A A n n n n u u a a l l R R e e p p o o r r t t 2 2 0 0 0 0 7 7 S S B B A A - - 4 4 : : R R I I C C E E

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

AAnnnnuuaall RReeppoorrtt 22000077
SSBBAA--44:: RRIICCEE

Output 1: Enhanced gene pools1A. IRRIGATED RICE………………………………………………………………….. Pag. 1
1.A.1 Breeding Strategies to increase the content of iron and zinc in the rice grain ……………………………………………………………………... Pag. 1 1.A.2. Progress Report on Broadening the Genetic Base of Rice In Latin America… Pag. 12 1.A.3. Yield evaluation in large panicle type lines………………………………..……. Pag. 21 1.A.4. Partial diallel analysis for dark green leaves and its relationship with yield. …Pag. 27
1B. UPLAND RICE……………………………………………………………………..... Pag. 35
1.B.1. Rice Synthetic Population Improvement………………………………………… Pag. 35
1C. BIOTECHNOLOGY ACTIVITIES RELATED TO ENHANCED GENE POOLS ……………………………………………………………………………………………... Pag. 43
1.C.1 SNPs Variation of iron homeostasis genes in mutant and natural rice populations………………………………………………………………………………... Pag. 43 1.C.2 Identification of Snps Markers for Biofortification in Rice……………………. Pag. 49
1.C.3 Development of introgression lines with chromosome segments substitutions from the African cultivated rice species into Oryza sativa genetic background……… Pag. 53 1.C.4 Exploring wild diversity in rice……………………………………...……………. Pag. 60 1.C.5. Characterization of Genetic Diversity: Relationships and potential origin of the weedy rice complex inColombia…………………………………………………….. Pag. 67 1.C.6 Anther culture for advancing breeding populations of FLAR…………………. Pag. 84 1.C.7 Validation of SNP markers in Oryza sativa and wild relative species…………. Pag. 85 1.C.8 The Use of Anther Culture and Embryo Rescue for Enhancement of Gene Pools……………………………………………………………………………… Pag.87 1.C.9 Foreign genes as novel sources for increased efficiency of water use in rice…. Pag.90
Output 2: Integrated crop, pest and disease management 2A. RICE PATHOLOGY……….……………………….………………………………. Pag 98
2.A.1 Rice Sheath Blight (Rhizoctonia solani) ……...………………………………….. Pag. 98 2.A.2 Mite-fungus-bacterium disease complex in rice……...………………………... Pag. 106
2B. RICE VIROLOGY……….……………………….………………………………... Pag. 118
2.B.1 Screening Rice for Resistance to Rice Hoja Blanca Virus and Tagosodes orizicolus…………………………………………………………………………………………….. Pag 118 2.B.2 Developing Molecular Markers for Resistance to Rice Hoja Blanca Virus and Tagosodes orizicolus……….…………………………..…………………………… Pag 121

2C. Resistance RHBV and Selection of Advanced Breeding Generation from Crosses with Transgenic Rice Resistant to RHBV in the Field in 2007……….…………………….. Pag 127
Output 3: Intensification and diversification of rice cropping systems for small farmers. 3A. Activity title: Participatory breeding of upland rice in Nicaragua……………… Pag. 132

AAnnnnuuaall RReeppoorrtt 22000077 EExxeeccuuttiivvee ssuummmmaarryy
SSBBAA--44:: RRIICCEE
FFoorr IInntteerrnnaall CCiirrccuullaattiioonn aanndd DDiissccuussssiioonn OOnnllyy
MMaarrcchh 22000077

TABLE OF CONTENTS
1. Narrative Project Description…………………………………………….. 1 2. Project Log Frame (2007-2009)………………………………………….. 8 3. Research highlights ……………………………………………………….. 11 4. Project Outcome …………………………………………………………. 12 5. Publications………………………………………………………………. 13
• Refereed journal articles……………………………………………… 14 • In books……………………………………………………………… 15 • Posters………………………………………………………………… 15 • Oral presentations……………………………………………………. 16 • Workshops …………………………………………………………… 18 • Conferences and workshops………………………………………….. 18 • In Proceedings Scientific Meetings………………………………….. 19
6. List of proposals funded in 2007, dollar value of contract and donor…… 19 • List of Partners………………………………………………………. 20 • List of proposals submitted in 2007, value and donor……………….. 20
7. Principal and Support Staff……………………………………………….. 21 8. M. Sc. Thesis................................................................................................ 21
• Others Students ………………………………………………………… 22 9. Problems encountered and their solutions………………………………………. 23
• Indicators…………………………………………………………….. 23 • Special Project………………………………………………………….. 24 • Liaison Officer…………………………………………………………. 24
10. Summary Budget prepared by Finances……………………………………. 25

11.. CCIIAATT –– SSBBAA-- 44 RRIICCEE NARRATIVE PROJECT DESCRIPTION Rationale & Changes GGooaall:: To generate food security and employment associated with rice production with emphasis on improving the options for the small farmers. OObbjjeeccttiivvee:: To produce robust high yielding rice varieties requiring lower inputs, we will provide well-characterized progenitors and advanced materials with an ample genetic base as well as information and training. RReesseeaarrcchh OOuuttppuuttss:: Our research is organized around three major outputs. 1) Enhanced gene pools 2) Integrated crop, pest and disease management 3) Intensification and diversification of rice cropping systems for small farmers. RRaattiioonnaallee:: There are opportunities for growth in the rice sector, because land and water are more abundant in Latin America than in other rice growing regions. The rice sector faces risks because there is a trend for more open markets and many countries subsidize rice production. Most rice producers are small to mid size farmers, and the governments in this region tend not to subsidize rice production. The Rice Project focuses on strengthening the rice sector, in the low and mid altitude regions of Latin America and the Caribbean. Our activities focus on problems that are locally important, and especially in the area of pest and diseases many of these constraints are unique to Latin America and the Caribbean. Our rice breeding activities are for both irrigated and upland rice. We concentrate on developing materials with broad genetic diversity that incorporates a range of grain quality traits and resistance to the pest and diseases common in this region. To increase the genetic diversity, we work with interspecific crosses, composite populations and are developing biotechnology methods that allow the incorporation of traits more efficiently. These activities are generating both segregating populations and advanced lines, which are transferred to partners through either bilateral agreements or networks including FLAR, GRUMEGA and INGER. Our partners are then able to select advanced lines, or make their own selections from the segregating populations. These materials also serve as a source of parents in the breeding activities of our partners. Unlike Asia, most of the rice in Latin America is planted by direct seeding. Since many small farmers do not have irrigated infrastructure, we are increasing our research on rice that has better tolerance to water stress and / or has increased efficiency in water use. This is a global effort that is being coordinated through the Generation Challenge Program.
1

We have been developing upland rice breeding populations and advanced lines for many years. The tendency in Latin America has been a decline in total area of upland rice but an increase in yields in those areas that remain in production. Improved varieties have made excellent impact in the upland rice ecosystems. Jointly with the Biotechnology Research Unit and in collaboration with JIRCAS (Japan), gene technology is being explored as an alternative to incorporate increased efficiency in water use for the irrigated rice ecosystems targeting reduction of water consumption. For the small farmers, jointly with our partners, we put in place participatory variety selection and breeding schemes in different agro system of Central America. Early maturity, vigorous, high yielding varieties with adequate grain quality are some of the traits selected by farmers which gives their families the food security needed to experiment and adopt higher value crops. The recent activities to develop rice with higher levels of iron and zinc is being funded by CIDA-Canada in collaboration with Harvest Plus and should lead to the development of naturally biofortified varieties that will provide better nutrition for both the rural and urban rice consumers. To sustain yields preventing the pressure of pests and diseases, we work in collaboration with our partners to disseminate Integrated Crop Management practices. FLAR has been active in promoting Integrated Crop Management Practices. With water becoming a more expensive and/or scarce resource and the need to produce rice at competitive prices, we are working with FLAR and other partners to develop a comprehensive set of management guidelines to reduce inputs while increasing yields. These activities are information intensive and require local support to be successful. SSiiggnniiffiiccaanntt CChhaannggeess:: While the output targets for 2007 remain the same, the CIAT rice project is developing closer ties with both IRRI and WARDA. Part of the Output Target “Integrated Crop and Pest Management information developed and available in print and electronic media” in 2007 is the translation of documents from the IRRI Rice Knowledge Bank. In the area of genetic resources, CIAT and IRRI will increase our activities for INGER in Latin America.and the Caribbean. We have also agreed to the development of a common database for our genetic resources. Locally, GRUMEGA is developing a nursery that will include many of the best varieties throughout Latin America. Another area of change is the increase in cooperation between the FLAR integrated crop management activities and the integrated pest management activities of CIAT as well as in the breeding activities. In the area of Interspecific crosses, we are shifting our focus to O. latifolia which is a species that is native to the Americas. During 2007, the core resources for the rice project have been significantly reduced. These changes are reflected in the consolidation or elimination of several of the Output Targets. The 2007 Output 2 target “Characterization and development of markers for 6 major rice blast resistance genes” has been incorporated into the 2008 Output 1 target “Implementation of Marker Aided Selection techniques for quality traits, RHBV and Rice Blast in varietal development”.
2

The 2008 Output 2 target “Advanced sources of Rhizoctonia resistance available for at least five countries of LA” has been eliminated and activities in this area have been reduced. The rice project has decided to integrate the participatory rice breeding activities which were being reported in 2008 Output 3 target “Varieties and management practices developed for small rice farmers using participatory methods in 6 countries in LAC” with the Output 1 in 2008 “Integrated regional collaboration for the use and development of rice genetic resources through networking will result in at least 10 new varieties”. This output was previously reported for 2009. The marker aided selection Output 1 target has been moved from 2008 to 2009. CCGG SSyysstteemm PPrriioorriittiieess:: The Rice Project promotes the conservation and characterization of the relatives of rice. Red rice, which is a major weed is also being characterized, both to understand the origin of this pest and to consider using it as a new source of genetic diversity for selected traits. The following species: O. glaberrima, O. rufipogon, O. barthi, O. glumaepatula, O. meridionalis and most recently O. latifolia were crossed with cultivated rice (O. sativa) in efforts to increase the biodiversity of rice varieties and introduce traits of importance to Latin America. The Rice Project develops breeding populations and advanced lines with traits that include high yield, good grain quality, early vigor, strong tillers, tolerance to water stress, rice blast, Rhizoctonia, rice hoja blanca virus and the plant hopper T. orizicolus. More recently in collaboration with IRRI, we have started enhancing the nutritional quality of rice by developing lines that are higher in iron and zinc. This output is reflected in the CIAT project SB-2. The pest and disease traits that are incorporated into the new varieties are helping the farmer to reduce the use of pesticides. Using integrated crop and pest management is central to producing a sustainable agro-ecological system. The efforts to develop rice with better water use efficiency benefits the rice farmers with the least amount of infrastructure and should lead to the reduction of water in the irrigated systems. IImmppaacctt PPaatthhwwaayyss:: The Rice Project focuses on strengthening the rice sector, in the low and mid altitude regions of Latin America and the Caribbean. Our research is organized around three major outputs: 1) Enhanced gene pools; 2) Integrated crop, pest and disease management; 3) Intensification and diversification of rice systems for small farmers. Output 1: Enhanced Gene Pools This output is concerned with the characterization of genetic resources and how to use them efficiently. The Future Harvest (CGIAR) rice genetic resources are held in IRRI, WARDA and CIAT. We are collaborating to ensure that this important public good becomes characterized and catalogued in a manner that makes it more accessible to the community of rice researchers. INGER is a network, which facilitates access to these materials.
3

We maintain many activities with other advanced research institutions and these activities contribute to bring recent technologies to the regional partners. These generate knowledge and technologies that include functional genomics, marker aided selection, transgenic rice, gene flow studies, biosafety and biofortification. Many of these activities are housed in the Biotechnology Research Unit and Agrobiodiversity Project of CIAT. The CIAT contribution to regional varietal development has included populations that were derived from both O. sativa and other relatives. We attempt to develop populations with a high degree of diversity, yet include many important traits of agronomic importance. The parents of these populations normally include germplasm from our sister Future Harvest centers. CIRAD has also been a source of parental materials. We also have been in the forefront of new methods for rice breeding. The two principal methods are Recurrent Selection and Marker Aided Selection. Recurrent selection is an activity that has been promoted through GRUMEGA. During the last 10 years, it has held many Rice Breeder Workshops and many local partners have populations and advanced rice lines from these activities. This year, the second rice variety that was developed by recurrent selection was released in Bolivia by CIAT Santa Cruz and ASPAR. We are a member of FLAR and most of the FLAR germplasm is developed using some of the CIAT germplasm. FLAR includes some of the strongest rice research institutions in Latin America and this is a valuable source for Germplasm Enhancement as well as other forms of collaboration. The Regional Rice partners are responsible for the release of varieties, which is the main impact of Output 1. A very high percentage of the new rice varieties contain CIAT germplasm. Many of the varieties were developed into advanced lines before they were selected by our partners. The need for germplasm is highly variable and depends on the amount of rice production in a given country. In general, the less rice that is produced the more these programs need advanced materials. The larger rice programs use germplasm and segregating populations to make their own selections. Output 2: Integrated Crop, Pest and Disease Management. The breeding for resistance activities that are part of this output are integrated with the activities of Output 1 and achieve impacts through the same mechanisms and partners. The characterization of the pest and diseases are an essential part of this output. This work is often done in collaboration with ARI or national partners. An example of the collaboration is the analysis of why the variety Llanos 5 has maintained stable (durable) resistance to rice blast for more than 15 years. This study was conducted with Kansas State University and has elucidated that both major and minor genes are contributing to the stable resistance. This information is being used in the breeding activities and in the development of molecular markers for this disease. A molecular analysis of resistance to rice hoja blanca and its vector are also being done.
4

The counterpart of the plant resistance is the diversity of the pathogens and pests. We have conducted extensive analysis of the diversity of the rice blast fungus. Similar characterization is being done for the causal agent of sheath blight. This can help determine which varieties could be deployed to mitigate losses or applications of pesticides. The management of pest and diseases cannot be separated from crop management. FLAR is active in promoting Crop Management practices that are helping to increase the yields and lower the cost of rice production. We are working to combine IPM with the Crop Management to maximize the benefits. These activities depend on extension activities and high levels of collaboration. We have initiated in 2006, a project to study in selected rice pathogens, the development of resistance to fungicides. The objective is to develop management strategies that mitigate the probability of pathogen resistance to fungicides through the judicious use of the pesticides. This should lead to lower pesiticide use which benefits the farmers and environment. Output 3: Intensification and diversification of rice cropping systems for small farmers. This activity includes participatory rice breeding and the main mechanism is working with farmer organizations. The small farmers generally have the least amount of land, equipment, irrigation systems, and credits (infrastructure) and need upland or aerobic rice varieties that use water and fertilizers efficiently. This activity integrates the advances in breeding methodology (recurrent selection), the use of diverse germplasm including the interspecifics and in the future the high iron and zinc rice lines using participatory methods to focus on the needs of the small rice farmers. These activities help the farmers by developing their organizational skills and can aid in their eligiblity for credits and other assistance. These farmers need to be aware of other opportunities to include other crops into their agro ecosystems especially high value crops. Rice is a food security crop, that also contributes to the farmer’s income. This output involves the training activities of the project. Many of these activities are integrated with the Outputs 1 and 2. CIAT has been a source of training for many of the scientists in the rice community and we continue to play a role in the development of rice researchers, extension agents and students in LAC. Access to information is extremely important for a competitive rice sector and this is an area in which more resources need to be devoted. The CIAT site strives not to duplicate information that is on other sites but to be a resource to help find the most useful information. IInntteerrnnaattiioonnaall PPuubblliicc GGooooddss:: The “International Treaty on Plant Genetic Resources for Food and Agriculture” is an international agreement governing many of the world’s most important crop diversity collections. The treaty will ensure that this diversity, which is critical for the rice crop improvement will remain in the public domain. In the area of germplasm, CIAT has decided to place most of its elite lines into this system. To do this, we will use the database format of IRRI and these should become part of the Future Harvest genetic resources.
5

Most of the technologies including database management programs, breeding methodologies, rice lines which are developed at CIAT enter into the public domain as international public goods. One of the most relevant and important outputs of the CIAT Rice Project is the development and deployment of interspecific rice lines derived from crosses between wild rice species and cultivated rice. Most of our partners and NARs in LAC are not in a position to carry out this type of breeding work since they lack the expertise, resources and funding to do it. Besides, they are more concerned with the development of improved lines to address production problems that impinge on today’s rice production but not on broadening the genetic base of rice or on problems for which no sources of genetic resistance are known. PPaarrttnneerrss:: IRRI and WARDA are CGIAR institutions working on rice and with whom we collaborate in germplasm exchange and on problems of global importance. The Generation Challenge Program and INGER are two of the major joint activities. We have an alliance with CIRAD and IRD of France, which is vital to our research activities. Two CIRAD scientists and one IRD scientist hold joint appointments with the CIAT rice project and contributed extensively to activities in Output 1 and Output 3. To increase our impact, we are a member of FLAR. This network includes members from fourteen countries. FLAR is a partnership of the private and public sectors for the international research of rice. Its mission is to generate new technologies to allow the Latin American rice sector to become more competitive, profitable and efficient with low environmental impact practices that propitiate lower unit costs and, as a consequence, lower rice prices to consumers. It generates both genetic resources (contributing to Output 1) and technology transfer of integrated crop management practices (contributing to Output 2 and 3). The AgroSalud Project (housed in SB-2) is aiming at increasing the iron and zinc content in the rice grain includes partners throughout the region. Brazil EMBRAPA & IRGA, Colombia FEDEARROZ, CORPOICA, U. Nacional U. del Tolima & U. de Antioquia, Peru INIA, Venezuela INIA, IVIC, FUNDARROZ & DANAC, Cuba IIA, Nicaragua INTA, Costa Rica CONARROZ, SENUMISA, INTA & U. Costa Rica, Guatemala ARROZGUA, Mexico Consejo Mexicano del Arroz, Bolivia CIAT Santa Cruz, ASPAR & CONARROZ, Dominican Republic IDIAF, Chile INIA, Panama U. de Panama, Uruguay INIA, Argentina INTA, CIB-FIBA, U. Corrientes & U. Tucuman are national institutions and we have activities many of which are carried out using the networks of FLAR, GRUMEGA, INGER and Biofortification. Many of these institutions develop rice varieties while other are more involved in the transfer of technologies to the rice farmers. Universities including KSU, Cornell, Purdue, LSU, U. Arkansas, Texas A&M, U. Missouri, Rutgers, and Yale. We have collaborative projects and students that work on research of mutual interest. IAEA collaborates in the use of induced mutations for crop improvement.
6

PPrroojjeecctt FFuunnddiinngg::
Budgeting 2005-2009
Year 2005 (actual)
2006 (estimated)
2007 (proposal)
2008 (plan)
2009 (plan)
US Dollars (millions)
2.621
2.450
2.017
1.946
1.920
7

22.. CCIIAATT:: SSBBAA--44 PPRROOJJEECCTT LLOOGG FFRRAAMMEE ((22000077--22000099))
PPRROOJJEECCTT:: IIMMPPRROOVVEEDD RRIICCEE FFOORR LLAATTIINN AAMMEERRIICCAA AANNDD TTHHEE CCAARRIIBBBBEEAANN PPRROOJJEECCTT MMAANNAAGGEERR:: Outputs Intended users Outcome ImpactOutput 1: Enhanced gene pools
A robust rice sector will generate employment and maintain low rice prices for the consumers. The expansion of the genetic base of rice is leading to yield stability and better adaptability for abiotic and biotic stresses.
Output Targets 2007 .
Advanced lines arising from interspecific crosses and recurrent selection will have been widely distributed and tested in more than 11 countries throughout the region
FLAR and rice breeding programs throughout the region
These interspecific crosses and recurrent selection populations will be a major basis for expanding the genetic diversity of cultivated rice in LAC.
Output Targets 2008
Integrated regional collaboration for the use and development of rice genetic resources through networking will result in at least 10 new varieties.
FLAR, GRUMEGA, INGER-LAC and Rice breeding programs throughout the region
Increase interactions and efficiency of Rice Breeding Programs throughout LAC.
Output Targets 2009 Implementation of Marker Aided Selection techniques for quality traits, RHBV and Rice Blast in varietal development.
FLAR and Rice breeding programs throughout the region
Rice breeding strategies for evaluation and selection of promising rice lines that result in more and better varieties released by the at a faster rate by rice sector.
8

CCIIAATT:: SSBBAA--44 PPRROOJJEECCTT LLOOGG FFRRAAMMEE ((22000077--22000099))
PPRROOJJEECCTT:: IIMMPPRROOVVEEDD RRIICCEE FFOORR LLAATTIINN AAMMEERRIICCAA AANNDD TTHHEE CCAARRIIBBBBEEAANN PPRROOJJEECCTT MMAANNAAGGEERR:: FFEERRNNAANNDDOO CCOORRRREEAA ((CCEESSAARR MMAARRTTÍÍNNEEZZ AASS OOFF 0022//22000088)) Outputs Intended users Outcome ImpactOutput 2: Integrated crop, pest and disease Management
Advanced techniques to develop resistant varieties faster and better mananagement practices will lower the use of agrochemicals and mitigate contamination to the farmers and environment. The farmers will be able toproduce rice at a lower cost.
Output Targets 2007 .
Control strategies implemented for the pest and disease complex associated with the invasive mite, S. spinki.
Rice scientists, extension agents and rice farmers
The use ofpest management practices to reduce losses caused by the complex of S. spinki and diseases.
Output Targets 2008
Identification of rice hoja blanca and planthopper resistance genes
Rice pathologists and breeders
The development of rice varieties with increased resistance to rice hoja blanca virus and T. orizicolus.
Output Targets 2009
Implementation of strategies to prevent development of fungicide resistance in rice pathogens
Rice scientists, extension agents, rice farmers and regulatory agencies
Implementation of strategies for controlling and managing problems associated with the development of resistance to pesticides
9

CCIIAATT:: SSBBAA--44 PPRROOJJEECCTT LLOOGG FFRRAAMMEE ((22000077--22000099))
PPRROOJJEECCTT:: IIMMPPRROOVVEEDD RRIICCEE FFOORR LLAATTIINN AAMMEERRIICCAA AANNDD TTHHEE CCAARRIIBBBBEEAANN PPRROOJJEECCTT MMAANNAAGGEERR:: FFEERRNNAANNDDOO CCOORRRREEAA ((CCEESSAARR MMAARRTTÍÍNNEEZZ AASS OOFF 0022//22000088)) Outputs Intended users Outcome ImpactOutput 3: Intensification and diversification of rice systems for small farmers
More competitive rice production so that the sector thrives even when faced with more open markets. This will lead to a dynamic and robust rice sector that improves the livelihoods of small farmers.
Output Targets 2007 .
Integrated Crop and Pest Management information developed and available in print and electronic media.
Rice scientists, extension agents and rice farmers.
The information will be used in participatory farm groups as well as other rice farmers to reduce the yield gap and intensify the farming system. Also a large volume of scientific literature will be published.
Output Targets 2008
Output Targets 2009
10

3. Research highlights • Two mapping populations were evaluated for their sheath blight reaction using
two different greenhouse screening methods. Preliminary QTL analysis using more than 100 microsatellite markers identified several QTLs on at least 7 rice chromosomes. Several of the QTLs identified are similar to other previously mapped QTLs.
• We have corroborated the presence of the bacterial panicle blight in Panama, Costa Rica, Nicaragua, and Colombia. Our studies have provided convincing evidence of the association of this bacterium to the disease symptoms observed in all these countries. Bacterial panicle blight has the potential to become a major constraint to rice production in Latin America. We have identified potential sources of resistance to the disease, which will probably offer good management alternatives in the near future.
• Among 3189 lines analyzed, 38 and 32 were found to reliably meet or exceed the iron 6-8 ppm target range and the zinc 22-25 ppm target range for milled rice, respectively. In collaboration with Harvest Plus, it was determine that NIRs technology can be successfully used to estimate iron and zinc content in rice grain.
• In close collaboration with US-Universities and RiceTec, high yield QTLs identified from O.rufipogon were verified to have a yield enhancing impact when introgressed into Jefferson.
• Collaborative work by FLAR-CIAT showed that large panicles alone do not automatically result in high yield; high spikelet fertility and lodging resistance are required. Additionally, dark green leaf color could be an important secondary trait for yield.
• The web page of the GRUMEGA network (http://www.grumega.org ) was launched and a breeding network Red-Mega was also established with FAO support.
• A systematic morphological, phenological and molecular analysis of 154 accessions of wild Oryza species from diverse environments in Venezuela was carried out in collaboration with Venezuelan scientists.
• Further work done in relation to RHBV and T.orizicolus confirmed that in Fedearroz 50 and Fedearroz 2000 there is a QTL associated with resistance to RHBV on the short arm of chromosome 4. In Fedearroz 2000, there is a QTL associated with resistance to RHBV on chromosome 5. In Fedearroz 50 there is a QTL associated with resistance to T.orizicolus on chromosome 7.
• After five years of work in Nicaragua, the PVS strategy succeeded in identifying four high performing lines that will be released by our Nicaraguan partners in 2008 covering a wide range of production systems in Nicaragua.
• Ten varieties were released by our partners in Colombia, Venezuela, Costa Rica, and Bolivia, Chile and El Salvador.
11

4. Project Outcome Output 1: Enhanced gene pools in IP-4 and OUTPUT2 in SB-2:Genomes modified: genes and gene combinations used to broaden the genetic base of crops (rice ). Where was the achievement of the output/output target documented? : Poster presented at the Rice Technical Working Group Meeting: Enhancement of yield through chromosomal introgressions from Oryza rufipogon, 2008.Anna McClung , Shanon Moon, Georgia Eizenga, Susan Mc Couch. 32nd Rice Technical Working Gropup Meeting, San Diego, California. February 18-21,2008, Another paper to be presented at the PCCMCA meeting; AVANCES DEL PROYECTO AGROSALUD EN EL CULTIVO DE ARROZ, SANTA CRUZ-BOLIVIA. R. Taboada1, J. Viruez1, V.H Callaú1 C. Martinez2 . San Jose. Costa Rica. Abril 14-19,2008. Who used/adopted or was influenced by the output? Identify the kinds of people or entities affected and their location by country.: USDA-Dale Bumpers National Rice Research Center, Texas A& M University, Cornel University, University of Arkansas, Rice Tec, Rice Program of CIAT-Santa Cruz and ASPAR, Bolivia, and INTA-Nicaragua. How was the output used or adopted? What was the nature of the influence of the output? The introduction of selected “wild QTLs” into commercial varieties/elite lines has the potential to enhance the performance of rice varieties. It was demonstrated that specific O.rufipogon introgressions confer superior performance for an array of agronomic and yield related traits. In the case of Bolivia, two inter-specific breeding lines derived form the cross Oryzica3 X O.rufipogon have gone through two years of field evaluations in farmer’s fields showing superior performance compared to available commercial varities in Bolivia and are being considered for release. Another breeding line from the cross Caiapox O.glaberrima went through a participatory breeding selection with farmers in Nicaragua and is being considere for varietal release by INTA. What is the magnitude (in terms of, for example, geographic coverage or reach of intended user groups) of the outcome relative to the intended recommendation domain? In the case of “wild QTLs”from the cross Jefferson x O.rufipogon, rice breeding programs in USA will benefit from this discovery as well as RiceTec(hybrid rice company) based in Texas is considering to introgress these QTLs into their hybrids. In the case of Bolivia, farmers growing rice under irrigated and rainfed conditions will benefir, along as small-resource farmers growing rice under upland conditions in Nicaragua. What is the evidence for the outcome? Specifically, what kind of data were collected or study was conducted? Who collected the data and/or conducted the study? If not included in the outcome evidence, what is the evidence that shows that the outcome is derived from the output/output target? Anna McClung , Shanon Moon, Georgia Eizenga, and Susan Mc Couch collected yield and agronomic data on 70 NILs planted in comparison with three commercial varieties and one hybrid in four locations in Texas and Arkansas in replicated yield trials in 2007. In the case of Bolivia , Roger Taboada , Juana Viruez and V.H. Callau collected yield and agronomic data in regional yield trials planted in different rice growing regions in Boliobia during 2006 and 2007. In Nicaragua, Gilles Trouche(CIRAD) , Zildghen Chow, Lazaro Narvaez, Jose Corrales and Marlon Ortega from INTA conducted and collected the data via participatory breeding.
12

5. Publications Nandakumar, R., Rush, M.C., and Correa-Victoria, F.J. 2007. Association of Burkholderia glumae and B. gladioli with Panicle Blight Symptoms on Rice in Panama. Plant Disease 91:767 Correa-Victoria, F.J., Perez, C.R., y Saavedra, E. 2007. Añublo bacterial de la panicula del arroz Burkholderia glumae. ARROZ 57(468):26-32 Fuentes, J.L., Correa-Victoria, F.J., Escobar, F., Prado, G., Aricapa, G., Duque, M.C., and Tohme, J. 2007. Microsatellite markers linked to the blast resistance gene Pi-1 in rice for use in marker assisted selection. Euphytica (accepted 28 June 2007. DOI 10.1007/s10681-007-9497-0) Correa-Victoria, F.J., 2007. Identification of resistance gene combinations conferring durable blast resistance in Colombia. Phytopathology 97:S24 Jia, Y., Correa-Victoria, F.J., McClung, A., Zhu, L., Wamishe, Y., Xie, J., Marchetti, M., Pinson, S., Rutger, N., and Correll. J. 2006. Rapid determination of rice cultivar responses to the sheath blight pathogen Rhizoctonia solani using a micro-chamber screening method. Plant Disease 91:485-489 Calvert, L.A., Cuervo, M., Lozano, I., Villareal, N., and Arroyave, J., 2008. Identification of three strains of a virus associated with cassava plants affected by frogskin disease. Journal of Phytopathology (In press) McCouch, S.R.,M.Sweeney, Jimming Li,Hui Jiang, M.Thomson, E.Septiningsih, J.Edwards, P.Moncada, Jinhua Xian, A.Harris, Tom Tai, Cesar Martinez, J.Tohme, M.Sugiono, Anna McClung, Long Pin Yuan, San Nag Ahn, . 2007. Through the bottleneck : O.rufipogon as a source of trait-enhancing alleles for O.sativa. Euphytica 154:317-339. Cesar P. Martinez, J.Borrero, J.Tohme, Myriam C.Duque, Silvio James Carabali, James Silva. 2007. Variedades de arroz con mayor valor nutricional para combatir la desnutricion en América Latina. Revista ASIAVA 76:5-9. Alvarez A., Fuentes JL, Puldon V, Gomez PJ, Moral L, Duque MC, Gallego G, and Tohme J ,2007, “Genetic diversity analysis of Cuban traditional rice (Oryza sativa L.) varieties based on microsatellite markers” Genet. Mol. Biol., 2007, vol.30, no.4, p.1109- 1117. ISSN 1415-4757. Pantoja A, Triana M, Bastidas H, García C, and Duque MC, 2007, “Damage by Tibraca limbativentris (Hemiptera: Pentatomidae) to rice in Southwestern Colombia”;J. Agric. Univ. P.R. 91(1-2):11-18 (2007)
13

Nandakumar, R., Rush, M.C., and Correa-Victoria, F.J. 2007. Association of Burkholderia glumae and B. gladioli with Panicle Blight Symptoms on Rice in Panama. Plant Disease 91:767 Larmande P, Gay C, Lorieux M, Perin C, Bouniol M, Droc G, Sallaud C, Perez P, Barnola I, Biderre-Petit C, Martin J, Morel J, Johnson A, Bourgis F, Ghesquière A, Ruiz M, Courtois B, Guiderdoni E (2008) Oryza Tag Line, a phenotypic mutant database for the Génoplante rice insertion line library. Nucleic Acids Res 36 :D1022-D1027 Torres, EA. and Geraldi IO. 2007. Partiall diallel analysis of agronomic characters in rice (Oryza sativa L.). Genetics and Molecular Biology 30 (3). 605 -613. - Red de Mejoramiento Genético de Arroz en América (Red-MeGA): Development of the “Estatutos y Reglamento para la Creación y el Funcionamiento de la Red-MeGA”. - Slide Show: Mejoramiento Genético de Arroz. Selección Recurrente utilizando Androesterilidad Genética: un nuevo Método de Selección. Refereed journal articles
McCouch, S.R.,M.Sweeney, Jimming Li,Hui Jiang, M.Thomson, E.Septiningsih, J.Edwards, P.Moncada, Jinhua Xian, A.Harris, Tom Tai, Cesar Martinez, J.Tohme, M.Sugiono, Anna McClung, Long Pin Yuan, San Nag Ahn, . 2007. Through the bottleneck : O.rufipogon as a source of trait-enhancing alleles for O.sativa. Euphytica 154:317-339. Trouche G., Aguirre Acuña S., Hocdé H., Obando Solís R., Gutiérrez Palacios N., Chow Wong Z. Valorisation de la diversité génétique du sorgho par des approches de sélection participative au Nicaragua. Valorisation of the genetic diversity of sorghum through methodologies of participatory breeding in Nicaragua. Cahiers Agricultures. Numéro spécial Agrobiodiversités. In press. Vom Brocke K., Trouche G., Zongo S., Bitié A., Barro-Kondombo C., Weltzien E., Chantereau J. Création et amélioration de populations de sorgho à base large avec les agriculteurs au Burkina Faso. Cahiers Agricultures. Numéro spécial Agrobiodiversités. Lentini, Z. 2007. Estimating Likelihood and Exposure. Environmental Biosafety Research 5 (4): pp. 193-195 Lentini Z, D. Debouck, A.M. Espinoza, and R. Araya. 2007. Gene flow analysis into wild/weedy relatives from crops with center origin/ diversity in tropical America. Environmental Biosafety Research 5 (4)
14

In books Calvert, L., 2008. Tenuiviruses, In: Compendium of Wheat Disease Second Edition. Eds. Bill Bock, Published by The American Phytopathological Society, St. Paul, Minnesota, USA. (In Press) Calvert L.A. and Z. Lentini. 2007. Rice Hoja Blanca Virus. In: Characterization, Diagnosis and Management of Plant Viruses. Vol. 4: Grain Crops and Ornamentals. Govind P. Rao, Claude Bragard and Benedicte S.M. Lebas (Editors). Stadium Press ILLC, Texas, USA. ISBN 1-933699-34-5. p: 85-99. Correa-Victoria, Fernando., 2007. Using rice differentials with known blast resistance genes for pathogen characterization and improvement of rice cultivars in Latin America. P.123. In: A differential system for blast resistance for stable rice production environment. Ed: Y. Fukuta, C.M. Vera-Cruz, and N. Kobayashi. JIRCAS Working Report No. 53. 123p. Correa-Victoria, F.J., 2007. Algunas consideraciones: Protección química de los cultivos. Ventana al Campo 3: 4-8 Online publications Correa-Victoria, F.J., 2007. The rice Tarsonemid mite: Steneotarsonemus spinki Smiley. RiceCAP Fact Sheet. 4 pages. www.ricecap.uark.edu/outreach_downloads.html - Web Page Development : http://www.grumega.org Posters Luisa. F. Fory, Mabel. Morales, Alicia Velazquez, Kiliany Arcia, Andrés E. Blanco, Eliana González, Myriam C. Duque, and Zaida Lentini. 2007. Gene flow and introgression analysis from rice into wild/weedy relatives in center of diversity in tropical America. VI Encuentro Latinoamericano y del Caribe de Biotecnología Agropecuaria REDBIO 2007. Viña del Mar, Chile. October 22-26, 2007. Eliana González, Luisa. F. Fory, Kiliany Arcia, Andrés E. Blanco, Aida Ortiz, Iris Pérez, Myriam C. Duque, and Zaida Lentini. 2007. Genetic diversity analysis of weedy rice and wild oryza species collected in crop-natural ecosystems contact zones. VI Encuentro Latinoamericano y del Caribe de Biotecnología Agropecuaria REDBIO 2007. Viña del Mar, Chile. October 22-26, 2007.
.
Manuel Quintero, Luisa Fory, Eddie Tabares, César P. Martínez and Zaida Lentini 2007. Transgenic rhbv resistance and breeding selection of advanced lines in the field. VI Encuentro Latinoamericano y del Caribe de Biotecnología Agropecuaria REDBIO 2007. Viña del Mar, Chile. October 22-26, 2007. *
15

Sanabria, Y; Carabalí, J; Giraldo, O ; Martínez, C; Tohme, J. Using molecular markers SSR to search wild introgressions from a relative tetraploid species in the diploid Oryza sativa L. VI Encuentro Latinoamericano de Biotecnología Agropecuaria REDBIO/2007. Viña del Mar – Chile, 22 - 26 Octubre 2007. Giraldo O. X*., Quintero C., Plata G., Rodriquez F., Borrero J., Martinez C. P. y Tohme J.2007. Identification of SNPs Markers for Biofortification in Rice . VI Encuentro Latinoamericano de Biotecnología Agropecuaria REDBIO/2007. Viña del Mar – Chile, 22 - 26 Octubre 2007. Sanabria, Y; Olaya, C; Carabalí, J; Martínez, C; Tohme, J. Eliminación cromosómica como estrategia para la recuperación de la fertilidad en híbridos interespecíficos de arroz. II Simposio Latinoamericano de Citogenética y evolución. Palmira – Valle. Agosto 2007. Sanabria, Y; Carabalí, J; Olaya, C; Martínez, C; Tohme, J. Cruzamientos interespecíficos entre una especie de arroz tetraploide (Oryza latifolia) y el diploide (Oryza sativa). X congreso colombiano de fitomejoramiento. Pasto – Nariño. Junio 5-7. 2007. Sanabria, Y; Carabalí, J; Giraldo, O ; Martínez, C; Tohme, J. Introducción de genes de una especie de arroz tetraploide (Oryza latifolia) en cultivares diploides (Oryza sativa) por medio de cruzamientos. IV Congreso Colombiano de Botánica. Medellín – Antioquia, Abril 2007. Castilla Lozano Luis Armando( FEDEARROZ) ,Martínez, Cesar P. Evaluación de líneas interespecíficas de arroz (Oryza sp) a la inoculación con bacterias fijadoras de nitrógeno azotobacter chroococcum y azospirillum amazonense en un suelo typic haplustalf de la meseta de Iibagué, colombia. XVII Cogreso Latinoamericano de la Ciencia del Suelo, Leon ,Guanajuato,Mexico, Septiembre 17-21,2007.
Péricles de Carvalho F. Neves, Priscila Zaczuk Bassinello, Orlando Peixoto de Morais, Jaime R. Fonseca, José Almeida Pereira, José Luíz Viana De Carvalho, Marília R. Nutti, Cesar Martínez, Helena Pachon. 2007. Germplasm identification and development of upland rice cultivars with high zinc contents. Zinc Crops 2007, Improving Crop Production And Human Health. Maio 24-26, 2007, Istanbul, Turquia.
Oral presentations Correa-Victoria, F.J. 2006. Situación del complejo acaro-hongo-bacteria en el arroz. Segundo Congreso Arrocero. San José, Costa Rica, Junio 29-30, 2006. (invited speaker) Correa-Victoria, F.J., and Martinez, C. 2007. Breeding rice cultivars with durable blast resistance in Colombia. 4th. International Rice Blast Conference. Changsha, China. Oct. 9-14, 2007 (invited speaker)
16

Correa-Victoria, F.J., 2007. Identification of resistance gene combinations conferring durable blast resistance in Colombia. APS Annual Meeting. San Diego, CA. July 27-August 2. McClung, A.M., Groth, D.E., Oard, J.H., Utomo, H., Moldenhauer, K.A.K., Boza, E., Scheffer, B., Jia, Y., Liu, G., Correa-Victoria, F., and Fjellstrom, R.G. 2007. Development and characterization of RiceCap QTL mapping population for sheath blight resistance. ASA Meeting, New Orleans, LA. Nov 3-9. 2007. Lentini, Z. 2007. Gene flow analysis in centers of origin and diversity. VI Encuentro Latinoamericano y del Caribe de Biotecnología Agropecuaria REDBIO 2007. Viña del Mar, Chile. October 22-26, 2007. Lentini, Z. 2007. Development and evaluation of transgenic rice in tropical America. University of Tokyo. Japan. July 12, 2007. Lentini, Z. 2007. Environmental safety of crops with center origin/ diversity in tropical America. NIAS. Tsukuba, Japan. July 11, 2007. Lentini, Z. 2007. Development of Drought tolerant rice via transgenesis. JIRCAS. Tsukuba, Japan. July 9, 2007 Lorieux M. 2007. MapDisto, a free user-friendly program for computing genetic maps. Computer demonstration given at the Plant and Animal Genome XV conference, Jan 13-17 2007, San Diego, CA. URL: http://mapdisto.free.fr/.
Martinez, César P. 2007.Development of high iron and zinc rice lines to combat malnutrition in Latin America and the Caribbean. 2nd Annual Harvest Plus and AgroSalud BIOFORTIFICATION MEETING.11 –14 November 2007, Brazil.
Martínez, César. P., Jaime Borrero, J. Tohme, Myriam. C. Duque, Silvio. J. Carabalí y James. Silva.2007. Desarrollo de germoplasma de arroz con mayor valor nutricional para combatir la desnutrición en América Latina . X Congreso Colombiano de Fitomejoramiento. San Juan de Pasto – Junio 5-7 , 2007. Martínez, C.P., Borrero, J., Carabalí, J., Duque, M.C., Correa, F., Sanabria, Y., Giraldo, O., Silva, J. 2007. Contribución de las especies silvestres en el mejoramiento del arroz cultivado de América Latina. X Congreso Colombiano de Fitomejoramiento. San Juan de Pasto – Junio 5-7 , 2007. Jaime Borrero C. and César P. Martínez. 2007. Contenido de hierro y zinc en arroz integral y pulido en algunos países de América Latina y el Caribe. XII Congreso Colombiano de nutrición y dietetica .Armenia-Quindío 2,3 y 4 de Agosto de 2007
17

César P. Martinez .2007. Development of high iron and zinc rice lines to combat malnutrition in Latin America and the Caribbean. HARVEST PLUS RICE CROP TEAM/BIOFORTIFICATION MEETING.2 – 6 November 2007, Thailand Martínez César P. 2007.Evaluation of Drought Tolerance of Transformed Upland Rice at CIAT . Kick-off Meeting of the MAFF Funded Project .JIRCAS,TSUKUBA, JAPAN
JULY 9-11,2007
Mathias Lorieux, 2007. Exploring Rice Diversity. An update on wild introgression populations and related tools Generation CP ARM.Benoni – Sept 2007 Gutiérrez, Andrés Gonzalo, César Pompilio Martínez ,Olga Ximena Giraldo, Joe Tohme,and Mathias Lorieux,2007. Desarrollo y evaluación de líneas de introgresión interespecíficas de arroz (Oryza sativa L.). . X Congreso Colombiano de Fitomejoramiento. San Juan de Pasto – Junio 5-7 , 2007. Martinez, César P ,Helena Pachón, Jaime Borrero.2007.Cultivos básicos biofortificados para abordar deficiencias nutricionales y la inseguridad alimenticia nutricional en Latinoamérica y el Caribe. Conference presented at the 5th Rice Producers Meeting, Santa Cruz, Bolivia, Octuber 12, 2007
Workshops Training on Advanced Rice Pathology. Palmira-Colombia. November 26-30, 2007. 10 participants from the Minister of Agriculture from Panama. Training course on Rice breeding : Project TCP/RLA/3102 (A). Chillán – Chile. January 22 - 25 del 2007. 30 participants from several NARs Training course on Rice Breeding Project TCP/RLA/3102 (A). Managua- Nicaragua. May 27 – 31, 2007.: 30 participants from different NARs. Lentini, Z. 2007. América Latina: Construcción de Capacidad Multi-País en Bioseguridad (Brasil, Colombia, Costa Rica y Perú). Centro Internacional de Agricultura Tropical, CIAT. Cali, Colombia. Miércoles Junio 13, 2007. Funded by GEF-World Bank. Lentini, Z. 2007. Workshop on Knowledge Generation for Biosafety Risk Assessment and Management. June 14-15, 2007. CIAT Cali, Colombia. Funded by GTZ Germany. Conferences and workshops Correa-Victoria, F.J. 2006. Situación del complejo acaro-hongo-bacteria en el arroz. Segundo Congreso Arrocero. San José, Costa Rica, Junio 29-30, 2006. (invited speaker)
18

Correa-Victoria, F.J., and Martinez, C. 2007. Breeding rice cultivars with durable blast resistance in Colombia. 4th. International Rice Blast Conference. Changsha, China. Oct. 9-14, 2007 (invited speaker) Correa-Victoria, F.J., 2007. Identification of resistance gene combinations conferring durable blast resistance in Colombia. APS Annual Meeting. San Diego, CA. July 27-August 2. McClung, A.M., Groth, D.E., Oard, J.H., Utomo, H., Moldenhauer, K.A.K., Boza, E., Scheffer, B., Jia, Y., Liu, G., Correa-Victoria, F., and Fjellstrom, R.G. 2007. Development and characterization of RiceCap QTL mapping population for sheath blight resistance. ASA Meeting, New Orleans, LA. Nov 3-9. 2007. Martinez, César P. , Helena Pachon. Taller Biofortificacion con diferentes Instituciones del sector salud y nutrición . La Habana, Cuba. Agosto 20-21, 2007. In Proceedings Scientific Meetings Lentini, Z. 2007. Gene flow analysis in centers of origin and diversity. VI Encuentro Latinoamericano y del Caribe de Biotecnología Agropecuaria REDBIO 2007. Viña del Mar, Chile. October 22-26, 2007. Lentini, Z. 2007. Development and evaluation of transgenic rice in tropical America. University of Tokyo. Japan. July 12, 2007. Lentini, Z. 2007. Environmental safety of crops with center origin/ diversity in tropical America. NIAS. Tsukuba, Japan. July 11, 2007. Lentini, Z. 2007. Development of Drought tolerant rice via transgenesis. JIRCAS. Tsukuba, Japan. July 9, 2007. 6. List of proposals funded in 2007, dollar value of contract and donor
• Gene Flow Analysis for Environmental safety in the Tropics. CIAT – University of Costa Rica – Hannover University and BBA, Germany. Donor: EURO 450,000 (2005-2008).
• Impacto ambiental de la adopción del arroz resistente a las imidazolinoas en sistemas productivos contrastantes de América Latina (AL). INIA-UCV-CIAT. USD 420,000. Donor: Fontagro. (2007-2010).
MAFF Funded Project “Promotion of Research Targeting Stable Supply of Global Food”. Japan, US$ 300,000/year , 3 years.
• AgroSalud: High iron and zinc rice lines. CIDA- Canada, US$ 238,000/year
19

List of Partners
• COLCIENCIAS-Colombia • CONAGRO, CALESA, IDIAP, MAG-Panama • CNPAF-EMBRAPA-Brazil • DALE-BUMPERS-NATIONAL RICE RESEARCH CENTER-USA • FEDEARROZ-Colombia • INTA-Nicaragua • INIA-Venezuela • INIA-Peru • IRRI-Philippines • IAEA-Austria • Louisiana State University-USA • SENUMISA, CONARROZ-Costa Rica • USDA-USA • University of Arkansas, Texas A &M U, Cornell U, and Yale U,USA • WARDA-Benin • CIBIOGEM, UNAM, Mexico • University of Costa Rica, Costa Rica • ICA, Corpoica, Institute von Humboldt, Colombia • UCV, IVIC, INIAP, Venezuela • CONAM, INIA, Peru • EMBRAPA, Brazil • University of Hanover, BBA, University of Braunschweig, Germany • ETH, Switzerland • PRI, University of Wageningen, The Netherlands • JIRCAS, Japan • CIAT-Santa Cruz , ASPAR, and Mision Alianza Noruega, Bolivia • Instituto Investigaciones del Arroz, Cuba • IDIAF. Dominican Republic • IDIAP, Panama • Harvest Plus-Rice • CIRAD,IRD, Genoplante, France • INIA.Chile
List of proposals submitted in 2007, value and donor Germoplasma mejorado de Arroz de Riego de Amplia Base Genética y Nuevas Características de Interés” MADR, Colombia, Total US 246,944. Introgresión asistida por marcadores de genes de resistencia al virus de la hoja blanca en líneas elites de arroz. MADR, Colombia, Total Col$ 627,7 millions.
20

7. Principal and Support Staff
• Principal Staff
Principal Staff Allocation of time Affiliations Location
IP-4 Other Dr. Lee Calvert 70% CIAT CIAT HQ Dr. Marc Chatel 100% CIRAD/CIAT CIAT HQ Dr. Fernando Correa 100% CIAT CIAT HQ Dr. Zaida Lentini 20% 80% SB-2 CIAT CIAT HQ Dr. Mathias Lorieux 50% 50% SB-2 IRD/CIAT CIAT HQ Dr. César Martínez 25% 50% SB-2 CIAT CIAT HQ Dr. Gilles Trouche 50% 50% PE-3 CIRAD/CIAT CIAT HQ Principal Staff positions in IP-4: 4.9 Associated projects 2.3 Total 7.2 Dr. Carlos Bruzzone 50% 50% (INIA) INIA/CIAT Peru Works as a consultant
• Support Staff Jaime Borrero, Genetics Myrian Cristina Duque, Biometry Fabio Escobar, Biotechnology/Pathology Iván Lozano, Virology Yolima Ospina, genetics Gustavo Prado, Pathology Manuel Quintero, Tissue Culture James Silva, Biometry Silvio James Carabalí, Genetics Jaime Lozano, Genoplant Jairo Barona, Database Eddy Tabares, Transformation 8. M. Sc. Thesis:
• Olga Higuera, FEDEARROZ Supervisor: Fernando Correa University: Universidad Nacional de Colombia Title: Correlation of blast resistance selected in early generations (F2-F3) and stability of resistance in advanced generations.
• Mabel Morales. B.Sc. Identificación y Caracterización de especies de los
complejos Oryza sativa y Oryza officinalis. Tesis en Curso. Universidad Javeriana, Bogotá, Colombia. Graduated January 2007
21

• Alicia Milena Velásquez. B.Sc. Rastreo de flujo de genes en campos comerciales
de arroz utilizando marcadores moleculares y resistencia a herbicidas. Tesis en Curso. Universidad Javeriana, Bogotá, Colombia. Graduated Decemeber 2007.
• Kiliany Andrea Arcia Moreno. M.Sc. Tesis: Caracterización de especies silvestres
del género Oryza colectadas en condiciones naturales y campos de arroz en el Estado Portuguesa, Venezuela. Tesis en Curso. Universidad Nacional, Sede Palmira.
• Andrés Eloy Blanco. M.Sc. Tesis: Caracterización de especies silvestres del
género Oryza colectadas en condiciones naturales y campos de arroz en el Estado Guárico, Venezuela. Tesis en Curso. Programa sándwich Universidad Nacional, Sede Palmira/ Universidad Central de Venezuela, Maracay, Venezuela..
• Francisco Amella MSc Thesis: Inheritance of the rice panicle size (Oryza Sativa
L.) Thesis research finished but statisctical analysis of data is on going. Universidad Nacional of Colombia, Sede Palmira.
• Olga Ximena Giraldo. Ms Thesis. Identification of SNPs Markers for
Biofortification in Rice. Research activities on –going. Universidad Nacional de Colombia. Palmira.
• Natalia Labrin MSc CATIE, Costa Rica Estudio de la resistencia en variedades de
arroz (Oryza sativa L.) venezolanas al virus de la hoja blanca. (Degree completed Feb. 2007)
Others Students
• Erique Bravo PhD U. of Valle, Colombia. The complete nucleotide sequence of the genomic RNA of Bean common mosaic virus strain NL4. (Degree to be completed March 2008)
• Monica Fernandez BSc. U. Nacional de Palmira, Colombia.
• Herminio Paredes. Universidad Nacional, Palmira, 6 months training in rice
agronomy. • Liliana Gil. Universidad Javeriana, Bogota. 6 months training in molecular
markers associated with rice blast.
22

9. Problems encountered and their solutions • We are developing two Projects funded by FONTAGRO. At the time the Projects
were developed, the Rice Pathology section had operational funds, which were offered as our matching funds to the funds of the FONTAGRO Projects. Today, our section has no operational funds and several charges, including technical and research support were charged to the FONTAGRO Projects in 2007, affecting the normal budget execution, and with the risk that FONTAGRO could not accept the financial report. No solutions were given to this problem, which will continue in 2008.
• Two well known, experienced, and respected scientists will be departing in 2008 reducing drastically our critical mass of scientists. This fact has already increased the level of anxiety and concern in our rice people. CIAT management has made some commitments to fill these positions with post-doctoral fellows. However, there will be some lead-time for the new scientists to get established and be productive. Therefore, there is a risk that the productivity of the Rice Product Line could be affected.
• Core funding for the Rice Product Line is still not clear and is not adequate. As a matter of fact we lost the administrative assistance position in 2007, which has created some logistic problems for us. A possible solution could be to bring more money through special projects. However, our senior staff, research assistants, and technicians are over-committed and over-work Human Resources is calling our attention because our people are not getting due rest on time.
• Future activities of the Rice Anther Culture laboratory are uncertain since this laboratory lost all its financial support in 2007. Activities will depend on third financial support.
Indicators List of technologies, methods and tools
• Two sheath blight greenhouse screening methods named micro-chamber and mist-chamber were improved for reliable phenotyping of rice mapping populations for the identification of QTLs controlling sheath blight resistance
• Five rice mapping populations with more than 250 advanced lines are available for penotyping and identification/corroboration of new QTLs controlling sheath blight resistance
• A laboratory and greenhouse method including morphological characteristics, pathogenicity tests and use of PCR-specific primers was developed for the identification of the bacterial panicle blight pathogen Burkholderia glumae
• A greenhouse screening method to evaluate and identify potential sources of tolerance to the bacterial panicle blight pathogen was developed and used for screening more than 200 Latin American rice cultivars and advanced breeding lines
23

• Six potential sources exhibiting high tolerance to the bacterial panicle blight pathogen, Burkholderia glumae, were identified in greenhouse inoculations for field testing and use in breeding for resistance
• In collaboration with Harvest Plus, it has been shown that NIRs technology can be successfully used to estimate iron and zinc content in rice grain.
• A marker assisted selection program for resistance to RHBV and T.oryzicolus is being established.
Rice Varietal Improvement: Ten varieties for commercial rice production were released in the region by our partners from germplam developed by the CIAT-FLAR breeding programs.
- “ESPERANZA” Upland rice variety in Bolivia coming from population breeding was released in Bolivia for both manual and mechanized rice ecosystems.
- “RQUILIA 23” First irrigated rice variety coming from population breeding in Chile, adapted to the temperate rice ecosystem.
- Fedearroz 174 and Fedearroz 60 were released by FEDARROZ in Colombia for irrigated and favored upland conditions.
- Cetauro was released for irrigated conditions by FUNDARROZ in Venezuela. - Coprosem 304 and Improarroz 420 were released by COPROSEM and
IMPROARROZ, respectively for irrigated and favored upland conditions in Colombia.
- Palmar 18 and Cabuyo were released by SENUMISA for irrigated/favored upland conditions in Costa Rica.
- CENTA –A8 was released by CENTA , El Salvador for irrigated-favored uplnad conditions.
Special Project
• FAO Regional Tecnical Cooperation Project (TCP/RLA/3102). Capacitación en Fitomejoramiento Genético e Intercambio de Germoplasma para utilizar los Recursos Genéticos del Arroz en América Latina y el Caribe. The Regional TCP Project involves 6 Countries: Argentina, Chile, Cuba, Guatemala, Nicaragua and Dominican Republic and 3 International Institutions: FAO, CIAT and Cirad. Liaison Officer
• Liaison between FAO (regional LAC Office and Rome Headquarters), CIAT and Cirad:
- Funding by FAO of a Regional TCP Project. - Implementation of the proposed activities (Breeders Workshops and Training courses)
24

• Inter-institutional liaison between CIRAD and CIAT:+
- Implementing new research activities linking Population Breeding and Eco-physiology
(3 weeks visit of a Cirad Scientist at CIAT and field experiment at Villavicencio-
Colombia).- Implementation of a rice population breeding project in France (Camargue)
targeting the development of aromatic varieties.
- New Cirad Scientist to be posted at CIAT in mid-2008 to reinforce the actual 10. Summary Budeet prepared by Finances
ACTUAL EXPENDITURES 2007
Outcome Line SBA-4: Rice
Rice Program SOURCE
HQ+LAC Biotech Total US$ (%)
Unrestricted Core 134,845 222,140 356,985 7% Restricted Core C.E 342,502 342,502 7% Sub-total Core 477,347 222,140 699,487 14% Restricted
Special Projects 1,515,514
1,628,741 3,144,255 65%
Generation Challenge Program 323,550 323,550 7%
Sub Total Restricted 1,515,514
1,952,291 3,467,805 72%
Direct Expenditures 1,992,861
2,174,431
4,167,292 86%
Non Research Cost 314,817
343,500
658,317 14%
Total Expenditures 2,307,678
2,517,931
4,825,609 100%
(1) Excluding Non Operational expenses: Phase-out and Fixed Assets adjustment.
25

1
AANNNNUUAALL RREEPPOORRTT 22000077
1. CIAT – SBA- 4 RICE Project Description
Goal To generate food security and employment associated with rice production with emphasis on improving the options for the small farmers.
Objective To produce robust high yielding rice varieties requiring lower inputs, we will provide well-characterized progenitors and advanced materials with an ample genetic base as well as information and training. Research Outputs: Our research is organized around three major outputs. 1) Enhanced gene pools
a. Irrigated Rice b. Upland Rice
2) Integrated crop, pest and disease management 3) Intensification and diversification of rice cropping systems for small farmers.
Output 1: Enhanced gene pools
1A. IRRIGATED RICE
Broadening the genetic base of irrigated rice in Latin America.
1.A.1 Breeding Strategies to increase the content of iron and zinc in the rice grain César P. Martínez, Jaime Borrero, Silvio James Carabali, Yamid Sanabria, Olga X. Giraldo, Myriam C. Duque, James Silva, Jairo Barona and J.Tohme Funding: CIDA-Canada and CIAT_Core Abstract
In the fast-track rice breeding program at CIAT, landraces and breeding lines conserved in germplasm banks are screened for mineral content to identify products that could have immediate utility as potential varieties or donors. Among 3189 rice lines analyzed in 2007, 38 and 32 were found to reliably meet or exceed the iron 6-8 ppm target range and

2
the zinc 22-25 ppm target range for milled rice, respectively. Additionally, 400 rice samples (200 brown rice and 200 milled rice samples) representing 200 genotypes were sent to CIP for establishing a NIRS evaluation method for rice iron and zinc in collaboration with Harvest Plus. Based on this information and further work done by Wolfgang Gruneberg and Thomas Zum Felde at CIP it was possible to determine that NIRS can be successfully used to estimate rice iron and zinc. Nevertheless additional work will be done in 2008 to get familiar with NIRS before using it as a high-throughput technique. With an eye to the medium- and long-term, a crossing program was also started to combine high-iron and -zinc with high yield potential, tolerance to main biotic and abiotic stresses, and good grain quality. During 2007 priority was given to the field evaluation and selection of lines derived from interspecific crosses. From 2789 single plant selections and 5,798 advanced lines, a total of 1821 selections were made for iron and zinc analysis. Most of these lines are combining good agronomic traits. The 83 best lines were included in a nursery that was distributed to AgroSalud partners in Bolivia, Cuba and Venezuela. This nursery is currently being dispatched to partners in Colombia, Nicaragua and the Dominican Republic.
Background
Our aim is to increase the content of iron and zinc in milled rice, using conventional methods of breeding, including methods of bulk and pedigree selection, backcrossing, population breeding and mutagenesis. On a fast track(short term phase), landraces and breeding lines conserved in the germplasm banks are screened for mineral content to identify products that could have immediate utility, as potential varieties or donors. A crossing program (medium- long term phase) was also started to combine high-iron and zinc with high yield potential, tolerance to main biotic and abiotic stresses, and good grain quality. This project is carried out in close partnership with research institutions in Colombia, Bolivia, Cuba, Brazil, Dominican Republic, and Nicaragua. More recently, Panamá and Venezuela also requested to be members of AgroSalud. We also work in the identification of molecular markers associated with high iron and zinc content in the rice grain to be used in a marker assisted breeding program. Breeding materials developed by the AgroSalud Project will be shared with our collaborators for evaluation and selection by collaborators under local conditions best lines will be released as varieties. This report presents the results of several activities carried out in 2007. Materials and Methods
Several breeding strategies are being used for the development and deployment of high iron(6-8 ppm) and zinc(22-25 ppm) rice lines for micro-nutrient deficiency areas in Colombia, Cuba, Bolivia, Brazil, Nicaragua, Panama, and Dominican Republic. Different activities including evaluation of rice germplasm and breeding lines maintained in our germplam banks, assessment of NIR technology to speed up analysis of iron and zinc in the rice grain, development of segregating populations via recurrent selection, correlation of the iron and zinc content in brown and milled rice, use of the statistical design “

3
increased blocks by Federer” in the analysis of iron and zinc content in the rice grain, comparison of iron and zinc data obtained by different labs, and development of a database for iron and zinc data were carried out.
Results and Discussions
1. Test of existing rice germplasm and breeding lines for increased iron and zinc content.
Several improvements were implemented in our clean lab. to reduce contamination at milling and grinding facilities, including setting up a double entry door at the lab. Additionaly, distilled and dionizided water is used for washing glassware, teflon chambers, zirconium balls, and vials used to prepare rice flour. A lab manual was prepared contaning instructions for lab. users. A total of 5090 rice lines were evaluated under field conditions and samples of 563 of them were prepared and sent to CIAT’s Analitycal Service Lab.for iron and zinc analysis via atomic adsortion method.(Table1). As expected most of the lines had low iron and zinc content in milled rice; however, about 38 rice lines were identified presenting between 5 and 8.3 ppm of iron. These are lines derived from crosess between O. sativa and O. glaberrima, and O.barthii. In terms of zinc, some lines were also identified as having between 22 and 25 ppm. Our reference check, Fedearroz 50 had on the average 4.3 ppm of iron and 16.1 ppm of zinc based on 40 independent evaluations. These results indicate that we are on track in terms of iron content, considering that our base line is 2-3 ppm of iron and 17-18 ppm of zinc. Figures 1 and 2 show genetic progress made in terms of iron content over time. A database is being developed to include all agronomic, breeding and iron/zinc data to make it available to the Agrosalud community via internet.
Table1. Breeding material evaluated and selected for iron and zinc analysis. CIAT.2007
Population Generation Lines Evaluated Evaluated
for Total Iron-zinc
Lines derived from IRRI material (HP Project) F6 1015 36 Lines derived from O.latifoliaxO.sativa F6 2529 7 Lines evaluated/selected in Montería F5 44 64 Lines derived from IRRI material/CTs) F4 960 63 Línes S4 (PCT 8, 19,21,22) S4 135 133 Línes S1 Biofortificación S1 160 39 Introductions 247 221 Total 5090 563

4
Development of a iron and zinc Database
A database is being developed to facilitate access of iron and zinc data as well as to main agronomic data of rice materials in a fast and structured form. This database is in Excel and constitutes a very easy, simple and friendly tool that can be handled by users; it is a dynamic database, where the stored information can be modified and updated as new data becomes available. Main objectives are:
-Fast and agile consultations by a series of agronomics characteristics including values of the content of micronutrients iron and zinc.
-It allows operations like updating, addition of data and consultations.
- It provides minimum and maximum values, averages values for iron and zinc of evaluated materials.
- It allows sorting the materials by anyone of the registered characteristics.
-It displays in graphic form the values of iron and zinc of the evaluated materials.
At present, the database has a record of 3552 materials, and is being fine tuned with new applications aimed at offering more information to the users. In the future this database will be in the webpage of the project available of all the cooperators. 2. Assessment of NIR technology to speed up analysis of iron and zinc in the rice grain.
A total of 400 rice samples (200 brown rice and 200 milled rice samples) representing 200 genotypes were sent to CIP ( W.J.Gruneberg) for establishing a NIRS evaluation method for iron and zinc in rice in collaboration with Harvest Plus. These samples were scanned twice by NIRS, the spectra stored and samples sent to Adelaide to get reference values for iron and zinc. Based on this information and furher work done by W.J. Gruneberg and Thomas Zum Felde at CIP it was possible to determine that NIRS can be successfully used to estimate iron and zinc in rice. Two programs (ricemeal and riceseed) were established and validated. For ricemeal it is possible to estimate protein content very well whilst for Fe, Zn and the other minerals the standard error of prediction (SEP) is acceptable, and the estimation can be used for screening. For complete seeds ("riceseed.eqa" contains the equations for proteine, Fe, Zn, Ca, Mg, K, P, S in rice grain) it is also possible to estimate protein very well. For Fe, Zn and the other minerals the prediction is less accurate as indicated by an higher SEP but still acceptable for a high-through-put, nondestructive screening. Protein is estimated in %, the unit for all minerals is mg/kg(ppm)in dry matter, using polished white rice or rice flour. Best results were obtained for iron compared to zinc, as shown in Figures 3, 4 and 5. Unfortunately the data for minerals in unpolished brown rice showed some contamination. The two programs have been installed at CIAT and some additional work will done in 2008 to get familiar with NIRS before using it as a high throughput technique.

5
3. Use of recurrent selection to increase iron and zinc content in the rice grain.
Four populations (PCT-8CG/1/CG/1, PCT-19, PCT-21, and PCT-22) developed by the CIAT Rice project which are carrying a cytoplasmic male-sterility gene were selected to start a population improvement program through recurrent selection for high iron and zinc.This is based on the successful use of recurrent selection by Dudley et al, 1974 to increase protein content in maize from 10.0% to 26,6%. Male -sterile plants were selected in each population and crossed to Azucena, Madhukar, Ketan Lumbu, Gundil Kuning, Perurutong, IR68552-100-1-2-2, and IR71703-657-3-1-2 used as donor parents. This activity is carried out in collaboration with Fedearroz, our partner in Colombia. F1 seed from each cross was planted and evaluated in 2006 and F2 seed was mixed in equal proportion to form new populations( PCTBF1,PCTBF2,PCTBF3, and PCTBF4);these populations went trough the first recombination cycle in 2007 and single plants selections were be made for iron and zinc analysis.
Populations PCTBF1/0/0/1, PCTBF2/0/0/1, PCTBF3/0/0/1, and PCTBF4/0/0/1, were planted at CIAT for their evaluation, 115 plants S0 were selected from PCTBF1, 24 plant S0 from PCTBF2, 64 plants S0 of PCTBF3 and 25 plants S0 of PCTBF4. The S1 seed of the selected S0 plants was evaluated for its content to iron and zinc and materials with higher micronutrient content were identified; seed of these plants will be mixed in homogenous form for a new cycle of recombination. This seed would be planted in isolated form and all the male sterility plants would be harvested, which will be mixed to constitute the new selection cycle (Table 3). Data suggest that, compared to our standard control (Fedearroz 50) gains in zinc content were greater in zinc than in the case of iron. Higher increase is expected as more recombination cycles are allowed.
Table 3. Number of plants S0 evaluated and S1 seed selected for its iron and zinc content in milled rice in populations PCTBF-1 PCTBF-2, PCTBF-3, PCTBF-4. CIAT 2007. Populations Plant S0
EvaluatedSeeds S1selected
Range (mg/kg) Fe Zn
PCTBF1 115 37 2.55 – 4.74 ; 8.89 – 21.94 PCTBF2 24 14 3.10 – 4.59 ; 11.29 – 27.26 PCTBF3 64 33 2.52 – 4.68 ; 8.95 – 22.61 PCTBF4 25 15 2.08 – 4.41 ; 10.62 – 19.72
Fedearroz50 2.82 ; 18.93
New progenitors (see Section 1) with high iron and zinc content will be introduced in these populations with the objective of enriching their variability and increasing the possibility of accumulating new genes associated with high iron and zinc content.(Borrero et al., 1997).

6
4. Influence of milling time on the content of iron and zinc in the grain rice . Ten materials with different grain types (extra long, long and slender, medium bold and short) were selected to determine the influence of milling time (seconds) in the content of iron and zinc in milled rice; this information is important to determine the best milling time for the evaluation of rice samples of breeding lines having different grain types under our laboratory conditions. Three replicates/line/milling time were used Figures 1 and 2 show values for iron and zinc of each material and milling time; mean value for iron and zinc, respectively, for brown rice (0 seconds milling time) was 6.41 and 24,37 mg/Kg.; milling time of ten seconds resulted in 3.96 and 20.33 mg/Kg., while 20 seconds of milling time gave 3.18 and 20.80 mg/Kg; with 30 seconds of milling time values were 3.84 y 19.26mg/Kg. Significant differences in the content of iron and zinc were observed between 0 seconds milling time (brown rice) versus different milling times, but not significant differences were found among milling times (10, 20, and 30 seconds). Data suggest that for iron the longer the milling time the lower is the iron content in milled rice, irrespective of the grain type; however, milling time of 20 seconds is more appropiate. Similar finding was reported by Harvest Plus at IRRI. (Figure1). Data for zinc are presented in Figure 2. There was a tendency for zinc content to decrease as milling time increases, specially for extra-long and long/slender grain types. Medium-bold and short grain types behaved differently and zinc losses were lover. Data suggests that in the case of zinc, losses in zinc content due to milling time depend on grain type. 5. Correlation between iron and zinc content in brown and milled rice in segregating breeding lines. The objective was to determine if values for iron and zinc in brown rice are good indicators of iron and zinc in milled rice. Harvest Plus and our own data indicate that most rice materials have low to very low values of iron and zinc in milled rice.These values are very near to the resolution value of the atomic absorption method we are using and could give false data.One of the problems we are facing is related to the inconsistent values we are getting when we compare values obtained from the same seed source but analyzed at different times. Since brown rice contain more iron and zinc, using brown rice data as indicators for these micronutrients in milled rice could be a good alternative and improve the sensitivity of the technique. A total of 25 breeding lines were evaluated for iron and zinc content in both brown and milled rice. Figure 3 shows the dispersion diagram of the results of the iron analysis in brown rice and milled grain. Not a defined trend it’s observed since (r=0.32) indicates that a weak correlation between the values obtained in these two conditions exists, which suggests that contains of iron in brown rice are not good indicators of the final content after milling. For zinc (Figure 4) in the dispersion diagram a better grouping of the results is observed (r=0.65), suggesting that the zinc content in the brown grain is a good indicator of the content in the milled grain. However, from a practical and cost perspective, it is not a good option to implement this results.

7
6. Analysis of iron and zinc content in the rice grain using the statistical design of “increased blocks by Federer (1961)
We know that significant differences in the results of the laboratory analyzes exist, due to contamination from different sources in the processes of harvesting, drying and sample preparation, to inherent variability of the soil where rice is grown, and to the sensitivity of the atomic absorption method used to estimate the content of iron and zinc in the rice grain. A field trial was set up using 64 materials to evaluate the efficiency of the statistical design of increased blocks (Federer and Raghavarao, 1975) in controlling variability in iron and zinc due to soil heterogeneity. This is very important since large number of lines are planted under field conditions but growing in different plots. Each blok contained 64 entries representing materials of diverse origen including four checks(repeated three times in a given block as a control for iron and zinc).Representatives samples of breeding materials from each block was sent for iron and zinc analysis on different times. Results of the statistical analysis showed that the coefficient of variation of the checks for iron was 27,5%, which indicates that the checks had readings different in every week (series) from evaluation. For zinc the coefficient of variation was smaller than 10,3%, but variability of the checks was also observed in each series of analysis. (Figure 5 and 6). The design did not fulfill the expectations previously defined due to the variability obtained in the estimation of the content of iron and zinc in the checks.
7. Comparison of results for iron and zinc content in rice samples processed and analyzed at EMBRAPA and CIAT Laboratories
One of the concerns of the researchers working in the analysis on micronutrients is the reliability of the data reported by the different laboratories. This raises questions about how comparable are these data and about what kind of corrective measures must be taken to make data comparable. During 2007, a scientist( Priscila Zaczuk) from EMBRAPA-CNPAF/Brasil) visited CIAT with the objective of correlating the data collected in laboratories of CNPAF and CIAT, to evaluate degree of reliability of the data reported by each laboratory and to identify the sources of variation. Both labs. are using similar techniques in the estimation of iron and zinc and we worked with samples processed in CNPAF- Brazil and CIAT; 7 materials were evaluated and the technique used for the analysis of the elements was Atomic absorption Spectrophotometer (AAS). The results in Table 4 indicate that there are great differences between the values found in each sample. The coefficient of variation for iron are between (14, 3 and 22,1%) and for zinc (16-17%.) indicating that there is greater difficulty in the iron determinations perhaps by its lower concentration. The differences found in this trial are attributed to the milling degree of grains in each laboratory, to the percentage of humidity of the samples at the time of the analysis and to the sensitivity of methodology used (AAS).

8
Table 4. Evaluation of 7 varieties combining treatments of processing flour at CNPAF and CIAT.2007.
Figure 1. Content of iron for ten materials in different milling time. CIAT 2007
2
3
4
5
6
7
8
0 10 20 30
Milling time (seg)
Fe (m
g/Kg
)
IR69428-6-1-1-3-3IR75862-206-2-8-3-B-B-BIR75862-221-2-1-2-B-B-BIR68144-2B-2-2-3-1-120CT18247-12-8-2-2-1IR68144-2B-2-2-3-1-166FEDEARROZ 50O. rufipogonCEYSVONIGROVENI
2
3
4
5
6
7
8
0 10 20 30
Milling time (seg)
Fe (m
g/Kg
)
IR69428-6-1-1-3-3IR75862-206-2-8-3-B-B-BIR75862-221-2-1-2-B-B-BIR68144-2B-2-2-3-1-120CT18247-12-8-2-2-1IR68144-2B-2-2-3-1-166FEDEARROZ 50O. rufipogonCEYSVONIGROVENI
2
3
4
5
6
7
8
0 10 20 30
Milling time (seg)
Fe (m
g/Kg
)
IR69428-6-1-1-3-3IR75862-206-2-8-3-B-B-BIR75862-221-2-1-2-B-B-BIR68144-2B-2-2-3-1-120CT18247-12-8-2-2-1IR68144-2B-2-2-3-1-166FEDEARROZ 50O. rufipogonCEYSVONIGROVENI
Variety
Fe Zn Fe Zn Fe Zn Fe Zn
Pepita 4.17 28.0 4.76 16.0 4.86 16.5 5.74 15.7Carajás 3.33 13.0 5.69 20.5 6.34 19.8 3.93 19.5Primavera 4.50 25.3 6.77 24.4 7.73 23.6 3.75 22.3Monarca 4.17 23.0 6.78 27.8 5.67 26.9 3.98 27.2Curinga 4.17 20.3 6.96 24.6 6.77 24.2 4.41 21.9Sertaneja 3.67 23.2 5.21 21.8 4.28 21.7 4.01 21.4Guarani 5.67 25.5 6.48 24.7 4.56 24.4 3.84 23.2C.V. 14.3 16.6 22.1 16.0 15.5 17.1
Prepared and analyzed at
CNPAF
Milled, polished and rice flour done at
CNPAF but analyzed at CIAT
Milled and polished at CNPAF but rice flour and analysis at CIAT
Milled at CNPAF; polishing, rice
flour and analysis at CIAT

9
Figure 2. Content of Zinc for ten materials at different milling time.
Figure 3. Dispersion diagram for iron in brown and milled rice.
2.
2.
3.
3.
3.
6 7 8 9 1 1 1
Fe Brown
Fe M
ille
d (
mg
/Kg
)
15
18
21
24
27
0 10 20 30Milling time (seg)
Zn(m
g/Kg
)
IR69428-6-1-1-3-3IR75862-206-2-8-3-B-B-BIR75862-221-2-1-2-B-B-BIR68144-2B-2-2-3-1-120CT18247-12-8-2-2-1IR68144-2B-2-2-3-1-166FEDEARROZ 50O. rufipogonCEYSVONIGROVENI
15
18
21
24
27
0 10 20 30Milling time (seg)
Zn(m
g/Kg
)
IR69428-6-1-1-3-3IR75862-206-2-8-3-B-B-BIR75862-221-2-1-2-B-B-BIR68144-2B-2-2-3-1-120CT18247-12-8-2-2-1IR68144-2B-2-2-3-1-166FEDEARROZ 50O. rufipogonCEYSVONIGROVENI
15
18
21
24
27
0 10 20 30Milling time (seg)
Zn(m
g/Kg
)
IR69428-6-1-1-3-3IR75862-206-2-8-3-B-B-BIR75862-221-2-1-2-B-B-BIR68144-2B-2-2-3-1-120CT18247-12-8-2-2-1IR68144-2B-2-2-3-1-166FEDEARROZ 50O. rufipogonCEYSVONIGROVENI

10
Figure 4. Dispersion diagram for zinc in brown and white rice.
1
1
1
1
2
1 2 2 2 2
Zn Integral
Zn
Bla
nco
(m
g/K
g)
Lower values
1
2
3
4
5
1 2 3 4Weeks
Fe (
mg
/K
g)
Higher values
1
2
3
4
5
1 2 3 4Weeks
Fe (
mg
/K
g)
Average
1
2
3
4
5
1 2 3 4Weeks
Fe (
mg
/Kg
)
Variance
0.0
0.5
1.0
1.5
1 2 3 4Weeks
Fe (
mg
/Kg
)
CT17334 -2-1-1-2-4-5-M-1 CT17334 -13-7-1-5-3-9-M-1 Fedearroz 50 CT17365 -21-3-6-1-2-M-1
Lower values
1
2
3
4
5
1 2 3 4Weeks
Fe (
mg
/K
g)
Higher values
1
2
3
4
5
1 2 3 4Weeks
Fe (
mg
/K
g)
Average
1
2
3
4
5
1 2 3 4Weeks
Fe (
mg
/Kg
)
Variance
0.0
0.5
1.0
1.5
1 2 3 4Weeks
Fe (
mg
/Kg
)
CT17334 -2-1-1-2-4-5-M-1
Lower values
1
2
3
4
5
1 2 3 4Weeks
Fe (
mg
/K
g)
Higher values
1
2
3
4
5
1 2 3 4Weeks
Fe (
mg
/K
g)
Average
1
2
3
4
5
1 2 3 4Weeks
Fe (
mg
/Kg
)
Variance
0.0
0.5
1.0
1.5
1 2 3 4Weeks
Fe (
mg
/Kg
)
CT17334 -2-1-1-2-4-5-M-1 CT17334 -13-7-1-5-3-9-M-1 Fedearroz 50 CT17365 -21-3-6-1-2-M-1
Figure 5. Variability of the checks in each series of analysis for iron.
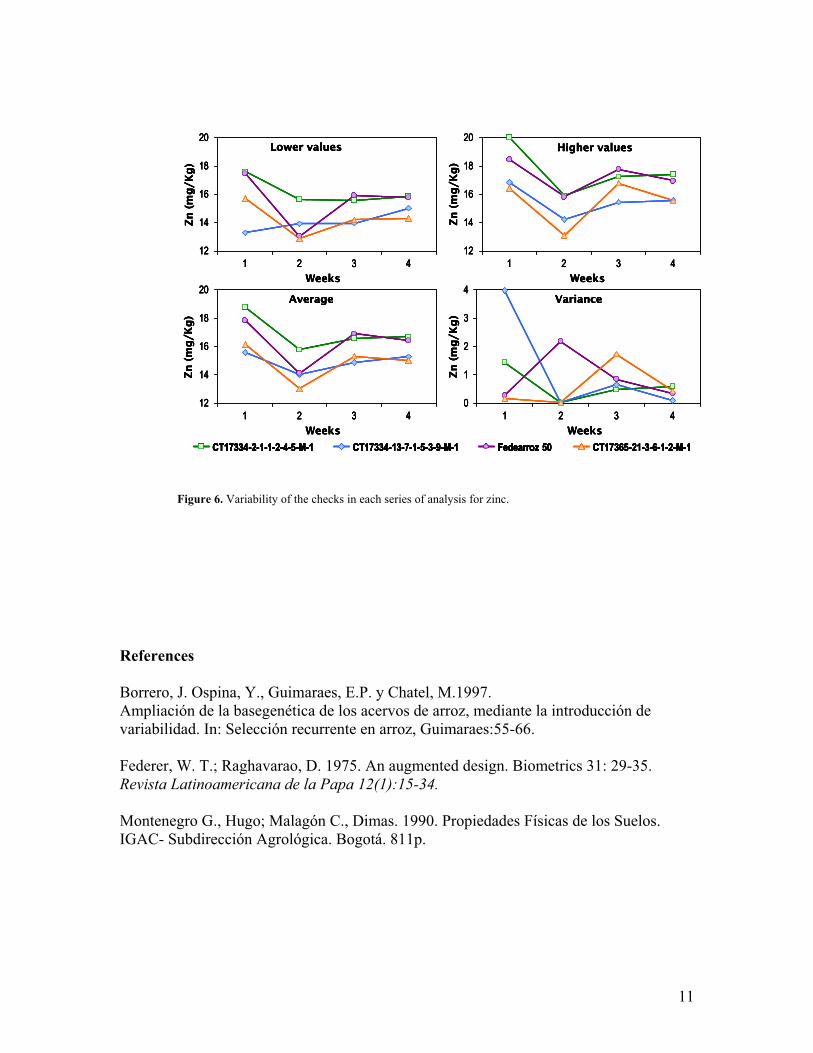
11
References
Borrero, J. Ospina, Y., Guimaraes, E.P. y Chatel, M.1997. Ampliación de la basegenética de los acervos de arroz, mediante la introducción de variabilidad. In: Selección recurrente en arroz, Guimaraes:55-66.
Federer, W. T.; Raghavarao, D. 1975. An augmented design. Biometrics 31: 29-35. Revista Latinoamericana de la Papa 12(1):15-34. Montenegro G., Hugo; Malagón C., Dimas. 1990. Propiedades Físicas de los Suelos. IGAC- Subdirección Agrológica. Bogotá. 811p.
Lower values
12
14
16
18
20
1 2 3 4Weeks
Zn
(mg
/Kg
)
Higher values
12
14
16
18
20
1 2 3 4Weeks
Zn
(mg
/Kg
)
Average
12
14
16
18
20
1 2 3 4Weeks
Zn
(mg
/Kg
)
Variance
0
1
2
3
4
1 2 3 4Weeks
Zn
(mg
/Kg
)CT17334-2-1-1-2-4-5-M-1 CT17334-13-7-1-5-3-9-M-1 Fedearroz 50 CT17365-21-3-6-1-2-M-1CT17334-2-1-1-2-4-5-M-1 CT17334-13-7-1-5-3-9-M-1 Fedearroz 50 CT17365-21-3-6-1-2-M-1
Lower values
12
14
16
18
20
1 2 3 4Weeks
Zn
(mg
/Kg
)
Higher values
12
14
16
18
20
1 2 3 4Weeks
Zn
(mg
/Kg
)
Average
12
14
16
18
20
1 2 3 4Weeks
Zn
(mg
/Kg
)
Variance
0
1
2
3
4
1 2 3 4Weeks
Zn
(mg
/Kg
)CT17334-2-1-1-2-4-5-M-1 CT17334-13-7-1-5-3-9-M-1 Fedearroz 50 CT17365-21-3-6-1-2-M-1CT17334-2-1-1-2-4-5-M-1 CT17334-13-7-1-5-3-9-M-1 Fedearroz 50 CT17365-21-3-6-1-2-M-1
Lower values
12
14
16
18
20
1 2 3 4Weeks
Zn
(mg
/Kg
)
Higher values
12
14
16
18
20
1 2 3 4Weeks
Zn
(mg
/Kg
)
Average
12
14
16
18
20
1 2 3 4Weeks
Zn
(mg
/Kg
)
Variance
0
1
2
3
4
1 2 3 4Weeks
Zn
(mg
/Kg
)CT17334-2-1-1-2-4-5-M-1
Lower values
12
14
16
18
20
1 2 3 4Weeks
Zn
(mg
/Kg
)
Higher values
12
14
16
18
20
1 2 3 4Weeks
Zn
(mg
/Kg
)
Average
12
14
16
18
20
1 2 3 4Weeks
Zn
(mg
/Kg
)
Variance
0
1
2
3
4
1 2 3 4Weeks
Zn
(mg
/Kg
)CT17334-2-1-1-2-4-5-M-1 CT17334-13-7-1-5-3-9-M-1 Fedearroz 50 CT17365-21-3-6-1-2-M-1CT17334-2-1-1-2-4-5-M-1 CT17334-13-7-1-5-3-9-M-1 Fedearroz 50 CT17365-21-3-6-1-2-M-1
Figure 6. Variability of the checks in each series of analysis for zinc.

12
1.A.2. Progress Report on Broadening the Genetic Base of Rice In Latin America Contributors: C. P. Martínez ,S. J. Carabalí and J. Borrero, Anna McClung, Georgia Eizenga, Susan McCouch and Shannon Moon(CIAT, USDA-Dale Bumpers National Rice Researc Center, Cornell University, RiceTec). Funding: CIDA-Canada; CIAT-core;USDA and NSF-USA Abstract.
High yield QTLs identified from O. rufipogon were verified to have a yield enhancing impact when introgressed into Jefferson. Six lines in particular, had improved yield potential and will be further tested in field trials in 2008. Out of 7679 breeding lines in different stages of development evaluated in 2007, a total of 5268 plant selections were made for further testing in 2008. Breeding nurseries were sent to collaborators in 8 countries. Additionally, CIAT contributed sets of advanced lines to INGER-IRRI and WARDA. Key words: wild rice, QTLs, inter-specific crosses
Background
Wild species from genus Oryza are a great alternative as a source of new genes for the improvement of cultivated rice. However, the transfer of useful traits from wild species to those cultivars has been very limited. Some examples like the Grassy Stunt virus resistance gene from O. nivara (Khush et to the one. 1977) and the CytoplasmaticSterility genes from O. perennis to generated hybrid rice (Lin and Yuan 1978) are reported as successful transfer of interest traits. Several wild species are highly resistant to all well-known biotypes of Brown Plant-hopper (BPH), White Backed Plant-hopper (WBPH), and Green Leaf-hopper (Heinrichs et al.1985). Recently Brar et al (2002) reported that genes associated with resistance to bacteria, Pyricularia oryzae, tungro virus and acid conditions tolerance can be transferred from wild species to cultivated rice. Recent data (Xiao et to the, 1996) suggest that some wild species of rice contains genes that are able to increase yield and improve the grain quality of modern rice varieties. The use of wild parentals in wheat, by CIMMYT, has given high levels of resistance and tolerance to Septoria, Fusarium, drought, heat, salinity and flooding. They have also found higher concentration of Iron and Zinc in the wild wheat types than in the average of the cultivated wheat. The increase of diseases and insect’s attacks during the cultivation of rice, like Rhizoctonia solani, Sarocladium oryzae, Bipolaris oryzae, and Polymyxa graminis (fungus that transmits the rice stripe necrosis virus RSNV) that used to be considered as low impact diseases, are now being associated with high yield losses in several areas of Latin America and the Caribbean. This is why it is so important to search for new sources of resistance. For all these reasons, CIAT has proposed a strategy to transfer resistance

13
and tolerance genes from wild species to improved elite germplasm using conventional method and new molecular tools. Through a network of international collaborators(McCouch et al. 2007), including the CIAT Rice Project, research has been conducted to explore genetic gains of introgressing genes from wild rice relatives into cultivated rice.Advanced backcross populations were developed so that transgressive variation could be dissected. Using a common set of molecular markers to identify QTLs associeted with enhanced performance in wild x elite commercial varieties it has been demonstrated by several groups that specific ontrogressions from O.rufipogon confers superior performance for diverse agronomic and yield related traits. Furher evidence will be shown in this report through a collaborative multi-institutional effort. (Mc Clung et al, 2008) Materials and Methods Cultivated rice varieties like; Fedearroz 50, Fedearroz 2000, Perla, Oryzica Caribe 8, Fanny, Epagri 108, Cica 8, Caiapó, Curinga, as weel as elite lines from IRRI, CIAT, and WARDA are being crossed with wild species such as O. rufipogon, O.glaberrima, O. barthii, O. latifolia, O.meridionalis to develop improved and broad-base populations for breeding purposes. This plant material is evaluated for trait of interest in Santa Rosa, CIAT and some Fedearroz´s field stations (Montería). Improved lines are shared with our multiple partners to be used as breeding tools in their breeding programs. In the first semester of 2007 a total of 2429 segreganting lines (F2-F3, F5) were evaluated in Santa Rosa (Table 1) under high disease pressure; of which 2717 plants were selected and harvested in order to be sown at CIAT (Palmira) by the end of 2007. These materials were preliminary evaluated for good agronomic characteristic and from this selection 235 lines were evaluated for the VHB virus.
Table 1. Evaluation and selection of segregating breeding populations at Santa Rosa 2007A.
Population Generation Evaluated lines Selected
lines Inter-specific crosses
F2 665 1333 Combination of genes for rice blast
F2 187 312 Three way crosses
F2 66 89 Combination of Pi1,Pi2,Pi33,Pi9 genes
F2 106 230 Nericax commercial varieties crosses
F2 80 57 Aromatic/ commercial varieties
F2 52 63 Resistance to Tagosodes oryzicola
F3 230 213 93-11 / commercial varieties F3 528 259 Nerica / commercial varieties
F3 154 15 Introgresiones(Moroberekan)
F5 361 142
Total 2429 2713

14
In the second semester of 2007, 5250 lines in different generation and derived from differents interspecific crosses and lines from IRRI and CIAT-FLAR germplasm banks were sown. Plants with good characteristic agronomic were selected. This material was later will sown at Santa Rosa for Pyricularia stress in the first semester of the 2008 (Table 2). In this same semester populations from interspecific crosses such as BC2F4 and BC2F5 from Fedearroz 50 x O. rufipogon, inbreed F6 lines derived from the cross with O.latifolia (Salahondita) and other populations (see table 2) were evaluated to determine iron and zinc content (results not shown in this report), Later in this semesters this lines will be evaluated for diseases resistance.
Table 2 Evaluation and selection of segregating populations at CIAT Palmira 2007B.
Population GenerationEvaluated
linesSelected
lines
Biofortification(germplasm. IRRI/CTs) F4 970 749 Germplasm IRRI (Biofortificatión) F6 1015 180 BC2 rufipogon / Fedearroz 50 F5 266 152 Salahondita (O.latifolia) F6 2529 1231 Línes S4 (PCT 8, 19,21,22) S4 135 133 Línes S1 Biofortificación S1 160 39 BC2 O. rufipogon / Fedearroz 50 (Montería) F4 49 33 BIOFORTIFICATION (Montería) F5 44 69 Recurrent Selection. (Biof.) S0 4 2 Germplasm Nicaragua 78 Total 5250 2555
Yield Enhancement via Chromosomal Introgressions from O.rufipogon: A set of 70 NILs that had been developed by McCouch’s group at Cornell University from a Jefferson 2/O.rufipogon cross originally developed at CIAT and identified to posses QTLs associated with improved yield were evaluated in replicated field trials in 2007 at two locations in TX and two locations in AR. NILs were evaluated in standard yield plots replicated three times; agronomic performance of NILs was comapred to Jeffereson(Jeff) and three new cultivars Cocodrie(CCDR), Trenasse(TRNS) and XL723(hybrid from RiceTec). The study consisted of six families of introgression lines, each possessing a yield QTL located on either chromosome 1, 2, 3, 6, 8, or 9. Each family was represented by 4 to 12 independent introgressions of the targeted QTLs and several control NILs, having a common background but lacking the introgression. The NILs were evaluated in standard yield plots (approx. 6 rows x 20’), replicated three

15
times, at each of four locations. The agronomic performance of the NILs was compared with the Jefferson (JEFF) parent, Cocodrie (CCDR), Trenasse (TRNS), and XL723 (hybrid). The objective of the study was to determine if purified near inbred recombinant lines (NILs) possessing individual O. rufipogon QTL that have previously been identified as improving productivity, impacted main crop yield when back crossed into Jefferson (i.e. Jeff//Jeff/O. rufi), (McClung et al.2008).
Results:
Pyricularia susceptibility. Figure 1 shows responses from different populations to the Pyricularia fungus at Santa Rosa experimental station; 23.6% of the materials were resistant, 39.6% intermediates and the 36.8% susceptible. The most resistance population was the F5 with Moroberekan introgressions with 49.44% of resistance. The F2 interspecific cross population, showed the less number of resistant lines; with a 39.25% of lines with intermediate response. VHB susceptibility. Figure shows 2 the susceptibility of different populations to VHB; 15.8% of the lines were resistant, 18.% had an intermediate susceptibility and 66.were susceptible. The population with the higher number of resistant lines was the F4 of 93 11/improved varieties with the 46% of resistant lines, while F3 lines from interspecific cross showed the less number of resistant lines with the 6.% .
Figure 1. Susceptibility of segregating populations to rice blast at Santa Rosa 2007.
0%10%20%30%40%50%60%70%80%90%
100%
Ceysv
oni
F2 piri(gen
esPi1,
Pi2,Pi33
, Pi9)
F2 Basmati
/Commerc
ial va
rietie
s
F2 Triple
Cross
es(S
t. Rosa
)
F2 Resist
ance
piri
F2 WAB/Commerc
ialva
rietie
s
F3 93-11
/ Commerc
ialva
rietie
s
F3 Dam
age Mec
hanic
F3 WAB C
ommercial
varie
ties
F5 Introgres
sions (
Morobere
kan)
Fedea
rroz 50
Oryzica
1
SRI

16
Figure 2. Susceptibility of breeding materials to VHB at CIAT Palmira 2007.
0%
10%
20%
30%
40%
50%
60%
70%
80%
90%
100%
Bbt 50
Colombia 1
F3 Res
istan
ce piri
F4 Dam
age M
echan
ic
Fedea
rroz 2
000
Fedea
rroz 5
0
F6 Intro
gressio
n (Moro
verek
an)
F4 93
-11/C
ommercial
varie
ties
SRI
Distribution of germplasm. Distributing germplasm to Latin America’s partners is a key activity of Ciat Rice Program. In 2007 different nurseries were distributed to collaborators and partners at local and regional level in 8 countries. (Table 3) Table 3 Type of breeding nurseries, countries and collaborators receiving CIAT-ION 2007
Order Institution Country Type of nursery #Line Collaborators
1 IVIC Venezuela CIAT-ION Biofortification 83 Thaura Glenn
2 CIAT Bolivia CIAT-ION Biofortification 23 Roger Taboada
3 IAC Cuba CIAT-ION Biofortification 23 Violeta Puldor
4 BAHA Belize CIAT-ION 200 Robert Shank
5 FLAR Colombia CIAT-ION 90 Gonzalo Zorrilla
6 IDIAF Rep. Dominicana
CIAT-ION Biofortification 47
Angel Adames F.
7 COOPROSEM Colombia CIAT-ION 384 Carmen Lasserna

17
8 FEDEARROZ Colombia Biofortification 83 Alejandro Vargas
9 INIAP Perú CIAT-ION 490 Orlando Palacios
10Molino Villavicencio Colombia CIAT-ION 459 Alberto Davalos
11 INTA Nicaragua CIAT-ION Biofortification 83 Lazaro Narvaez
In addition through the IRRI-CIAT-WARDA agreement nurseries of advanced breeding lines were sent to the IRRI- INGER Network and for evaluation in West Africa . Yield Enhancement through chromosomal introgressions from O.rufipogon
Main data (McClung et al.2008) are presented in Figures 3,4 and 5, and Table 4. Individual families possessing QTL introgressions from chromosomes 2 and 6 demonstrated significant improvement in yield potential as compared to the Jefferson parent whereas the impact of QTLs from chromosomes 3 and 8 were more modest. QTLs from chromosomes 2 and 6 sufficiently increased the yield potential of Jefferson to be competitive with more recently released cultivars. Yield increases are sometimes a result of increased biomass associated with taller plants and a longer growth cycle. However, these QTLs improved yield while maintaining the early maturity and short stature of Jefferson.(Figure 3) The highest yielding NILs possessing the introgression from chromosome 2 (43-2-1, 43-2-12, and 43-1-2) had significantly better yield than Jefferson and their controls (C-). Because these also have background introgressions from chromosome 12, futher purification is required to isolate the contributing QTL.(Figure 4) Two of the NILs (219-2-9 and 219-1-5) possessing QTls from chromosome 6 had significantly higher yield than Jefferson, their controls (C-), and check cultivars Trenasse and Cocodrie. However, further recombinants will need to be developed to fine map the specific region resulting in enhanced yield.(Figure 5). Among the 70 NILs and QTL controls evaluated, grain chalk was determined to be the variable with the largest impact on yield. This suggests that a change in the source/sink distribution of carbohydrate may be influenced by QTLs on these six chromosomes.(Table4) Finally, breeding activities done in close collaboration with our partners in Nicaragua(Ciat-Cirad-Inta) and in Bolivia(CIAT-Santa Cruz and ASPAR) led to the identification of breeding lines derived from inter-specific crosses between O.sativa/O.rufipogon and O.sativa/O.glaberrima having good performance under irrigated and upland conditions in farmer’s fields in 2006-2007. These lines are being recommended for varietal release in 2008 due to its superior performance compared to commercial varieties.
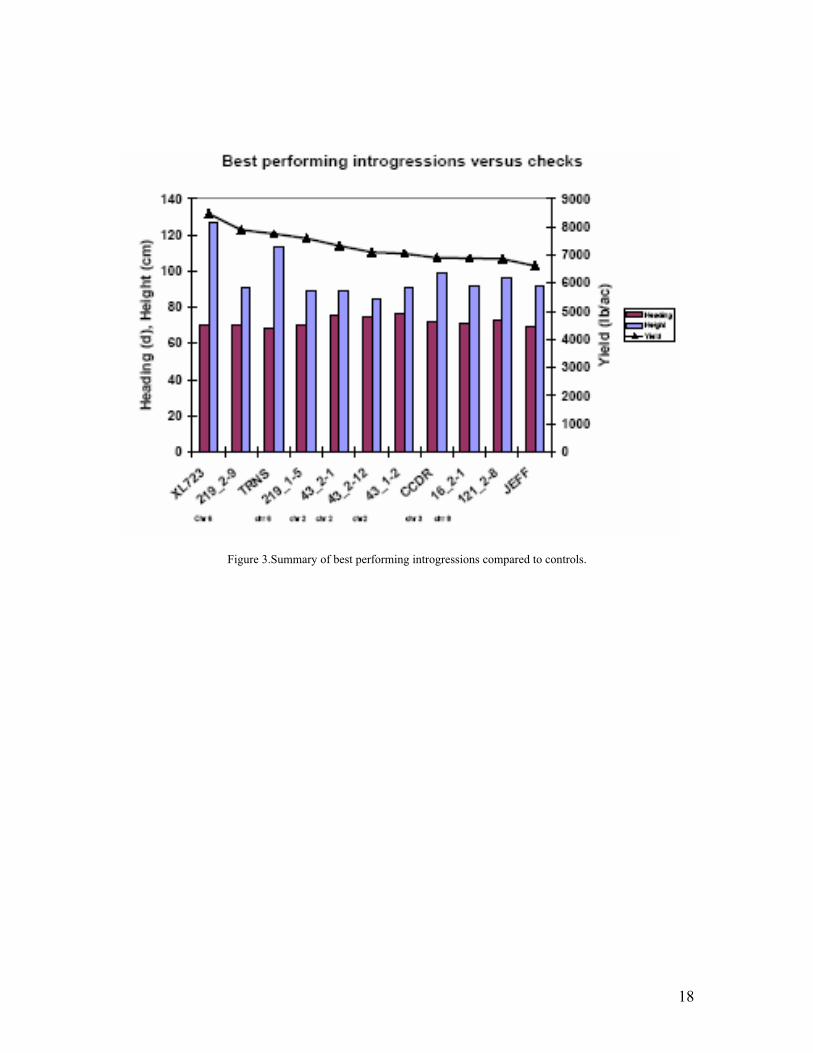
18
Figure 3.Summary of best performing introgressions compared to controls.

19
Figure 4. Yield of NIL 2.1 compared to controls.
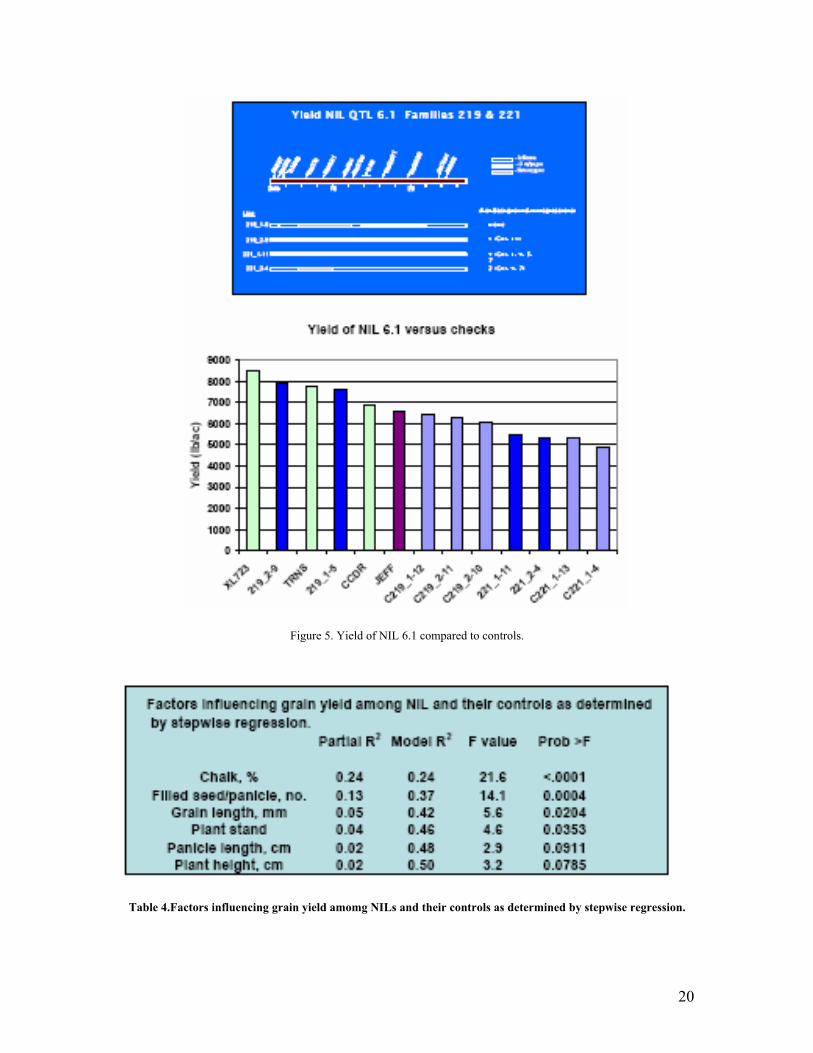
20
Figure 5. Yield of NIL 6.1 compared to controls.
Table 4.Factors influencing grain yield amomg NILs and their controls as determined by stepwise regression.

21
Future plans -Continue the development and characterization of populations derived from inter-
specific crosses.
-Evaluation of selected breeding lines for agronomic, disease and insect resistance and
grain quality.
-Evaluation of the segregating breeding populations in different environments in
collaboration with Fedearroz-Colombia
- Distribution of breeding nurseries to our collaborators
References.
Brar D.S., Bui Chi Bun, B. Nguyen, Z. Li, M. Jones and G.S. Khush. 2002. . Gene transfer from wild species and molecular characterization of alien introgression in rice. Abstracts International Rice Congress. 16-20 September, 2002. Beijing, China. p.69 Heinrichs, E. A., F. G. Medrano and H. R. Rapusas, 1985. Genetic evaluation for insect resistance in rice. International Rice Research Institute, P. 0. Box 933, Manila, Philippines. p. 356. Khush GS, Ling KC, Aquino RC, Aguiero VM (1977). Breeding for resistance to grassy stunt in rice. In: 3rd Int. Cong. SABRAO, Canberra, Australia. Plant Breeding Papers. 1-4: 3-9. Xiao, J. et al. 1998. Identification of trait-improving quantitative trait loci alleles from a wild rice relative, Oryza rufipogon. Genetics 150:899-909. McCouch, S.R. et al. 2007. Through the bottleneck : O.rufipogon as a source of trait-enhancing alleles for O.sativa. Euphytica 154:317-339 McClung , Anna, Shanon Moon, Georgia Eizenga, and Susan Mc Couch. 2008. Enhancement of yield through chromosomal introgressions from Oryza rufipogon. Proc. 32nd Rice Technical Working Group Meeting , San Diego, Ca . February 18-21 2008. 1.A.3. Yield evaluation in large panicle type lines. Collaborators: Torres, E.A. and Berrio L.E.
Source of funding: CIAT and FLAR Abstract: Twenty-three F5 Flar high yielding elite lines representing low, intermediate and high

22
combinations of tillering ability and number of grains per panicle and three checks were evaluated for yield and yield components in CIAT during 2007. In general, yields were good and high yielding lines were observed. Two lines with the lowest number of grains per panicles and the highest number of effective panicles by square meter surpassed the best check FL01028. Other two lines, with a low number of panicles per square meter and a high number of grains per panicles, were statistically similar to the two firsts and superior to Oryzica 1. Grain yield was related positively with the number of filled grains per square meter and 1000-grain weight; negatively related with percent of sterility; and non-related with the number of effective culms per square meter or the number of grains per panicle. The number of grains per panicle was related negatively with the number of effective panicles per square meter and the 1000-grain weight and positively with sterility, indicating compensation between yield components. It was concluded that large panicles alone do not automatically give high yield; to achieve the yield potential offered by large panicles filled spikelet percentage and lodging resistance require improvement. Key words: Yield potential, large panicles, yield components Background: In recent years, there have been several attempts to increase yield potential in rice by modifying the yield components. According to Yoshida (1981) rice yields are determined by: the grain number/unit of area, the 1000-grain weight and the percentage of filled spikelets. In this equation, the grains number can be separated into the number of effective panicles per unit of area and the number of spikelets per panicle. Because modern semi-dwarf varieties produce excessive unproductive tillers and leaf area which cause mutual shading and reduced canopy photosynthesis and sink size, the International Rice Research Institute (IRRI) proposed the concept of a new plant type. This new ideotype is characterized by low tillering capacity (9-10 tillers), no unproductive tillers, 200 – 250 grains per panicle, dark green leaves, and a vigorous and deep root system (Kush, 2005). However, after intensive evaluation of these materials, the yield results were disappointing because of low biomass production and poor grain filling (Peng, et al 1999). Japanese researchers also attempted to increase yield potential by modifying plant type, modeled after the Korean Tongil type varieties, characterized by short culms, relatively fewer panicles with a very large number of grains per panicle. This plant type deviates greatly from the traditional framework: short culms and many panicle type versus long culms and large panicle types (Yonezawa, 1997). Hybrid rice was also developed to break the yield potential barrier. Hybrid rice technology was developed by in China, where it has spread to about 50% of 30 million of ha, contributing 60% of national paddy production. Hybrid cultivars have a yield advantage of 10 – 20% over the best inbred varieties (Cheng, et al 2007). One recent study found that the yield advantage in hybrids was associated with large sink size due to large panicles and the capacity to maintain a balance between panicles number per unit of area and spikelet number per panicles (Yang, 2007).

23
The Flar strategy to increase rice productivity has two sequential approaches. The first is to close the gap existing between potential and actual yield in existing varieties by better agronomic practices. These practices reduce six major constraints: inappropriate seeding dates, heavy seeding densities, deficient insect control, deficient weed control, poor fertilization practices, and late establishment of permanent irrigation. The second approach is to develop more productive varieties by increasing sink size through large panicles while maintaining high tillering capacity, associated with dark- green, slow senescing leaves (Jennings PR, 2007). These characters are the result of many years of breeding and the first genotypes were developed in 2006. Plant type and high tillering capacity came from the introduction of Asian semi-dwarfs. Dark green slow senescening leaves is a japonica character derived from tropical japonicas as sources of ¨Hoja Blanca¨ virus resistance. Large panicles came from intercrossing indica by japonicas from Africa. One line was discovered in segregating populations having a very high number of grains per panicle (large panicles=PL). This line was used extensively as the PL donor in many three-way crosses. Several advanced lines with huge panicles combined with grain quality and disease resistance were developed. The objective of this work was to compare the yield of new lines having large panicles with normal varieties, and to identify the importance of yield components on the yield in those lines. Material and Methods: Twenty-five rice (Oryza sativa L.) genotypes were used in this study. Of these twenty-three were F5 Flar high yielding elite lines representing low, intermediate and high combinations of tillering ability and number of grains per panicle. The three checks were Oryzica 1, released in Colombia in 1982; high yielding Fedearroz 50, released in 1997 and FL01028 the large panicle donor in the FLAR breeding program. The experiment was planted May 10, 2007 on the experimental farm of the International Center for Tropical Agriculture CIAT in Palmira, Colombia (3°30´N, 75°22´21´´W, 965 m altitude). The seeding rate was 100 kg. ha-1, using pre-germinated seeds in puddled soil. Weeds were controlled with 3,36 kg ia ha-1 propanil, 3kg ia ha-1 butaclor and 0.19 kg ia ha-1 bentazon 10 days after seeding (dds). Chemical fertilizers applied were: 5 gr. m-2 N, 6 gr. m-2 P2O5, 6 gr. m-2 K2O and 1.5 gr. m-2 Zn, as basal after weed control at 12 dds; 8 gr. m-2 N. and 4 gr. m-2 K2O in dry soil (30 dds); 8 gr. m-2 N and 3 gr. m-2 K2O at panicle initiation (58 dds) and 3 gr. m-2 N at flowering. The total N application was 24 gr. m-2. Permanent flooding was applied immediately after fertilizer application at 30 dds and maintained until near harvest. Rice genotypes were arranged in a completely randomized block design with three replications. The size of the plot was 10 m2 (5x2). At maturity two sub-plots each 0.25 m2 within each plot were sampled by cutting plants at the soil level. The two sub-samples were mixed to make a composite sample representing a 0.5 m2 area. Plants were threshed by hand and separated into grain and

24
straw. In each straw sample the total and effective (bearing a panicle) culms were counted. The grain sample was oven dried at 70 °C for three days and filled and empty spikelets were separated and counted. Finally, filled grains per square meter, total grain per square meter, spikelets per panicle, 1000-grains weight, sterility percentage, and the ratio of effective panicles to total tillers were calculated. Grain yield was determined for 6.75 m2 area in each replication and adjusted to a moisture content of 14 percent. Genotypic differences for all characters and relationships between yield and yield components were determined by ANOVA and correlation analysis. Results and discussion: Climatic conditions were not optimum for high rice yields (Fig 1). According to Yoshida (1981), temperatures below 20 °C at about the reduction stage of the pollen mother cells usually induce high percentage spikelet sterility. Also, solar radiation over 400 cal/cm2/day during the reproductive and ripening stages is necessary to achieve high yields. In this experiment, from 55 days after seeding to maturity, there were some periods with low minimum temperature and low solar radiation. They contributed sterility in some genotypes. The variance analysis showed that genotypes used in the study were different for all characters measured excepting the ratio of effective to total culms (Table 1). In general, yields were good and high yielding lines were observed. Two lines surpassed the best check FL01028: FL06890-2P-9-2P-3P-M and FL06885-4P-3-2P-2P-M. Interestedly these lines had the lowest number of grains per panicles and the highest number of effective panicles by square meter. Four lines were superior to Oryzica 1: FL06890-2P-9-2P-3P-M, FL06885-4P-3-2P-2P-M, FL06733-16P-1-3P-2P-M and FL06562-3P-5P-6P-1P-M; the last two had a low number of panicles per square meter and a high number of grains per panicles. Among these four there was no difference in yield, indicating that a high number of grains per square meter giving a high grain yield can be achieved by increasing the number of grains per panicle and maintaining an adequate number of effective panicles. Two lines had an excellent number of grains per panicle indicating genetics gains for this character in the FLAR breeding program: FL06733-2P-1-2P-1P-M and FL06605-7P-4P-1P-1P-M. Their counts of 176.5 and 163.7 grains per panicle were superior to FL01028 (130.38). However, because of high sterility and light grains they had moderately low yield. Fedearroz 50 had low yield because of high sterility and low 1000-grains weight, indicating adverse low minimum temperatures and low light conditions during the reproductive stage. Oryzica 1, also was partially sterile. Consequently, the sterility observed in the large panicles lines could be due to bad climatic conditions. Phenotypic correlations analysis indicated that grain yield was related positively with the number of filled grains per square meter and 1000-grain weight, but was negatively related with percent of sterility. Grain yield was not related with the number of effective
25
culms per square meter or number of grains per panicle. Some well-known relationships between yield components were found. For example, the number of grains per panicle was related negatively with the number of effective panicles per square meter and the 1000-grain weight and positively with sterility, indicating compensation between yield components. These results indicate that FLAR large panicle lines are similar in sink capacity to hybrids and that breeding for large panicles had been effective. Katsura et al (2007) comparing a ¨super hybrid¨ with two normal varieties in transplanted rice, found in the 2004 dry season that hybrid had a yield of 11.8 tn ha-1, 217 panicles/m2, 50.1x103 grains/m2, 231 spikelets per panicle, 33% of sterility and 24.0 gr. for 1000-grains weight. Yang et al (2007) comparing indica inbreds, second generation NTP lines and hybrids found in the 2003 dry season in transplanted rice that means across hybrids were: yield 10,1 tn ha-1, 405 panicles/m2, 48.9x103 grains/m2, 121.4 spikelets per panicle, 22,7% of sterility and 24.7 gr. for 1000-grains weight. However, due to sterility, a large sink size was not related with yield. This sterility probably was caused by low temperature and low light, so that is possible to achieve higher yields with large panicles under better climatic conditions. But, sterility could be related to large panicles due to source limitations. Conclusions: In summary, large panicles alone do not automatically give high yield. To achieve the yield potential offered by large panicles filled spikelet percentage and lodging resistance require improvement.
Future activities: The same trial is actually being conducted in Costa Rica during dry season. Also, eight selected lines and four checks are under evaluation on four locations
Table 1. Square means and significance for: Grain yield (YLD), effective culms (EFC), total grains per panicle (TGP), one thousand grains weight (W1000), sterility percentage (SP), total grains number per square meter (TG) and ratio of effective to total culms (ETT). Sources df YLD EFC TGP W1000 SP TG ETT Blocks 2 1517137,38ns 3370,29ns 409,83ns 2,11ns 0,006ns 36676776ns 0,0004ns
Genotypes 24 3485548,55** 10781,44** 1385,83** 7,41** 0,003** 160564283** 0,0021ns
Error 48 743814,4 2680,07 304,54 0,74 0,008 5359773 0,0014 Total 74 Mean 8016,87 511,87 128,80 23,89 33,21 64725,34 0,8922 CV (%) 10,76 10,11 13,55 3,59 15,07 11,31 4,24 **, *, ns: Significant at 1%, 5% and non-significant, respectively, by F-test.

26
Table 2. Means values for: Grain yield (YLD), effective culms (EFC), total grains per panicle (TGP), one thousand grains weight (W1000), sterility percentage (SP), filled grains per square meter (FG) and total grains number per square meter (TG). Genotype YLD EFC TGP W1000 SP FG x103 TG x103
FL06890-2P-9-2P-3P-M 10199.10 655.33 95.35 26.46 22.67 49.7 63.9
FL06885-4P-3-2P-2P-M 10003.60 611.30 84.32 26.21 27.67 37.1 51.2
FL06733-16P-1-3P-2P-M 9716.80 449.33 149.01 24.62 20.00 53.2 66.5
FL06562-3P-5P-6P-1P-M 9326.70 505.33 130.31 23.53 21.67 51.4 65.9
FL06779-1P-1-3P-2P-M 8966.90 471.33 136.60 25.42 29.33 45.4 64.4
FL06613-20P-2AI-2P-2P-M 8828.90 519.33 156.18 21.62 38.00 49.7 81.1
FL06609-1P-8P-3P-3P-M 8784.50 532.67 135.54 21.67 26.33 52.1 70.3
FL01028 8527.00 510.67 130.38 23.67 28.67 46.9 65.7
FL06706-1P-7-1P-5P-M 8520.20 475.33 127.24 25.93 28.33 43.1 60.4
FL06629-7P-7-6P-3P-M 8323.70 443.33 140.40 24.21 24.67 46.5 62.1
FL06581-1P-6-1P-3P-M 7859.30 500.00 120.68 23.66 28.00 42.7 60.6
FL06733-12P-19-2P-3P-M 7849.30 551.33 121.91 24.41 35.67 42.8 66.7
FL06580-3P-6-6P-3P-M 7838.50 582.00 109.32 25.65 38.67 39.1 63.5
ORYZICA 1 7686.50 459.33 119.24 23.89 24.00 41.4 54.3
FL06605-7P-4P-1P-1P-M 7668.70 490.00 163.71 22.59 40.33 47.4 80.0
FL06742-8P-2-1P-2P-M 7534.80 564.67 105.15 26.20 42.00 34.5 59.3
FL06609-11P-12-3P-3P-M 7283.40 534.00 108.04 22.46 26.00 42.3 57.6
FL06733-2P-1-2P-1P-M 7247.40 408.67 176.56 23.57 45.67 38.7 71.3
FL06612-6P-11-3P-3P-M 7210.60 439.33 151.18 22.91 32.67 43.3 65.5
FL06883-9P-6-5P-5P-M 7185.50 471.33 137.63 23.24 40.67 37.7 64.4
FL06609-21P-8-5P-1P-M 7141.50 482.67 127.20 23.37 38.67 37.3 61.2
FL06741-6P-4-1P-2P-M 7112.20 506.00 114.24 23.68 33.67 37.2 57.0
FL06562-6P-21P-8P-2P-M 7109.60 472.00 133.49 25.85 47.33 32.5 62.8
FEDEARROZ 50 6504.10 603.33 105.44 22.92 39.67 38.1 63.6 FL06608-8P-4-2P-1P-M 5993.00 558.00 140.97 20.27 50.00 39.1 78.4
LSD 0.05 1415.9 84.99 28.65 2.01 13.9 8.3 12.0
Table 3. Phenotypic correlations between grain yield (YLD), effective panicles (EFC), total grains per panicle (TGP), 1000- grains weight (W1000), sterility percentage (SP) and filled grains per square meter (FG). YLD EFC TGP W1000 SP FGYLD 1,00 0,15ns -0,07ns 0,40** -0,56** 0,59** EFC 0,15ns 1,00 -0,68** 0,11ns 0,02ns 0,03ns
TGP -0,07ns -0,68** 1,00 -0,42** 0,32** 0,24* W1000 0,40** 0,11ns -0,42** 1,00 -0,26* -0,12 SP -0,56** 0,02ns 0,32** -0,26* 1,00 -0,63** FG 0,59** 0,03ns 0,24* -0,12 -0,63** 1,00 **, *, ns: Significant at 1%, 5% and non-significant

27
Figure 1. Changes of daily temperatures, solar radiation and solar brightness; each data point represents the average value of 10 days.
References
Cheng, S.H.; Zhuan, JY.; Fang, Y.; Du, JH.; and Cao, LY. 2007. Progress in research and development on hybrid rice: A super domesticate in China. Annals of Botany. 100: 959 – 966. Jennings, PR. 2007. Rice revolutions in Latin-America. Rice Today, April – June 2007, 38. Katsura, K.; Maeda, S.; Horie, T. and Shiraiwa, T. 2007. Analysis of yield attributes and crop physiological traits of Liangyoupeijiu, a hybrid recently bred in China. Field Crops Research 103: 170 -177. Kush, G.S. 2005. What it will take to feed 5,0 billion rice consumers in 2030. Plant Molecular Biology 59: 1 – 6. Peng, S.; Cassman, K.G.; Virmani, S.S.; Sheehy, J. and Kush G.S. 1999. Yields potential trends of tropical rice since the release of IR8 and the challenge of increasing yield potential. Crop Science 39: 1552 – 1559. Yang, W.; Peng, S.; Laza, R.; Visperas, R. and Dionisio-Sese M. 2007. Grain yield and yield attributes of new plant type and hybrid rice. Crops Science 47: 1393 – 1400. Yonezawa, K. 1997. Yield components. In: Matsuo, T.; Futsuhara, Y.; Kikuchi, F. and Yamaguchi, H. (Eds). 1997. The science of rice plant. Volume Three: Genetics. Food and Agriculture Policy Research Center, Tokyo, Japan. p. 1003.
1.A.4. Partial diallel analysis for dark green leaves and its relationship with yield.Collaborators: Torres, E.A.
Source of Funding: CIAT and FLAR Abstract: To study the relationship between dark green foliage and yield and to analyze the genetic architecture of dark green and slow senescence, a diallel analysis was conducted in CIAT during 2007. There were four parents by group and inter-group crosses were made

28
according to partial diallel design. Dark green parents had higher yield, heavier 1000-grain weight, less sterility, and darker green leaves than light green parents. Light green parents had more grains per panicles. The relative rate of senescence was similar in both groups; but as dark green parents and hybrids were greener at flowering, their leaf color at maturity was darker. Flag leaf and second leaf greenness at flowering and maturity were positively related with plant productivity, indicating that dark green leaves contribute to more productive plants and heavier grain. The relative rate of senescence of both leaves was negatively correlated with plant production, i.e., faster senescence, less plant production. Grain number per panicle was positively related with the percentage of sterility and negatively with 1000-grain weight. Heterosis effects were positive for plant production, 1000-grain weight, flag leaf color at flowering and second leaf color at maturity, showing directional dominance increasing these traits. General combining ability analysis showed that with the exception of percentage of sterility, the additive effects of the genes was more important than non additive effects for all traits. Light green parents were a more diverse group than dark green parents. It was concluded that dark green leaf color could be an important secondary trait in selection for yield.
Key words: Rice, yield, slow senescence, partial diallel, combining ability Background: Source capacity, the basis for plant productivity, depends on light reception determined by plant type and photosynthesis efficiency (photosynthetic capacity per unit leaf area, photosynthetic rate and duration, translocation rate, etc). (Yonezawa, 1997). According to Yoshida (1981) erect leaves allow deeper penetration and even distribution of light resulting in increased crop photosynthesis. Erect leaf angle is a desirable trait for high-yielding varieties because light is used more efficiently at high leaf area index. Semi-dwarf plant type was one of the most important characters in the green revolution varieties. Various physiological selection criteria have been tried in plant breeding including single-leaf net photosynthetic rate (Pn) and other traits. However, breeding for increased yield potential by improving photosynthesis efficiency has had little success (Peng, 2000). Maintaining photosynthesis during the grain filling period could increase yield capacity, because photosynthesis during ripening contributes to grain carbohydrate by 60 -100% (Yoshida, 1981). It could be achieved by delaying the senescence process for example, Kropff, et al (2000) using the ORYZA model found that an increase from 9.3 to 11.7 tn ha-1 is possible by an increase in grain filling duration from 24 to 31 days. This authors concluded that an increase to 13 – 15 tn ha-1 in the tropics without an increase in the in the maximum rate of net carbon assimilation, will require considerable delay in leaf senescence and maintenance of leaf N status during an extended grain filling period. Delayed senescence can be viewed as a consequence of the balance between N demand by the grain and N supply during grain filling. During the grain filling period, there are two sources of N for grain growth: concurrently absorbed N from the soil and

29
remobilized N from vegetative tissues (Borrel and Hammer, 2000). During senescence, proteins are degraded and amino acids are transported out of the leaf. Its yellowing indicates the loss of chlorophyll. However, delayed remobilization of N maintains photosynthetic capacity. The leaves serve as the source of nitrogen only at the cost of their source capacity for carbon (Jiang, 1999). Delayed senescence per se does not contribute to yield potential. Four classes of delayed senescence have been identified and only two are functional. These are related to the onset of senescence and the regulation of its rate of progress (Borrel, et al 2000). A healthy root system is necessary to maintain nitrogen absorption during grain filling. And, finally, loss of proteins during senescence is strongly influenced by light environments of leaves (Jiang, et al 1999). Dark green leaves with delayed senescence could be a secondary trait to use in selection for high yield capacity. Fedearroz 50 a commercial variety released in 1998 has dark green slow senescing leaves and a very high yielding capacity under favorable light conditions. One preliminary study showed that Fedearroz 50 flag leaf had a higher chlorophyll concentration, higher Pn and higher N status than Cica 4 (Jennings et al, 2002). The objectives of this research were to study the relationship between dark green color and delayed senescence with grain production and to analyze the genetic architecture of dark green color and delayed senescence. Materials and methods: The materials used in this study were four dark green slow senescening elite FLAR lines FL06784-5P-5-4P-M, FL06892-10P-5-2P-M, FL06899-1P-6-1P-M and FL06562-3P-5-5P-M (the DG group) and four light green early senescening parents Centauro, FL05372-7P-1-2P-1P-M, FL02768-18P-2-2P-1P-M-1P-F12 and CT16658-4-1-1SR-3-2-1-1-4-M the LG group; along with the 16 inter-group F1 hybrids. Inter-group crosses were made according to the partial diallel design (Miranda Filho and Geraldi, 1984). Parents and hybrids were planted in a completely randomized design with two replicates in the CIAT experimental field in May 2007. Entries were grown in three-row plots with ten plants per transplanted row. Plants and rows were separated by 30 cm. Chemical fertilization (25 gr./m2 N, 6 gr./m2 P2O5, 13 gr./m2 K2O and 0.8 gr./m2Zn), chemical weed control (3,36 kg ia ha-1 propanil, 3kg ia ha-1 butaclor and 0.19 kg ia ha-1 bentazon 5 days after transplanting DAT) and permanent flooding after 12 DAT were applied. Five plants in the middle row were tagged at heading and the degree of greenness of the flag leaf and the second leaf were measured using a Minolta Chlorophyll Meter SPAD-502 (Minolta Camera, Japan). Two readings were done at flowering and 38 days after flowering (maturity). The values were averages of three readings from five leaves: flag leaf color at flowering (FLC1), second leaf color at flowering (SLC1), flag leaf color at maturity (FLC7) and second leaf color at maturity (SLC7). The relatives rates of senescence in the flag leaf and second leaf were calculated by using the expression (Borrell, et al 1999). TRS=(((1-(leaf color at maturity/leaf color at heading))*100)/38

30
Other traits measured were: plant production (PP) averaged from three tagged plants in the central row, the grain number per panicle (GP) averaged from three panicles, one thousand grain weight (WG) and percentage of sterility (SP). Data analysis was made by the ¨partial ST¨ model (Geraldi and Miranda Filho, 1988) as explained by Torres and Geraldi (2007). According with that model, the total genotypes sum of squares was partitioned into parents, crosses and parents vs crosses. Additionally, parents were partitioned into dark green parents DG, light green parents LG and dark green vs light green. Variation due to crosses was partitioned into general DG (gi), general LG (gj) and specific (sij) combining ability. Significance of general and specific combining ability was tested with an error term considering only hybrid evaluation. Mid-parent heterosis was tested for significance using F-test for the contrast parents vs crosses in the analysis of variance. Results and discussion: Analysis of variance indicated the existence of differences among parents groups (Table 1). Dark green parents had higher yield, heavier 1000-grain weight, less sterility, and darker green leaves than light green parents. Light green parents had more grains per panicles. Also, there were differences between groups for several characters. There were no differences in the relative rate of senescence, indicating that senescence occurs at same rate in dark green parents, light green parents and hybrids. As dark green parents and hybrids are greener at flowering, their leaf color at maturity is darker. Hybrids were also different for all characters with exception of the relative rates of senescence and the color of the second leaf at maturity, indicating the release of genetic variability when dark and light groups were crossed and the feasibility of genetic analysis. General combining ability analysis showed that with the exception of percentage of sterility, the additive effects of the genes are more important than non additive effects. Light green parents were a more diverse group and had differences for breeding values for PP, GN, GW, CLF1, CSL1 and CLF7. Dark green parents were less diverse with differences for general combining ability for GN, GW and CSL1. The contrast of hybrids vs parents was significant for plant production, 1000-grain weight, flag leaf color at flowering and second leaf color at maturity, indicating the existence of mid-parent heterosis for this traits. Heterosis effects were positive for these characters showing directional dominance increasing these traits. The general combining effects (Table 2) for a parent determine its progeny average performance and is an estimate of its breeding value. In both of groups of parents there was a clear negative relationship between positive effects for grains number and negative effects for 1000-grain weight and vice versa. The best parent for high plant production was FL05372-7P-1-2P-1P-M and the worst was Centauro. Interestedly, DG parents were similar on their genetic effects for flag leaf greenness and plant production, suggesting they have the same genes for these traits.

31
For plant production and other characters, crossing positive GCA with average parents made good hybrids. For example, high production hybrids were obtained using FL05372-7P-1-2P-1P-M and CT16658-4-1-1SR-3-2-1-1-4-M. Hybrids with high grain number per panicle came from Centauro. A similar trend is observed for 1000-grain weight, crossing with FL06899-1P-6-1P-M and FL06562-3P-5-5P-M (Table 3). Flag leaf and second leaf greenness at flowering and maturity were positively correlated with plant productivity, indicating that dark green leaves contribute to more productive plants and heavier grain. The relative rate of senescence of both leaves was negatively correlated with plant production, i.e., faster senescence, less plant production. These results indicate that dark green slow senescent leaves can be a useful character in selecting for high yield (Table 4). Grain number per panicle was positively correlated with the percentage of sterility and negatively with 1000-grain weight, indicating that large panicles are more sterile and have lighter grains. Interestingly, grain number has an opposite relationship with leaf greenness and relatives rates of senescence than 1000-grain weight. It is negatively correlated with leaf greenness and positively correlated with the rate of senescence. It probably is associated with parents having and large panicles but light green rapid senescening leaves like Centauro. In applied breeding, it is better to select for dark green leaves at flowering, a visual trait, than only for slow senescening leaves. A dark green leaf probably has a higher photosynthetic efficiency, can accumulate more carbohydrates and have more stored nitrogen to export during the grain filling. At the same senescence rate, dark green leaves can maintain more greenness at maturity. The negative relationship between grain number and greenness can be simply the effect of parents like Centauro. There are other sources of large panicles as FL01028 with dark green foliage. Plant productivity and grain number were not correlated possibly because more grains compete for a given quantity of stored nitrogen and carbohydrates. This issue needs to be clarified. More greenness means more protein, which suggests a better food source for insect and pathogens. Dark green leaves may be an advantage only in high light environments. Non-improved indicas have a light green foliage probably because their adaptation to rainy seasons with low light.
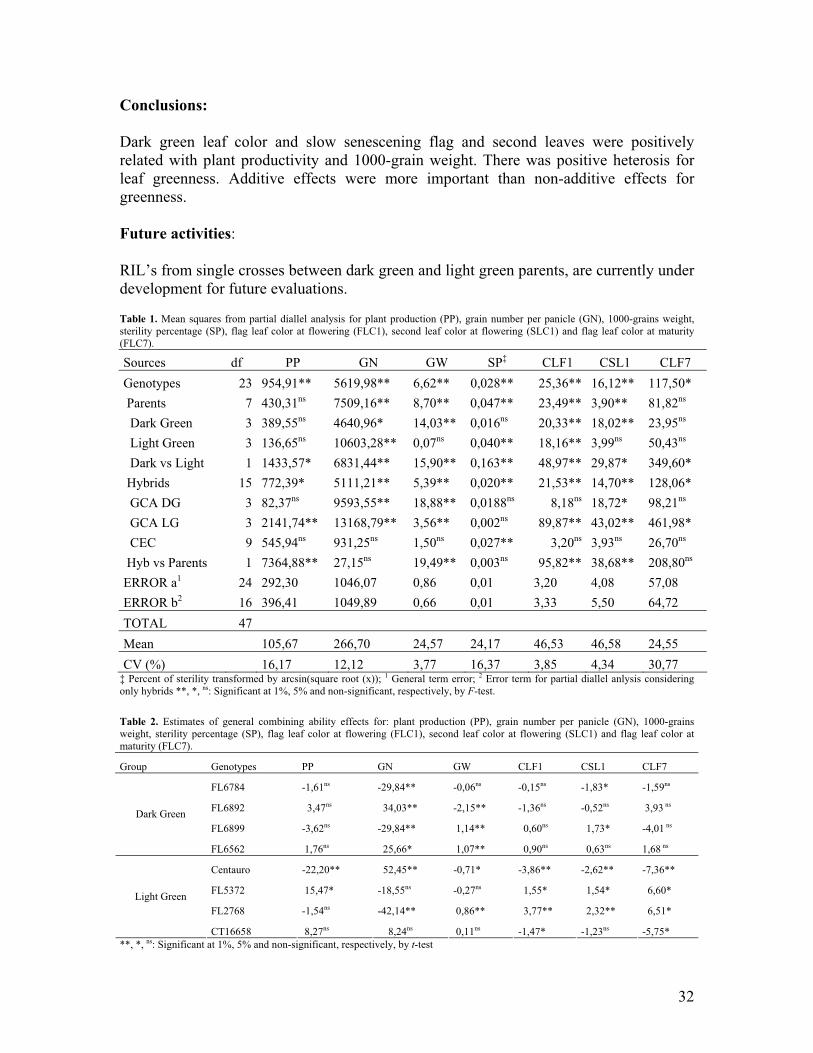
32
Conclusions: Dark green leaf color and slow senescening flag and second leaves were positively related with plant productivity and 1000-grain weight. There was positive heterosis for leaf greenness. Additive effects were more important than non-additive effects for greenness. Future activities: RIL’s from single crosses between dark green and light green parents, are currently under development for future evaluations.
Table 1. Mean squares from partial diallel analysis for plant production (PP), grain number per panicle (GN), 1000-grains weight, sterility percentage (SP), flag leaf color at flowering (FLC1), second leaf color at flowering (SLC1) and flag leaf color at maturity (FLC7).
Sources df PP GN GW SP‡ CLF1 CSL1 CLF7 Genotypes 23 954,91** 5619,98** 6,62** 0,028** 25,36** 16,12** 117,50* Parents 7 430,31ns 7509,16** 8,70** 0,047** 23,49** 3,90** 81,82ns Dark Green 3 389,55ns 4640,96* 14,03** 0,016ns 20,33** 18,02** 23,95ns Light Green 3 136,65ns 10603,28** 0,07ns 0,040** 18,16** 3,99ns 50,43ns Dark vs Light 1 1433,57* 6831,44** 15,90** 0,163** 48,97** 29,87* 349,60* Hybrids 15 772,39* 5111,21** 5,39** 0,020** 21,53** 14,70** 128,06* GCA DG 3 82,37ns 9593,55** 18,88** 0,0188ns 8,18ns 18,72* 98,21ns GCA LG 3 2141,74** 13168,79** 3,56** 0,002ns 89,87** 43,02** 461,98* CEC 9 545,94ns 931,25ns 1,50ns 0,027** 3,20ns 3,93ns 26,70ns Hyb vs Parents 1 7364,88** 27,15ns 19,49** 0,003ns 95,82** 38,68** 208,80ns ERROR a1 24 292,30 1046,07 0,86 0,01 3,20 4,08 57,08 ERROR b2 16 396,41 1049,89 0,66 0,01 3,33 5,50 64,72 TOTAL 47 Mean 105,67 266,70 24,57 24,17 46,53 46,58 24,55 CV (%) 16,17 12,12 3,77 16,37 3,85 4,34 30,77
‡ Percent of sterility transformed by arcsin(square root (x)); 1 General term error; 2 Error term for partial diallel anlysis considering only hybrids **, *, ns: Significant at 1%, 5% and non-significant, respectively, by F-test.
Table 2. Estimates of general combining ability effects for: plant production (PP), grain number per panicle (GN), 1000-grains weight, sterility percentage (SP), flag leaf color at flowering (FLC1), second leaf color at flowering (SLC1) and flag leaf color at maturity (FLC7).
Group Genotypes PP GN GW CLF1 CSL1 CLF7
FL6784 -1,61ns -29,84** -0,06ns -0,15ns -1,83* -1,59ns
FL6892 3,47ns 34,03** -2,15** -1,36ns -0,52ns 3,93 ns
FL6899 -3,62ns -29,84** 1,14** 0,60ns 1,73* -4,01 ns Dark Green
FL6562 1,76ns 25,66* 1,07** 0,90ns 0,63ns 1,68 ns
Centauro -22,20** 52,45** -0,71* -3,86** -2,62** -7,36**
FL5372 15,47* -18,55ns -0,27ns 1,55* 1,54* 6,60*
FL2768 -1,54ns -42,14** 0,86** 3,77** 2,32** 6,51* Light Green
CT16658 8,27ns 8,24ns 0,11ns -1,47* -1,23ns -5,75* **, *, ns: Significant at 1%, 5% and non-significant, respectively, by t-test

33
Table 3. Means values for: plant production (PP), grain number per panicle (GN), 1000-grains weight, sterility percentage (SP), flag leaf color at flowering (FLC1), second leaf color at flowering (SLC1) and flag leaf color at maturity (FLC7).
Genotype PP GN GW SP FLC1 SLC1 FLC7
FL6784 112,08 217,84 25,39 11,50 47,65 48,00 26,55
FL6892 79,05 291,84 21,04 17,00 44,90 44,00 22,10
FL6899 96,67 191,67 26,36 12,00 49,95 50,90 30,55
FL6562 103,05 279,17 25,89 26,50 42,64 43,84 25,90
Dark green parents 97,71 245,13 24,67 16,75 46,29 46,68 26,28
Centauro 68,61 331,17 22,52 52,00 42,20 43,90 14,90
FL5372 83,80 217,67 22,95 27,00 47,15 45,85 24,40
FL2768 75,74 231,65 22,58 21,00 41,50 43,60 14,85
CT16658 86,97 365,00 22,67 35,50 40,30 42,45 13,55
Light green parents 78,78 286,37 22,68 33,88 42,79 43,95 16,93
Parents 88,24 265,75 23,67 25,31 44,54 45,32 21,60
FL6784xCentauro 87,80 268,17 24,29 20,50 44,35 43,35 18,95
FL6784xFL5372 136,92 226,00 24,74 17,50 48,90 48,70 33,00
FL6784xFL2768 120,44 212,00 25,30 14,50 51,30 45,80 30,75
FL6784xCT16658 106,46 243,67 25,50 19,50 45,00 43,70 15,05
FL6892xCentauro 97,78 369,00 22,42 24,00 40,75 43,10 22,45
FL6892xFL5372 116,40 267,33 23,18 32,00 48,10 47,70 38,95
FL6892xFL2768 127,54 233,50 24,24 13,00 51,42 50,85 32,30
FL6892xCT16658 130,28 335,50 21,66 32,50 44,40 45,15 26,10
FL6899xCentauro 93,41 301,83 24,93 24,50 43,90 46,35 16,10
FL6899xFL5372 111,94 237,50 26,40 27,00 49,00 49,85 22,20
FL6899xFL2768 116,52 193,34 26,22 15,50 52,50 52,45 31,30
FL6899xCT16658 121,74 217,17 27,12 23,00 47,15 47,15 18,45
FL6562xCentauro 90,29 340,00 25,61 21,00 45,70 45,60 17,15
FL6562xFL5372 154,72 264,17 24,71 25,00 50,35 48,80 36,35
FL6562xFL2768 87,43 261,83 27,79 49,00 50,00 49,05 35,80
FL6562xCT16658 132,70 305,84 26,29 18,50 47,70 47,95 21,50
Hybrids 114,52 267,30 25,02 23,56 47,53 47,22 26,03

34
Table 4. Phenotypic correlations between plant production (PP), grain number per panicle (GN), 1000-grains weight (WP), sterility percentage (SP), flag leaf color at flowering (FLC1), second leaf color at flowering (SLC1), flag leaf color at maturity (FLC7), second leaf color at maturity (SLC7), relative rate of senescence of flag leaf (RRSFL) and relative rate of senescence of second leaf (RRSSL).
PP GN SP WP FLC1 SLC1 FLC7 SLC7 RRSFL RRSSL
PP 1,00 -0,21ns -0,37** 0,29* 0,56** 0,37** 0,43** 0,42** -0,35* -0,38*
GN -0,21ns 1,00 0,46** -0,43** -0,56** -0,38** -0,41** -0,41** 0,32* 0,37*
SP -0,37** 0,46** 1,00 -0,18ns -0,31* -0,16ns 0,00ns 0,00ns -0,06ns -0,03ns
WP 0,29* -0,43* -0,18ns 1,00 0,52** 0,46** 0,14ns 0,31* -0,03ns -0,24ns
FLC1 0,56** -0,56** -0,31* 0,52** 1,00 0,79** 0,58** 0,58** -0,40** -0,48**
SLC1 0,37* -0,38** -0,16ns 0,46** 0,79** 1,00 0,40** 0,45** -0,26ns 0,32*
FLC7 0,43** -0,41** 0,00ns 0,14ns 0,58** 0,40** 1,00 0,67** -0,98** -0,63**
SLC7 0,42** -0,41** 0,00ns 0,31* 0,58** 0,45** 0,67** 1,00 -0,60** -0,99**
RRLS -0,35* 0,33* -0,06ns -0,03ns -0,40** -0,26ns -0,98** -0,60** 1,00 0,58**
RRSS -0,38* 0,37* -0,03ns -0,24ns -0,48** -0,32ns -0,63** -0,99** 0,58** 1,00 **, *, ns: Significant at 1%, 5% and non-significant References Borrell, A.; Bidinger, F. and Sunitha K. 1999. Stay Green associated with yield in recombinant inbred sorghum lines varying in rate of senescence. Crop Science. 1999. Borrell, A.; Hammer, G. and Douglas, A. 2000. Does maintaining green leaf area in Sorghum improve yield under drought?. I. Leaf growth and senescence. Crop Science 40: 1026 – 1037. BorrelL, A. and Hammer, G. 2000. Nitrogen dynamics and the physiological basis of stay-green in Sorghum. Crop Science 40: 1295 – 1307. Geraldi, IO and Miranda Filho JB. 1988. Adapted models for the analysis of combining ability of varieties in partial diallel crosses. Brazilian Journal of Genetics. 11(2): 419 – 430. Jiang, C.; Ishihara, K.; Satoh, K. and Katoch, S. 1999. Loss of the photosynthetic capacity and proteins in senescing leaves at top positions of two cultivars of rice in relation to the source capacities of the leaves for carbon and nitrogen. Plant Cell Physiology 40(5): 496 – 503. Miranda Filho, JB. and Geraldi IO. 1984. An adapted model for the analysis of partial diallel analysis. Brazilian Journal of Genetics 7: 677 – 688. Jennings, PR.; Berrio, LE.; Torres, EA. and Corredor EA. 2002. Una estrategia de mejoramiento para incrementar el potencial de rendimiento en el arroz. Foro Arrocero Latinoamericano 8(2): 10-13. Kropff, M.; Cassman, K.; Peng, S.; Matthews R. and Setter T. 1994. Quantitative understanding of yiel potential. In: Cassman, K. et al. 1994. Breaking the yield barrier: Proceedings of a workshop on rice yield potential in favorable environments. International Rice Research Institute, 29-Nov to 4 Dec 1993. International Rice Research Institute. Manila Philippines. Peng, S. 2000. Single-leaf canopy photosynthesis of rice. In: Seehy, J.; Mitchell, P. and Hardy, B (eds). 2000. Redesigning rice photosynthesis to increase yield. Proceedings of the workshop on the quest to reduce hunger: Redesigning Rice Photosynthesis, 30 Nov - 3 Dec 1999. Los Baños Phillipines. International Rice Research Institute and Amsterdam: Elsevier Science B.V. 293 p.

35
Torres, EA. And Geraldi IO. 2007. Partiall diallel analysis of agronomic characters in rice (Oryza sativa L.). Genetics and Molecular Biology 30 (3). 605 -613. Yonezawa, K. 1997. Yield components. In: Matsuo, T.; Futsuhara, Y.; Kikuchi, F. and Yamaguchi, H. 1997. The science of rice plant. Volume three Genetics. Food and Agriculture Policy Research Center, Tokyo, Japan. p. 1003. Yoshida, S. 1981. Fundamentals of rice crop science. International Rice Research Institute. Los Baños, Laguna, Phillipines. p. 269.
1B. UPLAND RICE
1.B.1. Rice Synthetic Population Improvement
M. Châtel, Y. Ospina, A. Audebert, F. Rodríguez, D. Guzman and LAC NARS
Funding: Cirad, CIAT and Cooperators
Abstract The Cirad/CIAT rice collaborative project concentrates on broadening the genetic base of rice through the development and improvement of synthetic rice population using recurrent selection and conventional breeding methods. Recurrent selection is not intended to replace conventional breeding that have been successful. Recurrent selection is a complement to others breeding methods and has to be considered as another type of breeding method in the breeders’ arsenal available for developing improved varieties. The project conducts centralized upland breeding and pre-breeding activities in Colombia, and decentralized upland-aerobic and lowland rice breeding projects at regional level with cooperators in Latin America and the Caribbean (LAC). Basic synthetic populations are developed, shared and enhanced with regional cooperators. In Colombia, basic upland synthetic populations are enhanced using two recurrent selection-breeding methods: mass and S2 progenies evaluation. At each step of enhancement, fertile plants are selected for the development of segregating lines and progeny selection using the conventional pedigree method. The most advanced upland lines are shared with LAC cooperators through nurseries and are locally screened and selected. The best promising lines are evaluated at countries’ regional yield trials. The first upland rice commercial variety coming from population breeding was released in Bolivia in January 2006. For lowland irrigated rice ecosystems, the activities are completely decentralized and conducted in very close collaboration with LAC cooperators who adopted the novel breeding method. Several candidate advanced lines are in the process of becoming varieties in different countries. Leadership in networking permits to maintain the “Working Group on Advanced Rice Breeding” (GRUMEGA its Spanish acronym) strong and efficient with impact on research, capacity building and the development of the rice sector.

36
Key words: Rice synthetic population; Recurrent selection; Centralize and decentralize breeding; Rice ecosystems
Background Since the 60’s rice commercial cultivars released were developed by conventional crossbreeding methods such as pedigree, usually restricted to breeding populations often coming from the cross of 2 inbred lines. The genetic base of breeding materials is therefore becoming more restricted and the breeding method emphasizes inbreeding. Composite populations are expected to express greater variability than individual crosses, and recurrent selection breeding applied to composite populations that minimizes the effects of inbreeding, can contribute to broaden the genetic base of breeding programs. Genetic uniformity, or lack of genetic diversity, is of major concern to breeders, geneticists, and the agricultural community in general. In LAC, the genetic diversity of rice cultivars depends on a small genetic landraces core as described by different authors (Cuevas-Pérez et al., 1992; Rangel et al., 1996 and Montalván et al., 1998.) A way of broadening the genetic base of Latin American rice and assessing the genotype-by-environment interaction is to identify specific potential parents and pool them to develop new, genetically broad-based, breeding material (Guimarães, 2005) From 1996, with LAC regional cooperators, we concentrate on the development and enhancement of upland rice composite populations (Châtel and Guimarães, 1998.) The breeding strategy uses a recessive male-sterile gene (ms) from a mutant of IR36 (Singh & Ikehashi, 1981) easing the development of the multiple-parent populations as well as natural recombination during the enhancement process. Results
Upland and aerobic rice At CIAT-Colombia, upland rice composite basic populations are enhanced using two recurrent selection breeding methods; mass and S2 progenies evaluated using the statistical design described by Federer (1956) The main outputs of the project are the creation of breeding populations with broad genetic diversity, their enhancement by recurrent selection and the selection of superior genotypes as potential improved varieties but also as parental stocks for conventional crossbreeding programs. To do so, fertile plants are selected from the basic composite populations and at each one of their enhancement steps. They are the starting point for the selection of segregating progenies by conventional pedigree method and the identification of segregating and advanced lines to be shared with LAC cooperators. In Colombia, the main characteristics we select for are: early vigor, tolerance to soil acidity, resistance to diseases (mainly rice blast: Pyricularia grisea Sacc.), grain quality (translucent, long-slender grain), early maturity with a seed to seed cycle about 100 days, and yield potential.

37
Population breeding by recurrent selection is efficient for traits that show low heritability. Through short cycles of selection and recombination, linkage barriers are broken down and favorable genes are accumulated. This is a smooth process of continuous improvement. Basic populations were improved using recurrent selection in centralized pre-breeding activities (Châtel et al. 2005; Ospina et al. 2005). Upland composite populations are observed, characterized and improved by recurrent selection in Colombia, and improved lines are distributed to national programs in the region for local testing.
Lowland irrigated rice Lowland irrigated rice improvement is carried out in close collaboration with partners in the LAC region. In Colombia, the Cirad-CIAT rice project started developing basic populations targeting the various lowland rice ecosystems present in LAC, in partnership with scientists in Colombia, Venezuela and Cuba for the tropical ecosystem; Argentina for the subtropics; and Chile and France for the temperate zone. The basic populations were shipped to regional partners and evaluated locally. Most of the cooperators used this material to develop site-specific populations by introgressing additional variability to meet their specific breeding objectives. They then used these populations in their rice-improvement programs by recurrent selection.
Integrating population breeding and applied breeding The main purpose of a breeding project is to create variability and develop breeding materials that may lead to identifying promising lines and new cultivars for release. Recurrent selection methods can contribute to meeting the goals of continuous genetic improvement but should be integrated with other breeding methods to deliver superior breeding materials and improved varieties. Recurrent selection breeding should not be considered a separate phase of an applied breeding program (Hallauer and Miranda 1982). Selfed progenies extracted from recurrent populations are evaluated and then recombined to obtain improved populations. Superior progenies also have to be including in the applied breeding program, passing through cycles of selection and agronomic evaluation. Advanced lines are the starting point for developing commercial varieties and are donors in crossbreeding programs.
Commercial line release
Upland and aerobic rice
In Bolivia, the Cirad-CIAT project has been collaborating with both the public (CIAT Santa Cruz) and the private institutions of rice sector (Consejo Nacional del Arroz – CONARROZ). The first upland/aerobic commercial variety selected from the enhanced synthetic population PCT-4 was officially released as ESPERANZA. The variety is adapted both to manual upland and mechanized aerobic rice ecosystems. For the smallholders, earliness and drought tolerance associated with good yield potential and grain quality are important traits the variety has. The new variety ease rotation with other food or cash crops. Earliness also allows farmers to place rice to the market early in the

38
season, when prices are high. The variety shows a long and slender grain, very much praised by the rice industry (Taboada et al. 2005.)
Promising line for release
Upland and aerobic rice
In Cuba, with the Instituto de Investigaciones en Arroz (IIA), one advanced line from the population PCT-4 has completed the process of evaluation and selection and is now in regional yield trials (Pérez Polanco et al. 2005). It shows good yield potential, good resistance to rice blast and lodging, and broad adaptation to the various cropping situations of the so called “popular rice” ecosystem that ranges from upland and aerobic conditions to irrigated.
In Nicaragua, with the Instituto Nicaraguense de Tecnología Agropecuaria (INTA) and farmers groups, the project has evaluated upland synthetic populations as well as advanced and segregating lines shipped from Colombia. Promising lines selected from the population PCT-4 are being evaluated in regional participatory varietal selection yield trials, and one (PCT-4\SA\1\1>1479-M-1-M-1) was identified as very promising. Introduced synthetic populations were used to start participatory plant breeding as well as the source of male-sterile gene and good genetic background. New site-specific populations for Nicaragua and Central America are being developed and participatory enhanced (Trouche 2005).
In Venezuela, the Instutito Nacional de Investigación Agrícola (INIA-Guárico) applied for the release next year of two (2) Cirad/CIAT upland rice lines as an output of the “Proyecto FCI – 01 (PR11-01) titled “Obtención de Cultivares de Arroz (Oryza sativa L.) con Bajos Requerimientos de Agua”. These lines are well adapted to the irrigated system prevalent in the region, are high yielding promising cultivars and required less irrigation water that leads to better water use efficiency.
Commercial line release
Irrigated rice In Chile, the Instituto de Investigaciones Agropecuarias (INIA Quilamapu) is using site-specific populations developed from the introgression of locally adapted material into the population GPIRAT-10 (Hernaiz et al. 2005). The Chilean populations were being improved for cold tolerance at the vegetative stage and for other agronomic traits. Segregating lines were developed during the improvement process, and passed through regional yield trials. The first commercial variety, RQuila 23, adapted to the temperate irrigated rice ecosystem, coming from the enhancement and selection of the population PQUI-1 was proposed for official released in 2007.

39
Promising line for release
Irrigated rice
In Venezuela, at the Fundación para la Investigación Agricola (DANAC) rice breeding project, 43% of the advanced lines come from introduced and site-specific composite populations. A line from the population PCT-16 was identified as candidate cultivar for release in 2007 (Jayaro et al. 2005).
In France, the aromatic population PACQ-1 specially designed for the temperate rice ecosystem is the fruit of the close collaboration between the Cirad-Camargue and INIA Quilamapu (Chile) rice breeding projects. This site-specific population was synthesized in Colombia by the Cirad/CIAT rice project. Selected parents identified by Dr. Guy Clément, from the Cirad/CFR rice collaborative breeding project (France), were crossed with male-sterile plants from the Cirad/INIA Chile population PQUI-1 well adapted to cold temperature. After several cycles of recombination, progenies were extracted from the population and evaluated, both for agronomic characteristics and presence of aroma. The first results are very promising (Guy Clément, personal communication), S3 progenies show a great percentage of aromatic lines. And some are as scented as the best scented parent used to synthesize the aromatic population.
Networking LAC Breeding Projects and Breeders
GRUMEGA Network This is a framework for collaborative research built on five pillars: (i) capacity building, (ii) germplasm development and sharing, (iii) workshops for germplasm evaluation and selection, (iv) conferences to present results and advances, and (v) publications with and by collaborators. The IV GRUMEGA Conference was held in January 2006 in Chile and was organized by INIA-Quilamapu, Cirad/CIAT and FAO. In 2007 we published the web page of the GRUMEGA network (please have a look at the site http://www.grumega.org)
Regional Technical Cooperation Project (TCP) The regional TCP “Capacitación en fitomejoramiento genético e intercambio de germoplasma para utilizar los recursos genéticos del arroz en América Latina y el Caribe” involving 7 countries (Argentina, Bolivia, Chile, Cuba, Guatemala, Nicaragua and Dominican Republic), Cirad and CIAT was accepted and funded by FAO for 2 years, starting January 2006. (for more details see the web site: http://agr.unne.edu.ar/fao/index.htm) In this framework two (2) Breeders’ Workshops were organized, one in Colombia (2006) and the other one in Argentina (2007). Three (3) training courses were also organized in 2006 and 2007 in Chile, Cuba and Nicaragua. In 2006, the regional workshop, organized by Cirad-CIAT was held in Villavicencio Colombia in August. Sixty (60) rice breeders from 14 LAC countries and one 1 from

40
Cirad/SCRID Madagascar (Louis Marie Raboin) attended the workshop and selected upland and irrigated segregating and fixed lines. The participants selected a total of 983 and 1310 upland and irrigated lines respectively. These numbers show the importance for the region in having access to public genetic resources developed by international centers.
The way forward in rice synthetic population improvement Since 1996, improving rice biodiversity has become an important breeding alternative in LAC because of investment in capacity building, breeders’ confidence in the outcomes, and regional networking between international and national programs. The next step is to take advantage of new tools (Eco-physiology and Molecular) to increase the efficiency of recurrent selection breeding. In 2005, molecular tools were used, for example, to better assess the level of genetic diversity in a synthetic population (Ramis et al. 2005). Starting in 2006, Cirad and CIAT developed new activities in breeding for drought tolerante, implemented thanks to three (3) field missions of Dr. Alain Audebert, eco-physiologist at Cirad Montpellier. By mid-2008, the Cirad/CIAT Rice Breeding Project will be reinforced by the posting at CIAT of a new Cirad Scientist (Cécile Grenier) who will be in charge of implementing Marker Assisted Recurrent Selection (MARecS) and scientific prospects and approaches for the development of japonica’s hybrid rice. As mentioned above, the Cirad rice project at CIAT will be a more integrated and efficient project for rice breeding for drought tolerance and water use efficiency using: (1) recurrent selection breeding; (2) tools in eco-physiology and modeling and (3) molecular markers. The integration of these scientific disciplines are in accordance with the activities to be implemented by the new Cirad Research Unit AIVA (UPR 59+8+6 Merging) and the Cirad ATPd Bios “Diversité des caractères d’adaptation aux contraintes hydriques et thermiques chez le riz (Oryza sativa L.) : Phénotypage à grande échelle dans le cadre d'études d'association”.
Rice Synthetic Population Improvement : Highlights 2007
Line release - “ESPERANZA” Upland rice variety in Bolivia coming from population breeding was released in Bolivia for both manual and mechanized rice ecosystems.- “RQUILIA 23” First irrigated rice variety coming from population breeding in Chile, adapted to the temperate rice ecosystem.
Publications- Web Page Development : http://www.grumega.org- Red de Mejoramiento Genético de Arroz en América (Red-MeGA): Development of the “Estatutos y Reglamento para la Creación y el Funcionamiento de la Red-MeGA”. - Slide Show: Mejoramiento Genético de Arroz. Selección Recurrente utilizando Androesterilidad Genética: un nuevo Método de Selección.

41
Special Project - FAO Regional Tecnical Cooperation Project (TCP/RLA/3102). Capacitación en Fitomejoramiento Genético e Intercambio de Germoplasma para utilizar los Recursos Genéticos del Arroz en América Latina y el Caribe. The Regional TCP Project involves 6 Countries: Argentina, Chile, Cuba, Guatemala, Nicaragua and Dominican Republic and 3 International Institutions: FAO, CIAT and Cirad.
Training Courses: - II Curso de capacitación en fitomejoramiento genético de arroz: Proyecto TCP/RLA/3102 (A). Chillán – Chile. 22 al 25 de Enero del 2007. Marc Châtel, Yolima Ospina. Mejoramiento Genético de Arroz. Selección Recurrente utilizando Androesterilidad Genética: un nuevo Método de Selección. - III Curso de capacitación en fitomejoramiento genético de arroz: Proyecto TCP/RLA/3102 (A). Managua- Nicaragua. 27 al 31 de Mayo del 2007. Marc Châtel, Yolima Ospina. Mejoramiento Genético de Arroz. Selección Recurrente utilizando Androesterilidad Genética: un nuevo Método de Selección.
Liaison Officer: n between FAO (regional LAC Office and Rome Headquarters), CIAT and Cirad: - Funding by FAO of a Regional TCP Project. - Implementation of the proposed activities (Breeders Workshops and Training courses) Inter-institutional liaison between CIRAD and CIAT: - Implementing new research activities linking Population Breeding and Eco-physiology (3 weeks visit of a Cirad Scientist at CIAT and field experiment at Villavicencio-Colombia). - Implementation of a rice population breeding project in France (Camargue) targeting the development of aromatic varieties.
- New Cirad Scientist to be posted at CIAT in mid-2008 to reinforce the actual collaborative breeding project
References Châtel M, Guimarães EP. 1996. Selección recurrente con androesterilidad en arroz (Motta F, ed.). CIAT publication No. 246. CIAT, Cali, Colombia. 70 p. Châtel M, Ospina Y, Rodríguez F, Lozano VH. 2005. CIRAD/CIAT Rice Project: Population improvement and obtaining rice lines for the savannah ecosystem. In: Population improvement: A way of exploiting the rice genetic resources of Latin America (Guimarães EP, ed.). FAO, Rome, Italy. pp 237-253. Cuevas-Pérez FE, Guimarães EP, Berrio LE, González DI. 1992. Genetic base of irrigated rice in Latin America and the Caribbean, 1971 to 1989. Crop Sci. 32:1054-1059. Guimarães EP, Châtel M. 2005. Exploiting rice genetic resources through population

42
improvement. In: Population improvement: A way of exploiting the rice genetic resources of Latin America (Guimarães EP, ed.). FAO, Rome, Italy. pp 3-17. Hallauer AR, Miranda JB. 1982. Quantitative genetics in maize breeding (2nd edition). Iowa State University Press, Ames, Iowa, USA. pp. 468. Hernaiz S, Alvarado JR, Châtel M, Castillo D, Ospina Y. 2005. Improving irrigated rice populations of the temperate climate in Chile. In: Population improvement: A way of exploiting the rice genetic resources of Latin America (Guimarães EP, ed.). FAO, Rome, Italy. pp 129-143. Jayaro Y, Gamboa C, Graterol E. 2005. Avances del programa de mejoramiento poblacional de arroz mediamte selección recurrente en la Fundación DANAC, Venezuela. Proceedings of the III Encuentro Internacional del Arroz, Havana, Cuba, 6-10 June 2005. p 2. Ospina Y, Guimarães EP, Châtel M, Duque MC. 2005. Effects of selection and of recombinations on an upland-rice population. In: Population improvement: A way of exploiting the rice genetic resources of Latin America (Guimarães EP, ed.). FAO, Rome, Italy. pp 331-350. Pérez Polanco R, Châtel M, Guimarães EP. 2005. Acquiring a basic understanding of rice population improvement for use in Cuba. In: Population improvement: A way of exploiting the rice genetic resources of Latin America (Guimarães EP, ed.). FAO, Rome, Italy. pp 205-220. Ramis C, Badan AC de C, Díaz A, Gamboa CE. 2005. Molecular markers as tools for rice population improvement. In: Population improvement: A way of exploiting the rice genetic resources of Latin America (Guimarães EP, ed.). FAO, Rome, Italy. pp 75-93. Singh RJ, Ikehashi HI. 1981. Monogenic male-sterility in rice: Introduction, identification and inheritance. Crop Sci. 21:286-289. Taboada R, Guzman R, Viruez J, Callaú VH, Châtel M, Ospina Y, Rodriguez F, Lozano VH. 2005. Bolivia lanza la primera variedad de arroz de secano originada del mejoramiento poblacional. Proceedings of the III Encuentro Internacional del Arroz, Havana, Cuba, 6-10 June 2005. p 1. Trouche G. 2005. Participatory rice breeding, using population improvement: A new methodology adapted to the needs of small farmers in Central America and the Caribbean. In: Population improvement: A way of exploiting the rice genetic resources of Latin America (Guimarães EP, ed.). FAO, Rome, Italy. pp 99-109.

43
1C. BIOTECHNOLOGY ACTIVITIES RELATED TO ENHANCED GENE POOLS
1.C.1 SNPs Variation of iron homeostasis genes in mutant and natural rice populations.
Yamid Sanabria, Jaime Borrero, James Carabalí, Matthieu Bouniol, César Martínez, Joe Tohme.
FundingCIDA, CIAT CORE International Center for Tropical Agriculture, CIAT. Cali Colombia.
Background
Iron deficiency is a serious problem in developing countries affecting over 30% of world population (WHO, 2004). Iron deficiency is related to food diets low in iron content. Meet and others food presents high iron amounts but it is not always affordable for resource-poor populations. Furthermore, the iron content of rice, representing the major calorific supply in developing countries is not sufficient to provide recommended daily amounts of iron. A way to combat deficiencies is to increase nutritional values of staple food crops of major importance. Many genes are involved in iron transport and signalling pathways in plants (Curie and Briat, 2003). Knowledge of genes involved in iron homeostasis, from uptake to storage in rice, is a key step to understanding iron transport, distribution and accumulation in the rice grain. This knowledge should facilitate the development of varieties with iron enriched seeds. Several genes involved in iron homeostasis have been identified in rice, such as OsIRO2 (Ogo et al., 2007), OsYSL2 (Koike et al., 2004), and in other species, such as VIT1 in Arabidopsis (Kim et al., 2006). Gross and collaborators (2003) assigned functions to thirty-nine rice new genes, by means of comparing previously reported sequences for others species with the rice genomic sequence; however, experimental information and mutants are necessary to validate these putative genes. Mutant population development facilitates the study of gene functions. A mutant population of 8000 individuals has been developed in IAEA (International Atomic Energy Agency, Vienna, Austria), by gamma rays mutagenesis. Two different doses of radiation were used, 150 and 250 GY. We are searching for mutations in iron homeostasis genes in this mutant rice population and among varieties. The TILLING technology (Targeting Induced Local Lesions in Genomes) (Comai and Henikoff, 2006) is used to reveal mutations in target genes. This technique consist in the amplification of the targeted gene sequence, its heteroduplex formation, digestion with the CEL I endonuclease (extracted from celery), and finally visualization in gels that reveals mutation sites. We are aware of the limitations of this technique in mutant populations derived by gamma rays treatment. Unlike other mutagenesis methods such as T-DNA insertion, TILLING does not always inhibit gene expression. Moreover, it permits the

44
identification of allelic series in genes of interest, which is important in the study of gene function. In our population, low doses of gamma rays produce changes in simple nucleotide but can also result in sequence deletions.
Materials and Methods
Mutant population
Seed of five materials originated from CIAT and FLAR breeding programs (Bg 90, CT11275-4-M-1-M, FL03191-5P-10-1P-3P-M-M-M, FL04577-3P-11-4P-1P-M, and FL04577-3P-11-4P-1P-M) were irradiated with gamma rays with doses of 150 and 250 GY, at the International Atomic Energy Agency, Vienna, Austria (IAEA). All treated seed (M1) were sowed at CIAT in 2006. From each material 1600 plants were selected randomly, and M2 seed was harvested. One M2 seed per line was sowed to evaluate the effect of gamma ray over plants. Natural population
Based on iron content data, 51 lines, including wild species, with some variation were selected to search for natural changes in gene sequences involved in iron homeostasis. Search for SNP variation
To discover polymorphisms in mutant and natural populations TILLING method was used. First, we amplified the sequence target on our genes of interest (Table 1); second, heteroduplexes were formed. In mutant populations, evaluation is achieved in bulks of 6 individuals, therefore, heteroduplex is formed between the mutant and no mutants individuals. In natural populations is necessary to use a sequence of reference to detect mutation; in our case, we used as reference the variety Nipponbare. In each PCR reaction, Nipponbare DNA and the line DNA to evaluate is mixed. Amplification of sequences of these two varieties and later heteroduplexes lets us to detect mismatches sites by the endonuclease CEL 1 extracted from celery. Final digestions were observed in agarosa gels 2% (Figure 1).
Phenotype Data
Phenotypic data collection has been completed in about 6000 M2 mutant plants. Data were taken 15 days after germination and after flowering. Information was stored on RiceTilling Database that is based in ExcelTM files. Database allows recording and utilization of phenotypic and molecular information, including images.

45
Results and Discusion Mutations in seven genes involved in iron homeostasis: OsYSL1, OsNRAMP, OsFER1, OsFER2, OsVIT1, OsVIT2, and OsIRO2 have been searched in a population of 4000 M2 individuals. The study revealed mutations in four of these genes especially in OsYSL1. However, allelic variation is not high, possibly some regions in the genome could be hot spots for mutations. SSR markers are being used to check if these individuals are not contaminations. The iron content in seeds of mutant plants is being estimated by atomic absorption in order to evaluate the effect of genetic variations on iron accumulation in seeds. Besides that, natural variation of these genes is being evaluated in varieties and in wild relatives with this same methodology using CEL I. Important SNP variations have been found in wild species as well as in Indica and Japonica varieties (Figure 2). These natural populations show a major allelic variation (Table 2); for example, six different alleles were detected for the sequence OsIRO 2A and these variations could be important as molecular markers in genetic mapping. Phenotypic variation also has been observed under field conditions. Observations done 15 days after germination showed 0.8 % of albino plantlets, but these phenotypes did not survive more than 30 days. After flowering, others mutant phenotypes were observed related mainly with plant height (dwarf and tall plants), days to flower, leaf shape, awns, tiller number and spikelet sterility. All these phenotypes can be considered for others works in functional genomics. The rice improvement program of CIAT also targets other objectives. Thus, this mutant population is also being evaluated for herbicide resistance, pathogen resistance and yield traits. Molecular and phenotypic results are saved in the Rice Tilling Database (Figure 3), which, in the future, will be available on the internet. The whole population could be further explored. It is highly desirable to develop a new population obtained by chemical mutagenesis to increase the number of available mutations. Including others varieties for natural variation could be interesting too, in order to do relations between mutations and iron contents in the rice germplasm.
Figure 1. SNPs detection for OsNRAMP gene in a plant of the rice mutant population. After heteroduplex formation, mutant and wild amplified sequences are join, Cel I enzyme cuts on the loop originated from no paired bases. Cutting permits show 2 bands in mutants individuals (arrow).

46
Table 1. Primer sequences for Iron homeostasis genes. Primers were designed based on putative genes sequences from gene bank. For VIT genes, alignments with VIT gene Arabidopsis sequence (Accesion number at gene bank: At2g01770) were achieved, these reveled 2 homologous sequences on rice.
Gene Primer Sequence OsYSL 6 OsYSL6n BF CCTAATGATGTGCATGACGTTT OsYSL6n BR TAAGAGGGAAGAAAACGTGGAA OsYSL6n CF TAGCTGAAGCTGAGTGAATTGC OsYSL6n CR TAAGTGATGGTTTGCTTTGTGC OsFER 1 OsFer1n BF GTGCGCATTTCTTCTTCTTCTT OsFer1n BR GGAGGGAGTAATTGACATCAGG OsFER 2 OsFer2n AF GACCAATCATCAGAGGAAGGAG
OsFer2n AR GGAGGAGGAGGAAAACAGACTT OsNRAMP1 OsNramp1 BF CCCTACCATTGTCGCATTTATT OsNramp1 BR CACACATTAACAACTGGGGGTA OsVIT 1 OsVIT1nBF TAGGAAAGCCCATAGTGTTCGT OsVIT1nBR TTTTAGGGTCTGGTTTCTCCAA OsVIT1nCF TCGATTATCCTGTATGCGTTTG OsVIT1nCR AGTATTGGCAACCTGATTCGTT OsVIT 2 OsVIT2nCF ATAGGTTTGAGTTGGGACTGGA OsVIT2nCR ACTCCCTTTCTCTCTCGCTTTT OsIRO2 OsIRO2 AF ACTGGCCCCCAGAAAGTAGT OsIRO2 AR AGAGCTCGTTGAGCTGCTTC OsIRO2 BF CCCAGCATTTTGTGAAAGGT OsIRO2 BR TCAGGATGTGCGTCCTCATA
Figure 2. Natural variation in some rice varieties. Two different alleles for OsFER 2 gene, allele 1 is present in lines 2 (FED 50-1,BCF 1973,FED) and 10 (IR75862-206-2-8-3-B-B-B); allele 2 is present in line 6 (African rice, Oryza glaberrima)
1 2 3 4 5 6 7 8 9 10
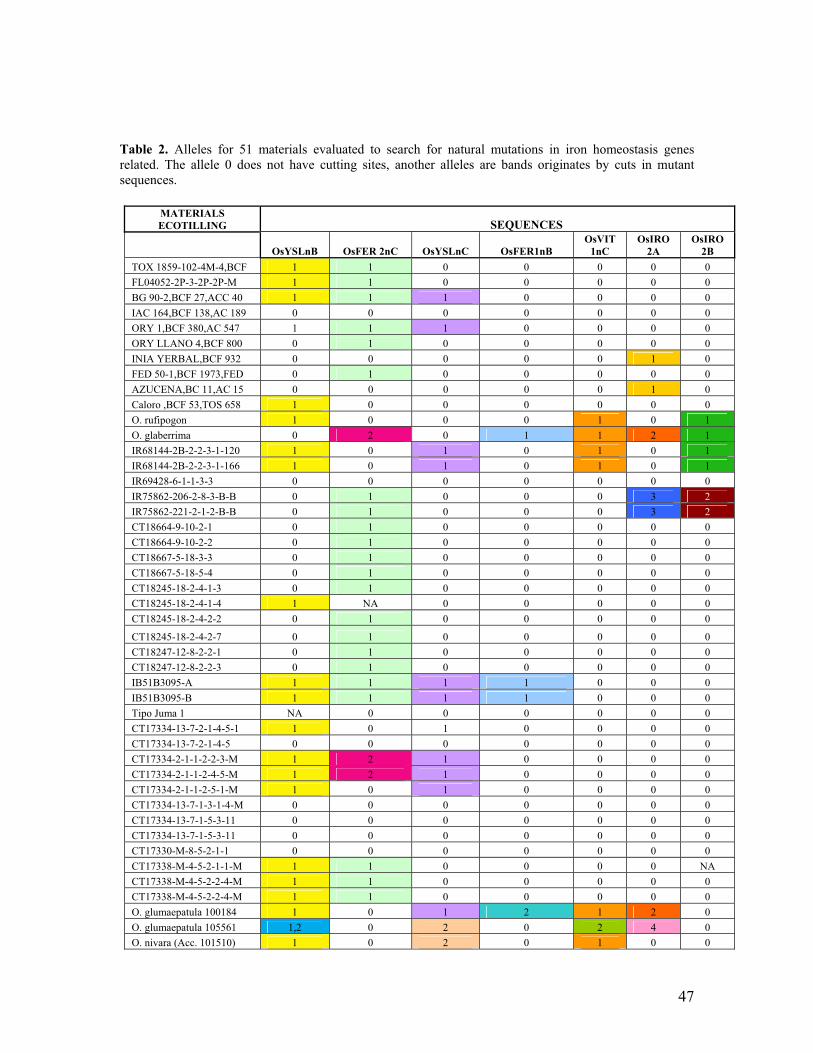
47
Table 2. Alleles for 51 materials evaluated to search for natural mutations in iron homeostasis genes related. The allele 0 does not have cutting sites, another alleles are bands originates by cuts in mutant sequences.
MATERIALSECOTILLING SEQUENCES
OsYSLnB OsFER 2nC OsYSLnC OsFER1nB OsVIT
1nCOsIRO
2AOsIRO
2B TOX 1859-102-4M-4,BCF 1 1 0 0 0 0 0 FL04052-2P-3-2P-2P-M 1 1 0 0 0 0 0 BG 90-2,BCF 27,ACC 40 1 1 1 0 0 0 0 IAC 164,BCF 138,AC 189 0 0 0 0 0 0 0 ORY 1,BCF 380,AC 547 1 1 1 0 0 0 0 ORY LLANO 4,BCF 800 0 1 0 0 0 0 0 INIA YERBAL,BCF 932 0 0 0 0 0 1 0 FED 50-1,BCF 1973,FED 0 1 0 0 0 0 0 AZUCENA,BC 11,AC 15 0 0 0 0 0 1 0 Caloro ,BCF 53,TOS 658 1 0 0 0 0 0 0 O. rufipogon 1 0 0 0 1 0 1 O. glaberrima 0 2 0 1 1 2 1 IR68144-2B-2-2-3-1-120 1 0 1 0 1 0 1 IR68144-2B-2-2-3-1-166 1 0 1 0 1 0 1 IR69428-6-1-1-3-3 0 0 0 0 0 0 0 IR75862-206-2-8-3-B-B 0 1 0 0 0 3 2 IR75862-221-2-1-2-B-B 0 1 0 0 0 3 2 CT18664-9-10-2-1 0 1 0 0 0 0 0 CT18664-9-10-2-2 0 1 0 0 0 0 0 CT18667-5-18-3-3 0 1 0 0 0 0 0 CT18667-5-18-5-4 0 1 0 0 0 0 0 CT18245-18-2-4-1-3 0 1 0 0 0 0 0 CT18245-18-2-4-1-4 1 NA 0 0 0 0 0 CT18245-18-2-4-2-2 0 1 0 0 0 0 0
CT18245-18-2-4-2-7 0 1 0 0 0 0 0 CT18247-12-8-2-2-1 0 1 0 0 0 0 0 CT18247-12-8-2-2-3 0 1 0 0 0 0 0 IB51B3095-A 1 1 1 1 0 0 0 IB51B3095-B 1 1 1 1 0 0 0 Tipo Juma 1 NA 0 0 0 0 0 0 CT17334-13-7-2-1-4-5-1 1 0 1 0 0 0 0 CT17334-13-7-2-1-4-5 0 0 0 0 0 0 0 CT17334-2-1-1-2-2-3-M 1 2 1 0 0 0 0 CT17334-2-1-1-2-4-5-M 1 2 1 0 0 0 0 CT17334-2-1-1-2-5-1-M 1 0 1 0 0 0 0 CT17334-13-7-1-3-1-4-M 0 0 0 0 0 0 0 CT17334-13-7-1-5-3-11 0 0 0 0 0 0 0 CT17334-13-7-1-5-3-11 0 0 0 0 0 0 0 CT17330-M-8-5-2-1-1 0 0 0 0 0 0 0 CT17338-M-4-5-2-1-1-M 1 1 0 0 0 0 NA CT17338-M-4-5-2-2-4-M 1 1 0 0 0 0 0 CT17338-M-4-5-2-2-4-M 1 1 0 0 0 0 0 O. glumaepatula 100184 1 0 1 2 1 2 0 O. glumaepatula 105561 1,2 0 2 0 2 4 0 O. nivara (Acc. 101510) 1 0 2 0 1 0 0

48
O. nivara (Acc. 102169) 1 0 2 0 1 0 0 O.meridionalis (OR-35) 1 0 3 2 3 4 0 93-11 1 1 1 0 0 0 NA AZUCENA 0 0 0 0 0 5 0 Basmati 0 0 0 0 0 0 0 O. barthii (3239) 1 2 0 0 1 6 0
Figure 3. RiceTilling Database, based on ExcelTM files. Left, main menu with access to date, search option for interesting traits. Right, results for a specific individual, database has complete information about each plant, including images of mutants, molecular dates from TILLING, and basic information as variety, plant number, treatment for mutagenesis, etc.
References
Comai and Henikoff. TILLING: practical single-nucleotide mutation discovery. The Plant Journal. 2006. Curie and Briat. Iron transport and signaling in plants. Annual Review of Plant Biology. 2003 Gross et al. Iron homeostasis related genes in rice. Genetics and Molecular Biology. 2003 Kim et al. Localization of iron in Arabidopsis seed requires the vacuolar membrane transporter VIT1. Science. November, 2006. Koike et al. OsYSL2 is a rice metal-nicotiamine transporter that is regulated by iron and expressed in the phloem. The plant journal. 2004.
Ogo et al. The rice bHLH protein OsIRO2 is an essential regulator of the genes involved in Fe uptake under Fe-deficient conditions. The Plant Journal. 2007. World Health Organization (WHO). Assessing the iron status of populations. Geneva, Switzerland. 2004.

49
1.C.2 Identification Of Snps Markers For Biofortification In RiceContributors: O.X. Giraldo1, C. Quintero1, F. Rodríguez1, G. Plata1, C.P. Martínez2
and J. Tohme1.
1SB-2 Project; 2IP-4 Project.
FundingCIDA, CIAT CORE
BackgroundIron deficiency is probably the most widespread micronutrient insufficiency in humans. Micronutrients are effectively assimilated by humans through their daily diet; we aim to use staple food in order to increase the levels of iron intake in average diets. As rice is the main staple food for more than half of the global population we choose it as our delivery vehicle. Various studies have evaluated the iron content in different rice seeds both polished and unpolished. The results indicate that the iron content is evenly low across the studied populations. Due to this fact several research groups have attempted to increase the iron content in rice; breeding, genetic engineering, biochemical and physical methods have been applied. Our approach is based upon marker-assisted selection; we have identified and validated SNPs located in 28 of 43 iron homeostasis candidate genes reported by Quintero et al, 2006. Rice lines have being genotyped and high iron content SNPs have been tagged. We are currently using the data thus gathered in marker-assisted selection breeding programs.
Materials and methods The process we followed is illustrated in figure 1:
Plant material
Gathering rice flour (Standardization of methodology)
Fe and Zn evaluation (Atomic absorption method)
DNA isolation
SNP identification and primer design
SNP Validation (Single base extension (SBE) method)
Allele calling (Masterplex GT (Miraibio, Inc))

50
Results Gathering rice flour This methodology successfully minimizes the contamination produced by metal agents during the milling process. The rice flour thus obtained does not present contamination. This methodology was implemented by the Laboratorio de Alta Calidad de Arroz (rice high quality laboratory) at CIAT, this process is illustrated in figure 2.
Figure 2. Initially the rice is milled and polished for 18 seconds in Zatake grain mill, see (1) and (2) in the figure above. Then, 5 grams of polished rice grain (without husk and bran) are added to the Teflon chambers (3) in the figure above. A zirconium ball is then added in to the Teflon chamber, which must be closed hermetically. The samples are then collected and placed inside wooden boxes; these boxes are shaken in a uniform manner for 7 minutes until a fine rice flour is obtained (4) in figure 2. In order to evaluate the content of Iron and Zinc the rice flour thus obtained is then sent to the Laboratorio de Servicios Analiticos at CIAT (5) and (6) in figure 2. The atomic absorption method is used (2). Content of Iron and Zinc in the vegetal material Rice genotypes with a higher iron (�5.0 mg/Kg) and zinc (�9.0 mg/Kg) concentration were evaluated. The analytical technique was described by Isaac and Kerber (1971); it an atomic absorption based technique. Genotypes selected include some from CIATs germoplasm bank, one wild rice species (O.rufipogon), IRRI samples and also accession from the Latin America bank of germoplasm. Table 1 presents the eleven genotypes selected.
(1)
(6)
(3)
(1
(2)
(4)
(5)
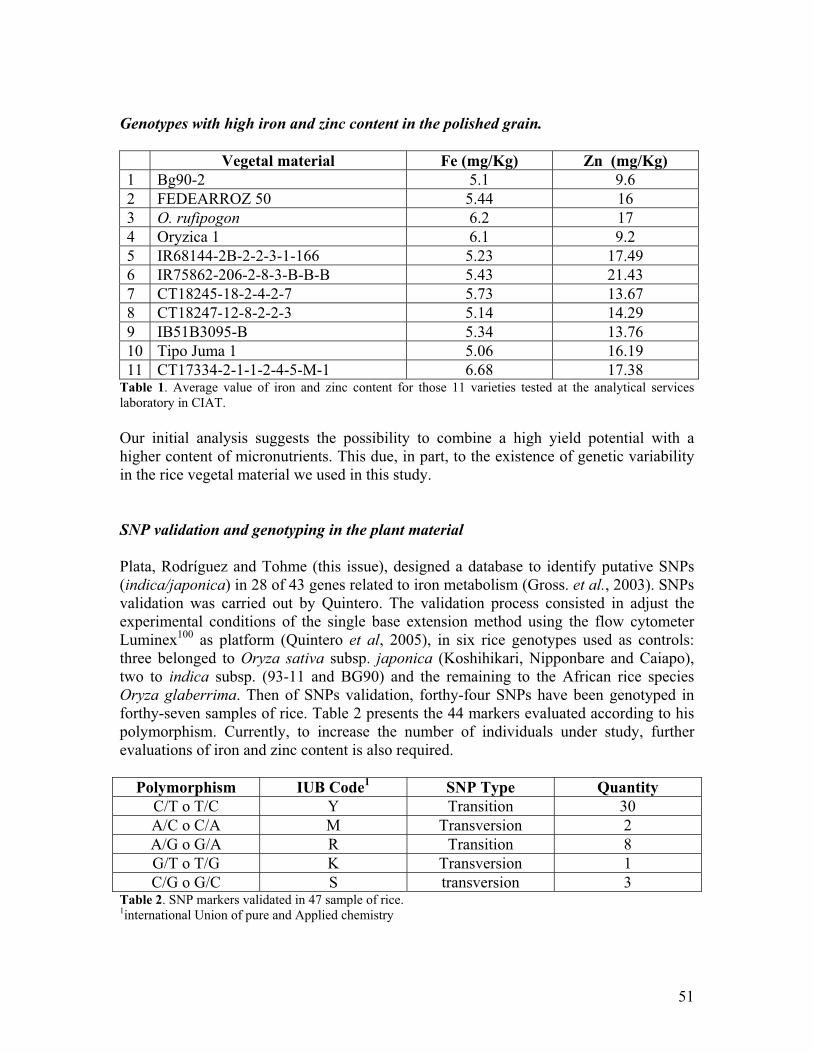
51
Genotypes with high iron and zinc content in the polished grain.
Vegetal material Fe (mg/Kg) Zn (mg/Kg) 1 Bg90-2 5.1 9.6 2 FEDEARROZ 50 5.44 16 3 O. rufipogon 6.2 17 4 Oryzica 1 6.1 9.2 5 IR68144-2B-2-2-3-1-166 5.23 17.49 6 IR75862-206-2-8-3-B-B-B 5.43 21.43 7 CT18245-18-2-4-2-7 5.73 13.67 8 CT18247-12-8-2-2-3 5.14 14.29 9 IB51B3095-B 5.34 13.76 10 Tipo Juma 1 5.06 16.19 11 CT17334-2-1-1-2-4-5-M-1 6.68 17.38
Table 1. Average value of iron and zinc content for those 11 varieties tested at the analytical services laboratory in CIAT. Our initial analysis suggests the possibility to combine a high yield potential with a higher content of micronutrients. This due, in part, to the existence of genetic variability in the rice vegetal material we used in this study. SNP validation and genotyping in the plant material Plata, Rodríguez and Tohme (this issue), designed a database to identify putative SNPs (indica/japonica) in 28 of 43 genes related to iron metabolism (Gross. et al., 2003). SNPs validation was carried out by Quintero. The validation process consisted in adjust the experimental conditions of the single base extension method using the flow cytometer Luminex100 as platform (Quintero et al, 2005), in six rice genotypes used as controls: three belonged to Oryza sativa subsp. japonica (Koshihikari, Nipponbare and Caiapo), two to indica subsp. (93-11 and BG90) and the remaining to the African rice species Oryza glaberrima. Then of SNPs validation, forthy-four SNPs have been genotyped in forthy-seven samples of rice. Table 2 presents the 44 markers evaluated according to his polymorphism. Currently, to increase the number of individuals under study, further evaluations of iron and zinc content is also required.
Polymorphism IUB Code1 SNP Type Quantity C/T o T/C Y Transition 30 A/C o C/A M Transversion 2 A/G o G/A R Transition 8 G/T o T/G K Transversion 1 C/G o G/C S transversion 3
Table 2. SNP markers validated in 47 sample of rice. 1international Union of pure and Applied chemistry

52
Future work
� To continue the validation of the remaining 57 SNPs from which PCR and SBE primers have been already designed.
� To continue the genotyping of rice lines with high and low iron content. Further
validation is also required.
� To identify SNPs markers associated with some of the genes pertaining to the YSL family. We would also like to identify other not-so-well-known genes which may play major roles in the iron and zinc content in the grain of rice.
Bibliography Gross J, R.J. Stein, A.G. Fett-Neto and J. Palma Fett. 2003. Iron homeostasis related genes in rice. Genetics and Molecular Biology, 26, 4, 477-497. Isaac, R. D and J. D. Kerber. 1971. Atomic absorption and flamephotometry: techniques and uses in soil, plant and water analysis. In: L.M Walsh (ed.) Instrumental methods for analysis of soils and plant tissue. SSA, Madison, Wis. Quintero C., F. Rodríguez, G. Plata, O.X. Giraldo, C.P. Martínez and J. Tohme. Validation of SNP markers located in rice iron homeostasis candidate genes. Annual Report 2006, SB-02 project Quintero C., E. Gaitán-Solís, C. Quigley, P. Cregan and J. Tohme. Genetic mapping and use of single nucleotide polymorphisms (SNPs) for the characterization of common bean (Phaseolus vulgaris L.) germplasm. Annual Report 2005, SB-02 project. 107-113p.

53
1.C.3 Development of introgression lines with chromosome segments substitutions from the African cultivated rice species into Oryza sativa genetic background A. G. Gutiérrez – M. F. Alvarez – A. Garavito – L. Melgarejo – S. J. Carabalí – C. P. Martinez – J. Tohme – M. Lorieux
Partners: IRD-UMR5096 Project funded by: The Generation Challenge Program IRD CIAT The French Ministry of Foreign Affairs
Background Although the methodologies for mapping of genes underlying quantitative traits have progressed considerably, development of new population structures have been poorly documented so far (Eshed and Zamir, 1995). The most used populations for QTL mapping in self pollinated crops have been recombinant inbred lines (RILs), doubled haploids (DH), backcross (BC) or F2/F3, which have several limitations such as low resolution power for QTLs with minor effects (Eshed and Zamir, 1995; Koumproglou etal., 2002). In the past twelve years, researchers have developed novel mapping populations such as introgression lines (ILs) in tomato (Eshed and Zamir, 1995), Brassicanapus (Howell et al., 1996) and Brassica oleracea (Ramsay et al., 1996), stepped aligned inbred recombinant strains (STAIRS) in Arabidopsis (Koumproglou et al., 2002), recombinant chromosome substitution lines (RCSLs) in barley (Matus et al., 2003) and chromosome segment substitution lines (CSSLs) in rice (Jena et al., 1992; Doi et al., 1997; Sobrizal et al., 1999; Kubo et al., 2002; Yu et al., 2004; Wan et al., 2004; Li et al., 2005; Ebitani et al., 2005). The CSSLs materials contain one or few contiguous chromosomal segments of the donor genotype in the background of a recurrent genotype, limiting the interactions between donor alleles to those homozygous substituted tracts, reducing the effects of interferences with the genetic background (Howell et al., 1996). These materials allow to make detailed analyses, as far as, marker assisted selection and map-based cloning (Li et al., 2005; Ebitani et al., 2005). Finally, CSSLs represent a small number of lines that can be evaluated in replicated trials and they provide a very good alternative to understand the genetic bases of reproductive barriers between species. The aim of this work is to describe the development and selection of two populations of Chromosome Segment Substitution Lines (CSSLs) between the two cultivated species of rice, Oryza sativa L. and O. glaberrima Steud. Furthermore, we present a QTLs detection analysis for yield and yield components and resistance to RSNV (Rice Stripe Necrosis Virus) in order to illustrate the advantages of using this kind of materials in genetic analysis and breeding of rice.

54
Methodology and Results Genotyping of the IR64 x TOG5681 BC2F4 & BC3F3 populations Interspecific cross: O. sativa ssp. indica cv. IR64 x O. glaberrima acc. TOG5681 A population made of BC2F4 and BC3F3 lines (Pre-CSSLs) was developed at IRD, Montpellier, France through marker-assisted backcrossing from the cross IR64 (O. sativa ssp. indica) x TOG5681 (O. glaberrima). This population is currently analyzed for its genomic content at CIAT. Twenty-seven microsatellite markers selected from Rice Universal Core Map data base and corresponding to eight chromosomes were added to the previous data set. Altogether, 128 SSRs markers have been evaluated for the two sub-populations. The search for a set of lines as candidate for CSSLs was done using program CSSL Finder v. 0.8a11 (Lorieux 2005). The following parameters were taken into account: size of introgressed segments, minimum number of segments that cover the genome and treat heterozygotes as donor homozygotes. As a result, 57 lines were selected using 107 of 128 SSRs markers that showed an even distribution across the twelve rice chromosomes. These lines covered the complete O. glaberrima genome with introgressions, except for some small regions on chromosomes 3, 4 and 9. New BC1F1 lines derived of the same cross have been developed in order to 1) validate the genetic map and 2) to recover the lost segments (see next section). A preliminary QTL analysis was carried out for various traits scored in the field at CIAT. Using the program CSSL Finder, we could identify several QTLs for tillering, panicle size and plant height. Each one of these QTLs are being compared to those obtained in the Caiapo x IRGC103544 population (see next section) and with QTLs for yield and yield components found in the literature and in the databases like Gramene (www.gramene.org). Finally, this population has been sent to WARDA, Cotonou, Benin in order to be evaluated in 2007 for response to drought stress in the field. Development of a new IR64 x TOG5681 BC1F1 population Interspecific cross: O. sativa ssp. indica cv. IR64 x O. glaberrima acc. TOG5681 As some gaps were observed in the genome coverage of the IR64 x TOG5681 BC2F4 and BC3F3 populations, we decided to develop a new BC1F1 population from the same cross. This will let us monitor the derivation of BC2 and BC3 lines that will complete the first set of lines. A total of 125 BC1F1 lines were produced and 62 of them could be successfully backcrossed to IR64 to advance to the next generation. The fertility of the BC1F1 lines was also evaluated with the scope of tagging the interspecific sterility genes. As a result, two lines showed a high fertility (> 50 %),

55
several showed intermediate fertility while the majority of the lines showed almost complete sterility. Construction of a new IR64 x TOG5681 genetic map Interspecific cross: O. sativa ssp. indica cv. IR64 x O. glaberrima acc. TOG5681
The new population derived from the cross (IR64 x TOG5681) x IR64 (125 individuals) has been evaluated for 156 anchors belonging to the Universal Core Genetic Map distributed every 15 cM along the twelve chromosomes that constitute the rice genome.
This allowed us to compute a new interspecific genetic map. The advantages of obtaining this new map are 1) to confirm and to improve the previous O. sativa x O. glaberrima interspecific genetic map (Lorieux et al., 2000), 2) to derive graphical genotypes of the lines to monitor the derivation of BC2 lines, 3) to validate the SSRs of the Core Map for their genetic location, 4) to provide information about the interspecific sterility loci 5) to allow a direct comparison of the location of the O. glaberrima introgressions to the location of wild species introgressions obtained from this project.
The data were analyzed with the MapDisto v. 1.7b126 program (Lorieux 2007). Of the 156 polymorphic SSRs markers that have been evaluated in the BC1F1, 136 mapped to the expected position based on the Nipponbare pseudomolecule, leading to a genetic map of about 1941.67 cM (Kosambi mapping function). Eight regions showing segregation distortion (SD) have been found on chromosomes 1, 2, 3, 6, 7 and 11. Of these regions, six matched with the twelve O. sativa/O. glaberrima sterility loci described so far: S30(t) (Li et al., 2005), S29(t) (Hu et al., 2004), S19 (Taguchi et al., 1999), S1 (Sano 1990), S21 (Doi et al., 1999) and S3 (Sano 1983). Two new regions with SD that have not been described for the cross were found in the long arms of chromosomes 3 and 6. As expected, the strongest SD was found to be located at the short arm of chromosome 6, corresponding to the expected position for the locus S1 Using the program CSSL Finder v.0.8a11 (Lorieux 2005), we selected 62 BC1F1 lines and a next generation (BC2F1) was carried out and completely evaluated with SSR markers. This population was used for the identification of donor lines for the development of a new library of CSSLs. The pre-CSSLs library showed a homogeneous distribution of contiguous overlapping chromosomal segments, which represented the nearly whole O. glaberrima genome in the O. sativa genetic background. However, nine fragments of O. glaberrima genome were lost in the process, but can be found in the first population developed at IRD.

56
Figure 1. Graphical genotypes of the 50 IR64 x TOG5681 BC2F1 lines selected with the help of the program CSSL Finder. The 12 chromosomes of rice are displayed vertically. The introgression (CSSL) lines are displayed horizontally. The brown rectangles indicate homozygous introgressions from O. glaberrima. The light pink rectangles indicate homozygous fragment of the recurrent genotype IR64. Nine small fragments of O. glaberrima genome were lost (gaps).
Genotyping of the Caiapo x IRGC103544 BC3F1DH population Interspecific cross: O. sativa ssp. tropical japonica cv. Caiapo x O. glaberrima acc. IRGC103544 A BC3F1 population was obtained at CIAT HQs from the cross between Caiapo (an elite tropical japonica from Brazil) and O. glaberrima (IRGC103544) (César P. Martinez). From these lines, anthers were collected and a population of 695 lines BC3DH was obtained through in vitro culture of anthers (Zaida Lentini). A subset of 312 BC3DH lines were genotyped using 199 SSRs. Sixty-four lines that cover the O. glaberrima genome were selected as candidate for CSSL development by means of the program CSSL Finder using the 125 best markers in terms of their distribution across the twelve rice chromosomes (Figure 2). The overlapping targeted chromosomal segment size was 10 cM on average. New markers from intervals between RM71-RM300 (Chr. 2) and RM185-RM241 (Chr. 4) have been evaluated to fill the gaps in these regions. A new BC1F1 population has been generated to validate the genetic map and to recover lost segments. This population will be mapped in 2007. A preliminary QTL analysis allowed to detect several QTLs for plant height, yield, tillering, 1000-grain weight and sterility located on chromosomes 1, 3, 4, 5, 6, 9 and 11. One highly significant QTL could be located on Chr. 11 for resistance to the Rice Stripe Necrosis Virus (RSNV) and fine mapping of this major QTL can be envisaged using BC4F2/F3 lines. All the analyses were performed considering 1) the ANOVA1 F-test value and 2) the graphical genotypes of the lines that showed extreme phenotypes for the trait.

57
In order to optimize and to purify the development of the CSSLs, 59 BC3DH lines of the 64 candidates were backcrossed to Caiapo and selfed to obtain 59 BC4F2 populations. These materials will be evaluated with microsatellite markers for their genetic background and for the presence of the targeted O. glaberrima segment. Also, 36 BC3DH lines were selected and backcrossed to both parental accessions, Caiapo and IRGC103544, in order to study the genetic basis of sterility in interspecific crosses.
Figure 2. Graphical genotypes of the 64 CSSLs lines selected with the help of the program CSSL Finder. The 12 chromosomes of rice are displayed vertically. They are covered by 125 evenly dispersed SSRs marker. The introgression (CSSL) lines are displayed horizontally. The black rectangles indicate homozygous introgressions from O. glaberrima. The light pink rectangles indicate homozygous fragment of the recurrent genotype Caiapo, the red rectangles indicate possible alien alleles and the gray rectangles indicate missing data.
Conclusion This work allowed us to advance significantly in the construction of CSSLs between the two cultivated species of rice, in both indica and japonica genetic backgrounds. Specifically, the results may be summarized as follows: - A new interspecific genetic maps were developed, that will allow us to complete the IR64 x TOG5681 CSSL population. - Two O. sativa x O. glaberrima populations are almost finished and ready for distribution to partners. - Four cultivated x wild BC1F1 populations were genotyped. - Four cultivated x wild BC2F1 populations were derived.

58
- The use of Universal Core Genetic Map for rice, combined with the data analysis using the CSSL Finder program, will allow us to easily compare the gene or QTL locations discovered with those two populations, to the results obtain with the new cultivated x wild CSSL populations (see the companion article “Exploring wild introgressions in rice”). - Several students and research assistants were trained. References
Doi K., Taguchi K. and Yoshimura A. (1999) RFLP mapping of S20 and S21 for F1 semi-sterility found in backcross progeny of O. sativa and O. glaberrima. Rice Genet. Newsl. 16: 65-68
Doi K., Iwata N. and Yoshimura A. (1997) The construction of chromosome substitution lines of African rice (Oryza glaberrima Steud.) in the background of Japonica rice (0ryza sativa L.). Rice Genet. Newsl. 14: 39-41
Ebitani T., Takeuchi Y., Nonoue Y., Yamamoto T., Takeuchi K. and Yano M. (2005)Construction and Evaluation of Chromosome Segment Substitution Lines Carrying Overlapping Chromosome Segments of indica Rice Cultivar “Kasalath” in a Genetic Background of japonica Elite Cultivar “Koshihikari”. Breeding Science 55: 65-73
Eshed Y. and Zamir D. (1995) An Introgression Line Population of Lycopersiconpennellii in the Cultivated Tomato Enables the Identification and Fine Mapping of Yield-Associated QTL. Genetics 141: 1147-1162
Howell P.M., Marshall D.F. and Lydiate D.J. (1996) Towards developing inter-varietal substitution lines in Brassica napus using marker-assisted selection. Genome 39:348-358 Hu F.Y., Xu P., Deng X.N., Zhou J.W., Li J. and Tao D.Y. (2004) Molecular mapping of a new pollen killer gene S29(t) on chromosome 2 in Oryza glaberrima. Rice Genet. Newsl. 21: 31-33 Jena K., Kochert G., and Khush G.S. (1992) RFLP analysis of rice (Oryza sativa L.) introgression lines. Theoretical and Applied Genetics 84: 608-616 Koumproglou R., Wilkes T.M., Townson P., Wang X.Y., Beynon J., Pooni H.S., Newbury H.J. and Kearsey M.J. (2002) STAIRS: a new genetic resource for functional genomic studies of Arabidopsis. The Plant Journal 31(3): 355–364 Kubo T., Aida Y., Nakamura K., Tsunematsu H., Doi K. and Yoshimura A. (2002) Reciprocal Chromosome Segment Substitution Series Derived from Japonica and Indica Cross of Rice (Oryza sativa L.). Breeding Science 52: 319-325

59
Li J., Xu P., Hu F., Zhou J., Deng X., Wan J. and Tao D. (2005) Mapping of QTLs for pollen stertility in Oryza sativa-O. glaberrima interspecific hybrid. 5th International Rice Genetics Symposium and 3rd International Rice Functional Genomics Symposium. IRRI P155: 131 Li J., Xiao J., Grandillo S., Jiang L., Wan Y., Deng Q., Yuan L. and McCouch S.R. (2004) QTL detection for rice grain quality traits using an interspecific backcross population derived from cultivated Asian (O. sativa L.) and African (O. glaberrima S.) rice. Genome 47: 697-704
Lorieux M. (2005) CSSL Finder, a free program for managing introgression lines. URL: http://mapdisto.free.fr/
Lorieux M. (2007) MapDisto, a free user-friendly program for computing genetic maps. Computer demonstration given at the Plant and Animal Genome XV conference, Jan 13-17 2007, San Diego, CA. URL: http://mapdisto.free.fr/
Lorieux M., Ndjiondjop M.N., Ghesquière A. (2000) A first interspecific O. sativa x O. glaberrima microsatellite-based genetic linkage map. Theoretical and Applied Genetics 100: 593-601
Matus I., Corey A., Filichkin T., Hayes P.M., Vales M.I., et al (2003) Development and Characterization of Recombinant Chromosome Substitution Lines (RCSLs) using Hordeum vulgare subsp. spontaneum as a source of donor alleles in a Hordeum vulgare subsp. vulgare background. Genome 46: 1010-1023
Ramsay L.D., Jennings D.E., Bohuon E.J.R., Arthur A.E., Lydiate D.J., Kearsey M.J. and Marshall D.F. (1996) The construction of a substitution library of recombinant backcross lines in Brassica oleracea for the precision mapping of quantitative trait loci. Genome 39: 558-567
Sano Y. (1983). A new gene controlling sterility in F1 hybrids of two cultivated rice species: Its association with photoperiod sensitivity. The Journal of Heredity 1983:74(6): 435-439
Sano Y. (1990) The genic nature of gamete eliminator in rice. Genetics 125: 183-191
Sobrizal K.I., Sanchez P.L., Doi K., Angeles E.R., Khush G.S. and Yoshimura A. (1999) Development of Oryza glumaepatula introgression lines in rice, O. sativa L. Rice Genet. Newsl. 16: 107-108
Taguchi K., Doi K. and Yoshimura A. (1999) RFLP mapping of S19, a gene for F1 pollen semi-sterility found in backcross progeny of Oryza sativa and O. glaberrima. RiceGenet. Newsl. 16: 70-71
Wan X.Y., Wan J.M., Su C.C., Wang C.M., Shen W.B., Li J.M., Wang H.L., Jiang L., Liu S.J., Chen L.M., Yasui H., and Yoshimura A. (2004) QTL detection for eating quality of cooked rice in a population of chromosome segment substitution lines. Theoretical and Applied Genetics 110:71-79

60
Yu J., Hu S., Wang J., Wong G.K., Li S., Liu B., Deng Y., Dai L., Zhou Y., et al. (2004)A draft sequence of the rice genome (Oryza sativa L. ssp. Indica). Science 296: 79-92
1.C.4 Exploring wild diversity in rice J. D. Arbelaez* – L. Moreno* – P. Rangel* – G. Djedatin* – J. Kimball – C. P. Martinez – J. Carabali – S.R. Mc Couch – J. Tohme – M. Lorieux *These authors contributed equally to the work PartnersIRD-UMR5096 Cornell University Embrapa-CNPAF Fedearroz WARDA
FundingThe Generation Challenge Program IRD CIAT Background The future of crop improvement depends on the availability of genetic variation. Most modern crop varieties have undergone a genetic bottleneck associated with the process of domestication resulting in a restriction of the genetic options that are available to plant breeders. There is a larger pool of genetic variation available in landraces and wild relatives of crops. These resources are known to contain many interesting traits for breeding, including good to strong tolerance to abiotic and biotic stresses and various nutritional traits of interest (Sun et al 2001). However, it is often difficult to utilize these natural sources of genetic diversity because of fertility barriers, linkage drag, the time and resources required to recover useful recombinants. To take advantage of the unexploited reservoir that exists in the wild relatives of cultivated rice (Oryza sativa L.), we started to develop interspecific introgression lines that will be of immediate use to breeders and will simultaneously serve to enhance our understanding of the “wild alleles” that contribute favorably to plant performance under drought stress. These lines are called Chromosome Segment Substitution Lines (CSSLs). CSSLs are particularly valuable when complex, quantitatively inherited phenotypes are the breeding target. Because they represent permanent (inbred) genetic resources that can be easily replicated by seed and distributed to collaborators working in different environments. Each set of CSSLs consists of a relatively small number of lines that can be evaluated in replicated trials. They are constructed to provide maximum power of statistical analysis because each line can be compared to all others or may simply be compared to the recurrent parent, making it possible to extract a great deal of valuable information from a relatively small number of lines crops. For phenotypes that are difficult to measure, or require repeated evaluation over years and environments, the ability to focus quickly on a small number of lines is a critical component of success (Ghesquière et al, 1997).

61
In addition to the targeted introgression of traits that can be identified phenotypically in the wild material, such as biotic or abiotic stress tolerance, it has been demonstrated that alleles hidden in low yielding, agronomically undesirable ancestors can enhance the productivity of many of the world’s most important crop varieties. These yield-enhancing alleles are the basis of ‘transgressive variation’ and may confer an advantage in both favorable (irrigated) and unfavorable conditions (drought and weed competition) (Moncada et al., 2000; Gur and Zamir, 2004). Thus, the use of wild and exotic germplasm for CSSLs construction carries with it the possibility that favorable transgressive segregants will be identified, providing the basis for studies aimed at understanding the genetic basis of transgressive variation associated with the trait of interest. Wide spread utilization of O. sativa relatives remains limited due to the fact that: (1) no extensive study has been carried out to explore the range of allelic diversity in any of the Oryza AA genome relatives, (2) the genetic basis of heterosis or transgressive variation in interspecific crosses remains largely unknown, (3) interspecific crossing barriers have hampered full utilization of rice relatives for breeding and genetic studies, (4) very few genomic resources have yet been developed to facilitate breeding efforts using O. sativa relatives. In particular, the lack of a cost effective, high throughput marker system that targets gene-based polymorphisms impedes efforts to efficiently and systematically select the best introgression lines and to evaluate the gene content of those lines in the context of comparative cereal genomics. Methodology and Results We are currently developing introgression lines from four cultivated x wild crosses, where the wild species are O. meridionalis, O. rufipogon, O. glumaepatula and O. barthii. O. sativa x O. meridionales Laura T Moreno – CIAT Interspecific cross: O. sativa ssp. tropical japonica cv. Curinga x O. meridionalis acc. OR44 We have worked at the construction of a genetic map based on unique-site microsatellite markers (SSRs) screened on a backcross population of 87 BC1F1 lines obtained from an interspecific backcross between the cultivated rice O. sativa BRSMG Curinga and its wild relative O. meridionalis accession OR44 (or W2112) from Queensland, Australia. The donor parent was the wild accession. A total of 387 SSRs were screened and 123 markers showed polymorphism between the two parents. These 123 SSRs were evaluated on the BC1F1 population. The genotyping data of this population was used for the construction of a genetic map using the MapDisto beta version 1.7 (121) software (Lorieux 2007).

62
From the development of the F1 and the BC1F1 population, we can conclude that it exists a strong hybrid inviability as a consequence of sterility barriers between the species O. sativa and O. meridionalis, though these barriers are hoped to be broken during the next backcross generations. From the screening of the SSRs, we could observed a high degree of polymorphism between these two species with 66% (81 SSR) out of 123 evaluated with an allelic variation of 10 or more base pairs. The construction of the linkage map allowed the identification of 12 linkage groups with a distance coverage of 2178 cM (Kosambi function). The colinearity of the map was good with only a few inversions that seem to be related to segregation distortion. The markers of these regions are being verified to fix possible scoring errors and data manipulation. A set of 12 seeds of each line were sown at CIAT staring December 2006. DNA was extracted and sent to Cornell University at the end of March 2007. Screening of 123 SSR conforming the set of polymorphic markers initially optimized during the BC1F1 genotyping, was done using either a 3730 xl ABI sequencer, agarose or polyacrylamide gels. A total of 3 to 7 SSR markers were screened at a 12-plant set of 42 families during this genotyping, which conformed a population of 504 individuals. All 123 SSR markers were rechecked on the parents and population in order to verify their efficiency of allele detection and visualization. The availability of a set of around 600 Miniature Inverted transposable Elements, MITEs for rice at Cornell University, allowed us to screen some of these markers on our cross at those genomic gap positions mentioned above, to improve the resolution of the genomic map. From this screening, a total of 10 MITEs were selected to form the set of polymorphic markers that will continue to be used on the next backcross generation. These MITEs markers ended up to be easy and fast to screen, and for this particular cross, a good degree and frequency of polymorphism could be found. We thus count with a set of of 133 polymorphic markers have been optimized for this cross, including the 10 MITEs markers. These additional selected markers were already screened on the BC2F1 population, in order to give a better estimate of the length of each introgression fragment. Backcrosses onto the BC2F1 lines were carried out at CIAT simultaneously with the molecular genotyping at Cornell. A set of 66 BC2F1 lines were selected based on a minimum fragment length for each chromosomal segment, and on the background selection done by our colleagues from Cornell University during the second semester of 2007. The desired fragment length could be optimized at this stage ranking from 7 to 20 Mb. Some longer fragments have been associated to low recombination regions as seen, for example, in chromosome 6 and 12. Some additional lines were selected in several regions with higher recombination frequency in order to try to avoid the possibility of losing the selected introgression fragment during the next generations. We expect to keep the maximum of minimal length fragments introgressions in order to have a final set of lines at the end of the breeding program, which could cover the whole genome with a good resolution.

63
Seeds from the set of the obtained BC3F1 families have been already checked to confirm their quality and quantity for the next sowing. Sowing of seeds will start in March 2008 in order to grow 6 plants from each line to create a population of 396 individuals. We plan to create a double haploid (DH) population through anther culture, using immature flowers of at least one DH plant from each of these 396 lines. At the same time, we will backcross the lines in order to obtain a BC4F1 population. Genotyping of the BC3DH population will be done during the fourth quarter of 2008. On the basis of the genotyping data, we will select the final set of lines that will conform the CSSL library for this interspecific cross. Additionally, a new mapping population of 111 lines from the same cross was obtained during September 2007. We will use this population verify some marker positions on the linkage map of this specific cross. The data will be available soon and will be published during this first semester. O. sativa x O. rufipogon Juan David Arbelaez – Fedearroz Interspecific cross: O. sativa ssp. tropical japonica cv. Curinga x O. rufipogon acc. IRGC105491 Rice wild relatives represent an abundant and a valuable source of new genes available for plant breeding programs. In order to explore this natural genetic variation, a systematic approach involving marker-assisted introgression, development of advance backcross populations and tissue culture techniques, have been proposed in this project to construct four new introgressions lines known as Chromosome Segment Substitution Lines (CSSL). From four F1 populations, generated between a single recurrent parent BRSMG Curinga (O. sativa) and 4 donor parents belonging from different wild rice relatives: O. rufipogon, O. meridionalis, O. barthii and O. glumaepatula, a series of backcrossed are going to be done to obtain the entire donor genome in small overlapping fragments in a number of lines with the recurrent parent genetic background, assisted by the use of molecular markers microsatellites (SSR´s). This report shows the construction of one of these populations. At the beginning of 2007 a genetic linkage map was constructed using the mapping program Map Disto v.b17 (Lorieux, 2007), a set of 128 unique-site microsatellite markers, chosen from the Universal Core Map, and a BC1F1 population of 80 plants, derived from an interspecific cross between the cultivar BRSMG Curinga (Oryza sativa) and the wild accession IRGC105491 (Oryza rufipogon), used them as a recurrent and a donor parent respectively. This map has total genetic distance of 1869 cM, very similar to that calculated using the physical distance for each SSR (using Map Disto v.b17), and to those reported for others maps between rice interspecific crosees O. sativa x O. glaberrima (Lorieux, 2000) and O.

64
sativa x O. meridionalis (Moreno, 2008). It also shows high synteny with the calculated physical map, confirming the use of these markers for fragment and background selection during the development of the CSSL library.With the genotyping work done during the genetic map construction with the BC1F1 population, fragments from the donor parent in each line were identified. Using this information and the CSSL finder program (Lorieux, 2007), 50 BC1F1 lines were selected to be backcrossed with the recurrent parent during the first semester of 2007, obtaining 50 BC2F1 families, 12 plants per family, representing a 600 plants BC2F1 population. This 50 BC1F1 chosen plants represent the entire donor genome, IRGC 105491, in small desirable overlapping chromosomal segments. These 50 BC2F1 families were genotyped at Cornell University between May and July 2007, using the previous set of SSR markers, tracking the introgressed fragment of interest for each line. 160 BC2F1 plants that represent 63 overlapping fragments with an average length of 12 Mbp, covering the entire donor genome, were chosen to be background selected. At the same time these plants were being backcrossed at CIAT to generate the BC3F1 population. During the last semester of 2007 these 160 BC2F1 plants have been genotyped at Cornell University to check each plant genetic background, to select between 70 and 90 plants that contain the fragment of interest and had recovered most of the recurrent background. Once this genotyping work is finished, the selected plants will generate a BC3F1 population, each BC3F1 family with 4 plants. That will be genotyped and selected to be taken for tissue culture, in order to generated a BC3F1 double haploid population that contain a number of overlapping homologous donor fragments covering the entire genome of IRG 105491 in a genetic background of the recurrent parent BRSMG Curinga. O. sativa x O. barthii Gustave Djedatin – WARDA Interspecific cross: O. sativa ssp. tropical japonica cv. Curinga x O. barthii acc. IRGC101937 The cultivar Curinga, used as female parent, was crossed at CIAT with the O. barthii IRGC101937 accession. The F1 plants were backcrossed to Curinga to produce the population of BC1F1 lines. A total of 80 BC1F1 seeds have been obtained and sown at two times. Among them, 64 have germinated. A total of 233 SSR markers from the Core Map have been used to survey the polymorphism between the two parents on each anchor in order to cover the whole genome through the 12 chromosomes. A set of 131 polymorphic markers well distributed on all 12 chromosomes were used to evaluate the population. They have been set up in 17 multiplex. For each multiplex, the markers have been amplified and PCR products checked on 4 % polyacrylamide or 4 % Metaphor® agarose gels. The separation of PCR fragments of the BC1F1 DNAs analysis was done on an Applied Biosystems 3730x1 capillary DNA Analyzer. The ABI results have been analyzed by GenMapper® version 3.0 program.

65
The linkage map was computed and drawn using the MapDisto program. The total length of the first draft of the map was of 2646 cM, which is significantly longer than the expected size. Most of the markers were regularly distributed along the chromosomes. The genetic distance between 2 markers varied from 14.81 cM on chromosome 7 to 28.0 cM on chromosome 4 with a mean interval length of 22.0 cM. A significant part of the map expansion observed in this population is supposed to originate from genotyping errors. We are currently double-checking the genotyping data using the “Show error candidates” option of the MapDisto program. This will allow us to correct the data in order to attain a correct map size, and to make sure that we are actually targeting the right chromosomal segments. Using the program CSSL Finder, several O. barthii introgressions could be detected in each line, while the proportion of introgressed regions varied across chromosomes and individuals. The proportion of introgressed O. barthii genome varied among the 12 chromosomes. Introgressions at several loci were observed at high frequency. A BC2F2 population was produced at CIAT and the seeds have been sent to WARDA for sowing and genotyping. The BC3F1 population are currently be produced at WARDA. The BC2F2s were genotyped, both for foreground and background, using the set of SSRs described above. O. sativa x O. glumaepatula Priscila Rangel – Embrapa-CNPAF
Interspecific cross: O. sativa ssp. tropical japonica cv. Curinga x O. glumaepatula acc. GEN1233 A genetic map was obtained for the BC1F1 population derived from the interspecific cross O. sativa (Curinga) x O. glumaepatula (GEN1233). The map was constructed with 127 SSR markers using the software MapDisto v. 1.6. The CIAT Core Map was used as a reference to establish markers orders. The genetic distances and genotyping data were used to detect wild chromosomal segments introgressed to the BC1F1 plants using the software CSSL Finder v. 0.8. A total of 60 BC1F1 plants were genotyped and one plant per targeted wild fragment was selected. All BC1F1 plants were backcrossed to Curinga and 10 seeds from each plant were sown in pots in the greenhouse. Ten BC2F1 plants, representative of each selected BC1F1 plant, were genotyped with three to four markers that covered the wild fragments. The detection of flanking markers was done using the software QGene version 2.3 for Macintosh (Nelson 1997). The BC2F1 plant that showed the wild allele for all three or four markers on each fragment was selected for the third backcross. A total of 60 BC2F1 plants were selected and are being backcrossed to Curinga to generate the BC3 population. The BC3 crosses have been made ready and the BC3F1 plants have been genotyped at Embrapa-CNPAF.

66
Conclusion CSSLs were proven as very a powerful tool for gene discovery in different crops. They are of particular value for studies involving wild progenitors as they 1) often permit to overcome interspecific sterility barriers as a large part of the cultivated species is recovered in advanced generations, 2) allow a direct comparison of the introgressed lines to the cultivated parent, permitting to display the effect of the wild progenitor on the phenotype. We hope that the development of full-genome coverage CSSL populations will contribute significantly to the set of genetic and genomic tools available for breeding and gene discovery in rice. To date, the project allowed us to obtain many important results. Among them, we may want to mention in particular the following: - Four interspecific genetic maps were developed, - Four cultivated x wild BC1F1 populations were genotyped, - Four cultivated x wild BC2F1 populations were derived, - A software (CSSL Finder) was designed for the specific purpose of helping at developing CSSL lines. - Several students and research assistants were trained, - Four students do shuttle research between their respective centers and Cornell University, - The international collaboration between several ARIs, CG centers and NARS was strengthened, - Several publications are in preparation. References
Ghesquière, A., J. Séquier, et al. (1997). "First steps towards a rational use of African rice, Oryza glaberrima, in rice breeding through a 'contig line' concept." Euphytica 96: 31-39.
Gur A and D Zamir (2004) Unused natural variation can lift yield barriers in plant breeding. Public Library of Science 2(10):1610-1615
Lorieux M, Ndjiondjop MN, Ghesquière A (2000) A first interspecific O. sativa x O. glaberrima microsatellite-based genetic linkage map. Theoretical and Applied Genetics 100: 593-601.
Lorieux M (2005) CSSL Finder, a free program for managing introgression lines. URL: http://mapdisto.free.fr/
Lorieux M (2007) MapDisto, a free user-friendly program for computing genetic maps. Computer demonstration given at the Plant and Animal Genome XV conference, Jan 13-17 2007, San Diego, CA. URL: http://mapdisto.free.fr/
Moncada M.P., C.P. Martínez, J. Tohme, E. Guimaraes, M. Chatel, J. Borrero, H. Gauch Jr. and S.R. McCouch (2001) Quantitative trait loci for yield and yield components in an O. sativa x O. rufipogon BC2F2 population evaluated in an upland environment. Theor Appl Genet 102:41–52

67
Nelson, JC (1997) QGENE: software for marker-based genomic analysis and breeding. Mol. Breed. 3:239–245
Sun, C.Q., X. K. Wang, Z. C. Li, A. Yoshimura and N. Iwata (2001) Comparison of the genetic diversity of common wild rice (O. rufipogon Griff.) and cultivated rice (O. sativa L.) using RFLP markers Theo. Appl. Genet. 102:157-162
1.C.5 Characterization of Genetic Diversity: Relationships and potential origin of the weedy rice complex in Colombia E. González
1, L. F. Fory
1, J. J. Vásquez
1, P. Ruiz
1, J. Silva
2, M. C. Duque
1, 2, and Z.
Lentini1, 2.
1 Biotecnolog y,
2Rice. Funding from GTZ, Germany and CIAT.
AbstractWe had previously reported the molceular characterization of Colombian weedy rice populations collected from commerical farmers fields using 19 SSR markers (Gonzalez etal., 2003, SB2 Annual Report 2003). This report includes the analysis of additional 7 SSR markers and 13 Oryza rufipogon accessions. The objective is to have a more complete analysis on the genetic relationships between weedy rice and cultivated rice (japonica and indica varieties); Colombian rice landraces; and wild Oryza species. This information will increase the understanding on the genetic diversity and potential origin of the weedy rice complex in this region, and potential scenarios of gene flow for the development of gene flow effect mitigation management approaches. Key Words: weedy rice, genetic diversity, molecular markers, SSR
Background Weedy rice is a worldwide common weed in rice cropping systems. It is commonly considered a de-domesticated population of the Oryza genus showing a red pericarp, seed shattering and dormancy and with high competitive ability. In the tropics, the weedy rice complex may also include other annual as Oryza species such as O. rufipogon, O. barthii, O. longistaminata, O. rufipogon, O. perennis, or O. punctata (Kwon et al. 1991). There are different hypothesis about its origin. Some hypotheses suggest that weedy rice originated from the invasion of wild species, such as O.rufipogon species from Asia in the agro ecosystem. This species has a high environmental adaptation capacity. Another alternative is that weedy rice is derived from inter-specific crossing between wild Oryza species and cultivated rice. But it is also thought that weedy rice may have endo-ferally evolved through the dedomestication of cultivated rice to weedy types, where wild rice is not present (Vaughan et al., 2003; Kane and Baack, 2007). Different reports show that weedy rice may have some characteristics of wild O. rufipogon, Oryza sativa indica or japonica variety types (Bres-Patry et al., 2001 Lentini and Espinoza, 2005) or intermediate characteristics from both groups (Cho et al., 1995; Lentini and Espinoza, 2005). Most of the rice in the United States relates to O. sativa whereas in other regions, other species from the O. nivara and O. rufipogon complex may also be involved (Vaughan et al. 2001). Recent studies on the origin of weedy rice in the USA, analyzes the main weedy types strawhull and blackhull using SSR markers and the sequence of the seudogen p-VATPase (Kane and Baack, 2007;

68
London and Schaal, 2007). The results show that weedy rice has likely evolved multiple times in such different ways as a crossing between O. sativa and O. rufipogon supporting the hypothesis that weedy rice may derives from inter-specific hybridization between these two species. Evidences also show that some weedy rice accessions appear to be derived from the crossing of O. sativa Aus varieties cultivated in India and Bangladesh suggesting they evolved mainly from escaped domesticated plants, but other accessions may derived from wild O. rufipogon supporting the hypothesis that this weed may also evolved from the wild species. We are interested in understanding the genetic diversity and potential origin of the Colombian rice complex as a model for establishing environmental safety measurements on the mitigation of gene flow effects applied to the cultivation of transgenic rice in the neo-tropics. Our previous reports show that the Colombian weedy rice populations, collected from commercial rice fields, is genetically associated with indica rice varieties. Nevertheless, few weedy rice accessions are also genetically associated with some accessions of O. nivara, O. rufipogon and a cross O. nivara/ O. rufipogon. None of the accessions analyzed so far are japonica type, in contrast to the Colombian landraces that all appear to be of japonica type origin. This report shows the complete analysis with 26 microsatellites markers showing high polymorphism in the weedy rice population type and the comparison of this population with a large collection of indica and japonica commercial varieties, Colombian land races and Oryza species
Materials and Methods
Plant Material and Genetic Analysis using Microsatellites Markers: 148 weedy rice accessions collected in Colombia were used. Controls consisted of: 19 O. sativa indica rice commercial varieties and 12 japonica type; 16 hand-made manual crosses between the RHBV-resistant transgenic Cica 8 line and non-transgenic variety purple or selected weedy rice accessions; 20 accessions of wild Oryza species AA genome, and 15 Colombian landraces. Twenty-six SSR primers derived from rice were amplified in all samples. The PCR products were resolved on silver-stained poly-acrylamide gels and microsatellites alleles band sizes were determined by comparison to 10 bp molecular weight standards (Promega).
Multiple correspondence analyses (MCA). The MCA includes the molecular markers data using 26 SSR. The individuals analyzed were first clustered in 13 groups based on its genetic relatedness, and then identified in the three major groups characterized by presence-absence of seed awn and seed main color (straw or brown-black). Structure. This program implements a model-based clustering method for inferring population structure using genotypes data consisting of unlinked marker (Pritchard, and Donnelly (2000). The number of subpopulation (K) was set from 2 to 8, and each was run three times. Each run started with 10.000 burn-ins followed by 50.000 interactions. When K was set at 5. The ACM groups were assigned to classify the individuals at 10.000 length Burn and MCMC 50.000 and K = 2 to 5.

69
Results and Discussion Total 162 alleles (an average of 6 alleles per locus) were detected using the selected 26 SSRs. The allelic size ranged from 96 to 268 bp. The RM251, RM222 and RM241 showed 10 alleles whereas RM349 detected 3 alleles. All SSRs were polymorphic and ranked from a PIC (polymorphic information contained) of 0.75 for the RM 212 and RM 252, whereas the RM 220 and RM 234 showed the lowest PIC that is 0.274 and 0.30 respectively. The heterozygosis per each marker oscillated from 0.00 for RM 234 and 0.081 for RM246 and RM 11, suggesting a predominant homozygosis in the population. Genetic diversity was 0.545. Fis oscillated from 0.831 and 0.972. The molecular variance suggests a variation of about 13% within the weedy rice population.
Three main clusters are identified when the seed awn presence and color is overlapped with the genetic clusters derived from the ACM analysis (Figure 1). The first group (variety type) contains 30% of the weedy rice accessions of which 68% are awn less straw seeds similar to rice varieties. The largest group (64% of the samples) contains 57% accessions with straw-awn seeds (intermediate type). The smallest group (6% of accessions) is characterized by a predominance (86%) black-brown straw seeds which also includes accessions of the wild species O. rufipogon (O. rufipogon type )(Figure 1). Additional analysis using the structure program also classifies the population in into these three distinct groups (K=3 or K=2), showing an association of awn less accessions with commercial varieties and black awn accessions with O. rufipogon species. Our work confirms that gene flow and introgression occurs freely in the crop-weedy rice complex in farmer’s fields. Results from last year using nuclear and chloroplast sequences also indicate that the weedy rice complex in Colombia is AA genome and related to indica rice type, and some individuals are genetic related to few accession of O. rufipogon and O. nivara. Similar results were reported recently for the weedy rice complex in the USA (London and Schaal, 2007). These Asian wild species may had been introduced in the Americas via seed contamination. Thus, part of the variation found in weedy rice maybe the product of continuous crossing in the crop/ weedy rice/ O. rufipogon complex in addition to a de-domestication process from O.sativa. Current work includes a comprehensive analysis with a broad range of AA genome species and the Latin American Oryza species aimed to elucidate the possible origin of weedy rice in Colombia.

70
References
Bres-Patry, C., Lorieux, M., Clément, G., Bangratz, M. and Ghesquière, A. 2001. Heredity and genetic mapping of domestication-related traits in a temperate japonica weedy rice. Theor. Appl. Genet. 102: 118-126.
Cho, Y-C., T-Y, Chung and H.-S. Suh. 1995. Genetics characteristics of Korean weedy rice (Oryza sativa L.)by RFLP analysis. Euphitica 86: 103-110.
Lentini Z and A.M. Espinoza. 2005. Coexistence of Weedy Rice and Rice in Tropical America: Gene Flow and Genetic Diversity. Chapter 19. p: 303-319. In: J. Gressel (Ed.). “Crop Ferality and Volunteerism: A Threat to Food Security in the Transgenic Era?”. CRC Press. Boca Raton, FL
London, J and Shaal. (2007). Origins and population genetics of weedy rice in the USA. Molecular Ecology 16:4523-4535.
McCouch, S. R., G. Kochert, Z. Y. Wang G. S., Khush, W. R. Coffman & S. D. Tanksley. 1988. Molecular mapping of rice chromosomes. Theoretical Applied of Genetics 76: 815-829.
Ruiz, P. 2003. Morphological, phenological and genetic characterization of weedy rice collected in rice farmers fields of Colombia. Part II. Fields planted with Fedearroz 50, Coprosem and Cimarrón rice commercial varieties. B.Sc Thesis. Universidad Javeriana, Bogota. 186 p
SAS, I.I. 1989. SAS/STAT. User Guide, Version 6, Fourth edition, Volumen 2. Cary, N.C: S.A.S. Institute Inc. p 864.
Vasquez, J.J. 2002. Morphological, phenological and genetic characterization of weedy rice collected in rice farmers fields of Colombia. Part I: Field planted with Oryzica 1 rice commercial variety. B.Sc Thesis. Universidad de Los Andes, Bogota. 210 p
Vaughan L.K., Ottis B.V., Prazak-Havey A.M., Bormans C.A., Sneller C., Chandler J.M and Park W.D. 2001. Is all red rice found in commercial rice rally Oryza sativa?. Weed Science 49: 468-476.
Vaughan, D., Morishima, H. and Kadowaki, K. 2003. Diversity in the Oryza genus. Current Opinion in Plant Biology. 6: 139-146.
Figure 1.- Multiple Correspondence Analysis (SSR) weedy rice in Colombia

71
Tracking gene flow at landscape. Dissemination of IMI-herbicide resistance-gene from rice into weedy rice.A. Velazquez
1, L. Fory
1, M. Quintero
2, T. Agrono
2, C. Ordoñez
2, E. Bolaños, P. Solarte.
E. Corredor3 M. Duque1. J. Silva
1 and Z. Lentini
1, 2. 1
SB2,2IP4, 3FLAR. Funding from
GTZ, Germany and CIAT
Abstract
Here we report a methodology at large scale to track hybridization and introgression of Clearfield CF205® herbicide-resistant gene into weedy rice in commercial rice fields. Crop-weedy rice hybrids are first identified using a lethal herbicide dose of imazapic. Putative herbicide resistant plants under field conditions are subsequently analyze at the molecular level to detect the single nucleotide ALS-gene mutation associated with the imazapic resistance using a SNP marker. The methodology described in this work eases its implementation at large scale in commercial fields, and it could be useful for understanding gene flow impact at regional level. Key Words: weedy rice, herbicide tolerance, IMI, Clearfield CF205®, ALS
Background In Colombia Fedearroz used the Clearfield system created by BASF and the Louisiana State University via mutagenesis to develop Clearfield CF205® variety tolerant to the imazapic Masterkey DG® herbicide system. In this work, we are monitoring the transfer and introgression of this CF205® non-transgenic herbicide resistance source as a model to assess the gene flow impact from rice into weedy rice. Total 1,113 accessions of weedy rice and individual CF205® plants were collected from commercial Clearfield® fields in Tolima and the Valle del Cauca. The sampled plots have one to three cycles of CF205® variety with or without rotation with other crop. Last year, we reported the standardization for IMI-tolerance evaluation in the field and laboratory conditions using the Masterkey DG® system recommended by BASF. In addition, the CF205® ALS gene was sequenced and specific primers were designed to detect the point mutation (SNP) conferring the herbicide resistance in this variety. We also showed the use of tilling as an alternative method to trace gene flow from rice into weedy rice at large scale. This year we report the evaluation of about 50,000 plants for herbicide tolerance in the field and for the ALS mutation confirming the presence of the CF205® ALS gene. This work is part of the 2nd phase of a project entitled “Gene Flow Analysis for Environmental Safety In the Tropics”, which main goal is to generate baseline genetic information for the development of guidelines on the safe introduction and use of novel agriculture traits (biotechnology derived or not native from the place of introduction) in the Tropics.

72
Materials and Methods Plant Material. Original plant materials consisted of 959 accessions of weedy rice and 188 accessions Clearfield CF205 ® variety collected from commercial rice field in Tolima and Valle del Cauca. Controls consisted of six weedy biotypes collected in Tolima in 2001, four commercial indica type rice varieties (Fedearroz-50, Oryzica I, Cimarrón and Coprosem II), four accessions of wild Oryza species (O.rufipogon IRGC-105491 Malasia, O.rufipogon IRGC-100916 China, O. barthii IRGC-104119 Chad, O.glumaepatula Costa Rica) and the cultivated rice O. glaberrima IRGC-103544.
Field evaluation for resistance to Imazapic herbicide. The first generation of self-progeny seeds (959 accessions) derived from the original weedy rice accessions collected in Tolima and Valle del Cauca were tested for herbicide tolerance. Fifty (50) seeds per each accession were sown per row of 10 cm between rows, in beds of 14 m long by 1.2 m wide. Herbicide treatment consisted of one application of imazapic at pre-emergence (PRE) followed by one at post-emergence (POST) 15 days after seed sown (DAS) in the case of the weedy rice population from Valle del Cauca, whereas it was necessary to apply one PRE and two POS applications at 15 and 35 DAS for the Tolima population. Imazapic was applied using a concentration of 115g/ha according to the manufacturer recommendation (BASF, 2003). Herbicide was diluted in water, and a nonionic surfactant was added to the solution at 0.5 %. The herbicide was applied by aspersion with a backpack sprayer A-Z at a height of 1.20 m. Three rows per bed of Clearfield CF205 ® (resistant control) and the susceptible rice commercial varieties (Coprosem 2, Cica 8, Fedearroz 50 and Yacú 9) were included. The number of alive and dead plants was recorded at 30 days, and 45 days after the herbicide final application. Plant toxicity was evaluated at 15 and 25 DAA, following the ordinal herbicide susceptibility scale established by Finol et al. (1999). Score 1 refers to zero damage, plants similar to non-herbicide application control. Score 3, refers to moderate damage, characterized by general plant chlorosis; and score 5 refer to severe damage indicated stunted plants or death. The methodology of evaluation was optimized to avoid potential escapes (Fory et al., 2006, Velásquez, 2008). Detection of Clearfield CF 205®-weedy rice hybrids. Herbicide tolerant plants were analyzed in bulk for the presence of the ALS gene mutation. DNA was extracted from bulks of seven plants. The ALS gene was sequenced (Fory and Lentini, 2006 SB2 Annual Report 2006) and 26 gene specific primers were designed to detect the ALS single point mutation (single nucleotide polymorphism, SNP) associated with the herbicide resistance using the SNAPER program (Drenkar et al. 2000). Two primers were selected from the 26 originally designed and used to trace this gene in the weedy populations at landscape level using molecular bulk analysis. The PCR was performed as follows: one cycle at 94°C for 2 min, 27 cycles following the sequence of 94°C for 15s; 60°C for 15 s 72°C for 15 s; and then 72°C for 5 min. The PCR reaction was carried out at a final volume of 25 �L (DNA 20 ng, PCR 1 X buffer MgCl2 1.8 mM, dNTP’s 0.25 mM, concentration of

73
primers forward and reverse of 0.1 �M, and 0.1 �L Taq polymerase). Results and discussion Progeny (49,883 plants) derived from the 959 weedy rice accessions collected from the commercial CF205® rice fields were first assayed for herbicide resistance (Figure 1). DNA of herbicide resistant plants was bulked, and then analyzed for the presence of the ALS resistant gene from CF205®. Resistant bulk samples were subsequently analyzed to identify the number of individual plant(s) per bulk containing the ALS resistant gene. In the case of Valle del Cauca population which comes from fields with just 1 (one) commercial cycle of CF205®, 9% (17.566 plants) of the samples showed tolerance to imazapic, but of these herbicide tolerant plants only 0.4 % also contained the ALS gene from Clearfield CF 205®. In contrast to Valle del Cauca, the Tolima population was collected from fields with 3 cycles of CF205®. In this case 23.1% (32.317 plants) of the plants show tolerance after three applications of lethal herbicide concentration. The molecular analysis of these plants indicates that 2.1 % (3 fold higher than Valle del Cauca) contained the CF205® ALS gene. The herbicide tolerance in those plants that do not contained the CF205® ALS gene mutation suggest that in these cases tolerance may be conferred by other mutation(s) in the ALS gene due to exposure of other herbicide sharing the same mode of action as the IMI. This phenomenon is commonly known as cross-resistance. Cross herbicide resistance occurs when plants acquire resistance to more than one herbicide because they share the same mode of action, therefore resistance evolves from different point mutations in the same gene (Webster and Masson 2001, Usui 2001). Cross-herbicide resistance has been already demonstrated in other Clearfield rice varieties planted in the USA(Tan et al., 2005) that were derived from the same original mutant used to develop CF205® . Our results also show that resistant weedy rice are established in few generations. This means that the technology Clearfield will not last too long in the market unless stringent weedy management measurements are applied with the technology including crop / variety and herbicide rotation, cultural and chemical control, and most important the use of the certified seed free of weedy rice. The use of IMI herbicide resistance as a model may allow developing crop management approaches applicable to the efficient control of weedy rice and safe use of transgenic crops in tropical environments.
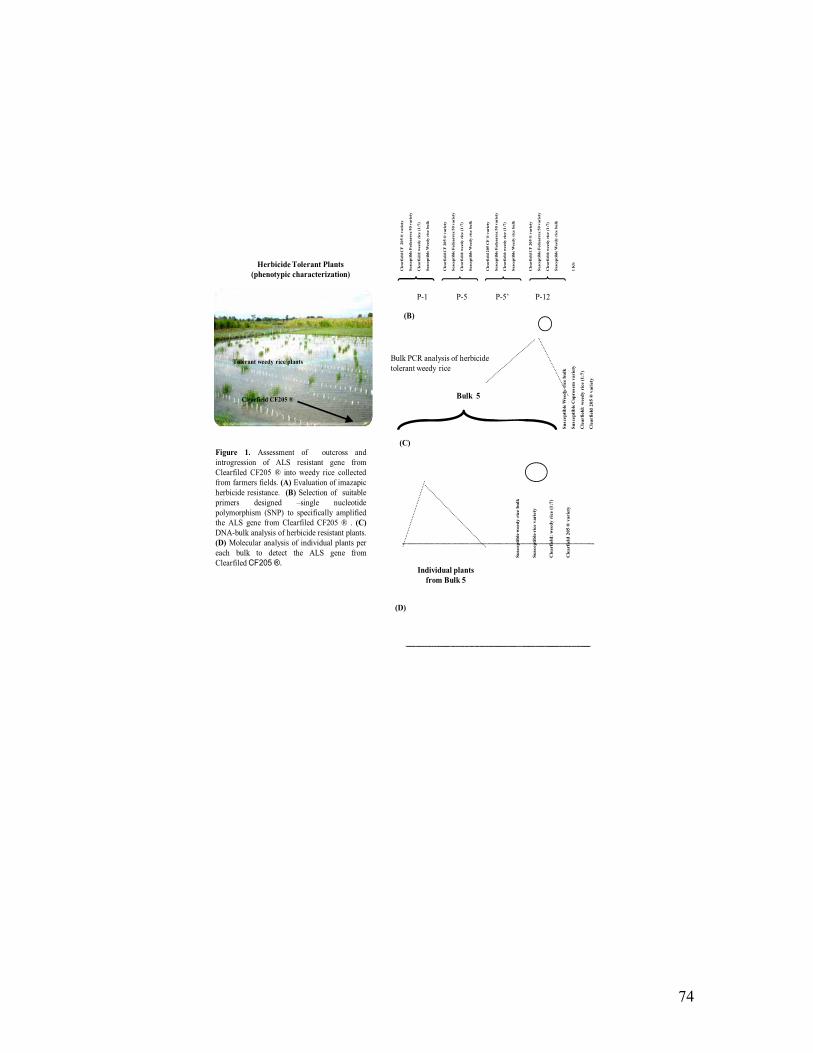
74
Cle
arfi
eld
CF
205
® v
arie
ty
Susc
epti
ble
Fede
arro
z 50
var
iety
Cle
arfi
eld:
wee
dy r
ice
(1:7
)
Susc
epti
ble
Wee
dy r
ice
bulk
Cle
arfi
eld
CF
205
® v
arie
ty
Susc
epti
ble
Fede
arro
z 50
var
iety
Cle
arfi
eld:
wee
dy r
ice
(1:7
)
Susc
epti
ble
Wee
dy r
ice
bulk
Cle
arfi
eld
205
CF
® v
arie
ty
Susc
epti
ble
Fede
arro
z 50
var
iety
Cle
arfi
eld:
wee
dy r
ice
(1:7
)
Susc
epti
ble
Wee
dy r
ice
bulk
Cle
arfi
eld
CF
205
® v
arie
ty
Susc
epti
ble
Fede
arro
z 50
var
iety
Cle
arfi
eld:
wee
dy r
ice
(1:7
)
Susc
epti
ble
Wee
dy r
ice
bulk
1 K
b
P-1 P-5 P-5’ P-12
Figure 1. Assessment of outcross andintrogression of ALS resistant gene fromClearfiled CF205 ® into weedy rice collectedfrom farmers fields. (A) Evaluation of imazapicherbicide resistance. (B) Selection of suitableprimers designed –single nucleotidepolymorphism (SNP) to specifically amplifiedthe ALS gene from Clearfiled CF205 ® . (C)DNA-bulk analysis of herbicide resistant plants.(D) Molecular analysis of individual plants pereach bulk to detect the ALS gene fromClearfiled CF205 ®.
Bulk PCR analysis of herbicide tolerant weedy rice
Bulk 5
Individual plants from Bulk 5
Susc
eptib
le W
eedy
ric
e bu
lk
Susc
eptib
le C
opro
sem
var
iety
Cle
arfie
ld: w
eedy
ric
e (1
:7)
Cle
arfie
ld 2
05 ®
var
iety
Susc
eptib
le w
eedy
ric
e bu
lk
Susc
eptib
le ri
ce v
arie
ty
Cle
arfie
ld: w
eedy
ric
e (1
:7)
Cle
arfie
ld 2
05 ®
var
iety
Herbicide Tolerant Plants(phenotypic characterization)
Tolerant weedy rice plants
Clearfield CF205 ®
(D)
(C)
(B)

75
References BASF. 2003. Folleto técnico del sistema Clearfield CF 205 ®. Sistema de Producción de arroz que
optimiza sus recursos. Pp30 Drenkar, E., Richter, B.G., Rozen, S., Stutius, L. M., Angell, N.A., Mindrinos, M., Cho, R. J., Oefner, P. J.,
Davis, R. W. and Ausubel, F. M. 2000. Nucleotide Polymorphisms Facilitates Map-Based Cloning in Arabidopsis. Plant Physiology 124: 1483–1492.
Fory L, Corredor E, Agrono T, Gonzalez E, Ordoñez C, Duque M. and Z. Lentini. 2005. Scaling up
analysis gene flow analysis from rice into weedy rice at landscape under farmers’ commercial conditions. SB-02 Project. International Center for Tropical Agriculture.
Fory, L and Lentini, L. 2006, Scaling up analysis gene flow analysis from rice into weedy rice at landscape under farmers’ commercial conditions.Part II. Molecular detection of IMI herbicide resistance gene in the Clearfield CF205 ® variety. SB-02 Project. International Center for Tropical Agriculture
Lentini Z. and Espinoza A. 2005. Coexistence of weedy rice in tropical America-gene flow and genetic diversity. Edited by Jonathan Gressel, USA. In : Crop ferality and volunteerism. Chapter 19: 306-307.
Tan S, Evans R, Dahmer M, Singh B. and Shaner D. 2005. Imidazolinone-tolerant crops: history, current status and future. Pest Manag. Sci. 61: 246-57.
Tranel, P and Wright, T. 2002. Resistance of weeds to ALS-hinhibiting herbicide: what have we learned? Weed Sci: 50 700-712.
Usui, K. 2001. Metabolism and selectivity of rice herbicides in plants. Weed Biology and Mangement. 1: 137-146.
Webster , E. P. and Masson, J. A. 2001. Acetolactate sinthase-inhibiting herbicides on imidazolinone-tolerant rice. Weed Technology 16:627-630.
Velásquez, 2008 Evaluación del flujo de genes desde una variedad de arroz (oryza sativa l.) resistente al herbicida imazapic hacia arroz rojo (oryza sativa f. spontanea) proveniente de los departamentos de tolima y valle del cauca. Tesis Unviersidad Javeriana p 171.
Zhang, W., Linscombe, S., Webster, E., Tan, S. and Oard, T. 2006. Risk assessment of the transfer of imazethapir herbicide tolerance from Clearfield rice to red rice (Oryza sativa). Euphytica 152:75-86.
Morphological and phenological characterization of wild Oryza speciesfrom Venezuela A. Eloy
1, L. Fory
1, C. Ordoñez
2, K. Arcia
1, E. González
1, T. Agrono
2, and I. Perez3, A.
Ortiz4, Z. Lentini1, 2
. 1
SB2,2IP4, 3 INIA 4Universidad Central de Venezuela. Funding
from GTZ, Germany.
Abstract
This work reports the morphological and phenological characterization of 154 accessions of wild Oryza species collected from Guarico and Portuguesa States in Venezuela. The materials analyzed under greenhouse conditions and compared with a set of accessions used as reference including Latin America wild Oryza species obtained from IRRI germplasm bank. This work is the first systematic analysis of wild Oryza species from diverse environments (natural and agricultural) in Venezuela.

76
Key Words: wild Oryza species, morphological characterization, weedy rice, genetic diversity Background The genus Oryza is composed of two cultivated species and 21 wild relatives (Vaughan et al., 2003). The cultivated species are Oryza sativa and Oryza glaberrima of Asian and African origin, respectively. In Central and South America, four wild Oryza species have been reported: three tetraploids of CCDD genome (O. alta, O. grandiglumis and O. latifolia) (Vaughan, 1994), and one diploid O. glumaepatula, with AA genome sometimes referred as O. rufipogon americana (Vaughan, 1994; Lentini and Espinoza, 2005). The study and collection of Oryza wild rice species in Central and South America began with Oka (1961). Thirty one years later Morishima and co-workers (NIAS, Japan) re-initiated collections in Brazil They studied 103 sites from Manaus to upstream Barcelos including the Solimoes river during 1992 and 1993 (Morishima and Martins 1994). Subsequent expeditions continue until 1999. (Morishima et al. 1999; Akimoto, 1999). Studies on composition, characterization and distribution of wild species are fundamental for the development of appropriate in situ conservation strategies. In the Americas, Brazil and Costa Rica are the only countries that had conducted throughout analysis of their wild Oryza species populations. In the case of Venezuela, so far it is reported 64 herbarium specimens of wild Oryza in the National Herbarium (Vaughan and Tomooka, 1999). In Venezuela, some well preserve natural Oryza niche environments are in vicinity to agricultural areas. These scenarios facilitate the analysis of the wild species-cultivated continuum and of potential hybrid swarms. This work is part of the 2nd phase of a project entitled “Gene Flow Analysis for Environmental Safety In the Tropics”, which main goal is to generate baseline genetic information for the development of guidelines on the safe introduction and use of novel agriculture traits (biotechnology derived or not native from the place of introduction) in the Tropics. This analysis serves as first step for an in depth study on gene flow-introgession dynamics between rice and compatible wild species in this region.. Materials and Methods
Collection of wild samples in the field. Accessions (seeds per individual plants) of wild Oryza species were collected from natural and agricultural environments, and ditches along roadsides in the States of Guarico (196 accessions) and Portuguesa (127 accessions), the two main rice cropping areas in the country. Each collecting site was registered by GPS. Seed germination and seedling growth. De-husked seeds from each plant collected in the field were germinated in vitro and then transplanted under greenhouse conditions during two consecutive generations. Plants were grown to maturity. The morphological characterization was conducted using the main qualitative, quantitative and phenology descriptors to distinguish Oryza species. Accessions of wild Oryza species obtained from the IRRI Germplasm Banck including O. rufipogon (22), O. glumaepatula (8), O. alta,

77
(4), O. grandiglumis (6) and O. latifolia (20), weedy rice accessions (15), and commercial rice varieties indica (7) were used as control.
Results and Discussion Seeds (5,373) were placed for germination in vitro. Germination increased when seeds were dehusked. In general, the accessions from Camaguan natural marsh showed the lowest germination rate (14% first generation harvested from field plants, and 53% second generation harvested from greenhouse grown plants) than the rest of the samples collected (about 80% germination).
Populations of putative O. latifolia (tetraploids, CCDD genome) were collected in natural environments and disturbed areas (farmland and irrigation canals) in two rice farms (Clavellinas and El Esfuerzo) located in the Portuguesa State. It appears that this species is plastic with a broad environmental adaptation. In the case of Clavellinas, this used to be a traditional maize farm with the cultivation of the first cycle of certified rice seeds (variety Fedearroz 50) at the moment of the plant collection. Some individual plants of O.latifolia were located in the middle of the rice field. These plants appeared to come from a natural population (about 200 individuals) located nearby the rice field in the margins of the Morrocoy stream, which derives from the Guanare River. The other population of O. latifolia was collected in the irrigation canal at “El Esfuerzo” farm. The taxonomic morphological descriptors for these accessions agree with those of the IRRI accessions used as reference. All plants have a typical truncated ligule. However, a clear distinction was noted between the Clavellinas and el Esfuerzo populations. The plants from the former were homogenous; intermediate plant habit, some plants are as high as 3 meters, broad flag leaves with descendent angle, open panicles with small spiklets (5.3 mm), and small black hulled seeds with short straw awns. Spiklets < 7mm are used to distinguish O. latifolia from O. alta and O. grandiglumis in Venezuela (Vaughan, 2003). In the case of “El Esfuerzo” plants have larger spiklets of about 5.9 mm, and glabrous leaves in contrast to Clavellinas which has pubescent leaves. In addition, two clearly distinct biotypes are noted in El Esfuerzo population, one type is characterized by short plants (about 1 m height) with narrow leaves and small panicles; and the other type is distinguished by tall plants (up to 2 m height), broad leaves and large-open panicles (Figure 1). Similar plant differentiation was also reported for populations of O. latifolia in Argentina (Vaughan, 2003).
The diploid (AA genome) populations were collected from four locations, three in the Guarico State (Estero de Camaguán, Via Herrera, Lecherito and Km 133), and two in the Portuguesa State (El Esfuerzo and Santa Lucia)(Figure 1). Estero de Camaguán is a natural marsh that is dry during the dry season and the water table can rise up to 3 meters or more during rainy season. Via Herrera, Km 133, and Santa Lucia accessions were collected from damped road ditches, whereas accessions form Lecherito were taken from an abandoned field nearby a commercial rice field.
The Estero de Camaguan population was collected using a boat, and plants were collected from shallow to deep-water areas separated from more than 5 meters apart. Plants are floating forming small colonies (plant-islands), and clonal propagation via rhizomes is predominant. Other species such as Ceraptoteris thalitroides, Echinornia crassipes,

78
Hymenanne amplexicaule, Caperonia palustres, Ipomeae sp, Dalechampia tilifolia are also floating in this zone. This wild Oryza population showed late and asynchrony flowering. Plant height ranges from 50 cm to 130 cm height, plants showed acuminate ligules and narrow leaves, plant type is open, with coriaceous and glabrous or low pubescent leaves, purple auricles, about 50% show red seed awns, purple stems or with purple stripes. Two plant types are distinguished (short and tall plants) collected from shallow and deep-water areas respectively. The tall type, flowers earlier, has open panicles, whereas the short type is late flowering with compact panicles.
The other AA genome populations collected are clearly morphologically distinct from that of Estero de Camaguan. Plants from Estero de Camaguan have longer anthers (2,9 mm ) and spiklets (7.6 mm) than those from the location nearby via Herrera (1.8 mm and 7.1 mm, respectively), and the flag leaf has 90° angle; whereas those from via Herrera have obtuse-angle flag leaf, have green auricle and green or purple stripes stems. Two biotypes are distinguished in this population, plants with straw or red seed-awns.
The diploid populations from Lecherito and El Esfuerzo are the most alike although there are some morphological differences. These two populations are characterized for showing intermediate plant habit, acuminate ligule, most panicles with red long awns (some with straw awns) in Lecherito and with anthers and spikelets of 2,1 mm and 7,1mm, whereas in El Esfuerzo all panicles are red awns and longer spikelets of 7, 8 mm long. The main difference between these two populations is that in the case of the Esfuerzo, panicles are widely open, the flag leaf angle is obtuse, the leaves are highly pubescent and some individuals showed green purple auricles. In contrast, in the Lecherito population plants showed compact panicles, the flag leaf angle is 90°, leaves and all individuals have green auricles.
A clearly distinct AA population from the others is the one collected in Santa Lucia. All the plants have straw awn, stems with purple stripes and green auricles. The anthers and the spikelets are the smallest of all the diploid wild populations collected (1,6 mm and 6,1 mm). Plants have acuminate ligules, the plant habit is intermediate, the panicle is open and the flag leaf angle is obtuse. Although the individuals of this population are not as high as the tetraploids, some individuals can reach up to 1.30 m at flowering.
In Lecherito, in addition to the AA wild Oryza that was collected in an abandoned field, several plants of putative weedy rice with long red awns were intermingled within a Fedearroz 50 commercial field. Some of the weedy rice accessions collected is segregating for the red awn color and others look alike the variety type. These accessions were labeled tentatively as putative hybrids (to be confirmed by molecular markers), since morphologically they appear to be derived from a potential cross between the red awn wild Oryza sp and cultivated rice. The hull coloration of these weedy rice are straw, straw with brown stripes, or solid black-brown likewise wild Oryza species. These putative hybrids have the largest anthers (2,1 mm ) and spikelets (8,1 mm) likewise cultivated rice.

79
Flowering period and life cycle are two important parameters for the characterization of wild Oryza species. Cultivated Oryza are annual plants that has synchronous flowering within 3 to 4 months after planting, produce mature grains and senesce. In contrast, wild Oryza is characterized by asynchronous flowering, the same plant may flower more than once and different individuals may flower at different times, and plants may be perennials (evergreen) or have extended life cycle more than 8 months. All the tetraploid populations collected are perennials, have continues cycles of flowering that start as early as 78 to 130 days after planting. According to the IRRI reports, O. latifolia flowers all year long. The O. latifolia accessions (IRRI) used as reference were also early flowering (72 to 90 days) whereas the O. alta (IRRI) flower much later (125-157 days). Some O. grandiglumis accessions never flowered or showed a little flowering. O. grandiglumis tends to flower from March to July under natural conditions whereas O. alta also flowers from September to October (Vaughan, 1994). The “Clavellinas and “Esfuerzo” tetraploids populations are perennials. Plants had already produced several cycles of seeds, and new tillers emerged profusely even during the seed filling process.
The population from Estero de Camaguan is also perennial with asynchronous flowering. Some plants start flowering as early as 80 after planting but other about 170 days. This population looks alike the Venezuelan O glumaepatula accession obtained from IRRI.Similarly, the population from Santa Lucia is also perennial but in this case, the population shows a synchronic two distinct flowering periods during the evaluation time (less than a year). In the case of the diploid populations from Via Herrera, Lecherito and El Esfuerzo although they showed various flowering cycles they are not profuse, and most plants show weak re-growth (few new tillers) and clear indications of senescence during the grain filling period. However, they are more vigorous and have a more extended life cycle compared to cultivated rice. These population appeared to have a life cycle intermediate between cultivated and perennials wild Oryza. All the putative hybrids show life cycle similar to cultivated rice (annual, one flowering cycle, not spontaneous new tillers formation after flowering, and senescence during grain filling). These morphological characterization will be complemented with molecular analysis using specific markers to dissect the taxonomy of plant species collected.

80
Reference
Lentini Z. and Espinoza A. 2005. Coexistence of weedy rice in tropical America-gene flow and genetic diversity. Edited by Jonathan Gressel, USA. In : Crop ferality and volunteerism. Chapter 19: 306-307
Vaughan D A. 1994. Wild relatives of rice: Genetic resources handbook. Int. Rice Res. Inst., Manila, Philippines, p.137.
Vaughan D. and Tomoka, 1999. Wild rice in Venezuela D. Research Notes. I. Varietal Differentiation and Evolution. National Institute of Agrobiological Resources, 2-1-2 Kannondai, Tsukuba, 305-8602 Japan.
Vaughan D. 2003. Gene pool of genus Oryza. In Monograph on genus Oryza. Nanda. S.D. Sharma S. D., eds. P 113-138.

81
Molecular characterization of rice wild species from Venezuela K. Arcia
1, A. Eloy
1, L. Fory
1, E. González
1, C. Ordoñez
2, P. González
1, A. Velásquez,
T. Agrono2, and I. Perez3, A. Ortiz4, Z. Lentini
1, 2. 1
SB2,2IP4, 3 INIA 4Universidad
Central de Venezuela. Funding from GTZ, Germany Abstract
Here we report the molecular characterization of the 154 accessions of wild Oryzaspecies collected in Guarico and Portuguesa using chloroplast markers (ORF100, TrnL-Trn and TrnS-TrnT regions), one nuclear gene (ADH) and SSRs markers. The morphological characterization of these accessions is described in the Section 1.1.13 of this Annual Report. Preliminary results indicate that wild accessions collected in Clavellinas and some in El Esfuerzo are tetraploids CCDD genome and close related to all the O.latifolia accessions analyzed from IRRI. In contrast, the diploids wild Oryza populations from Guarico and Portuguesa are AA genome. With the exception of the Santa Lucia population and the putative wild/cultivated rice hybrid that is close related to the O. rufipogon/ O. nivara/ O. sativa complex, all the other AA populations collected in Venezuela so far appear related to O. glumaepatula. Key Words: wild Oryza species, O. latifolia, O. glumaepatula, O. rufipogon, cultivated/ wild rice hybrids, chloroplast markers, SSR markers, nuclear markers Background Rice (Oryza sativa of Asian origin, AA genome) is an introduced domesticated species that has become one of the most important staple grains for human consumption in tropical America in recent decades. The rice genus, Oryza has a pan-tropical distribution. Four species have been recorded in tropical America. Oryza glumaepatula (diploid, AA genome) classifies within the primary gene pool (Akimoto, 1998; Vaughan, 1994), whereas Oryza grandiglumis, O. alta and O. latifolia are allotetraploid (CCDD) and included in the secondary gene pool. Due to its morphological similarity with O. rufipogon, O. glumaepatula was originally classified as the American strain of O.rufipogon (Vaughan, 1994). Nevertheless, O. glumaepatula is a distinct AA species based on morphological traits, molecular markers, has compatibility barriers with O. rufipogon, and is closer related to the African species O. glaberrima, O. barthii and O.longistaminata than to the Asian O. rufipogon (Akimoto 1998, Ge et al., 2001; Juliano et al., 1998). With the exception of Costa Rica and Brazil that had conducted a complete analysis of the Oryza wild relatives composition and spatial frequency distribution, the information for the rest of the region is incomplete and scattered in few herbarium records (Lentini and Espinoza, 2005). Previous results from experiments conducted under controlled-confined conditions showed that gene flow occurred predominantly from the crop into weedy rice (Lentini and Espinoza, 2005). This work also describes the use of chloroplast and nuclear molecular markers for the characterization of weedy and wild rice populations collected in commercial rice fields and natural environments in Colombia and Venezuela, and their utility for tracking gene flow at landscape level (rate and direction) in weedy/ wild Oryza species populations .

82
Materials and Methods Plant materials. Accessions of weedy rice or wild Oryza collected in Portuguesa (86) and Guarico (68), commercial rice varieties (4), and 60 wild Oryza species obtained from IRRI germplasm bank as reference, were used. Total DNA was isolated according to Lorieux (2002). The wild Oryza species included all the accessions at IRRI reported for the four Latin American species (O.glumapepatula, O. latifolia, O. grandiglumis, O. alta). Other Oryza species were also included (O. rufipogon, O. nivara, O. galberrima, O. barthii, O. punctata, O. australiensis, O. officinalis, O. granulata, O. brachyanta, O. minuta and O. rhizomatis) to confirm the use of specific molecular markers for genome type and taxonomy confirmation.
Genetic analysis using microsatellite markers. The characterization was carried out using four highly polymorphic SSRs that clearly distinguish five wild Oryza species (O.rufipogon, O. barthii, O. glaberrima, O. glumaepatula, O. latifolia, O. alta and O. grandiglumis from IRRI). These SSRs had also been used successfully for the molecular characterization of large populations of weedy rice collected in Colombia and Venezuela conducted by our research team. The PCR products were resolved on silver-stained polyacrylamide gels and microsatellite alleles distinguish by size using a 10-to-25 bp molecular weight standard (Promega).
Chloroplast and nuclear marker genes. Three chloroplast (cp) regions (ORF100; non-coding regions between the amino acid trnS [tRNA-Ser- (GGA)] and trnT [tRNA-Thr (UGU)]; and TrnL-Trnf F, and one nuclear gene regions ( Adh-2 gene) were amplified using universal forward and reverse primers (Adh F1 and adh RR) reported by Ge et. (2001). Results and Discussion
Molecular and statistical analyses of these populations are still in progress. Preliminary results suggest that molecular analysis using the four SSR markers generates seven clusters (Figure 1). O. glaberrima and O. barthii are clearly separated from the other the Latin American Oryza species, O. nivara, O. rufipogon, O sativa, weedy rice and putative hybrids. Tetraploid accession from IRRI and the CCDD experimental wild accessions identified according to cpDNA TrnL-TrnF sequence (Figure 1) were also clear-cut separated from the AA genome samples. Twenty nine of the CCDD wild accessions are closely related to O. latifolia. These wild CCDD accessions were characterized by being tall plants with truncated ligules, small seeds with short awns and the spikes were <7 mm, taxonomy traits used reproducibly to distinguish O. latifolia from O. alta and O. grandiglumis in Venezuela (D. Vaughan, NIAS, Japan, personal communication). The remaining 9 wild CCDD appeared to be closer to O. grandiglumis and O. alta accessions. About 89% of the AA genome wild accessions collected from Guárico and Portuguesa (99 of 111 accessions) and identified as O. glumaepatula according to cpDNA trnS [TRNA-Ser- (GGA)] and trnT [tRNA-Thr (UGU)] sequences clustered with 7 of the 8 (88%) O. glumaepatula accessions from IRRI. All the weedy

83
rice, putative hybrids and the remaining (115) AA genome wild accessions clustered with all the O. rufipogon IRRI accessions (15) from a diverse number of Asian countries (Figure 1). All the weedy rice accessions analyzed so far are diploid AA genome and most of them are of indica type according to cp ORF 100 sequence. Analysis with chloroplast (cp) and nuclear DNA specific sequences indicate that weedy rice accession collected in Colombia and Venezuela are AA genome. Likewise in Colombia, in Venezuela there is some weedy rice accessions closely related to O. rufipogon and the genetic identity of the putative weedy rice hybrids collected in the farmers fields still needs further analysis. Most of the CCDD wild accessions are closely related to O. latifolia. AA genome wild accessions from “Estero de Camaguán”, “Via Herrera”,“Lecherito” and some from Portuguesa are genetically related to O. glumaepatula according to the cpDNA sequenceand SSR analysis. Specific SSRs and cpDNA markers are being used to identify potential hybrids between O. sativa and wild Oryza, to determine the reproductive biology and direction of gene flow.
Dice Similarity Coefficient0.0 0.2 0.4 0.6 0.8
A
B
Fig 1. Similarity analysis (UPGMA).
O. nivara (1)O. rufipogon /O. nivara (2)O. sativa (4)O. rufipogon (10)Wild accessions Portuguesa (21)Wild accessions Guárico (2)Weedy rice putative hybrids Guárico (12)
O. rufipogon (1)O. nivara (1)Wild accession Guárico (1)
O. latifolia (20)Wild accessions from Portuguesa (29)
O. alta (4)O. grandiglumis (4)
Wild accessions from Portuguesa (9)
O. rufipogon (6)O. barthii (1)O. glaberrima (1)
O. rufipogon (4)Weedy rice (4)Weedy rice putative hybrids (5)
Wild accessions Guárico (33)O. glumaepatula (1)
Wild accessions Guárico (55)Wild accessions Portuguesa (11)O. glumaepatula (6)

84
ReferenceAkimoto M, Shimamoto Y. and Morishima. 1998. Genetic differentiation in Oryza glumaepatula and its
phylogenetic relationships with other AA genome species. Rice Genet. Newsl 14:37-39. Buso G., Rangel P. and Ferreira M. 1998. Analysis of genetic variability of South American wild rice
populations (Oryza glumaepatula) with isozyme and RAPD markers. Molecular Ecology 7. 107-117 Buso G, Nakano R. and Ferreira M. 2001. Analysis of random and specific sequences of nuclear and
cytoplasmic DNA in diploid and tetraploid American wild rice species (Oryza spp.). Genome 44: 476-494.
Kuroda Y, Sato Y, Bounphanousay C, Kono Y. and Tanaka K. 2005. Gene Flow from cultivated rice (Oryza sativa L.) to wild Oryza species (O. rufipogon Griff. And O. nivara Sharma and Shastry) on the Vientiane plain of Laos. Euphytica 142: 75-83.
Pereira R, Brondani V, Zucchi M, Brondani C, Rangel N, Borba T, Rangel P, Magalhaes M. and Vencovsky R. 2005. Genetic structure of wild rice Oryza glumaepatula populations in three Brazilian biomes using microsatellite markers. Genetic 125: 115-123.
Lorieux, M., Ndjiondjop, M.-N., and Ghesquiere, A. 2002. A first interspecific Oryza sativa x Oryza glaberrima microsatellite-based genetic map. In : Theorical and Applied Genetics. Vol. 100; p. 591–601.
Vaughan D. and Tomoka, 1999. Wild rice in Venezuela D. Research Notes. I. Varietal Differentiation and Evolution. National Institute of Agrobiological Resources, 2-1-2 Kannondai, Tsukuba, 305-8602 Japan.
Zamora. 2005. Diversity of native rice (Oryza Poaceae) species of Costa Rica. In : Genetic Resourses and Crop Evolution. 50: 855- 870.
1.C.6 Anther culture for advancing breeding populations of FLAR
M. Quintero (IP4), G. Delgado (IP4), T. Agrono (IP4), C. Ordóñez (IP4), J. Ortega (FLAR), E. Torres (CIAT), M. Cruz (FLAR), E. Pulver (FLAR), Z. Lentini (IP4, SB2)
1.IP-4 Project; 2. FLAR;
Doubled haploids are used as a tool to ease the development of lines combining high yield, cold tolerance and commercial grain quality traits. Crosses processed by CA in 2001 were sent to Santa Victoria do Palmar (Brazil) for further crossing. In 2007, of the original lines sent FL04223-CA-9P, FL04225-CA-2P and FL04225-CA-20P-A are in preliminary evaluations; FL04225-CA-20P-B is in advanced evaluations; FL04225-CA-22P in compact evaluation and FL04225-CA-1P is in observations plots. Another set of DH lines are used as parents. In relation to the hybrid vigor exploration program by FLAR, 14 lines DH lines from FL06963 were evaluated by grain quality and were selected last year. These lines were used as progenitors in triple crosses to combine good plant type, extra-large panicle length of panicle, and cold tolerance. These crosses are in F3 and will be evaluated in Argentina, Brazil and Uruguay next year. Conclusions
The Anther Culture Laboratory had demonstrated a solid record of service for about 2 decades generating doubled haploids and/or embryo rescue from broad crosses. These plants are distributed to the breeding programs of CIAT and FLAR, as well as for developing populations for strategic research, including the development of population for molecular genome introgression analysis, at Headquarters by various partners (CIAT, IRD, and CIRAD) and elsewhere NARS from Latin America, IRRI, Texas A&M,

85
Dupont, Cornell University, among others. The proof of concept of this efficiency was transferred to WARDA during the 90s, capacity that was used to generate the NERICA lines currently evaluated throughout Africa. The efficiency at CIAT has been maintained not only due to the earlier research investment improving a protocol adapted to Latin American indica and japonica germplasm, but also by incorporating new knowledge in tissue culture procedures such temporary immersion systems and for plant regeneration.
Future Activities
Future activities are uncertain. Activities will depend on third party financial support since due to the current restructure scenario of CIAT this laboratory lost all its financial support in 2007. References Castaño, R. C. 2002. Identification of QTLs for yield and yield components using microsatellites in a doubled haploid population derived from advanced back-cross between the variety Caiapo and the wild species Oryza glaberrima. Under-graduate Thesis. Universidad de Los Andes. Department of Biological Sciences. Snape, J.W. 1989. Doubled haploid breeding: theoretical basis and practical applications. p.19-33. In A. Mujeeb-Kazi & L.A. Stich (eds.). Review of Advances in Plant Biotechnology. 2nd. International Symposium on Genetic Manipulation in Crops. CIMMYT and IRRI. Snape, J.W., and E. Simpson. 1981. The genetic expectations of doubled haploid lines derived from different filial generations. Theory. Apple. Genet. 60:123-128. Lentini Z., P. Reyes, C.P. Martínez, and W.M. Roca. 1995. Androgenesis of highly recalcitrant
1.C.7 Validation of SNP markers in Oryza sativa and wild relative species C. Quintero1, G. Plata1
, F. Rodríguez1, J. Tohme1 and M. Lorieux1
1CIAT, Agrobiodiversity and Biotechnology Project, SB-02. FundingGPC, CIAT CORE
Background Single nucleotide polymorphisms (SNPs) are biallelic markers, and together with insertions/deletions are the most abundant sources of polymorphisms in human genome. The potential of these markers has been proposed for association studies and several studies related to SNP identification in plants such as maize, barley and soybean have been initiated. In rice, in silico SNP discovery is been the most desirable method to have access to these markers. Some of the main objectives of this project are to develop a set of 140 SNPs identified in genes associated with tolerance to abiotic stresses (drought, acid soils, mineral deficiencies or toxicities), to validate their utility by using them in the development of the CSSLs and to explore their value in breeding programs for other cereals.

86
Materials and methods
Plant material Genomic DNA from 15 rice genotypes was extracted following Dellaporta (1983) protocol with some modifications. Plant material belonged to Oryza sativa subsps. japonica/indica and to the wild rice species O. barthii, O. glumaepatula, O. rufipogon, O glaberrima and O. meridionalis used as donor parents in the development of CSSLs populations.
SNP identification and genotyping
345 SNPs were detected along the 12 rice chromosomes. 12 PCR-multiplex experiments were designed by Plata and Lorieux. Information relevant to SNPs markers was retrieved such as PCR primer pairs and single base extension (SBE) primer. SBE reactions were designed by Quintero and SNP allele scoring was done as previously reported using the flow cytometer Luminex100 as platform and Nipponbare and 93-11 as checks (Quintero etal., Annual Report 2006). Results SNP Validation.
Although the basics of the methodology was determined in previous assays, some modifications are still being introduced in order to decrease the cost per data point without loosing the high resolution of the single base extension method and the Luminex platform. A dramatic reduction in the AccuPrime� Taq Polymerase (Invitrogen) was performed together with the use of fewer amounts of beads per sample, producing consistent results across different assays. No changes in detected fluorescence intensity were observed and confident SNP genotyping was done under the new standardized conditions. All SNP markers located in rice chromosomes 1, 2, 3, 5, 6 and 7 were validated. PCR-multiplex reactions were then carried out in six sets of 30 markers, one for each chromosome. Single base extension reactions were carried out in multiplex of 13 to 19 SBE primers and then joined (up to 36 extended products) for the quantification of their mean fluorescent intensity in the Luminex flow cytometer. No fluorescent signal was observed for 47 SNPs, 11 were monomorphic and 7 were classified as no-specific since one allele could not be recognized alone and was observed always together with the alternate allele as a heterozygote. The remaining 114 SNPs (64%) were scored with high fluorescent signals and were also polymorphic, although 7 of them did not reproduced the polymorphism between Nipponbare and 93-11. For four markers, alleles could be scored only in O. sativa and O. rufipogon but not in the other species.
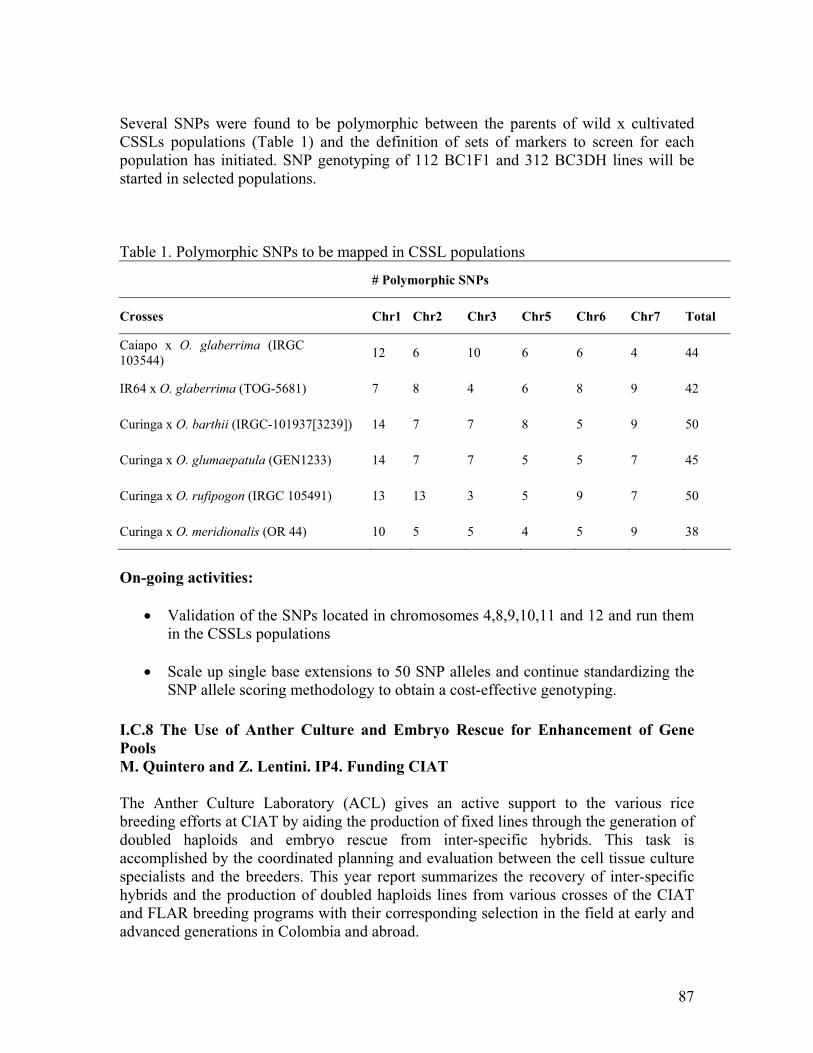
87
Several SNPs were found to be polymorphic between the parents of wild x cultivated CSSLs populations (Table 1) and the definition of sets of markers to screen for each population has initiated. SNP genotyping of 112 BC1F1 and 312 BC3DH lines will be started in selected populations. Table 1. Polymorphic SNPs to be mapped in CSSL populations
# Polymorphic SNPs
Crosses Chr1 Chr2 Chr3 Chr5 Chr6 Chr7 Total
Caiapo x O. glaberrima (IRGC 103544) 12 6 10 6 6 4 44
IR64 x O. glaberrima (TOG-5681) 7 8 4 6 8 9 42
Curinga x O. barthii (IRGC-101937[3239]) 14 7 7 8 5 9 50
Curinga x O. glumaepatula (GEN1233) 14 7 7 5 5 7 45
Curinga x O. rufipogon (IRGC 105491) 13 13 3 5 9 7 50
Curinga x O. meridionalis (OR 44) 10 5 5 4 5 9 38
On-going activities:
� Validation of the SNPs located in chromosomes 4,8,9,10,11 and 12 and run them in the CSSLs populations
� Scale up single base extensions to 50 SNP alleles and continue standardizing the
SNP allele scoring methodology to obtain a cost-effective genotyping.
I.C.8 The Use of Anther Culture and Embryo Rescue for Enhancement of Gene PoolsM. Quintero and Z. Lentini. IP4. Funding CIAT The Anther Culture Laboratory (ACL) gives an active support to the various rice breeding efforts at CIAT by aiding the production of fixed lines through the generation of doubled haploids and embryo rescue from inter-specific hybrids. This task is accomplished by the coordinated planning and evaluation between the cell tissue culture specialists and the breeders. This year report summarizes the recovery of inter-specific hybrids and the production of doubled haploids lines from various crosses of the CIAT and FLAR breeding programs with their corresponding selection in the field at early and advanced generations in Colombia and abroad.

88
Key Words: doubled haploids, anther culture Background Homozygous doubled haploids (DH) lines derived from spontaneous chromosome doubling of the microspore haploid genome of rice can be obtained through anther culture (AC) in less than one year, saving time in evaluation trials (DH vs. F6) and in building up pure stocks. It is also possible to gain efficiency with DH populations when selecting for qualitative traits because of the absence of dominance, and for quantitative traits due to a greater additive variance, no intra-family segregation, and no interplant competition (Snape and Simpson, 1981; and Snape, 1989). At CIAT, AC has proved to be useful in accelerating the development of germplasm tolerant to low temperatures and having excellent grain quality, increasing the recovery of useful recombinants from wide crosses for disease and pest resistance, drought tolerance; and facilitating the production of materials suitable for molecular markers gene tagging. The CIAT rice anther culture laboratory (ACL) currently focus on developing doubled haploid lines for the various breeding efforts stationed at CIAT. In the case of CIAT, the work has been mainly directed to advanced populations adapted to the irrigated and upland savanna ecosystems, as well as backcross populations derived from crosses between cultivated rice and wild rice species (Castaño, 2002). In the case of FLAR, CIAT has given a support service. The laboratory has generated lines from FLAR crosses targeting the sub-tropical and cold tolerant breeding lines pools for the Southern cone, and produced somaclone lines for Tropical Latin America. In addition to this support, the laboratory also aids the generation of broad crosses through the rescue of immature embryos from inter-specific hybridizations that otherwise abort few days after pollination.
Materials and Methods For anther culture, plants are planted in the field, panicles harvested and cold pre-treated, and anthers dissected and cultured in vitro according to Lentini et al (1995). Regenerated plants are delivered to the breeders to continue with the selection process under field conditions. In the case of embryo rescue, embryos are cultured in vitro few days after pollination when the endosperm is still in milky stage (liquid). Rooted and elongated seedlings gown in vitro are transferred to the greenhouse and 25-day-old plants are grown in the field. Selected plants are then processed through anther culture.
Results and Discussion Embryo rescue from inter-specific hybrids M. Quintero (IP4), G. Delgado (IP4), S. Sánchez (IP4), S.J. Carabalí (IP4), J. Borrero (IP4), C. Martínez (IP4, SB2), Z. Lentini (IP4, SB2) Plants recovered by embryo rescue from Inter-specific crosses and backcross with

89
advances lines BCF1720, BCF1658 and varieties Fedearroz 50, Pi9 and Fanny were evaluated for RHBV resistance and Tagosodes a mechanical damage, and other agronomic characteristics in the last years. These backcrosses are in a generation F7, represented by 1,231 selected lines. The advanced selection will be evaluated for resistance to different pathogens in Santa Rosa experimental station. Doubled haploids lines aiding gene mappingM. Quintero (IP4), G. Delgado (IP4), T. Agrono (IP4), C. Ordóñez (IP4), J. Borrero (IP4),C. Martínez (IP4, SB2), Z. Lentini (IP4, SB2)
Triple crosses from the japonica X wild (O. barthii)/ indica were processed through anther culture, and then selected for RHBV and Pyricularia resistance, yield potential and grain quality. Advanced lines were evaluated in replicated trials during the first semester of 2006 for yield potential at CIAT experimental station in Villavicencio, and on the second semester at CIAT station in Palmira. This year the selected lines were distributed to National Programs via different nurseries.
Genetic fixation through the doubled haploids generation M. Quintero (IP4), G. Delgado (IP4), J.D. Arbelaez (Fedearroz), M. Lorieaux (SB2), C. Martinez (IP4, SB2), Z. Lentini (IP4, SB2)
Curinga (elite Brazilian variety) is a tropical japonica variety tolerant to drought, which was chosen as recurrent parent for the development of 4 populations of introgression lines with 4 AA genome wild Oryza species (O. rufipogon, O. bartii, O. meridionalis and O. glumaepatula). The response to anther culture of this variety was evaluated as preliminary step for developing DH form the inter-specific crosses. Curinga showed intermediate response to AC, and of the 211 R1 plants generated about 36% were DH. These results suggest that AC is an alternative technique to induce rapid homozygosis in Curinga crosses.
Anther culture for advancing breeding populations of FLAR M. Quintero (IP4), G. Delgado (IP4), T. Agrono (IP4), C. Ordóñez (IP4), J. Ortega (FLAR), E. Torres (CIAT), M. Cruz (FLAR), E. Pulver (FLAR), Z. Lentini (IP4, SB2) Doubled haploids are used as a tool to ease the development of lines combining high yield, cold tolerance and commercial grain quality traits. Crosses processed by CA in 2001 were sent to Santa Victoria do Palmar (Brazil) for further crossing. In 2007, of the original lines sent FL04223-CA-9P, FL04225-CA-2P and FL04225-CA-20P-A are in preliminary evaluations; FL04225-CA-20P-B is in advanced evaluations; FL04225-CA-22P in compact evaluation and FL04225-CA-1P is in observations plots. Another set of DH lines are used as parents. In relation to the hybrid vigor exploration program by FLAR, 14 lines DH lines from FL06963 were evaluated by grain quality and were selected last year. These lines were used as progenitors in triple crosses to combine good plant type, extra-large panicle length of panicle, and cold tolerance. These crosses are in

90
F3 and will be evaluated in Argentina, Brazil and Uruguay next year. ConclusionsThe Anther Culture Laboratory had demonstrated a solid record of service for about 2 decades generating doubled haploids and/or embryo rescue from broad crosses. These plants are distributed to the breeding programs of CIAT and FLAR, as well as for developing populations for strategic research, including the development of population for molecular genome introgression analysis, at Headquarters by various partners (CIAT, IRD, and CIRAD) and elsewhere NARS from Latin America, IRRI, Texas A&M, Dupont, Cornell University, among others. The proof of concept of this efficiency was transferred to WARDA during the 90s, capacity that was used to generate the NERICA lines currently evaluated throughout Africa. The efficiency at CIAT has been maintained not only due to the earlier research investment improving a protocol adapted to Latin American indica and japonica germplasm, but also by incorporating new knowledge in tissue culture procedures such temporary immersion systems and for plant regeneration. Future Activities Future activities are uncertain. Activities will depend on third party financial support since due to the current restructure scenario of CIAT this laboratory lost all its financial support in 2007. References Castaño, R. C. 2002. Identification of QTLs for yield and yield components using microsatellites in a doubled haploid population derived from advanced back-cross between the variety Caiapo and the wild species Oryza glaberrima. Under-graduate Thesis. Universidad de Los Andes. Department of Biological Sciences. Snape, J.W. 1989. Doubled haploid breeding: theoretical basis and practical applications. p.19-33. In A. Mujeeb-Kazi & L.A. Stich (eds.). Review of Advances in Plant Biotechnology. 2nd. International Symposium on Genetic Manipulation in Crops. CIMMYT and IRRI. Snape, J.W., and E. Simpson. 1981. The genetic expectations of doubled haploid lines derived from different filial generations. Theory. Apple. Genet. 60:123-128. Lentini Z., P. Reyes, C.P. Martínez, and W.M. Roca. 1995. Androgenesis of highly recalcitrant 1.C.9 Foreign genes as novel sources for increased efficiency of water use in rice
E. Tabares1, T. Sakai1, G. Delgado2, A. Salcedo1, L. Galindo1., T. Agrono2, C. Ordóñez 2M. Ishitani1 and Z. Lentini 1,2. 1SB2 project, 2 IP4 project. Funding from MAFF, Japan Background Rice yield potential (irrigated and upland rice) is highly dependent of adequate availability of water throughout the plant growth cycle. Irrigated rice currently accounts for most productivity worldwide. Irrigated rice requires large amount of water competing with water usage for other human activities. Flooded paddy rice fields significantly contribute with methane emission associated with the earth greenhouse effect and consequently climate change. In addition, water is becoming a scarce resource, also in Latin America which currently accounts for the largest ratio of water availability per

91
person, thus breeding rice for increased efficiency in water use must be a priority. Knowledge on molecular metabolism associated with drought tolerance in rice is limited. The physiological response to water stress is driven by changes in gene expression at the cellular level. Several genes had been demonstrated to be associated with tolerance to drought, salinity and cold, and the proteins encoded by these genes are thought to protect cells from these stresses (Shinozaki and Yamaguchi- Shinozaki, 1997). Some genes respond to water stress very rapidly, whereas others are induced after the accumulation of abcisic acid (ABA). Analyses in Arabidopsis thaliana of gene promoters induced by dehydration and cold have identified several cis-acting elements that are involved in ABA dependent and ABA independent responses to water stress. The DRE element (Dehydration Responsive Element) has been implicated in the regulation of dehydration responsive gene expression and found in promoter regions of dehydration and cold stress inducible genes (Kasuga et al., 1999). The main gene controlling the expression of these stress inducible genes is DREB (Dehydration Responsive Element Binding protein), which has been characterized as an early response transcription factor controlling the expression of multiple genes under drought stress. DREB (also known as CBF) genes have been isolated and characterized from Arabidopsis thaliana (Liu et al., 1998), and rice (Oryza sativa, Dubouzet et al., 2003). The Arabidospis DREB have been used in heterologous system to test its transgenic expression in tomato (Hsieh et al., 2002). Other transgenic approaches have been tested to improve stress tolerance in plants using genes encoding for enzymes involved in the biosynthesis of different osmo-protectants or encoding for modified membrane lipids, such as LEA protein and detoxification enzymes (cited by Kasuga et al., 1999). The main objective of this work is to test DREB genes, and other sequences associated with tolerance to drought stress, in order to confer increased water use efficiency in commercial rice lines adapted to Latin American tropical conditions, and to understand the molecular mechanisms underlining the stress tolerance in these plants. The gene constructs were kindly provided by Dr. Yamaguchi-Shinozaki (from the Japan International Research Center for Agricultural Sciences, JIRCAS) and Dr. Shinozaki (from RIKEN, Japan). Here we report the progress made in rice during the past six months.
Materials and Methods
Plant Material: A set of 9 upland genotypes released as commercial varieties (from which 60% are derived from CIAT germplasm) were selected by the rice breeder and tested for their plant regeneration response in order to identify the best experimental material to generate the transgenic lines carrying the various gene constructs for the project. The tropical japonica materials included 2 lines released in Nicaragua and grown by small farmers (Inta Chinandega, and the line CT15944-10-4-3-3 inter-specific cross Caiapo / O.glaberrima (Trouche G, et al. 2006); 6 commercial varieties released in Brazil (Curinga, Can-s 9025, Caiapo, Libranza, Bonanza, and Primavera); and one commercial variety released in the Colombian Llanos (Linea 30). The Japonica variety of Nipponbare was used as a control.
Selection for hygromycin resistance: Previous experience at CIAT indicates that not all

92
rice varieties have the same sensitivity to hygromycin. Some Latin American varieties are either highly susceptible or more tolerant to the standard hygromycin concentration of 30 -50 mg/l usually used for most rice genotypes worldwide. Therefore the selected materials were evaluated for susceptibility on various hygromycin concentrations (0, 10, 30, 50 and mg/L hygromycin) at callus and plant regeneration stage.
Rice genetic transformation: CIAT’s protocol takes about 3-4 months to generate 500 transgenic events for six construct combinations from indica and upland rice. In order to evaluate if it is possible to reduce the time span in the laboratory and generate higher numbers of transgenic Latin American upland rice independent events with a larger number of constructs (> 6), experiments were conducted comparing CIAT’s protocol with other protocols developed for high speed transformation of Nipponbare (Toki et al., 2006). The comparison included the evaluations of the effect of high temperature (34°C as in Toki et al., 2006) during callus induction and selection compared to 24-26°C used at CIAT, of the different medium compositions, and of the Agrobacterium strains [Agl1 (Wang et al., 1997) Vs EHA105 (Toki et al., 2006)] on the in vitro response and transformation efficiency. Plasmid constructions. Six gene cassette constructs : pBIH-Lip9:DREB1A, pBIH:osnac6:DREB1A, pBIH:ubi:SRK2C, pBIH:ubi:DREB1A, pBIH :ubi:NCED3, and pBIH:ubi:At-GOLS2 were used for plant Agrobacterium mediated transformation .The pBIH-Lip9:DREB1A contains the Arabidopsis thaliana DREB1A transcription factor conferring tolerance to drought and low temperatures (Lui et al., 1998),
Results Production of transgenic rice CIAT has well-established and efficient rice genetic transformation protocol adapted to generate a large number of events and healthy fertile plants from commercial varieties adapted to upland and irrigated Latin American ecosystems (Lentini et al., 2003). The protocol takes about 3-4 months to generate 500 events of six constructs combinations from indica and upland rice ready for transfer to the biosafety greenhouse conditions. Using this protocol we generated 188 events Lip9::AtDREB1A and 228 events Lip9::OsDREB1B using Palmar, Cica 8 and CT6241 Latin American genotypes adapted to lowland and upland conditions. We had identified enough single copy events (19 from Lip9::AtDREB1A) and (58 from Lip 9::OsDREB1B), and 2-copy events (20 from Lip9::AtDREB1A) and (11 from Lip 9::OsDREB1B). This material will be useful to conduct Lip 9 promoter dosage analysis and preliminary analysis of gene construct efficiency for drought tolerance with large-scale population screening in the field (more 300 seeds per homozygous event) at CIAT-Head Quarters experimental station under biosafety conditions. This information will be useful for establishing the Standard Operation Procedures (SOP) for the field handling and evaluation of Curinga upland transgenic events, the target variety selected for this project. In evaluating to the possibility of reducing the time span in the laboratory to generate a high number of transgenic Latin American upland rice independent events with a large

93
number (6-10) of constructs, a preliminary experiment was conducted testing other protocols developed for high speed transformation of Nipponbare (Toki et al., 2006). In addition to differences in medium composition, a major difference between CIAT’s and Toki’s protocol is that callus induction and hygromycin resistance callus selection in performed at a high temperature (34C) instead at 24-26°C. The high temperature is the key factor in Toki’s protocol to speed up the process for generating transgenic plants. However, it is well known that high temperature may reduce embryogenesis in rice depending on the genotype, and therefore reduce the plant regeneration capacity, and in some genotypes such us upland rice, high temperature may increase the regeneration of albino plants. For these reasons, a series of experiments were conducted to systematically compare CIAT’s and Toki’s protocol to identify any potential modifications that could be introduced into CIAT’s protocol for a higher speed transformation.
Selection of experimental upland genotypeDe-husked mature seeds were used to induce callus, and 1–2 mm in diameter embryogenic callus were transferred to evaluate plant regeneration about 4 weeks after treatment. Experiments included the evaluation of the response from of 6 upland Brazilian commercial varieties (Curinga, Can-s 9025, Caiapo, Libranza, Bonanza, and Primavera), the Colombian variety Linea 30; the Central American variety Inta Chinandenga, an inter-specific hybrid O. sativa/ O. glalberrima and Nipponbare, using CIAT’s and Toki’s protocol.. Linea 30 show a high callus embryogenesis response from 65% to 96% at 24-26°C on NBA medium (CIAT’s protocol). Plant regeneration from these materials is in progress in order to make the final decision of the upland genotype selected for the project. Curinga showed the highest in vitro response on callus induction and plant regeneration (about 80%) when using CIAT’s protocol (Figure 1) in contrast to 40% when using 34°C, N6D and RIII media (as in Toki et al., 2006). In all the cases, high temperature (34°C) significantly accelerated the process but the callus induced from the upland materials was non-embryogenic and sometimes yielded albino plants, whereas Nipponbare showed formation of highly embryogenic in vitro response. The other Brazilian upland genotypes show high callus embryogenesis (65% to 96%) at 24-26C and on NBA medium (CIAT’s protocol). Bonanza and Primavera showed high plant regeneration (70-75%) likewise Curinga. Of the upland materials showing high in vitro response, Curinga has was selected as the target variety because it has a boarder adapatability under favored and ufavored upland conditions, higher yield potential, better tolerance to rice blast, good grain quality. Curinga is rapidly replacing other upland commercial varieties in Brazil, Brazilian National Breeding Program (CNPAF EMBRAPA) is more confident on Curinga’s performance towards a higher potential impact in terms of area planted. This genotype could also have high adapatibility in other upland regions of Latin America, including Central America, Bolivia, Colombia, and Peru, where upland rice is preferentially grown by small farmers. An additional advantage is that Curinga is the genotype of choice for a project on functional genomics on drought tolerance between CIAT and EMBRAPA (Brazil) funded by the Generation Challenge Program, thus information generated may complement the one generated in the MAFF project. Optimal selection for hygromycin resistance of experimental upland genotypes

94
Results indicate that callus proliferation of most genotypes evaluated (i.e.Inta Chinandenga) starts being inhibited at 30 mg/L and optimal selection can be obtained at 30-50 mg/L likewise Nipponbare (Figure 2). Similar results are obtained for Linea 30 (data not shown). In contrast, Curinga appears to be highly susceptible to hygromycin and concentrations as low as 10 mg/L reduces callus induction by 90% (Figure 2). This high susceptibility of Curinga to hygromycin may cause difficulties in the selection of agro-infected callus, thus various combinations for hygromycin selection were tested to standardize an optimal protocol for generating transgenic plants. Generation of transgenic plants In order to gain time for establishing the optimal genetic transformation protocol for Curinga (there was not previous experience with this variety at CIAT laboratory), preliminary experiments were conducted transforming rice callus with Agro-strains harboring the pCAMBIA 1305.2 plasmid which contains the gus-intron and hygromycin resistance genes driven by the CaMV 35S promoter prior receiving the set of constructs from JIRCAS and RIKEN in September 2007. The protocol developed for Nipponbare (Toki et al., 2006) for higher speed transformation using 34�C for callus induction and selection on hygromycin is highly efficient, about 100% of hygromycin resistant callus showed stable gus expression (Table 1 and Figure 3). Curinga showed low or no stable transformation at such high temperatures (Table 1), but in contrast about 70% to 100% stable gus expression on hygromycin resistant callus is obtained at 24-26 �C independent of the Agrobacterium strain used (Table 2). Based on these results the first set of transgenic activities with Curinga-Lip9::AtDREB1A cassette-construct were initiated by mid-September and the first set of transgenic events were generated and transferred to biosafety greenhouse conditions (Figure 4). The first round of transformation activities using pBIH-osnac6:DREB1A; pBIH-ubi:SRK2C; pBIH-ubi:NCED3; pBIH-ubi:OsNAC6; and pBIH-ubi:AtGolS2 were conducted, and those using the first 4 constructs are at advanced hygromycin regeneration selection stage. Acquisition of a biosafety permit for field release of transgenic rice:CIAT already has a permit from the Colombian biosafety authorities to conduct 2 Ha field trials with transgenic virus resistant rice (since 2000, Lentini et al., 2003), and 5 Ha field trials with T-DNA transgenic rice mutant populations from IRD (France). Based on recent changes in the National Regulation and the long history of CIAT researchers conducting field trials in compliance with biosafety regulations, Colombian authorities advised CIAT to first request a generic permit for the evaluation of drought tolerance of the different transgenic events to be generated under the MAFF project. Right after receiving the first set of gene constructs for MAFF project in September 2007, the generic permit was submitted to the Colombian National Biosafety Committee in October 2007, this request was evaluated and approved by the Committee in December 2007, and the official permit is being issued and will be delivered to CIAT by the end of January-February 2008 according to personal communication with the Competent authorities. This official generic permit will allow conducting field trails under biosafety conditions at CIAT Headquarters’ experimental station for all the MAFF project constructs. This type of permit will significantly facilitate and accelerate the field trials evaluation since there is not need to request a permit in each case because the Competent authorities only

95
requires to be notified and provided with the corresponding specific gene constructs sequence and required molecular transgenic events information prior the first field trial.
Figure 1. High plant regeneration from embryogenic callus of Curinga following CIAT protocol at 24-26 °C
0
20
40
60
80
100
0 10 30 50 75Hygromycin concentration (mg/L)
Callus survival (%)
NipponbareINTA CHINANDEGA CURINGA
Figure 2. Hygromycin susceptibility of embryogenic callus (1-2mm in diameter) from Nipponbare, Inta Chinandega and Curinga genotypes.

96
Figure 3. Stable gus expression in Curinga-pCAMBIA 1305.2 hygromycin resistant callus from experiments establishing optimal transformation protocol for this new genotype
Figure 4. First set of Curinga plants from pBIH-lip9:DREB1A transformation activities currently under molecular analysis
Table 1. Gus expression of callus induced and selected on hygromycin contained medium at 34�C (Toki et al, 2006)
Ricegenotype
Protocol for agroinfection
Agrobacteriumstrain
Selectionmedium
Hygromycin mg/ L
Callus hygromycin resistant(%)
Callus w/stable gus expression (%)
Curinga CIAT AGL 1 NBA 20 95 11 EHA105 NBA 20 83 0
AGL 1 NBA 50 100 100 Nipponbare EHA105 NBA 50 75 100 Curinga TOKI AGL 1 N6D 20 85 0 EHA105 N6D 20 70 0

97
Nipponbare AGL 1 N6D 50 50 100
EHA105 N6D 50 100 95
Table 2. Gus expression of Curinga callus induced and selected on hygromycin contained medium at 24-26 �C
Ricegenotype
Protocol for agroinfection
Agrobacteriumstrain
Selectionmedium
Hygromycinmg/ L
Callus hygromycin resistant(%)
Callus w/ stable gusexpression (%)
Curinga CIAT AGL 1 NBA 10 83 70 NBA 20 67 100
EHA105 NBA 10 40 69 NBA 20 25 100 Curinga TOKI AGL 1 NBA 10 88 61 NBA 20 40 25 EHA105 NBA 10 58 55 NBA 20 20 75 References
Bray E A. 1997. Plant response to water deficit. Trends Plant Sci. 2:48-54 Dubouzet J G, Sakuma Y, Ito Y, Kasuga M, Dubouzet E G, Miura S, Seki M, Shinozaki K, and Yamaguchi-Shinozaki K. 2003. OsDREB genes in rice, Oryza sativa L., encode transcription activators that function in drought-, high-salt- and cold-responsive gene expression. The plant Journal 33: 751-763 Ingham D J. 2004. The study of transgene copy number and organization. In: Methods in Molecular Biology, Vol. 286: Transgenic Plants: Methods and Protocols. Edited by L. Peña. Humana Press Inc. Totowa, NJ. Kasuga M, Liu Q, Miura S, Yamaguchi-Shinozaki K, and Shinozaki K. 1999. Improving plant drought, salt, and freezing tolerance by gene transfer of a single stress-inducible transcription factor. Nature Biotechnology, Vol. 17: 287-291. Kohli A, Leech M, Vain P, Laurie D A, and Christou P. 1998. Transgene organization in rice engineered through direct DNA transfer supports a two-phase integration mechanism mediated by the establishment of integration hot spots. Plant Biology Vol. 95: 7203-7208. Liu, Q., Kasuga, M., Sakuma, Y., Abe, H., Miura, S., Yamaguchi -Shinozaki, K. and Shinozaki, K. 1999. Two transcription factors DREB1 and DREB2, with an EREBP/AP2 DNA-binding domain separate two cellular signal transduction pathways in drought and low temperature responsive gene expression respectively in Arabidopsis. Plant Cell 10: 1391-1406.

98
Output 2: Integrated crop, pest and disease management
2A. RICE PATHOLOGY
2.A.1 Rice Sheath Blight (Rhizoctonia solani)
� Analysis of two mapping populations for the identification of QTLs controlling sheath blight tolerance in rice (RiceCap Project)
Contributors: Fernando Correa1, Gustavo Prado1, Yulin Jia2, Guangjie Liu2, James Correll3, Anna McClung2. (1CIAT, 2USDA-Dale Bumpers National Rice Research Center, 3University of Arkansas) Funding:
USDA – CSREES, U. of ARKANSAS, CIAT
Abstract Sheath blight caused by Rhizoctonia solani is one of the most important and devastating rice diseases in the world. Sheath blight tolerance is a complex quantitative trait controlled by many genes located on different chromosomes for which some QTLs have been identified. In this study, two mapping populations were evaluated for their sheath blight reaction using two different greenhouse screening methods. Preliminary QTL analysis using more than 100 microsatellite markers identified several QTLs on at least 7 rice chromosomes. Several of the QTLs identified are similar to other previously mapped QTLs. Key words: QTL, sheath blight tolerance, screening methods, mapping populations Background Sheath blight caused by Rhizoctonia solani is one of the most important and devastating rice diseases in the world. In Latin America yield losses up to 50% can occur from this disease. Qualitative resistance genes capable of conferring complete resistance to the pathogen (R genes) have not been identified in rice. Instead, sheath blight resistance, or tolerance, is a complex quantitative trait controlled by many genes located on different chromosomes for which some QTLs have been identified. The slow progress in developing sheath blight resistant cultivars is partly due to the difficulty in identifying true resistance in cultivars. Disease development can be greatly influenced by climatic factors and plant traits, which may confound the host-pathogen interaction in the field. To accurately quantify sheath blight resistance, a study was conducted to refine a greenhouse screening method for sheath blight resistance in Arkansas, Louisiana, Texas, and CIAT-

99
Colombia, as part of a USDA-RiceCap Project. Two screening methods, micro-chamber (soft drink bottle) and mist-chamber (high temperature and humidity conditions) were selected after being adapted, modified, and tested for screening sheath blight resistance and obtaining similar observed field reactions of known rice cultivars. Materials and Methods For identifying and using molecular markers linked to quantitative trait loci (QTLs) which control resistance to sheath blight disease, two greenhouse screening-methods (micro-chamber and mist-chamber) were used to phenotype two breeding populations at CIAT-Colombia. One population consisted of 250 recombinant inbreed lines (RIL) developed from the cross between the cultivars JASMINE 85 (tolerant) and LEMONT (susceptible), and the other consisted of 325 double haploid lines (DH) developed from the cross between MCR 10277 (tolerant) and COCODRIE (susceptible). For the micro-chamber method (soft-drink bottle), inoculum of the pathogen R. solani was produced on culture medium (PDA) and used to inoculate 3 week old rice seedlings growing in 25 cm diameter pots. A small section of agar colonized by the pathogen was placed at the base of each seedling. The plants were covered with a 3 L clear plastic bottle (bottom and cap removed) and incubated for 8-10 days, after which they were scored for sheath blight severity evaluating the percentage of plant area affected. Each line was evaluated in four replications with three plants per replication. For the mist chamber method (high temperature and high humidity), inoculum of the pathogen grown on PDA was used to inoculate 50 days old plants growing in 25 cm diameter pots. A small section of agar colonized by the pathogen was placed at the base of each plant. The inoculated plants were incubated at high RH (> 95%) for 10 days. The plants were then moved to the greenhouse for 5 days and then returned to the high humidity chamber for 5 more days. The plants were scored for sheath blight severity evaluating the percentage of plant area affected. Each line was evaluated in five replications with five plants per replication. Results and Discussion The sheath blight segregating pattern for the recombinant inbred line population inoculated using the micro-chamber screening method is presented in Figure 1. Results indicate a pretty well distribution of lines with a mean average for plant area affected of 73.3%. The two parents exhibited a contrasting sheath blight reaction of 50.7% plant area affected for JASMINE 85, and 91.7% for LEMONT. This contrasting reaction observed for the two parents makes this population very appropriate for mapping potential QTLs controlling sheath blight resistance. The average correlation coefficient between replications was in the range of 0.39-0.49. A total of 8.4% of the lines exhibited more tolerance than the Jasmine 85 parent while 13.2% of the lines were more susceptible than Lemont. The sheath blight segregating pattern for this RIL population evaluated using the mist-chamber method is shown in Figure 2, again showing a pretty well distribution of the sheath blight reaction. Comparison of the sheath blight reaction of Jasmine 85 and Lemont with local rice cultivars included in the experiment is presented in Figure 33. The

100
two parents of the population were as tolerant or susceptible as the local rice cultivars where there were significant differences between the tolerant and susceptible groups. The sheath blight segregating pattern for the double haploid lines inoculated using the mist-chamber screening method is presented in Figure 4. Results indicate also a good distribution pattern of the reaction to the pathogen, despite the low contrasting reaction of the two parents, MCR 10277 (46.2% plant area affected) and 55.0% plant area affected for Cocodrie. The mean average for all 325 double haploid lines was 64.6%, which is higher than the mean PAA exhibited by the susceptible parent Cocodrie. Just 2.15% of the lines were more tolerant than MCR 10277 while 78.15% of the lines were more susceptible than Cocodrie, with 31 lines exhibiting a highly susceptible sheath blight reaction above 85% plant area affected. We consider that the broad reaction pattern observed for the double haploid breeding population makes it still appropriate for QTL analysis of the sheath blight tolerance, despite the low contrasting reaction of the two parents observed in this greenhouse inoculation. The average correlation coefficient between replications was in the range of 0.34-0.50, comparable to the correlations observed using the micro-chamber method with the recombinant inbreed population. The sheath blight segregating pattern for this DH population evaluated using the micro-chamber method is shown in Figure 5, again showing a pretty well distribution of the sheath blight reaction. The percentage of lines within each one of the two populations classified in the same group as tolerant or non-tolerant using each of the two screening methods and based on the confidence intervals of 95% identified for each tolerant progenitor ranged between 85-90% (Figure 6 and Figure 7). The confidence intervals calculated for Jasmine 85 were 56.5% for the micro-chamber method and 48.1% for the mist-chamber screening method, while the confidence intervals for MCR 10277 were 57.7% and 50.8% for the micro-chamber and mist-chamber screening methods, respectively (Figure 6 and Figure 7). These results indicate a high correlation between the two screening methods in the evaluation of the sheath blight reaction for each rice population. Preliminary analysis for the identification of possible QTLs controlling the tolerant reaction to sheath blight was conducted in the RILs of the Jasmine 85 x Lemont population evaluated with the mist-chamber screening method and using more than 100 microsatellite markers. Several QTLs associated with sheath blight tolerance were identified on chromosomes 1, 2, 3, 6, 9, 10, and 12 (Figure 8). Two major QTLs (qSB-9 and qSB-2-1) were identified on chromosomes 9 and 2, respectively. Several of the QTLs found are located similarly to other QTLs reported in other studies, indicating a good commonality between the results being obtained in this study and previously mapped SB-QTLs using field evaluations. This commonality also indicates the suitability of the greenhouse screening methods used for the evaluation and phenotyping of the sheath blight reaction for the identification of QTLs controlling tolerance to the Rhizoctoniasolani pathogen in different mapping populations.

101
Conclusion Phenotyping of the sheath blight reaction of two breeding populations using two different greenhouse screening methods has potentially yielded reliable information that can be used for mapping QTLs controlling resistance to the pathogen (overall goal of this project), which can be used in the future for improving the tolerance to the disease in commercial cultivars introgressing such resistance genes with the help of molecular markers. We are continuing our research to complete the genotypic analysis of the sheath blight evaluation of each population with each screening method and to compare results with similar trials conducted in Arkansas and Louisiana using the micro-chamber method as well as field evaluations. Figure 1. Distribution of the percentage of number of lines from 250 RIL from the cross LEMONT (L) x JASMINE 85 (J) inoculated with R. solani using the micro-chamber screening method to evaluate the percentage plant area affected (PAA).
Average
0
10
20
30
40
50
5 15 25 35 45 55 65 75 85 95% PAA
% o
f Lines
J L

102
Figure 2. Distribution of the percentage of number of lines from 250 RIL from the cross LEMONT (L) x JASMINE 85 (J) inoculated with R. solani using the mist-chamber screening method to evaluate the percentage plant area affected (PAA).
Figure 3. Comparison of the sheath blight reaction (% plant area affected, PAA) of the progenitors JASMINE 85 and LEMONT with local rice cultivars using the micro-chamber screening method
0
25
50
75
100
Aruare 3
Oryzica
3
Jasmine
85
Remadj
aPalm
ar
Colombia
1
Fedearr
oz 200
0Lem
ont
Cultivar
% P
AA
bb b b b
aa
a
0
10
20
30
5 15 25 35 45 55 65 75 85 95% Plant Area Affected
% o
f Lin
es
J
L

103
Figure 4. Distribution of the percentage of number of lines (325 DH) from the cross COCODRIE (C) x MCR 10277(M) inoculated with R. solani using the mist-chamber screening method to evaluate the percentage of plant area affected (PAA)
Figure 5. Distribution of the percentage of number of lines (325 DH) from the cross COCODRIE (C) x MCR 10277(M) inoculated with R. solani using the micro-chamber screening method to evaluate the percentage of plant area affected (PAA)
0
10
20
30
40
0-10 10-20 20-30 30-40 40-50 50-60 60-70 70-80 80-90 90-100
% Plant Area Affected
% o
f Lin
es
M
C
Global average
0
10
20
30
5 15 25 35 45 55 65 75 85 95% PAA
% o
f Li
nes
M
C

104
30
40
50
60
70
80
90
100
30 40 50 60 70 80 90 100% PAA (micro chamber)
% P
AA
(mis
t cha
mbe
r)
Tolerant in micro chamber/Tolerant inmist chamberTolerant in micro chamber/No Tolerantin mist chamberNo tolerant in micro chamber/Tolerantin mist chamberNo tolerant in micro chamber/NoTolerant in mist chamberJasmine 85
Lemont
30
40
50
60
70
80
90
100
30 40 50 60 70 80 90 100% PAA (micro chamber)
% P
AA
(mis
t cha
mbe
r)
Tolerant in micro chamber/Tolerant inmist chamberTolerant in micro chamber/No Tolerantin mist chamberNo tolerant in micro chamber/Tolerantin mist chamberNo tolerant in micro chamber/NoTolerant in mist chamberJasmine 85
Lemont
Figure 6. Sheath blight tolerant and non-tolerant classification of 250 RILs of the cross Jasmine 85 (tolerant) x Lemont (susceptible) using a mist chamber screening method (Jasmine 85=48.1% PAA) and micro chamber screening method (Jasmine 85=56.5%). PAA=Plant Area Affected.
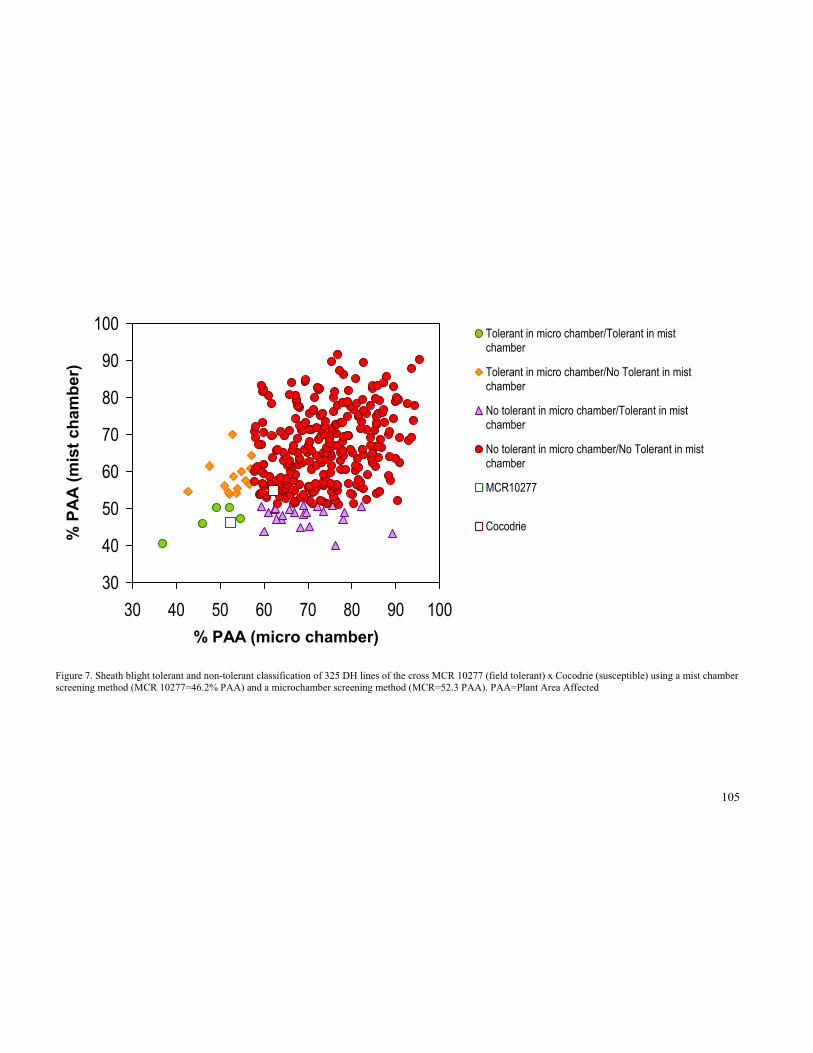
105
30
40
50
60
70
80
90
100
30 40 50 60 70 80 90 100% PAA (micro chamber)
% P
AA
(mis
t cha
mbe
r)
Tolerant in micro chamber/Tolerant in mistchamber
Tolerant in micro chamber/No Tolerant in mistchamber
No tolerant in micro chamber/Tolerant in mistchamber
No tolerant in micro chamber/No Tolerant in mistchamber
MCR10277
Cocodrie
Figure 7. Sheath blight tolerant and non-tolerant classification of 325 DH lines of the cross MCR 10277 (field tolerant) x Cocodrie (susceptible) using a mist chamber screening method (MCR 10277=46.2% PAA) and a microchamber screening method (MCR=52.3 PAA). PAA=Plant Area Affected

106
Figure 8. Quantitative Trait Loci (QTLs) controlling rice sheath blight tolerance in the genetic cross Jasmine 85 x Lemont using a mist chamber screening method. References Jia, Y., Correa-Victoria, F.J., McClung, A., Zhu, L., Wamishe, Y., Xie, J., Marchetti, M., Pinson, S., Rutger, N., and Correll. J. 2006. Rapid determination of rice cultivar responses to the sheath blight pathogen Rhizoctonia solani using a micro-chamber screening method. Plant Disease 91:485-489
2.A.2 Mite-fungus-bacterium disease complex in rice
� Identification of bacterial pathogens associated to the mite-fungus-bacterium disease complex
Contributors: Fernando Correa1, Gustavo Prado1, Edilia Mejia1, Girlena Aricapa1, Paola Fory1, Ismael Camargo2, Norman Oviedo3, Lazaro Narvaez4. (1CIAT-Colombia, 2IDIAP-Panama, 3SENUMISA-Costa Rica, 4INTA-Nicaragua)
Chromosome
AP28820.0
RM42815.2
RM116724.1RM22025.7RM126.3RM28327.5
RM535941.9RM49043.6RM25945.4
RM58063.2RM31268.1
RM44987.5
RM334198.9
RM5108.0
RM488113.7
RM246129.2
RM443134.5RM403139.3
RM128146.0
RM5501160.3
RM1361194.0
RM104202.4
qSB
-1
1
RM1090.0
RM27913.8
RM42323.7RM55527.5
RM7155.3RM691157.3RM45259.2RM42464.5
RM56183.0RM34186.3
RM47592.7
RM542798.1
RM3730131.5
RM263136.8RM526140.3
RM221148.9
RM530160.3
RM112167.3
RM250176.9
RM166184.7
RM208194.1RM498195.8
qSB-2-1
qSB-2-2
2
RM220.0
RM2317.2
RM48927.9
RM54536.1
RM51742.4OSR1346.3
RM759.0RM23259.4RM25160.0
RM28272.2
RM33886.7
M RG6395107.2M RG4864109.0RM156110.1RM16112.8RM5626118.2
RM426134.2
RM55142.0RM15824142.8
RM293159.6
RM468165.8RM422168.9RM143169.5
RM514179.9RM442184.1RM85185.1
qSB-3-1
qSB-3-2
3
RM5510.0RM3354.3
RM595320.9
RM18550.8
RM41759.4
RM14268.3
RM11975.5
RM355886.6
RM27393.3
RM3288107.9RM451110.2RM303110.5RM317111.0RM6748117.4RM255117.8
RM348132.0
RM124149.6
4
RM5070.0
RM57968.3RM41312.4RM1314.2
RM734924.7RM43729.7
RM28950.3RM16953.4
RM50959.9
RM16480.8
RM45989.1RM16193.8
RM540199.4
RM421107.6
RM26121.6
RM87127.5
qSB
-5
5
RM1330.0RM4351.7
RM1909.5RM58714.2RM51016.6RM22521.5
RM25337.1
RM13655.5RM343157.3
RM318364.7
RM54169.9RM719372.6
RM45485.2
RM16291.3
RM5371105.4
RM340121.2
RM103135.3qS
B-6
6
RM2950.0
RM575215.2
RM571121.7
RM12543.5
RM21453.0
RM258.9RM41863.4RM1166.2RM340469.6
RM45582.1
RM50595.5
RM234100.9RM478104.0
RM118110.0
RM1362125.8
RM248131.9
7
RM4080.0
RM1525.3
RM370212.0RM686313.6
RM114827.7RM2532.1RM31034.9
RM7242.1
RM40454.8
RM28485.8
RM21094.1
RM149109.1
RM230118.9RM433119.2RM477125.1RM447127.2
8
RM3160.0
RM2198.4
AP559316.4RM29619.7RM10522.8
RM40940.9
RM43448.8
RM25758.2
RM10867.2RM10771.1
RM21578.9
RM24590.3
qSB-9
9
RM4740.0
RM2225.9RM2169.7RM31112.5RM2528917.1
RM46727.7
RM27144.4
RM25851.7
RM22875.4RM48479.6RM14781.8RM59084.7
qSB
-10
10
RM2860.0
RM72039.9
RM20275.7RM53676.6
RM28783.4
RM22994.7RM2199.9RM3605101.8
RM206115.7
RM254125.3
RM224137.1
11
RM4150.0
RM34838.5
RM1915.5
RM24729.4
RM699844.8
OSM8951.2
RM27763.1
RM51969.8
RM46382.1
RM1300111.1RM270114.7RM17117.3
qSB
-12
12
Identification of the Quantitative Trait Loci (QTLs) responsible to sheath blight resistance based on the diseased plant area affected (PAA) in percentage in greenhouse using mist-chamber method
QTLs in red and green indicate the contributions from the resistant parent Jasmine 85 and the susceptible Lemont, respectively.
Guangjie et al., 2008

107
Funding
FONTAGRO, CIAT
Abstract The rice tarsonemid mite, Steneotarsonemus spinki has been recognized as an important rice pest in several Asian. The mite was detected in Cuba in 1997 causing severe yield losses, and subsequently was reported in the Dominican Republic, Haiti, Nicaragua, Costa Rica, and Panama causing 30 to 90% yield reduction. The mite was detected in Colombia in 2005 but in low populations with no significant yield reductions. The rice tarsonemid mite is normally found associated with the fungus Sarocladium oryzae, however, the main damage is caused in association with the bacterial panicle blight pathogen (Burkholderia glumae), which is detected during panicle emergence resulting in symptoms known as empty head or grain sterility. We have corroborated the presence of the bacterial panicle blight in Panama (2005 and 2006) and Colombia (2007). Bacterial Panicle Blight has been a recurrent problem in Panama, Costa Rica, Nicaragua, Colombia, and other rice producing areas of Central American and Caribbean countries in recent years. Our studies have provided convincing evidence of the association of this bacterium to the disease symptoms observed in all these countries. The incidence of the disease has increased in recent years probable due to the favorable climatic conditions and to the high percentage of infected seed being planted by rice farmers. Bacterial panicle blight has the potential to become a major constraint to rice production in Latin America. According to our studies, chemical seed treatment as well as improvement of tolerance to the disease will probably offer good management alternatives in the near future.
Keywords: bacterial panicle blight, Burkholderia glumae, Steneotarsonemus spinki, mite Background
The rice tarsonemid mite, Steneotarsonemus spinki was first recorded in Baton Rouge, La., USA in 1960. It has been recognized as an important rice pest in several Asian countries such as China, India, Taiwan, Korea, Philippines, and Thailand. The mite was detected in Cuba in 1997 causing severe yield losses, and subsequently was reported in the Dominican Republic, Haiti, Nicaragua, Costa Rica, and Panama causing 30 to 90% yield reduction. The mite was detected in Colombia in 2005 but in low populations with no significant yield reductions. Larvae and adults of S. spinki feed on the rice plant tissue causing browning of leaf sheaths and grain hulls. The rice tarsonemid mite is normally found associated with the fungus Sarocladium oryzae, however, the main damage is caused in association with the bacterial panicle blight pathogen (Burkholderia glumae), which is detected during panicle emergence resulting in symptoms known as empty head or grain sterility. We have corroborated the presence of the bacterial panicle blight in Panama (2005 and 2006) and Colombia (2007). Due to the severe economic losses caused by this disease complex in Central America and the Caribbean, FONTAGRO is funding the Project “Management of the disease complex mite-fungus-bacterium new challenge for Central American rice farmers”. We are reporting

108
this year the progress made on the identification and characterization of the bacterial pathogen present in this disease complex and identification of potential sources of resistance.
Materials and Methods Several visits to rice farmers’ fields in Panama, Costa Rica, Nicaragua and Colombia were made between 2005 and 2007 to observe and evaluate the presence of the disease complex in different rice farmer’s fields. Different plant parts including affected sheaths and rice grains were collected at different locations in each country and brought to the corresponding phytopathology or entomology labs for the detection and identification of possible organisms associated to the disease complex. To isolate the bacterial pathogen, diseased leaf sheaths or grains were rinsed in sterilized distilled water, and crushed in 5 ml of sterilized distilled water with a sterilized mortar and pestle. A dilution series of the resulting suspension was prepared and plated onto King’s B agar-medium and incubated at 28C for 2 days. Individual bacterial colonies were purified on the same medium and used for pathogenicity tests by suspending bacterial cells in sterile distilled water at 108 CFU/ml and injecting them into sheaths/stems of 20 days old seedlings or by spraying the bacterial suspension on the grains of recently emerged panicles. Inoculated plants were incubated under high humidity conditions for 24 hour in the case of stem injection or 4-6 days for panicle spraying. Screening for disease resistance was performed by injecting a bacterial suspension on 20 days old plants of different commercial rice cultivars and evaluated using a 1-9 scale developed in these studies on the basis of plant area affected. For the identification of the bacterium, two sets of Burkholderia glumae-specific primers were used: G1-13F (5’ACACGGAACACCTGGGTA3’) and G1-14R (5’TCGCTCTCCCGAAGAGAT3’) Takeuchi et al., 1997; and F (5’ACGTTCAGGGATRCTGAGCAG3’) and R (5’AGTCTGTCTCGCTCTCCCGA3’) Sayler et al., 2006. DNA extraction was performed using bacterial isolates grown on liquid King’s B medium for 24 hours. For the extraction, 1 ml of the bacterial suspension was used with the Wizard Genomic DNA purification kit Plus (Promega) following manufacturer instructions. The PCR reaction was performed according to the following description: 20 ng/ul of bacterial DNA, 5.0 ul of Buffer 10X, 2.5 Mm MgCL2, 200 uM dNTPs, 0.2 uM of each primer, and 1.25 U Taq polymerase, completing the final volume to 50ul with water. Amplification was performed in a thermal cycler PTC 100 with the following program: initial denaturation at 94C for 2 minutes, followed by 29 cycles consisting of denaturation at 94C for 1 minute, annealing at 56C for 1.25 minutes, extension at 72C for 1 minute, and an additional extension step of 72C for 7 minutes. The amplified products were visualized on 2 % agarose gels and stained with ethidium bromide.

109
Results and Discussion Field observations of infected plants included the following symptoms: small tan lesions with brown margins on the leaves and spikelets, resulting in abortion of infected kernels before completing fill. Infected florets may exhibit dark gray or brown bases, but the rachis of the panicle remains green. Prominent sheath lesions several centimeters long with a tan center and reddish brown margin may form on individual flag leaf sheaths of infected tillers. The panicles often remain erect, due to a loss of grain weight, instead of bending over (Figure 9). Affected panicles contain a mixture of green, tan, and brown kernels. The color of the spikelets is initially changed to grayish-brown and then straw color with sterility or abortion of kernels. Grain quality and milling are severely affected causing significant yield losses. All the symptoms observed are known as panicle blight. The observed grain discoloration, sterility, and abortion are thought to be due to the mite, Steneotarsonemus spinki, which is normally found in association to the disease complex, especially in the inner part of the leaf sheaths where high populations of nymphs and adults can be found. Small to medium size brown spots on the sheath are indicators of mite presence (Figure 10). The mite can be disseminated on seeds, by wind, water, insects, agricultural machinery, and survives on plant debris after harvesting. Heavy infestations of the mite occur during the rice reproductive stage helping to disseminate the bacterial panicle blight pathogen B. glumae causing erect or deformed panicles and turning the surface of grain brown to dark black. The sheath rot symptoms observed are similar to sheath rot symptoms caused by the fungus Sarocladium oryzae; however, this fungus is not always associated to the disease complex and with the bacterial disease a deep brown border surrounds the lesions. Our studies on microorganisms inhabiting the surface of infected leaves, sheaths and grains of rice cultivars under farmer fields’ conditions found many bacterial species as reported in similar studies conducted on bacterial panicle blight in the USA by Dr. M.C. Rush from Louisiana State University and who suggested the possible presence of the bacterial panicle blight pathogen, B. glumae in Panama. In collaborative work with Dr. Rush, the presence of B. gluame was corroborated. We isolated several strains of the bacterium which developed non-fluorescing colonies producing toxoflavin on King’s B medium (green pigment on medium) and which were found to be B. glumae by PCR analysis using two different sets of primers (Figure 11). All bacterial isolates identified as B. glumae were pathogenic and highly virulent in greenhouse inoculations causing typical disease rot symptoms on seedlings, leaves, sheaths and stems (Figure 12) and grains on inoculated panicles (Figure 13). All observed symptoms were similar to the symptoms observed in the field. We have isolated the pathogen from seeds from diseased panicles suggesting that it is seed borne. This bacterium is known in Japan as a grain-rotting bacterium that causes seedling rot and grain rot. Twenty antibacterial compounds were tested for their efficacy against B. glumae. These included antibiotics, copper containing compounds, organic compounds of acid characteristics, and fungicides used on rice. The pesticide that appeared to give the best control either in in-vitro tests inhibiting bacterial growth or panicle blight control on

110
inoculated panicles was oxolinic acid (Figure 14). This antibiotic is being tested on field trials conducted in several farmer fields in Panama, although initial tests gave no conclusive results on the effectiveness of this chemical, probably due to the high percentage of infected seeds being planted. Further tests will be conducted under the Project to determine the effectiveness of oxolinic acid as seed treatment given that most farmers plant their own seed, which is normally highly infested with B. glumae. Development of severe disease symptoms appears to be associated with unusually hot weather and especially warm nights with increases of the minimal temperature by 1C. In Colombia, symptoms of bacterial panicle blight have been observed since 2005 although the presence of the bacterial panicle pathogen was only demonstrated in 2007. Comparison of the historical minimum temperature between 1993 and 2005 made by FEDEARROZ (Enrique Saavedra and Cristo Perez) in Montería, Cordoba in Colombia where the disease caused severe yield losses between 2005-2007, demonstrates that the minimal temperature in 2005 has in average 1C increase compared to the historical record (Figure 15). This observation could suggest that B. glumae is the cause of sporadic occurrence of panicle blight and that it has the potential to become a major constraint to rice production where record of high temperatures might occur. Because bacterial panicle blight is a new disease affecting rice production in Latin America, this Project considered the development of a screening method, development of a disease evaluating scale, and the identification of potential sources of resistance as an alternative for future management of the disease. A scale with disease scores from 1 to 9 was developed for green house inoculations, where 1-3 represents resistance to the disease, 4-5 intermediate levels of resistance, and 6-9 susceptibility to the pathogen (Figure 16). The bacterial inoculation method consisted of injecting a bacterial suspension of 108 CFU/ml on the stems of 20 days old rice plants and incubating them for 24 hours under high humidity conditions. Inoculated plants were left on greenhouse benches until evaluation 4-6 days after inoculation. We have tested more than 200 commercial rice cultivars from Latin America and breeding lines in greenhouse inoculations for resistance to B. glumae. The bacterium was highly virulent on most of the rice cultivars and lines tested. None of the entries exhibited complete resistance to the pathogen; however some of them showed intermediate reactions to the infection (Table 1, Table 2). It has been reported that rice varieties differ in how this disease develops and in yield losses. Most Colombian commercial rice cultivars exhibited a susceptible reaction; however the cultivars Oryzica Llanos 5 and Selecta 320 and the elite breeding lines from FLAR (FL 03174 and FL 03188) had an intermediate reaction (Table 1). Similarly, although most commercial rice cultivars from Panama, Costa Rica and Nicaragua, countries participating in this FONTAGRO Project, were susceptible to the bacterial pathogen, some cultivars exhibited an intermediate reaction (Table 2). Other rice cultivars with intermediate reaction to the bacterial panicle blight that could be potential sources of tolerance to the pathogen were also found in greenhouse inoculations of germplasm form Bolivia, Brazil, Cuba, Ecuador, Guatemala, Mexico, Peru, Salvador, Venezuela, Sri Lanka, Guyana Francesa, Filipinas and Japan, and some inter-specific lines (IRGC 100967) from the CIAT rice breeding program (Table 3). Some of these
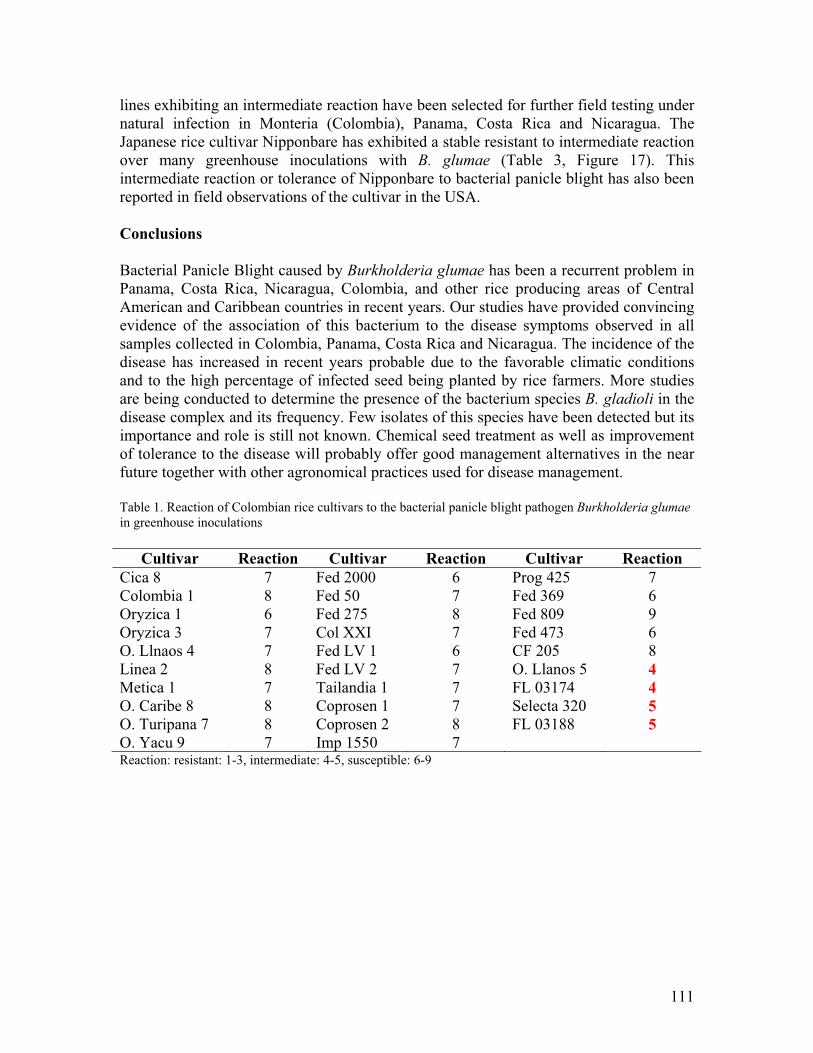
111
lines exhibiting an intermediate reaction have been selected for further field testing under natural infection in Monteria (Colombia), Panama, Costa Rica and Nicaragua. The Japanese rice cultivar Nipponbare has exhibited a stable resistant to intermediate reaction over many greenhouse inoculations with B. glumae (Table 3, Figure 17). This intermediate reaction or tolerance of Nipponbare to bacterial panicle blight has also been reported in field observations of the cultivar in the USA. Conclusions Bacterial Panicle Blight caused by Burkholderia glumae has been a recurrent problem in Panama, Costa Rica, Nicaragua, Colombia, and other rice producing areas of Central American and Caribbean countries in recent years. Our studies have provided convincing evidence of the association of this bacterium to the disease symptoms observed in all samples collected in Colombia, Panama, Costa Rica and Nicaragua. The incidence of the disease has increased in recent years probable due to the favorable climatic conditions and to the high percentage of infected seed being planted by rice farmers. More studies are being conducted to determine the presence of the bacterium species B. gladioli in the disease complex and its frequency. Few isolates of this species have been detected but its importance and role is still not known. Chemical seed treatment as well as improvement of tolerance to the disease will probably offer good management alternatives in the near future together with other agronomical practices used for disease management. Table 1. Reaction of Colombian rice cultivars to the bacterial panicle blight pathogen Burkholderia glumae in greenhouse inoculations
Cultivar Reaction Cultivar Reaction Cultivar Reaction Cica 8 7 Fed 2000 6 Prog 425 7 Colombia 1 8 Fed 50 7 Fed 369 6 Oryzica 1 6 Fed 275 8 Fed 809 9 Oryzica 3 7 Col XXI 7 Fed 473 6 O. Llnaos 4 7 Fed LV 1 6 CF 205 8 Linea 2 8 Fed LV 2 7 O. Llanos 5 4Metica 1 7 Tailandia 1 7 FL 03174 4O. Caribe 8 8 Coprosen 1 7 Selecta 320 5O. Turipana 7 8 Coprosen 2 8 FL 03188 5O. Yacu 9 7 Imp 1550 7 Reaction: resistant: 1-3, intermediate: 4-5, susceptible: 6-9

112
Table 2. Reaction of rice cultivars from Panama, Costa Rica and Nicaragua to the bacterial panicle blight pathogen Burkholderia glumae in greenhouse inoculations
Cultivar Reaction Cultivar Reaction Cultivar Reaction Panama Costa Rica Nicaragua
Anayansi 6 CR 1113 7 Altamira 9 6 Damaris 7 CR 201 7 FL 01119 6 Pan 1048 8 CR 1821 7 Altamira 7 5Pan 1537 6 CR 5272 7 Altamira 10 5Pan 3621 6 FL 03160 6 Uni 3189 6 CR 1707 5 Toc 5430 7 FL 00447 7 Idiap L-7 7 Pan 4721 5 Reaction: resistant: 1-3, intermediate: 4-5, susceptible: 6-9 Table 3. Potential sources of tolerance to the bacterial panicle blight pathogen Burkholderia glumae in greenhouse inoculations
Cultivar Country Reaction Cultivar Country Reaction Saavedra Bolivia 5 Bamoa A 75 Mexico 5 San Pedro Bolivia 5 Culiacan A 82 Mexico 5 Paititi Bolivia 5 Chetumal A 86 Mexico 5 Br Irga 410 Brasil 5 Palizada A 86 Mexico 5 Epagri 108 Brasil 5 Altamira 7 Nicaragua 5 O. Llanos 5 Colombia 4 Altamira 10 Nicaragua 5 Selecta 320 Colombia 5 Panama 4721 Panama 5 FL 03174 Colombia 4 Inti Peru 5 FL 03188 Colombia 5 Alto Mayo 88 Peru 4CR 1707 Costa Rica 5 San Martin 86 Peru 4Perla Cuba 5 Centa A-6 Salvador 5 Iniap 11 Ecuador 5 Venezuela 21 Venezuela 5 Tikal 2 Guatemala 5 BG 90-2 Sri Lanka 5 Icta Pasos Guatemala 5 FL 00867 G. Francesa 5 I. Motagua Guatemala 5 IR 36 Filipinas 5 FL 03197 Guatemala 5 Nipponbare Japan 4FL 03232 Guatemala 5 IRGC 100967 CIAT1 4Reaction: resistant: 1-3, intermediate: 4-5, susceptible: 6-9. 1Interspecific line

113
Figure 9. Bacterial panicle blight symptoms caused by Burkholderia glumae in rice fields. Affected panicles contain a mixture of green, tan, and brown grains and panicles often remain erect. Sheath lesions with a tan center and reddish-brown margin may form on individual leaf sheaths
Figure 10. The rice tarsonemid mite Steneotarsonemus spinki feeds on the rice plant tissue causing browning of leaf sheaths and grain hulls. High populations of adults of the mite can be observed in the inner part of infested leaf sheaths

114
Figure 11. Typical non-fluorescent colonies of Burkholderia glumae (upper-left) and toxiflavin pigment produced by the bacterium on King’s B medium (botton-left). PCR amplification products of 400 bp and 280 bp used for identification of B. glumae (upper-right; 17: B. glumae, 70: B. gladioli, 83: Pseudomonas fluorescens, 95: B. glumae, 107: Acidovorax avenae). PCR amplification product (1492 bp) with universal Burkholderia primers (17: B. glumae, 60: B. glumae, 68: B. gladioli, 70: B. gladioli, 73: B. glumae, 94: Acidovorax avenae, 98: B. glumae)
17 60 68 70 73 94 98 (-)
1650pb
1kb 17 70 83 95 107 (-) 1kb 17 70 83 95 107 (-) 1kb
400pb 300pb

115
Figure 12. Reproduction of typical symptoms caused by the bacterium Burkholderia glumae pathogen on leaves, sheaths, and stems in green house inoculation of young rice plants by injecting a bacterial cell suspension
Figure 13. Reproduction of typical symptoms caused by the bacterium Burkholderia glumae pathogen on seeds in green house inoculation of emerging rice panicles by spraying a bacterial cell suspension

116
Figure 14. In-vitro inhibition of the bacterium Burkholderia glumae (left) and control of disease development on bacterial inoculated panicles (right) by the antibiotic oxolinic acid Figure 15. Comparison of minimal temperatures in 2005 and historical record between 1993 and 2004. FEDEARROZ, Monteria, Colombia
T° MINIMA 2005 E HISTORICO 1993-2005
22,0
22,5
23,0
23,5
24,0
24,5
25,0
25,5
Enero
Febrer
oMarz
oAbri
lMay
oJu
nio Julio
Agosto
Septie
mbre
Octubre
Noviembre
Diciem
bre
MES
°C
2005PROM 1933-2005

117
Figure 16. Evaluating scale of disease symptoms caused by Burkholderia glumae on rice where 1-3 is resistant, 4-5 is intermediate reaction and 6-9 is susceptible
Figure 17. Consistent resistant to intermediate reaction of the rice cultivar Nipponbare (left) compared to the susceptible reaction of the control (right) References Nandakumar, R., Rush, M.C., and Correa-Victoria, F.J. 2007. Association of Burkholderia glumae and B. gladioli with Panicle Blight Symptoms on Rice in Panama. Plant Disease 91:767 Correa-Victoria, F.J., Perez, C.R., y Saavedra, E. 2007. Añublo bacterial de la panicula del arroz Burkholderia glumae. ARROZ 57(468):26-32 Correa-Victoria, F.J. 2006. Situación del complejo acaro-hongo-bacteria en el arroz. Segundo Congreso Arrocero. San José, Costa Rica, Junio 29-30, 2006. (invited speaker) Correa-Victoria, F.J., 2007. The rice Tarsonemid mite: Steneotarsonemus spinki Smiley. RiceCAP Fact Sheet. 4 pages. www.ricecap.uark.edu/outreach_downloads.htm

118
Takeuchi, T., Sawada, H., Suzuki, F., and Matsuda, I. 1997. Specific detection of Burkholderia plantarii and B. glumae using primers selected from the 16S-23S rDNA spacer regions. Ann. Phytopathol. Soc. Jpn 63: 455-462 Sayler, R.J., Cartwright, R.D., and Yang, Y. 2006. Genetic characterization and real-time PCR detection of Burkholderia glumae, a newly emerging bacterial pathogen of rice in the United States. Plant Disease 90: 603-610 2.B RICE VIROLOGY
2.B.1 Screening Rice for Resistance to Rice Hoja Blanca Virus and Tagosodes orizicolus Lee Calvert and Luis Reyes Bullet highlight
1. Many varieties with resistance to RHBV have been develop using two stage field screening method. Even if molecular markers are developed to replace the mass screening, the small plot screening using different levels of infestation will be needed for advanced lines.
2. This has been a transition year for the field evaluation for resistance to RHBV and the screenhouse evaluation for tolerance to mechanical damage to T. orizicoulus. The transition from a core activity to a self supporting service is nearly complete.
Abstract
The varieties that have proven to be resistance to RHBV also have good resistance to T.orizicolus. Currently this involves mass screening of many lines with few or no replications in field trials using viruliferous vectors of RHBV. Screening the apparently resistance lines for tolerance to mechanical damage caused by T. orizicolus. Finally the advanced lines, which have been selected for many different characteristics, are tested under a small plot design with multiple replications and infestation levels. The mass screening is the least precise and it is difficult to maintain an intermediate level of disease pressure. High levels of disease pressure eliminate too many of the breeders materials and intermediate resistance to RHBV has proven to be a valuable deterrent to epidemics of RHBV. Until an alternative method using molecular markers is proven select RHBV resistance rice lines, the screening capacity needs to be maintained. Even after molecular markers are implemented advanced lines will need to be tested using this biological assay.
Background
The planthopper, Tagosodes orizicolus, is one of the most important pests of the rice in the tropical America. It causes direct damage to the plant through phloem feeding and oviposition of its eggs. Also, it is the vector Rice Hoja Blanca Virus (RHBV), which has

119
caused epidemics in 1935, 1939, 1956-1961, 1982, 1990 and 1996-98. In 1982, a major epidemic caused losses of 50%-70% in the variety Cica 8 (Vargas, 1985). In the decade of the nineties epidemics occurred in Peru, Costa Rica, Colombia, Dominican Republic and Venezuela. When RHBV is present, farmers often use excessive insecticides to control the vector T. orizicolus. Indiscriminate application of insecticides eliminates the natural enemies of T. orizicolus, which leaves the crop vulnerable to attacks by various insects and increases the likelihood of severe epidemics of RHBV.
Within rice germplasm, there is resistance to both RHBV and T. orizicolus. Most of the varieties with resistance to RHBV also are tolerant to mechanical damage by T.orizicolus. Since 1998, more than ten varieties with resistance to both RHBV and T.orizicolus have been released. The first evaluations are made in the early generations when the populations are segregating. Since the RHBV massive field evaluation use only one to three replications, the data is best used to eliminate the most susceptible lines. Those materials are then screened in the greenhouse to determine their resistance to mechanical damage caused by the planthopper. Further testing using fixed populations must be done using replicated small plots to assure that the selected lines are resistant to RHBV.
CIAT evaluates materials for RHBV and planthopper resistance for the varietal development activities for CIAT, FLAR, FEDEARROZ, commercial producers of seeds and national programs. This year, through training at CIAT, Danac of Venezula has set up its own colonies of T. orizicolus and is implementing this system to evaluate their materials for both RHBV and T. orizicolus resistance.
Materials and Methods
Prescreening of the vector colonies. One month before the main RHBV field evaluation, two colonies were tested to determine the best dosage for the RHBV field evaluation. The susceptible variety Bluebonnet 50, the intermediate resistant variety Fedearroz 50, the resistant variety Colombia 1 and the highly resistant variety Fedearroz 2000 were used in the prescreening and as checks in the main evaluation. These materials were planted in rows of 50 cm long and 15 cm apart, with a density of 1.5 grams per row in small plots.
Evaluations of resistance to RHBV. Evaluations to incorporate sources of resistance to RHBV were conducted twice in 2007. The vector colony was determined to contain approximate 65-78% viruliferous insects. One and half grams of seed of each test line (ca 50 seeds/line) are sown in 0.5 mts row spaced 15 cm apart. Bluebonnet 50, Fedearroz 50, Colombia 1 and Fedearroz 2000 were planted as the control lines. At 18 days after planting, the insects were distributed evenly onto the lines. Five days after the infestation for the first semester and three days in the second semester, an application of imidacloroprid was applied. Plants were evaluated for RHBV symptoms 30- 35 days after infestation using a visual damage scale (0-9). The controls were used to calibrate the results.

120
Screening for resistance to mechanical damage: Evaluations for resistance to T. orizicolus mechanical damage were conducted in the greenhouse. The lines were planted in trays containing sterilized soil in rows of 10 plants spaced 3 cm apart. In each tray the resistant check Makalioka, the intermediate check Cica 8, and the susceptible check Bluebonnet 50 were planted in a random order. After 15 days, the trays were transferred to cages and infested with nymphs of T. orizicolus, with an average of ten insects per plant. The insects fed on the plants until the death of Bluebonnet 50. At that time, the other materials were evaluated according to the following scale: Resistant of 0-1 dead plants; Intermediate of 2-5 dead and Susceptible 6-10 dead plants. The final evaluation is made using a visual scale 1: No symptoms or slight discoloration; 2: chlorosis of the tip and borders of the leaves; 3: half of the plant with chlorosis or necrosis; 4: Stunting with more than half of the plant necrotic and the percentage of dead plants. Results and Discussion Evaluations of resistance to RHBV and T. orizicolus
Through the prescreening, it was determined that the vector colony was highly viruliferous. The first semester, 1.2 insects per plant were used for the infestation. The plots were treated with insecticide at five days after infestation. The pressure was rated as high and this can cause the elimination of lines with intermediate resistance. Since there was not time to start a new colony for the second semester, the level of infestation was 1.0 insect per plant and the plots were treated at three days after infestation. This reduced the pressure although the level of disease pressure was still considered to be high. The lines that were identified as resistant during this year should be excellent candidates for varieties with resistance. The RHBV and T. orizicolus screening are in the process of becoming a self supporting service activity. A user fee was charged to everyone who used the service. This is reflected in the reduction in the number of lines that were evaluated. In the first semester 6856 lines were evaluated, and in the second semester 7890 lines were evaluated. The number of lines that were evaluated for resistance to mechanical damage was less than 60% of the level of previous years. A total of 867 lines were evaluated. The high disease pressure in the RHBV screening were similar to the levels that were used from 1997-2000. This is the time when many of new varieties released were resistant to RHBV. These varieties included Fedearroz 50, Fedearroz Victoria 1, Fedearroz Victoria 2, and Fedearroz 2000. The resistance scale is adjusted each season to reflect the disease pressure and to include lines with intermediate resistance. In 2007, all lines rate 1-5 were rated as highly resistance. Normally the range is 1-3 for highly resistant lines. A new colony has been established and this should allow the lowering of disease pressure in the subsequent semesters. Also, experiments in the prescreening will be done to determine the effectiveness of using different infestation periods as another means of controlling the levels of disease pressure.

121
Conclusions and Future Activities
This is a transition year for the evaluation of resistance to RHBV and mechanical damage caused by T. orizicolus. The total cost of the activity has been reduced by more than 50%. The external demand by private companies, which have been charged for the service for many years remained constant. The number of lines evaluated for FLAR, FEDEARROZ and CIAT all were reduced. During 2008, it is expected that the demand for resistance screening will be higher than 2007, and this will allow this activity to be self sustaining.
2.B.2 Developing Molecular Markers for Resistance to Rice Hoja Blanca Virus and Tagosodes orizicolus. L.Calvert, I. Lozano, L. Romero, M. Lorieux, Natalia Villareal,
Luis Reyes and Cesar Martinez. Bullet highlights
1. In Fedearroz 50 and Fedearroz 2000, there is a QTL associated with resistance to RHBV on the short arm of chromosome 4.
2. In Fedearroz 2000, there is a QTL associated with resistance to RHBV on chromosome 5. This QTL is not present in Fedearroz 50.
3. In Fedearroz 50, there is a QTL associated with resistance to T. orizicolus on chromosome 7.
Abstract Rice hoja blanca disease occurs in cyclic epidemics that can cause very severe losses. Since rice hoja blanca virus (RHBV) can only be transmitted using its planthopper vector Tagosodes orizicolus, it has made breeding for resistance difficult and expensive. The variety Fedearroz 2000, which is highly resistant to RHBV and has intermediate resistant to T. orizicolus, was crossed the highly susceptible line WC366, and 235 F2 lines were generated. The variety Fedearroz 50, which has intermediate resistance to RHBV and is highly resistant to T. orizicolus, was crossed with WC366, and 291 F2 lines were generated. To phenotype the F2 plants, their F3 populations were tested for their resistance to mechanically damage by T. orizicolus and to their resistance to RHBV. Bulk analyses of F3 populations were evaluated using microsatellites. For Fedearroz 2000, two QTLs were associated for resistance to RHBV. For Fedearroz 50, one QTL was associated for resistance to RHBV on the short arm of chromosome 4 and one QTL was associated for resistance to T. orizicolus on chromosome 7. Further saturation with microsatellites and completion of analysis of all the lines is being done to determine if several additional potential QTL are real. This comparative study using two distinct populations is allowing a better understanding the genetics of resistance to RHBV and its vector.

122
Key words: Rice Hoja Blanca Virus, T. orizicolus, resistance, micro-satellites, QTLs
Background Rice hoja blanca virus causes cyclic epidemics which cause great losses. In order to develop varieties with resistance, colonies of viruliferous T. orizicolus were established in the mid 1980s. Breeding for RHBV method is a two-step process. In the early generations the segregating populations are tested using up to three rows of plants. We recommend using short rows with more repetitions so that two levels of vectors can be applied to the mass screening. This only eliminates the most susceptible lines but does not assure that the subsequent progeny will be resistant to RHBV. After the lines are more advanced and the traits are fixed, the materials are tested in a randomized block design using different ages and vector pressures. The screening done at CIAT has been part of the rice breeding for Fedearroz, FLAR as well as other organizations. Fedearroz has successfully released several varieties with resistance to RHBV including Fedarroz 50, Fedarroz 2000, Fedearroz Victoria 1, and Fedearroz Victoria 2. While the mass screening for resistance to RHBV has proven a useful method for developing resistance varieties, there are problems with consistency of results because of lack of replications of each line and variability of the colonies. To properly characterize the level of resistance, replicated randomized block trials using different levels of infestation pressure need to be done. This is only feasible for a limited number of nearly finished lines, and is recommended before the lines are released as varieties. Also, the colonies are costly to maintain. Molecular studies were made to identify QTLs associated with resistance to RHBV and T. orizicolus. Crosses were made between the highly susceptible line WC 366 and Fedearroz 2000. A second cross was made between WC 366 and Fedearroz 50. The aim of this investigation is to develop a molecular-marker assisted program to replace the massive RHBV screening. Even if we are highly successful in developing effective markers, the advanced lines will still need to be screened biologically in replicated trials to determine their resistance to RHBV. To better understand the genetics of resistance and identify potential markers for resistance to RHBV and its vector T. orizicolus. Material and Methods
Plant populations.
The parents for the crosses were chosen for their genetic distant as well as their reaction to RHBV an to T. orizicolus. The line WC366 is susceptible to both RHBV and to T.orizicolus. The variety Fedearroz 50 has intermediate resistance to RHBV and is highly resistant to T. orizicolus. The variety Fedearroz 2000 is resistant to RHBV and has intermediate resistance to T. orizicolus.

123
Cross 1. The F1, F2 y F3 populations of crosses between Fedearroz 2000 (CT10323-29-4-1-1T-20) was crossed with WC 366 (IR65598-27-3-1). From 235 plants of the F2 population, the F3 seed was collected. Cross 2. The F1, F2 y F3 populations of crosses between Fedearroz 50 was crossed with the susceptible line WC 366 (IR65598-27-3-1). From 291 plants of the F2 population, the F3 seed was collected.
Evaluation for RHBV.
Ten rows each with 20 plants for a population of 200 were used for the evaluation of the F2 generation as well as for 235 lines of the F3 generation. Twenty-five days after planting, the rice lines were infested with a dosage of 1.5 insects per plant using viruliferous insects from the colony “Costa-CIAT”. Five days after the infestation, the insects were collected and tested using ELISA in order to determine the virulence of the insects used in the evaluation. The remaining insects were eliminated and each week for four weeks the plants were evaluated for symptoms of RHBV.
Evaluation for resistance to T. orizicolus.
Each row consists of twenty plants of a line, and after 15 days, they are infested with approximately 10 T. orizicolus nymphs per plant. When the susceptible check Bluebonnet 50 dies, the lines are evaluated . Cica 8 was the intermediate control and Makalioka was the resistant control. The evaluation scale was a standard scale developed by IRRI (1996).
Micro-satellite analysis.
For the micro-satellite analysis, 20 F3 plants from each of the 235 lines were grown. From each of the 231 populations that successfully germinated, 200 mg of tissue from each plant was collected at 20 days after planting. For each line the tissue sampled were placed together to make up the bulk samples. The samples were frozen in liquid nitrogen and stored at –80C until processed. The DNA was isolated and quantified. The PCR reactions for the micro-satellites were made with the commercial primers and the PCR products visualized with silver staining of the 6% polyacrylamide, 7M urea gels. The analysis calculated the distances with interval map using the program MapDistov17b.
Results and Discussion Evaluation of the crosses. Bulk analysis of at least 20 F3 plants of both crosses were evaluated using microsatellites. The analysis reported was done using the 30 most susceptible and 30 most resistance lines from each cross.

124
To date, 78 microsatellite markers (figure 1A) have been used to evaluate the Fedearroz 50 X WC366 cross. For resistance to mechanical damage by T. orizicolus, a highly significant QTL has been identified. A single point analysis was made to identify and quantify the strength of potential QTL for resistance to RHBV and T. orizicolus (figure 2). Even with the addition of more markers, the only QTL associated for resistance to RHBV is on the short arm of chromosome 4. For resistance to T. orizicolus, the only QTL identified is on chromosome 7. To date, 108 microsatellite markers have been used to evaluate the Fedearroz 2000 X WC366 cross. A single point analysis was made to identify and quantify the strength of potential QTL for resistance to RHBV and T. orizicolus (figure 3). For resistance to mechanical damage caused by T. orizicolus, no strong association has been made with any of the microsatillites tested. On chromosome 5 and 6, there is some weak associations, but no QTL has yet to be identified. For resistance to RHBV, a QTL on the short arm of QTL has been identified. Also a weak association has been made for resistance on chromosome 5. Conclusions and Future Activities
The main objective of this research is to develop a marker aided selection for resistance to RHBV. To better localize the QTL in the short arm of chromosome 4 additional markers have been used for both crosses. Still more needs to be done to fine map the QTL with the objective of identifying the resistance gene. Selected References
Calvert, L.A., Lozano I., Villareal N., Romero L., Lorieuz M., Martinez C., Garavito A., 2006. Molecular analysis of the genetics of resistance to rice hoja blanca virus and its vector Tagosodes orizicolus. Phytopathology 96:S18 D.A. Cartwright, M. Troggio, R. Velasco, A. Gutin. 2007. Genetic Mapping in the Presence of Genotyping Errors. Genetics: Published Articles Ahead of Print, published on February 4, 2007 as 10.1534/genetics.106.063982. Lorieux, M. 2007. MapDisto, A Free User-Friendly Program For Computing Genetic Maps. Computer demonstration given at the Plant and Animal Genome XV conference, Jan 13-17 2007, San Diego, CA. URL: http://mapdisto.free.fr. Lincoln S, Daly M, Lander ES (1992) Constructing genetic maps with MAPMAKER/EXP 3.0, 3rd edn. Whitehead Institute Technical Report, Cambridge.
McCouch SR, Teytelman L, Xu YB, Lobos KB, Clare K, Walton M, Fu BY, Maghirang R, Li ZK, Xing YZ, Zhang QF, Kono I, Yano M, Fjellstom R, Declerck G, Scheider D, . Wang, S., C. J. Basten, P. Gaffney and Z-B. Zeng, 2004 Windows QTL Cartographer 2.0 User Manual (http://statgen.ncsu.edu/ qtlcart/WQTLCart.html).

125
Figure 1. The 78 markers that were used to analyse the cross Fedearroz 50 X WC366.
Figure 2. The 108 markers that were used to analyse the cross Fedearroz 2000 X WC366.
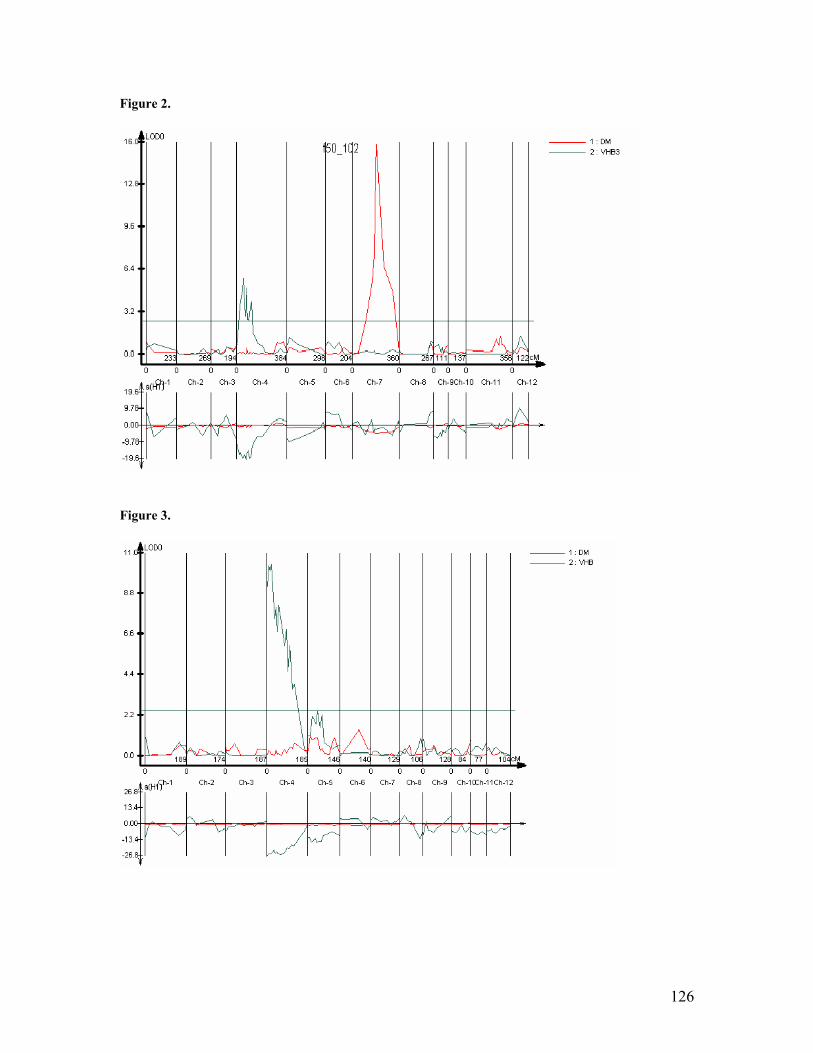
126
Figure 2.
Figure 3.

127
2.C Resistance RHBV and Selection of Advanced Breeding Generation from Crosses with Transgenic Rice Resistant to RHBV in the Field in 2007M. Quintero1, L. Fory2. J. Borrero1, E. Bolaños1, T. Agrono1, E. Tabares2 C. Martínez 1, 2,and Z. Lentini1, 2 .1IP4, 2SB2. Funding The Rockefeller Foundation and CIAT Abstract The Rice Biotechnology Project had generated transgenic rice lines with resistance to RHBV (Lentini et al., 2003) an endemic disease and one of the main constrains of rice in Tropical America. Resistant transgenic lines carrying different source of resistance than the one already deployed in commercial varieties would allow increased production to a lower cost by reducing the need of insecticide applications commonly used to control Tagosodes oryzicolus (the virus vector). After stepwise selection from the original transgenic plants, including elimination of un-stable inheritance of RHBV resistance and plants with undesirable agronomic traits, the best lines were crossed with various varieties and derived progenies had been subjected to a breeding selection process in the field including evaluation for yield potential, other disease resistance (Rhizoctonia and Pyricularia), agronomic and grain quality traits. This report describes the final selection of the best lines, step after which these lines should be evaluated by third party and decision should be made on the fate of these materials for potential deployment to farmer fields. Key Words: transgenic rice, RHBV, yield potential, field, anther culture
Background
The efficiency and scope of research programs in rice can be increased by access to the tools of biotechnology as transformation. Transgenic rice lines highly resistance to RHBV were generated at CIAT (Lentini et al., 2003). The transgenic RHBV resistant rice was generated by splicing in genes from the RHBV virus into Cica 8 variety (Lentini etal., 2003). The breeders selected Cica 8 as potential parent donor of the transgene for future conversion of modern varieties, because of its good grain quality, high productivity, and broad adaptation including low soil humidity, and acceptance to large and small resource farmers. Large farmers because of its high susceptibility to RHBV do not currently grow this variety commercially. In spite of its high susceptibility to RHBV, this variety is still of preference by small farmers due to its good agronomic qualities and resilience to stress conditions. The transgenic resistance incorporated in Cica 8 is RNA-mediated and some plants show hypersensitive reaction when challenged with the RHBV virus (Lentini et al., 2003). Some of the transgenic lines outperform in RHBV resistance to most currently grown commercial rice varieties, except to variety Fedearroz 2000 that although is highly resistant to the virus, it has poor environmental adaptability, which has restrain its commercial adoption by farmers. Attempts to transfer the RHBV transgenic resistance into other modern varieties through regular crossing indicated that the RHBV-N transgene is inherited and expressed independently of the genotype background. This transgenic RHBV resistance source might complement the breeding resistance source that was released through conventional breeding. The transgenic rice is resistant even when plants are younger than 5 to 20 day-old. By combining different RHBV-resistance

128
sources, it might be possible to lower production costs by reducing the need of insecticide applications common in the control this viral disease transmitted by the sogata (Tagosodes oryzicolus). In addition to selection for RHBV resistance and yield potential, the advanced transgenic lines were evaluated for tolerance to Rhizoctonia, resistance to Pyricularia, and grain quality traits. Progress in previous years led to a shift from the proof of concept towards breeding, targeting the selection of improved materials for an eventual commercial release. Previous reports described the stepwise breeding selection in the field. Last year we reported the evaluation and selection for RHBV resistance in the field, and final selection based on their RHBV resistance, tolerance to sheath blight and blast, grain quality and agronomic traits, and yield potential of T7 and T10 advanced transgenic lines, and F6 progeny plants derived from crosses between the transgenic lines and commercial varieties. This year we report the final evaluation for RHBV resistance of the lines selected in 2006. The selected lines will be evaluated for their performance in replicated yield trials, and processed through anther culture. Materials and Methods Evaluation of RHBV resistance in the field in 2007. Field evaluations were conducted using 8 T8 and T11 advanced transgenic lines, and 5 F7 families derived from crosses between the transgenic lines and the commercial variety Oryzica 1. The selection of lines was based on its agronomic performance in the field in 2006. A row with 40 plants per line with 5 replicates was used for the RHBV evaluation. Controls consisted of RHBV resistant plants (Colombia 1, and Fedearroz 2000) and susceptible checks (Bluebonnet 50, Cica 8, Oryzica 1, Fedearroz 50, a cross Cica 8/ Fedearroz 50, and Cica 8/ Oryzica 1). Eighteen days after planting, plants were infested with 1.5 viruliferous insects per plant using insects from the colony “Jamundi-CIAT”. Five days after the infestation, the insects were killed, and the plants were evaluated for disease symptoms development at 30 and 45 after infestation. Agronomic evaluation and yield potential test of advanced generations of transgenic events and derived progeny plant from crosses. Sister plants of the same lines evaluated for RHBV resistance were also evaluated for agronomic performance and yield potential in the field. Plants were transplanted in the field using 12 rows per lines of 21 plants per rows with 3 replicates. Agronomic traits are evaluated throughout the life cycle up to maturity, including tiller number, plant height, plant vigor, days to flowering, fertility, and yield. Agronomic traits are evaluated according the IRRI scale (1996). Yield potential will be evaluated by analyzing the harvest from four central rows and taking into account the grain humidity (approximately 20%).
Results and Discussion Cluster analysis of advanced crosses or self-cross transgenic lines using principal coordinates were conducted using data from the RHBV resistance evaluations with one

129
dosage of 1.5 insects per plant and 5 replicates. Only the F7 generation line A3-49-101-18-19/Oryzica 1-14-M-10-2-4-6 derived from crosses with Oryzica 1 was clustered in Group 4 jointly with Fedearroz 2000 showing the highest level of RHBV resistance (score � 3.8) (Figure 1 and Table 1). In addition, several lines derived from crosses and lines derived from backcross to Cica 8, were grouped in cluster 3 and 5 near to Fedearroz 2000 with average values � 4.6. The RHBV resistance of these crosses and transgenic lines were significantly different from their corresponding non-transgenic controls (the controls clustered in other groups). The non-transgenic cross Cica 8/Oryzica 1 score � 6.6, and the non-transgenic control varieties Cica 8, score � 8.2 and Oryzica 1, score � 7 were clustered in other groups with susceptible materials likewise the controls Cica 8/Fedearroz 50, score � 7; and Fedearroz 50, score � 7.4 (Figure 1 and Table 1). The susceptible transgenic control line A3-78-24 (which does not contain the RHBV transgene, transgene null internal control) show the highest susceptibility as in earlier evaluations (score = 8.6). This RHBV test in the field was more stringent than other evaluations from previous years, in general control showed higher levels of susceptibility. Colombia 1 (usually with intermediate resistance) was highly affected (score 5.8), and it was grouped with non-transgenic line Cica 8 (susceptible, score 8.2). The higher levels of susceptibility seen this year may have been due to a higher virulence level of the insect colony. Based on this RHBV resistance profile, and the agronomic performance including yield potential, previous evaluations for sheath blight and blast resistance as well as for grain quality traits, the best lines will be selected and process Anther Culture. Future Activity
The selected lines are currently being processed through anther culture to generate doubled haploid (complete homozygous) lines. The DH transgenic RHBV resistant lines will be multiply and use in replicated multi-location field trials and molecular genotyping. Anther culture offers a rapid method of generating homozygous lines of rice (Oryza sativa L.) for breeding program and genetic analysis. For anther culture, the immature panicles re harvested and cold pre-treated, then the anthers are dissected and cultured in vitro according to Lentini et al (1995). Anthers are culture in liquid medium M1 (Quintero et al., 2003) contained in baby food jars closed with perforated plastic caps with a foam plug in a hole for aeration.

130
Table 1. RHBV resistance in the field in 2007 of selected families derived from crosses between Cica 8-RHBV transgenic resistant lines and commercial variety Oryzica 1, and Cica 8-RHBV transgenic resistant progeny derived from self-cross.
Pedigree Cluster 1RHBV 1RHBV 2007 2006
A3-49-101-18-19/Oryzica 1-14-M-10-1-3-1 1 4.6 2,3 A3-49-101-18-19/Oryzica 1-14-M-10-1-3-2 3 3.8 A3-49-101-18-19/Oryzica 1-14-M-10-1-3-3 5 4.2 A3-49-101-18-19/Oryzica 1-14-M-10-1-3-4 5 4.6 A3-49-101-18-19/Oryzica 1-14-M-10-2-4-3 3 5.0 4,3 A3-49-101-18-19/Oryzica 1-14-M-10-2-4-4 3 3.8 A3-49-101-18-19/Oryzica 1-14-M-10-2-4-5 3 4.2 A3-49-101-18-19/Oryzica 1-14-M-10-2-4-6 4 3.8 A3-49-101-18-19/Oryzica 1-14-M-7-2-M-1-2 3 4.2 3,0 A3-49-101-18-19/Oryzica 1-14-M-7-2-M-1-3 3 4.6 A3-49-101-18-19/Oryzica 1-14-M-7-2-M-1-4 5 5.0 A3-49-60-4-13/ Oryzica 1-13III-6-5-M-1-1 5 4.6 3,0 A3-49-60-4-13/ Oryzica 1-13III-6-5-M-1-2 5 5.0 A3-49-60-4-5/ Oryzica 1-15-15-11-M-3-3 3 4.2 3,7 A3-49-60-13-69-M-1-4-1-1 5 4.6 3,7 A3-49-60-13-69-M-1-4-1-3 5 4.6 A3-49-60-13-69-M-1-4-2-1 3 5.0 3,0 A3-49-60-13-69-M-1-4-2-3 3 4.2 A3-49-60-13-69-M-1-4-2-4 5 4.6 A3-49-60-13-69-M-1-4-3-1 3 4.6 3,0 A3-49-60-13-69-M-1-4-3-4 3 4.6 A3-49-60-12-3-20-M-13-2-1-M-1-4 6 5.4 4,3 A3-49-60-12-3-20-M-13-2-1-M-2-1 3 5.0 3,7 A3-49-60-12-3-20-M-13-2-1-M-2-5 5 4.6 A3-49-60-12-3-20-M-13-2-1-M-2-6 5 4.2 A3-49-60-12-3-20-M-13-2-2-M-2-1 3 4.2 4,3 A3-49-60-12-3-20-M-13-2-2-M-2-2 3 5.0 A3-49-60-12-3-20-M-8-4-4-M-3-2 5 4.6 3,0 A3-49-60-12-3-20-M-8-4-4-M-3-3 5 4.6 A3-49-60-12-3-20-M-8-4-4-M-3-4 3 5.0 A3-49-60-12-3-20-M-8-4-4-M-3-5 3 3.8 A3-49-60-12-3-20-M-8-4-4-M-5-1 3 4.2 3,7 Cica 8/ Fedearroz 50 6 7.0 5,7 Cica 8 / Oryzica 1 6 6.6 5,7 Cica 8 6 8.2 7,7 Fedearroz 2000 4 2.2 2,3 Fedearroz 50 6 7.4 7,7 Oryzica 1 1 7.0 7,0 Colombia 1 6 5.8 4,3 A3-78-24 6 8.6 9,0

131
Figure 1. Cluster analyses for RHBV resistance of transgenic advanced lines and crosses between selected transgenic lines and Oryzica 1. Controls consisted of varieties Fedearroz 2000 (highly resistant, score <2.3); susceptible checks of Oryzica 1 (score � 7); Cica 8 (score � 8.2); Fedearroz 50 (score � 7.4) and Bluebonnet 50 (highly susceptible, score = 9); and the non-transgenic cross Cica 8/Oryzica 1 (score � 6.6) and Cica 8/Fedearroz 50 (score � 7.7). References.
Lentini Z., Lozano I, Tabares E., Fory L., Domínguez J., Cuervo M., Calvert L. 2003. Expression and inheritance of hypersensitive resistance to rice hoja blanca virus mediated by the viral nucleocapsid protein gene in transgenic rice. Theoretical and Applied Genetics 106: 1018-1026. IRRI (1996). Standard evaluation system for rice. 4th edition. 52p
1 2 3 4 5
Clusters Fd2000
Col1
Ory1
Cica8
Fd50
Blub50
6

132
Output 3: Intensification and diversification of rice cropping systems for small farmers. 3A. Activity title: Participatory breeding of upland rice in Nicaragua Authors: Gilles Trouche, Zildghean Chow
Collaborators: - CIRAD: Marc Châtel, Henri Hocdé - CIAT: Cesar Martinez, Jaime Borrero, Fernando Correa, Myriam Duque, James
Silva - INTA Nicaragua: Lázaro Narváez, José Corrales, Marlon Ortega - NGOs, farmers’ organizations and other institutions in Nicaragua: UNAG/PcaC,
UCA Siuna, SERVITECA, CIPRES, PPB-MA program.
Source of funding: CIRAD, CIAT MOFA France Abstract: The goal of CIRAD-CIAT project on participatory breeding of upland rice is to develop and select germplasm that respond to the needs of small farmers for the diverse upland rice cropping systems found specially in Nicaragua and other countries in Central America(AC). This project strongly involves the national agricultural research institution (INTA) and several NGOs and farmers’ organizations. The strategy used has a focus on participatory and decentralized varietal selection (PVS), and takes advantage of the improved germplasm recently developed by CIAT and CIRAD, which represents a great diversity of phenotypes and genetic background. The strategy of in situ participatory plant breeding (PPB) started in 2003, and used broad-based and site-specific synthetic populations as sources of genetic diversity and pedigree selection for line development. After five years, the PVS strategy succeded in identifying four high performing lines that will be released by partners in 2008. IRAT 364-Kilambé and IRAT 366-Flora will be released by the extension agency SERVITECA for the low inputs upland cropping systems in the North and Atlantic regions. The CT 15679-17-1-1-1-4 and the very early WAB 758-1-1-HB-4 lines will also be released by the INTA institution, for mechanized cropping systems in favourable upland conditions and for areas with drought stress, respectively. The promising CT 15944-10-4-3-3, derived from a japonica/glaberrima cross(Caipo/O.glaberrima), will go through a second year of validation tests in 2008 for a probable launching in 2009. Among the lines developed with the PPB approach, several lines derived from the PCT-4 and PCT-18 populations gave in 2007 promising results in their preliminaries evaluations for yield and farmers appraisal; some progenies from PCT-11 and PCTNic-3 also showed interesting traits. This research activity has generated other relevant outcomes concerning the development of methods, knowledge acquisition and capacity building.

133
Key words Participatory varietal selection, Participatory plant breeding, upland rice, small and medium-scale farmers, Nicaragua.
Background This collaborative research project between CIAT and CIRAD started in Nicaragua in May 2002. The project proposes to develop and to apply participatory crop improvement (PCI) approaches and methods for two « model plants » -- i.e., rice and sorghum – for small and medium-scale farmers in low inputs cropping systems but undergoing progressive crop intensification and increased access to markets. The specific objectives of the project are as follow: 1. To develop and apply new Participatory Variety Selection (PVS) and Participatory
Plant Breeding (PPB) methods including population enhancement and recurrent selection.
2. To identify and develop new germplasm matching the needs of small and medium-scale farmers.
3. To enhance the partners’ capacity on the participatory breeding approaches and methods.
Upland rice, otherwise known as aerobic rice, is an important food and cash crop in Central America. The total area of rice, upland and irrigated, in Central America totalizes 255,000 ha with a global paddy production of approx. 850,000 tons [1]. In Nicaragua, aerobic rice covers 55,000 ha and represents 66% of the total rice production area [2]. The most important constraints for upland rice production in Central America are: - The lack of modern and performing varieties adapted to the diverse cropping systems, particularly for less favourable upland conditions and low inputs no mechanized cropping systems; - Deficient weed control; - Drought stress in some areas of the Pacific region; - Acid soils and low solar radiation in the Atlantic regions; - Pest and diseases, particularly blast, grain discoloration and mite complex; - Insufficient grain quality for industry requirements and competitiveness with imported rice. Materials and Methods 1. Germplasm enhancement and selection using participatory approaches The introduction and evaluation in Nicaragua of exotic genetic resources from CIAT and CIRAD as well as the local development of new germplasm (lines and site-specific synthetic populations using a male-sterile gene) form the core of the participatory breeding project.

134
1.1 Development of New Lines using Participatory Breeding (PPB) At the Centro Experimental del Occidente (CEO) in Posoltega, Chinandega department, the participatory breeding schemes were started in 2003 from the PCT-4, PCT-11, CNA-7 and PCT-18 synthetic populations (CIRAD-CIAT program in Colombia) and from F2s populations derived from specific crosses to match the needs of the mechanized cropping systems for the favourable areas of the Pacific area. This work was followed-on during the 2004-2007 rain seasons with a stable group of trained expert-farmers. In 2005, a new PPB scheme was started using two new synthetic site-specific populations, PCTNic-1\0\0\1 and PCTNic-2\0\0\1, created in Nicaragua. In 2007, 251 progenies derived from these PPB schemes + 85 progenies derived conventional breeding schemes from PCT-11 were planted for continuing the selection process. At the Siuna and Bocay-Wiwili sites representing the manual cropping systems of the Jinotega and RAAN regions, the participatory breeding schemes were started in 2004 with two F2s populations from crosses incorporating the local varieties Raizora Amarillo and Criolla Siuna and 63 S1-S3 progenies derived from the CNA-7 and PCT-11 populations. This work was then continued from 2005 to 2007 at the Siuna site because of better experimental conditions and institutional framework. In 2006, a new PBB scheme was started using the site-specific synthetic population PCTNic-3\0\0\1 developed in Nicaragua for this rice agro-systems. Finally, in 2006 and 2007, progenies firstly selected at the Posoltega site but presenting plant types and agronomic traits more interesting for the manual cropping systems were included in the breeding nurseries. At the CIAT referential site for hillsides of San Dionisio, a PPB scheme was performed from 2002 to 2005 from the PCT-18 populations. Then the best S3 lines have followed their selection and evaluation process at the Bocay-Wiwili and Siuna sites.
1.2 Preliminary and advanced participatory variety trials (PVS) In 2007, three nurseries of new lines and progenies introduced from CIAT and CIRAD were planted at the CEO station for a first screening, representing 385 lines. Moreover, six advanced trials including 100 introduced and new developed lines and commercial varieties as checks and 32 validation trials were established in four upland rice areas. 2. Experimental designs and cropping conditions The breeding nurseries and PVS trials planted at the Posoltega and Siuna sites were managed following the common cultural practices and fertilizer doses applied by the farmers in each area. For the preliminary yield trials with a high number of lines, Federer incomplete block designs were used. For the more advanced yield trials, randomized complete block designs with three to four replications are commonly used.

135
3. Approaches and methods for PCI activities
The global approach, methods and tools for managing the PVS and PPB activities has been described and discussed in different recent publications [3] [4] [5].
Main Results 1. Germplasm enhancement using participatory approach Following a 2006 season with extreme drought problems, the 2007 rainy season in Nicaragua (and a great part of the Central America region) has been marked by very abundant rainfall and low radiation during the months of September and October (reproductive and early maturity phases). These climatic conditions induced high diseases pressure and sterility problems on the upland rice experiments and production fields, especially at the Posoltega site and close on-farm trials, and so adequate conditions for an effective selection for these constraints.
1.1. Development of new lines using Participatory Breeding At Posoltega, among the 89 S4 or F5 progenies derived from PPB work using the PCT-4, PCT-11 and PCT-18 populations, the breeder-farmers and the breeders selected 38 S5 and F6 progenies with good plant types, high resistance to leaf disease and grain discoloration and good grain format. In a preliminary evaluation for yield and other agronomic traits of the 28 of these S4 progenies, three have got higher yield than the best check INTA N-1 with a high farmers’ acceptation (Table 1). Among the 127 S2 progenies derived from the site-specific populations PCTNic-1 and PCTNic-2, the same breeder-farmers and the breeders have only selected 46 S3 new progenies, almost exclusively from the first population. At Siuna, among the 392 different progenies derived from the single crosses PCT-18/Raizora Amarillo and PCT-18/Criolla, and the PCT-11 and PCTNic-3 populations, the expert-farmers and the breeders have selected 275 new progenies. The progenies derived from the PCTNic-3 population particularly present excellent plant types (adequate plant height, strong stems, good tillering and long panicles) for these manual low inputs cropping conditions. In an advanced yield trial planted in two localities, at least three PCT-18>SD lines, developed at the San Dionisio CIAT referential site, over-yielded the improved variety IRAT 366 and received good farmer appraisal at the Kitri site, Jinotega (Table 2) but just obtained passable results at the Siuna site (data not shown), probably due to a deficient adaptation to the more acid and less fertile soils of the latter site.

136
1.2 Participatory Variety Selection Mechanized cropping systems in favourable conditions (medium-scale farmers)
For this target cropping system, the most promising line selected with farmers with the PVS approach, POBL 1-38 (BC2 Epagri 108/O. Llanos 5) which associates the good agronomic performances and grain quality of Epagri 108 and the durable blast resistance of O. llanos 5, was included during the 2006 season in a first round of on-farm validation (pre-launching) trials. In these trials, with some drought constraints, POBL 1-38 showed an excellent productivity with an average yield of 4.9 t/ha (19% more than the commercial variety INTA Chinandega). In a second year of validation, POBL 1-38 has been evaluated in 2007 in seven localities of the Chinandega region in comparison with two new promising lines CT 15679-17-1-1-4 and CT 15679-17-1-2-3-5, selected through a mixed conventional and PVS approach, and the same commercial variety. In these 2007 tests, CT 15679-17-1-1-1-4-M obtained higher yields (5.1 t/ha), surpassing CT 15679-17-1-2-3-5-M (4.8 t/ha) and POBL 1-38 (4.6 t/ha); moreover the latter line presents a slightly better grain quality regarding the milling yield. Considering these data and additional data in irrigated conditions, INTA decided to release the CT 15679-17-1-1-1-4-M for presenting high yield potential in both favorable upland and irrigated conditions and high level of resistance to main diseases (blast, sheath blight and others) and an appropriate grain quality. Low inputs cropping systems in climatic favourable conditions In the North-East Jinotega region, the two CIRAD varieties, IRAT 364 and IRAT 366, identified as the most promising cultivars for the local manual cropping systems, based on the agronomic results at on-farm level and the farmers’ appreciation for agronomic and post-harvest traits including grain quality [5] [6], are now in the final steps of formal registration and release. During the 2007 season, the Serviteca institution, partner of this work, carried out a very successful field day for presenting the two varieties to local authorities and farmers (Photos 1 and 2); the two varieties have received new launching names (Flora and Kilambé for IRAT 366 and IRAT 364, respectively), the foundation seed production is in progress and it is hoped that the two varieties will be officially registered in the first 2008 semester for official release (launching) during the 2008 season. On request of the farmers, a set of the available very early lines was evaluated in this area with the purpose of giving new options for rice production (new planting dates and double cropping). According to the yield results and the farmers’ preferences for plant type and other agronomic traits in the 2005-2006 on-farm trials, the line PCT-4\SA\1\1>1479-M-1-M-1 has been included in validation trials during the 2007 season; this activity must be repeated in 2008 to get sufficient data for a possible release of this early line in this region. In both the Jinotega and Siuna regions, the best two lines tested since 2004, CT 15944-10-4-3-3 and CIRAD 401, were evaluated during the 2007 season in the first cycle of on-

137
farm validation trials; despite the lost of 50% of the trials because of the Felix hurricane damages, preliminary data confirmed the quality and high farmers appraisal of the CT 15944-10-4-3-3 line. This line is derived from a japonica/glaberrima cross (BC2 Caiapo/GC 103544) and could be the first variety with a glaberrima background to be released in Latin America. Nevertheless, due to the small number of testing localities during this 2007 season, the local partner, the farmer organization U.C.M. Siuna and INTA, decided to continue the two lines evaluations during the 2008 season to get the required data for the official registration and release process. Low inputs cropping systems in areas with drought problems
In some less favourable areas of the Pacific plains of Nicaragua, very early materials (90 days or less to maturity) are of interest because they can achieve their growing cycle only during the “primera” (May-July) or the “postrera” (mid-August to end of November) rain seasons and so escape the common drought stress due to the “canícula” short intermediate dry season (mid-July to mid-August), so giving new cropping options, compared with the current medium-cycle varieties, planted in June. Following the participatory preliminary and advanced variety trials carried-out between 2003 and 2004, the best lines have been evaluated in validation trials from 2005 to 2007 in three departments of the Pacific regions, enduring contrasted climatic conditions. In 2007, only the best two lines of the preceding phase, PCT-4\SA\1\1>1479-M-1-M-1 and WAB758-1-1-HB-4, were included in this last year of evaluation. On the average of 13 environments, WAB 758-1-1-HB-4 (2.7 t/ha) achieved better yield than PCT-4>1479 and the local check, even the latter was significantly later (Table 3). WAB 758-1-1-HB-4 also presents a good industrial grain quality (Table 3). For the small farmers of the marginal rice areas, the adoption of these very early varieties will permit a more secure production with an earliest harvest, so contributing to the food security. WAB 758-1-1-HB-4 will be release by INTA in May-June 2008. 2. Strengthening partners’ capacities on Participatory Breeding In 2006, the project team had contributed to a special issue of the Agronomía Mesoamericana journal dedicated to participatory breeding researches in Latin America, on the initiative of the PPB-Meso American network. In 2007, collaborative actions were achieved with this PPB-Ma network for different training, fund-raising and communications activities. The project also administers the Spanish version of a CIRAD-INRA web site on participatory plant breeding http://fitomejoramiento-participativo.cirad.fr/. , which aims to share information, news, publications and tools and methods for scientists and students. 3. Seed distribution
In 2007, 152 progenies and advanced lines were distributed to CENTA El Salvador, Arrozgua Guatemala and the private society Altamira Nicaragua. Moreover 30 lines and the PCTNic-2 and PCTNic-3 populations were sent to the CIRAD-FOFIFA program, Madagascar.

138
Furthermore 48 new lines with high tolerance to cool temperature developed by the CIRAD-FOFIFA program in Madagascar has been introduced in CIAT for using as source of cold tolerance and were sent to Bolivia for their evaluation in hillsides conditions in the La Paz department (collaboration with Man-Dideca NGO and CIAT Bolivia).
4. Participation to conferences and meetings
- Meeting of the INTSORMIL program for Centro-America, March 12-13, San Salvador, El Salvador: one communication (G. Trouche).
- International conference Agrobiodiversity-Management of genetic resources in the savannah areas of West Africa, May 15-18, Bamako, Mali. CIRAD-ICRISAT-IER-INERA-AOPP: one communication (G. Trouche)
- Annual PCCMCA meeting at Antigua, Guatemala (April 16-20): 1 communication (S. Aguirre)
- Regional training course on Rice breeding, May 28-30, Managua, Nicaragua: course on methods for participatory breeding with specific focus on rice breeding (G. Trouche).
- EPMR review to the CIAT projects in Centro-America: two presentations of participatory breeding activities on rice and sorghum (Z. Chow, rice and S. Aguirre CIPRES, sorghum).
5. Resource mobilization
In 2007, a CORPOICA-CIAT proposal on sweet sorghum breeding was presented at the research call of the Colombian Agricultural Ministry; the project was approved in October 2007 for four years (184,000,000 Colombian pesos for CIAT-CIRAD activities). Three concept notes were elaborated and sent to possible donors. A proposal focused on developing and implementing training courses on PPB approaches and methods for Latin America to be submitted to the FAO-Technical Collaborative Program finally didn’t receive sufficient support of the countries to be admissible by FAO. Conclusion
The diversified new rice germplasm from CIAT and CIRAD associated with PVS and PPB approaches is offering interesting results for improving and diversifying rice varieties for the upland cropping systems of Nicaragua and Central America. The participatory varietal selection strategy permitted to identify fairly fast better-performing lines for matching the needs of existing upland cropping systems (manual as well as mechanized systems) and for giving new variety options to avoid drought constraints or to allow new cropping systems including rice crop. Among these lines, IRAT 364-Kilambé and IRAT 366-FLORA for the low inputs cropping systems of the North-East and Atlantic regions, CT 15679-17-1-1-1-4 for the mechanized cropping systems in favourable upland conditions and the early line WAB 758-1-1-HB-4 should be officially

139
released in Nicaragua in 2008. New lines developed in Nicaragua using a decentralized PPB strategy also gave in 2007 very promising results, For information, as result of the sorghum component of the project, the white-grain tortillero sorghum variety, Blanco Tortillero, was officially released in Nicaragua in 2007. Other three elites sorghum lines are in the pipeline to be released in 2008.
Future activities This CIAT-CIRAD project normally will be closed in June 2008. CIAT and CIRAD are thinking about new perspectives of collaboration in this theme, probably in other regions. Concerning the current project in Nicaragua, the priority activities for the last six months will be to support the partners in the registration of the varieties, to write a scientific publication and an end-project document, to share the results and lessons of this experience with partners in a workshop to be held in Nicaragua and to distribute the developed elite lines and progenies between the partners and to the regional NARS which show interest. In order to get more consistent data for the publication about the PPB work, an additional growing cycle, in particular at the Siuna site, will be useful but depends of funds availability. References [1] FAO 2005. Datos estadísticos de producción agrícola, FAOSTAT, http://faostat.fao.org/ [2] Estadísticas de producción de granos básicos de Nicaragua. MAG-FOR 2002. [3] Trouche G. et al. 2004. CIAT Annual report for the research activity on upland rice participatory plant breeding in Central America. [4] Trouche G. 2005. “Participatory Rice Breeding using Population Improvement: A New Methodology Adapted to the Needs of Small Farmers in Central America and the Caribbean. In E.P. Guimarães, ed. Population improvement: a way of exploiting the rice genetic resources of Latin America, pp. 99-110. Rome, Italie. FAO, CIRAD, CIAT, Embrapa Arroz e Feijao, DANAC. [5] Trouche, G.; Narváez-Rojas, L.; Chow-Wong, Z.; Corrales-Blandón, J. 2006. Fitomejoramiento participativo del arroz de secano en Nicaragua: metodologías, resultados y lecciones aprendidas. Agronomía Mesoamericana (CR) 17(3): 307-322. [6] Trouche, G.; Chow Wong, Z. 2007. CIAT 2006 Annual report for the research activity on upland rice participatory plant breeding in Central America. 11 p. 4 tab.

140
Last Publications
Refereed journal articles Trouche G., Aguirre Acuña S., Hocdé H., Obando Solís R., Gutiérrez Palacios N., Chow Wong Z. Valorisation de la diversité génétique du sorgho par des approches de sélection participative au Nicaragua. Valorisation of the genetic diversity of sorghum through methodologies of participatory breeding in Nicaragua. Cahiers Agricultures. Numéro spécial Agrobiodiversités. In press. vom Brocke K., Trouche G., Zongo S., Bitié A., Barro-Kondombo C., Weltzien E., Chantereau J. Création et amélioration de populations de sorgho à base large avec les agriculteurs au Burkina Faso. Cahiers Agricultures. Numéro spécial Agrobiodiversités. In press.

141
Table 1: Days to flowering, plant height, grain yield and farmers’ appreciation of the most promising lines developed by PPB methodology for the mechanized cropping systems in favourable upland conditions, Posoltega, Chinandega 2007.
Line Daysto
flowering
Plantheight(cm)
GrainYield(t/ha)
Index of overall
farmers’ appreciation 1
Frequencyof farmers
selection (%)
PCT-18>SG4-1G-2F-M 72 95 5.09 3.0 75 PCT-4>LM8-2P-2F-M 72 90 4.85 ne ne PCT-4>SG9-1G-29-M 83 99 4.36 3.5 100 PCT-4>BE8-1P-2F-M 72 110 4.36 ne ne PCT-18>SG5-1G-2F-M 83 110 4.31 ne ne PCT-4>LM4-2G-2F-M 79 84 4.30 3.0 50 INTA N-1 (Commercial variety) 75 102 3.87 2.8 31 INTA Chinandega (Commercial variety) 75 91 3.64 3.0 50 1 Farmers’ appreciation of the variety concerning the five most important plant traits using a 1-4 evaluation scale where 1 =bad, 2 = acceptable, 3 = good and 4 = excellent ne: not evaluated because harvest was done before the participatory evaluation exercise

142
Table 2: Agronomic results and farmers appreciation of the best PCT-18>SD lines developed by PPB methodology for manual low inputs cropping systems, Kitri, Jinotega, Nicaragua, 2007.
Line Days to flowering
Plantheight (cm)
Lodging! Grain Yield(t/ha)
Index of farmers’ appreciation 1
Frequency of farmers
selection (%) PCT-18\0\0\1>SD21-2-2-M-M 79 110 1 6.36 3.3 50 PCT-18\0\0\1>SD34-1-1-M-M 75 124 1 5.45 3.4 50 PCT-18\0\0\1>SD52-1-4-M-M 78 101 1 5.19 3 0 PCT-18|0\0\1>SD3-1-4-M-M 75 114 1.7 4.93 2.8 33 IRAT 366 (check) 80 97 1 4.74 3.9 83 PCT-18|0\0\1> 76 112 3 4.46 2.5 0 PCT-18|0\0\1> 75 109 1 4.16 2.7 0 INTA Dorado (check) 93 90 1 3.99 3.4 16 Trial Mean 78 107 4.36 CV (%) 21 LSD 1.56 ! IRRI scale 1 Farmers’ appreciation of the variety concerning the five most important plant traits using a 1-4 evaluation scale where 1 =bad, 2 = acceptable, 3 = good and 4 = excellent Table 3: Agronomic results and milling grain quality of two very-early lines in validation trials in upland conditions with drought constraints, Chinandega, Masaya and Carazo regions, Nicaragua 2006-2007
Line Days to flowering 1
Plant height 1(cm)
GrainYield 1 (t/ha)
Milling yield (%)
% of head rice
PCT-4\SA\1\1>1479-M-1-M-1 66 94 2333 67 55 WAB 758-1-1-HB-4 61 94 2695 74 69.5 Farmer variety 80 110 2594
1 average for 13 environments, mainly during the postrera season with delayed planting in late August-beginning of September; the check data are only for six localities where farmer used to grow an fairly early variety with delayed planting.

143
Photo 1: Field day of presentation of the two upland varieties IRAT 364-kilambé and IRAT 366-Flora under release process for the North and Atlantic regions of Nicaragua (C Serviteca)

132
Output 3: Intensification and diversification of rice cropping systems for small farmers. 3A. Activity title: Participatory breeding of upland rice in Nicaragua Authors: Gilles Trouche, Zildghean Chow
Collaborators: - CIRAD: Marc Châtel, Henri Hocdé - CIAT: Cesar Martinez, Jaime Borrero, Fernando Correa, Myriam Duque, James
Silva - INTA Nicaragua: Lázaro Narváez, José Corrales, Marlon Ortega - NGOs, farmers’ organizations and other institutions in Nicaragua: UNAG/PcaC,
UCA Siuna, SERVITECA, CIPRES, PPB-MA program.
Source of funding: CIRAD, CIAT MOFA France Abstract: The goal of CIRAD-CIAT project on participatory breeding of upland rice is to develop and select germplasm that respond to the needs of small farmers for the diverse upland rice cropping systems found specially in Nicaragua and other countries in Central America(AC). This project strongly involves the national agricultural research institution (INTA) and several NGOs and farmers’ organizations. The strategy used has a focus on participatory and decentralized varietal selection (PVS), and takes advantage of the improved germplasm recently developed by CIAT and CIRAD, which represents a great diversity of phenotypes and genetic background. The strategy of in situ participatory plant breeding (PPB) started in 2003, and used broad-based and site-specific synthetic populations as sources of genetic diversity and pedigree selection for line development. After five years, the PVS strategy succeded in identifying four high performing lines that will be released by partners in 2008. IRAT 364-Kilambé and IRAT 366-Flora will be released by the extension agency SERVITECA for the low inputs upland cropping systems in the North and Atlantic regions. The CT 15679-17-1-1-1-4 and the very early WAB 758-1-1-HB-4 lines will also be released by the INTA institution, for mechanized cropping systems in favourable upland conditions and for areas with drought stress, respectively. The promising CT 15944-10-4-3-3, derived from a japonica/glaberrima cross(Caipo/O.glaberrima), will go through a second year of validation tests in 2008 for a probable launching in 2009. Among the lines developed with the PPB approach, several lines derived from the PCT-4 and PCT-18 populations gave in 2007 promising results in their preliminaries evaluations for yield and farmers appraisal; some progenies from PCT-11 and PCTNic-3 also showed interesting traits. This research activity has generated other relevant outcomes concerning the development of methods, knowledge acquisition and capacity building.

133
Key words Participatory varietal selection, Participatory plant breeding, upland rice, small and medium-scale farmers, Nicaragua.
Background This collaborative research project between CIAT and CIRAD started in Nicaragua in May 2002. The project proposes to develop and to apply participatory crop improvement (PCI) approaches and methods for two « model plants » -- i.e., rice and sorghum – for small and medium-scale farmers in low inputs cropping systems but undergoing progressive crop intensification and increased access to markets. The specific objectives of the project are as follow: 1. To develop and apply new Participatory Variety Selection (PVS) and Participatory
Plant Breeding (PPB) methods including population enhancement and recurrent selection.
2. To identify and develop new germplasm matching the needs of small and medium-scale farmers.
3. To enhance the partners’ capacity on the participatory breeding approaches and methods.
Upland rice, otherwise known as aerobic rice, is an important food and cash crop in Central America. The total area of rice, upland and irrigated, in Central America totalizes 255,000 ha with a global paddy production of approx. 850,000 tons [1]. In Nicaragua, aerobic rice covers 55,000 ha and represents 66% of the total rice production area [2]. The most important constraints for upland rice production in Central America are: - The lack of modern and performing varieties adapted to the diverse cropping systems, particularly for less favourable upland conditions and low inputs no mechanized cropping systems; - Deficient weed control; - Drought stress in some areas of the Pacific region; - Acid soils and low solar radiation in the Atlantic regions; - Pest and diseases, particularly blast, grain discoloration and mite complex; - Insufficient grain quality for industry requirements and competitiveness with imported rice. Materials and Methods 1. Germplasm enhancement and selection using participatory approaches The introduction and evaluation in Nicaragua of exotic genetic resources from CIAT and CIRAD as well as the local development of new germplasm (lines and site-specific synthetic populations using a male-sterile gene) form the core of the participatory breeding project.

134
1.1 Development of New Lines using Participatory Breeding (PPB) At the Centro Experimental del Occidente (CEO) in Posoltega, Chinandega department, the participatory breeding schemes were started in 2003 from the PCT-4, PCT-11, CNA-7 and PCT-18 synthetic populations (CIRAD-CIAT program in Colombia) and from F2s populations derived from specific crosses to match the needs of the mechanized cropping systems for the favourable areas of the Pacific area. This work was followed-on during the 2004-2007 rain seasons with a stable group of trained expert-farmers. In 2005, a new PPB scheme was started using two new synthetic site-specific populations, PCTNic-1\0\0\1 and PCTNic-2\0\0\1, created in Nicaragua. In 2007, 251 progenies derived from these PPB schemes + 85 progenies derived conventional breeding schemes from PCT-11 were planted for continuing the selection process. At the Siuna and Bocay-Wiwili sites representing the manual cropping systems of the Jinotega and RAAN regions, the participatory breeding schemes were started in 2004 with two F2s populations from crosses incorporating the local varieties Raizora Amarillo and Criolla Siuna and 63 S1-S3 progenies derived from the CNA-7 and PCT-11 populations. This work was then continued from 2005 to 2007 at the Siuna site because of better experimental conditions and institutional framework. In 2006, a new PBB scheme was started using the site-specific synthetic population PCTNic-3\0\0\1 developed in Nicaragua for this rice agro-systems. Finally, in 2006 and 2007, progenies firstly selected at the Posoltega site but presenting plant types and agronomic traits more interesting for the manual cropping systems were included in the breeding nurseries. At the CIAT referential site for hillsides of San Dionisio, a PPB scheme was performed from 2002 to 2005 from the PCT-18 populations. Then the best S3 lines have followed their selection and evaluation process at the Bocay-Wiwili and Siuna sites.
1.2 Preliminary and advanced participatory variety trials (PVS) In 2007, three nurseries of new lines and progenies introduced from CIAT and CIRAD were planted at the CEO station for a first screening, representing 385 lines. Moreover, six advanced trials including 100 introduced and new developed lines and commercial varieties as checks and 32 validation trials were established in four upland rice areas. 2. Experimental designs and cropping conditions The breeding nurseries and PVS trials planted at the Posoltega and Siuna sites were managed following the common cultural practices and fertilizer doses applied by the farmers in each area. For the preliminary yield trials with a high number of lines, Federer incomplete block designs were used. For the more advanced yield trials, randomized complete block designs with three to four replications are commonly used.

135
3. Approaches and methods for PCI activities
The global approach, methods and tools for managing the PVS and PPB activities has been described and discussed in different recent publications [3] [4] [5].
Main Results 1. Germplasm enhancement using participatory approach Following a 2006 season with extreme drought problems, the 2007 rainy season in Nicaragua (and a great part of the Central America region) has been marked by very abundant rainfall and low radiation during the months of September and October (reproductive and early maturity phases). These climatic conditions induced high diseases pressure and sterility problems on the upland rice experiments and production fields, especially at the Posoltega site and close on-farm trials, and so adequate conditions for an effective selection for these constraints.
1.1. Development of new lines using Participatory Breeding At Posoltega, among the 89 S4 or F5 progenies derived from PPB work using the PCT-4, PCT-11 and PCT-18 populations, the breeder-farmers and the breeders selected 38 S5 and F6 progenies with good plant types, high resistance to leaf disease and grain discoloration and good grain format. In a preliminary evaluation for yield and other agronomic traits of the 28 of these S4 progenies, three have got higher yield than the best check INTA N-1 with a high farmers’ acceptation (Table 1). Among the 127 S2 progenies derived from the site-specific populations PCTNic-1 and PCTNic-2, the same breeder-farmers and the breeders have only selected 46 S3 new progenies, almost exclusively from the first population. At Siuna, among the 392 different progenies derived from the single crosses PCT-18/Raizora Amarillo and PCT-18/Criolla, and the PCT-11 and PCTNic-3 populations, the expert-farmers and the breeders have selected 275 new progenies. The progenies derived from the PCTNic-3 population particularly present excellent plant types (adequate plant height, strong stems, good tillering and long panicles) for these manual low inputs cropping conditions. In an advanced yield trial planted in two localities, at least three PCT-18>SD lines, developed at the San Dionisio CIAT referential site, over-yielded the improved variety IRAT 366 and received good farmer appraisal at the Kitri site, Jinotega (Table 2) but just obtained passable results at the Siuna site (data not shown), probably due to a deficient adaptation to the more acid and less fertile soils of the latter site.

136
1.2 Participatory Variety Selection Mechanized cropping systems in favourable conditions (medium-scale farmers)
For this target cropping system, the most promising line selected with farmers with the PVS approach, POBL 1-38 (BC2 Epagri 108/O. Llanos 5) which associates the good agronomic performances and grain quality of Epagri 108 and the durable blast resistance of O. llanos 5, was included during the 2006 season in a first round of on-farm validation (pre-launching) trials. In these trials, with some drought constraints, POBL 1-38 showed an excellent productivity with an average yield of 4.9 t/ha (19% more than the commercial variety INTA Chinandega). In a second year of validation, POBL 1-38 has been evaluated in 2007 in seven localities of the Chinandega region in comparison with two new promising lines CT 15679-17-1-1-4 and CT 15679-17-1-2-3-5, selected through a mixed conventional and PVS approach, and the same commercial variety. In these 2007 tests, CT 15679-17-1-1-1-4-M obtained higher yields (5.1 t/ha), surpassing CT 15679-17-1-2-3-5-M (4.8 t/ha) and POBL 1-38 (4.6 t/ha); moreover the latter line presents a slightly better grain quality regarding the milling yield. Considering these data and additional data in irrigated conditions, INTA decided to release the CT 15679-17-1-1-1-4-M for presenting high yield potential in both favorable upland and irrigated conditions and high level of resistance to main diseases (blast, sheath blight and others) and an appropriate grain quality. Low inputs cropping systems in climatic favourable conditions In the North-East Jinotega region, the two CIRAD varieties, IRAT 364 and IRAT 366, identified as the most promising cultivars for the local manual cropping systems, based on the agronomic results at on-farm level and the farmers’ appreciation for agronomic and post-harvest traits including grain quality [5] [6], are now in the final steps of formal registration and release. During the 2007 season, the Serviteca institution, partner of this work, carried out a very successful field day for presenting the two varieties to local authorities and farmers (Photos 1 and 2); the two varieties have received new launching names (Flora and Kilambé for IRAT 366 and IRAT 364, respectively), the foundation seed production is in progress and it is hoped that the two varieties will be officially registered in the first 2008 semester for official release (launching) during the 2008 season. On request of the farmers, a set of the available very early lines was evaluated in this area with the purpose of giving new options for rice production (new planting dates and double cropping). According to the yield results and the farmers’ preferences for plant type and other agronomic traits in the 2005-2006 on-farm trials, the line PCT-4\SA\1\1>1479-M-1-M-1 has been included in validation trials during the 2007 season; this activity must be repeated in 2008 to get sufficient data for a possible release of this early line in this region. In both the Jinotega and Siuna regions, the best two lines tested since 2004, CT 15944-10-4-3-3 and CIRAD 401, were evaluated during the 2007 season in the first cycle of on-

137
farm validation trials; despite the lost of 50% of the trials because of the Felix hurricane damages, preliminary data confirmed the quality and high farmers appraisal of the CT 15944-10-4-3-3 line. This line is derived from a japonica/glaberrima cross (BC2 Caiapo/GC 103544) and could be the first variety with a glaberrima background to be released in Latin America. Nevertheless, due to the small number of testing localities during this 2007 season, the local partner, the farmer organization U.C.M. Siuna and INTA, decided to continue the two lines evaluations during the 2008 season to get the required data for the official registration and release process. Low inputs cropping systems in areas with drought problems
In some less favourable areas of the Pacific plains of Nicaragua, very early materials (90 days or less to maturity) are of interest because they can achieve their growing cycle only during the “primera” (May-July) or the “postrera” (mid-August to end of November) rain seasons and so escape the common drought stress due to the “canícula” short intermediate dry season (mid-July to mid-August), so giving new cropping options, compared with the current medium-cycle varieties, planted in June. Following the participatory preliminary and advanced variety trials carried-out between 2003 and 2004, the best lines have been evaluated in validation trials from 2005 to 2007 in three departments of the Pacific regions, enduring contrasted climatic conditions. In 2007, only the best two lines of the preceding phase, PCT-4\SA\1\1>1479-M-1-M-1 and WAB758-1-1-HB-4, were included in this last year of evaluation. On the average of 13 environments, WAB 758-1-1-HB-4 (2.7 t/ha) achieved better yield than PCT-4>1479 and the local check, even the latter was significantly later (Table 3). WAB 758-1-1-HB-4 also presents a good industrial grain quality (Table 3). For the small farmers of the marginal rice areas, the adoption of these very early varieties will permit a more secure production with an earliest harvest, so contributing to the food security. WAB 758-1-1-HB-4 will be release by INTA in May-June 2008. 2. Strengthening partners’ capacities on Participatory Breeding In 2006, the project team had contributed to a special issue of the Agronomía Mesoamericana journal dedicated to participatory breeding researches in Latin America, on the initiative of the PPB-Meso American network. In 2007, collaborative actions were achieved with this PPB-Ma network for different training, fund-raising and communications activities. The project also administers the Spanish version of a CIRAD-INRA web site on participatory plant breeding http://fitomejoramiento-participativo.cirad.fr/. , which aims to share information, news, publications and tools and methods for scientists and students. 3. Seed distribution
In 2007, 152 progenies and advanced lines were distributed to CENTA El Salvador, Arrozgua Guatemala and the private society Altamira Nicaragua. Moreover 30 lines and the PCTNic-2 and PCTNic-3 populations were sent to the CIRAD-FOFIFA program, Madagascar.

138
Furthermore 48 new lines with high tolerance to cool temperature developed by the CIRAD-FOFIFA program in Madagascar has been introduced in CIAT for using as source of cold tolerance and were sent to Bolivia for their evaluation in hillsides conditions in the La Paz department (collaboration with Man-Dideca NGO and CIAT Bolivia).
4. Participation to conferences and meetings
- Meeting of the INTSORMIL program for Centro-America, March 12-13, San Salvador, El Salvador: one communication (G. Trouche).
- International conference Agrobiodiversity-Management of genetic resources in the savannah areas of West Africa, May 15-18, Bamako, Mali. CIRAD-ICRISAT-IER-INERA-AOPP: one communication (G. Trouche)
- Annual PCCMCA meeting at Antigua, Guatemala (April 16-20): 1 communication (S. Aguirre)
- Regional training course on Rice breeding, May 28-30, Managua, Nicaragua: course on methods for participatory breeding with specific focus on rice breeding (G. Trouche).
- EPMR review to the CIAT projects in Centro-America: two presentations of participatory breeding activities on rice and sorghum (Z. Chow, rice and S. Aguirre CIPRES, sorghum).
5. Resource mobilization
In 2007, a CORPOICA-CIAT proposal on sweet sorghum breeding was presented at the research call of the Colombian Agricultural Ministry; the project was approved in October 2007 for four years (184,000,000 Colombian pesos for CIAT-CIRAD activities). Three concept notes were elaborated and sent to possible donors. A proposal focused on developing and implementing training courses on PPB approaches and methods for Latin America to be submitted to the FAO-Technical Collaborative Program finally didn’t receive sufficient support of the countries to be admissible by FAO. Conclusion
The diversified new rice germplasm from CIAT and CIRAD associated with PVS and PPB approaches is offering interesting results for improving and diversifying rice varieties for the upland cropping systems of Nicaragua and Central America. The participatory varietal selection strategy permitted to identify fairly fast better-performing lines for matching the needs of existing upland cropping systems (manual as well as mechanized systems) and for giving new variety options to avoid drought constraints or to allow new cropping systems including rice crop. Among these lines, IRAT 364-Kilambé and IRAT 366-FLORA for the low inputs cropping systems of the North-East and Atlantic regions, CT 15679-17-1-1-1-4 for the mechanized cropping systems in favourable upland conditions and the early line WAB 758-1-1-HB-4 should be officially

139
released in Nicaragua in 2008. New lines developed in Nicaragua using a decentralized PPB strategy also gave in 2007 very promising results, For information, as result of the sorghum component of the project, the white-grain tortillero sorghum variety, Blanco Tortillero, was officially released in Nicaragua in 2007. Other three elites sorghum lines are in the pipeline to be released in 2008.
Future activities This CIAT-CIRAD project normally will be closed in June 2008. CIAT and CIRAD are thinking about new perspectives of collaboration in this theme, probably in other regions. Concerning the current project in Nicaragua, the priority activities for the last six months will be to support the partners in the registration of the varieties, to write a scientific publication and an end-project document, to share the results and lessons of this experience with partners in a workshop to be held in Nicaragua and to distribute the developed elite lines and progenies between the partners and to the regional NARS which show interest. In order to get more consistent data for the publication about the PPB work, an additional growing cycle, in particular at the Siuna site, will be useful but depends of funds availability. References [1] FAO 2005. Datos estadísticos de producción agrícola, FAOSTAT, http://faostat.fao.org/ [2] Estadísticas de producción de granos básicos de Nicaragua. MAG-FOR 2002. [3] Trouche G. et al. 2004. CIAT Annual report for the research activity on upland rice participatory plant breeding in Central America. [4] Trouche G. 2005. “Participatory Rice Breeding using Population Improvement: A New Methodology Adapted to the Needs of Small Farmers in Central America and the Caribbean. In E.P. Guimarães, ed. Population improvement: a way of exploiting the rice genetic resources of Latin America, pp. 99-110. Rome, Italie. FAO, CIRAD, CIAT, Embrapa Arroz e Feijao, DANAC. [5] Trouche, G.; Narváez-Rojas, L.; Chow-Wong, Z.; Corrales-Blandón, J. 2006. Fitomejoramiento participativo del arroz de secano en Nicaragua: metodologías, resultados y lecciones aprendidas. Agronomía Mesoamericana (CR) 17(3): 307-322. [6] Trouche, G.; Chow Wong, Z. 2007. CIAT 2006 Annual report for the research activity on upland rice participatory plant breeding in Central America. 11 p. 4 tab.

140
Last Publications
Refereed journal articles Trouche G., Aguirre Acuña S., Hocdé H., Obando Solís R., Gutiérrez Palacios N., Chow Wong Z. Valorisation de la diversité génétique du sorgho par des approches de sélection participative au Nicaragua. Valorisation of the genetic diversity of sorghum through methodologies of participatory breeding in Nicaragua. Cahiers Agricultures. Numéro spécial Agrobiodiversités. In press. vom Brocke K., Trouche G., Zongo S., Bitié A., Barro-Kondombo C., Weltzien E., Chantereau J. Création et amélioration de populations de sorgho à base large avec les agriculteurs au Burkina Faso. Cahiers Agricultures. Numéro spécial Agrobiodiversités. In press.

141
Table 1: Days to flowering, plant height, grain yield and farmers’ appreciation of the most promising lines developed by PPB methodology for the mechanized cropping systems in favourable upland conditions, Posoltega, Chinandega 2007.
Line Daysto
flowering
Plantheight(cm)
GrainYield(t/ha)
Index of overall
farmers’ appreciation 1
Frequencyof farmers
selection (%)
PCT-18>SG4-1G-2F-M 72 95 5.09 3.0 75 PCT-4>LM8-2P-2F-M 72 90 4.85 ne ne PCT-4>SG9-1G-29-M 83 99 4.36 3.5 100 PCT-4>BE8-1P-2F-M 72 110 4.36 ne ne PCT-18>SG5-1G-2F-M 83 110 4.31 ne ne PCT-4>LM4-2G-2F-M 79 84 4.30 3.0 50 INTA N-1 (Commercial variety) 75 102 3.87 2.8 31 INTA Chinandega (Commercial variety) 75 91 3.64 3.0 50 1 Farmers’ appreciation of the variety concerning the five most important plant traits using a 1-4 evaluation scale where 1 =bad, 2 = acceptable, 3 = good and 4 = excellent ne: not evaluated because harvest was done before the participatory evaluation exercise

142
Table 2: Agronomic results and farmers appreciation of the best PCT-18>SD lines developed by PPB methodology for manual low inputs cropping systems, Kitri, Jinotega, Nicaragua, 2007.
Line Days to flowering
Plantheight (cm)
Lodging! Grain Yield(t/ha)
Index of farmers’ appreciation 1
Frequency of farmers
selection (%) PCT-18\0\0\1>SD21-2-2-M-M 79 110 1 6.36 3.3 50 PCT-18\0\0\1>SD34-1-1-M-M 75 124 1 5.45 3.4 50 PCT-18\0\0\1>SD52-1-4-M-M 78 101 1 5.19 3 0 PCT-18|0\0\1>SD3-1-4-M-M 75 114 1.7 4.93 2.8 33 IRAT 366 (check) 80 97 1 4.74 3.9 83 PCT-18|0\0\1> 76 112 3 4.46 2.5 0 PCT-18|0\0\1> 75 109 1 4.16 2.7 0 INTA Dorado (check) 93 90 1 3.99 3.4 16 Trial Mean 78 107 4.36 CV (%) 21 LSD 1.56 ! IRRI scale 1 Farmers’ appreciation of the variety concerning the five most important plant traits using a 1-4 evaluation scale where 1 =bad, 2 = acceptable, 3 = good and 4 = excellent Table 3: Agronomic results and milling grain quality of two very-early lines in validation trials in upland conditions with drought constraints, Chinandega, Masaya and Carazo regions, Nicaragua 2006-2007
Line Days to flowering 1
Plant height 1(cm)
GrainYield 1 (t/ha)
Milling yield (%)
% of head rice
PCT-4\SA\1\1>1479-M-1-M-1 66 94 2333 67 55 WAB 758-1-1-HB-4 61 94 2695 74 69.5 Farmer variety 80 110 2594
1 average for 13 environments, mainly during the postrera season with delayed planting in late August-beginning of September; the check data are only for six localities where farmer used to grow an fairly early variety with delayed planting.

143
Photo 1: Field day of presentation of the two upland varieties IRAT 364-kilambé and IRAT 366-Flora under release process for the North and Atlantic regions of Nicaragua (C Serviteca)
Related Documents











