Effects of harmful dinoflagellate Ostreopsis cf. ovata exposure on immunological, histological and oxidative responses of mussels Mytilus galloprovincialis S. Gorbi a, * , G.C. Avio a , M. Benedetti a , C. Totti a , S. Accoroni a , S. Pichierri a , S. Bacchiocchi b , R. Orletti b , T. Graziosi b , F. Regoli a a Dipartimento di Scienze della Vita e dell’Ambiente, Università Politecnica delle Marche, Via Brecce Bianche, 60131 Ancona, Italy b Istituto Zooprofilattico Umbria e Marche, Via Cupa di Posatora, 3, 60100 Ancona, Italy article info Article history: Received 28 March 2013 Received in revised form 10 June 2013 Accepted 5 July 2013 Available online 13 July 2013 Keywords: Ostreopsis cf. ovata Immune response Haemocytes Antioxidant response Mytilus galloprovincialis abstract In the last decade massive blooms of the Ostreopsis cf. ovata have occurred in the Mediterranean basin, posing great concern to both environmental and human health. Biotoxicological and chemical studies demonstrated that O. cf. ovata produces palytoxin and ovatoxins; besides direct respiratory effects on humans due to inhalation of marine toxic aerosols, O. cf. ovata blooms can cause adverse effects on benthic invertebrates. The main aim of this study was to highlight the role of immunological, cellular and oxidative mech- anisms in modulating the toxicity induced by O. cf. ovata in mussels Mytilus galloprovincialis. Organisms were exposed in laboratory condition to O. cf. ovata and analysed after 7 and 14 days of exposure. Obtained results demonstrated a clear involvement of the immune system with a significant decre- ment of granulocytes respect to the hyalinocytes type cells, a diminished phagocytosis activity and a reduced lysosomal membrane stability in haemocytes, after both 7 and 14 days of exposure. Histological analyses showed a decrease of the digestive gland wall thickness, dilatation of the tubules, haemocytes infiltration into the digestive gland and a decrement of neutral lipid levels in exposed mussels; similar results suggest a possible inhibition of the feeding activity, with a consequent induction of authophagic phenomena and utilization of stored reserve products such as neutral lipids. Antioxidant parameters revealed a limited role of O. cf. ovata to induce oxidative stress in mussels exposed under laboratory conditions excepting for a certain increase of catalase, glutathione reductase and glutathione peroxidases activities, and a significantly higher capability to neutralize peroxyl radicals in mussels exposed for 14 days. Although the obtained results suggest a non-specific response of mussels to the O. cf. ovata exposure, observed effects on the general health status of exposed mussels should be adequately considered when assessing the ecological relevance of these algal blooms. Ó 2013 Elsevier Ltd. All rights reserved. 1. Introduction Toxic blooms of the benthic dinoflagellate Ostreopsis cf. ovata represent an emerging problem in the Mediterranean basin [1,2]. The first appearance along the Italian coasts occurred in the ’90s [3] and since then several blooms have been recorded in Ligurian Sea [4], Tyrrhenian Sea [5], Sicily [6] and Adriatic basin [7e9]. Recent investigations revealed that O. ovata species includes different strains having similar morphology; however the lack of molecular information and the relative uncertainty on the taxo- nomic status of this species suggested to use the designation O. cf. ovata when referring to isolates from the Mediterranean Sea [10]. Beside ecological implications, possibly leading to the mass mortality events of benthic organisms, O. cf. ovata blooms can also have adverse effects on human health do to inhalation of marine contaminated aerosol; moreover several economic losses are caused on local economies, when local Authorities prohibit bathing, fishing, and shellfish commerce during these algal blooms [8,11e 13]. The toxicity of O. cf. ovata has been ascribed to palytoxin-like compounds represented by more than 90% by ovatoxins and approximately 1% by a putative-palytoxin [4,14]. Due to the * Corresponding author. Tel.: þ39 0712204142; fax: þ39 0712204609. E-mail address: [email protected] (S. Gorbi). Contents lists available at SciVerse ScienceDirect Fish & Shellfish Immunology journal homepage: www.elsevier.com/locate/fsi 1050-4648/$ e see front matter Ó 2013 Elsevier Ltd. All rights reserved. http://dx.doi.org/10.1016/j.fsi.2013.07.003 Fish & Shellfish Immunology 35 (2013) 941e950

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

at SciVerse ScienceDirect
Fish & Shellfish Immunology 35 (2013) 941e950
Contents lists available
Fish & Shellfish Immunology
journal homepage: www.elsevier .com/locate / fs i
Effects of harmful dinoflagellate Ostreopsis cf. ovata exposure onimmunological, histological and oxidative responses of musselsMytilus galloprovincialis
S. Gorbi a,*, G.C. Avio a, M. Benedetti a, C. Totti a, S. Accoroni a, S. Pichierri a, S. Bacchiocchi b,R. Orletti b, T. Graziosi b, F. Regoli a
aDipartimento di Scienze della Vita e dell’Ambiente, Università Politecnica delle Marche, Via Brecce Bianche, 60131 Ancona, Italyb Istituto Zooprofilattico Umbria e Marche, Via Cupa di Posatora, 3, 60100 Ancona, Italy
a r t i c l e i n f o
Article history:Received 28 March 2013Received in revised form10 June 2013Accepted 5 July 2013Available online 13 July 2013
Keywords:Ostreopsis cf. ovataImmune responseHaemocytesAntioxidant responseMytilus galloprovincialis
* Corresponding author. Tel.: þ39 0712204142; faxE-mail address: [email protected] (S. Gorbi).
1050-4648/$ e see front matter � 2013 Elsevier Ltd.http://dx.doi.org/10.1016/j.fsi.2013.07.003
a b s t r a c t
In the last decade massive blooms of the Ostreopsis cf. ovata have occurred in the Mediterranean basin,posing great concern to both environmental and human health.
Biotoxicological and chemical studies demonstrated that O. cf. ovata produces palytoxin and ovatoxins;besides direct respiratory effects on humans due to inhalation of marine toxic aerosols, O. cf. ovatablooms can cause adverse effects on benthic invertebrates.
The main aim of this study was to highlight the role of immunological, cellular and oxidative mech-anisms in modulating the toxicity induced by O. cf. ovata in mussels Mytilus galloprovincialis. Organismswere exposed in laboratory condition to O. cf. ovata and analysed after 7 and 14 days of exposure.
Obtained results demonstrated a clear involvement of the immune system with a significant decre-ment of granulocytes respect to the hyalinocytes type cells, a diminished phagocytosis activity and areduced lysosomal membrane stability in haemocytes, after both 7 and 14 days of exposure.
Histological analyses showed a decrease of the digestive gland wall thickness, dilatation of the tubules,haemocytes infiltration into the digestive gland and a decrement of neutral lipid levels in exposedmussels; similar results suggest a possible inhibition of the feeding activity, with a consequent inductionof authophagic phenomena and utilization of stored reserve products such as neutral lipids. Antioxidantparameters revealed a limited role of O. cf. ovata to induce oxidative stress in mussels exposed underlaboratory conditions excepting for a certain increase of catalase, glutathione reductase and glutathioneperoxidases activities, and a significantly higher capability to neutralize peroxyl radicals in musselsexposed for 14 days.
Although the obtained results suggest a non-specific response of mussels to the O. cf. ovata exposure,observed effects on the general health status of exposed mussels should be adequately considered whenassessing the ecological relevance of these algal blooms.
� 2013 Elsevier Ltd. All rights reserved.
1. Introduction
Toxic blooms of the benthic dinoflagellate Ostreopsis cf. ovatarepresent an emerging problem in the Mediterranean basin [1,2].The first appearance along the Italian coasts occurred in the ’90s [3]and since then several blooms have been recorded in Ligurian Sea[4], Tyrrhenian Sea [5], Sicily [6] and Adriatic basin [7e9].
Recent investigations revealed that O. ovata species includesdifferent strains having similar morphology; however the lack of
: þ39 0712204609.
All rights reserved.
molecular information and the relative uncertainty on the taxo-nomic status of this species suggested to use the designation O. cf.ovata when referring to isolates from the Mediterranean Sea [10].
Beside ecological implications, possibly leading to the massmortality events of benthic organisms, O. cf. ovata blooms can alsohave adverse effects on human health do to inhalation of marinecontaminated aerosol;moreover several economic losses are causedon local economies,when local Authorities prohibit bathing,fishing,and shellfish commerce during these algal blooms [8,11e13].
The toxicity of O. cf. ovata has been ascribed to palytoxin-likecompounds represented by more than 90% by ovatoxins andapproximately 1% by a putative-palytoxin [4,14]. Due to the

S. Gorbi et al. / Fish & Shellfish Immunology 35 (2013) 941e950942
chemical similarity, molecular effects of ovatoxins were oftenassimilated to those exerted by palytoxins (PlTX) [15], including theinhibition of Naþ/Kþ-ATPase pump [16] and the increase ofinflammation-related proteins [17]. However several differencescan also be recognized, especially considering toxicological effectsat human level. The ingestion of fish and shellfish contaminated bypalytoxins have caused severe intoxications and deaths [18,19],while less toxic events characterized by skin irritation, respiratoryinfections and fever have been reported in humans exposed tomarine aerosol, during O. cf. ovata blooms [12]; no human intoxi-cation events have been yet ascribed to consumption of marineorganisms contaminated by ovatoxins.
At the present, the real impact of O. cf. ovata on benthic or-ganisms has not been clarified: although a causative effect of algalblooms has been hypothesized on mass mortality episodes [8,20],the main factors involved, such as the indirect role of mucilagemat and/or direct effects of the produced toxins, are stillunknown.
Studies on molecular and cellular effects of palytoxin-likecompounds produced by O. cf. ovata in mussels are scarce dueto the difficulty to obtain toxins from these algal cells [14], anddo not allowed to distinguish the different effects of putativepalytoxin and ovatoxins. Sublethal responses were recentlyinvestigated in mussels sampled from field condition during abloom occurred in the Northern Adriatic sea in 2009 [21]; themain results indicated a clear osmolregulatory unbalance inmussels with a significant inhibition of Naþ/Kþ ATPase andacetylcholinesterase activities, while more variable time- anddose-dependent alterations were suggested for the antioxidantand lipid metabolism.
The main aim of this study was to highlight the role of immu-nological, cellular and oxidative mechanisms in modulating thetoxicity of O. cf. ovata in mussels Mytilus galloprovincialis. In thisrespect, organisms were exposed in laboratory conditions to anenvironmentally realistic concentration of algal cells, isolated fromnorthern Adriatic Sea strain.
Accumulation of toxins in mussels was evaluated throughbiotoxicological methods, including the mouse bioassay (MBA)and the haemolytic neutralization assay (HNA). The MBA iscurrently adopted by European Sanitary Agencies for monitoringthe risk of algal toxins in shellfish, while the HNA is applied toreveal the presence of a retarded haemolytic capacity, a typicaleffect of PlTX-like compounds. The HNA has been recommendedby recent EFSA opinion [13] as possible alternative to the complexchemical methods for measurement of O. cf. ovata biotoxins[13,14], due to its specificity and the ban of Mouse Bioassay from2014 [22].
The activation of an immune strategy to counteract the toxiceffects of O. cf. ovata, was evaluated in terms of haemocytesmorphology and functionality as granulocytesehyalinocytes ratioand phagocytosis capacity [21,23]. Considering the sensitivity oflysosomal alterations as predictive biomarkers of adverse healtheffects [24], such responses were evaluated in terms of membranestability both in haemocytes and digestive cells; neutral lipidsaccumulation, lipofuscin in the tertiary lysosoms and epitheliadegeneration were further evaluated in mussels digestive gland.The possibility of O. cf. ovata to induce toxicological effects throughan oxidative pathway was assessed by the analyses of oxidativedisturbance biomarkers including both the study of individual an-tioxidants (catalase, glutathione reductase, glutathione S-trans-ferases, glutathione peroxidases, total glutathione) and themeasurement of total oxyradical scavenging capacity toward bothperoxyl and hydroxyl radicals.
The overall results of the present study were expected to pro-vide new insights on the toxicological mechanisms of O. cf. ovata
exposure on mussels, thus contributing to a better assessment ofecotoxicological risk of algal blooms.
2. Materials and methods
2.1. Algal cultures
Monoclonal cultures of Ostreopsis cf. ovata and Skeletonemamarinoi were obtained by isolating cells through the capillarypipette method from water samples collected along the Marchecoast (Italy, NW Adriatic Sea) in August 2007 and February 2010,respectively. After an initial growth in microplates, cells werecultured at 21 � 0.1 �C under a 12:12 h L:D photoperiod and anirradiance of 90e100 mmol m�2 s�1. The diatom S. marinoi wasused in the control as common species occurring in the phyto-plankton community of northern Adriatic Sea during winterblooms and used in aquaculture applications [25]. O. cf. ovatawas cultured in natural seawater with macronutrients added ata two-fold diluted f/2 concentration plus selenium, while forS. marinoi a replete f/2 growth medium was used.
O. cf. ovata was made identified by epifluorescence microscopeafter staining with Calcofluor-White M2R [26]. Previous molecu-lar analyses revealed that strains of O. cf. ovata isolated from thesame area belong to the Atlantic/Mediterranean clade [10], andpossess a high toxin content per cell, mostly characterized byovatoxin-a followed by ovatoxin-b and ovatoxin-d, whileovatoxin-c and putative palytoxin were present at very low levels[8]. One strain of O. cf. ovata and one of S. marinoi were used inthis experiment.
2.2. Mussel exposure
Mussels M. galloprovincialis (5 � 1 cm shell length) were ob-tained from a local farm (Ancona, Adriatic Sea); one hundred fifty(150) mussels were gently cleanedwith a brush to remove any algalcell from the shell, randomly distributed into ten 6-l glass-beakers(15 mussels per beaker), and acclimatised for 10 days to laboratoryconditions with filtered aerated seawater, 18 � 1 �C, salinity at 28e30, and daily fed with S. marinoi at 1 � 106 cells l�1. Five beakerswere used in this experiment for each of two treatments (Controland Exposed).
During the exposure period, control mussels were fed only withS. marinoi 1 � 106 cells l�1 (Ctrl), while exposed organisms withS. marinoi 1 � 106 cells l�1 and Ostreopsis cf. ovata 1 � 105 cells l�1
(Exp). Water renewal and microalgae additions were performeddaily. Before the beginning of the exposures and after 7 and 14 days(namely T0, T1 and T2 respectively) organisms from both controland exposed beakers were sampled.
At the three sampling times (T0, T1, T2), 2 specimens weresampled from each of the 5 beakers, for both Control andExposed groups to obtain approximately 100 g fresh tissues,immediately used to evaluate the presence of PlTX-like com-pounds with the mouse biological assay (MBA) and the hae-molysis neutralization assay (HNA). For immunological andantioxidant analyses, 5 replicates of haemolymph and digestivegland tissues were obtained collecting 3 specimens from eachbeaker at each sampling time. Haemolymph was withdrawnfrom the adductor muscle, partly frozen in liquid nitrogen andmaintained at �80 �C for the acetylcholinesterase analysis, andpartly used for the haemocyte analyses (i.e. granulocytesehya-linocytes ratio, phagocytosis rate, lysosomal membrane stabil-ity). Digestive glands were dissected and, before freezing thetissues at �80 �C, small pieces were excised from 5 specimensfor both control and exposed groups (1 from each beaker), and

S. Gorbi et al. / Fish & Shellfish Immunology 35 (2013) 941e950 943
frozen in hexane precooled to �70 �C in liquid nitrogen, forhistochemical analyses.
2.3. Toxin evaluation in mussels tissue
2.3.1. Mouse bioassay (MBA)Themouse biological assay (MBA) is themethod adopted by IZS-
National Veterinary Institutes to evaluate the presence of algaltoxins in bivalves for human consumption; the assay is based on theacute toxicity induced in adult mice after intraperitoneal injectionof specific extracts of bivalve molluscs. Lipophilic toxins wereextracted from an aliquot of 100 g of mussel tissues added with300 ml of acetone, homogenized for 2 min with an Ultraturrax andfiltered under reduced pressure; the filtrate was collected while theresidue was added to 300 ml of methanol (MeOH), and furtherhomogenized and filtered; the obtained filtrate was mixed to thefirst and the residue eliminated. The mixture was evaporated todryness and the residue dissolved in 30 ml of dichloromethane(CH2Cl2) and evaporated to dryness (Step 1); alternatively it wasextracted thricewith a double volume of aMeOH/H2O 60% solution,and adjusted to 200 ml of methanol before an aliquot of 16 ml wasevaporated to dryness (Step 2).
Both the dichloromethane (CH2Cl2) and methanolic evaporatedresidues were resuspended in 4 ml of 1% Tween-60 and for eachsolution 1 ml was intraperitoneally injected in three mice (AlbinoSwiss, 18e20 g b.w.). The death of two out of three micewithin 24 h(for the dichloromethane extract, Step 1) and within 5 h (for themethanolic extract, Step 2) is considered as a positive result for thefollowing toxins: okadaic acid and analogs, dinophysistoxins, pec-tenotoxins and azaspiracids (Step 1), yessotoxins and palytoxins(Step 2).
All mice manipulations were performed by the researchers ofthe IZS-National Veterinary Institute of Ancona, in accordance withthe UE Directive 86/609/EC, under official licence from NationalVeterinary Service.
2.3.2. Haemolysis neutralization assay (HNA)Haemolysis neutralization assay (HNA)was performed following
slight modification from previous methods [1]. Sheep erythrocyteswere separated from plasma by centrifugation, washed three timesin Dulbecco’s phosphate buffer (D-PBS), pH 7.0e7.2, with 1 mMCaCl2 0.5mMH3BO3, and finally diluted in D-PBS (1:100) to obtain afinal concentration of 1.7 � 107 erythrocytes/ml. Cells were incu-bated with methanolic resuspended residues (Step 2, see above) orwith palytoxin standard (0e15 mg/l Wako Chemicals GmbH, Ger-many) for 4 h at 37 �C.
The haemolytic activity was determined on supernatantsmeasuring absorbance at 405 nm. Blank values were obtained bymixing only erythrocytes with D-PBS solution while total (100%)haemolysis was achieved by adding distilled water to the cell sus-pension (1:1). Ouabain (C29H44O12, SIGMA O-3125) was used as aknown PlTX antagonist to demonstrate the presence of PlTX in thetested materials: for this reason all mussels extracts were alsotested on red blood cells pretreated for 30 min at 37 �C with 1 mMouabain.
2.4. Immunological analyses
Immunological responses were evaluated in terms of gran-ulocyteehyalinocytes ratio, phagocytosis capacity and lysosomalmembrane stability in haemocytes.
For the analysis of granulocyteehyalinocytes ratio, aliquotsof haemolymph were dispersed on glass slides and, afterdrying, fixed in Beker’s fixative (þ2.5% NaCl). The slides werewashed and stained with May Grunwald Giemsa before
mounting in glycerol gelatin. Observations were carried outwith a light microscope (1000�) and percentage of gran-ulocytes was evaluated after counting at least 200 cells foreach sample [21,27].
Phagocytosis capacity assay was performed following slightmodification from previous methods [28]; 100 ml of haemolymphwere dispersed on glass slides and allowed to adhere incubatingthem for 15 min at 15 �C in the dark. Fluorescein-labelledZymosan A bioparticles (Invitrogen) were added at 10:1 targe-t:haemocyte ratio. After 2 h incubation at 15 �C in the dark,uninternalized particles were removed by washing with physio-logical solution and slides were finally fixed in Beker’s fixative(þ2.5% NaCl) and mounted in glycerol gelatin. Phagocytosis wasexpressed as percentage of cells that internalized at least 3fluorescent particle (positive cells), observed under a fluorescencemicroscope, after counting at least 200 cells for each sample[21,27].
The cationic probe Neutral Red (NR) was used to evaluate thecapability of the lysosomal membranes to retain the dye [29]. Atleast 10 mussels were individually analysed for each sampling timeand treatment. Haemocytes were incubated on a glass slide with afreshly prepared NR working solution (2 ml/ml filtered seawater)from a stock solution of 20 mg neutral red dye dissolved in 1 ml ofdimethyl sulfoxide, and microscopically examined at 15 min in-tervals, to determine the time at which 50% of cells had lost to thecytosol the dye previously taken up by lysosomes.
2.5. Histological analyses
Cryostat sections (8 mm thick) of digestive gland were fixed inBeker’s fixative (þ2.5% NaCl) for 15 min, washed twice in 2.5% NaCl,then stained with haematoxylin-eosin and examined under a lightmicroscope.
The Latency Period (LP) of the lysosomal membranes wascytochemically measured on the digestive cells [30]. Serial cryostatsections (10 mm thick) were obtained from digestive glands of 5animals from each sampling time and treatment. Slides wereincubated in 0.1 M citrate buffer (pH 4.5) at 37 �C in a shaking waterbath for different time intervals (0, 3, 5, 10, 15, 20, 30 and 40 min).For visualization of lysosomal hexosaminidase, sections wereincubated for 20 min at 37 �C in 50 ml 0.1 M citrate buffer (pH 4.5)containing 20 mg naphthol AS-BI N-acetyl-b-glucosaminide(Sigma) dissolved in 2.5 ml 2-methoxyethanol, 2.5% NaCl (w:v) and3.5 g of low viscosity Polypep (Sigma) acting as a section stabilizer.Slides were washed twice in 3% NaCl, and an azo-coupling reactionwas performed for 9 min in 0.1% Fast Violet B salt dissolved in 0.1 Mphosphate buffer, pH 7.4. Slides were then rapidly rinsed in runningtap water for 5 min, fixed for 10 min in 10% formaldehyde con-taining 2% calcium acetate, before being mounted in glycerolgelatin. The determination of lysosomal membrane stability wasbased on the time required for acid labilization and thereforeproducing the maximum staining intensity and assessed under alight microscope.
Lipofuscin content of tertiary lysosomes was determined oncryostat sections (8 mm thick) of digestive glands [31]. Slides werefixed in Beker’s fixative (þ2.5% NaCl) and stained by Schmrol re-action before mounting in glycerol gelatin. For analyses of neutrallipids, cryostat sections (8 mm thick) were fixed as above andstained with the Oil Red O (ORO) method before mounting inglycerol gelatin [31]. For both lipofuscin and neutral lipids, fivemeasurements were made on digestive tubules of each section(two sections for mussel, 10 mussels for sampling period). Quan-tification of staining intensity was performed with Image-Pro� Plus6.2 Analysis Software and then normalized to the area of digestivetubules.

Table 1Results on Mouse Biological Assay (MBA) and Haemolysis Neutralization Assay(HNA) in mussels exposed to Ostreopsis cf. ovata for 7 and 14 days. þ þ þ: 3/3 micedead; � � �: no deaths.
Exposureconditions
MBA HNA
Step 1 Step 2 Ouabainsuppression
Type/Rapidity
T0 � � � � � � No NoneT1 Ctrl � � � þ � � No NoneT1 Exp � � � þ þ þ Yes DelayedT2 Ctrl � � � � � � No NoneT2 Exp � � � þ þ � Yes Delayed
S. Gorbi et al. / Fish & Shellfish Immunology 35 (2013) 941e950944
2.6. Antioxidant analyses
For the biochemical parameters (including enzymatic antioxi-dants, glutathione levels and total oxyradical scavenging capacity),the sample preparation and analytical methods have been reportedelsewhere [21]. For enzymatic antioxidants, samples of digestivegland were homogenized (1:5 w:v ratio) in 100 mM K-phosphatebuffer (pH 7.5), 0.1 mM phenylmethylsulphonyl fluoride (PMSF),0.1 mg/ml bacitracin, 0.008 TIU/ml aprotinin, 1 mg/ml leupeptin,0.5mg/mlpepstatin, NaCl 2.5%, and centrifuged at 110,000� g for 1 hat 4 �C. Measurements were made with a Varian (model Cary 3)spectrophotometer at a constant temperature of 18 �C. Catalase(CAT) was measured by the decrease in absorbance at 240 nm(extinction coefficient, 3¼ 0.04 mM�1 cm�1) due to the consump-tion of hydrogen peroxide, H2O2 (12 mM H2O2 in 100 mM K-phos-phate buffer pH 7.0). Glutathione reductase (GR) was determinedfromNADPHoxidationduring the reduction of oxidizedglutathione,GSSG (l ¼ 340 nm, 3¼ 6.22 mM�1 cm�1). The final assay conditionwas 100 mM K-phosphate buffer pH 7.0, 1 mM GSSG, and 60 mMNADPH. Glutathione peroxidases (GPx) activities were assayed in acoupled enzyme systemwhere NADPH is consumed by glutathionereductase to convert the formedGSSG to its reduced form (GSH). Thedecrease of absorbance was monitored at 340 nm( 3¼ 6.22 mM�1 cm�1) in 100 mM K-phosphate buffer pH 7.5, 1 mMEDTA,1 mMdithiothreitol, 2 mMGSH,1 unit glutathione reductase,0.24 mM NADPH, and 0.8 mM cumene hydroperoxide as substratesfor the sum of Se-dependent and Se-independent forms. The rate ofthe blank reactionwas subtracted from the total rate. Glutathione S-transferases (GST) were determined at 340 nm using 1-chloro-2,4-dinitrobenzene as substrate (CDNB). The assay was carried out in100 mM K-phosphate buffer pH 6.5, 1.5 mM CDNB, 1 mM GSH( 3¼ 9.6 mM�1 cm�1). Total glutathione was analysed in sampleshomogenized (1:5 w:v ratio) in 5% sulfosalicylic acid with 4 mMEDTA,maintained for 45min on ice and centrifuged at 37,000� g for15 min. The resulting supernatants were enzymatically assayed.
The total oxyradical scavenging capacity (TOSC) assaymeasure theoverall capability of cellular antioxidants to absorb different forms ofartificially generated oxyradicals, thus inhibiting the oxidation of0.2mMa-keto-g-methiolbutyric acid (KMBA) to ethylenegas. Peroxylradicals (ROO�)were generated by the thermalhomolysis of 20mM2-20-azo-bis-(2-methylpropionamidine)-dihydrochloride (ABAP) in100 mM K-phosphate buffer, pH 7.4. Hydroxyl radicals (�OH) wereproduced by the Fenton reaction of iron-EDTA (1.8 mM Fe3þ, 3.6 mMEDTA) plus ascorbate (180 mM) in 100 mM K-phosphate buffer.
Under these conditions the different oxyradicals produced quan-titatively similar yields of ethylene in control reactions, thus allowingto compare the relative efficiency of cellular antioxidants toward aquantitatively similar radical flux. Ethylene formation in control andsample reactions was analysed at 10e12 min time intervals by gas-chromatographic analyses and the TOSC values are quantified fromthe equation: TOSC¼ 100� (ʃSA/ʃCA� 100),where ʃSA and ʃCAare theintegrated areas calculated under the kinetic curves for samples (SA)and control (CA) reactions. For all the samples, a specific TOSC(normalized to content of protein) was calculated by dividing theexperimental TOSC values by the relative protein concentrationcontained in the assay. Levels of malondialdehyde (MDA) werespectrophotometrically measured after derivatization in 1-methyl-2-phenylindole with HCl and calibrated against a malondialdehydestandard curve. Protein concentrations were measured according toLowry method, using bovine serum albumin (BSA) as standard.
2.7. Statistical analyses
Analysis of variance (ANOVA) was applied for all of the analisedparameters to test differences between the experimental
conditions. Level of significance was set at p< 0.05, homogeneity ofvariance was checked by Cochram C and mathematical trans-formation applied if necessary; post-hoc comparison (NewmaneKeuls) was used to discriminate between means of values (n ¼ 5).The results of phagocytosis rate were analysed with the non-parametric KruskaleWallis test. Descriptive multivariate statisti-cal analysis (PCA, principal component analyses) was applied todiscriminate between different exposure conditions; multiplePearson correlations were performed between all measured vari-ables and significant relationship was assumed at the 0.05 level.
3. Results
The mouse biological assay revealed the accumulation of algaltoxins in tissues of mussels exposed to O. cf. ovata (Table 1). Thepositivity to PlTX-like compounds was expressed by the mortalitywithin 1 h of almost two of three mice intraperitoneally injectedwith the methanolic extracts obtained by the Step 2. The survival ofmice intraperitoneally injected with the dichloromethane extracts(Step 1), excluded the co-presence of other algal toxins in musseltissues (Table 1). Haemolytic activity was observed in all positivemussels sampled (Table 1); the ouabain suppression and thedelayed effect on erythrocytes haemolysis confirmed the specificityof PlTX-like compounds and the absence of other lipophilic toxinswhich usually induce a not complete ouabain suppression or a veryrapid haemolysis.
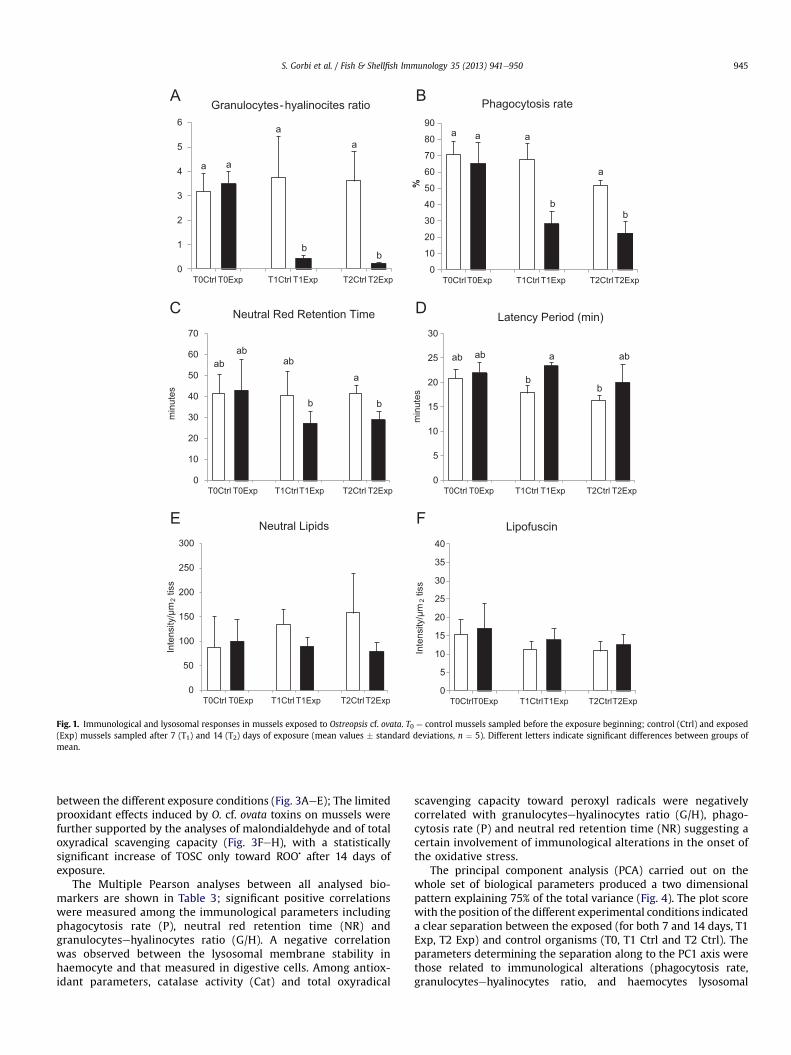
Immunological responses in exposed mussels revealed a sig-nificant decrement of granulocytes respect to the hyalinocytes typecells, a diminished phagocytosis activity and reduced lysosomalmembrane stability in haemocytes after both 7 and 14 days ofexposure (Fig. 1AeC). Lysosomal membrane stability was analysedalso in digestive gland where, contrarily to haemocytes, the latencyperiod showed a certain enhancement in mussels with a significantvariation after 7 days of O. cf. ovata exposure (Fig. 1D).
A limited not significant, decrease was observed for neutrallipids content in digestive gland of exposed organisms, while levelsof lipofuscins were more uniform among control and treatedgroups (Fig. 1E, F).
Histological alterations of digestive tissues were observed inmussels after 7 and 14 days of exposure to O. cf. ovata (Fig. 2 andTable 2). The predominant symptoms were the thinning of thedigestive tubules which appeared dilated in respect to the healthycondition. In most of the samples the tubules were disaggregatedand a general tissue degeneration was observed after 14 day ofexposure: haemocytes infiltration appeared into the digestivegland after both 7 and 14 days of exposure.
Variations of antioxidant defenses, lipid peroxidation levels andtotal oxyradical scavenging capacity (TOSC) are shown in Fig. 3AeH. Activities of catalase, glutathione reductase, glutathione perox-idases, glutathione S-transferases and levels of glutathione showedonly a few and usually not statistically significant variations
0
1
2
3
4
5
6
T0Ctrl T0Exp T1Ctrl T1Exp T2Ctrl T2Exp
Granulocytes-hyalinocites ratio
0
5
10
15
20
25
30
T0Ctrl T0Exp T1Ctrl T1Exp T2Ctrl T2Exp
min
utes
Latency Period (min)
0
50
100
150
200
250
300
T0Ctrl T0Exp T1Ctrl T1Exp T2Ctrl T2Exp
Inte
nsity
/µm
2tis
s
Neutral Lipids
0
10
20
30
40
50
60
70
80
90
T0CtrlT0Exp T1CtrlT1Exp T2CtrlT2Exp
%
Phagocytosis rate
0
10
20
30
40
50
60
70
T0Ctrl T0Exp T1CtrlT1Exp T2Ctrl T2Exp
min
utes
Neutral Red Retention Time
0
5
10
15
20
25
30
35
40
T0CtrlT0Exp T1CtrlT1Exp T2CtrlT2Exp
Inte
nsity
/µm
2tis
s
Lipofuscin
a
a
b
a
b
bb
a
aa
ab ab
b
a
b
ab
b
a
b
ab
A B
C D
E F
a
a
ab ab
Fig. 1. Immunological and lysosomal responses in mussels exposed to Ostreopsis cf. ovata. T0 ¼ control mussels sampled before the exposure beginning; control (Ctrl) and exposed(Exp) mussels sampled after 7 (T1) and 14 (T2) days of exposure (mean values � standard deviations, n ¼ 5). Different letters indicate significant differences between groups ofmean.
S. Gorbi et al. / Fish & Shellfish Immunology 35 (2013) 941e950 945
between the different exposure conditions (Fig. 3AeE); The limitedprooxidant effects induced by O. cf. ovata toxins on mussels werefurther supported by the analyses of malondialdehyde and of totaloxyradical scavenging capacity (Fig. 3FeH), with a statisticallysignificant increase of TOSC only toward ROO� after 14 days ofexposure.
The Multiple Pearson analyses between all analysed bio-markers are shown in Table 3; significant positive correlationswere measured among the immunological parameters includingphagocytosis rate (P), neutral red retention time (NR) andgranulocytesehyalinocytes ratio (G/H). A negative correlationwas observed between the lysosomal membrane stability inhaemocyte and that measured in digestive cells. Among antiox-idant parameters, catalase activity (Cat) and total oxyradical
scavenging capacity toward peroxyl radicals were negativelycorrelated with granulocytesehyalinocytes ratio (G/H), phago-cytosis rate (P) and neutral red retention time (NR) suggesting acertain involvement of immunological alterations in the onset ofthe oxidative stress.
The principal component analysis (PCA) carried out on thewhole set of biological parameters produced a two dimensionalpattern explaining 75% of the total variance (Fig. 4). The plot scorewith the position of the different experimental conditions indicateda clear separation between the exposed (for both 7 and 14 days, T1Exp, T2 Exp) and control organisms (T0, T1 Ctrl and T2 Ctrl). Theparameters determining the separation along to the PC1 axis werethose related to immunological alterations (phagocytosis rate,granulocytesehyalinocytes ratio, and haemocytes lysosomal
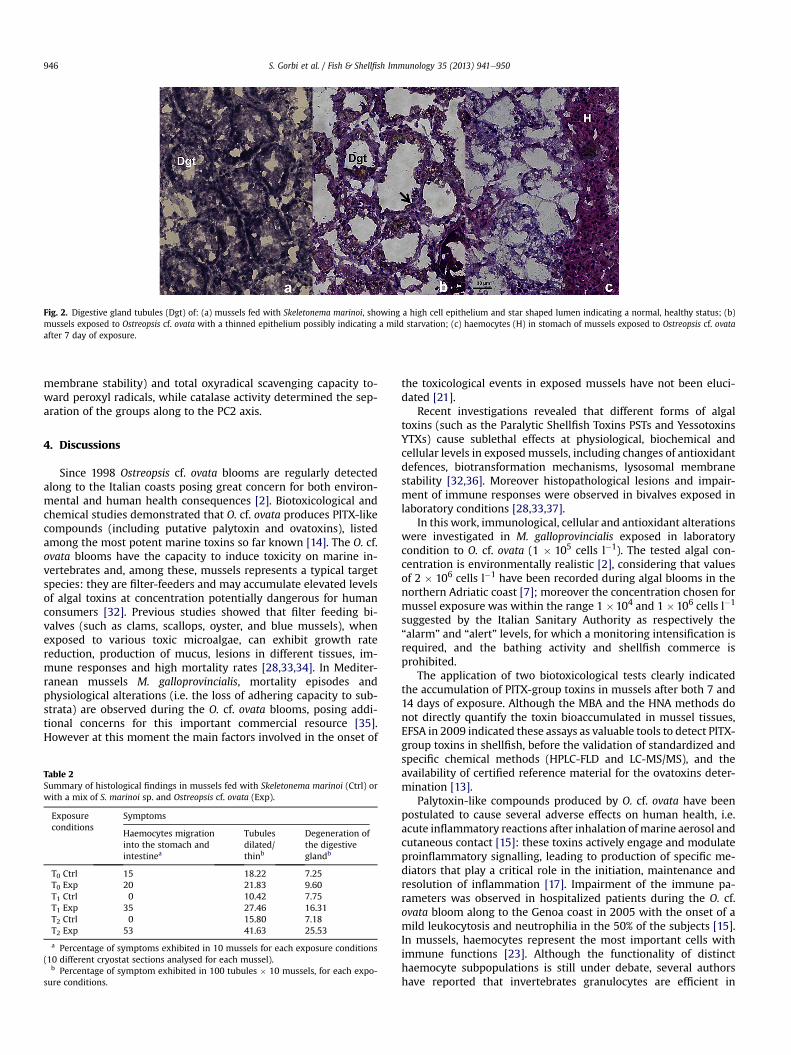
Fig. 2. Digestive gland tubules (Dgt) of: (a) mussels fed with Skeletonema marinoi, showing a high cell epithelium and star shaped lumen indicating a normal, healthy status; (b)mussels exposed to Ostreopsis cf. ovata with a thinned epithelium possibly indicating a mild starvation; (c) haemocytes (H) in stomach of mussels exposed to Ostreopsis cf. ovataafter 7 day of exposure.
S. Gorbi et al. / Fish & Shellfish Immunology 35 (2013) 941e950946
membrane stability) and total oxyradical scavenging capacity to-ward peroxyl radicals, while catalase activity determined the sep-aration of the groups along to the PC2 axis.
4. Discussions
Since 1998 Ostreopsis cf. ovata blooms are regularly detectedalong to the Italian coasts posing great concern for both environ-mental and human health consequences [2]. Biotoxicological andchemical studies demonstrated that O. cf. ovata produces PlTX-likecompounds (including putative palytoxin and ovatoxins), listedamong the most potent marine toxins so far known [14]. The O. cf.ovata blooms have the capacity to induce toxicity on marine in-vertebrates and, among these, mussels represents a typical targetspecies: they are filter-feeders and may accumulate elevated levelsof algal toxins at concentration potentially dangerous for humanconsumers [32]. Previous studies showed that filter feeding bi-valves (such as clams, scallops, oyster, and blue mussels), whenexposed to various toxic microalgae, can exhibit growth ratereduction, production of mucus, lesions in different tissues, im-mune responses and high mortality rates [28,33,34]. In Mediter-ranean mussels M. galloprovincialis, mortality episodes andphysiological alterations (i.e. the loss of adhering capacity to sub-strata) are observed during the O. cf. ovata blooms, posing addi-tional concerns for this important commercial resource [35].However at this moment the main factors involved in the onset of
Table 2Summary of histological findings in mussels fed with Skeletonema marinoi (Ctrl) orwith a mix of S. marinoi sp. and Ostreopsis cf. ovata (Exp).
Exposureconditions
Symptoms
Haemocytes migrationinto the stomach andintestinea
Tubulesdilated/thinb
Degeneration ofthe digestiveglandb
T0 Ctrl 15 18.22 7.25T0 Exp 20 21.83 9.60T1 Ctrl 0 10.42 7.75T1 Exp 35 27.46 16.31T2 Ctrl 0 15.80 7.18T2 Exp 53 41.63 25.53
a Percentage of symptoms exhibited in 10 mussels for each exposure conditions(10 different cryostat sections analysed for each mussel).
b Percentage of symptom exhibited in 100 tubules � 10 mussels, for each expo-sure conditions.
the toxicological events in exposed mussels have not been eluci-dated [21].
Recent investigations revealed that different forms of algaltoxins (such as the Paralytic Shellfish Toxins PSTs and YessotoxinsYTXs) cause sublethal effects at physiological, biochemical andcellular levels in exposedmussels, including changes of antioxidantdefences, biotransformation mechanisms, lysosomal membranestability [32,36]. Moreover histopathological lesions and impair-ment of immune responses were observed in bivalves exposed inlaboratory conditions [28,33,37].
In this work, immunological, cellular and antioxidant alterationswere investigated in M. galloprovincialis exposed in laboratorycondition to O. cf. ovata (1 � 105 cells l�1). The tested algal con-centration is environmentally realistic [2], considering that valuesof 2 � 106 cells l�1 have been recorded during algal blooms in thenorthern Adriatic coast [7]; moreover the concentration chosen formussel exposure was within the range 1 �104 and 1 �106 cells l�1
suggested by the Italian Sanitary Authority as respectively the“alarm” and “alert” levels, for which a monitoring intensification isrequired, and the bathing activity and shellfish commerce isprohibited.
The application of two biotoxicological tests clearly indicatedthe accumulation of PlTX-group toxins in mussels after both 7 and14 days of exposure. Although the MBA and the HNA methods donot directly quantify the toxin bioaccumulated in mussel tissues,EFSA in 2009 indicated these assays as valuable tools to detect PlTX-group toxins in shellfish, before the validation of standardized andspecific chemical methods (HPLC-FLD and LC-MS/MS), and theavailability of certified reference material for the ovatoxins deter-mination [13].
Palytoxin-like compounds produced by O. cf. ovata have beenpostulated to cause several adverse effects on human health, i.e.acute inflammatory reactions after inhalation of marine aerosol andcutaneous contact [15]: these toxins actively engage and modulateproinflammatory signalling, leading to production of specific me-diators that play a critical role in the initiation, maintenance andresolution of inflammation [17]. Impairment of the immune pa-rameters was observed in hospitalized patients during the O. cf.ovata bloom along to the Genoa coast in 2005 with the onset of amild leukocytosis and neutrophilia in the 50% of the subjects [15].In mussels, haemocytes represent the most important cells withimmune functions [23]. Although the functionality of distincthaemocyte subpopulations is still under debate, several authorshave reported that invertebrates granulocytes are efficient in

0
200
400
600
800
1000
1200
T0Ctrl T0Exp T1Ctrl T1Exp T2Ctrl T2Exp
UTo
sc/m
g pr
ot
TOSC (•OOR)
0
100
200
300
400
500
600
T0Ctrl T0Exp T1Ctrl T1Exp T2Ctrl T2Exp
Uto
sc/m
g pr
ot
TOSC (•OH)
0102030405060708090
100
T0Ctrl T0Exp T1Ctrl T1Exp T2Ctrl T2Exp
nmol
/g ti
ss
MDA
0
5
10
15
20
25
30
35
40
T0Ctrl T0Exp T1Ctrl T1Exp T2Ctrl T2Exp
µmol
/min
/mg
prot
Catalase
0
10
20
30
40
50
60
70
T0Ctrl T0Exp T1Ctrl T1Exp T2Ctrl T2Exp
nmol
/min
/mg
prot
Glutathione Reductase
0
5
10
15
20
25
30
T0Ctrl T0Exp T1Ctrl T1Exp T2Ctrl T2Exp
nmol
/min
/mg
prot
Glutathione Peroxidases
020406080
100120140160180
T0Ctrl T0Exp T1Ctrl T1Exp T2Ctrl T2Expnm
ol/m
in/m
g pr
ot
Glutathione S-transferases
0
0,2
0,4
0,6
0,8
1
1,2
1,4
1,6
T0Ctrl T0Exp T1Ctrl T1Exp T2Ctrl T2Exp
µmol
/g ti
ss
Total Glutathione
ab abab
b
a
a aa
a
b
A B
C D
E F
G H
a
a
Fig. 3. Antioxidant defenses, malondialdehyde levels and total oxyradical scavenging capacity (TOSC) toward peroxyl (ROO�) and hydroxyl (HO�) radicals in mussels exposed toOstreopsis cf. ovata. T0 ¼ control mussels sampled before the exposure beginning; control (Ctrl) and exposed (Exp) mussels sampled after 7 (T1) and 14 (T2) days of exposure (meanvalues � standard deviations, n ¼ 5). Different letters indicate significant differences between groups of mean.
S. Gorbi et al. / Fish & Shellfish Immunology 35 (2013) 941e950 947
phagocytosis [38,39], while hyalinocytes, which present charac-teristics of undifferentiated cells, would be specialized in coagula-tion and encapsulation processes [23].
One of the main evidence of this work is related to the strongdecrement of circulating granulocytes and phagocytosis capacity inhemolymph of exposedmussels. These effects were already evidentafter 7 days of exposure, corroborating the hypothesis that immunesystems is one of the first targets of algal toxins, also in marineorganisms. A decrease of granulocyte cells was demonstrated inhaemolymph of mussels Mytilus edulis exposed to the toxic Karlo-dinium veneficum, suggesting the movement of these cells from the
circulating systems into the digestive tissues, in order to preventand counteract damages possibly due to the ingestion of the toxicalgae [33]. A decline in haemocyte complexity was revealed inmussels exposed to the toxic dinoflagellates Prorocentrum mini-mum, indicating the hemocyte degranulation processes as part ofthe immune response [28].
In oyster, Crassostrea virginica, and scallop, Argopecten irradiansexposed to P. minimum the percentage of granulocytes decreased atthe very beginning of the treatment, but increased after a few days[37]. Similarly a certain modulation of granulocytes percentage wasconfirmed in hemolymph of mussels exposed to O. cf. ovata in field
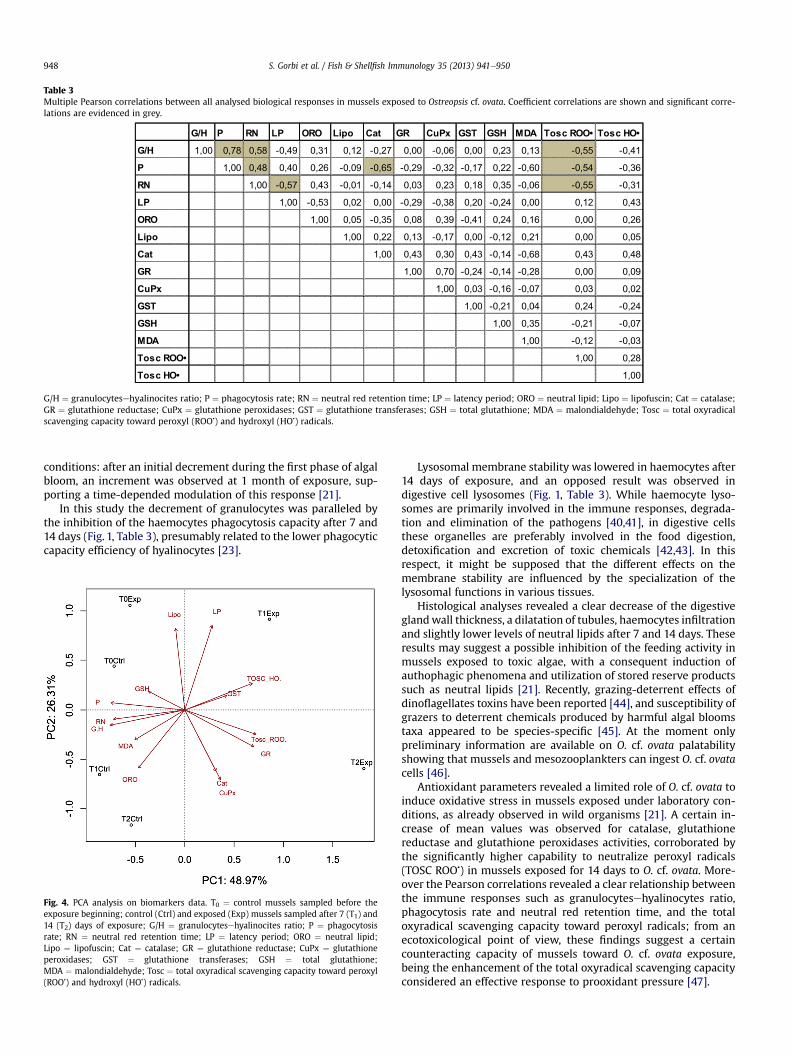
Table 3Multiple Pearson correlations between all analysed biological responses in mussels exposed to Ostreopsis cf. ovata. Coefficient correlations are shown and significant corre-lations are evidenced in grey.
G/H ¼ granulocytesehyalinocites ratio; P ¼ phagocytosis rate; RN ¼ neutral red retention time; LP ¼ latency period; ORO ¼ neutral lipid; Lipo ¼ lipofuscin; Cat ¼ catalase;GR ¼ glutathione reductase; CuPx ¼ glutathione peroxidases; GST ¼ glutathione transferases; GSH ¼ total glutathione; MDA ¼ malondialdehyde; Tosc ¼ total oxyradicalscavenging capacity toward peroxyl (ROO�) and hydroxyl (HO�) radicals.
S. Gorbi et al. / Fish & Shellfish Immunology 35 (2013) 941e950948
conditions: after an initial decrement during the first phase of algalbloom, an increment was observed at 1 month of exposure, sup-porting a time-depended modulation of this response [21].
In this study the decrement of granulocytes was paralleled bythe inhibition of the haemocytes phagocytosis capacity after 7 and14 days (Fig. 1, Table 3), presumably related to the lower phagocyticcapacity efficiency of hyalinocytes [23].
Fig. 4. PCA analysis on biomarkers data. T0 ¼ control mussels sampled before theexposure beginning; control (Ctrl) and exposed (Exp) mussels sampled after 7 (T1) and14 (T2) days of exposure; G/H ¼ granulocytesehyalinocites ratio; P ¼ phagocytosisrate; RN ¼ neutral red retention time; LP ¼ latency period; ORO ¼ neutral lipid;Lipo ¼ lipofuscin; Cat ¼ catalase; GR ¼ glutathione reductase; CuPx ¼ glutathioneperoxidases; GST ¼ glutathione transferases; GSH ¼ total glutathione;MDA ¼ malondialdehyde; Tosc ¼ total oxyradical scavenging capacity toward peroxyl(ROO�) and hydroxyl (HO�) radicals.
Lysosomal membrane stability was lowered in haemocytes after14 days of exposure, and an opposed result was observed indigestive cell lysosomes (Fig. 1, Table 3). While haemocyte lyso-somes are primarily involved in the immune responses, degrada-tion and elimination of the pathogens [40,41], in digestive cellsthese organelles are preferably involved in the food digestion,detoxification and excretion of toxic chemicals [42,43]. In thisrespect, it might be supposed that the different effects on themembrane stability are influenced by the specialization of thelysosomal functions in various tissues.
Histological analyses revealed a clear decrease of the digestiveglandwall thickness, a dilatation of tubules, haemocytes infiltrationand slightly lower levels of neutral lipids after 7 and 14 days. Theseresults may suggest a possible inhibition of the feeding activity inmussels exposed to toxic algae, with a consequent induction ofauthophagic phenomena and utilization of stored reserve productssuch as neutral lipids [21]. Recently, grazing-deterrent effects ofdinoflagellates toxins have been reported [44], and susceptibility ofgrazers to deterrent chemicals produced by harmful algal bloomstaxa appeared to be species-specific [45]. At the moment onlypreliminary information are available on O. cf. ovata palatabilityshowing that mussels and mesozooplankters can ingest O. cf. ovatacells [46].
Antioxidant parameters revealed a limited role of O. cf. ovata toinduce oxidative stress in mussels exposed under laboratory con-ditions, as already observed in wild organisms [21]. A certain in-crease of mean values was observed for catalase, glutathionereductase and glutathione peroxidases activities, corroborated bythe significantly higher capability to neutralize peroxyl radicals(TOSC ROO�) in mussels exposed for 14 days to O. cf. ovata. More-over the Pearson correlations revealed a clear relationship betweenthe immune responses such as granulocytesehyalinocytes ratio,phagocytosis rate and neutral red retention time, and the totaloxyradical scavenging capacity toward peroxyl radicals; from anecotoxicological point of view, these findings suggest a certaincounteracting capacity of mussels toward O. cf. ovata exposure,being the enhancement of the total oxyradical scavenging capacityconsidered an effective response to prooxidant pressure [47].

S. Gorbi et al. / Fish & Shellfish Immunology 35 (2013) 941e950 949
The PCA analyses applied on the whole set of biomarkers pro-vided a clear separation between control and exposed mussels:particularly useful in this discrimination on the PC1 appeared theimmunological responses (i.e. granulocytesehyalinocytes ratio,phagocytosis rate and neutral red retention time) and level of totaloxyradical scavenging capacity toward peroxyl radical in musselsexposed for 14 days. Among the two times of exposure, catalaseactivity, one of the most sensible antioxidant responses, was usefulto distinguish between 7 and 14 days of exposure along to the PC2.
5. Conclusions
The changes at immunological, histological and oxidative levelsobserved in mussels exposed to O. cf. ovata revealed that the im-mune system is clearly stimulated as confirmed by the anomalousdecrement of granulocytes in haemolymph and the accumulationof haemocytes in the digestive gland. Moreover a general thinningand disaggregation of digestive tubules and lower levels of neutrallipids suggest that O. cf. ovata can influence feeding behaviour ofmussels, possibly leading to autophagic phenomena and con-sumption of stored reserves. The O. cf. ovata exposure induced alimited prooxidant pressure, evidenced after 14 days of exposure bya significant enhancement of oxyradical scavenging capacity to-ward peroxyl radicals.
Overall these biological effects suggest a non-specific responseto O. cf. ovata in mussels, with effects similar to those previouslydescribed in other bivalves exposed to various harmful algal toxins[28,33,37]. Obtained results further support the necessity that thegeneral health status of mussels is adequately considered whenassessing the ecological relevance of O. cf. ovata algal blooms inMediterranean.
References
[1] Aligizaki K, Katikou P, Nikolaidis G, Panou A. First episode of shellfishcontamination by palytoxin-like compounds from Ostreopsis species (AegeanSea, Greece). Toxicon 2008;51(3):418e27.
[2] Mangialajo L, Ganzin N, Accoroni S, Asnaghi V, Blanfunè A, Cabrini M, et al.Trends in Ostreopsis proliferation along the Northern Mediterranean coasts.Toxicon 2011;57:408e20.
[3] Tognetto L, Bellato S, Moro I, Andreoli C. Occurrence of Ostreopsis ovata(Dinophyceae) in the Tyrrhenian Sea during summer 1994. Bot Mar 1995;38:291e5.
[4] Ciminiello P, Dell’Aversano C, Fattorusso E, Forino M, Tartaglione L, Grillo C,et al. Putative palytoxin and its new analogue, ovatoxin-a, in Ostreopsis ovatacollected along the Ligurian coasts during the 2006 toxic outbreak. J Am SocMass Spectrom 2008;19(1):111e20.
[5] Bianco I, Sangiorgi VC, Penna A, Guerrini F, Pistocchi R, Zaottini E, et al.Ostreopsis ovata in benthic aggregates along the Latium coast (middleTyrrhenian Sea). In: International symposium on algal toxins, Trieste 2007.p. 27e9.
[6] Barone R, Prisinzano A. Peculiarità comportamentale del dinoflagellatoOstreopsis ovata Fukuyo (Dinophyceae) la strategia del ragno. Naturalista Sicil2006;30:401e18.
[7] Totti C, Accoroni S, Cerino F, Cucchiari E, Romagnoli T. Ostreopsis ovata bloomalong the Conero Riviera (northern Adriatic Sea): relationships with envi-ronmental conditions and substrata. Harmful Algae 2010;9:233e9.
[8] Accoroni S, Romagnoli T, Colombo F, Pennesi C, Di Camillo CG, Marini M, et al.Ostreopsis cf. ovata bloom in the northern Adriatic Sea during summer 2009:ecology, molecular characterization and toxin profile. Mar Pollut Bull 2011;62:2512e9.
[9] Accoroni S, Colombo F, Pichierri S, Romagnoli T, Marini M, Battocchi C, et al.Ecology of Ostreopsis cf. ovata blooms in the northwestern Adriatic Sea.Cryptogamie Algol 2012;33:191e8.
[10] Penna A, Fraga S, Battocchi C, Casabianca S, Riobò P, Giacobbe MG, et al.A phylogeography study of the toxic benthic genus Ostreopsis Schmidt.J Biogeogr 2010;37:830e41.
[11] Amzil Z, Sibat M, Chomerat N, Grossel H, Marco-Miralles F, Lemee R, et al.Ovatoxin-a and palytoxin accumulation in seafood in relation to Ostreopsis cf.ovata blooms on the French Mediterranean coast. Mar Drugs 2012;10(2):477e96.
[12] Gallitelli M, Ungaro N, Addante LM, Procacci V, Silver NG, Sabbà C. Respiratoryillness as a reaction to tropical algal blooms occurring in a temperate climate.J Am Med Assoc 2005;293(21):2595e600.
[13] EFSA Panel on Contaminants in the Food Chain (CONTAM). Scientific opinionon marine biotoxins in shellfish e palytoxin group. EFSA J 2009;7(12):1393.http://dx.doi.org/10.2903/j.efsa.2009.1393 [38 pp.]. Available online: http://www.efsa.europa.eu.
[14] Ciminiello P, Dell’Aversano C, Dello Iacovo E, Fattorusso E, Forino M,Tartaglione L, et al. Unique toxin profile of a Mediterranean Ostreopsis cf. ovatastrain: HR LC-MSn characterization of Ovatoxin-f, a new palytoxin congener.Chem Res Toxicol 2012;25(6):1243e52.
[15] Tubaro A, Durando P, Del Favero G, Ansaldi F, Icardi G, Deeds JR, et al. Casedefinitions for human poisonings postulated to palytoxins exposure. Toxicon2011;57:478e95.
[16] Rossini GP, Bigiani A. Palytoxin action on the Naþ, Kþ-ATPase and thedisruption of ion equilibria in biological systems. Toxicon 2011;7:429e39.
[17] Crinelli R, Carloni E, Giacomini E, Penna A, Dominici S, Battocchi C, et al.Palytoxin and an ostreopsis toxin extract increase the levels of mRNAsencoding inflammation-related proteins in human macrophages via p38MAPK and NF-kB. PLoS One 2012;7(6) art. no. e38139.
[18] Alcala AC, Alcala LC, Garth JS, Yasumura D, Yasumoto T. Hauman fatality dueto ingestion of the crab Demania reynaudii that contained a palytoxin-liketoxin. Toxicon 1988;26:105e7.
[19] Taniyama S, Mahmud Y, Terada M, Takatani T, Arakawa O, Noguchi T.Occurrence of a food poisoning incident by palytoxin from a serranid Epi-nephelus sp. in Japan. J Nat Toxins 2002;11(4):277e82.
[20] Sansoni G, Borghini B, Camici G, Casatti M, Righini P, Rustighi C. Fioriture algalidi Ostreopsis ovata (Gonyaulacales: Dinophyceae): un problema emergente.Biol Ambient 2003;17(1):17e23.
[21] Gorbi S, Bocchetti R, Binelli A, Bacchiocchi S, Orletti R, Nanetti L, et al. Bio-logical effects of palytoxin-like compounds from Ostreopsis cf. ovata: a mul-tibiomarkers approach with mussels Mytilus galloprovincialis. Chemosphere2012;89(5):623e32.
[22] Commission Regulation (EU) No 15/2011 of 10 January 2011 amendingRegulation (EC) No 2074/2005 as regards recognised testing methods fordetecting marine biotoxins in live bivalve mollusks.
[23] García-García E, Prado-Álvarez M, Novoa B, Figueras A, Rosales C. Immuneresponses of mussel hemocyte subpopulations are differentially regulated byenzymes of the PI 3-K, PKC, and ERK kinase families. Dev Comp Immunol2008;32:637e53.
[24] Hegseth MN, Regoli F, Gorbi S, Bocchetti R, Gabrielsen GW, Camus L. Lyso-somal and lipid-associated parameters in the livers of three species of arcticseabird chicks: species differences and relationships with contaminant levels.Mar Pollut Bull 2011;62(8):1652e60.
[25] Totti C, Cangini M, Ferrari C, Kraus R, Pompei M, Pugnetti A, et al. Phyto-plankton size-distribution and community structure in relation to mucilageoccurrence in the northern Adriatic Sea. Sci Total Environ 2005;353:204e17.
[26] Steidinger K, Tangen K. Dinoflagellates. In: Tomas CR, editor. Identifyingmarine diatoms and dinoflagellates. Academic Press; 1997. p. 387e584.
[27] Ciacci C, Barmo C, Fabbri R, Canonico B, Gallo G, Canesi L. Immunomodulationin Mytilus galloprovincialis by non-toxic doses of hexavalent Chromium. FishShellfish Immunol 2011;31:1026e33.
[28] Galimany E, Sunila I, Hégaret H, Ramon M, Wikfors GH. Pathology andimmune response of the blue mussel (Mytilus edulis L.) after an exposure tothe harmful dinoflagellate Prorocentrum minimum. Harmful Algae 2008;7:630e8.
[29] Lowe DM, Fossato VU, Depledge MH. Contaminant induced lysosomal mem-brane damage in blood cells of mussels Mytilus galloprovincialis from theVenice Lagoon: an in vitro study 1995;129:189e96.
[30] Moore MN, Lowe D, Köhler A. In: ICES techniques in marine environmentalsciences. Biological effects of contaminants: measurement of lysosomalmembrane stability, vol. 36. ICES; 2004. p. 31.
[31] Moore MN. Cytochemical responses of the lysosomal system and NADPH-ferrihemoprotein reductase in molluscan digestive cells to environmentaland experimental exposure to xenobiotics. Mar Ecol Prog Ser 1988;46:81e9.
[32] Buratti S, Franzellitti S, Poletti R, Ceredi A, Montanari G, Capuzzo A, et al.Bioaccumulation of algal toxins and changes in physiological parameters inMediterranean mussels from the North Adriatic Sea (Italy). Environ Toxicol2013;28:451e70.
[33] Galimany E, Place AR, Ramón M, Jutson M, Pipe RK. The effects of feedingKarlodinium veneficum (PLY # 103; Gymnodinium veneficum Ballantine) to theblue mussel Mytilus edulis. Harmful Algae 2008;7(1):91e8.
[34] Wikfors GH. A review and new analysis of trophic interactions betweenProrocentrum minimum and clams, scallops, and oysters. Harmful Algae2005;4:585e92.
[35] FAO con dati riferiti all’anno. http://www.fao.org/fishery/countrysector/naso_italy/en; 2004.
[36] Haberkorn H, Lambert C, Le Goïc N, Moal J, Suquet M, Guéguen M, et al. Effectsof Alexandrium minutum exposure on nutrition-related processes and repro-ductive output in oysters Crassostrea gigas. Aquat Toxicol 2010;97:96e108.
[37] Hégaret H, Wikfors GH. Time-dependent changes in hemocytes of easternoysters, Crassostrea virginica, and northern bay scallops, Argopecten irradiansirradians, exposed to a cultured strain of Prorocentrum minimum. HarmfulAlgae 2005;4(2):187e99.
[38] Carballal MJ, Lopez MC, Azevedo C, Villalba A. Hemolymph cell types of themussel Mytilus galloprovincialis. Dis Aquat Org 1997;29:127e35.
[39] Giulianini PG, Bertolo F, Battistella S, Amirante GA. Ultrastructure of the he-mocytes of Cetonischema aeruginosa larvae (Coleoptera, Scarabaeidae):

S. Gorbi et al. / Fish & Shellfish Immunology 35 (2013) 941e950950
involvement of both granulocytes and oenocytoids in vivo phagocytosis.Tissue Cell 2003;35:243e51.
[40] Pryor SC, Facher E. Immune response in mussels to environmental pollution.Am Biol Teach 1997;59:328e30.
[41] Canesi L, Gallo G, Gavioli M, Pruzzo C. Bacteria-hemocyte interactions andphagocytosis in marine bivalves. Microsc Res Tech 2002;57(6):469e76.
[42] Garmendia L, Izagirre U, Cajaraville MP, Marigómez I. Application of a batteryof biomarkers in mussel digestive gland to assess long-term effects of thePrestige oil spill in Galicia and the Bay of Biscay: lysosomal responses.J Environ Monit 2011;13:901e14.
[43] Marigómez I, Lekube X, Cajaraville MP, Domouhtsidou G, Dimitriadis V.Comparison of cytochemical procedures to estimate lysosomal biomarkers inmussel digestive cells. Aquat Toxicol 2005;75(1):86e95.
[44] Hong J, Talapatra S, Katz J, Tester PA, Waggett RJ, Place AR. Algal toxins altercopepod feeding behaviour. PLoS One 2012;7(5) art. no. e36845.
[45] Landsberg JH. The effects of harmful algal blooms on aquatic organisms. RevFish Sci 2002;10(2):113e390.
[46] Furlan M, Antonioli M, Zingone A, Sardo A, Blason C, Pallavicini A, et al. Mo-lecular identification of Ostreopsis cf. ovata in filter feeders and putativepredators. Harmful Algae 2013;21e22:20e9.
[47] Monserrat J, Letts RE, Ribas Ferreira JL, Ventura-Lima J, Amado LL, Rocha AM,et al. Biomarkers of oxidative stress: benefits and drawbacks for their appli-cation in biomonitoring of aquatic environments. In: Abele Doris, Vazquesz-Medina Josè Pablo, Zenteno-Savin Tania, editors. Oxidative stress in aquaticecosystems. Wiley-Blackwell A John Wiley & Sons, Ltd. Publication; 2011.p. 317e26.
Related Documents











