970 J. Parasitol., 90(5), 2004, pp. 970–979 q American Society of Parasitologists 2004 AFRICAN TRYPANOSOME INTERACTIONS WITH AN IN VITRO MODEL OF THE HUMAN BLOOD–BRAIN BARRIER Dennis J. Grab, Olga Nikolskaia, Yuri V. Kim, John D. Lonsdale-Eccles*, Susumu Ito , Tatsuru Hara‡, Toshihide Fukuma‡, Elvis Nyarko‡, Kee Jun Kim, Monique F. Stins, Michael J. Delannoy§, Jean Rodgers\, and Kwang Sik Kim Department of Pediatrics, The Johns Hopkins School of Medicine, Baltimore, Maryland 21287. e-mail: [email protected] ABSTRACT: The neurological manifestations of sleeping sickness in man are attributed to the penetration of the blood–brain barrier (BBB) and invasion of the central nervous system by Trypanosoma brucei gambiense and Trypanosoma brucei rhode- siense. However, how African trypanosomes cross the BBB remains an unresolved issue. We have examined the traversal of African trypanosomes across the human BBB using an in vitro BBB model system constructed of human brain microvascular endothelial cells (BMECs) grown on Costar Transwelly inserts. Human-infective T. b. gambiense strain IL 1852 was found to cross human BMECs far more readily than the animal-infective Trypanosoma brucei brucei strains 427 and TREU 927. Tsetse fly–infective procyclic trypomastigotes did not cross the human BMECs either alone or when coincubated with bloodstream- form T. b. gambiense. After overnight incubation, the integrity of the human BMEC monolayer measured by transendothelial electrical resistance was maintained on the inserts relative to the controls when the endothelial cells were incubated with T. b. brucei. However, decreases in electrical resistance were observed when the BMEC-coated inserts were incubated with T. b. gambiense. Light and electron microscopy studies revealed that T. b. gambiense initially bind at or near intercellular junctions before crossing the BBB paracellularly. This is the first demonstration of paracellular traversal of African trypanosomes across the BBB. Further studies are required to determine the mechanism of BBB traversal by these parasites at the cellular and molecular level. African trypanosomes are major pathogens of humans and animals. Trypanosoma congolense and Trypanosoma vivax are pathogens of cattle but do not infect humans. Two subspecies of Trypanosoma brucei, Trypanosoma brucei rhodesiense and Trypanosoma brucei gambiense, are human pathogens, whereas the closely related subspecies Trypanosoma brucei brucei is noninfective to humans. The latter is frequently used as a model for trypanosomiasis in laboratory animals. Each of the above parasites plays an important role, sometimes devastating in the health and welfare of people and cattle throughout large areas of sub-Saharan Africa. Although there were indications that hu- man trypanosomiasis (commonly called African sleeping sick- ness) was under control during the middle of the 20th century, the World Health Organization reports that several countries are seeing a resurgence of trypanosomiasis of epidemic propor- tions. Because death is inevitable if a patient is untreated (Mo- lyneux et al., 1996), human trypanosomiasis has been claimed to be more deadly than other vector-borne diseases, such as malaria (Rogers and Williams, 1993). In its terminal stage, human sleeping sickness is character- ized by muscle wasting and a general increase in nighttime insomnia and daytime drowsiness. Because the neurological symptoms of human sleeping sickness frequently occur late in the course of the disease, i.e., just before death, it is often as- sumed that the invasion of the central nervous system (CNS) occurs late in the course of the infection. However, T. b. rho- desiense can be found in the cerebrospinal fluid (CSF) of pa- Received 26 November 2003; revised 17 February 2004; accepted 23 February 2004. * Biochemistry & Molecular Genetics, University of Alabama, Bir- mingham, Alabama 35294. ² Department of Cell Biology, Harvard Medical School, Boston, Mas- sachusetts 02115. ‡ Department of Parasitology, Kurume University School of Medicine, Fukuoka 830-0011, Japan. § Department of Cell Biology and Anatomy, The Johns Hopkins School of Medicine, Baltimore, Maryland 21287. \ University of Glasgow Veterinary School, Glasgow G11 6NU, Scot- land, U.K. tients as early as 7–10 days after commencement of illness (Banks, 1980). Early infections of the CNS have also been seen in laboratory animals (Morrison et al., 1983; Schultzberg et al., 1988; Jennings et al., 1989). Interestingly, parasites that are injected directly into the brain do not survive (Schmidt and Bafort, 1987), and the CSF alone is insufficient to maintain the parasites in vitro (Hawking and Greenfield, 1941). Yet, the par- asites that cross the blood–brain barrier (BBB) in vivo remain viable. Once inside the brain, the parasites are safe from many of the most effective trypanocidal drugs (Jennings and Gray, 1983; Jennings et al., 1993). Indeed, the brain is probably the source for many relapse infections (Jennings et al., 1979). Con- sequently, if the parasites do cause any damage to the barrier, such damage must be either minimal or transient. Despite its importance, the mechanism by which the try- panosomes enter the CNS remains an unresolved issue (Dumas and Bouteille, 1997; Lonsdale-Eccles and Grab, 2002). Deter- mination of how trypanosomes get into the human brain has been hampered by the fact that the presence of parasites in the brain at autopsy has been reported only infrequently (Mott, 1907). This may be because most patients received some form of trypanocide therapy before death and that the in situ dem- onstration of parasites is only possible if the brain is fixed with- in 4–5 hr after death (Calwell, 1937). Because most autopsy specimens are obtained after 5 hr after death, the chances of finding trypanosomes are significantly diminished. The question of where trypanosomes enter the brain has therefore relied on direct animal studies or on inference from pathologic findings in human and animal tissues (or both). Trypanosoma brucei brucei infections in dogs and cattle cause severe disruptive and degenerative changes in the affected tissues. However, the na- ture and distribution of the lesions are different (Morrison et al., 1983). In dogs, severe lesions with large numbers of try- panosomes are found in the choroid plexus, and only small numbers of trypanosomes are found in meninges (Morrison et al., 1983). In cattle, there is extensive involvement within the brain. Bovine meningoencephalitis is characterized by cellular infiltration into the meninges and perivascularly throughout the

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

970
J. Parasitol., 90(5), 2004, pp. 970–979q American Society of Parasitologists 2004
AFRICAN TRYPANOSOME INTERACTIONS WITH AN IN VITRO MODEL OF THE HUMANBLOOD–BRAIN BARRIER
Dennis J. Grab, Olga Nikolskaia, Yuri V. Kim, John D. Lonsdale-Eccles*, Susumu Ito†, Tatsuru Hara‡,Toshihide Fukuma‡, Elvis Nyarko‡, Kee Jun Kim, Monique F. Stins, Michael J. Delannoy§, Jean Rodgers\, andKwang Sik KimDepartment of Pediatrics, The Johns Hopkins School of Medicine, Baltimore, Maryland 21287. e-mail: [email protected]
ABSTRACT: The neurological manifestations of sleeping sickness in man are attributed to the penetration of the blood–brainbarrier (BBB) and invasion of the central nervous system by Trypanosoma brucei gambiense and Trypanosoma brucei rhode-siense. However, how African trypanosomes cross the BBB remains an unresolved issue. We have examined the traversal ofAfrican trypanosomes across the human BBB using an in vitro BBB model system constructed of human brain microvascularendothelial cells (BMECs) grown on Costar Transwelly inserts. Human-infective T. b. gambiense strain IL 1852 was found tocross human BMECs far more readily than the animal-infective Trypanosoma brucei brucei strains 427 and TREU 927. Tsetsefly–infective procyclic trypomastigotes did not cross the human BMECs either alone or when coincubated with bloodstream-form T. b. gambiense. After overnight incubation, the integrity of the human BMEC monolayer measured by transendothelialelectrical resistance was maintained on the inserts relative to the controls when the endothelial cells were incubated with T. b.brucei. However, decreases in electrical resistance were observed when the BMEC-coated inserts were incubated with T. b.gambiense. Light and electron microscopy studies revealed that T. b. gambiense initially bind at or near intercellular junctionsbefore crossing the BBB paracellularly. This is the first demonstration of paracellular traversal of African trypanosomes acrossthe BBB. Further studies are required to determine the mechanism of BBB traversal by these parasites at the cellular and molecularlevel.
African trypanosomes are major pathogens of humans andanimals. Trypanosoma congolense and Trypanosoma vivax arepathogens of cattle but do not infect humans. Two subspeciesof Trypanosoma brucei, Trypanosoma brucei rhodesiense andTrypanosoma brucei gambiense, are human pathogens, whereasthe closely related subspecies Trypanosoma brucei brucei isnoninfective to humans. The latter is frequently used as a modelfor trypanosomiasis in laboratory animals. Each of the aboveparasites plays an important role, sometimes devastating in thehealth and welfare of people and cattle throughout large areasof sub-Saharan Africa. Although there were indications that hu-man trypanosomiasis (commonly called African sleeping sick-ness) was under control during the middle of the 20th century,the World Health Organization reports that several countries areseeing a resurgence of trypanosomiasis of epidemic propor-tions. Because death is inevitable if a patient is untreated (Mo-lyneux et al., 1996), human trypanosomiasis has been claimedto be more deadly than other vector-borne diseases, such asmalaria (Rogers and Williams, 1993).
In its terminal stage, human sleeping sickness is character-ized by muscle wasting and a general increase in nighttimeinsomnia and daytime drowsiness. Because the neurologicalsymptoms of human sleeping sickness frequently occur late inthe course of the disease, i.e., just before death, it is often as-sumed that the invasion of the central nervous system (CNS)occurs late in the course of the infection. However, T. b. rho-desiense can be found in the cerebrospinal fluid (CSF) of pa-
Received 26 November 2003; revised 17 February 2004; accepted 23February 2004.
* Biochemistry & Molecular Genetics, University of Alabama, Bir-mingham, Alabama 35294.
† Department of Cell Biology, Harvard Medical School, Boston, Mas-sachusetts 02115.
‡ Department of Parasitology, Kurume University School of Medicine,Fukuoka 830-0011, Japan.
§ Department of Cell Biology and Anatomy, The Johns Hopkins Schoolof Medicine, Baltimore, Maryland 21287.
\ University of Glasgow Veterinary School, Glasgow G11 6NU, Scot-land, U.K.
tients as early as 7–10 days after commencement of illness(Banks, 1980). Early infections of the CNS have also been seenin laboratory animals (Morrison et al., 1983; Schultzberg et al.,1988; Jennings et al., 1989). Interestingly, parasites that areinjected directly into the brain do not survive (Schmidt andBafort, 1987), and the CSF alone is insufficient to maintain theparasites in vitro (Hawking and Greenfield, 1941). Yet, the par-asites that cross the blood–brain barrier (BBB) in vivo remainviable. Once inside the brain, the parasites are safe from manyof the most effective trypanocidal drugs (Jennings and Gray,1983; Jennings et al., 1993). Indeed, the brain is probably thesource for many relapse infections (Jennings et al., 1979). Con-sequently, if the parasites do cause any damage to the barrier,such damage must be either minimal or transient.
Despite its importance, the mechanism by which the try-panosomes enter the CNS remains an unresolved issue (Dumasand Bouteille, 1997; Lonsdale-Eccles and Grab, 2002). Deter-mination of how trypanosomes get into the human brain hasbeen hampered by the fact that the presence of parasites in thebrain at autopsy has been reported only infrequently (Mott,1907). This may be because most patients received some formof trypanocide therapy before death and that the in situ dem-onstration of parasites is only possible if the brain is fixed with-in 4–5 hr after death (Calwell, 1937). Because most autopsyspecimens are obtained after 5 hr after death, the chances offinding trypanosomes are significantly diminished. The questionof where trypanosomes enter the brain has therefore relied ondirect animal studies or on inference from pathologic findingsin human and animal tissues (or both). Trypanosoma bruceibrucei infections in dogs and cattle cause severe disruptive anddegenerative changes in the affected tissues. However, the na-ture and distribution of the lesions are different (Morrison etal., 1983). In dogs, severe lesions with large numbers of try-panosomes are found in the choroid plexus, and only smallnumbers of trypanosomes are found in meninges (Morrison etal., 1983). In cattle, there is extensive involvement within thebrain. Bovine meningoencephalitis is characterized by cellularinfiltration into the meninges and perivascularly throughout the

GRAB ET AL.—BLOOD–BRAIN BARRIER TRAVERSAL BY AFRICAN TRYPANOSOMES 971
brain and spinal cord, whereas accumulation of trypanosomesin the choroid plexus is focal and less marked. Morrison et al.(1983) also point out that certain serodemes (groups of try-panosome isolates) in cattle produce more lesions in the CNSand cardiac tissue (T. b. brucei TREU 667) than others (T. b.brucei 427). Interestingly, the pathological changes in the brainsof cattle (Morrison et al., 1983) are similar to those in fatalhuman trypanosomiasis (Mott, 1907; Buchanan, 1929; Calwell,1937; Manuelidis et al., 1965).
In the mouse, T. b. rhodesiense has been observed in highlyvascularized areas of the brain, e.g., cerebral cortex, the hy-pocampus, and the nuclei of the brain stem, suggesting thatmeningoencephalitis may start from the capillaries (Fink andSchmidt, 1979). But, as in the dog (Morrison et al., 1983),trypanosomes have also been found in areas where the endo-thelial BBB is incomplete (Schmidt, 1983; Schultzberg et al.,1988); however, the surface area of the microvascular endothe-lium of BBB is approximately 5,000-fold greater than thoseareas of the brain were the barrier is incomplete such as thechoroid plexus and other circumventricular organs (http://us-ers.ahsc.arizona.edu/davis/bbbhistory.htm).
In vitro models of the BBB have clearly become importanttools for identifying the cellular and molecular elements thatmay be possible targets for interventions for the transmigrationof many pathogens into the CNS (Tuomanen, 1996; Zhang andTuomanen, 1999; Kim, 2000, 2003). Because it is not possibleto undertake in vivo experiments, an in vitro model of the BBBis essential if we are to understand how trypanosomes cross thehuman brain microvascular endothelial cells (BMECs) thatcomprise the functional unit of the human BBB. Using an ex-tensively tested in vitro model for the human BBB developedby this laboratory, we examined how African trypanosomes tra-verse the human BBB.
MATERIALS AND METHODS
Materials and biochemicals
The Transwelly tissue culture inserts (6.5 mm diameter, 3.0mm pore size) and 24-well microwell plates were purchasedfrom Corning Costar Corp (Corning, New York). Both the En-dohm Chamber and EVOM meter, used to measure transen-dothelial electrical resistance (TEER) of cell monolayers grownon tissue culture inserts, were obtained from World PrecisionInstruments (WPI, Inc, Sarasota, Florida). Electric cell substrateimpedance sensing (ECIS) was conducted using an ECIS Model1600R system from Applied BioPhysics (Troy, New York).Olympus fluorescence microscopy system (Olympus AmericaInc., Melville, New York) was equipped with an inverted Olym-pus microscope IX-70, cooled CCD camera OlymPix (modelTE3/A/S, AstroCam, U.K.), 340, 1.3–numerical aperture oil-immersion objective and computer-controlled Sutter FilterWheel (Sutter Instrument Co., Novato, California). The PKH67Green Fluorescent Cell Linker Kits for general cell membranelabeling were from Sigma (St. Louis, Missouri). Iscove’s Me-dium (used to make HMI-9), Medium 199, and Hams F-12were from Invitrogen (Carlsbad, California). Fura-2/AM andPluronic-123 were from Molecular Probes (Eugene, Oregon).Heat-inactivated fetal bovine serum (FBS) was from Hyclone(Logan, Utah) or Omega (Irvine, California). Mouse anti-hu-man ZO-1 was from Zymed (San Diego, California).
The human BBB model
The human BMEC primary cell cultures that are used for ourstudy have been previously described (Persidsky et al., 1997;Stins, Gilles et al., 1997; Stins, Prasadarao et al., 1997). Afterstudying nonendothelial markers such as glial fibrillary acidicprotein for astrocytes, smooth muscle actin to identify pericytes,galactocerebroside C for oligoglia, cytokeratin for epithelialcells, and macrophage antigens for microglia, fluorescence-ac-tivated cell sorted (FACS) human BMECs are found to be.99% pure (Persidsky et al., 1997; Stins, Gilles et al., 1997;Stins, Prasadarao et al., 1997). The tumor necrosis factor-a in-duced an increase in intracellular adhesion molecule-1 expres-sion on the apical side of the human BMEC, which is in agree-ment with data from Dorovini-Zis et al. (1992) and indicatesthat our monolayers are polarized.
Because the morphological characteristics of human BMECsare known to be passage dependent, we used a human BMECcell line whose phenotypic expression was stabilized by im-mortalizing the cells with pSVT, a pBR322-based plasmid con-taining DNA sequence encoding the SV40-large T antigen(Stins, Prasadarao et al., 1997). As in the primary humanBMEC cell line from which they were derived, the transfectedBMECs were positive for FVIII-Rag, carbonic anhydrase IV,Ulex europeus agglutinin I, take up acetylated low-density li-poprotein, and express gamma glutamyl transpeptidase (Stins,Gilles et al., 1997; Stins, Prasadarao et al., 1997). By FACSanalysis, transfected human BMECs are positive for both factorVIII-Rag and the drug-transporting (or mdr1-type) P-glycopro-tein (D. Grab and M. F. Stins, unpubl. data) known to be presentin endothelial cells that form the BBB (Schinkel, 1999; Gaillardet al., 2001). In addition, tight junctional proteins such as ZO-1 are also expressed (Fig. 5C). The high TEER of humanBMECs, relative to nonbrain endothelial cells, is dependent onthe clone and isolation (Zink et al., 1996; Lo et al., 1997; Nizetet al., 1997). It is of related interest that recent genomics datahave revealed other unique BMEC features not found or de-tected in nonbrain endothelial cells. For example, BMECs spe-cifically express BBB-specific anion transporter 1 (BSAT1), ex-pressed sequence tags; angiogenic factors like vascular endo-thelial growth factor, follistatin, fibroblast growth factor-1 andfibroblast growth factor-5; as well as CXC chemokines withELR-motifs like ELR-78 and GRO-a (Li et al., 2001; Kallmannet al., 2002).
To create in vitro models of the human BBB, transfectedhuman BMECs are seeded on top of the Transwelly tissue cul-ture inserts that have been coated with type I collagen. Afterreaching confluency, this in vitro model of the BBB forms acontinuous lining (Perisidsky et al., 1997) that allows us sepa-rate access to the upper compartment (blood side) and lowercompartment (brain side). Our in vitro BBB model has beenused to study transmigration of monocytes, bacteria, and fungi(Huang et al., 1995; Fiala et al., 1997; Nizet et al., 1997; Per-isidsky et al., 1997; Giri et al., 2000; Hoffman et al., 2000;Huang et al., 2000; Reddy et al., 2000; Jong et al., 2001; Stinset al., 2001).
The trypanosomes
Bloodstream-form T. b. gambiense IL 1852, a strain origi-nally obtained from ILRI (Nairobi, Kenya), was isolated more

972 THE JOURNAL OF PARASITOLOGY, VOL. 90, NO. 5, OCTOBER 2004
that 50 yr ago from the CSF of a Kenyan woman. Bloodstream-form T. b. brucei TREU 927 (van Deursen et al., 2001) and T.b. brucei 427 (expressing 221 VSG), as well as T. b. brucei427 and T. b. brucei TREU procyclics, were from the labora-tory of Dr. P. Englund. All bloodstream- and procyclic-formtrypanosomes in this study were maintained in culture in eitherHMI-9 (H. Hirumi and K. Hirumi, 1989) or SDM-79 (Giffinand McCann, 1993), respectively. In some experiments, blood-stream-form parasites were fluorescently tagged with the celltracker dye PKH67 according to the manufacturer’s instructions(Sigma). Green fluorescent protein (GFP)–transformed procycl-ic trypanosomes were made using a pTbsc plasmid vector usingthe method of Hara et al. (2002).
Incubation of human BMECs with African trypanosomes
Transfected human BMECs (isolate XIII) (Stins, Prasadaraoet al., 1997) were maintained in Medium 199 supplementedwith 10% heat-inactivated FBS. The cells were grown to con-fluency on 6.5-mm, collagen-coated Costar Transwelly insertswith a 3.0-mm pore size. Before the experiments, the BMECswere equilibrated for several hours in Experimental Mediumconsisting of 1 part Medium 199, 1 part Hams F-12, and 2 partsHMI-9 supplemented with 10% heat-inactivated FBS. To de-termine the ability of trypanosomes to cross our in vitro BBBmodel, the parasites were centrifuged, washed, and resuspendedin the Experimental Medium, then approximately 1 3 105 par-asites were added to the BMEC-coated Transwelly inserts (up-per compartment mimicking the blood side) of our in vitro hu-man BBB model. Trypanosomes added to Transwelly insertswithout BMECs to measure the ability of viable parasites tocross the inserts without the endothelial barrier, as well as intowells without inserts to monitor trypanosome growth under theincubation conditions, served as controls. The samples wereincubated at 37 C in 5% CO2/95% air in a humidified environ-ment. Aliquots from the wells beneath the inserts (bottom com-partment mimicking the brain side) were removed at selectedtimes. The numbers of parasites that traversed the inserts werecounted by phase-contrast microscopy using a hemocytometerand were compared with the total number of parasites presentin the assay at the times studied. Each sample was assayed intriplicate, and the data were represented as the mean 6 SEM.The viability of the trypanosomes at the end of the experimentwas unchanged, as determined by parasite motility and cell dou-bling times.
Measurements of BMEC barrier function by TEER
To assess the integrity of the human BMEC monolayersgrown on the inserts, we determined the TEER of the BMECmonolayers using an Endohm Chamber coupled with an EVOMmeter. Although TEER is often expressed as ohms/cm2, i.e.,human BMECs .300 ohm/cm2, because resistance is inverselyproportional to unit area, the unit area resistance was obtainedby multiplying the EVOM meter readings by the effective sur-face area of the Transwelly filter membranes, a value that isindependent of the area of the membrane used (WPI manual,2001). Using this system with 6.5-mm diameter inserts, we cal-culated that intact human BMEC monolayers have TEER values.25 ohm 3 cm2 ([resistance in ohms of the inserts with humanBMEC minus the resistance in ohms of the inserts alone] 3
0.33 cm2), values comparable with epithelial cells grown oninserts (Matter and Balda, 2003). For comparison, TEER fornonbrain vascular endothelium, i.e., human umbilical vein en-dothelial cells (HUVEC), measured in the same way reachedmuch lower TEER values of ,12 ohm 3 cm2 when confluencywas attained.
Statistical analyses
The sign test was used to test the null hypothesis that thepopulation median of the distribution of the paired differencesof the percent trypanosomes that will cross the BBB model iszero, versus the 2-sided alternative hypothesis. For comparingthe TEER (ohms 3 cm2) in the paired experiments, the Wil-coxon matched-pairs signed-rank test (the nonparametric analogof the paired t-test) was used to test the null hypothesis that themedian population of the paired differences in TEER is zero,versus the 2-sided alternative hypothesis that the distribution ofone member of the pair is shifted relative to that of the othermember.
Measurements of real-time TEER by ECIS
The ECIS system was used to assess the barrier function ofhuman BMEC monolayers during active trypanosome invasionin real time. ECIS measures the resistance and impedance ofsmall gold electrodes that serve as substrates for cell attachmentand growth. Time course changes of barrier function of theendothelial cells can be made (Keese, 2001) by using weak ACcurrents and computer-controlled instrumentation. In our study,human BMECs were grown on collagen-coated gold film elec-trodes in disposable 8-well microelectrode arrays. Initially,ECIS was used to monitor the time it takes the human BMECto reach confluency before equilibration in Experimental Me-dium and incubation with the trypanosomes.
Human BMEC intracellular [Ca21]i measurements
Human BMECs were grown to confluency in M199 mediumsupplemented with 10% FBS on 22-mm, collagen-coatedsquare glass coverslips. The coverslips were washed with 15mM N-2-hydroxyethylpiperazine-N9-ethane sulfonic acid (HE-PES)–buffered Hank’s balanced salt solution (HBSS) contain-ing 137 mM NaCl, 4.2 mM NaHCO3, 0.4 mM Na2HPO4, 5.4mM KCl, 0.4 mM KH2PO4, 1.3 mM CaCl2, 0.5 mM MgCl2, 0.4mM MgSO4, 5.6 mM D-glucose, 2 mM Na pyruvate, and 15mM HEPES buffered at pH 7.4 and incubated for 40 min with3 mM Fura-2/AM and 0.04% Pluronic-123 in the dark at roomtemperature. After loading, the cells were washed from extra-cellular Fura-2/AM and incubated in the same medium for anadditional 20 min. Cells loaded with Fura-2 were mounted inrecording chamber (Warner Instruments Corp., Hamden, Con-necticut) on the microscope stage, and fluorescence imageswere captured using an Olympus fluorescence microscopy sys-tem. Before starting fluorescence measurements, 15–30 regionsof interest of individual cells were selected on the field of view,and the Fura-2 fluorescent images were captured at 2-sec inter-vals by alternating excitation of cells at 340 and 380 nm wave-lengths using a reflection off dichroic mirror with a cutoffwavelength at 510 nm and bandpass emission filtering centeredat 530 nm. The real-time fluorescent images were displayed on
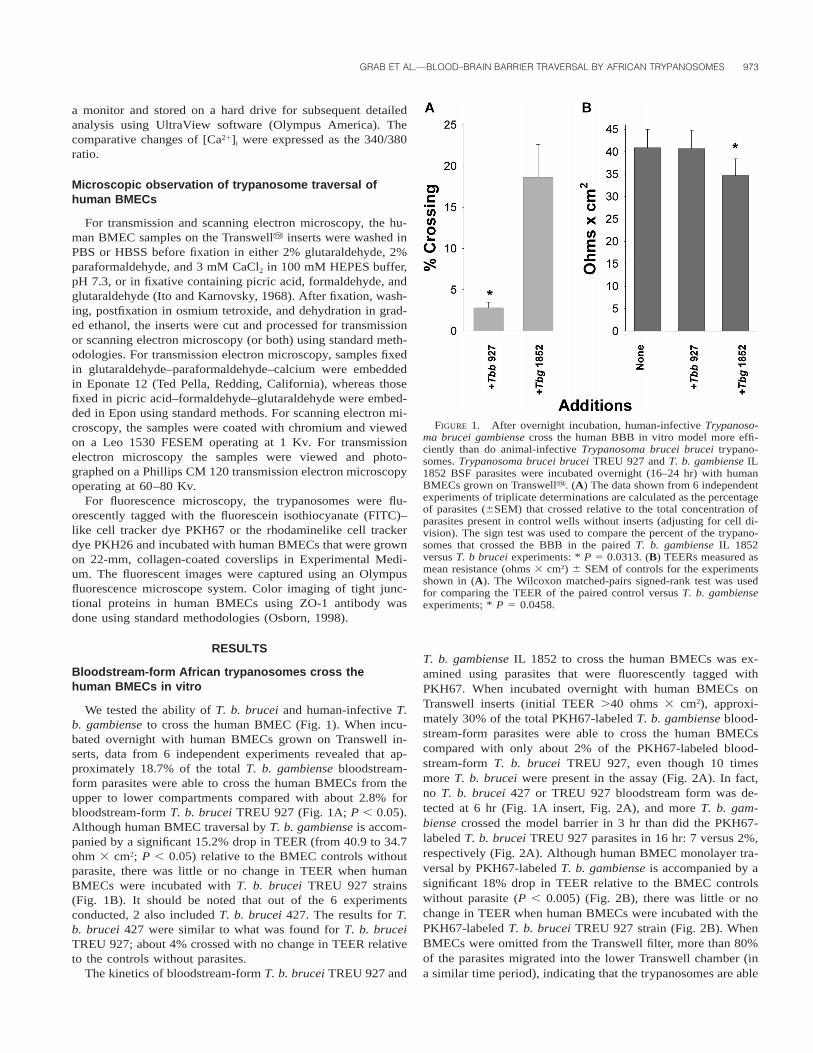
GRAB ET AL.—BLOOD–BRAIN BARRIER TRAVERSAL BY AFRICAN TRYPANOSOMES 973
FIGURE 1. After overnight incubation, human-infective Trypanoso-ma brucei gambiense cross the human BBB in vitro model more effi-ciently than do animal-infective Trypanosoma brucei brucei trypano-somes. Trypanosoma brucei brucei TREU 927 and T. b. gambiense IL1852 BSF parasites were incubated overnight (16–24 hr) with humanBMECs grown on Transwelly. (A) The data shown from 6 independentexperiments of triplicate determinations are calculated as the percentageof parasites (6SEM) that crossed relative to the total concentration ofparasites present in control wells without inserts (adjusting for cell di-vision). The sign test was used to compare the percent of the trypano-somes that crossed the BBB in the paired T. b. gambiense IL 1852versus T. b brucei experiments: * P 5 0.0313. (B) TEERs measured asmean resistance (ohms 3 cm2) 6 SEM of controls for the experimentsshown in (A). The Wilcoxon matched-pairs signed-rank test was usedfor comparing the TEER of the paired control versus T. b. gambienseexperiments; * P 5 0.0458.
a monitor and stored on a hard drive for subsequent detailedanalysis using UltraView software (Olympus America). Thecomparative changes of [Ca21]i were expressed as the 340/380ratio.
Microscopic observation of trypanosome traversal ofhuman BMECs
For transmission and scanning electron microscopy, the hu-man BMEC samples on the Transwelly inserts were washed inPBS or HBSS before fixation in either 2% glutaraldehyde, 2%paraformaldehyde, and 3 mM CaCl2 in 100 mM HEPES buffer,pH 7.3, or in fixative containing picric acid, formaldehyde, andglutaraldehyde (Ito and Karnovsky, 1968). After fixation, wash-ing, postfixation in osmium tetroxide, and dehydration in grad-ed ethanol, the inserts were cut and processed for transmissionor scanning electron microscopy (or both) using standard meth-odologies. For transmission electron microscopy, samples fixedin glutaraldehyde–paraformaldehyde–calcium were embeddedin Eponate 12 (Ted Pella, Redding, California), whereas thosefixed in picric acid–formaldehyde–glutaraldehyde were embed-ded in Epon using standard methods. For scanning electron mi-croscopy, the samples were coated with chromium and viewedon a Leo 1530 FESEM operating at 1 Kv. For transmissionelectron microscopy the samples were viewed and photo-graphed on a Phillips CM 120 transmission electron microscopyoperating at 60–80 Kv.
For fluorescence microscopy, the trypanosomes were flu-orescently tagged with the fluorescein isothiocyanate (FITC)–like cell tracker dye PKH67 or the rhodaminelike cell trackerdye PKH26 and incubated with human BMECs that were grownon 22-mm, collagen-coated coverslips in Experimental Medi-um. The fluorescent images were captured using an Olympusfluorescence microscope system. Color imaging of tight junc-tional proteins in human BMECs using ZO-1 antibody wasdone using standard methodologies (Osborn, 1998).
RESULTS
Bloodstream-form African trypanosomes cross thehuman BMECs in vitro
We tested the ability of T. b. brucei and human-infective T.b. gambiense to cross the human BMEC (Fig. 1). When incu-bated overnight with human BMECs grown on Transwell in-serts, data from 6 independent experiments revealed that ap-proximately 18.7% of the total T. b. gambiense bloodstream-form parasites were able to cross the human BMECs from theupper to lower compartments compared with about 2.8% forbloodstream-form T. b. brucei TREU 927 (Fig. 1A; P , 0.05).Although human BMEC traversal by T. b. gambiense is accom-panied by a significant 15.2% drop in TEER (from 40.9 to 34.7ohm 3 cm2; P , 0.05) relative to the BMEC controls withoutparasite, there was little or no change in TEER when humanBMECs were incubated with T. b. brucei TREU 927 strains(Fig. 1B). It should be noted that out of the 6 experimentsconducted, 2 also included T. b. brucei 427. The results for T.b. brucei 427 were similar to what was found for T. b. bruceiTREU 927; about 4% crossed with no change in TEER relativeto the controls without parasites.
The kinetics of bloodstream-form T. b. brucei TREU 927 and
T. b. gambiense IL 1852 to cross the human BMECs was ex-amined using parasites that were fluorescently tagged withPKH67. When incubated overnight with human BMECs onTranswell inserts (initial TEER .40 ohms 3 cm2), approxi-mately 30% of the total PKH67-labeled T. b. gambiense blood-stream-form parasites were able to cross the human BMECscompared with only about 2% of the PKH67-labeled blood-stream-form T. b. brucei TREU 927, even though 10 timesmore T. b. brucei were present in the assay (Fig. 2A). In fact,no T. b. brucei 427 or TREU 927 bloodstream form was de-tected at 6 hr (Fig. 1A insert, Fig. 2A), and more T. b. gam-biense crossed the model barrier in 3 hr than did the PKH67-labeled T. b. brucei TREU 927 parasites in 16 hr: 7 versus 2%,respectively (Fig. 2A). Although human BMEC monolayer tra-versal by PKH67-labeled T. b. gambiense is accompanied by asignificant 18% drop in TEER relative to the BMEC controlswithout parasite (P , 0.005) (Fig. 2B), there was little or nochange in TEER when human BMECs were incubated with thePKH67-labeled T. b. brucei TREU 927 strain (Fig. 2B). WhenBMECs were omitted from the Transwell filter, more than 80%of the parasites migrated into the lower Transwell chamber (ina similar time period), indicating that the trypanosomes are able
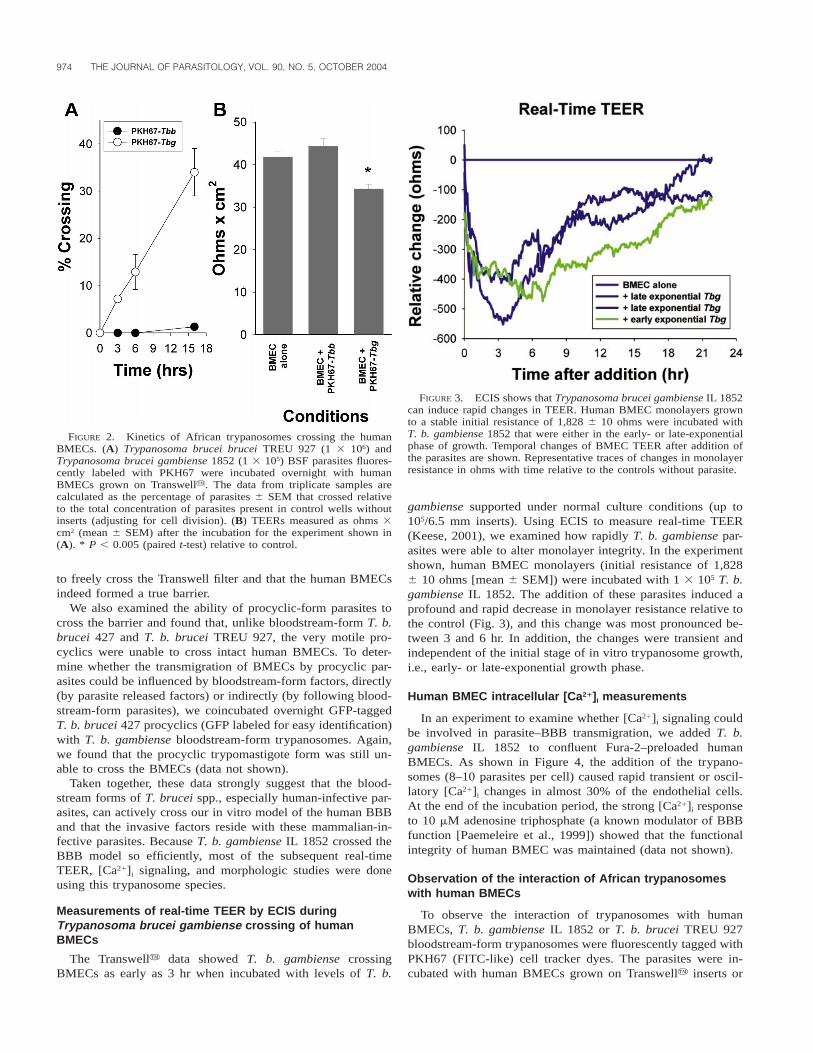
974 THE JOURNAL OF PARASITOLOGY, VOL. 90, NO. 5, OCTOBER 2004
FIGURE 2. Kinetics of African trypanosomes crossing the humanBMECs. (A) Trypanosoma brucei brucei TREU 927 (1 3 106) andTrypanosoma brucei gambiense 1852 (1 3 105) BSF parasites fluores-cently labeled with PKH67 were incubated overnight with humanBMECs grown on Transwelly. The data from triplicate samples arecalculated as the percentage of parasites 6 SEM that crossed relativeto the total concentration of parasites present in control wells withoutinserts (adjusting for cell division). (B) TEERs measured as ohms 3cm2 (mean 6 SEM) after the incubation for the experiment shown in(A). * P , 0.005 (paired t-test) relative to control.
FIGURE 3. ECIS shows that Trypanosoma brucei gambiense IL 1852can induce rapid changes in TEER. Human BMEC monolayers grownto a stable initial resistance of 1,828 6 10 ohms were incubated withT. b. gambiense 1852 that were either in the early- or late-exponentialphase of growth. Temporal changes of BMEC TEER after addition ofthe parasites are shown. Representative traces of changes in monolayerresistance in ohms with time relative to the controls without parasite.
to freely cross the Transwell filter and that the human BMECsindeed formed a true barrier.
We also examined the ability of procyclic-form parasites tocross the barrier and found that, unlike bloodstream-form T. b.brucei 427 and T. b. brucei TREU 927, the very motile pro-cyclics were unable to cross intact human BMECs. To deter-mine whether the transmigration of BMECs by procyclic par-asites could be influenced by bloodstream-form factors, directly(by parasite released factors) or indirectly (by following blood-stream-form parasites), we coincubated overnight GFP-taggedT. b. brucei 427 procyclics (GFP labeled for easy identification)with T. b. gambiense bloodstream-form trypanosomes. Again,we found that the procyclic trypomastigote form was still un-able to cross the BMECs (data not shown).
Taken together, these data strongly suggest that the blood-stream forms of T. brucei spp., especially human-infective par-asites, can actively cross our in vitro model of the human BBBand that the invasive factors reside with these mammalian-in-fective parasites. Because T. b. gambiense IL 1852 crossed theBBB model so efficiently, most of the subsequent real-timeTEER, [Ca21]i signaling, and morphologic studies were doneusing this trypanosome species.
Measurements of real-time TEER by ECIS duringTrypanosoma brucei gambiense crossing of humanBMECs
The Transwelly data showed T. b. gambiense crossingBMECs as early as 3 hr when incubated with levels of T. b.
gambiense supported under normal culture conditions (up to105/6.5 mm inserts). Using ECIS to measure real-time TEER(Keese, 2001), we examined how rapidly T. b. gambiense par-asites were able to alter monolayer integrity. In the experimentshown, human BMEC monolayers (initial resistance of 1,8286 10 ohms [mean 6 SEM]) were incubated with 1 3 105 T. b.gambiense IL 1852. The addition of these parasites induced aprofound and rapid decrease in monolayer resistance relative tothe control (Fig. 3), and this change was most pronounced be-tween 3 and 6 hr. In addition, the changes were transient andindependent of the initial stage of in vitro trypanosome growth,i.e., early- or late-exponential growth phase.
Human BMEC intracellular [Ca21]i measurements
In an experiment to examine whether [Ca21]i signaling couldbe involved in parasite–BBB transmigration, we added T. b.gambiense IL 1852 to confluent Fura-2–preloaded humanBMECs. As shown in Figure 4, the addition of the trypano-somes (8–10 parasites per cell) caused rapid transient or oscil-latory [Ca21]i changes in almost 30% of the endothelial cells.At the end of the incubation period, the strong [Ca21]i responseto 10 mM adenosine triphosphate (a known modulator of BBBfunction [Paemeleire et al., 1999]) showed that the functionalintegrity of human BMEC was maintained (data not shown).
Observation of the interaction of African trypanosomeswith human BMECs
To observe the interaction of trypanosomes with humanBMECs, T. b. gambiense IL 1852 or T. b. brucei TREU 927bloodstream-form trypanosomes were fluorescently tagged withPKH67 (FITC-like) cell tracker dyes. The parasites were in-cubated with human BMECs grown on Transwelly inserts or
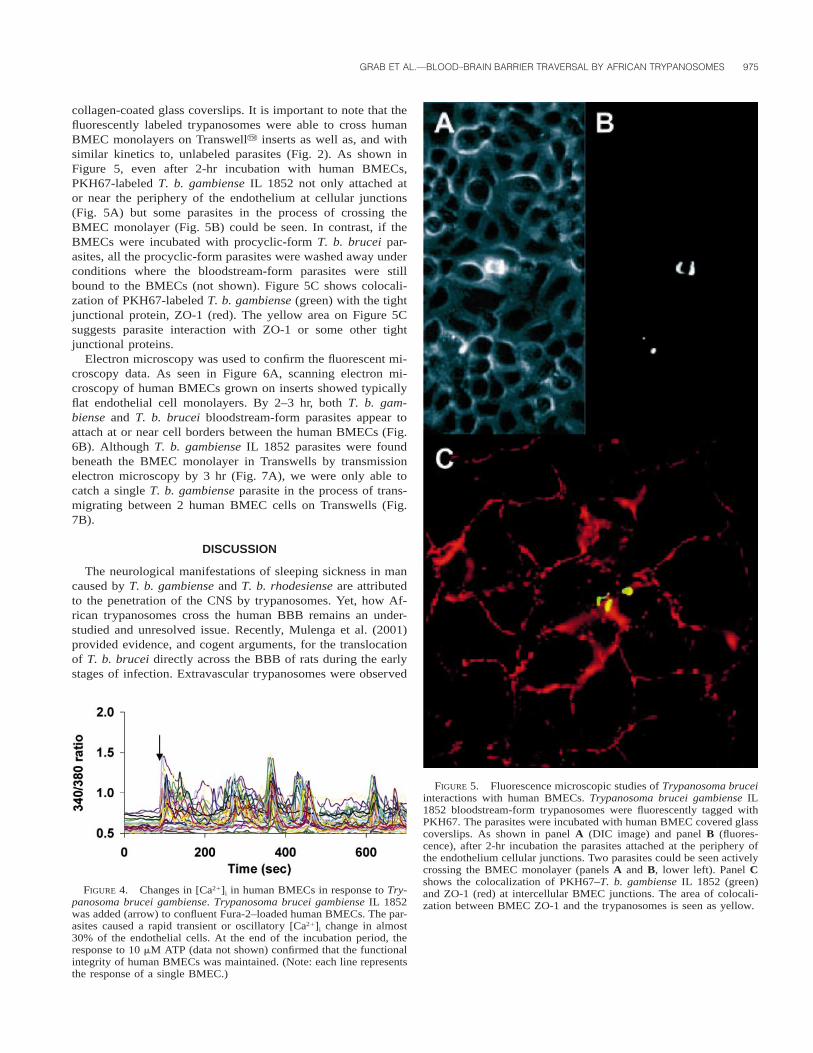
GRAB ET AL.—BLOOD–BRAIN BARRIER TRAVERSAL BY AFRICAN TRYPANOSOMES 975
FIGURE 5. Fluorescence microscopic studies of Trypanosoma bruceiinteractions with human BMECs. Trypanosoma brucei gambiense IL1852 bloodstream-form trypanosomes were fluorescently tagged withPKH67. The parasites were incubated with human BMEC covered glasscoverslips. As shown in panel A (DIC image) and panel B (fluores-cence), after 2-hr incubation the parasites attached at the periphery ofthe endothelium cellular junctions. Two parasites could be seen activelycrossing the BMEC monolayer (panels A and B, lower left). Panel Cshows the colocalization of PKH67–T. b. gambiense IL 1852 (green)and ZO-1 (red) at intercellular BMEC junctions. The area of colocali-zation between BMEC ZO-1 and the trypanosomes is seen as yellow.
FIGURE 4. Changes in [Ca21]i in human BMECs in response to Try-panosoma brucei gambiense. Trypanosoma brucei gambiense IL 1852was added (arrow) to confluent Fura-2–loaded human BMECs. The par-asites caused a rapid transient or oscillatory [Ca21]i change in almost30% of the endothelial cells. At the end of the incubation period, theresponse to 10 mM ATP (data not shown) confirmed that the functionalintegrity of human BMECs was maintained. (Note: each line representsthe response of a single BMEC.)
collagen-coated glass coverslips. It is important to note that thefluorescently labeled trypanosomes were able to cross humanBMEC monolayers on Transwelly inserts as well as, and withsimilar kinetics to, unlabeled parasites (Fig. 2). As shown inFigure 5, even after 2-hr incubation with human BMECs,PKH67-labeled T. b. gambiense IL 1852 not only attached ator near the periphery of the endothelium at cellular junctions(Fig. 5A) but some parasites in the process of crossing theBMEC monolayer (Fig. 5B) could be seen. In contrast, if theBMECs were incubated with procyclic-form T. b. brucei par-asites, all the procyclic-form parasites were washed away underconditions where the bloodstream-form parasites were stillbound to the BMECs (not shown). Figure 5C shows colocali-zation of PKH67-labeled T. b. gambiense (green) with the tightjunctional protein, ZO-1 (red). The yellow area on Figure 5Csuggests parasite interaction with ZO-1 or some other tightjunctional proteins.
Electron microscopy was used to confirm the fluorescent mi-croscopy data. As seen in Figure 6A, scanning electron mi-croscopy of human BMECs grown on inserts showed typicallyflat endothelial cell monolayers. By 2–3 hr, both T. b. gam-biense and T. b. brucei bloodstream-form parasites appear toattach at or near cell borders between the human BMECs (Fig.6B). Although T. b. gambiense IL 1852 parasites were foundbeneath the BMEC monolayer in Transwells by transmissionelectron microscopy by 3 hr (Fig. 7A), we were only able tocatch a single T. b. gambiense parasite in the process of trans-migrating between 2 human BMEC cells on Transwells (Fig.7B).
DISCUSSION
The neurological manifestations of sleeping sickness in mancaused by T. b. gambiense and T. b. rhodesiense are attributedto the penetration of the CNS by trypanosomes. Yet, how Af-rican trypanosomes cross the human BBB remains an under-studied and unresolved issue. Recently, Mulenga et al. (2001)provided evidence, and cogent arguments, for the translocationof T. b. brucei directly across the BBB of rats during the earlystages of infection. Extravascular trypanosomes were observed

976 THE JOURNAL OF PARASITOLOGY, VOL. 90, NO. 5, OCTOBER 2004
FIGURE 6. Scanning electron microscopic studies of Trypanosoma brucei interactions with human BMEC. Panel A shows a scanning electronmicroscopic image of the flat human BMEC monolayer (Transwelly insert). Panel B shows a trypanosome binding at or near cell borders ofhuman BMECs. Samples fixed in buffered glutaraldehyde–paraformaldehyde–calcium.
near capillaries. A transcellular rather than a paracellular routewas proposed because they failed to see a generalized loss oftight junction proteins (Mulenga et al., 2001). However, tightjunctions form an active component of cell–cell interactions andmay be rapidly repaired after the parasites have passed throughas can be seen after transendothelial migration of monocytes(Giri et al., 2000) and neutrophils (Burns et al., 2000). Asshown here, as a precursory event to crossing, the trypano-somes appear to first attach at or near the periphery of theendothelial cells near cellular junctions. Such binding is similarto that of T. congolense to vascular endothelial cells (Banks,1978, 1979, 1980; Hemphill et al., 1994; Shakibaei et al.,1994); these parasites also seem to prefer binding near the pe-riphery of vascular endothelial cells (Hemphill and Ross, 1995).After initial binding, a paracellular translocation of T. b. gam-biense across the in vitro model human BBB is seen with min-imal, transient breakdown of the BBB integrity. More to thepoint, according to Huber et al. (2001) ‘‘by allowing the ‘loos-ening’ of TJs and an increase in paracellular permeability, theBBB is able to ‘bend without breaking;’ thereby, maintainingstructural integrity.’’
Using an in vitro BBB system constructed of human BMECsgrown on inserts or glass coverslips, we initiated studies aimedat deciphering the mechanisms used by African trypanosomesto cross the BBB. We found that some African trypanosomes,such as the human-infective T. b. gambiense IL 1852, have ahigh potential for transendothelial migration, whereas other try-panosomes, such as animal-infective T. b. brucei 427 or TREU927, only poorly cross the human BBB. Since the clinicallyrelevant human-infective bloodstream-form T. b. gambiense IL1852 was found to efficiently cross our human BMEC model,why study T. b. brucei as this parasite is not infective to hu-mans? It is known that T. b. brucei parasites are unable to live
in human serum because of the action of 2 subfractions of se-rum high-density lipoprotein, together known as the trypano-some lytic factor (Milner and Hajduk, 1999). However, it stillwas possible that T. b. brucei possess the machinery necessaryto traverse a human BBB. Unfortunately, neither did T. b. bru-cei TREU 927, which has been chosen for complete sequencingof the African trypanosome genome (van Deursen et al., 2001),efficiently cross the barrier nor did the commonly used labo-ratory strain T. b. brucei 427. Thus, unidentified species-strain–specific factors appear to dictate why bloodstream-form T. b.gambiense crosses human BBB so efficiently, whereas animal-infective T. b. brucei cross far less effectively.
Although the pathological changes in the brains of T. b. bru-cei infected cattle (Morrison et al., 1983) are similar to thosein fatal human trypanosomiasis cases (Mott, 1907; Calwell,1937; Manuelidis et al., 1965), the finding that T. b. brucei 427was less invasive than T. b. gambiense IL 1852 may not besurprising because T. b. brucei 427 causes few CNS lesions incattle (Morrison, 1983). Furthermore, it has also been observedthat a high number of trypanosomes in the CNS of cattle areonly seen with mixed T. b. brucei and T. congolense infectionsbut not with either T. brucei or T. congolense alone (Masakeet al., 1984). This suggests that, unlike the situation for humans,cerebral trypanosomiasis in cattle involves an interdependenceof T. brucei and T. congolense and that passage across the BBBmust be a complex process. A bovine BBB model could helpresolve this issue.
Although we found that the monolayer appears to be intactwhen human BMECs were incubated overnight with T. b. bru-cei, there were often changes in Transwelly TEERs after over-night incubation with T. b. gambiense. Furthermore, changes inmonolayer integrity measurements in real time by ECIS clearlyshowed that T. b. gambiense is able to also induce early changes

GRAB ET AL.—BLOOD–BRAIN BARRIER TRAVERSAL BY AFRICAN TRYPANOSOMES 977
FIGURE 7. Transmission electron microscopic studies of Trypanosoma brucei interactions with human BMECs. Panel A shows a Trypanosomabrucei gambiense IL 1852 beneath the human BMEC monolayer (Transwelly insert) by 3 hr. Sample fixed in buffered glutaraldehyde–parafor-maldehyde–calcium. Panel B shows a transmission electron microscopic analysis of T. b. gambiense IL 1852 interactions with the human BMECmonolayer (Transwelly insert) after an overnight incubation. The entry of a trypanosome between 2 BMEC cells is shown. Sample fixed inbuffered picric acid–formaldehyde–glutaraldehyde. Tryp, trypanosome; endo, endothelial cell.
in monolayer dynamics and that these changes are transient innature. Whether the drop in TEER for T. b. gambiense foundin this study is physiologic depends on whether these changescan be linked to clinical levels of parasitemia. Because of con-straints on the number of live trypanosomes that can be countedin a hemocytometer, parasite loads between 1 3 105 and 1 3106 trypanosomes per sample were initially added into the in-serts where necessary to be able to detect sufficient numbers ofparasites crossing in the wells below the inserts. Parasitemiasof T. b. brucei of up to 1 3 106/ml blood are common in cattle,whereas in rodents parasitemias 1,000-fold higher than this arethe norm (often more than 1 3 109/ml). However, parasitemiasin humans rarely reach 1 3 102/ml. Together with ECIS-basedreal-time TEER, the use of sensitive quantitative polymerasechain reaction and fluorescence-based methodologies is cur-rently under investigation to determine the ability of low par-asite loads of T. b. gambiense as well as T. b. rhodesiense tocross the BBB model.
Although the real-time TEER data strongly support a tran-sient paracellular mode of parasite traversal across BMEC, thetrypanosome coincubation data using procyclic trypanosomesalso suggest that the changes in the monolayer integrity of theendothelium when the trypanosomes traverse the BBB may betransient in nature. We determined whether or not tsetse-infec-tive procyclic T. b. brucei could cross human BMECs in thepresence of T. b. gambiense bloodstream forms. Unlike T. b.brucei bloodstream forms, the procyclic forms express a sur-face-bound trans-sialidase that transfers sialic acid to the gly-cosylphosphatidylinositol (GPI) anchor of procyclin, a proteinthat contributes to the establishment of strong infections in thefly vector (Montagna et al., 2002). Although procyclic trypo-
mastigotes in nature would never infect a mammal, they maystill have the potential to cross the human BMEC, at least invitro, especially if the passage was a passive, nonspecific pro-cess involving passage through holes in the barrier. Apparentlythey do not, thereby indicating that the barrier at least is com-plete. It was also surmised that if T. b. gambiense bloodstream-form parasites released factors that directly affect BMEC integ-rity, the procyclics in the presence of these released factors maynow be able to cross the BMECs. Alternatively, procyclic try-panosomes could cross the BMEC paracellularly by followingbloodstream-form parasites as they traversed the monolayer. Wetested these last 2 possibilities by coincubating GFP–T. b. bru-cei 427 procyclic trypanosomes with T. b. gambiense or T. b.brucei bloodstream-form trypanosomes. Again, the inability ofprocyclic trypanosomes to cross the BMEC was not altered.Taken together, even with high numbers of T. b. gambiense, theevidence suggests that alteration of human BMEC monolayerintegrity compromised by the passage of bloodstream-form T.b. gambiense is a transient event. If it were not, then we wouldhave expected to see more procyclic-form parasites crossing theBMECs when they were coincubated with the bloodstream-form trypanosomes. This is not surprising because the BBB iscapable of rapid modulation in response to physiological stimuliat the cytoskeletal level, which enables it to protect the brainparenchyma and maintain a homeostatic environment (Huber etal., 2001).
In response to T. b. gambiense IL 1852, the subsequent earlychanges in BMEC monolayer integrity from measurements ofreal-time TEER by ECIS, though rapid, occur after the ob-served changes in intracellular Ca21. It has been shown thatcalcium response as a result of mechanical stimulation of a

978 THE JOURNAL OF PARASITOLOGY, VOL. 90, NO. 5, OCTOBER 2004
single BMEC cell is able to trigger an intercellular propagatingcalcium wave in passaged BMEC cultures (Paemeleire et al.,1999). Whether similar BBB dynamics, i.e., intracellular Ca21
and TEER, will extend to other T. b. gambiense, T. b. brucei,and T. b. rhodesiense isolates is currently under investigation.
Based in part on our findings, we propose the followingworking hypothesis as to how the trypanosomes cross the BBB.We envision that mechanical stimulation of human BMECs oc-curs during initial contact with actively swimming trypano-somes. This event appears to be coupled with signaling path-ways responsible for inducing rapid changes in [Ca21] in humanBMECs, which results in endothelial cell contraction, possiblythrough (RhoA directed?) cytoskeletal rearrangements that cre-ate tension at human BMEC tight junctions. With the possiblehelp of parasite-associated proteases and other hydrolases, ashypothesized by Lonsdale-Eccles and Grab (2002), the try-panosomes then cross through the barrier between the cells.
Our in vitro model of the human BBB mimics many of theimportant features of in vivo trypanosome interactions with theBBB. Hence, this model should be an important tool for iden-tifying the cellular and molecular elements implicated in T. b.gambiense (and other African trypanosome species) interactionswith BMECs as well as helping characterize the biochemicalmechanisms by which these parasites cross the BBB. It mayalso help identify possible targets for intervening in the trans-migration of African trypanosomes into the CNS.
ACKNOWLEDGMENTS
We thank P. David Wilson (University of Maryland, Baltimore, Mary-land) for statistical analysis and J. Stephen Dumler, Paul Englund, Kim-berly Paul, Soo Hee Lee (Johns Hopkins School of Medicine, Balti-more, Maryland), and Arthur C. Cooper (Cornell University School ofMedicine, New York, New York) for critical reading of the manuscript,many helpful comments, and suggestions during the course of thisstudy. This work, presented in part at the XIII Molecular ParasitologyMeeting, Woods Hole, Massachusetts, was supported by grant supportfrom the National Institutes of Health to D.J.G. (1 RO AI1464-01),K.S.K. (RO1-AI47225, NS26310, HL61951, AA13858), and M.F.S. (1-RO-MH63850).
LITERATURE CITED
BANKS, K. L. 1978. Binding of Trypanosoma congolense to the wallsof small blood vessels. Journal of Protozoology 25: 241–245.
———. 1979. In vitro binding of Trypanosoma congolense to eryth-rocytes. Journal of Protozoology 26: 103–108.
———. 1980. Injury induced by Trypanosoma congolense adhesion tocell membranes. Journal of Parasitolgy 66: 34–37.
BUCHANAN, J. C. R. 1929. Some clinical aspects of TrypanosomiasisRhodesiensis. Transactions of the Royal Society of Tropical Med-icine and Hygiene 23: 81–88.
BURNS, A. R., R. A. BOWDEN, S. D. MACDONELL, D. C. WALKER, T. O.ODEBUNMI, E. M DONNACHIE, S. I. SIMON, M. I. ENTMAN, AND C.W. SMITH. 2000. Analysis of tight junctions during neutrophil tran-sendothelial migration. Journal of Cell Science 113: 45–57.
CALWELL, H. G. 1937. The pathology of the brain in Rhodesian Try-panosomiasis. Transactions of the Royal Society of Tropical Med-icine and Hygiene 30: 611–624.
DOROVINI-ZIS, K., P. D. BOWMAN, AND R. PRAMEYA. 1992. Adhesion andmigration of human polymorphonuclear leukocytes across culturedbovine brain microvessel endothelial cells. Journal of Neuropa-thology and Experimental Neurology 51: 194–205.
DUMAS, M., AND B. BOUTEILLE. 1997. Current status of trypanosomiasis.Medecine Tropicale 57: 65–69.
FIALA, M., D. J. LOONEY, M. STINS, D. D. WAY, L. ZHANG, X. GAN, F.CHIAPPELLI, E. S. SCHWEITZER, P. SHAPSHAK, M. WEINAND, M. C.
GRAVES, M. WITTE, AND K. S. KIM. 1997. TNF-alpha opens a par-acellular route for HIV-1 invasion across the blood-brain barrier.Molecular Medicine 3: 553–564.
FINK, E., AND H. SCHMIDT. 1979. Meningoencephalitis in chronic Try-panosoma brucei rhodesiense infection of the white mouse. Tro-penmedizin und Parasitologie 30: 206–211.
GAILLARD, P. J., L. H. VOORWINDEN, J. L. NIELSEN, A. IVANOV, R. AT-SUMI, H. ENGMAN, C. RINGBOM, A. G. DE BOER, AND D. D. BREIMER.2001. Establishment and functional characterization of an in vitromodel of the blood-brain barrier, comprising a co-culture of braincapillary endothelial cells and astrocytes. European Journal ofPharmacological Sciences 12: 215–222.
GIFFIN, B. F., AND P. P. MCCANN. 1993. Altered intracellular polyaminesin bloodstream form Trypanosoma brucei brucei: Transformationto procyclic trypomastigotes. Acta Tropica 55: 181–190.
GIRI, R., Y. SHEN, M. STINS, S. DU YAN, A. M. SCHMIDT, D. STERN, K.S. KIM, B. ZLOKOVIC, AND V. K. KALRA. 2000. Beta-amyloid-in-duced migration of monocytes across human brain endothelial cellsinvolves RAGE and PECAM-1. American Journal Physiology. CellPhysiology 279: C1772–C1781.
HARA, T., K. YASUDA, AND T. FUKUMA. 2002. Effective gene transferinto Trypanosoma brucei bloodstream forms by particle bombard-ment. Molecular Biochemical Parasitology 119: 117–119.
HAWKING, F., AND J. D. GREENFIELD. 1941. Two autopsies on Rhode-siense sleeping sickness; visceral lesions and significance of chang-es in cerebrospinal fluid. Transactions of the Royal Society of Trop-ical Medicine and Hygiene 35: 155–164.
HEMPHILL, A., I. FRAME, AND C. A. ROSS. 1994. The interaction of Try-panosoma congolense with endothelial cells. Parasitology 109:631–641.
———, AND C. A. ROSS. 1995. Flagellum-mediated adhesion of Try-panosoma congolense to bovine aorta endothelial cells. Parasitol-ogy Research 81: 412–420.
HIRUMI, H., AND K. HIRUMI. 1989. Continuous cultivation of Trypano-soma brucei blood stream forms in a medium containing a lowconcentration of serum protein without feeder cell layers. Journalof Parasitology 75: 985–989.
HOFFMAN, J. A., J. L. BADGER, Y. ZHANG, S. H. HUANG, AND K. S. KIM.2000. Escherichia coli K1 aslA contributes to invasion of brainmicrovascular endothelial cells in vitro and in vivo. Infection andImmunity 68: 5062–5067.
HUANG, S. H., M. F. STINS, AND K. S. KIM. 2000. Bacterial penetrationacross the blood-brain barrier during the development of neonatalmeningitis. Microbes and Infection 2: 1237–1244.
———, C. WASS, Q. FU, N. V. PRASADARAO, M. STINS, AND K. S. KIM.1995. Escherichia coli invasion of brain microvascular endothelialcells in vitro and in vivo: Molecular cloning and characterizationof invasion gene ibe10. Infection and Immunity 63: 4470–4475.
HUBER, J. D., R. D. EGLETON, AND T. P. DAVIS. 2001. Molecular phys-iology and pathophysiology of tight junctions in the blood–brainbarrier. Trends in Neuroscience 24: 719–725.
ITO, S., AND M. J. KARNOVSKY. 1968. Formaldehyde-glutaraldehyde fix-atives containing trinitro compounds. Journal of Cell Biology 34:168a.
JENNINGS, F. W., AND G. D. GRAY. 1983. Relapsed parasitaemia follow-ing chemotherapy of chronic T. brucei infections in mice and itsrelation to cerebral trypanosomes. Contributions to Microbiologyand Immunology 7: 147–154.
———, C. A. HUNTER, P. G. KENNEDY, AND M. MURRAY. 1993. Che-motherapy of Trypanosoma brucei infection of the central nervoussystem: The use of a rapid chemotherapeutic regimen and the de-velopment of post-treatment encephalopathies. Transactions of theRoyal Society of Tropical Medicine and Hygiene 87: 224–226.
———, P. E. MCNEIL, J. M. NDUNG’U, AND M. MURRAY. 1989. Try-panosomiasis and encephalitis: Possible aetiology and treatment.Transactions of the Royal Society of Tropical Medicine and Hy-giene 83: 518–519.
———, D. D. WHITELAW, P. H. HOLMES, H. G. CHIZYUKA, AND G. M.URQUHART. 1979. The brain as a source of relapsing Trypanosomabrucei infection in mice after chemotherapy. International Journalfor Parasitology 9: 381–384.
JONG, A. Y., M. F. STINS, S. H. HUANG, S. H. CHEN, AND K. S. KIM.

GRAB ET AL.—BLOOD–BRAIN BARRIER TRAVERSAL BY AFRICAN TRYPANOSOMES 979
2001. Traversal of Candida albicans across human blood-brain bar-rier in vitro. Infection and Immunity 69: 4534–4536.
KALLMANN, B. A., S. WAGNER, B. HUMMEL, M. BUTTMANN, A. BAYAS,J. C. TONN, AND P. RIECKMANN. 2002. Characteristic gene expres-sion profile of primary human cerebral endothelial cells. FASEBJournal 16: 589–591.
KEESE, C. 2001. Monitoring of cells during signal transduction. GeneticEngineering News 21: 51.
KIM, K. S. 2000. E. coli invasion of brain microvascular endothelialcells as a pathogenetic basis of meningitis. Subcellular Biochem-istry 33: 47–59.
———. 2003. Pathogenesis of bacterial meningitis: From bacteraemiato neuronal injury. Nature Reviews. Neuroscience 4: 376–385.
LI, J. Y., R. J. BOADO, AND W. M. PARDRIDGE. 2001. Blood-brain barriergenomics. Journal of Cerebral Blood Flow and Metabolism 21: 61–68.
LO, H. J., J. R. KOHLER, B. DIDOMENICO, D. LOEBENBERG, A. CACCIAP-UOTI, AND G. R. FINK. 1997. Nonfilamentous C. albicans mutantsare avirulent. Cell 90: 939–949.
LONSDALE-ECCLES, J. D., AND D. J. GRAB. 2002. Trypanosome hydro-lases and the blood-brain barrier. Trends in Parasitology 18: 17–19.
MANUELIDIS, E. E., D. H. ROBERTSON, J. M. AMBERSON, M. POLAK, AND
W. HAYMAKER. 1965. Trypanosoma rhodesiense encephalitis. Clin-icopathological study of five cases of encephalitis and one of melB hemorrhagic encephalopathy. Acta Neuropathologica (Berlin) 5:176–204.
MASAKE, R. A., V. M. NANTULYA, G. W. AKOL, AND A. J. MUSOKE.1984. Cerebral trypanosomiasis in cattle with mixed Trypanosomacongolense and T. brucei brucei infections. Acta Tropica 41: 237–246.
MATTER, K., AND M. S. BALDA. 2003. Functional analysis of tight junc-tions. Methods 30: 228–234.
MILNER, J. D., AND S. L. HAJDUK. 1999. Expression and localization ofserum resistance associated protein in Trypanosoma brucei rho-desiense. Molecular and Biochemical Parasitology 104: 271–283.
MOLYNEUX, D. H., V. PENTREATH, AND F. DOUA. 1996. African trypano-somiasis in man. In Manson’s tropical diseases, G. C. Cook (ed.).W. B. Saunders, London, U.K., p. 1171–1196.
MONTAGNA, G., M. L. CREMONA, G. PARIS, M. F. AMAYA, A. BUSCHIAZ-ZO, P. M. ALZARI, AND A. C. FRASCH. 2002. The trans-sialidase fromthe African trypanosome Trypanosoma brucei. European Journalof Biochemistry 269: 2941–2950.
MORRISON, W. I., M. MURRAY, D. D. WHITELAW, AND P. D. SAYER. 1983.Pathology of infection with Trypanosoma brucei: Disease syn-dromes in dogs and cattle resulting from severe tissue damage.Contributions to Microbiology and Immunology 7: 103–119.
MOTT, F. W. 1907. Histological observations on the changes in the ner-vous system in trypanosome infections especially sleeping sicknessand their relation to syphilitic lesions of the nervous system. Ar-chives of Neurology 3: 581–646.
MULENGA, C., J. D. MHLANGA, K. KRISTENSSON, AND B. ROBERTSON.2001. Trypanosoma brucei brucei crosses the blood-brain barrierwhile tight junction proteins are preserved in a rat chronic diseasemodel. Neuropathology and Applied Neurobiology 27: 77–85.
NIZET, V., K. S. KIM, M. STINS, M. JONAS, E. Y. CHI, D. NGUYEN, AND
C. E. RUBENS. 1997. Invasion of brain microvascular endothelialcells by group B streptococci. Infection and Immunity 65: 5074–5081.
OSBORN, M. 1998. Immunofluorescence microscopy of cultured cells.
In Cell biology: A laboratory handbook, 2nd ed., vol. 2, Julio E.Celis (ed.). Academic Press, San Diego, California, p. 462–468.
PAEMELEIRE, K., A. DE HEMPTINNE, AND L. LEYBAERT. 1999. Chemically,mechanically, and hyperosmolarity-induced calcium responses ofrat cortical capillary endothelial cells in culture. ExperimentalBrain Research 126: 473–481.
PERSIDSKY, Y., M. STINS, D. WAY, M. H. WITTE, M. WEINAND, K. S.KIM, P. BOCK, H. E. GENDELMAN, AND M. FIALA. 1997. A modelfor monocyte migration through the blood-brain barrier duringHIV-1 encephalitis. Journal of Immunology 158: 3499–3510.
REDDY, M. A., C. A. WASS, K. S. KIM, D. D. SCHLAEPFER, AND N. V.PRASADARAO. 2000. Involvement of focal adhesion kinase in Esch-erichia coli invasion of human brain microvascular endothelialcells. Infection and Immunity 68: 6423–6430.
ROGERS, D. J., AND B. G. WILLIAMS. 1993. Monitoring trypanosomiasisin space and time. Parasitology 106: S77–S92.
SCHINKEL, A. H. 1999. P-Glycoprotein, a gatekeeper in the blood-brainbarrier. Advanced Drug Delivery Reviews 36: 179–194.
SCHMIDT, H. 1983. The pathogenesis of trypanosomiasis of the CNS.Studies on parasitological and neurohistological findings in Try-panosoma rhodesiense-infected vervet monkeys. Virchows Ar-chives. A, Pathological Anatomy and Histopathology 399: 333–343.
———, AND J. M. BAFORT. 1987. African trypanosomiasis: Haemato-genic brain parasitism early in experimental infection through by-passing the blood-brain barrier, with considerations on brain try-panosomiasis in man. Parasitology Research 73: 15–21.
SCHULTZBERG, M., M. AMBATSIS, E. B. SAMUELSSON, K. KRISTENSSON,AND N. VAN MEIRVENNE. 1988. Spread of Trypanosoma brucei tothe nervous system: Early attack on circumventricular organs andsensory ganglia. Journal of Neuroscience Research 21: 56–61.
SHAKIBAEI, M., M. MILANINEZHAD, AND H. J. RISSE. 1994. Immunoelec-tron microscopic studies on the specific adhesion of Trypanosomacongolense to cultured vascular endothelial cells. Journal of Struc-tural Biology 112: 125–135.
STINS, M. F., J. BADGER, AND K. S. KIM. 2001. Bacterial invasion andtranscytosis in transfected human brain microvascular endothelialcells. Microbial Pathogenesis 30: 19–28.
———, F. GILLES, AND K. S. KIM. 1997. Selective expression of ad-hesion molecules on human brain microvascular endothelial cells.Journal of Neuroimmunology 76: 81–90.
———, N. V. PRASADARAO, J. ZHOU, M. ARDITI, AND K. S. KIM. 1997.Bovine brain microvascular endothelial cells transfected withSV40-large T antigen: Development of an immortalized cell lineto study pathophysiology of CNS disease. In Vitro Cellular & De-velopmental Biology. Animal 33: 243–247.
TUOMANEN, E. 1996. Entry of pathogens into the central nervous sys-tem. FEMS Microbiology Reviews 18: 289–299.
VAN DEURSEN, F. J., S. K. SHAHI, C. M. TURNER, C. HARTMANN, C.GUERRA-GIRALDEZ, K. R. MATTHEWS, AND C. E. CLAYTON. 2001.Characterisation of the growth and differentiation in vivo and invitro of bloodstream-form Trypanosoma brucei strain TREU 927.Molecular and Biochemical Parasitology 112: 163–171.
WPI MANUAL. 2001. EVOM & EVOMX epithelial voltohmmetersWorld Precision Instruments instruction manual. World PrecisionInstruments, Inc., Sarasota, Florida, p. 23.
ZHANG, J. R., AND E. TUOMANEN. 1999. Molecular and cellular mecha-nisms for microbial entry into the CNS. Journal of Neurovirology5: 591–603.
ZINK, S., T. NASS, P. ROSEN, AND J. F. ERNST. 1996. Migration of thefungal pathogen Candida albicans across endothelial monolayers.Infection and Immunity 64: 5085–5091.
Related Documents











