10.1128/JB.182.3.812-817.2000. 2000, 182(3):812. DOI: J. Bacteriol. Wen-Tyng Li, John W. Shriver and John N. Reeve Mesophilic and Hyperthermophilic Archaea Thermostability between Histones from Mutational Analysis of Differences in http://jb.asm.org/content/182/3/812 Updated information and services can be found at: These include: REFERENCES http://jb.asm.org/content/182/3/812#ref-list-1 This article cites 15 articles, 3 of which can be accessed free at: CONTENT ALERTS more» articles cite this article), Receive: RSS Feeds, eTOCs, free email alerts (when new http://journals.asm.org/site/misc/reprints.xhtml Information about commercial reprint orders: http://journals.asm.org/site/subscriptions/ To subscribe to to another ASM Journal go to: on February 8, 2014 by guest http://jb.asm.org/ Downloaded from on February 8, 2014 by guest http://jb.asm.org/ Downloaded from

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

10.1128/JB.182.3.812-817.2000.
2000, 182(3):812. DOI:J. Bacteriol. Wen-Tyng Li, John W. Shriver and John N. Reeve Mesophilic and Hyperthermophilic ArchaeaThermostability between Histones from Mutational Analysis of Differences in
http://jb.asm.org/content/182/3/812Updated information and services can be found at:
These include:
REFERENCEShttp://jb.asm.org/content/182/3/812#ref-list-1This article cites 15 articles, 3 of which can be accessed free at:
CONTENT ALERTS more»articles cite this article),
Receive: RSS Feeds, eTOCs, free email alerts (when new
http://journals.asm.org/site/misc/reprints.xhtmlInformation about commercial reprint orders: http://journals.asm.org/site/subscriptions/To subscribe to to another ASM Journal go to:
on February 8, 2014 by guest
http://jb.asm.org/
Dow
nloaded from
on February 8, 2014 by guest
http://jb.asm.org/
Dow
nloaded from

JOURNAL OF BACTERIOLOGY,0021-9193/00/$04.0010
Feb. 2000, p. 812–817 Vol. 182, No. 3
Copyright © 2000, American Society for Microbiology. All Rights Reserved.
Mutational Analysis of Differences in Thermostability betweenHistones from Mesophilic and Hyperthermophilic Archaea
WEN-TYNG LI,1 JOHN W. SHRIVER,2 AND JOHN N. REEVE1*
Department of Microbiology, The Ohio State University, Columbus, Ohio 43210,1 and Department of Biochemistry andMolecular Biology, School of Medicine, Southern Illinois University, Carbondale, Illinois 629012
Received 7 September 1999/Accepted 26 October 1999
Amino acid residues responsible for the large difference in thermostability between HMfB and HFoB,archaeal histones from the hyperthermophile Methanothermus fervidus and the mesophile Methanobacteriumformicicum, respectively, have been identified by site-specific mutagenesis. The thermal denaturation of ;70archaeal histone variants has been monitored by circular dichroism, and the data generated were fit to atwo-state unfolding model (dimer3two random coil monomers) to obtain a standard-state (1M) meltingtemperature for each variant dimer. The results of single-, double-, and triple-residue substitutions reveal thatthe much higher stability of rHMfB dimers, relative to rHFoB dimers, is conferred predominantly by improvedintermolecular hydrophobic interactions near the center of the histone dimer core and by additional favorableion pairs on the dimer surface.
Histones from mesophilic, thermophilic, and hyperthermo-philic Archaea have similar sequences and folds but very dif-ferent thermodynamic stabilities (5, 8, 9, 14, 18, 20). Thesesmall proteins (66 to 69 amino acid residues) exhibit fullyreversible temperature-, salt-, and pH-dependent unfoldingand refolding and therefore provide an experimentally tracta-ble system to relate primary sequences, and three-dimensionalstructures, to inherent protein stability. For example, HMfBand HFoB from the hyperthermophile Methanothermus fervi-dus (17) and mesophile Methanobacterium formicicum (2), re-spectively, have amino acid sequences that are 78% identical(Fig. 1a), and the tertiary structures of recombinant (r)(HMfB)2 and (rHFoB)2 dimers have a root-mean-square de-viation for backbone atoms of only 0.65 6 0.13 Å2 (18, 20).However, under identical solution conditions, they have max-imum free energies of unfolding of 14.6 and 7.2 kcal/mol,respectively, and unfold at temperatures that differ by .30°C(8). To identify the residues responsible for this large differ-ence in structural stability, site-specific mutagenesis, followedby synthesis and purification from Escherichia coli, has beenused to obtain rHMfB and rHFoB variants with residue sub-stitutions at all of the sites at which rHMfB and rHFoB differ(Fig. 1a and b). Here we report the thermostability of eachvariant based on circular dichroism (CD) measurements ofthermal unfolding transitions and the interpretation of thesedata in terms of the difference in stability of the (rHMfB)2 and(rHFoB)2 histone folds.
MATERIALS AND METHODS
Site-directed mutagenesis and recombinant histone purification. Specific mu-tations were introduced into hmfB and hfoB by using the Altered Sites I and II(Promega Corp., Madison, Wis.) and QuikChange (Stratagene, La Jolla, Calif.)kits. The manufacturers’ protocols were followed, with mutagenic oligonucleo-tide primers (sequences available on request) purchased from Ransom HillBiosciences (Romana, Calif.). Each construction was confirmed by DNA se-quencing, and then cloned into pKK223-3 and transformed into E. coli JM105 forrHMfB synthesis (15, 16) or cloned into pRAT4 and transformed into E. coliB834(DE3) for rHFoB synthesis (9, 13). Isopropyl-b-D-thiogalactopyranoside(400 mM; IPTG) was added to cultures of the E. coli transformants growing at
37°C in Luria-Bertani medium containing 100 mg of ampicillin/ml to inducevariant synthesis (13, 15), and incubation was continued for 3 h at 37°C. Aliquotswere removed at 30-min intervals, and the polypeptide content of the E. coli cellswas visualized by Coomassie blue staining after cell lysis and sodium dodecylsulfate-polyacrylamide gel electrophoresis (15). Accumulation of an archaealhistone was indicated by an increase in the intensity of a stained band thatmigrated faster than almost all other polypeptides present. The E. coli cells wereharvested by centrifugation, and lysed by passage through a French pressure cell,and the variant was then purified from the lysate as previously described forrHMfB and rHFoB (8, 9, 16). The composition and concentration of the archaealhistone preparations were determined by acid hydrolysis and amino acid analysis(8), and DNA binding and archaeal nucleosome formation were documented byagarose gel shift assays (16, 17). In cases where there was no detectable accu-mulation of an archaeal histone after IPTG addition, the accuracy of the recom-binant DNA construction was reconfirmed by sequencing, and additional at-tempts were made to induce synthesis of the variant, but generally withoutsuccess.
CD spectropolarimetry. CD spectra of the recombinant archaeal histone vari-ants were obtained at 25°C using an AVIV 62A-DS spectropolarimeter (Aviv,Lakewood, N.J.) with a 1-mm-path-length cylindrical quartz cell and averagingtimes of 2 to 5 s. Temperature-induced changes in the CD measurements at 222nm (u222) were determined at 1°C intervals from 0 to 99°C using a 10-mm-path-length quartz cell with an averaging time of 5 s. The temperature was maintainedto within 60.2°C with a 1-min equilibrium time between each temperatureincrement. In terms of histone monomers, the rHMfB and rHFoB variant solu-tions investigated ranged in concentration from 2.6 to 5.3 mM and from 1.0 to 5.6mM, respectively. Baseline measurements were determined using deionized wa-ter and subtracted from the experimental data. As previously described for botharchaeal and eucaryal histones (7, 8), a two-state model in which a histone dimerunfolds directly into two random coil monomers (D32M) with negligibly pop-ulated intermediate states fitted the experimental data and was used to calculatestandard state (1M) midpoint unfolding temperatures (T° values).
RESULTS AND DISCUSSION
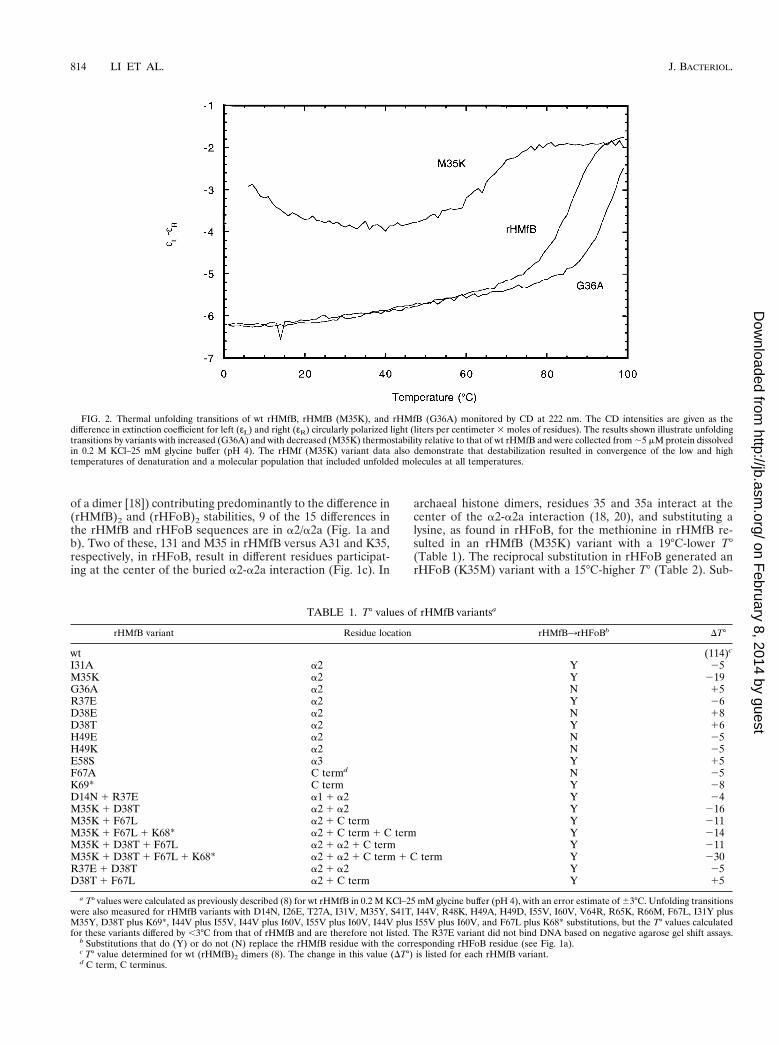
Thermal unfolding transitions and T° values. (rHMfB)2 and(rHFoB)2 have secondary structures that are over 70% a-he-lical, and changes in ellipticity at 222 nm were therefore mea-sured to monitor protein unfolding transitions (6). To observecomplete unfolding transitions below 99°C, the highest oper-ating temperature of the CD spectropolarimeter, measure-ments of (rHMfB)2 and (rHFoB)2 and their variants weremade in solutions containing 0.2 M and 1 M KCl, respectively(8). The thermal transition mid-point temperatures (Tm) ob-served for unfolding were dependent on the protein concen-tration, as expected for dimers, and were extrapolated to 1Mstandard-state values (T°) for comparative purposes. Examplesof the unfolding transitions observed are shown in Fig. 2, andwhen assayed, reduction of the temperature subsequently re-
* Corresponding author. Mailing address: Department of Microbi-ology, The Ohio State University, Columbus, OH 43210. Phone: (614)292-2301. Fax: (614) 292-8120. E-mail: [email protected].
812
on February 8, 2014 by guest
http://jb.asm.org/
Dow
nloaded from
sulted in the histone variant refolding. As described in detailpreviously for wild-type (wt) archaeal histones (8), based onthe excellent fit of the two-state model (D32M) to the data,this model was used to calculate T° values for each variant fromthe thermal unfolding transition data (Tables 1 and 2).
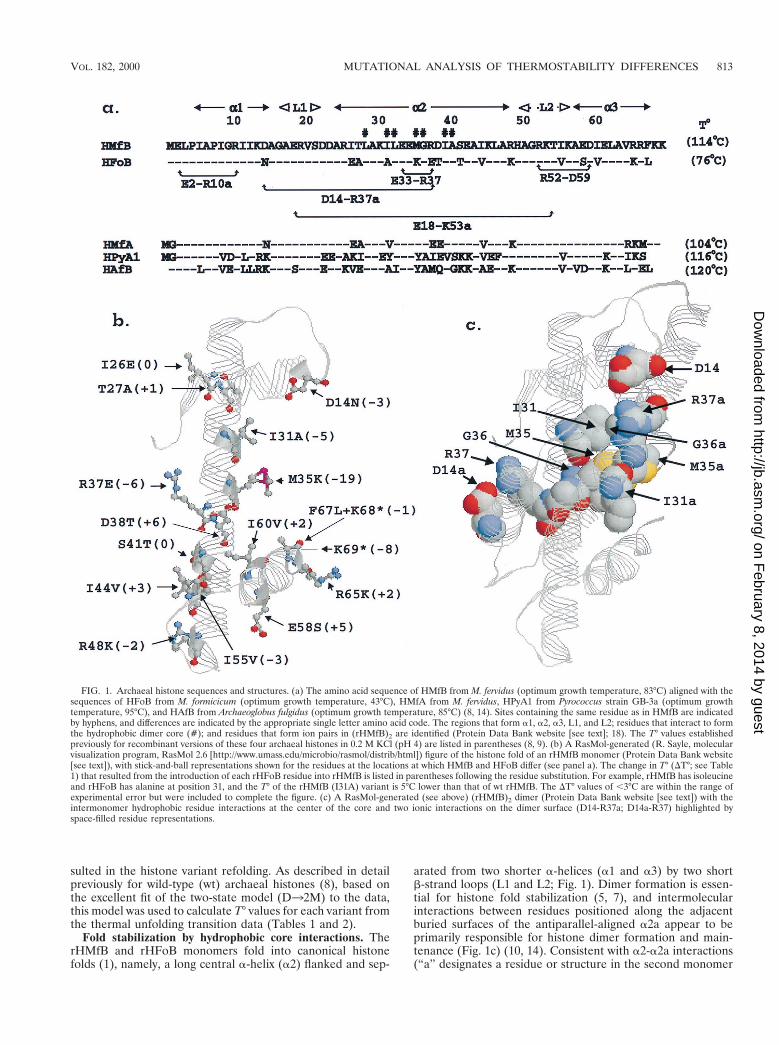
Fold stabilization by hydrophobic core interactions. TherHMfB and rHFoB monomers fold into canonical histonefolds (1), namely, a long central a-helix (a2) flanked and sep-
arated from two shorter a-helices (a1 and a3) by two shortb-strand loops (L1 and L2; Fig. 1). Dimer formation is essen-tial for histone fold stabilization (5, 7), and intermolecularinteractions between residues positioned along the adjacentburied surfaces of the antiparallel-aligned a2a appear to beprimarily responsible for histone dimer formation and main-tenance (Fig. 1c) (10, 14). Consistent with a2-a2a interactions(“a” designates a residue or structure in the second monomer
FIG. 1. Archaeal histone sequences and structures. (a) The amino acid sequence of HMfB from M. fervidus (optimum growth temperature, 83°C) aligned with thesequences of HFoB from M. formicicum (optimum growth temperature, 43°C), HMfA from M. fervidus, HPyA1 from Pyrococcus strain GB-3a (optimum growthtemperature, 95°C), and HAfB from Archaeoglobus fulgidus (optimum growth temperature, 85°C) (8, 14). Sites containing the same residue as in HMfB are indicatedby hyphens, and differences are indicated by the appropriate single letter amino acid code. The regions that form a1, a2, a3, L1, and L2; residues that interact to formthe hydrophobic dimer core (#); and residues that form ion pairs in (rHMfB)2 are identified (Protein Data Bank website [see text]; 18). The T° values establishedpreviously for recombinant versions of these four archaeal histones in 0.2 M KCl (pH 4) are listed in parentheses (8, 9). (b) A RasMol-generated (R. Sayle, molecularvisualization program, RasMol 2.6 [http://www.umass.edu/microbio/rasmol/distrib/html]) figure of the histone fold of an rHMfB monomer (Protein Data Bank website[see text]), with stick-and-ball representations shown for the residues at the locations at which HMfB and HFoB differ (see panel a). The change in T° (DT°; see Table1) that resulted from the introduction of each rHFoB residue into rHMfB is listed in parentheses following the residue substitution. For example, rHMfB has isoleucineand rHFoB has alanine at position 31, and the T° of the rHMfB (I31A) variant is 5°C lower than that of wt rHMfB. The DT° values of ,3°C are within the range ofexperimental error but were included to complete the figure. (c) A RasMol-generated (see above) (rHMfB)2 dimer (Protein Data Bank website [see text]) with theintermonomer hydrophobic residue interactions at the center of the core and two ionic interactions on the dimer surface (D14-R37a; D14a-R37) highlighted byspace-filled residue representations.
VOL. 182, 2000 MUTATIONAL ANALYSIS OF THERMOSTABILITY DIFFERENCES 813
on February 8, 2014 by guest
http://jb.asm.org/
Dow
nloaded from
of a dimer [18]) contributing predominantly to the difference in(rHMfB)2 and (rHFoB)2 stabilities, 9 of the 15 differences inthe rHMfB and rHFoB sequences are in a2/a2a (Fig. 1a andb). Two of these, 131 and M35 in rHMfB versus A31 and K35,respectively, in rHFoB, result in different residues participat-ing at the center of the buried a2-a2a interaction (Fig. 1c). In
archaeal histone dimers, residues 35 and 35a interact at thecenter of the a2-a2a interaction (18, 20), and substituting alysine, as found in rHFoB, for the methionine in rHMfB re-sulted in an rHMfB (M35K) variant with a 19°C-lower T°(Table 1). The reciprocal substitution in rHFoB generated anrHFoB (K35M) variant with a 15°C-higher T° (Table 2). Sub-
FIG. 2. Thermal unfolding transitions of wt rHMfB, rHMfB (M35K), and rHMfB (G36A) monitored by CD at 222 nm. The CD intensities are given as thedifference in extinction coefficient for left (εL) and right (εR) circularly polarized light (liters per centimeter 3 moles of residues). The results shown illustrate unfoldingtransitions by variants with increased (G36A) and with decreased (M35K) thermostability relative to that of wt rHMfB and were collected from ;5 mM protein dissolvedin 0.2 M KCl–25 mM glycine buffer (pH 4). The rHMf (M35K) variant data also demonstrate that destabilization resulted in convergence of the low and hightemperatures of denaturation and a molecular population that included unfolded molecules at all temperatures.
TABLE 1. T° values of rHMfB variantsa
rHMfB variant Residue location rHMfB3rHFoBb DT°
wt (114)c
I31A a2 Y 25M35K a2 Y 219G36A a2 N 15R37E a2 Y 26D38E a2 N 18D38T a2 Y 16H49E a2 N 25H49K a2 N 25E58S a3 Y 15F67A C termd N 25K69* C term Y 28D14N 1 R37E a1 1 a2 Y 24M35K 1 D38T a2 1 a2 Y 216M35K 1 F67L a2 1 C term Y 211M35K 1 F67L 1 K68* a2 1 C term 1 C term Y 214M35K 1 D38T 1 F67L a2 1 a2 1 C term Y 211M35K 1 D38T 1 F67L 1 K68* a2 1 a2 1 C term 1 C term Y 230R37E 1 D38T a2 1 a2 Y 25D38T 1 F67L a2 1 C term Y 15
a T° values were calculated as previously described (8) for wt rHMfB in 0.2 M KCl–25 mM glycine buffer (pH 4), with an error estimate of 63°C. Unfolding transitionswere also measured for rHMfB variants with D14N, I26E, T27A, I31V, M35Y, S41T, I44V, R48K, H49A, H49D, I55V, I60V, V64R, R65K, R66M, F67L, I31Y plusM35Y, D38T plus K69*, I44V plus I55V, I44V plus I60V, I55V plus I60V, I44V plus I55V plus I60V, and F67L plus K68* substitutions, but the T° values calculatedfor these variants differed by ,3°C from that of rHMfB and are therefore not listed. The R37E variant did not bind DNA based on negative agarose gel shift assays.
b Substitutions that do (Y) or do not (N) replace the rHMfB residue with the corresponding rHFoB residue (see Fig. 1a).c T° value determined for wt (rHMfB)2 dimers (8). The change in this value (DT°) is listed for each rHMfB variant.d C term, C terminus.
814 LI ET AL. J. BACTERIOL.
on February 8, 2014 by guest
http://jb.asm.org/
Dow
nloaded from

stitution of either tyrosine or phenylalanine at position 35 alsoresulted in rHFoB (K35Y; K35F) variants with much-in-creased T° values (Table 2), whereas the rHMfB (M35Y) vari-ant had essentially the same T° as that of wt rHMfB. Substi-tuting a larger hydrophobic residue at position 31 alsogenerated rHFoB (A31I; A31Y) variants with increased ther-mal stabilities, and the rHFoB (A31I plus K35M; A31I plusK35Y) variants with large hydrophobic residues at both posi-tions had even higher thermal stabilities (Table 2). Maintain-ing hydrophobicity but decreasing the size of the residue atposition 31 in rHMfB (I31A) gave a reduced T° (Table 1). Aplasmid was constructed to generate the rHMfB (I31A plusM35K) variant, but when the construct was expressed in E. coli,the variant did not accumulate, suggesting that combining A31and K35 resulted in a protein that folded inadequately and wastherefore degraded rapidly in vivo. As several of the archaealhistones from hyperthermophiles have tyrosines naturally atpositions 31 and/or 35 (Fig. 1a) (8, 14), an rHMfB (I31Y plusM35Y) variant was constructed with tyrosines at both positionsand found to have a T° very similar to that of wt rHMfB.
Glycines 36 and 36a flank the 35-35a interaction at thecenter of both (rHMfB)2 and (rHFoB)2 dimers, and the pres-ence of these small residues results in cavities in both hydro-phobic cores (Protein Data Bank [http://www.rcsb.org/pdb/cgi/explore.cgi?pid55348933077468&page50&pdbId51A7W];18, 20). Larger alanine residues are present at these locationsin some of the archaeal histones from hyperthermophiles (Fig.1a) (8, 14). Both the rHMfB (G36A) and rHFoB (G36A)variants had increased T° values relative to those of wt rHMfBand wt rHFoB, respectively (Tables 1 and 2), consistent withreducing the size of internal cavities adding stability (11, 19).The side chain of residue 67 extends from a3 into the central
hydrophobic core, and both rHMfB and rHFoB have a large,hydrophobic residue at this location, phenylalanine andleucine, respectively. The rHFoB (L67F) variant had a stabilityvery similar to that of wt rHFoB, and the reciprocal substitu-tion generated an rHMfB (F67L) variant with marginally in-creased thermal stability. Combining the F67L substitutionwith M35K resulted in an rHMfB (M35K plus F67L) variantthat was more thermally stable than rHMfB (M35K) but stillmuch less stable than wt rHMfB. As expected, the rHMfB(F67A) variant, with a much smaller residue at position 67, hadreduced thermal stability (Table 1).
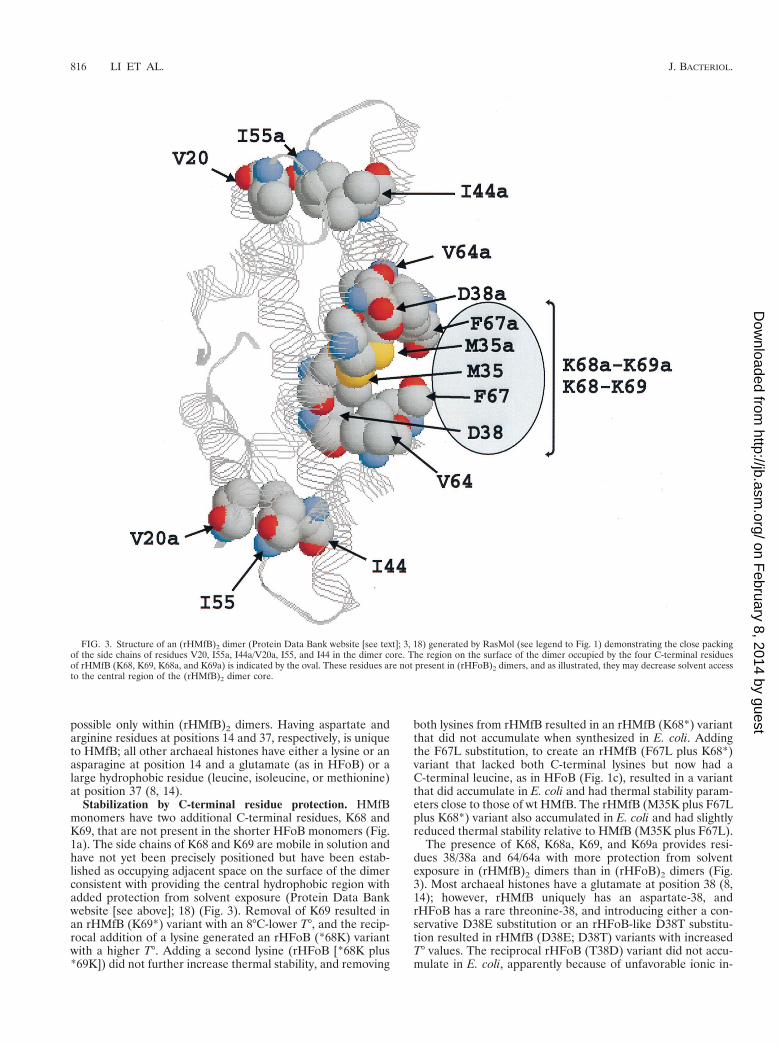
There are three isoleucines in HMfB (I44, I55, and I60) thatare replaced by three smaller valines in HFoB, and I44V, I55V,and I60V and all possible combinations of these substitutionswere therefore introduced into rHMfB to determine if valine-for-isoleucine substitutions would systematically decrease sta-bility (19). The results obtained, however, show no such con-sistent trend, and introducing all three substitutions resulted inan rHMfB (I44V plus I55V plus I60V) variant with a T° almostidentical to that of wt rHMfB. The side chains of I44/I44a arepartially buried by dimer formation and interact with the I55/I55a side chains that are fully buried and tightly packed withthe side chains of V20a/V20 by facing inwards from the L1-L2aand L1a-L2 regions (Fig. 3). All archaeal histones exceptHMfB, including many from hyperthermophiles, have a valineor alanine at position 44, almost all have valine at position 20,and only HMfB and HMfA (also from M. fervidus) have anisoleucine, not a valine, at position 55 (8, 14). It appears likelytherefore that smaller valine side chains are more readily ac-commodated within the space available surrounding positions20/20a, 44/44a, and 55/55a in an archaeal histone dimer, andconsistent with this, the rHMfB (I55V) and rHFoB (V55I)variants had increased and decreased thermal stabilities, re-spectively. The side chains of isoleucines 60/60a are also par-tially buried, adjacent to the side chains of leucines 28/28a andalanines 43/43a, forming hydrophobic core interactions be-tween a3/a3a and a2/a2a. Most archaeal histones do have anisoleucine at position 60 (8, 14), and as the rHMfB (I60V) andrHFoB (V60I) variants had slightly reduced and increasedthermal stabilities, respectively, having an isoleucine residue atposition 60 is preferred for stability.
Stabilization by ionic interactions. The histone fold is sta-bilized by a buried, intramolecular arginine-aspartate interac-tion in all archaeal (R52-D59 in Fig. 1a) (14) and eucaryal (10)histones, and E2-R10a and E18-K53a intermolecular saltbridges appear similarly to be conserved on the surface ofalmost all archaeal histone dimers (Fig. 1a). The presence ofD14 and R37 in HMfB versus N14 and E37, respectively, inHFoB (Fig. 1a), however, provides (rHMfB)2 dimers with theopportunity to form four additional ion pairs, D14-R27a,D14a-R37, E33-R37, and E33a-R37a, that cannot be formedby (rHFoB)2 dimers (Fig. 1a and c). Preventing this pair for-mation, by substituting glutamate for arginine at position 37,resulted in an rHMfB (R37E) variant with a 6°C-lower T°(Table 1). The reciprocal substitution generated an rHFoB(E37R) variant with increased thermal stability (Table 2) ap-parently primarily by replacing the two potentially repulsiveE33-E37 interactions along the solvent-exposed surfaces of a2and a2a with two attractive E33-R37 interactions. rHFoB(N14D; N14K; N14K plus E37R) variants were also con-structed with the idea of creating repulsive D14-E37a andN14K-R37a interactions and an attractive K14-E37a interac-tion, but these variants had thermal stabilities only marginallydifferent from that of wt HFoB. The orientation of a1 relativeto a2a is different in (rHMfB)2 and (rHFoB)2 dimers (20), anddirect residue 14-37a and 14a-37 interactions may therefore be
TABLE 2. T° values of rHFoB variantsa
rHFoB variant Residue location rHFoB3rHMfBb DT°
wt (92)c
E26I a2 Y 16A31I a2 Y 111A31Y a2 N 112K35M a2 Y 115K35Y a2 N 117K35F a2 N 120G36A a2 N 18E37R a2 Y 14V55I L2 Y 25S58E a3 Y 14V60I a3 Y 15V64R a3 N 28A31I 1 K35M a2 1 a2 Y 125A31I 1 K35Y a2 1 a2 N 119A31I 1 K35M 1 V64R a2 1 a2 1 a3 N 113K35M 1 V64R a2 1 a3 N 18K35M 1 L67F a2 1 a3 Y 112K35M 1 *68K a2 1 C termd Y 16K35M 1 *68K 1 *69K a2 1 C term 1 C term Y 121E37R 1 T38D a2 1 a2 Y 24
a T° values were calculated as previously described (8) for wt rHFoB in 1 MKCl–25 mM glycine buffer (pH 4), with an error estimate of 63°C. Unfoldingtransitions were also measured for rHFoB variants with N14D, N14K, N14K plusE37R, E37R plus T38D, L67F, *68K, *68K plus *69K, and L67F plus *68K plus*69K substitutions, but the T° values calculated for these variants differed by,3°C from that of wt rHFoB and are therefore not listed.
b Substitutions that do (Y) or do not (N) replace the rHFoB residue with thecorresponding rHMfB residue (see Fig. 1a).
c T° value determined for wt (rHFo)2 dimers (8). The change in this value(DT°) is listed for each rHFoB variant.
d C term, C terminus.
VOL. 182, 2000 MUTATIONAL ANALYSIS OF THERMOSTABILITY DIFFERENCES 815
on February 8, 2014 by guest
http://jb.asm.org/
Dow
nloaded from
possible only within (rHMfB)2 dimers. Having aspartate andarginine residues at positions 14 and 37, respectively, is uniqueto HMfB; all other archaeal histones have either a lysine or anasparagine at position 14 and a glutamate (as in HFoB) or alarge hydrophobic residue (leucine, isoleucine, or methionine)at position 37 (8, 14).
Stabilization by C-terminal residue protection. HMfBmonomers have two additional C-terminal residues, K68 andK69, that are not present in the shorter HFoB monomers (Fig.1a). The side chains of K68 and K69 are mobile in solution andhave not yet been precisely positioned but have been estab-lished as occupying adjacent space on the surface of the dimerconsistent with providing the central hydrophobic region withadded protection from solvent exposure (Protein Data Bankwebsite [see above]; 18) (Fig. 3). Removal of K69 resulted inan rHMfB (K69*) variant with an 8°C-lower T°, and the recip-rocal addition of a lysine generated an rHFoB (*68K) variantwith a higher T°. Adding a second lysine (rHFoB [*68K plus*69K]) did not further increase thermal stability, and removing
both lysines from rHMfB resulted in an rHMfB (K68*) variantthat did not accumulate when synthesized in E. coli. Addingthe F67L substitution, to create an rHMfB (F67L plus K68*)variant that lacked both C-terminal lysines but now had aC-terminal leucine, as in HFoB (Fig. 1c), resulted in a variantthat did accumulate in E. coli and had thermal stability param-eters close to those of wt HMfB. The rHMfB (M35K plus F67Lplus K68*) variant also accumulated in E. coli and had slightlyreduced thermal stability relative to HMfB (M35K plus F67L).
The presence of K68, K68a, K69, and K69a provides resi-dues 38/38a and 64/64a with more protection from solventexposure in (rHMfB)2 dimers than in (rHFoB)2 dimers (Fig.3). Most archaeal histones have a glutamate at position 38 (8,14); however, rHMfB uniquely has an aspartate-38, andrHFoB has a rare threonine-38, and introducing either a con-servative D38E substitution or an rHFoB-like D38T substitu-tion resulted in rHMfB (D38E; D38T) variants with increasedT° values. The reciprocal rHFoB (T38D) variant did not accu-mulate in E. coli, apparently because of unfavorable ionic in-
FIG. 3. Structure of an (rHMfB)2 dimer (Protein Data Bank website [see text]; 3, 18) generated by RasMol (see legend to Fig. 1) demonstrating the close packingof the side chains of residues V20, I55a, I44a/V20a, I55, and I44 in the dimer core. The region on the surface of the dimer occupied by the four C-terminal residuesof rHMfB (K68, K69, K68a, and K69a) is indicated by the oval. These residues are not present in (rHFoB)2 dimers, and as illustrated, they may decrease solvent accessto the central region of the (rHMfB)2 dimer core.
816 LI ET AL. J. BACTERIOL.
on February 8, 2014 by guest
http://jb.asm.org/
Dow
nloaded from

teractions, as the addition of E37R to create the adjacentsequence R37-D38 that occurs in rHMfB (Fig. 1a) resulted inan rHFoB (R37E plus T38D) variant that did accumulate in E.coli and had thermal stability parameters similar to those of wtrHFoB. Combining the D38T substitution in rHMfB withM35K and F67L generated rHMfB (M35K plus D38T) and(M35K plus D38T plus F67L) variants with T° values interme-diate between those of rHMfB (M35K) and wt rHMfB, butwhen the terminal lysine residues were also removed, the re-sulting rHMfB (M35K plus D38T plus F67L plus K68*) varianthad a 30°C-lower T° (Table 1).
A valine occupies position 64 in both HMfB and HFoB, anda hydrophobic residue is present at this location in all archaealhistones, except rHMfA, which has an arginine at position 64(Fig. 1a). (rHMfA)2 and (rHMfB)2 have very similar structures(3; Protein Data Bank website [see above]), but (rHMfA)2dimers unfold at temperatures ;10°C lower than (rHMfB)2 (8,9), and replacing the arginine at position 64 with valine re-sulted in an rHMfA (R64V) variant with an ;10°C-higher T°(results not shown). Introducing a reciprocal V64R substitu-tion into rHMfB did not result in decreased thermal stability,although the T° of the rHFoB (V64R) variant was 8°C lowerthan that of rHFoB (Table 2). Combining V64R with M35Kresulted in an rHMfB (V64R plus M35K) variant that did notaccumulate in E. coli, possibly due to repulsive K35-R64 andK35a-R64a interactions, although such interactions would alsobe predicted to occur in the core of the rHFoB (V64R) variant.
Conclusions. Based on the ;30 archaeal histone sequencesso far established (8, 14), conserved residues can be identifiedthat are presumably essential for histone fold formation and/orDNA binding (10, 14), and predictions can be made for resi-dues that might confer differences in thermostability and saltand pH dependence. Here we have investigated the basis fordifferences in thermostability by measuring the effects of resi-due substitutions on temperature-induced unfolding by focus-ing on two archaeal histones with very different stabilities butsimilar structures (Protein Data Bank website [see above]; 8,18, 20). The increased stability of proteins from thermophilescompared with that of proteins from mesophiles is frequentlyattributed to improved hydrophobic core packing (11, 19)and/or an increased number of attractive ionic interactions (4,12), and the results reported here are consistent with differ-ences in intermonomer hydrophobic core interactions domi-nating in determining the difference in the (rHMfB)2 and(rHFoB)2 fold stabilities. Introducing large hydrophobic resi-dues and decreasing cavity sizes within the cores increasedthermostability, combinations of such substitutions had addi-tive effects, and introducing smaller or potentially polar resi-dues decreased stability. Most substitutions that added or re-moved a potentially attractive or repulsive ionic interactionalso had the predicted positive or negative effect on thermo-stability, but this was not always the case, underlining thelimitations of predicting stability even for such very simpleproteins. Approximately 70 variants have so far been con-structed, with one to four substitutions, and assayed for ther-mostability, and residue differences that together account formost of the difference in the stabilities of (rHFoB)2 and(rHMfB)2 have been identified. Some results, for example, theincreased thermostability of the rHMfB (D38E) variant, how-ever, remain inexplicable, and it has been assumed that thelack of accumulation in E. coli identifies a variant that is somisfolded that it is rapidly degraded when synthesized in E.coli, but this has not been systematically proven. The rHFoBand rHMfB sequences differ at 15 locations (Fig. 1a), andconstructing and assaying the thermodynamic stabilities of allpossible rHMfB variants with all combinations of rHFoB res-
idues, and/or vice versa, would be a monumental task. As somesubstitutions would probably also change the overall fold, thiswould then also be a misguided undertaking if all argumentswere based on only the wt (rHMfB)2 and (rHFoB)2 structures.With this in mind, the structures of selected archaeal histonevariants must now be determined.
ACKNOWLEDGMENTS
We thank K. Sandman for construction, purification, and DNAbinding assays of several of the archaeal histone variants used in thisstudy.
This research was supported by a grant from the National Institutesof Health (GM53185).
REFERENCES
1. Arents, G., and E. N. Moudrianakis. 1995. The histone fold: a ubiquitousarchitectural motif utilized in DNA compaction and protein dimerization.Proc. Natl. Acad. Sci. USA 92:11170–11174.
2. Darcy, T. J., K. Sandman, and J. N. Reeve. 1995. Methanobacterium formici-cum, a mesophilic methanogen, contains three HFo histones. J. Bacteriol.177:858–860.
3. Decanniere, K., K. Sandman, J. N. Reeve, and U. Heinemann. 1996. Crys-tallization and preliminary X-ray characterization of the Methanothermusfervidus histones HMfA and HMfB. Proteins Struct. Funct. Genet. 24:269–271.
4. Elcock, A. H. 1998. The stability of salt bridges at high temperatures: impli-cations for hyperthermophilic proteins. J. Mol. Biol. 284:489–502.
5. Grayling, R. A., W. J. Becktel, and J. N. Reeve. 1995. Structure and stabilityof histone HMf from the hyperthermophilic archaeon Methanothermus fer-vidus. Biochemistry 34:8441–8448.
6. Hirst, J. D., and C. L. Brooks III. 1994. Helicity, circular dichroism andmolecular dynamics of proteins. J. Mol. Biol. 243:173–178.
7. Karantza, V., A. D. Baxevanis, E. Freire, and E. N. Moudrianakis. 1995.Thermodynamic studies of the core histones: ionic strength and pH depen-dencies of H2A-H2B dimer stability. Biochemistry 35:5988–5996.
8. Li, W.-T., R. A. Grayling, K. Sandman, S. Edmondson, J. W. Shriver, andJ. N. Reeve. 1998. Thermodynamic stability of archaeal histones. Biochem-istry 30:10563–10572.
9. Li, W.-T., K. Sandman, S. L. Pereira, and J. N. Reeve. MJ1647, an openreading frame in the genome of the hyperthermophile Methanococcus jan-naschii, encodes a very thermostable archaeal histone with a C-terminalextension. Extremophiles, in press.
10. Luger, K., A. W. Mader, R. K. Richmond, D. F. Sargent, and T. J. Richmond.1997. Crystal structure of the nucleosome core particle at 2.8 Å resolution.Nature 389:251–260.
11. Pace, C. N. 1992. Contribution of the hydrophobic effect to globular proteinstability. J. Mol. Biol. 226:29–35.
12. Pappenberger, G., H. Schurig, and R. Jaenicke. 1997. Disruption of an ionicnetwork leads to accelerated thermal denaturation of D-glyceraldehyde-3-phosphate dehydrogenase from the hyperthermophilic bacterium Thermo-toga maritima. J. Mol. Biol. 274:676–683.
13. Peranen, J., M. Rikkonen, M. Hyvonen, and L. Kaariainen. 1996. T7 vectorwith a modified T7lac promoter for expression of proteins in E. coli. Anal.Biochem. 236:371–373.
14. Reeve, J. N., K. Sandman, and C. J. Daniels. 1997. Archaeal histones,nucleosomes and transcription initiation. Cell 87:999–1002.
15. Sambrook, J., E. F. Fritsch, and T. Maniatis. 1989. Molecular cloning: alaboratory manual, 2nd ed., p. 6.46–6.48. Cold Spring Harbor LaboratoryPress, Cold Spring Harbor, N.Y.
16. Sandman, K., R. A. Grayling, and J. N. Reeve. 1995. Improved N-terminalprocessing of recombinant proteins synthesized in Escherichia coli. Bio/Tech-nology 13:504–506.
17. Sandman, K., J. A. Krzycki, B. Dobrinski, R. Lurz, and J. N. Reeve. 1990.HMf, a DNA-binding protein isolated from the hyperthermophilic archaeonMethanothermus fervidus, is most closely related to histones. Proc. Natl.Acad. Sci. USA 87:5788–5791.
18. Starich, M. R., K. Sandman, J. N. Reeve, and M. F. Summers. 1996. NMRstructure of HMfB from the hyperthermophile, Methanothermus fervidus,confirms that this archaeal protein is a histone. J. Mol. Biol. 255:187–203.
19. Takano, K., Y. Yamagata, and K. Yutani. 1998. A general rule for therelationship between hydrophobic effects and conformational stability of aprotein: stability and structure of a series of hydrophobic mutants of humanlysozyme. J. Mol. Biol. 280:749–761.
20. Zhu, W., K. Sandman, G. E. Lee, J. N. Reeve, and M. F. Summers. 1998.NMR structure and comparison of the archaeal histone HFoB from themesophile Methanobacterium formicicum with HMfB from the hyperthermo-phile Methanothermus fervidus. Biochemistry 37:10573–10580.
VOL. 182, 2000 MUTATIONAL ANALYSIS OF THERMOSTABILITY DIFFERENCES 817
on February 8, 2014 by guest
http://jb.asm.org/
Dow
nloaded from
Related Documents











