Secretion of macrophage urokinase plasminogen activator is dependent on proteoglycans Gunnar Pejler 1 , Jan-Olof Winberg 2 , Tram T. Vuong 3 , Frida Henningsson 1 , Lars Uhlin-Hansen 2 , Koji Kimata 4 and Svein O. Kolset 5 1 Department of Veterinary Medical Chemistry, Swedish University of Agricultural Sciences, Uppsala, Sweden; 2 Department of Biochemistry, Institute of Medical Biology, University of Tromsø, Norway; 3 Department of Biochemistry, University of Oslo, Norway; 4 Institute for Molecular Science of Medicine, Aichi Medical University, Japan; 5 Institute for Nutrition Research, University of Oslo, Norway The importance of proteoglycans for secretion of proteolytic enzymes was studied in the murine macrophage cell line J774. Untreated or 4b-phorbol 12-myristate 13-acetate (PMA)-stimulated macrophages were treated with hexyl-b-D-thioxyloside to interfere with the attachment of glycosaminoglycan chains to their respective protein cores. Activation of the J774 macrophages with PMA resulted in increased secretion of trypsin-like serine proteinase activity. This activity was completely inhibited by plasminogen acti- vator inhibitor 1 and by amiloride, identifying the activity as urokinase plasminogen activator (uPA). Treatment of both the unstimulated or PMA-stimulated macrophages with xyloside resulted in decreased uPA activity and Western blotting analysis revealed an almost complete absence of secreted uPA protein after xyloside treatment of either control- or PMA-treated cells. Zymography analyses with gels containing both gelatin and plasminogen confirmed these findings. The xyloside treatment did not reduce the mRNA levels for uPA, indicating that the effect was at the post-translational level. Treatment of the macrophages with xylosides did also reduce the levels of secreted matrix met- alloproteinase 9. Taken together, these findings indicate a role for proteoglycans in the secretion of uPA and MMP-9. Keywords: proteoglycan; xyloside; matrix metalloprotein- ase; urokinase; secretion. The capacity to secrete various compounds is an important property of cells in the monocytoid–macrophage lineage, in addition to the phagocytic and antigen presenting functions [1]. The secretory repertoire includes such molecules as tumor necrosis factor-a, lipoprotein lipase, proteoglycans, leukotrienes, and various proteases [2]. The proteoglycans expressed by monocytes and macrophages have been characterized to some extent. The major product seems to be serglycin, as shown by N-terminal sequencing of proteoglycans released from the cultured monocytic cell lines U937 and THP-1 [2,3]. Moreover, it has been shown that activated murine and human macrophages express syndecan-4 [4] and syndecan-2 [5], respectively, on the cell surface. The release of serglycin from monocytes and macro- phages is the subject of regulation by inflammatory signaling molecules such as interferon-c, transforming growth factor-b, and platelet derived growth factor [2,6]. It is therefore likely that the secretion of proteoglycans in these cells is linked to inflammatory reactions and that its function(s) may be linked to the binding, transport and regulation of other secretory products. Indeed, recent data indicate that mice lacking functional heparin chains attached to their serglycin proteoglycans show severe defects in their capacities to store mast cell proteases in the secretory granules [7,8], clearly demonstrating the importance of intact proteoglycans for normal storage of proteases in these cells. Serglycin proteoglycans have also been implicated in the regulation of mast cell protease activities [9–11]. The biological functions of proteoglycans from activated monocytes and macrophages have not been outlined in any detail. It has however, been shown that serglycin may be associated with chemokines and enzymes after release from the cells [12]. It has furthermore been demonstrated that serglycin may interact with CD44 [13], and possibly engage in cell interactions between immune cells. Considering that serglycin proteoglycans are of critical importance for the secretory granule proteases in mast cells it is reasonable to assume that serglycin proteoglycans may also affect proteases in other cell types. In the present study we have investigated the possible role of proteoglycans in the secretion of proteolytic enzymes by macrophages. For this purpose we made use of b-D-xylosides. These com- pounds have been widely used to study proteoglycan biosynthesis and the role of proteoglycans in different biological processes. b-D-Xylosides will compete with endogenous core protein for access to the glycosaminogly- can biosynthesis machinery [14], resulting in the biosynthesis Correspondence to S.O. Kolset, Institute for Nutrition Research, University of Oslo, Box 1046 Blindern, 0316 Oslo, Norway. Fax: + 47 2285 1398, Tel.: + 47 2285 1383, E-mail: [email protected] Abbreviations: C-ABC, chondroitinase ABC; MMP, matrix metallo- proteinase; HX-xyl, hexyl-b-D-thioxyloside; PMA, 4b-phorbol 12-myristate 13-acetate; uPA, urokinase plasminogen activator; SBTI, soy bean trypsin inhibitor; DMEM, Dulbecco’s modified Eagles medium; PAI-1, plasminogen activator inhibitor 1; tPA/uPA, tissue type/urokinase type plasminogen activators. Enzymes: chondroitinase ABC (EC 4.2.2.4) (Received 17 June 2003, accepted 7 August 2003) Eur. J. Biochem. 270, 3971–3980 (2003) Ó FEBS 2003 doi:10.1046/j.1432-1033.2003.03785.x

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Secretion of macrophage urokinase plasminogen activatoris dependent on proteoglycans
Gunnar Pejler1, Jan-Olof Winberg2, Tram T. Vuong3, Frida Henningsson1, Lars Uhlin-Hansen2,Koji Kimata4 and Svein O. Kolset5
1Department of Veterinary Medical Chemistry, Swedish University of Agricultural Sciences, Uppsala, Sweden; 2Department of
Biochemistry, Institute of Medical Biology, University of Tromsø, Norway; 3Department of Biochemistry, University of Oslo,
Norway; 4Institute for Molecular Science of Medicine, Aichi Medical University, Japan; 5Institute for Nutrition Research,
University of Oslo, Norway
The importance of proteoglycans for secretion of proteolyticenzymes was studied in the murine macrophage cell lineJ774. Untreated or 4b-phorbol 12-myristate 13-acetate(PMA)-stimulated macrophages were treated withhexyl-b-D-thioxyloside to interfere with the attachment ofglycosaminoglycan chains to their respective protein cores.Activation of the J774 macrophages with PMA resulted inincreased secretion of trypsin-like serine proteinase activity.This activity was completely inhibited by plasminogen acti-vator inhibitor 1 and by amiloride, identifying the activity asurokinase plasminogen activator (uPA). Treatment of boththe unstimulated or PMA-stimulated macrophages withxyloside resulted in decreased uPA activity and Western
blotting analysis revealed an almost complete absence ofsecreted uPA protein after xyloside treatment of eithercontrol- or PMA-treated cells. Zymography analyses withgels containing both gelatin and plasminogen confirmedthese findings. The xyloside treatment did not reduce themRNA levels for uPA, indicating that the effect was at thepost-translational level. Treatment of the macrophages withxylosides did also reduce the levels of secreted matrix met-alloproteinase 9. Taken together, these findings indicate arole for proteoglycans in the secretion of uPA andMMP-9.
Keywords: proteoglycan; xyloside; matrix metalloprotein-ase; urokinase; secretion.
The capacity to secrete various compounds is an importantproperty of cells in the monocytoid–macrophage lineage, inaddition to the phagocytic and antigen presenting functions[1]. The secretory repertoire includes such molecules astumor necrosis factor-a, lipoprotein lipase, proteoglycans,leukotrienes, and various proteases [2]. The proteoglycansexpressed by monocytes and macrophages have beencharacterized to some extent. The major product seems tobe serglycin, as shown by N-terminal sequencing ofproteoglycans released from the cultured monocytic celllines U937 and THP-1 [2,3]. Moreover, it has been shownthat activated murine and human macrophages expresssyndecan-4 [4] and syndecan-2 [5], respectively, on the cellsurface.The release of serglycin from monocytes and macro-
phages is the subject of regulation by inflammatorysignaling molecules such as interferon-c, transforming
growth factor-b, and platelet derived growth factor [2,6].It is therefore likely that the secretion of proteoglycans inthese cells is linked to inflammatory reactions and that itsfunction(s) may be linked to the binding, transport andregulation of other secretory products. Indeed, recent dataindicate that mice lacking functional heparin chainsattached to their serglycin proteoglycans show severe defectsin their capacities to storemast cell proteases in the secretorygranules [7,8], clearly demonstrating the importance ofintact proteoglycans for normal storage of proteases in thesecells. Serglycin proteoglycans have also been implicated inthe regulation of mast cell protease activities [9–11].The biological functions of proteoglycans from activated
monocytes and macrophages have not been outlined in anydetail. It has however, been shown that serglycin may beassociated with chemokines and enzymes after release fromthe cells [12]. It has furthermore been demonstrated thatserglycin may interact with CD44 [13], and possibly engagein cell interactions between immune cells.Considering that serglycin proteoglycans are of critical
importance for the secretory granule proteases in mast cellsit is reasonable to assume that serglycin proteoglycans mayalso affect proteases in other cell types. In the present studywe have investigated the possible role of proteoglycans inthe secretion of proteolytic enzymes by macrophages. Forthis purpose we made use of b-D-xylosides. These com-pounds have been widely used to study proteoglycanbiosynthesis and the role of proteoglycans in differentbiological processes. b-D-Xylosides will compete withendogenous core protein for access to the glycosaminogly-can biosynthesismachinery [14], resulting in the biosynthesis
Correspondence to S.O. Kolset, Institute for Nutrition Research,
University of Oslo, Box 1046 Blindern, 0316 Oslo, Norway.
Fax: + 47 2285 1398, Tel.: + 47 2285 1383,
E-mail: [email protected]
Abbreviations: C-ABC, chondroitinase ABC; MMP, matrix metallo-
proteinase; HX-xyl, hexyl-b-D-thioxyloside; PMA, 4b-phorbol12-myristate 13-acetate; uPA, urokinase plasminogen activator;
SBTI, soy bean trypsin inhibitor; DMEM, Dulbecco’s modified
Eagles medium; PAI-1, plasminogen activator inhibitor 1;
tPA/uPA, tissue type/urokinase type plasminogen activators.
Enzymes: chondroitinase ABC (EC 4.2.2.4)
(Received 17 June 2003, accepted 7 August 2003)
Eur. J. Biochem. 270, 3971–3980 (2003) � FEBS 2003 doi:10.1046/j.1432-1033.2003.03785.x

of free glycosaminoglycan chains attached to the b-D-xyloside rather than intact proteoglycans. Depending on theconcentration of xylosides used, endogenous proteoglycanexpression may be completely abrogated. b-D-Xylosidesseem to be more efficient in abrogating the expression ofchondroitin sulfate proteoglycans than heparan sulfateproteoglycans. Results presented here show that the treat-ment of macrophages with b-D-xylosides results in impairedsecretion of urokinase plasminogen activatior (uPA), indi-cating that uPA is dependent on proteoglycans. Thesecretion of matrix metalloproteinase 9 (MMP-9) was alsodecreased by the xyloside treatment.
Materials and methods
Materials
Sephadex G50 Fine and Superose 6 were from Amer-sham Pharmacia, Uppsala, Sweden. [35S]Sodium sulfatewas obtained from Amersham. The chromogenic peptidesubstrates S-2288 (H-D-Ile-Pro-Arg-p-nitroanilide), S-2444(pyroGlu-Gly-Arg-p-nitroanilide), S-2390 (H-D-Val-Phe-Lys-p-nitroanilide) and S-2586 (MeO-Suc-Arg-Pro-Tyr-p-nitroanilide) were from Chromogenix, Molndal, Sweden.S-2288 is a general substrate for trypsin-like serineproteinases, whereas S-2444 and S-2390 are relativelyspecific substrates for plasminogen activators and plas-min, respectively. S-2586 is a substrate for chymotrypsin-like serine proteinases. Hexyl-b-D-thioxyloside (HX-xyl)was used as described previously. This particular xylosidewas shown to be one of the most efficient abrogatorsof proteoglycan biosynthesis in comparison withother xylosides [14,15]. Chondroitinase ABC (C-ABC,EC 4.2.2.4) was bought from Seikagaku Kogyo Co.,Tokyo, Japan. Amiloride, soy bean trypsin inhibitor(SBTI), phenylmethanesulfonyl fluoride and gelatin wereobtained from Sigma Chemical Co. Plasminogen, humanplasminogen activator inhibitor 1 (PAI-1), a1-anti-chymo-trypsin, a1-protease inhibitor were from Calbiochem-Novabiochem.
Cells
The murine macrophage cell line, J774 A1 (hereafter calledJ774), was from the American Type Culture Collection,Rockville, MD, USA. The cells were routinely kept inDulbecco’s modified Eagles medium (DMEM) with 2 mM
L-glutamine and gentamycin (0.1 mgÆmL)1), all from BioWhittaker, Verviers, Belgium. The medium was fortifiedwith 10% fetal bovine serum from Sigma Chemical Co.The human histiocytic lymphoma cell line U937 clone 1(U937-1) was cultured in RPMI medium with 10% fetalbovine serum, 2 mM L-glutamine and gentamycin(0.1 mgÆmL)1), all from Bio Whittaker.
Enzyme assays
J774 cells were established in medium with serum in16 mm wells at cell densities between 0.5 and 1.0 · 106 cellsper well, or in 96-well plates at densities of approximately1.5 · 105 cells per well. After reaching confluency, J774cells were washed three times in medium without supple-
ments to remove serum proteins. The cells were thereaftercultured in the serum-free medium QBSF 51 (Sigma). Cellswere incubated with or without 50 ngÆmL)1 of PMA in theabsence or presence of 0.1–2.0 mM HX-xyl. No differencein cell numbers could be measured after the differenttreatments by cell counting after 24 h incubation in serumfree media. Maximum effect on the abrogation of proteo-glycan biosynthesis was observed at the 2 mM concentra-tion. This concentration was used in studies on enzymesecretion. After 20 h the conditioned media were harvested,centrifuged to remove nonadherent cells and frozen beforefurther analyses. Media to be used for zymographyanalyses were frozen after adding Hepes buffer pH 7.4and CaCl2 to final concentrations of 0.1 M and 10 mM,respectively.Trypsin-like activities were measured in the recovered
conditioned media. 50–100 lL conditioned medium wasadded to wells of 96-well microtiter plates followed by theaddition of 100–150 lL of NaCl/Pi (200 lL final volume)and 20 lL of either substrate S-2288 or S-2444, dissolvedin distilled water at stock concentrations of 20 mM. Theenzyme activities were recorded by reading the absorbanceat 405 nm at different time points using a TitertekMultiscan spectrophotometer (Flow Laboratories, Irvine,Scotland). The increase in absorbance showed linearkinetics over a time period of 5 h, indicating that theenzyme was stable for at least this period of time insolution.For inhibition studies, 50 lL of conditioned medium was
mixed with 150 lL of NaCl/Pi in 96-well plates. Next, eitherof the following protease inhibitors was added at a finalconcentration of 0.2 lM: PAI-1, a1-anti-chymotrypsin,a1-protease inhibitor or soybean trypsin inhibitor. Theeffect of phenylmethanesulfonyl fluoride at a final con-centration of 1 mM was also tested. After 30 min ofincubation, 20 lL of S-2288 (20 mM in H2O) was addedfollowed by monitoring of residual trypsin-like activity.The effect of amiloride was tested in a similar fashion.50 lL of conditioned medium was mixed with 150 lL ofNaCl/Pi and with amiloride at 0.001–10 mM final con-centration (amiloride was diluted from a 100-mM stocksolution in dimethylsulfoxide). Residual activity towardsS-2288 was determined after 30 min.Enzymatic determinations were performed in triplicates.
Results shown represent the mean ± SD.
Zymography
SDS/PAGE was performed as described previously [16].Gels (7.5 cm · 8.5 cm · 0.75 mm) contained 0.1% (w/v)gelatin in both the stacking and the separating gel, whichcontained 4 and 7.5% (w/v) of polyacrylamide, respectively.In some cases, the separating gel also contained plasmino-gen [16] (10 lgÆmL)1) in addition to gelatin that allowed thedetection of plasminogen activators [17]. Serum-free med-ium from the monocytic cell line THP-1 was used as astandard because it contains proMMP-9 monomer, givingrise to a main band at 92 kDa and the proMMP-9homodimer (a minor band at 225 kDa) [16]. In addition,serum-free conditioned medium from normal human skinfibroblasts [18] was used as a source for pro-MMP-2standard (72 kDa). Ten microlitres of conditioned medium
3972 G. Pejler et al. (Eur. J. Biochem. 270) � FEBS 2003

was mixed with 3 lL of loading buffer (333 mM Tris/HCl,pH 6.8, 11% SDS, 0.03% bromophenol blue and 50%glycerol). Six microlitres of this nonheated mixture wasapplied to the gel, which was run at 20 mA/gel at 4 �C.Thereafter, the gel was washed twice in 50 mL 2.5% (v/v)Triton X-100, and then incubated in 50 mL of assaybuffer (50 mM Tris/HCl, pH 7.5, 5 mM CaCl2, 0.2 M
NaCl and 0.02% Brij-35) for approximately 20 h at37 �C. In some cases 10 mM of EDTA was added to boththe washing and assay buffers to block potential metallo-proteinase activity, but not serine proteinase activity. Inother cases samples were incubated with 10 mM ofpefabloc (a serine proteinase inhibitor) for 60 min atroom temperature. Thereafter the samples were treated asdescribed above. Gels were stained with 0.2% CoomassieBrilliant Blue R-250 (30% methanol) and destained in asolution containing 30% methanol and 10% acetic acid.Gelatinase activity was evident as cleared (unstained)regions. The area of the cleared zones and Mr determin-ation of unknown bands was analyzed with the GELBASE/GELBLOT
TMPRO computer program from Ultra Violet
Products (Cambridge, UK).In some cases, the serum-free conditioned medium from
J774 cells was incubated with either 0.1 M Hepes buffer or24 lgÆmL)1 of trypsin for 15 min at 37 �C prior toelectrophoresis. Trypsin was thereafter inactivated by theaddition of 7 mgÆmL)1 of SBTI. In these experiments,0.2% of SBTI was also incorporated in both the stackingand separating gels to prevent degradation of the incor-porated gelatin substrate by trace amounts of trypsin thatmay escape from the inhibitor complex during electro-phoresis.
Western blotting
Media (5 mL) from nontreated cells (control) and cellstreated with PMA and xyloside, respectively, were concen-trated 10 times on Millipore ultrafree-15, NMWL 10 000(Biomax-10) centrifugal filter device. The concentratedsamples were mixed with SDS/PAGE sample buffer,without 2-mercaptoethanol. Cells (1 · 106) were solubilizedby adding 100 lL of SDS/PAGE sample buffer followed byboiling for 3 min. Samples (40 lL) from medium- or cellfractions were subjected to SDS/PAGE on 12% polyacryl-amide gels under reducing conditions. Proteins were subse-quently blotted onto nitrocellulose membranes, followed byblocking with 5%milk powder in NaCl/Pi for 1 h at 20 �C.Next, the membranes were incubated with antiserum(1 : 200) in 5%milk powder/Tris/NaCl/Pi/0.1% Tween 20,at 4 �C for 20 h. The rabbit anti-(mouse urokinase) Ig wasa kind gift from K. Danø, Rigshospitalet, Copenhagen,University Hospital, Denmark. After extensive washingwith Tris/NaCl/Pi/0.1% Tween 20, the membranes wereincubated with secondary Ig conjugated to horseradishperoxidase (Amersham Pharmacia Biotech; 1 : 3000 dilu-tion in TBS/0.1% Tween 20). After 45 min of incubation at20 �C, the membranes were again washed extensively withTris/NaCl/Pi/0.1% Tween 20, followed by washing withTris/NaCl/Pi without detergent. The membranes weredeveloped with the ECL system (Amersham PharmaciaBiotech) according to the protocol provided by the manu-facturer.
Transmission electron microscopy
Cells were fixed in 2% glutaraldehyde, incubated in 1%OsO4/NaCl/Pi, dehydrated and embedded in TAAB-B12resin. Sections were analyzed at 60 kV in a Philips CM10microscope and photographed.
Isolation of RNA and Northern blotting
J774 cells were lysed with Trizol and RNA was extractedwith chloroform and precipitated in isopropanol. mRNAwas isolated from the precipitate using Dynabeads witholigo dT25 magnetic beads (Dynal, Oslo Norway), andseparated on 1% agarose gels containing formaldehydeand blotted to Hybond N nylon membranes (AmershamPharmacia Biotech). After prehybridization the blots werehybridized in 0.5 M sodium phosphate buffer with 7% SDSand 1 mMEDTA and 32P-labelled probes at 65 �C for 16 h.The blots were washed three times at 65 �C with 40 mM
sodium phosphate containing 1% SDS, sealed and exposedto phosphorimage screen over night. The obtained screenswere analyzed in a phosphorimager (Molecular Dynamics,Amersham Pharmacia Biotech). Probe for murine urokin-ase was a kind gift from L. Hellman, Uppsala University. Aprobe for the housekeeping gene, 36B4, obtained fromH. Nebb, University of Oslo, was used to compare mRNAlevels in different samples.
Proteoglycan expression
To analyze the effects of PMAandHX-xyl treatment on theexpression of proteoglycans, J774 cells were labelled with[35S]sodium sulfate for 24 h. PMAandHX-xyl were presentonly during the labeling period. The media were harvestedand loose cells pelleted by centrifugation. The cell fractionswere recovered by adding 0.05 M Tris/HCl, pH 8.0 with0.15 M NaCl and 1% Triton X-100. Both medium and cellfractions were subjected to Sephadex G50 Fine gel chro-matography to remove free [35S]sulfate. The chromatograhywas performed in 0.05 M Tris/HCl, pH 8.0 with 0.15 M
NaCl and 0.1% Triton X-100. Material eluting in the voidvolume was frozen before further analyses. Both mediumand cell fractions were analysed by gel chromatographyusing a Superose 6 column (Pharmacia). Fractions of 1 mLwere collected and analysed for content of radioactivity byscintillation counting using a Wallac TriCarb scintillationcounter. [35S]Sodium sulfate samples were subjected tochondroitinase ABC treatment to depolymerize chondro-itin sulfate and deaminative cleavage using HNO2 todegrade heparan sulfate, as previously described [19].
Results
Xyloside and proteoglycan expression
To analyze the possible importance of proteoglycanexpression for the secretion of proteolytic enzyme activitiesin activated macrophages, J774 cells were treated with HX-xyl or PMA alone or with PMA and HX-xyl in combina-tion. As can be seen in Table 1, PMA treatment resulted in a50–80% increase in total proteoglycan synthesis. Further,treatment of the cells with HX-xyl, both in the presence or
� FEBS 2003 Proteoglycans and urokinase (Eur. J. Biochem. 270) 3973

absence of PMA, resulted in a marked (� threefold)increase in the synthesis of 35S-labelled macromolecules(Table 1). After HX-xyl treatment, the major part of the35S-labelled macromolecules expressed was recovered in theculture medium, regardless if PMA was present or not(Table 1). In contrast, control cells and cells treated withPMA retained a major portion of the 35S-labelled macro-molecules in the cell fraction (Table 1). 35S-labelled macro-molecules recovered from the medium fractions wereanalyzed by gel chromatography to discriminate betweenintact proteoglycans and free glycosaminoglycan chains.Further, samples were analyzed both before and aftertreatment with alkali (NaOH), a treatment that is known torelease glycosaminoglycans from their respective proteincores. In agreement with a previous study [14], treatmentwith HX-xyl resulted in a shift from synthesis of predomi-nantly intact proteoglycans to an almost exclusive synthesisof free glycosaminoglycan chains (Fig. 1). Note the com-plete shift in elution pattern after alkali treatment in theupper and third panel, showing that the 35S-labelledmacromolecules released from control and PMA-treatedcells are almost exclusively in proteoglycan form. Note alsothat the 35S-labelled macromolecules in the panels corres-ponding to HX-xyl-treated cells are resistant to alkalitreatment, demonstrating the predominance of free glycos-aminoglycan chains.Control- and PMA-treated cells secreted proteoglycans of
both chondroitin sulfate and heparan sulfate type, as shownby the partial susceptibility of the secreted 35S-labelledmacromolecules to either chondroitinase ABC or deamin-ative cleavage (HNO2), respectively (first and third panel).In contrast, cells subjected to HX-xyl treatment, in thepresence or absence of PMA, secreted predominantly freechondroitin sulfate chains. This was demonstrated by thedepolymerization of most of the medium 35S-labelledmacromolecules after treatment with chondroitinase ABC(Fig. 1; panels two and four). However, small amounts ofHSPGs can also found in the medium of these cultures.Both heparan and chondroitin sulfate proteoglycans
could be detected in the cell fractions of control- and PMA-treated cells, as well as in cells treated with HX-xyl or PMA/HX-xyl. When these fractions were analyzed by gel
chromatography, they displayed almost identical elutionprofiles (results not shown), irrespective of treatment. Theratio between heparan sulfate and chondroitin sulfate in thecell fractions was therefore not affected by the xylosidetreatment. The shift from chondroitin sulfate/heparansulfate proteoglycans to mostly free chondroitin sulfatechains is, accordingly, only seen in the medium fractionsafter HX-xyl or PMA/HX-xyl treatment.
Xyloside and serine proteinases
Conditioned medium collected after 20 h incubationunder serum-free conditions did not contain any chymo-trypsin-like activity, as no cleavage of the chromogenic
Table 1. [35S]-labelled macromolecules recovered from medium and cell
fractions of J774 cells. Cells were labelled with [35S]sodium sulfate for
20 h with the indicated treatments. [35S]-Labelled macromolecules
were recovered from cell and medium fractions and the amount
determined by scintillation counting. The results presented are the
mean values ±SD of three separate measurements. Total incorpor-
ated [35S]-radioactivity is from one experiment. Four separate experi-
ments showed the same trend.
Treatment
Percentage of [35S]-
labelled macromoleculesTotal incorporated
[35S]-radioactivity
(c.p.m.)
Cell
fraction
Medium
fraction
Control 65 ± 5 35 ± 3 265 000
PMA 60 ± 19 40 ± 6 331 000
HX-xyl 24 ± 5 76 ± 3 723 000
PMA + HX-xyl 20 ± 1 80 ± 18 748 000
Fig. 1. Superose 6 gel chromatography of medium fractions.35S-La-
belled macromolecules recovered from medium fractions of control
cells (Control), HX-xyl-treated cells (HX-xyl), PMA-treated cells
(PMA) and cells treated with PMA and HX-xyl (PMA + HX-xyl)
were subjected to Superose 6 gel chromatography. Aliquots were also
subjected to deaminative cleavage (HNO2) to degrade heparan sulfate,
chondroitinase ABC treatment to depolymerize chondroitin/dermatan
sulfate or alkali treatment to release free GAG chains and also ana-
lyzed by gel chromatography. Equal amounts of radioactivity were
taken from the different fractions for analyses by gel chromatography.
3974 G. Pejler et al. (Eur. J. Biochem. 270) � FEBS 2003

chymotrypsin substrate S-2586 was observed (result notshown). Considerable activity, however, could be detectedwhen the chromogenic substrate S-2288 was used. Thissubstrate is cleaved by enzymes with trypsin-like substratespecificities. From Fig. 2 it is evident that the secretion oftrypsin-like activity was increased approximately twofoldwhen the cells were treated with PMA.When proteoglycan expression was compromised by
treatment with HX-xyl, the levels of trypsin-like activitiesrecovered in the conditioned media were reduced both inuntreated and in PMA-stimulated cells by � 50%. Theeffects of xyloside varied somewhat between differentexperiments using different cell batches. In some experi-ments the HX-xyl treatment reduced the secretion oftrypsin-like activities to an even larger extent, both incontrol and PMA-stimulated cells (not shown). The reduc-tion in trypsin-like activity in the medium upon HX-xyltreatment was most pronounced after extended periods ofincubation. However, time course studies revealed a clearlynoticeable effect already 1 h after the addition of HX-Xyl,with a gradually increased effect up to 20 h of incubation(not shown). Furthermore, in experiments with the humanmonocytic cell line U937 the presence of trypsin-like activityin supernatants from serum-free cultures could also bedemonstrated. The activity was stimulated more thantwofold with PMA and was inhibited to a large extent withHX-xyl (not shown). Hence, secretion of trypsin-like pro-teases seems todependonproteoglycans inbothmurine J774macrophage-like cells and in humanmonocytic U937 cells.Macrophages secrete a wide range of enzymes active at
neutral pH, many of which are serine proteinases [1].However, one prominent serine proteinase in the monocyte/macrophage system is plasminogen activator (PA). Thechromogenic substrate, S-2444, (pyrGlu-Gly-Arg-pNA) isconsidered to be a relatively specific PA substrate. FromFig. 2 it is apparent that the conditioned media from theJ774 cells contained S-2444-cleaving activity, and that theactivity towards S-2444 was higher than the activity against
S-2288. Further, the S-2444-cleaving activity was stimulatedto the same extent by PMA as was the activity towardsS-2288, and HX-xyl caused similar inhibitory effects onsecretion of S-2444-hydrolyzing activity as was observed forthe secretion of activity towards S-2288. These results arethus compatible with the possibility that the cleavage ofS-2288 and S-2444 are carried out by the same enzymeactivity, and that this activitymay be related to plasminogenactivator. To characterize the activity further, conditionedmedia were incubated with various protease inhibitorsfollowed by the measurement of residual trypsin-likeactivity. The S-2288-hydrolyzing activity, both from controland PMA-stimulated cells, was completely inhibited byphenylmethanesulfonyl fluoride, demonstrating that it wasa serine proteinase. Further, the activity was completelyinhibited by plasminogen activator inhibitor 1 (PAI-1), butnot to any significant extent by neither a1-protease inhibitor,a1-anti-chymotrypsin nor soybean trypsin inhibitor (Fig. 3).This pattern of inhibition was seen in conditioned mediaboth from control- and PMA-stimulated cells.To verify that the murine macrophage cell line J774
produced plasminogen activators, cell conditioned serum-freemediumwas subjected to substrate zymography [17]. Asshown in Fig. 4 (left panel), a band at approximately24 kDa was detected in the gel that contained bothplasminogen and gelatin, but not in the control gel thatcontained only gelatin. This indicates that this band is aplasminogen activator.The figure also shows that the presence of PMA resulted
in a slight increase in the intensity of this plasminogenactivator band, whichwas verified in other experiments withdiluted conditioned medium (data not shown). Figure 4(left panel) also shows that HX-xyl treatment of the cellsresulted in a reduction in the intensity of the plasminogenactivator band. This band was also drastically reducedin conditioned medium (control as well as PMA- and
Fig. 2. Trypsin-like activities in conditioned media from J774 macro-
phages. Equal number of J774 macrophages were incubated with
PMA, HX-xyl or both. Conditioned media were harvested and the
levels of trypsin-like activities were assayed using the chromogenic
substrates, S-2288 or S-2444 (see Materials and methods).
Fig. 3. The effect of protease inhibitors on plasminogen activator
activity in supernatants from J774 cells. Conditioned media from equal
number of untreated and PMA-treated J774 macrophages were incu-
bated for 30 min with 0.2 lM of the various macromolecular protease
inhibitors, or 1 mM of phenylmethanesulfonyl fluoride, followed by
determination of residual trypsin-like activities.
� FEBS 2003 Proteoglycans and urokinase (Eur. J. Biochem. 270) 3975
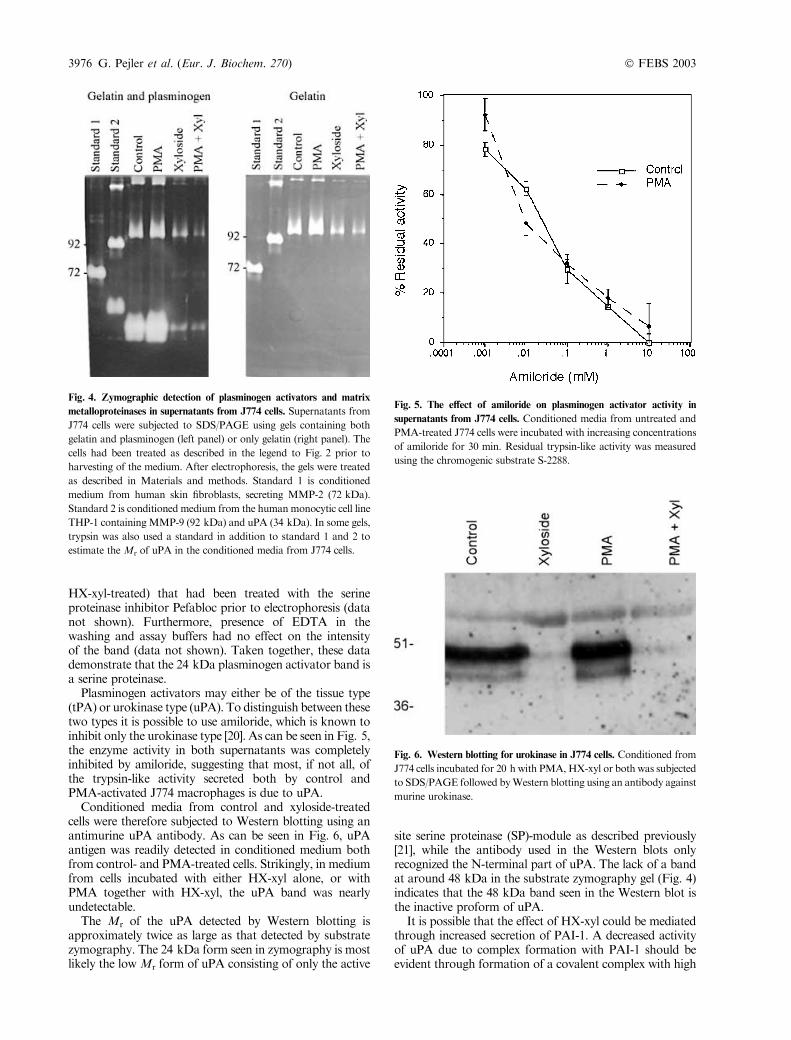
HX-xyl-treated) that had been treated with the serineproteinase inhibitor Pefabloc prior to electrophoresis (datanot shown). Furthermore, presence of EDTA in thewashing and assay buffers had no effect on the intensityof the band (data not shown). Taken together, these datademonstrate that the 24 kDa plasminogen activator band isa serine proteinase.Plasminogen activators may either be of the tissue type
(tPA) or urokinase type (uPA). To distinguish between thesetwo types it is possible to use amiloride, which is known toinhibit only the urokinase type [20]. As can be seen in Fig. 5,the enzyme activity in both supernatants was completelyinhibited by amiloride, suggesting that most, if not all, ofthe trypsin-like activity secreted both by control andPMA-activated J774 macrophages is due to uPA.Conditioned media from control and xyloside-treated
cells were therefore subjected to Western blotting using anantimurine uPA antibody. As can be seen in Fig. 6, uPAantigen was readily detected in conditioned medium bothfrom control- and PMA-treated cells. Strikingly, in mediumfrom cells incubated with either HX-xyl alone, or withPMA together with HX-xyl, the uPA band was nearlyundetectable.The Mr of the uPA detected by Western blotting is
approximately twice as large as that detected by substratezymography. The 24 kDa form seen in zymography is mostlikely the lowMr form of uPA consisting of only the active
site serine proteinase (SP)-module as described previously[21], while the antibody used in the Western blots onlyrecognized the N-terminal part of uPA. The lack of a bandat around 48 kDa in the substrate zymography gel (Fig. 4)indicates that the 48 kDa band seen in the Western blot isthe inactive proform of uPA.It is possible that the effect of HX-xyl could be mediated
through increased secretion of PAI-1. A decreased activityof uPA due to complex formation with PAI-1 should beevident through formation of a covalent complex with high
Fig. 4. Zymographic detection of plasminogen activators and matrix
metalloproteinases in supernatants from J774 cells. Supernatants from
J774 cells were subjected to SDS/PAGE using gels containing both
gelatin and plasminogen (left panel) or only gelatin (right panel). The
cells had been treated as described in the legend to Fig. 2 prior to
harvesting of the medium. After electrophoresis, the gels were treated
as described in Materials and methods. Standard 1 is conditioned
medium from human skin fibroblasts, secreting MMP-2 (72 kDa).
Standard 2 is conditioned medium from the human monocytic cell line
THP-1 containingMMP-9 (92 kDa) and uPA (34 kDa). In some gels,
trypsin was also used a standard in addition to standard 1 and 2 to
estimate theMr of uPA in the conditioned media from J774 cells.
Fig. 5. The effect of amiloride on plasminogen activator activity in
supernatants from J774 cells. Conditioned media from untreated and
PMA-treated J774 cells were incubated with increasing concentrations
of amiloride for 30 min. Residual trypsin-like activity was measured
using the chromogenic substrate S-2288.
Fig. 6. Western blotting for urokinase in J774 cells. Conditioned from
J774 cells incubated for 20 h with PMA,HX-xyl or both was subjected
to SDS/PAGE followed byWestern blotting using an antibody against
murine urokinase.
3976 G. Pejler et al. (Eur. J. Biochem. 270) � FEBS 2003
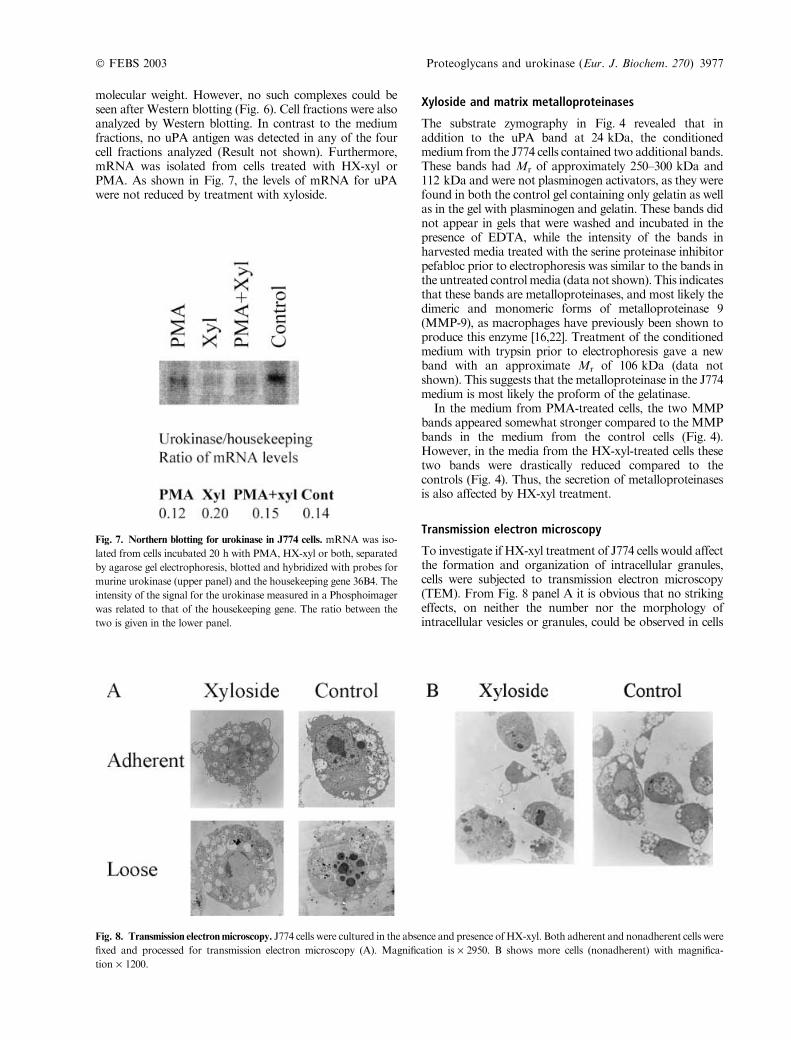
molecular weight. However, no such complexes could beseen after Western blotting (Fig. 6). Cell fractions were alsoanalyzed by Western blotting. In contrast to the mediumfractions, no uPA antigen was detected in any of the fourcell fractions analyzed (Result not shown). Furthermore,mRNA was isolated from cells treated with HX-xyl orPMA. As shown in Fig. 7, the levels of mRNA for uPAwere not reduced by treatment with xyloside.
Xyloside and matrix metalloproteinases
The substrate zymography in Fig. 4 revealed that inaddition to the uPA band at 24 kDa, the conditionedmedium from the J774 cells contained two additional bands.These bands had Mr of approximately 250–300 kDa and112 kDa and were not plasminogen activators, as they werefound in both the control gel containing only gelatin as wellas in the gel with plasminogen and gelatin. These bands didnot appear in gels that were washed and incubated in thepresence of EDTA, while the intensity of the bands inharvested media treated with the serine proteinase inhibitorpefabloc prior to electrophoresis was similar to the bands inthe untreated controlmedia (data not shown). This indicatesthat these bands are metalloproteinases, and most likely thedimeric and monomeric forms of metalloproteinase 9(MMP-9), as macrophages have previously been shown toproduce this enzyme [16,22]. Treatment of the conditionedmedium with trypsin prior to electrophoresis gave a newband with an approximate Mr of 106 kDa (data notshown). This suggests that the metalloproteinase in the J774medium is most likely the proform of the gelatinase.In the medium from PMA-treated cells, the two MMP
bands appeared somewhat stronger compared to the MMPbands in the medium from the control cells (Fig. 4).However, in the media from the HX-xyl-treated cells thesetwo bands were drastically reduced compared to thecontrols (Fig. 4). Thus, the secretion of metalloproteinasesis also affected by HX-xyl treatment.
Transmission electron microscopy
To investigate if HX-xyl treatment of J774 cells would affectthe formation and organization of intracellular granules,cells were subjected to transmission electron microscopy(TEM). From Fig. 8 panel A it is obvious that no strikingeffects, on neither the number nor the morphology ofintracellular vesicles or granules, could be observed in cells
Fig. 7. Northern blotting for urokinase in J774 cells. mRNA was iso-
lated from cells incubated 20 h with PMA, HX-xyl or both, separated
by agarose gel electrophoresis, blotted and hybridized with probes for
murine urokinase (upper panel) and the housekeeping gene 36B4. The
intensity of the signal for the urokinase measured in a Phosphoimager
was related to that of the housekeeping gene. The ratio between the
two is given in the lower panel.
Fig. 8. Transmission electronmicroscopy. J774 cells were cultured in the absence and presence of HX-xyl. Both adherent and nonadherent cells were
fixed and processed for transmission electron microscopy (A). Magnification is · 2950. B shows more cells (nonadherent) with magnifica-
tion · 1200.
� FEBS 2003 Proteoglycans and urokinase (Eur. J. Biochem. 270) 3977

treated with HX-xyl. In panel B more cells are shown at asmaller magnification.
Discussion
In the present paper we show that proteoglycans areimportant for secreted uPA activity in J774 macrophages.uPA activity has previously been demonstrated in severalmacrophage cell lines [23] and in human macrophages [22].Mice lacking uPA expression are not able to recruitsufficient number of macrophages during inflammation[24], suggesting that the enzyme is important in the cellularimmune system. Indeed, uPA activity was increased in themedium after PMA treatment, in agreement with the notionthat uPA secretion is a characteristic feature of activatedmacrophages [25]. Additionally, secretion of proteoglycansin monocytes and macrophages increases when the cells areactivated [6], as was also apparent in this study. Accord-ingly, secretion of both uPA and proteoglycans increase inactivated monocytes and macrophages. Plasmin, generatedfrom the precursor plasminogen through the action of uPA,can cleave matrix proteins such as fibronectin, laminin andaggrecan, and also activate matrix- and membrane associ-ated MMPs, fibroblast growth factor and transforminggrowth factor b [26]. In atherosclerosis, lipid-rich macro-phages increase uPA and plasmin expression and the releaseof growth factors from the extracellular matrix [27]. Clearly,the regulation of plasmin formation is important formacrophages and metastasizing tumor cells, and cellsinvolved in tissue repair. Likewise, secretion of MMP-9from macrophages is important in immune reactions andatherosclerosis [28]. The results presented here thus indicatethat proteoglycans secreted from macrophages, e.g. sergly-cin, may regulate the activity or availability of uPA andMMP-9. However, HX-xyl treatment does not lead to acomplete inhibition of uPA release from the cells, despite anessentially total abrogation of the synthesis of intactproteoglycans. The reason for this is not known. However,it is possible that preformed uPA and intact proteoglycansare present in the cells and are being released during thecourse of the experiments. Alternatively, uPA secretionmaybe only partly dependent on the intact proteoglycans.Control and PMA-stimulated J774 macrophages release
proteoglycans of both chondroitin sulfate and heparansulfate type. In the present study we show that xylosidetreatment of both control and PMA-stimulated J774 cellscompletely blocks the assembly of intact heparan sulfateand chondroitin sulfate proteoglycans that are destined forsecretion. Which of the two proteoglycans, heparan sulfateor chondroitin sulfate that is important for the uPA activity/secretion is at present not known. Importantly, we did notsee any reduction in mRNA levels for uPA upon xylosidetreatment, indicating that the inhibitory effect of xylosideson extracellular uPA was caused by post-translationalmechanisms. However, we do not know at which level uPAis dependent on proteoglycans. One possibility is that uPA isdependent on proteoglycans after release from the cellswhere the lack of intact proteoglycans may affect theactivity or half-life of uPA. It is conceivable that uPA orMMP-9 released to the medium in the J774 systemmight beinactivated either by other proteases or by proteaseinhibitors, if no proteoglycans are simultaneously secreted
to the medium. In this context it is interesting to note thatheparan sulfate has been shown to both protect plasminfrom inactivation by protease inhibitors and to stimulate itsenzyme activity [29]. In addition, recent findings show thatthe interaction between serglycin and granzymes in cyto-toxic granules is important to mediate apoptosis in targetcells [30]. Granzymes have also been shown to circulate inplasma bound to proteoglycans, whereby they are protectedfrom inactivation by protease inhibitors [31]. Accordingly,based on the findings presented here, one possible functionof secreted proteoglycans in macrophages may be to protectand regulate the activity of uPA andMMP-9 expressed andsecreted by the same cells. A second possibility could be thatthe proteoglycans may be important intracellularly in theformation of the secretory vesicles. Each of these twopossibilities implies that the protein core of the proteogly-can, or the intact proteoglycan molecule, is an importantcomponent of the secretory process, as the xylosidetreatment did not reduce the amount of secreted glycos-aminoglycan chains available. The mechanism by which theprotein core could influence the secretion of proteolyticenzymes is uncertain. It is possible, for example, that theprotein core in some way is involved in intracellular sortingof uPA and MMP-9. Another possibility could be that theprotein core is attached to the vesicle membrane, and thatsuch a linkagemay be important for formation or structuralintegrity of the secretory vesicles. In this context it is notedthat proteoglycans, possibly GPI-linked to the granulemembrane, are important for the formation of zymogengranules in pancreatic acinar cells [32]. Further, proteogly-cans have been suggested to be important for the intracel-lular transport of enzymes to the lysosomes in monocytes[33]. A third possibility would be that the cell-surfaceproteoglycans participate in the regulation of uPA. HX-xyl-treated cells have reduced levels of cell surface-associatedproteoglycans compared to control macrophages. Possibly,this may affect the cell association of uPA after release and/or the level of activity. In fact, it has been shown previouslythat cell association of uPA-generating activity enhances therate of formation of active uPA [34].An alternative explanation for the effect of the xyloside
on uPA and MMP-9 secretion could be that the xylosidetreatment reduces the amount of heparan sulfate chainssynthesized in favor of chondroitin sulfate, and that uPAand MMP-9 may be specifically dependent on glycosami-noglycans of the heparan sulfate type. In line with such anexplanation, it was recently shown that mast cell carboxy-peptidase A expressed by bone marrow-derivedmast cells isstrictly dependent on heparin glycosaminoglycan for stor-age and processing, whereas mast cell tryptase can be storedand processed also in cells lacking heparin but containingchondroitin sulfate of equal charge density [35].A dependence of uPA on proteoglycans has to our
knowledge not been described previously. However, it hasbeen shown recently that serglycin and tPA colocalize inintracellular granules of endothelial cells, thus giving furthersupport for a role of proteoglycans in the regulation ofplasminogen activators [36].The activity of uPA can be regulated through several
mechanisms, including the expression levels, uPA receptorbinding and regulation by PAI-1. The expression levels arethe subject of regulation through the actions of growth
3978 G. Pejler et al. (Eur. J. Biochem. 270) � FEBS 2003

factors and inflammatory mediators [26]. Tumor-associatedmacrophages have, e.g. been demonstrated to increase theexpression of uPA when exposed to transforming growthfactor-b [37]. It has also been shown that the expression levelof uPA in J774 cells can be regulated through interactions ofthe cells with extracellular laminin through the integrinreceptor a6b1 [26]. Data presented here suggest an additionallevel of regulation of uPA, and also MMP-9, activity inmacrophages, through the dependence of cellular proteo-glycan expression and secretion.
Acknowledgements
The expert technical assistance of Eli Berg and Annicke Stranda is
acknowledged.
This work was supported by grants from The Norwegian Cancer
Society, The Throne-Holst Fund, The Swedish Medical Research
Council (grant no. 9913) and from King Gustaf V’s 80th anniversary
Fund.
References
1. Nathan, C.F. (1987) Secretory products of macrophages. J. Clin.
Invest. 79, 319–326.
2. Uhlin-Hansen, L., Wik, T., Kjellen, L., Berg, E., Forsdahl, F. &
Kolset, S.O. (1993) Proteoglycan metabolism in normal and
inflammatory human macrophages. Blood 82, 2880–2889.
3. Oynebraten, I., Hansen, B., Smedsrod, B. & Uhlin-Hansen, L.
(2000) Serglycin secreted by leukocytes is efficiently eliminated
from the circulation by sinusoidal scavenger endothelial cells in the
liver. J. Leukoc. Biol. 67, 183–188.
4. Yeaman, C. & Rapraeger, A.C. (1993) Membrane-anchored
proteoglycans of mouse macrophages: P388D1 cells express a
syndecan-4-like heparan sulfate proteoglycan and a distinct
chondroitin sulfate form. J. Cell Physiol. 157, 413–425.
5. Clasper, S., Vekemans, S., Fiore, M., Plebanski, M., Wordsworth,
P., David, G. & Jackson, D.G. (1999) Inducible expression of the
cell surface heparan sulfate proteoglycan syndecan-2 (fibroglycan)
on human activated macrophages can regulate fibroblast growth
factor action. J. Biol. Chem. 274, 24113–24123.
6. Uhlin-Hansen, L., Eskeland, T. &Kolset, S.O. (1989)Modulation
of the expression of chondroitin sulfate proteoglycan in stimulated
human monocytes. J. Biol. Chem. 264, 14916–14922.
7. Forsberg, E., Pejler, G., Ringvall, M., Lunderius, C., Tomasini-
Johansson, B., Kusche-Gullberg, M., Eriksson, I., Ledin, J.,
Hellman, L. & Kjellen, L. (1999) Abnormal mast cells in mice
deficient in a heparin-synthesizing enzyme. Nature 400, 773–776.
8. Humphries, D.E., Wong, G.W., Friend, D.S., Gurish, M.F., Qiu,
W.T., Huang, C., Sharpe, A.H. & Stevens, R.L. (1999) Heparin is
essential for the storage of specific granule proteases in mast cells.
Nature 400, 769–772.
9. Pejler, G. & Sadler, J.E. (1999) Mechanism by which heparin
proteoglycan modulates mast cell chymase activity. Biochemistry
38, 12187–12195.
10. Hallgren, J., Spillmann, D. & Pejler, G. (2001) Structural
requirements and mechanism for heparin-induced activation of a
recombinant mouse mast cell tryptase, mouse mast cell protease-6:
formation of active tryptase monomers in the presence of low
molecular weight heparin. J. Biol. Chem. 276, 42774–42781.
11. Tchougounova, E. & Pejler, G. (2001) Regulation of extravascular
coagulation and fibrinolysis by heparin-dependent mast cell chy-
mase. FASEB J. 15, 2763–2765.
12. Kolset, S.O., Mann, D.M., Uhlin-Hansen, L., Winberg, J.O. &
Ruoslahti, E. (1996) Serglycin-binding proteins in activated
macrophages and platelets. J. Leukoc. Biol. 59, 545–554.
13. Toyama-Sorimachi, N., Kitamura, F., Habuchi, H., Tobita, Y.,
Kimata, K. & Miyasaka, M. (1997) Widespread expression of
chondroitin sulfate-type serglycins with CD44 binding ability in
hematopoietic cells. J. Biol. Chem. 272, 26714–26719.
14. Kolset, S.O., Sakurai, K., Ivhed, I., Overvatn, A. & Suzuki, S.
(1990) The effect of beta-D-xylosides on the proliferation and
proteoglycan biosynthesis of monoblastic U-937 cells. Biochem.
J. 265, 637–645.
15. Halvorsen, B., Aas, U.K., Kulseth, M.A., Drevon, C.A.,
Christiansen, E.N. & Kolset, S.O. (1998) Proteoglycans in mac-
rophages: characterization and possible role in the cellular uptake
of lipoproteins. Biochem. J. 331, 743–752.
16. Winberg, J.O., Kolset, S.O., Berg, E. & Uhlin-Hansen, L. (2000)
Macrophages secrete matrix metalloproteinase 9 covalently linked
to the core protein of chondroitin sulfate proteoglycans. J. Mol.
Biol. 304, 669–680.
17. Heussen, C. & Dowdle, E.B. (1980) Electrophoretic analysis of
plasminogen activators in polyacrylamide gels containing sodium
dodecyl sulfate and copolymerized substrates.Anal. Biochem. 102,
196–202.
18. Svendsrud, D.H., Loennechen, T. & Winberg, J.O. (1997) Effect
of adenosine analogues on the expression of matrix metallopro-
teinases and their inhibitors from human dermal fibroblasts.
Biochem. Pharmacol. 53, 1511–1520.
19. Shively, J.E. & Conrad, H.E. (1976) Formation of anhydrosugars
in the chemical depolymerization of heparin. Biochemistry 15,
3932–3942.
20. Vassalli, J.D. & Belin, D. (1987) Amiloride selectively inhibits the
urokinase-type plasminogen activator. FEBS Lett. 214, 187–191.
21. Novokhatny, V., Medved, L., Mazar, A., Marcotte, P., Henkin, J.
& Ingham, K. (1992) Domain structure and interactions of
recombinant urokinase-type plasminogen activator. J. Biol. Chem.
267, 3878–3885.
22. Shapiro, S.D., Campbell, E.J., Senior, R.M. & Welgus, H.G.
(1991) Proteinases secreted by human mononuclear phagocytes.
J. Rheumatol. 27, 95–98.
23. Jones, C.M., Goldfarb, R.H. & Holden, H.T. (1983) Macrophage
cell lines behave as activated macrophages in the production and
regulation of plasminogen activator. Cancer Invest. 1, 207–213.
24. Carmeliet, P. & Collen, D. (1996) Gene manipulation and transfer
of the plasminogen and coagulation system in mice. Semin.
Thromb. Hemost. 22, 525–542.
25. Vassalli, J.D. & Pepper, M.S. (1994) Tumour biology. Membrane
proteases in focus. Nature 370, 14–15.
26. Khan, K.M. & Falcone, D.J. (1997) Role of laminin in matrix
induction of macrophage urokinase-type plasminogen activator
and 92-kDa metalloproteinase expression. J. Biol. Chem. 272,
8270–8275.
27. Falcone, D.J., McCaffrey, T.A., Haimovitz-Friedman, A.,
Vergilio, J.A. & Nicholson, A.C. (1993) Macrophage and foam
cell release of matrix-bound growth factors. Role of plasminogen
activation. J. Biol. Chem. 268, 11951–11958.
28. Opdenakker, G., Van den Steen, P.E. & Van Damme, J. (2001)
Gelatinase B: a tuner and amplifier of immune functions. Trends
Immunol. 22, 571–579.
29. Brunner, G., Reimbold, K., Meissauer, A., Schirrmacher, V. &
Erkell, L.J. (1998) Sulfated glycosaminoglycans enhance tumor
cell invasion in vitro by stimulating plasminogen activation. Exp.
Cell Res. 239, 301–310.
30. Metkar, S.S., Wang, B., Aguilar-Santelises, M., Raja, S.M.,
Uhlin-Hansen, L., Podack, E., Trapani, J.A. & Froelich, C.J.
(2002) Cytotoxic cell granule-mediated apoptosis: perforin delivers
granzyme B-serglycin complexes into target cells without plasma
membrane pore formation. Immunity 16, 417–428.
31. Spaeny-Dekking, E.H., Kamp, A.M., Froelich, C.J. &Hack, C.E.
(2000) Extracellular granzyme A, complexed to proteoglycans, is
� FEBS 2003 Proteoglycans and urokinase (Eur. J. Biochem. 270) 3979

protected against inactivation by protease inhibitors. Blood 95,
1465–1472.
32. Schmidt, K., Dartsch, H., Linder, D., Kern, H.F. & Kleene, R.
(2000) A submembranous matrix of proteoglycans on zymogen
granule membranes is involved in granule formation in rat pan-
creatic acinar cells. J. Cell Sci. 113, 2233–2242.
33. Lemansky, P. & Hasilik, A. (2001) Chondroitin sulfate is involved
in lysosomal transport of lysozyme in U937 cells. J. Cell Sci. 114,
345–352.
34. Duval-Jobe, C. & Parmely, M.J. (1994) Regulation of plasmino-
gen activation by human U937 promonocytic cells. J. Biol. Chem.
269, 21353–21357.
35. Henningsson, F., Ledin, J., Lunderius, C., Wilen, M., Hellman, L.
& Pejler, G. (2002) Altered storage of proteases in mast cells from
mice lacking heparin: a possible role for heparin in carboxy-
peptidase A processing. Biol. Chem. 383, 793–801.
36. Schick, B.P., Gradowski, J.F. & San Antonio, J.D. (2001)
Synthesis, secretion, and subcellular localization of serglycin pro-
teoglycan in human endothelial cells. Blood 97, 449–458.
37. Hildenbrand, R., Jansen, C., Wolf, G., Bohme, B., Berger, S., von
Minckwitz, G., Horlin, A., Kaufmann, M. & Stutte, H.J. (1998)
Transforming growth factor-beta stimulates urokinase expression
in tumor-associated macrophages of the breast. Laboratory Invest.
78, 59–71.
3980 G. Pejler et al. (Eur. J. Biochem. 270) � FEBS 2003
Related Documents











