The Journal of Immunology Evidence That the Erythrocyte Invasion Ligand PfRh2 is a Target of Protective Immunity against Plasmodium falciparum Malaria Linda Reiling,* Jack S. Richards,* ,† Freya J. I. Fowkes,* Alyssa E. Barry, ‡ Tony Triglia,* Watcharee Chokejindachai, x Pascal Michon, { Livingstone Tavul, { Peter M. Siba, { Alan F. Cowman,* Ivo Mueller, { and James G. Beeson* Abs targeting blood-stage Ags of Plasmodium falciparum are important in acquired immunity to malaria, but major targets remain unclear. The P. falciparum reticulocyte-binding homologs (PfRh) are key ligands used by merozoites during invasion of erythrocytes. PfRh2a and PfRh2b are functionally important members of this family and may be targets of protective immunity, but their potential role in human immunity has not been examined. We expressed eight recombinant proteins covering the entire PfRh2 common region, as well as PfRh2a- and PfRh2b-specific regions. Abs were measured among a cohort of 206 Papua New Guinean children who were followed prospectively for 6 mo for reinfection and malaria. At baseline, Abs were associated with increasing age and active infection. High levels of IgG to all PfRh2 protein constructs were strongly associated with protection from symptomatic malaria and high-density parasitemia. The predominant IgG subclasses were IgG1 and IgG3, with little IgG2 and IgG4 detected. To further understand the significance of PfRh2 as an immune target, we analyzed PfRh2 sequences and found that polymorphisms are concentrated in an N-terminal region of the protein and seem to be under diversifying selection, suggesting immune pressure. Cluster analysis arranged the sequences into two main groups, suggesting that many of the hap- lotypes identified may be antigenically similar. These findings provide evidence suggesting that PfRh2 is an important target of protective immunity in humans and that Abs act by controlling blood-stage parasitemia and support its potential for vaccine development. The Journal of Immunology, 2010, 185: 000–000. P lasmodium falciparum malaria remains a major global health problem, with ∼500 million cases and 1 million deaths annually (1, 2), and is a leading cause of death among children younger than 5 years of age (3). Protective immunity de- velops after repeated exposure and prevents severe disease and symptomatic episodes by control of blood-stage parasitemia (4). During blood-stage replication, P. falciparum merozoites invade erythrocytes, and Abs to merozoite Ags are believed to be im- portant in mediating acquired immunity and immunity generated by candidate blood-stage vaccines (5). However, the targets of protective human Abs are largely undefined. More than 40 mer- ozoite proteins have been identified that may play a role in in- vasion and/or represent potentially important targets of acquired immunity (6). However, very few of these have been studied in detail as targets of protective immunity against P. falciparum (7, 8). Abs to merozoite Ags are thought to act by inhibition of invasion, Ab-dependent cellular inhibition, and opsonization for phagocytosis or neutrophil-mediated killing (9–15). The P. falciparum reticulocyte-binding homologs (PfRh; also known as P. falciparum normocyte-binding proteins) are located in the rhoptries of merozoites and are thought to be released to bind erythrocyte receptors for invasion after initial interactions of the merozoite with the erythrocyte surface. The members of this protein family have a region of 500 aa that has identity with the P. vivax reticulocyte-binding proteins 1 and 2; these are thought to form a heterocomplex involved in the direct binding of retic- ulocytes (16). In P. falciparum, PfRh1, PfRh2a, PfRh2b, PfRh4, and PfRh5 are expressed members of this family, whereas PfRh3 seems to be a pseudogene (17–28). PfRh2a and PfRh2b are identical for the first 2776 aa (88%) of the protein. After a breakpoint, the sequence diverges into PfRh2a- and PfRh2b- specific sequences (19). Substantial evidence suggests the PfRh proteins play important roles in erythrocyte invasion, although their precise functions are yet to be elucidated. PfRh1, PfRh4, and PfRh5 were shown to bind erythrocytes, and Abs generated in *Walter and Eliza Hall Institute of Medical Research; † Department of Medical Bi- ology, University of Melbourne; ‡ Center for Population Health, Burnet Institute, Melbourne, Victoria, Australia; x Faculty of Tropical Medicine, Mahidol University, Rajthewe, Bangkok, Thailand; and { Papua New Guinea Institute of Medical Re- search, Madang, Papua New Guinea Received for publication May 10, 2010. Accepted for publication September 8, 2010. This work was supported by the National Health and Medical Research Council of Australia (Project Grant and Career Development Award to J.B., Postgraduate Re- search Scholarship to J.R., Training Award to F.F., Project Grant to A.B., Program Grant to A.C., Infrastructure for Research Institutes Support Scheme Grant 361646); the Miller Fellowship of the Walter and Eliza Hall Institute (to J.B.); an Austra- lian Research Council Future Fellowship (to J.B.); an Innovation Fellowship from the Victorian Endowment for Science Knowledge and Innovation (to A.B.); the Australia–India Strategic Research Fund of the Australian Government; a Victorian State Government Operational Infrastructure Support grant; and the International Nutrition Foundation/Ellison Medical Foundation Fellowship (to W.C.). The sequences presented in this article have been submitted to GenBank (http://www. ncbi.nlm.nih.gov/) under accession numbers HM802474–HM802488. Address correspondence and reprint requests to Dr. James Beeson, Walter and Eliza Hall Institute of Medical Research, 1G Royal Parade, Parkville, Victoria 3050, Aus- tralia. E-mail address: [email protected] The online version of this article contains supplemental material. Abbreviations used in this paper: aHR, adjusted hazard ratio; AMA1, apical mem- brane Ag 1; CI, confidence interval; d n , number of nonsynonymous changes per nonsynonymous site; d s , number of synonymous changes per synonymous site; EBA, erythrocyte binding Ag; fl-, full length; HR, hazard ratio; HvL, high versus low levels of Abs; IQR, interquartile range; K, number of clusters; MSP, merozoite surface protein; MvL, medium versus low levels of Abs; PfRh, Plasmodium falcipa- rum reticulocyte-binding homolog; PNG, Papua New Guinea; SA, sialic acid; SNP, single nucleotide polymorphism; TM, transmembrane domain. Copyright Ó 2010 by The American Association of Immunologists, Inc. 0022-1767/10/$16.00 www.jimmunol.org/cgi/doi/10.4049/jimmunol.1001555 Published October 20, 2010, doi:10.4049/jimmunol.1001555 on November 2, 2010 www.jimmunol.org Downloaded from

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

The Journal of Immunology
Evidence That the Erythrocyte Invasion Ligand PfRh2 isa Target of Protective Immunity against Plasmodiumfalciparum Malaria
Linda Reiling,* Jack S. Richards,*,† Freya J. I. Fowkes,* Alyssa E. Barry,‡ Tony Triglia,*
Watcharee Chokejindachai,x Pascal Michon,{ Livingstone Tavul,{ Peter M. Siba,{
Alan F. Cowman,* Ivo Mueller,{ and James G. Beeson*
Abs targeting blood-stage Ags of Plasmodium falciparum are important in acquired immunity to malaria, but major targets
remain unclear. The P. falciparum reticulocyte-binding homologs (PfRh) are key ligands used by merozoites during invasion of
erythrocytes. PfRh2a and PfRh2b are functionally important members of this family and may be targets of protective immunity,
but their potential role in human immunity has not been examined. We expressed eight recombinant proteins covering the entire
PfRh2 common region, as well as PfRh2a- and PfRh2b-specific regions. Abs were measured among a cohort of 206 Papua New
Guinean children who were followed prospectively for 6 mo for reinfection and malaria. At baseline, Abs were associated with
increasing age and active infection. High levels of IgG to all PfRh2 protein constructs were strongly associated with protection
from symptomatic malaria and high-density parasitemia. The predominant IgG subclasses were IgG1 and IgG3, with little IgG2
and IgG4 detected. To further understand the significance of PfRh2 as an immune target, we analyzed PfRh2 sequences and
found that polymorphisms are concentrated in an N-terminal region of the protein and seem to be under diversifying selection,
suggesting immune pressure. Cluster analysis arranged the sequences into two main groups, suggesting that many of the hap-
lotypes identified may be antigenically similar. These findings provide evidence suggesting that PfRh2 is an important target
of protective immunity in humans and that Abs act by controlling blood-stage parasitemia and support its potential for
vaccine development. The Journal of Immunology, 2010, 185: 000–000.
Plasmodium falciparum malaria remains a major globalhealth problem, with ∼500 million cases and 1 milliondeaths annually (1, 2), and is a leading cause of death among
children younger than 5 years of age (3). Protective immunity de-
velops after repeated exposure and prevents severe disease andsymptomatic episodes by control of blood-stage parasitemia (4).During blood-stage replication, P. falciparum merozoites invade
erythrocytes, and Abs to merozoite Ags are believed to be im-
portant in mediating acquired immunity and immunity generated
by candidate blood-stage vaccines (5). However, the targets of
protective human Abs are largely undefined. More than 40 mer-
ozoite proteins have been identified that may play a role in in-
vasion and/or represent potentially important targets of acquired
immunity (6). However, very few of these have been studied in
detail as targets of protective immunity against P. falciparum
(7, 8). Abs to merozoite Ags are thought to act by inhibition of
invasion, Ab-dependent cellular inhibition, and opsonization for
phagocytosis or neutrophil-mediated killing (9–15).The P. falciparum reticulocyte-binding homologs (PfRh; also
known as P. falciparum normocyte-binding proteins) are located
in the rhoptries of merozoites and are thought to be released to
bind erythrocyte receptors for invasion after initial interactions of
the merozoite with the erythrocyte surface. The members of this
protein family have a region of 500 aa that has identity with the
P. vivax reticulocyte-binding proteins 1 and 2; these are thought
to form a heterocomplex involved in the direct binding of retic-
ulocytes (16). In P. falciparum, PfRh1, PfRh2a, PfRh2b, PfRh4,
and PfRh5 are expressed members of this family, whereas PfRh3
seems to be a pseudogene (17–28). PfRh2a and PfRh2b are
identical for the first 2776 aa (88%) of the protein. After
a breakpoint, the sequence diverges into PfRh2a- and PfRh2b-
specific sequences (19). Substantial evidence suggests the PfRh
proteins play important roles in erythrocyte invasion, although
their precise functions are yet to be elucidated. PfRh1, PfRh4, and
PfRh5 were shown to bind erythrocytes, and Abs generated in
*Walter and Eliza Hall Institute of Medical Research; †Department of Medical Bi-ology, University of Melbourne; ‡Center for Population Health, Burnet Institute,Melbourne, Victoria, Australia; xFaculty of Tropical Medicine, Mahidol University,Rajthewe, Bangkok, Thailand; and {Papua New Guinea Institute of Medical Re-search, Madang, Papua New Guinea
Received for publication May 10, 2010. Accepted for publication September 8, 2010.
This work was supported by the National Health and Medical Research Council ofAustralia (Project Grant and Career Development Award to J.B., Postgraduate Re-search Scholarship to J.R., Training Award to F.F., Project Grant to A.B., ProgramGrant to A.C., Infrastructure for Research Institutes Support Scheme Grant 361646);the Miller Fellowship of the Walter and Eliza Hall Institute (to J.B.); an Austra-lian Research Council Future Fellowship (to J.B.); an Innovation Fellowship fromthe Victorian Endowment for Science Knowledge and Innovation (to A.B.); theAustralia–India Strategic Research Fund of the Australian Government; a VictorianState Government Operational Infrastructure Support grant; and the InternationalNutrition Foundation/Ellison Medical Foundation Fellowship (to W.C.).
The sequences presented in this article have been submitted to GenBank (http://www.ncbi.nlm.nih.gov/) under accession numbers HM802474–HM802488.
Address correspondence and reprint requests to Dr. James Beeson, Walter and ElizaHall Institute of Medical Research, 1G Royal Parade, Parkville, Victoria 3050, Aus-tralia. E-mail address: [email protected]
The online version of this article contains supplemental material.
Abbreviations used in this paper: aHR, adjusted hazard ratio; AMA1, apical mem-brane Ag 1; CI, confidence interval; dn, number of nonsynonymous changes pernonsynonymous site; ds, number of synonymous changes per synonymous site;EBA, erythrocyte binding Ag; fl-, full length; HR, hazard ratio; HvL, high versuslow levels of Abs; IQR, interquartile range; K, number of clusters; MSP, merozoitesurface protein; MvL, medium versus low levels of Abs; PfRh, Plasmodium falcipa-rum reticulocyte-binding homolog; PNG, Papua New Guinea; SA, sialic acid; SNP,single nucleotide polymorphism; TM, transmembrane domain.
Copyright� 2010 by The American Association of Immunologists, Inc. 0022-1767/10/$16.00
www.jimmunol.org/cgi/doi/10.4049/jimmunol.1001555
Published October 20, 2010, doi:10.4049/jimmunol.1001555 on N
ovember 2, 2010
ww
w.jim
munol.org
Dow
nloaded from

experimental animals to PfRh1, PfRh2, PfRh4, and PfRh5 caninhibit invasion (22, 24–27). Furthermore, genetic disruption ofspecific members alters the erythrocyte receptor preference ofmerozoites during invasion (6, 13).Merozoites can use different pathways for erythrocyte invasion
mediated by variation in the expression and/or use of erythro-cyte binding Ags (EBAs) and PfRh proteins (6, 13). Invasion phe-notypes can be broadly classified into two main groups: sialicacid (SA)-dependent invasion, demonstrated by poor invasion ofneuraminidase-treated erythrocytes (neuraminidase cleaves SAon the erythrocyte surface) and SA-independent invasion, demon-strated by efficient invasion of neuraminidase-treated erythro-cytes. SA-dependent (neuraminidase-sensitive) invasion involvesthe EBAs and PfRh1. PfRh2 and PfRh4 are important in SA-independent invasion (27–29); however, receptors for these li-gands are unknown. Recent studies showed that the use of differentinvasion pathways through variation in EBA and PfRh utilizationmediates evasion of human invasion inhibitory Abs (13).The important role of the PfRh proteins in invasion, as well as
the ability of Abs raised against PfRh ligands to inhibit invasion,suggests that they could be important targets of protective im-munity and may be suitable for vaccine development. However,very little is known about immune responses to PfRh proteins. Aninitial study in Kenya reported that Abs to PfRh2 and PfRh4 wereacquired in an age-dependent manner, reflecting the acquisition ofimmunity in the population (13). Differential inhibition by humanAbs of P. falciparum isolates that varied in their use of PfRhproteins pointed to this ligand family as a potentially importanttarget of inhibitory Abs (13). Although the expression of PfRhproteins may vary, PfRh2 seems to be expressed in most isolates,although the expression of gene transcripts was reported to vary(27, 30–33).In this study, we examined the acquisition of Abs to PfRh2 and
their association with protective immunity. We expressed multiplerecombinant proteins covering the entire ectodomain of PfRh2 andmeasured Abs in a longitudinal cohort study of children exposed toP. falciparum. The study design enabled prospective examinationof Abs in relation to the risk for reinfection, symptomatic malaria,and high-density parasitemia. IgG subclasses were assessed as onemeasure of potential Ab function. To further investigate whetherPfRh2 may be an important target of protective immune respon-ses, we performed sequence analysis to determine whether there isevidence of diversifying selection in the pfrh2 gene.
Materials and MethodsProtein expression
Eight recombinant proteins of PfRh2a/b were expressed in Escherichia coli.Proteins were named according to the starting amino acid in the full-lengthprotein sequence. The PfRh2-297 fragment (i.e., commencing at aminoacid 297) was amplified by PCR from a codon-optimized 3D7 PfRh2 genesegment. All other PfRh2a and PfRh2b gene fragments were amplifiedby PCR from 3D7 genomic DNA (European Molecular Biology Labora-tory nucleotide sequence database [http://www.ebi.ac.uk/embl/] accessionnumbers: AY138496 [P. falciparum normocyte-binding protein 2a gene(3D7)] and AY138500 [P. falciparum normocyte-binding protein 2b gene(3D7)]). The following oligonucleotide primers were used (restriction sitesare denoted by lower case letters): PfRh2-34: 59-CAATCAAGTggatcc-CATGGAGCATCTTCAG-39, 59-CTAATAGCTCTTTctcgagttaTAAGTA-TAAATCAATAGGTGT-39; PfRh2-297: 59-agctggatcccGAAAGCTATGT-GATGAAC-39, 59-agctctcgagttaGCTGGTGTTCAGAATGG-39; PfRh2-673:59-aggaacgatcatcatttgaatggatccaaaatacatg-39, 59-CATCTATTATATTTTG-TTGTTCTGActcgagttaATATAAGTT-39; PfRh2-1288: 59-ATAAATGATT-TTggatccGAAAAGAATATATCACAAG-39, 59-CTTCATAAAAAAAAG-ActcgagttaTTTATCTCCATT-39; PfRh2a: 59-agctggatcccCACATAAAAA-GTAAACTAGAATC-39, 59-AgctctcgagttaTGATCGAGAAAAATTTCTA-TC-39; and PfRh2b: 59-AGAAATATCCAAGAAggatccGAGCAAAAAA-AG-39, 59-CTCCAGCATTATATACctcgagttaCATTTTGTTATG-39.
Gene fragments for PfRh2a/b-34, -673, and -1288 and PfRh2b weresubcloned into pPROEX HTb (Life Technologies, Sydney, Australia).Proteins were expressed as N-terminal (His)6 fusion proteins in E. coli andaffinity purified on NiNTA columns (Invitrogen, Melbourne, Australia),following the manufacturer’s instructions. Insoluble protein (PfRh2-34 andPfRh2-673) was refolded by rapid dilution (0.1 M Tris [pH 8], 0.4 M L-Arginine, 0.2 mM PMSF, 0.5 mM glutathione disulfide, 5 mM reducedglutathione). Residual insoluble protein was precipitated by centrifugation(10,000 3 g, 20 min 4˚C). All proteins were dialyzed against PBS. Genefragments for PfRh2-297 and PfRh2a were subcloned into pET45b andwere expressed as (His)6 fusion proteins. Purified PfRh2-297 was refoldedby dilution in 2 M urea/20 mM Tris (pH 8)/100 mM NaCl. n-Octyl glu-coside (Sigma-Aldrich, St. Louis, MO) detergent (20 mM) was added todissociate the fusion protein from a contaminating protein. The resultingprecipitate contained relatively pure fusion protein and was resuspended inPBS/2 M urea for use in ELISA assays. PfRh2a was expressed as a solublefusion protein, purified on a NiNTA column, and further purified bysize fractionation. PfRh2-2030 and PfRh2-2530 [previously published asPfRh2A9 and PfRh2A11 (19)], were expressed as GST fusion proteinsin E. coli and purified on glutathione agarose (Sigma-Aldrich, Sydney,Australia), following the manufacturer’s instructions, and then dialyzedovernight against PBS. All recombinant proteins were run in reducing andnonreducing sample buffer in SDS-PAGE gels (Supplemental Fig. 1) tocheck for purity and integrity. Coomassie-stained protein bands were cutout and evaluated by mass spectrometry to verify their identity. PfRh2-1288 occurred as two clear bands on reducing and nonreducing gels; bothbands were labeled by anti-His tag Abs in Western blots, and analysis bymass spectrometry confirmed that both bands were PfRh2 recombinantproteins. Based on their Mr, the lower molecular mass species seems tohave resulted from truncation at the C terminus.
Study population and ethics statement
Details of the cohort were described elsewhere (34). Briefly, plasmasamples were obtained from a prospective treatment reinfection study of206 children from Madang Province on the north coast of Papua NewGuinea (PNG). The median age at enrollment was 9.3 y (range, 5–14 y;interquartile range, 8.1–10.3 y). Baseline clinical observations and venoussampling were performed. The prevalence of P. falciparum and P. vivax,detected by PCR, at enrollment was 67.5 and 33.9%, respectively. Allchildren received artesunate orally for 7 d to clear parasitemia. Over thesubsequent 6 mo, the cohort was followed by active surveillance every 2wk, including finger-prick blood collection for parasitemia and passivecase detection. Merozoite surface protein (MSP)2-based genotyping wasused to distinguish between reinfection and treatment failure. A clinicalepisode of P. falciparum malaria was defined as the presence of fever andparasitemia .5000/ml, but all children with fever and any parasitemia bylight microscopy were treated with antimalarials. Plasma samples used forthis study were those taken at enrollment, prior to treatment with anti-malarials. Sera for positive controls were obtained from adults within theMadang area. Sera for negative controls were obtained from anonymousAustralian blood donors who were malaria naive.
Ethics approval was obtained from the Medical Research AdvisoryCommittee, PNG, and the Walter and Eliza Hall Institute of MedicalResearch Ethics Committee. Written informed consent was obtained fromall study participants or their guardians.
ELISA
Abs to recombinant proteins were measured by ELISAs using establishedmethods (13, 35). Briefly, Ags were coated onto Maxisorp microtiter plates(Nunc, Roskilde, Denmark) at 1–2 mg/ml (schizont lysate at 4.6 mg/ml).Sera samples were tested at 1/500 for total IgG. Polyclonal goat anti-human IgG HRP-conjugated Ab (Chemicon, Melbourne, Australia) wasused at 1/5000. For measurement of IgG subclasses, secondary Abs wereadded at a dilution of 1/1000 using monoclonal anti-human IgG1, IgG2,IgG3, and IgG4 (Zymed, Melbourne, Australia). We used an HRP-conjugated sheep anti-mouse Ab (Chemicon, Melbourne, Australia) at 1/2500 as a tertiary Ab for IgG subclass detection. ABTS liquid substrate(Sigma, Castle Hill, Australia) was used to measure enzymatic reactivity,and the reaction was stopped with 1% SDS. OD was determined at 405 nm.Nine nonimmune serum donors were used as negative controls, and serafrom three malaria-exposed adults were used as positive controls for val-idation and standardization purposes. All sera were tested in duplicate,which was repeated if there was a discrepancy .25% between duplicates.PfRh2-2030 and PfRh2-2530 were expressed as GST-fusion proteins.Therefore, reactivity to GST alone was assessed and deducted from allvalues for these Ags; however, reactivity to GST was very low or negli-gible among samples. In addition, we tested for IgG to the NANP repeats
2 PfRh2 AS A TARGET OF IMMUNITY TO MALARIA
on Novem
ber 2, 2010 w
ww
.jimm
unol.orgD
ownloaded from

of circumsporozoite protein. A sample was classified as Ab+ if the OD wasgreater than the mean plus three SDs of the reactivity observed withnonexposed samples. Data on IgG to MSP1–19, MSP2, and apical mem-brane Ag 1 (AMA1) were determined previously (35) and included in thisstudy for comparison with PfRh2.
Statistical analysis
Statistical analysis was performed using STATA 9.2 (STATACorp, CollegeStation, TX). Differences in seroprevalence and IgG levels between cate-gorical variables were assessed using x2 tests or Kruskal–Wallis tests,respectively. Correlations between Ab responses were examined usingSpearman’s rho. To determine the association between Ab levels andP. falciparum infection and symptomatic malaria, OD values for each Agwere stratified into three equal groups (tertiles) reflecting low, medium,and high responders. Kaplan–Meier curves were generated for time-to-event (infection or clinical episode) data and compared using the Wil-coxon test. Assumptions of proportional hazards were assessed, and Coxproportional hazards models were used to calculate hazard ratios andevaluate the effects of possible confounders. For Ab variables that showednonproportional hazards, results from Cox regression including an in-teraction term between Ab variable and time are reported. Hazard ratios forAb variables were adjusted by multivariate Cox regression using pre-defined covariates of age (binary variable: ,9 y, $9 y) and location ofresidence, which were reported previously (34, 35). To understand theextent of differences in relative IgG levels to PfRh2 among protectedversus malaria-susceptible children, they were stratified into three groupsaccording to their history of symptomatic malaria episodes in the 6-moperiod after drug treatment, and median Ab levels between groups werecompared. For this analysis, we only included children who had recordedexposure to P. falciparum during follow-up, as determined by a detectableparasitemia by PCR or light microscopy of any density; this included 95%of children. Children who had no detected parasitemia during follow-upwere regarded as not exposed and were excluded from this analysis.Children were classified as “protected” if they had no recorded symp-tomatic P. falciparum malaria and low-grade parasitemia only (parasitemia, 5000 parasites/ml), “susceptible 1” if they had one symptomatic episode(and a parasitemia .5000 parasites/ml), and “susceptible 2” if they two ormore symptomatic episodes.
Sequence analysis
P. falciparum genomic DNA was extracted from the cell pellet of bloodsamples from children in the cohort using the QIAamp 96 DNA Bloodkit (34). A 1662-bp fragment was amplified in a PCR reaction with thefollowing oligonucleotides: Rh2_1aF: 59-AATGAAGAACAATGTTTAG-TTGGTGGGAAAACAG-39 andRh2_1R: 59-TTTCTTTAAGACTTAATA-GATGACTTAATTCAG-39.
PCR fragments were cloned into pCRII-TOPO (TOPO TA Cloning,Invitrogen) and used to transform E. coli (TOP10, Invitrogen). Forward-and reverse-sequencing reactions for several clones were done by the se-quencing service of the Applied Genetic Diagnostics Unit, Department ofPathology, University of Melbourne. Single nucleotide polymorphismswere identified by visual inspection of high-quality raw chromatogramdata. SNPs were considered valid if they were observed in multiple clonesfrom the same isolate and found in more than one isolate or verified bysequencing a second independent PCR product from the same isolate.
Sequences were submitted to GenBank (http://www.ncbi.nlm.nih.gov/)under new accession numbers HM802474–HM802488.
For comparison with the PNG sequences, we downloaded the 3D7, 7G8,and Dd2 reference sequences (36) from the public database, as well as thosefrom an additional 12 isolates from worldwide origins being sequencedin genome projects at the Broad (U.S.) and Sanger (U.K.) Institutes. BasicLocal Alignment Search Tool homology searching identified PfRh2 se-quences for Senegal, RO33 and Ghana clinical (Ghana), RAJ116 and ICH-CR14 (India), HB3 (Honduras), IT (Brazil), SL (El Salvador), K1 (Thai-land), FCC-2 (China), and FVO and VS/1 (both from Vietnam). Only par-tial sequences covering the 1600-bp region sequenced were available forisolates K1 and FCC-2. Sequence data for the IT clone and the Ghana fieldisolate were produced by the pathogen genomics group at the WellcomeTrust Sanger Institute and are available at http://www.sanger.ac.uk/Projects/P_falciparum/. Sequences for all other P. falciparum laboratory clones weregenerated by the P. falciparum sequencing project at the Broad Institute ofHarvard and Massachusetts Institute of Technology (http://www.broad.mit.edu) and are available at http://www.broadinstitute.org/annotation/genome/plasmodium_falciparum_spp/Blast.html. Sequences of the reference lines3D7, Dd2, FVO, and 7G8 were downloaded from the European Molecu-lar Biological Laboratory nucleotide database (http://www.ebi.ac.uk/embl/;
accession numbers AY138500, AY138502, AY138503, and AY138501, re-spectively). We also included sequences that we generated from recentlaboratory-adapted isolates from Thailand [HCS3 (37); http://www.ncbi.nlm.nih.gov/; new GenBank accession number is HM802473] and West Africa[Pf2004 and Pf2006 (38); http://www.ncbi.nlm.nih.gov/; new GenBank ac-cession numbers are HM802472 and HM802471, respectively], giving a totalof 18 sequences from diverse geographic origins.
Tajima’s D test was used to test for departure from selective neutralityby comparing S (number of segregating sites) and p (the average pairwisenucleotide diversity). Balancing selection increases p compared with S;thus, the value for Tajima’s D becomes positive. Additionally, Fu and Li’stest was used to test for selective neutrality. It compares hs, the number ofsingletons, with h, the total number of mutations, expressed as Dp, or hs
and p (the average number of nucleotide differences between pairs ofsequences), expressed as Fp. The average evolutionary divergence over allsequence pairs was estimated using the Tamura three-parameter model,which corrects for multiple hits and takes into account differences intransitional and transversional rates and guanine and cytosine contentbias. Codon-based tests for neutrality or positive selection were performedusing the Nei–Gojobori method (39). All tests were performed usingDNASp5.00.04 (40) or MEGA v4 (41). Cluster analysis of amino acidhaplotypes was done using Structure v. 2.2 software (42, 43). Structure 2.2assigns individual multilocus haplotypes probabilistically to a user-definednumber of clusters (K) (43). For PfRh2 amino acid haplotypes, Structurewas run 20 times for K = 1–10 for 10,000 Monte Carlo Markov Chainiterations after a burn-in period of 10,000 (44) using the admixture modeland correlated allele frequencies. The estimated value of K was defined bycalculating DK, according to the method of Evanno et al. (44). Ancestrycoefficients were then plotted for each PfRh2 sequence.
ResultsAcquisition of Abs to PfRh2
To determine the acquisition of Abs to PfRh2, we expressedconsecutive regions of nearly the entire PfRh2a and PfRh2bectodomain (Fig. 1). In total, we expressed eight proteins: fiveproteins covered the common region shared by the full-length (fl-)PfRh2a and PfRh2b (referred to as PfRh2-34, PfRh2-297, PfRh2-673, PfRh2-1288, and PfRh2-2030; numbers indicate startingamino acid); one protein covered the last 246 aa of the commonregion and the first 256 aa of the PfRh2a-specific region (PfRh2a-2530); and proteins representing each of the PfRh2a-specific andPfRh2b-specific sequences (referred to as PfRh2a and PfRh2bthroughout this paper). Analysis of total IgG among the cohortsamples showed that all proteins were recognized by the majorityof serum samples (Table I), whereas there was little reactivityamong nonexposed Melbourne residents. Seroprevalence rangedfrom 66.5% for PfRh2-673 to 94.2% for PfRh2-2030 and PfRh2-2530. Median OD was highest for PfRh2-2030, PfRh2-2530,and PfRh2a (Table I). Among the entire cohort, 123 individuals(60.6%) were seropositive for all PhRh2 proteins tested, and only2 individuals (0.99%) were seronegative to all PfRh2 proteins.IgG levels were significantly higher in children who had PCR-
detectable parasitemia at the time of sample collection for allPfRh2 proteins tested (Table I; p# 0.001). Despite a restricted agerange in the cohort (median age, 9.3 y; range: 5–14 y), higher IgGlevels or higher prevalence of Abs were observed among olderchildren (.9 y of age) compared with younger children (#9 y ofage) for all proteins, which was significant for PfRh2-34, PfRh2-1288, PfRh2-2030, PfRh2-2530, and PfRh2b (Table I). IgG levelsto all rPfRh2 proteins were positively correlated with IgG to P.falciparum schizont protein extract, used as a marker of exposureto blood-stage infection (p # 0.001; Table II). This observation,together with the acquisition of Abs with age, higher IgG levels inparasite-positive children, and lack of reactivity among samplesfrom nonexposed donors, suggest that the responses measured aremalaria specific.There was a modest to strong correlation between Abs against
a particular protein and Abs against the adjacent proteins (Table II).
The Journal of Immunology 3
on Novem
ber 2, 2010 w
ww
.jimm
unol.orgD
ownloaded from

The highest correlation was found between Abs against PfRh2-2030 and PfRh2-2530 (Spearman’s rho = 0.844; p , 0.001).Correlations between Abs against N-terminal proteins PfRh2-34and PfRh-297 were less strong but were still significant (Spear-man’s rho = 0.470; p , 0.001).
Abs to PfRh2 are associated with reduced risk for malaria andhigh-density parasitemia
The design and longitudinal nature of the cohort study, incor-porating initial antimalarial treatment and active follow-up withthe application of sensitive molecular methods to detect P. falci-parum infections, enabled us to investigate the relationship be-tween levels of PfRh2-specific Abs (high, medium, and low) andreinfection, parasitemia of different densities, and symptomaticmalaria in the same cohort of children.Comparing children with the highest versus lowest Ab levels
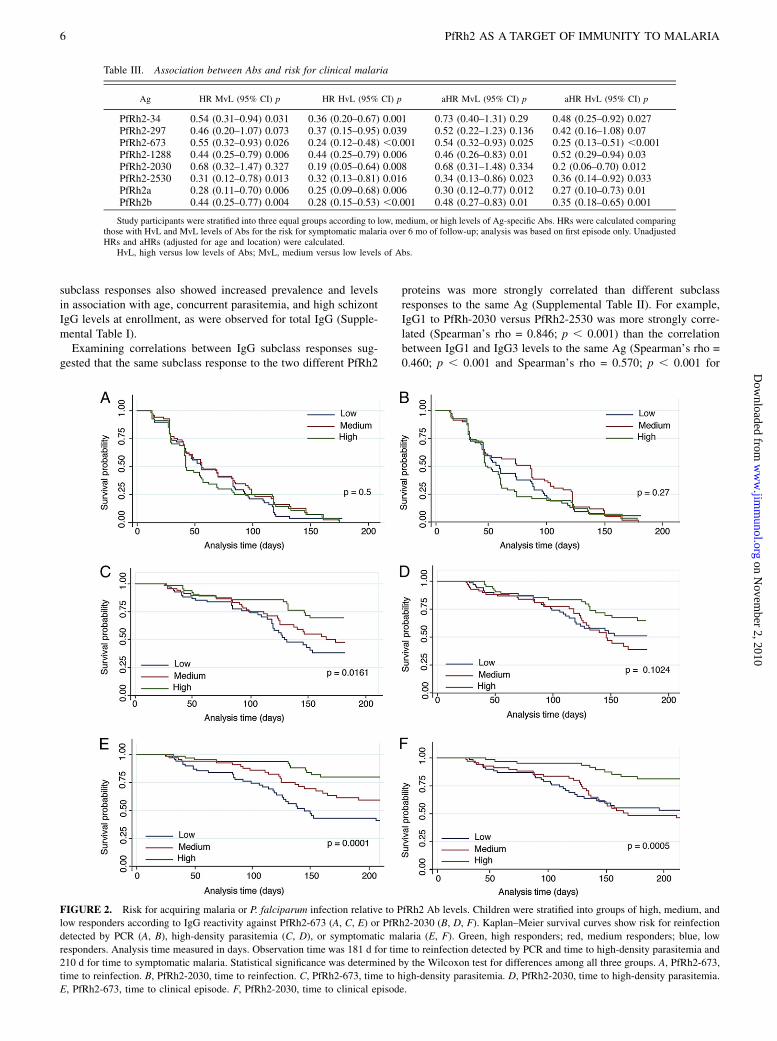
showed that IgG against PfRh2 was associated with a significantlydecreased risk for symptomatic malaria (defined as parasitemia.5000 parasites/ml plus fever) for all eight PfRh2 proteins (TableIII). Even after adjustments for age and spatial confounders, highIgG responses to all PfRh2 proteins showed a .50% reduction inrisk for symptomatic malaria (Table III). Protective associationswere strongest for PfRh2-673, PfRh2-2030, and PfRh2a (adjustedhazard ratio [aHR], 0.25, p, 0.001; aHR, 0.2, p = 0.012; and aHR,0.27, p = 0.01, respectively). Of note, aHRs for all PfRh2 proteinswere lower than or comparable to those for 3D7-AMA1 (aHR, 0.39;95% confidence interval [CI]: 0.143–1.089; p = 0.073), 3D7-MSP1-19 (aHR, 0.60; 95% CI: 0.240–1.505; p = 0.276), MSP2-3D7 (aHR,0.56; 95% CI: 0.211–1.489; p = 0.246), and MSP2-FC27 (aHR,0.72; 95% CI: 0.263–1.994; p = 0.532), which are leading blood-stage vaccine candidates. Furthermore, Abs to the pre-erythrocyticAg circumsporozoite protein or the control protein GST were notassociated with risk for malaria (data not shown).We also examined associations between Abs and reinfection with
a parasitemia of any density (as detected by PCR or light micro-scopy) or episodes of moderate (.500 parasites/ml) or high-density(.5000 parasites/ml) parasitemia. High Ab levels to all PfRh2proteins were significantly associated (p , 0.05) with protectionagainst high-density parasitemia (.5000 parasites/ml), except forPfRh2-297, which was of borderline significance (hazard ratio[HR], 0.51; p = 0.065). Abs to PfRh2-673 (HR, 0.39; p = 0.002),
PfRh2-2030 (HR, 0.44; p = 0.023), and PfRh2-34 (HR, 0.45; p =0.005) showed the strongest association with protection from high-density parasitemia. After adjusting for covariates, a high level ofIgG to PfRh2-673 (aHR, 0.42; p = 0.005) and PfRh2-2030 (aHR,0.46; p = 0.034) remained strongly predictive of protection againsthigh-density parasitemia, whereas associations for PfRh2-2530(aHR, 0.53; p = 0.051) and PfRh2a (aHR 0.52; p = 0.056) were ofborderline significance, and the aHRs for PfRh2-34, PfRh2-297,PfRh2-1288, and PfRh2b were no longer significant. No signifi-cant associations were observed between high levels of Abs againstany PfRh2 protein and protection from parasitemia detected byPCR or light microscopy (p . 0.2) or against moderate-densityparasitemia (.500/ml; p . 0.19). Fig. 2 shows representativeexamples of Kaplan–Meier survival analyses for two of the PfRh2proteins (PfRh2-673 and PfRh2-2030) that showed the strongestassociation with protection against clinical malaria. These dem-onstrate the protective effect of Abs against clinical disease (Fig.2E, 2F) and high-density parasitemia (Fig. 2C, 2D) but not againstreinfection per se (Fig. 2A, 2B).
To further understand the extent of differences in relative IgGlevels to PfRh2 among protected versus malaria-susceptible chil-dren, we stratified children into three groups according to theirhistory of symptomatic malaria episodes in the follow-up pe-riod. Children were classified as “protected,” “susceptible 1” (onesymptomatic episode), and “susceptible 2” (at least two symp-tomatic episodes). IgG levels were highest among the protectedgroup of children compared with susceptible children (Table IV).The difference in median Ab values was 1.3–2.5-fold higher forPfRh2a and PfRh2-673, respectively, in protected children com-pared with susceptible group 2.
IgG subclass responses to PfRh2 are predominantly IgG1 andIgG3
Prior studies suggested that IgG1 and IgG3 are the predominantsubclasses that respond to merozoite Ags (35, 45, 46, 47); however,to the best of our knowledge, no study has examined IgG subclassresponses to any PfRh protein. IgG1 and IgG3 are cytophilic Abswith a high potential to activate complement and with high affinityfor FcgRs, thus having the ability to activate T cell-mediated cy-totoxicity and Ab-dependent phagocytosis. To gain insights intothe potential function of Abs to PfRh2, we evaluated IgG sub-
FIGURE 1. Full-length and recombinant PfRh2 proteins. Recombinant proteins expressed in E. coli are shown in relation to their position within the full-
length PfRh2a and PfRh2b (fl-PfRh2a and fl-PfRh2b) protein sequences. Numbers represent amino acid positions in the PfRh2 sequence. Amino acid 2276
indicates the position where the PfRh2a-and PfRh2b-specific sequences commence; fl-PfRh2a and fl-PfRh2b share the same sequence from aa 1–2276. The
500-aa region (1701–2221) that has identity with P. vivax reticulocyte-binding protein 2 and Py235 is indicated. Recombinant proteins are shown below the
fl-PfRh2 proteins. PfRh2a and PfRh2a refer to the specific regions of the fl-PfRh2a and fl-PfRh2b proteins. Recombinant proteins corresponding to the
common region are named according to the start amino acid (e.g., PfRh2–34). TM, transmembrane domain.
4 PfRh2 AS A TARGET OF IMMUNITY TO MALARIA
on Novem
ber 2, 2010 w
ww
.jimm
unol.orgD
ownloaded from

classes to two representative proteins: PfRh2-2030 and PfRh2-2530. We observed predominantly IgG1 and IgG3 subclass re-sponses to PfRh2, with very little IgG2 and IgG4 detected (Sup-
plemental Table I). The seroprevalence of IgG1 against PfRh2-2030 and PfRh2-2530 was higher compared with IgG3 (85 ver-sus 63.1% for PfRh2-2030; 85 versus 78.6% for PfRh2-2530). IgG
Table I. IgG seroprevalence and levels for PfRh2 Ags
AgeaEnrollment P. falciparum
Parasitemic Status
AgAll Individuals
(n = 206)#9.0 y(n = 91)
.9.0 y(n = 115) p Valueb
PCR2(n = 67)
PCR+(n = 139) p Valueb
PfRh2-34Seropositivec 171
83.0%68
74.7%103
89.6%0.005 46
68.7%125
89.9%,0.001
Median OD(IQR)d
0.2525(0.087–0.490)
0.169(0.061–0.383)
0.313(0.126–0.602)
0.0026 0.117(0.037–0.382)
0.293(0.135–0.559)
,0.001
PfRh2-297Seropositivec 161
78.2%67
73.6%94
81.7%0.162 45
67.2%116
83.5%0.008
Median OD(IQR)d
0.242(0.064–0.550)
0.242(0.052–0.526)
0.237(0.075–0.626)
0.3387 0.128(0.042–0.373)
0.316(0.111–0.715)
,0.001
PfRh2-673Seropositivec 137
66.5%58
63.7%79
68.7%0.454 35
52.2%102
73.4%,0.001
Median OD(IQR)d
0.402(0.156–0.863)
0.324(0.109–0.748)
0.46(0.180–0.925)
0.082 0.236(0.074–0.648)
0.522(0.224–0.917)
,0.001
PfRh2-1288Seropositivec 190
92.2%78
85.7%112
97.4%0.002 57
85.1%133
95.7%0.008
Median OD(IQR)d
0.584(0.266–1.015)
0.484(0.108–0.856)
0.675(0.335–1.168)
0.003 0.389(0.098–0.803)
0.675(0.344–1.050)
,0.001
PfRh2-2030Seropositivec 194
94.2%80
87.9%114
99.1%0.001 59
88.1%135
97.1%0.009
Median OD(IQR)d
1.111(0.587–1.557)
1.137(0.461–1.557)
1.064(0.609–1.560)
0.768 0.772(0.188–1.403)
1.111(0.707–1.647)
,0.001
PfRh2-2530Seropositivec 194
94.2%8189%
11398.3%
0.005 5988.1%
13597.1%
0.009
Median OD(IQR)d
2.087(1.390–2.352)
2.089(0.883–2.323)
2.084(1.534–2.374)
0.227 1.832(0.446–2.196)
2.189(1.626–2.369)
0.001
PfRh2ae
Seropositivec 19194.1%
8291.1%
10996.5%
0.108 5887.9%
13397.1%
0.009
Median OD(IQR)d
1.256(0.775–1.477)
1.261(0.656–1.493)
1.256(0.825–1.449)
0.889 1.062(0.290–1.322)
1.347(0.966–1.512)
,0.001
PfRh2bSeropositivec 175
85.0%72
79.1%103
89.6%0.037 47
70.2%128
92.1%,0.001
Median OD(IQR)d
0.610(0.367–0.977)
0.513(0.268–0.806)
0.745(0.453–1.053)
,0.001 0.457(0.208–0.675)
0.759(0.453–0.998)
,0.001
a#9.0 y indicates individuals younger than 9 years of age; .9.0 y indicates individuals older than 9 years of age.bThe p values were calculated using the x2 test for comparison of proportions or the Wilcoxon rank-sum test for comparison of medians.cSeropositive (number of seropositive individuals) and percentage (%) of seropositive individuals were defined by IgG reactivity that was higher than
the mean plus three SDs of control sera (unexposed donors) measured by ELISA. Mean OD of nine Melbourne control samples for each run were 0.0212(PfRh2-34), 0.0205 (PfRh2-297), 0.0701 (PfRh2-673), 0.0169 (PfRh2-1288), 0.0135 (PfRh2-2030), 0.0313 (PfRh2-2530), 0.049 (PfRh2a) and 0.0710 forPfRhb.
dMedian ODs are displayed.eBecause of insufficient samples, the number of samples tested for IgG to Rh2a was 90 (,9 y), 113 (.9 y), 66 (PCR2), and 137 (PCR+).PCR2, P. falciparum not detected by PCR; PCR+, P. falciparum detected by PCR.
Table II. Correlations between IgG responses to PfRh2 proteins and schizont protein extract
Ag PfRh2-34 PfRh2-297 PfRh2-673 PfRh2-1288 PfRh2-2030 PfRh2-2530 PfRh 2a PfRh 2b
PfRh2-297 0.470PfRh2-673 0.604 0.706PfRh2-1288 0.658 0.693 0.827PfRh2-2030 0.528 0.633 0.758 0.780PfRh2-2530 0.548 0.662 0.762 0.789 0.844PfRh2a 0.498 0.645 0.745 0.723 0.772 0.808PfRh2b 0.529 0.410 0.534 0.555 0.407 0.463 0.406Schizont extract 0.662 0.535 0.695 0.685 0.604 0.656 0.546 0.665
Correlation coefficients are Spearman’s rho. All correlations are significant (p # 0.001).
The Journal of Immunology 5
on Novem
ber 2, 2010 w
ww
.jimm
unol.orgD
ownloaded from
subclass responses also showed increased prevalence and levelsin association with age, concurrent parasitemia, and high schizontIgG levels at enrollment, as were observed for total IgG (Supple-mental Table I).Examining correlations between IgG subclass responses sug-
gested that the same subclass response to the two different PfRh2
proteins was more strongly correlated than different subclassresponses to the same Ag (Supplemental Table II). For example,IgG1 to PfRh-2030 versus PfRh2-2530 was more strongly corre-lated (Spearman’s rho = 0.846; p , 0.001) than the correlationbetween IgG1 and IgG3 levels to the same Ag (Spearman’s rho =0.460; p , 0.001 and Spearman’s rho = 0.570; p , 0.001 for
Table III. Association between Abs and risk for clinical malaria
Ag HR MvL (95% CI) p HR HvL (95% CI) p aHR MvL (95% CI) p aHR HvL (95% CI) p
PfRh2-34 0.54 (0.31–0.94) 0.031 0.36 (0.20–0.67) 0.001 0.73 (0.40–1.31) 0.29 0.48 (0.25–0.92) 0.027PfRh2-297 0.46 (0.20–1.07) 0.073 0.37 (0.15–0.95) 0.039 0.52 (0.22–1.23) 0.136 0.42 (0.16–1.08) 0.07PfRh2-673 0.55 (0.32–0.93) 0.026 0.24 (0.12–0.48) ,0.001 0.54 (0.32–0.93) 0.025 0.25 (0.13–0.51) ,0.001PfRh2-1288 0.44 (0.25–0.79) 0.006 0.44 (0.25–0.79) 0.006 0.46 (0.26–0.83) 0.01 0.52 (0.29–0.94) 0.03PfRh2-2030 0.68 (0.32–1.47) 0.327 0.19 (0.05–0.64) 0.008 0.68 (0.31–1.48) 0.334 0.2 (0.06–0.70) 0.012PfRh2-2530 0.31 (0.12–0.78) 0.013 0.32 (0.13–0.81) 0.016 0.34 (0.13–0.86) 0.023 0.36 (0.14–0.92) 0.033PfRh2a 0.28 (0.11–0.70) 0.006 0.25 (0.09–0.68) 0.006 0.30 (0.12–0.77) 0.012 0.27 (0.10–0.73) 0.01PfRh2b 0.44 (0.25–0.77) 0.004 0.28 (0.15–0.53) ,0.001 0.48 (0.27–0.83) 0.01 0.35 (0.18–0.65) 0.001
Study participants were stratified into three equal groups according to low, medium, or high levels of Ag-specific Abs. HRs were calculated comparingthose with HvL and MvL levels of Abs for the risk for symptomatic malaria over 6 mo of follow-up; analysis was based on first episode only. UnadjustedHRs and aHRs (adjusted for age and location) were calculated.
HvL, high versus low levels of Abs; MvL, medium versus low levels of Abs.
FIGURE 2. Risk for acquiring malaria or P. falciparum infection relative to PfRh2 Ab levels. Children were stratified into groups of high, medium, and
low responders according to IgG reactivity against PfRh2-673 (A, C, E) or PfRh2-2030 (B, D, F). Kaplan–Meier survival curves show risk for reinfection
detected by PCR (A, B), high-density parasitemia (C, D), or symptomatic malaria (E, F). Green, high responders; red, medium responders; blue, low
responders. Analysis time measured in days. Observation time was 181 d for time to reinfection detected by PCR and time to high-density parasitemia and
210 d for time to symptomatic malaria. Statistical significance was determined by the Wilcoxon test for differences among all three groups. A, PfRh2-673,
time to reinfection. B, PfRh2-2030, time to reinfection. C, PfRh2-673, time to high-density parasitemia. D, PfRh2-2030, time to high-density parasitemia.
E, PfRh2-673, time to clinical episode. F, PfRh2-2030, time to clinical episode.
6 PfRh2 AS A TARGET OF IMMUNITY TO MALARIA
on Novem
ber 2, 2010 w
ww
.jimm
unol.orgD
ownloaded from

PfRh2-2030 and PfRh2-2530, respectively). The same was ob-served for the correlation between IgG3 responses to the differentPfRh2 Ags (Spearman’s rho = 0.725; p , 0.001). IgG2 and IgG4against each of the proteins were less strongly correlated (Spear-man’s rho = 0.429; p , 0.001 and Spearman’s rho = 0.487; p ,0.001, respectively). There was a moderate correlation betweenIgG2 and IgG4 against PfRh2-2030 (Spearman’s rho = 0.353; p ,0.001) as well as between IgG2 and IgG4 against PfRh2-2530(Spearman’s rho = 0.552; p , 0.001).
Associations between IgG subclass responses and protectiveimmunity
IgG1 and IgG3 against PfRh2-2030 and PfRh2-2530 were asso-ciated with a reduced risk for symptomatic malaria episodes whencomparing children with high versus low Ab levels (Table V). Thestrongest association was for IgG3 to PfRh2-2530 (HR = 0.21; p =0.005). Although the reduction in risk was greater for IgG3 (79%reduction) versus IgG1 (66% reduction) to PfRh2-2530, it was
similar for IgG1 and IgG3 to PfRh2-2030 (70 and 64% reduction,respectively). After adjustment for age and location, HRs for IgG1and IgG3 against PfRh2-2530 remained significantly reduced,whereas only IgG1 remained significant for PfRh2-2030. Therewas some evidence of a protective association for IgG2 againstPfRh2-2530 (aHR, 0.38; p = 0.064). However, because levels ofPfRh2-2530–specific IgG2 were very low (median OD, 0.022;interquartile range: 0.006–0.067, Supplemental Table I), it is un-clear whether this association is biologically significant. No sig-nificant associations were observed for IgG4. There is evidencethat high levels of IgG1 against PfRh2-2530 are also protectiveagainst high-density parasitemia (p = 0.008). High levels of IgG3showed an association with protection against high-density para-sitemia with borderline significance (p = 0.06). There were nosignificant associations between Abs of any IgG subclass and riskfor low-grade parasitemia or light microscopy- or PCR-detectableparasitemia (i.e., reinfection per se; data not shown).Ab levels among children who were classified as protected were
significantly higher for IgG1 and IgG3 against PfRh2-2530 as wellas for IgG3 against PfRh2-2030 compared with children who hadmalaria episodes; levels were increased 1.63–3.75-fold in protectedchildren compared with those who had two or more episodes(Supplemental Table III). No significant differences were foundfor IgG2 or IgG4. This further suggests that IgG1 and IgG3 are themain contributors to the protective effect of PfRh2-specific Abs.
Sequence analysis of the N-terminal region of PfRh2a/b
To obtain further evidence that PfRh2 may be a target of protectiveimmunity, we generated PfRh2 gene sequences from the P. fal-ciparum isolates derived from the participants of the study cohort.Previous sequence analysis of four reference isolates (3D7, 7G8,Dd2, FVO) that were available in the public database suggestedthat polymorphisms are concentrated in the N-terminal region ofPfRh2 and that this region might be under diversifying selectivepressure (36). To determine the number of SNPs and the rate andratio of synonymous versus nonsynonymous changes within thisregion, we amplified, cloned, and sequenced an N-terminal frag-ment of PfRh2 that corresponded to the most polymorphic regionbased on analysis of P. falciparum reference isolates. This se-quence is contained within the PfRh2-34, PfRh2-297, and PfRh2-673 recombinant proteins (Fig. 1).We obtained 15 PfRh2 sequences from genomic DNA samples
obtained from 12 infected individuals in the cohort, including fourdifferent sequences from one individual. Among these gene se-quences, we identified a total of 20 polymorphic sites; 19 werenonsynonymous. For comparison, we also obtained sequences from18 isolates from different worldwide locations. Among these, weidentified 28 nonsynonymous polymorphisms and 1 synonymouspolymorphism. Two of the nonsynonymous polymorphisms werecontained within the same codon, which resulted in coding for three
Table IV. Ab levels against PfRh2 proteins among protected versussusceptible children
Protein Groupa IgG Levelb p Valuec
PfRh2-34 Protected 0.33 (0.18–0.61) 0.0011Susceptible 1 0.22 (0.07–0.26)Susceptible 2 0.14 (0.05–0.37)
PfRh2-297 Protected 0.28 (0.07–0.60) 0.0944Susceptible 1 0.24 (0.07–0.49)Susceptible 2 0.13 (0.04–0.4)
PfRh2-673 Protected 0.58 (0.18–0.99) 0.0059Susceptible 1 0.25 (0.14–0.74)Susceptible 2 0.23 (0.07–0.49)
PfRh2-1288 Protected 0.67 (0.29–1.21) 0.0109Susceptible 1 0.51 (0.25–1.02)Susceptible 2 0.39 (0.09–0.71)
PfRh2-2030 Protected 1.26 (0.59–1.74) 0.0237Susceptible 1 1.11 (0.61–1.39)Susceptible 2 0.81 (0.29–1.31)
PfRh2-2530 Protected 2.2 (1.70–2.37) 0.0052Susceptible 1 1.96 (1.47–2.27)Susceptible 2 1.75 (0.59–2.29)
PfRh2a Protected 1.26 (0.82–1.5) 0.0398Susceptible 1 1.26 (0.77–1.47)Susceptible 2 0.97 (0.24–1.4)
PfRh2b Protected 0.74 (0.56–1.06) 0.0001Susceptible 1 0.52 (0.28–0.76)Susceptible 2 0.39 (0.22–0.67)
aThe cohort was divided into three groups according to whether individualsexperienced episodes of symptomatic malaria: protected, no recorded symptomaticP. falciparum malaria and low-grade parasitemia only (,5000 parasites/ml); suscep-tible 1, one episode of symptomatic malaria; and susceptible 2, two or more episodesof symptomatic malaria. Children who had no detected parasitemia during follow-up(5%) were regarded as not exposed and were excluded from this analysis.
bValues represent median OD (interquartile range).cThe p values were calculated using the Kruskall–Wallis test.
Table V. Association between IgG subclasses and risk for clinical malaria
Ag IgG Subclass HR MvL (95% CI) p HR HvL (95% CI) p aHR MvL (95% CI) p aHR HvL (95% CI) p
PfRh2-2030 IgG1 0.57 (0.25–1.28) 0.172 0.30 (0.11–0.82) 0.019 0.70 (0.31–1.60) 0.398 0.35 (0.13–0.97) 0.044PfRh2-2030 IgG2 0.73 (0.31–1.72) 0.475 0.65 (0.27–1.56) 0.333 0.66 (0.28–1.55) 0.345 0.69 (0.29–1.68) 0.417PfRh2-2030 IgG3 0.48 (0.09–1.13) 0.21 0.36 (0.03–0.92) 0.17 0.59 (0.25–1.41) 0.237 0.49 (0.19–1.30) 0.153PfRh2-2030 IgG4 1.18 (0.51–2.73) 0.698 0.75 (0.31–1.84) 0.534 0.95 (0.41–2.21) 0.898 0.71 (0.29–1.75) 0.46PfRh2-2530 IgG1 0.13 (0.04–0.44) 0.001 0.34 (0.14–0.79) 0.013 0.15 (0.04–0.52) 0.003 0.34 (0.14–0.82) 0.016PfRh2-2530 IgG2 0.71 (0.32–1.59) 0.407 0.35 (0.13–0.96) 0.041 0.67 (0.30–1.49) 0.323 0.38 (0.14–1.06) 0.064PfRh2-2530 IgG3 0.41 (0.18–0.95) 0.036 0.21 (0.07–0.62) 0.005 0.43 (0.19–1.01) 0.052 0.24 (0.08–0.72) 0.011PfRh2-2530 IgG4 1.14 (0.50–2.57) 0.761 0.64 (0.25–1.65) 0.356 1.17 (0.51–2.66) 0.711 0.63 (0.24–1.62) 0.337
Study participants were stratified into three equal groups based on low, medium, or high levels of Ag-specific Abs. HRs and aHRs were calculatedcomparing those with HvL and MvL Abs for the risk for symptomatic malaria over 6 mo of follow-up; analysis was based on the first episode only.
HvL, high versus low levels of Abs; MvL, medium versus low levels of Abs.
The Journal of Immunology 7
on Novem
ber 2, 2010 w
ww
.jimm
unol.orgD
ownloaded from

different amino acids. The synonymous site resulted from a thirdnucleotide variant at one position found only in two isolates (HCS3and RAJ116), in addition to the more common nonsynonymousvariant at the same site. Combining the PNG and worldwidedatasets, there were 35 nonsynonymous (located within 34 codons)and 2 synonymous polymorphisms; only 12 polymorphisms wereshared between the two sets of isolates, and all of these werenonsynonymous. Therefore, there were many polymorphismsunique to each dataset, suggesting that there may be more poly-morphisms still to be sampled for this gene. A multiple align-ment of the DNA sequences is shown in Supplemental Fig. 2. Bycontrast, the 1497-bp gene fragment coding for PfRh2-2030 fur-ther downstream within the PfRh2 gene showed markedly less di-versity compared with the N-terminal region with only one non-synonymous and one synonymous polymorphism among 10 iso-lates (RAJ116, K1, IGH-CR14, HB3, FVO, Dd2, 7G8, 3D7, IT,Ghana; data not shown).To investigate whether the pattern of polymorphisms in PfRh2
N-terminal coding region were consistent with immune selec-tion, we determined the number of nonsynonymous changes pernonsynonymous site (dn) and the number of synonymous changesper synonymous site (ds) among the 15 sequences from PNG. Wefound that v, the ratio between dn and ds, was positive (4.95),pointing toward diversifying selection (Table VI). Neither Fu andLi’s Dp and Fp statistics (0.89 and 1.021, respectively) nor Taji-ma’s D (1.021) reached significance, most likely as a result of thelimited sample size. Therefore, a more sensitive sliding windowanalysis was performed, which revealed highly positive values forTajima’s D and Fu and Li’s statistics at various window mid-points, suggesting a departure from neutrality in certain regions ofthe analyzed sequence (Fig. 3). We also compared dn and ds usinga codon-based test (Z-test of selection, using the Nei–Gojoborimethod, p-distance model), and found a significant departure fromneutrality (p = 0.014) in favor of positive selection (p = 0.008).Together, these findings point toward diversifying selective pres-sure acting on the N-terminal region of PfRh2 proteins in clinicalisolates among the cohort, consistent with it being a target of pro-tective immune responses.If the PfRh2 N-terminal domain is a target of immune selection,
we expect high levels of diversity to be found within the PNGparasite population, with important implications for immunity andits development as a vaccine candidate. Among the 15 PNG PfRh2sequences, there were 13 haplotypes, with only two pairs of PNGisolates sharing the same haplotype, showing an extremely highdegree of haplotype diversity for PfRh2 in this parasite popula-tion. Because there are a variable number of polymorphisms amongdifferent haplotypes (Supplemental Fig. 2), and this is likely to beassociated with the degree of antigenic diversity, we also performeda Bayesian cluster analysis on the combined dataset of 33 PfRh2sequences (28 haplotypes) to identify groups of similar haplotypes.The distribution of polymorphisms among sequences was mostconsistent with two major clusters, each with a broad geographicdistribution (Fig. 4). Both clusters were found in PNG, suggestingthat the full spectrum of diversity is found in this local sample.Importantly, several PNG haplotypes clustered with the 3D7 hap-lotype, showing that 3D7-like sequences are present within thePNG parasite population. Thus, the measurement of Abs using3D7-derived PfRh2 protein seems to be appropriate for this cohort.
DiscussionAn important and major finding of this study was that Abs to PfRh2were strongly associated with a reduced risk for symptomaticmalaria using a prospective study design. Remarkably, this asso-ciation was observed for all eight PfRh2 protein fragments, and T
able
VI.
Sequence
diversity
intheN-terminalregionofPfh
Na
Sites
bS
hhs
hHd(SD)
p(SD)c
d(SE)d
uTajim
a’sDe
FuandLi’sD*e
FuandLi’sF*e
PfRh2-(bp634-bp2233)
SNS
dn
ds
dn/d
s(v)
15
1600
20
20
313
0.981
(0.00095)
0.00477(0.00037)
0.00480(0.00107)
0.00384(0.00072)
0.990**
0.849**
1.021**
119
0.00564
0.00114
4.95
aOnly
sequencesfrom
MadangProvince,PNG
wereincluded
inthisanalysis.
bTotalnumber
ofbasepairs
per
sequence,excludingalignmentgaps.
cCalculatedusingJukes
andCantorcorrection.
dObtained
usingtheTam
ura
3param
eter
model
obtained
bybootstrap
procedure.
eCalculatedusingthetotalnumber
ofsegregatingsites.
ppp.
0.1
(notsignificant).
v,Ka/K
s;h,totalnumber
ofmutations;
hs,number
ofsingletons(m
utationsappearingonly
once
amongallsequences);p,averagepairw
isenucleotidediversity;u,expectednucleotidediversity
under
neutralityderived
from
thenumber
of
segregatingsites;
d,averageevolutionarydistance
over
allsequence
pairs;h,totalnumber
ofhaplotypes;Hd,diversity
ofhaplotypes;N,totalnumber
ofsequencesincluded
inanalysis;
S,totalnumber
ofsegregating(polymorphic)sites.
8 PfRh2 AS A TARGET OF IMMUNITY TO MALARIA
on Novem
ber 2, 2010 w
ww
.jimm
unol.orgD
ownloaded from

associations remained significant after adjusting for age and lo-cation for all proteins, except PfRh2-297, which was of borderlinesignificance. Furthermore, the strength of association betweenPfRh2 total IgG and protection from malaria was similar to orgreater than that for the leading vaccine candidates MSP1-19,MSP2, and AMA1. We extended our analysis by comparing Ablevels among children who did or did not have episodes of malariaduring the 6-mo follow-up period. Consistent with the survivalanalysis, Ab levels were significantly higher among protectedchildren than among those who had malaria episodes.An important question in understanding human immunity to
malaria is whether Abs to merozoite Ags contribute to protectionagainst parasitization per se or only act to prevent symptomaticmalaria. Our results demonstrate that Abs to PfRh2 are significantlyassociated with protection from high-density parasitemia andsymptomatic malaria but not against parasitization per se. Very fewstudies have examined associations of immune responses with riskfor reinfection and clinical malaria in the same study (7). To ad-dress these important questions in our study, we cleared para-sitemia among participants at enrollment with a highly effectivetreatment (.90% cure rate), actively screened for reparasitizationduring follow-up using sensitive molecular methods, and geno-typed infecting parasites to distinguish reinfection from treatmentfailure (34). No PfRh2 IgG or subclass response was associatedwith a reduced risk for reinfection as detected by PCR or lightmicroscopy. Instead, our results suggest that Abs to PfRh2 me-diate their protective effect by control of blood-stage parasitemia,thereby preventing high-density infections. High-density para-sitemia and overall parasite biomass are linked with the patho-
genesis of severe malaria (48). Therefore, these Abs may con-tribute to protection from severe malaria. Abs to PfRh2 would beexpected to function, in part, by inhibiting erythrocyte invasionand blood-stage parasite replication because Abs raised againstPfRh2 in laboratory animals have growth-inhibitory activityin vitro (27). Our findings also demonstrated antigenicity acrossthe entire PfRh2a and PfRh2b proteins. Abs to all PfRh2 proteinswere higher among those with active parasitemia at the time oftesting and were generally higher among older children, and Abscorrelated with Abs to schizont protein extract, used as a markerof exposure to blood-stage infection. In contrast, there was little orno reactivity among samples from nonexposed donors, and controlproteins, such as GST, showed little reactivity among cohortsamples. A single prior study showed age-associated acquisitionof Abs to PfRh2-2030 among a Kenyan population (13), but otherregions of PfRh2 were not studied.Analysis of IgG subclass responses revealed a predominance of
IgG1 and IgG3, as found for other Ags in this cohort and morebroadly (35, 46, 47, 49, 50). A significant age-associated increasein IgG1 and IgG3 levels and associations with parasitemia andexposure (reactivity to schizont extract) point toward malaria-specific responses. IgG1 and IgG3 against PfRh2-2030 andPfRh2-2530 were significantly associated with protection againstclinical malaria. Although IgG2 to PfRh2-2530 was associatedwith protection, the low levels of IgG2 detected suggest that thecontribution of these Abs to immunity is small. Examining cor-relations between IgG subclass responses revealed that the samesubclass response to the two different PfRh2 proteins was morestrongly correlated than were different subclass responses to thesame Ag. This suggests that subclass responses among individualsmay be related and that there is a tendency for individuals toproduce a similar IgG subclass response to different Ags (35). Thecytophilic nature of IgG1 and IgG3 suggests that Abs to PfRh2may interact with macrophages, granulocytes, and/or NK cells oractivate complement to mediate parasite clearance.Analysis of PfRh2 sequence diversity provided additional evi-
dence that PfRh2 is an important target of protective immunity.Prior analysis of four reference lines indicated that polymorphismsare predominantly located in an N-terminal region of the pro-tein (36). Our sequence analysis demonstrated significant poly-morphism in this region and suggested it is under diversifyingselection, presumably resulting from immune pressure. Manypolymorphic residues were common among isolates from differentgeographic regions, although some polymorphisms were restrictedto specific isolates and, thus, geographic locations. Codon-basedtests for neutrality and positive selection were in favor of di-versifying selection. Although values for Tajima’s D, Fu and Li’s
FIGURE 3. Sliding-window analysis of PfRh2 sequence poly-
morphisms. Analysis was performed with a window length of 100 bp and
a step size of 3. x-axis, Nucleotide position within gene fragment. Data
points represent midpoint values of sliding window. y-axis, Relative values
for Tajima’s D and Fu and Li’s D* and F*. Positive values indicate evi-
dence of diversifying selection.
FIGURE 4. Cluster analysis of PfRh2 sequences. Amino acid haplotypes for each of the PfRh2 N-terminal sequences were analyzed using Structure v2.2.
to identify the number of clusters (K) and the ancestry coefficients for each of the clusters. Individual PfRh2 sequences are represented by a vertical column,
which is partitioned into K-colored segments that represent that sequence’s ancestry within a particular cluster. This plot was derived from ancestry
coefficients obtained for K = 2, which was estimated for this dataset. Isolates shown from left to right: 3D7, IT, HB3, 7G8, SL, Senegal, RO33, Ghana,
Pf06.6 (isolate Pf2006), Pf04.7 (isolate Pf2004), RAJ116, IGH-CR14, VS/1, HCS3, FVO, Dd2, K1, and FCC-2, followed by the PNG sequences. Geo-
graphic origins for each of the isolates are indicated below the x-axis. The origins of 3D7 and IT are ambiguous.
The Journal of Immunology 9
on Novem
ber 2, 2010 w
ww
.jimm
unol.orgD
ownloaded from

D*, and Fu and Li’s F* were not significantly associated withdeparture from neutrality, the sliding-window analysis showedareas with elevated values. This is further supported by the excessof nonsynonymous changes compared with synonymous changesand a resulting high dn/ds ratio observed within all P. falciparumsequences that we analyzed in this study. The average pairwisenucleotide diversity (p = 0.00477) was greater than that reportedfor EBA175 region II [p = 0.003 (51); p = 0.004 (52)] but lessthan AMAI domains I, II, or III or the entire sequence [p = 0.025,0.006, 0.014, and 0.014, respectively (53)]. By contrast, region IIof EBA140 and EBA181 reportedly have limited diversity (p =0.001 and 0.0003, respectively) and no evidence for selection (51,52). Consistent with these findings, when we investigated how thesequence diversity was distributed, we found that the majority ofparasite isolates had unique PfRh2 haplotypes. The identificationof only two main sequence clusters shows that many of thesePfRh2 haplotypes may be antigenically similar, and both of thesewere found among the PNG PfRh2 sequences that we obtained.This pattern of high within-population diversity for PfRh2 in thisstudy was also found in several other merozoite Ags known to beimportant targets of host immunity (38, 54). Merozoite Ags seemto have little regional diversification, but diversification has beenobserved for other Ags (38, 54). Our analysis did not clarifywhether the observed changes occurred in PfRh2a or PfRh2bbecause the polymorphic region is contained within the commonregion of PfRh2. The 14 nonsynonymous changes described pre-viously in the reference lines were shown to be identical in PfRh2aand PfRh2b, and it was suggested that this might be due to con-tinual gene conversion (36). Therefore, we consider it safe toassume that the changes that we observed are the same in PfRh2aand PfRh2b.Our findings suggest that PfRh2 is an important target of pro-
tective immune responses and support PfRh2 as a potential vaccinecandidate. We found that the protein is naturally immunogenic, andAbs are associated with protection from malaria and high-densityparasitemia. Sequence diversity also supports PfRh2 as an im-portant target of immunity, and there is some evidence that thePfRh ligands may be targets of human inhibitory Abs (13). Im-portantly, Abs raised against the PfRh2-2030 protein in rabbitswere shown to inhibit invasion (27), and Abs to this region werestrongly associated with a reduced risk for malaria in our study. Asis the case for many P. falciparum merozoite Ags, there are noproteins that are similar to PfRh ligands in structure and functionin rodent malarias. Therefore, responses to PfRh ligands cannot beeasily studied in animal models, emphasizing the importance ofhuman studies to evaluate these Ags as targets of protective im-munity and to aid in the identification and prioritization of can-didates for vaccine development.
AcknowledgmentsWe thank all study participants and Papua New Guinea Institute of Medical
Research staff involved in the study and Danny Wilson and Jack Taraika for
assistance with sample processing.We thank Jake Baum and John Donelson
for helpful discussions regarding sequence analysis. We also thank the
Broad Institute and the Sanger Institute for permission to use the PfRh2
sequence data.
DisclosuresThe authors have no financial conflicts of interest.
References1. Snow, R. W., C. A. Guerra, A. M. Noor, H. Y. Myint, and S. I. Hay. 2005. The
global distribution of clinical episodes of Plasmodium falciparum malaria. Na-ture 434: 214–217.
2. World Health Organization. 2008. World malaria report. WHO Press, Geneva,Switzerland, p. vii–x.
3. Elliott, S. R., and J. G. Beeson. 2008. Estimating the burden of global mortalityin children aged ,5 years by pathogen-specific causes. Clin. Infect. Dis. 46:1794–1795.
4. Langhorne, J., F. M. Ndungu, A. M. Sponaas, and K. Marsh. 2008. Immunity tomalaria: more questions than answers. Nat. Immunol. 9: 725–732.
5. Beeson, J. G., F. H. Osier, and C. R. Engwerda. 2008. Recent insights into hu-moral and cellular immune responses against malaria. Trends Parasitol. 24: 578–584.
6. Cowman, A. F., and B. S. Crabb. 2006. Invasion of red blood cells by malariaparasites. Cell 124: 755–766.
7. Fowkes, F. J., J. S. Richards, J. A. Simpson, and J. G. Beeson. 2010. The re-lationship between anti-merozoite antibodies and incidence of Plasmodiumfalciparum malaria: A systematic review and meta-analysis. PLoS Med. 7:e1000218.
8. Richards, J. S., and J. Beeson. 2009. The future for blood-stage vaccines againstmalaria. Immunol. Cell Biol. 87: 377–390.
9. Cohen, S., G. A. Butcher, and R. B. Crandall. 1969. Action of malarial antibodyin vitro. Nature 223: 368–371.
10. McCallum, F. J., K. E. Persson, C. K. Mugyenyi, F. J. Fowkes, J. A. Simpson,J. S. Richards, T. N. Williams, K. Marsh, and J. G. Beeson. 2008. Acquisition ofgrowth-inhibitory antibodies against blood-stage Plasmodium falciparum. PLoSONE 3: e3571.
11. Egan, A. F., J. Morris, G. Barnish, S. Allen, B. M. Greenwood, D. C. Kaslow,A. A. Holder, and E. M. Riley. 1996. Clinical immunity to Plasmodium falci-parum malaria is associated with serum antibodies to the 19-kDa C-terminalfragment of the merozoite surface antigen, PfMSP-1. J. Infect. Dis. 173: 765–769.
12. Hodder, A. N., P. E. Crewther, and R. F. Anders. 2001. Specificity of the pro-tective antibody response to apical membrane antigen 1. Infect. Immun. 69:3286–3294.
13. Persson, K. E., F. J. McCallum, L. Reiling, N. A. Lister, J. Stubbs,A. F. Cowman, K. Marsh, and J. G. Beeson. 2008. Variation in use of erythrocyteinvasion pathways by Plasmodium falciparum mediates evasion of human in-hibitory antibodies. J. Clin. Invest. 118: 342–351.
14. Bouharoun-Tayoun, H., P. Attanath, A. Sabchareon, T. Chongsuphajaisiddhi, andP. Druilhe. 1990. Antibodies that protect humans against Plasmodium falciparumblood stages do not on their own inhibit parasite growth and invasion in vitro, butact in cooperation with monocytes. J. Exp. Med. 172: 1633–1641.
15. Kumaratilake, L. M., A. Ferrante, T. Jaeger, and C. M. Rzepczyk. 1992. Effectsof cytokines, complement, and antibody on the neutrophil respiratory burst andphagocytic response to Plasmodium falciparum merozoites. Infect. Immun. 60:3731–3738.
16. Galinski, M. R., C. C. Medina, P. Ingravallo, and J. W. Barnwell. 1992. Areticulocyte-binding protein complex of Plasmodium vivax merozoites. Cell 69:1213–1226.
17. Rayner, J. C., M. R. Galinski, P. Ingravallo, and J. W. Barnwell. 2000. TwoPlasmodium falciparum genes express merozoite proteins that are related toPlasmodium vivax and Plasmodium yoelii adhesive proteins involved in host cellselection and invasion. Proc. Natl. Acad. Sci. USA 97: 9648–9653.
18. Rayner, J. C., E. Vargas-Serrato, C. S. Huber, M. R. Galinski, and J. W. Barnwell.2001. A Plasmodium falciparum homologue of Plasmodium vivax reticulocytebinding protein (PvRBP1) defines a trypsin-resistant erythrocyte invasion path-way. J. Exp. Med. 194: 1571–1581.
19. Triglia, T., J. Thompson, S. R. Caruana, M. Delorenzi, T. Speed, andA. F. Cowman. 2001. Identification of proteins from Plasmodium falciparum thatare homologous to reticulocyte binding proteins in Plasmodium vivax. Infect.Immun. 69: 1084–1092.
20. Kaneko, O., J. Mu, T. Tsuboi, X. Su, and M. Torii. 2002. Gene structure andexpression of a Plasmodium falciparum 220-kDa protein homologous to thePlasmodium vivax reticulocyte binding proteins. Mol. Biochem. Parasitol. 121:275–278.
21. Rodriguez, M., S. Lustigman, E. Montero, Y. Oksov, C. A. Lobo, and D. Ojcius.2008. PfRH5: a novel reticulocyte-binding family homolog of Plasmodiumfalciparum that binds to the erythrocyte, and an investigation of its receptor.PLoS ONE 3: e3300.
22. Baum, J., L. Chen, J. Healer, S. Lopaticki, M. Boyle, T. Triglia, F. Ehlgen,S. A. Ralph, J. G. Beeson, and A. F. Cowman. 2009. Reticulocyte-bindingprotein homologue 5 - an essential adhesin involved in invasion of humanerythrocytes by Plasmodium falciparum. Int. J. Parasitol. 39: 371–380.
23. Taylor, H. M., T. Triglia, J. Thompson, M. Sajid, R. Fowler, M. E. Wickham,A. F. Cowman, and A. A. Holder. 2001. Plasmodium falciparum homologue ofthe genes for Plasmodium vivax and Plasmodium yoelii adhesive proteins, whichis transcribed but not translated. Infect. Immun. 69: 3635–3645.
24. Gao, X., K. P. Yeo, S. S. Aw, C. Kuss, J. K. Iyer, S. Genesan, R. Rajamanonmani,J. Lescar, Z. Bozdech, and P. R. Preiser. 2008. Antibodies targeting the PfRH1binding domain inhibit invasion of Plasmodium falciparum merozoites. PLoSPathog. 4: e1000104.
25. Gaur, D., S. Singh, S. Singh, L. Jiang, A. Diouf, and L. H. Miller. 2007.Recombinant Plasmodium falciparum reticulocyte homology protein 4 binds toerythrocytes and blocks invasion. Proc. Natl. Acad. Sci. USA 104: 17789–17794.
26. Tham, W. H., D. W. Wilson, L. Reiling, L. Chen, J. G. Beeson, andA. F. Cowman. 2009. Antibodies to reticulocyte binding protein-like homologue4 inhibit invasion of Plasmodium falciparum into human erythrocytes. Infect.Immun. 77: 2427–2435.
10 PfRh2 AS A TARGET OF IMMUNITY TO MALARIA
on Novem
ber 2, 2010 w
ww
.jimm
unol.orgD
ownloaded from

27. Duraisingh, M. T., T. Triglia, S. A. Ralph, J. C. Rayner, J. W. Barnwell,G. I. McFadden, and A. F. Cowman. 2003. Phenotypic variation of Plasmodiumfalciparum merozoite proteins directs receptor targeting for invasion of humanerythrocytes. EMBO J. 22: 1047–1057.
28. Stubbs, J., K. M. Simpson, T. Triglia, D. Plouffe, C. J. Tonkin, M. T. Duraisingh,A. G. Maier, E. A. Winzeler, and A. F. Cowman. 2005. Molecular mechanism forswitching of P. falciparum invasion pathways into human erythrocytes. Science309: 1384–1387.
29. Gaur, D., T. Furuya, J. Mu, L. B. Jiang, X. Z. Su, and L. H. Miller. 2006. Up-regulation of expression of the reticulocyte homology gene 4 in the Plasmodiumfalciparum clone Dd2 is associated with a switch in the erythrocyte invasionpathway. Mol. Biochem. Parasitol. 145: 205–215.
30. Triglia, T., M. T. Duraisingh, R. T. Good, and A. F. Cowman. 2005.Reticulocyte-binding protein homologue 1 is required for sialic acid-dependentinvasion into human erythrocytes by Plasmodium falciparum. Mol. Microbiol.55: 162–174.
31. Jennings, C. V., A. D. Ahouidi, M. Zilversmit, A. K. Bei, J. Rayner, O. Sarr,O. Ndir, D. F. Wirth, S. Mboup, and M. T. Duraisingh. 2007. Molecular analysisof erythrocyte invasion in Plasmodium falciparum isolates from Senegal. Infect.Immun. 75: 3531–3538.
32. Gomez-Escobar, N., A. Amambua-Ngwa, M. Walther, J. Okebe, A. Ebonyi, andD. J. Conway. 2010. Erythrocyte invasion and merozoite ligand gene expressionin severe and mild Plasmodium falciparum malaria. J. Infect. Dis. 201: 444–452.
33. Taylor, H. M., M. Grainger, and A. A. Holder. 2002. Variation in the expressionof a Plasmodium falciparum protein family implicated in erythrocyte invasion.Infect. Immun. 70: 5779–5789.
34. Michon, P., J. L. Cole-Tobian, E. Dabod, S. Schoepflin, J. Igu, M. Susapu,N. Tarongka, P. A. Zimmerman, J. C. Reeder, J. G. Beeson, et al. 2007. The riskof malarial infections and disease in Papua New Guinean children. Am. J. Trop.Med. Hyg. 76: 997–1008.
35. Stanisic, D. I., J. S. Richards, F. J. McCallum, P. Michon, C. L. King, S. Schoepflin,P. R. Gilson, V. J. Murphy, R. F. Anders, I. Mueller, and J. G. Beeson. 2009.Immunoglobulin G subclass-specific responses against Plasmodium falciparummerozoite antigens are associated with control of parasitemia and protection fromsymptomatic illness. Infect. Immun. 77: 1165–1174.
36. Rayner, J. C., T. M. Tran, V. Corredor, C. S. Huber, J. W. Barnwell, andM. R. Galinski. 2005. Dramatic difference in diversity between Plasmodiumfalciparum and Plasmodium vivax reticulocyte binding-like genes. Am. J. Trop.Med. Hyg. 72: 666–674.
37. Beeson, J. G., and G. V. Brown. 2004. Plasmodium falciparum-infected eryth-rocytes demonstrate dual specificity for adhesion to hyaluronic acid and chon-droitin sulfate A and have distinct adhesive properties. J. Infect. Dis. 189: 169–179.
38. Hommel, M., S. R. Elliott, V. Soma, G. Kelly, F. J. Fowkes, J. M. Chesson,M. F. Duffy, J. Bockhorst, M. Avril, I. Mueller, et al. 2010. Evaluation of theantigenic diversity of placenta-binding Plasmodium falciparum variants and theantibody repertoire among pregnant women. Infect. Immun. 78: 1963–1978.
39. Nei, M., and T. Gojobori. 1986. Simple methods for estimating the numbers ofsynonymous and nonsynonymous nucleotide substitutions. Mol. Biol. Evol. 3:418–426.
40. Librado, P., and J. Rozas. 2009. DnaSP v5: a software for comprehensiveanalysis of DNA polymorphism data. Bioinformatics 25: 1451–1452.
41. Tamura, K., J. Dudley, M. Nei, and S. Kumar. 2007. MEGA4: Molecular Evo-lutionary Genetics Analysis (MEGA) software version 4.0. Mol. Biol. Evol. 24:1596–1599.
42. Duan, J., J. Mu, M. A. Thera, D. Joy, S. L. Kosakovsky Pond, D. Diemert,C. Long, H. Zhou, K. Miura, A. Ouattara, et al. 2008. Population structure of thegenes encoding the polymorphic Plasmodium falciparum apical membrane an-tigen 1: implications for vaccine design. Proc. Natl. Acad. Sci. USA 105: 7857–7862.
43. Pritchard, J. K., M. Stephens, and P. Donnelly. 2000. Inference of populationstructure using multilocus genotype data. Genetics 155: 945–959.
44. Evanno, G., S. Regnaut, and J. Goudet. 2005. Detecting the number of clusters ofindividuals using the software STRUCTURE: a simulation study. Mol. Ecol. 14:2611–2620.
45. Polley, S. D., D. J. Conway, D. R. Cavanagh, J. S. McBride, B. S. Lowe,T. N. Williams, T. W. Mwangi, and K. Marsh. 2006. High levels of serum anti-bodies to merozoite surface protein 2 of Plasmodium falciparum are associatedwith reduced risk of clinical malaria in coastal Kenya. Vaccine 24: 4233–4246.
46. Roussilhon, C., C. Oeuvray, C. Muller-Graf, A. Tall, C. Rogier, J. F. Trape,M. Theisen, A. Balde, J. L. Perignon, and P. Druilhe. 2007. Long-term clinicalprotection from falciparum malaria is strongly associated with IgG3 antibodiesto merozoite surface protein 3. PLoS Med. 4: e320.
47. Taylor, R. R., D. B. Smith, V. J. Robinson, J. S. McBride, and E. M. Riley. 1995.Human antibody response to Plasmodium falciparum merozoite surface protein2 is serogroup specific and predominantly of the immunoglobulin G3 subclass.Infect. Immun. 63: 4382–4388.
48. Gravenor, M. B., M. B. van Hensbroek, and D. Kwiatkowski. 1998. Estimatingsequestered parasite population dynamics in cerebral malaria. Proc. Natl. Acad.Sci. USA 95: 7620–7624.
49. Shi, Q., M. M. Lynch, M. Romero, and J. M. Burns, Jr. 2007. Enhanced pro-tection against malaria by a chimeric merozoite surface protein vaccine. Infect.Immun. 75: 1349–1358.
50. Metzger, W. G., D. M. Okenu, D. R. Cavanagh, J. V. Robinson, K. A. Bojang,H. A. Weiss, J. S. McBride, B. M. Greenwood, and D. J. Conway. 2003. SerumIgG3 to the Plasmodium falciparum merozoite surface protein 2 is stronglyassociated with a reduced prospective risk of malaria. Parasite Immunol. 25:307–312.
51. Baum, J., A. W. Thomas, and D. J. Conway. 2003. Evidence for diversifyingselection on erythrocyte-binding antigens of Plasmodium falciparum andP. vivax. Genetics 163: 1327–1336.
52. Verra, F., W. Chokejindachai, G. D. Weedall, S. D. Polley, T. W. Mwangi,K. Marsh, and D. J. Conway. 2006. Contrasting signatures of selection on thePlasmodium falciparum erythrocyte binding antigen gene family. Mol. Biochem.Parasitol. 149: 182–190.
53. Polley, S. D., W. Chokejindachai, and D. J. Conway. 2003. Allele frequency-based analyses robustly map sequence sites under balancing selection ina malaria vaccine candidate antigen. Genetics 165: 555–561.
54. Barry, A. E., L. Schultz, C. O. Buckee, and J. C. Reeder. 2009. Contrastingpopulation structures of the genes encoding ten leading vaccine-candidateantigens of the human malaria parasite, Plasmodium falciparum. PLoS ONE4: e8497.
The Journal of Immunology 11
on Novem
ber 2, 2010 w
ww
.jimm
unol.orgD
ownloaded from
Related Documents











