Separation of phasic arousal and expectancy effects in a speeded reaction time task via fMRI STEVEN A. HACKLEY, a ROBERT LANGNER, b,c BETTINA ROLKE, c MICHAEL ERB, b WOLFGANG GRODD, b and ROLF ULRICH c a Department of Psychological Sciences, University of MissouriFColumbia, Columbia, Missouri, USA b Department of Neuroradiology, University of Tuebingen, Tuebingen, Germany c Department of Cognitive & Biological Psychology, University of Tuebingen, Tuebingen, Germany Abstract Sixteen neurologically normal volunteers performed a 2-choice speeded reaction time (RT) task in which the imperative was the change in color of a clock hand. During trial blocks with low temporal uncertainty (good clock condition), this imperative stimulus occurred at a fixed location (e.g., 2:00). In the bad clock condition, the clock was unpredictive of imperative onset. On half of both good and bad clock trials, a task-irrelevant, cutaneous accessory stimulus accom- panied the imperative. The speeding of reactions by the accessory was associated with activation primarily in and near the supramarginal gyrus of the parietal lobe. Contrasts of good- versus bad-clock conditions revealed activation in a variety of perceptual, motor, and executive control regions. Apart from interactions within the cerebellum and left anterior insula, there was little overlap between structures influenced by the arousal and expectancy manipulations. Descriptors: attention, arousal, expectancy, motor, reaction time, functional magnetic resonance imaging Previous research indicates that warning effects depend not just on directing attention to the point in time at which an imperative is expected but also on the ability of the warning signal to trigger a brief burst of arousal. Behavioral evidence for this assumption includes the finding that, at short intervals (foreperiods) between warning and imperative stimuli, the intensity of the warning signal affects the force of response (Ulrich & Mattes, 1996). Warning signals associated with brief foreperiods also cause a shortening of reaction time (RT) that is often coupled with an increased error rate (Bernstein, Rose, & Ashe, 1970; Posner, Klein, Summers, & Bugge, 1973). An explanation of the RT effect based on distraction can be excluded because, if capture of attention by the warning signal were involved, this should slow rather than speed responses. Considering the fact that arousal-based effects can occur even if the warning signal actually follows the imperative stimulus by a brief interval (Bernstein et al., 1970), the word ‘‘warning’’ is something of a misnomer. We follow conventional usage in pre- ferring the term ‘‘accessory stimulus’’ when foreperiods are below 500 ms (Welch & Warren, 1986). Warning and accessory stimuli facilitate reflexive as well as voluntary reactions (for review, see Hackley & Valle-Incla´ n, 2003). The underlying mechanisms for the two types of motor behavior can be presumed to overlap because there are a number of parallels: For example, effects for both types of reactions vary as a function of foreperiod duration (Sollers & Hackley, 1997) and the peak force of both is facilitated at short foreperiods (Scheirs & Brunia, 1985; Ulrich & Mattes, 1996). Invasive neu- robiological research in animals indicates that the speeding of reflexive saccades by a warning signal is due to enhancement of early motor processes, not faster sensory analysis (Fecteau & Munoz, 2007). Similarly, potentiation of reflexive blink ampli- tude by an accessory stimulus is due to facilitation of the motor limb of the reflex arc (Young, Cegavske, & Thompson, 1976). In the case of the masseteric H reflex, this motor facilitation is known to be noradrenergic in nature (Stafford & Jacobs, 1990). Similarly, a study of phasic arousal effects on voluntary reactions in monkeys implicated a noradrenergic mechanism (Witte & Marrocco, 1997). With this in mind, Coull, Nobre, and Frith (2001) employed noradrenergic blockers in an fMRI study to identify arousal- related components of warning effects in the human brain. Sub- jects performed a choice RT task in which a foveally presented symbol precued either the spatial location of the target or its time of arrival. There was also a third condition, designed to isolate phasic arousal effects, in which the precue was uninformative. The spatial and temporal precues were valid on some trials, in- valid on others. When scans were compared between clonidine (a nor- adrenergic a2 agonist) and placebo sessions, an attenuation of This research was supported in part by sabbatical grants to the first author from Tuebingen University and the Max Planck Institute for Human Cognitive and Brain Sciences. We thank Dr. Christoph Braun for the loan of the Braille stimulator, Ju¨ rgen Dax for assistance with the use of this device, and Luisa Frei for help with the data collection. Robert Langner is currently with the Neuropsychology Section, Department of Neurology, RWTH Aachen University (Germany). Address reprint requests to: Steven A. Hackley, Department of Psy- chological Sciences, University of Missouri, Columbia, 210 McAlester Hall, Columbia, MO 65211, USA. E-mail: [email protected] Psychophysiology, 46 (2009), 163–171. Wiley Periodicals, Inc. Printed in the USA. Copyright r 2008 Society for Psychophysiological Research DOI: 10.1111/j.1469-8986.2008.00722.x 163

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Separation of phasic arousal and expectancy effects in a
speeded reaction time task via fMRI
STEVEN A. HACKLEY,a ROBERT LANGNER,b,c BETTINA ROLKE,c MICHAEL ERB,b
WOLFGANG GRODD,b and ROLF ULRICHc
aDepartment of Psychological Sciences, University of MissouriFColumbia, Columbia, Missouri, USAbDepartment of Neuroradiology, University of Tuebingen, Tuebingen, GermanycDepartment of Cognitive & Biological Psychology, University of Tuebingen, Tuebingen, Germany
Abstract
Sixteenneurologically normal volunteers performed a 2-choice speeded reaction time (RT) task inwhich the imperative
was the change in color of a clock hand. During trial blocks with low temporal uncertainty (good clock condition), this
imperative stimulus occurred at a fixed location (e.g., 2:00). In the bad clock condition, the clock was unpredictive of
imperative onset. On half of both good and bad clock trials, a task-irrelevant, cutaneous accessory stimulus accom-
panied the imperative. The speeding of reactions by the accessory was associated with activation primarily in and near
the supramarginal gyrus of the parietal lobe. Contrasts of good- versus bad-clock conditions revealed activation in a
variety of perceptual, motor, and executive control regions. Apart from interactions within the cerebellum and left
anterior insula, there was little overlap between structures influenced by the arousal and expectancy manipulations.
Descriptors: attention, arousal, expectancy, motor, reaction time, functional magnetic resonance imaging
Previous research indicates that warning effects depend not just
on directing attention to the point in time at which an imperative
is expected but also on the ability of the warning signal to trigger
a brief burst of arousal. Behavioral evidence for this assumption
includes the finding that, at short intervals (foreperiods) between
warning and imperative stimuli, the intensity of the warning
signal affects the force of response (Ulrich & Mattes, 1996).
Warning signals associated with brief foreperiods also cause a
shortening of reaction time (RT) that is often coupled with an
increased error rate (Bernstein, Rose, & Ashe, 1970; Posner,
Klein, Summers, & Bugge, 1973). An explanation of the RT
effect based on distraction can be excluded because, if capture of
attention by the warning signal were involved, this should slow
rather than speed responses.
Considering the fact that arousal-based effects can occur even
if the warning signal actually follows the imperative stimulus by a
brief interval (Bernstein et al., 1970), the word ‘‘warning’’ is
something of a misnomer. We follow conventional usage in pre-
ferring the term ‘‘accessory stimulus’’ when foreperiods are below
500 ms (Welch & Warren, 1986).
Warning and accessory stimuli facilitate reflexive as well as
voluntary reactions (for review, see Hackley & Valle-Inclan,
2003). The underlying mechanisms for the two types of motor
behavior can be presumed to overlap because there are a number
of parallels: For example, effects for both types of reactions vary
as a function of foreperiod duration (Sollers & Hackley, 1997)
and the peak force of both is facilitated at short foreperiods
(Scheirs & Brunia, 1985; Ulrich & Mattes, 1996). Invasive neu-
robiological research in animals indicates that the speeding of
reflexive saccades by a warning signal is due to enhancement of
early motor processes, not faster sensory analysis (Fecteau &
Munoz, 2007). Similarly, potentiation of reflexive blink ampli-
tude by an accessory stimulus is due to facilitation of the motor
limb of the reflex arc (Young, Cegavske, & Thompson, 1976). In
the case of the masseteric H reflex, this motor facilitation is
known to be noradrenergic in nature (Stafford & Jacobs, 1990).
Similarly, a study of phasic arousal effects on voluntary reactions
in monkeys implicated a noradrenergic mechanism (Witte &
Marrocco, 1997).
With this in mind, Coull, Nobre, and Frith (2001) employed
noradrenergic blockers in an fMRI study to identify arousal-
related components of warning effects in the human brain. Sub-
jects performed a choice RT task in which a foveally presented
symbol precued either the spatial location of the target or its time
of arrival. There was also a third condition, designed to isolate
phasic arousal effects, in which the precue was uninformative.
The spatial and temporal precues were valid on some trials, in-
valid on others.
When scans were compared between clonidine (a nor-
adrenergic a2 agonist) and placebo sessions, an attenuation of
This research was supported in part by sabbatical grants to the first
author from Tuebingen University and the Max Planck Institute for
HumanCognitive and Brain Sciences.We thankDr. Christoph Braun for
the loan of the Braille stimulator, Jurgen Dax for assistance with the use
of this device, and Luisa Frei for help with the data collection. Robert
Langner is currently with the Neuropsychology Section, Department of
Neurology, RWTH Aachen University (Germany).Address reprint requests to: Steven A. Hackley, Department of Psy-
chological Sciences, University of Missouri, Columbia, 210 McAlesterHall, Columbia, MO 65211, USA. E-mail: [email protected]
Psychophysiology, 46 (2009), 163–171. Wiley Periodicals, Inc. Printed in the USA.Copyright r 2008 Society for Psychophysiological ResearchDOI: 10.1111/j.1469-8986.2008.00722.x
163

task-induced activation was found in five regions. For visuospa-
tial orienting, clonidine reduced effects in the right superior pa-
rietal cortex (BA 7). For temporal expectancy, a reduction was
observed in the left dorsolateral prefrontal cortex (BA 9), left
anterior insula, and left ventral premotor cortex (BA 6). For the
neutral precue, a reduction in task-induced changes in blood
oxygenation levels was observed in the left temporo-parietal
junction (BA 39). A follow-up study (Coull, Jones, Egan, Frith,
& Maze, 2004) showed that another noradrenergic a2 agonist,
dexmedotomidine, enhanced activity in the pulvinar and in a
region in or slightly above the left supramarginal gyrus (BA 40)
when phasic arousal was augmented via task-irrelevant acoustic
stimulation.
Rationale
These results extend Nobre and Coull’s influential corpus of re-
search on the topic of temporal orienting. However, the gener-
ality of their findings is limited by the fact that neuromodulators
other than norepinephrine are likely to be involved in generating
the diffuse brain activation that constitutes arousal (e.g., Rob-
bins & Everitt, 1995). The generality of their work has been
further limited by their more-or-less exclusive use of the valid/
invalid precue method. The use of alternative paradigms is es-
sential if the full range of cognitive processes that underlie warn-
ing effects is to be characterized.
The goal of our study was to distinguish brain activation
patterns associated with the temporal expectancy and phasic
arousal mechanisms engaged by warning signals. Expectancy
was manipulated using two variants of an analog clock display
(Requın, Brener, & Ring, 1991). During blocks of ‘‘good clock’’
trials, the participant knew exactly when the imperative would
occur (e.g., as soon as the hand pointed toward the four o’clock
position). To discourage a spatial attention strategy (focusing at
the four o’clock point), the clock ‘‘hand’’ was actually a wide,
short arrow that rotated about its center of gravity. Temporal
uncertainty was enhanced during the ‘‘bad clock’’ condition, in
that the orientation and speed of the clock hand were unpredic-
tive of imperative onset. Comparing the two types of trials al-
lowed us to examine brain activation associated with expectancy,
but in the absence of any need for the subject to actively estimate
the passage of time or prepare to reorient in the case of an invalid
cue, as in the Coull and Nobre studies. The good- versus bad-
clock contrast also addresses the less extensively investigated
topic of externalized time keeping (e.g., Jennings, van derMolen,
& Terezis, 1987).
An accessory stimulus was used to elicit phasic arousal. On
half of the blocks of trials, an intense but task-irrelevant vibra-
tory stimulus was delivered to the soles of the subject’s bare feet.
The bilateral cutaneous stimuli began 100 ms prior to the im-
perative stimulus, which was a change in color of the clock hand.
The salience of the accessory is illustrated by the fact that
on the first familiarization trial it triggered a whole-body startle,
typically accompanied by laughter, in each of our 16 partici-
pants. (Eyeblinks were not monitored, but overt startle was oth-
erwise completely habituated by the third familiarization trial.)
In planned contrasts designed to assess phasic arousal, the ac-
cessory and no-accessory blocks were compared in the good
clock condition. Because participants could accurately predict
imperative onset in these conditions, the foot vibration would
have been uninformative as well as task irrelevant. There was
presumably no temporal uncertainty whatsoever on good clock
trials. The subjects knew exactly when the imperative would
occur.
Predictions
On the assumption that stimulus-induced arousal engages mech-
anisms not limited to the noradrenergic system studied by Coull
and colleagues (Coull et al., 2001; Coull, Jones, et al., 2004), we
predicted that a broader range of brain regions would be impli-
cated than in their experiments. These regions might include the
various arousal-sensitive regions that have been identified in
emotion research. For example, a recent study at this laboratory
found that phenomenological and electrodermal indices of
arousal during affective picture viewing were correlated with ac-
tivity in the thalamus, right supramarginal gyrus, and medial
orbitofrontal cortex (Anders, Lotze, Erb, Grodd, & Birbaumer,
2004). Imaging studies by Critchley and colleagues highlight the
importance of the insula for representing bodily states of arousal
(Critchley, Mathias, & Dolan, 2002; Critchley, Melmed, Feath-
erstone, Mathias, & Dolan, 2002).
The range of areas related to expectancy in the good clock
condition might be more constrained than that reported by
Nobre, Coull, and colleagues because our participants did not
need to prepare to reorient following invalid cues or to internally
estimate foreperiod duration. It is possible that keeping track of
time bymeans of explicit, external cues could involve some of the
same brain structures as internal time estimation. A large number
of structures have been associated with the internal time estima-
tion, but by far the most consistently implicated have been the
cerebellum and frontostriatal regions (Mattell & Meck, 2004;
Stevens, Kiehl, Pearlson, & Calhoun, 2007). Using warning sig-
nals effectively requires not just tracking the passage of time, but
also orienting to the point in time at which the imperative is
expected to arrive. Coull and Nobre (e.g., 1998) have convinc-
ingly argued that the left superior parietal cortex is critical for this
function.
Some incidental predictions should be noted. These follow
from the manipulations employed but are not particularly rel-
evant to distinguishing phasic arousal and temporal expectancy
effects. Because our accessory stimulus was cutaneous, it can be
assumed to activate the various somatosensory cortices. As noted
above, it was also mildly reflexogenic and so might recruit in-
hibitory motor control processes. To monitor the analog clock
countdown, frontal executive and visual perception areas would
be needed.
The above predictions were tested using a 2 � 2 blocked an-
alyses, with good clock versus bad clock and accessory versus no
accessory as factors. In addition, event-related analyses were
employed on an exploratory basis to assess which portions of the
stimulus–response (S-R) pathway might be modulated by ex-
pectancy and arousal. Our interest in this topic comes from a
series of studies using electrophysiological and dual-task behav-
ioral methods to demonstrate that warning/accessory effects on
reaction time are not due to the speeding of late motoric pro-
cesses (Bausenhart, Rolke, Hackley, & Ulrich, 2006; Hackley,
Schankin, Wohlschlaeger, & Wascher, 2007; Hackley & Valle-
Inclan, 1998, 1999; Muller-Gethmann, Ulrich, & Rinkenauer,
2003). However, other aspects of response execution certainly are
affected (e.g., Coull & Nobre, 1998; Hasbroucq et al., 1999).
Therefore, we evaluated the sensitivity of basal ganglia, motor
cortex, and other relevant structures to arousal and expectancy.
Similarly, both behavioral and electrophysiological measures in-
dicate that warning signals influence sensory-perceptual analyses
164 S.A. Hackley et al.

(e.g., Correa, Lupianez, Madrid, & Tudela, 2006; Rolke & Ho-
fmann, 2007), although perhaps only late perceptual processes
(Hackley et al., 2007). Consequently, we predicted that event-
related analyses of the S-R interval would reveal activations
within visual areas necessary for discriminating the color of the
imperative. Finally, because accessory stimulation can enhance
early movement-related potentials (Hackley & Valle-Inclan,
1999) and trigger a speed–accuracy trade-off (Posner et al.,
1973), we predicted that structures involved in decision making
would be activated by phasic arousal (e.g., SMA,ACC, striatum;
the theory underlying this prediction is developed more fully in
Hackley & Valle-Inclan, 2003).
Methods
Participants
Sixteen neurologically normal, adult volunteers served as partici-
pants. Ten were female, 14 were right-handed by self-report, and
the mean age was 29.4 years. Because our experimental
manipulation of phasic arousal could easily be eclipsed by sit-
uational anxiety, we preferentially recruited individuals who
had previous experience inMR scanners (N5 14). The study was
compliant with the code of ethics of the World Medical Asso-
ciation (Declaration of Helsinki), and the protocol of human
research was approved by the local ethics committee. Prior to
fMRI data acquisition, participants gave their informed written
consent.
Stimuli, Task, and Experimental Design
Subjects viewed a 12.51 wide clock face, similar to a European
Union flag, that was displayed in the center of a rear-projection
screen near their feet. The letter ‘‘G’’ or ‘‘S’’ was positioned
above the 12 o’clock point to indicate whether, for the current
block of eight trials, the clock was good (G5 ‘‘Gut’’) or bad
(S5 ‘‘Schlecht’’). A large, wide, short, white arrow centered
within the 12-point clock face constituted the clock hand. The
imperative for the two-choice RT task was a brief change in the
color of the arrow from white to either red or green. Participants
responded to red with a left-hand key press and to green with a
right-hand press.
The cutaneous accessory stimuli were produced by two com-
mercially available Braille stimulators (Metec AG, Stuttgart)
strapped onto the arches of the participant’s left and right bare
feet. The stimulating element consists of a 2 � 4 matrix of blunt
plastic pins, each generating a force of about 5 mN. The upper
four and lower four pins were extended in alternation at a rate of
10 Hz for a duration of 600 ms, beginning 100 ms prior to im-
perative onset. The left and right feet were stimulated simulta-
neously. As noted in the Introduction, the sensation was at first
startling and tickling. These characteristics habituated quickly,
but the accessory stimuli retained a high degree of salience
throughout the approximately 50-min-long experiment.
A block of eight trials began with the hand at the 12 o’clock
position. The speed of movement varied randomly from trial to
trial among four possible values: 1200, 1800, 2400, and 3000 ms
per clock cycle. Each speed occurred twice per block. In the good
clock condition, the hand always stopped and changed color at
the same position. This position stayed the same for any given
subject but, across subjects, it varied among the 2:00, 4:00, 8:00,
and 10:00 positions. In the bad clock condition, the imperative
could occur at any of the 12 hour positions. When the motion of
the hand stopped, the color changed to either red or green for 500
ms. The clock hand remained stationary for another 500 ms, and
then resumed its course. When it reached the 12:00 position, a
new trial began. Each block lasted 24.8 s in total.
These experimental blocks of eight trials were separated by
no-task control blocks, 20 s in length, to allow hemodynamic
activity to return to baseline. The clock was still displayed,
but the hand remained motionless at 12:00. Participants rested
with their eyes open and fixated on the clock hand during these
no-task blocks.
Therewere four types of trial blocks: good clock, no accessory
(GCnA); good clock with accessory (GCA); bad clock, no ac-
cessory (BCnA), and bad clock with accessory (BCA). The ex-
periment consisted of two 20-min-long sessions, each with 24
experimental blocks. The sessions were separated by a 2-min
break.
The order of conditions was counterbalanced both within and
across subjects. For one group of participants the order of con-
ditions for the initial third of the first session was ABCDDCBA
(letter designations are arbitrary). This sequence of eight blocks
was repeated two more times to complete the session. The eight-
block sequences employed in the second session followed the
reverse order, DCBA ABCD. This scheme was used for all sub-
jects, but the initial four conditions used to create the double-
mirror pattern varied across four groups of four participants
according to a Latin square. There were 384 total trials per sub-
ject, comprising 96 replicates of each of the four conditions.
Recording and Analyses
Participants lay supine in a 1.5-Tesla MRI scanner (Siemens Av-
anto), viewing the clock display through amirrormounted on the
head coil. Their heads were immobilized with cushions to min-
imize movements. A T1-weighted structural image was used for
anatomical reference (MPRAGE, TI5 660 ms, TE5 3.93 ms,
TR5 1300 ms, flip angle5 151, matrix size5 256 � 256, 176
axial slices, voxel size5 1 � 1 � 1 mm3). Blood oxygenation
level-dependent (BOLD) signals were acquired using echo-pla-
nar imaging (EPI) across the entire brain in 36 transverse slices
(TE5 40 ms, TR5 3.0 s, flip angle5 901, matrix size5 64 � 64,
field of view5 192 mm, voxel size5 3 � 3 � 3.2 mm3, 0.8-mm
gaps between slices).
In addition to the 2 � 2 analyses with clock and accessory as
factors, two planned comparisons were employed. To minimize
the predictive value of the foot vibration, accessory and no-ac-
cessory blocks were compared only for good clock conditions.
Specifically, Student t tests comparing GCA minus GCnA were
used to identify ‘‘activations,’’ whereas the reverse comparisons,
GCnA–GCA, detected ‘‘deactivations.’’ Similarly, expectancy
was evaluated by comparing good versus bad clock conditions
only for trials in which the accessory was omitted. This would
rule out the possibility that subjects might wait for the vibration
and then shift attention to the clock hand. The relevant t tests can
be described as GCnA–BCnA and the reverse, BCnA–GCnA.
Data were analyzed with the Statistical Parameter Mapping
software package (SPM2, Wellcome Department of Imaging
Neuroscience, London). Each individual scan was realigned to
the first scan of the first session to correct for movement artifacts.
Normalization of brain size, shape, and orientation was per-
formed using the SPM template image (3 � 3 � 3 mm3). The
EPI data were smoothed with a Gaussian filter of 12 mm.
The experiment was designed for a blocked analysis in that
each block of trials contained only one of the four types of trials,
Decomposition of warning effects 165
the trials were densely packed, and there was little event jittering.
This was the primary statistical method, but, as noted earlier, it
was supplemented with event-related analyses time-locked to
imperative onset and, separately, to the subject’s key press. These
exploratory analyses were intended to help determine which
processes within the S-R pathway weremodulated by expectancy
and arousal. The blocked analyses, by contrast, were assumed to
be sensitive to activity during the preparatory (clock count-
down), S-R, and intertrial intervals.
For second-level group statistics, t tests were used with a
general height threshold of t5 3.73 (p5 .001) and a cluster ex-
tent threshold of k5 5 adjacent voxels. The exceptions were two
preliminary analyses that used higher thresholds. A blocked
analysis compared each of the four experimental conditions to
the no-task, rest interval, in order to give a general idea of the
brain regions involved in task performance. Also, an event-re-
lated analysis compared left- to right-hand key presses so as to
localize motor areas that might appear in the main analyses. For
ease of comparison with other studies, t scores have been trans-
formed into z values.
Results
Behavioral Data
TheRTanalysiswas restricted to trialswith latencies between 200
and 1000 ms, which resulted in the exclusion of 2.7% of the data.
Erroneous responses, which averaged 4.1%, were also excluded
from the 2 � 2 analysis of variance. Confirming the effectiveness
of our manipulations, both factors yielded significant main
effects, F(1,15)5 84.9 and 43.3, for clock type and accessory
presence, respectively, ps o.0001. The differences in means un-
derlying these two effects averaged 42ms and 22ms, respectively.
A significant interaction reflected the fact that the accessory effect
on RT was larger under conditions of temporal uncertainty,
presumably because the vibration provided a useful temporal
cue, F(1,15)5 5.5, p5 .033; Mean GCA5 420, GCnA5 436,
BCA5 456, BCnA5 485 ms. This supports the a priori decision
to use a planned comparison limited to the good clock trials for
evaluating phasic arousal effects on the BOLD response.
Error analyses yielded no significant results. However, con-
sistent with previous findings mentioned in the Introduction, the
accessory tended to trigger a speed–accuracy trade-off,
F(1,15)5 2.6, p5 .13. The behavioral results have been repli-
cated in a follow-up study involving healthy older adults and
patients with Parkinson’s disease (Lee, Valle-Inclan, Rolan, &
Hackley, 2008).
General Task-Related Brain Activity
Activation maps comparing bad clock, no accessory blocks to
the no-task control condition are shown in Figure 1, left panel,
and Table 1a. In Figure 1, increases in BOLD signal relative to
the no-task control condition are portrayed as red, decreases, as
blue. The findings document a pattern of regional brain activity
that is typical for visual choice-RTtasks, with increased signal for
visual, motor, and executive control regions. To isolate move-
ment-related activity, an event-related analysis was performed
comparing left- and right-hand trials, collapsing across all other
conditions. These results are shown in the right column of Figure
1, with significant clusters listed in Table 1b.
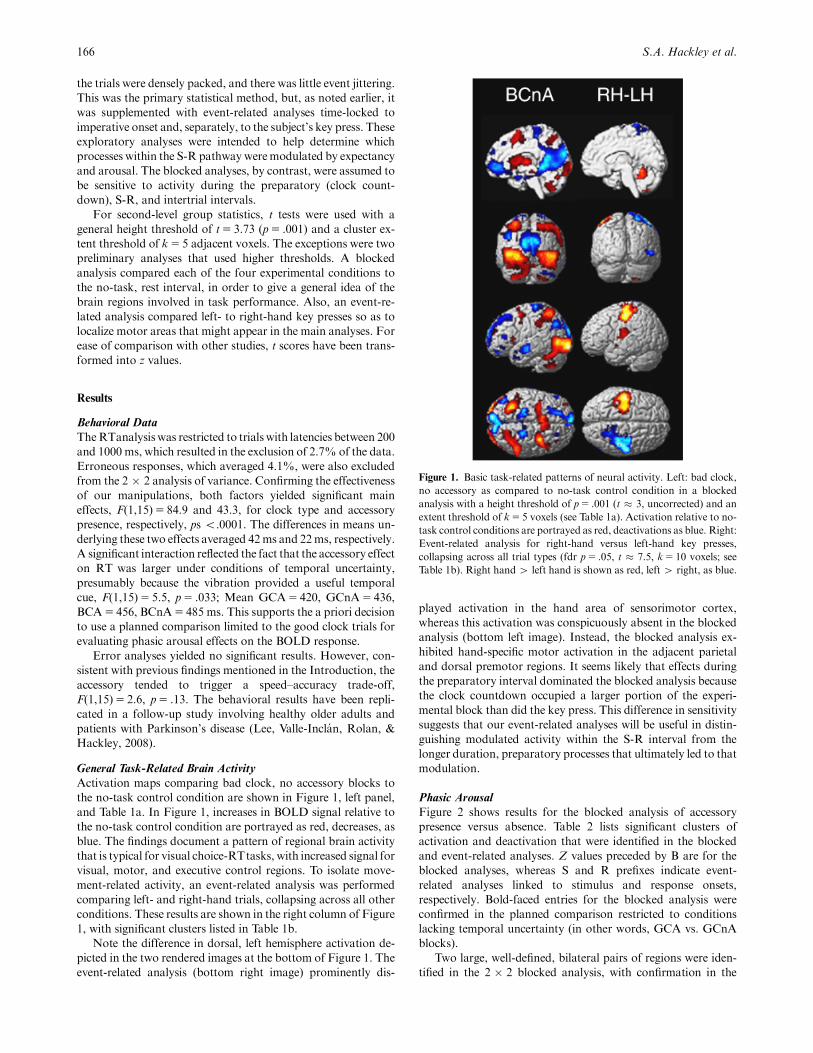
Note the difference in dorsal, left hemisphere activation de-
picted in the two rendered images at the bottom of Figure 1. The
event-related analysis (bottom right image) prominently dis-
played activation in the hand area of sensorimotor cortex,
whereas this activation was conspicuously absent in the blocked
analysis (bottom left image). Instead, the blocked analysis ex-
hibited hand-specific motor activation in the adjacent parietal
and dorsal premotor regions. It seems likely that effects during
the preparatory interval dominated the blocked analysis because
the clock countdown occupied a larger portion of the experi-
mental block than did the key press. This difference in sensitivity
suggests that our event-related analyses will be useful in distin-
guishing modulated activity within the S-R interval from the
longer duration, preparatory processes that ultimately led to that
modulation.
Phasic Arousal
Figure 2 shows results for the blocked analysis of accessory
presence versus absence. Table 2 lists significant clusters of
activation and deactivation that were identified in the blocked
and event-related analyses. Z values preceded by B are for the
blocked analyses, whereas S and R prefixes indicate event-
related analyses linked to stimulus and response onsets,
respectively. Bold-faced entries for the blocked analysis were
confirmed in the planned comparison restricted to conditions
lacking temporal uncertainty (in other words, GCA vs. GCnA
blocks).
Two large, well-defined, bilateral pairs of regions were iden-
tified in the 2 � 2 blocked analysis, with confirmation in the
166 S.A. Hackley et al.
Figure 1. Basic task-related patterns of neural activity. Left: bad clock,
no accessory as compared to no-task control condition in a blocked
analysis with a height threshold of p5 .001 (t � 3, uncorrected) and an
extent threshold of k5 5 voxels (see Table 1a). Activation relative to no-
task control conditions are portrayed as red, deactivations as blue. Right:
Event-related analysis for right-hand versus left-hand key presses,
collapsing across all trial types (fdr p5 .05, t � 7.5, k5 10 voxels; see
Table 1b). Right hand4 left hand is shown as red, left 4 right, as blue.

planned comparison. The strongest of these reflected bilateral
increases of BOLD signal within secondary somatosensory
cortex (S-II, which lies along the upper rear, inner surface of
the lateral fissure) and adjacent motor structures, the supramar-
ginal gyrus (SMG) and Rolandic operculum (the gyrus sur-
rounding the inferolateral terminus of the central sulcus). The
second most intense signal originated in the left and right middle
insulae (emotion-sensitive areas) and in the overlying superior
temporal gyrus. In addition, an activation was observed in the
foot area of primary somatosensory cortex (left mesial parietal
area), as would be expected for our cutaneous accessory stimulus.
Significant deactivation for blocks of trials with accessory stim-
ulation relative to those without was observed bilaterally in the
parahippocampal/fusiform region (not illustrated).
Event-related analysis of the S-R interval (‘‘S’’ and ‘‘R’’
entries) generated a similar pattern of results. However, an
additional activation was observed in the right superior/middle
frontal gyrus near the frontal pole, and a deactivation was noted
in the right angular gyrus.
Expectancy
In contrast to accessory stimulation, the good clock manipula-
tion produced clear-cut evidence for enhanced activity within
hand area of primary sensorimotor cortex (see Figure 3 and Ta-
ble 3). This confirms findings from previous neuroimaging stud-
ies that employed other methods, principally the valid/invalid
precue paradigm (e.g., Coull & Nobre, 1998). Increased signal
was also observed in the right superior/middle frontal gyrus, near
Decomposition of warning effects 167
Table 1a. Basic Task-Related, Brain Activity Patterns: Bad Clock, No-Accessory Blocks Compared to No-Task
Control Blocks
Brain region Coordinates Analysis/Z score
Bad clock, no accessory 4 no task (Figure 1, left)L & R Putamen/Caudate (� 24, 15, 12) B5.45, R5.42
(21, 9, 6) R5.03L Mid/Sup Frontal, Precentral G (� 27, 0, 51) B5.29, R5.87L & R Supp Motor Area, L Sup Frontal G (� 12, 9, 57) B5.15, R5.80R Mid Frontal, Precentral G (33, 0, 54) B5.95, R6.22L Sup/Inf Parietal Cx (� 33, � 54, 54) B5.58, R5.64L Mid/Inf Occipital G, L Lingual, Calcarine G (� 27, � 99, � 9) B6.11, R5.90R Mid/Inf Occip G, R Inf/Mid Temp, Fusiform G (42, � 81, � 6) B5.04, R5.15L & R Cerebellum (� 33, � 54, � 30) B4.95, R5.57
(30, � 57, � 36) R5.40L Thalamus (� 18, � 12, 3) R5.42L Mid/Ant Insula (� 24, 21, 6) R5.42
No task 4 bad clock, no accessory (Figure 1, left)L & R Sup/Med/Orb Frontal G, R Ant Cingulate, L Rectus G (3, 57, � 3) B5.05, R5.42L & R Cuneus, Precuneus, Calcarine, Sup Occip, Parahipp, Hipp G. (� 12, � 99, 24) B5.07, R5.77
(� 9, � 60, 18) B4.94, R5.77L Angular, Mid Occip G (� 48, � 75, 39) R5.07L Mid/Sup Temp G (� 66, � 9, � 12) R4.88
Table 1b. Basic Task-Related, Brain Activity Patterns: Hand-Specific Motor Activity Identified via Event-Related Analysis of Key Press
Responses
Brain region Coordinates Analysis/Z score
Hand-specific motor activity (R hand 4 L hand; red in Figure 1, right)L Pre/Postcentral G (hand) Inf Parietal Lobule (� 54, � 27, 60) R4.68L Rolandic Operc, Insula, Postcentral, Sup Temporal, Supramarginal, Heschls G (� 45, � 21, 21) R4.23L Putamen/Pallidum, Mid/Posterior Insula (� 39, � 6, � 9) R4.12R Cerebellum, Lingual, Fusiform G. (12, � 54, � 9) R4.45
Hand-specific motor activity (L hand 4 R hand; blue in Figure 1, right)R Pre/Postcentral G (hand) Sup/Mid Frontal, Supramarginal G. (60, � 18, 42) R4.80R Rolandic Operc, Insula, Sup Temporal, Inf Parietal G Heschls G, Putamen (45, � 27, 18) R4.44L Cerebellum (� 18, � 54, � 24) R3.74R Inf/ Mid Temporal, Mid Occip G (54, � 66, 0) R3.97L Sup/Mes Frontal G, Ant Cing Cx (� 12, 48, 9) R3.38
Note. Coordinates followMontreal Neurological Institute standardswith lateral, anterior, and axial distance from the anterior commissure to the clustercenter reported inmillimeters. Under the heading Analysis, the z score for the most significant voxel within that brain region is listed preceded by a letterdesignating the analysis: The letter B indicates the blocked analysis; R signifies the event-related analysis of key press responses (stimulus-locked analyseswere not included). If a given brain region was significant in more than one analysis, then coordinates are given for the leftmost analysis reachingsignificance (e.g., if both B andRwere significant, then the coordinates for B are given). For the entries in Table 1a, a height threshold of p5 .05 (family-wise error; Student’s t � 3.67) and an extent threshold of k5 5 voxels were employed. The entries in Table 1b are corrected formultiple comparisons at aFalse Discovery Rate of p5 .05 (t5 7.5) and an extent threshold of k5 10 voxels. The vertical ordering of entries is intended to convey groupings acrossthe B and R analyses, not effect size or anatomical arrangement. Ant5 anterior, Cing5 cingulated, Cx5 cortex, G5 gyrus, Hipp5hippocampus,Inf5 inferior, L5 left, Mid5middle, Mes5mesial, Occip5 occipital, Operc5operculum, Orb5orbital, R5 right, SI5primary somatosensorycortex, Sup5 superior, Temp5 temporal.

the frontal pole. This right frontal area also achieved significance
in the event-related analysis, as did a region in the left, middle
temporal gyrus.
The bad clock condition was more demanding in that subjects
had to continuously monitor the clock hand in order to rapidly
detect the unpredictable change in color. As can be seen in Figure
3, this perceptual activity generated an extended region of BOLD
signal in the left occipital and ventral temporal regions. Only the
left occipital cluster retained significance when the analysis was
restricted to conditions in which there was no accessory to use as
a temporal cue (i.e., the planned comparison BCnA–GCnA).
Several additional regions were identified in the event-related
analyses (Table 3). The caudate and cerebellum were among
those whose activity was time-locked to the imperative stimulus.
As noted in the Introduction, these structures have been impli-
cated in tasks for which internal time keeping is required. Greater
BOLD signal in the BCnA as compared to the GCnA condition
could indicate suppression when temporal uncertainty is elimi-
nated by external cues.
Discussion
The results suggest that distinct patterns of regional brain activity
underlie the expectancy and phasic arousal processes that underlie
warning effects. There was, in fact, little overlap. Only the right
frontal polar area (broadly defined) and left middle/superior tem-
poral gyrus showed significant variation with both the clock and
accessory manipulations (cf. Tables 2 and 3). Evidence for a right
frontal polar activation is congruent with prior studies showing
that this region is involved in holding an intention to act online
during a delay (Burgess, Scott, & Frith, 2003). The left middle/
superior temporal region is generally regarded as a language area.
Hence, it might have been involved in verbal aspects of task per-
formance (e.g., ‘‘Press left for red when it gets to 4:00’’; ‘‘Ignore
the buzzer and try not to jerk’’). Interactions between clock type
and accessory presence in the left anterior insula and right cer-
ebellum (Table 2) might indicate regions of shared influence.
However, these interactions could equally well reflect use of the
accessory as a time cue on bad clock trials, an anticipated but
undesired strategy thatwas dealt with using planned comparisons.
Phasic Arousal
The regions found to be specific to phasic arousal were well de-
fined and consistent with prior research. The most prominent
was the supramarginal gyrus and nearby sensorimotor areas such
as S-II. It has long been known that lesions of the supramarginal
gyrus cause apraxia and deficits in haptic perception (Brown,
1972). Interestingly, a neuroimaging experiment has shown that
this region is particularly involved in motor memory (Russ,
Mack, Grama, Lanfermann, & Knopf, 2003). During the study
phase participants either enacted a movement (such as cutting
168 S.A. Hackley et al.
Figure 2. Effects of accessory stimulation on regional hemodynamic
activity (see Table 2). These images show activity with po.001, k � 5
voxels, in the blocked analysis.
Table 2. Regional Brain Activity Related to Phasic Arousal Triggered by a Task-Irrelevant Accessory Stimulus
Brain region Coordinates Analysis
Accessory 4 no accessory (Figure 2)L & R Supramarginal, Postcentral, Sup Temp G, Rolandic Operc (� 57, � 24, 21) B4.46, S3.62, R3.34
(51, � 30, 24) B4.36, S3.39, R3.29L & R Mid Insula, Sup Temp G (� 36, 0, � 12) B4.40, S3.87, R4.03
(39, � 9, � 6) B4.02R Sup/Mid Frontal G (30, 45, 21) R3.43R Rolandic Operculum, Sup Temp G (69, 6, 6) B3.86L Postcentral G (foot SI) (� 12, � 45, 75) B4.08
No accessory 4 accessory (not illustrated)L & R Parahipp, R Lingual, L & R Fusiform, L Hipp (30, � 48, � 9) B3.35, S3.29, R3.19
(� 27, � 18, � 21) B3.30L Mid Occip (� 39, � 93, 15) B4.00R Inf Occip (33, � 87, � 9) B3.39L Postcentral G (� 57, � 3, 45) B3.86R Precentral G (42, � 15, 66) B3.23R Angular Gyrus (57, � 66, 42) S3.50, R3.34
Accessory � Clock (not illustrated)L Ant Insula (� 30, 24, 6) B3.30L Cerebellum (� 45, � 60, � 33) B3.48
Note. The entries in this table meet a height criterion of p5 .001 (uncorrected, t5 3.73, z5 3.09) and cluster extent criterion of k5 5 voxels. Types ofanalyses (B5blocked, S5 imperative stimulus, R5key press response) are described in the text. Bold face entries for the blocked analysis indicate thatthe cluster was also significant for a planned comparison restricted to conditions in which participants were unlikely to use the accessory as a time cue(GCA–GCnA and GCnA–GCA). Activations that were significant in the blocked, planned comparison, but not in the main analysis, are not listed. SeeTable 1 footnote for abbreviations.
bread) or read aloud a verbal description of a movement. When
scanned during the recall phase, greater activation of the SMG
was observed for previously enacted movements.
The association of this motor area with phasic arousal has
also been previously observed. Asmentioned in the Introduction,
an earlier study at this laboratory showed that SMG activation
during the viewing of affective pictures was correlated with both
electrodermal and verbal report measures of arousal (Anders
et al., 2004). Of even greater relevance is an accessory stimulus
experiment by Coull, Jones, and colleagues (2004): Comparing
BOLD signal in a visual discrimination task with and without
task-irrelevant acoustic stimuli, they found that accessory-in-
duced activation centered in or just above the supramarginal
gyrus (BA 40; � 57, � 48, 39) was enhance when participants
were administered an a2 noradrenergic agonist, dexmedotomi-
dine. This change in SMG activation was significantly correlated
with the performance effects of accessory stimulation.
The second most prominent activation in the accessory-ver-
sus-no accessory contrast was centered on the lower, middle in-
sulae (� 36, 0, � 12, and 39, � 9, � 6). Consistent with this is a
finding from the 2001 study by Coull and colleagues in which
activation of the left anterior insula (� 51, 18, � 3) during tem-
poral orienting was reduced by noradrenergic blockade. Simi-
larly, Nagai and coworkers (2004) observed that BOLD signals
from the left and right mid/anterior insulae (42, 6, 0) were cor-
related with both electrodermal and contingent negative varia-
tion (CNV) amplitudes during a warned RT task. These phasic
arousal effects are congruent with the well-established role in-
sular cortex plays in emotional processes (e.g., Anders et al.,
2004) and autonomic arousal (Critchley, Mathias, et al., 2002;
Critchley, Melmud, et al., 2002).
Based on amplitude modulations of the early, anterior por-
tion of the lateralized readiness potential (Hackley & Valle-
Inclan, 1999) and on induced speed–accuracy trade-offs (Posner
et al., 1973), it was predicted (Hackley & Valle-Inclan, 2003,
following Posner, Nissen, & Klein, 1976) that the locus of
accessory effects within the S-R pathway would lie within struc-
tures involved in decision making, specifically, response selec-
tion. Although the supplementary motor area (SMA), anterior
cingulate cortex (ACC), and dorsal premotor areas were acti-
Decomposition of warning effects 169
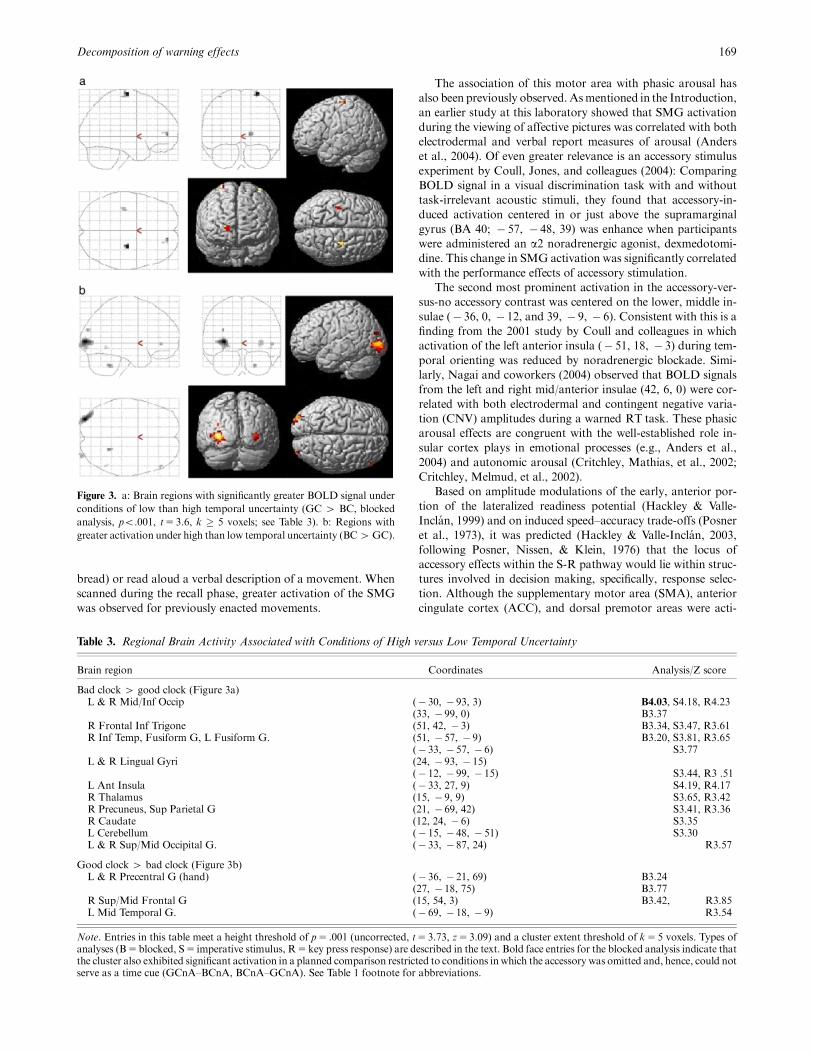
Figure 3. a: Brain regions with significantly greater BOLD signal under
conditions of low than high temporal uncertainty (GC 4 BC, blocked
analysis, po.001, t5 3.6, k � 5 voxels; see Table 3). b: Regions with
greater activation under high than low temporal uncertainty (BC4GC).
Table 3. Regional Brain Activity Associated with Conditions of High versus Low Temporal Uncertainty
Brain region Coordinates Analysis/Z score
Bad clock 4 good clock (Figure 3a)L & R Mid/Inf Occip (� 30, � 93, 3) B4.03, S4.18, R4.23
(33, � 99, 0) B3.37R Frontal Inf Trigone (51, 42, � 3) B3.34, S3.47, R3.61R Inf Temp, Fusiform G, L Fusiform G. (51, � 57, � 9) B3.20, S3.81, R3.65
(� 33, � 57, � 6) S3.77L & R Lingual Gyri (24, � 93, � 15)
(� 12, � 99, � 15) S3.44, R3 .51L Ant Insula (� 33, 27, 9) S4.19, R4.17R Thalamus (15, � 9, 9) S3.65, R3.42R Precuneus, Sup Parietal G (21, � 69, 42) S3.41, R3.36R Caudate (12, 24, � 6) S3.35L Cerebellum (� 15, � 48, � 51) S3.30L & R Sup/Mid Occipital G. (� 33, � 87, 24) R3.57
Good clock 4 bad clock (Figure 3b)L & R Precentral G (hand) (� 36, � 21, 69) B3.24
(27, � 18, 75) B3.77R Sup/Mid Frontal G (15, 54, 3) B3.42, R3.85L Mid Temporal G. (� 69, � 18, � 9) R3.54
Note. Entries in this table meet a height threshold of p5 .001 (uncorrected, t5 3.73, z5 3.09) and a cluster extent threshold of k5 5 voxels. Types ofanalyses (B5blocked, S5 imperative stimulus, R5key press response) are described in the text. Bold face entries for the blocked analysis indicate thatthe cluster also exhibited significant activation in a planned comparison restricted to conditions inwhich the accessory was omitted and, hence, could notserve as a time cue (GCnA–BCnA, BCnA–GCnA). See Table 1 footnote for abbreviations.

vated during the present task (Figure 1; Table 1a), the signal from
these regions did not vary as a function of accessory presence/
absence (Figure 2; Table 2). Instead, the data point toward SMG
as the most likely site for accessory effects on RT.
Decision making in a choice RTtask also involves perceptual
discrimination, but enhanced activity in visual cortex was not
observed. The present failure to identify any visual areas as loci
for the accessory effect might be due to the fact that our stimuli
were designed to minimize cross-modal interactions: If the cu-
taneous accessory and visual imperative had been consistently
linked in space and time so as to form a meaningful perceptual
object, evidence for cross-modal sensory integration might have
been obtained.
Taken as a whole, the data are consistent with the theory of
accessory stimulus effects originally proposed by Posner and
colleagues in 1976: ‘‘Rather than increasing the discriminability
of the imperative stimulus, the accessory causes the subject to
respond sooner to the information building up in his memory
system. The earlier the response, the less accurate it is’’ (p. 161).
The present data in conjunction with those of Russ, Coull, and
their respective colleagues (Coull, Jones, et al., 2004; Russ et al.,
2003) indicate where this memory system liesFin the supra-
marginal gyrus.
Expectancy
As noted in the Results section, the regions that were more
strongly engaged in the good than bad clock conditions were
limited to an executive control structure, the right frontal pole,
a language-related area, the left middle temporal gyrus, and the
left and right hand areas of sensorimotor cortex. Our previous
studies using chronometric methods (Bausenhart et al., 2006;
Hackley et al., 2007; Muller-Gethmann et al., 2003) led us to
conclude that the source of the RT effect lies prior to hand-
specific activation within sensorimotor cortex. By contrast, the
present study did document modulation of this region, consistent
with prior research (e.g., Coull & Nobre, 1998). This diverging
pattern of results for time and intensitymeasures is reminiscent of
the finding that the latency of a voluntary reaction can vary
independently from its force (Mattes & Ulrich, 1997; Miller,
Franz, & Ulrich, 1999).
Based on evoked potential and psychophysics research (e.g.,
Correa et al., 2006; Hackley et al., 2007; Rolke & Hofmann,
2007), we predicted that processing in late visual areas would be
sensitive to temporal expectancy. The blocked analysis docu-
mented extensive activation of visual areas during bad clock
trials, due to the need to continuously monitor the arrow. Effects
were also noted in the event-related analysis, possibly reflecting a
preset bias within the S-R pathway. For example, the ventral
occipito-temporal area, which varies parametrically as a function
of attention to color (Coull, Vidal, Nazarian, & Macar, 2004),
produced significant BOLD signal in both of the blocked event-
related analyses (Table 3).
Two brain structures that are widely recognized (e.g., Mattell
& Meck, 2004) for their involvement in time estimation, the cer-
ebellum and striatum, exhibited reduced activation when tem-
poral uncertainty was eliminated (Table 3). These structures have
other functions, of course, but the fact that their suppression was
time-locked to imperative onset rather than key press is sugges-
tive of a nonmotoric role. Also relevant in this context is the
conspicuous absence of modulation within the left intraparietal
sulcus, a cortical region that has been repeatedly associated with
temporal orienting by Coull and Nobre (e.g., 1998). These find-
ings highlight the difference between tracking time by means of
external rather than internal cues.
Conclusion
The goal of this study was to distinguish the neuronatomical
correlates of two general mechanisms underlying warning
effectsFphasic arousal and temporal expectancy. With regard
to structures within the S-R pathway that are modulated by
phasic arousal, the evidence suggested that a motor memory
area, SMG, is a likely candidate. With regard to temporal ex-
pectancy, the most likely site of modulation appeared to be mo-
tor cortex (e.g., hand area ofM1). As discussed above, the motor
cortex finding must be reconciled with chronometric evidence
that the speed of late motor processes is little affected by warning
signals.
The source of modulatory, top-down signals for both arousal
and expectancy appeared to include the right frontal pole (an
executive region responsible for holding online an intention to
act) and the left middle and superior temporal gyri (which might
maintain verbal representations of task instructions). An addi-
tional source for phasic arousal may be the anterior/middle in-
sula, a region implicated in emotional processes. Parallel studies
using neuroimaging and traditional psychophysiological mea-
sures would be useful for testing these possibilities.
REFERENCES
Anders, S., Lotze, M., Erb, M., Grodd, W., & Birbaumer, N. (2004).Brain activity underlying emotional valence and arousal: A response-related fMRI study. Human Brain Mapping, 23, 200–209.
Bausenhart, K. M., Rolke, B., Hackley, S. A., & Ulrich, R. (2006). Thelocus of temporal preparation effects: Evidence from the psycholog-ical refractory period paradigm. Psychonomic Bulletin & Review, 13,536–542.
Bernstein, I. H., Rose, R., & Ashe, V. (1970). Preparatory state effects inintersensory facilitation. Psychonomic Science, 19, 113–114.
Brown, J. W. (1972). Aphasia, apraxia, and agnosia: Clinical and theo-retical aspects. Springfield, IL: Charles C. Thomas.
Burgess, P. W., Scott, S. K., & Frith, C. D. (2003). The role of rostralfrontal cortex (area 10) in prospective memory: A lateral versus me-dial dissociation. Neuropsychologia, 41, 906–918.
Correa, A., Lupianez, J., Madrid, E., & Tudela, P. (2006). Tem-poral attention enhances early visual processing: A review andnew evidence from event-related potentials. Brain Research, 1076,116–128.
Coull, J. T., Jones, M. E. P., Egan, T. D., Frith, C. D., & Maze, M.(2004). Attentional effects of noradrenaline vary with arousal level:Selective activation of thalamic pulvinar in humans. NeuroImage, 22,315–322.
Coull, J. T., & Nobre, A. C. (1998). Where and when to pay attention:The neural systems for directing attention to spatial locations and totime intervals as revealed by both PET and fMRI. Journal ofNeuroscience, 18, 7426–7435.
Coull, J. T., Nobre, A. C., & Frith, C. D. (2001). The noradrenergic a2agonist clonidine modulates behavioural and neuroanatomical cor-relates of human attentional orienting and alerting. Cerebral Cortex,11, 73–84.
Coull, J. T., Vidal, F., Nazarian, B., & Macar, F. (2004). Functionalanatomy of the attentional modulation of time estimation. Science,303, 1506–1508.
Critchley, H., Mathias, C., & Dolan, R. (2002). Fear conditioning inhumans: The influence of awareness and autonomic arousal on func-tional neuroanatomy. Neuron, 33, 653–663.
170 S.A. Hackley et al.

Critchley, H. D., Melmed, R. N., Featherstone, E., Mathias, C. J., &Dolan, R. J. (2002). Volitional control of arousal: A functional mag-netic resonance study. NeuroImage, 16, 909–919.
Fecteau, J. H., & Munoz, D. P. (2007). Warning signals influence motorprocessing. Journal of Neurophysiology, 97, 1600–1609.
Hackley, S. A., Schankin, A., Wohlschlaeger, A., & Wascher, E. (2007).Localization of temporal preparation effects via trisected reactiontime. Psychophysiology, 44, 334–338.
Hackley, S. A., & Valle-Inclan, F. (1998). Automatic alerting does notspeed late motoric processes in a reaction time task. Nature, 391,786–788.
Hackley, S. A., & Valle-Inclan, F. (1999). Accessory stimulus effects onresponse selection: Does arousal speed decision making? Journal ofCognitive Neuroscience, 11, 321–329.
Hackley, S. A., & Valle-Inclan, F. (2003). Which stages of informationprocessing are speeded by awarning signal?Biological Psychology, 64,27–45.
Hasbroucq, T.,Osman,A., Possamaı, C.A., Burle, B.,Carron, S.,Depy,D.,et al. (1999). Cortico-spinal inhibition reflects time but not event prep-aration: Neural mechanisms of preparation dissociated by transcra-nial magnetic stimulation. Acta Psychologica, 101, 243–266.
Jennings, J. R., van der Molen, M. W., & Terezis, C. (1987). Primarybradycardia and vagal inhibition as two manifestations of the timingof vagal influence on the heart beat. Journal of Psychophysiology, 1,361–374.
Lee, E. Y., Valle-Inclan, F., Rolan, T., & Hackley, S. A. (2008). Ab-normalities of the phasic arousal and temporal expectancy compo-nents of warning effects in Parkinson’s disease. Manuscript inpreparation.
Mattell, M. S., & Meck, W. H. (2004). Cortico-striatal circuits and in-terval timing: Coincidence detection of oscillatory processes. Cogni-tive Brain Research, 21, 139–170.
Mattes, S., & Ulrich, R. (1997). Response force is sensitive to the tem-poral uncertainty of response stimuli.Perception & Psychophysics, 59,1089–1097.
Miller, J., Franz, V., & Ulrich, R. (1999). Effects of auditory stimulusintensity on response force in simple, go/no-go, and choice RT tasks.Perception & Psychophysics, 61, 107–119.
Muller-Gethmann, H., Ulrich, R., & Rinkenauer, G. (2003). Locus ofthe effect of temporal resolution: Evidence from the lateralized readi-ness potential. Psychophysiology, 40, 597–611.
Nagai, Y., Critchley, H. D., Featherstone, E., Fenwick, P. B. C.,Trimble, M. R., & Dolan, R. J. (2004). Brain activity relating to thecontingent negative variation: An fMRI investigation. NeuroImage,21, 1232–1241.
Posner, M. I., Klein, R., Summers, J., & Buggie, S. (1973). On the se-lection of signals. Memory and Cognition, 1, 2–12.
Posner, M. I., Nissen, M. J., & Klein, R. M. (1976). Visual dominance:An information-processing account of its origins and significance.Psychological Review, 83, 157–171.
Requın, J., Brener, J., & Ring, C. (1991). Preparation for action. InJ. R. Jennings & M. G. H. Coles (Eds.), Handbook of cognitive psy-chophysiology: Central and autonomic nervous system approaches(pp. 357–448). New York: John Wiley.
Robbins, T. M., & Everitt, B. J. (1995). Arousal systems and attention.In M. S. Gazzaniga (Ed.), The cognitive neurosciences (pp. 243–262).Cambridge, MA: MIT Press.
Rolke, B., & Hofmann, P. (2007). Temporal uncertainty degrades per-ceptual processing. Psychonomic Bulletin and Review, 14, 522–526.
Russ, M. O., Mack, W., Grama, C. R., Lanfermann, H., & Knopf, M.(2003). Enactment effect in memory: Evidence concerning the func-tion of the supramarginal gyrus. Experimental Brain Research, 149,497–504.
Scheirs, J. G.M., & Brunia, C. H.M. (1985). Achilles tendon and surfaceEMG activity during anticipation of a significant event and prepa-ration for a voluntary movement. Journal of Motor Behavior, 17, 96–109.
Sollers, J. J. III, & Hackley, S. A. (1997). Effects of foreperiod durationon reflexive and voluntary responses to intense noise bursts. Psycho-physiology, 34, 518–526.
Stafford, I. L., & Jacobs, B. L. (1990). Noradrenergic modulation of themasseteric reflex in behaving cats. II. Physiologic studies. Journal ofNeuroscience, 10, 99–107.
Stevens, M. C., Kiehl, K. A., Pearlson, G., & Calhoun, V. D. (2007).Functional neural circuits for mental timekeeping. Human BrainMapping, 28, 394–408.
Ulrich, R., & Mattes, S. (1996). Does immediate arousal enhance re-sponse force in simple reaction time tasks? The Quarterly Journal ofExperimental Psychology, 49A, 972–990.
Welch, R. B., &Warren, D. H. (1986). Intersensory interactions. In K. R.Boff, L. Kaufman, & J. P. Thomas (Eds.), Handbook of perceptionand human performance (vol. 1, pp. 25.1–25.36). New York: Wiley.
Witte, E. A., & Marrocco, R. T. (1997). Alteration of brain nor-adrenergic activity in rhesus monkeys affects the alerting componentof covert orienting. Psychopharmacology, 132, 315–323.
Young,R.A., Cegavske, C. F., & Thompson, R. F. (1976). Tone-inducedchanges in excitability of abducens motoneurons and of the reflexpath of nictitating membrane response in rabbit. Journal of Compar-ative and Physiological Psychology, 90, 424–434.
(Received July 16, 2007; Accepted April 7, 2008)
Decomposition of warning effects 171
Related Documents