6 1,1 5513m Gravettien 5 3,4 11 7,5 119 81,0 7 4,8 5 3,4 14710 Aurignac. 6 1 2 911 Aurignac. 4 1,8 2 0,9 15 6,8 1 0,4 128 58,4 17 7,8 15 6,8 9 4,1 28 12,8 21912 Aurignac. 10 2,3 8 1,9 14 3,3 2 0,5 1 0,2 213 49,7 62 14,5 44 10,3 22 5,1 52 12,1 42813 Aurignac. 5 2,2 7 3,0 12 5,2 4 1,7 132 57,4 42 18,3 8 3,5 10 4,3 10 4,3 23014 Uluzzien 1 0,3 18 5,2 16 4,6 3 0,9 4 1,2 2 0,6 58 16,7 152 43,8 34 9,8 13 3,7 46 13,2 347Benini et al. 1997, 70; Boscato et al. 1997, 167.
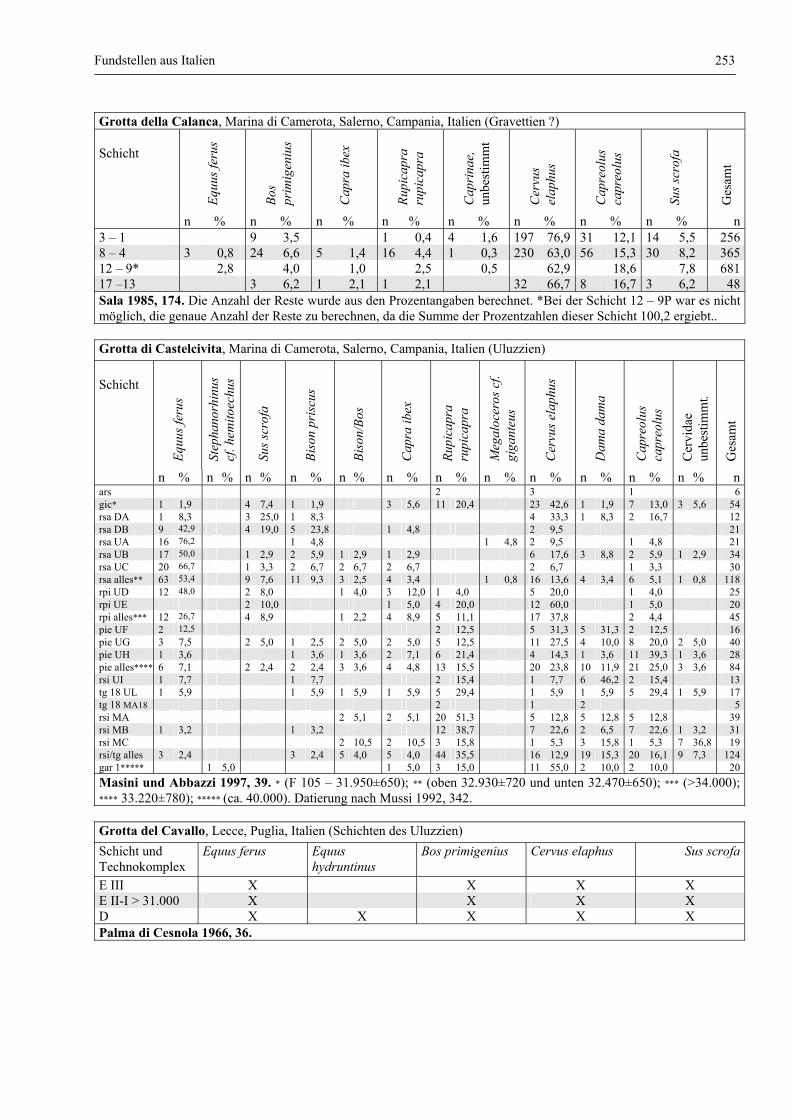
Fundstellen aus Italien 253
Grotta della Calanca, Marina di Camerota, Salerno, Campania, Italien (Gravettien ?) Schicht
Equu
s fer
us
Bos
prim
igen
ius
Capr
a ib
ex
Rupi
capr
a ru
pica
pra
Capr
inae
, un
best
imm
t
Cer
vus
elap
hus
Capr
eolu
s ca
preo
lus
Sus s
crof
a
Ges
amt
n % n % n % n % n % n % n % n % n3 – 1 9 3,5 1 0,4 4 1,6 197 76,9 31 12,1 14 5,5 2568 – 4 3 0,8 24 6,6 5 1,4 16 4,4 1 0,3 230 63,0 56 15,3 30 8,2 36512 – 9* 2,8 4,0 1,0 2,5 0,5 62,9 18,6 7,8 68117 –13 3 6,2 1 2,1 1 2,1 32 66,7 8 16,7 3 6,2 48Sala 1985, 174. Die Anzahl der Reste wurde aus den Prozentangaben berechnet. *Bei der Schicht 12 – 9P war es nicht möglich, die genaue Anzahl der Reste zu berechnen, da die Summe der Prozentzahlen dieser Schicht 100,2 ergiebt..
Grotta di Castelcivita, Marina di Camerota, Salerno, Campania, Italien (Uluzzien) Schicht
Equu
s fer
us
Step
hano
rhin
us
cf. h
emito
echu
s
Sus s
crof
a
Biso
n pr
iscus
Biso
n/Bo
s
Capr
a ib
ex
Rupi
capr
a ru
pica
pra
Meg
aloc
eros
cf.
giga
nteu
s
Cerv
us e
laph
us
Dam
a da
ma
Capr
eolu
s ca
preo
lus
Cer
vida
e un
best
imm
t.
Ges
amt
n % n % n % n % n % n % n % n % n % n % n % n % nars 2 3 1 6gic* 1 1,9 4 7,4 1 1,9 3 5,6 11 20,4 23 42,6 1 1,9 7 13,0 3 5,6 54rsa DA 1 8,3 3 25,0 1 8,3 4 33,3 1 8,3 2 16,7 12rsa DB 9 42,9 4 19,0 5 23,8 1 4,8 2 9,5 21rsa UA 16 76,2 1 4,8 1 4,8 2 9,5 1 4,8 21rsa UB 17 50,0 1 2,9 2 5,9 1 2,9 1 2,9 6 17,6 3 8,8 2 5,9 1 2,9 34rsa UC 20 66,7 1 3,3 2 6,7 2 6,7 2 6,7 2 6,7 1 3,3 30rsa alles** 63 53,4 9 7,6 11 9,3 3 2,5 4 3,4 1 0,8 16 13,6 4 3,4 6 5,1 1 0,8 118rpi UD 12 48,0 2 8,0 1 4,0 3 12,0 1 4,0 5 20,0 1 4,0 25rpi UE 2 10,0 1 5,0 4 20,0 12 60,0 1 5,0 20rpi alles*** 12 26,7 4 8,9 1 2,2 4 8,9 5 11,1 17 37,8 2 4,4 45pie UF 2 12,5 2 12,5 5 31,3 5 31,3 2 12,5 16pie UG 3 7,5 2 5,0 1 2,5 2 5,0 2 5,0 5 12,5 11 27,5 4 10,0 8 20,0 2 5,0 40pie UH 1 3,6 1 3,6 1 3,6 2 7,1 6 21,4 4 14,3 1 3,6 11 39,3 1 3,6 28pie alles**** 6 7,1 2 2,4 2 2,4 3 3,6 4 4,8 13 15,5 20 23,8 10 11,9 21 25,0 3 3,6 84rsi UI 1 7,7 1 7,7 2 15,4 1 7,7 6 46,2 2 15,4 13tg 18 UL 1 5,9 1 5,9 1 5,9 1 5,9 5 29,4 1 5,9 1 5,9 5 29,4 1 5,9 17tg 18 MA18 2 1 2 5rsi MA 2 5,1 2 5,1 20 51,3 5 12,8 5 12,8 5 12,8 39rsi MB 1 3,2 1 3,2 12 38,7 7 22,6 2 6,5 7 22,6 1 3,2 31rsi MC 2 10,5 2 10,5 3 15,8 1 5,3 3 15,8 1 5,3 7 36,8 19rsi/tg alles 3 2,4 3 2,4 5 4,0 5 4,0 44 35,5 16 12,9 19 15,3 20 16,1 9 7,3 124gar 1***** 1 5,0 1 5,0 3 15,0 11 55,0 2 10,0 2 10,0 20Masini und Abbazzi 1997, 39. * (F 105 – 31.950±650); ** (oben 32.930±720 und unten 32.470±650); *** (>34.000); **** 33.220±780); ***** (ca. 40.000). Datierung nach Mussi 1992, 342.
Grotta del Cavallo, Lecce, Puglia, Italien (Schichten des Uluzzien) Schicht und Technokomplex
Equus ferus Equus hydruntinus
Bos primigenius Cervus elaphus Sus scrofa
E III X X X X E II-I > 31.000 X X X X D X X X X X Palma di Cesnola 1966, 36.
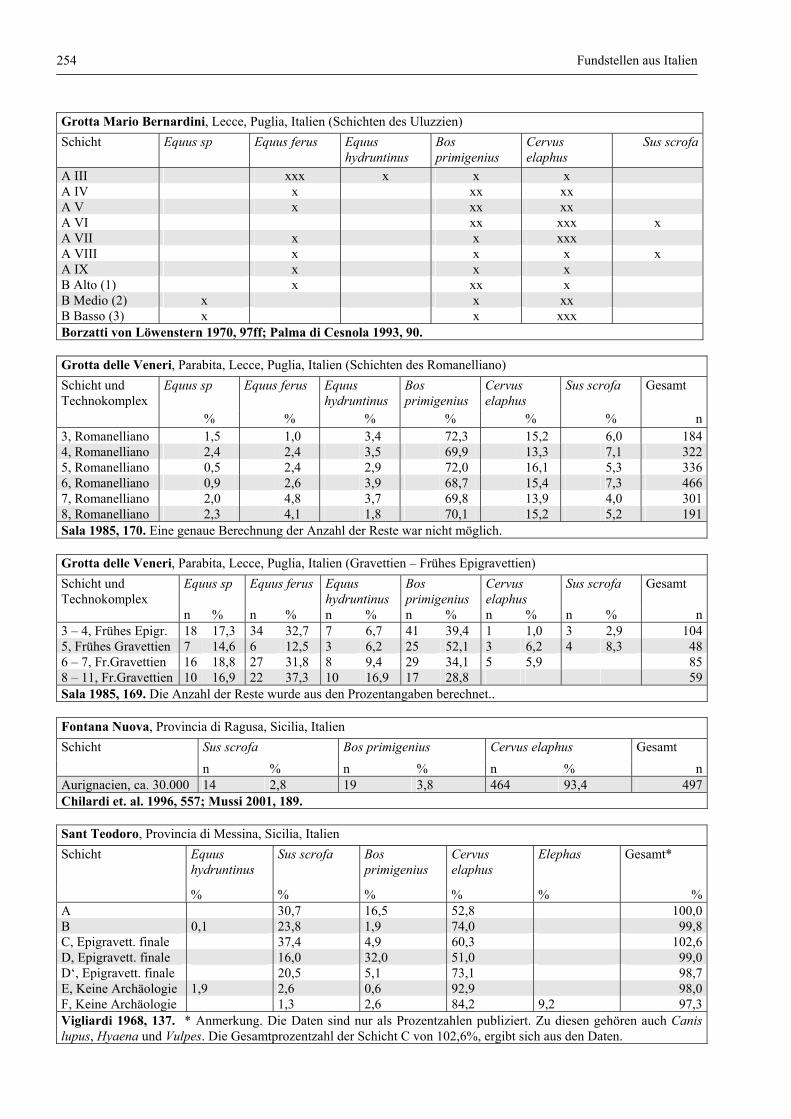
Fundstellen aus Italien
254
Grotta Mario Bernardini, Lecce, Puglia, Italien (Schichten des Uluzzien) Schicht Equus sp Equus ferus Equus
hydruntinus Bos primigenius
Cervus elaphus
Sus scrofa
A III xxx x x x A IV x xx xx A V x xx xx A VI xx xxx x A VII x x xxx A VIII x x x x A IX x x x B Alto (1) x xx x B Medio (2) x x xx B Basso (3) x x xxx Borzatti von Löwenstern 1970, 97ff; Palma di Cesnola 1993, 90.
Grotta delle Veneri, Parabita, Lecce, Puglia, Italien (Schichten des Romanelliano) Schicht und Technokomplex
Equus sp Equus ferus Equus hydruntinus
Bos primigenius
Cervus elaphus
Sus scrofa Gesamt
% % % % % % n3, Romanelliano 1,5 1,0 3,4 72,3 15,2 6,0 1844, Romanelliano 2,4 2,4 3,5 69,9 13,3 7,1 3225, Romanelliano 0,5 2,4 2,9 72,0 16,1 5,3 3366, Romanelliano 0,9 2,6 3,9 68,7 15,4 7,3 4667, Romanelliano 2,0 4,8 3,7 69,8 13,9 4,0 3018, Romanelliano 2,3 4,1 1,8 70,1 15,2 5,2 191Sala 1985, 170. Eine genaue Berechnung der Anzahl der Reste war nicht möglich.
Grotta delle Veneri, Parabita, Lecce, Puglia, Italien (Gravettien – Frühes Epigravettien) Schicht und Technokomplex
Equus sp Equus ferus Equus hydruntinus
Bos primigenius
Cervus elaphus
Sus scrofa Gesamt
n % n % n % n % n % n % n3 – 4, Frühes Epigr. 18 17,3 34 32,7 7 6,7 41 39,4 1 1,0 3 2,9 1045, Frühes Gravettien 7 14,6 6 12,5 3 6,2 25 52,1 3 6,2 4 8,3 486 – 7, Fr.Gravettien 16 18,8 27 31,8 8 9,4 29 34,1 5 5,9 858 – 11, Fr.Gravettien 10 16,9 22 37,3 10 16,9 17 28,8 59Sala 1985, 169. Die Anzahl der Reste wurde aus den Prozentangaben berechnet..
Fontana Nuova, Provincia di Ragusa, Sicilia, Italien Schicht Sus scrofa Bos primigenius Cervus elaphus Gesamt n % n % n % nAurignacien, ca. 30.000 14 2,8 19 3,8 464 93,4 497Chilardi et. al. 1996, 557; Mussi 2001, 189.
Sant Teodoro, Provincia di Messina, Sicilia, Italien Schicht
Equus hydruntinus
Sus scrofa Bos primigenius
Cervus elaphus
Elephas Gesamt*
% % % % % %A 30,7 16,5 52,8 100,0B 0,1 23,8 1,9 74,0 99,8C, Epigravett. finale 37,4 4,9 60,3 102,6D, Epigravett. finale 16,0 32,0 51,0 99,0D‘, Epigravett. finale 20,5 5,1 73,1 98,7E, Keine Archäologie 1,9 2,6 0,6 92,9 98,0F, Keine Archäologie 1,3 2,6 84,2 9,2 97,3Vigliardi 1968, 137. * Anmerkung. Die Daten sind nur als Prozentzahlen publiziert. Zu diesen gehören auch Canis lupus, Hyaena und Vulpes. Die Gesamtprozentzahl der Schicht C von 102,6%, ergibt sich aus den Daten.
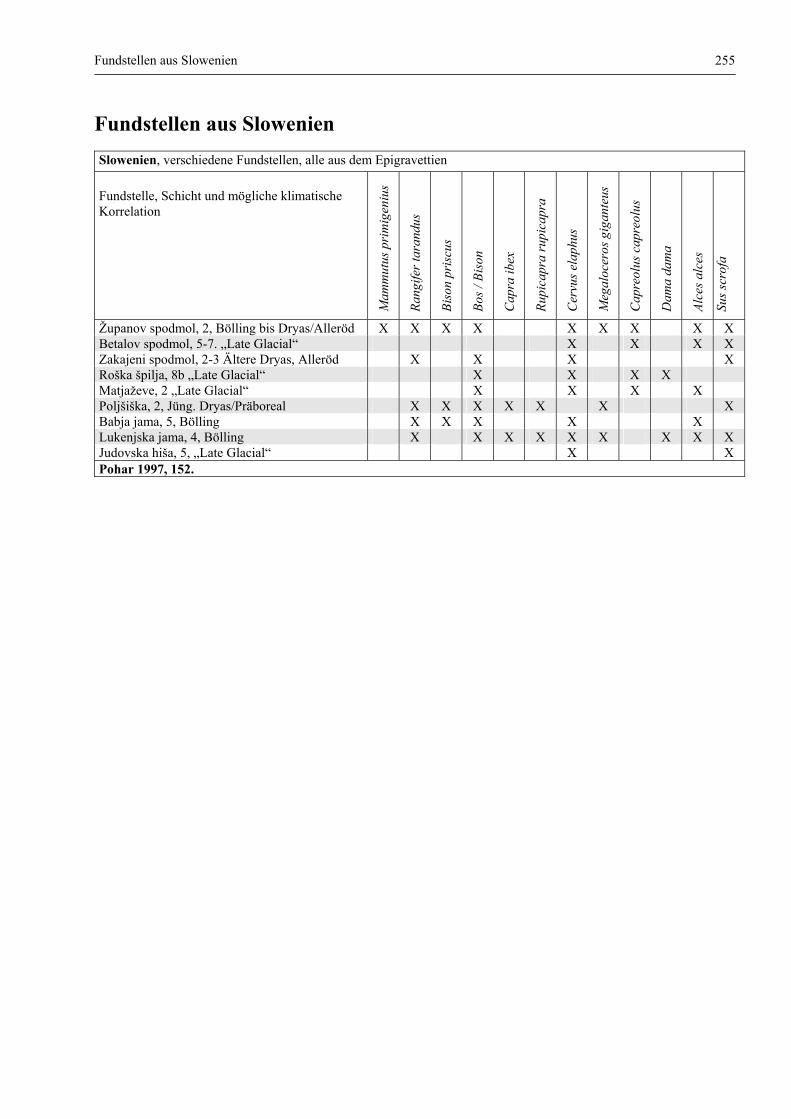
Fundstellen aus Slowenien 255
Fundstellen aus Slowenien Slowenien, verschiedene Fundstellen, alle aus dem Epigravettien Fundstelle, Schicht und mögliche klimatische Korrelation
Mam
mut
us p
rim
igen
ius
Rang
ifer t
aran
dus
Biso
n pr
iscus
Bos /
Bis
on
Capr
a ib
ex
Rupi
capr
a ru
pica
pra
Cerv
us e
laph
us
Meg
aloc
eros
gig
ante
us
Capr
eolu
s cap
reol
us
Dam
a da
ma
Alce
s alc
es
Sus s
crof
a
Županov spodmol, 2, Bölling bis Dryas/Alleröd X X X X X X X X X Betalov spodmol, 5-7. „Late Glacial“ X X X X Zakajeni spodmol, 2-3 Ältere Dryas, Alleröd X X X X Roška špilja, 8b „Late Glacial“ X X X X Matjaževe, 2 „Late Glacial“ X X X X Poljšiška, 2, Jüng. Dryas/Präboreal X X X X X X X Babja jama, 5, Bölling X X X X X Lukenjska jama, 4, Bölling X X X X X X X X X Judovska hiša, 5, „Late Glacial“ X X Pohar 1997, 152.
Related Documents











