New hominoid specimens from the Middle Miocene site at Paqalar, Turkey Introduction ‘l’he fossil site at Pagalar is situated on the edge of the Giinen basin in IVestern Anatolia about 75 km west south west of Bursa (see Andrews 8r Alpagut, 1990). ‘T’he sediments art exposed in the side of a road cut, which led to their discovery by the Turkish-German lignite survey in 1968. Fossils collected there during the 1960s were described b), Sickenberg (1975), and the primates by Andrews & Tobicn (1977). who assigned the hominoids to Ramupithecus wickeri and S’irxzpitherus darwini for the small and large specimens respectively. Subsequently, Ramapithecus has been synonymized with Sizapithecus (e.g., Greenfield, 1979; Andrews & Cronin, 1982), with the specimens from East Africa no& distinguished from SirlapithecuslRamapithecus and returned to their original genus Keryupithecus. Synonymy is maintained with Siwpithccus alpani (Tekkaya, 1974) based on the type specimen from Candor. Both small and large specimens are now attributed to .S. ciurzelini (Abel, 1902) and grouped with S. alpani 2~s 21 single highly dimorphic species. ‘l’hrre is evidence of a second species at Pagalar, which will be left unnamed in the present work. In 1983, fieldwork was recommenced at Pagalar and has proceeded annually since that time. This has led to the recovery of many fossil mammals and to a more complete understanding of the geology of the site (Andrews & Alpagut, 1990), the nature (~1‘ the sedimentary environment (Bestland, 1990) and the taphonomic processes that have influenced the composition of the Pagalar fauna (Andrews 8r Erso);, 1990). For the * -I‘<, whom corrrapondencc should hr addrcsscd

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

New hominoid specimens from the Middle Miocene site at Paqalar, Turkey
Introduction
‘l’he fossil site at Pagalar is situated on the edge of the Giinen basin in IVestern Anatolia
about 75 km west south west of Bursa (see Andrews 8r Alpagut, 1990). ‘T’he sediments art
exposed in the side of a road cut, which led to their discovery by the Turkish-German
lignite survey in 1968. Fossils collected there during the 1960s were described b),
Sickenberg (1975), and the primates by Andrews & Tobicn (1977). who assigned the
hominoids to Ramupithecus wickeri and S’irxzpitherus darwini for the small and large specimens
respectively. Subsequently, Ramapithecus has been synonymized with Sizapithecus (e.g.,
Greenfield, 1979; Andrews & Cronin, 1982), with the specimens from East Africa no&
distinguished from SirlapithecuslRamapithecus and returned to their original genus
Keryupithecus. Synonymy is maintained with Siwpithccus alpani (Tekkaya, 1974) based on
the type specimen from Candor. Both small and large specimens are now attributed to .S.
ciurzelini (Abel, 1902) and grouped with S. alpani 2~s 21 single highly dimorphic species. ‘l’hrre
is evidence of a second species at Pagalar, which will be left unnamed in the present work.
In 1983, fieldwork was recommenced at Pagalar and has proceeded annually since that
time. This has led to the recovery of many fossil mammals and to a more complete
understanding of the geology of the site (Andrews & Alpagut, 1990), the nature (~1‘ the
sedimentary environment (Bestland, 1990) and the taphonomic processes that have
influenced the composition of the Pagalar fauna (Andrews 8r Erso);, 1990). For the
* -I‘<, whom corrrapondencc should hr addrcsscd

flurposes of the present paper two majnr conclusions have rcsultctl. First. the \r~tlimc~nt~
and incfudcd fossils were all deposited during a single depositional r\.cnt ot‘rxtrcmc.l\ shlJrt
duration (Andrcws Sr Afpagut. 1990: Xntfrr\~s 8i Erso)-. 1990). Scconcf, the c\itf&,~ ot
regional geology (AndreLvs Br Xfpagut. 1990) and from the sediments (Bestland. 1990~
indicates that the fossils were derived from no more than a few kilomctcra distant Ii-om tllc,
present site. These conclusions arc significant for the tasonomic interpretation of thts
hominoids as it indicates that they represent a nearly synchronous group of animals, so
that the effects of time a\,eraging on increasing sample \-ariahifit) is minimal. and th<
restricted geographical. and probahl>- ecological extent of the catchment area alacj
minimizes the effects of geographic variation. Both of these facts mean that the Paaafar
fossils represent a very close approximation to a living community of animals. \Z’e feel that
this has implications for the selection of appropriate models of species definition.
Specimen numbers used in this paper ha\-e two kinds of prelises. First, tht, prefix BP
denotes material collected prior to our project. which is currently at the British Museum
(Natural History). Second, the material that \ve have collected since 1983 is prcfised by the
fetters A through G. with A denoting 1983, B 1984 and so on up to the 1989 collection.
which is prefixed G. This matriaf is housed in the Department of Pafeoanthropoloq. at tht
University of Ankara. Abbreviations used in the paper are as foffoMs: km = kilomettars:
mm = miflimcters; MD = mesio-distal length: BL = hucco-lingual breadth; max. L =
maximum length: pcrp B = perpendicular breadth.
Description of morphology
The morphology of the Pagalar hominoids is described with reference to grout’s of‘
homologous elements that are morphologically homogeneous. Individual specimens will
not be described unless they provide information not available from the general
descriptions, and the descriptions themselves will be based on the kvhofe range of
specimens available for that body part. The reasons for this will become clear as \ve
proceed, for although there is considerable variation present in the sample of hominoids
from Paaalar, it is not such as to make it obvious either how many species are present at the
site or how they should be defined. In this paper we have concentrated on providing a
description of morphology expanded from that provided by Andrews & Tobien ( 1977) that
is possible as a result of the greatly enlarged sample of material now available (Table 1).
We also mention aspects of metrical and morphological variation that relate to
considerations of alpha taxonomy. Our numbers of specimens are based on complete,
measurable specimens, and we should emphasize that there is an additional sample of
several hundred tooth fragments that can be clearly identified as hominoid and to gross
tooth types (upper molar, lower molar, etc.) but whose exact determination is not possible.
Such specimens may have considerable value for other studies such as dental microw-ear or
dental growth and development.
Ranges of variation are compared numerically by the index of maximum and minimum
values (Martin, 1983; Martin & Andrews. 1984). This has the advantage that varying
sized samples can be compared with only minor loss of information, and there is no
arbitrary division into males and females. The largest specimen in a sample is, however.
most likely to be a male, and the smallest is most likely to be a female (which has been
confirmed empirically, Martin, 1983). and so by computing maximum/minimum x 100,
an index that parallels the sexual dimorphism index is obtained. The values for this index
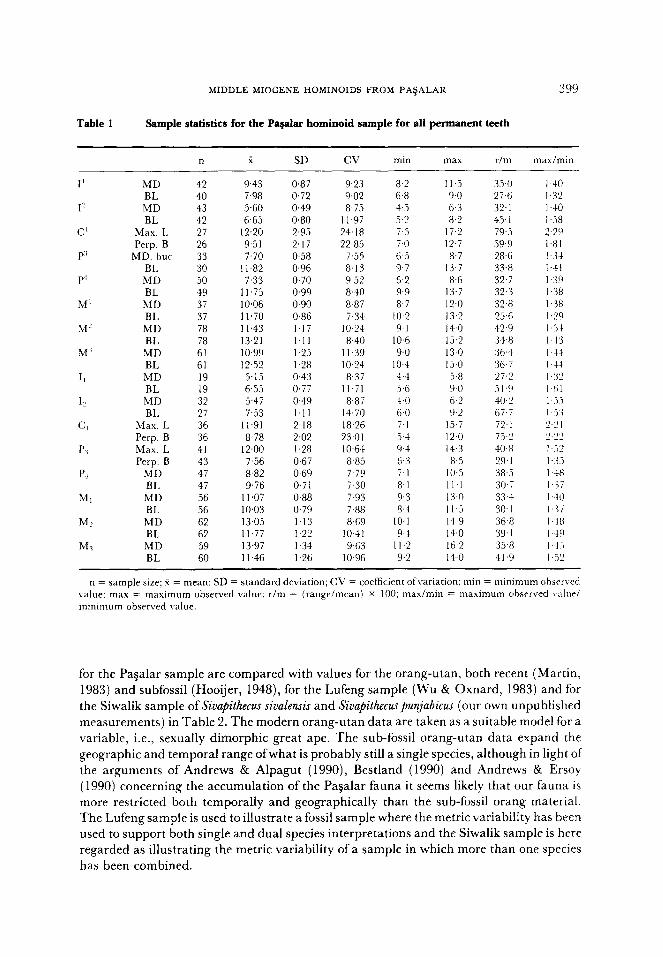
Table 1
MIDDLE MIOCENE HOMINOIDS FROM PA$ALAR 399
Sample statistics for the Pagalar hominoid sample for all permanent teeth
n x SD cv min max r/m max/min
I’
I-’
C’
P”
P’
M’
W
M’
I,
12
c ‘I
PI
P,
Ml
MC
M1
MD 42 BL 40 MD 43 BL 42
Max. I, 27 Perp. B 26
MD. hut 33 BL 30 MD 50 BL 49 MD 37 BL 37 MD 78 BL 78
MD 61 BL 61 MD 19 BL 19 MD 32 BI. 27
Max. L 36 Perp. B 36 Max. L 41 Perp. B 43
MD 47 BL 47 MD 56 BL 56 MD 62 BL 62 MD 59 BL 60
9.43 0.87 9.23 8.2 11.5 35.0 7.98 0.72 9.02 6.8 9.0 27.6 5.60 0.49 8.75 4.5 6.3 32.1 6.65 0.80 11.97 5.2 8.2 45.1
12.20 2.95 24.18 7.5 17.2 79.5 9.51 2.17 22.85 7.0 12.7 59.9 7.70 0.58 7.55 6.5 8.7 28.6
I I .82 0.96 8.13 9.7 13.7 33.8 7.33 0.70 9.5? 6.2 8.6 32.7
11.75 0.99 8.40 9.9 13.7 32.3 10.06 0.90 8.87 8.7 I?,0 32.8 11.70 0.86 7.34 10.2 13.2 25.6 11.43 1.17 10.24 9.1 14.0 42.9 13.21 1.11 8.40 IO.6 15.2 34.8 10.99 1.25 11.39 9.0 13.0 36.4 12.52 1.28 IO.24 10.4 15.0 36.7 5.15 0.43 8.37 4.4 5.8 27.1 6.55 0.77 11.71 3.6 9.0 5 1 9 5.47 0.49 8.87 4.0 6.2 40.2 7.53 1.11 14.70 6.0 9.2 6J.i
11.91 2.18 18.26 7.1 15.7 72, I 8.78 2.02 23.01 5.4 12.0 73.2
12.00 1.28 1 0+x 9-l 14.3 40.8 7.56 0.67 8.85 6.3 8.5 29.1 8.82 0.69 7.79 7.1 IO.5 38.3 9.76 0.71 i.30 8.1 Il.1 30.7
Il.07 0.88 7.93 9.3 13.0 33.4 10.03 0.79 J.88 8.4 11.5 30.1 13.05 1.13 8.69 10.1 14.9 36.8 11.77 1.22 1041 9.4 14.0 39.1 13.97 1.34 9.63 Il.2 16.2 35.8 11.46 1.26 10.96 9.2 14.0 41.9
I ,40 1.32 l-&O 1 ..%3 2.29
l-al I-i.+ I.41 1 .‘W I.38 I.38 I.29 I ..iS 1.411 1 A l-44 I.‘32 l.lil
1 ,.i.j I i I 2.2 I ‘).‘)‘J _ -_ I i9 I..(i I 4 I -.ii I .+I! I.‘<7 l-k8 I ,*9 14-1 1 .il’
n = sample size; 4 = mean; SD = standard deviation; CV = coefficient ofvariation; min = minimum obscrvrd value: max = maximum observed value; r/m = (range/mean) x 100: maximin = maximum ohsrrved vale/ minimum observed value.
for the Pagalar sample are compared with values for the orang-utan, both recent (Martin, 1983) and subfossil (Hooijer, 1948), for the Lufeng sample (Wu & Oxnard, 1983) and for the Siwalik sample of Sivapithecus sivalensis and Sivapithecuspunjabicus (our own unpublished
measurements) in Table 2. The modern orang-utan data are taken as a suitable model for a variable, i.e., sexually dimorphic great ape. The sub-fossil orang-utan data expand the geographic and temporal range ofwhat is probably still a single species, although in light of the arguments of Andrews & Alpagut (1990), Bestland (1990) and Andrews & Ersoy
(1990) concerning the accumulation of the Pagalar fauna it seems likely that our fauna is more restricted both temporally and geographically than the sub-fossil orang material. The Lufeng sample is used to illustrate a fossil sample where the metric variability has been used to support both single and dual species interpretations and the Siwalik sample is here regarded as illustrating the metric variability of a sample in which more than one species has been combined.
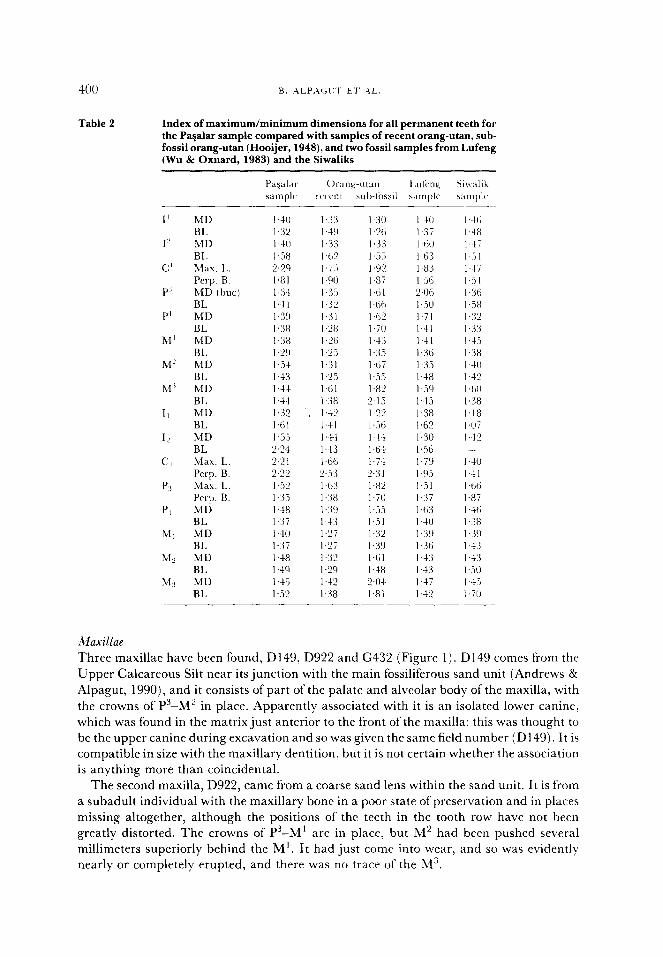
Table 2 Index of maximum/minimum dimensions for all permanent teeth for the Pavalar sample compared with samples of recent orang-utan, sub- fossil orang-utan (Hooijer, 1948), and two fossil samples from Lufeng (Wu & Oxnard, 1983) and the Siwaliks
I’ MD BL
I MD BL
c’ Max. 1~. Pcrp. B.
P’ MD (but) BL
P’ MD BL
M’ MD BL
M’ MD BL
M” MD BL
II MD BL
IL MD BL
c:, Max. L. Perp. B.
Pi Max. I,. Perp. B.
PI MD BL
Ml MD BL
Mz MD BL
MS MD BL
I .30
1.X I ,3:i I ,.i.i I.92 I.8i I.til 1.61; I.62 I.70 143 1~3.‘, l,tii I.55 I.82 2.15 1 .“‘j I ,.Xi 144 14s 1.7-l 2.3 I I.82 I.70 1,.X I.51 1.3’ I39 I.61 1.48 “al 1.81
Maxillae
Three maxillae have been found, D149, D922 and G432 (Figure 1) . D 149 comes from the
Upper Calcareous Silt near its junction with the main fossiliferous sand unit (Andrews &
Alpagut, 1990), and it consists of part of the palate and alveolar body of the maxilla, with
the crowns of Ps-M’ in place. Apparently associated with it is an isolated lower canine,
which was found in the matrix just anterior to the front of the maxilla: this was thought to
be the upper canine during excavation and so was given the same field number (D149). It is
compatible in size with the maxillary dentition, but it is not certain whether the association
is anything more than coincidental.
The second maxilla, D922, came from a coarse sand lens within the sand unit. It is from
a subadult individual with the maxillary bone in a poor state of preservation and in places
missing altogether, although the positions of the teeth in the tooth row have not been
greatly distorted. The crowns of P3-M’ are in place, but M’ had been pushed several
millimeters superiorly behind the M’. It had just come into wear, and so was evidently
nearly or completely erupted, and there was no trace of the Ms.

MIDDLE MIOCENE HOMINOIDS FROM PXj”L”R
Figure I. Three maxillary specimens from Pa&w, from top tu bottom specimen numbers G432. II149 CYC DYE. G432 has the crowns of P’ to Ms. the palatal process to the midline, and part of the maxilIar\ sinus. DIN has the crowns of P’ to bf’ in a ~wy distorted palate. D922 has the same teeth. tvith mo$t of’ thr maxillarv bone now destroved.

Thr third maxilla, G432, is both the one found most recently and the moat c~omplctc~. 1 t ih from the right side and preserves the crowns of P’, Pt and M’ in a portion of a palate, \% itI+
M’ and M” present but detached from the palate. The palate is complete to the palatal
suture from P+ to M’, and the floor of the maxillary sinus is exposed above P+ to hl’. The
palate is shallow, 6.7 mm deep at the level of M’. and palatal width between the innet
border of M’ and the midline is 12.4 mm, giving an estimated palatal width of 24.8 mm
The floor of the maxillary sinus is high and gently concave, with the roots of the molars not
intruding into it.
In addition to these three maxillae, an upper tooth row has been fijund, G239. This
consists of the crowns of MI-M” with no trace of maxillary bone or tooth roots. The three
teeth were associated in the sediments, their contact wear facets match perfectly, and they
are of comparable size and morphology, all of which provides good support for their
association. Other isolated teeth have also been matched on the basis of size and state of
wear, but these cannot be used with confidence for between-tooth comparisons.
The morphology of the teeth in these specimens will be described together with the
individual dental descriptions. They help to establish differences between M’ and M’.
which are not always easy to distinguish in isolated teeth in the absence of dual mesial
contact facets (see below), and they also help to show association between premolar/molar
morphotypes. \$Xthin the molars, M2 is considerably longer mesiodistally than M’ but
only marginally broader buccolingually, and it is always larger than M’. It is also usually
larger than M”. P3 is fra t c ionally larger than P”: crown module ratios for the three maxillae
range from 1 .Ol to 1.04. Ratios of P3 to M’ for the same specimens are 0.86 to 0.89; and of
P” to M’ are 0.83 and 0.86. These are well within the normal range for Simpithecus and most
other hominoids, and they differ from the derived condition seen in Ken~apithecm- and
Afropithecus, two genera of African hominoids of similar age to, or slightly older than the
Pagalar hominoids (Andrews & Martin, 1987).
Mandible
Two mandibles have been recovered from the Pagalar deposits (Figure 2). One has a
fragment ofbone with the crowns of P+ and Mt (numbered E697), but the other is an intact
symphysis with the left mandibular body and crowns of It to P+ and part of the right
mandibular body (Gl313). The crown morphology of the teeth will be described in their
place with the isolated teeth, but a brief description of the mandibular morphology of
Gl313 will be given here.
The symphysis is very long and shallow sloping. Its inner surface is intact except for the
inferior edge, which has been slightly crushed and broken (Figure 2). The outer surface is
broken around the alveolar region of the incisors, so that only the extreme tip of the 12 is
covered by bone. Similarly, the inferior border of the outer plane of the symphysis is
damaged. The upper plane of the internal surface is flat, with a gentle slope to a superior
transverse torus at the level of P+. At this point the symphyseal contour plunges steeply
downward and then flattens out again to form an extremely prominent and extended
inferior transverse torus, which extends to beyond the distal limit of the M+ (the alveolar
border in this region is damaged, and the crown is not preserved, but the inferior border of
the mandible is preserved intact). It seems likely that the inferior torus extends to about the
middle of MP, which is greatly in excess of the posterior extension of the torus in most
hominoids. The orang-utan is the only living hominoid that shows this combination of
shallow sloping symphyseal plane and posterior extension, and it has also been described

MIDDLE MIOCENE HOMINOIDS FROM P:\~“IAR 4)3
as a characteristic feature of Ken~upithecus (Andrews, 197 1). The conformation of the inner
plane of the sy-mphysis is remarkably similar to that of &yapithecu.r and differs from other
fossil hominoids. and this emphasizes the similarity of the Pasalar hominoids to this genus
already observed for the Candor mandible (Andrews & Tekkaya, 1976).
The body of the mandible is also high and slender. The estimate ofdepth in the region of
Ml is 35 mm, with the comparable breadth being 16.2 mm. This compares with the
symphysis dimensions of 39.7 and 21.6 mm. The total length of the symphysis is 54.2 mm.
Roth symphysis and mandibular body are more slender than is usual in later S’irv7~ithrc~u.r
from Indo-Pakistan (Kay. 1982: Kelley. 1988).
0 I
cm ! 0 . ! cm?
Fiqurr 2. TMYJ mandible fragments from Pagalar, on the left G13 I3 with the symphysis prcsrrwd and tht,
crown\ of 1, tcl P,. and on the right E697, a fragment of mandiblr nit11 crams of P, tu hI,.
I ;bpes central incisors
There is a sample of42 upper central incisors from the Pasalar deposits. Five of these came
from the earlier German excavations, and were described briefly by Andrews & Tobien
(1977), but the remainder have come from our excavations. None is associated with the
posterior dentition or with any mandibles or maxillae. There are two distinct size classes
with two different morphologies in each size class.
In morphology I the tooth is moderately high crowned and spatulate (Figure 3). It has a

a Figure 3. Pagalar upper central incisors showing ranges of size and morphological variation.
Morphology I is shown on BP32 top left. All thr uther specimens show morphology II in varying degrws
of wear: BP1301, top row middle, is the smallest and C t-16 bottom left, is thr largest.
diffuse lingual swelling at the base of the crown which appears to be an outgrowth of the
lingual cingulum in that traces of its origin from a pillar-like structure may still be seen.
The lingual swelling is V-shaped in outline, broader towards the apex of the crown than
cervically but becoming less distinct as it broadens. Cervically, the swelling narrows to a
point that is either continuous with the narrow lingual cingulum, which is also raised at
this point, or is in close contact with it. The swelling is most sharply defined at this point.

MIDDLE MIOCENE HOMINOIDS FROM PA~ALAR 10.-l
and the narrowness of the swelling and the adjoining cingulum gives the crown an
appearance of being relatively narrow when viewed from the lingual side. This impression
is not justified by the overall measurements of the crown, and because of the lack of
repeatable landmarks on this part of the crown, it has not been possible to demonstrate this
feature with more precise measurements. FVc have tried measuring height and breadth of
the swelling for comparison with similar measurements of the lingual pillar present in
morphology II, but this has proved ineffective. The swelling becomes shallower ah it
broadens out towards the top of the crown, so much so that it rarely has any trace ofwcar
on it. In the teeth with this morphology, wear is restricted to the incisiveedge, extent1in.e 011
to the lingual surface as far as but not usually on to the linqual swelling.
Morphology II is present on more than 90% of the teeth in the PaSalar sample. The
(‘rowns are relatively more robust and ha1.c a prominrnt lingual pillar (Fi,gurc 3). The
length/breadth dimensions of the crown show it to be huccolingually deep compared M.ith
tnesiodistal length, but it is not certain whether this is because ofa reduction in mesiodistal
length or an expansion of the buccolingual dimension (Table 1). The comparison of‘ crown
height shows that the crown is higher relative to mesioclistal length than is the casr li)l
morphology I, and it may be that both this and the apparent greater robusticity can IW
attributed to the same phenomenon, namely reduction in mediodistal length. The IiIlgual
pillar is well defined throughout its length, and it runs f?om the base of the crown. M hcrc it
is continuous with the very narrow lingual cingulum, to close to the incisive ed,qc gradualI>
becoming narrower in the process. This is the opposite of rhc shape of the lingual swelling
in morphology I, which gets broader towards the incisix-e ed,ge of the crown. It is also
different in that the pillar becomes more prominent incisally as well. and as a rt*suit it
nearly always shows signs of wear as it encroaches from the incisive edge on the lingual
surface of the crown. The degree ofwear is frequently so strong as to completeI)- oblitc’ratc,
the whole of the pillar, leaving a pillar-shaped concave dentine pit in its place (set* Figure%
3).
In comparison with other hominoids, there is a striking similarity between morpholog)-
II and that described for Proconsul (Andrews. 1978). Roth have a prominent lingual pillar.
although the Pagalar teeth are considerably more robust. They are more robust also than
the single I’ known for Dyopithecus fontani (an undescribed specimen from La Gri\-c.
currently on loan to the B.M.N.H.), which also has a prominent lingual pillar. It seems
quite possible that this morphology represents the retention in the Pasalar material of an
ancestral hominoid morphology. It may also be that the other morphology seen in the
PaSalar sample, with a lingual swelling rather than a pillar. could be deri\,ed from this
samr ancestral morphology. It has not bepn observed in any other hominoid. fossil 01’
modern, so that it appears to be unique to this part of the Paqalar sample. This
morphological variation is not within the normal variation limits present in other primates.
The two morphologies are quite distinct. with no intermediates found so far, and since the
sample size is better than is available for most samples of fossil hominoid. the lack ot
intermediates must be considered significant.
Size Lrariation in the PaSalar upper central incisors is of the same order as the size
\.ariation ofthe same teeth in recent orang-utans, although considerably smaller in terms 01‘
absolute size, and it exceeds the variation in sub-fossil orang-utans. This variation is
comparable in overall extent to that of the whole of the Siwalik sample of central incisors.
which on most interpretations contains at least two species (Greenfield, 1979; Kay, 1982:
Kelley & Pilbeam. 1986). The size variation does not correspond with morphological
406 B. ALPACU? E7‘.4L. T
differences: i.e., both morphologies are present in both size classes. Tasonomic
interpretation of this variation is problematic, for if two species are recognized in the
Pagalar sample on the basis of morphological differences, they would have completely
overlapping ranges of metrical variation; on the other hand, if only one species is
represented, it would have a degree of variation considerably greater and different from
that seen in other hominoids.
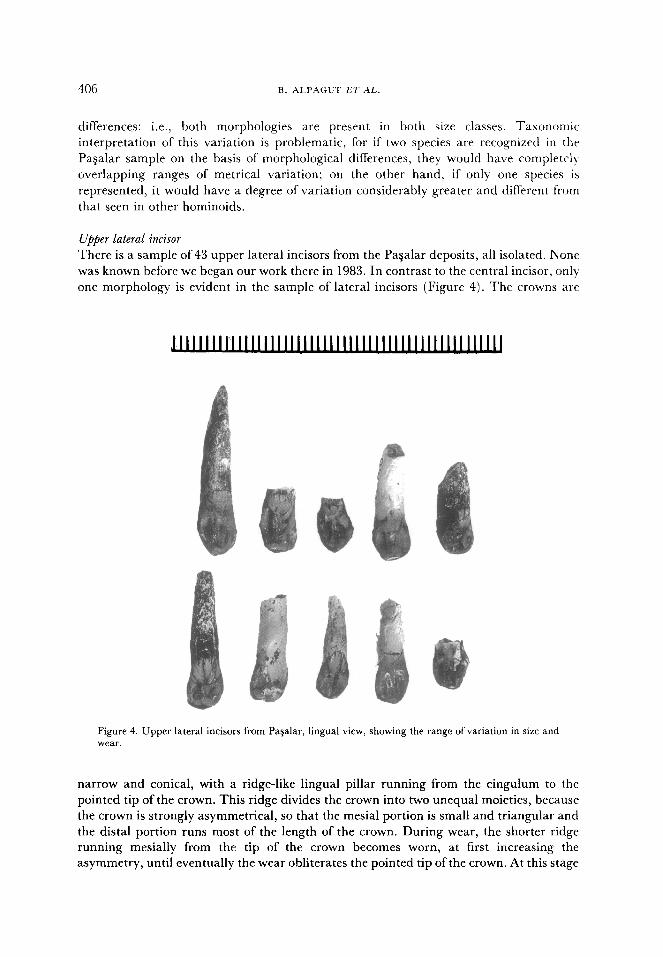
Upper lateral incisor
There is a sample of 43 upper lateral incisors from the Pagalar deposits, all isolated. None
was known before we began our work there in 1983. In contrast to the central incisor, only
one morphology is evident in the sample of lateral incisors (Figure 4). The crowns are
Figure 4. Upper lateral incisors from Pa$alar, lingual view, showing the range of variation in size and wear.
narrow and conical, with a ridge-like lingual pillar running from the cingulum to the
pointed tip of the crown. This ridge divides the crown into two unequal moieties, because
the crown is strongly asymmetrical, so that the mesial portion is small and triangular and
the distal portion runs most of the length of the crown. During wear, the shorter ridge
running mesially from the tip of the crown becomes worn, at first increasing the
asymmetry, until eventually the wear obliterates the pointed tip of the crown. At this stage

MlDDLE MIOCENE HOMINO’DS FROM PA$ALAR 405
it appears more incisiform than in the unworn state, but since all intermediate wear states
are present in the sample it is clear that there is no difference in the original morphology.
There are no lateral incisors in association with centrals. Based on average medio-distal
dimensions of I’ and I2 (Table l), the ratio of the first to the second is 1.68, or 168%.
Similar percentages are obtained when the two smallest I’ and I” are compared, giving :I
value of 182%, or when the two largest are compared, gi\-ing 183%. The largest I’
compared to the smallest I2 gives a value of 256%, while the smallest I’ compared to the
largest I’ gives 130%. These figures represent the possible range of II/I” size comparisons.
and the probable size relationship is of the order of 168%. This is a high value, comparable
with the orang-utan, which for a sample of 20 individuals has a mean value of 159% and a
range of 140-187%. It is lower than the values obtained for later Miocene species 01’ Sirlapithecus, but higher than values for Proconsul (Andrews. 1978).
It appears from the size distribution of I’ that the degree of robusticity of the crowns is
slightly greater than that of the orang-utan, but much less than that of Sizrapithecus. The size,
relationship of the Pa?alar sample to that of the orang-utan appears identical for both I’
and I’? so that it would appear from this that overall the I’ bears the same order of size
relationship to I’ in both samples. Since this relationship has been shown to be one of the
diagnostic features ofthe orang-utan dentition (Andrews & Cronin, 1982), this result ma)
be of taxonomic importance.
There is no separation of the Pagalar sample of upper lateral incisors into distinct size
classes. The range of variation is little greater than that of the living or subfossil orang-
utans (Table 2) and it is comparable with the ranges of the Lufcng and Siwalik samples.
although at these sites the sample variation is based on more than one species. Thcrr is
therefore no reason to recognize more than one species based on I” size or morpholoq.
l@er canine
There is a sample of 27 upper canines from the Payalar deposits. All are isolated and all
represent new collections made since 1983. The crown is simple in construction. It is robust
rather than bilaterally flattened, and although functionally a projecting tooth the crown of
the upper canine is relatively low: maximum crown height is 19.6 mm on the largest
specimen. The buccal side of the crown is rounded, the lingual side gently convex with a
very narrow cingulum. There may be one or two rounded ridges running down the lingual
surface, but the main feature is the mesial ridge, which forms the mesiobuccal side of the
deep mesial groove. This groove is well developed on large canines and shallow on small
canines, but the mesial ridge is always well developed (Figure 5). The groove terminatrs
cervically against the furthest mesial extent of the lingual cingulum, which also, therefore.
forms the end of the mesial groove where it is present. Distally, the crown ends in a rounded
distal ridge, which occludes against the lower third premolar. Mesial wear cuts across thf
mesial groove. but it appears never to be extreme: a large flattened wear facet is never seen
across the mesial surface of the crown, nor is dentine ever exposed. This is in marked
contrast to wear on the distal surface of the lower canine (see below) where crown wc’ar is
highly unusual.
The only morphological variation observed seems to be directly related to size. This
concerns the variation in the development of the mesial groove just described, and it is the
kind of difference that is commonly seen in males and females of the same species of living
hominoids. The size variation is very great (Table 2), with the canines from PaQalar falling
into two completely distinct size groups (Figure 5). Some of the canines from Pagalar are at
the exteme small end of the range of Siwalik fossils, but most are beyond the bottom end of
the Siwalik range, while the large canines are at the top of the Siwalik range, with several
specimens beyond that range. The values of the maximum to minimum index exceed those
for recent and sub-fossil orang-utans and those of the comparative fossil samples. The
coefficients of variation for canine length and breadth for the pooled Pagalar sample arc
24.2 and 22.9 respectively, remarkably high values compared with variation in living
primates (Kay, 1982). It seems clear from this that more than one species is indicated by
this extreme degree of metrical variation.
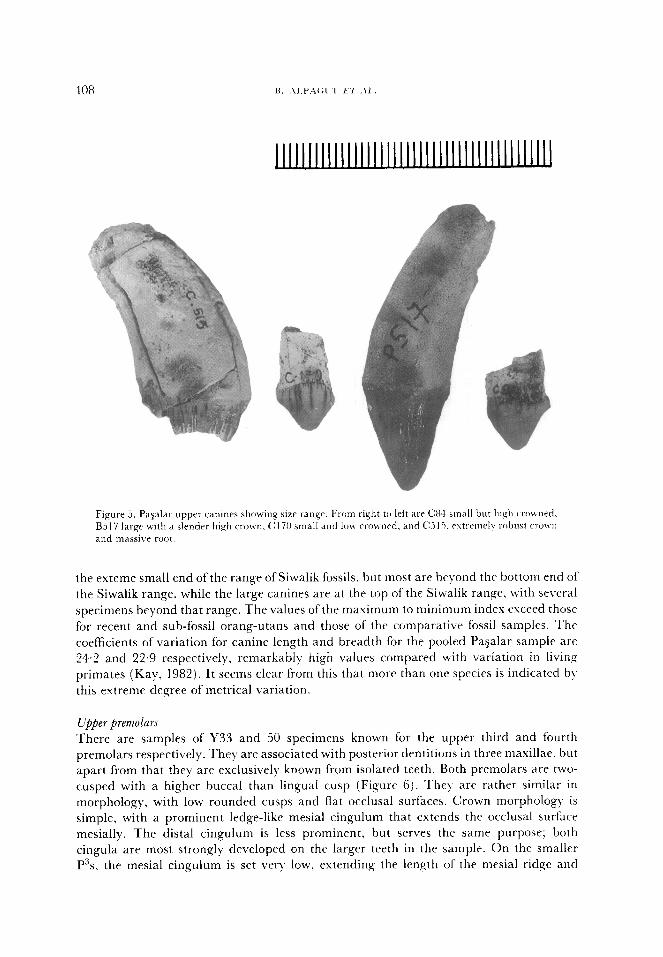
Upper premolars
There are samples of Y33 and 50 specimens known for the upper third and fourth
premolars respectively. They are associated with posterior dentitions in three maxillae, but
apart from that they are exclusively known from isolated teeth. Both premolars are two-
cusped with a higher buccal than lingual cusp (Figure 6). They are rather similar in
morphology, with low rounded cusps and flat occlusal surfaces. Crown morphology is
simple, with a prominent ledge-like mesial cingulum that extends the occlusal surface
mesially. The distal cingulum is less prominent, but serves the same purpose; both
cingula are most strongly developed on the larger teeth in the sample. On the smaller
P”s, the mesial cingulum is set very low, extending the length of the mesial ridge and
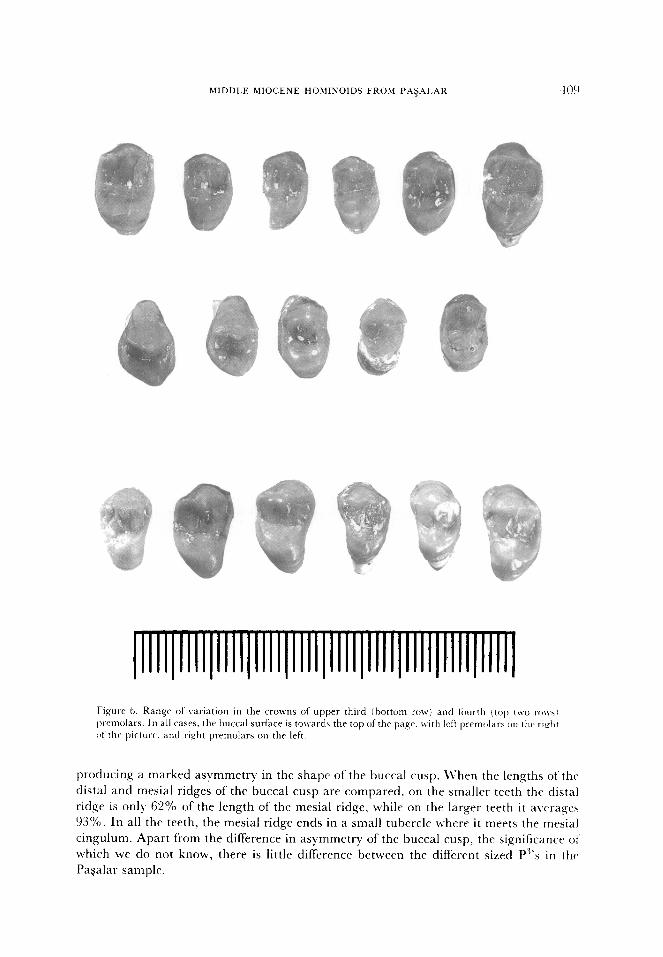
MIDDLE MIOCENE HOhlINOIDS FROM PA$AI.AR 409
II .
~lllI)Il;I~IIll)IIIl(IIIl~llll~llll~llll~llll~llll~
Fi,qurA 6. Range of variation in the CTOWIIS of upper third j tmttom row) and tiiur-th (top t\vo ro~\~ Ixelnolars. In a11 cases. the huccal surface is tolvards the top of the pa,qe. with left prcmtkws on thv ri&t
01 tht‘ picture. and right premolars on thr left.
producing a marked asymmetry in the shape of the buccal cusp. LVhen the lengths of the
distal and mesial ridges of the buccal cusp are compared. on the smaller teeth the distal
ridge is only 62% of the length of the mesial ridge, while on the larger teeth it a\rrrages
93%. In all the teeth, the mesial ridge ends in a small tubercle where it meets the mesial
cingulum. Apart from the difference in asymmetry of the buccal cusp, the significance 01‘
which we do not know, there is little difference between the different sized P”‘s in the
Pagalar sample.

The main differences bctwcen P” and P’ are in the slightly grcatcr cspansic~n (,I’ IIN.
buccal cusp in P”, expansion both in height and in mesiodistal length relative to the lingual
cusp, and the complete lack of P-’ cingulum (Figure 6). There is usually no ridge
development on the occlusal surface of P’, and the buccal and lingual cusps are nearI!-
equal in size. In size, the P3 is larger than P’ when the masimum dimensions are counted.
because of the buccal expansion of P”, but when medial lengths are compared the two terth
are much the same size (Table 1). There is a marked division into large and small size
classes for both premolars, with little overlap between them. This contrasts with thr ranges
of variation seen in living orang-utans, and with the variation in the sample offbssil terth
from the Siwaliks where at least two species arc supposed to be present. This could either
be attributed to sexual dimorphism of an unusual kind in the PaSalar sample, or it could
indicate the presence of more than one species.
The total range of variation of both upper premolars is very similar to that of the entire
Siwalik sample. In both cases the PaSalar premolars are slightly larger overall than those
from the Siwaliks, but the ranges are almost identical. The ranges of variation are also
similar to those for the orang-utan, although the teeth in the living ape are considerabl)
bigger than those in both sets of fossils. Comparing the maximum/minimum dimensions
(Table 2), the indices for lengths and breadths for the PaTalar sample are greater than
those for modern orang-utans, comparable with the ranges for the pooled Siwalik sample.
but contained within the range of variation for sub-fossil orang-utans and the Lufeng
sample.
Upper molars
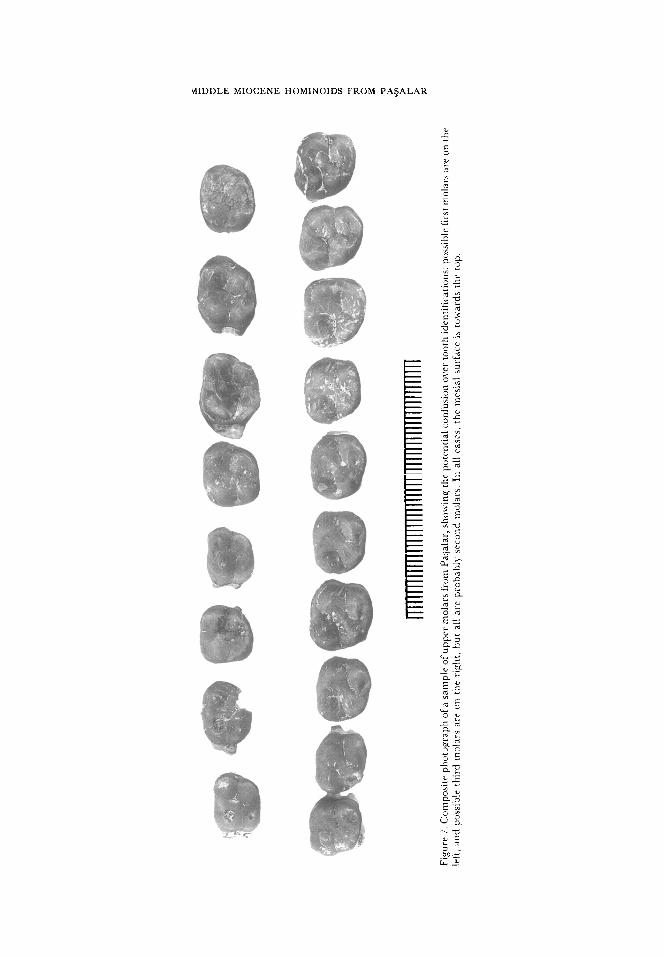
The upper molars in the Pagalar sample will be described together to avoid unnecessar)
repetition of description. Samples include associated molars in the three maxillae D149,
D922 and G432, and an associated tooth row of Ml to M”, C239 (Figure 3). Available
sample sizes are:
M’ - 37 specimens
M’- 78 specimens
M3 -6 1 specimens
The molars are similar to each other in crown morphology and shape, and it has
sometimes been difficult to distinguish between them (Figure 7). Positive identifications
have been possible for the first molars in the associated dentitions. Positive identifications
of the third molar have also been possible on the basis of extreme distal abbreviation of the
crown and absence of distal contact facets on heavily worn teeth. Beyond this,
identifications have been made on the basis of minor morphological features, and we
recognize that there are probably still some misidentifications, particularly between M’
and M*. Double mesial contact facets are present on some lower first molars, where they
contacted first dp+ and then Pq, but we found no evidence for double facets on upper
molars, presumably as there is less difference in distal crown height between dpt and P”
than is the case for dp4 and P+ We feel, therefore, that we have included some M’s in the
M* sample, and we are undertaking further more detailed analyses of micromorphology to
remedy this problem.
The molars from Pagalar, both upper and lower, have been shown to have thick enamel
comparable with that seen in Siwalik Siuapithecus and in modern humans (Martin, 1983,
1985). This enamel is dominated by pattern 3 prism packing and cross-striation intervals
MIDDLE MIOCENE HOMINOIDS FROM PA$ALAR
-
0

are in the range of 5-7 pm. Hunter-Schreyer bands cxtcnd from clo.sc- to the, c~~amc*l-
dentine junction through most of the enamel thickness (Martin, 198:i I. This condition has
been interpreted as the shared derived condition fbr the great apt and human cl&t* alltl
indicates that the Payalar hominoids are more closely related to extant memht~rs ot’ that
clade than are, for example, D;yvopithecu.r, Proconsul, .-?fropithecus and Hdobute.,. The Pasalar
hominoids have molar teeth in which the enamel-dentinejunction shows little topographic
relief (Martin, 1983). This condition is found only in orang-utans and Siwalik ,S~UZ~~&YUY
among other hominoids, and Martin (1983) suggested that it represents ;I dcri~t~tl
character state by which to define a Pongol.S’izlapitheczIs clade.
The crowns of the upper molars are ofsimple construction, with poorly developed ridgea.
infrequent accessory cusps, and variably developed cingula. Nearly half the teeth ha\-? a
slight development ofthe buccal cingulum in the interval between paracone and metaconc.
A smaller number of teeth also have traces of a lingual cingulum. The four main cusps are
developed on all molars, including the third, and the only consistently developed ridge is
the crista obliqua. which is low and rounded. This lack ofdefinition is likely to be caused b\
the thick enamel on all the molars. On M’ the four cusps are approximately equal in size,
although the hypocone may be slightly offset lingually and distally, and the hypocone ma)
be slightly smaller than the other cusps. As a result, the M’ is usually square in outline. On
M’, the hypocone is usually more reduced, and the metacone may also be reduced, so that
the crown narrows distally and may appear slightly elongated. The M” has even grratcr
reduction of the distal cusps. although it is never carried so far as to lose the cusps
altogether. In all these features, there is no consistent difference in morpholog)-, either
related to size or independent of size, that might indicate the presence at Pagalar of more
than one species of hominoid.
The three molars do not vary greatly in size. The second molar is the largest, M’ and M”
are approximately equal in size. In the early Miocene genus Proconsul, by contrast, the M”
is always small relative to M2, and there is greater size disparity between M’ and M’. It is
of interest that the size distribution of the molar sample from PaSalar overlaps only part of
that of the Siwalik sample, in the case of M’ the bottom half of the Siwalik range, and for
M’ the bottom 85%. In other words, the molars from the PaSalar sample are slight])
smaller than those from the Siwaliks, but in comparison the premolars range the other way,
overlapping the top of the Siwalik range and extending beyond it. This indicates a
difference in premolar/molar proportions not readily apparent from the indices, nameI)-
that on average, the premolars in the Pagalar sample are larger relative to the molars than
is the case in the Siwalik sample. The total estent of the Pagalar range, however. is similar
to that of the Siwalik range for the molars, as it is for the premolars.
The size distributions for the M’ dimensions show a division into two distinct size classes
that is not present in the other two molars. It would certainly be unusual for any one
species sample to have a bimodal distribution for the length/breadth dimensions of the first
molar, and so some weight must be attached to this distribution. The total range of
variation for all of the upper molars in the Pagalar sample exceeds the variability of modern
orang-utans, is less than or equal to the Lufeng and Siwalik samples, and is contained
within the variation of sub-fossil orang-utans (Table 2). The lower level of variation in M’
compared to M2 and M3 (Table 2) could be taken to indicate that the first molar range for
the Pagalar sample is artificially truncated and thus be an indication of a bias in this part of
the sample, perhaps due to misidentification of some of the isolated teeth as discussed
above. This bias could be the cause of the bimodalitv.
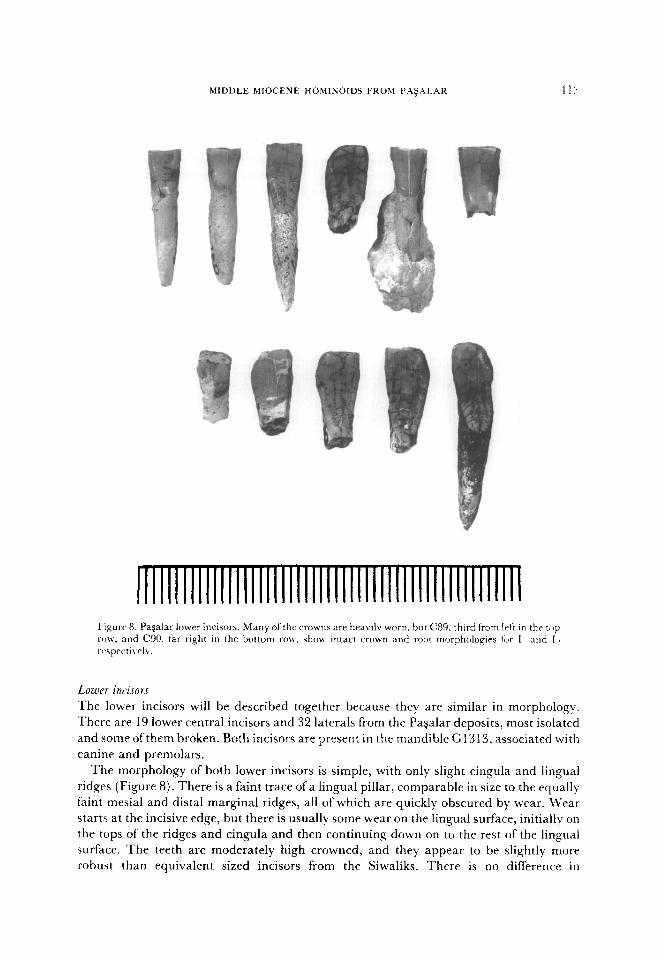
MIDDLE MIOCENE HOMINOIDS FROM PA+41..4R 41:
Lowrr incisors
The lower incisors will be described together because they are similar in morphology.
There are 19 lower central incisors and 32 laterals from the Pagalar deposits, most isolated
and some of them broken. Both incisors are present in the mandible G13 13, associated with
canine and premolars.
The morphology of both lower incisors is simple, with only slight cingula and lingual
ridges (Figure 8). There is a faint trace of a lingual pillar, comparable in size to the equally
faint mesial and distal marginal ridges, all of which are quickly obscured by wear. Wear
starts at the incisive edge, but there is usually some wear on the lingual surface, initially on
the tops of the ridges and cingula and then continuing down on to the rest of the lingual
surface. The teeth are moderately high crowned, and they appear to be slightly more
robust than equivalent sized incisors from the Siwaliks. There is no difference in

414 B. .ALPAt:t '1' E7 .-\L.
morphology apparent within the Pagalar sample, and certainly no differences between sizt classes. The main variable is the greater asymmetry of the lateral incisors, the distal
surfaces ofwhich are rounded so that the incisive edge is offset mesially. The central incisor is almost perfectly symmetrical. This is the usual condition for hominoid lower incisors, but the difference between central and laterals is perhaps less than is usual, or at any rate it is
more than usually difficult to distinguish between them, especially when there is some wear.
Figure 9. Lower canines from Pagalar, showing range of variation of a small sample of crowns.
The central incisor appears small relative to the Siwalik sample but this may be
sampling error because of the small size of the Siwalik sample. The largest central incisors from Pagalar are equivalent in size to the smallest incisors from the Siwaliks, with only slight overlap. The same is true of the mesiodistal dimension of the lateral incisors, but because of breakage of the bases of their crowns, the buccolingual dimensions of the Siwalik specimens cannot be estimated. Both incisors are considerably smaller than those from the orang-utan samples, and their size relationships to the orang-utan samples are similar. In fact, when all four incisors are compared with those of the orang-utan, the sample ranges of the Pagalar samples have a similar size relationship with the orang-man sample.
There is no separation of the Pasalar lower incisors into distinguishable size classes. The range of variation in the lateral incisors is greater than that seen in modern and sub-fossil orang-utans, and in the Lufeng and Siwalik samples (Table 2). This metrical variation is taken to indicate the presence of more than one species at Pagalar.

MIDDLE MIOCENE HOMINOIDS FROM PA?ALAR 415
Lower canine
There is a sample of 36 lower canines known from the Pagalar deposits. All are isolated,
except for the canine in the mandible G 13 13, and the lower canine D 149, which was found associated with the maxilla of the same number in circumstances that suggest they might
have come from the same individual. The original description of the Pagalar hominoids included five lower canines, all of similar large size (Andrews & Tobien, 1977) and in fact
still among the largest specimens of the present expanded sample. The crowns are rounded on their buccal surface and nearly flat lingually (Figure 9). The
lingual surface is bounded by a long and prominent mesial ridge, sometimes so prominent as to impart a superficial resemblance to upper canines because of the groove-like
depression immediately behind the ridge. In general, the larger the overall crown size, the larger the mesial ridge and the deeper the post-ridge depression. The ridge meets the
lingual cingulum at its base, and at this point the cingulum is always well developed. As it passes distally, however, it diminishes in size and may be notched by wear, but it rarely
completely disappears before it reaches the rounded and indistinct distal ridge, where it forms a distinct distobuccal heel. The cingulum forms the lower limit of the flattened
lingual face.
Although the buccal face of the crown is gently rounded overall, the distal ridge has a concavity on its buccal side so that the distal part of the buccal face can be strikingly concave. This forms the only distinguishing mark on the buccal surface and increases the prominence of the ridge.
Where preserved, the root of the lower canine is massive. It is twice the height of the crown and has greater length and breadth dimensions as well. It is similar to the root
proportions of the sample of “Ouranopithecus” macedoniensis, although the molars of that
species are much larger than those from PaSalar (Martin & Andrews, 1984). There is
therefore a disproportion here between the two samples. The morphology of the distal surface of the crown of the lower canine results in a highly
characteristic form of wear. All stages of wear are present in the Paaalar sample, and the description that follows is a generalized account drawing on the different stages
represented by these specimens. To begin with, wear starts as a flattened facet
distobuccally at the tip of the crown where it meets the mesial edge of the upper canine. This begins to extend downward towards the base of the crown. Shortly afterwards, a compound wear facet develops independently towards the base of the crown, one part on the distal ridge and the one on the buccal surface on the other side of the disto-buccal
concavity just described. As wear progresses, these two facets coalesce around the edges, leaving an island of enamel between them in the buccal concavity, and eventually they join up with the growing wear facet at the tip of the canine. Finally, further wear removes even
the island of enamel left between the two lower facets so that a single large distal wear facet remains over the whole of the distobuccal face of the tooth. Even then, however, evidence of the origin of the facet remains in the tongue-like projection of enamel from the base of the
crown up into the wear facet. A similar pattern of wear has been observed on the orang- utan lower canine, with both the final wear facet and the intemediate stages being similar, but it is not known if this has any taxonomic significance. Both this morphology and pattern of wear are characteristic of all the Pagalar specimens of lower canine. There is no observed morphological variation that would lead to their separation into two samples or species. In size, however, there is considerable variation. There are two very distinct size classes, rather far removed from each other. The range of variation of the larger size group
approximates that of the Siwalik sample, ancl it also coincides with the range of temale
recent and sub-fossil orang-utans. The smaller size class is beyond the range of both the
orang-utan and Siwalik samples. The degree ofvariability in canine length of the combined
sample of small and large size classes from Pagalar is much greater than that of living
orang-utans and all other comparative samples (Table 2). Ganinc breadth variation is
similar to orang-utan variation but esceeds that of the two comparative fossil samples.
particularly that of the combined Siwalik sample. This is also seen in the high coefficients of
variation (Table l), which are 18.3 for length and 23.0 for breadth, and as for the uppet
canines and lower incisors these figures suggest the presence at PaSalar of more than one
species.
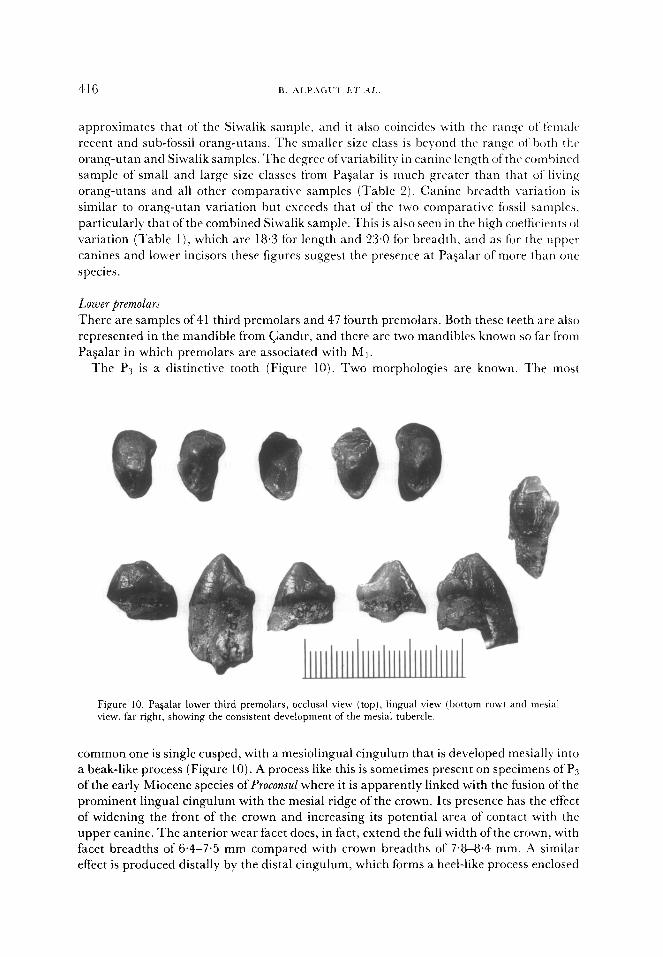
Lower premolar5
There are samples of 41 third premolars and 47 fourth premolars. Both these teeth are also
represented in the mandible from Gander, and there are two mandibles known so far from
Pagalar in which premolars are associated with Mt.
The P3 is a distinctive tooth (Figure 10). Two morphologies are known. The most
Figure 10. Pa&u lower third premolars, occlusal view (top), lingual view (bottom row) and mesial view. far right, showing the consistent development of the mesial tubercle.
common one is single cusped, with a mesiolingual cingulum that is developed mesially into
a beak-like process (Figure 10). A process like this is sometimes present on specimens of PLs
of the early Miocene species of Proconsul where it is apparently linked with the fusion of the
prominent lingual cingulum with the mesial ridge of the crown. Its presence has the effect
of widening the front of the crown and increasing its potential area of contact with the
upper canine. The anterior wear facet does, in fact, extend the full width of the crown, with
facet breadths of 6.4-7.5 mm compared with crown breadths of 7.8-8.4 mm. A similar
effect is produced distally by the distal cingulum, which forms a heel-like process enclosed

MIDDLE MIOCENE HOMINOIDS FROM PAQALAR 417
by the distolingual and distal ridges. The distal cingulum actually crosses the distal ridge
to form a transient buccal cingulum, but this does not extend far on to the buccal surface of
the crown. The distal cingulum does not cross the distolingual ridge, so that at the mid-
point of the lingual surface of the crown, no cingulum is present. but the crown is expanded
lingually at this point by a marked convexity of the lingual surface. This convexity,
together with the mesial and distal processes, combine to increase the surface area of the
crown, and this may be seen as a development from the more parallel sided, sectorial P3’s
that characterize the ancestral hominoid morphotype (Delson & Andrews, 1975: Andrrws,
1978). In addition to this common morphology, a small number of teeth show a difI&ent
suite of characters: the lingual cingulum is extremely prominent, the mesial tuber& is
poorly developed or absent, the crown is relatively broad, and there may be incipient
development of a lingual cusp on the lingual ridge/cingulum. The specimens on which this
morphology occurs (e.g., (31141) are all at the top end of the range of trariation of the
sample.
Although not as developed as the P3 morphology seen in the living great apes and
humans, many of these characters are the same as in the living species and may be seen as
advances towards the shared great ape and human morphotype. This morphological shift
is not apparent from the breadth/length indices of P.3. which for the Pasalar samplta are
little different from the Proconsul range and far short of the broad. low crowned Pi’s of the
living great apes. This apparent anomaly only serves to illustrate the inadequacies of
simple length-breadth measurements.
In terms of overall size, the PaSalar Ps sample falls into two distinct size classes that are
himodally distributed when numbers of specimens are divided into size classes. The t\vo
size distributions correspond closely to the two common species currently recognized from
the Siwaliks, with the overall range very close to that of the Siwalik sample. but therta is
nothing approaching the size of the recently named Sir~a$thecus pawada (Kelley, 1988).
Overall variation is less than that ofthe modern and sub-fossil orang-utans and the Siwalik
sample but is similar to that of the Lufeng sample (Table 2).
1lllllllnTmllllllllllllllllllllTlllllllllllmll Figure 1 I. Occlusal views of the fourth lower premolar, mesial side towards the top. from the left sidr in the top row, right side in the bottom row.

The P+ is a variable tooth, as is commonly the case in hominoids. Part of this \,ariabilit!
is due to difficulties in orienting the specimen, and this may be true even for spccimcan still
in place in mandibles. The trigonid basin is transversely restricted, and the talonid baain is
deeply excavated and also relatively narrow (Figure 11). As a result, the occlusal surface of‘
the crown appears relatively narrow, but this is offset by the great buccal Rare of the cro\vn,
which exaggerates the buccolingual dimension. Some specimens have traces of buccal
cingulum, and these have reduced buccal flare and also tend to have lower brradthilcngth
ratios.
The size distribution of the Pagalar sample of P4 is similar to that ofP,s, although the two
size classes are not quite as distinct. The degree of variation is comparable to the orang-
utan samples and to the two fossil samples.
Lower molars
There are samples of 56 first lower molars, 62 second molars and 60 third molars. These
teeth are also present on both sides of the Gander mandible, and there is a first molar in onr
of the mandibles found at Pagalar (E697).
The lower molars are among the best represented teeth in the PaSalar collection, but
there are sometimes problems in distinguishing first from second molars. Obviously it
helps to have associated material, where there is no ambiguity, but because of the close
similarity between Ml and M2 there will always be a problem with isolated teeth.
However, many of the Ml’s retain signs of a double mesial contact facet where the)
connected first with the deciduous premolar and then with P+, but in the absence of this it
becomes more difficult to distinguish a large M1 from a small M?. The closer balance of
specimen numbers of Ml/My/M3 than was the case for the upper molars (Table 1)
indicates that our attributions have been generally correct.
The lower molars increase in size from M1 to M3, mainly as a result of length increase.
They also increase in breadth variability (Figure 12). a phenomenon they share with the
Siwalik Sivapithecus and subfossil orang-utans, but not with recent orang-utans or with the
Lufeng hominoids. In other words, the breadth dimensions of the Pagalar sample are more
variable in M3 than in M,. A small proportion of the crowns of the first and second molars
are characterized by their extreme breadth in some cases with breadth actually exceeding
length (e.g., specimen G1303). In the M:+ most ofthe specimens show some degree ofdistal
taper of the crown, although it varies widely (Figure 12). but a small proportion show no
distal narrowing, and may even get wider distally, a very unusual characteristic of third
molars. The extreme expression of this broadening in all the molars, but especially on M:j,
may have taxonomic significance.
The lower molars have five cusps and two roots. in the usual hominoid molar pattern
(Figure 12). They have a mesiodistally compressed trigonid basin, which on aomr
specimens is almost non-existent. The metaconid is set transversely to the protoconid, and
they are connected by a low rounded ridge that delimits the trigonid basin (mesial fovea of
some authors). The talonid basin is large, and there is usually a small but distinct distal
fovea between the entoconid and the hypoconulid. There is a buccal cingulum present on
rather more than half of the specimens from Pagalar (Figure 12). There is no apparent
difference between size categories in cingulum development, which varies uidel) Lvithout
any apparent taxonomic significance. It forms a wide and distinct ledge on a small number
of specimens, a discontinuous shelf on rather more, and on more still it is a discontinuous
line, often hardly visible. On about one third of specimens there is no tract of cingulum at
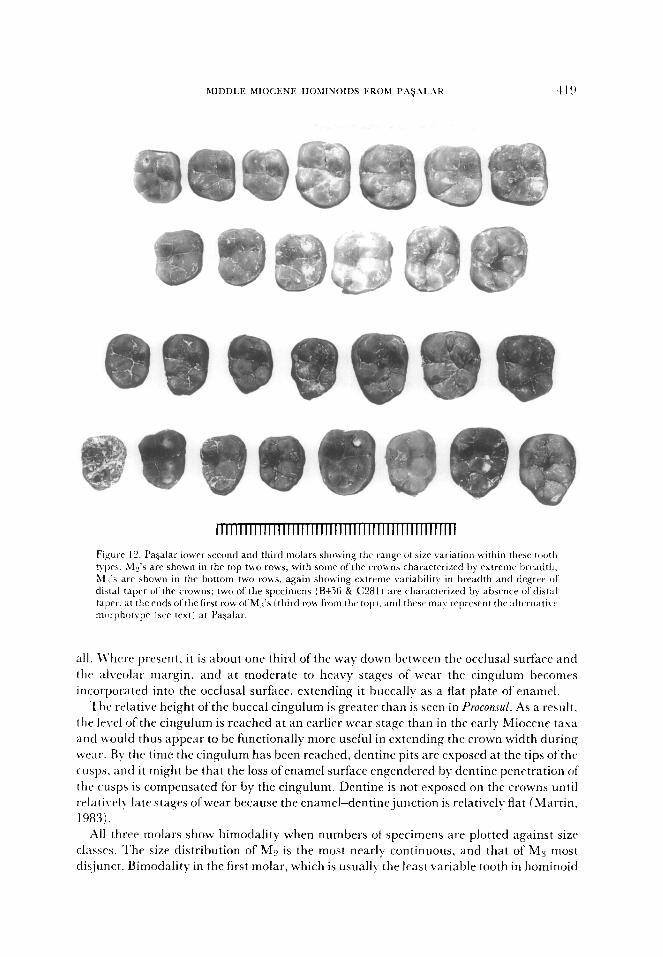
MIDDLE MIOCENE HOhlINOIDS FROhl PAFjALAK 4, I Cl
@QwBl,Q@
IIIIIIIIIlIIlllllllm
Figurr 12. Pagalar lower second and third molars showing thr rangr of size variation within these tooth typr>. My’s are shown in the top two rows, with somr of the crou~~s characterized by extreme breadth; &I 3.5 arr shown in the bottom two rows. again showing extreme variability in breadth and degree ol distal taper of the crowns; two of the specimens (B4.56 & C28 I i are characterized by absence of distal taper. at the ends ofthe first row of Mi’s (third row from the top I. and these may repwsent thr altunatiw nxirphotvpe lsee text) at Pagalar.
all. U’here present, it is about one third of the way down between the occlusal surface and
the al\reolar margin, and at moderate to heavy stages of wear the cingulum becomes
incorporated into the occlusal surface, extending it buccally as a flat plate of enamel.
The relative height of the buccal cingulum is greater than is seen in Proconsul. As a result,
the level of the cingulum is reached at an earl& wear stage than in the early Miocene tasa
and would thus appear to be functionally more useful in extending the crown width during
wear. By the time the cingulum has been reached, dentine pits are exposed at the tips ot‘thtk
cusps. and it might be that the loss ofenamel surface engendered by dentine penetration ot
the cusps is compensated for by the cingulum. Dentine is not exposed on the crowns until
relati\.ply late stages ofwear because the enamel-dentinejunction is relatively flat (Martin.
1983).
All three molars show bimodality when numbers of specimens are plotted against size
classes. The size distribution of ML’ is the most nearly continuous, and that of M3 most
dis,junct. Bimodality in the first molar, which is usually the least variable tooth in hominoid
dentitions, is unusual for a single species. although the greater dcgrec of bimoclality in ;I1 ,, could be more easily explained as it is usually more variable. This pattern of variation
would tend to indicate the presence of two species at Pagalar. Degrees ofvariation for all tht,
molars exceed those for modern orang-utans but are similar to ranges for sub-fossil orang-
utans and the comparative fossil samples (Table 2).
Deciduous teeth
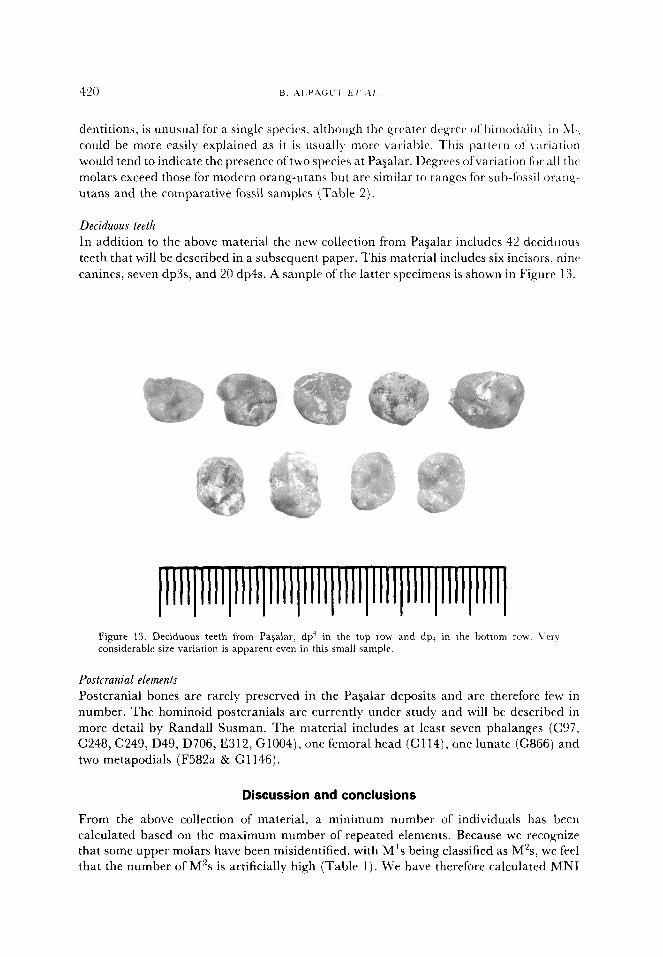
In addition to the above material the new collection from Paaalar includes 42 deciduous
teeth that will be described in a subsequent paper. This material includes six incisors, ninr
canines, seven dp3s, and 20 dp4s. A sample of the latter specimens is shown in Figure 13.
Figure 13. Deciduous teeth from Pagalar. dp ’ in the top row and dp, in the bottom row. Very considerable size variation is apparent even in this small sample.
Postcranial elements
Postcranial bones are rarely preserved in the Pagalar deposits and are therefore few in
number. The hominoid postcranials are currently under study and will be described in
more detail by Randall Susman. The material includes at least seven phalanges (C97,
C248, C249, D49, D706, E312, G1004), one femoral head (Cl 14), one lunate (G866) and
two metapodials (F582a & G1146).
Discussion and conclusions
From the above collection of material, a minimum number of individuals has been
calculated based on the maximum number of repeated elements. Because we recognize
that some upper molars have been misidentified, with M’s being classified as M’s, we feel
that the number of M’s is artificially high (Table 1). We have therefore calculated MN1

MIDDLE MIOCENE HOMINOIDS FROM PASAL.AR 421
basrd on LMT (35) and LM3 (32) as more realistic estimates compared with MN1 based
on LM’ (42). There are also seven juveniles based on right or left (7 each) dp’. \Ve can
conclude reliably that 35 adults and 7 juveniles are present in the Pagalar assemhla,qe.
These figures are significant in light of the evidence from depositional and taphonomic
studies (Andrews & Alpagut, 1990; Andrews & Ersoy, 1990) that indicates that the fauna
might closely approximate a life assemblage as it accumulated during a very short period ot‘
time and was sampled from a restricted geographical area. More detailed calculations of‘
life stages that take account of dental wear stages are in preparation and from thesr it i\
hoped to build up a more accurate representation of the life history stages of the PaSalar
hominoid assemblage.
It seems clear from the discussion of the degree of variation in the hominoid sample that
any attempt to impose a single species interpretation on the Pagalar hominoids ignores
significant morphological as well as metric variation. Since the metric variation extends to
the deciduous dentition a strong case can be made for division of the sample into two
species solely on metric grounds. More detailed morphological and metrical analyses arc
required to provide definitive answers to questions concerning the pattern and nature of‘
sexual dimorphism of the Pagalar hominoids. but present levels ofanalysis indicate that at
leasr two species are represented. One of the two species, and hy far the most abundant in
terms of specimen numbers (approximately 90% of the available sample) is recogniztld
here as being conspecific with the holot)-pe of [email protected] a/pani (Tekkaya, 1974) as
previously recognized by Andrews & Tekka)-a (1976) and Andrews & Tobien (19771, but
this hypodigm is now viewed as being distinct from that of Keqapithecus africanus (SW
Martin. 1983). This species is synonymized here with .Simpithecus darwini (Abel. 1902;
Andrews & Tobien, 1977), and it is extremely sexually dimorphic. The second species is
presently unnamed. It is a large species, although tvith extensivtl overlap in size range: it
differs demonstrably in the morphology of- the upper central incisor, the lower third
premolar and the lower third molar: and it probably also differs in the morphology ol‘
several other teeth as well. It seems probable that these two taxa arc sister species but it is
by no means clear to which genus they should be attributrd. On present evidenct= the!
could be assigned provisionally either to Keylapithecus or to Sirwpithecus, but more detailed
analysis or more complete specimens are required to resolve this issue. Should it bc
determined that the Paaalar hominoids do not belong to either genus, the genus
Griphopithecus (Abel, 1902) is available, but we hesitate to resurrect this genus because the
type specimen is an incomplete isolated molar and it is not even certain ifit is permanrnt or
deciduous.
M’hatever the nomenclatorial outcome, there is preliminary evidence for the association
of the Pa$alar hominoids with the orang-utanlSizla@ithecus clade. This is based on incisor
proportions, the morphology of P.1, and the topography of the enamel-dentine junction. In
all of these features, the Pagalar hominoids share the derived character state of the
PongolSirlapithecus clade (Andrews & Cronin, 1982; Martin. 1983).
Acknowledgements
\Ve are grateful to the many people who have helped in the field work at Pagalar.
particularly Ayhan Ersoy, Mikael Fortelius and Songiil Alpaslan, and to Libby Andrews
and Wendy Martin who have contributed so greatly to the collection of the primate

422 B. ALPAGUT ET AL.
specimens by organizing the screening operation at the site. We thank Eric Delson, Terry
Harrison and Jeff Schwartz and anonymous reviewers for helpful comments on the
manuscript. The Pasalar excavation was carried out on behalf of the T.C. Ministry of Culture and Tourism, Directorate of Antiquities, and the University of Ankara, Faculty of
Language, History and Geography, Department of Paleoanthropology. The excavation was funded by the Directorate of Antiquities and the participation of non-Turkish personnel was made possible through the generous support of the L. S. B. Leakey
Foundation with support for specific projects from the Boise Fund, Oxford University.
References
Abel, 0. (1902). Zwei neue Menscenaffen aus den Leithakalkbildungen des Wiener Beckens. S. Ber. Akad. JViss. Wien, math.-nat. Kl. 111, 1171-1207.
Andrews, P. (1971). Ramapithecus zuickeri mandible from Fort Ternan, Kenya. Nature 231, 192-194. Andrews, P. (1978). A Revision of the Miocene Hominoidea of East Africa. Bull. Br. Mu. nut. Hist. (Geol.) 30,
85-224. Andrews, P. & Alpagut, B. (1990). Description of the fossiliferous units at PaSalar, Turkey. J. hum. Evol. 19,
343-36 1. Andrews, P. & Cronin, J. (1982). The relationships of.Sivapithecus and Ramapithecus and the evolution of the orang-
utan. Nature 297, 541-546. Andrews, P. & Ersoy, A. (1990). Taphonomy ofthe Miocene bone accumulations at Pasalar, Turkey.J. hum. Evol.
19, 379-396. Andrews, P. & Martin, L. (1987). The phyletic position of the Ad Dabityah hominoid. Bull. Br. Mu. nat. Hist.
(Geol.) 41, 383-393. Andrews, P. & Tekkaya, I. (1976). Ramapitheclcr from Kenya and Turkey. In (P. V. Tobias & Y. Coppens, Eds)
Les Plus Antiens Hominides, Colloque VI, IX Congr. Union Internat. Sci. Prehist. Protohist, pp. 7-25, Nice. Andrews, P. & Tobien, H. (1977). New Miocene locality in Turkey with evidence on the origin of&zmapi&-us and
Sivapithecus. Nature 268, 699-701. Bestland, E. (1990). Sedimentology and paleopedology of Miocene alluvial deposits at PaSalar hominoid site,
western Turkey. J. hum. Evol. 19, 363-378. Delson, E. & Andrews, P. (1975). Evolution and interrelationships ofthe catarrhine primates. In (W. P. Luckett
& F. S. Szalay, Eds) Phylogeny of the Primates, an Interdisciplinary Approach, pp. 405446. New York: Plenum. Greenfield, L. (1979). On the adaptive pattern of Ramapithecus. Am. J. Phys. Anthrop. 50, 527-548. Hooijer, D. (1948). The prehistoric teeth of man and orang-utan from Central Sumatra with notes on the fossil
orang-utan from southern China. 2001. Med., Leiden, 29, 175-301. Kay, R. F. (1982). Sivapithecus simonri, a new species of Miocene hominoid with comments on the phylogenetic
status of the Ramapithecinae. Inst. J. primat. 3, 113-173. Kelley, J. (1988). A new large species of Sivapithecus from the Siwaliks of Pakistan. J. hum. Evol. 17, 305-324. Kelley, J. & Pilbeam, D. (1986). The dryopithecines: taxonomy, comparative anatomy and phylogeny of
Miocene large hominoids. In (D. R. Swindler &J. Erwin, Eds) Comparative Primate Biology, vol. 1, pp. 361--111. New York: Alan R. Liss.
Martin, L. (1983). The Relatiomhips of Later Miocene Hominoidea. Ph.D. thesis, University of London. Martin, L. & Pilbeam, D. (1985). Significance of enamel thickness in hominoid evolution. Nature 314,260-263. Martin, L. & Andrews, P. (1984). The phyletic position of Graecopithecusfreybergi Koenigswald. Cour. Forsch. Inst.
Seckenberg 69, 25-40. Sickenberg, 0. (1975). Die Gliederung des hoheren Jungtertiars und Altquartiars in der Tiirkei nach Vertebraten
und ihre Bedeutung ftir die internationale Neogen-Stratigraphie. Geol. Jb. B15, l-167. Tekkaya, I. (1974). A new species of anthropoid (Primates, Mammalia) from Anatolia. Bull. Miner. Res. Expl.
Imt., Ankara 83, 148-165. Wu Rukang & Oxnard, C. (1983). Ramapithecines from China: eqidence from tooth dimensions. Nature 306,
258-260.
Related Documents











