Behavioural Brain Research 274 (2014) 137–142 Contents lists available at ScienceDirect Behavioural Brain Research jou rn al hom epage: www.elsevier.com/locate/bbr Research report BDNF val66met genotype and schizotypal personality traits interact to influence probabilistic association learning Ashley Jayne Skilleter a,b,c , Cynthia Shannon Weickert a,b,c , Ahmed Abdelhalim Moustafa d , Rasha Gendy e , Mico Chan e , Nur Arifin e , Philip Bowden Mitchell a,f , Thomas Wesley Weickert a,b,c,∗ a School of Psychiatry, University of New South Wales, Randwick, NSW, Australia b Schizophrenia Research Institute, Darlinghurst, NSW, Australia c Neuroscience Research Australia, Randwick, NSW, Australia d School of Social Sciences and Psychology, Marcs Institute for Brain and Behaviour, University of Western Sydney, Penrith, NSW, Australia e School of Medicine, University of New South Wales, Randwick, NSW, Australia f Black Dog Institute, Randwick, NSW, Australia h i g h l i g h t s • BDNF val66met interacts with schizotypal personality traits to influence learning. • BDNF met-carriers displaying few schizotypal personality traits performed best. • BDNF met-carriers displaying high schizotypal personality traits performed worst. • Fronto-striatal related probabilistic learning deficits may be intrinsic to schizophrenia. a r t i c l e i n f o Article history: Received 17 March 2014 Received in revised form 24 July 2014 Accepted 27 July 2014 Available online 11 August 2014 Keywords: Brain derived neurotrophic factor val66met polymorphism Probabilistic association learning Schizotypal personality a b s t r a c t The brain derived neurotrophic factor (BDNF) val66met polymorphism rs6265 influences learning and may represent a risk factor for schizophrenia. Healthy people with high schizotypal personality traits display cognitive deficits that are similar to but not as severe as those observed in schizophrenia and they can be studied without confounds of antipsychotics or chronic illness. How genetic variation in BDNF may impact learning in individuals falling along the schizophrenia spectrum is unknown. We predicted that schizotypal personality traits would influence learning and that schizotypal personality- based differences in learning would vary depending on the BDNF val66met genotype. Eighty-nine healthy adults completed the Schizotypal Personality Questionnaire (SPQ) and a probabilistic association learning test. Blood samples were genotyped for the BDNF val66met polymorphism. An ANOVA was performed with BDNF genotype (val homozygotes and met-carriers) and SPQ score (high/low) as grouping variables and probabilistic association learning as the dependent variable. Participants with low SPQ scores (fewer schizotypal personality traits) showed significantly better learning than those with high SPQ scores. BDNF met-carriers displaying few schizotypal personality traits performed best, whereas BDNF met-carriers displaying high schizotypal personality traits performed worst. Thus, the BDNF val66met polymorphism appears to influence probabilistic association learning differently depending on the extent of schizotypal personality traits displayed. Crown Copyright © 2014 Published by Elsevier B.V. This is an open access article under the CC BY-NC-ND license (http://creativecommons.org/licenses/by-nc-nd/3.0/). ∗ Corresponding author at: School of Psychiatry, University of New South Wales, Neuroscience Research Australia, Barker Street, Randwick, NSW 2031, Australia. Tel.: +61 2 9399 1730; fax: +61 2 9399 1034. E-mail address: [email protected] (T.W. Weickert). 1. Introduction Brain-derived neurotrophic factor (BDNF) is a widely distributed neurotrophin in the central nervous system and is important for synaptic plasticity, neuronal differentiation, and survival [1–4]. A single nucleotide polymorphism (SNP) in the human BDNF http://dx.doi.org/10.1016/j.bbr.2014.07.041 0166-4328/Crown Copyright © 2014 Published by Elsevier B.V. This is an open access article under the CC BY-NC-ND license (http://creativecommons.org/ licenses/by-nc-nd/3.0/).

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

R
Bt
ARTa
b
c
d
e
f
h
••••
a
ARRAA
KBpPS
NT
h0l
Behavioural Brain Research 274 (2014) 137–142
Contents lists available at ScienceDirect
Behavioural Brain Research
jou rn al hom epage: www.elsev ier .com/ locate /bbr
esearch report
DNF val66met genotype and schizotypal personality traits interacto influence probabilistic association learning
shley Jayne Skilletera,b,c, Cynthia Shannon Weickerta,b,c, Ahmed Abdelhalim Moustafad,asha Gendye, Mico Chane, Nur Arifine, Philip Bowden Mitchell a,f,homas Wesley Weickerta,b,c,∗
School of Psychiatry, University of New South Wales, Randwick, NSW, AustraliaSchizophrenia Research Institute, Darlinghurst, NSW, AustraliaNeuroscience Research Australia, Randwick, NSW, AustraliaSchool of Social Sciences and Psychology, Marcs Institute for Brain and Behaviour, University of Western Sydney, Penrith, NSW, AustraliaSchool of Medicine, University of New South Wales, Randwick, NSW, AustraliaBlack Dog Institute, Randwick, NSW, Australia
i g h l i g h t s
BDNF val66met interacts with schizotypal personality traits to influence learning.BDNF met-carriers displaying few schizotypal personality traits performed best.BDNF met-carriers displaying high schizotypal personality traits performed worst.Fronto-striatal related probabilistic learning deficits may be intrinsic to schizophrenia.
r t i c l e i n f o
rticle history:eceived 17 March 2014eceived in revised form 24 July 2014ccepted 27 July 2014vailable online 11 August 2014
eywords:rain derived neurotrophic factor val66metolymorphismrobabilistic association learningchizotypal personality
a b s t r a c t
The brain derived neurotrophic factor (BDNF) val66met polymorphism rs6265 influences learning andmay represent a risk factor for schizophrenia. Healthy people with high schizotypal personality traitsdisplay cognitive deficits that are similar to but not as severe as those observed in schizophrenia andthey can be studied without confounds of antipsychotics or chronic illness. How genetic variation inBDNF may impact learning in individuals falling along the schizophrenia spectrum is unknown. Wepredicted that schizotypal personality traits would influence learning and that schizotypal personality-based differences in learning would vary depending on the BDNF val66met genotype. Eighty-nine healthyadults completed the Schizotypal Personality Questionnaire (SPQ) and a probabilistic association learningtest. Blood samples were genotyped for the BDNF val66met polymorphism. An ANOVA was performedwith BDNF genotype (val homozygotes and met-carriers) and SPQ score (high/low) as grouping variablesand probabilistic association learning as the dependent variable. Participants with low SPQ scores (fewer
schizotypal personality traits) showed significantly better learning than those with high SPQ scores. BDNFmet-carriers displaying few schizotypal personality traits performed best, whereas BDNF met-carriersdisplaying high schizotypal personality traits performed worst. Thus, the BDNF val66met polymorphismappears to influence probabilistic association learning differently depending on the extent of schizotypalpersonality traits displayed.Crown Copyright © 2014 Published by Elsevier B.V. This is an open access article under the CCBY-NC-ND license (http://creativecommons.org/licenses/by-nc-nd/3.0/).
∗ Corresponding author at: School of Psychiatry, University of New South Wales,euroscience Research Australia, Barker Street, Randwick, NSW 2031, Australia.el.: +61 2 9399 1730; fax: +61 2 9399 1034.
E-mail address: [email protected] (T.W. Weickert).
ttp://dx.doi.org/10.1016/j.bbr.2014.07.041166-4328/Crown Copyright © 2014 Published by Elsevier B.V. This is an open
icenses/by-nc-nd/3.0/).
1. Introduction
Brain-derived neurotrophic factor (BDNF) is a widely distributedneurotrophin in the central nervous system and is important forsynaptic plasticity, neuronal differentiation, and survival [1–4].A single nucleotide polymorphism (SNP) in the human BDNF
access article under the CC BY-NC-ND license (http://creativecommons.org/

1 Brain
gifiphsonw[tgi
ortaiaelpabalad
crfldhtppisttmgtatssps
tdBGgBifftasdd
38 A.J. Skilleter et al. / Behavioural
ene exists which produces a valine to methionine substitutionn the protein prodomain that can impact intracellular traf-cking, packaging and regulated secretion of BDNF [5]. Theresence of a met allele may attenuate episodic memory andippocampal function in healthy adult humans [5–7]. However,ome studies examining the val66met polymorphism (rs6265)f the BDNF gene in patients with schizophrenia report a sig-ificant over-transmission of the val allele [8,9], in the face oforse episodic memory performance overall in schizophrenia
10]. Thus, this discrepancy in the literature may suggest thathe advantageous versus deleterious effects of the BDNF val66metenotype may vary on the basis of context, e.g., health versusllness.
While the effect of the BDNF genotype on episodic mem-ry and hippocampal function has been studied, little is knownegarding the role of the BDNF genotype in probabilistic associa-ion learning, which requires a gradual learning of cue-outcomessociations that is dependent on normal frontal–striatal activ-ty [11–13]. The relationship between BDNF and probabilisticssociation learning is of interest as peripheral blood BDNF lev-ls correlate with brain activity during probabilistic associationearning in healthy adults [14] and the BDNF val66met polymor-hism also influences prefrontal cortex function in healthy peoplend those at high genetic risk for schizophrenia [15–18]. Proba-ilistic association learning is impaired in schizophrenia [19–22]nd is associated with abnormal fronto-striatal activity [23]. Sib-ings of people with schizophrenia who may share risk genesre also more likely to show probabilistic association learningeficits [21].
Schizotypal personality traits, such as magical ideation, per-eptual aberration, inappropriate affect, and social anxiety, closelyesemble symptoms characterising schizophrenia but are lessorid and debilitating [24,25]. While schizotypal personalityisorder is a formal diagnosis [26], examining those with aigh expression of schizotypal personality traits (i.e., schizo-ypy) in an otherwise healthy sample is important as thisopulation may reveal clues to neurobiological routes thatredispose a person to schizophrenia [27] and allows the exam-
nation of behavioural and cognitive processes associated withchizophrenia without the potential confounds of antipsychoticreatment and a history of chronic illness. Schizotypal personalityraits are displayed along a spectrum in which some individuals
ay display attenuated traits without debilitating effects. Someenetic polymorphisms, such as in Dysbindin, have been showno act as a moderating factor on both schizotypal personality traitsnd cognition [28]. Schizotypal personality traits may be relatedo BDNF genotype, given that healthy met carriers show moretress-induced paranoid thoughts than val homozygotes [29], andchizophrenia-like traits are more often observed in relatives ofeople with schizophrenia who may share genes associated withchizophrenia [30–33].
The aim of the present study was to determine the extento which schizotypal personality traits influence frontal–striatalependent probabilistic association learning in the context ofDNF val66met genotype in a sample of healthy young adults.iven that BDNF in the striatum is derived largely by antero-rade transport from cortical afferents [34], alterations in theDNF genotype may also affect frontal–striatal circuitry and
n turn probabilistic association learning as assayed by per-ormance on the ‘weather prediction’ test, a reliable assay ofrontal–striatal function [11,13,23]. We predicted that schizo-ypal personality traits would negatively influence probabilistic
ssociation learning in otherwise healthy young adults and thatchizotypal personality-based differences in learning would varyepending on the BDNF val66met genotype allelic combinationisplayed.Research 274 (2014) 137–142
2. Materials and methods
2.1. Participants
Eighty-nine healthy young adults without a history of mentalillness were recruited from the local university and the commu-nity. All participants were screened for the following exclusionfactors: a personal or family history of psychiatric disorder, historyof alcohol or substance abuse/dependence within past 5 years, headinjury with loss of consciousness, seizures, central nervous systeminfection, developmental disorder, and uncontrolled diabetes orhypertension. A four subtest version of the Wechsler Adult Intel-ligence Scale, 3rd Edition (WAIS-III) [35] was administered to allparticipants to obtain estimates of current IQ. The Depression, Anx-iety, and Stress Scales [36] and the SF36v2 Health Survey [37] wereadministered to assess negative emotional states and health status,respectively (see Supplemental Table 1 for demographic character-istics of the entire sample). Written informed consent was obtainedfrom all participants before beginning the study and all procedureswere approved by the University of New South Wales and the SouthEastern Sydney and Illawarra Area Health Service Human ResearchEthic Committees.
2.2. Schizotypal personality traits
The self-report Schizotypal Personality Questionnaire (SPQ) isa psychometric and multidimensional questionnaire for assessingschizotypal traits developed from the DSM-III-R criteria for schizo-typal personality disorder [38]. Participants completed the SPQ,which contains subscales for nine schizotypal traits (ideas of ref-erence, excessive social anxiety, odd beliefs or magical thinking,unusual perceptual experiences, odd or eccentric behaviour, noclose friends, odd speech, constricted affect, suspiciousness) [39].The questionnaire comprises 74 items with each being scored 1 fora ‘yes’ and 0 for a ‘no’ response.
2.3. Probabilistic association learning assessment
Participants were tested on the probabilistic association learn-ing “weather prediction” test described in detail elsewhere[19–22,40]. In this test, participants learn the relationship betweentwo equally occurring outcomes (rain or shine) and combinationsof four cue cards composed of simple geometric shapes. Partici-pants were instructed to predict an outcome of rain or shine onthe basis of four cue cards presented individually or in combina-tions of up to 3 cards. They were told that they should guess at firstbut they would gradually improve at determining which cue cardcombinations predicted each outcome. The relationships betweencue card combinations and their outcomes (“rain” or “shine”)were predetermined on a probabilistic basis (see SupplementalTable 2). Card combinations were presented pseudo-randomlywith the constraint that identical cue combinations would notappear consecutively and identical outcomes were limited to fiveconsecutive occurrences. The experimental paradigm consistedof 15 blocks of weather prediction trials (with 10 trials/block),yielding 150 weather prediction trials in total. Testing wasadministered individually in a quiet room by a trained researchassistant.
2.4. Genetic assay
Blood samples were collected in 9 ml EDTA tubes and DNA was
isolated from the buffy coat and genotyped for the BDNF val66metpolymorphism (rs6265) using Applied Biosystems TaqMan SNPassays with our ABI Prism 7900HT Fast Real Time quantitativePCR system and standard procedures. Genomic DNA (gDNA) from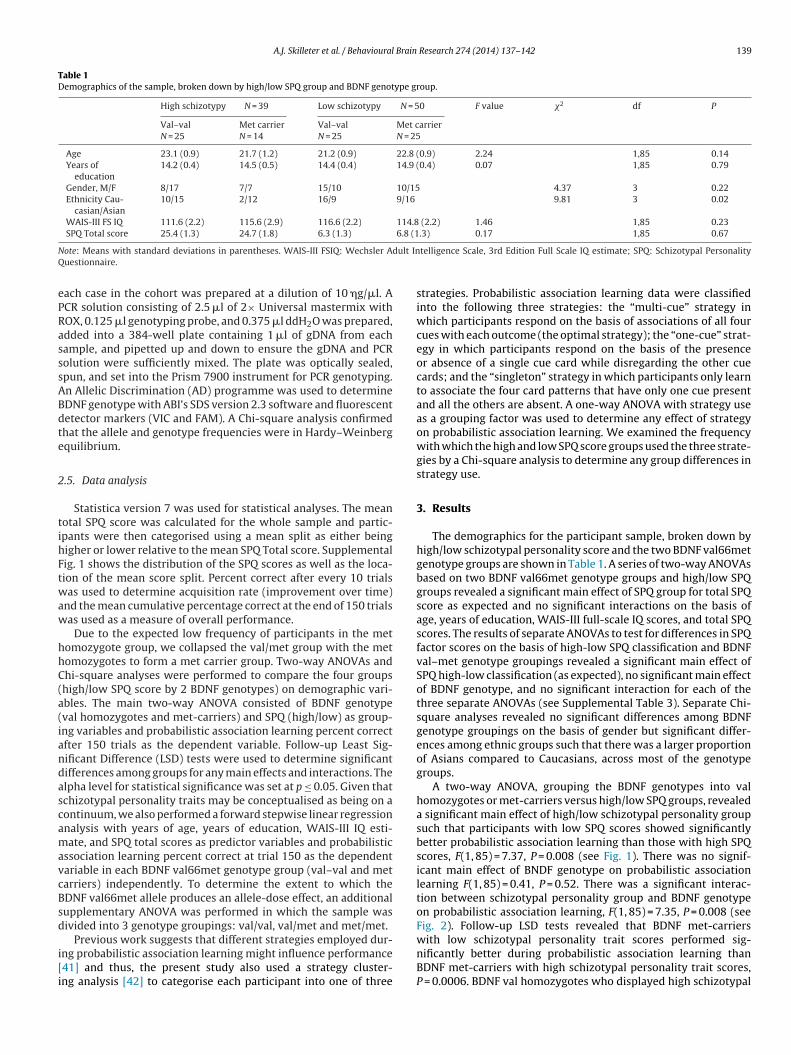
A.J. Skilleter et al. / Behavioural Brain Research 274 (2014) 137–142 139
Table 1Demographics of the sample, broken down by high/low SPQ group and BDNF genotype group.
High schizotypy N = 39 Low schizotypy N = 50 F value �2 df P
Val–valN = 25
Met carrierN = 14
Val–valN = 25
Met carrierN = 25
Age 23.1 (0.9) 21.7 (1.2) 21.2 (0.9) 22.8 (0.9) 2.24 1,85 0.14Years of
education14.2 (0.4) 14.5 (0.5) 14.4 (0.4) 14.9 (0.4) 0.07 1,85 0.79
Gender, M/F 8/17 7/7 15/10 10/15 4.37 3 0.22Ethnicity Cau-
casian/Asian10/15 2/12 16/9 9/16 9.81 3 0.02
WAIS-III FS IQ 111.6 (2.2) 115.6 (2.9) 116.6 (2.2) 114.8 (2.2) 1.46 1,85 0.23SPQ Total score 25.4 (1.3) 24.7 (1.8) 6.3 (1.3) 6.8 (1.3) 0.17 1,85 0.67
N dult InQ
ePRasssABdte
2
tihFtwaw
hhC(a(iandascamavcBsd
i[i
ote: Means with standard deviations in parentheses. WAIS-III FSIQ: Wechsler Auestionnaire.
ach case in the cohort was prepared at a dilution of 10 �g/�l. ACR solution consisting of 2.5 �l of 2× Universal mastermix withOX, 0.125 �l genotyping probe, and 0.375 �l ddH2O was prepared,dded into a 384-well plate containing 1 �l of gDNA from eachample, and pipetted up and down to ensure the gDNA and PCRolution were sufficiently mixed. The plate was optically sealed,pun, and set into the Prism 7900 instrument for PCR genotyping.n Allelic Discrimination (AD) programme was used to determineDNF genotype with ABI’s SDS version 2.3 software and fluorescentetector markers (VIC and FAM). A Chi-square analysis confirmedhat the allele and genotype frequencies were in Hardy–Weinbergquilibrium.
.5. Data analysis
Statistica version 7 was used for statistical analyses. The meanotal SPQ score was calculated for the whole sample and partic-pants were then categorised using a mean split as either beingigher or lower relative to the mean SPQ Total score. Supplementalig. 1 shows the distribution of the SPQ scores as well as the loca-ion of the mean score split. Percent correct after every 10 trialsas used to determine acquisition rate (improvement over time)
nd the mean cumulative percentage correct at the end of 150 trialsas used as a measure of overall performance.
Due to the expected low frequency of participants in the metomozygote group, we collapsed the val/met group with the metomozygotes to form a met carrier group. Two-way ANOVAs andhi-square analyses were performed to compare the four groupshigh/low SPQ score by 2 BDNF genotypes) on demographic vari-bles. The main two-way ANOVA consisted of BDNF genotypeval homozygotes and met-carriers) and SPQ (high/low) as group-ng variables and probabilistic association learning percent correctfter 150 trials as the dependent variable. Follow-up Least Sig-ificant Difference (LSD) tests were used to determine significantifferences among groups for any main effects and interactions. Thelpha level for statistical significance was set at p ≤ 0.05. Given thatchizotypal personality traits may be conceptualised as being on aontinuum, we also performed a forward stepwise linear regressionnalysis with years of age, years of education, WAIS-III IQ esti-ate, and SPQ total scores as predictor variables and probabilistic
ssociation learning percent correct at trial 150 as the dependentariable in each BDNF val66met genotype group (val–val and metarriers) independently. To determine the extent to which theDNF val66met allele produces an allele-dose effect, an additionalupplementary ANOVA was performed in which the sample wasivided into 3 genotype groupings: val/val, val/met and met/met.
Previous work suggests that different strategies employed dur-ng probabilistic association learning might influence performance41] and thus, the present study also used a strategy cluster-ng analysis [42] to categorise each participant into one of three
telligence Scale, 3rd Edition Full Scale IQ estimate; SPQ: Schizotypal Personality
strategies. Probabilistic association learning data were classifiedinto the following three strategies: the “multi-cue” strategy inwhich participants respond on the basis of associations of all fourcues with each outcome (the optimal strategy); the “one-cue” strat-egy in which participants respond on the basis of the presenceor absence of a single cue card while disregarding the other cuecards; and the “singleton” strategy in which participants only learnto associate the four card patterns that have only one cue presentand all the others are absent. A one-way ANOVA with strategy useas a grouping factor was used to determine any effect of strategyon probabilistic association learning. We examined the frequencywith which the high and low SPQ score groups used the three strate-gies by a Chi-square analysis to determine any group differences instrategy use.
3. Results
The demographics for the participant sample, broken down byhigh/low schizotypal personality score and the two BDNF val66metgenotype groups are shown in Table 1. A series of two-way ANOVAsbased on two BDNF val66met genotype groups and high/low SPQgroups revealed a significant main effect of SPQ group for total SPQscore as expected and no significant interactions on the basis ofage, years of education, WAIS-III full-scale IQ scores, and total SPQscores. The results of separate ANOVAs to test for differences in SPQfactor scores on the basis of high-low SPQ classification and BDNFval–met genotype groupings revealed a significant main effect ofSPQ high-low classification (as expected), no significant main effectof BDNF genotype, and no significant interaction for each of thethree separate ANOVAs (see Supplemental Table 3). Separate Chi-square analyses revealed no significant differences among BDNFgenotype groupings on the basis of gender but significant differ-ences among ethnic groups such that there was a larger proportionof Asians compared to Caucasians, across most of the genotypegroups.
A two-way ANOVA, grouping the BDNF genotypes into valhomozygotes or met-carriers versus high/low SPQ groups, revealeda significant main effect of high/low schizotypal personality groupsuch that participants with low SPQ scores showed significantlybetter probabilistic association learning than those with high SPQscores, F(1, 85) = 7.37, P = 0.008 (see Fig. 1). There was no signif-icant main effect of BNDF genotype on probabilistic associationlearning F(1, 85) = 0.41, P = 0.52. There was a significant interac-tion between schizotypal personality group and BDNF genotypeon probabilistic association learning, F(1, 85) = 7.35, P = 0.008 (seeFig. 2). Follow-up LSD tests revealed that BDNF met-carriers
with low schizotypal personality trait scores performed sig-nificantly better during probabilistic association learning thanBDNF met-carriers with high schizotypal personality trait scores,P = 0.0006. BDNF val homozygotes who displayed high schizotypal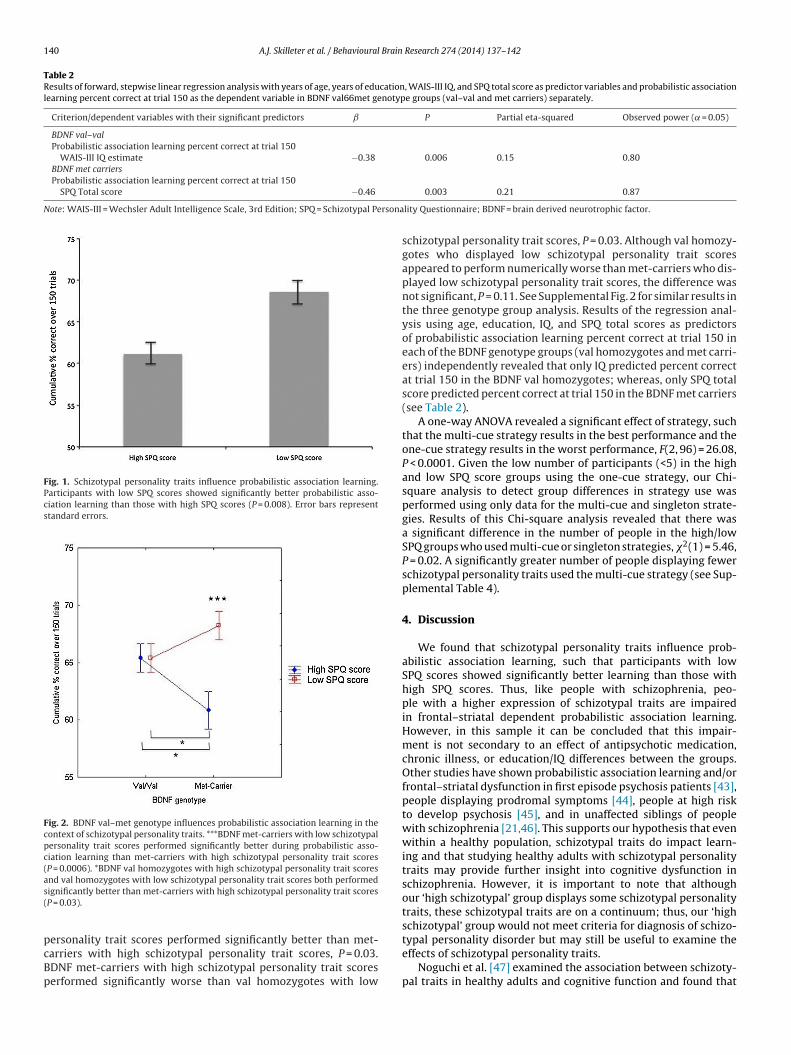
140 A.J. Skilleter et al. / Behavioural Brain Research 274 (2014) 137–142
Table 2Results of forward, stepwise linear regression analysis with years of age, years of education, WAIS-III IQ, and SPQ total score as predictor variables and probabilistic associationlearning percent correct at trial 150 as the dependent variable in BDNF val66met genotype groups (val–val and met carriers) separately.
Criterion/dependent variables with their significant predictors ̌ P Partial eta-squared Observed power ( ̨ = 0.05)
BDNF val–valProbabilistic association learning percent correct at trial 150
WAIS-III IQ estimate −0.38 0.006 0.15 0.80BDNF met carriersProbabilistic association learning percent correct at trial 150
SPQ Total score −0.46
Note: WAIS-III = Wechsler Adult Intelligence Scale, 3rd Edition; SPQ = Schizotypal Persona
Fig. 1. Schizotypal personality traits influence probabilistic association learning.Participants with low SPQ scores showed significantly better probabilistic asso-ciation learning than those with high SPQ scores (P = 0.008). Error bars representstandard errors.
Fig. 2. BDNF val–met genotype influences probabilistic association learning in thecontext of schizotypal personality traits. ***BDNF met-carriers with low schizotypalpersonality trait scores performed significantly better during probabilistic asso-ciation learning than met-carriers with high schizotypal personality trait scores(P = 0.0006). *BDNF val homozygotes with high schizotypal personality trait scoresand val homozygotes with low schizotypal personality trait scores both performeds(
pcBp
typal personality disorder but may still be useful to examine the
ignificantly better than met-carriers with high schizotypal personality trait scoresP = 0.03).
ersonality trait scores performed significantly better than met-
arriers with high schizotypal personality trait scores, P = 0.03.DNF met-carriers with high schizotypal personality trait scoreserformed significantly worse than val homozygotes with low0.003 0.21 0.87
lity Questionnaire; BDNF = brain derived neurotrophic factor.
schizotypal personality trait scores, P = 0.03. Although val homozy-gotes who displayed low schizotypal personality trait scoresappeared to perform numerically worse than met-carriers who dis-played low schizotypal personality trait scores, the difference wasnot significant, P = 0.11. See Supplemental Fig. 2 for similar results inthe three genotype group analysis. Results of the regression anal-ysis using age, education, IQ, and SPQ total scores as predictorsof probabilistic association learning percent correct at trial 150 ineach of the BDNF genotype groups (val homozygotes and met carri-ers) independently revealed that only IQ predicted percent correctat trial 150 in the BDNF val homozygotes; whereas, only SPQ totalscore predicted percent correct at trial 150 in the BDNF met carriers(see Table 2).
A one-way ANOVA revealed a significant effect of strategy, suchthat the multi-cue strategy results in the best performance and theone-cue strategy results in the worst performance, F(2, 96) = 26.08,P < 0.0001. Given the low number of participants (<5) in the highand low SPQ score groups using the one-cue strategy, our Chi-square analysis to detect group differences in strategy use wasperformed using only data for the multi-cue and singleton strate-gies. Results of this Chi-square analysis revealed that there wasa significant difference in the number of people in the high/lowSPQ groups who used multi-cue or singleton strategies, �2(1) = 5.46,P = 0.02. A significantly greater number of people displaying fewerschizotypal personality traits used the multi-cue strategy (see Sup-plemental Table 4).
4. Discussion
We found that schizotypal personality traits influence prob-abilistic association learning, such that participants with lowSPQ scores showed significantly better learning than those withhigh SPQ scores. Thus, like people with schizophrenia, peo-ple with a higher expression of schizotypal traits are impairedin frontal–striatal dependent probabilistic association learning.However, in this sample it can be concluded that this impair-ment is not secondary to an effect of antipsychotic medication,chronic illness, or education/IQ differences between the groups.Other studies have shown probabilistic association learning and/orfrontal–striatal dysfunction in first episode psychosis patients [43],people displaying prodromal symptoms [44], people at high riskto develop psychosis [45], and in unaffected siblings of peoplewith schizophrenia [21,46]. This supports our hypothesis that evenwithin a healthy population, schizotypal traits do impact learn-ing and that studying healthy adults with schizotypal personalitytraits may provide further insight into cognitive dysfunction inschizophrenia. However, it is important to note that althoughour ‘high schizotypal’ group displays some schizotypal personalitytraits, these schizotypal traits are on a continuum; thus, our ‘highschizotypal’ group would not meet criteria for diagnosis of schizo-
effects of schizotypal personality traits.Noguchi et al. [47] examined the association between schizoty-
pal traits in healthy adults and cognitive function and found that

Brain
Satsiatiti
fsshtapCtiTmpitpisig
w(thpppathipd(hmc
iBpstrtwiaiibllS
A.J. Skilleter et al. / Behavioural
PQ scores showed a significant inverse correlation with verbal IQnd the information, comprehension and similarities subtests fromhe Wechsler Adult Intelligence Scale-Revised. McClure et al. [48]howed that a cognitive composite score was significantly impairedn schizotypal personality disorder relative to healthy controls andvoidant personality disorder. The results of the current study fur-her suggest that a relatively high expression of schizotypal traitsn healthy adults are associated with learning deficits, suggestinghat schizotypal traits themselves, even at a non-clinical level, maynterfere with cognition and co-occur with some cognitive deficits.
To our knowledge, the impact of the BDNF polymorphism onrontal–striatal learning in humans has not previously been demon-trated. We found that BDNF met homozygotes displaying fewchizotypal personality traits performed best and those BDNF metomozygotes displaying a high degree of schizotypal personalityraits performed worst on probabilistic association learning. Thus,
high ‘load’ of schizotypal personality traits appears to predictoorer learning, which may be accentuated by the BDNF met allele.onsistent with previous studies of healthy adults, the presence ofhe BDNF val allele appears to provide a protective effect on learn-ng, but only in the context of high schizotypal personality traits.his suggests that there may be a deleterious impact of the BDNFet allele for learning requiring frontal–striatal circuitry and sup-
orts the negative effect of the BDNF met allele on memory shownn previous studies of healthy adults [5–7] but only in the con-ext of a high load of schizotypal personality traits. However, theossible association of the met allele to better frontal–striatal learn-
ng performance in low schizotypal personality trait individualsuggests that genotype effects can vary depending on personal-ty characteristics that are likely influenced by variation in otherenes.
Eisenberg et al. [49] found that the met allele was associatedith significantly less hippocampal regional cerebral blood flow
rCBF) in people with schizophrenia during a working-memoryask. However, the opposite effect of BDNF genotype was seen inealthy controls, such that the met-carriers showed increased hip-ocampal rCBF, suggesting that variation in the BDNF val66metolymorphism interacts differentially with aspects of hippocam-al and frontotemporal function in schizophrenia and healthydults. The SPQ high–low group by BDNF genotype interaction inhe present study showing significantly impaired learning in metomozygotes with high schizotypal personality scores and signif-
cantly better learning in met homozygotes with low schizotypalersonality scores supports the Eisenberg et al. [49] finding of aifferential effect for the BDNF met genotype on brain physiologyrCBF) that is dependent on diagnostic status (schizophrenia versusealthy control) and shows that this differential genotype effectay not be simply secondary to antipsychotic effects or to other
onfounds of having a chronic debilitating mental illness.Egan et al. [5] demonstrated that when the BDNF met allele
s transfected into rat hippocampal neurons, it results in bluntedDNF trafficking, activity-dependent secretion, and less BDNFrotein release. The current study found that within the highchizotypy group, val homozygotes perform significantly betterhan met homozygotes, which suggests that when there is moreeleasable BDNF protein (e.g., in vals), there is better learning inhose with more schizotypal traits relative to met homozygotesith more schizotypal traits. This is supported by previous stud-
es demonstrating that the met allele is detrimental for learningnd memory [5,6] as well as IQ [50] in healthy adults. However,n the current study the exact opposite effect was found in thosendividuals with low SPQ scores, where the met allele appeared to
e beneficial (although not significantly greater) for frontal–striatalearning relative to BDNF val homozygotes. This suggests that ear-ier studies may have recruited individuals on the high end of thePQ spectrum and that the BDNF val/met polymorphism has quite
Research 274 (2014) 137–142 141
different effects depending on the extent of schizotypal personalitytraits expressed.
While using a dichotomous model and breaking schizotypal per-sonality traits into high and low categories for analysis may notbe most consistent with the continuous nature of the schizotypalpersonality trait concept and scale, we obtained similar results inboth an ANOVA using the dichotomous model and our regressionanalyses in which SPQ scores were treated as a continuous vari-able. The regression analyses demonstrated that a change in thedegree of schizotypal personality traits contributed significantly tothe extent of probabilistic association learning only in BDNF metcarriers. These results show that in this case whether the analysisis performed based on cut off scores or using SPQ scores as a con-tinuous variable, the effect in BDNF met carriers is robust enoughto yield significant effects with either model.
Finally, we confirmed that the multicue strategy was the optimalstrategy, which supports previous findings on strategy use duringprobabilistic association learning [41,51]. We also found that thosewith a low expression of schizotypal personality traits used themore optimal multi-cue strategy which is consistent with display-ing a significantly better performance.
There are limitations to the current study. In the original sampleon which the SPQ was developed and normed [38] the mean SPQtotal score was 26.9, as opposed to our study in which the meanSPQ total score was 14.7. Thus, our sample showed a lesser extentof schizotypal traits than the original study. In a similar manner,many of the people in our high schizotypal personality group wouldnot have been classified as deviant, having an SPQ score > 42 [52].Second, we recruited mainly students from the local university andthus a sampling bias may exist such that our sample has a highercognitive functioning capacity as reflected in the above averageWAIS-III IQ scores. In spite of these limitations, in the 39 people whoscored above 14.7, their mean total SPQ score was 25.2 and was sub-stantially different from the mean score of 6.6 in the low SPQ group.We did not find a main effect of BDNF val–met polymorphism onprobabilistic association learning. However, our finding of a BDNFval–met genotype by SPQ high–low group interaction suggests thata genotype effect on learning may be revealed in the context ofschizotypal personality traits. There was also a higher proportion ofAsians than Caucasians in our study sample, particularly in the methomozygote groups. However, because they represent the major-ity in both the high and low SPQ score met homozygote groups, theethnicity is relatively balanced across these schizotypal personal-ity trait and genotype groups and the proportion is consistent withprevious research showing that the frequency of the met allele issignificantly higher in Asians [53].
In conclusion, this study helps refine our understanding of thecontributions of the BDNF genotype and illness-associated traits ondysfunctional frontal–striatal dependant learning in schizophrenia.These results provide the first evidence for a specific genetic effectof the BDNF val66met polymorphism in the context of a sub-clinicallevel of pathology in relation to schizotypal personality traits onprobabilistic association learning. This study contributes to thegrowing body of evidence on the role of alterations of BDNF in thepathophysiology of schizophrenia by showing worse performancein a group of healthy people who were BDNF met carriers withincreased schizotypal personality traits and an absence of antipsy-chotics and history of chronic illness, suggesting that probabilisticassociation learning deficits may be intrinsic to frontal–striatal dys-function associated with schizophrenia.
Acknowledgments
This work was supported by the University of New SouthWales School of Psychiatry, and Neuroscience Research Australia.

1 Brain
CRMvCcrH
A
t
R
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
[
42 A.J. Skilleter et al. / Behavioural
ynthia Shannon Weickert’s work was supported by Schizophreniaesearch Institute (utilising infrastructure funding from the NSWinistry of Health and the Macquarie Group Foundation), the Uni-
ersity of New South Wales, and Neuroscience Research Australia.SW is a recipient of a National Health and Medical Research Coun-il (Australia) Senior Research Fellowship (#1021970). AJS is aecipient of the Ian Scott Scholarship awarded by Australian Rotaryealth.
ppendix A. Supplementary data
Supplementary data associated with this article can be found, inhe online version, at http://dx.doi.org/10.1016/j.bbr.2014.07.041.
eferences
[1] Bekinschtein P, Cammarota M, Katche C, Slipczuk L, Rossato JI, Goldin A, et al.BDNF is essential to promote persistence of long-term memory storage. ProcNatl Acad Sci U S A 2008;105(7):2711–6.
[2] Durany N, Thome J. Neurotrophic factors and the pathophysiology ofschizophrenic psychoses. Eur Psychiatry 2004;19(6):326–37.
[3] Lu B. BDNF and activity-dependent synaptic modulation. Learn Mem2003;10(2):86–98.
[4] Lu B, Chow A. Neurotrophins and hippocampal synaptic transmission and plas-ticity. J Neurosci Res 1999;58(1):76–87.
[5] Egan MF, Kojima M, Callicott JH, Goldberg TE, Kolachana BS, BertolinoA, et al. The BDNF val66met polymorphism affects activity-dependentsecretion of BDNF and human memory and hippocampal function. Cell2003;112(2):257–69.
[6] Dempster E, Toulopoulou T, McDonald C, Bramon E, Walshe M, Filbey F, et al.Association between BDNF val66 met genotype and episodic memory. Am JMed Genet B Neuropsychiatr Genet 2005;134B(1):73–5.
[7] Hariri AR, Goldberg TE, Mattay VS, Kolachana BS, Callicott JH, Egan MF, et al.Brain-derived neurotrophic factor val66met polymorphism affects humanmemory-related hippocampal activity and predicts memory performance. JNeurosci 2003;23(17):6690–4.
[8] Neves-Pereira M, Cheung JK, Pasdar A, Zhang F, Breen G, Yates P, et al. BDNFgene is a risk factor for schizophrenia in a Scottish population. Mol Psychiatry2005;10(2):208–12.
[9] Rosa A, Cuesta MJ, Fatjo-Vilas M, Peralta V, Zarzuela A, Fananas L. The Val66Metpolymorphism of the brain-derived neurotrophic factor gene is associated withrisk for psychosis: evidence from a family-based association study. Am J MedGenet B Neuropsychiatr Genet 2006;141B(2):135–8.
10] Weickert TW, Goldberg TE, Gold JM, Bigelow LB, Egan MF, Weinberger DR.Cognitive impairments in patients with schizophrenia displaying preservedand compromised intellect. Arch Gen Psychiatry 2000;57(9):907–13.
11] Fera F, Weickert TW, Goldberg TE, Tessitore A, Hariri A, Das S, et al. Neural mech-anisms underlying probabilistic category learning in normal aging. J Neurosci2005;25(49):11340–8.
12] Poldrack RA, Clark J, Pare-Blagoev EJ, Shohamy D, Creso Moyano J,Myers C, et al. Interactive memory systems in the human brain. Nature2001;414(6863):546–50.
13] Poldrack RA, Prabhakaran V, Seger CA, Gabrieli JD. Striatal activation duringacquisition of a cognitive skill. Neuropsychology 1999;13(4):564–74.
14] Skilleter AJ, Weickert CS, Vercammen A, Lenroot R, Weickert TW. PeripheralBDNF. A candidate biomarker of healthy neural activity during learning is dis-rupted in schizophrenia. Psychol Med 2014 (in press).
15] Pezawas L, Verchinski BA, Mattay VS, Callicott JH, Kolachana BS, Straub RE, et al.The brain-derived neurotrophic factor val66met polymorphism and variationin human cortical morphology. J Neurosci 2004;24(45):10099–102.
16] Rybakowski JK, Borkowska A, Czerski PM, Skibinska M, Hauser J. Polymorphismof the brain-derived neurotrophic factor gene and performance on a cognitiveprefrontal test in bipolar patients. Bipolar Disord 2003;5(6):468–72.
17] Rybakowski JK, Borkowska A, Skibinska M, Szczepankiewicz A, Kapelski P,Leszczynska-Rodziewicz A, et al. Prefrontal cognition in schizophrenia andbipolar illness in relation to Val66Met polymorphism of the brain-derivedneurotrophic factor gene. Psychiatry Clin Neurosci 2006;60(1):70–6.
18] Whalley HC, Baig BJ, Hall J, Job DE, McIntosh AM, Cunningham-Owens DG,et al. Effects of the BDNF val66met polymorphism on prefrontal brain func-tion in a population at high genetic risk of schizophrenia. Am J Med Genet BNeuropsychiatr Genet 2010;153B(8):1474–82.
19] Foerde K, Poldrack RA, Khan BJ, Sabb FW, Bookheimer SY, Bilder RM, et al.Selective corticostriatal dysfunction in schizophrenia: examination of motorand cognitive skill learning. Neuropsychology 2008;22(1):100–9.
20] Horan WP, Green MF, Knowlton BJ, Wynn JK, Mintz J, Nuechterlein KH.
Impaired implicit learning in schizophrenia. Neuropsychology 2008;22(5):606–17.21] Weickert TW, Goldberg TE, Egan MF, Apud JA, Meeter M, Myers CE, et al. Relativerisk of probabilistic category learning deficits in patients with schizophreniaand their siblings. Biol Psychiatry 2010;67(10):948–55.
[
[
Research 274 (2014) 137–142
22] Weickert TW, Terrazas A, Bigelow LB, Malley JD, Hyde T, Egan MF, et al. Habitand skill learning in schizophrenia: evidence of normal striatal processing withabnormal cortical input. Learn Mem 2002;9(6):430–42.
23] Weickert TW, Goldberg TE, Callicott JH, Chen Q, Apud JA, Das S, et al. Neuralcorrelates of probabilistic category learning in patients with schizophrenia. JNeurosci 2009;29(4):1244–54.
24] Boyle GJ. Schizotypal personality traits: an extension of previous psychometricinvestigations. Aust J Psychol 1998;50:114–8.
25] Vollema MG, van den Bosch RJ. The multidimensionality of schizotypy.Schizophr Bull 1995;21(1):19–31.
26] Diagnostic and statistical manual of mental disorders. 4th ed. Washington, DC:American Psychiatric Association; 1994.
27] Miller P, Byrne M, Hodges A, Lawrie SM, Owens DG, Johnstone EC. Schizotypalcomponents in people at high risk of developing schizophrenia: early findingsfrom the Edinburgh High-Risk Study. Br J Psychiatry 2002;180:179–84.
28] Stefanis NC, Trikalinos TA, Avramopoulos D, Smyrnis N, Evdokimidis I, NtzaniEE, et al. Impact of schizophrenia candidate genes on schizotypy and cognitiveendophenotypes at the population level. Biol Psychiatry 2007;62(7):784–92.
29] Simons CJ, Wichers M, Derom C, Thiery E, Myin-Germeys I, Krabbendam L,et al. Subtle gene–environment interactions driving paranoia in daily life. GenesBrain Behav 2009;8(1):5–12.
30] Kendler KS, McGuire M, Gruenberg AM, O’Hare A, Spellman M, Walsh D. TheRoscommon Family Study. III. Schizophrenia-related personality disorders inrelatives. Arch Gen Psychiatry 1993;50(10):781–8.
31] Kotsaftis A, Neale JM. Schizotypal personality disorder: 1. The clinical syn-drome. Clin Psychol Rev 1993;13:451–72.
32] Lenzenweger MF, Loranger AW. Detection of familial schizophrenia using apsychometric measure of schizotypy. Arch Gen Psychiatry 1989;46(10):902–7.
33] Tsuang MT, Stone WS, Faraone SV. Schizophrenia: a review of genetic studies.Harv Rev Psychiatry 1999;7(4):185–207.
34] Altar CA, Cai N, Bliven T, Juhasz M, Conner JM, Acheson AL, et al. Anterogradetransport of brain-derived neurotrophic factor and its role in the brain. Nature1997;389(6653):856–60.
35] Wechsler D. Wechsler Adult Intelligence Scale. San Antonio, TX: The Psycho-logical Corporation; 1997.
36] Lovibond SH, Lovibond PF. Manual for depression anxiety stress scales. Sydney:Psychology Foundation Monograph; 1995.
37] Ware JE, Kosinski M, Gandek B. SF36 health survey: manual and interpretationguide. Lincoln: Quality Metric; 2005.
38] Raine A. The SPQ: a scale for the assessment of schizotypal personality basedon DSM-III-R criteria. Schizophr Bull 1991;17(4):555–64.
39] Raine A, Reynolds C, Lencz T, Scerbo A, Triphon N, Kim D. Cognitive-perceptual,interpersonal, and disorganized features of schizotypal personality. SchizophrBull 1994;20(1):191–201.
40] Knowlton BJ, Mangels JA, Squire LR. A neostriatal habit learning system inhumans. Science 1996;273(5280):1399–402.
41] Gluck MA, Shohamy D, Myers C. How do people solve the weather prediction”task?: individual variability in strategies for probabilistic category learning.Learn Mem 2002;9(6):408–18.
42] Meeter M, Myers CE, Shohamy D, Hopkins RO, Gluck MA. Strategies in proba-bilistic categorization: results from a new way of analyzing performance. LearnMem 2006;13(2):230–9.
43] Murray GK, Corlett PR, Clark L, Pessiglione M, Blackwell AD, Honey G, et al.Substantia nigra/ventral tegmental reward prediction error disruption in psy-chosis. Mol Psychiatry 2008;13(3):239, 67–76.
44] Howes OD, Montgomery AJ, Asselin MC, Murray RM, Valli I, Tabraham P, et al.Elevated striatal dopamine function linked to prodromal signs of schizophrenia.Arch Gen Psychiatry 2009;66(1):13–20.
45] Fusar-Poli P, Howes OD, Allen P, Broome M, Valli I, Asselin MC, et al. Abnor-mal prefrontal activation directly related to pre-synaptic striatal dopaminedysfunction in people at clinical high risk for psychosis. Mol Psychiatry2011;16(1):67–75.
46] Wagshal D, Knowlton BJ, Cohen JR, Poldrack RA, Bookheimer SY, Bilder RM,et al. Impaired automatization of a cognitive skill in first-degree relatives ofpatients with schizophrenia. Psychiatry Res 2014;215:294–9.
47] Noguchi H, Hori H, Kunugi H. Schizotypal traits and cognitive function inhealthy adults. Psychiatry Res 2008;161(2):162–9.
48] McClure MM, Harvey PD, Bowie CR, Iacoviello B, Siever LJ. Functional outcomes,functional capacity, and cognitive impairment in schizotypal personality dis-order. Schizophr Res 2013;144(1–3):146–50.
49] Eisenberg DP, Ianni AM, Wei SM, Kohn PD, Kolachana B, Apud J, et al. Brain-derived neurotrophic factor (BDNF) Val(66)Met polymorphism differentiallypredicts hippocampal function in medication-free patients with schizophrenia.Mol Psychiatry 2013;18(6):713–20.
50] Tsai SJ, Hong CJ, Yu YW, Chen TJ. Association study of a brain-derived neu-rotrophic factor (BDNF) Val66Met polymorphism and personality trait andintelligence in healthy young females. Neuropsychobiology 2004;49(1):13–6.
51] Moustafa AA, Gluck MA. A neurocomputational model of dopamine andprefrontal–striatal interactions during multicue category learning by Parkinsonpatients. J Cogn Neurosci 2011;23(1):151–67.
52] Barkus E, Lewis S. Schizotypy and psychosis like experiences from recreationalcannabis in a non-clinical sample. Psychol Med 2008;38(9):1267–76.
53] Pivac N, Kim B, Nedic G, Joo YH, Kozaric-Kovacic D, Hong JP, et al. Ethnicdifferences in brain-derived neurotrophic factor Val66Met polymorphism inCroatian and Korean healthy participants. Croat Med J 2009;50(1):43–8.
Related Documents











