MICROVASCULAR RESEARCH 41, 92-110 (1991) Penetration of the Systemic Blood Pressure into the Microvasculature of Rat Skeletal Muscle F. A. DELANO, G. W. SCHMID-SCH~NBEIN, T. C. SKALAK,* AND B. W. ZWEIFACH Department of AMES-Bioengineering, University of California, San Diego, La Jolla, CA 92093; and *Department of Biomedical Engineering, Health Sciences Center, University of Virginia, Charlottesville, Virginia 22908 Received January 18, 1990 A series of arterial micropressure measurements in different skeletal muscles of the Wistar- Kyoto and spontaneously hypertensive rat is presented. The micropunctures were carried out with minimal surgical intervention through small skin incisions and the micropressures were recorded simultaneously with femoral artery pressures. The measurement sites were located at the entry points into the muscles for the proximal and distal supply arteries and at the midpoint of the arteriolar arcade bridge which directly connects these two supply arteries in the center of the muscle parenchyma. In contrast to feed artery pressure values from exteriorized muscles, which in the past have been reported to be as low as 40 mm Hg, the current mean pressure values are substantially higher and in the range between 70 and 100 mm Hg, equivalent to 70 to 90% of the mean systemic pressure. Systolic and diastolic values exhibit comparable trends to the mean pressures and they are similar in muscles at different locations in the body. Although in spontaneously hypertensive rats the absolute pressures were significantly higher compared with their controls, the normalized pressures were virtually identical at the locations used in this study. These data indicate that the absolute pressure in the central arteries of spontaneously hypertensive animals is reduced to a greater degree than in Wistar-Kyoto rats, while in both strains the major pressure reduction in skeletal muscle still occurs in the microcirculation. o 1991 Academic press, IX. INTRODUCTION One of the key unresolved issues in cardiovascular research concerns the extent to which the blood pressure is reduced along the circulatory network. In textbooks the blood pressure is depicted as a continuously decreasing function along the schematic “vascular tree,” without a clear indication to what proportion of the pressure reduction takes place in large and small arteries and in the microcir- culation. The descriptions of the pressure profiles are qualitative without due consideration of the real arrangement of the vascular network, especially in regard to different organs. There are excellent reports that deal with the pressure dis- tribution in the central arteries (summary by Stettler et al., 1987), but the pressures during the transition into the microcirculation are usually based on extrapolations without regard to the microanatomy and the flow field. In this report we propose to focus on several details of the pressures proximal to the microcirculation in a single organ, the skeletal muscle of rats, with specific attention to the terminal 92 002~286Z91 $3.00 Copyright 0 1991 by Academic Press, Inc. All rights of reproduction in any form reserved. Printed in U.S.A.

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

MICROVASCULAR RESEARCH 41, 92-110 (1991)
Penetration of the Systemic Blood Pressure into the Microvasculature of Rat Skeletal Muscle
F. A. DELANO, G. W. SCHMID-SCH~NBEIN, T. C. SKALAK,* AND B. W. ZWEIFACH
Department of AMES-Bioengineering, University of California, San Diego, La Jolla, CA 92093; and *Department of Biomedical Engineering, Health Sciences Center, University of
Virginia, Charlottesville, Virginia 22908
Received January 18, 1990
A series of arterial micropressure measurements in different skeletal muscles of the Wistar- Kyoto and spontaneously hypertensive rat is presented. The micropunctures were carried out with minimal surgical intervention through small skin incisions and the micropressures were recorded simultaneously with femoral artery pressures. The measurement sites were located at the entry points into the muscles for the proximal and distal supply arteries and at the midpoint of the arteriolar arcade bridge which directly connects these two supply arteries in the center of the muscle parenchyma. In contrast to feed artery pressure values from exteriorized muscles, which in the past have been reported to be as low as 40 mm Hg, the current mean pressure values are substantially higher and in the range between 70 and 100 mm Hg, equivalent to 70 to 90% of the mean systemic pressure. Systolic and diastolic values exhibit comparable trends to the mean pressures and they are similar in muscles at different locations in the body. Although in spontaneously hypertensive rats the absolute pressures were significantly higher compared with their controls, the normalized pressures were virtually identical at the locations used in this study. These data indicate that the absolute pressure in the central arteries of spontaneously hypertensive animals is reduced to a greater degree than in Wistar-Kyoto rats, while in both strains the major pressure reduction in skeletal muscle still occurs in the microcirculation. o 1991 Academic press,
IX.
INTRODUCTION
One of the key unresolved issues in cardiovascular research concerns the extent to which the blood pressure is reduced along the circulatory network. In textbooks the blood pressure is depicted as a continuously decreasing function along the schematic “vascular tree,” without a clear indication to what proportion of the pressure reduction takes place in large and small arteries and in the microcir- culation. The descriptions of the pressure profiles are qualitative without due consideration of the real arrangement of the vascular network, especially in regard to different organs. There are excellent reports that deal with the pressure dis- tribution in the central arteries (summary by Stettler et al., 1987), but the pressures during the transition into the microcirculation are usually based on extrapolations without regard to the microanatomy and the flow field. In this report we propose to focus on several details of the pressures proximal to the microcirculation in a single organ, the skeletal muscle of rats, with specific attention to the terminal
92
002~286Z91 $3.00 Copyright 0 1991 by Academic Press, Inc. All rights of reproduction in any form reserved. Printed in U.S.A.

SYSTEMIC-MICROVASCULAR PRESSURE PROFILE 93
vascular network and to differences in muscles positioned in various regions of the body.
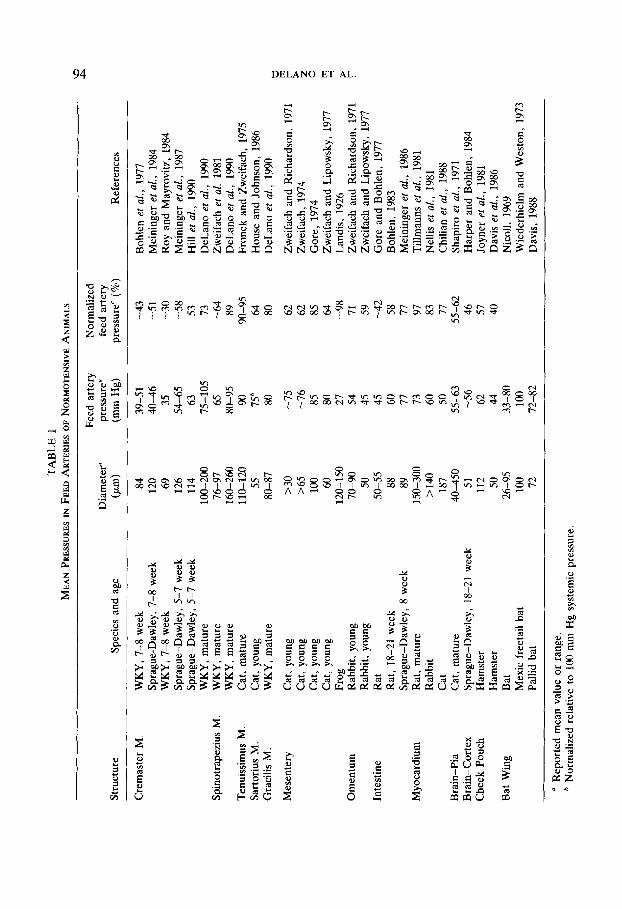
In the last 2 decades several pressure profiles in the skeletal muscle microcir- culation of different species have been reported (Smaje et al., 1970; Fronek and Zweifach, 1975; Bohlen et al., 1977; Zweifach et al., 1981; Roy and Mayrovitz, 1984; Meininger et al., 1984; Meininger et al., 1987; Hill et al., 1990). A common observation in all these studies is the fact that in laboratory mammals, the pressure decreases progressively along the arteries and arterioles so as to reach in the entry region of the capillaries values in the range between 20 and 30 mm Hg, apparently independent of the location of a particular skeletal muscle or the animal species. In contrast, the published values for the pressure at the point where the small arteries feed into the skeletal muscle microcirculation differ considerably (see summary in Table 1). For example in the cremaster and spinotrapezius muscle preparations, average pressures as low as about 40 mm Hg have been reported (Top, Table l), levels which constitute only about 50% of the mean systemic (typically the femoral) blood pressure and even lower values. In contrast, the mean pressure in 110 to 120 ,um feeder arterioles to the cat tenuissimus muscle is almost 90% of the systemic blood pressure (Fronek and Zweifach, 1975), so that on average the reduction in pressure along the systemic arteries that supply the microcirculation is about 10% in one region and about 50% in another. Why such a discrepancy?
In part these discrepancies may rest in the differences of vascular anatomy. However, before we can accept such an explanation, we have to weigh the lim- itations of current measurement techniques. One of the difficulties in evaluating micropressure measurements in skeletal muscle stems from the fact that prepa- ration of the organ for intravital microscopy and micropipette puncture frequently entails interruption of blood flow from ancillary supply arteries. Skeletal muscles have multiple supply arteries’ leading from the central circulation and also in some cases smaller arterioles connecting with vessels in neighboring muscle (Spal- teholz, 1888; Anderson et al., 1988). The spinotrapezius muscle has two relatively large arteries, one from the thoracodorsal artery the other from the 11th intercostal artery (Schmid-Schonbein et al., 1986). They connect head-on in the muscle to form a large arcade arteriole, a vessel denoted as the arcade bridge (Schmid- Schonbein et al., 1987). The arteriolar arcade bridge in most muscles constitutes the arteriole with the largest diameter and it is usually paired with a large venule (Myrhage, 1977; Lindbom et al., 1983). Although the arteriolar arcade bridge serves as a supply vessel for the remainder of the arcade arterioles and the microcirculation proper, it is anatomically positioned inside the organ parenchyma, thus leaving it exposed to locally derived mediators. The specific question under consideration is the pressure level along the course of the long arteriolar arcade bridges under conditions where all its blood supplies are left intact.
In the present report detailed measurements are given for pressures in small arteries at sites where they feed into the skeletal muscle microcirculation and at selected points in the arcade bridge. A number of skeletal muscles located at different distances from the heart are compared. The micropunctures were carried
’ In this report we define arteries to be vessels outside the muscle proper, arterioles are vessels within the muscle, such as arcade or transverse arterioles.
TABL
E 1
MEAN
PR
ESSU
RES
IN
FEED
AR
TERI
ES
OF
NORM
OTEN
SIVE
AN
IMAL
S
Stru
ctur
e Sp
ecie
s an
d ag
e D
iam
eteP
6.
4
Feed
ar
tery
No
rmal
ized
pres
sure
” fe
ed a
rtery
(m
m
Hd
pres
sure
’ (%
) R
efer
ence
s
Cre
mas
ter
M.
Spin
otra
peziu
s M
Tenu
issim
us
M.
Sarto
rius
M.
Gra
cilis
M.
Mes
ente
ry
Om
entu
m
Inte
stin
e
Myo
card
ium
Brai
n-Pi
a Br
ain-
Cor
tex
Che
ek
Pouc
h
Bat
Win
g
WKY
, 7-
8 we
ek
84
39-5
1 -4
3 Sp
ragu
e-D
awle
y,
7-8
week
12
0 40
-46
-51
WKY
, 7-
8 we
ek
69
35
-30
Spra
gue-
Daw
ley,
5-
7 we
ek
126
54-6
5 -5
8 Sp
ragu
e-D
awle
y,
5-7
week
11
4 63
53
W
KY,
mat
ure
100-
200
75-1
05
73
WKY
, m
atur
e 76
-97
65
-64
WKY
, m
atur
e 16
0-26
0 80
-95
89
Cat
, m
atur
e 11
0-12
0 90
90
-95
Cat
, yo
ung
55
75b
64
WKY
, m
atur
e 80
-87
80
80
Cat
, yo
ung
>30
-75
Cat
, yo
ung
>65
-76
Cat
, yo
ung
100
85
Cat
, yo
ung
60
80
Frog
12
0-15
0 27
R
abbi
t, yo
ung
70-9
0 54
R
abbi
t, yo
ung
50
45
Rat
50
-55
45
Rat
, 18
-21
week
88
60
Sp
ragu
e-D
awle
y,
8 we
ek
89
77
Rat
, m
atur
e 15
0-30
0 73
R
abbi
t >1
40
60
Cat
18
7 50
C
at,
mat
ure
40-4
50
55-6
3 Sp
ragu
e-D
awle
y,
18-2
1 we
ek
51
-56
Ham
ster
11
2 62
H
amst
er
50
44
Bat
26-9
5 33
-80
Mex
ic fre
etai
l ba
t 10
0 10
0 Pa
llid
bat
72
72-8
2
62
62
85
64
-98 71
59
-42 58
77
97
83
77
55
-62
46
57
40
Bohl
en
et al.
, 19
77
Mei
ning
er
et al.
, 19
84
Roy
and
May
rovi
tz,
1984
M
eini
nger
et
al.,
1987
Hill
et al.
, 19
90
DeLa
no
et al.
, 19
90
Zwei
fach
et
al.
1981
De
Lano
et
al.,
1990
Fr
onek
an
d Zw
eifa
ch,
1975
H
ouse
an
d Jo
hnso
n,
1986
De
Lano
et
al.,
1990
u
Zwei
fach
an
d R
ichar
dson
, 19
71
Zwei
fach
, 19
74
F
Gor
e,
1974
3
Zwei
fach
an
d Li
pow
sky,
19
77
2 La
ndis,
19
26
Zwei
fach
an
d R
ichar
dson
, 19
71
:: . Zw
eifa
ch
and
Lipo
wsk
y,
1977
Go
re an
d Bo
hlen
, 19
77
Bohl
en,
1983
M
eini
nger
et
al.,
1986
Tillm
anns
et
al.,
1981
N
ellis
et
al.,
1981
Ch
ilian
et al.
, 19
88
Shap
iro
et al.
, 19
71
Harp
er
and
Bohl
en,
1984
Jo
yner
et a
l.,
1981
D
avis
el
al.,
1986
N
icol
l, 19
69
Wie
derh
ielm
an
d W
esto
n,
1973
D
avis
, 19
88
’ Re
porte
d m
ean
valu
e or
ra
nge.
b
Norm
alize
d re
lativ
e to
10
0 m
m H
g sy
stem
ic p
ress
ure.

SYSTEMIC-MICROVASCULAR PRESSURE PROFILE 95
out through the smallest possible skin incisions to minimize the effects of surgery and to preserve the muscle circulation, while at the same time the systemic blood pressure was recorded. All measurements were carried out in spontaneously hy- pertensive rats (SHR) and in Wistar-Kyoto rats (WKY), their normotensive control strain. Portions of this work have previously been reported in abstract form (DeLano et al., 1989).
METHODS
Male SHR and WKY rats (Charles River Breeding Laboratories, Wilmington, MA) between 12 and 18 weeks of age with body weights between 300 and 400 g were used for the study. Control femoral pressures were obtained in each animal about 1 h before induction of general anesthesia by insertion of a small catheter (PE-50 tubing) under local anesthesia (4% lidocaine, SC) (Zweifach et al., 1981). Mean and pulsatile arterial pressures were recorded continuously for about 20 min using a digital laboratory computer. The mean pressures (f standard de- viation) under local anesthesia were 110 + 6 mm Hg in WKY and 160 + 12 mm Hg in SHR, with no overlap of the pressure histograms between the two strains.
General anesthesia was maintained in the current study with alfaxalone (Al- fathesin; Glaxovet, Boronia, Australia), a short-acting steroid-type agent, by con- tinuous infusion of 7.5 to 10 mg/kg/hr iv. Pressures under such conditions are lower than those in the control period and are stable for several hours. The necessary measurements were completed within 2 hr.
The study was focused on three muscles, the spinotrapezius m. (Gray, 1973), the cremaster m. (Baez, 1973), and the gracilis m. (Henrich and Hecke, 1978). Additional measurements were also obtained in the tibialis m., which was selected because its blood supply pathway is one of the most distant from the heart. With the exception of a separate study in the cremaster m., none of the muscles were exteriorized as for microscopy. Instead the locations of the posterior and anterior supply arterioles at the point where they enter the muscle were identified (about 0.5 to 0.1 mm from the actual entry point) by a series of exploratory experiments and only small skin incisions were made (about 0.5 to 0.1 cm) to expose these vessels for micropipette puncture. The distal feeder of the cremaster m. was exposed after removal of the skin around the tip of the testis and penetration through the fascia and muscle per se via a small (<0.5 cm) incision. Interruption of blood flow to surrounding tissues was kept to a minimum. The measurements along the arteriolar arcade bridge were carried out at the midway point between the proximal and distal supply arteries. Since the feeder arteries are usually outside the muscle fibers they require only minimal dissection of fascia, whereas the arcade bridge is more deeply embedded in the muscle. Exposure of these arcades for micropuncture requires the separation of a thin layer of muscle fibers about 0.5 to 1.0 mm in depth in which the edges were kept about 1 mm apart by the placement of two sutures (6-O silk, Ethicon) without significant interruption of the blood flow in the contiguous arcade arterioles. At exposed tissue sites a superfusion of a bicarbonate buffered Krebs-Henseleit solution (saturated with 5% 02, 95% Nz, pH 7.4, 36”) was maintained.
Micropressures were measured with the servo-null technique by Wiederhielm et al. (1964), as implemented by Intaglietta et al. (1970). Pipettes were pulled out
96 DELANO ET AL.
rMIDPOINT ARCADE BRIDGE A.
,’
/I’ PROXIMAL SUPPLY A.
/ / / i i
‘\ ‘\
/
‘xx ..__- fee-----/ ’ --THORACODORSAL A.
1 ’ DISTAL SUPPLY A. ( from INTERCOSTAL A )
DISTAL ARCADE
BRIDGE A.
in zi TIME ( seconds )
MIDPOINT ARCADE PROXIMAL ARCADE
BRIDGE A. BRIDGE A.
240
TIME ( seconds ) TIME ( seconds )
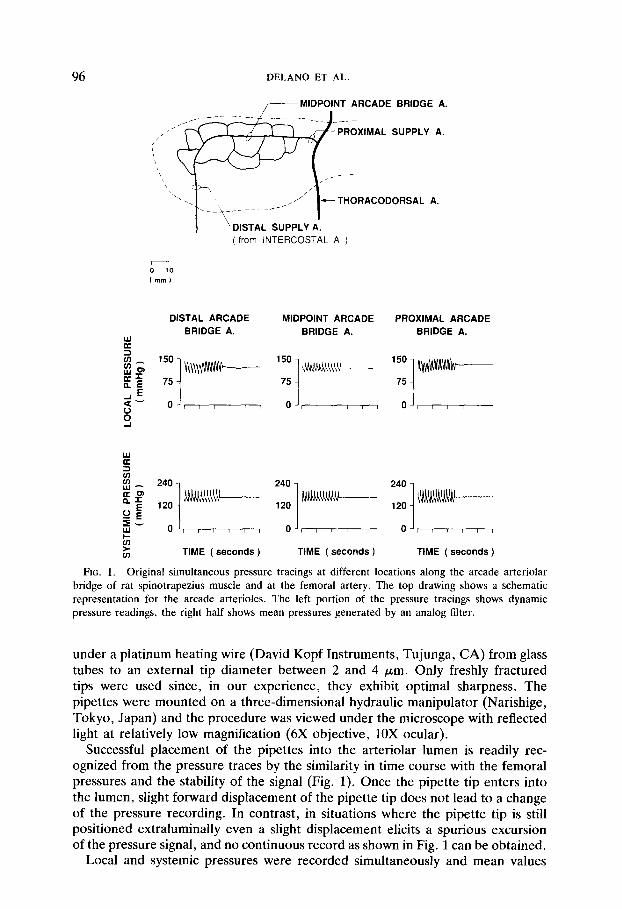
FIG. 1. Original simultaneous pressure tracings at different locations along the arcade arteriolar bridge of rat spinotrapezius muscle and at the femoral artery. The top drawing shows a schematic representation for the arcade arterioles. The left portion of the pressure tracings shows dynamic pressure readings, the right half shows mean pressures generated by an analog filter.
under a platinum heating wire (David Kopf Instruments, Tujunga, CA) from glass tubes to an external tip diameter between 2 and 4 pm. Only freshly fractured tips were used since, in our experience, they exhibit optimal sharpness. The pipettes were mounted on a three-dimensional hydraulic manipulator (Narishige, Tokyo, Japan) and the procedure was viewed under the microscope with reflected light at relatively low magnification (6X objective, 10X ocular).
Successful placement of the pipettes into the arteriolar lumen is readily rec- ognized from the pressure traces by the similarity in time course with the femoral pressures and the stability of the signal (Fig. 1). Once the pipette tip enters into the lumen, slight forward displacement of the pipette tip does not lead to a change of the pressure recording. In contrast, in situations where the pipette tip is still positioned extraluminally even a slight displacement elicits a spurious excursion of the pressure signal, and no continuous record as shown in Fig. 1 can be obtained.
Local and systemic pressures were recorded simultaneously and mean values

SYSTEMIC-MICROVASCULAR PRESSURE PROFILE
INTACT MUSCLE EXTERIORIZED
E
i- 150, 1501
97
MUSCLE
i w - 240 240
EI” OEE 120 120
11 2 0 0
in ?i TIME ( seconds ) TIME ( seconds )
FIG. 2. Original pressure tracings at the midpoint arcade bridge arteriole of the same cremaster muscle (see Fig. 3 top for the exact location) before and in the same animal (SHR) after muscle exteriorization according to the procedure by Baez (1973). Simultaneous systemic pressure tracings are shown. In this case the mean pressure reduction is 22 mm Hg, the pulse pressure reduction is 35 mm Hg.
were determined electronically by an analog low pass filter with a cutoff frequency at about 0.3 Hz. Steady-state measurements are accurate to within about 3%, and pulsatile pressures are recorded at 100% amplitude up to about 2 Hz; at 5 Hz there is about a 20% amplitude reduction (Lipowsky, 1975), but no correction was made since most pressure signals were at or below 5 Hz basic frequency. Statistical analysis for comparison between WKY and SHR was carried out with Student’s t test, since most pressure histograms exhibit symmetric shapes so that the mean pressure is representative. P < 0.05 was considered to be significant.
RESULTS
Mean Arterial Pressure after Muscle Exteriorization
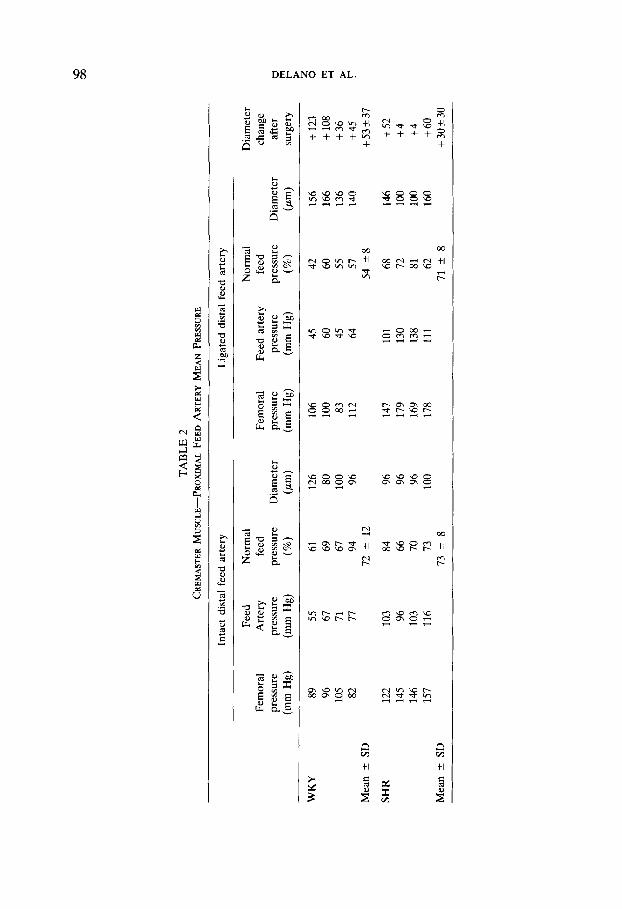
We have found that a basic principle for faithful measurements of pressure in skeletal muscle, is that all feeder arterioles should be left intact. Figure 2 compares pressure tracings at the midpoint of the arcade bridge in the cremaster muscle in situ (left tracing) and in the same muscle at exactly the same location after exposure (right tracing) by the method of Baez (1973). Surgical exposure for microscopy interrupts the blood supply from the distal arteriole which originates from the deferential artery in the region of the cauda epididymis. Typically, with exter- iorization of the cremaster muscle there is a reduction in mean pressure of about 20 mm Hg (Table 2), and pulse pressure at the midpoint arcade bridge is narrowed from about 40 mm Hg in intact muscle to about 5 mm Hg after exteriorization. When the pressure in the exteriorized muscle is measured at a site closer to the distal supply artery, mean and pulse pressures may even be lower. On the other hand, when arcade pressures are measured closer to the proximal feeder after the exteriorization, less of an effect is seen.
TABL
E 2
CREM
ASTE
R MU
SCLE
-PRO
XIMA
L FE
ED
ARTE
RY
MEAN
PR
ESSU
RE
Inta
ct
dist
al
feed
ar
tery
Liga
ted
dist
al
feed
ar
tery
Fem
oral
pres
sure
(mm
W
WKY
Mea
n *
SD
89
55
96
67
105
71
82
77
Feed
No
rmal
Arter
y fe
ed
pres
sure
pr
essu
re
(mm
W
(%
I 61
69
67
94
72
* 12
Diam
eter
km)
Fem
oral
Feed
ar
tery
pres
sure
pr
essu
re
(mm
W
(m
m
Hg)
126
106
45
80
100
60
100
83
45
96
112
64
Norm
al fe
ed
pres
sure
(%I 42
60
55
57
54
28
Diam
eter
ch
ange
Diam
eter
af
ter
0 km
) su
rger
y F
+ 12
3 %
15
6 0
166
+ 10
8 z
136
+36
140
+45
F
+53r
37
SHR
122
103
84
96
147
101
68
146
i-52
145
96
66
96
179
130
72
100
+4
146
103
70
96
169
138
81
100
f4
157
116
73
100
178
111
62
160
+60
Mea
n 2
SD
73
t 8
71
f 8
+30,
30

SYSTEMIC-MICROVASCULAR PRESSURE PROFILE 99
TABLE 3. ARTERIOLAR DIAMETERS AT SITE OF PRESSURE MEASUREMENTS (pm)
Spinotrapezius muscle (n = 3) Proximal supply artery Midpoint arcade bridge A Distal supply artery
Cremaster muscle (n = 3) Proximal supply artery Midpoint arcade bridge A Distal supply artery
Tibialis muscle (n = 3) Distal supply artery
Gracilis muscle (n = 4) Proximal supply artery Proximal arcade bridge Midpoint arcade bridge A Distal arcade bridge A Distal supply artery
WKY SHR
262 -c 33 (225-285) 345 k 54 (268-385) 155 2 24 (130-186) 108 ” 28 (74-139) 167 rt 58 (99-200) 184 k 3 (180-187)
197 2 12 (185-210) 203 2 15 (190-220) 92 k 15 (70-110) 82 2 22 (65-125) 82 k 12 (70-95) 100 k 10 (90-110)
205 2 9 (200-215) 283 k 21 (260-300)
211 L 44 (155-276) 162 f 15 (141-180) 87 k 19 (70-119) 85 ” 9 (76-101) 69 2 16 (50-97) 73 k 18 (45-90) 80 f 6 (72-90) 73 k 13 (65-96)
129 k 50 (90-218) 288 I 67 (167-370)
y Mean 2 SD (Range).
Vessel Diameter
The diameters m the current set of vessels were estimated under reflected light conditions and therefore may be associated with an error of about lo-20% of the mean value. More accurate measurements are possible under different light conditions using more complete exposure of the vessels, a step that was not desired in this study. Therefore the values listed in Table 3 are intended only to serve as a guideline for the size of vessels selected in this study. They are not intended to serve as a basis for a comparison between strains, since their accuracy and numbers are limited.
Mean Pressures in Intact Muscles
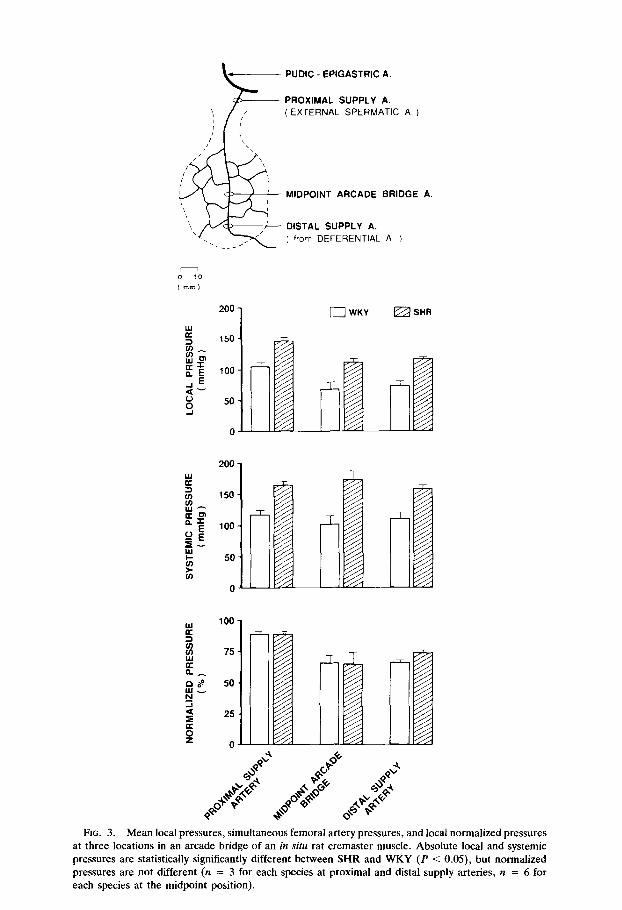
In contrast to the exteriorized muscle specimen, the pressures in the arteries and arteriolar arcade bridge of intact muscle were substantially higher. Figures 3, 4, and 5 display the mean pressures for SHR and WKY rats in the intact cremaster m., spinotrapezius m., and gracilis m., respectively. In spite of the fact that these three muscles are located in widely different regions of the circulation, the mean pressures in the supply artery and arcade bridge are only 10 to 30% below the femoral artery pressures. At the distal supply from the deferential artery and at the midpoint of the arcade bridge in the cremaster muscle pressure is slightly lower, about 65 to 75% of the femoral pressure. Comparatively high feed pressures were also found in the distal supply artery of the tibialis muscle, 89 k 11 mm Hg in WKY (n = 3) and 123 r 12 mm Hg in SHR (n = 3), levels which are 79 k 2% in WKY and 75 2 4% in SHR relative to the mean femoral pressure.
Pulse Pressures
In the majority of cases accurate readings of systolic and diastolic pressures could readily be obtained in the arteriolar arcade bridges and were found to be
PROXIMAL SUPPLY A. ( EXTERNAL SPERMATIC A )
MIDPOINT ARCADE BRIDGE A.
DISTAL SUPPLY A.
200
5 150
%- :I”
“E 100
i-
8 50
200,
, : f from DEFERENTIAL A i
c 2 150 b- gP og
100
H- w
cl 50
$I 0
5
100
B 75
Fe
0 WKY @j SHR
FIG. 3. Mean local pressures, simultaneous femoral artery pressures, and local normalized pressures at three locations in an arcade bridge of an in situ rat cremaster muscle. Absolute local and systemic pressures are statistically significantly different between SHR and WKY (P < 0.05), but normalized pressures are not different (n = 3 for each species at proximal and distal supply arteries, n = 6 for each species at the midpoint position).
~MIOPOINT ARCADE BRIDGE A. 101
KIMAL SUPPLY A.
%AL A. ‘\ \ ’ -\
;\ ------
/’ - THORACODC _-- -__ -- l
DISTAL SUPPLY A. ( from INTERCOSTAL A. )
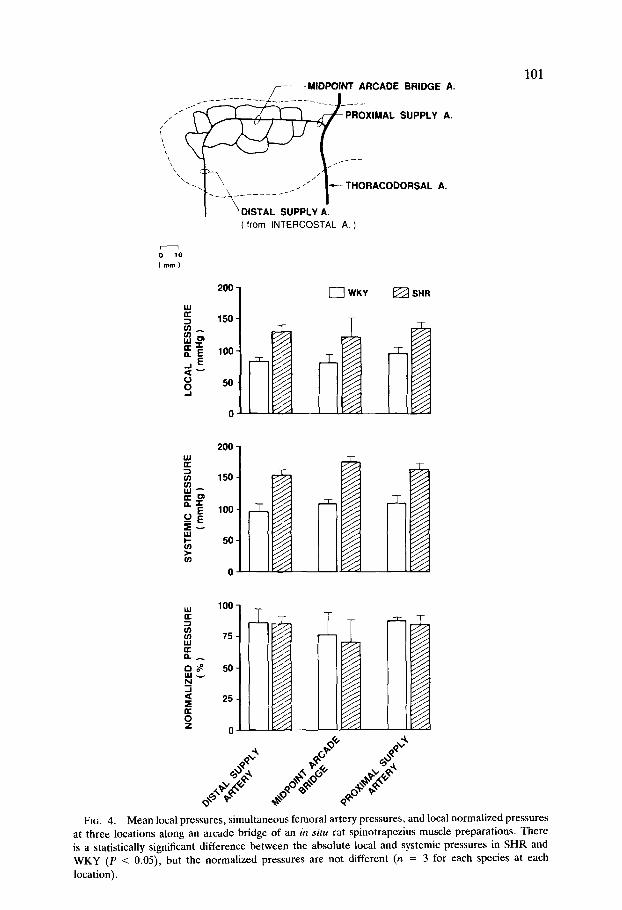
FIG. 4. Mean local pressures, simultaneous femoral artery pressures, and local normalized pressures at three locations along an arcade bridge of an in situ rat spinotrapezius muscle preparations. There is a statistically significant difference between the absolute local and systemic pressures in SHR and WKY (P < 0.05), but the normalized pressures are not different (n = 3 for each species at each location).
102 DISTAL SUPPLY A. ( SAPHENOUS A
FEMORAL A.
, 1 I I 1 DISTAL ARCADE I , / \
/
/
I ( MUSCULAR A. )
PROXIMAL ARCADE BRIDGE A.
MIDPOINT ARCADE BRIDGE A.
BRIDGE A.
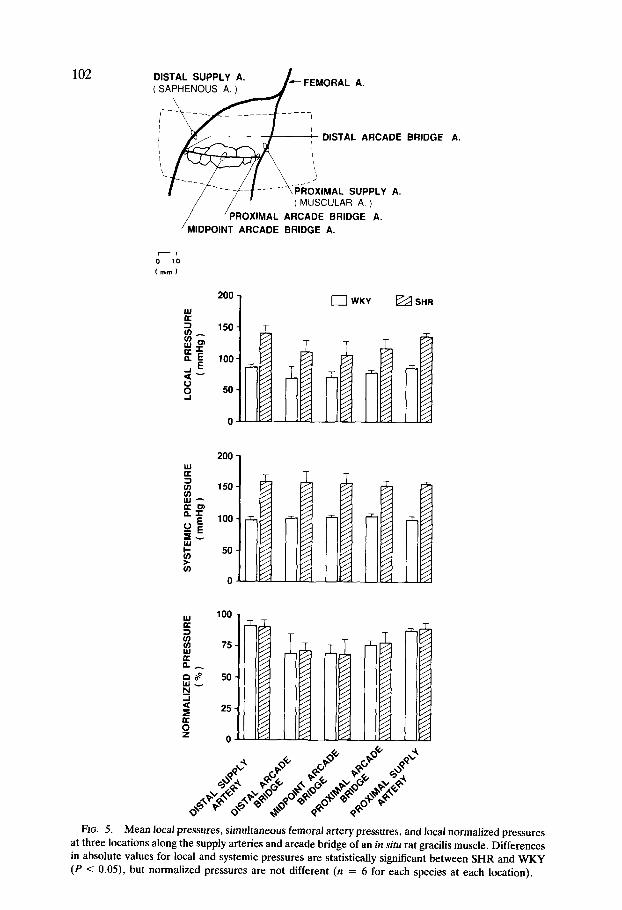
FIG. 5. Mean local pressures, simultaneous femoral artery pressures, and local normalized pressures at three locations along the supply arteries and arcade bridge of an in situ rat gracilis muscle. Differences in absolute values for local and systemic pressures are statistically significant between SHR and WKY (P < 0.05), but normalized pressures are not different (n = 6 for each species at each location).

SYSTEMIC.MICROVASCULAR PRESSURE PROFILE 103
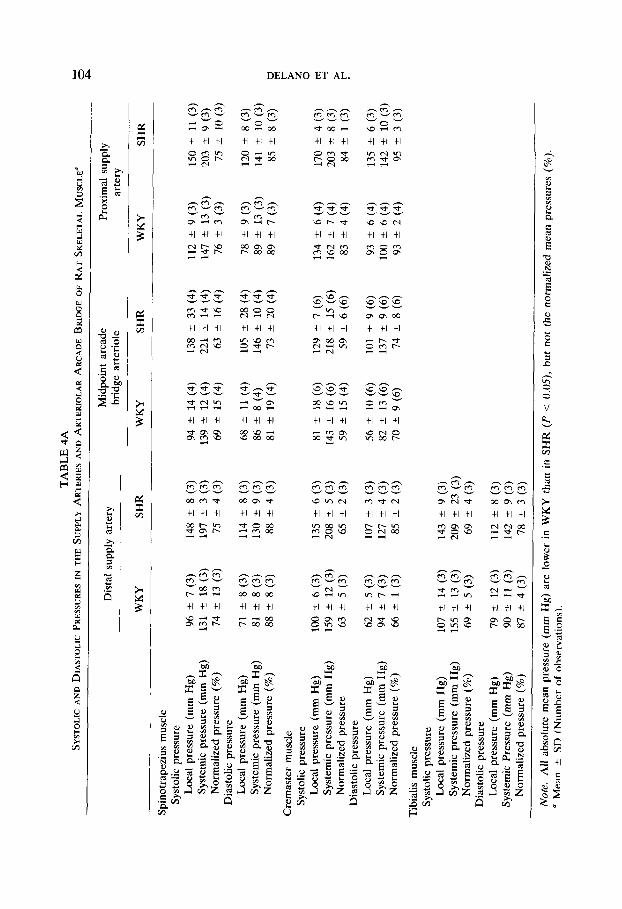
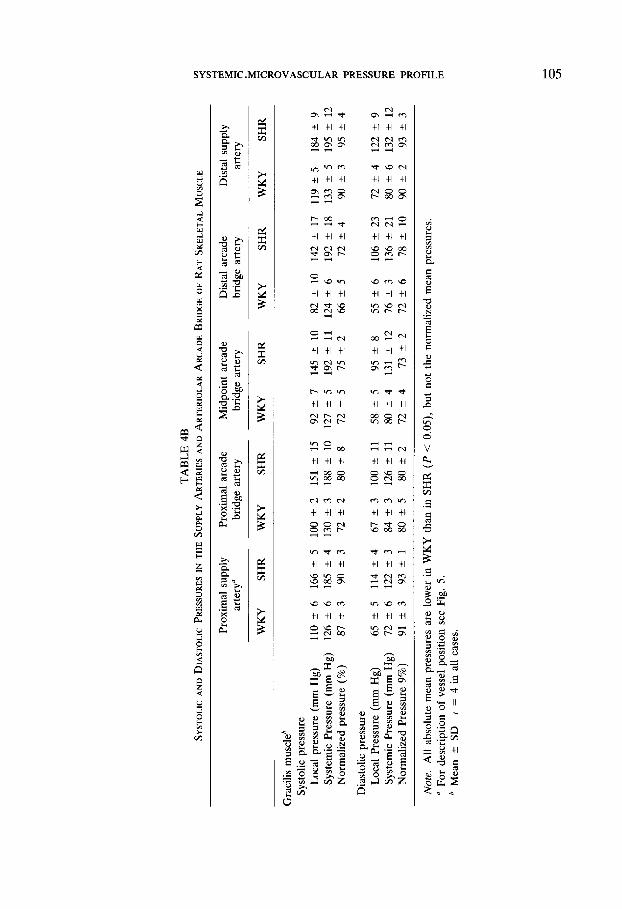
closely synchronized with the femoral artery pressure traces. The absolute and normalized values are summarized in Tables 4A, and 4B for the four different muscles. The normalized systolic and diastolic pressures (%) are comparable to those for the mean pressures, with no significant difference between systolic or diastolic normalized values. Pulse pressures which typically range between 30 and 40 mm Hg in the feeder arterioles, are still on average 28 and 37 mm Hg, respectively, at the midpoint of the arcade bridge for WKY and SHR. At the distal supply artery and midpoint of the arteriolar arcade bridge in the cremaster muscle, the pulse pressure is on average somewhat lower, about 25 mm Hg in WKY and about 28 mm Hg in SHR. None of the pulsatile pressures in the supply arteries fall below 60% of the systemic values (Table 4). In the majority of cases, measurements of the normalized systolic and diastolic pressures are at least 80% of systemic values. The pulsatile pressures near the center of the skeletal muscle, at the midpoint of the arcade bridges, typically range between 65 and 80% of the femoral pressures.
The Pressure in SHR Hypertensives
Despite some variation from animal to animal, there is no overlap of systemic or local absolute pressures (mm Hg) in the WKY and SHR strains and their mean values are significantly different. Nonetheless, when the pressure data are nor- malized, mean pressure values (%) (Figs. 3,4,5) or the normalized systolic or diastolic pressures (Table 4) are not significantly different in SHR and WKY rats. The pulsatile pressures follow a trend similar to the normalized mean pressures along the arteriolar arcade bridges, the proximal and distal values being quite similar, but with lower midpoint values (Table 4).
DISCUSSION
The objective of this study was to provide a detailed set of pressure measure- ments at the entry point to the microcirculation of skeletal muscle under conditions of minimal interference with circulation. Thus, all surgical procedures to expose the arteries were kept to a minimum and measurements were made while all supply arteries to the arcade arterioles were left intact. The principal conclusion drawn from this study is that feed pressures for the skeletal muscle microcirculation are on average only about 10 to 25% below the mean systemic pressure, irre- spective of the particular location of the skeletal muscle. Pressures in the feed arteries for a relatively remote structure, the tibialis muscle, are similar to those in the spinotrapezius muscle that lies in closer proximity to the heart. Systolic and diastolic pressures follow similar trends. Furthermore, the current results show relatively high arteriolar pressures over the entire length of the skeletal muscle along the arcade bridge. Since the arcade bridge is an integral part of the arteriolar arcade system, which in turn spans the entire muscle and serves to supply the transverse arterioles and their capillaries, such an arrangement ensures a remarkably uniform arteriolar pressure over the extent of the muscle, one of the key requirements for uniform perfusion of the vast capillary network.
Our values agree in general with previous measurements carried out in intact muscle specimens (Fronek and Zweifach, 1975; Hill et al., 1990) but are higher than those obtained in exteriorized muscle specimens (Table 1). Since exterior-
104 DELANO ET AL.
wmhl r?bN
tl fl $1 $1 fl tl
TABL
E 4B
SY
STO
LIC
AND
DIAS
TOLI
C PR
ESSU
RES
IN
THE
SUPP
LY
ARTE
RIES
AN
D AR
TERI
OLA
R AR
CADE
BR
IDGE
O
F RA
T SK
ELET
AL
MUSC
LE
2 2 Pr
oxim
al su
pply
Prox
imal
arca
de
Midp
oint
arca
de
Dist
al
arca
de
Dist
al
supp
ly E
arter
y” br
idge
arter
y br
idge
arter
y br
idge
arter
y ar
tery
8
WKY
SH
R W
KY
SHR
WKY
SH
R k
WKY
SH
R W
KY
SHR
ij T1
Gra
cilis
mus
cleb
Syst
olic
pres
sure
s %
Lo
cal
pres
sure
(m
m
Hg)
110
k 6
166
k 5
100
+-
2 15
1 2
15
92
2 7
145
k 10
82
+
10
142
2 17
11
9 “_
5
184
k 9
5 Sy
stem
ic Pr
essu
re
(mm
Hg
) 12
6 k
6 18
5 k
4 13
0 k
3 18
8 +
10
127
c 5
192
2 11
12
4 k
6 19
2 +
18
133
f 5
195
k 12
$
Norm
alize
d pr
essu
re
(%)
87
t 3
90
” 3
72
+-
2 80
k
8 72
2
5 75
k
2 66
k
5 72
2
4 90
-c
3
95
k 4
z Di
asto
lic
pres
sure
B
Loca
l Pr
essu
re
(mm
Hg
) 65
t
5 11
4 2
4 67
2
3 10
0 t
11
58
* 5
95
? 8
55
t 6
106
k 23
72
zt
4 12
2 k
9 2
Syst
emic
Pres
sure
(m
m
Hg)
72
? 6
122
k 3
84
k 3
126
k 11
80
k
4 13
1 k
12
76
k 3
136
k 21
80
2
6 13
2 k
12
g No
rmali
zed
Pres
sure
9%
) 91
t
3 93
2
1 80
k
5 80
k
2 72
k
4 73
k
2 72
k
6 78
k
10
90
2 2
93
r 3
z
Note
. Al
l ab
solu
te
mea
n pr
essu
res
are
lower
in
W
KY
than
in
SH
R (P
<
0.05
), bu
t no
t th
e no
rmali
zed
mea
n pr
essu
res.
8
a Fo
r de
scrip
tion
of
vess
el
posit
ion
see
Fig.
5.
E
’ M
ean
2 SD
1
= 4
in
all
case
s.

106 DELANO ET AL.
ization requires ligation of one or several of the supply arteries to the muscle, a situation is set up whereby the muscle remains well perfused but the pressures in the more distal arcade arterioles may fall. Ligation of the distal feeder in skeletal muscle results in a progressive fall of pressures along the arcade arterioles from the level prevailing at the proximal feeder down to its distal segment along the length of the muscle. The presence of even a relatively small feed arteriole serves effectively to raise the arcade arteriolar pressure even though it may not provide a large volume flow to the muscle microcirculation. Intravital studies that are carried out on exteriorized muscle specimen but are limited to a region close to the proximal feed artery are expected to encounter only a small compromise in the microhemodynamics due to the ligation of the distal feeder. On the other hand, if the studies are carried out in the vicinity of the ligated distal feeder, altered hemodynamics and vascular tone have to be expected.
Hill et al. (1990) have shown recently that the exteriorization of the cremaster muscle in young Sprague-Dawley rats leads to only a modest reduction of the proximal feed artery mean pressure (about 10 mm Hg), which is somewhat less than observed in the mature WKY rats (Table 2). The exact pressure reduction after such vessel ligation depends on the number of collateral vessels that feed the arteriolar arcades. In the SHR, which have high numbers of collateral vessels (Schmid-Schbnbein et al., 1986), the pressure reduction is much lower than in the WKY rats (Table 2). More importantly, Hill et aE. (1990) showed that the ligation of the distal feed artery leads to substantially reduced organ flow.
The high pressures indicated here point to the need to investigate the extent to which autoregulatory processes affect the arterioles of the microcirculation. Autoregulatory mechanisms should be investigated over a pressure range that includes systemic values. Myogenic control mechanisms may be most effective in the range between 50 and 120 mm Hg even in small arterioles, where the vessel wall media in these rat muscles contains only a single smooth muscle layer (Schmid- Schijnbein et al, 1990). The high mean and pulse pressures in these arterioles have important implications with respect to lymph flow. A remarkable charac- teristic of the larger arcade arterioles and the arcade bridges is their location closely paired with lymphatics, the two being deeply embedded in multiple muscle fibers (Skalak et al., 1984). Pressure pulsations associated with periodic elastic expansion and compression of these arterioles cause an inverse periodic volume change of the adjacent lymphatics. Due to the absence of lymphatic smooth muscle, the vessels cannot provide intrinsic spontaneous contractions needed to pump lymph fluid out of the muscle. Lymph flow in skeletal muscle can be facilitated by skeletal muscle contraction, but in light of the high pulse pressure and relatively high heart rates in this species, arcade arteriolar pulsations can serve as a lymph pump, even when the skeletal muscle is at rest.
The precise mechanism of the segmental reduction in the blood pressure remains a key unresolved issue in hypertension research. Is there a particular site along the central arteries or in the microcirculation for the major reduction in pressure‘? Such data are obviously basic to the understanding of the mechanism responsible for the elevated peripheral resistance in which the microcirculation appears to make a major contribution (Zweifach et al., 1981). There is a remarkable similarity of the pressure profiles in the SHR and WKY rats. When the arteriolar pressures are normalized with respect to the simultaneous systemic pressures, no detectable

SYSTEMIC-MICROVASCULAR PRESSURE PROFILE 107
differences are evident in supply arteries in the two animal strains. The data indicate that although the absolute pressures (mm Hg) in the hypertensive animals are reduced to a somewhat greater degree in the arteries proximal to the micro- circulation as compared with those in WKY, the principal pressure reduction in both species occurs in the microcirculation of the skeletal muscle per se. Previous measurements have shown that the pressures in the entry region to the capillaries, although not identical, are quite similar in WKY and SHR, suggesting that the elevation in systemic pressure in SHR is largely the result of changes at the arteriolar level (see summary in Schmid-Schonbein and Chien, 1986). In light of such observations it becomes evident that although the pressure control mecha- nisms in WKY and SHR are similar, absolute pressure reduction is higher in the SHR along the artery-arteriolar segment of the circulation, but the major burden of this activity still falls more on the microcirculation than on the systemic arteries.
The pressure distributions may in part be dependent on the particular choice of the anesthetic agent. In a separate study we examined the effect of an alternative anesthetic agent, 1% chloralose/l3.3% urethane, on the normalized pressures in the feeder artery and arteriolar arcade bridge of the gracilis muscle. Although this anesthetic agent decreases systemic arterial pressure in both WKY and SHR (6 and 13-week-old) by about 15%, the normalized pressures in both strains in the proximal and distal feeder arteries were 80 and 60%, respectively. These values are slightly but not significantly lower than those with alfaxalone in the major part of this study. In a comparison of the normalized pressures we also could not detect a difference with respect to age in the relative narrow range between 12 and 18 weeks.
Although the current investigation was limited to skeletal muscle, other organs may show similar pressure reduction profiles. The values reported for the large feeding arterioles, usually denoted as Al arterioles, are summarized at the bottom of Table 1. In the omentum, cheek pouch, and in the brain, the reported pressures in these large arterioles supplying the microcirculation are on average lower than the current values. Pressures in the modules and feed arteries of the mesentery are consistently higher than in the intestine (Table l), and both constitute a relatively reliable data set, since they have been derived from preparations with intact blood vessels. Even in the frog with its low central pressure, Landis (1926) reports only a small pressure reduction in the mesentery arteries compared to the systemic values. In the myocardium the pressures in the larger feed arteries (between 100 and 300 pm in diameter) are relatively high and comparable to those in skeletal muscle arcade bridges (Tillmanns et al., 1981; Nellis et al., 1981; Chilian et al., 1988). The pressure values in the wing of the unanesthetized bat show within a wide scatter no consistent trend. This feature may reflect the high density of the anastomosing arterioles and strong vasomotor tone of both the arterioles and the venules in this organ. Although the comparison summarized in Table 1 is incomplete due to a lack of precise specification of the measurement site within the vascular network, the data suggest that the feed pressures to the heart and skeletal muscle microcirculation are higher than those to the intestinal organs, like mesentery, intestine, omentum, or the brain. An important implication of such an observation with respect to hypertension is that different types of microcirculatory control mechanisms for pressure reduction may indeed be in- volved in muscles and in abdominal organs.

108 DELANO ET AL.
It may be argued that the high pressures in the arcade bridge of rat skeletal muscle are a phenomenon limited to the rat or other small mammalian creatures, and may not necessarily be representative of a large species, like man, where the central arteries are much longer. Such vessels, however, have considerably larger diameters. There are currently no comparable measurements in man, but if we compare current estimates of the pressure drops in the central circulation with realistic models of the arteries (Stettler et al., 1987) we are impressed that the large central conduits impose only a comparatively small pressure drop, suggesting that the general situation may not be unlike that reported here for rats.
ACKNOWLEDGEMENTS
This work was supported by NSF Grant DCB 88-19346 and by USPHS Grant HL-10881.
REFERENCES
ANDERSON, G. L., ACKLAND, R. D., SIEMINOW, M., AND MCCABE, S. J. (1988). Vascular isolation of the rat cremaster muscle. Microvasc. Rex 36, 56-63.
BAEZ, S. (1973). An open cremaster muscle preparation for the study of blood vessels by in vivo microscopy. Microvasc. Res. 5, 384-394.
BOHLEN, H. G. (1983). Intestinal microvascular adaptation during maturation of spontaneously hy- pertensive rats. Hypertension 5, 739-745.
BOHLEN, H. G., GORE, R. W., AND HUTCHINS, P. M. (1977). Comparison of microvascular pressure in normal and hypertensive rats. Microvasc. Res. 13, Z-130.
CHILIAN, W. M., EASTHAM, C. L., LAYNE, S. M., AND MARCUS, M. L. (1988). Small vessel phenomena in the coronary microcirculation: Phasic intramyocardial perfusion and coronary microvascular dy- namics. Prog. Cardiovasc. Disease 31, 17-38.
DAVIS, J. J. (1988). Control of bat wing capillary pressure and blood flow during reduced perfusion pressure. Amer. J. Physiol. 255, H1114-1129.
DAVIS, M. J., FERRER, P. M., AND GORE, R. W. (1986). Vascular anatomy and hydrostatic pressure profile in the hamster cheek pouch. Amer. J. Physiof. 250, H291-H303.
DELANO, F. A., SCHMID-SCH~NBEIN, G. W., AND ZWEIFACH, B. W. (1989). “Penetration” of the central blood pressure into the microcirculation of intact skeletal muscle in SHR and WKY rats. FASEB J. 3, 1383.
FRONEK, K., AND ZWEIFACH, B. W. (1975). Microvascular pressure distribution in skeletal muscle and the effect of vasodilation. Amer. J. Physiol. 228, 791-796.
GORE, R. W. (1974). Pressures in cat mesenteric arterioles and capillaries during changes in systemic arterial blood pressure. Circ. Res. 34, 581-591.
GORE, R. W., AND BOHLEN, H. G. (1977). Microvascular pressure in rat intestinal muscle and mucosal villi. Amer. J. Physiol. 233, H685-H693.
GORE, R. W., AND BOHLEN, H. G., (1982). Microvascular pressure in rat intestinal muscle. Amer. /. Physiol. 242, H268-H287.
GRAY, S. D. (1973). Rat spinotrapezius muscle preparation for microscopic observation of the terminal vascular bed. Microvasc. Res. 5, 395-400.
HARPER, S. L., AND BOHLEN, H. G. (1984). Microvascular adaptation in the cerebral cortex of adult spontaneously hypertensive rats. Hypertension 6, 408-419.
HENRICH, H., AND HECKE, A. (1978) A gracihs muscle preparation for quantitative microcirculatory studies in the rat. Microvasc. Res. 15, 349-356.
HILL, M. A., SIMPSON, B. E., AND MEININGER, G. A. (1990). Altered cremaster muscle hemodynamics due to disruption of the deferential feed vessel. Microvasc. Res. 39, 349-363.
HOUSE, S. D., AND JOHNSON, P. C. (1986). Microvascular pressure in venules of skeletal muscle during arterial pressure reduction. Amer. J. Physiol. 250, H834-H845.

SYSTEMIC-MICROVASCULAR PRESSURE PROFILE 109
INTAGLIETTA, M., PAWULA, R. F., AND TOMPKINS, W. R. (1970). Pressure measurement in the mam- malian microvasculature. Microvasc. Rex 2, 212-220.
JOYNER, W. L., DAVIS, M. J., AND GILMORE, J. P. (1981). Intravascular pressure distribution and dimensional analysis of microvessels in hamster with renovascular hypertension. Microvasc. Res. 22, 190-198.
LANDIS, E. M. (1926). The capillary pressure in frog mesentery as determined by micro-injections method. Amer. J. Physiol. 75, 546-570.
LINDBOM, L. (1983). Microvascular blood flow distribution in skeletal muscle. Acta Physiol. Stand. Suppl. 52, 525-532.
LIPOWSKY, H. H. (1975). In vivo studies of the rheology of blood in the microcirculation. Ph.D. Thesis, University of California, San Diego, University Microfilm No. 7.529446.
MEININGER, G. A., FEHR, K. L., AND YATES, M. B. (1987). Anatomic and hemodynamic characteristics of the blood vessels feeding the cremaster skeletal muscle in the rat. Microvasc. Res. 33, 81-97.
MEININGER, G. A., FEHR, K. L., YATES, M. B., BORDERS, J. L., AND GRANGER, H. J. (1986). Hemo- dynamic characteristics of the intestinal microcirculation in renal hypertension Hypertension 8, 66- 75.
MEININGER, G. A., HARRIS, P. D., AND JOSHUA, I. G. (1984). Distributions of microvascular pressure in skeletal muscle of one-kidney, one-clip, two-kidney, one-clip and deoxycorticosterone-salt hy- pertensive rats. Hypertension 6, 27-34.
MYRHAGE, R. (1977). Microvascular supply of skeletal muscle fibers. Actu Orthop. Stand. Suppl. 168. NELLIS, S. H., LIEDTKE, A. J., AND WHITESELL, L. (1981). Small coronary vessel pressure and diameter
in an intact beating rabbit heart using fixed-position and free-motion techniques. Circ. Res. 49, 342-353.
NICOLL, P. A. (1969). Intrinsic regulation in the microcirculation based on direct pressure measure- ments. In “Microcirculation,” (W. L. Winters and A. N. Best, Eds.), Chap. 6, pp. 89-101. Thomas, Springfield, IL.
ROY, J. W., AND MAYROVITZ, H. N. (1984). Microvascular pressure, flow and resistance in sponta- neously hypertensive rats. Hypertension 6, 877-886.
SCHMID-SCH~NBEIN, G. W., AND CHIEN, S. (1986). The microcirculation in hypertension. In “Handbook of Hypertension,” (A. Zanchetti, and R. C. Tarazi, Eds.), Vol. 7, Chap. 25. Elsevier Amsterdam.
SCHMID-SCH~NBEIN, G. W., DELANO, F. A., CHU, S., AND ZWEIFACH, B. W. (1990). Wall structure of arteries and arterioles feeding the spinotrapezius muscle of normotensive and spontaneously hypertensive rats. Int. J. Microcirc. Clin. Exp. 9, 47-66.
SCHMID-SCH~NBEIN, G. W., FIRESTONE, G., AND ZWEIFACH, B. W. (1986). Network anatomy of arteries feeding the spinotrapezius muscle in normotensive and hypertensive rats. Blood Vessels 23, 34-49.
SCHMID-SCHBNBEIN, G. W., SKALAK, T. C., AND FIRESTONE, G. (1987). The microvasculature in skeletal muscle. V. The arteriolar and venular arcades in normotensive and hypertensive rats. Microvasc. Res. 34, 385-393.
SHAPIRO, H. M., STROMBERG, D. D., LEE, D. R., AND WIEDERHIELM C. A. (1971). Dynamic pressures in the pial artery microcirculation. Amer. J. Physiol. 221, 279-283.
SKALAK, T. C., SCHMID-SCHBNBEIN, G. W., AND ZWEIFACH, B. W. (1984). New morphological evidence for a mechanism of lymph formation in skeletal muscle. Microvasc. Res. 28, 95-112.
SMAJE, L., ZWEIFACH, B. W., AND INTAGLIETTA, M. (1970). Micropressure and capillary filtration coefficients in single vessels of the cremaster muscle of the rat. Microvasc. Res. 2, 96-110.
SPALTEHOLZ, W. (1888). Die Verteilung der Blutgefasse in Muskel. Abh. Siichs. Ges. Wiss. Math. Phys. Kl. 14, 509-528.
STETTLER, J. C., NIEDERER, P., AND ANLIKER, M. (1987). Nonlinear mathematical models of the arterial system: Effects of bifurcations, wall elasticity, stenoses, and counterpulsation on pressure and flow pulses. In “Handbook of Bioengineering,” (R. Skalak and S. Chien, Eds.), Chap 17, pp 17.1- 17.26. McGraw-Hill, New York.
SUTTON, D., AND SCHMID-SCH~NBEIN, G. W. (1989). Hemodynamics at low flow in the resting, va- sodilated rat skeletal muscle. Amer. J. Physiol. 257, H1419-H1427.
TILLMANNS, H., STEINHAUSEN, M., LEINBERGER, H., THEDERAN, H., AND K~~BLER, W. (1981). Pressure measurements in the terminal vascular bed of the epicardium of rats and cats. Circ. Res. 49, 1202- 1211.
WIEDERHIELM, C. A., AND WESTON, B. V., (1973). Microvascular, lymphatic and tissue pressure in the unanesthetized mammal. Amer. J. Physiol. 225, 992-996.

110 DELANO ET AL.
WIEDERHIELM, C. A., WOODBURY, J. W., KIRK, S., AND RUSHMER, R. F. L. (1964). Pulsatile pressure in microcirculation of the frog’s mesentery. Amer. J. Physiol. 207, 173-176.
ZWEIFACH, B. W. (1974). Quantitative studies of microcirculatory structure and function. I. Analysis of pressure distribution in the terminal vascular bed of cat mesentery. Circ. Res. 34, 843-857.
ZWEIFACH, B. W., KOVALCHECK, S., DELANO, F., AND CHEN, P. (1981). Micropressure-flow rela- tionships in a skeletal muscle of spontaneously hypertensive rats. Hypertension 3, 601-614.
ZWEIFACH, B. W., AND LIPOWSKY, H. H. (1977). Quantitative studies of microcirculatory structure and function. III. Microvascular hemodynamics of cat mesentery and rabbit omentum. Circ. Res. 41, 380-390.
ZWEIFACH, B. W., AND RICHARDSON, D. R. (1971). “Microcirculatory adjustment of pressure in mesentery. 6th Europ. Conf. Microcirc., Aalborg 1970,” pp. 248-253, Karger, Basel.
Related Documents