21. Amino acid metabolism: nitrogen fixation, transamination and NH 3 transport

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

21. Amino acid metabolism: nitrogen fixation, transamination and NH3 transport
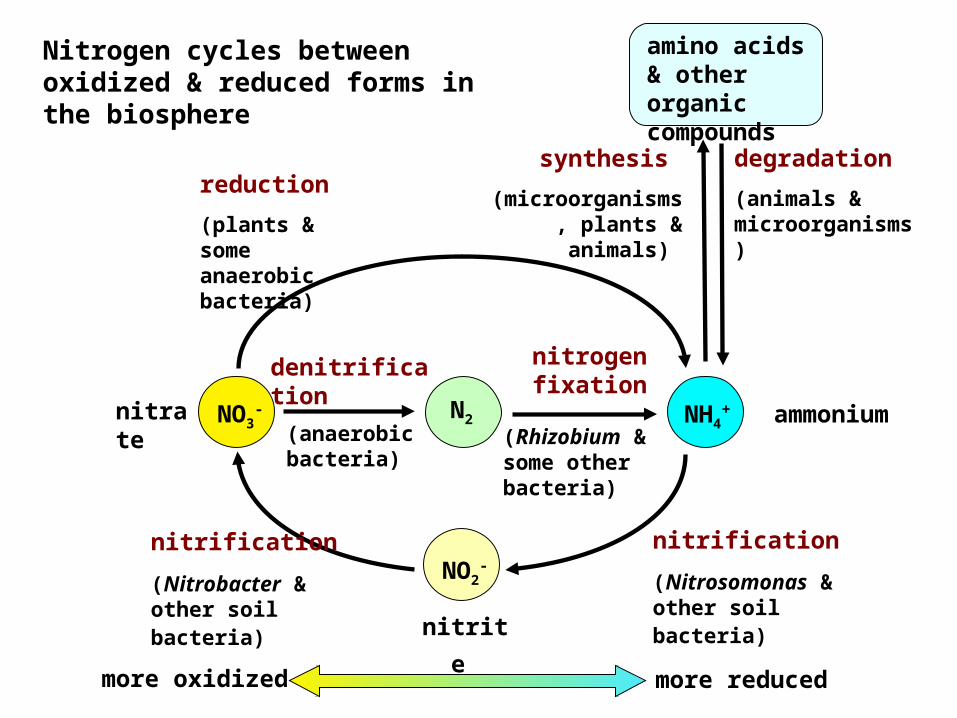
Nitrogen cycles between oxidized & reduced forms in the biosphere
degradation (animals & microorganisms)
synthesis (microorganisms
, plants & animals)
amino acids & other organic compounds
(Rhizobium & some other bacteria)
nitrification (Nitrosomonas & other soil bacteria)
NO2-
nitrite
(anaerobic bacteria)
nitrogen fixationdenitrifica
tionnitrate
NH4+ ammonium N2
reduction (plants & some anaerobic bacteria)
NO3-
more oxidized more reduced
nitrification (Nitrobacter & other soil bacteria)
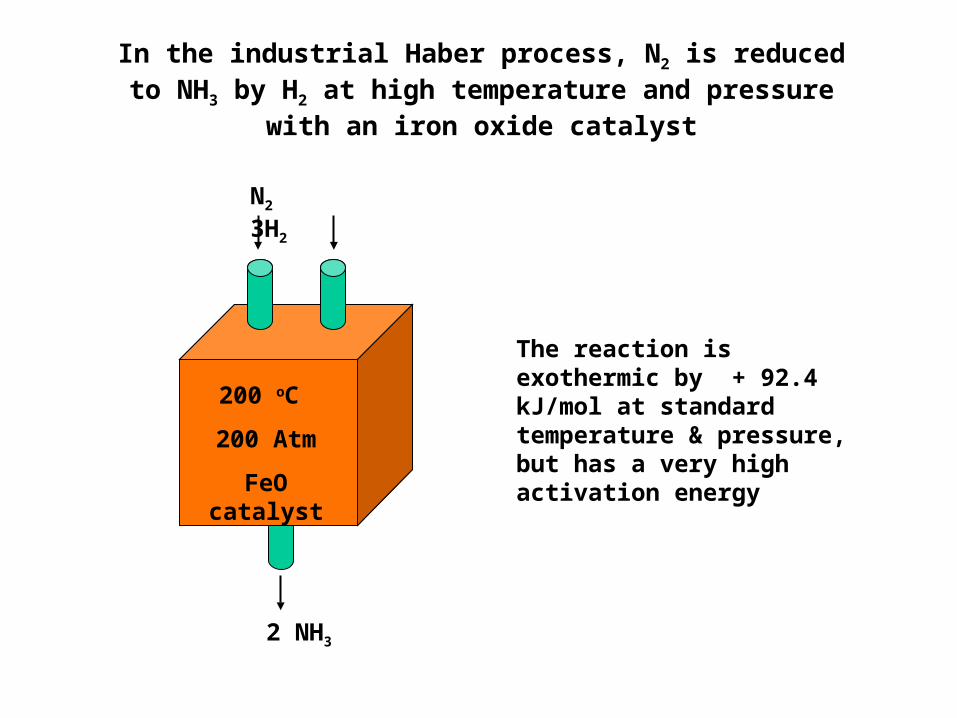
In the industrial Haber process, N2 is reduced to NH3 by H2 at high temperature and pressure
with an iron oxide catalyst
2 NH3
N2 3H2
200 oC 200 Atm
FeO catalyst
The reaction is exothermic by + 92.4 kJ/mol at standard temperature & pressure, but has a very high activation energy

The roots of leguminous plants have nodules that contain N2-fixing bacteria Bacteroids (rod-like bacteria) containing nitrogenase live inside the nodule cells
2 m
bacteroids
nodule cell nucleus
(This electron micrograph is colorized artificially.)
Nitrogenase is very sensitive to O2. It is protected in the nodules by leghemoglobin, a heme protein with a strong affinity for O2. Leghemoglobin is produced by the plant, but carries O2 for reduction by the bacterial respiratory chain, keeping the O2 concentration low.
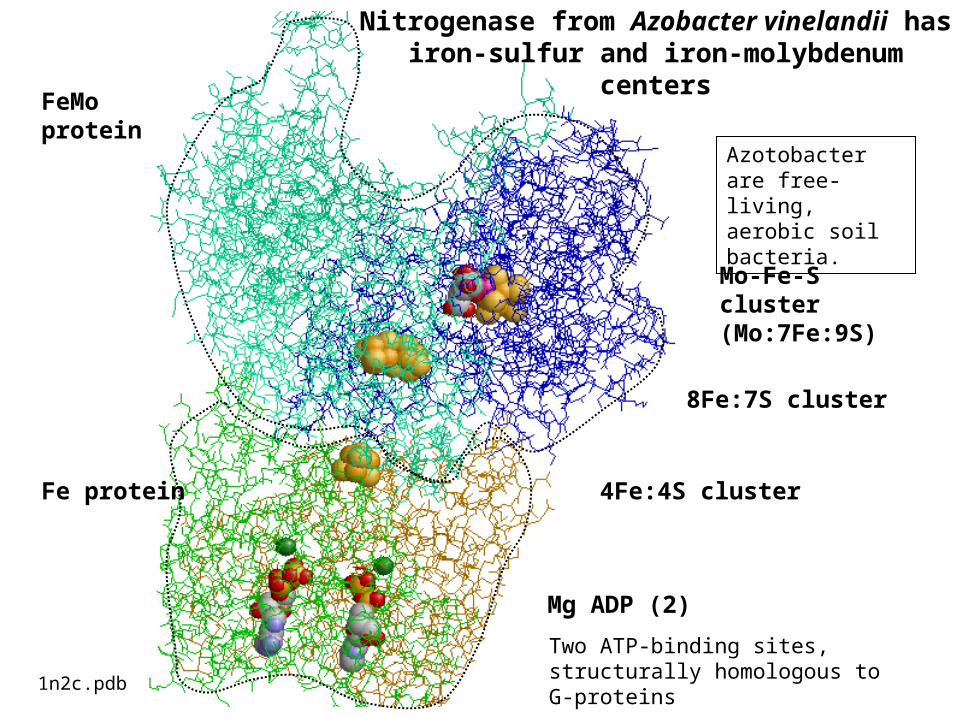
Mo-Fe-S cluster (Mo:7Fe:9S)
8Fe:7S cluster
4Fe:4S cluster
Mg ADP (2)
FeMo protein
Fe protein
1n2c.pdb
Nitrogenase from Azobacter vinelandii has iron-sulfur and iron-molybdenum
centers
Two ATP-binding sites, structurally homologous to G-proteins
Azotobacter are free-living, aerobic soil bacteria.
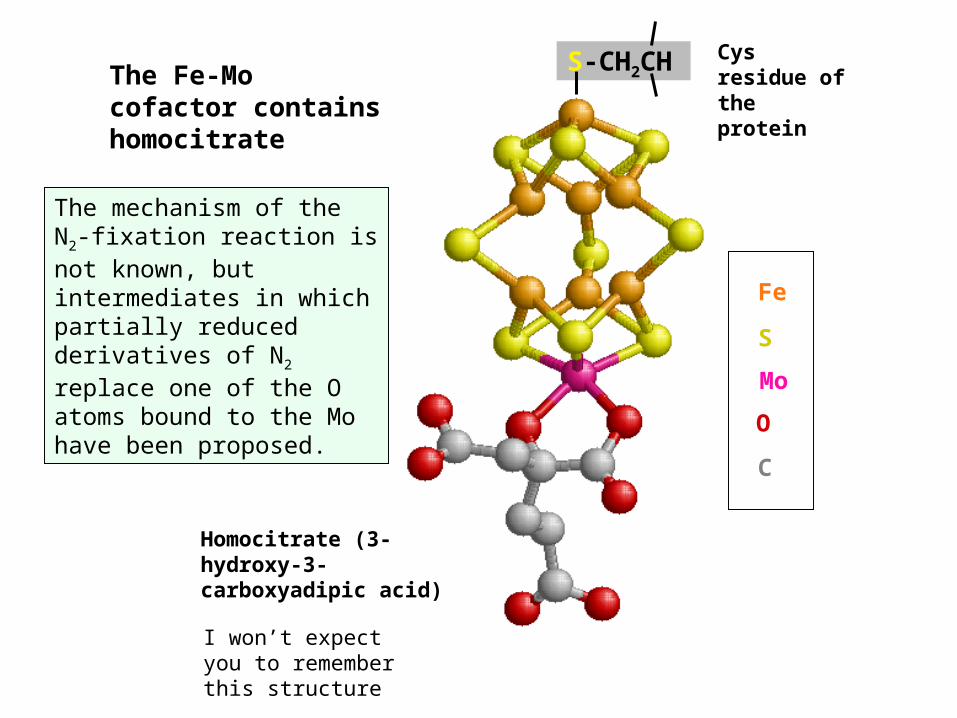
The mechanism of the N2-fixation reaction is not known, but intermediates in which partially reduced derivatives of N2 replace one of the O atoms bound to the Mo have been proposed.
Homocitrate (3-hydroxy-3-carboxyadipic acid)
MoSFe
OC
Cys residue of the protein
The Fe-Mo cofactor contains homocitrate
S-CH2CH
I won’t expect you to remember this structure
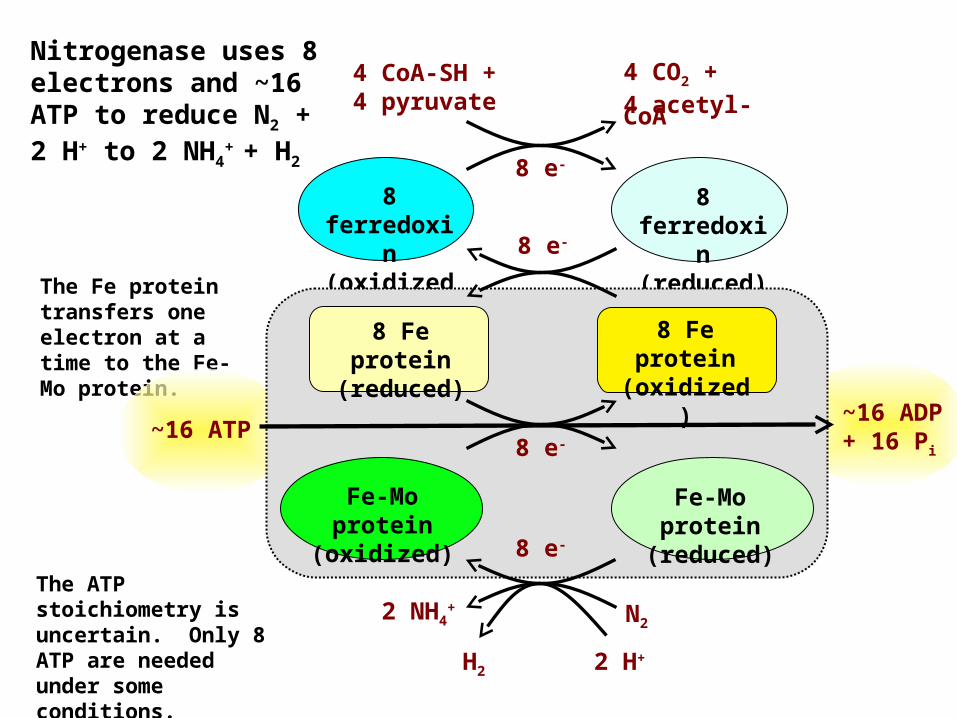
Nitrogenase uses 8 electrons and ~16 ATP to reduce N2 + 2 H+ to 2 NH4
+ + H2
The ATP stoichiometry is uncertain. Only 8 ATP are needed under some conditions.
The Fe protein transfers one electron at a time to the Fe-Mo protein.
8 ferredoxi
n (oxidized
)
8 ferredoxi
n (reduced)
Fe-Mo protein (reduced)
Fe-Mo protein
(oxidized)
8 Fe protein
(reduced)
8 Fe protein (oxidized
)
4 CO2 +4 acetyl-CoA
4 CoA-SH + 4 pyruvate
8 e-
8 e-
8 e-
8 e-
2 H+H2
2 NH4+ N2
~16 ADP + 16 Pi
~16 ATP
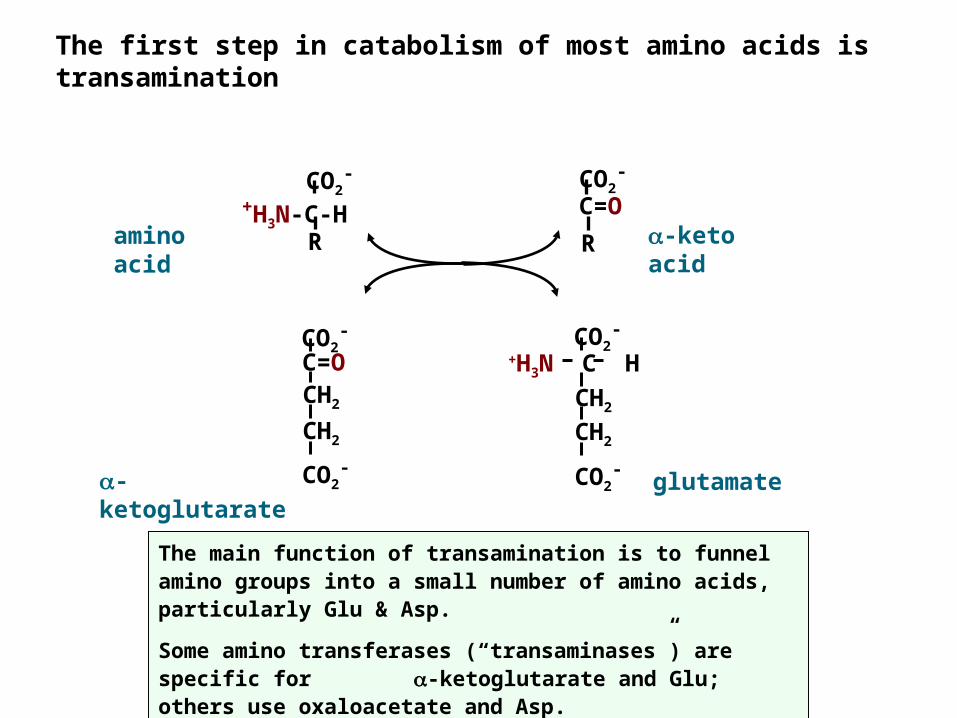
The first step in catabolism of most amino acids is transamination
-ketoglutarate
glutamate
-keto acid
amino acid
The main function of transamination is to funnel amino groups into a small number of amino acids, particularly Glu & Asp.Some amino transferases (“transaminases”) are specific for -ketoglutarate and Glu; others use oxaloacetate and Asp.
CO2-
CO2-
C=OCH2
CH2
CO2-
CO2-
+H3N C HCH2
CH2
CO2-
+H3N-C-HR
CO2-
RC=O
CH3
CH2OH
HO
+HOCH2 Cl-NH
pyridoxine hydrochloride (vitamin B6)
pyridoxamine phosphate
CH3
CH2O- P
HO
H3NCH2+
NH +
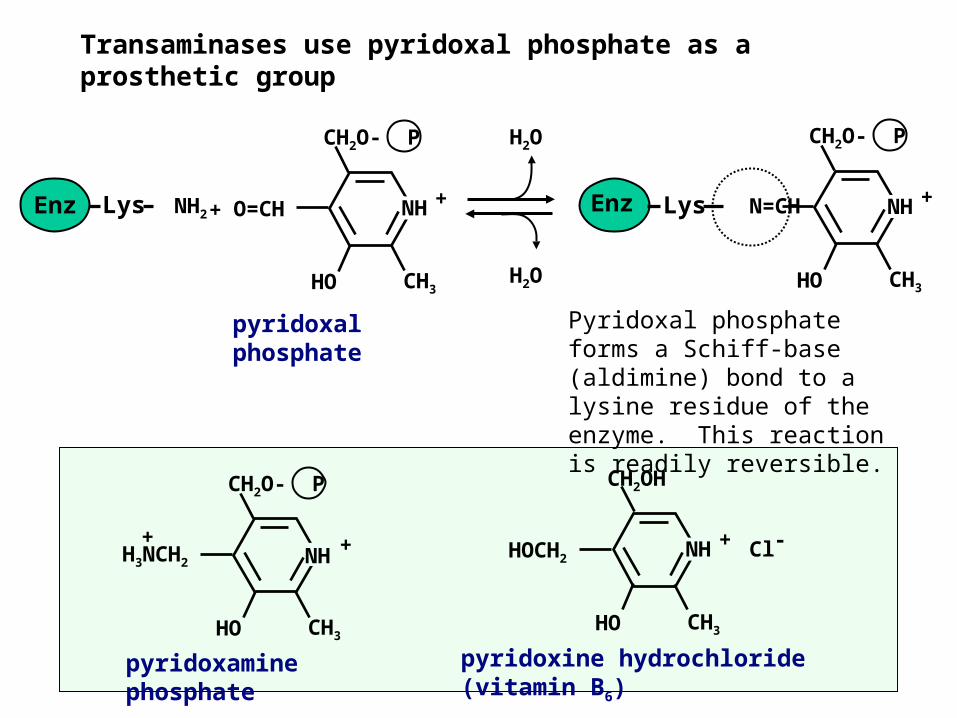
Transaminases use pyridoxal phosphate as a prosthetic group
Pyridoxal phosphate forms a Schiff-base (aldimine) bond to a lysine residue of the enzyme. This reaction is readily reversible.
pyridoxal phosphate
CH3
CH2O- P
HO
NH +O=CHLys NH2Enz
CH3
CH2O- P
HO
NH +Lys N=CHEnz+
H2O
H2O
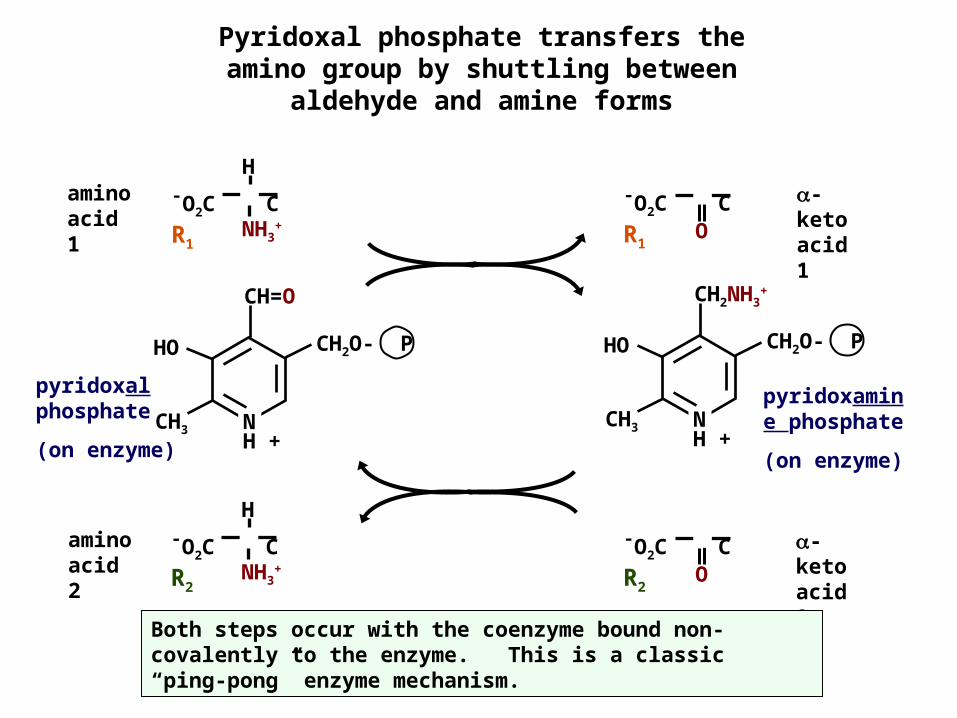
Pyridoxal phosphate transfers the amino group by shuttling between
aldehyde and amine forms
amino acid 2
-keto acid 2
pyridoxamine phosphate(on enzyme)
CH=O
CH3
CH2O- PHO
NH +
CH2NH3+
CH3
CH2O- PHO
NH +
pyridoxal phosphate (on enzyme)
amino acid 1
-keto acid 1
Both steps occur with the coenzyme bound non-covalently to the enzyme. This is a classic “ping-pong” enzyme mechanism.
-O2C C R1 ONH3
+
H-O2C C R1
NH3+
H-O2C C R2
-O2C C R2 O
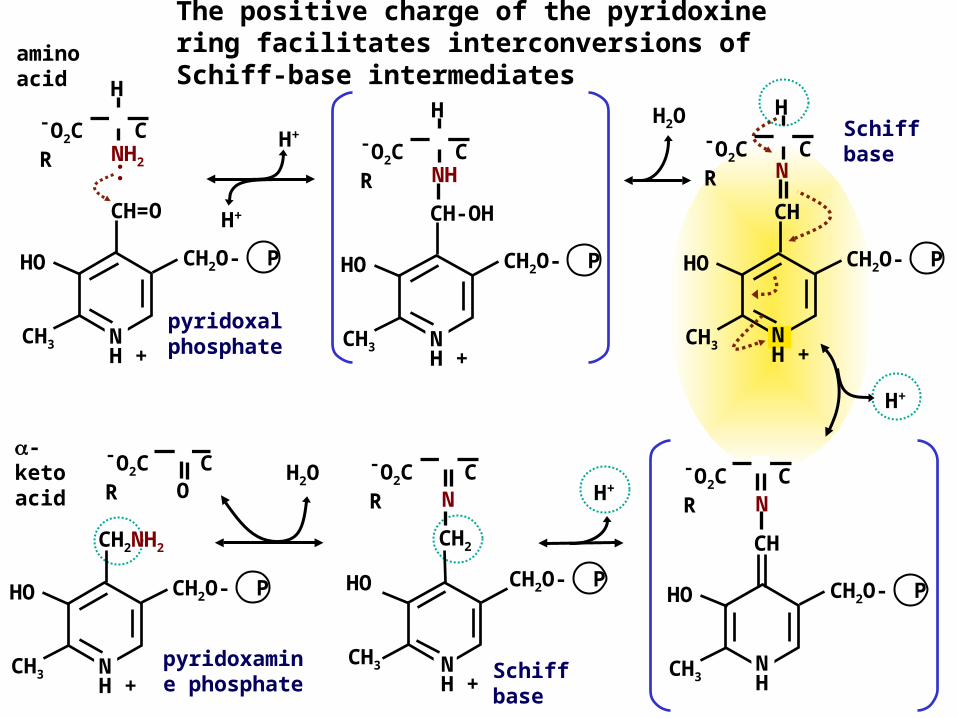
The positive charge of the pyridoxine ring facilitates interconversions of Schiff-base intermediates
Schiff base
pyridoxal phosphate
H2O
H+
CH=O
CH3
CH2O- PHO
NH +
NH2
H-O2C C R ..
-keto acid
amino acid
H+
H+NCH
CH3
CH2O- PHO
NH +
H-O2C C R
Schiff base
pyridoxamine phosphate
H+
CH2NH2
CH3
CH2O- PHO
NH +
H2O
CHN
CH3
CH2O- PHO
NH
-O2C C R
CH2
CH3
CH2O- PHO
N
NH +
-O2C C R
CH-OH
CH3
CH2O- PHO
NH +
NH
H-O2C C R
O-O2C C R
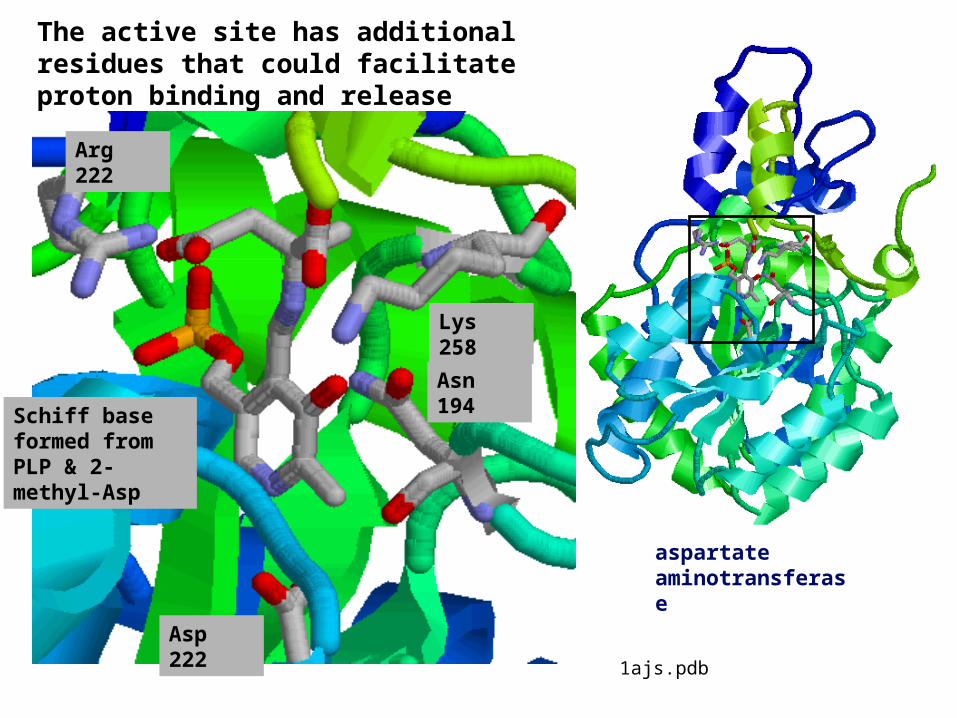
The active site has additional residues that could facilitate proton binding and release
Asp 222
Asn 194
Arg 222
Schiff base formed from PLP & 2-methyl-Asp
aspartate aminotransferase
Lys 258
1ajs.pdb
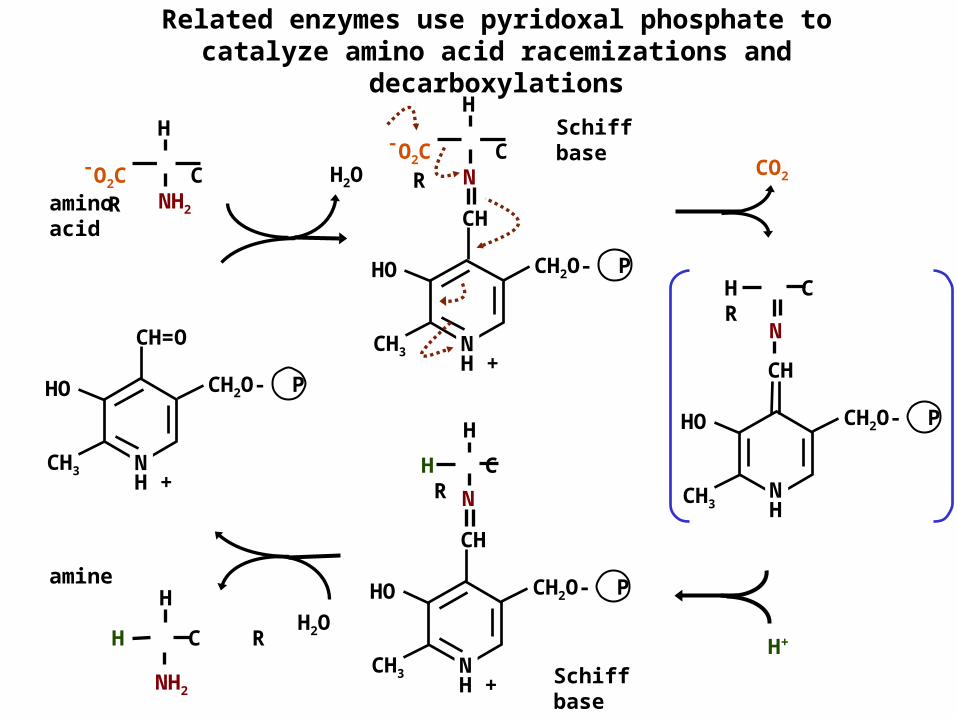
Related enzymes use pyridoxal phosphate to catalyze amino acid racemizations and
decarboxylationsSchiff base
Schiff base
H2O
CH=O
CH3
CH2O- PHO
NH +
NH2
H-O2C C R
CHN
H C R
CH3
CH2O- PHO
NH
NH2
H C RH
CO2
H+
amino acid
amine
H C R
CH3
CH2O- PHO
NH +
CHN
H
CH
CH3
CH2O- PHO
NH +
N
H-O2C C R
H2O
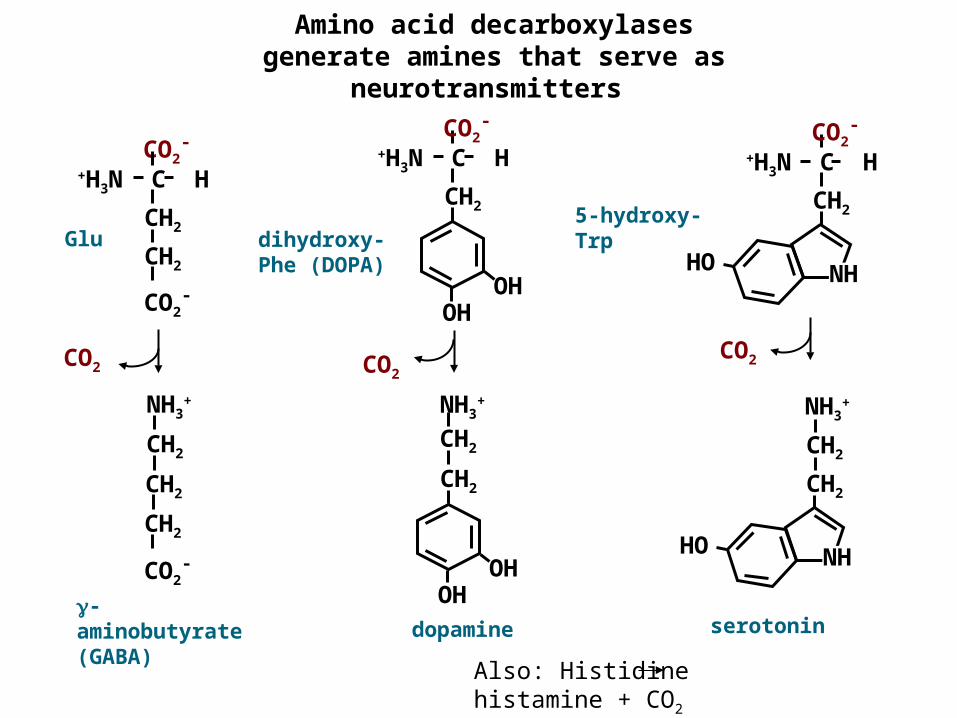
Amino acid decarboxylases generate amines that serve as
neurotransmitters
CO2-
CO2-
+H3N C HCH2
CH2dihydroxy-Phe (DOPA)
Glu5-hydroxy-Trp
CO2-
+H3N C HCH2
HO NH
CO2
CO2-
+H3N C HCH2
OHOH
CO2-
CH2
CH2
CH2
NH3+
dopamine-aminobutyrate (GABA)
serotonin
CH2
HO NH
CH2
NH3+
CH2
CH2
OHOH
NH3+
CO2CO2
Also: Histidine histamine + CO2
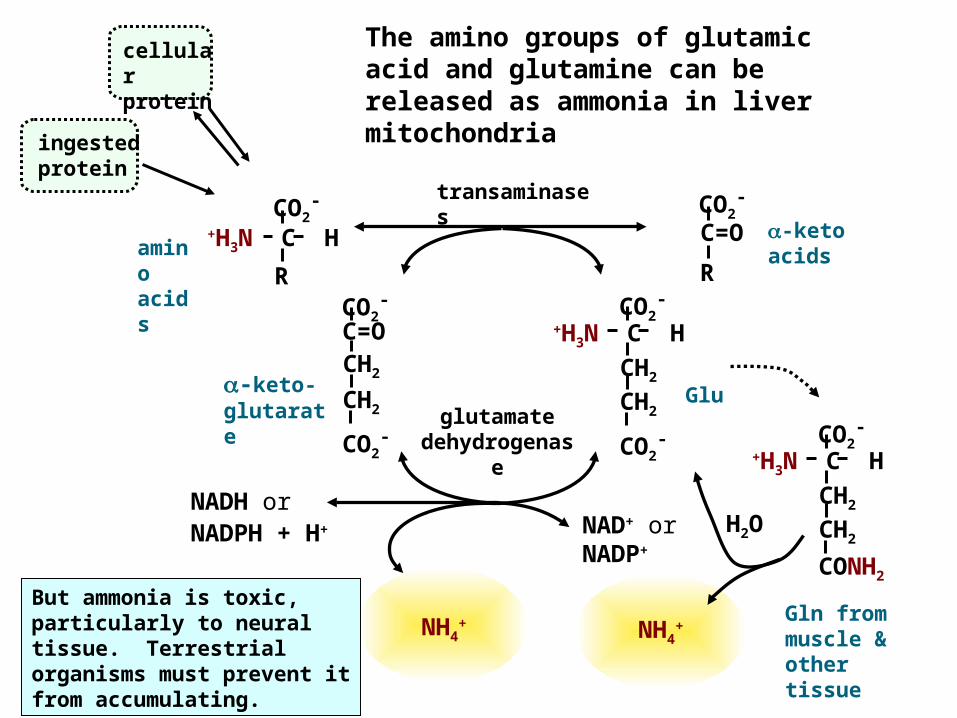
The amino groups of glutamic acid and glutamine can be released as ammonia in liver mitochondria
H2ONAD+ or NADP+
Glu
-keto acids
cellular protein
ingested protein
NADH or NADPH + H+
-keto-glutarate
CO2-
+H3N C HR
CO2-
CO2-
C=OCH2
CH2
CO2-
CO2-
+H3N C HCH2
CH2
CO2-
C=OR
amino acids
Gln from muscle & other tissue
NH4+
But ammonia is toxic, particularly to neural tissue. Terrestrial organisms must prevent it from accumulating.
CONH2
CO2-
+H3N C HCH2
CH2
transaminases
glutamate dehydrogenas
e
NH4+
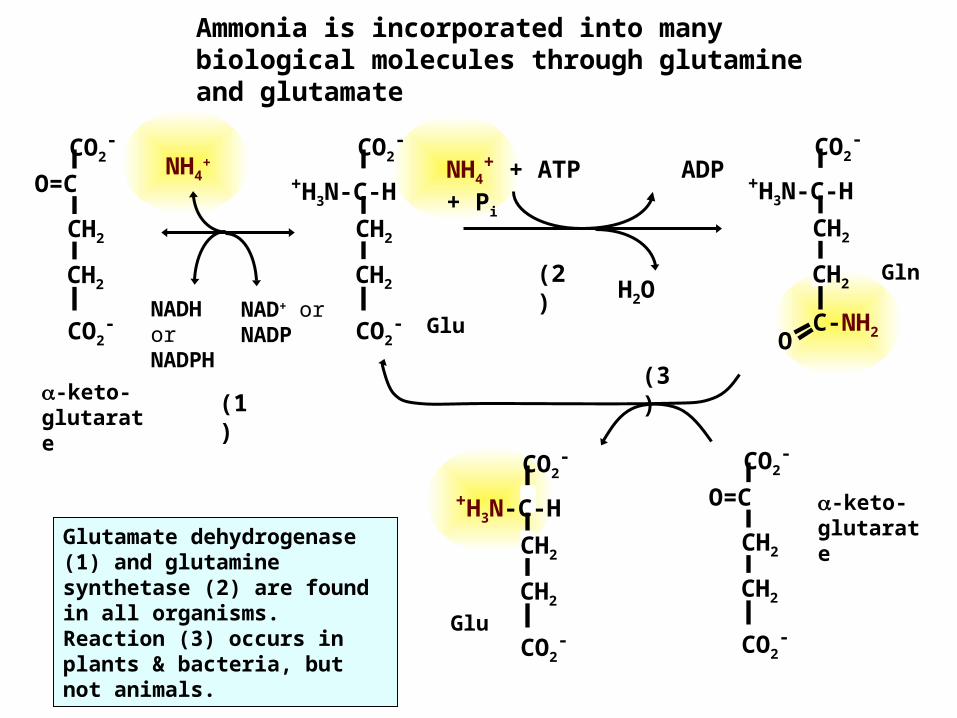
Ammonia is incorporated into many biological molecules through glutamine and glutamate
Glu
NH4+ + ATP ADP
+ Pi
H2OGln
O C-NH2
CO2-
+H3N-C-HCH2
CH2
-keto-glutarate
CO2-
CO2-
O=CCH2
CH2
CO2-
CO2-
+H3N-C-HCH2
CH2Glu
CO2-
CO2-
O=CCH2
CH2
CO2-
CO2-
+H3N-C-HCH2
CH2
-keto-glutarate
NADH or NADPH
NAD+ or NADP
NH4+
(2)
(3)(1
)
Glutamate dehydrogenase (1) and glutamine synthetase (2) are found in all organisms. Reaction (3) occurs in plants & bacteria, but not animals.
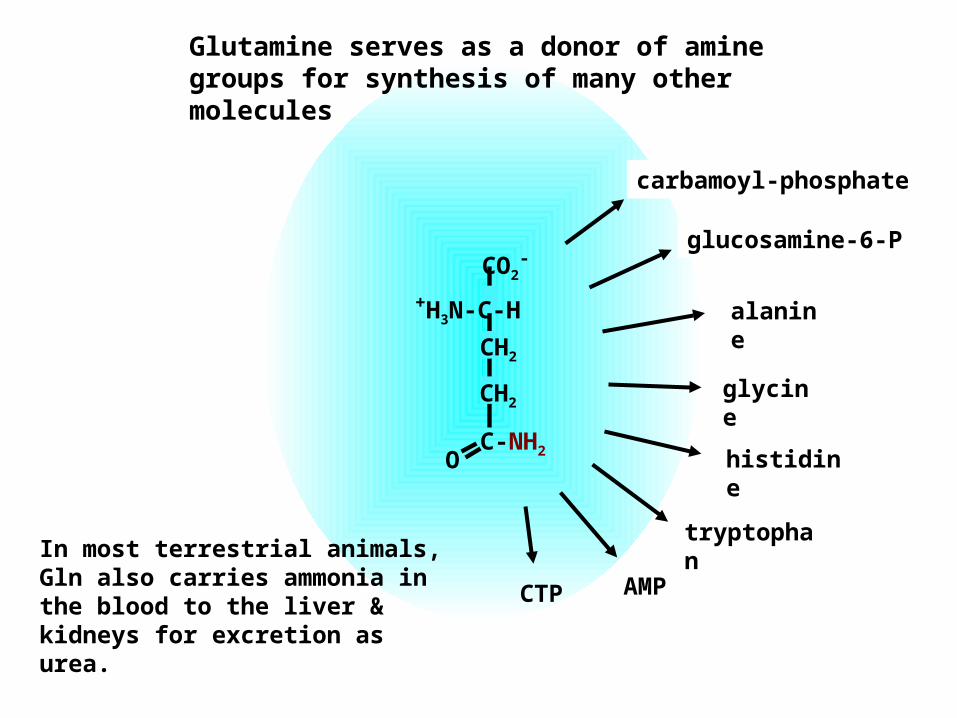
glycine
alanine
tryptophan
histidine
glucosamine-6-P
carbamoyl-phosphate
CTP AMP
O C-NH2
CO2-
+H3N-C-HCH2
CH2
Glutamine serves as a donor of amine groups for synthesis of many other molecules
In most terrestrial animals, Gln also carries ammonia in the blood to the liver & kidneys for excretion as urea.
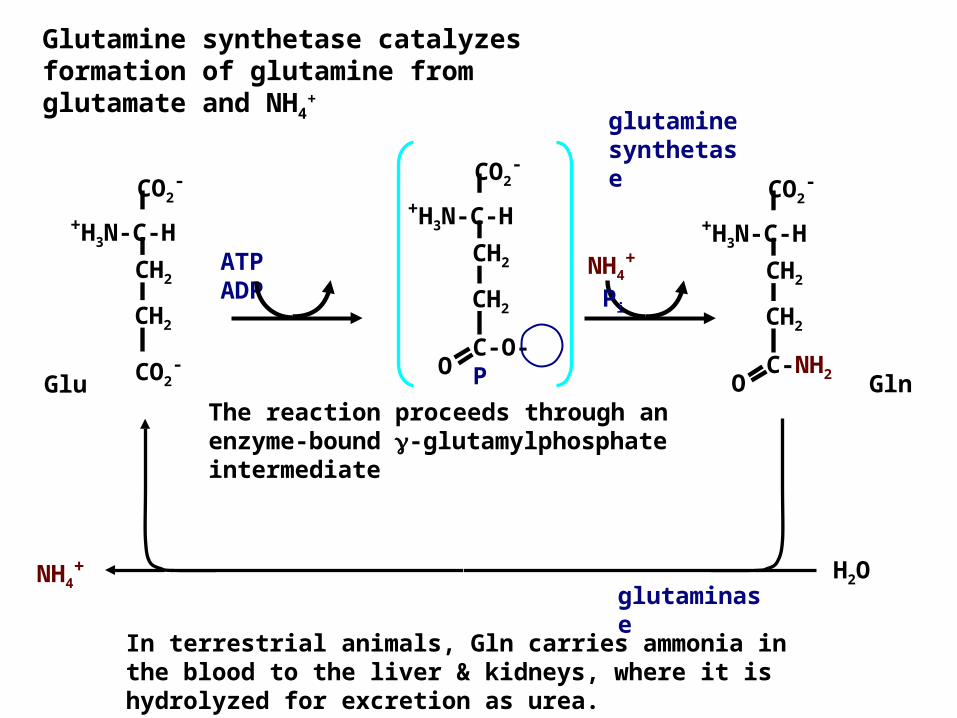
Glutamine synthetase catalyzes formation of glutamine from glutamate and NH4
+
The reaction proceeds through an enzyme-bound -glutamylphosphate intermediate
Glu GlnCO2-
CO2-
+H3N-C-HCH2
CH2
O C-O- P
CO2-
+H3N-C-HCH2
CH2
NH4+
Pi
ATP ADP
O C-NH2
CO2-
+H3N-C-HCH2
CH2
In terrestrial animals, Gln carries ammonia in the blood to the liver & kidneys, where it is hydrolyzed for excretion as urea.
H2ONH4+
glutaminase
glutamine synthetase
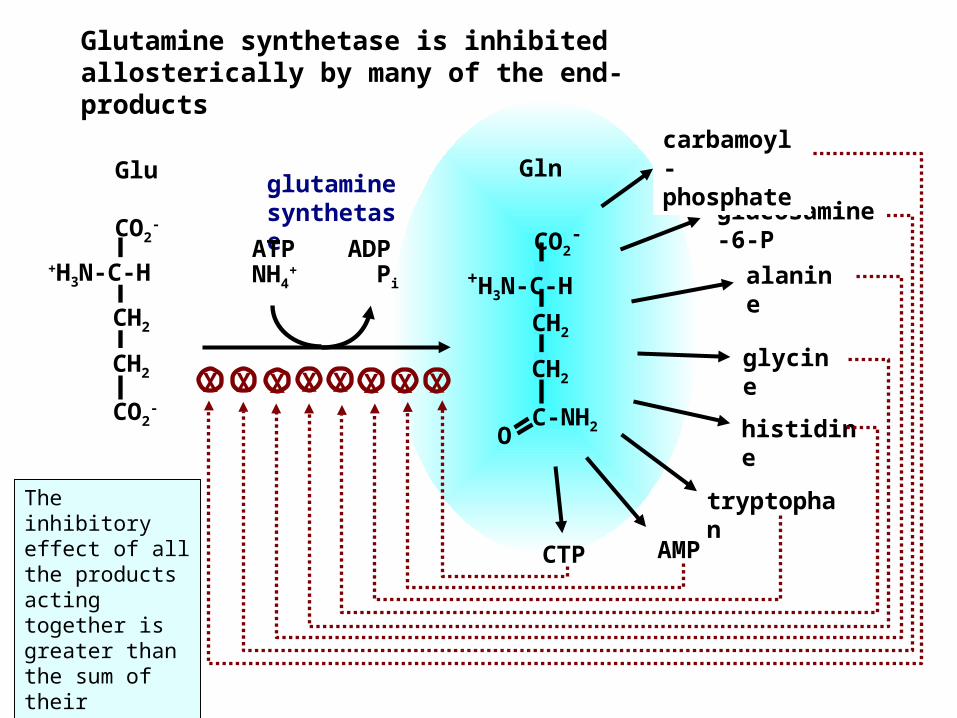
O C-NH2
CO2-
+H3N-C-HCH2
CH2glycine
alanine
tryptophan
histidine
AMP
Glutamine synthetase is inhibited allosterically by many of the end-products
GlnGluglucosamine-6-P
carbamoyl-phosphateglutamine
synthetaseATP ADP
NH4+ Pi
CTP
X X X X X X X XCO2
-
CO2-
+H3N-C-HCH2
CH2
The inhibitory effect of all the products acting together is greater than the sum of their individual effects.
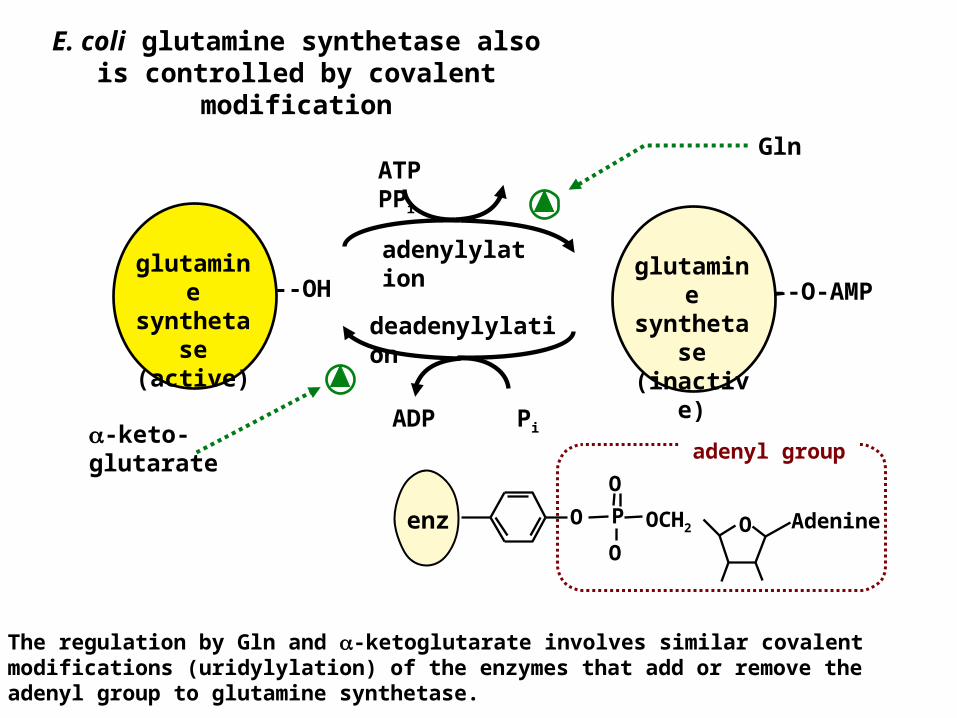
E. coli glutamine synthetase also is controlled by covalent
modification
adenylylation --O-AMP
glutamine
synthetase
(inactive)
--OHglutamin
e syntheta
se (active)
ATP PPi
deadenylylation
ADP Pi-keto-glutarate
Gln
The regulation by Gln and -ketoglutarate involves similar covalent modifications (uridylylation) of the enzymes that add or remove the adenyl group to glutamine synthetase.
enz OO OCH2
O
OP Adenine
adenyl group
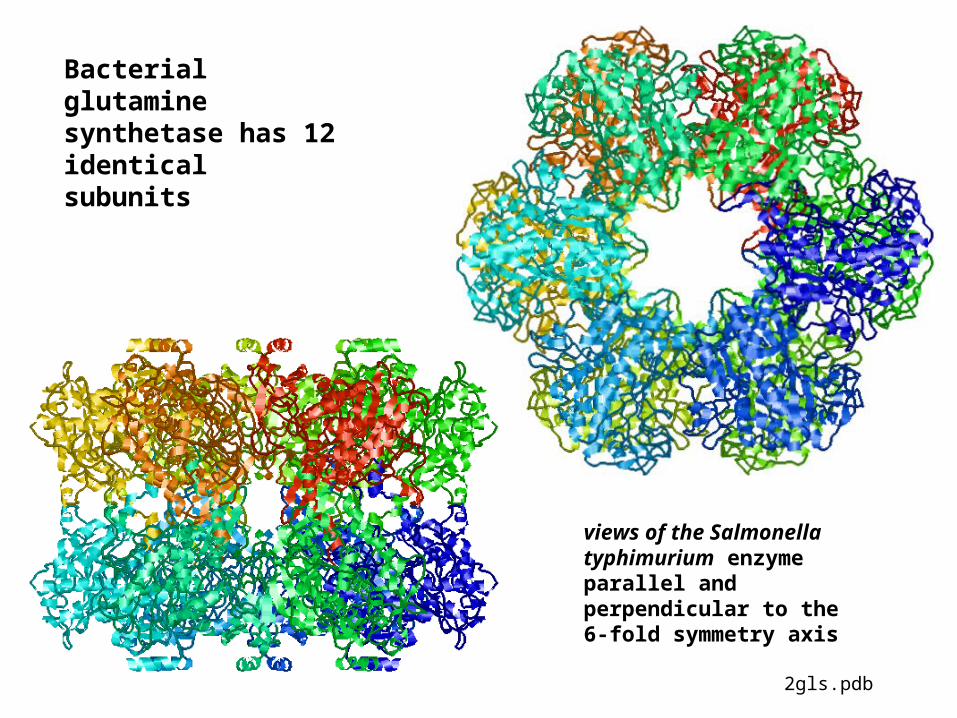
Bacterial glutamine synthetase has 12 identical subunits
views of the Salmonella typhimurium enzyme parallel and perpendicular to the 6-fold symmetry axis
2gls.pdb
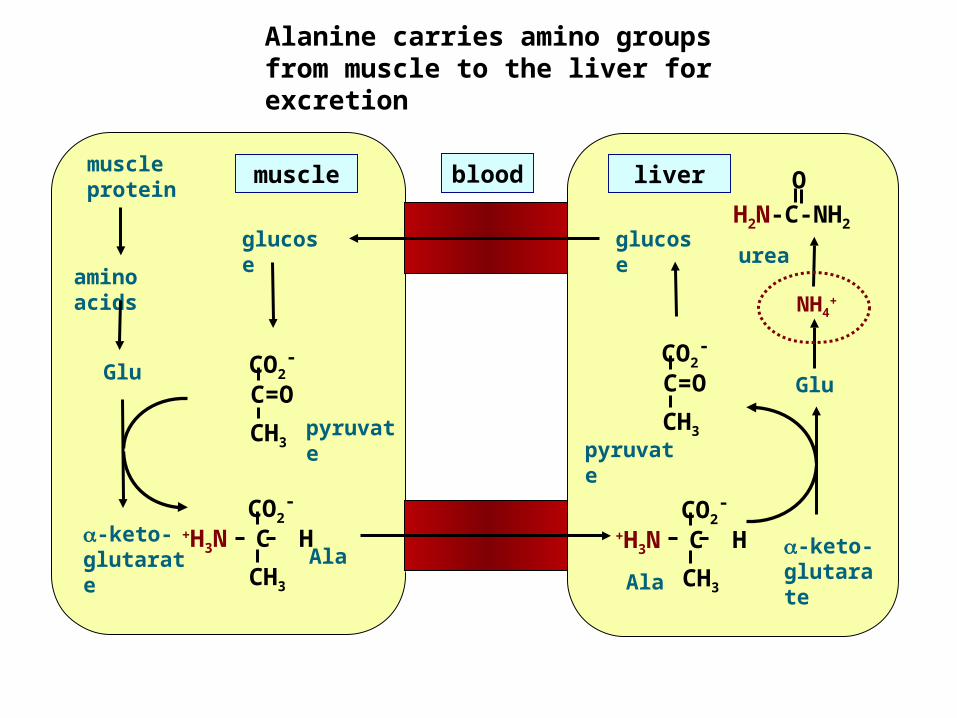
Alanine carries amino groups from muscle to the liver for excretion
Ala-keto- glutarate
pyruvate
CO2-
C=OCH3
Glu
NH4+
glucose
CO2-
+H3N C HCH3
urea
liver
Ala-keto- glutarate
muscle protein
pyruvate
CO2-
C=OCH3
Glu
amino acids
glucose
CO2-
+H3N C HCH3
muscle blood OH2N-C-NH2
Related Documents











