Molecular & Biochemical Parasitology 133 (2004) 1–14 Common trafficking pathway for variant antigens destined for the surface of the Plasmodium falciparum-infected erythrocyte Malin Haeggström a,b , Fred Kironde c , Klavs Berzins d , Qijun Chen a,b , Mats Wahlgren a,b,∗ , Victor Fernandez a,b a Microbiology and Tumorbiology Center, Karolinska Institutet, Nobels väg 16, Box 280, SE-171 77 Stockholm, Sweden b Swedish Institute for Infectious Disease Control, SE-171 82 Solna, Sweden c Department of Biochemistry, University of Makerere, Kampala, Uganda d Department of Immunology, Stockholm University, SE-106 91 Stockholm, Sweden Received 12 May 2003; received in revised form 22 July 2003; accepted 22 July 2003 Abstract Intraerythrocytic Plasmodium falciparum exports proteins to the cytosol and to the plasma membrane of the host cell. We here present data revealing the existence of a unique common pathway for the surface bound traffic of the clonally variant antigens, repeated-interspersed- antigen (RIFINS) and P. falciparum erythrocyte-membrane-protein-1 (PfEMP1). RIFIN- and PfEMP1-specific antibodies were found to stain single small vesicles (SSV) that bud off from the parasitophorus vacuolar membrane (PMV) at 6–10 h post-invasion. Large multimeric vesicle (LMV) assemblies, composed of subunits each of a similar size to that of a SSV, appeared as the dominant vesicle type carrying the variant antigens in the cytosol as the parasites developed into early trophozoite stages (≥16 h post-invasion). Later, more than 24 h post-invasion, large spinle-like vesicles (LSLV) built up as the LMV approached and accumulated underneath the erythrocyte membrane. LMV were found to associate both with the Maurer’s cleft antigen Pf332 and with lipids as seen by fluorescent BODIPY-Ceramide staining. Co-traffic of Pf332 with RIFINS and PfEMP1 occurred in sub-compartmentalized LMV, as the variant antigens co-localized at the outer rim while Pf332 occupied the core of the vesicle complex. Formation of LMV for the trafficking of RIFINS and PfEMP1 is a prominent feature of freshly isolated P. falciparum and of in vitro propagated K + as well as K − parasites, seemingly independent of the knob-associated histidine-rich protein (KAHRP). In vitro cultured 3D7 clones lack LMV formation and traffic the variant antigens in vesicles of a similar size to that of the SSV. © 2003 Elsevier B.V. All rights reserved. Keywords: Plasmodium falciparum; Protein trafficking; Variant surface antigens; RIFIN; PfEMP1; Pf332 1. Introduction During the asexual intraerythrocytic stage, as the Plas- modium falciparum parasite develops inside the para- sitophorous vacuole (PV), the composition and function of the infected erythrocyte (IE) cytosol and plasma membrane are extensively altered. Membranous structures such as the Maurer’s clefts and a tubovesicular network are formed Abbreviations: ATS, acidic-terminal-sequence; DBL, Duffy-binding- like; IE, infected erythrocyte; IEM, immunoelectron microscopy; IFL, im- munofluorescence; KAHRP, knob-associated histidine-rich protein; LMV, large multimeric vesicle; LSLV, large spindle-like vesicle; PfEMP1, Plasmodium falciparum erythrocyte membrane protein 1; PVM, para- sitophorous vacuole membrane; PV, parasitophorous vacuole; rif, repeti- tive interspersed family; SSV, small single vesicle ∗ Corresponding author. Tel.: +46-8-4572510; fax: +46-8-310525. E-mail address: [email protected] (M. Wahlgren). beyond the parasitophorous vacuole membrane (PVM) [1,2]. Several parasite-derived proteins are translocated and sorted to compartments in the IE cytosol and to different locations near or at the plasma membrane where they can be associated with the cytoskeleton as well as anchored in the lipid bilayer and displayed at the surface [3–5]. A num- ber of studies indicate that the traffic of parasite molecules through the IE cytosol is, at least in part, vesicle-mediated [6,7]. Homologues of proteins involved in trafficking path- ways, vesicle coating and fusion have been identified in the P. falciparum genome [8–11] but very little is known about the mechanisms involved in the transport of parasite proteins to the surface of the IE surface. Two groups of polypeptides are known to be expressed by P. falciparum at the IE surface. Both consist of anti- genically variant proteins encoded by large gene families. The best studied comprise the 200–400 kDa adhesins gener- 0166-6851/$ – see front matter © 2003 Elsevier B.V. All rights reserved. doi:10.1016/j.molbiopara.2003.07.006

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Molecular & Biochemical Parasitology 133 (2004) 1–14
Common trafficking pathway for variant antigens destined for thesurface of thePlasmodium falciparum-infected erythrocyte
Malin Haeggströma,b, Fred Kirondec, Klavs Berzinsd, Qijun Chena,b,Mats Wahlgrena,b,∗, Victor Fernandeza,b
a Microbiology and Tumorbiology Center, Karolinska Institutet, Nobels väg 16, Box 280, SE-171 77 Stockholm, Swedenb Swedish Institute for Infectious Disease Control, SE-171 82 Solna, Sweden
c Department of Biochemistry, University of Makerere, Kampala, Ugandad Department of Immunology, Stockholm University, SE-106 91 Stockholm, Sweden
Received 12 May 2003; received in revised form 22 July 2003; accepted 22 July 2003
Abstract
IntraerythrocyticPlasmodium falciparumexports proteins to the cytosol and to the plasma membrane of the host cell. We here presentdata revealing the existence of a unique common pathway for the surface bound traffic of the clonally variant antigens, repeated-interspersed-antigen (RIFINS) andP. falciparumerythrocyte-membrane-protein-1 (PfEMP1). RIFIN- and PfEMP1-specific antibodies were found tostain single small vesicles (SSV) that bud off from the parasitophorus vacuolar membrane (PMV) at 6–10 h post-invasion. Large multimericvesicle (LMV) assemblies, composed of subunits each of a similar size to that of a SSV, appeared as the dominant vesicle type carryingthe variant antigens in the cytosol as the parasites developed into early trophozoite stages (≥16 h post-invasion). Later, more than 24 hpost-invasion, large spinle-like vesicles (LSLV) built up as the LMV approached and accumulated underneath the erythrocyte membrane.LMV were found to associate both with the Maurer’s cleft antigen Pf332 and with lipids as seen by fluorescent BODIPY-Ceramidestaining. Co-traffic of Pf332 with RIFINS and PfEMP1 occurred in sub-compartmentalized LMV, as the variant antigens co-localized atthe outer rim while Pf332 occupied the core of the vesicle complex. Formation of LMV for the trafficking of RIFINS and PfEMP1 isa prominent feature of freshly isolatedP. falciparumand of in vitro propagated K+ as well as K− parasites, seemingly independent ofthe knob-associated histidine-rich protein (KAHRP). In vitro cultured 3D7 clones lack LMV formation and traffic the variant antigens invesicles of a similar size to that of the SSV.© 2003 Elsevier B.V. All rights reserved.
Keywords: Plasmodium falciparum; Protein trafficking; Variant surface antigens; RIFIN; PfEMP1; Pf332
1. Introduction
During the asexual intraerythrocytic stage, as thePlas-modium falciparumparasite develops inside the para-sitophorous vacuole (PV), the composition and function ofthe infected erythrocyte (IE) cytosol and plasma membraneare extensively altered. Membranous structures such as theMaurer’s clefts and a tubovesicular network are formed
Abbreviations: ATS, acidic-terminal-sequence; DBL, Duffy-binding-like; IE, infected erythrocyte; IEM, immunoelectron microscopy; IFL, im-munofluorescence; KAHRP, knob-associated histidine-rich protein; LMV,large multimeric vesicle; LSLV, large spindle-like vesicle; PfEMP1,Plasmodium falciparumerythrocyte membrane protein 1; PVM, para-sitophorous vacuole membrane; PV, parasitophorous vacuole;rif , repeti-tive interspersed family; SSV, small single vesicle
∗ Corresponding author. Tel.:+46-8-4572510; fax:+46-8-310525.E-mail address:[email protected] (M. Wahlgren).
beyond the parasitophorous vacuole membrane (PVM)[1,2]. Several parasite-derived proteins are translocated andsorted to compartments in the IE cytosol and to differentlocations near or at the plasma membrane where they canbe associated with the cytoskeleton as well as anchored inthe lipid bilayer and displayed at the surface[3–5]. A num-ber of studies indicate that the traffic of parasite moleculesthrough the IE cytosol is, at least in part, vesicle-mediated[6,7]. Homologues of proteins involved in trafficking path-ways, vesicle coating and fusion have been identified inthe P. falciparumgenome[8–11] but very little is knownabout the mechanisms involved in the transport of parasiteproteins to the surface of the IE surface.
Two groups of polypeptides are known to be expressedby P. falciparum at the IE surface. Both consist of anti-genically variant proteins encoded by large gene families.The best studied comprise the 200–400 kDa adhesins gener-
0166-6851/$ – see front matter © 2003 Elsevier B.V. All rights reserved.doi:10.1016/j.molbiopara.2003.07.006

2 M. Haeggström et al. / Molecular & Biochemical Parasitology 133 (2004) 1–14
ically termedP. falciparumerythrocyte membrane protein 1(PfEMP1) which are encoded by 59var-genes in the haploidgenome[12,13]. PfEMP1 proteins are cytoadhesive ligandsthought to mediate the sequestration of IE in various or-gans, a mechanism by which parasites escape splenic clear-ance[14]. Expression of PfEMP1 polypeptides at the cellsurface takes place as the parasite develops from the earlyring form into the pigmented trophozoite stage, simultane-ously with the onset of adhesive capacity and antigenic-ity of the IE [15]. The RIFIN polypeptides, 30–40 kDa insize encoded by 150–200rif (repetitive interspersed family)genes, constitute the second family of parasite antigens ex-pressed on the IE surface[16–19]. Expression of PfEMP1and RIFINS exhibits clonal variation, a feature thought tohave evolved as a means of immune evasion, improving theodds for prolonged parasite survival, generation of gameto-cytes and transmission to the next host.
We here present data revealing the existence, in wildtypeP. falciparumparasites as well as in laboratory-adaptedstrains, of a unique common trafficking pathway for the sur-face bound antigens RIFINS and PfEMP1. We further showthat the Maurer’s cleft antigen Pf332 transiently co-trafficswith RIFINS and PfEMP1, although Pf332 and the variantantigens segregate to different compartments in the vesiclecomplex. We also show that several other parasite proteinsexported outside of the PV, including the knob-associatedhistidine-rich protein (KAHRP), traffic across the host cellcytosol following pathways distinct from that used by thesurface variant antigens.
2. Materials and methods
2.1. Parasites and cultures
Parasites used in this study were theP. falciparumrosetting clone FCR3S1.2(K−), sub-cloned by microma-nipulation from the rosetting FCR3S1[20] which in turnwas cloned by limiting dilution from FCR3S, a parasiteoriginating from the FCR3 strain isolated in The Gambia,West Africa [21]. The rosetting clone TM284S2(K+) wasderived by micromanipulation from TM284, which was
Table 1Antibodies used in immunofluorescence studies ofPlasmodium falciparumantigens
Antigen Antibody Host/type Reference
RIFIN (repetitive interspersed family (rif )-gene product) #565 Rabbit serum [18]PfEMP1 (P. falciparumerythrocyte membrane protein 1) �-ATS-GST Rat serum [24]
�-DBL1�-GST Rat serum�-DBL1�-GST Rabbit serum
Pf332 (P. falciparum332 antigen) mAb 33G2 Human IgM [25]KAHRP (knob-associated histidine-rich protein) mAb 89 Mouse IgG2a [28]HRP2 (histidine-rich protein 2) mAb 2G12 Mouse IgG1 [29]Exp1 (exported protein 1) McAb 5.1 Mouse IgG1 [32,33]Exp2 (exported protein 2) McAb 7.7 Mouse IgG1 [31]PfEMP2 (P. falciparumerythrocyte membrane protein 2) mAb 4H9.1 Mouse IgM [30]
isolated from a malaria patient in Thailand. The rosettingclone 3D7S8(K+) was derived by micromanipulation from3D7AH1, a clone previously obtained by limiting dilutioncloning from the isolate NF54, originally derived from apatient who acquired malaria at the airport area in Ams-terdam, The Netherlands. The laboratory-adapted parasiteswere kept in continuous culture according to standard pro-cedures with erythrocytes (O+) at 5% hematocrit and 10%AB+ Rh+ serum in buffered culture medium[22]. Cultureswere synchronized in 5% sorbitol for 10 min as previouslydescribed[22,23]. The fresh clinicalP. falciparumisolatesU9, U12, U13, U15, U16, U26, U27 and U31 were obtainedfrom venous blood samples taken from malaria patientsat the Mulago Hospital, Kampala, Uganda. In all cases,informed consent was obtained from the patients and/ortheir parents. The erythrocytes were immediately separatedfrom mononuclear cells on Polymorphprep (Axis-shield)and washed in RPMI. Non-immune AB+ serum to a finalhematocrit of 40% and freezing media (28% Glycerol, 3%Sorbitol, 0.65% NaCl) to a 1:1 ratio were added beforefreezing the cells in liquid N2. For the immunofluorescenceanalysis, the freshly frozen isolates were thawed and main-tained in culture, in their own blood with 15% AB+ serumin buffered culture medium, until parasites developed intotrophozoite stages.
2.2. Sera and specific antibodies
Sera and antibodies used for the detection ofP. fal-ciparum proteins are listed inTable 1. Anti-RIFIN serawere produced in rabbits immunized with synthetic 20amino acids-long peptides corresponding to the conservedC-terminal end of RIFINS. All peptides were conjugated tothe carrier protein KLH. Serum #565 was generated usingas immunogen, the peptide LVLRYLRKKKMKKKLEYIK-LLKE as predicted from the ORF PfB1035w (rif ) in chro-mosome 2 of 3D7[16,18]. Antibodies to PfEMP1 wereobtained by immunizing rabbits and rats with GST-fusionproteins comprising fragments from the highly conservedC-terminal acidic-terminal-sequence (ATS) or the semicon-served N-terminal Duffy-binding-like-1 (DBL-1) domainsof the PfEMP1 encoded by thevar type FCR3S1.2var1

M. Haeggström et al. / Molecular & Biochemical Parasitology 133 (2004) 1–14 3
[24]. The human monoclonal antibody 33G2 recognizes thesequential epitope VTEEI present in theP. falciparumanti-gens Pf332 and Pf155/RESA[25,26]. The lack of overlap inthe expression of these two proteins in the parasite enablesthe use of the 33G2 for the specific detection of Pf332[27].The mAb89 reacts with KAHRP, also called histidine-richprotein 1 (PfHRP1)[28]. The mAb 2G12 was used for thespecific detection of histidine-rich protein 2 (PfHRP2), andthe mAb 4H9.1 as well as the rabbit serum 1054 to detect theerythrocyte membrane protein 2 (PfEMP2)[29,30]. The ex-ported protein 1 (Exp1) and exported protein 2 (Exp2) weredetected with mAb5.1 and mAb7.7, respectively[31–33].
2.3. Indirect immunofluorescence assay for the detectionof intracellular proteins
Plasmodium falciparumclinical isolates and laboratoryparasites at early to mid-trophozoite stages were harvested,washed three times with phosphate-buffered saline (PBS)and diluted to a density of 106 cells ml−1. Monolayers wereprepared by adding 15�l per well of the cell suspensionto 15-well multitest slides (ICN) pre-treated 30 min withpoly-l-Lysine (Sigma). After sedimentation, the excess ofcells was washed away with PBS. The monolayers were air-dried at room temperature and stored in boxes with silicagel (KEBO Lab) for up to 4 weeks. When needed, rosettesand auto-agglutinates were disrupted by adding 100 IU ml−1
Heparin (Lovens) to the culture and by passing it six timesthrough a syringe (0.6 mm)[34].
Monolayers were incubated 30 min with primary antibod-ies (Table 1) diluted in PBS, washed three times in PBS,and incubated 30 min with secondary antibodies conjugatedwith FITC, TRITC or CY3 fluorophores. All incubationswere at RT in a humid chamber. Antibody concentrationswere titrated to achieve minimal leakage over filters whentwo-color staining was performed, and secondary antibodies(DAKO and Jackson Immuno Research), made in suitablehosts, were selected to avoid any possible cross-reactivity.Cover slips were mounted with an anti-fading solution con-sisting of 20% DABCO (Sigma) in glycerol, and the slideswere analyzed with a 100× oil immersion lens in a NikonOptiphot 2 UV microscope equipped with suitable barrierfilters.
2.4. Kinetic studies of synchronous parasite populations
Time-course studies were performed on synchronousFCR3S1.2(K−) parasite cultures, harvested with 2–4 h in-tervals and prepared as above. Four independent studieswere performed, out of which two were on the whole 48-herythrocytic cycle with 4-h intervals, and two focused on theearlier time-points of 10–24 h post-invasion with 2 h inter-vals. For detail studies of vesicle-types (SSV versus LMV)at the time-points between 10 and 24 h post-invasion, 100individual infected cells were analyzed for their content ofsmall vesicles, large multiple vesicles or both types, using
both RIFIN and PfEMP1 reagents. For the study of patternsof transport of the different proteins, cells were analyzed forthe pattern shown in a majority of all cells with cytosolicfluorescence (>100 cells for each time point and reagent).
2.5. Labeling of membrane-containing cytosolicstructures
BODIPY-TR-Ceramide (Molecular Probes) was usedfor the identification of membranous structures withinP.falciparum-infected erythrocytes. BODIPY-TR-Ceramideis a fluorescent lipid-analogue, which freely fuses into lipidstructures. Live cells of FCR3S1.2 cultures were washedin serum-free culture medium, incubated with 25�M ofBODIPY-TR-Ceramide in PBS at 37◦C for 30 min, andwashed with PBS. The culture was examined under UV-microscope for successful incorporation of the lipid marker,and dried cell monolayers were prepared in the dark asdescribed above.
3. Results
3.1. Formation of single small vesicles (SSV), largemultimeric vesicles (LMV) and large spindle-like vesicles(LSLV): a default trafficking pathway
We have studied the transport of the variant surface anti-gens RIFINS and PfEMP1 inP. falciparum-infected erythro-cytes directly isolated from patients as well as in in vitrocultured parasites. For this purpose, air-dried monolayersof infected red cells with parasites grown from ring stagesto mature trophozoites were probed with antibodies to themost conserved portions of the target proteins, i.e. the C-terminal end of the RIFINS, the N-terminal DBL-1� domainof PfEMP1 as well as C-terminal ATS of PfEMP1. Currenttopology models place the C-terminal domains of RIFINSand PfEMP1 at the cytoplasmic side of the IE plasma mem-brane and the DBL-1 moiety on the outside.
The RIFINS and PfEMP1 were detected with identicalimmunofluorescence (IFL) patterns, budding off from thePVM between 6 and 10 h post-invasion and entering thecytosol as single vesicles of≤0.3�m in size (Fig. 1A).In the cytoplasm of erythrocytes infected with maturetrophozoites, RIFINS and PfEMP1 proteins were mainlydetected in large vesicle-like structures with a diameterof 0.8–1.0�m. These structures seemed to consist of anumber of smaller subunits of≤0.3�m in diameter, con-catenated to form an apparently “hollow” doughnut-likering or horseshoe-like assembly of multiple compartmentswith a coarse granular outside (Fig. 1A). In some casesthe large structures appeared rounder and with a smoothersurface. Later, more than 24 h post-invasion, heterogeneousand increasingly enlarged spindle-like structures loadedwith RIFINS and PfEMP1 built up at the periphery of theIE beneath the plasma membrane, suggesting that SSV
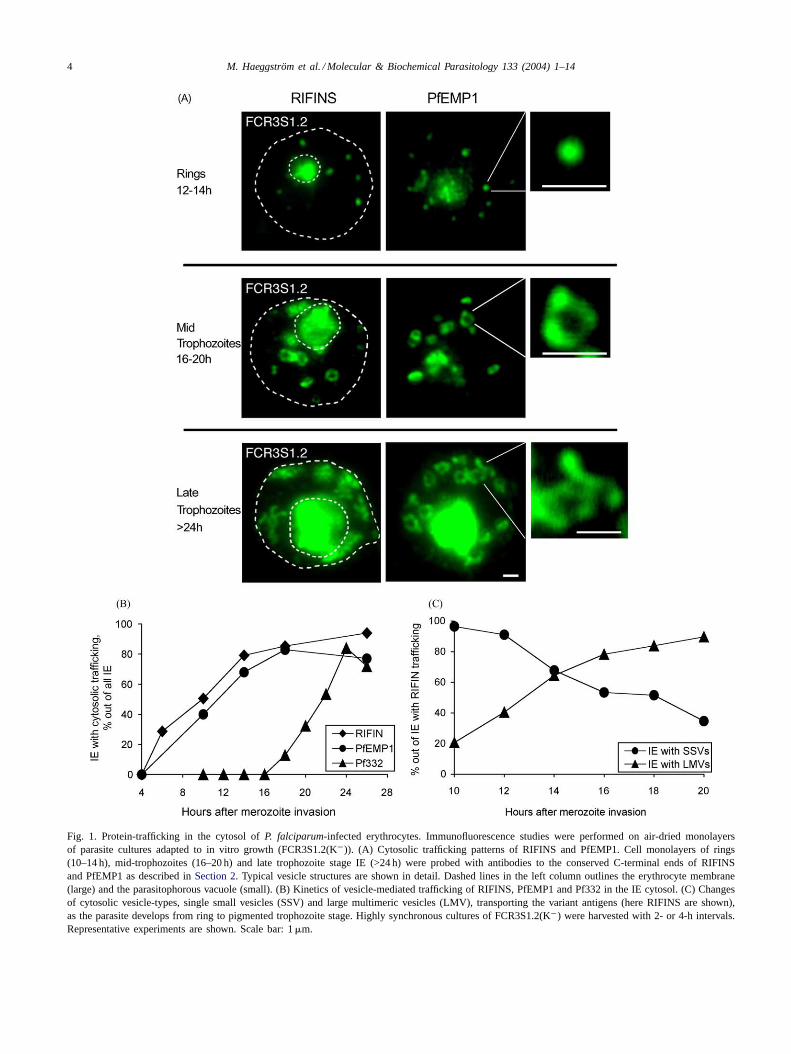
4 M. Haeggström et al. / Molecular & Biochemical Parasitology 133 (2004) 1–14
Fig. 1. Protein-trafficking in the cytosol ofP. falciparum-infected erythrocytes. Immunofluorescence studies were performed on air-dried monolayersof parasite cultures adapted to in vitro growth (FCR3S1.2(K−)). (A) Cytosolic trafficking patterns of RIFINS and PfEMP1. Cell monolayers of rings(10–14 h), mid-trophozoites (16–20 h) and late trophozoite stage IE (>24 h) were probed with antibodies to the conserved C-terminal ends of RIFINSand PfEMP1 as described inSection 2. Typical vesicle structures are shown in detail. Dashed lines in the left column outlines the erythrocyte membrane(large) and the parasitophorous vacuole (small). (B) Kinetics of vesicle-mediated trafficking of RIFINS, PfEMP1 and Pf332 in the IE cytosol. (C) Changesof cytosolic vesicle-types, single small vesicles (SSV) and large multimeric vesicles (LMV), transporting the variant antigens (here RIFINS are shown),as the parasite develops from ring to pigmented trophozoite stage. Highly synchronous cultures of FCR3S1.2(K−) were harvested with 2- or 4-h intervals.Representative experiments are shown. Scale bar: 1�m.
M. Haeggström et al. / Molecular & Biochemical Parasitology 133 (2004) 1–14 5
assembled in LMV that accumulate and possibly further as-sociate into LSLV (Fig. 1A). Identical patterns of PfEMP1fluorescence were obtained with anti-ATS and anti-DBL-1antibodies (data not shown).
3.2. The kinetics of RIFIN and PfEMP1 export concur andthe two antigens co-localize in the IE cytosol
A number of independent experiments suggested thatvesicle-size-differences corresponded to distinct develop-mental stages of the parasites. Rigorous time-course studieswere performed by studying RIFIN- and PfEMP1-specificfluorescence and the formation of trafficking vesicles (SSVversus LMV) in highly synchronous parasite populations,from merozoite invasion to the rupture of schizont IE.
RIFIN and PfEMP1 polypeptides first emerged at the ringstage, 6–10 h post-invasion. Transport of these two antigensreached a maximum, both in terms of the number of vesi-cles per IE and the fraction of IEs displaying transport vesi-
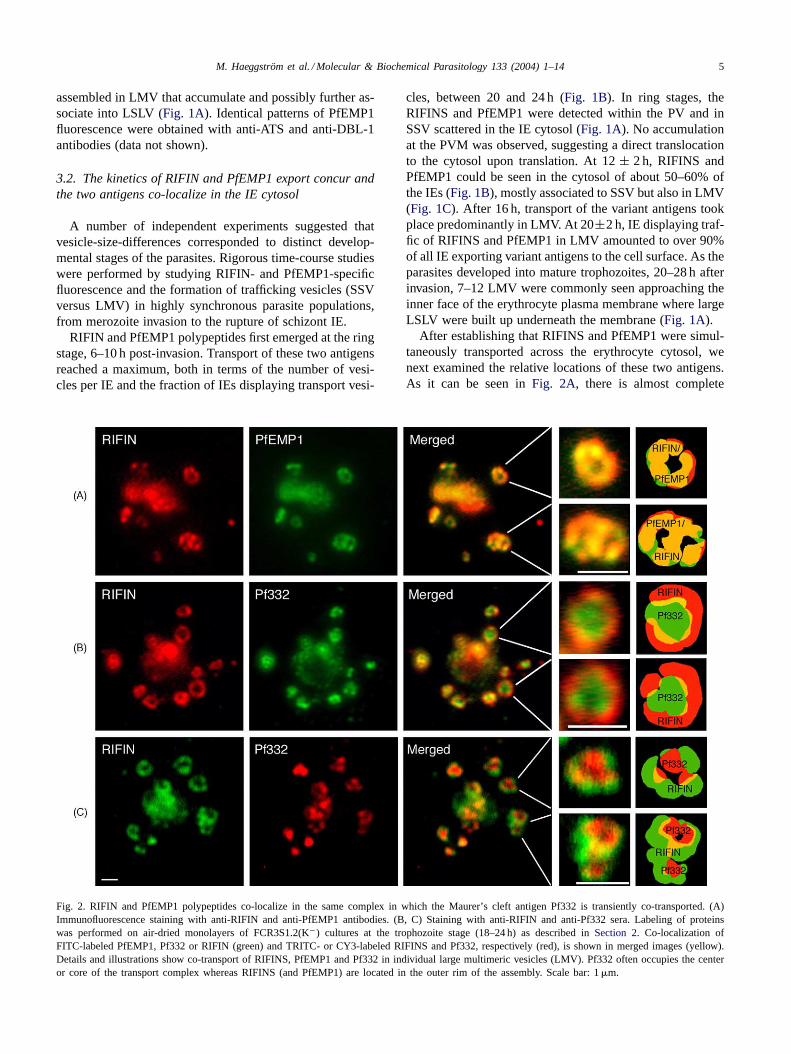
Fig. 2. RIFIN and PfEMP1 polypeptides co-localize in the same complex in which the Maurer’s cleft antigen Pf332 is transiently co-transported. (A)Immunofluorescence staining with anti-RIFIN and anti-PfEMP1 antibodies. (B, C) Staining with anti-RIFIN and anti-Pf332 sera. Labeling of proteinswas performed on air-dried monolayers of FCR3S1.2(K−) cultures at the trophozoite stage (18–24 h) as described inSection 2. Co-localization ofFITC-labeled PfEMP1, Pf332 or RIFIN (green) and TRITC- or CY3-labeled RIFINS and Pf332, respectively (red), is shown in merged images (yellow).Details and illustrations show co-transport of RIFINS, PfEMP1 and Pf332 in individual large multimeric vesicles (LMV). Pf332 often occupies the centeror core of the transport complex whereas RIFINS (and PfEMP1) are located in the outer rim of the assembly. Scale bar: 1�m.
cles, between 20 and 24 h (Fig. 1B). In ring stages, theRIFINS and PfEMP1 were detected within the PV and inSSV scattered in the IE cytosol (Fig. 1A). No accumulationat the PVM was observed, suggesting a direct translocationto the cytosol upon translation. At 12± 2 h, RIFINS andPfEMP1 could be seen in the cytosol of about 50–60% ofthe IEs (Fig. 1B), mostly associated to SSV but also in LMV(Fig. 1C). After 16 h, transport of the variant antigens tookplace predominantly in LMV. At 20±2 h, IE displaying traf-fic of RIFINS and PfEMP1 in LMV amounted to over 90%of all IE exporting variant antigens to the cell surface. As theparasites developed into mature trophozoites, 20–28 h afterinvasion, 7–12 LMV were commonly seen approaching theinner face of the erythrocyte plasma membrane where largeLSLV were built up underneath the membrane (Fig. 1A).
After establishing that RIFINS and PfEMP1 were simul-taneously transported across the erythrocyte cytosol, wenext examined the relative locations of these two antigens.As it can be seen inFig. 2A, there is almost complete
6 M. Haeggström et al. / Molecular & Biochemical Parasitology 133 (2004) 1–14
co-localization of RIFINS and PfEMP1 in the LMV. Lessfrequently, SSV carrying either PfEMP1 or RIFINS wereobserved in the cytoplasm of individual IEs. These resultsprovide strong evidence that a unique transport pathwayaccounts for the translocation of translated products ofthe two variant antigen families, from the parasite to theerythrocyte membrane. This pathway involves single andmultimeric vesicles, the latter possibly resulting from theorganized interaction of the individual smaller vesicles.
3.3. LMV formation in fresh isolates and in vitro grownparasites
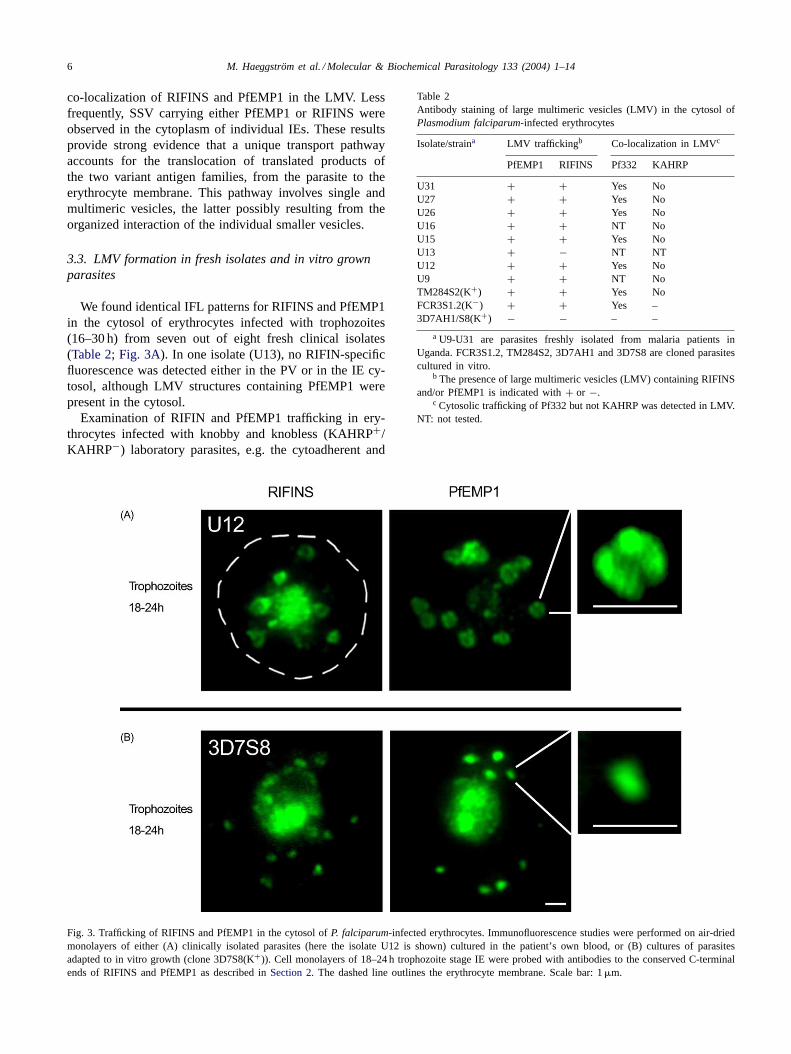
We found identical IFL patterns for RIFINS and PfEMP1in the cytosol of erythrocytes infected with trophozoites(16–30 h) from seven out of eight fresh clinical isolates(Table 2; Fig. 3A). In one isolate (U13), no RIFIN-specificfluorescence was detected either in the PV or in the IE cy-tosol, although LMV structures containing PfEMP1 werepresent in the cytosol.
Examination of RIFIN and PfEMP1 trafficking in ery-throcytes infected with knobby and knobless (KAHRP+/KAHRP−) laboratory parasites, e.g. the cytoadherent and
Fig. 3. Trafficking of RIFINS and PfEMP1 in the cytosol ofP. falciparum-infected erythrocytes. Immunofluorescence studies were performed on air-driedmonolayers of either (A) clinically isolated parasites (here the isolate U12 is shown) cultured in the patient’s own blood, or (B) cultures of parasitesadapted to in vitro growth (clone 3D7S8(K+)). Cell monolayers of 18–24 h trophozoite stage IE were probed with antibodies to the conserved C-terminalends of RIFINS and PfEMP1 as described inSection 2. The dashed line outlines the erythrocyte membrane. Scale bar: 1�m.
Table 2Antibody staining of large multimeric vesicles (LMV) in the cytosol ofPlasmodium falciparum-infected erythrocytes
Isolate/straina LMV traffickingb Co-localization in LMVc
PfEMP1 RIFINS Pf332 KAHRP
U31 + + Yes NoU27 + + Yes NoU26 + + Yes NoU16 + + NT NoU15 + + Yes NoU13 + − NT NTU12 + + Yes NoU9 + + NT NoTM284S2(K+) + + Yes NoFCR3S1.2(K−) + + Yes –3D7AH1/S8(K+) − − – –
a U9-U31 are parasites freshly isolated from malaria patients inUganda. FCR3S1.2, TM284S2, 3D7AH1 and 3D7S8 are cloned parasitescultured in vitro.
b The presence of large multimeric vesicles (LMV) containing RIFINSand/or PfEMP1 is indicated with+ or −.
c Cytosolic trafficking of Pf332 but not KAHRP was detected in LMV.NT: not tested.

M. Haeggström et al. / Molecular & Biochemical Parasitology 133 (2004) 1–14 7
rosetting clones TM284S2(K+) and FCR3S1.2(K−), re-vealed a similar transport pattern as in freshP. falciparumisolates (Table 2; Fig 5B). This was not the case withthe 3D7AH1 strain or its cytoadherent and rosetting clone3D7S8, as large multimeric structures (LMV, LSLV) werenever observed in the cytosol of IE bearing these parasites.In 3D7 parasites, transport of RIFIN and PfEMP1 in bothyoung and mature stages occurred in small fluorescentpackets of≤0.5�m in diameter (Fig. 3B).
3.4. The Maurer’s cleft marker Pf332 is transientlyco-transported by LMV carrying RIFINS and PfEMP1
The P. falciparum antigen Pf332 is a high molecularweight protein transported to the plasma membrane of theIE in association with Maurer’s clefts in the erythrocytecytoplasm[27,35]. We studied the time-course of Pf332transport, and its location in the FCR3S1.2 IE with respectto RIFINS and PfEMP1. In contrast to the variant anti-gens, Pf332 was not detected in SSV in the cytosol butaccumulated in the PV/PVM in young ring stages. First,at approximately 18–24 h, LMVs carrying Pf332 (Figs. 1Band 2B, C) as well as RIFINS and PfEMP1 (Fig. 2A),could be detected in the cytosol. In late trophozoiteand schizont stages the fluorescence pattern again dif-fers between Pf332 and RIFINS/PfEMP1, as Pf332 wasfound to associated with the red cell plasma membranebut not seen in the parasite or in the cytosol (data notshown).
3.5. LMV are sub-compartmentalized
The variant antigens (RIFIN/PfEMP1) and Pf332 seg-regate into distinct compartments of the LMV complexes.RIFINS and PfEMP1 were found located in the outer rimof the assembly (Fig. 2A), whereas Pf332 was often seen tooccupy the center or core of the vesicular complex (Fig. 2Band C). Spatial dissociation of the variant antigens and
Fig. 4. Demonstration of the lipid content of the vesicular cytosolic structures involved in trafficking of variant antigens to the IE surface. FCR3S1.2(K−)cultures at trophozoite stage (18–24 h) were pre-incubated with BODIPY-TR-Ceramide that label membrane-containing structures (red). Air-driedmonolayers were prepared in the dark and proteins were FITC-labeled as described inSection 2. Scale bar: 1�m.
Pf332 in different compartments of the same complex wasfound to be highly prevalent, while in a few LMV core ver-sus peripheral localization of the antigens was not evident.These results further suggest that the transport pathway ofRIFIN and PfEMP1 antigens across the IE cytosol com-prises a compartment formed by primary vesicles (small,≤0.3�m) loaded with RIFINS and/or PfEMP1 whichdock to and perhaps eventually fuse with an outboundvesicular complex carrying the export machinery and pre-sumably other proteins, among them Pf332 in a transientmanner.
3.6. LMV associate with lipids as seen byBODIPY-Ceramide staining
To investigate whether trafficking of the variant antigensin the IE cytosol was due to classical transport of vesic-ular nature we studied their association with membranousstructures. Upon labeling of parasites with the fluorescentlipid BODIPY-Ceramide, intensive staining of the PV andof membranes budding from the vacuole was evident. Prob-ing of the ceramide-labeled IE with anti-RIFIN or anti-PfEMP1 antibodies showed co-localization of lipid- andprotein-associated fluorescence in the cytosol (Fig. 4). Thesedata indicated that vesicles, SSV and LMV, loaded withRIFIN and PfEMP1 polypeptides, mediate the export ofthese proteins from the PV to the erythrocyte plasma mem-brane.
3.7. The knob-associated protein KAHRP is exported in aseparate trafficking pathway
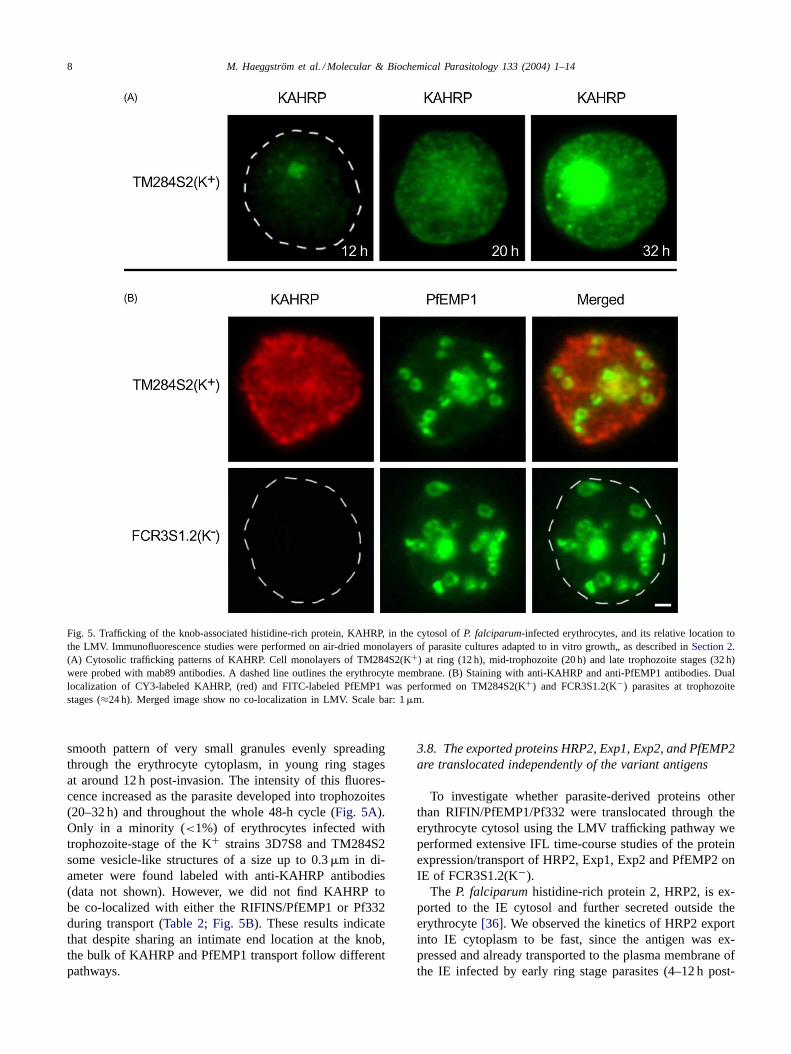
The finding of surface variant antigens (RIFIN/PfEMP1)transported similarly in K+ and K− IE motivated a de-tailed analysis of KAHRP translocation across the ery-throcyte cytosol. IFL detection of KAHRP protein in IEfrom fresh clinical isolates and K+ laboratory-adaptedparasites showed parasite-specific staining, as well as a
8 M. Haeggström et al. / Molecular & Biochemical Parasitology 133 (2004) 1–14
Fig. 5. Trafficking of the knob-associated histidine-rich protein, KAHRP, in the cytosol ofP. falciparum-infected erythrocytes, and its relative location tothe LMV. Immunofluorescence studies were performed on air-dried monolayers of parasite cultures adapted to in vitro growth„ as described inSection 2.(A) Cytosolic trafficking patterns of KAHRP. Cell monolayers of TM284S2(K+) at ring (12 h), mid-trophozoite (20 h) and late trophozoite stages (32 h)were probed with mab89 antibodies. A dashed line outlines the erythrocyte membrane. (B) Staining with anti-KAHRP and anti-PfEMP1 antibodies. Duallocalization of CY3-labeled KAHRP, (red) and FITC-labeled PfEMP1 was performed on TM284S2(K+) and FCR3S1.2(K−) parasites at trophozoitestages (≈24 h). Merged image show no co-localization in LMV. Scale bar: 1�m.
smooth pattern of very small granules evenly spreadingthrough the erythrocyte cytoplasm, in young ring stagesat around 12 h post-invasion. The intensity of this fluores-cence increased as the parasite developed into trophozoites(20–32 h) and throughout the whole 48-h cycle (Fig. 5A).Only in a minority (<1%) of erythrocytes infected withtrophozoite-stage of the K+ strains 3D7S8 and TM284S2some vesicle-like structures of a size up to 0.3�m in di-ameter were found labeled with anti-KAHRP antibodies(data not shown). However, we did not find KAHRP tobe co-localized with either the RIFINS/PfEMP1 or Pf332during transport (Table 2; Fig. 5B). These results indicatethat despite sharing an intimate end location at the knob,the bulk of KAHRP and PfEMP1 transport follow differentpathways.
3.8. The exported proteins HRP2, Exp1, Exp2, and PfEMP2are translocated independently of the variant antigens
To investigate whether parasite-derived proteins otherthan RIFIN/PfEMP1/Pf332 were translocated through theerythrocyte cytosol using the LMV trafficking pathway weperformed extensive IFL time-course studies of the proteinexpression/transport of HRP2, Exp1, Exp2 and PfEMP2 onIE of FCR3S1.2(K−).
The P. falciparumhistidine-rich protein 2, HRP2, is ex-ported to the IE cytosol and further secreted outside theerythrocyte[36]. We observed the kinetics of HRP2 exportinto IE cytoplasm to be fast, since the antigen was ex-pressed and already transported to the plasma membrane ofthe IE infected by early ring stage parasites (4–12 h post-
M. Haeggström et al. / Molecular & Biochemical Parasitology 133 (2004) 1–14 9
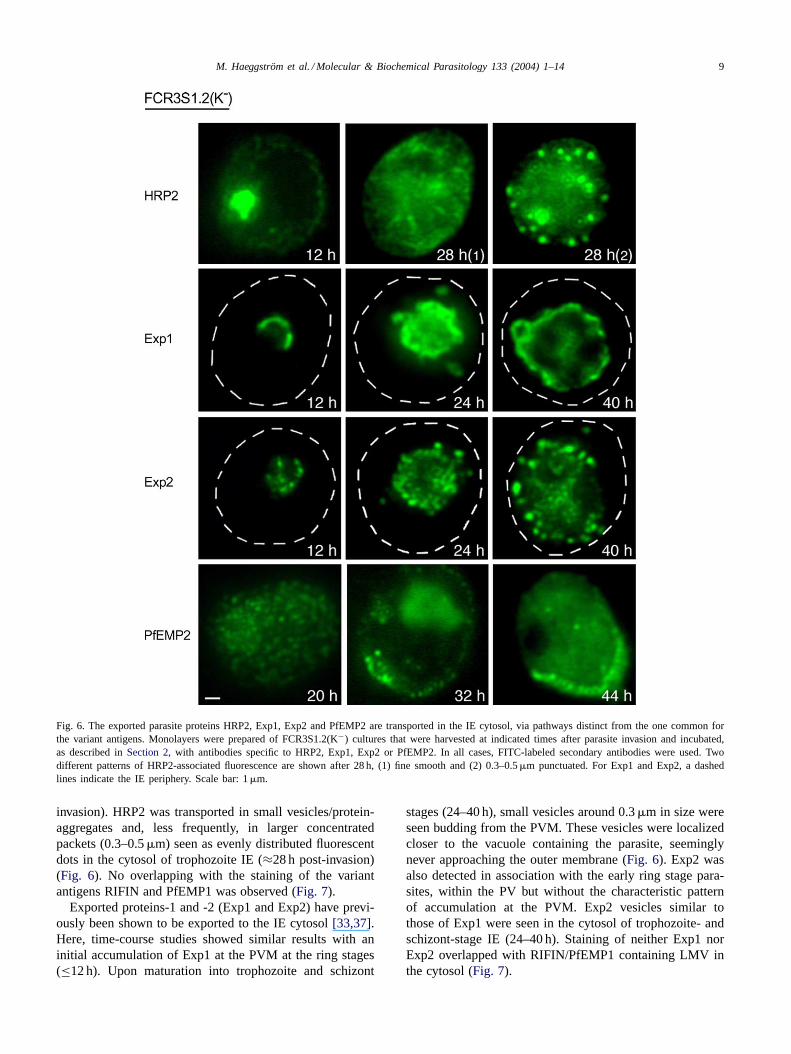
Fig. 6. The exported parasite proteins HRP2, Exp1, Exp2 and PfEMP2 are transported in the IE cytosol, via pathways distinct from the one common forthe variant antigens. Monolayers were prepared of FCR3S1.2(K−) cultures that were harvested at indicated times after parasite invasion and incubated,as described inSection 2, with antibodies specific to HRP2, Exp1, Exp2 or PfEMP2. In all cases, FITC-labeled secondary antibodies were used. Twodifferent patterns of HRP2-associated fluorescence are shown after 28 h, (1) fine smooth and (2) 0.3–0.5�m punctuated. For Exp1 and Exp2, a dashedlines indicate the IE periphery. Scale bar: 1�m.
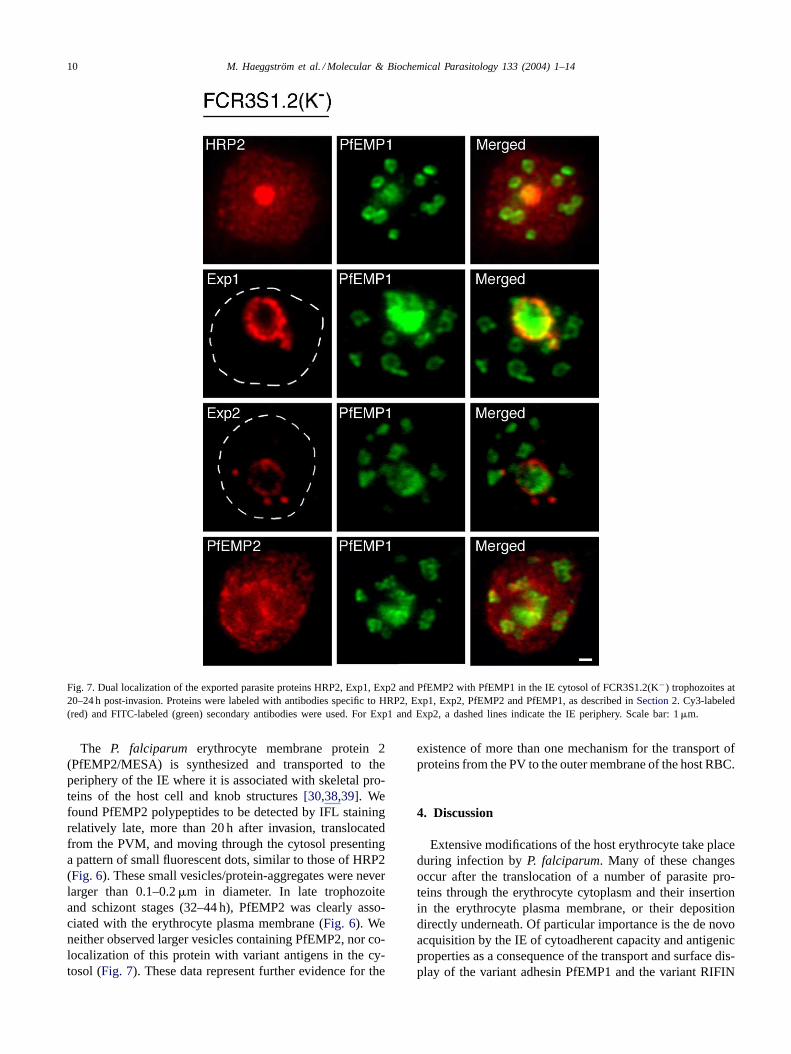
invasion). HRP2 was transported in small vesicles/protein-aggregates and, less frequently, in larger concentratedpackets (0.3–0.5�m) seen as evenly distributed fluorescentdots in the cytosol of trophozoite IE (≈28 h post-invasion)(Fig. 6). No overlapping with the staining of the variantantigens RIFIN and PfEMP1 was observed (Fig. 7).
Exported proteins-1 and -2 (Exp1 and Exp2) have previ-ously been shown to be exported to the IE cytosol[33,37].Here, time-course studies showed similar results with aninitial accumulation of Exp1 at the PVM at the ring stages(≤12 h). Upon maturation into trophozoite and schizont
stages (24–40 h), small vesicles around 0.3�m in size wereseen budding from the PVM. These vesicles were localizedcloser to the vacuole containing the parasite, seeminglynever approaching the outer membrane (Fig. 6). Exp2 wasalso detected in association with the early ring stage para-sites, within the PV but without the characteristic patternof accumulation at the PVM. Exp2 vesicles similar tothose of Exp1 were seen in the cytosol of trophozoite- andschizont-stage IE (24–40 h). Staining of neither Exp1 norExp2 overlapped with RIFIN/PfEMP1 containing LMV inthe cytosol (Fig. 7).
10 M. Haeggström et al. / Molecular & Biochemical Parasitology 133 (2004) 1–14
Fig. 7. Dual localization of the exported parasite proteins HRP2, Exp1, Exp2 and PfEMP2 with PfEMP1 in the IE cytosol of FCR3S1.2(K−) trophozoites at20–24 h post-invasion. Proteins were labeled with antibodies specific to HRP2, Exp1, Exp2, PfEMP2 and PfEMP1, as described inSection 2. Cy3-labeled(red) and FITC-labeled (green) secondary antibodies were used. For Exp1 and Exp2, a dashed lines indicate the IE periphery. Scale bar: 1�m.
The P. falciparum erythrocyte membrane protein 2(PfEMP2/MESA) is synthesized and transported to theperiphery of the IE where it is associated with skeletal pro-teins of the host cell and knob structures[30,38,39]. Wefound PfEMP2 polypeptides to be detected by IFL stainingrelatively late, more than 20 h after invasion, translocatedfrom the PVM, and moving through the cytosol presentinga pattern of small fluorescent dots, similar to those of HRP2(Fig. 6). These small vesicles/protein-aggregates were neverlarger than 0.1–0.2�m in diameter. In late trophozoiteand schizont stages (32–44 h), PfEMP2 was clearly asso-ciated with the erythrocyte plasma membrane (Fig. 6). Weneither observed larger vesicles containing PfEMP2, nor co-localization of this protein with variant antigens in the cy-tosol (Fig. 7). These data represent further evidence for the
existence of more than one mechanism for the transport ofproteins from the PV to the outer membrane of the host RBC.
4. Discussion
Extensive modifications of the host erythrocyte take placeduring infection byP. falciparum. Many of these changesoccur after the translocation of a number of parasite pro-teins through the erythrocyte cytoplasm and their insertionin the erythrocyte plasma membrane, or their depositiondirectly underneath. Of particular importance is the de novoacquisition by the IE of cytoadherent capacity and antigenicproperties as a consequence of the transport and surface dis-play of the variant adhesin PfEMP1 and the variant RIFIN

M. Haeggström et al. / Molecular & Biochemical Parasitology 133 (2004) 1–14 11
polypeptides. We now demonstrate that a common traffick-ing pathway mediates the transport of RIFINS and PfEMP1through the IE cytosol. PfEMP1 has previously been local-ized to single and multiple string-like vesicles in the erythro-cyte cytosol[7,9,40,41]. Our data show that the cytosoliclocation of RIFINS in early single vesicles (SSV,≈0.3�m)budding off the PVM and subsequent vesicle-like struc-tures of increasing complexity (LMV and LSLV, >0.8�m)overlaps with the location of PfEMP1. Fine analysis of thefluorescence distribution in LMV shows an almost completeco-localization of RIFIN and PfEMP1 polypeptides in aperipherally located compartment of these vesicular assem-blies. Further, we found the secreted polypeptide Pf332 tobe co-transported with RIFIN and PfEMP1 in LMV. Pf332has previously been shown both by IFL and immunoelectronmicroscopy (IEM) studies to localize in large membrane-containing cytosolic structures, referred to as Maurer’sclefts, before associating with the erythrocyte membrane[27,35]. Translocation of Pf332, a 600 kDa polypeptidewith one predicted transmembrane region, from the parasiteto the IE cytosol is completely inhibited by Brefeldin A, afungal metabolite which induces redistribution of proteinsfrom Golgi to ER and consequently blocks protein transportin eukaryotic cells, suggesting classical vesicle-mediatedtransport of Pf332. Another exported protein found in largecytosolic vesicles resembling LMV or Maurer’s clefts isthe P. falciparum skeleton binding protein 1, PfSbp1, apredicted transmembrane polypeptide which is associatedwith the erythrocyte cytoskeleton[42]. No function hasyet been ascribed to Pf332 or PfSbp1 although the latterhas been postulated to mediate binding to the cytoskeleton.P. falciparumhomologues of proteins involved in vesiclecoating, docking and trafficking as well as in membrane fu-sion, PfNSF, PfSar1p and Sec31p, have also been identifiedin cytosolic compartments with the features of LMV andMaurer’s clefts[8–10]. Co-localization with PfEMP1 hasbeen determined for PfSbp1, PfSar1p and PfSec31p. Yetanother parasite protein shown to traffic in association withPfEMP1 co-localizing at the Maurer’s clefts is theP. falci-parumerythrocyte membrane protein 3, PfEMP3, which hasstructural similarity to yeast factors involved in protein sort-ing and vesicle targeting[41]. The evidence here presented,taken together with the data from others, is consistent with asystem of mobile vesicles within the erythrocyte cytoplasmcarrying the variant antigens to the IE surface and a numberof other proteins necessary for (i) the formation of the indi-vidual vesicles at the PVM, (ii) their further complexing orassociation of the individual vesicles, (iii) directionalityof transport, (iv) interaction with cytoskeleton and plasmamembrane, and (v) adequate delivery at the final targetlocation.
Notably, parasite proteins such asP. falciparum ery-throcyte membrane protein 2, PfEMP2/MESA, which isexported to the vicinity of the knob where it associateswith the electron-dense structures and the cytoskeleton,and P. falciparum histidine-rich protein 2, HRP2, which
is exported and secreted outside the IE, do not traffic theerythrocyte cytosol via the LMV (Maurer’s cleft) pathway.This fact underscores the common export route/mechanismfor proteins destined to insertion in the outer membrane,i.e. PfEMP1, RIFINS, and presumably Pf332.
The KAHRP protein is exported by the parasite to re-gions underlying the IE plasma membrane[43–45]. Oncethere, in addition to an essential role in the formation ofknobs, KAHRP is believed to be involved in the anchor-ing of PfEMP1[40,46,47]. Despite the close interaction ofKAHRP and PfEMP1 at their end location in the knob, thesetwo proteins do not co-localize in SSV or LMV. In agree-ment with earlier reports[4,27,48], our IFL data obtainedusing laboratory strains and freshly isolated parasites indi-cates that the cytosolic traffic of KAHRP essentially takesplace via a vesicle-independent pathway, presumably in adiffusible fashion. Although a sporadic transit of KAHRPthrough Maurer’s clefts cannot be ruled out[49], our data isnot consistent with co-localized trafficking of KAHRP andPfEMP1 along the LMV pathway.
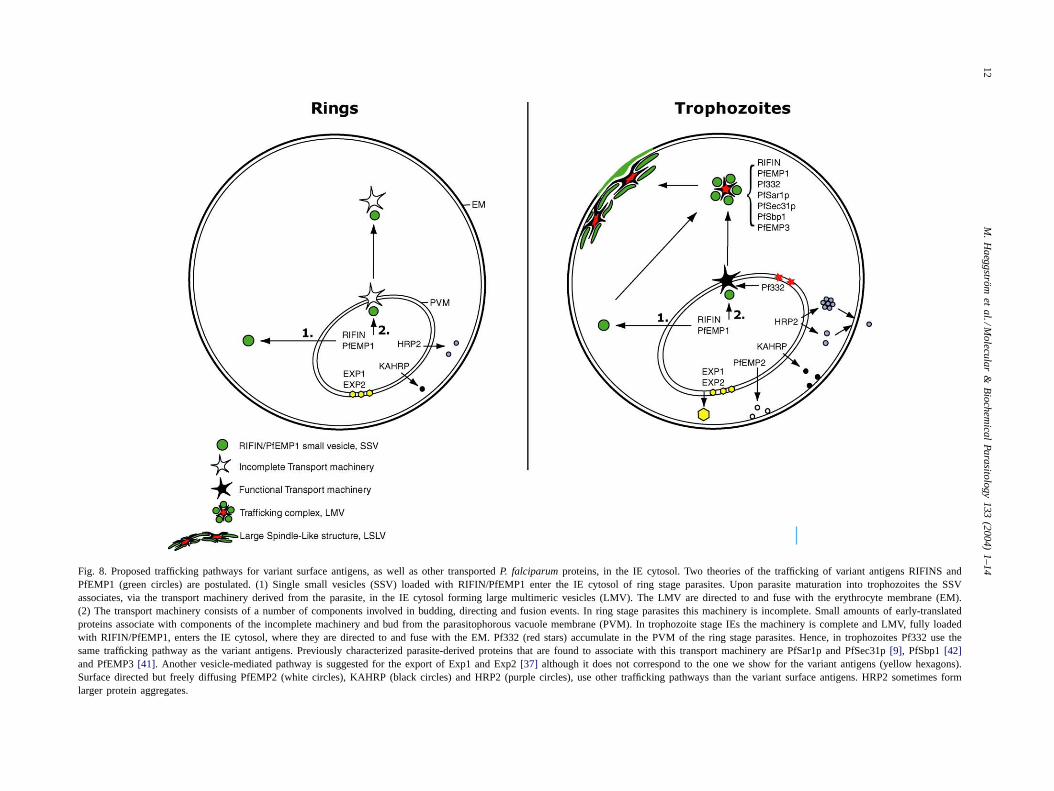
The transport of proteins, lipids and other molecules inthe cytosol ofP. falciparum-infected erythrocytes has beenwidely investigated. The existence of classical and alter-native trafficking pathways has been suggested, althoughpoorly defined. The classical trafficking pathways, definedby vesicle-mediated transport, include vesicle budding andfusion with membranes, whereas alternative pathways in-clude all non-vesicle-mediated transport mechanisms, suchas the movement of lipid-free protein aggregates within theerythrocyte cytosol. Focus has been on the classical path-way, with major efforts conducted on the characterizationand understanding of the mechanisms involved[9,11,50].Here we show that single vesicles transporting RIFIN andPfEMP1 proteins bud off from the PVM and enter the IEcytosol at 6–10 h post-invasion. These early compartments,the SSV, may represent either (1) actual independent vesi-cles moving in the cytosol or (2) incomplete assembliesof the transport machinery (see alternatives 1 and 2 in thescheme for trafficking of proteins in the IE cytosol,Fig. 8).Upon parasite maturation into trophozoites at 16–20 h post-invasion, a functional transport unit including all auxiliarytransport proteins is assembled, to which further singlevesicles may dock and conceivably fuse. This functionalassembly, the LMV, can then move towards the periphery ofthe IE, approaching the cytoskeleton and inner leaflet of thered cell membrane. The morphological evidence providedby IFL, the relative localization of single and multimericvesicles in the IE cytoplasm, and the mutually exclusivekinetics of trafficking between these two vesicle archetypesstrongly suggest that LMV result from the association ofSSV. It is not clear whether LMV/Maurer’s clefts representa sorting station for exported proteins, which eventuallyresume their trafficking to the membrane contained insmaller secondary vesicles, or a compartment that physi-cally reaches the plasma membrane. A possibility is thatboth events may take place. The IFL observed suggests that
12M
.H
aegg
ström
et
al./M
ole
cula
r&
Bio
che
mica
lP
ara
sitolog
y1
33
(20
04
)1
–1
4
Fig. 8. Proposed trafficking pathways for variant surface antigens, as well as other transportedP. falciparumproteins, in the IE cytosol. Two theories of the trafficking of variant antigens RIFINS andPfEMP1 (green circles) are postulated. (1) Single small vesicles (SSV) loaded with RIFIN/PfEMP1 enter the IE cytosol of ring stage parasites. Upon parasite maturation into trophozoites the SSVassociates, via the transport machinery derived from the parasite, in the IE cytosol forming large multimeric vesicles (LMV). The LMV are directed toand fuse with the erythrocyte membrane (EM).(2) The transport machinery consists of a number of components involved in budding, directing and fusion events. In ring stage parasites this machinery is incomplete. Small amounts of early-translatedproteins associate with components of the incomplete machinery and bud from the parasitophorous vacuole membrane (PVM). In trophozoite stage IEs the machinery is complete and LMV, fully loadedwith RIFIN/PfEMP1, enters the IE cytosol, where they are directed to and fuse with the EM. Pf332 (red stars) accumulate in the PVM of the ring stage parasites. Hence, in trophozoites Pf332 use thesame trafficking pathway as the variant antigens. Previously characterized parasite-derived proteins that are found to associate with this transport machinery are PfSar1p and PfSec31p[9], PfSbp1[42]and PfEMP3[41]. Another vesicle-mediated pathway is suggested for the export of Exp1 and Exp2[37] although it does not correspond to the one we show for the variant antigens (yellow hexagons).Surface directed but freely diffusing PfEMP2 (white circles), KAHRP (black circles) and HRP2 (purple circles), use other trafficking pathways than the variant surface antigens. HRP2 sometimes formlarger protein aggregates.

M. Haeggström et al. / Molecular & Biochemical Parasitology 133 (2004) 1–14 13
several LMV may associate in larger networks, the LSLV,which are detected in close proximity to the red blood cellmembrane. Whether these structures are part of an extendednetwork or a reflection of LVM approaching or fusing withthe membrane is not determined here.
It has previously been suggested that a continuous mem-brane network, extending from the PVM across the erythro-cyte cytoplasm, could be involved in the uptake of serumcomponents, as well as in the trafficking and export ofparasite proteins outside the PV[51–54]. According to thismodel, surface-bound antigens such as PfEMP1 and RIFINSwould traffic most or the entire gap between the parasite andthe erythrocyte plasma membrane by lateral diffusion, aftertheir insertion in the membrane network. The existence oflipid-rich tubovesicular structures in the cytoplasmic spaceof the IE has been demonstrated by fluorometry and electronmicroscopy[54,55]. Our time-course data clearly show that,as the parasite develops from ring to mature trophozoite, thevast majority of the antigens recognized by anti-PfEMP1and anti-RIFIN antibodies are associated with discrete struc-tures, in contrast to a continuous network. These structures,which initially are of dimensions compatible with transportvesicles, subsequently are apparent as highly organized com-plexes. Vesicle-like structures within the host erythrocytecytoplasm have been shown by others, both by immunoflu-orescence and electron microscopy analysis[7,9,11,27,42].The interplay between early transport vesicles and laterhighly compartmentalized complexes, perhaps forming partof an extended cytosolic network, remains to be elucidated.
The finding of prominent LMV structures in the IE cytosolof all P. falciparum fresh isolates examined could reflecta crucial role of the export pathway for parasite survivalduring residence in the vertebrate host. It could be speculatedthat this feature of wild parasites might be related to theenhanced surface expression of RIFIN proteins in freshlyisolatedP. falciparum [18]. We do not know whether theabsence of LMV in the cytosol of erythrocytes infected withparasites such as 3D7AH1 and its rosetting clone 3D7S8 isof decisive importance for the export of variant antigens andtheir surface expression. Further investigations are needed todissect the molecular interactions and mechanisms operatingin the export pathway of the surface-bound variant antigens.
Acknowledgements
We wish to thank Berit Schmidt-Aydin for the collec-tion and processing of patient blood samples. We thankDiane Taylor (Georgetown University, Washington, DC)for kindly providing the Mab89; Russel J. Howard (Maxy-gen, Redwood City, CA) for the kind gift of MabID6ascites, Mab4H9.1 ascites and rabbit serum 1054; and JanaS. McBride (University of Edinburgh, Edinburgh, Scot-land) for the kind gift of Mab 5.1 and 7.7. This work wasfunded by grants from the United Nations Developing Pro-gramme/World Bank/World Health Organization Special
Programme for Research and Training in Tropical Dis-eases (TDR), the European Union (QLRT-PL-1999-30109(ADMAL) and QLK2-2002-01197 (Euromalvac 2)), theSwedish Research Council and the Swedish InternationalDevelopment Authority (Sida/SAREC) to Mats Wahlgrenand Klavs Berzins.
References
[1] Aikawa M, Miller LH, Rabbege J. Caveola–vesicle complexes inthe plasmalemma of erythrocytes infected byPlasmodium vivaxandP. cynomolgi. Unique structures related to Schuffner’s dots. Am JPathol 1975;79:285–300.
[2] Atkinson CT, Aikawa M, Perry G, et al. Ultrastructural localization oferythrocyte cytoskeletal and integral membrane proteins inPlasmod-ium falciparum-infected erythrocytes. Eur J Cell Biol 1988;45:192–9.
[3] Barnwell JW. Vesicle-mediated transport of membrane and proteinsin malaria-infected erythrocytes. Blood Cells 1990;16:379–95.
[4] Gormley JA, Howard RJ, Taraschi TF. Trafficking of malarial proteinsto the host cell cytoplasm and erythrocyte surface membrane involvesmultiple pathways. J Cell Biol 1992;119:1481–95.
[5] Deitsch K, Wellems T. Membrane modifications in erythrocytesparasitized by Plasmodium falciparum. Mol Biochem Parasitol1996;76:1–10.
[6] Hibbs AR, Saul AJ.Plasmodium falciparum: highly mobile smallvesicles in the malaria-infected red blood cell cytoplasm. Exp Para-sitol 1994;79:260–9.
[7] Trelka DP, Schneider TG, Reeder JC, Taraschi TF. Evidence forvesicle-mediated trafficking of parasite proteins to the host cell cy-tosol and erythrocyte surface membrane inPlasmodium falciparuminfected erythrocytes. Mol Biochem Parasitol 2000;106:131–45.
[8] Albano FR, Berman A, La Greca N. A homologue of Sar1p localisesto a novel trafficking pathway in malaria-infected erythrocyte. Eur JCell Biol 1999;78:453–62.
[9] Adisa A, Albano FR, Reeder J, Foley M, Tilley L. Evidence for arole for aPlasmodium falciparumhomologue of Sec31p in the exportof proteins to the surface of malaria parasite-infected erythrocytes.J Cell Sci 2001;114:3377–86.
[10] Hayashi M, Taniguchi S, Ishizuka Y, et al. A homologue ofN-ethylmaleimide-sensitive factor in the malaria parasitePlasmodiumfalciparum is exported and localized in vesicular structures in thecytoplasm of infected erythrocytes in the Brefeldin A-sensitive path-way. J Biol Chem 2001;276:15249–55.
[11] Taraschi TF, Trelka DP, Martinez S, Schneider TG, O’Donnell ME.Vesicle-mediated trafficking of parasite proteins to the host cellcytosol and erythrocyte surface membrane inPlasmodium falciparuminfected erythrocytes. Int J Parasitol 2001;31:1381–91.
[12] Baruch DI, Pasloske BL, Singh HB, et al. Cloning theP. falci-parum gene endcoding PfEMP1, a malarial variant antigen and ad-herence receptor on the surface of parasitized human erythrocytes.Cell 1995;82:77–87.
[13] Su X, Heatwole VM, Wertheimer SP. The large diverse genefamily var encodes proteins involved in cytoadherence and anti-genic variation ofPlasmodium falciparum-infected erythrocytes. Cell1995;82:89–100.
[14] Chen Q, Schlichtherle M, Wahlgren M. Molecular aspects of severemalaria. Clin Microbiol Rev 2000;13:439–50.
[15] Howard RJ. Malaria proteins at the membrane ofPlasmodiumfalciparum-infected erythrocytes and their involvement in cytoadher-ence to endothelial cells. In: Perlmann P, Wigzell H, editors. Malariaimmunology, Vol. 41. Progress in allergy. Basel: Karger AG; 1988.p. 98–147.
[16] Gardner MJ, Tettelin H, Carucci DJ, et al. Chromosome 2 se-quence of the human malaria parasitePlasmodium falciparum. Sci-ence 1998;282:1126–32.

14 M. Haeggström et al. / Molecular & Biochemical Parasitology 133 (2004) 1–14
[17] Helmby H, Cavelier L, Pettersson U, Wahlgren M. RosettingPlas-modium falciparum-infected erythrocytes express unique antigens ontheir surface. Infect Immun 1993;61:284–8.
[18] Fernandez V, Hommel M, Chen Q, Hagblom P, Wahlgren M. Small,clonally variant antigens expressed on the surface of thePlasmod-ium falciparum-infected Erythrocyte are encoded by therif genefamily and are the target of human immune responses. J Exp Med1999;190:1393–404.
[19] Kyes S, Rowe A, Kriek N, Newbold C. Rifins: a second familyof clonally variant proteins expressed on the surface of red cellsinfected with Plasmodium falciparum. Proc Natl Acad Sci USA1999;96:9333–8.
[20] Fernandez V, Treutiger CJ, Nash GB, Wahlgren M. Multiple adhesivephenotypes linked to rosetting binding of erythrocytes inPlasmodiumfalciparum malaria. Infect Immun 1998;66:2969–75.
[21] Udomsangpetch R, Wåhlin B, Carlson J, et al.Plasmod-ium falciparum-infected erythrocytes form spontaneous erythrocyterosettes. J Exp Med 1989;169:1835–40.
[22] Schlichtherle M, Wahlgren M, Perlmann H, Scherf A. Meth-ods in malaria research; 2000,http://www.malaria.mr4.org/MR4Protocols.htlm.
[23] Lambros C, Vanderberg JP. Synchronization ofPlasmodium falci-parum erythrocytic stages in culture. J Parasitol 1979;65:418–20.
[24] Chen Q, Barragan A, Fernandez V, et al. Identification ofPlasmod-ium falciparum erythrocyte membrane protein 1 (PfEMP1) as therosetting ligand of the malaria parasiteP. falciparum. J Exp Med1998;187:15–23.
[25] Udomsangpetch R, Lundgren K, Berzins K, et al. Human monoclonalantibodies to Pf155, a major antigen of malaria parasitePlasmodiumfalciparum. Science 1986;231:57–9.
[26] Ahlborg N, Berzins K, Perlmann P. Definition of the epitope rec-ognized by the human monoclonal antibody 33G2. Mol BiochemParasitol 1991;46:89–96.
[27] Hinterberg K, Scherf A, Gysin J, et al.Plasmodium falciparum:the Pf332 antigen is secreted from the parasite by a Brefeldin A-dependent pathway and is translocated to the erythrocyte membranevia the Maurer’s clefts. Exp Parasitol 1994;79:279–91.
[28] Taylor DW, Parra M, Chapman GB, et al. Localisation ofPlasmodiumfalciparumhistidine-rich protein 1 in the erythrocyte skeleton underknobs. Mol Biochem Parasitol 1987;25:165–74.
[29] Rock EP, Marsh K, Saul AJ, et al. Comparative analysis of thePlas-modium falciparumhistidine-rich proteins HRP-I, HRP-II and HRP-III in malaria parasites of diverse origin. Parasitology 1987;95:209–27.
[30] Howard RJ, Lyon JA, Uni S, et al. Transport of an Mr≈300,000Plasmodium falciparumprotein (PfEMP2) from the intraerythrocyticasexual parasite to the cytoplasmic face of the host cell membrane.J Cell Biol 1987;104:1269–80.
[31] Hall R, McBride J, Morgan G, et al. Antigens of the erythrocyticstages of the human malaria parasitePlasmodium falciparumdetectedby monoclonal antibodies. Mol Biochem Parasitol 1983;7:247–65.
[32] Hope IA, Hall R, Simmons DL, Hyde JE, Scaife JG. Evidence forimmunological cross-reaction between sporozoites and blood stagesof a human malaria parasite. Nature 1984;308:191–4.
[33] Simmons D, Woollett G, Bergin-Cartwright M, Kay D, Scaife J. Amalaria protein exported into a new compartment within the hosterythrocyte. EMBO J 1987;6:485–91.
[34] Barragan A, Spillmann D, Wahlgren M, Carlson J.Plasmodiumfalciparum: molecular background of strain specific rosette disruptionby glycosaminoglycans and sulfated glycoconjugates. Exp Parasitol1999;91:133–43.
[35] D , Scherf A. The Pf332 gene ofPlasmodium falciparumcodes fora giant protein that is translocated from the parasite to the membraneof infected erythrocytes. Gene 1992;1:71–9.
[36] Howard RJ, Uni S, Aikawa M, et al. Secretion of a malarial histidine-rich protein (PfHRPII) fromPlasmodium falciparum-infected ery-throcytes. J Cell Biol 1986;103:1269–77.
[37] Johnson D, Gunther K, Ansorge I, et al. Characterization of mem-brane proteins exported fromPlasmodium falciparuminto the hostcell erythrocyte. Parasitology 1994;109:1–9.
[38] Bennett BJ, Mohandas N, Coppel RL. Defining the minimal domainof the Plasmodium falciparumprotein MESA involved in the inter-action with the red cell membrane skeletal protein 4.1. J Biol Chem1997;272:15299–306.
[39] Kun JFJ, Waller KL, Coppel RL.Plasmodium falciparum: structuraland functional domains of the mature-parasite-infected erythrocytesurface antigen. Exp Parasitol 1999;91:258–67.
[40] Crabb B, Cooke B, Reeder J, et al. Targeted gene disruption showsthat knobs enable malaria-infected red cells to cytoadhere underphysiological shear stress. Cell 1997;89:287–96.
[41] Waterkeyn JG, Wickham ME, Davern KM, et al. Targeted muta-genesis ofPlasmodium falciparumerythrocyte membrane protein 3(PfEMP3) disrupts cytoadherence of malaria-infected red blood cells.EMBO J 2000;19:2813–23.
[42] Blisnick T, Morales Betoulle ME, Barale J-C, et al. Pfsbp1 a Maurer’scleft Plasmodium falciparumprotein, is associated with the erythro-cyte skeleton. Mol Biochem Parasitol 2000;111:107–21.
[43] Kilejian A. Characterization of a protein correlated with the pro-duction of knob-like protrusions on membranes of erythrocytesinfected with Plasmodium falciparum. Proc Natl Acad Sci USA1979;76:4650–3.
[44] Pologe LG, Pavlovec A, Shio H, Ravetch JV. Primary structure andsubcellular localization of the knob-associated histidine-rich proteinof Plasmodium falciparum. Proc Natl Acad Sci USA 1987;84:7139–43.
[45] Triglia T, Stahl HD, Crewther PE, et al. The complete sequence of thegene for the knob-associated histidine-rich protein fromPlasmodiumfalciparum. EMBO J 1987;6:1413–9.
[46] Waller KL, Cooke BM, Nunomura W, Mohandas N, Coppel RL.Mapping the binding domains involved in the interaction be-tween thePlasmodium falciparumknob-associated histidine-rich pro-tein (KAHRP) and the cytoadherence ligandP. falciparum ery-throcyte membrane protein 1 (PfEMP1). J Biol Chem 1999;274:23808–13.
[47] Oh SS, Voigt S, Fisher D, et al.Plasmodium falciparumerythrocytemembrane protein 1 is anchored to the actin–spectrin junction andknob-associated histidine-rich protein in the erythrocyte skeleton.Mol Biochem Parasitol 2000;108:237–47.
[48] Mattei D, Berry L, Couffin S, Richard O. The transport of thehistidine-rich protein I fromPlasmodium falciparumis insensitiveto Brefeldin A. Novartis Found Symp 1999;226:215–30.
[49] Wickham ME, Rug M, Ralph SA, et al. Trafficking and assemblyof the cytoadherence complex inPlasmodium falciparum-infectedhuman erythrocytes. EMBO J 2001;20:5636–49.
[50] Nacer A, Berry L, Slomianny C, Mattei D.Plasmodium falciparumsignal sequences: simply sequences or special signals? Int J Parasitol2001;31:1371–9.
[51] Pouvelle B, Spiegel R, Hsiao L, et al. Direct access to serummacromolecules by intraerythrocytic malaria parasites. Nature1991;353:73–5.
[52] Lauer S, Rathod P, Ghori N, Haldar K. A membrane network fornutrient import in red cells infected with the malaria parasite. Science1997;276:1122–5.
[53] Elmendorf H, Haldar K. Identification and localization of ERD2 inthe malaria parasitePlasmodium falciparum: separation from sitesof sphingomyelin synthesis and implications for organization of theGolgi. EMBO J 1993;12:4763–73.
[54] Wickert H, Wissing F, Andrews KT, Stich A, Krohne G, Lanzer M.Evidence for trafficking of PfEMP1 to the surface ofP. falciparum-infected erythrocytes via a complex membrane network. Eur J CellBiol 2003;82:271–84.
[55] Elmendorf H, Haldar K.Plasmodium falciparumexports the Golgimarker sphingomyelin synthase into a tubovesicular network in thecytoplasm of mature erythrocytes. J Cell Biol 1994;124:449–62.
Related Documents











