THE JOURNAL OF BIOLOGICAL CHEMISTRY Vol. 267, No. 15, Issue of May 25, pp. 10925-10930,1992 Q 1992 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in U. S. A. Dietary Regulation of Stearoyl-CoA Desaturase 1 Gene Expression in Mouse Liver* (Received for publication, August 7, 1991) James M. NtambiS From the Department of Biochemistry and Molecular Biology, the Georgetown University School of Medicine, Washington, D. C. 20007 Expression of the murine liver stearoyl-CoA desat- urase gene (SCD1) is induced upon feeding fasted mice a fat-free, high carbohydrate diet (Ntambi, J. M., Buhrow, S. A., Kaestner, K. H., Christy, R. J., Sibley, E., Kelly, T. J., and Lane, M. D. (1988) J. Bid. Chern. 263, 17291-17300). In the present study, SCD1-spe- cific RNA riboprobes and cDNA probes were used to study the mechanism for the induction of SCDl mRNA. Based on the time course of induction, the SCDl mRNA increased from 2-fold within 6 h to 46-fold within 36 h. Nuclear run-on transcription studies showed that the accumulation of SCDl mRNA after refeeding starved mice a fat-free, high carbohydrate diet was a consequence of the transcriptional activation of the SCDl gene. The SCDl mRNA level decreased rapidly (tlI2 = -4 h) within 24 h when mice fed the fat-free, high carbohydrate diet were switched to a regular chow diet. Furthermore, when the fat-free diet was supplemented with triacylglycerides containing poly- unsaturated fattyacids, the transcriptionof the SCDl gene and the induction of the SCDl mRNA were sig- nificantly blunted. Triacylglycerides containing satu- rated and monounsaturated fatty acids were without dramatic effects. In contrast, synthesis of liver albumin mRNA was little affected by any of these dietary var- iations, indicating that the observed changes in the transcription of the SCDl gene and mRNA levels were specific. These data demonstrate that both dietary car- bohydrates and polyunsaturated fatty acids or their metabolites directly or indirectly regulate the expres- sion of the SCDl gene in mouse liver. Stearoyl-CoA desaturase is a key enzyme involved in the synthesis of unsaturated fatty acids, as well as the regulation of this process. This enzyme catalyzes the A’-cis desaturation of fatty acids (1). The preferred substrates are palmitoyl- and stearoyl-CoA (l), which are converted to palmitoleoyl- and oleoyl-CoA, respectively. Palmitoleic and oleic acids are the major (58% of total) storage form of unsaturated fatty acids in mouse depot fat (2). Hepatic stearoyl-CoA desaturase levels in rats can be increased 40-fold by a shift from a natural diet containing unsaturated triacylglycerides to a regimen of fast- ing followed by refeeding a fat-free, high carbohydrate diet (3). This increase in enzyme activity correlates positively with * This work wassupported by National Institutes of Health Grant DK42825. The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “advertisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact. $ To whom correspondence should be addressed Dept. of Biochem- istry and Molecular Biology, Georgetown University School of Med- icine, 3900 Reservoir Rd., Washington, D. C. 20007. an abundance of stearoyl-CoA desaturase mRNA in total liver (4). We have demonstrated previously that hepatic SCD1’ mRNA levels in mice are also induced by a similar feeding schedule (5). Recently, we isolated and characterized two mouse stearoyl-CoA desaturase genes (SCD1 and SCD2) that encode two isoforms of stearoyl-CoA desaturase enzyme (5, 6). We have generated DNA probes that are specific for each gene and have shown that, whereas both genes are expressed in adipose tissue under normal dietary conditions, SCDl mRNA but not SCD2 mRNA is induced in liver when fasted mice are fed a fat-free, high carbohydrate diet (6). This difference in dietary induction in liver tissue demands distinct control mechanisms for the expression of each of the two genes and can be exploited for the study of tissue-specific, as well as nutritional and hormonal regulation, of gene expres- sion. This study sought to examine the nutritional regulation of expression of the SCDlgene in mouse liver. It is shown here that feeding fasted mice a fat-free, high carbohydrate diet causes induction of the SCDl mRNA. The induction of the SCDl mRNA is a consequence of the transcriptional activa- tion of the gene. In addition, the induction of the SCDl mRNA caused by fasting/refeeding is repressed by switching from a fat-free diet to a regular chow diet or by supplementing the fat-free diet with triacylglycerides containing polyunsat- urated fatty acids. These results suggest positive control by carbohydrate and negative control by polyunsaturated fatty acids on SCDl gene expression. EXPERIMENTAL PROCEDURES Materials-Restriction enzymes and other nucleic acid-modifying enzymes were obtained from Boehringer Mannheim, Stratagene, and Promega Biotec. Fat-free, high carbohydrate diet was from U. S. Biochemical Corp. Tripalmitin (99% pure),tristearin (99% pure), triolein (95% pure), trilinolein (95% pure), trilinolenin (95% pure), and triarachidonin (95% pure) were purchased from Sigma. cDNA probes were obtained as follows: mouse liver albumin cDNA (pmalb2) from K. Zaret, Division ofBiology & Medicine, Brown University (7), rat liver cytosolic phosphoenolpyruvate carboxykinase cDNA (pPck-10) from R. Hanson, Department of Biochemistry, Case West- ern Reserve University School of Medicine @), mouse fatty acid synthase cDNA (pFAS-3) from H. Sul, Harvard University School of Public Health (9), rat liver malic enzyme cDNA (pMEB) from N. Nikodem, National Institutes of Health (lo), human j3-actin from D. Cleveland, Johns Hopkins University School of Medicine, and rat liver S14 protein cDNA (pS14 EXO PE 11-8) from D. Jump, Michigan State University (15). All plasmid DNAs were isolated by a modifi- cation of the SDS-NaOH method (11). Animals and Treatments-Male CD-1 mice, used throughout this study, were obtained from Charles River Breeding Laboratories. Mice The abbreviations used are: SCD, stearoyl-CoA desaturase; bp, base pairs; SDS, sodium dodecyl sulfate; EGTA, [ethylene- bis(oxyethylenenitrilo)]tetraacetic acid; HEPES, 4-(2-hydroxyethyl)- 1-piperazineethanesulfonic acid. 10925

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

THE JOURNAL OF BIOLOGICAL CHEMISTRY Vol. 267, No. 15, Issue of May 25, pp. 10925-10930,1992 Q 1992 by The American Society for Biochemistry and Molecular Biology, Inc. Printed in U. S. A.
Dietary Regulation of Stearoyl-CoA Desaturase 1 Gene Expression in Mouse Liver*
(Received for publication, August 7, 1991)
James M. NtambiS From the Department of Biochemistry and Molecular Biology, the Georgetown University School of Medicine, Washington, D. C. 20007
Expression of the murine liver stearoyl-CoA desat- urase gene (SCD1) is induced upon feeding fasted mice a fat-free, high carbohydrate diet (Ntambi, J. M., Buhrow, S. A., Kaestner, K. H., Christy, R. J., Sibley, E., Kelly, T. J., and Lane, M. D. (1988) J. Bid. Chern. 263, 17291-17300). In the present study, SCD1-spe- cific RNA riboprobes and cDNA probes were used to study the mechanism for the induction of SCDl mRNA. Based on the time course of induction, the SCDl mRNA increased from 2-fold within 6 h to 46-fold within 36 h. Nuclear run-on transcription studies showed that the accumulation of SCDl mRNA after refeeding starved mice a fat-free, high carbohydrate diet was a consequence of the transcriptional activation of the SCDl gene. The SCDl mRNA level decreased rapidly (tlI2 = -4 h) within 24 h when mice fed the fat-free, high carbohydrate diet were switched to a regular chow diet. Furthermore, when the fat-free diet was supplemented with triacylglycerides containing poly- unsaturated fatty acids, the transcription of the SCDl gene and the induction of the SCDl mRNA were sig- nificantly blunted. Triacylglycerides containing satu- rated and monounsaturated fatty acids were without dramatic effects. In contrast, synthesis of liver albumin mRNA was little affected by any of these dietary var- iations, indicating that the observed changes in the transcription of the SCDl gene and mRNA levels were specific. These data demonstrate that both dietary car- bohydrates and polyunsaturated fatty acids or their metabolites directly or indirectly regulate the expres- sion of the SCDl gene in mouse liver.
Stearoyl-CoA desaturase is a key enzyme involved in the synthesis of unsaturated fatty acids, as well as the regulation of this process. This enzyme catalyzes the A’-cis desaturation of fatty acids (1). The preferred substrates are palmitoyl- and stearoyl-CoA (l), which are converted to palmitoleoyl- and oleoyl-CoA, respectively. Palmitoleic and oleic acids are the major (58% of total) storage form of unsaturated fatty acids in mouse depot fat (2). Hepatic stearoyl-CoA desaturase levels in rats can be increased 40-fold by a shift from a natural diet containing unsaturated triacylglycerides to a regimen of fast- ing followed by refeeding a fat-free, high carbohydrate diet (3). This increase in enzyme activity correlates positively with
* This work was supported by National Institutes of Health Grant DK42825. The costs of publication of this article were defrayed in part by the payment of page charges. This article must therefore be hereby marked “advertisement” in accordance with 18 U.S.C. Section 1734 solely to indicate this fact.
$ To whom correspondence should be addressed Dept. of Biochem- istry and Molecular Biology, Georgetown University School of Med- icine, 3900 Reservoir Rd., Washington, D. C. 20007.
an abundance of stearoyl-CoA desaturase mRNA in total liver (4). We have demonstrated previously that hepatic SCD1’ mRNA levels in mice are also induced by a similar feeding schedule (5). Recently, we isolated and characterized two mouse stearoyl-CoA desaturase genes (SCD1 and SCD2) that encode two isoforms of stearoyl-CoA desaturase enzyme (5, 6). We have generated DNA probes that are specific for each gene and have shown that, whereas both genes are expressed in adipose tissue under normal dietary conditions, SCDl mRNA but not SCD2 mRNA is induced in liver when fasted mice are fed a fat-free, high carbohydrate diet (6). This difference in dietary induction in liver tissue demands distinct control mechanisms for the expression of each of the two genes and can be exploited for the study of tissue-specific, as well as nutritional and hormonal regulation, of gene expres- sion.
This study sought to examine the nutritional regulation of expression of the SCDl gene in mouse liver. It is shown here that feeding fasted mice a fat-free, high carbohydrate diet causes induction of the SCDl mRNA. The induction of the SCDl mRNA is a consequence of the transcriptional activa- tion of the gene. In addition, the induction of the SCDl mRNA caused by fasting/refeeding is repressed by switching from a fat-free diet to a regular chow diet or by supplementing the fat-free diet with triacylglycerides containing polyunsat- urated fatty acids. These results suggest positive control by carbohydrate and negative control by polyunsaturated fatty acids on SCDl gene expression.
EXPERIMENTAL PROCEDURES
Materials-Restriction enzymes and other nucleic acid-modifying enzymes were obtained from Boehringer Mannheim, Stratagene, and Promega Biotec. Fat-free, high carbohydrate diet was from U. S. Biochemical Corp. Tripalmitin (99% pure), tristearin (99% pure), triolein (95% pure), trilinolein (95% pure), trilinolenin (95% pure), and triarachidonin (95% pure) were purchased from Sigma. cDNA probes were obtained as follows: mouse liver albumin cDNA (pmalb2) from K. Zaret, Division of Biology & Medicine, Brown University (7), rat liver cytosolic phosphoenolpyruvate carboxykinase cDNA (pPck-10) from R. Hanson, Department of Biochemistry, Case West- ern Reserve University School of Medicine @), mouse fatty acid synthase cDNA (pFAS-3) from H. Sul, Harvard University School of Public Health (9), rat liver malic enzyme cDNA (pMEB) from N. Nikodem, National Institutes of Health (lo), human j3-actin from D. Cleveland, Johns Hopkins University School of Medicine, and rat liver S14 protein cDNA (pS14 EXO PE 11-8) from D. Jump, Michigan State University (15). All plasmid DNAs were isolated by a modifi- cation of the SDS-NaOH method (11).
Animals and Treatments-Male CD-1 mice, used throughout this study, were obtained from Charles River Breeding Laboratories. Mice
The abbreviations used are: SCD, stearoyl-CoA desaturase; bp, base pairs; SDS, sodium dodecyl sulfate; EGTA, [ethylene- bis(oxyethylenenitrilo)]tetraacetic acid; HEPES, 4-(2-hydroxyethyl)- 1-piperazineethanesulfonic acid.
10925
10926 Stearoyl-CoA Desaturase 1 Gene Regulation (3-4 weeks old) were maintained on a complete diet of Purina Chow for 2 days before the start of an experiment. Experimental animals were fed according to the modification of the protocol of Thiede and Strittmatter (3) to induce the expression of hepatic stearoyl-CoA desaturase. Briefly, mice were starved for 24 h, refed Purina Chow diet for 24 h, starved for 24 h and then refed a fat-free diet for the time intervals shown in the figures and legends. This feeding schedule ensured that the mice ingested the food. In some experiments the fat- free diet was supplemented with triacylglycerides (l-lO%) and 0.1% butylated hydroxytoluene as an antioxidant. Control animals received Purina Chow instead of the fat-free diet. Mice were given free access t o tap water a t all times and the meals were prepared fresh daily.
Isolation and Analysis of Liver RNA-Livers were rapidly removed from sacrificed animals and minced by Polytron for 10 s into 5 M guanidinium thiocyanate containing 0.5% 8-mercaptoethanol. After adding 0.5% Sarkosyl and heating at 65 "C for 5 min, the RNA was isolated according to Chirgwin et al. (12). RNase protection analysis was performed according to Melton et al. (13) as described previously (6) using antisense RNA probes for the SCDl and albumin. These were synthesized in vitro using linearized plasmid DNA templates, [a-"P]CTP (Amersham Corp.), and either T7 or SP6 RNA polym- erase, respectively. The DNA template used for the albumin probe was generated by subcloning the 600-base pair Hind111 fragment of pmalb2 (7) into the pKKl6O plasmid (6). Probes (4.0 X lo5 cpm each) were hybridized to RNA samples (15 pg) at 54 "C in 80% formamide followed by digestion with RNase A and T1. The protected RNA probe fragments were ethanol-precipitated, analyzed on 8% polyacryl- amide, 7 M urea sequencing gels, and visualized by autoradiography. Relative levels of hybridization were quantitated by densitometry. The integrity of the RNA was verified by fractionating 15 pg of total liver RNA on a 1.5% agarose gel containing 6.7% formaldehyde and visualizing the RNA by ethidium bromide staining.
Isolation of Nuclei for Run-on Transcription-Nuclei for gene tran- scription were isolated from fresh liver. The method used is a modi- fication of the procedure described by Jump et al. (14). Briefly, tissue (2 g) was homogenized in 10 ml of buffer A (15 mM Tris-HC1, pH 7.5, 60 mM KCl, 15 mM NaCl, 0.15 mM spermidine, 0.5 mM dithiothreitol, 20 mM EDTA, 0.5 mM EGTA) containing 0.34 M sucrose in a glass Teflon homogenizer. The homogenate was centrifuged a t 600 X g for 10 min and resuspended in 10 ml of buffer B (buffer A with 1.0 M sucrose). The suspension was adjusted to 0.5% Triton X-100 and centrifuged at 2500 X g for 10 min. The pellet was resuspended in 10 ml of buffer C (buffer A containing 0.25 M sucrose, 0.1 mM EDTA, 0.1 mM EGTA), adjusted to 0.5% Triton X-100, and centrifuged a t 600 X g for 10 min. This step was repeated once with buffer C without Triton X-100. Nuclei were then resuspended in 2 ml of Nuclear Storage Buffer (40% glycerol, 75 mM HEPES, pH 7.5, 60 mM KCl, 15 mM NaCl, 0.15 mM spermidine, 0.5 mM spermine, 0.5 mM dithio- threitol, 0.1 mM EDTA, 0.1 mM EGTA) and used immediately or adjusted to 50 Anm units/ml and stored a t -70 "C until use. All steps during the isolation of the nuclei were carried out a t 4 "C, and all buffers were adjusted to 1 mM phenylmethylsulfonyl fluoride just before use.
In Vitro Nuclear Run-on Assay for SCDl Gene Activity-The run- on transcription assay used in this study is a modification of the method described by Jump et al. (15). Briefly, nuclei (10 A280 units) in nuclear storage buffer were added directly to transcription reac- tions (0.3 ml, total volume) containing 25% glycerol, 75 mM HEPES (pH 7.5), 3 mM MgC12, 0.1 M KCl, 0.1 mM EDTA, 0.05 mM EGTA, 1.0 mM spermidine, 1.0 mM dithiothreitol, 16 pg/ml creatine kinase, 100 pg/ml creatine phosphate, 1 mM ATP, 0.5 mM CTP, 0.5 mM GTP, 0.5 PM UTP, 250 pCi of [cY-~*P]UTP, and 120 units of RNasin. The mixtures were incubated a t 26 "C for 20 min, and the resulting radioactive RNA was isolated as previously described (15). Typically, yields of 5 x lo6 cpm/reaction were obtained.
Plasmid DNA was resuspended in 10 mM Tris-HC1 (pH 8.0), 1 mM EDTA (pH 8.0). NaOH was added to 30 mM, and the mixture was incubated at 65 "C for 30 min. After cooling on ice, ammonium acetate was added to 1 M. The DNA was then spotted on Hy-bond nylon membrane (Amersham Corp.), prewetted with 1 M ammonium ace- tate, using the Schleicher & Schull Manifold I1 Slot-blotter. Filters were hybridized with 2 X lo6 cpm of the 32P-labeled RNA/ml using a modification of the protocol of Thompson and Rosner (16) at 37 "C in 6 X SSC, 50 mM phosphate buffer (pH 7), 5 X Denhardt's solution, 0.1% SDS, 50% deionized formamide, and 250 pg/ml tRNA. Filters were washed at 60 "C in 0.1 X SSC, 0,1% SDS, and 1 mM EDTA and then autoradiographed.
RESULTS
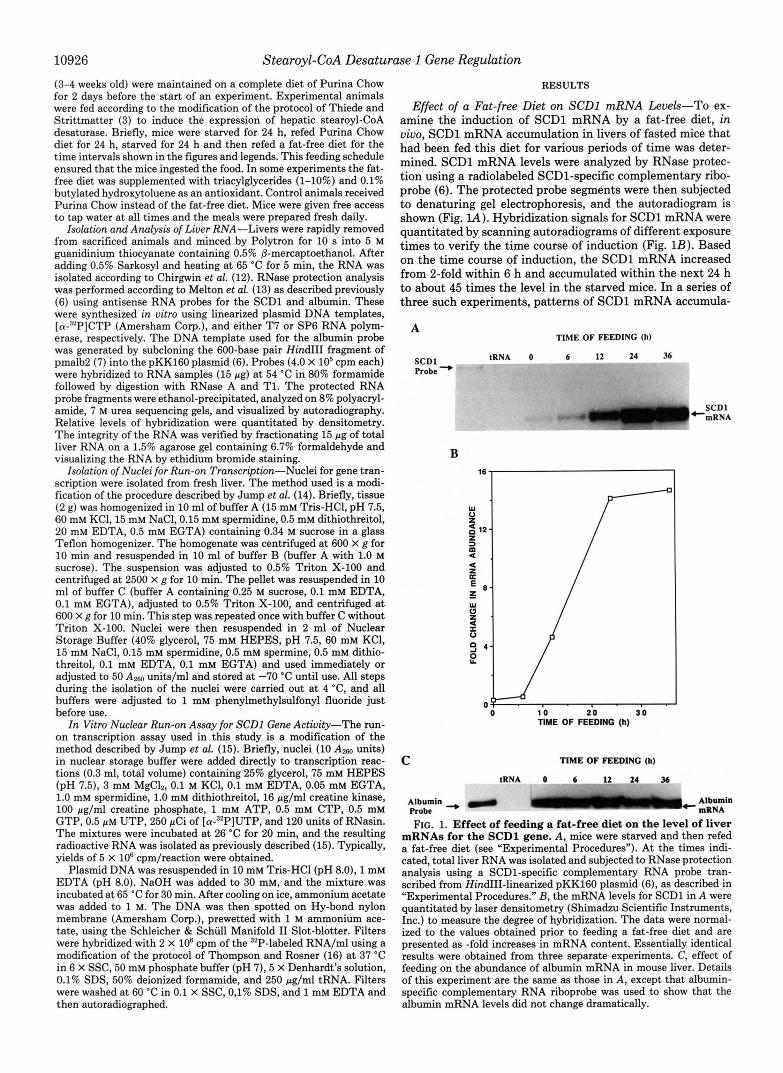
Effect of a Fat-free Diet on SCDl mRNA Levels-To ex- amine the induction of SCDl mRNA by a fat-free diet, in uiuo, SCDl mRNA accumulation in livers of fasted mice that had been fed this diet for various periods of time was deter- mined. SCDl mRNA levels were analyzed by RNase protec- tion using a radiolabeled SCD1-specific complementary ribo- probe (6). The protected probe segments were then subjected to denaturing gel electrophoresis, and the autoradiogram is shown (Fig. lA). Hybridization signals for SCDl mRNA were quantitated by scanning autoradiograms of different exposure times to verify the time course of induction (Fig. 1B). Based on the time course of induction, the SCDl mRNA increased from 2-fold within 6 h and accumulated within the next 24 h to about 45 times the level in the starved mice. In a series of three such experiments, patterns of SCDl mRNA accumula-
A TIME OF FEEDING (h)
SCDl Probe- '
tRNA 0 6 12 24 36
B
I
C TIME OF FEEDING (a)
Albumin ~
Probe FIG. 1. Effect of feeding a fat-free diet on the level of liver
mRNAs for the SCDl gene. A , mice were starved and then refed a fat-free diet (see "Experimental Procedures"). A t the times indi- cated, total liver RNA was isolated and subjected to RNase protection analysis using a SCD1-specific complementary RNA probe tran- scribed from HindIII-linearized pKK16O plasmid (6), as described in "Experimental Procedures." B, the mRNA levels for SCDl in A were quantitated by laser densitometry (Shimadzu Scientific Instruments, Inc.) to measure the degree of hybridization. The data were normal- ized to the values obtained prior to feeding a fat-free diet and are presented as -fold increases in mRNA content. Essentially identical results were obtained from three separate experiments. C, effect of feeding on the abundance of albumin mRNA in mouse liver. Details of this experiment are the same as those in A, except that albumin- specific complementary RNA riboprobe was used to show that the albumin mRNA levels did not change dramatically.
Stearoyl-CoA Desaturase 1 Gene Regulation 10927
tion were similar. When compared with levels of SCDl mRNA in control animals, starvation of mice for 24 h repressed SCDl mRNA by 85%. The level of liver albumin mRNA that served as a control for liver-specific gene expression was also ana- lyzed by RNase protection (Fig. IC) and did not change significantly during the time course of induction of SCDl mRNA. This control study shows that the opposing effects of fasting and a fat-free diet on SCDl mRNA levels in mouse liver are specific.
Effect of a Fat-free Diet on the Transcriptional Rate of the SCDl Gene-To determine whether the increase in steady state levels of SCDl mRNA in response to a fat-free diet was due to an increase in gene transcription, nuclear run-on experiments were performed using nuclei isolated from livers of mice that had been starved and then refed a fat-free diet for various periods of time. Fig. 2 demonstrates that an
A Time (h)
- 0 . . 6 12 24 36 PCEM
pmalbl
SCD:
PC3 (SCDI
1 0 2 0 3 0
TIME (h)
pmalb:
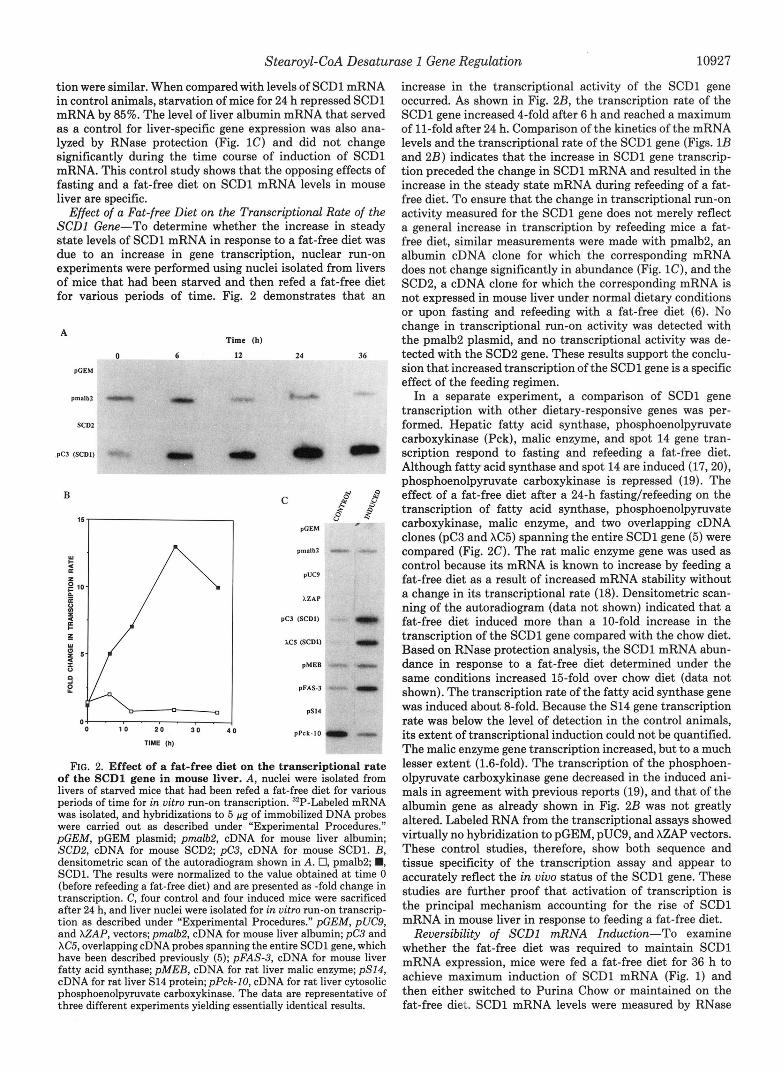
FIG. 2. Effect of a fat-free diet on the transcriptional rate of the SCDl gene in mouse liver. A, nuclei were isolated from livers of starved mice that had been refed a fat-free diet for various periods of time for in vitro run-on transcription. 32P-Labeled mRNA was isolated, and hybridizations to 5 pg of immobilized DNA probes were carried out as described under “Experimental Procedures.” pGEM, pGEM plasmid; pmlb2 , cDNA for mouse liver albumin; SCD2, cDNA for mouse SCD2; pC3, cDNA for mouse SCD1. B, densitometric scan of the autoradiogram shown in A. 0, pmalb2; W, SCD1. The results were normalized to the value obtained at time 0 (before refeeding a fat-free diet) and are presented as -fold change in transcription. C, four control and four induced mice were sacrificed after 24 h, and liver nuclei were isolated for in vitro run-on transcrip- tion as described under “Experimental Procedures.” pGEM, pUC9, and XZAP, vectors; p m l b 2 , cDNA for mouse liver albumin; pC3 and XC5, overlapping cDNA probes spanning the entire SCDl gene, which have been described previously (5); pFAS-3, cDNA for mouse liver fatty acid synthase; pMEB, cDNA for rat liver malic enzyme; pSI4, cDNA for rat liver S14 protein;pPck-10, cDNA for rat liver cytosolic phosphoenolpyruvate carboxykinase. The data are representative of three different experiments yielding essentially identical results.
increase in the transcriptional activity of the SCDl gene occurred. As shown in Fig. 2B, the transcription rate of the SCDl gene increased 4-fold after 6 h and reached a maximum of 11-fold after 24 h. Comparison of the kinetics of the mRNA levels and the transcriptional rate of the SCDl gene (Figs. 1B and 2B) indicates that the increase in SCDl gene transcrip- tion preceded the change in SCDl mRNA and resulted in the increase in the steady state mRNA during refeeding of a fat- free diet. To ensure that the change in transcriptional run-on activity measured for the SCDl gene does not merely reflect a general increase in transcription by refeeding mice a fat- free diet, similar measurements were made with pmalb2, an albumin cDNA clone for which the corresponding mRNA does not change significantly in abundance (Fig. lC), and the SCD2, a cDNA clone for which the corresponding mRNA is not expressed in mouse liver under normal dietary conditions or upon fasting and refeeding with a fat-free diet (6). No change in transcriptional run-on activity was detected with the pmalb2 plasmid, and no transcriptional activity was de- tected with the SCD2 gene. These results support the conclu- sion that increased transcription of the SCDl gene is a specific effect of the feeding regimen.
In a separate experiment, a comparison of SCDl gene transcription with other dietary-responsive genes was per- formed. Hepatic fatty acid synthase, phosphoenolpyruvate carboxykinase (Pck), malic enzyme, and spot 14 gene tran- scription respond to fasting and refeeding a fat-free diet. Although fatty acid synthase and spot 14 are induced (17,20), phosphoenolpyruvate carboxykinase is repressed (19). The effect of a fat-free diet after a 24-h fastinglrefeeding on the transcription of fatty acid synthase, phosphoenolpyruvate carboxykinase, malic enzyme, and two overlapping cDNA clones (pC3 and XC5) spanning the entire SCDl gene (5) were compared (Fig. 2C). The rat malic enzyme gene was used as control because its mRNA is known to increase by feeding a fat-free diet as a result of increased mRNA stability without a change in its transcriptional rate (18). Densitometric scan- ning of the autoradiogram (data not shown) indicated that a fat-free diet induced more than a 10-fold increase in the transcription of the SCDl gene compared with the chow diet. Based on RNase protection analysis, the SCDl mRNA abun- dance in response to a fat-free diet determined under the same conditions increased 15-fold over chow diet (data not shown). The transcription rate of the fatty acid synthase gene was induced about 8-fold. Because the S14 gene transcription rate was below the level of detection in the control animals, its extent of transcriptional induction could not be quantified. The malic enzyme gene transcription increased, but to a much lesser extent (1.6-fold). The transcription of the phosphoen- olpyruvate carboxykinase gene decreased in the induced ani- mals in agreement with previous reports (19), and that of the albumin gene as already shown in Fig. 2B was not greatly altered. Labeled RNA from the transcriptional assays showed virtually no hybridization to pGEM, pUC9, and XZAP vectors. These control studies, therefore, show both sequence and tissue specificity of the transcription assay and appear to accurately reflect the in vivo status of the SCDl gene. These studies are further proof that activation of transcription is the principal mechanism accounting for the rise of SCDl mRNA in mouse liver in response to feeding a fat-free diet.
Reversibility of SCDl mRNA Induction-To examine whether the fat-free diet was required to maintain SCDl mRNA expression, mice were fed a fat-free diet for 36 h to achieve maximum induction of SCDl mRNA (Fig. 1) and then either switched to Purina Chow or maintained on the fat-free diet. SCDl mRNA levels were measured by RNase
10928 Stearoyl-CoA Desaturase 1 Gene Regulation
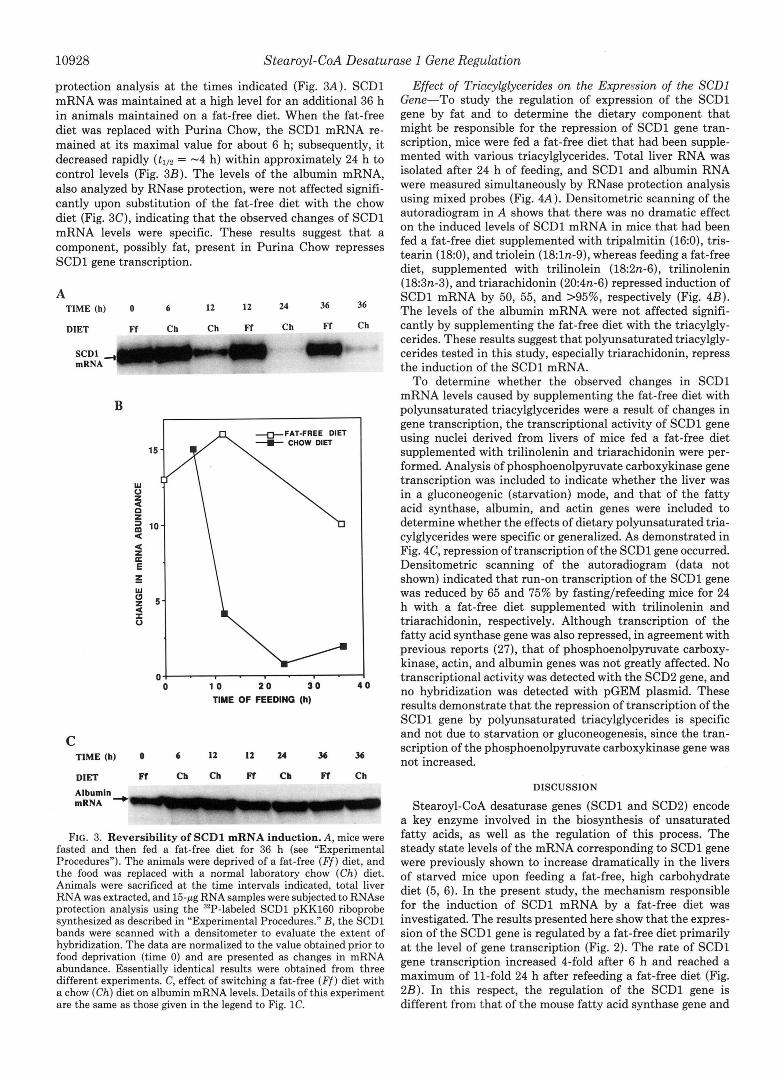
protection analysis at the times indicated (Fig. 3A). SCDl mRNA was maintained at a high level for an additional 36 h in animals maintained on a fat-free diet. When the fat-free diet was replaced with Purina Chow, the SCDl mRNA re- mained at its maximal value for about 6 h; subsequently, it decreased rapidly (tlI2 = -4 h) within approximately 24 h to control levels (Fig. 3B). The levels of the albumin mRNA, also analyzed by RNase protection, were not affected signifi- cantly upon substitution of the fat-free diet with the chow diet (Fig. 3C), indicating that the observed changes of SCDl mRNA levels were specific. These results suggest that a component, possibly fat, present in Purina Chow represses SCDl gene transcription.
A TIME (h) 0 6 12 12 24 36 36
DIET Ff Ch Ch Ff Ch Ff Ch
B I 1
0 1 0 2 0 3 0 4 0 TIME OF FEEDING (h)
C TIME (h) 0 6 I2 12 24 3 6 3 6
DIET Ff Cb Ch Ff Cb Ff Ch
FIG. 3. Reversibility of SCDl mRNA induction. A, mice were fasted and then fed a fat-free diet for 36 h (see “Experimental Procedures”). The animals were deprived of a fat-free ( F f ) diet, and the food was replaced with a normal laboratory chow (Ch) diet. Animals were sacrificed at the time intervals indicated, total liver RNA was extracted, and 15-jtg RNA samples were subjected to RNAse protection analysis using the 32P-labeled SCDl pKK16O riboprobe synthesized as described in “Experimental Procedures.” B, the SCDl bands were scanned with a densitometer to evaluate the extent of hybridization. The data are normalized to the value obtained prior to food deprivation (time 0) and are presented as changes in mRNA abundance. Essentially identical results were obtained from three different experiments. C, effect of switching a fat-free ( F f ) diet with a chow (Ch) diet on albumin mRNA levels. Details of this experiment are the same as those given in the legend to Fig. 1C.
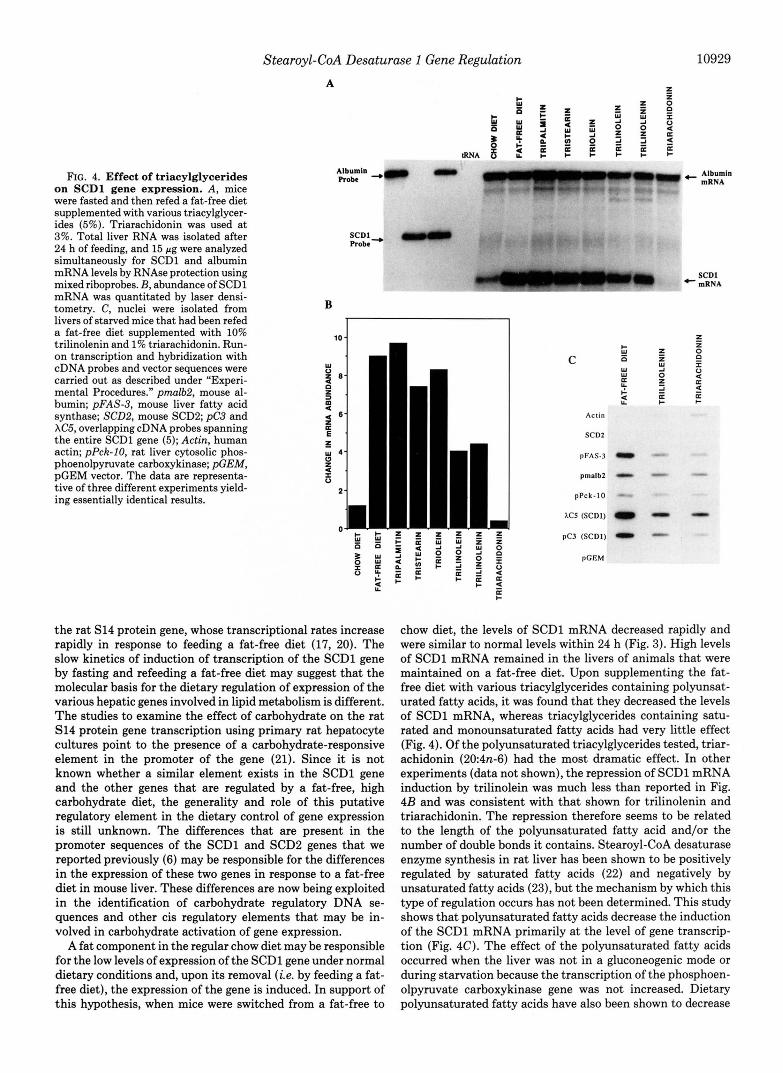
Effect of Trincylglycerides on the Expression of the SCDl Gene-To study the regulation of expression of the SCDl gene by fat and to determine the dietary component that might be responsible for the repression of SCDl gene tran- scription, mice were fed a fat-free diet that had been supple- mented with various triacylglycerides. Total liver RNA was isolated after 24 h of feeding, and SCDl and albumin RNA were measured simultaneously by RNase protection analysis using mixed probes (Fig. 4A). Densitometric scanning of the autoradiogram in A shows that there was no dramatic effect on the induced levels of SCDl mRNA in mice that had been fed a fat-free diet supplemented with tripalmitin (16:0), tris- tearin (18:0), and triolein (18ln-9), whereas feeding a fat-free diet, supplemented with trilinolein (182n-6), trilinolenin (183n-3), and triarachidonin (204n-6) repressed induction of SCDl mRNA by 50, 55, and >95%, respectively (Fig. 4B). The levels of the albumin mRNA were not affected signifi- cantly by supplementing the fat-free diet with the triacylgly- cerides. These results suggest that polyunsaturated triacylgly- cerides tested in this study, especially triarachidonin, repress the induction of the SCDl mRNA.
To determine whether the observed changes in SCDl mRNA levels caused by supplementing the fat-free diet with polyunsaturated triacylglycerides were a result of changes in gene transcription, the transcriptional activity of SCDl gene using nuclei derived from livers of mice fed a fat-free diet supplemented with trilinolenin and triarachidonin were per- formed. Analysis of phosphoenolpyruvate carboxykinase gene transcription was included to indicate whether the liver was in a gluconeogenic (starvation) mode, and that of the fatty acid synthase, albumin, and actin genes were included to determine whether the effects of dietary polyunsaturated tria- cylglycerides were specific or generalized. As demonstrated in Fig. 4C, repression of transcription of the SCDl gene occurred. Densitometric scanning of the autoradiogram (data not shown) indicated that run-on transcription of the SCDl gene was reduced by 65 and 75% by fasting/refeeding mice for 24 h with a fat-free diet supplemented with trilinolenin and triarachidonin, respectively. Although transcription of the fatty acid synthase gene was also repressed, in agreement with previous reports (27), that of phosphoenolpyruvate carboxy- kinase, actin, and albumin genes was not greatly affected. No transcriptional activity was detected with the SCD2 gene, and no hybridization was detected with pGEM plasmid. These results demonstrate that the repression of transcription of the SCDl gene by polyunsaturated triacylglycerides is specific and not due to starvation or gluconeogenesis, since the tran- scription of the phosphoenolpyruvate carboxykinase gene was not increased.
DISCUSSION
Stearoyl-CoA desaturase genes (SCD1 and SCD2) encode a key enzyme involved in the biosynthesis of unsaturated fatty acids, as well as the regulation of this process. The steady state levels of the mRNA corresponding to SCDl gene were previously shown to increase dramatically in the livers of starved mice upon feeding a fat-free, high carbohydrate diet (5, 6). In the present study, the mechanism responsible for the induction of SCDl mRNA by a fat-free diet was investigated. The results presented here show that the expres- sion of the SCDl gene is regulated by a fat-free diet primarily at the level of gene transcription (Fig. 2). The rate of SCDl gene transcription increased 4-fold after 6 h and reached a maximum of 11-fold 24 h after refeeding a fat-free diet (Fig. 2B). In this respect, the regulation of the SCDl gene is different from that of the mouse fatty acid synthase gene and
Stearoyl-CoA Desaturase 1 Gene Regulation A
10929
FIG. 4. Effect of triacylglycerides on SCDl gene expression. A, mice were fasted and then refed a fat-free diet supplemented with various triacylglycer- ides (5%). Triarachidonin was used a t 3%. Total liver RNA was isolated after 24 h of feeding, and 15 pg were analyzed simultaneously for SCDl and albumin mRNA levels by RNAse protection using mixed riboprobes. B, abundance of SCDl mRNA was quantitated by laser densi- tometry. C, nuclei were isolated from livers of starved mice that had been refed a fat-free diet supplemented with 10% trilinolenin and 1% triarachidonin. Run- on transcription and hybridization with cDNA probes and vector sequences were carried out as described under “Experi- mental Procedures.” pmalb.2, mouse al- bumin; pFAS-3, mouse liver fatty acid synthase; SCDB, mouse SCDP; pC3 and XC5, overlapping cDNA probes spanning the entire SCDl gene (5); Actin, human actin; pPck-IO, rat liver cytosolic phos- phoenolpyruvate carboxykinase; &EM, pGEM vector. The data are representa- tive of three different experiments yield- ing essentially identical results.
SCDl Robe-
B
+ I - O D u u z w p : “ 2
2
the rat S14 protein gene, whose transcriptional rates increase rapidly in response to feeding a fat-free diet (17, 20). The slow kinetics of induction of transcription of the SCDl gene by fasting and refeeding a fat-free diet may suggest that the molecular basis for the dietary regulation of expression of the various hepatic genes involved in lipid metabolism is different. The studies to examine the effect of carbohydrate on the rat S14 protein gene transcription using primary rat hepatocyte cultures point to the presence of a carbohydrate-responsive element in the promoter of the gene (21). Since it is not known whether a similar element exists in the SCDl gene and the other genes that are regulated by a fat-free, high carbohydrate diet, the generality and role of this putative regulatory element in the dietary control of gene expression is still unknown. The differences that are present in the promoter sequences of the SCDl and SCD2 genes that we reported previously (6) may be responsible for the differences in the expression of these two genes in response to a fat-free diet in mouse liver. These differences are now being exploited in the identification of carbohydrate regulatory DNA se- quences and other cis regulatory elements that may be in- volved in carbohydrate activation of gene expression.
A fat component in the regular chow diet may be responsible for the low levels of expression of the SCDl gene under normal dietary conditions and, upon its removal ( i e . by feeding a fat- free diet), the expression of the gene is induced. In support of this hypothesis, when mice were switched from a fat-free to
C E 0
W W U
2 2
Actin
SCD;
pFAS-.
pmslb:
pPck- l (
Albumin mRNA
+ mRNA SCDl
chow diet, the levels of SCDl mRNA decreased rapidly and were similar to normal levels within 24 h (Fig. 3). High levels of SCDl mRNA remained in the livers of animals that were maintained on a fat-free diet. Upon supplementing the fat- free diet with various triacylglycerides containing polyunsat- urated fatty acids, it was found that they decreased the levels of SCDl mRNA, whereas triacylglycerides containing satu- rated and monounsaturated fatty acids had very little effect (Fig. 4). Of the polyunsaturated triacylglycerides tested, triar- achidonin (204n-6) had the most dramatic effect. In other experiments (data not shown), the repression of SCDl mRNA induction by trilinolein was much less than reported in Fig. 4B and was consistent with that shown for trilinolenin and triarachidonin. The repression therefore seems to be related to the length of the polyunsaturated fatty acid and/or the number of double bonds it contains. Stearoyl-CoA desaturase enzyme synthesis in rat liver has been shown to be positively regulated by saturated fatty acids (22) and negatively by unsaturated fatty acids (23), but the mechanism by which this type of regulation occurs has not been determined. This study shows that polyunsaturated fatty acids decrease the induction of the SCDl mRNA primarily at the level of gene transcrip- tion (Fig. 4C). The effect of the polyunsaturated fatty acids occurred when the liver was not in a gluconeogenic mode or during starvation because the transcription of the phosphoen- olpyruvate carboxykinase gene was not increased. Dietary polyunsaturated fatty acids have also been shown to decrease

10930 Stearoyl-CoA Desaturase 1 Gene Regulation
mRNA abundance for yeast stearoyl-CoA desaturase (24) and several rat hepatic lipogenic proteins and enzymes, including S14 protein, fatty acid synthase, and glucose-6-phosphate dehydrogenase (25, 26). The suppression of rat hepatic fatty acid synthase and S14 mRNA levels by polyunsaturated fatty acids occurs primarily at the level of gene transcription (27). These data suggest that polyunsaturated fatty acids or their metabolic products regulate the expression of several genes involved in lipid metabolism and that these genes may share common cis regulatory elements and/or trans factors that mediate polyunsaturated fatty acid repression of gene tran- scription. We are currently studying the mechanisms by which the polyunsaturated fatty acids and their metabolites act to repress the transcription of the mouse SCDl gene both in vivo and in vitro.
The plasma signals that mediate the effects of starvation and feeding on the levels of the lipogenic enzymes in liver are generally considered to be insulin and glucagon. Insulin that is elevated in blood of well fed animals stimulates the synthe- sis of lipogenic enzymes, and glucagon, which is elevated in the blood of starved animals, inhibits the synthesis of lipo- genic enzymes (28). The nuclear run-on assays showed a decrease in the transcription of the phosphoenolpyruvate carboxykinase gene and an increase in that of the fatty acid synthase gene upon feeding starved mice a fat-free, high carbohydrate diet (Fig. 2C). The response in the transcription of the phosphoenolpyruvate carboxykinase and fatty acid synthase genes would be consistent with an increase in blood levels of insulin and a corresponding drop in hepatic levels of cAMP (17,19). Stearoyl-CoA desaturase enzyme synthesis in rat liver is known to be regulated positively by insulin (29- 31), but it is not clear from the present study whether the regulation of the mouse enzyme by insulin occurs at the level of gene transcription. The decrease in SCDl mRNA abun- dance during starvation may be due to its repression by polyunsaturated fatty acids in liver that are mobilized from adipose tissue in response to elevated cAMP and a drop in blood insulin levels. Thus, it is likely that the expression of the SCDl gene is under complex control mechanisms such as hormones and, possibly, intermediates of carbohydrate and fat metabolism. We are currently investigating the role of agents such as insulin and glucagon on the expression of SCDl gene in mouse liver. It is, however, hoped that more precise identification of the agents that regulate SCDl gene transcription and analysis of the mechanisms involved will be best approached with liver cells in culture.
Acknowledgments-I thank Kalpana Hassan for excellent technical assistance, and Jane Murphey, Drs. Dale Hereld, Mark Smulson, and Gihan Tennekoon for useful discussions and comments. I thank Drs. D. Jump, N. Nikodem, R. Hanson, H. Sul, S. Tilghman, D. Cleveland, and K. Zaret for providing me with some of the cDNA probes used in this study.
1.
2.
3.
4.
5.
6.
7.
8.
9.
10.
11.
12.
13.
14.
15.
16.
17.
18.
19.
20.
21.
22. 23.
24.
25.
26.
27. 28.
29.
REFERENCES Enoch, H. G., Catala, A., and Strittmatter, P. (1976) J. Biol.
Kasturi, R., and Joshi, V. C. (1982) J. Biol. Chem. 267 , 12224-
Thiede, M. A., and Strittmatter, P. (1985) J. Biol. Chem. 260 ,
Thiede, M. A., Ozols, J., and Strittmatter, P. (1986) J. Biol. Chem. 261,13230-13235
Ntambi, J. M., Buhrow, S. A., Kaestner, K. H., Christy, R. J., Sibley, E., Kelly, T. J., Jr., and Lane, M. D. (1988) J. Biol. Chem. 263,17291-17300
Kaestner, K. H., Ntambi, J. M., Kelly, T. J., Jr., and Lane M. D. (1989) J. Biol. Chem. 264 , 14755-14761
Kioussis, D., Eiferman, F., van de Rijn, P., Gorin, M. B., Ingram, R. S., and Tilghman, S. M. (1981) J. Biol. Chem. 266 , 1960- 1967
Yoo-Warren, H., Monahan, J., Short, H., Short, A., Bruzel, A., Wynshaw-Boris, A., Meisner, H., Samols, D., and Hanson, R. W. (1983) Proc. Natl. Acad. Sci. U. S. A. 80,3656-3660
Paulauskis, J. D., and SUI, S. H. (1989) Biochem. Biophys. Res.
Song, M.-K. H., Dozin, B., Grieco, D., Rall, E. J., and Nikodem,
Birnboim, H. C., and Doly, J. (1979) Nucleic Acids Res. 7 , 1513- 1523
Chirgwin, J. M., Przybyla, A. E., MacDonald, R. J., and Rutter, W. J. (1979) Biochemistry 18, 5294-5299
Melton, D. A., Krieg, P. A., Rebaghiati, M. R., Maniatis, T., Zinn, K., and Green, M. R. (1984) Nucleic Acids Res. 12,7035-7056
Jump, D. B., Wong, N. C. W., and Oppenheimer, J. H. (1987) J. Biol. Chem. 262,778-784
Jump, D. B., Veit, A., Santiago, V., Lepar, G., and Herberholz, L. (1988) J. Biol. Chem. 263,7254-7260
Thompson, K. L., and Rosner, R. M. (1989) J. Biol. Chem. 264 ,
Paulauskis, J. D., and Sul, H. S. (1989) J. Biol. Chem. 264 , 574-
Dozin, B., Rall, J. E., and Nikodem, V. M. (1986) Proc. Natl.
Lamers, W. H., Hanson, R. W., and Meisner, H. M. (1982) Proc.
Jump, D. B., Bell, A., and Santiago, V. (1990) J. Bid. Chem.
Jacobv. D. B.. Zilz. N. D.. and Towle. H. C. (1989) J. Biol. Chem.
Chem. 25,5095-5103
12230
14459-14463
Commun. 158,690-695
V. M. (1988) J. Biol. Chem. 2 6 3 , 17970-17974
3230-3234
577
Acad. Sci. U. S. A. 8 3 , 4705-4709
Natl. Acad. Sci. U. S. A. 79,5137-5141
266,3474-3478
264,’ 17623-17626 ‘
. .
Esner. M.. and Roberts. J. L. (1982) Biochem. J. 206.561-570 Jeffcoat, R., Roberts, P.’A., Ormesher, J., and James, A. T. (1979)
Bossie, M. A., and Martin, C. E. (1989) J. Bacteriol. 171, 6409-
Clarke, S. D., Armstrong, M. K., and Jump, D. B. (1990) J. Nutr.
Tomlinson, J. E., Nakayama, R., and Holten, D. (1988) J. Nutr.
Blake, W. L., and Clarke, S. D. (1990) J. Nutr. 120 , 1727-1729 Goodridee. A. G.. Fisch. J. E.. and Glvnias. M. J. (1983) Arch.
Eur. J. Biochem. 101,439-445
6413
120,225-231
118,408-415
Biophis.’ 227,54-63 ’ “ I . .
Prasad. M. R.. and Joshi. V. C. (1979) J. Biol. Chem. 254.6362- 6369’
11782
Biophys. 233 , 530-539
30. Joshi, V. C., and Aranda, L. P. (1979) J. Biol. Chem. 254,11779-
31. Kasturi, R., Joshi, V. C., and Wakil, S. J. (1984) Arch. Biochem.
Related Documents











