Fertilization and development of quail oocytes after intracytoplasmic sperm injection Anna Hrabia 3,4 , Soichi Takagi 3 , Tamao Ono 5 , Kiyoshi Shimada 2,3 Laboratory of Animal Physiology, 3 Graduate School of Bioagricultural Sciences, Nagoya University, Nagoya, Aichi 464-8601, Japan JSPS Postdoctoral Fellow on leave from Department of Animal Physiology, 4 Agricultural University of Cracow, Al. Mickiewicza 24/28, 30-059 Krakow, Poland Faculty of Agriculture, 5 Shinshiu University, Minamiminowa 399-4598, Japan 1 This research was supported by grant-in-aid from the Japan Society for the Promotion of Science, Postdoctoral Fellowship for Foreign Researchers awarded to A.H. (P 01319) and from the Japan Society for the Promotion of Science, Basic Research (C2) to K.S. (No. 13660286). 2 Correspondence and request for reprints: Kiyoshi Shimada, Ph.D. Laboratory of Animal Physiology, Graduate School of Bioagricultural Sciences, Nagoya University, Nagoya, Aichi 464-8601, Japan. tel. & fax. 052-789-4065 e-mail:[email protected] Running title: Fertilization of quail oocytes by ICSI Key Words: early development, in vitro fertilization, ovum, sperm, spermatid BOR Papers in Press. Published on July 9, 2003 as DOI:10.1095/biolreprod.103.019315

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Fertilization and development of quail oocytes after intracytoplasmic sperm
injection
Anna Hrabia3,4, Soichi Takagi3, Tamao Ono5, Kiyoshi Shimada2,3
Laboratory of Animal Physiology,3 Graduate School of Bioagricultural Sciences,
Nagoya University, Nagoya, Aichi 464-8601, Japan
JSPS Postdoctoral Fellow on leave from Department of Animal Physiology,4 Agricultural
University of Cracow, Al. Mickiewicza 24/28, 30-059 Krakow, Poland
Faculty of Agriculture,5 Shinshiu University, Minamiminowa 399-4598, Japan
1This research was supported by grant-in-aid from the Japan Society for the Promotion of
Science, Postdoctoral Fellowship for Foreign Researchers awarded to A.H. (P 01319) and from
the Japan Society for the Promotion of Science, Basic Research (C2) to K.S. (No. 13660286).
2Correspondence and request for reprints:
Kiyoshi Shimada, Ph.D.
Laboratory of Animal Physiology, Graduate School of Bioagricultural Sciences, Nagoya
University, Nagoya, Aichi 464-8601, Japan.
tel. & fax. 052-789-4065
e-mail:[email protected]
Running title: Fertilization of quail oocytes by ICSI
Key Words: early development, in vitro fertilization, ovum, sperm, spermatid
BOR Papers in Press. Published on July 9, 2003 as DOI:10.1095/biolreprod.103.019315

2
ABSTRACT
The present study was conducted to establish the intracytoplasmic sperm injection (ICSI)
method for in vitro fertilization and development in quail. The efficiency of fertilization of
oocytes was compared (1) between spontaneous and premature ovulation and (2) among
testicular round spermatids, elongated spermatids and immature and mature spermatozoa. The
oocytes were injected with a single spermatozoon or spermatid and cultured for 24 h. Cell
division was histologically observed with Hematoxylin-Eosin (HE) and a nuclear specific
fluorescent dye DAPI. Five out of 30 (16.6%) and 4 out of 30 (13.3%) oocytes injected with
mature sperm were fertilized in the spontaneous and induced ovulation group, respectively. Those
embryos showed development at stages II-VII. Half of the number (3/6) of the oocytes injected
with testicular spermatozoa were fertilized and developed to stages IV-VII and 2 out of 5 oocytes
injected with elongated spermatids were fertilized and developed to stage VI. All oocytes injected
with round spermatids were unfertilized. The results demonstrate that intracytoplasmic injection
of a single sperm into quail oocyte can activate the oocyte and lead to fertilization. Oocytes
prematurely ovulated are capable of fertilizing with mature sperm as those spontaneously
ovulated. In addition, the results suggest that the testicular round spermatids may not possess
sufficient oocyte-activating potency but the elongated spermatids and immature spermatozoa are
competent to participate in fertilization and early embryonic development in quail.

3
INTRODUCTION
Fertilization in birds takes place in the infundibulum, the most upper region of the oviduct,
soon after ovulation. This must occur before the outer layer of vitelline membrane (outer
perivitelline layer) is laid down since spermatozoa are not able to penetrate this layer [1, 2] and
bind to sperm receptors in the inner layer of the vitelline membrane [3]. Unlike mammals, but
common with reptiles, some amphibians and most urodeles (newts and salamanders), birds
exhibit physiological polyspermy. Many sperm pass through the inner layer of vitelline
membrane into germinal disc of an oocyte, where the nuclei undergo decondensation and form
male pronuclei although only one sperm successfully fertilizes the oocyte [1, 4-7]. The
supernumerary pronuclei degenerate at the very early developmental stages of embryo [5-7].
However, it has been shown that the number of spermatozoa reaching the ovum is positively
correlated with ovum size in variety of birds suggesting that a larger ovum requires more
spermatozoa to ensure fertilization [8, 9]. While the physiological role of polyspermy is not
known, it is unclear whether or not a single sperm is capable of activating an oocyte in birds.
Compared to mammals, information about in vitro fertilization is very limited in birds.
Howarth [10] first demonstrated by Delafield’s hematoxylin staining of nuclei that the ovum
immediately after ovulation could be fertilized in vitro when the germinal disc of the ovum was
introduced to millions of sperm in medium. More recently, Nakanishi et al. [6] provided direct
evidence of in vitro fertilization (IVF) of chicken oocytes by showing male and female pronuclei
formation. They also compared details of early nuclear events between in vivo and in vitro
fertilization during 1-4 h of culture and concluded that the process of IVF is comparable to that
in vivo. Furthermore, Nakanishi et al. [11] demonstrated the fertility competency of multiple-

4
ovulated eggs in the chicken after IVF. Tanaka et al. [12] produced viable chicks by implanting
in vitro fertilized ova into the oviduct of recipient hens followed by incubation of the eggs.
Recently, Olszańska et al. [13] investigated development of quail embryos after IVF using in
vitro ovulated oocytes with a special emphasis on the link between the steps of ovulation and
fertilization and with the early cleavage stages under in vitro conditions. They demonstrated for
the first time that cytoplasmic segmentation can occur in the absence of nuclear divisions in the
germinal disc of the quail and showed the existence and significance of ooplasmic maternal
information in birds. These conventional IVF techniques gave an insight to the early nuclear
events to some extent in chicken and quail, but questions remain still unanswered.
The methods of the direct injection of a sperm into an oocyte called intracytoplasmic sperm
injection (ICSI) have been well developed in mammals and contributed to (1) the basic studies of
fertilization, (2) clinical practice in human, (3) production of normal offspring using
morphologically normal and abnormal spermatozoa and spermatogenic cells and (4) production
of transgenic animals. However, in birds no ICSI has been reported.
Establishment of the ICSI method in birds is advantageous to study the mechanism of
fertilization and the role of gametes in this process and to protect endangered species of birds. In
addition, the use of sperm as a vector to carry foreign DNA to the oocyte may help for
production of transgenic birds. Accordingly, the aims of present experiments were (1) to
establish a method of fertilization of quail ova using ICSI followed by short-term culture system
of embryo, (2) to compare fertilization ability of premature ovulated oocytes to those
spontaneously ovulated, and (3) to test the competency of testicular round spermatids, elongated
spermatids and immature spermatozoa in fertilization and embryonic development.

5
MATERIALS AND METHODS
Birds and chemicals
Male and female Japanese quail (Coturnix japonica) were purchased from Tokai Yuki
Company (Toyohashi, Japan) at 6 weeks of age and raised in environmentally controlled room
(room temperature at 24 ± 1ºC with 50 - 60% relative humidity). Birds were caged under a
photoperiod of 14L:10D (lights on at 5:00 and off at 19:00) with free access to food and water.
Total number of 159 quail was used and birds were divided into 3 groups. The first were oocyte
suppliers for ICSI. Female quail individually caged were used at the age of 11-19 weeks. Time of
egg laying was recorded for each quail by video camera-recorder system (camera – ExwaveHAD
SSC-DC430, camera adapter - YS-W170, VHS time lapse – SVT-S 960 ES; Sony, Tokyo,
Japan). They laid eggs consecutively for more than 5 days in a sequence between 14:00-18:00.
Time of ovulation was estimated by the time of oviposition. Ovulation was considered to occur
within 30 min after oviposition of the previous egg in the series [14]. The second group was used
to supply testicular spermatids, spermatozoa and semen for ICSI. Adult male quail (12-25 weeks
old; n=10) were kept individually. The third group supplied fertilized oocytes. Each female was
coupled with a male in a cage. The time of egg laying was recorded as described above.
All chemicals were obtained from Sigma-Aldrich (Tokyo, Japan) except ethanol, glacial
acetic acid and eosin which were obtained from Wako Pure Chemical Industries, Ltd. (Osaka,
Japan).
Collection of oocytes

6
a) Controls
For morphological standardization of early embryo development and examination of the
effect of culture medium and polyvinylpyrrolidone (PVP) on embryo development, the positive
and negative control ova were prepared. The positive control was as follows: 1) in vivo fertilized
ova collected from the oviduct within 50 min after ovulation and cultured in the same conditions
as those after ICSI (n=5), 2) in vivo fertilized ova collected from the oviduct within 50 min after
ovulation and PVP was injected, followed by culture (n=5), 3) in vivo fertilized ova were
collected from the oviduct at different times after ovulation i.e. 3.5, 4-5, 6, 7, 9, 10, 12, 13, 15,
16, 17 and 20 h, which reflect different stages of embryonic development (n=33), 4) in vivo
fertilized eggs immediately after spontaneous oviposition (n=10). As a negative control,
unfertilized ova were collected within 50 min after ovulation and PVP was injected, followed by
culture (sham-operated control; n=5). Unfertilized eggs were also collected from virgin quail
after oviposition (n=10).
b) Spontaneous ovulation
To obtain ova after spontaneous ovulation, birds (n=51) were killed 50-60 min after
oviposition and oocytes were recovered from the infundibulum or the upper part of the magnum.
c) Premature ovulation
Quail (n=30) were subcutaneously injected with aLHRH ([Gln8]-LHRH; 25 µg/ 0.1 ml of
0.9% NaCl/ quail; Sigma-Aldrich, Tokyo, Japan) 12 h before expected ovulation and sacrificed
9-11 h after the treatment i.e. about 1 h after premature oviposition. Oocytes were collected from
the infundibulum or the upper part of the magnum. Any oocytes present in the other region of the
oviduct were not used. Induced ovulation occurred 1.5 h-5 h prematurely.

7
Collection of semen sperm and testicular spermatids and spermatozoa
The isolated male was introduced to the isolated female for mating. Semen was collected with
swab from cloaca of female quail immediately after mating and washed 3 times by centrifugation
at 1000 x g for 5 min in 1 ml of 0.9% NaCl solution. The washed sperm were diluted with 12%
PVP in Dulbecco Modified Eagle Medium (DMEM). About 100 µl of this solution was placed
on Petri dish (35 x 10 mm; cat. no. 35 1008; Falcon, Becton Dickinson Labware, Franklin Lakes,
NJ, USA), covered with mineral oil and kept until injection at room temperature (25 - 26°C).
To collect round and elongated spermatids and testicular spermatozoa, male quail (n=4) were
killed by dislocation of atlas bone and the testes were excised. They were cut in pieces and a
piece of testis was placed in 0.9% NaCl and minced by scissors. One part of this suspension was
mixed thoroughly with 2 parts of PVP solution and kept for up to 5 h on Petri dish before
spermatozoa or spermatids were selected for injection into spontaneously ovulated oocytes.
Preparation of injection pipettes
To prepare pipettes for ICSI, borosilicate glass capillary tubing (1 mm o.d., 0.75 mm i.d., cat.
no. B100-75-10; Sutter Instrument Co., Novato, CA, USA) was drawn with a pipette puller (P-
97/IVF; Sutter Instrument Co.) and the tip of the pipette was broken with a microforge (MF-900;
Narishige Instrument, Tokyo, Japan ) such that the inner diameter at and near the tip was
approximately 10 µm. Next, the tip of the pipette was beveled at a 30° angle with a micropipette
beveler (BV-10; Sutter Instrument Co.), dipped in 70% ethanol and dried. Then the injection

8
pipette was connected to micromanipulator (ONM-1; Narishige Instrument), half-filled with
silicon oil and a few µl of 0.9% NaCl were sucked into the pipette. The injection pipettes for
round and elongated spermatids were prepared in the same way except beveling.
Intracytoplasmic injection of sperm and round and elongated spermatids
Immediately before injection, a single sperm or spermatid was isolated from PVP-sperm or
PVP-testicular tissue solution, respectively, into the injection pipette under the Hoffman
modulation contrast microscope (IX70; Olympus, Tokyo, Japan). The round and elongated
spermatids were recognized from other cells by their size and shape. The round spermatids in
diameter 6-8 µm have centrally located condensed chromatin, and elongated spermatids are easy
to distinguish by visible elongated tail (Fig. 1). The ovum was placed into DMEM in a plastic
dish (35 x 18 mm; multidish 6 wells, Nunclon Delta, Denmark) and sperm or spermatid was
injected into the central area of the germinal disc using a micromanipulator connected to the
injector (IM-9B; Narishige Instrument) with silicon tubing filled with silicon oil, under
stereomicroscope (SZ11; Olympus) (Fig. 1). This manipulation was performed with the aid of an
image-processor system (Image Σ-III, Nippon Avionics, Tokyo, Japan). It took 10-15 min from
sacrifice of female quail (oocyte supplier) to completion of sperm or spermatid injection into
oocyte.
Culture

9
The sperm or spermatid injected ova were cultured in plastic dishes (35 x 18 mm; multidish 6
wells) for 24 h at 41°C under 5% CO2 in air, in DMEM containing 3 mg/ ml BSA (fraction V;
Sigma-Aldrich, Tokyo, Japan) and 2 µl/ ml antibiotic-antimycotic solution (with 10,000 units
penicillin, 10 mg streptomycin and 25 µg amphotericin B per ml).
Observation of embryo
Fertilization and development of embryo were staged as described by Eyal-Giladi and Kochav
[15] under stereomicroscope (SZ11; Olympus). The blastoderm was cut and in conformity with
the method of Nakanishi et al. [6] fixed in ethanol/ glacial acetic acid (3:1), embedded in paraffin
and sectioned vertically. Half the sections were stained with a fluorescent dye 4,6-diamidino-2-
phenylindole (DAPI) for nuclei in order to examine the cell division by the method of
Waddington et al. [7] and observed under epifluorescent microscope (BX60; Olympus) within 20
h. Additionally, the other half of sections was stained with hematoxylin and eosin (HE) using
standard procedures. Stained sections were imaged using a light microscope.
RESULTS
Controls
Figure 2 shows examples of control blastoderms of in vivo fertilized, developing oocytes.
Quail embryo collected from the shell gland 9 h after ovulation was at stage III in which
individual blastomeres and cleavage furrows were seen (Fig. 2A). Much more advanced cleavage

10
was observed in the embryo collected from the shell gland 12 h after ovulation (stage V; Fig.
2B). In embryo at stage VIII (Fig. 2C) obtained 16-17 h after ovulation the area pellucida was
under a way of formation. In the blastoderm of the egg collected immediately after oviposition
(stage X) formation of the area pellucida was completed (Fig. 3A). In contrast, unfertilized
oviposited eggs from virgin quail always contained numerous vacuoles at the periphery of
germinal disc and a part of their cytoplasm was often segmented into pseudo cells (Fig. 3B). One
of sham-operated oocytes in which PVP was injected into unfertilized oocyte collected from the
infundibulum had cleaved cytoplasm similar to stage IV (pseudo blastoderm) (Fig. 3C).
Remained four sham-operated oocytes did not show any cleavage and had vacuoles at the
periphery of germinal disc. Naturally (in vivo) fertilized oocytes collected from the infundibulum
developed to stages IX-XI in DMEM and fertilized oocytes in vivo which were injected with
PVP and incubated in DMEM, developed to stages VII-X (data not shown).
DAPI-stained nuclei giving intensive blue fluorescence in fertilized oocytes were
distinguished by their round shape, size and location. The number of nuclei observed on sections
varied depending on stage of embryo development i.e. several at very early stages (Fig. 2D) and
numerous at advanced stages (Figs. 2F and 3D). Control embryos of in vivo fertilized oocytes
developed in the oviduct or in culture for 24 h always showed nuclei identified by DAPI (Figs.
2D, E, F and 3D) and HE (Figs. 2G, H, I and 3G) staining. In unfertilized eggs no nuclei were
seen after staining, although a part of their cytoplasm showed cleavage (Figs. 3E and H). Two
out of 5 sham-operated control oocytes showed irregularly fragmented DNA bodies with DAPI
(Fig. 3F) and HE (Fig. 3I) staining, while in 3 other oocytes no fragmented DNA was present.
HE staining showed individual cells with nuclei (Figs. 2H, I and 3G) as well as layers of the

11
germinal disc such as yolk, cytoplasm (Figs. 2G and 3I), epiblast and primary hypoblast (Fig.
3G). DAPI staining showed more distinguishable nuclei.
Spontaneously and prematurely ovulated oocytes
Several embryos after ICSI showed “normal” cleavage, but contained no fluorescent nuclei or
exhibited signs of degeneration of nuclei seen as bright irregular spots. They were considered as
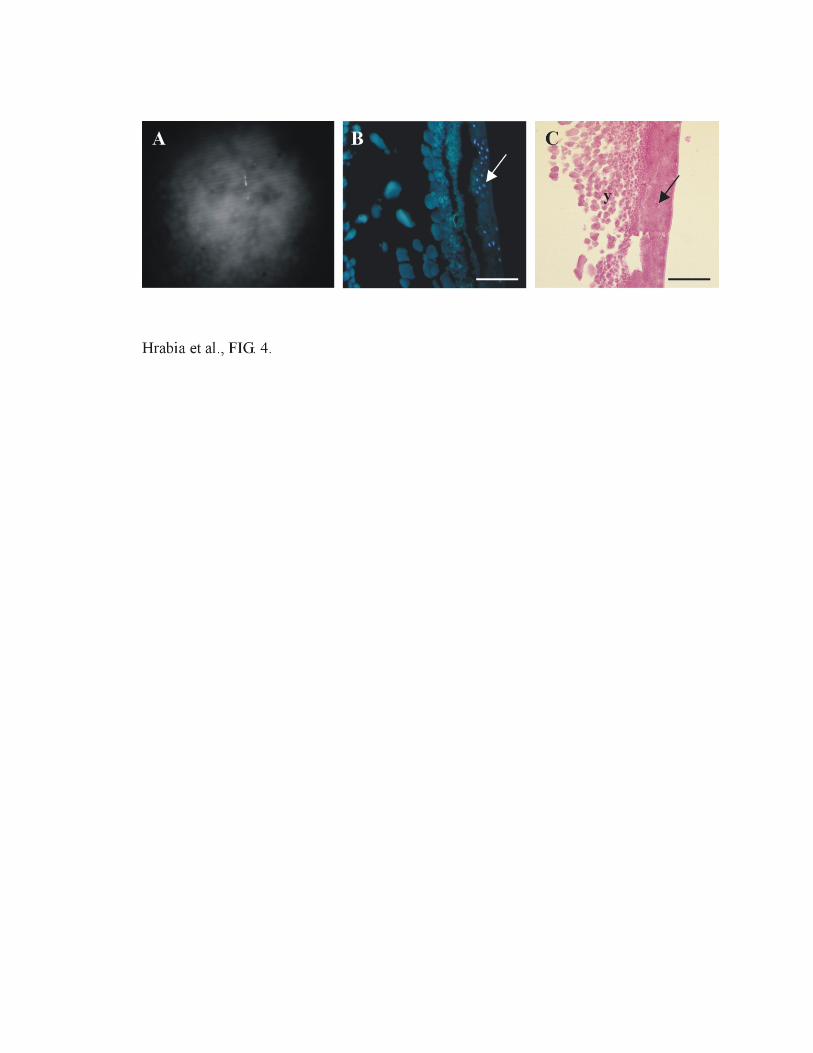
unfertilized eggs. Figure 4 depicts example of blastoderm after ICSI into spontaneously ovulated
oocyte. Blastoderms of the embryos developed from the ICSI of premature ovulated oocytes are
not shown. Table 1 presents fertilization and development of quail oocytes spontaneously and
prematurely ovulated followed by ICSI and culture for 24 h in DMEM. Five out of 30 (16.6%)
and 4 out of 30 (13.3%) oocytes injected with mature sperm were fertilized in the spontaneous
and induced ovulation groups, respectively. There was no clear difference in stages of embryo
development after ICSI between the two groups. Both groups showed maximum development at
stage VI.
Intracytoplasmic injection of testicular sperm and round and elongated spermatids
Figure 5 shows a representative blastoderm of the embryos from experiment in which a single
testicular sperm or spermatid was injected into an individual oocyte followed by culture for 24 h
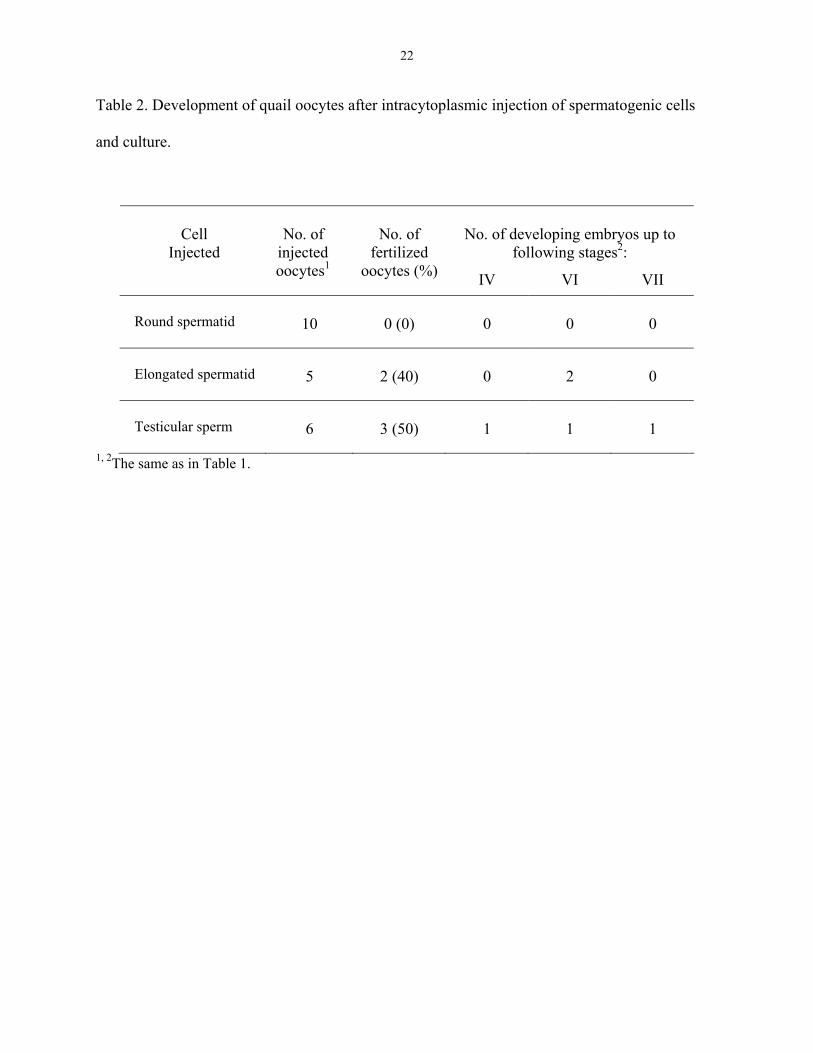
in DMEM. All data are summarized in Table 2. Half of the number (3/6) of oocytes injected with
testicular spermatozoa were fertilized and developed to stages IV-VII and 2 out of 5 oocytes

12
injected with elongated spermatids were fertilized and developed to stage VI. In contrast, all
oocytes injected with round spermatids were unfertilized.
DISCUSSION
To our knowledge, this is the first paper demonstrating by ICSI that a single sperm is capable
of activating the oocyte for further fertilization process and embryonic development in the quail.
The present study showed that the rate of fertilization after ICSI of spontaneously ovulated
oocytes, 16.6%, is comparable to that of Olszańska et al. [13] who demonstrated 15% of
efficiency after conventional IVF using 1-2 x 104 semen sperm of quail. The embryonic
development is also comparable between the two studies (stages II-VI 24 h after culture in the
present study vs stages IV-VI 18 h after culture in theirs). Higher rate of fertilization (75%) was
reported by Nakanishi et al. [6] when the development was observed with DAPI staining only 1-
4 h after culture in the conventional IVF in chicken oocytes. The conventional IVF may retain
more physiological conditions than ICSI, but the present study suggests that ICSI method is valid
to the same degree as the conventional IVF method. The ICSI omits some physiological steps
preceding the fertilization required in IVF such as sperm maturation or interaction of sperm
membrane with oocyte. Thus, this method simplifies the experimental conditions and provides
useful means to study mechanisms of fertilization and development in birds as in mammals. As
one of examples in this regard, this study provided evidence, that one sperm is capable of
activating an oocyte for fertilization in quail, although polyspermy naturally occurs in birds.
The present study also showed that oocytes collected from quail of prematurely induced-
ovulation are capable of fertilizing with a single mature sperm as those in spontaneous ovulation,
suggesting that oocytes are matured for fertilization several hours before the destined ovulation

13
time. Maturity of the ovarian follicle in birds has been characterized in terms of ovulability in
response to exogenous progesterone, luteinizing hormone or luteinizing hormone releasing
hormone injections [16, 17]. We propose the fertilizing ability of the oocyte as a new index for
follicle maturation.
Although both the conventional IVF [13] and ICSI in quail oocytes exhibited the formation of
embryonic blastoderms with a typical cleavage pattern of nucleated blastomeres at progressive
stages, the development was somehow delayed when compared to that in vivo conditions. As
demonstrated in the present study and by Stępinska and Olszańska [18], in natural conditions in
the oviduct quail embryos developed to stages III-IV 9-10 h, to stage V 12 h, to stage VI 13-14 h,
to stages VII-VIII 15-17 h and to stages X-XI 24 h after ovulation. Delay of embryo
development in our ICSI experiment may be attributable at least in a part, to high volume of PVP
injection as well as mechanical damage to the germinal disc due to injection. During this
manipulation the germinal disc was exposed to air, which in turn, may cause it to dryness.
However, these points cannot fully explain for the delay, because avian oocytes well tolerated
microsurgery as has been reported in the foreign DNA injection studies in quail [19] and chicken
[20, 21]. In these studies, already fertilized ova were microinjected with DNA, whereas the
unfertilized ova were used in the conventional IVF and in the present ICSI. Nevertheless, there is
room to improve the culture system for avian IVF.
Several embryos after ICSI showed a similar cleavage pattern to a “normal” one but these
embryos did not contain any nuclei or fragmented DNA bodies when determined by DAPI
staining method. Olszańska et al. [13] also reported this phenomenon in quail oocytes after IVF
and suggested that experimental procedures may have activated the egg, but fertilization did not
go to completion. The appearance of the external features of the embryo cannot be only criterion

14
as a base to estimate the early embryo development in culture. The examination of
presence/absence of the nuclei is important to determine true development of the quail embryo.
Kosin [22] observed degenerated nuclei in the germinal disc of the unfertilized eggs of chicken.
Some breeds of turkeys and strains of chicken exhibit limited parthenogenesis. In most cases this
spontaneous development without fertilization lead to very early degeneration. In a selected line
of turkeys a few percent of parthenogenetically derived embryos hatched and grew into adult
male [23, 24].
We also provided a new finding that a single elongated spermatid and immature sperm in the
testis can fertilize the oocyte in quail. In mammals, birth of normal offspring was shown by
direct injections of immature spermatozoa from the testis [25, 26]. The present study
demonstrates that injection of the testicular sperm or elongated spermatid into quail oocyte
resulted in the fertilization and the embryo development to stages VI-VII. The results clearly
indicate that these cells are competent to participate in fertilization and early embryonic
development. In contrast, the injection of round spermatid did not result in successful
fertilization and development. Kimura and Yanagimachi [25] reported that mouse round
spermatids could not activate the oocytes, unless the oocytes were activated artificially. The rate
of fertilization was improved by activating oocyte by electric current before injection of
spermatid nucleus. In quail, the reason for the failure of round spermatids is not clear but may
attributed to insufficient amounts of the oocyte-activating factor(s). Quail round spermatids may
need preactivation of quail oocytes similar to that in mammals.
To sum up, the present study demonstrates for the first time that (1) intracytoplasmic injection
of a single sperm isolated from semen into quail oocyte can activate the oocyte and lead to
fertilization, (2) using the employed ICSI and culture conditions, embryos can develop to

15
maximum stages of VI-VII, (3) spontaneously and prematurely ovulated oocytes possess similar
degree of fertilizing competency and (4) the testicular elongated spermatids and spermatozoa can
activate the oocyte like mature sperm in semen. Accordingly, microfertilization method in birds
has the potential to assist in the production of transgenic birds and protection of endangered
species of birds.

16
REFERENCES
1. Bakst MR, Howarth B. Hydrolysis of the hen’s perivitelline layer by cock sperm in vitro.
Biol Reprod 1977; 17:370-379.
2. Howarth B. Maturation of spermatozoa and mechanism of fertilization. In: Cunningham FJ,
Lake PE, Hewitt D (eds.), Reproductive Biology of Poultry. Oxford: British Poultry Science
Ltd; 1984:161-174.
3. Kuroki M, Mori M. Binding of spermatozoa to the perivitelline layer in the presence of a
protease inhibitor. Poult Sci 1997; 748-752.
4. Okamura F, Nishiyama H. Penetration of spermatozoa into the ovum and transformation of
the sperm nucleus into the male pronucleus in the domestic fowl, Gallus gallus. Cell Tissue
Res 1978; 497-508.
5. Perry MM. Nuclear events from fertilisation to the early cleavage in the domestic fowl
(Gallus domesticus). J Anat 1987; 150:99-109.
6. Nakanishi A, Utsumi K, Iritani A. Early nuclear events of in vitro fertilization in the
domestic fowl (Gallus domesticus). Mol Reprod Dev 1990; 26:217-221.
7. Waddington D, Gribbin C, Sterling RJ, Sang HM, Perry MM. Chronology of events in the
first cell cycle of the polyspermic egg of the domestic fowl (Gallus domesticus). Int J Dev
Biol 1998; 42:625-628.
8. Bramwell RK, Howarth B. Preferential attachment of cock spermatozoa to the perivitelline
layer directly over the germinal disc of the hen’s ovum. Biol Reprod 1992; 47:1113-1117.
9. Birkhead TR, Sheldon BC, Fletcher F. A comparative study of sperm-egg interaction in
birds. J Reprod Fert 1994; 101:353-361.

17
10. Howarth B Jr. An examination for sperm capacitation in the fowl. Biol Reprod 1971; 3:338-
341.
11. Nakanishi A, Miyake M, Utsumi K, Iritani A. Fertilizing competency of multiple ovulated
eggs in the domestic fowl (Gallus domesticus). Mol Reprod Dev 1991; 26:217-221.
12. Tanaka K, Wada T, Koga O, Nishio Y, Hertelendy F. Chick production by in vitro
fertilization of fowl ovum. J Reprod Fert 1994; 100:447-449.
13. Olszańska B, Stępińska U, Perry M. Development of embryos from in vitro ovulated and
fertilized oocytes of the quail (Coturnix coturnix japonica). J Exp Zool 2002; 292:580-586.
14. Woodard AE, Mather FB. The timing of ovulation, movement of the ovum through the
oviduct, pigmentation and shell deposition in Japanese quail (Coturnix coturnix japonica).
Poult Sci 1964; 43:1427-1432.
15. Eyal-Giladi H, Kochav S. From cleavage to primitive streak formation: a complementary
normal tables and a new look at the first stages of the development of the chick. I. General
morphology. Dev Biol 1976; 49:321-337.
16. Sekiguchi S, Imai K. Induction of ovulation by LH, progesterone and LHRH administered at
various times during the ovulatory cycle in Japanese quail. Japan Poultry Sci 1987; 24:79-85.
17. Onagbesan OM, Peddie MJ. Induction of ovulation and oviposition in female quail with
luteinizing hormone, luteinizing hormone releasing hormone, or progesterone. Gen Comp
Endocrinol 1988; 71:124-131.
18. Stępińska U, Olszańska B. Cell Multiplication and blastoderm development in relation to egg
envelope formation during uterine development of quail (Coturnix coturnix japonica)
embryo. J Exp Zool 1983; 228:505-510.

18
19. Mizuarai S, Ono K-I, Yamaguchi K, Nishijima K-I, Kamihira M, Iijima S. Production of
transgenic quails with high frequency of germ-line transmission using VSV-G pseudotyped
retroviral vetor. Biochem Biophys Res Commun 2001; 286:456-463.
20. Love J, Gribbin C, Mather Ch, Sang H. Transgenic birds by DNA microinjection.
Bio/Technology 1994; 12:60-63.
21. Naito M, Sasaki E, Ohtaki M, Sakurai M. Introduction of exogenous DNA into somatic and
germ cells of chicken by microinjection into germinal disc of fertilized ova. Mol Reprod Dev
1994; 37:167-171.
22. Kosin IL. Abortive parthenogenesis in the domestic chicken. Anat Rec 1945; 91:245-251.
23. Olsen MW. Frequency of parthenogenesis in chicken egg. J Hered 1966; 57:23-25.
24. Olsen MW. Frequency and cytological aspect of diploid parthenogenesis in turkey eggs.
Theoret Appl Genetics 1974; 44:216-221.
25. Kimura Y, Yanagimachi R. Mouse oocytes injected with testicular spermatozoa or round
spermatids can develop into normal offspring. Development 1995; 121:2397-2405.
26. Hewitson L, Martinovich C, Simerly C, Takahashi D, Schatten G. Rhesus offspring produced
by intracytoplasmic injection of testicular sperm and elongated spermatids. Fertil Steril 2002;
77:794-801.
FIGURE LEGENDS
FIG. 1. ICSI system for identification, isolation and injection of testicular spermatid and sperm
of quail. A - section of quail testis stained with HE. Small and large rectangles indicate areas for
magnification in B. Ba, b - left and right photos are magnification from A, respectively. Sperm

19
(sp); elongated spermatid (esd); round spermatid (rsd); spermatocyte (stc). C - quail sperm,
elongated spermatid, round spermatid and spermatocyte in PVP solution under Hoffman
modulation contrast microscope. Legends are the same as B. D - isolation of single sperm from
PVP solution by injection pipette. E - single sperm isolated in the pipette as indicated by arrow.
F - apparatus for sperm injection into oocyte in medium. G - close-up of ICSI into quail oocyte.
H - close-up of ICSI into the center of germinal disc.
FIG. 2. Quail blastoderms of in vivo fertilized, developing oocytes under stereomicroscope (A,
B, C) and after DAPI (D, E, F) and HE (G, H, I) staining. A, D, G - blastoderm at stage III. B, E,
H - blastoderm at stage V. C, F, I - blastoderm at stage VIII. a.o. - area opaca; a.p. - area
pellucida; c – cytoplasm; c.f. - cleavage furrows; y - yolk; arrows indicate the representative
nuclei. Bar = 0.1 mm
FIG. 3. Quail blastoderms of control group under stereomicroscope (A, B, C) and after DAPI (D,
E, F) and HE (G, H, I) staining. A, D, G - blastoderm of in vivo fertilized and developed oocyte
after oviposition at stage X. B, E, H - germinal disc of unfertilized egg after oviposition, with
segmentation of cytoplasm and numerous vacuoles at the periphery, absence of nuclei after
DAPI and HE staining in pseudo cells. C, F, I - sham-operated control, DAPI-stained irregular
fragmented DNA bodies. a.o.- area opaca; a.p. - area pellucida; c – cytoplasm; e – epiblast; h –
primary hypoblast; p.c. - pseudo cells; v – vacuoles; y – yolk; arrows indicate the presence of
nuclei, arrow heads indicate fragmented DNA bodies. Bar = 0.1 mm.

20
FIG. 4. Quail blastoderm at stage V after ICSI into spontaneously ovulated oocyte under
stereomicroscope (A) and after DAPI (B) and HE (C) staining. Arrows indicate representative
nuclei; y-yolk. Bar = 0.1 mm.
FIG. 5. Quail blastoderm at stage VI in oocyte injected with elongated spermatid under
stereomicroscope (A) and after DAPI (B) and HE (C) staining. Other explanation is the same as
in Fig. 4.

21
Table 1. Development of quail oocytes spontaneously and prematurely ovulated after
intracytoplasmic sperm injection and culture.
No. of developing embryos up to
following stages2:
Ovulation of the
oocyte
No. of
injected oocytes1
No. of
fertilized oocytes (%) II III IV V VI
Spontaneous
30
5 (16.6)
1
0
1
2
1
Induced (premature)
30
4 (13.3)
1
1
1
0
1
1Only one oocyte was collected from one female quail
2Developmental stages of embryos were estimated according to those fertilized and developed in vivo.
22
Table 2. Development of quail oocytes after intracytoplasmic injection of spermatogenic cells
and culture.
No. of developing embryos up to
following stages2:
Cell
Injected
No. of
injected oocytes1
No. of
fertilized oocytes (%) IV VI VII
Round spermatid
10
0 (0)
0
0
0
Elongated spermatid
5
2 (40)
0
2
0
Testicular sperm
6
3 (50)
1
1
1
1, 2The same as in Table 1.




Related Documents