American Journal of Primatology 68:777–788 (2006) RESEARCH ARTICLE Arterial Vascularization of the Mandible and Maxilla of Neotropical Primates CRISTIANE SCHILBACH PIZZUTTO 1 , MARCELO ALCINDO DE BARROS VAZ GUIMARA ˜ ES 1 , AND ARANI NANCI BOMFIM MARIANA 2 1 Departamento de Reproduc - a˜o Animal, Faculdade de Medicina Veterina´ria e Zootecnia, Universidade de Sa˜o Paulo, Sa˜o Paulo, Brazil 2 Departamento de Cirurgia, Faculdade de Medicina Veterina´ria e Zootecnia, Universidade de Sa˜o Paulo, Sa˜o Paulo, Brazil The objective of the present investigation was to conduct a comparative macroscopic study of the arterial vascularization of the mandible and maxilla of neotropical primates of the genera Cebus, Alouatta, Callithrix, and Leontopithecus. After vinyl was injected into the arterial system of the head of each specimen, the pieces were macerated and corroded. The level of the bifurcation of the common carotid artery into the internal and external carotids varied between the first and third cervical vertebrae. The external carotid artery accounts for most of the vascularization of the facial structures. The actual vessels responsible for the supply of this region are the sublingual, facial, angular, lingual, submandibular, submental, inferior and superior labial, maxillary, inferior alveolar, infraorbital, superior posterior alveolar, palatine major, and sphenopa- latine arteries. We conclude that although the arterial vascular pattern was similar in all the genera studied, and resembles the human pattern, there are notable variations in the vasculature of the mandible and maxilla among these four neotropical genera. Am. J. Primatol. 68:777–788, 2006. c 2006 Wiley-Liss, Inc. Key words: comparative anatomy; neotropical primates; arteries; mandible; maxilla INTRODUCTION Dental health is an important factor in the general well-being of zoo animals [Wenker et al., 1999]. According to Hillden et al. [1989] and Robinson [1986], tooth pain, reduced feeding, body weight loss, and impairment of organic functions due to infections have all been attributed to dental pathologies, including fractures, periodontal diseases, caries, impaction, and tooth retention [Robinson, 1986]. Depending on the severity of the pathology in the affected tooth, adequate treatment should take into account the anatomical structures Published online in Wiley InterScience (www.interscience.wiley.com). DOI 10.1002/ajp.20278 Received 2 March 2003; revised 26 September 2005; revision accepted 18 October 2005 Contract grant sponsor: Fundac - a ˜o de Amparo a Pesquisa do Estado de Sa ˜o Paulo. Correspondence to: Cristiane Schilbach Pizzutto, Rua Caraı ´bas 1342, apto 33, Pompe ´ia, Sa ˜o Paulo, SP, Brazil CEP 05020-000. E-mail: [email protected] r r 2006 Wiley-Liss, Inc.

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

American Journal of Primatology 68:777–788 (2006)
RESEARCH ARTICLE
Arterial Vascularization of the Mandible and Maxillaof Neotropical Primates
CRISTIANE SCHILBACH PIZZUTTO1�, MARCELO ALCINDO DE BARROS VAZGUIMARAES1, AND ARANI NANCI BOMFIM MARIANA2
1Departamento de Reproduc- ao Animal, Faculdade de Medicina Veterinaria e Zootecnia,Universidade de Sao Paulo, Sao Paulo, Brazil2Departamento de Cirurgia, Faculdade de Medicina Veterinaria e Zootecnia, Universidadede Sao Paulo, Sao Paulo, Brazil
The objective of the present investigation was to conduct a comparativemacroscopic study of the arterial vascularization of the mandible andmaxilla of neotropical primates of the genera Cebus, Alouatta, Callithrix,and Leontopithecus. After vinyl was injected into the arterial system ofthe head of each specimen, the pieces were macerated and corroded. Thelevel of the bifurcation of the common carotid artery into the internal andexternal carotids varied between the first and third cervical vertebrae.The external carotid artery accounts for most of the vascularization of thefacial structures. The actual vessels responsible for the supply of thisregion are the sublingual, facial, angular, lingual, submandibular,submental, inferior and superior labial, maxillary, inferior alveolar,infraorbital, superior posterior alveolar, palatine major, and sphenopa-latine arteries. We conclude that although the arterial vascular patternwas similar in all the genera studied, and resembles the human pattern,there are notable variations in the vasculature of the mandible andmaxilla among these four neotropical genera. Am. J. Primatol.68:777–788, 2006. �c 2006 Wiley-Liss, Inc.
Key words: comparative anatomy; neotropical primates; arteries;mandible; maxilla
INTRODUCTION
Dental health is an important factor in the general well-being of zoo animals[Wenker et al., 1999]. According to Hillden et al. [1989] and Robinson [1986],tooth pain, reduced feeding, body weight loss, and impairment of organicfunctions due to infections have all been attributed to dental pathologies,including fractures, periodontal diseases, caries, impaction, and tooth retention[Robinson, 1986]. Depending on the severity of the pathology in the affectedtooth, adequate treatment should take into account the anatomical structures
Published online in Wiley InterScience (www.interscience.wiley.com).DOI 10.1002/ajp.20278
Received 2 March 2003; revised 26 September 2005; revision accepted 18 October 2005
Contract grant sponsor: Fundac- ao de Amparo a Pesquisa do Estado de Sao Paulo.
�Correspondence to: Cristiane Schilbach Pizzutto, Rua Caraıbas 1342, apto 33, Pompeia, Sao Paulo,SP, Brazil CEP 05020-000. E-mail: [email protected]
rr 2006 Wiley-Liss, Inc.

where the teeth are inserted. Tooth extraction is often indicated, requiringsurgical procedures that are based on an understanding of the vascularizationand innervation of the underlying structures.
The arterial trunk responsible for the vascularization of the teeth andperiodontium in humans is the maxillary artery, which originates from theexternal carotid artery [Brand & Isselhard, 1994; Serra & Ferreira, 1970;Widdowson, 1952]. This vessel sends out numerous collateral branches along itscourse. The inferior alveolar artery supplies the teeth and periodontal structuresof the mandible, while the superior alveolar and infraorbital branches areresponsible for the supply of similar structures in the upper jaw.
In view of the lack of information about the vascular anatomy of the head ofthe brown capuchin monkey, Madeira and Watanabe [1978] carried out acomparative study of the facial arteries of that primate and humans. Veterinarytreatments based on data about humans can be problematic when applied to otherprimates, since the dental structures of nonhuman primates often differ fromthose of humans. This highlights the need for research involving other primates.
Given this scarcity of information, the aims of the present study were toobtain more extensive data from neotropical primates and to describe thevascularization of their mandibular and maxillary periodontal structures.
MATERIALS AND METHODS
We examined a total of 15 individuals from six species: six brown capuchinmonkeys (Cebus apella), two white ear-tufted marmosets (Callithrix jacchusjacchus), three black howler monkeys (Alouatta caraya), one golden lion tamarin(Leontopithecus rosalia), two gold-and-black lion tamarins (L. chrysomelas), andone golden-rumped lion tamarin (L. chrysopygus). All of the specimens wereobtained following routine autopsy procedures carried out at Fundac- ao ParqueZoologico de Sao Paulo. For better conservation of the material, the specimenswere immediately frozen after autopsy and kept frozen for about 2 months priorto preparation.
After the specimens were thawed, the vascular system was injected asfollows: Both the right and left common carotid arteries were cannulated andperfused with physiological saline at ambient temperature, followed by perfusionwith acetone P.A. After perfusion the arteries were injected with vinyl acetate(Solvent Vinyl VMCH B-1099; Chemical and Plastic Union Carbide Corp., NewYork, NY) and stained with Laca Duco Nitrocellulose red (Glasurit do Brasil S.A.,Sao Paulo, SP). About 2 hr later the samples were placed in water for 24 hr fortotal resin polymerization, and then macerated in water for approximately2 months. The samples were then incubated in 20% potassium hydroxide forabout 2 hr for final corrosion of the remaining soft tissues.
The terms used to designate the vessels follow the classification system of theInternational Human Nomina Anatomica [Feneis, 1976].
RESULTS
All results include (whenever possible) the origin, distribution, destination,ramifications, and anastomoses of the vessels, and correlations between thesepathways and the bony structures in the left and right sides of the animal’s face.The sample size in this study precluded us from obtaining conclusive evidence of adirect correlation between vessel size and the size of the foramen and/or itsaccompanying nerve.
778 / Pizzutto et al.
Am. J. Primatol. DOI 10.1002/ajp
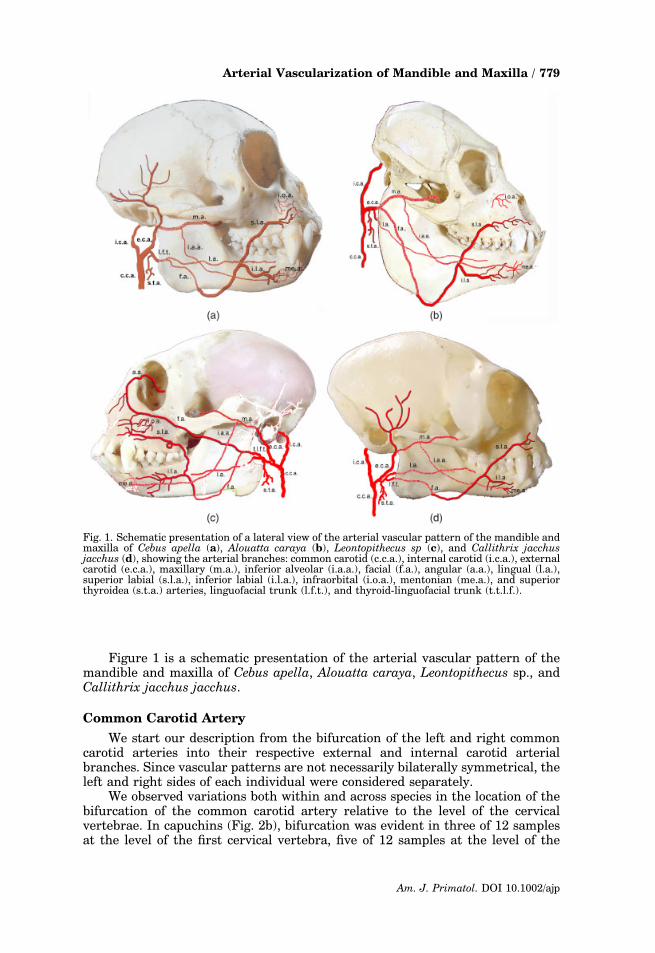
Figure 1 is a schematic presentation of the arterial vascular pattern of themandible and maxilla of Cebus apella, Alouatta caraya, Leontopithecus sp., andCallithrix jacchus jacchus.
Common Carotid Artery
We start our description from the bifurcation of the left and right commoncarotid arteries into their respective external and internal carotid arterialbranches. Since vascular patterns are not necessarily bilaterally symmetrical, theleft and right sides of each individual were considered separately.
We observed variations both within and across species in the location of thebifurcation of the common carotid artery relative to the level of the cervicalvertebrae. In capuchins (Fig. 2b), bifurcation was evident in three of 12 samplesat the level of the first cervical vertebra, five of 12 samples at the level of the
Fig. 1. Schematic presentation of a lateral view of the arterial vascular pattern of the mandible andmaxilla of Cebus apella (a), Alouatta caraya (b), Leontopithecus sp (c), and Callithrix jacchusjacchus (d), showing the arterial branches: common carotid (c.c.a.), internal carotid (i.c.a.), externalcarotid (e.c.a.), maxillary (m.a.), inferior alveolar (i.a.a.), facial (f.a.), angular (a.a.), lingual (l.a.),superior labial (s.l.a.), inferior labial (i.l.a.), infraorbital (i.o.a.), mentonian (me.a.), and superiorthyroidea (s.t.a.) arteries, linguofacial trunk (l.f.t.), and thyroid-linguofacial trunk (t.t.l.f.).
Arterial Vascularization of Mandible and Maxilla / 779
Am. J. Primatol. DOI 10.1002/ajp
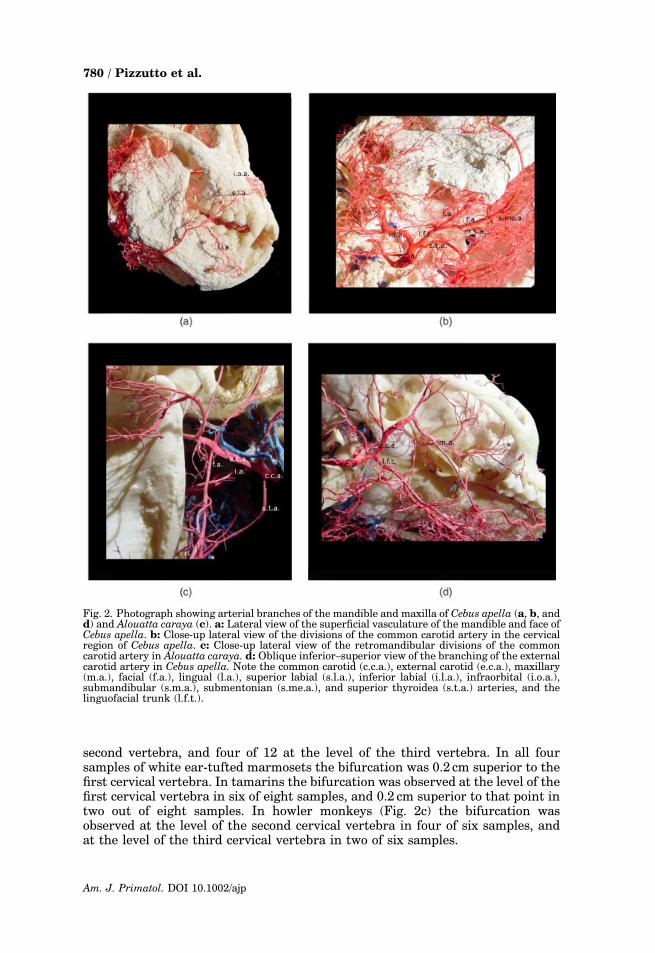
second vertebra, and four of 12 at the level of the third vertebra. In all foursamples of white ear-tufted marmosets the bifurcation was 0.2 cm superior to thefirst cervical vertebra. In tamarins the bifurcation was observed at the level of thefirst cervical vertebra in six of eight samples, and 0.2 cm superior to that point intwo out of eight samples. In howler monkeys (Fig. 2c) the bifurcation wasobserved at the level of the second cervical vertebra in four of six samples, andat the level of the third cervical vertebra in two of six samples.
Fig. 2. Photograph showing arterial branches of the mandible and maxilla of Cebus apella (a, b, andd) and Alouatta caraya (c). a: Lateral view of the superficial vasculature of the mandible and face ofCebus apella. b: Close-up lateral view of the divisions of the common carotid artery in the cervicalregion of Cebus apella. c: Close-up lateral view of the retromandibular divisions of the commoncarotid artery in Alouatta caraya. d: Oblique inferior–superior view of the branching of the externalcarotid artery in Cebus apella. Note the common carotid (c.c.a.), external carotid (e.c.a.), maxillary(m.a.), facial (f.a.), lingual (l.a.), superior labial (s.l.a.), inferior labial (i.l.a.), infraorbital (i.o.a.),submandibular (s.m.a.), submentonian (s.me.a.), and superior thyroidea (s.t.a.) arteries, and thelinguofacial trunk (l.f.t.).
780 / Pizzutto et al.
Am. J. Primatol. DOI 10.1002/ajp

External Carotid Artery
The external carotid artery is one of two branches that originate from thebifurcation of the common carotid artery, and is mainly responsible for thevascularization of facial structures (Fig. 2b–d). Its ramifications are described indetail below according to their importance for the blood supply to the mandible,maxilla, and periodontium.
Linguofacial Trunk
In all 12 brown capuchin monkey and all four marmoset samples, a commonlinguofacial trunk was present and originated as the first branch from theexternal carotid artery (Fig. 2b and d). The superior thyroid artery branched offof the linguofacial truck prior to its bifurcation.
The linguofacial trunk was present in all of the lion tamarin samples, withthis trunk being the first branch of the external carotid artery. In all four liontamarins, regardless of species, the right superior thyroid artery was a branch offthe linguofacial truck. All the left sides, however, showed a trifurcating thyroid-linguofacial trunk that gave rise to the lingual, facial, and superior thyroidarteries simultaneously (Fig. 1c).
No linguofacial trunk was observed in the six howler monkey specimens, withthe first three branches of the external carotid artery arising separately as thesuperior thyroid, lingual, and facial arteries, respectively (Fig. 2c).
The linguofacial trunk, when present, bifurcates into two large vessels–thelingual and facial arteries (Figs. 2b–d, and 3a). The lingual artery gives origin tothree branches: the dorsal lingual, deep lingual, and sublingual arteries. The firsttwo not were studied here, since the main emphasis of the present study was thearterial branches of the mandible and maxilla destined to reach the periodontium.
Sublingual Artery
The sublingual artery originates from the lingual artery and is responsiblefor the vascular supply to the mandibular gingival on the lingual side. It runsanteriorly along the medial side of the ventral border of the mandible, branchingon the floor of the mouth and supplying the mandibular periodontal membraneand bone tissue, and anastomoses with the facial, submental, mandibularalveolar, and inferior labial arteries.
In the current study the sublingual artery was present in eight of 12 browncapuchin monkey samples, five of six howler monkey samples, three of fourmarmoset samples, and all eight of the tamarin samples.
Facial Artery
The facial artery originated from the linguofacial trunk in all of the browncapuchin (Fig. 2d), tamarin, and marmoset specimens, while in howler monkeys itconsistently arose separately from the external carotid artery (Fig. 2c). In all ofthe animals this artery ran medial to the mandibular angle, hidden by its lowerborder until it reached the mid-portion of the body of the mandible, when itemerged inferiorly to continue more superficially in the direction of the face,laterally crossing the angle of the mouth (Fig. 2d). From this point on, differencesin the trajectory of this artery were observed both between and within species.
In the present study the facial artery was found to give origin to fiveimportant collateral branches: the submandibular, superior and inferior labial,angular, and submental arteries. These branches differed among the species
Arterial Vascularization of Mandible and Maxilla / 781
Am. J. Primatol. DOI 10.1002/ajp

studied (Figs. 2d and 3a). In all of the capuchins, marmosets, and howlermonkeys, the facial artery in the mid-region of the body of the mandible gave riseto a branch (the inferior labial artery) that continued on toward the mentalprotuberance. In these species the facial artery then crossed the angle of themouth, where it gave origin to the superior labial artery. There were numerousanastomoses of the facial artery, especially with vessels on the opposite side of theface and neck, with particular emphasis on the labial ones.
In four of 12 capuchin samples, the facial artery ran superior to the angle ofthe mouth, traversing the side of the frontal process of the nose and ending in themedial commissure of the eye (where it is called the angular artery).
Angular Artery
The angular artery, when present in capuchin, marmoset, and howlermonkeys, is the terminal part of the facial artery (Fig. 1a, b, and d). It ascends tothe medial angle of the eye and anastomoses with branches that are not describedin this work.
In contrast, a double facial artery was observed in all of the tamarin samples(Figs. 1c and 3a). Two main branches are sent out from its origin in thelinguofacial trunk. One runs approximately 0.5 cm below the condylar andcoronoid processes of the mandible on its lateral side, passing inferior to thezygomatic bone, continuing on the side of the frontal process of the nose, andending in the medial commissure of the eye. Here it is called angular artery andgives origin to some supraorbital branches above the supraorbital margin. Theother main branch of the tamarin facial artery follows the description of the facialartery of the other animals, and ends in the superior and inferior labial arteries. Itshould be noted this dual facial artery was present only in tamarins, and wasconsistent in all eight specimens.
Submandibular Artery
This artery was considered only as an important branch of the facial artery(Fig. 2d) and was present at a frequency equal to that of the submental arterydescribed below.
Submental Artery
This vessel was found to originate from the facial artery close to the middle ofthe body of the mandible in all of the samples (Fig. 2d). It runs to the symphysisof the chin and anastomoses with the sublingual artery. It then ascends abovethe border of the mandible and divides into a superficial branch and a deepbranch that connect with the inferior labial and mental arteries, respectively. Weidentified this artery in 10 of 12 brown capuchins, five of six howler monkeys, twoof four marmosets, and seven of eight tamarin samples.
It should be noted that when present the tamarin submental artery had anextremely wide caliber and contained numerous tiny ramifications.
Inferior Labial Artery
This vessel originates close to the angle of the mouth, continuing in awinding manner until the base of the mandible above the mental protuberance(Figs. 2a and 3a), where it anastomoses with the artery of the opposite side andwith the mental branch of the inferior alveolar artery. Some branches of theinferior labial artery extend up to the inferior border of the mandible, connecting
782 / Pizzutto et al.
Am. J. Primatol. DOI 10.1002/ajp
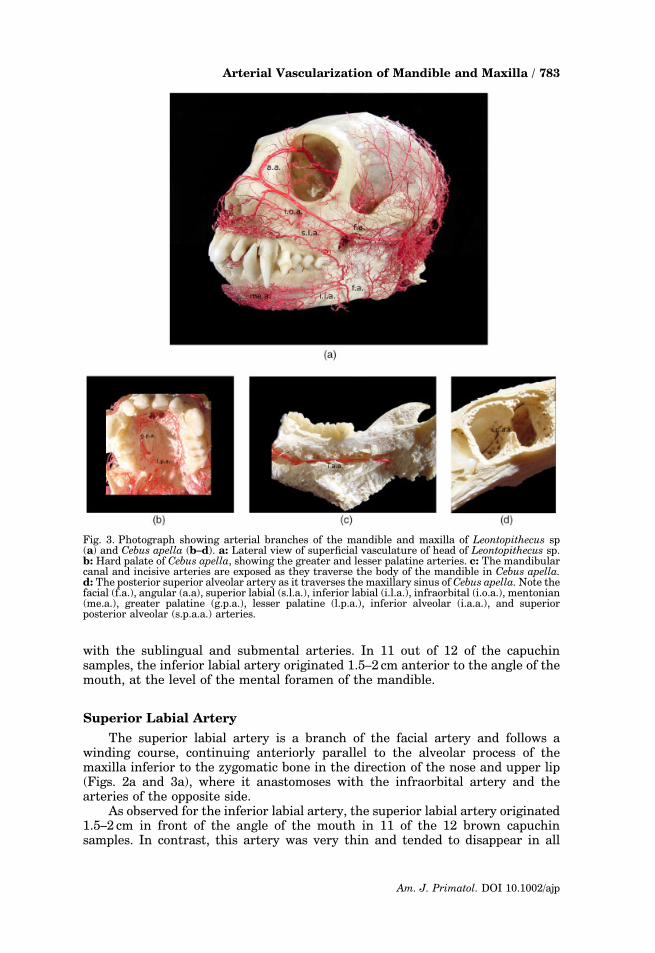
with the sublingual and submental arteries. In 11 out of 12 of the capuchinsamples, the inferior labial artery originated 1.5–2 cm anterior to the angle of themouth, at the level of the mental foramen of the mandible.
Superior Labial Artery
The superior labial artery is a branch of the facial artery and follows awinding course, continuing anteriorly parallel to the alveolar process of themaxilla inferior to the zygomatic bone in the direction of the nose and upper lip(Figs. 2a and 3a), where it anastomoses with the infraorbital artery and thearteries of the opposite side.
As observed for the inferior labial artery, the superior labial artery originated1.5–2 cm in front of the angle of the mouth in 11 of the 12 brown capuchinsamples. In contrast, this artery was very thin and tended to disappear in all
Fig. 3. Photograph showing arterial branches of the mandible and maxilla of Leontopithecus sp(a) and Cebus apella (b–d). a: Lateral view of superficial vasculature of head of Leontopithecus sp.b: Hard palate of Cebus apella, showing the greater and lesser palatine arteries. c: The mandibularcanal and incisive arteries are exposed as they traverse the body of the mandible in Cebus apella.d: The posterior superior alveolar artery as it traverses the maxillary sinus of Cebus apella. Note thefacial (f.a.), angular (a.a), superior labial (s.l.a.), inferior labial (i.l.a.), infraorbital (i.o.a.), mentonian(me.a.), greater palatine (g.p.a.), lesser palatine (l.p.a.), inferior alveolar (i.a.a.), and superiorposterior alveolar (s.p.a.a.) arteries.
Arterial Vascularization of Mandible and Maxilla / 783
Am. J. Primatol. DOI 10.1002/ajp

howler monkey samples. In tamarins, the only interesting difference between theinferior and superior labial arteries was the caliber of the latter. The superiorlabial artery was much thicker and showed a larger number of ramifications,as well as anastomoses with the angular artery.
Maxillary Artery (Arteria maxillaris)
This vessel was responsible for the blood supply to the deep facial structuresin all of the animals and was the main terminal branch of the external carotidartery (Fig. 2b and d). It ran medial to the respective condylar and coronoidprocesses of the mandible, from which point it continued medially up to thepterygopalatine fossa. At the anterior end of the pterygopalatine fossa themaxillary artery emits two branches at almost the same point: the infraorbitaland posterior superior alveolar arteries. After these ramifications, the descendingpalatine artery branches off the maxillary artery. The sphenopalatine artery isthe real terminal branch of the maxillary artery and passes across the back of thenose to the nasal septum.
Inferior Alveolar Artery
The inferior alveolar artery arose anteriorly close to the origin of themaxillary artery in all samples studied. It penetrated the mandibular canalthrough the mandibular foramen and continued as an incisive artery within themandibular canal (Fig. 3c) until it reached the midline anteriorly, providingbranches to the incisor and canine teeth and anastomoses with its homologue onthe opposite side.
The inferior alveolar artery gave off mental branches through the mentalforamen in all of the animals, supplying blood to the region of the chin andanastomoses with the submental and inferior labial arteries. The largest numberof anastomoses was observed in tamarins.
Infraorbital Artery
The infraorbital artery represents one of the last branches of the maxillaryartery at the anterior end of the pterygopalatine fossa. In all samples studied itwas found to run along the sulcus and infraorbital canal through the infraorbitalforamen.
Within the canal the infraorbital artery gave origin to dental branches tothe maxillary teeth, and emerged on the face through the infraorbital foramenin all of the animals (Figs. 2a and 3a).
Posterior Superior Alveolar Artery
The posterior superior alveolar artery originates from the maxillary arteryand is almost always associated with the infraorbital artery when the vessel trunkpasses through the pterygopalatine fossa. Descending along the maxillarytuberosity, it divides into numerous branches, some of which penetrate thealveolar canals in order to supply blood to the molar and premolar teeth (Fig. 3d),and to the lining of the maxillary sinus, whereas others continue anteriorly to thealveolar process to irrigate the gingivae.
It was not possible to observe all of the branches sent out to the teeth.
784 / Pizzutto et al.
Am. J. Primatol. DOI 10.1002/ajp

Anterior Superior Alveolar Artery
The anterior superior alveolar artery is a continuation of the posteriorsuperior alveolar artery. It is responsible for the blood supply to the canine andincisor teeth through the alveolar branches that descend along the anterioralveolar canals to irrigate incisors, canines, and the mucosal membrane of themaxillary sinus.
Descending Palatine Artery
The descending palatine artery is another branch of the maxillary artery atthe anterior end of the pterygopalatine fossa. It continues through thepterygopalatine canal, emerging at the greater palatine foramen, and givesorigin to two collateral branches (the greater and lesser palatine arteries).
Lesser Palatine Artery
The lesser palatine artery arises from the descending palatine artery in thepterygopalatine canal and descends through the lesser palatine foramen to supplythe soft palate (Fig. 3b).
Greater Palatine Artery
The greater palatine artery is the main arterial vessel responsible for thevascularization of the hard palate. This artery originated from a directcontinuation of the trunk of the descending palatine artery after it emergedfrom the greater palatine foramen in all of the samples studied. It ran anteriorlyin the palatine sulcus on the medial side of the alveolar border of the hardpalate to the incisive canal (Fig. 3b). Some branches were found to be distributedto the gingiva and the mucosal membrane of the roof of the mouth. Posterior tothe incisor teeth, it anastomosed with its homologue on the opposite side,thus forming an arterial arch. Small branches arose from this arch supplying themucosa, teeth, and periosteum. A terminal branch anastomosed with thenasopalatine branch of the sphenopalatine artery in the incisive canal.
In eight of the 12 capuchin samples, the greater palatine artery sent out awide-caliber branch that continued until it reached the alveolar process of theincisor teeth. This branch was also highly ramified. In two of the 12 samples, thisramification of the greater palatine artery emerged from the greater palatineforamen and continued in the direction of the choanae.
Sphenopalatine Artery
The sphenopalatine artery is a direct continuation, and thus the terminalbranch, of the maxillary artery. It penetrates the nasal cavity after it passes thesphenopalatine foramen and anastomoses with the branches of the greaterpalatine artery.
DISCUSSION
The preservation of exotic species requires adequate knowledge about thehealth of these animals in captivity. It is necessary to provide good dental care forsuch animals [Wenker et al., 1999], since ignorance can result in severe problems,including loss of appetite, body weight loss, and impairment of organic functionsthrough the occurrence of infections [Hillden et al., 1989; Robinson, 1986].
Arterial Vascularization of Mandible and Maxilla / 785
Am. J. Primatol. DOI 10.1002/ajp

It has long been known that the teeth play an important role in masticationand that therefore their preservation is fundamental for survival. Toothextraction requires a refined knowledge about the anatomical structures,particularly the vessels and nerves, in the region. Extraction often provokesvascular alterations in the mandibular and maxillary branches, as demonstratedby Castelli and Nasjleti [1975] in rhesus monkeys.
In primates the main blood supply to the head comes from the commoncarotid artery, which bifurcates into external and internal carotid arteries. Theexternal carotid artery is responsible for supplying the most extracranialstructures of the head and neck. Therefore, we started our discussion with thebifurcation of the common carotid artery in these different species. Since we useda technique that preserves only bony structures and resin-filled vessels, thereference to soft structures (e.g., muscle and cartilage) was lost; however,this method permitted us to establish direct relationships with bony landmarks inthe head.
Bugge [1974] showed in his study of the cephalic pattern of supply in diversemammals that primates can be allocated into different levels of developmentwithin an expanded primate order, thereby expressing the order in which thegroups in question are presumed to have departed from a line of development thatleads from protoinsectivores to anthropomorphs (including humans).
The level of the bifurcation of the common carotid artery into the internaland external carotids showed significant variation in previous studies of primates.In chimpanzees, Sperino [1987] observed the same pattern as that found inhumans, i.e., bifurcation occurring at the thyroid cartilage level. In studies ofbaboons [Krieger, 1982], rhesus [Castelli & Huekle, 1965; Dyrud, 1944], andgorillas [Raven & Hill, 1950] the bifurcation was observed more cranially, nearthe hyoid bone. In the present study, the bifurcation of the common carotid arterywas observed at an even higher level, ranging from above the first to the thirdcervical vertebra.
In most primates the facial artery normally emerges in a common trunk withthe lingual artery [Castelli & Huelke, 1965; Hill, 1960, 1962; Krieger, 1982;Livini, 1903; Madeira & Watanabe, 1978]. Hill [1962] did not mention thepresence of a linguofacial trunk in the genus Alouatta, which confirms thefindings of the present study that the external carotid artery dorsally sends outseparate lingual and facial arteries. The present results also show that howlermonkeys (Alouatta caraya) do not possess a linguofacial trunk, in contrast to thefindings of Madeira and Watanabe [1978]. This point may be related tointraspecific variation resulting from the small number of individuals studiedin these analyses.
The presence of a thyroid-linguofacial trunk, as evidenced by a trifurcation,was observed only in half the animals in the genus Leontopithecus, and never inCallithrix jacchus. However, Madeira and Watanabe [1978] also identified thepresence of this trunk in C. penicillata. In Leontopithecus and C. jacchus, when athyroid-linguofacial trunk was absent the superior thyroid artery was observed tobe a branch of the linguofacial trunk.
With respect to the facial artery, Hill [1962] reported that in capuchinmonkeys it disappeared almost immediately after it crossed the mandibular angle.In a study of the same primate, Madeira and Watanabe [1978] found that thisartery crossed the angle of the mouth and gave origin to the superior and inferiorlabial arteries. They also reported that the facial artery showed a completetrajectory in only 0.5% of the animals in their study, ending in the so-calledangular artery at the level of the eyes. In the present study a wide variation
786 / Pizzutto et al.
Am. J. Primatol. DOI 10.1002/ajp

regarding this result was observed, with four of the 12 brown capuchin monkeyspossessing an angular artery.
No reports are available in the literature concerning the facial artery inLeontopithecus. Our results show that these animals have a double facial arteryof the same origin. The angular artery of these animals continues on the side ofthe frontal process of the nose and ends in the medial commissure of the eye, andwas identified in all the samples studied.
Previous studies of capuchin monkeys [Madeira & Watanabe, 1978] andrhesus monkeys [Castelli & Huelke, 1965] reported that the superior and inferiorlabial arteries are ramifications of the facial artery. Our results are in agreementwith those findings, with the frequency of the inferior and superior labial arteriesbeing 94.7% in the study of Madeira and Watanabe [1978], and 11 of 12 in thepresent investigation on brown capuchin monkeys.
Of interest, we observed that the superior labial artery in howler monkeyswas very thin and almost disappeared. Given the presence of such reducedalveolar processes in the maxilla, we believe that the branches of the superiorlabial artery have practically no bone surface available to perform theiranastomoses.
With respect to tamarins, the only interesting difference was the greaterthickness of the superior labial artery compared to the inferior labial artery.This variation was also observed by Madeira and Watanabe [1978] incapuchin monkeys.
According to Serra and Ferreira [1970], the arterial trunk responsible for thevascularization of the teeth and periodontium in humans is the maxillary artery(A. maxillaris). This finding is in agreement with our results, since the branchesof this artery were found to irrigate the deep facial structures. Grasse [1972]reported that in capuchin monkeys the maxillary artery ends its trajectory in twobranches: the descending palatine artery and the sphenopalatine artery. Thisobservation is in contrast to our findings, which showed that the maxillary arteryended only in the sphenopalatine artery in all of the neotropical primates studied.
Cohen [1960] considered the dental arteries to be the main vesselsresponsible for the nutrition of the mandible. In another study, Cohen [1959]showed that the lower borders of the mandible are mainly supplied by vessels ofthe periosteum. Subsequently, Castelli [1963] confirmed that the angle of themandible and its lower borders are irrigated by the inferior alveolar artery.The present study demonstrates that the inferior alveolar artery plays animportant role in supplying blood to structures of the periosteum and gingiva inneotropical primates.
According to Gray [1988], the greater palatine artery in humans originatesfrom the maxillary artery in the pterygopalatine fossa and descends to thepterygopalatine canal. After it emerges from the greater palatine foramen, it runsanteriorly in a sulcus on the medial side of the alveolar border of the hard palate.That finding differs from the present results in neotropical primates, in that wefound the greater palatine artery to be a direct continuation of the trunk of thedescending palatine artery after it emerged from the greater palatine foramen.This pattern was also observed by Castelli and Huelke [1965] in rhesus monkeys.
Based on the present results, we conclude that the arterial vascular patternof the maxilla and mandible of neotropical primates is closely similar to that ofhumans. However, differences among species do exist and should be taken intoconsideration during dental and/or surgical interventions in captive animals.Bifurcation of the common carotid artery appears to occur at a higher level inneotropical primates as compared to humans, apes, and Old World monkeys.
Arterial Vascularization of Mandible and Maxilla / 787
Am. J. Primatol. DOI 10.1002/ajp

Other notable variations in facial vasculature among the neotropical primateswere that Alouatta caraya did not possess a linguofacial trunk, a thyroid-linguofacial trunk was frequently observed in the genus Leontopithecus, and thegenus Leontopithecus possessed a double facial artery, with the angular arteryrepresenting the terminal branch of the more superior of two facial arteries.
ACKNOWLEDGMENTS
The authors thank the Fundac- ao Parque Zoologico de Sao Paulo forproviding the study animals. The authors also thank the reviewers for comments,suggestions, and corrections that significantly improved the quality of this paper.
REFERENCES
Brand RW, Isselhard DE. 1994. Anatomy oforofacial structures. 5th ed. St. Louis:Mosby Yearbook. p 189–197.
Bugge JA. 1974. The cephalic arterial systemin insectivores, primates, rodents andlagomorphs, with special reference to thesystematic classification. Acta Anat 87:99–159.
Castelli W. 1963. Vascular architecture ofthe human adult mandible. J Dental Res42:786–792.
Castelli WA, Huelke DF. 1965. The arterialsystem of head and neck of the rhesusmonkey with emphasis on the externalcarotid system. Am J Anat 116:149–170.
Castelli WA, Nasjleti CE. 1975. Vascularchanges in the monkey mandible and max-illa after multiple extraction of teeth: aradiographic study. J Dental Res 54:1237.
Cohen L. 1959. Methods of investigatingthe vascular architecture of the mandible.J Dental Res 38:920–931.
Cohen L. 1960. Further studies into thevascular architecture of the mandible.J Dental Res 39:936–946.
Dyrud J. 1944. The external carotid artery ofrhesus monkey (Macaca mulatta). Anat Rec90:17–22.
Feneis H. 1976. Dicionario ilustrado de anato-mia (nomenclatura internacional). 4th ed.Rio de Janeiro: Cultura Medica. p 191–197.
Grasse PP. 1972. Traite de zoologie, anatomiesystematique, biologie. Vol. 16. Paris: Mas-son. p 755–759.
Gray HFRS. 1988. Gray anatomia. 29th ed.Rio de Janeiro: Guanabara Koogan.p 476–485.
Hill WCO. 1960. Primates–comparative anat-omy and taxonomy. Vol. 4, Part A. Cebidae.Edinburgh: Edinburgh University Press.p 63–66, 281–282, 369–371.
Hill WCO. 1962. Primates–ComparativeAnatomy and Taxonomy. Vol. 5, Part B.Cebidae. Edinburgh: Edinburgh UniversityPress. p 65–67.
Hillden N, Carlsson B, Lundebergh A. 1989.Behavioral disorders in wolves caused bydental problems. Verh Int Symp ErkrankZoo 31:231–233.
Livini. 1903. L’arteria carotis externa. ArchItal Anat Embriol 2:653–741.
Madeira MC, Watanabe I. 1978. Dados anato-micos sobre a arteria facial no homem (fetose adultos) e no macaco-prego (Cebus apella).Rev Faculdade Odontol Arac-atuba 7:59–67.
Raven HC, Hill JH. 1950. Regional anatomyof the gorilla. Part II. In: Raven HC,editor. The anatomy of the gorilla. NewYork: Columbia University Press. p 24–26,114–115.
Robinson PT. 1986. Dentistry in zoo animals.In: Fowler ME, editor. Zoo and wild animalmedicine. 2nd ed. Philadelphia: W.B. Saun-ders. p 534–547.
Serra OD, Ferreira FV. 1970. Anatomiadental. Sao Paulo: Artes Medicas. p 259–262.
Sperino G. 1987. Anatomia do chimpanze.Torino: Unione Tipografico-Editrice. p 264–267.
Swindler DR, Wood CD. 1982. An atlas ofprimate gross anatomy–baboon, chimpan-zee and man. Malabar: Robert E. Krieger.p 58–73.
Wenker CJ, Stich H, Muller M, Lussi A. 1999.A retrospective study of dental conditions ofcaptive brown bears (Ursus arctos spp.)compared with free-ranging Alaskan grizz-lies (Ursos arctos horribilis). J Zoo WildlMed 30:208–221.
Widdowson TW. 1952. Special or dentalanatomy and physiology and dental histo-logy. 8th ed. London: Staples. 23p.
788 / Pizzutto et al.
Am. J. Primatol. DOI 10.1002/ajp
Related Documents











