Acid-sensing ion channels are involved in epithelial Na uptake in the rainbow trout Oncorhynchus mykiss Agnieszka K. Dymowska, Aaron G. Schultz, Salvatore D. Blair, Danuta Chamot, and Greg G. Goss Department of Biological Sciences, University of Alberta, Edmonton, Alberta, Canada Submitted 30 December 2013; accepted in final form 27 May 2014 Dymowska AK, Schultz AG, Blair SD, Chamot D, Goss GG. Acid- sensing ion channels are involved in epithelial Na uptake in the rainbow trout Oncorhynchus mykiss. Am J Physiol Cell Physiol 307: C255–C265, 2014. First published June 4, 2014; doi:10.1152/ajpcell.00398.2013.—A role for acid-sensing ion channels (ASICs) to serve as epithelial channels for Na uptake by the gill of freshwater rainbow trout was investigated. We found that the ASIC inhibitors 4=,6-diamidino-2- phenylindole and diminazene decreased Na uptake in adult rainbow trout in a dose-dependent manner, with IC 50 values of 0.12 and 0.96 M, respectively. Furthermore, we cloned the trout ASIC1 and ASIC4 homologs and demonstrated that they are expressed differen- tially in the tissues of the rainbow trout, including gills and isolated mitochondrion-rich cells. Immunohistochemical analysis using cus- tom-made anti-zASIC4.2 antibody and the Na -K -ATPase ( 5 - subunit) antibody demonstrated that the trout ASIC localizes to Na /K -ATPase-rich cells in the gill. Moreover, three-dimensional rendering of confocal micrographs demonstrated that ASIC is found in the apical region of mitochondrion-rich cells. We present a revised model whereby ASIC4 is proposed as one mechanism for Na uptake from dilute freshwater in the gill of rainbow trout. sodium uptake; gill; acid-sensing ion channels; mitochondrion-rich cells; fish; ionoregulation FISHES LIVING IN FRESHWATER (FW) must actively take up Na against a steep concentration gradient. Na uptake occurs via specialized mitochondrion-rich cells (MRCs), located on the fish gill epithelium (26, 29), and was initially proposed to be linked to NH 4 /H excretion (26). Subsequent studies have described two models of Na transport (for review see Refs. 8 and 18). In the first proposed model, Na is exchanged for H via an electroneutral Na /H exchanger (NHE) located on the apical membrane of MRCs in fish gill epithelia. The identifi- cation of multiple NHEs in the gills of zebrafish (Danio rerio) (51), Osorezan dace (Tribolodon hakonensis) (15), rainbow trout (Oncorhynchus mykiss) (7, 19), and tilapia (Oreochromis mossambicus) (48) by immunocytochemistry, Western blot analysis, and RT-PCR, supports this model. However, signif- icant thermodynamic constraints associated with the function of NHEs at very low ion concentrations (Na 0.1 mM) and low environmental pH (pH 5) (1, 34) suggest that fishes living in very soft and poorly buffered water would not be able to rely on a NHE-based mechanism for sufficient Na uptake. Recently, the NHE model was revised, whereby the ammonia transporter [rhesus (Rh) protein] present on the apical mem- brane of MRCs (30, 31) forms a functional metabolon with NHE2/3 (50). The revised model does alleviate the thermody- namic constraints associated with a low-pH environment, but not those imposed by low Na concentrations in the FW aquatic environment (6). Therefore, it is unlikely that this mechanism is the sole contributor to Na uptake by FW fishes living in low-ionic-strength and poorly buffered waters. The second proposed model for Na uptake is based on the Na uptake models in frog skin (for review see Ref. 14). In this model, an epithelial Na channel (ENaC) is electrochemically linked to an apical vacuolar-type H -ATPase (VHA) (1). Several studies have provided functional evidence supporting the role of VHA in Na uptake in FW fish gills (9, 39) and apical localization of VHA in gill MRCs (44). However, a major drawback of this model has been the inability to identify the channel responsible for Na uptake, since no ENaC ho- mologs have been found in the available teleost fish genomes. Therefore, we hypothesized that other channels related to ENaCs may perform this function. ENaC is a member of the amiloride-sensitive ENaC/degen- erin family of ion channels (23). In vertebrates, the closest relatives to ENaC are the acid-sensing ion channels (ASICs), with which they share 25% identity (24). Therefore, we investigated the possibility that ASICs may be responsible for Na uptake in FW fishes. ASICs are voltage-insensitive Na channels gated by extracellular H (47) and are associated with the nervous system of mammals and fishes (16, 35, 40, 42, 46). They have been implicated in mechanoreception and sensory transduction of taste and pain (for review see Ref. 23). In mammals, at least seven different ASIC subunits (ASIC1a, ASIC1b, ASIC2a, ASIC2b, ASIC3, ASIC4, and ASIC5) are encoded by five ASIC genes, while in zebrafish, six ASIC subunits (ASIC1.1, ASIC1.2, ASIC1.3, ASIC2, ASIC4.1, and ASIC4.2) are encoded by six different genes (13, 35, 40). No ASIC3 or ASIC5 has been demonstrated in zebrafish. Recently, Chen and colleagues (5) described a group of potent ASIC inhibitors, the diarylamidines. Diarylamidines are a non- amiloride-derived ASIC blockers class of compounds that inhibit ASIC currents at very low concentrations but do not have an effect on ENaC. This finding can be exploited to pharmacologically investigate a potential role for ASICs in Na uptake in FW fishes. In this study we used a variety of whole-animal, cellular, and molecular approaches to investigate a potential role for ASICs in Na uptake in FW fishes. We used FW rainbow trout as our model organism, since their natural habitat includes very-low- ionic-strength waters with Na concentrations and pH levels below the theoretical limits of the NHE/Rh metabolon model. We identified two trout subunits of ASICs (ASIC1 and ASIC4) that are expressed in gill MRCs and demonstrated an apical MRC localization for trout ASIC4. This is the first study to identify a role for ASICs in Na uptake in FW fishes and is also the first study to demonstrate a nonneuronal function for ASICs in an organism. Address for reprint requests and other correspondence: G. G. Goss, Dept. of Biological Sciences, Univ. of Alberta, Edmonton, AB, Canada T6G 2E9 (e-mail: [email protected]). Am J Physiol Cell Physiol 307: C255–C265, 2014. First published June 4, 2014; doi:10.1152/ajpcell.00398.2013. 0363-6143/14 Copyright © 2014 the American Physiological Society http://www.ajpcell.org C255

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Acid-sensing ion channels are involved in epithelial Na uptake in therainbow trout Oncorhynchus mykiss
Agnieszka K Dymowska Aaron G Schultz Salvatore D Blair Danuta Chamot and Greg G GossDepartment of Biological Sciences University of Alberta Edmonton Alberta Canada
Submitted 30 December 2013 accepted in final form 27 May 2014
Dymowska AK Schultz AG Blair SD Chamot D Goss GG Acid-sensing ion channels are involved in epithelial Na uptake in the rainbowtrout Oncorhynchus mykiss Am J Physiol Cell Physiol 307 C255ndashC2652014 First published June 4 2014 doi101152ajpcell003982013mdashArole for acid-sensing ion channels (ASICs) to serve as epithelialchannels for Na uptake by the gill of freshwater rainbow trout wasinvestigated We found that the ASIC inhibitors 4=6-diamidino-2-phenylindole and diminazene decreased Na uptake in adult rainbowtrout in a dose-dependent manner with IC50 values of 012 and 096M respectively Furthermore we cloned the trout ASIC1 andASIC4 homologs and demonstrated that they are expressed differen-tially in the tissues of the rainbow trout including gills and isolatedmitochondrion-rich cells Immunohistochemical analysis using cus-tom-made anti-zASIC42 antibody and the Na-K-ATPase (5-subunit) antibody demonstrated that the trout ASIC localizes toNaK-ATPase-rich cells in the gill Moreover three-dimensionalrendering of confocal micrographs demonstrated that ASIC is foundin the apical region of mitochondrion-rich cells We present a revisedmodel whereby ASIC4 is proposed as one mechanism for Na uptakefrom dilute freshwater in the gill of rainbow trout
sodium uptake gill acid-sensing ion channels mitochondrion-richcells fish ionoregulation
FISHES LIVING IN FRESHWATER (FW) must actively take up Na
against a steep concentration gradient Na uptake occurs viaspecialized mitochondrion-rich cells (MRCs) located on thefish gill epithelium (26 29) and was initially proposed to belinked to NH4
H excretion (26) Subsequent studies havedescribed two models of Na transport (for review see Refs 8and 18) In the first proposed model Na is exchanged for H
via an electroneutral NaH exchanger (NHE) located on theapical membrane of MRCs in fish gill epithelia The identifi-cation of multiple NHEs in the gills of zebrafish (Danio rerio)(51) Osorezan dace (Tribolodon hakonensis) (15) rainbowtrout (Oncorhynchus mykiss) (7 19) and tilapia (Oreochromismossambicus) (48) by immunocytochemistry Western blotanalysis and RT-PCR supports this model However signif-icant thermodynamic constraints associated with the functionof NHEs at very low ion concentrations (Na 01 mM) andlow environmental pH (pH 5) (1 34) suggest that fishesliving in very soft and poorly buffered water would not be ableto rely on a NHE-based mechanism for sufficient Na uptakeRecently the NHE model was revised whereby the ammoniatransporter [rhesus (Rh) protein] present on the apical mem-brane of MRCs (30 31) forms a functional metabolon withNHE23 (50) The revised model does alleviate the thermody-namic constraints associated with a low-pH environment butnot those imposed by low Na concentrations in the FW
aquatic environment (6) Therefore it is unlikely that thismechanism is the sole contributor to Na uptake by FW fishesliving in low-ionic-strength and poorly buffered waters Thesecond proposed model for Na uptake is based on the Na
uptake models in frog skin (for review see Ref 14) In thismodel an epithelial Na channel (ENaC) is electrochemicallylinked to an apical vacuolar-type H-ATPase (VHA) (1)Several studies have provided functional evidence supportingthe role of VHA in Na uptake in FW fish gills (9 39) andapical localization of VHA in gill MRCs (44) However amajor drawback of this model has been the inability to identifythe channel responsible for Na uptake since no ENaC ho-mologs have been found in the available teleost fish genomesTherefore we hypothesized that other channels related toENaCs may perform this function
ENaC is a member of the amiloride-sensitive ENaCdegen-erin family of ion channels (23) In vertebrates the closestrelatives to ENaC are the acid-sensing ion channels (ASICs)with which they share 25 identity (24) Therefore weinvestigated the possibility that ASICs may be responsible forNa uptake in FW fishes ASICs are voltage-insensitive Na
channels gated by extracellular H (47) and are associatedwith the nervous system of mammals and fishes (16 35 40 4246) They have been implicated in mechanoreception andsensory transduction of taste and pain (for review see Ref 23)In mammals at least seven different ASIC subunits (ASIC1aASIC1b ASIC2a ASIC2b ASIC3 ASIC4 and ASIC5) areencoded by five ASIC genes while in zebrafish six ASICsubunits (ASIC11 ASIC12 ASIC13 ASIC2 ASIC41 andASIC42) are encoded by six different genes (13 35 40) NoASIC3 or ASIC5 has been demonstrated in zebrafish RecentlyChen and colleagues (5) described a group of potent ASICinhibitors the diarylamidines Diarylamidines are a non-amiloride-derived ASIC blockers class of compounds thatinhibit ASIC currents at very low concentrations but do nothave an effect on ENaC This finding can be exploited topharmacologically investigate a potential role for ASICs inNa uptake in FW fishes
In this study we used a variety of whole-animal cellular andmolecular approaches to investigate a potential role for ASICsin Na uptake in FW fishes We used FW rainbow trout as ourmodel organism since their natural habitat includes very-low-ionic-strength waters with Na concentrations and pH levelsbelow the theoretical limits of the NHERh metabolon modelWe identified two trout subunits of ASICs (ASIC1 and ASIC4)that are expressed in gill MRCs and demonstrated an apicalMRC localization for trout ASIC4 This is the first study toidentify a role for ASICs in Na uptake in FW fishes and isalso the first study to demonstrate a nonneuronal function forASICs in an organism
Address for reprint requests and other correspondence G G Goss Dept ofBiological Sciences Univ of Alberta Edmonton AB Canada T6G 2E9(e-mail greggossualbertaca)
Am J Physiol Cell Physiol 307 C255ndashC265 2014First published June 4 2014 doi101152ajpcell003982013
0363-614314 Copyright copy 2014 the American Physiological Societyhttpwwwajpcellorg C255
MATERIALS AND METHODS
Animal holding Rainbow trout embryos were obtained from theRaven Brood Trout Station (Caroline AB Canada) and grown to theappropriate size for experimentation (2 g for flux experiments and200ndash500 g as adults for other experiments) Fish were maintainedindoors in flow-through 450-liter fiberglass tanks supplied with aer-ated and dechlorinated Edmonton tap water (hardness as CaCO3 16mmoll alkalinity 120 mgl NaCl 05 mmoll pH 82 15degC) Fishwere fed ground dry commercial trout pellets (Purina trout chow)once daily and kept on a 1410-h light-dark photoperiod All animaluse was approved by the University of Alberta Animal Care Com-mittee under Protocol AUP00000072
Pharmacological inhibition of Na uptake For flux studies juve-nile trout were first acclimated to low-ionic-strength pH 60 water for4 days prior to Na flux experiments The duration of acclimation wasdeemed sufficient on the basis of a previous soft water exposure studyin rainbow trout (Salmo gairdneri) (43) Fish were transferred to60-liter plastic tubs supplied with aerated recirculating low-ionic-strength water (Na 300 30 moll Ca2 24 moll Cl 780 20 moll) and kept at a constant temperature of 15degC Afteracclimation two series of flux experiments were completed The firstexperiment investigated concentration-dependent effects of two ASIC-inhibiting diarylamidines 4=6-diamidino-2-phenylindole (DAPI 0001001 01 10 and 10 moll Sigma) and diminazene (00003 0003003 03 and 30 moll Sigma) on Na uptake in juvenile troutThis experiment established the 50 inhibitory concentrations (IC50Hill function) and optimal concentrations (80 inhibition) forDAPI and diminazene The second flux experiment compared theeffects of two well-known pharmacological Na uptake inhibitors[amiloride (500 moll) and phenamil (50 moll) Sigma] and thetwo ASIC inhibitors [DAPI (1 moll) and diminazene (3 moll)] onNa uptake in juvenile trout
For all flux experiments fish were transferred to individual dark-ened 60-ml flux chambers supplied with a constant flow of aeratedlow-ionic-strength low-pH water and acclimated to the chambers for24 h prior to experimentation Unidirectional Na influx was mea-sured using radiolabeled 22Na as described previously (12) Brieflyflow of water to chambers was turned off 01 Cil 22Na was addedto each chamber and allowed to mix for 5 min and either the DMSOcontrol (005 DMSO) or pharmacological agents dissolved inDMSO were added to the chambers After a 5-min equilibrationperiod 6-ml water samples were collected at time 0 and 90 and 180min as appropriate At the completion of the experiment fish wereterminally anesthetized (MS-222 1 gl) and weighed Water samples(3 ml) were analyzed for 22Na radioactivity using a gamma counter(Packard Cobra II Auto Gamma model 5010 Perkin ElmerWaltham MA) and total concentration of Na was measured usingatomic absorption spectrophotometry (model 3300 Perkin ElmerShelton CT) Unidirectional 22Na influx (molmiddotkg1middoth1) was cal-culated for each flux period according to the following equation JNain cpm V(SA t M) where cpm is the difference between theinitial and final radioactivities in the water (cpmml) V is the waterflux volume (ml) SA is the average specific activity (cpmmol Na)as measured at the beginning and end of the flux period t is the timeelapsed (h) and M is the mass of the fish (kg)
Tissue collection and preparation RNA isolation for expressionanalysis was performed on adult fish Briefly fish were euthanized asdescribed above a blood sample was withdrawn from the caudal archand the brain head kidney and trunk kidney were dissected out andimmediately freeze-clamped in liquid N2 for later processing For gilltissue the animal was first perfused with ice-cold heparinized (15mg) phosphate-buffered saline (PBS in mM 137 NaCl 27 KCl 43Na2HPO4 and 14 NaH2PO4 pH 78) and gill tissue or MRCs (asappropriate) were obtained according to the protocols described else-where (10) After perfusion gill arches were processed for MRCisolation freeze-clamped in liquid N2 for RNA isolation or placed in
fixative for immunohistochemistry or scanning electron microscopy(SEM) (see below)
MRC isolation and cellular imaging Adult rainbow trout (300ndash500 g) gills were perfused with PBS to remove blood according tooriginal protocols (10 33) Subsequently gill filaments were removedfrom the rakers cut into sections (2ndash5 mm 3ndash6 filaments) rinsed inPBS (in mM 137 NaCl 27 KCl 43 Na2HPO4 and 14 NaH2PO4pH 78) and subjected to three (20-min) incubations in 5 ml of 005trypsin-EDTA with shaking (200 rpm) at room temperature Thesubsequent cellular suspensions following each incubation werepassed through a 64-m nylon mesh filter into 10 ml of ice-cold fetalbovine serum and rinsed through with PBS to halt trypsin activity Thecells were then centrifuged (5 min 1500 g 4degC) and washed threetimes with 25 ml of PBS The final pellet was resuspended in 2 ml ofice-cold PBS and applied to a three-step Percoll density gradient (2ml 109 gml 2 ml 105 gml and 3 ml 103 gml) and centrifuged(45 min 2000 g 4degC 0-brake) After centrifugation the cell layer atthe 109-105 interface was removed washed three times in ice-coldPBS and resuspended in 800 l of ice-cold PBS
Isolated MRCs (500000) in PBS with an additional 2 l ofMgCl2 (1 M) and CaCl2 (1 M) to aid in cell attachment were placedon pretreated (1 M HCl acid-washed 01 poly-L-lysine-coated andrinsed with double-distilled H2O and 70 ethanol) 15-mm roundglass coverslips (catalog no CS-15R Warner Instruments) and al-lowed to attach for 2 h at 4degC After attachment coverslips wereplaced at room temperature and after careful removal of PBS thecells were briefly rinsed in Na-containing buffer (in mM 1425NaCl 50 CaCl2 10 MgCl2 40 KCl 15 HEPES and 25 NaHCO3pH 78) The cells were then incubated in 200 l of Na-containingbuffer which contained 2 l of 5 mM pH-sensitive BCECF-AM (50g in 16 l of DMSO and 20 pluronic acid) for 30 min at 18degCCoverslips were placed into a 70-l imaging chamber (model RC-20H Warner Instruments) for the perfusion experiments The cham-ber was fixed to an inverted fluorescence microscope (Nikon EclipseTE300) and the cells were subjected to differential interferencecontrast microscopy and fluorescent imaging To allow forBCECF-AM excitation at 495 and 440 nm the microscope was fittedwith a xenon arc lamp (Lambda DG-4 Sutter Instruments NovatoCA) Images were digitally obtained at 440 and 495 nm on a mono12-bit charge-couple device camera (Retiga EXi QImagingBurnaby BC Canada) every 17 s during perfusion experimentsRatios of fluorescence at 495 nm to fluorescence at 440 nm weredigitally compiled using Northern Eclipse software (MississaugaON Canada)
Cell perfusion Solutions were perfused across the attached MRCsin the holding chamber by gravity feed A six-input manifold (modelMp-6 Warner Instruments) attached to 60-ml syringe holder blockscontrolled by pinch valves (model VE-6 Warner Instruments) mon-itored manually with VC-6 valve controllers (Warner Instruments)was used to perfuse the solutions at a rate of 05 mlmin Cells weremonitored from original resting state for changes in intracellular pH(pHi) when exposed to Na-free and Na-containing solutions Start-ing in Na-free conditions (in mM 1425 N-methyl-D-glucamine-Cl25 C5H14NOmiddotHCO3
5 CaCl2 1 MgCl2 4 KCl and 15 HEPES pH78) followed by replacement with a Na-containing solution wemonitored the activity of the NHE in trout MRCs following a pHi
disturbance An identical transition between Na-free and Na-containing solutions was made but this time in the presence of 1 MDAPI or 100 M ethylisopropylamiloride (EIPA) EIPA was used inthis experiment since it potently inhibits NHE but has a low affinityfor Na channels (25) Perfusion solutions were bubbled with 03CO2 balanced with O2 throughout the experiments At the end of eachexperimental perfusion cells were subjected to a final high-K
solution calibration protocol (in mM 120 potassium gluconate 20KCl 2 MgCl2 and 20 HEPES) in which four solutions were adjustedto pH levels (840 780 720 and 660) by the addition of theionophore nigericin (5 M) to equilibrate pHi and extracellular pH
C256 INVOLVEMENT OF ASICs IN FISH GILL Na TRANSPORT
AJP-Cell Physiol bull doi101152ajpcell003982013 bull wwwajpcellorg
The ratios of fluorescence at 495 nm to fluorescence at 440 nm at eachstep of the pH calibration were used to form a regression equation foreach individual cell trace This equation was extrapolated to the rawratio data obtained over the entire time course yielding a calibratedpHi for each individual cell monitored during the perfusion experi-ment Data were collected and analyzed for each calibrated individualcell by comparing the rate of alkalinization (pHit) under thecontrol parameters with that in the presence of the drug to determinepercent inhibition
Preparation of total RNA Total RNA was extracted from frozentissues using TRIzol reagent (Invitrogen Carlsbad CA) according tothe manufacturerrsquos instructions RNA was treated with DNase I(Ambion Austin TX) according to the manufacturerrsquos protocol thenan on-column cleanup was carried out using an RNeasy Mini Kit(Qiagen Mississauga ON Canada) The quality of the extractedRNA was assessed by a bioanalyzer (model 2100 Agilent Technol-ogies Palo Alto CA) or by visualization on a formaldehyde gel RNAconcentration was measured with a spectrophotometer (ND-1000UV-Vis NanoDrop Technologies Rockland DE)
Molecular cloning and phylogenetic analysis Rapid amplificationof cDNA ends (RACE)-ready cDNA was synthesized from gill RNAusing a SMARTer RACE cDNA amplification kit (Clontech Moun-tain View CA) according to the manufacturerrsquos instructions cDNAsolutions were diluted 110 with tricine-EDTA buffer provided in thekit and stored at 20degC
Oligonucleotides for cloning of ASIC1 and ASIC4 genes weredesigned with PrimerQuest (Integrated DNA Technology) (Table 1)on the basis of conserved regions in available ASIC mRNA sequencesfrom zebrafish tilapia pufferfish stickleback and cod (NationalCenter for Biotechnology Information Ensembl) Sequences werealigned with GeneDoc (version 2602 httpwwwpscedubiomedgenedoc) and ClustalX (version 181) software DNA sequencesencoding the 5=- and 3=-termini of the ASIC1 gene and the 5=-terminus of the ASIC4 gene were obtained using the Advantage 2PCR kit (Clonetech) with RACE-ready cDNA templates generatedfrom gill RNA The ASIC4 3=-terminus was obtained using degener-ate primer (Table 1) Resulting PCR products were excised from thegel and purified with a QIAquick gel extraction kit (Qiagen) Subse-quently amplicons of interest were sequenced directly or ligated intopJET12 vector using a CloneJET kit (Thermo Scientific) for verifi-cation Recombinant plasmids were transformed into competent cells
(One Shot TOP10 chemically competent Escherichia coli Invitro-gen) Colonies were screened for plasmids containing inserts of thecorrect size by agarose gel electrophoresis and sequenced withpJET12-specific primers provided in the CloneJET kit
The amino acid sequences derived from our cloned trout ASIC4and ASIC1 genes (accession numbers KF964646 and KF964645respectively) together with the previously annotated amino acidsequences of ASIC1 ASIC2 ASIC3 and ASIC4 from various fishspecies and other taxa were aligned and analyzed using Seaviewsoftware (version 4 httppbiluniv-lyon1frsoftwareseaview) Arooted phylogenetic tree showing evolutionary relationships betweenthe different ASIC proteins was generated using the maximum-likelihood method and LG model and 700 bootstrap resamplings
RT-PCR Tissue-specific cDNA templates utilized for RT-PCRwere synthesized from 3 g of RNA from gill MRCs kidney brainor blood using SuperScript III reverse transcriptase (Invitrogen) ac-cording to protocols provided by the manufacturer ASIC4 and ASIC1amplicons were obtained using primers designed with PrimerQuest(Table 1) PCR conditions were as follows 98degC for 1 min of initialdenaturation followed by 35 cycles of denaturation at 98degC for 10 sannealing at 62degC for 30 s and elongation at 72degC for 40 s with thefinal elongation at 72degC for 10 min PCR products were visualized by1 agarose gel electrophoresis followed by ethidium bromide stain-ing
Immunoprecipitation of ASIC4 and Western blot analysis Ananti-zebrafish ASIC4b (zASIC42) polyclonal antibody against twopeptides corresponding to regions near the NH2 terminus (aa 146ndash160 PKSRKGHRPSELQYP) and the COOH terminus (aa 519ndash533CFEEVKVKAANDVAQP) of zASIC42 protein (accession noQ708S31) was custom-made by 21st Century Biochemicals (Marl-boro MA) The polyserum was affinity-purified against each of thesepeptides independently and the NH2-terminal peptide-purified anti-body was used exclusively in this study Immunoprecipitation wasperformed according to the technique described by Goss and col-leagues (11) Briefly trout gills were perfused with heparinizedice-cold PBS (Ca2-free) and the second and third gill arches weredissected out and washed three times in ice-cold PBS Gill rakers wereremoved and the remaining gill arches were cut into 200- to 300-mgsections Each section was lysed for 30 min in 2ndash3 ml of lysis buffer(100 mM NaCl 4 mM KCl 50 mM HEPES 5 mM EDTA pH 74and 1 Triton-X) containing protease inhibitors (cOmplete MiniEDTA-free protease inhibitor tablets Roche Diagnostics MannheimGermany) After centrifugation to remove the debris (5000 g 5 min4degC) the lysates were incubated with 2ndash4 l of anti-zASIC42antibody at 4degC overnight on an end-over-end rotator On the nextday 60 l of protein A-Sepharose CL4b beads (Sigma Chemical StLouis MO) preswelled in immunoprecipitation (IP) buffer (in mM100 NaCl 4 KCl 50 HEPES and 5 EDTA pH 74) and preblockedwith 1 bovine serum albumin (fraction V Sigma) were added to thelysates which were incubated at 4degC for 4ndash6 h on the rotator Afterthe incubation the supernatant was removed and the beads werewashed three times with 1 ml of IP buffer After the washes the beadswere incubated in Laemmli buffer for 15 min at 65degC and centrifugedand the supernatant was retained for Western blot analysis as de-scribed previously (45) Briefly the samples were separated on a 75polyacrylamide minigel and transferred to a nitrocellulose (NC) mem-brane using a wet transfer system (Bio-Rad Laboratories HerculesCA) The NC membrane was blocked in 5 bovine serum albumin inTris-buffered saline with 02 Triton X-100 (TBST) for 30 min andincubated with rabbit anti-zASIC42 antibody (11000 dilution) on arocker at 4degC overnight The membrane was washed four times for 15min each with TBST blocked again with 5 bovine serum albuminfor 15 min and then incubated with a secondary horseradish peroxi-dase-conjugated goat anti-rabbit antibody (Santa Cruz BiotechnologyDallas TX) at 150000 dilution at room temperature for 1 h The NCmembrane was washed four times in TBST and the immunoreactivebands were visualized using a SuperSignal West Pico chemilumines-
Table 1 Primer sets for cloning and tissue distributionof ASICs
Gene Primer Sequence
CloningASIC1
5= 5=-TCACCCAGCAACCCTGCGAACTCGT-3=3= 5=-ACACCCTGCAACATGACGCGCT-3=
ASIC45= 5=-GATGTCCCGTCCGGTTGAAGATGTC-3=Forward 5=-CCCAGTAACATCAAGTGTGTCG-3=Reverse 5=-YYARCANGCRAARTCYTC-3=
RT-PCRASIC1
Forward 5=-AAGTCCACTCCCATAGA-3=Reverse 5=-CAGCCAGGTTATTCCTT-3=
ASIC4Forward 5=-CTTTCGTTTCTCTGCTCTCACC-3=Reverse 5=-CAAGACCAGGAAGTTGTCTCTG-3=
GAPDHForward 5=-AAGGGTGAGGTGAGCATGGA-3=Reverse 5=-GCTTTACCCCATGGGATCTCAT-3=
ASIC acid-sensing ion channel 5= 5=-rapid amplification of cDNA ends(RACE) primer 3= 3=RACE primer Y C T R A G N A G T C
C257INVOLVEMENT OF ASICs IN FISH GILL Na TRANSPORT
AJP-Cell Physiol bull doi101152ajpcell003982013 bull wwwajpcellorg
cent substrate kit (Thermo Scientific) following the manufacturerrsquosinstructions
SEM and immunohistochemistry For examination of the surface ofthe gill by SEM filaments were fixed in 3 glutaraldehyde and 2paraformaldehyde in 015 mM sodium cacodylate buffer (pH 74 290mosM) for 1 h at 4degC dehydrated in a graded series of ethanol andcritical point dried Samples were then mounted on an SEM stubsputter-coated with gold and examined with a scanning electronmicroscope (model JSM-6301FXV Jeol)
For immunohistochemistry gills were excised from adult rain-bow trout fixed in 4 paraformaldehyde in PBS (pH 74) over-night at 4degC dehydrated in a graded series of ethanol andembedded into paraffin blocks Serial sections (5 m) were cut andrehydrated in a decreasing ethanol series followed by double-distilled H2O Rehydrated sections were incubated in 10 mMcitrate buffer at 70degC for 1 h for epitope retrieval After incubationslides were washed in PBS and then blocked with 6 milk powderin a humidified chamber for 1 h Sections were then incubatedovernight at 4degC simultaneously with the anti-zASIC42 antibody(1400 dilution) and anti-Na-K-ATPase (NKA) monoclonalantibody (1400 dilution anti-chicken 5-subunit DevelopmentalStudies Hybridoma Bank University of Iowa) followed by sec-ondary FITC-conjugated anti-mouse antibody (1500 dilution In-vitrogen Oregon) and tetramethylrhodamine-conjugated anti-rab-bit antibody (1500 dilution Invitrogen Oregon) Slides incubatedwith preimmune serum from the anti-zASIC42 antibody produc-tion in place of primary antibody were used as a negative controlSlides were viewed with a laser scanning confocal microscope(model LSM 710 Zeiss Germany) at the Cross Cancer InstituteCell Imaging Facility (Edmonton AB Canada) Images wereprocessed with LSM Image Browser (version 420121 CarlZeiss) Surface rendering and three-dimensional reconstruction ofz-stack images were performed using Imaris software (version622 Bitplane Zurich Switzerland)
Statistical analysis Na flux data were subjected to statisticalanalysis and are reported as means SE All pharmacologicalexperiment data sets were tested for homogeneity of variance andcompared using one-way ANOVA (SigmaPlot version 11 SystatChicago IL) If significant differences (P 005) were found a posthoc multiple-comparisons Tukeyrsquos test was applied to determine thesedifferences A paired t-test (P 005) was used in the pHi imagingexperiments to compare the relative inhibition of pHit in isolatedcells under control conditions and after addition of each pharmaco-logical inhibitor
RESULTS
Pharmacological inhibition Exposure of juvenile rainbowtrout to increasing concentrations of DAPI resulted in a con-centration-dependent decrease in Na uptake with 90inhibition at 1 moll (Fig 1A) The DAPI concentration atwhich there was a 50 inhibition in Na uptake (IC50) wascalculated to be 012 moll Exposure to diminazene alsoresulted in a concentration-dependent decrease in Na uptakein juvenile trout with a maximum reduction of Na uptakefrom 303 43 (control) to 51 19 molmiddotkg1middoth1 (79)after exposure to 30 moll diminazene (Fig 1B) The IC50 fordiminazene was calculated to be 096 moll
Na uptake rates in juvenile rainbow trout were significantlylower in the first 90-min flux time period after exposure to 500M amiloride (17 185 molmiddotkg1middoth1) 1 M DAPI (29 189 molmiddotkg1middoth1) and 3 M diminazene (41 334molmiddotkg1middoth1) than in controls (539 1709 molmiddotkg1middoth1 Fig 2) while 50 M phenamil (299 1079 molmiddotkg1middoth1) did not significantly reduce Na uptake Howeverfor the second consecutive flux period between 90 and 180min of exposure to the pharmacological agents the inhibitionwas less pronounced but still significant for amiloride (104 334 molmiddotkg1middoth1) and DAPI (141 627 molmiddotkg1middoth1)while diminazene (297 415 molmiddotkg1middoth1) was no longersignificantly different from control (393 441 molmiddotkg1middoth1 Fig 2)
MRC pHi imaging Results presented from the whole-animalflux experiments showed that administration of DAPI at vari-ous concentrations significantly inhibited Na uptake (Fig1A) To demonstrate that the inhibition of Na uptake attrib-uted to DAPI was in fact due to the blocking of ASIC-type Na
channels rather than affecting NHE activity we conductedexperiments using pHi imaging NHE activity was measuredby examining the rate of pHi alkalization (pHit) uponexposure of MRCs to a Na-containing medium (145 mM) inthe absence or presence of DAPI (1 M) or EIPA (100 M)
A representative trace (Fig 3A) shows a Na-dependentalkalization following Na application (1st rise) while addi-tion of EIPA significantly decreased the rate of alkalization(2nd rise) The combined results (n 19) show a control of
0
50
100
150
200
250
300
350
400
450
0 0001 001 01 1 10
JNa I
N(μ
mol
kg-1
h-1 )
DAPI concentration (μmol l-1)
0
50
100
150
200
250
300
350
400
0 00003 0003 003 03 3
JNa I
N(μ
mol
k g-1
h-1 )
Diminazene concentration (μmol l-1)
A B
Fig 1 Dose-dependent decreases in Na uptake rates (JNain) in juvenile trout after exposure to increasing concentrations of 4=6-diamidino-2-phenylindole(DAPI) and diminazene in low-Na (30 M) and low-pH (pH 60) water Values are means SE (n 6) P 005 vs control
C258 INVOLVEMENT OF ASICs IN FISH GILL Na TRANSPORT
AJP-Cell Physiol bull doi101152ajpcell003982013 bull wwwajpcellorg
0642 0040 pHimin while the alkalization rate wasreduced by 62 in the presence of EIPA (0 240 0036pHimin P 0001 Fig 3B) For DAPI Na-dependentalkalization following Na application was similar in the
presence or absence of DAPI (Fig 3D) No differences inpHit were noted between control (0542 0042 pHimin) and DAPI-treated (0578 0047 pHimin) cells(P 0149 n 22 Fig 3D)
0
100
200
300
400
500
600
700
800
Cont Amil DAPI Dim Phen Cont Amil DAPI Dim Phen
JNa I
N(μ
mol
kg-1
h-1 )
Treatment
T0-90min T90-180min
Fig 2 Effects of pharmacological agents [control (Cont DMSO) amiloride (Amil 500 moll) DAPI (1 moll) diminazene (Dim 3 moll) and phenamil(Phen 50 moll)] during consecutive 90-min measurements of Na uptake in juvenile trout in low-Na (30 M) and low-pH (pH 60) water Values are means SE(n 6) P 005 vs control
65
7
75
8
85
9
0 100 200 300 400 500 600
pHi
Time (sec)
Na FreeNa Free Na
EIPA
Na
0
01
02
03
04
05
06
07
08
Na+ Na+EIPA
ΔpH
iΔM
in
Cell Alkalinization Treatments
65
7
75
8
85
9
0 100 200 300 400 500 600
pHi
Time (sec)
Na Free Na
DAPI
Na Na Free
0
01
02
03
04
05
06
07
08
Na+ Na+DAPI
ΔpH
iΔM
in
Cell Alkalinization Treatments
A
DC
B
Fig 3 Effect of ethylisopropylamiloride (EIPA) and DAPI on Na-induced alkalinization of peptide nucleic acid-positive (PNA) cells following Na-freeexposure at resting intracellular pH (pHi) A representative trace demonstrating inhibition of cell alkalinization by EIPA (100 M) B quantitative analysis ofcell alkalinization in the absence and presence of EIPA (62 inhibition) Values are means SE (n 19 cells) P 0001 (by paired t-test) C representativetrace demonstrating lack of effect on cell alkalinization in the presence of DAPI (1 M) D quantitative analysis of cell alkalinization in the absence and presenceof DAPI Values are means SE (n 22) P 005 (by paired t-test)
C259INVOLVEMENT OF ASICs IN FISH GILL Na TRANSPORT
AJP-Cell Physiol bull doi101152ajpcell003982013 bull wwwajpcellorg
Phylogenetic analysis and tissue distribution of ASIC1 andASIC4 Two distinct DNA fragments whose derived aminoacid sequences exhibit similarity to the ASIC family werecloned from the gill of rainbow trout To determine theiridentity we conducted a phylogenetic analysis by comparisonwith known ASIC homologs from a variety of species Thegenerated phylogenetic tree (Fig 4) showed that the twocloned rainbow trout ASIC genes (tASICs) were members ofthe ASIC1 and ASIC4 subunits respectively (Fig 4A) ThetASIC1 (partial clone) grouped most closely with ASIC1 fromsalmon (Salmo salar) while tASIC4 grouped with other iden-
tified ASIC4 family members from other fish and mammalspecies
Furthermore we conducted a tissue distribution analysis oftASIC1 and tASIC4 by RT-PCR We observed mRNA expres-sion of tASIC1 in all tested tissues gill MRCs head and trunkkidney brain and blood tASIC4 had a slightly differentdistribution it was present in the perfused gill MRCs brainand trunk kidney but it was absent in head kidney and bloodtissues
Immunoprecipitation and immunostaining with anti-zASIC42To determine if ASIC4 was present in the fish gill and to validate our
Fig 4 Phylogenetic analysis of amino acid sequences of cloned rainbow trout acid-sensing ion channels ASIC1 and ASIC4 and ASICs from various fish speciesPhylogenetic tree was constructed using the maximum-likelihood method and numbers indicate bootstrap values for 700 resamplings GenBank or Ensemblaccession numbers are as follows human ASIC2 NP_0010852 Western clawed frog ASIC2 XP_0029330862 Japanese pufferfish ASIC2 XP_0039648711zebrafish ASIC2 NP_9999531 three-spined stickleback ASIC2 ENSGACG00000009558 Atlantic cod ASIC2 ENSGMOT00000019145 spotted greenpufferfish ASIC2 ENSTNIG00000014943 Southern platyfish ASIC2 ENSXMAG00000008283 oyster toadfish ASIC2 AAP456331 coelacanth ASIC2ENSLACG00000007191 rat ASIC2 NP_0370242 Southern platyfish ASIC1c ENSXMAG00000018841 African clawed frog ASIC11 AEC486651 Africanclawed frog ASIC12 AEC486671 brown rat ASIC1 NP_0770681 human ASIC1 NP_0644232 three-spined stickleback ASIC1a ENSGACG00000003075Japanese pufferfish ASIC1 XP_0039635201 Southern platyfish ASIC1a XP_0058018011 Atlantic cod ASIC1a ENSGMOG00000009659 zebrafishASIC12 NP_9999551 rainbow trout ASIC1 KF964646 Atlantic salmon ASIC1 NP_0011334561 oyster toadfish ASIC1 Q7T1N41 elephant shark ASIC1AEC486681 three-spined stickleback AIC1c ENSGACG00000017974 Atlantic cod ASIC1c ENSGMOG00000003105 zebrafish ASIC13 NP_9999541river lamprey ASIC1 AAY289831 sea lamprey ASIC1b ENSPMAG00000003813 dogfish shark ASIC1 AAY289851 zebrafish ASIC11 NP_9999561human ASIC4 NP_8782672 rat ASIC4 NP_0715702 African clawed frog ASIC4 AEC486661 Japanese pufferfish ASIC4 XP_0039621271 spotted greenpufferfish ASIC4b ENSTNIG00000010994 Nile tilapia ASIC4 XP_0034454461 Southern platyfish ASIC4a ENSXMAG00000003624 three-spinedstickleback ASIC4a ENSGACG00000007052 Atlantic cod ASIC4a ENSGMOG00000012740 rainbow trout ASIC4 KF964645 zebrafish ASIC41NP_9999521 zebrafish ASIC42 NP_9999511
C260 INVOLVEMENT OF ASICs IN FISH GILL Na TRANSPORT
AJP-Cell Physiol bull doi101152ajpcell003982013 bull wwwajpcellorg
antibody for later immunocytochemistry we performed an im-munoprecipitation using an antibody raised against zASIC42on the basis of sequence similarly of 60 between zASIC42(aa 146 ndash160 PKSRKGHRPSELQYPP) and the clonedtASIC4 (aa 142ndash156 PKNREGHKPTDLDYPA) Using West-ern blotting we analyzed the precipitates for the presence ofASIC4 (see Fig 6A) and found a single band of 65 kDawhich approximates the predicted size for tASIC4 (604 kDa)The strong band at 55 kDa corresponds to the IgG heavychain
To determine the location of ASIC4 protein within therainbow trout gill tissue fixed sections were double-labeledwith anti-zASIC42 antibody (see Fig 6B) and anti-NKA andthen counterstained with the nuclear stain DAPI Immunore-activity was observed in cells located in the lamellae andinterlamellar region (see Fig 6B arrowhead) Moreover themajority of cells positive for zASIC42 also colocalized withNKA protein (Fig 6 C and D arrowhead) a known marker ofMRCs suggesting that in gill epithelium ASIC4 protein ismainly expressed in MRCs SEM analysis of the gill filamentdemonstrated that MRCs are clearly distinguished on the sur-face of the epithelium and have a broad apical convex openingto the environment (see Fig 7A arrowheads) The generatedthree-dimensional reconstructions of the staining immunoreac-tivity pattern for ASIC4 and NKA proteins revealed the pres-ence of ASIC4 and NKA in the same cells but in distinctregions of the cells NKA was present on the basolateralmembrane (arrows green staining) while ASIC4 was localizedto the broad apical surface of the cell (arrowheads red stain-ing) Figure 7B shows a generated image with a top view whileFig 7C is the same cell viewed from the side
DISCUSSION
The existence of an apical Na channel electrochemicallycoupled to a VHA mediating Na uptake from dilute FW hasbeen postulated on theoretical thermodynamic grounds for anumber of decades (1 36 34) However an inability todetermine the molecular identity of the Na channel has beena major drawback in the general acceptance of the modelUsing a combination of pharmacological blockade isotopicfluxes immunocytochemistry and molecular biology we pro-pose that an ASIC family member is potentially the long-sought-after apical Na channel responsible for Na uptake byrainbow trout acclimatized to water with low Na and low pHOur study provides the first evidence for a nonneuronal role forASICs in any vertebrate whereby in gills from FW fishesASICs are associated with the MRCs and tASIC4 localizes tothe apical membrane of NKA positive cells
ASICs are a broad family of Na-specific ion channels thathave been extensively studied in mammalian models (forreview see Ref 23) with only a few studies performed in anyfish species (35 42 46) ASICs have primarily been associatedwith neuronal tissues with studies focusing on a neuronalfunction for ASICs (23) In this study ASIC1 and ASIC4 werecloned in rainbow trout using PCR and they were expressed ina variety of tissues both neuronal and nonneuronal in origin(Fig 5) Importantly tASIC1 gene was present in all fiveinvestigated tissues including whole blood It appears that thetASIC4 gene has a more defined tissue distribution thantASIC1 with expression in the ionoregulatory tissues (trunk
kidney and perfused gill) and the brain Unlike tASIC1 therewas no apparent expression of tASIC4 in the head kidney orthe blood Therefore tASIC4 expression in MRCs and per-fused gill could not be attributed to blood contamination
Both tASIC1 and tASIC4 genes were expressed in the gillepithelium and more specifically were present in MRCs ofrainbow trout gills It is believed that similar to ENaCsfunctional ASICs are trimers (20) although their configurationhas not been resolved It has been demonstrated that with afew exceptions ASICs function as homomers (forming chan-nels of identical subunits) or heteromers (forming channelswith other ASIC subunits) (4 23) ASICs have been charac-terized by using oocyte expression in zebrafish (4 35) anddogfish shark (ASIC1b only) (42) It has been found thatcoexpression of zASIC41 with zASIC13 in Xenopus oocytesincreased the current amplitude and abundance (15-fold) ofthe channel at the cell surface indicating that these two ASICsubunits form a functional channel (4) Moreover thisASIC4113 heteromeric channel was more efficiently traf-ficked to the plasma membrane and had increased affinity forH Additionally zASIC41 when heterologously expressedin Xenopus oocytes has been shown to be also gated open bydecreases in extracellular Ca2 The signal responsible forgating of zASIC42 was unable to be determined in that study(4) It is therefore possible that tASIC1 and tASIC4 form aheteromeric channel in MRCs but this cannot be determined atthis time and would require confirmation by immunohisto-chemistry with an anti-ASIC1 antibody that is immunoreactivefor rainbow trout It is also possible that other ASIC subunitsandor regulatory elements may be involved in the function ofASICs in the FW trout gill A full characterization of tASIC4by oocyte expression and electrophysiology is required tounderstand the mechanisms of gating in low-Na low-Ca2
watersIn our study ASIC4 protein was colocalized with NKA and
was present on the apical surface of cells suggesting that it isfound primarily in MRCs (Fig 6 D and E) Previous studies onASICs in fishes have suggested that they are primarily associ-ated with neuronal tissue and sensory cells (27 35 42 46) Inadult and developing zebrafish morphological studies usingconfocal immunofluorescence techniques demonstrated thatgill MRCs come into direct contact with nerve fibers in thebasolateral region (21 22) However in our study ASIC4protein was clearly localized to the apical region of the MRCsas demonstrated by three-dimensional reconstruction of con-
Head k
idney
ASIC4
ASIC1
GAPDH
Gill MRCs
Brain
Trunk k
idney
Blood
No tem
plate
Fig 5 Tissue distribution analysis of ASIC4 and ASIC1 genes in adultrainbow trout as determined by PCR GAPDH was used as a reference geneMRCs mitochondrion-rich cells
C261INVOLVEMENT OF ASICs IN FISH GILL Na TRANSPORT
AJP-Cell Physiol bull doi101152ajpcell003982013 bull wwwajpcellorg
focal images ASIC4 immunoreactivity did not overlap withthe standard basolateral immunoreactive pattern associatedwith NKA (Fig 7 B and C) Therefore we conclude thatneuronally derived tissues cannot account for the expression ofASIC4 protein in MRCs We also corroborate the ASIC4staining pattern with the MRC external morphology whichshows that in FW rainbow trout raised in Edmonton tap water(500 M Na) the MRCs have wide apical openings (Fig7A) This apical localization of ASIC4 protein in the cells of atransporting epithelium is the first evidence for a nonneuronalfunction for any ASIC member and suggests that ASIC4 as aNa channel may be responsible for apical uptake of Na inFW fish gill
Pharmacological profiling has long been used to differentiatethe mechanisms of ion transport across epithelial tissues For Na
transport classical analysis has usually involved amiloride and itssubstituted derivatives (eg EIPA mercaptoisobutyric acid andphenamil) as a means to differentiate between the involvementof NHE isoforms and ENaC in Na transport (25) We inves-tigated two newly described ASIC-specific inhibitors dimina-zene and DAPI (5) for their potency in inhibiting unidirec-tional Na uptake in fish Diminazene and DAPI exhibited adose-dependent inhibitory relationship and both were veryeffective at low concentrations (Fig 1) Na uptake in rainbow
trout acclimatized to low-ionic-strength acidic water wasclearly inhibited by these diarylamidines at concentrationssimilar to those reported in a previous study of cultured rathippocampal neurons (5) However in our study DAPI provedto be more effective than diminazene in contrast to observa-tions in rat neurons The inhibition of 22Na uptake by DAPIand diminazene was also compared with other common Na
transport inhibitors amiloride and phenamil Amiloride wasobserved to be an effective inhibitor of Na uptake in rainbowtrout but only at relatively high concentrations (500 M)which is consistent with previous studies (2 37) Recentlyamiloride was also demonstrated to completely block ASIC-mediated currents (5 35 42) however it is also known toblock many other Na transporters including NHE and ENaC(25) Amiloride therefore cannot be used as a distinguishingpharmacological agent
While DAPI diminazene and amiloride almost completelyblocked Na transport in the present study phenamil had nosignificant effect on Na flux in rainbow trout This suggeststhat phenamil is a poor inhibitor of Na uptake in troutexposed to low-ionic-strength water (30 moll Na) Paststudies of the effects of phenamil on Na uptake in a variety ofFW fishes have reported inconsistent results For example inneon tetras acclimatized to low Na (50 moll) phenamil had
Fig 6 Immunoreactivity of anti-zebrafish ASIC42 (zASIC42) antibody in the gill of rainbow trout A immunoprecipitation Western blot from whole gillhomogenates with zASIC42 showing a distinct band of 65 kDa Lane 1 (L1) contains 300 mg of gill tissue and lane 2 (L2) contains 200 mg of gill tissueBndashE confocal images of gill sections labeled for ASIC4 and Na-K-ATPase (NKA) proteins B and C double-labeling with DAPI and anti-zASIC42 (B) andDAPI and anti-NKA (C) D merged image of B and C E higher-magnification image of a MRC showing that ASIC4 and NKA are present in distinct regionswithin the MRC F control micrograph with DAPI and no primary antibody Arrowheads (BndashD) indicate positive staining
C262 INVOLVEMENT OF ASICs IN FISH GILL Na TRANSPORT
AJP-Cell Physiol bull doi101152ajpcell003982013 bull wwwajpcellorg
no effect on Na transport (38) whereas it caused 70reduction in Na influx in Cyprinidon variegatus hubbsi (100moll Na) and 60 reduction in goldfish (300 moll Na)(2 38) These conflicting results from various phenamil studiessuggest that sensitivity to phenamil might be species- andorenvironment-specific Collectively the pharmacological anal-ysis suggests that the Na uptake in FW rainbow trout is mostlikely not mediated by ENaC given the lack of inhibition byphenamil but may be attributed to ASIC since Na uptakewas almost completely blocked by DAPI and diminazene bothof which inhibit ASICs but not ENaC
In our Na flux study with various pharmacological agentswe made an interesting observation that inhibition of 22Na
uptake was less pronounced for all tested inhibitors during thesecond consecutive measurement (90ndash180 min) (Fig 2) Toour knowledge this phenomenon where potency of the testeddrug diminishes over time has not been reported in anyprevious study on fish This observation may be explained bya decreased stabilityefficacy of the tested pharmacologicalagents in aqueous media metabolism of the drug by theanimal or activation of compensatory Na uptake mechanismsnot sensitive to amiloride DAPI or diminazene We believethat the decrease of the potency of amiloride DAPI anddiminazene over time demonstrates the importance of properexperimental design and choice of period of study when ionfluxes are examined using pharmacological inhibitors
While the two novel ASIC inhibitors have been tested fortheir potency in inhibiting ASICs and ENaC in neurons (5)they have not been tested for their potential effects on otherNa transporters implicated in Na uptake in fishes mostnotably the NHEs To verify that the Na uptake inhibitioncaused by DAPI and diminazene in rainbow trout was due toinhibition of ASICs rather than NHEs we tested the effect ofDAPI on MRCs isolated from the rainbow trout gill epithe-lium Our laboratory previously showed that rainbow troutpossess two populations of MRCs peanut lectin agglutinin(PNA) positive (PNA) and PNA negative (PNA) MRCs(10) It has been demonstrated that NHE3 and NHE2 areexpressed in PNA cells isolated from branchial epithelium of
FW rainbow trout (19) Therefore we investigated the effect ofDAPI on PNA cells and compared it with the effect of EIPAa potent and highly selective inhibitor of NHE (25) Earlierstudies in our laboratory using cell imaging determined thatPNA cells undergo NHE-dependent alkalinization (32 33)Here we demonstrate that while EIPA was effective at inhib-iting NHE function in PNA cells (Fig 3A) there was noeffect of high concentrations of DAPI on Na-dependent pHi
elevation demonstrating that NHE was not inhibited by DAPI(Fig 3B) Therefore the measured inhibition of Na uptake byDAPI that we see in whole animals (Fig 1) cannot be attrib-uted to inhibition of NHE
On the basis of the mRNA expression of tASIC1 andtASIC4 genes in the gill epithelium and isolated MRCs local-ization of ASIC4 protein to the apical region of the MRCs and
Fig 7 Three-dimensional views of freshwater(FW) rainbow trout MRCs A scanning elec-tron micrograph of FW rainbow trout gillepithelium showing broad apical openings ofMRCs (arrowheads) and mucous cells () Band C 3-dimensional reconstruction of z-stack images showing apical localization ofASIC4 protein (red arrowhead) and basolat-eral localization of NKA (green arrow)B top view C side view
PNA- MRC
Na+
ASIC
H+
VHA
NKA
Na+
K+
NBC
Na+ HCO3-
PNA+ MRC
NH3
Na+
H+
NHE Rhcg2
NKA
Na+
K+
Fig 8 Model for Na uptake in PNA and PNA MRCs in gill epithelium ofrainbow trout showing placement of proposed ASIC-mediated Na uptakemechanisms Rhcg2 rhesus glycoprotein NHE NaH exchanger VHAvacuolar H-ATPase NBC Na-HCO3
cotransporter
C263INVOLVEMENT OF ASICs IN FISH GILL Na TRANSPORT
AJP-Cell Physiol bull doi101152ajpcell003982013 bull wwwajpcellorg
pharmacological blockade of Na influx by ASIC inhibitors inadult rainbow trout we propose a revised model for transepi-thelial Na uptake in FW rainbow trout gills (Fig 8) wherebyone apical mode of Na entry is via an ASIC4-mediatedmechanism
A number of recent studies on Na uptake in FW fishes havedemonstrated the presence of the NHERh metabolon in thegills of various species (17 30 31) As mentioned previouslywe specifically targeted the rainbow trout as a model for thisstudy as it is native to oligotrophic Na-poor mountainstreams with low buffering capacity This type of environmentpresents key thermodynamic challenges to the NHERhmetabolon as Na restrictions cannot be alleviated by thepresence of an Rh protein (6) Moreover searches of rainbowtrout genomes and transcriptomes have failed to identify anyENaC subunits ( 13 or ) despite the fact that they are knownto actively take up Na from water with 50 M Na (49)Furthermore other research studies have suggested and dem-onstrated an apical localization of the VHA in the gills ofrainbow trout (44 48) and bafilomycin a VHA-specific in-hibitor has been demonstrated to reduce Na uptake in ze-brafish (2) tilapia and carp (9) Previous studies have pro-posed VHA to be a part of the NHERh metabolon (41 50 51)however the placement of an apically oriented VHA is coun-terintuitive to the function of an NHE as it would exacerbatethe noted thermodynamic constraints Placement of a VHA inconcert with an apical ASIC is more plausible since it wouldnot only facilitate Na uptake from very low ionic strengthwater but could also provide a mechanism for gating ofASIC-mediated Na conductance It remains to be determinedif ASIC and VHA proteins colocalize to the same type ofMRCs and this warrants further investigation
Perspectives In summary we propose that ASCI4 is in-volved in mediating Na uptake from FW in rainbow troutacclimatized to low ionic strength low pH media This is thefirst demonstration of a nonneuronal function for any ASICfamily member and simultaneously provides an attractive so-lution to the debate surrounding the thermodynamic limitationsof the other proposed Na transport mechanisms and resolvesthe conundrum of the role of an apical VHA in Na transportThe possibility that other fish models may use similar mecha-nisms under different environmental conditions where modelssuch as the NHERh metabolon cannot be expected to functionremains to be explored
ACKNOWLEDGMENTS
We thank the staff in the Biosciences Aquatic Facilities the MolecularBiology Service Unit and the Cross Cancer Institute Cell Imaging Facility atthe University of Alberta for outstanding help throughout the study We alsothank Alex Clifford for advice and help with phylogenetic analysis and JamesEde for help with confocal microscopy
GRANTS
This study was supported by a Natural Sciences and Engineering ResearchCouncil of Canada Discovery Grant to G G Goss and an Alberta IngenuityStudent Scholarship to A K Dymowska
DISCLOSURES
No conflicts of interest financial or otherwise are declared by the authors
AUTHOR CONTRIBUTIONS
AKD AGS and GGG are responsible for conception and design ofthe research AKD AGS SDB and DC performed the experiments
AKD AGS SDB DC and GGG analyzed the data AKD AGSSDB DC and GGG interpreted the results of the experiments AKDAGS SDB and GGG prepared the figures AKD AGS SDB DCand GGG drafted the manuscript AKD AGS SDB DC and GGGedited and revised the manuscript AKD AGS SDB and GGGapproved the final version of the manuscript
REFERENCES
1 Avella M Bornancin M A new analysis of ammonia and sodiumtransport through the gills of the freshwater rainbow trout (Salmo gaird-neri) J Exp Biol 142 155ndash175 1989
2 Boisen AM Amstrup J Novak I Grosell M Sodium and chloridetransport in soft water and hard water acclimatized zebrafish (Daniorerio) Biochim Biophys Acta 1618 207ndash218 2003
3 Brix KV Grosell M Comparative characterization of Na transport inCyprinodon variegatus variegatus and Cyprinodon variegatus hubbsi amodel species complex for studying teleost invasion of freshwater J ExpBiol 215 1199ndash1209 2012
4 Chen X Polleichtner G Kadurin I Grunder S Zebrafish acid-sensingion channel (ASIC) 4 Characterization of homo- and heteromeric chan-nels and identification of regions important for activation by H J BiolChem 282 30406ndash30413 2007
5 Chen X Qiu L Li M Durrnagel S Orser BA Xiong ZG MacDonaldJF Diarylamidines high potency inhibitors of acid-sensing ion channelsNeuropharmacology 58 1045ndash1053 2010
6 Dymowska AK Hwang PP Goss GG Structure and function of iono-cytes in the freshwater fish gill Respir Physiol Neurobiol 184 282ndash2922012
7 Edwards SL Tse CM Toop T Immunolocalization of NHE3-likeimmunoreactivity in the gills of the rainbow trout (Oncorhynchus mykiss)and the blue-throated wrasse (Pseudolabrus tetrious) J Anat 195 465ndash469 1999
8 Evans DH Piermarini PM Choe KP The multifunctional gill domi-nant site of gas exchange osmoregulation acid-base regulation andexcretion of nitrogenous waste Physiol Rev 85 97ndash177 2005
9 Fenwick JC Bonga SE Flik G In vivo bafilomycin-sensitive Na
uptake in young freshwater fish J Exp Biol 202 3659ndash3666 199910 Galvez F Reid SD Hawkings G Goss GG Isolation and characteriza-
tion of mitochondria-rich cell types from the gill of freshwater rainbowtrout Am J Physiol Regul Integr Comp Physiol 282 R658ndashR668 2002
11 Goss G Orlowski J Grinstein S Coimmunoprecipitation of a 24-kDaprotein with NHE1 the ubiquitous isoform of the NaH exchanger AmJ Physiol Cell Physiol 270 C1493ndashC1502 1996
12 Goss GG Wood CJ Na and Cl uptake kinetics diffusive effluxes andacidic equivalent fluxes across the gills of rainbow trout 1 Responses toenvironmental hyperoxia J Exp Biol 152 521ndash547 1990
13 Grunder S Geissler HS Bassler EL Ruppersberg JP A new memberof acid-sensing ion channels from pituitary gland Neuroreport 11 1607ndash1611 2000
14 Harvey BJ Energization of sodium absorption by the H-ATPase pumpin mitochondria-rich cells of frog skin J Exp Biol 172 289ndash309 1992
15 Hirata T Kaneko T Ono T Nakazato T Furukawa N Hasegawa SWakabayashi S Shigekawa M Chang MH Romero MF Hirose SMechanism of acid adaptation of a fish living in low pH 35 lake Am JPhysiol Regul Integr Comp Physiol 284 R1199ndashR1212 2003
16 Holzer P Acid-sensitive ion channels and receptors Hand Exp Pharma-col 194 283ndash332 2009
17 Hung CY Tsui KN Wilson JM Nawata CM Wood CM Wright PARhesus glycoprotein gene expression in the mangrove killifish Kryptole-bias marmoratus exposed to elevated environmental ammonia levels andair J Exp Biol 210 2419ndash2429 2007
18 Hwang PP Lee TH Lin LY Ion regulations in fish gills recentprogresses in the cellular and molecular mechanisms Am J Physiol RegulIntegr Comp Physiol 301 R28ndashR47 2011
19 Ivanis G Esbaugh AJ Perry SF Branchial expression and localizationof SLC9A2 and SLC9A3 sodiumhydrogen exchangers and their possiblerole in acid-base regulation in freshwater rainbow trout (Oncorhynchusmykiss) J Exp Biol 211 2467ndash2477 2008
20 Jasti J Furukawa H Gonzales EB Gouaux E Structure of acid-sensingion channel 1 at 19A resolution and low pH Nature 449 316ndash323 2007
21 Jonz MG Nurse CA Epithelial mitochondria-rich cells and associatedinnervation in adult and developing zebrafish J Comp Neurol 497817ndash832 2006
C264 INVOLVEMENT OF ASICs IN FISH GILL Na TRANSPORT
AJP-Cell Physiol bull doi101152ajpcell003982013 bull wwwajpcellorg
22 Jonz MG Nurse CA New developments on gill innervation insightsfrom a model vertebrate J Exp Biol 211 2371ndash2378 2008
23 Kellenberger S Epithelial sodium and acid-sensing ion channels InSensing With Ion Channels edited by Martinac B New York Springer2008 chapt 11 p 225ndash246
24 Kellenberger S Schild L Epithelial sodium channeldegenerin family ofion channels a variety of functions for a shared structure Physiol Rev 82735ndash767 2002
25 Kleyman TR Cragoe EJ Amiloride and its analogs as tools in the studyof ion-transport J Membr Biol 105 1ndash21 1988
26 Krogh A The active absorption of ions in some freshwater animals ZVergl Physiol 25 335ndash350 1938
27 Levanti MB Guerrera MC Calavia MG Ciriaco E Montalbano GCobo J Germana A Vega JA Acid-sensing ion channel 2 (ASIC2) inthe intestine of adult zebrafish Neurosci Lett 494 24ndash28 2011
28 Lin H Randall DJ Evidence for the presence of an electrogenic protonpump on the trout gill epithelium J Exp Biol 161 119ndash134 1991
29 Maetz J Garcia Romeu F The mechanism of sodium and chlorideuptake by the gills of a fresh-water fish Carassius auratus II Evidencefor NH4
Na and HCO3Cl exchanges J Gen Physiol 47 1209ndash1227
196430 Nakada T Hoshijima K Esaki M Nagayoshi S Kawakami K Hirose
S Localization of ammonia transporter Rhcg1 in mitochondrion rich cellsof yolk sac gill and kidney of zebrafish and its ionic strength-dependentexpression Am J Physiol Regul Integr Comp Physiol 293 R1743ndashR17532007
31 Nawata CM Hung CC Tsui TK Wilson JM Wright PA Wood CMAmmonia excretion in rainbow trout (Oncorhynchus mykiss) evidence forRh glycoprotein and H-ATPase involvement Physiol Genomics 31463ndash474 2007
32 Parks SK Tresguerres M Galvez F Goss GG Intracellular pH regu-lation in isolated trout gill mitochondrion-rich (MR) cell subtypes evi-dence for NaH activity Comp Biochem Physiol A 155 139ndash1452010
33 Parks SK Tresguerres M Goss GG Interactions between Na channelsand Na-HCO3
cotransporters in the freshwater fish gill MRC a modelfor transepithelial Na uptake Am J Physiol Cell Physiol 292 C935ndashC944 2007
34 Parks SK Tresguerres M Goss GG Theoretical considerations under-lying Na uptake mechanisms in freshwater fishes Comp BiochemPhysiol C 148 411ndash418 2008
35 Paukert M Sidi S Russell C Siba M Wilson SW A family ofacid-sensing ion channels from zebrafish widespread expression in thecentral nervous system suggests a conserved role in neuronal communi-cation J Biol Chem 279 18783ndash18791 2004
36 Perry SF The chloride cell structure and function in the gill of freshwaterfish Annu Rev Physiol 59 325ndash347 1997
37 Perry SF Randall DJ Effects of amiloride and SITS on branchial ionfluxes in rainbow trout Salmo gairdneri J Exp Zool 215 225ndash228 1981
38 Preest MR Gonzales RJ Wilson RW Pharmacological examination ofNa and Cl transport in two species of freshwater fish Physiol BiochemZool 78 259ndash272 2005
39 Reid SD Hawkings GS Galvez F Goss GG Localization and charac-terization of phenamil-sensitive Na influx in isolated rainbow trout gillepithelial cells J Exp Biol 206 551ndash559 2003
40 Sakai H Lingueglia E Champigny G Mattei MG Lazdunski MCloning and functional expression of a novel degenerin-like Na channelgene in mammals J Physiol 519 323ndash333 1999
41 Shih TH Horng JL Liu ST Hwang PP Lin LY Rhcg1 and NHE3b areinvolved in ammonium-dependent sodium uptake by zebrafish larvaeacclimated to low-sodium water Am J Physiol Regul Integr Comp Physiol302 R84ndashR93 2012
42 Springauf A Grunder S An acid-sensing ion channel from shark(Squalus acanthias) mediates transient and sustained responses to protonsJ Physiol 588 809ndash820 2010
43 Spry DJ Wood CM Ion flux rates acid-base status and blood gases inrainbow trout Salmo gairdneri exposed to toxic zinc in natural soft waterCan J Fish Aquat Sci 42 1332ndash1337 1985
44 Sullivan GV Fryer JN Perry SF Immunolocalization of proton pumps(H-ATPase) in pavement cells of rainbow trout gill J Exp Biol 1982619ndash2629 1996
45 Tresguerres M Parks SK Katoh F Goss GG Microtubule-dependentrelocation of branchial V-H-ATPase to the basolateral membrane in thePacific spiny dogfish (Squalus acanthias) a role in base secretion J ExpBiol 209 599ndash609 2006
46 Vina E Parisi V Cabo R Laura R Lopez-Velasco S Lopez-Muniz AGarcia-Suarez O Germana A Vega JA Acid-sensing ion channels(ASICs) in the taste buds of adult zebrafish Neurosci Lett 536 35ndash402013
47 Waldmann R Lazdunski M H-gated cation channels neuronal acidsensors in the NaCDEG family of ion channels Curr Opin Neurobiol 8418ndash424 1998
48 Wilson JM Laurent P Tufts BL Benos DJ Donowitz M Vogl AWRandall DJ NaCl uptake by the branchial epithelium in freshwater teleostfish an immunological approach to ion-transport protein localization JExp Biol 203 2279ndash2296 2000
49 Wood CM Randall DJ Sodium balance in rainbow-trout (Salmo-gaird-neri) during extended exercise J Comp Physiol 82 235ndash256 1973
50 Wright PA Wood CM A new paradigm for ammonia excretion inaquatic animals role of rhesus (Rh) glycoproteins J Exp Biol 2122303ndash2312 2009
51 Yan JJ Chou MY Kaneko T Hwang PP Gene expression of NaH
exchanger in zebrafish H-ATPase-rich cells during acclimation to low-Na and acidic environments Am J Physiol Cell Physiol 293 C1814ndashC1823 2007
C265INVOLVEMENT OF ASICs IN FISH GILL Na TRANSPORT
AJP-Cell Physiol bull doi101152ajpcell003982013 bull wwwajpcellorg

MATERIALS AND METHODS
Animal holding Rainbow trout embryos were obtained from theRaven Brood Trout Station (Caroline AB Canada) and grown to theappropriate size for experimentation (2 g for flux experiments and200ndash500 g as adults for other experiments) Fish were maintainedindoors in flow-through 450-liter fiberglass tanks supplied with aer-ated and dechlorinated Edmonton tap water (hardness as CaCO3 16mmoll alkalinity 120 mgl NaCl 05 mmoll pH 82 15degC) Fishwere fed ground dry commercial trout pellets (Purina trout chow)once daily and kept on a 1410-h light-dark photoperiod All animaluse was approved by the University of Alberta Animal Care Com-mittee under Protocol AUP00000072
Pharmacological inhibition of Na uptake For flux studies juve-nile trout were first acclimated to low-ionic-strength pH 60 water for4 days prior to Na flux experiments The duration of acclimation wasdeemed sufficient on the basis of a previous soft water exposure studyin rainbow trout (Salmo gairdneri) (43) Fish were transferred to60-liter plastic tubs supplied with aerated recirculating low-ionic-strength water (Na 300 30 moll Ca2 24 moll Cl 780 20 moll) and kept at a constant temperature of 15degC Afteracclimation two series of flux experiments were completed The firstexperiment investigated concentration-dependent effects of two ASIC-inhibiting diarylamidines 4=6-diamidino-2-phenylindole (DAPI 0001001 01 10 and 10 moll Sigma) and diminazene (00003 0003003 03 and 30 moll Sigma) on Na uptake in juvenile troutThis experiment established the 50 inhibitory concentrations (IC50Hill function) and optimal concentrations (80 inhibition) forDAPI and diminazene The second flux experiment compared theeffects of two well-known pharmacological Na uptake inhibitors[amiloride (500 moll) and phenamil (50 moll) Sigma] and thetwo ASIC inhibitors [DAPI (1 moll) and diminazene (3 moll)] onNa uptake in juvenile trout
For all flux experiments fish were transferred to individual dark-ened 60-ml flux chambers supplied with a constant flow of aeratedlow-ionic-strength low-pH water and acclimated to the chambers for24 h prior to experimentation Unidirectional Na influx was mea-sured using radiolabeled 22Na as described previously (12) Brieflyflow of water to chambers was turned off 01 Cil 22Na was addedto each chamber and allowed to mix for 5 min and either the DMSOcontrol (005 DMSO) or pharmacological agents dissolved inDMSO were added to the chambers After a 5-min equilibrationperiod 6-ml water samples were collected at time 0 and 90 and 180min as appropriate At the completion of the experiment fish wereterminally anesthetized (MS-222 1 gl) and weighed Water samples(3 ml) were analyzed for 22Na radioactivity using a gamma counter(Packard Cobra II Auto Gamma model 5010 Perkin ElmerWaltham MA) and total concentration of Na was measured usingatomic absorption spectrophotometry (model 3300 Perkin ElmerShelton CT) Unidirectional 22Na influx (molmiddotkg1middoth1) was cal-culated for each flux period according to the following equation JNain cpm V(SA t M) where cpm is the difference between theinitial and final radioactivities in the water (cpmml) V is the waterflux volume (ml) SA is the average specific activity (cpmmol Na)as measured at the beginning and end of the flux period t is the timeelapsed (h) and M is the mass of the fish (kg)
Tissue collection and preparation RNA isolation for expressionanalysis was performed on adult fish Briefly fish were euthanized asdescribed above a blood sample was withdrawn from the caudal archand the brain head kidney and trunk kidney were dissected out andimmediately freeze-clamped in liquid N2 for later processing For gilltissue the animal was first perfused with ice-cold heparinized (15mg) phosphate-buffered saline (PBS in mM 137 NaCl 27 KCl 43Na2HPO4 and 14 NaH2PO4 pH 78) and gill tissue or MRCs (asappropriate) were obtained according to the protocols described else-where (10) After perfusion gill arches were processed for MRCisolation freeze-clamped in liquid N2 for RNA isolation or placed in
fixative for immunohistochemistry or scanning electron microscopy(SEM) (see below)
MRC isolation and cellular imaging Adult rainbow trout (300ndash500 g) gills were perfused with PBS to remove blood according tooriginal protocols (10 33) Subsequently gill filaments were removedfrom the rakers cut into sections (2ndash5 mm 3ndash6 filaments) rinsed inPBS (in mM 137 NaCl 27 KCl 43 Na2HPO4 and 14 NaH2PO4pH 78) and subjected to three (20-min) incubations in 5 ml of 005trypsin-EDTA with shaking (200 rpm) at room temperature Thesubsequent cellular suspensions following each incubation werepassed through a 64-m nylon mesh filter into 10 ml of ice-cold fetalbovine serum and rinsed through with PBS to halt trypsin activity Thecells were then centrifuged (5 min 1500 g 4degC) and washed threetimes with 25 ml of PBS The final pellet was resuspended in 2 ml ofice-cold PBS and applied to a three-step Percoll density gradient (2ml 109 gml 2 ml 105 gml and 3 ml 103 gml) and centrifuged(45 min 2000 g 4degC 0-brake) After centrifugation the cell layer atthe 109-105 interface was removed washed three times in ice-coldPBS and resuspended in 800 l of ice-cold PBS
Isolated MRCs (500000) in PBS with an additional 2 l ofMgCl2 (1 M) and CaCl2 (1 M) to aid in cell attachment were placedon pretreated (1 M HCl acid-washed 01 poly-L-lysine-coated andrinsed with double-distilled H2O and 70 ethanol) 15-mm roundglass coverslips (catalog no CS-15R Warner Instruments) and al-lowed to attach for 2 h at 4degC After attachment coverslips wereplaced at room temperature and after careful removal of PBS thecells were briefly rinsed in Na-containing buffer (in mM 1425NaCl 50 CaCl2 10 MgCl2 40 KCl 15 HEPES and 25 NaHCO3pH 78) The cells were then incubated in 200 l of Na-containingbuffer which contained 2 l of 5 mM pH-sensitive BCECF-AM (50g in 16 l of DMSO and 20 pluronic acid) for 30 min at 18degCCoverslips were placed into a 70-l imaging chamber (model RC-20H Warner Instruments) for the perfusion experiments The cham-ber was fixed to an inverted fluorescence microscope (Nikon EclipseTE300) and the cells were subjected to differential interferencecontrast microscopy and fluorescent imaging To allow forBCECF-AM excitation at 495 and 440 nm the microscope was fittedwith a xenon arc lamp (Lambda DG-4 Sutter Instruments NovatoCA) Images were digitally obtained at 440 and 495 nm on a mono12-bit charge-couple device camera (Retiga EXi QImagingBurnaby BC Canada) every 17 s during perfusion experimentsRatios of fluorescence at 495 nm to fluorescence at 440 nm weredigitally compiled using Northern Eclipse software (MississaugaON Canada)
Cell perfusion Solutions were perfused across the attached MRCsin the holding chamber by gravity feed A six-input manifold (modelMp-6 Warner Instruments) attached to 60-ml syringe holder blockscontrolled by pinch valves (model VE-6 Warner Instruments) mon-itored manually with VC-6 valve controllers (Warner Instruments)was used to perfuse the solutions at a rate of 05 mlmin Cells weremonitored from original resting state for changes in intracellular pH(pHi) when exposed to Na-free and Na-containing solutions Start-ing in Na-free conditions (in mM 1425 N-methyl-D-glucamine-Cl25 C5H14NOmiddotHCO3
5 CaCl2 1 MgCl2 4 KCl and 15 HEPES pH78) followed by replacement with a Na-containing solution wemonitored the activity of the NHE in trout MRCs following a pHi
disturbance An identical transition between Na-free and Na-containing solutions was made but this time in the presence of 1 MDAPI or 100 M ethylisopropylamiloride (EIPA) EIPA was used inthis experiment since it potently inhibits NHE but has a low affinityfor Na channels (25) Perfusion solutions were bubbled with 03CO2 balanced with O2 throughout the experiments At the end of eachexperimental perfusion cells were subjected to a final high-K
solution calibration protocol (in mM 120 potassium gluconate 20KCl 2 MgCl2 and 20 HEPES) in which four solutions were adjustedto pH levels (840 780 720 and 660) by the addition of theionophore nigericin (5 M) to equilibrate pHi and extracellular pH
C256 INVOLVEMENT OF ASICs IN FISH GILL Na TRANSPORT
AJP-Cell Physiol bull doi101152ajpcell003982013 bull wwwajpcellorg
The ratios of fluorescence at 495 nm to fluorescence at 440 nm at eachstep of the pH calibration were used to form a regression equation foreach individual cell trace This equation was extrapolated to the rawratio data obtained over the entire time course yielding a calibratedpHi for each individual cell monitored during the perfusion experi-ment Data were collected and analyzed for each calibrated individualcell by comparing the rate of alkalinization (pHit) under thecontrol parameters with that in the presence of the drug to determinepercent inhibition
Preparation of total RNA Total RNA was extracted from frozentissues using TRIzol reagent (Invitrogen Carlsbad CA) according tothe manufacturerrsquos instructions RNA was treated with DNase I(Ambion Austin TX) according to the manufacturerrsquos protocol thenan on-column cleanup was carried out using an RNeasy Mini Kit(Qiagen Mississauga ON Canada) The quality of the extractedRNA was assessed by a bioanalyzer (model 2100 Agilent Technol-ogies Palo Alto CA) or by visualization on a formaldehyde gel RNAconcentration was measured with a spectrophotometer (ND-1000UV-Vis NanoDrop Technologies Rockland DE)
Molecular cloning and phylogenetic analysis Rapid amplificationof cDNA ends (RACE)-ready cDNA was synthesized from gill RNAusing a SMARTer RACE cDNA amplification kit (Clontech Moun-tain View CA) according to the manufacturerrsquos instructions cDNAsolutions were diluted 110 with tricine-EDTA buffer provided in thekit and stored at 20degC
Oligonucleotides for cloning of ASIC1 and ASIC4 genes weredesigned with PrimerQuest (Integrated DNA Technology) (Table 1)on the basis of conserved regions in available ASIC mRNA sequencesfrom zebrafish tilapia pufferfish stickleback and cod (NationalCenter for Biotechnology Information Ensembl) Sequences werealigned with GeneDoc (version 2602 httpwwwpscedubiomedgenedoc) and ClustalX (version 181) software DNA sequencesencoding the 5=- and 3=-termini of the ASIC1 gene and the 5=-terminus of the ASIC4 gene were obtained using the Advantage 2PCR kit (Clonetech) with RACE-ready cDNA templates generatedfrom gill RNA The ASIC4 3=-terminus was obtained using degener-ate primer (Table 1) Resulting PCR products were excised from thegel and purified with a QIAquick gel extraction kit (Qiagen) Subse-quently amplicons of interest were sequenced directly or ligated intopJET12 vector using a CloneJET kit (Thermo Scientific) for verifi-cation Recombinant plasmids were transformed into competent cells
(One Shot TOP10 chemically competent Escherichia coli Invitro-gen) Colonies were screened for plasmids containing inserts of thecorrect size by agarose gel electrophoresis and sequenced withpJET12-specific primers provided in the CloneJET kit
The amino acid sequences derived from our cloned trout ASIC4and ASIC1 genes (accession numbers KF964646 and KF964645respectively) together with the previously annotated amino acidsequences of ASIC1 ASIC2 ASIC3 and ASIC4 from various fishspecies and other taxa were aligned and analyzed using Seaviewsoftware (version 4 httppbiluniv-lyon1frsoftwareseaview) Arooted phylogenetic tree showing evolutionary relationships betweenthe different ASIC proteins was generated using the maximum-likelihood method and LG model and 700 bootstrap resamplings
RT-PCR Tissue-specific cDNA templates utilized for RT-PCRwere synthesized from 3 g of RNA from gill MRCs kidney brainor blood using SuperScript III reverse transcriptase (Invitrogen) ac-cording to protocols provided by the manufacturer ASIC4 and ASIC1amplicons were obtained using primers designed with PrimerQuest(Table 1) PCR conditions were as follows 98degC for 1 min of initialdenaturation followed by 35 cycles of denaturation at 98degC for 10 sannealing at 62degC for 30 s and elongation at 72degC for 40 s with thefinal elongation at 72degC for 10 min PCR products were visualized by1 agarose gel electrophoresis followed by ethidium bromide stain-ing
Immunoprecipitation of ASIC4 and Western blot analysis Ananti-zebrafish ASIC4b (zASIC42) polyclonal antibody against twopeptides corresponding to regions near the NH2 terminus (aa 146ndash160 PKSRKGHRPSELQYP) and the COOH terminus (aa 519ndash533CFEEVKVKAANDVAQP) of zASIC42 protein (accession noQ708S31) was custom-made by 21st Century Biochemicals (Marl-boro MA) The polyserum was affinity-purified against each of thesepeptides independently and the NH2-terminal peptide-purified anti-body was used exclusively in this study Immunoprecipitation wasperformed according to the technique described by Goss and col-leagues (11) Briefly trout gills were perfused with heparinizedice-cold PBS (Ca2-free) and the second and third gill arches weredissected out and washed three times in ice-cold PBS Gill rakers wereremoved and the remaining gill arches were cut into 200- to 300-mgsections Each section was lysed for 30 min in 2ndash3 ml of lysis buffer(100 mM NaCl 4 mM KCl 50 mM HEPES 5 mM EDTA pH 74and 1 Triton-X) containing protease inhibitors (cOmplete MiniEDTA-free protease inhibitor tablets Roche Diagnostics MannheimGermany) After centrifugation to remove the debris (5000 g 5 min4degC) the lysates were incubated with 2ndash4 l of anti-zASIC42antibody at 4degC overnight on an end-over-end rotator On the nextday 60 l of protein A-Sepharose CL4b beads (Sigma Chemical StLouis MO) preswelled in immunoprecipitation (IP) buffer (in mM100 NaCl 4 KCl 50 HEPES and 5 EDTA pH 74) and preblockedwith 1 bovine serum albumin (fraction V Sigma) were added to thelysates which were incubated at 4degC for 4ndash6 h on the rotator Afterthe incubation the supernatant was removed and the beads werewashed three times with 1 ml of IP buffer After the washes the beadswere incubated in Laemmli buffer for 15 min at 65degC and centrifugedand the supernatant was retained for Western blot analysis as de-scribed previously (45) Briefly the samples were separated on a 75polyacrylamide minigel and transferred to a nitrocellulose (NC) mem-brane using a wet transfer system (Bio-Rad Laboratories HerculesCA) The NC membrane was blocked in 5 bovine serum albumin inTris-buffered saline with 02 Triton X-100 (TBST) for 30 min andincubated with rabbit anti-zASIC42 antibody (11000 dilution) on arocker at 4degC overnight The membrane was washed four times for 15min each with TBST blocked again with 5 bovine serum albuminfor 15 min and then incubated with a secondary horseradish peroxi-dase-conjugated goat anti-rabbit antibody (Santa Cruz BiotechnologyDallas TX) at 150000 dilution at room temperature for 1 h The NCmembrane was washed four times in TBST and the immunoreactivebands were visualized using a SuperSignal West Pico chemilumines-
Table 1 Primer sets for cloning and tissue distributionof ASICs
Gene Primer Sequence
CloningASIC1
5= 5=-TCACCCAGCAACCCTGCGAACTCGT-3=3= 5=-ACACCCTGCAACATGACGCGCT-3=
ASIC45= 5=-GATGTCCCGTCCGGTTGAAGATGTC-3=Forward 5=-CCCAGTAACATCAAGTGTGTCG-3=Reverse 5=-YYARCANGCRAARTCYTC-3=
RT-PCRASIC1
Forward 5=-AAGTCCACTCCCATAGA-3=Reverse 5=-CAGCCAGGTTATTCCTT-3=
ASIC4Forward 5=-CTTTCGTTTCTCTGCTCTCACC-3=Reverse 5=-CAAGACCAGGAAGTTGTCTCTG-3=
GAPDHForward 5=-AAGGGTGAGGTGAGCATGGA-3=Reverse 5=-GCTTTACCCCATGGGATCTCAT-3=
ASIC acid-sensing ion channel 5= 5=-rapid amplification of cDNA ends(RACE) primer 3= 3=RACE primer Y C T R A G N A G T C
C257INVOLVEMENT OF ASICs IN FISH GILL Na TRANSPORT
AJP-Cell Physiol bull doi101152ajpcell003982013 bull wwwajpcellorg
cent substrate kit (Thermo Scientific) following the manufacturerrsquosinstructions
SEM and immunohistochemistry For examination of the surface ofthe gill by SEM filaments were fixed in 3 glutaraldehyde and 2paraformaldehyde in 015 mM sodium cacodylate buffer (pH 74 290mosM) for 1 h at 4degC dehydrated in a graded series of ethanol andcritical point dried Samples were then mounted on an SEM stubsputter-coated with gold and examined with a scanning electronmicroscope (model JSM-6301FXV Jeol)
For immunohistochemistry gills were excised from adult rain-bow trout fixed in 4 paraformaldehyde in PBS (pH 74) over-night at 4degC dehydrated in a graded series of ethanol andembedded into paraffin blocks Serial sections (5 m) were cut andrehydrated in a decreasing ethanol series followed by double-distilled H2O Rehydrated sections were incubated in 10 mMcitrate buffer at 70degC for 1 h for epitope retrieval After incubationslides were washed in PBS and then blocked with 6 milk powderin a humidified chamber for 1 h Sections were then incubatedovernight at 4degC simultaneously with the anti-zASIC42 antibody(1400 dilution) and anti-Na-K-ATPase (NKA) monoclonalantibody (1400 dilution anti-chicken 5-subunit DevelopmentalStudies Hybridoma Bank University of Iowa) followed by sec-ondary FITC-conjugated anti-mouse antibody (1500 dilution In-vitrogen Oregon) and tetramethylrhodamine-conjugated anti-rab-bit antibody (1500 dilution Invitrogen Oregon) Slides incubatedwith preimmune serum from the anti-zASIC42 antibody produc-tion in place of primary antibody were used as a negative controlSlides were viewed with a laser scanning confocal microscope(model LSM 710 Zeiss Germany) at the Cross Cancer InstituteCell Imaging Facility (Edmonton AB Canada) Images wereprocessed with LSM Image Browser (version 420121 CarlZeiss) Surface rendering and three-dimensional reconstruction ofz-stack images were performed using Imaris software (version622 Bitplane Zurich Switzerland)
Statistical analysis Na flux data were subjected to statisticalanalysis and are reported as means SE All pharmacologicalexperiment data sets were tested for homogeneity of variance andcompared using one-way ANOVA (SigmaPlot version 11 SystatChicago IL) If significant differences (P 005) were found a posthoc multiple-comparisons Tukeyrsquos test was applied to determine thesedifferences A paired t-test (P 005) was used in the pHi imagingexperiments to compare the relative inhibition of pHit in isolatedcells under control conditions and after addition of each pharmaco-logical inhibitor
RESULTS
Pharmacological inhibition Exposure of juvenile rainbowtrout to increasing concentrations of DAPI resulted in a con-centration-dependent decrease in Na uptake with 90inhibition at 1 moll (Fig 1A) The DAPI concentration atwhich there was a 50 inhibition in Na uptake (IC50) wascalculated to be 012 moll Exposure to diminazene alsoresulted in a concentration-dependent decrease in Na uptakein juvenile trout with a maximum reduction of Na uptakefrom 303 43 (control) to 51 19 molmiddotkg1middoth1 (79)after exposure to 30 moll diminazene (Fig 1B) The IC50 fordiminazene was calculated to be 096 moll
Na uptake rates in juvenile rainbow trout were significantlylower in the first 90-min flux time period after exposure to 500M amiloride (17 185 molmiddotkg1middoth1) 1 M DAPI (29 189 molmiddotkg1middoth1) and 3 M diminazene (41 334molmiddotkg1middoth1) than in controls (539 1709 molmiddotkg1middoth1 Fig 2) while 50 M phenamil (299 1079 molmiddotkg1middoth1) did not significantly reduce Na uptake Howeverfor the second consecutive flux period between 90 and 180min of exposure to the pharmacological agents the inhibitionwas less pronounced but still significant for amiloride (104 334 molmiddotkg1middoth1) and DAPI (141 627 molmiddotkg1middoth1)while diminazene (297 415 molmiddotkg1middoth1) was no longersignificantly different from control (393 441 molmiddotkg1middoth1 Fig 2)
MRC pHi imaging Results presented from the whole-animalflux experiments showed that administration of DAPI at vari-ous concentrations significantly inhibited Na uptake (Fig1A) To demonstrate that the inhibition of Na uptake attrib-uted to DAPI was in fact due to the blocking of ASIC-type Na
channels rather than affecting NHE activity we conductedexperiments using pHi imaging NHE activity was measuredby examining the rate of pHi alkalization (pHit) uponexposure of MRCs to a Na-containing medium (145 mM) inthe absence or presence of DAPI (1 M) or EIPA (100 M)
A representative trace (Fig 3A) shows a Na-dependentalkalization following Na application (1st rise) while addi-tion of EIPA significantly decreased the rate of alkalization(2nd rise) The combined results (n 19) show a control of
0
50
100
150
200
250
300
350
400
450
0 0001 001 01 1 10
JNa I
N(μ
mol
kg-1
h-1 )
DAPI concentration (μmol l-1)
0
50
100
150
200
250
300
350
400
0 00003 0003 003 03 3
JNa I
N(μ
mol
k g-1
h-1 )
Diminazene concentration (μmol l-1)
A B
Fig 1 Dose-dependent decreases in Na uptake rates (JNain) in juvenile trout after exposure to increasing concentrations of 4=6-diamidino-2-phenylindole(DAPI) and diminazene in low-Na (30 M) and low-pH (pH 60) water Values are means SE (n 6) P 005 vs control
C258 INVOLVEMENT OF ASICs IN FISH GILL Na TRANSPORT
AJP-Cell Physiol bull doi101152ajpcell003982013 bull wwwajpcellorg
0642 0040 pHimin while the alkalization rate wasreduced by 62 in the presence of EIPA (0 240 0036pHimin P 0001 Fig 3B) For DAPI Na-dependentalkalization following Na application was similar in the
presence or absence of DAPI (Fig 3D) No differences inpHit were noted between control (0542 0042 pHimin) and DAPI-treated (0578 0047 pHimin) cells(P 0149 n 22 Fig 3D)
0
100
200
300
400
500
600
700
800
Cont Amil DAPI Dim Phen Cont Amil DAPI Dim Phen
JNa I
N(μ
mol
kg-1
h-1 )
Treatment
T0-90min T90-180min
Fig 2 Effects of pharmacological agents [control (Cont DMSO) amiloride (Amil 500 moll) DAPI (1 moll) diminazene (Dim 3 moll) and phenamil(Phen 50 moll)] during consecutive 90-min measurements of Na uptake in juvenile trout in low-Na (30 M) and low-pH (pH 60) water Values are means SE(n 6) P 005 vs control
65
7
75
8
85
9
0 100 200 300 400 500 600
pHi
Time (sec)
Na FreeNa Free Na
EIPA
Na
0
01
02
03
04
05
06
07
08
Na+ Na+EIPA
ΔpH
iΔM
in
Cell Alkalinization Treatments
65
7
75
8
85
9
0 100 200 300 400 500 600
pHi
Time (sec)
Na Free Na
DAPI
Na Na Free
0
01
02
03
04
05
06
07
08
Na+ Na+DAPI
ΔpH
iΔM
in
Cell Alkalinization Treatments
A
DC
B
Fig 3 Effect of ethylisopropylamiloride (EIPA) and DAPI on Na-induced alkalinization of peptide nucleic acid-positive (PNA) cells following Na-freeexposure at resting intracellular pH (pHi) A representative trace demonstrating inhibition of cell alkalinization by EIPA (100 M) B quantitative analysis ofcell alkalinization in the absence and presence of EIPA (62 inhibition) Values are means SE (n 19 cells) P 0001 (by paired t-test) C representativetrace demonstrating lack of effect on cell alkalinization in the presence of DAPI (1 M) D quantitative analysis of cell alkalinization in the absence and presenceof DAPI Values are means SE (n 22) P 005 (by paired t-test)
C259INVOLVEMENT OF ASICs IN FISH GILL Na TRANSPORT
AJP-Cell Physiol bull doi101152ajpcell003982013 bull wwwajpcellorg
Phylogenetic analysis and tissue distribution of ASIC1 andASIC4 Two distinct DNA fragments whose derived aminoacid sequences exhibit similarity to the ASIC family werecloned from the gill of rainbow trout To determine theiridentity we conducted a phylogenetic analysis by comparisonwith known ASIC homologs from a variety of species Thegenerated phylogenetic tree (Fig 4) showed that the twocloned rainbow trout ASIC genes (tASICs) were members ofthe ASIC1 and ASIC4 subunits respectively (Fig 4A) ThetASIC1 (partial clone) grouped most closely with ASIC1 fromsalmon (Salmo salar) while tASIC4 grouped with other iden-
tified ASIC4 family members from other fish and mammalspecies
Furthermore we conducted a tissue distribution analysis oftASIC1 and tASIC4 by RT-PCR We observed mRNA expres-sion of tASIC1 in all tested tissues gill MRCs head and trunkkidney brain and blood tASIC4 had a slightly differentdistribution it was present in the perfused gill MRCs brainand trunk kidney but it was absent in head kidney and bloodtissues
Immunoprecipitation and immunostaining with anti-zASIC42To determine if ASIC4 was present in the fish gill and to validate our
Fig 4 Phylogenetic analysis of amino acid sequences of cloned rainbow trout acid-sensing ion channels ASIC1 and ASIC4 and ASICs from various fish speciesPhylogenetic tree was constructed using the maximum-likelihood method and numbers indicate bootstrap values for 700 resamplings GenBank or Ensemblaccession numbers are as follows human ASIC2 NP_0010852 Western clawed frog ASIC2 XP_0029330862 Japanese pufferfish ASIC2 XP_0039648711zebrafish ASIC2 NP_9999531 three-spined stickleback ASIC2 ENSGACG00000009558 Atlantic cod ASIC2 ENSGMOT00000019145 spotted greenpufferfish ASIC2 ENSTNIG00000014943 Southern platyfish ASIC2 ENSXMAG00000008283 oyster toadfish ASIC2 AAP456331 coelacanth ASIC2ENSLACG00000007191 rat ASIC2 NP_0370242 Southern platyfish ASIC1c ENSXMAG00000018841 African clawed frog ASIC11 AEC486651 Africanclawed frog ASIC12 AEC486671 brown rat ASIC1 NP_0770681 human ASIC1 NP_0644232 three-spined stickleback ASIC1a ENSGACG00000003075Japanese pufferfish ASIC1 XP_0039635201 Southern platyfish ASIC1a XP_0058018011 Atlantic cod ASIC1a ENSGMOG00000009659 zebrafishASIC12 NP_9999551 rainbow trout ASIC1 KF964646 Atlantic salmon ASIC1 NP_0011334561 oyster toadfish ASIC1 Q7T1N41 elephant shark ASIC1AEC486681 three-spined stickleback AIC1c ENSGACG00000017974 Atlantic cod ASIC1c ENSGMOG00000003105 zebrafish ASIC13 NP_9999541river lamprey ASIC1 AAY289831 sea lamprey ASIC1b ENSPMAG00000003813 dogfish shark ASIC1 AAY289851 zebrafish ASIC11 NP_9999561human ASIC4 NP_8782672 rat ASIC4 NP_0715702 African clawed frog ASIC4 AEC486661 Japanese pufferfish ASIC4 XP_0039621271 spotted greenpufferfish ASIC4b ENSTNIG00000010994 Nile tilapia ASIC4 XP_0034454461 Southern platyfish ASIC4a ENSXMAG00000003624 three-spinedstickleback ASIC4a ENSGACG00000007052 Atlantic cod ASIC4a ENSGMOG00000012740 rainbow trout ASIC4 KF964645 zebrafish ASIC41NP_9999521 zebrafish ASIC42 NP_9999511
C260 INVOLVEMENT OF ASICs IN FISH GILL Na TRANSPORT
AJP-Cell Physiol bull doi101152ajpcell003982013 bull wwwajpcellorg
antibody for later immunocytochemistry we performed an im-munoprecipitation using an antibody raised against zASIC42on the basis of sequence similarly of 60 between zASIC42(aa 146 ndash160 PKSRKGHRPSELQYPP) and the clonedtASIC4 (aa 142ndash156 PKNREGHKPTDLDYPA) Using West-ern blotting we analyzed the precipitates for the presence ofASIC4 (see Fig 6A) and found a single band of 65 kDawhich approximates the predicted size for tASIC4 (604 kDa)The strong band at 55 kDa corresponds to the IgG heavychain
To determine the location of ASIC4 protein within therainbow trout gill tissue fixed sections were double-labeledwith anti-zASIC42 antibody (see Fig 6B) and anti-NKA andthen counterstained with the nuclear stain DAPI Immunore-activity was observed in cells located in the lamellae andinterlamellar region (see Fig 6B arrowhead) Moreover themajority of cells positive for zASIC42 also colocalized withNKA protein (Fig 6 C and D arrowhead) a known marker ofMRCs suggesting that in gill epithelium ASIC4 protein ismainly expressed in MRCs SEM analysis of the gill filamentdemonstrated that MRCs are clearly distinguished on the sur-face of the epithelium and have a broad apical convex openingto the environment (see Fig 7A arrowheads) The generatedthree-dimensional reconstructions of the staining immunoreac-tivity pattern for ASIC4 and NKA proteins revealed the pres-ence of ASIC4 and NKA in the same cells but in distinctregions of the cells NKA was present on the basolateralmembrane (arrows green staining) while ASIC4 was localizedto the broad apical surface of the cell (arrowheads red stain-ing) Figure 7B shows a generated image with a top view whileFig 7C is the same cell viewed from the side
DISCUSSION
The existence of an apical Na channel electrochemicallycoupled to a VHA mediating Na uptake from dilute FW hasbeen postulated on theoretical thermodynamic grounds for anumber of decades (1 36 34) However an inability todetermine the molecular identity of the Na channel has beena major drawback in the general acceptance of the modelUsing a combination of pharmacological blockade isotopicfluxes immunocytochemistry and molecular biology we pro-pose that an ASIC family member is potentially the long-sought-after apical Na channel responsible for Na uptake byrainbow trout acclimatized to water with low Na and low pHOur study provides the first evidence for a nonneuronal role forASICs in any vertebrate whereby in gills from FW fishesASICs are associated with the MRCs and tASIC4 localizes tothe apical membrane of NKA positive cells
ASICs are a broad family of Na-specific ion channels thathave been extensively studied in mammalian models (forreview see Ref 23) with only a few studies performed in anyfish species (35 42 46) ASICs have primarily been associatedwith neuronal tissues with studies focusing on a neuronalfunction for ASICs (23) In this study ASIC1 and ASIC4 werecloned in rainbow trout using PCR and they were expressed ina variety of tissues both neuronal and nonneuronal in origin(Fig 5) Importantly tASIC1 gene was present in all fiveinvestigated tissues including whole blood It appears that thetASIC4 gene has a more defined tissue distribution thantASIC1 with expression in the ionoregulatory tissues (trunk
kidney and perfused gill) and the brain Unlike tASIC1 therewas no apparent expression of tASIC4 in the head kidney orthe blood Therefore tASIC4 expression in MRCs and per-fused gill could not be attributed to blood contamination
Both tASIC1 and tASIC4 genes were expressed in the gillepithelium and more specifically were present in MRCs ofrainbow trout gills It is believed that similar to ENaCsfunctional ASICs are trimers (20) although their configurationhas not been resolved It has been demonstrated that with afew exceptions ASICs function as homomers (forming chan-nels of identical subunits) or heteromers (forming channelswith other ASIC subunits) (4 23) ASICs have been charac-terized by using oocyte expression in zebrafish (4 35) anddogfish shark (ASIC1b only) (42) It has been found thatcoexpression of zASIC41 with zASIC13 in Xenopus oocytesincreased the current amplitude and abundance (15-fold) ofthe channel at the cell surface indicating that these two ASICsubunits form a functional channel (4) Moreover thisASIC4113 heteromeric channel was more efficiently traf-ficked to the plasma membrane and had increased affinity forH Additionally zASIC41 when heterologously expressedin Xenopus oocytes has been shown to be also gated open bydecreases in extracellular Ca2 The signal responsible forgating of zASIC42 was unable to be determined in that study(4) It is therefore possible that tASIC1 and tASIC4 form aheteromeric channel in MRCs but this cannot be determined atthis time and would require confirmation by immunohisto-chemistry with an anti-ASIC1 antibody that is immunoreactivefor rainbow trout It is also possible that other ASIC subunitsandor regulatory elements may be involved in the function ofASICs in the FW trout gill A full characterization of tASIC4by oocyte expression and electrophysiology is required tounderstand the mechanisms of gating in low-Na low-Ca2
watersIn our study ASIC4 protein was colocalized with NKA and
was present on the apical surface of cells suggesting that it isfound primarily in MRCs (Fig 6 D and E) Previous studies onASICs in fishes have suggested that they are primarily associ-ated with neuronal tissue and sensory cells (27 35 42 46) Inadult and developing zebrafish morphological studies usingconfocal immunofluorescence techniques demonstrated thatgill MRCs come into direct contact with nerve fibers in thebasolateral region (21 22) However in our study ASIC4protein was clearly localized to the apical region of the MRCsas demonstrated by three-dimensional reconstruction of con-
Head k
idney
ASIC4
ASIC1
GAPDH
Gill MRCs
Brain
Trunk k
idney
Blood
No tem
plate
Fig 5 Tissue distribution analysis of ASIC4 and ASIC1 genes in adultrainbow trout as determined by PCR GAPDH was used as a reference geneMRCs mitochondrion-rich cells
C261INVOLVEMENT OF ASICs IN FISH GILL Na TRANSPORT
AJP-Cell Physiol bull doi101152ajpcell003982013 bull wwwajpcellorg
focal images ASIC4 immunoreactivity did not overlap withthe standard basolateral immunoreactive pattern associatedwith NKA (Fig 7 B and C) Therefore we conclude thatneuronally derived tissues cannot account for the expression ofASIC4 protein in MRCs We also corroborate the ASIC4staining pattern with the MRC external morphology whichshows that in FW rainbow trout raised in Edmonton tap water(500 M Na) the MRCs have wide apical openings (Fig7A) This apical localization of ASIC4 protein in the cells of atransporting epithelium is the first evidence for a nonneuronalfunction for any ASIC member and suggests that ASIC4 as aNa channel may be responsible for apical uptake of Na inFW fish gill
Pharmacological profiling has long been used to differentiatethe mechanisms of ion transport across epithelial tissues For Na
transport classical analysis has usually involved amiloride and itssubstituted derivatives (eg EIPA mercaptoisobutyric acid andphenamil) as a means to differentiate between the involvementof NHE isoforms and ENaC in Na transport (25) We inves-tigated two newly described ASIC-specific inhibitors dimina-zene and DAPI (5) for their potency in inhibiting unidirec-tional Na uptake in fish Diminazene and DAPI exhibited adose-dependent inhibitory relationship and both were veryeffective at low concentrations (Fig 1) Na uptake in rainbow
trout acclimatized to low-ionic-strength acidic water wasclearly inhibited by these diarylamidines at concentrationssimilar to those reported in a previous study of cultured rathippocampal neurons (5) However in our study DAPI provedto be more effective than diminazene in contrast to observa-tions in rat neurons The inhibition of 22Na uptake by DAPIand diminazene was also compared with other common Na
transport inhibitors amiloride and phenamil Amiloride wasobserved to be an effective inhibitor of Na uptake in rainbowtrout but only at relatively high concentrations (500 M)which is consistent with previous studies (2 37) Recentlyamiloride was also demonstrated to completely block ASIC-mediated currents (5 35 42) however it is also known toblock many other Na transporters including NHE and ENaC(25) Amiloride therefore cannot be used as a distinguishingpharmacological agent
While DAPI diminazene and amiloride almost completelyblocked Na transport in the present study phenamil had nosignificant effect on Na flux in rainbow trout This suggeststhat phenamil is a poor inhibitor of Na uptake in troutexposed to low-ionic-strength water (30 moll Na) Paststudies of the effects of phenamil on Na uptake in a variety ofFW fishes have reported inconsistent results For example inneon tetras acclimatized to low Na (50 moll) phenamil had
Fig 6 Immunoreactivity of anti-zebrafish ASIC42 (zASIC42) antibody in the gill of rainbow trout A immunoprecipitation Western blot from whole gillhomogenates with zASIC42 showing a distinct band of 65 kDa Lane 1 (L1) contains 300 mg of gill tissue and lane 2 (L2) contains 200 mg of gill tissueBndashE confocal images of gill sections labeled for ASIC4 and Na-K-ATPase (NKA) proteins B and C double-labeling with DAPI and anti-zASIC42 (B) andDAPI and anti-NKA (C) D merged image of B and C E higher-magnification image of a MRC showing that ASIC4 and NKA are present in distinct regionswithin the MRC F control micrograph with DAPI and no primary antibody Arrowheads (BndashD) indicate positive staining
C262 INVOLVEMENT OF ASICs IN FISH GILL Na TRANSPORT
AJP-Cell Physiol bull doi101152ajpcell003982013 bull wwwajpcellorg
no effect on Na transport (38) whereas it caused 70reduction in Na influx in Cyprinidon variegatus hubbsi (100moll Na) and 60 reduction in goldfish (300 moll Na)(2 38) These conflicting results from various phenamil studiessuggest that sensitivity to phenamil might be species- andorenvironment-specific Collectively the pharmacological anal-ysis suggests that the Na uptake in FW rainbow trout is mostlikely not mediated by ENaC given the lack of inhibition byphenamil but may be attributed to ASIC since Na uptakewas almost completely blocked by DAPI and diminazene bothof which inhibit ASICs but not ENaC
In our Na flux study with various pharmacological agentswe made an interesting observation that inhibition of 22Na
uptake was less pronounced for all tested inhibitors during thesecond consecutive measurement (90ndash180 min) (Fig 2) Toour knowledge this phenomenon where potency of the testeddrug diminishes over time has not been reported in anyprevious study on fish This observation may be explained bya decreased stabilityefficacy of the tested pharmacologicalagents in aqueous media metabolism of the drug by theanimal or activation of compensatory Na uptake mechanismsnot sensitive to amiloride DAPI or diminazene We believethat the decrease of the potency of amiloride DAPI anddiminazene over time demonstrates the importance of properexperimental design and choice of period of study when ionfluxes are examined using pharmacological inhibitors
While the two novel ASIC inhibitors have been tested fortheir potency in inhibiting ASICs and ENaC in neurons (5)they have not been tested for their potential effects on otherNa transporters implicated in Na uptake in fishes mostnotably the NHEs To verify that the Na uptake inhibitioncaused by DAPI and diminazene in rainbow trout was due toinhibition of ASICs rather than NHEs we tested the effect ofDAPI on MRCs isolated from the rainbow trout gill epithe-lium Our laboratory previously showed that rainbow troutpossess two populations of MRCs peanut lectin agglutinin(PNA) positive (PNA) and PNA negative (PNA) MRCs(10) It has been demonstrated that NHE3 and NHE2 areexpressed in PNA cells isolated from branchial epithelium of
FW rainbow trout (19) Therefore we investigated the effect ofDAPI on PNA cells and compared it with the effect of EIPAa potent and highly selective inhibitor of NHE (25) Earlierstudies in our laboratory using cell imaging determined thatPNA cells undergo NHE-dependent alkalinization (32 33)Here we demonstrate that while EIPA was effective at inhib-iting NHE function in PNA cells (Fig 3A) there was noeffect of high concentrations of DAPI on Na-dependent pHi
elevation demonstrating that NHE was not inhibited by DAPI(Fig 3B) Therefore the measured inhibition of Na uptake byDAPI that we see in whole animals (Fig 1) cannot be attrib-uted to inhibition of NHE
On the basis of the mRNA expression of tASIC1 andtASIC4 genes in the gill epithelium and isolated MRCs local-ization of ASIC4 protein to the apical region of the MRCs and
Fig 7 Three-dimensional views of freshwater(FW) rainbow trout MRCs A scanning elec-tron micrograph of FW rainbow trout gillepithelium showing broad apical openings ofMRCs (arrowheads) and mucous cells () Band C 3-dimensional reconstruction of z-stack images showing apical localization ofASIC4 protein (red arrowhead) and basolat-eral localization of NKA (green arrow)B top view C side view
PNA- MRC
Na+
ASIC
H+
VHA
NKA
Na+
K+
NBC
Na+ HCO3-
PNA+ MRC
NH3
Na+
H+
NHE Rhcg2
NKA
Na+
K+
Fig 8 Model for Na uptake in PNA and PNA MRCs in gill epithelium ofrainbow trout showing placement of proposed ASIC-mediated Na uptakemechanisms Rhcg2 rhesus glycoprotein NHE NaH exchanger VHAvacuolar H-ATPase NBC Na-HCO3
cotransporter
C263INVOLVEMENT OF ASICs IN FISH GILL Na TRANSPORT
AJP-Cell Physiol bull doi101152ajpcell003982013 bull wwwajpcellorg
pharmacological blockade of Na influx by ASIC inhibitors inadult rainbow trout we propose a revised model for transepi-thelial Na uptake in FW rainbow trout gills (Fig 8) wherebyone apical mode of Na entry is via an ASIC4-mediatedmechanism
A number of recent studies on Na uptake in FW fishes havedemonstrated the presence of the NHERh metabolon in thegills of various species (17 30 31) As mentioned previouslywe specifically targeted the rainbow trout as a model for thisstudy as it is native to oligotrophic Na-poor mountainstreams with low buffering capacity This type of environmentpresents key thermodynamic challenges to the NHERhmetabolon as Na restrictions cannot be alleviated by thepresence of an Rh protein (6) Moreover searches of rainbowtrout genomes and transcriptomes have failed to identify anyENaC subunits ( 13 or ) despite the fact that they are knownto actively take up Na from water with 50 M Na (49)Furthermore other research studies have suggested and dem-onstrated an apical localization of the VHA in the gills ofrainbow trout (44 48) and bafilomycin a VHA-specific in-hibitor has been demonstrated to reduce Na uptake in ze-brafish (2) tilapia and carp (9) Previous studies have pro-posed VHA to be a part of the NHERh metabolon (41 50 51)however the placement of an apically oriented VHA is coun-terintuitive to the function of an NHE as it would exacerbatethe noted thermodynamic constraints Placement of a VHA inconcert with an apical ASIC is more plausible since it wouldnot only facilitate Na uptake from very low ionic strengthwater but could also provide a mechanism for gating ofASIC-mediated Na conductance It remains to be determinedif ASIC and VHA proteins colocalize to the same type ofMRCs and this warrants further investigation
Perspectives In summary we propose that ASCI4 is in-volved in mediating Na uptake from FW in rainbow troutacclimatized to low ionic strength low pH media This is thefirst demonstration of a nonneuronal function for any ASICfamily member and simultaneously provides an attractive so-lution to the debate surrounding the thermodynamic limitationsof the other proposed Na transport mechanisms and resolvesthe conundrum of the role of an apical VHA in Na transportThe possibility that other fish models may use similar mecha-nisms under different environmental conditions where modelssuch as the NHERh metabolon cannot be expected to functionremains to be explored
ACKNOWLEDGMENTS
We thank the staff in the Biosciences Aquatic Facilities the MolecularBiology Service Unit and the Cross Cancer Institute Cell Imaging Facility atthe University of Alberta for outstanding help throughout the study We alsothank Alex Clifford for advice and help with phylogenetic analysis and JamesEde for help with confocal microscopy
GRANTS
This study was supported by a Natural Sciences and Engineering ResearchCouncil of Canada Discovery Grant to G G Goss and an Alberta IngenuityStudent Scholarship to A K Dymowska
DISCLOSURES
No conflicts of interest financial or otherwise are declared by the authors
AUTHOR CONTRIBUTIONS
AKD AGS and GGG are responsible for conception and design ofthe research AKD AGS SDB and DC performed the experiments
AKD AGS SDB DC and GGG analyzed the data AKD AGSSDB DC and GGG interpreted the results of the experiments AKDAGS SDB and GGG prepared the figures AKD AGS SDB DCand GGG drafted the manuscript AKD AGS SDB DC and GGGedited and revised the manuscript AKD AGS SDB and GGGapproved the final version of the manuscript
REFERENCES
1 Avella M Bornancin M A new analysis of ammonia and sodiumtransport through the gills of the freshwater rainbow trout (Salmo gaird-neri) J Exp Biol 142 155ndash175 1989
2 Boisen AM Amstrup J Novak I Grosell M Sodium and chloridetransport in soft water and hard water acclimatized zebrafish (Daniorerio) Biochim Biophys Acta 1618 207ndash218 2003
3 Brix KV Grosell M Comparative characterization of Na transport inCyprinodon variegatus variegatus and Cyprinodon variegatus hubbsi amodel species complex for studying teleost invasion of freshwater J ExpBiol 215 1199ndash1209 2012
4 Chen X Polleichtner G Kadurin I Grunder S Zebrafish acid-sensingion channel (ASIC) 4 Characterization of homo- and heteromeric chan-nels and identification of regions important for activation by H J BiolChem 282 30406ndash30413 2007
5 Chen X Qiu L Li M Durrnagel S Orser BA Xiong ZG MacDonaldJF Diarylamidines high potency inhibitors of acid-sensing ion channelsNeuropharmacology 58 1045ndash1053 2010
6 Dymowska AK Hwang PP Goss GG Structure and function of iono-cytes in the freshwater fish gill Respir Physiol Neurobiol 184 282ndash2922012
7 Edwards SL Tse CM Toop T Immunolocalization of NHE3-likeimmunoreactivity in the gills of the rainbow trout (Oncorhynchus mykiss)and the blue-throated wrasse (Pseudolabrus tetrious) J Anat 195 465ndash469 1999
8 Evans DH Piermarini PM Choe KP The multifunctional gill domi-nant site of gas exchange osmoregulation acid-base regulation andexcretion of nitrogenous waste Physiol Rev 85 97ndash177 2005
9 Fenwick JC Bonga SE Flik G In vivo bafilomycin-sensitive Na
uptake in young freshwater fish J Exp Biol 202 3659ndash3666 199910 Galvez F Reid SD Hawkings G Goss GG Isolation and characteriza-
tion of mitochondria-rich cell types from the gill of freshwater rainbowtrout Am J Physiol Regul Integr Comp Physiol 282 R658ndashR668 2002
11 Goss G Orlowski J Grinstein S Coimmunoprecipitation of a 24-kDaprotein with NHE1 the ubiquitous isoform of the NaH exchanger AmJ Physiol Cell Physiol 270 C1493ndashC1502 1996
12 Goss GG Wood CJ Na and Cl uptake kinetics diffusive effluxes andacidic equivalent fluxes across the gills of rainbow trout 1 Responses toenvironmental hyperoxia J Exp Biol 152 521ndash547 1990
13 Grunder S Geissler HS Bassler EL Ruppersberg JP A new memberof acid-sensing ion channels from pituitary gland Neuroreport 11 1607ndash1611 2000
14 Harvey BJ Energization of sodium absorption by the H-ATPase pumpin mitochondria-rich cells of frog skin J Exp Biol 172 289ndash309 1992
15 Hirata T Kaneko T Ono T Nakazato T Furukawa N Hasegawa SWakabayashi S Shigekawa M Chang MH Romero MF Hirose SMechanism of acid adaptation of a fish living in low pH 35 lake Am JPhysiol Regul Integr Comp Physiol 284 R1199ndashR1212 2003
16 Holzer P Acid-sensitive ion channels and receptors Hand Exp Pharma-col 194 283ndash332 2009
17 Hung CY Tsui KN Wilson JM Nawata CM Wood CM Wright PARhesus glycoprotein gene expression in the mangrove killifish Kryptole-bias marmoratus exposed to elevated environmental ammonia levels andair J Exp Biol 210 2419ndash2429 2007
18 Hwang PP Lee TH Lin LY Ion regulations in fish gills recentprogresses in the cellular and molecular mechanisms Am J Physiol RegulIntegr Comp Physiol 301 R28ndashR47 2011
19 Ivanis G Esbaugh AJ Perry SF Branchial expression and localizationof SLC9A2 and SLC9A3 sodiumhydrogen exchangers and their possiblerole in acid-base regulation in freshwater rainbow trout (Oncorhynchusmykiss) J Exp Biol 211 2467ndash2477 2008
20 Jasti J Furukawa H Gonzales EB Gouaux E Structure of acid-sensingion channel 1 at 19A resolution and low pH Nature 449 316ndash323 2007
21 Jonz MG Nurse CA Epithelial mitochondria-rich cells and associatedinnervation in adult and developing zebrafish J Comp Neurol 497817ndash832 2006
C264 INVOLVEMENT OF ASICs IN FISH GILL Na TRANSPORT
AJP-Cell Physiol bull doi101152ajpcell003982013 bull wwwajpcellorg
22 Jonz MG Nurse CA New developments on gill innervation insightsfrom a model vertebrate J Exp Biol 211 2371ndash2378 2008
23 Kellenberger S Epithelial sodium and acid-sensing ion channels InSensing With Ion Channels edited by Martinac B New York Springer2008 chapt 11 p 225ndash246
24 Kellenberger S Schild L Epithelial sodium channeldegenerin family ofion channels a variety of functions for a shared structure Physiol Rev 82735ndash767 2002
25 Kleyman TR Cragoe EJ Amiloride and its analogs as tools in the studyof ion-transport J Membr Biol 105 1ndash21 1988
26 Krogh A The active absorption of ions in some freshwater animals ZVergl Physiol 25 335ndash350 1938
27 Levanti MB Guerrera MC Calavia MG Ciriaco E Montalbano GCobo J Germana A Vega JA Acid-sensing ion channel 2 (ASIC2) inthe intestine of adult zebrafish Neurosci Lett 494 24ndash28 2011
28 Lin H Randall DJ Evidence for the presence of an electrogenic protonpump on the trout gill epithelium J Exp Biol 161 119ndash134 1991
29 Maetz J Garcia Romeu F The mechanism of sodium and chlorideuptake by the gills of a fresh-water fish Carassius auratus II Evidencefor NH4
Na and HCO3Cl exchanges J Gen Physiol 47 1209ndash1227
196430 Nakada T Hoshijima K Esaki M Nagayoshi S Kawakami K Hirose
S Localization of ammonia transporter Rhcg1 in mitochondrion rich cellsof yolk sac gill and kidney of zebrafish and its ionic strength-dependentexpression Am J Physiol Regul Integr Comp Physiol 293 R1743ndashR17532007
31 Nawata CM Hung CC Tsui TK Wilson JM Wright PA Wood CMAmmonia excretion in rainbow trout (Oncorhynchus mykiss) evidence forRh glycoprotein and H-ATPase involvement Physiol Genomics 31463ndash474 2007
32 Parks SK Tresguerres M Galvez F Goss GG Intracellular pH regu-lation in isolated trout gill mitochondrion-rich (MR) cell subtypes evi-dence for NaH activity Comp Biochem Physiol A 155 139ndash1452010
33 Parks SK Tresguerres M Goss GG Interactions between Na channelsand Na-HCO3
cotransporters in the freshwater fish gill MRC a modelfor transepithelial Na uptake Am J Physiol Cell Physiol 292 C935ndashC944 2007
34 Parks SK Tresguerres M Goss GG Theoretical considerations under-lying Na uptake mechanisms in freshwater fishes Comp BiochemPhysiol C 148 411ndash418 2008
35 Paukert M Sidi S Russell C Siba M Wilson SW A family ofacid-sensing ion channels from zebrafish widespread expression in thecentral nervous system suggests a conserved role in neuronal communi-cation J Biol Chem 279 18783ndash18791 2004
36 Perry SF The chloride cell structure and function in the gill of freshwaterfish Annu Rev Physiol 59 325ndash347 1997
37 Perry SF Randall DJ Effects of amiloride and SITS on branchial ionfluxes in rainbow trout Salmo gairdneri J Exp Zool 215 225ndash228 1981
38 Preest MR Gonzales RJ Wilson RW Pharmacological examination ofNa and Cl transport in two species of freshwater fish Physiol BiochemZool 78 259ndash272 2005
39 Reid SD Hawkings GS Galvez F Goss GG Localization and charac-terization of phenamil-sensitive Na influx in isolated rainbow trout gillepithelial cells J Exp Biol 206 551ndash559 2003
40 Sakai H Lingueglia E Champigny G Mattei MG Lazdunski MCloning and functional expression of a novel degenerin-like Na channelgene in mammals J Physiol 519 323ndash333 1999
41 Shih TH Horng JL Liu ST Hwang PP Lin LY Rhcg1 and NHE3b areinvolved in ammonium-dependent sodium uptake by zebrafish larvaeacclimated to low-sodium water Am J Physiol Regul Integr Comp Physiol302 R84ndashR93 2012
42 Springauf A Grunder S An acid-sensing ion channel from shark(Squalus acanthias) mediates transient and sustained responses to protonsJ Physiol 588 809ndash820 2010
43 Spry DJ Wood CM Ion flux rates acid-base status and blood gases inrainbow trout Salmo gairdneri exposed to toxic zinc in natural soft waterCan J Fish Aquat Sci 42 1332ndash1337 1985
44 Sullivan GV Fryer JN Perry SF Immunolocalization of proton pumps(H-ATPase) in pavement cells of rainbow trout gill J Exp Biol 1982619ndash2629 1996
45 Tresguerres M Parks SK Katoh F Goss GG Microtubule-dependentrelocation of branchial V-H-ATPase to the basolateral membrane in thePacific spiny dogfish (Squalus acanthias) a role in base secretion J ExpBiol 209 599ndash609 2006
46 Vina E Parisi V Cabo R Laura R Lopez-Velasco S Lopez-Muniz AGarcia-Suarez O Germana A Vega JA Acid-sensing ion channels(ASICs) in the taste buds of adult zebrafish Neurosci Lett 536 35ndash402013
47 Waldmann R Lazdunski M H-gated cation channels neuronal acidsensors in the NaCDEG family of ion channels Curr Opin Neurobiol 8418ndash424 1998
48 Wilson JM Laurent P Tufts BL Benos DJ Donowitz M Vogl AWRandall DJ NaCl uptake by the branchial epithelium in freshwater teleostfish an immunological approach to ion-transport protein localization JExp Biol 203 2279ndash2296 2000
49 Wood CM Randall DJ Sodium balance in rainbow-trout (Salmo-gaird-neri) during extended exercise J Comp Physiol 82 235ndash256 1973
50 Wright PA Wood CM A new paradigm for ammonia excretion inaquatic animals role of rhesus (Rh) glycoproteins J Exp Biol 2122303ndash2312 2009
51 Yan JJ Chou MY Kaneko T Hwang PP Gene expression of NaH
exchanger in zebrafish H-ATPase-rich cells during acclimation to low-Na and acidic environments Am J Physiol Cell Physiol 293 C1814ndashC1823 2007
C265INVOLVEMENT OF ASICs IN FISH GILL Na TRANSPORT
AJP-Cell Physiol bull doi101152ajpcell003982013 bull wwwajpcellorg

The ratios of fluorescence at 495 nm to fluorescence at 440 nm at eachstep of the pH calibration were used to form a regression equation foreach individual cell trace This equation was extrapolated to the rawratio data obtained over the entire time course yielding a calibratedpHi for each individual cell monitored during the perfusion experi-ment Data were collected and analyzed for each calibrated individualcell by comparing the rate of alkalinization (pHit) under thecontrol parameters with that in the presence of the drug to determinepercent inhibition
Preparation of total RNA Total RNA was extracted from frozentissues using TRIzol reagent (Invitrogen Carlsbad CA) according tothe manufacturerrsquos instructions RNA was treated with DNase I(Ambion Austin TX) according to the manufacturerrsquos protocol thenan on-column cleanup was carried out using an RNeasy Mini Kit(Qiagen Mississauga ON Canada) The quality of the extractedRNA was assessed by a bioanalyzer (model 2100 Agilent Technol-ogies Palo Alto CA) or by visualization on a formaldehyde gel RNAconcentration was measured with a spectrophotometer (ND-1000UV-Vis NanoDrop Technologies Rockland DE)
Molecular cloning and phylogenetic analysis Rapid amplificationof cDNA ends (RACE)-ready cDNA was synthesized from gill RNAusing a SMARTer RACE cDNA amplification kit (Clontech Moun-tain View CA) according to the manufacturerrsquos instructions cDNAsolutions were diluted 110 with tricine-EDTA buffer provided in thekit and stored at 20degC
Oligonucleotides for cloning of ASIC1 and ASIC4 genes weredesigned with PrimerQuest (Integrated DNA Technology) (Table 1)on the basis of conserved regions in available ASIC mRNA sequencesfrom zebrafish tilapia pufferfish stickleback and cod (NationalCenter for Biotechnology Information Ensembl) Sequences werealigned with GeneDoc (version 2602 httpwwwpscedubiomedgenedoc) and ClustalX (version 181) software DNA sequencesencoding the 5=- and 3=-termini of the ASIC1 gene and the 5=-terminus of the ASIC4 gene were obtained using the Advantage 2PCR kit (Clonetech) with RACE-ready cDNA templates generatedfrom gill RNA The ASIC4 3=-terminus was obtained using degener-ate primer (Table 1) Resulting PCR products were excised from thegel and purified with a QIAquick gel extraction kit (Qiagen) Subse-quently amplicons of interest were sequenced directly or ligated intopJET12 vector using a CloneJET kit (Thermo Scientific) for verifi-cation Recombinant plasmids were transformed into competent cells
(One Shot TOP10 chemically competent Escherichia coli Invitro-gen) Colonies were screened for plasmids containing inserts of thecorrect size by agarose gel electrophoresis and sequenced withpJET12-specific primers provided in the CloneJET kit
The amino acid sequences derived from our cloned trout ASIC4and ASIC1 genes (accession numbers KF964646 and KF964645respectively) together with the previously annotated amino acidsequences of ASIC1 ASIC2 ASIC3 and ASIC4 from various fishspecies and other taxa were aligned and analyzed using Seaviewsoftware (version 4 httppbiluniv-lyon1frsoftwareseaview) Arooted phylogenetic tree showing evolutionary relationships betweenthe different ASIC proteins was generated using the maximum-likelihood method and LG model and 700 bootstrap resamplings
RT-PCR Tissue-specific cDNA templates utilized for RT-PCRwere synthesized from 3 g of RNA from gill MRCs kidney brainor blood using SuperScript III reverse transcriptase (Invitrogen) ac-cording to protocols provided by the manufacturer ASIC4 and ASIC1amplicons were obtained using primers designed with PrimerQuest(Table 1) PCR conditions were as follows 98degC for 1 min of initialdenaturation followed by 35 cycles of denaturation at 98degC for 10 sannealing at 62degC for 30 s and elongation at 72degC for 40 s with thefinal elongation at 72degC for 10 min PCR products were visualized by1 agarose gel electrophoresis followed by ethidium bromide stain-ing
Immunoprecipitation of ASIC4 and Western blot analysis Ananti-zebrafish ASIC4b (zASIC42) polyclonal antibody against twopeptides corresponding to regions near the NH2 terminus (aa 146ndash160 PKSRKGHRPSELQYP) and the COOH terminus (aa 519ndash533CFEEVKVKAANDVAQP) of zASIC42 protein (accession noQ708S31) was custom-made by 21st Century Biochemicals (Marl-boro MA) The polyserum was affinity-purified against each of thesepeptides independently and the NH2-terminal peptide-purified anti-body was used exclusively in this study Immunoprecipitation wasperformed according to the technique described by Goss and col-leagues (11) Briefly trout gills were perfused with heparinizedice-cold PBS (Ca2-free) and the second and third gill arches weredissected out and washed three times in ice-cold PBS Gill rakers wereremoved and the remaining gill arches were cut into 200- to 300-mgsections Each section was lysed for 30 min in 2ndash3 ml of lysis buffer(100 mM NaCl 4 mM KCl 50 mM HEPES 5 mM EDTA pH 74and 1 Triton-X) containing protease inhibitors (cOmplete MiniEDTA-free protease inhibitor tablets Roche Diagnostics MannheimGermany) After centrifugation to remove the debris (5000 g 5 min4degC) the lysates were incubated with 2ndash4 l of anti-zASIC42antibody at 4degC overnight on an end-over-end rotator On the nextday 60 l of protein A-Sepharose CL4b beads (Sigma Chemical StLouis MO) preswelled in immunoprecipitation (IP) buffer (in mM100 NaCl 4 KCl 50 HEPES and 5 EDTA pH 74) and preblockedwith 1 bovine serum albumin (fraction V Sigma) were added to thelysates which were incubated at 4degC for 4ndash6 h on the rotator Afterthe incubation the supernatant was removed and the beads werewashed three times with 1 ml of IP buffer After the washes the beadswere incubated in Laemmli buffer for 15 min at 65degC and centrifugedand the supernatant was retained for Western blot analysis as de-scribed previously (45) Briefly the samples were separated on a 75polyacrylamide minigel and transferred to a nitrocellulose (NC) mem-brane using a wet transfer system (Bio-Rad Laboratories HerculesCA) The NC membrane was blocked in 5 bovine serum albumin inTris-buffered saline with 02 Triton X-100 (TBST) for 30 min andincubated with rabbit anti-zASIC42 antibody (11000 dilution) on arocker at 4degC overnight The membrane was washed four times for 15min each with TBST blocked again with 5 bovine serum albuminfor 15 min and then incubated with a secondary horseradish peroxi-dase-conjugated goat anti-rabbit antibody (Santa Cruz BiotechnologyDallas TX) at 150000 dilution at room temperature for 1 h The NCmembrane was washed four times in TBST and the immunoreactivebands were visualized using a SuperSignal West Pico chemilumines-
Table 1 Primer sets for cloning and tissue distributionof ASICs
Gene Primer Sequence
CloningASIC1
5= 5=-TCACCCAGCAACCCTGCGAACTCGT-3=3= 5=-ACACCCTGCAACATGACGCGCT-3=
ASIC45= 5=-GATGTCCCGTCCGGTTGAAGATGTC-3=Forward 5=-CCCAGTAACATCAAGTGTGTCG-3=Reverse 5=-YYARCANGCRAARTCYTC-3=
RT-PCRASIC1
Forward 5=-AAGTCCACTCCCATAGA-3=Reverse 5=-CAGCCAGGTTATTCCTT-3=
ASIC4Forward 5=-CTTTCGTTTCTCTGCTCTCACC-3=Reverse 5=-CAAGACCAGGAAGTTGTCTCTG-3=
GAPDHForward 5=-AAGGGTGAGGTGAGCATGGA-3=Reverse 5=-GCTTTACCCCATGGGATCTCAT-3=
ASIC acid-sensing ion channel 5= 5=-rapid amplification of cDNA ends(RACE) primer 3= 3=RACE primer Y C T R A G N A G T C
C257INVOLVEMENT OF ASICs IN FISH GILL Na TRANSPORT
AJP-Cell Physiol bull doi101152ajpcell003982013 bull wwwajpcellorg
cent substrate kit (Thermo Scientific) following the manufacturerrsquosinstructions
SEM and immunohistochemistry For examination of the surface ofthe gill by SEM filaments were fixed in 3 glutaraldehyde and 2paraformaldehyde in 015 mM sodium cacodylate buffer (pH 74 290mosM) for 1 h at 4degC dehydrated in a graded series of ethanol andcritical point dried Samples were then mounted on an SEM stubsputter-coated with gold and examined with a scanning electronmicroscope (model JSM-6301FXV Jeol)
For immunohistochemistry gills were excised from adult rain-bow trout fixed in 4 paraformaldehyde in PBS (pH 74) over-night at 4degC dehydrated in a graded series of ethanol andembedded into paraffin blocks Serial sections (5 m) were cut andrehydrated in a decreasing ethanol series followed by double-distilled H2O Rehydrated sections were incubated in 10 mMcitrate buffer at 70degC for 1 h for epitope retrieval After incubationslides were washed in PBS and then blocked with 6 milk powderin a humidified chamber for 1 h Sections were then incubatedovernight at 4degC simultaneously with the anti-zASIC42 antibody(1400 dilution) and anti-Na-K-ATPase (NKA) monoclonalantibody (1400 dilution anti-chicken 5-subunit DevelopmentalStudies Hybridoma Bank University of Iowa) followed by sec-ondary FITC-conjugated anti-mouse antibody (1500 dilution In-vitrogen Oregon) and tetramethylrhodamine-conjugated anti-rab-bit antibody (1500 dilution Invitrogen Oregon) Slides incubatedwith preimmune serum from the anti-zASIC42 antibody produc-tion in place of primary antibody were used as a negative controlSlides were viewed with a laser scanning confocal microscope(model LSM 710 Zeiss Germany) at the Cross Cancer InstituteCell Imaging Facility (Edmonton AB Canada) Images wereprocessed with LSM Image Browser (version 420121 CarlZeiss) Surface rendering and three-dimensional reconstruction ofz-stack images were performed using Imaris software (version622 Bitplane Zurich Switzerland)
Statistical analysis Na flux data were subjected to statisticalanalysis and are reported as means SE All pharmacologicalexperiment data sets were tested for homogeneity of variance andcompared using one-way ANOVA (SigmaPlot version 11 SystatChicago IL) If significant differences (P 005) were found a posthoc multiple-comparisons Tukeyrsquos test was applied to determine thesedifferences A paired t-test (P 005) was used in the pHi imagingexperiments to compare the relative inhibition of pHit in isolatedcells under control conditions and after addition of each pharmaco-logical inhibitor
RESULTS
Pharmacological inhibition Exposure of juvenile rainbowtrout to increasing concentrations of DAPI resulted in a con-centration-dependent decrease in Na uptake with 90inhibition at 1 moll (Fig 1A) The DAPI concentration atwhich there was a 50 inhibition in Na uptake (IC50) wascalculated to be 012 moll Exposure to diminazene alsoresulted in a concentration-dependent decrease in Na uptakein juvenile trout with a maximum reduction of Na uptakefrom 303 43 (control) to 51 19 molmiddotkg1middoth1 (79)after exposure to 30 moll diminazene (Fig 1B) The IC50 fordiminazene was calculated to be 096 moll
Na uptake rates in juvenile rainbow trout were significantlylower in the first 90-min flux time period after exposure to 500M amiloride (17 185 molmiddotkg1middoth1) 1 M DAPI (29 189 molmiddotkg1middoth1) and 3 M diminazene (41 334molmiddotkg1middoth1) than in controls (539 1709 molmiddotkg1middoth1 Fig 2) while 50 M phenamil (299 1079 molmiddotkg1middoth1) did not significantly reduce Na uptake Howeverfor the second consecutive flux period between 90 and 180min of exposure to the pharmacological agents the inhibitionwas less pronounced but still significant for amiloride (104 334 molmiddotkg1middoth1) and DAPI (141 627 molmiddotkg1middoth1)while diminazene (297 415 molmiddotkg1middoth1) was no longersignificantly different from control (393 441 molmiddotkg1middoth1 Fig 2)
MRC pHi imaging Results presented from the whole-animalflux experiments showed that administration of DAPI at vari-ous concentrations significantly inhibited Na uptake (Fig1A) To demonstrate that the inhibition of Na uptake attrib-uted to DAPI was in fact due to the blocking of ASIC-type Na
channels rather than affecting NHE activity we conductedexperiments using pHi imaging NHE activity was measuredby examining the rate of pHi alkalization (pHit) uponexposure of MRCs to a Na-containing medium (145 mM) inthe absence or presence of DAPI (1 M) or EIPA (100 M)
A representative trace (Fig 3A) shows a Na-dependentalkalization following Na application (1st rise) while addi-tion of EIPA significantly decreased the rate of alkalization(2nd rise) The combined results (n 19) show a control of
0
50
100
150
200
250
300
350
400
450
0 0001 001 01 1 10
JNa I
N(μ
mol
kg-1
h-1 )
DAPI concentration (μmol l-1)
0
50
100
150
200
250
300
350
400
0 00003 0003 003 03 3
JNa I
N(μ
mol
k g-1
h-1 )
Diminazene concentration (μmol l-1)
A B
Fig 1 Dose-dependent decreases in Na uptake rates (JNain) in juvenile trout after exposure to increasing concentrations of 4=6-diamidino-2-phenylindole(DAPI) and diminazene in low-Na (30 M) and low-pH (pH 60) water Values are means SE (n 6) P 005 vs control
C258 INVOLVEMENT OF ASICs IN FISH GILL Na TRANSPORT
AJP-Cell Physiol bull doi101152ajpcell003982013 bull wwwajpcellorg
0642 0040 pHimin while the alkalization rate wasreduced by 62 in the presence of EIPA (0 240 0036pHimin P 0001 Fig 3B) For DAPI Na-dependentalkalization following Na application was similar in the
presence or absence of DAPI (Fig 3D) No differences inpHit were noted between control (0542 0042 pHimin) and DAPI-treated (0578 0047 pHimin) cells(P 0149 n 22 Fig 3D)
0
100
200
300
400
500
600
700
800
Cont Amil DAPI Dim Phen Cont Amil DAPI Dim Phen
JNa I
N(μ
mol
kg-1
h-1 )
Treatment
T0-90min T90-180min
Fig 2 Effects of pharmacological agents [control (Cont DMSO) amiloride (Amil 500 moll) DAPI (1 moll) diminazene (Dim 3 moll) and phenamil(Phen 50 moll)] during consecutive 90-min measurements of Na uptake in juvenile trout in low-Na (30 M) and low-pH (pH 60) water Values are means SE(n 6) P 005 vs control
65
7
75
8
85
9
0 100 200 300 400 500 600
pHi
Time (sec)
Na FreeNa Free Na
EIPA
Na
0
01
02
03
04
05
06
07
08
Na+ Na+EIPA
ΔpH
iΔM
in
Cell Alkalinization Treatments
65
7
75
8
85
9
0 100 200 300 400 500 600
pHi
Time (sec)
Na Free Na
DAPI
Na Na Free
0
01
02
03
04
05
06
07
08
Na+ Na+DAPI
ΔpH
iΔM
in
Cell Alkalinization Treatments
A
DC
B
Fig 3 Effect of ethylisopropylamiloride (EIPA) and DAPI on Na-induced alkalinization of peptide nucleic acid-positive (PNA) cells following Na-freeexposure at resting intracellular pH (pHi) A representative trace demonstrating inhibition of cell alkalinization by EIPA (100 M) B quantitative analysis ofcell alkalinization in the absence and presence of EIPA (62 inhibition) Values are means SE (n 19 cells) P 0001 (by paired t-test) C representativetrace demonstrating lack of effect on cell alkalinization in the presence of DAPI (1 M) D quantitative analysis of cell alkalinization in the absence and presenceof DAPI Values are means SE (n 22) P 005 (by paired t-test)
C259INVOLVEMENT OF ASICs IN FISH GILL Na TRANSPORT
AJP-Cell Physiol bull doi101152ajpcell003982013 bull wwwajpcellorg
Phylogenetic analysis and tissue distribution of ASIC1 andASIC4 Two distinct DNA fragments whose derived aminoacid sequences exhibit similarity to the ASIC family werecloned from the gill of rainbow trout To determine theiridentity we conducted a phylogenetic analysis by comparisonwith known ASIC homologs from a variety of species Thegenerated phylogenetic tree (Fig 4) showed that the twocloned rainbow trout ASIC genes (tASICs) were members ofthe ASIC1 and ASIC4 subunits respectively (Fig 4A) ThetASIC1 (partial clone) grouped most closely with ASIC1 fromsalmon (Salmo salar) while tASIC4 grouped with other iden-
tified ASIC4 family members from other fish and mammalspecies
Furthermore we conducted a tissue distribution analysis oftASIC1 and tASIC4 by RT-PCR We observed mRNA expres-sion of tASIC1 in all tested tissues gill MRCs head and trunkkidney brain and blood tASIC4 had a slightly differentdistribution it was present in the perfused gill MRCs brainand trunk kidney but it was absent in head kidney and bloodtissues
Immunoprecipitation and immunostaining with anti-zASIC42To determine if ASIC4 was present in the fish gill and to validate our
Fig 4 Phylogenetic analysis of amino acid sequences of cloned rainbow trout acid-sensing ion channels ASIC1 and ASIC4 and ASICs from various fish speciesPhylogenetic tree was constructed using the maximum-likelihood method and numbers indicate bootstrap values for 700 resamplings GenBank or Ensemblaccession numbers are as follows human ASIC2 NP_0010852 Western clawed frog ASIC2 XP_0029330862 Japanese pufferfish ASIC2 XP_0039648711zebrafish ASIC2 NP_9999531 three-spined stickleback ASIC2 ENSGACG00000009558 Atlantic cod ASIC2 ENSGMOT00000019145 spotted greenpufferfish ASIC2 ENSTNIG00000014943 Southern platyfish ASIC2 ENSXMAG00000008283 oyster toadfish ASIC2 AAP456331 coelacanth ASIC2ENSLACG00000007191 rat ASIC2 NP_0370242 Southern platyfish ASIC1c ENSXMAG00000018841 African clawed frog ASIC11 AEC486651 Africanclawed frog ASIC12 AEC486671 brown rat ASIC1 NP_0770681 human ASIC1 NP_0644232 three-spined stickleback ASIC1a ENSGACG00000003075Japanese pufferfish ASIC1 XP_0039635201 Southern platyfish ASIC1a XP_0058018011 Atlantic cod ASIC1a ENSGMOG00000009659 zebrafishASIC12 NP_9999551 rainbow trout ASIC1 KF964646 Atlantic salmon ASIC1 NP_0011334561 oyster toadfish ASIC1 Q7T1N41 elephant shark ASIC1AEC486681 three-spined stickleback AIC1c ENSGACG00000017974 Atlantic cod ASIC1c ENSGMOG00000003105 zebrafish ASIC13 NP_9999541river lamprey ASIC1 AAY289831 sea lamprey ASIC1b ENSPMAG00000003813 dogfish shark ASIC1 AAY289851 zebrafish ASIC11 NP_9999561human ASIC4 NP_8782672 rat ASIC4 NP_0715702 African clawed frog ASIC4 AEC486661 Japanese pufferfish ASIC4 XP_0039621271 spotted greenpufferfish ASIC4b ENSTNIG00000010994 Nile tilapia ASIC4 XP_0034454461 Southern platyfish ASIC4a ENSXMAG00000003624 three-spinedstickleback ASIC4a ENSGACG00000007052 Atlantic cod ASIC4a ENSGMOG00000012740 rainbow trout ASIC4 KF964645 zebrafish ASIC41NP_9999521 zebrafish ASIC42 NP_9999511
C260 INVOLVEMENT OF ASICs IN FISH GILL Na TRANSPORT
AJP-Cell Physiol bull doi101152ajpcell003982013 bull wwwajpcellorg
antibody for later immunocytochemistry we performed an im-munoprecipitation using an antibody raised against zASIC42on the basis of sequence similarly of 60 between zASIC42(aa 146 ndash160 PKSRKGHRPSELQYPP) and the clonedtASIC4 (aa 142ndash156 PKNREGHKPTDLDYPA) Using West-ern blotting we analyzed the precipitates for the presence ofASIC4 (see Fig 6A) and found a single band of 65 kDawhich approximates the predicted size for tASIC4 (604 kDa)The strong band at 55 kDa corresponds to the IgG heavychain
To determine the location of ASIC4 protein within therainbow trout gill tissue fixed sections were double-labeledwith anti-zASIC42 antibody (see Fig 6B) and anti-NKA andthen counterstained with the nuclear stain DAPI Immunore-activity was observed in cells located in the lamellae andinterlamellar region (see Fig 6B arrowhead) Moreover themajority of cells positive for zASIC42 also colocalized withNKA protein (Fig 6 C and D arrowhead) a known marker ofMRCs suggesting that in gill epithelium ASIC4 protein ismainly expressed in MRCs SEM analysis of the gill filamentdemonstrated that MRCs are clearly distinguished on the sur-face of the epithelium and have a broad apical convex openingto the environment (see Fig 7A arrowheads) The generatedthree-dimensional reconstructions of the staining immunoreac-tivity pattern for ASIC4 and NKA proteins revealed the pres-ence of ASIC4 and NKA in the same cells but in distinctregions of the cells NKA was present on the basolateralmembrane (arrows green staining) while ASIC4 was localizedto the broad apical surface of the cell (arrowheads red stain-ing) Figure 7B shows a generated image with a top view whileFig 7C is the same cell viewed from the side
DISCUSSION
The existence of an apical Na channel electrochemicallycoupled to a VHA mediating Na uptake from dilute FW hasbeen postulated on theoretical thermodynamic grounds for anumber of decades (1 36 34) However an inability todetermine the molecular identity of the Na channel has beena major drawback in the general acceptance of the modelUsing a combination of pharmacological blockade isotopicfluxes immunocytochemistry and molecular biology we pro-pose that an ASIC family member is potentially the long-sought-after apical Na channel responsible for Na uptake byrainbow trout acclimatized to water with low Na and low pHOur study provides the first evidence for a nonneuronal role forASICs in any vertebrate whereby in gills from FW fishesASICs are associated with the MRCs and tASIC4 localizes tothe apical membrane of NKA positive cells
ASICs are a broad family of Na-specific ion channels thathave been extensively studied in mammalian models (forreview see Ref 23) with only a few studies performed in anyfish species (35 42 46) ASICs have primarily been associatedwith neuronal tissues with studies focusing on a neuronalfunction for ASICs (23) In this study ASIC1 and ASIC4 werecloned in rainbow trout using PCR and they were expressed ina variety of tissues both neuronal and nonneuronal in origin(Fig 5) Importantly tASIC1 gene was present in all fiveinvestigated tissues including whole blood It appears that thetASIC4 gene has a more defined tissue distribution thantASIC1 with expression in the ionoregulatory tissues (trunk
kidney and perfused gill) and the brain Unlike tASIC1 therewas no apparent expression of tASIC4 in the head kidney orthe blood Therefore tASIC4 expression in MRCs and per-fused gill could not be attributed to blood contamination
Both tASIC1 and tASIC4 genes were expressed in the gillepithelium and more specifically were present in MRCs ofrainbow trout gills It is believed that similar to ENaCsfunctional ASICs are trimers (20) although their configurationhas not been resolved It has been demonstrated that with afew exceptions ASICs function as homomers (forming chan-nels of identical subunits) or heteromers (forming channelswith other ASIC subunits) (4 23) ASICs have been charac-terized by using oocyte expression in zebrafish (4 35) anddogfish shark (ASIC1b only) (42) It has been found thatcoexpression of zASIC41 with zASIC13 in Xenopus oocytesincreased the current amplitude and abundance (15-fold) ofthe channel at the cell surface indicating that these two ASICsubunits form a functional channel (4) Moreover thisASIC4113 heteromeric channel was more efficiently traf-ficked to the plasma membrane and had increased affinity forH Additionally zASIC41 when heterologously expressedin Xenopus oocytes has been shown to be also gated open bydecreases in extracellular Ca2 The signal responsible forgating of zASIC42 was unable to be determined in that study(4) It is therefore possible that tASIC1 and tASIC4 form aheteromeric channel in MRCs but this cannot be determined atthis time and would require confirmation by immunohisto-chemistry with an anti-ASIC1 antibody that is immunoreactivefor rainbow trout It is also possible that other ASIC subunitsandor regulatory elements may be involved in the function ofASICs in the FW trout gill A full characterization of tASIC4by oocyte expression and electrophysiology is required tounderstand the mechanisms of gating in low-Na low-Ca2
watersIn our study ASIC4 protein was colocalized with NKA and
was present on the apical surface of cells suggesting that it isfound primarily in MRCs (Fig 6 D and E) Previous studies onASICs in fishes have suggested that they are primarily associ-ated with neuronal tissue and sensory cells (27 35 42 46) Inadult and developing zebrafish morphological studies usingconfocal immunofluorescence techniques demonstrated thatgill MRCs come into direct contact with nerve fibers in thebasolateral region (21 22) However in our study ASIC4protein was clearly localized to the apical region of the MRCsas demonstrated by three-dimensional reconstruction of con-
Head k
idney
ASIC4
ASIC1
GAPDH
Gill MRCs
Brain
Trunk k
idney
Blood
No tem
plate
Fig 5 Tissue distribution analysis of ASIC4 and ASIC1 genes in adultrainbow trout as determined by PCR GAPDH was used as a reference geneMRCs mitochondrion-rich cells
C261INVOLVEMENT OF ASICs IN FISH GILL Na TRANSPORT
AJP-Cell Physiol bull doi101152ajpcell003982013 bull wwwajpcellorg
focal images ASIC4 immunoreactivity did not overlap withthe standard basolateral immunoreactive pattern associatedwith NKA (Fig 7 B and C) Therefore we conclude thatneuronally derived tissues cannot account for the expression ofASIC4 protein in MRCs We also corroborate the ASIC4staining pattern with the MRC external morphology whichshows that in FW rainbow trout raised in Edmonton tap water(500 M Na) the MRCs have wide apical openings (Fig7A) This apical localization of ASIC4 protein in the cells of atransporting epithelium is the first evidence for a nonneuronalfunction for any ASIC member and suggests that ASIC4 as aNa channel may be responsible for apical uptake of Na inFW fish gill
Pharmacological profiling has long been used to differentiatethe mechanisms of ion transport across epithelial tissues For Na
transport classical analysis has usually involved amiloride and itssubstituted derivatives (eg EIPA mercaptoisobutyric acid andphenamil) as a means to differentiate between the involvementof NHE isoforms and ENaC in Na transport (25) We inves-tigated two newly described ASIC-specific inhibitors dimina-zene and DAPI (5) for their potency in inhibiting unidirec-tional Na uptake in fish Diminazene and DAPI exhibited adose-dependent inhibitory relationship and both were veryeffective at low concentrations (Fig 1) Na uptake in rainbow
trout acclimatized to low-ionic-strength acidic water wasclearly inhibited by these diarylamidines at concentrationssimilar to those reported in a previous study of cultured rathippocampal neurons (5) However in our study DAPI provedto be more effective than diminazene in contrast to observa-tions in rat neurons The inhibition of 22Na uptake by DAPIand diminazene was also compared with other common Na
transport inhibitors amiloride and phenamil Amiloride wasobserved to be an effective inhibitor of Na uptake in rainbowtrout but only at relatively high concentrations (500 M)which is consistent with previous studies (2 37) Recentlyamiloride was also demonstrated to completely block ASIC-mediated currents (5 35 42) however it is also known toblock many other Na transporters including NHE and ENaC(25) Amiloride therefore cannot be used as a distinguishingpharmacological agent
While DAPI diminazene and amiloride almost completelyblocked Na transport in the present study phenamil had nosignificant effect on Na flux in rainbow trout This suggeststhat phenamil is a poor inhibitor of Na uptake in troutexposed to low-ionic-strength water (30 moll Na) Paststudies of the effects of phenamil on Na uptake in a variety ofFW fishes have reported inconsistent results For example inneon tetras acclimatized to low Na (50 moll) phenamil had
Fig 6 Immunoreactivity of anti-zebrafish ASIC42 (zASIC42) antibody in the gill of rainbow trout A immunoprecipitation Western blot from whole gillhomogenates with zASIC42 showing a distinct band of 65 kDa Lane 1 (L1) contains 300 mg of gill tissue and lane 2 (L2) contains 200 mg of gill tissueBndashE confocal images of gill sections labeled for ASIC4 and Na-K-ATPase (NKA) proteins B and C double-labeling with DAPI and anti-zASIC42 (B) andDAPI and anti-NKA (C) D merged image of B and C E higher-magnification image of a MRC showing that ASIC4 and NKA are present in distinct regionswithin the MRC F control micrograph with DAPI and no primary antibody Arrowheads (BndashD) indicate positive staining
C262 INVOLVEMENT OF ASICs IN FISH GILL Na TRANSPORT
AJP-Cell Physiol bull doi101152ajpcell003982013 bull wwwajpcellorg
no effect on Na transport (38) whereas it caused 70reduction in Na influx in Cyprinidon variegatus hubbsi (100moll Na) and 60 reduction in goldfish (300 moll Na)(2 38) These conflicting results from various phenamil studiessuggest that sensitivity to phenamil might be species- andorenvironment-specific Collectively the pharmacological anal-ysis suggests that the Na uptake in FW rainbow trout is mostlikely not mediated by ENaC given the lack of inhibition byphenamil but may be attributed to ASIC since Na uptakewas almost completely blocked by DAPI and diminazene bothof which inhibit ASICs but not ENaC
In our Na flux study with various pharmacological agentswe made an interesting observation that inhibition of 22Na
uptake was less pronounced for all tested inhibitors during thesecond consecutive measurement (90ndash180 min) (Fig 2) Toour knowledge this phenomenon where potency of the testeddrug diminishes over time has not been reported in anyprevious study on fish This observation may be explained bya decreased stabilityefficacy of the tested pharmacologicalagents in aqueous media metabolism of the drug by theanimal or activation of compensatory Na uptake mechanismsnot sensitive to amiloride DAPI or diminazene We believethat the decrease of the potency of amiloride DAPI anddiminazene over time demonstrates the importance of properexperimental design and choice of period of study when ionfluxes are examined using pharmacological inhibitors
While the two novel ASIC inhibitors have been tested fortheir potency in inhibiting ASICs and ENaC in neurons (5)they have not been tested for their potential effects on otherNa transporters implicated in Na uptake in fishes mostnotably the NHEs To verify that the Na uptake inhibitioncaused by DAPI and diminazene in rainbow trout was due toinhibition of ASICs rather than NHEs we tested the effect ofDAPI on MRCs isolated from the rainbow trout gill epithe-lium Our laboratory previously showed that rainbow troutpossess two populations of MRCs peanut lectin agglutinin(PNA) positive (PNA) and PNA negative (PNA) MRCs(10) It has been demonstrated that NHE3 and NHE2 areexpressed in PNA cells isolated from branchial epithelium of
FW rainbow trout (19) Therefore we investigated the effect ofDAPI on PNA cells and compared it with the effect of EIPAa potent and highly selective inhibitor of NHE (25) Earlierstudies in our laboratory using cell imaging determined thatPNA cells undergo NHE-dependent alkalinization (32 33)Here we demonstrate that while EIPA was effective at inhib-iting NHE function in PNA cells (Fig 3A) there was noeffect of high concentrations of DAPI on Na-dependent pHi
elevation demonstrating that NHE was not inhibited by DAPI(Fig 3B) Therefore the measured inhibition of Na uptake byDAPI that we see in whole animals (Fig 1) cannot be attrib-uted to inhibition of NHE
On the basis of the mRNA expression of tASIC1 andtASIC4 genes in the gill epithelium and isolated MRCs local-ization of ASIC4 protein to the apical region of the MRCs and
Fig 7 Three-dimensional views of freshwater(FW) rainbow trout MRCs A scanning elec-tron micrograph of FW rainbow trout gillepithelium showing broad apical openings ofMRCs (arrowheads) and mucous cells () Band C 3-dimensional reconstruction of z-stack images showing apical localization ofASIC4 protein (red arrowhead) and basolat-eral localization of NKA (green arrow)B top view C side view
PNA- MRC
Na+
ASIC
H+
VHA
NKA
Na+
K+
NBC
Na+ HCO3-
PNA+ MRC
NH3
Na+
H+
NHE Rhcg2
NKA
Na+
K+
Fig 8 Model for Na uptake in PNA and PNA MRCs in gill epithelium ofrainbow trout showing placement of proposed ASIC-mediated Na uptakemechanisms Rhcg2 rhesus glycoprotein NHE NaH exchanger VHAvacuolar H-ATPase NBC Na-HCO3
cotransporter
C263INVOLVEMENT OF ASICs IN FISH GILL Na TRANSPORT
AJP-Cell Physiol bull doi101152ajpcell003982013 bull wwwajpcellorg
pharmacological blockade of Na influx by ASIC inhibitors inadult rainbow trout we propose a revised model for transepi-thelial Na uptake in FW rainbow trout gills (Fig 8) wherebyone apical mode of Na entry is via an ASIC4-mediatedmechanism
A number of recent studies on Na uptake in FW fishes havedemonstrated the presence of the NHERh metabolon in thegills of various species (17 30 31) As mentioned previouslywe specifically targeted the rainbow trout as a model for thisstudy as it is native to oligotrophic Na-poor mountainstreams with low buffering capacity This type of environmentpresents key thermodynamic challenges to the NHERhmetabolon as Na restrictions cannot be alleviated by thepresence of an Rh protein (6) Moreover searches of rainbowtrout genomes and transcriptomes have failed to identify anyENaC subunits ( 13 or ) despite the fact that they are knownto actively take up Na from water with 50 M Na (49)Furthermore other research studies have suggested and dem-onstrated an apical localization of the VHA in the gills ofrainbow trout (44 48) and bafilomycin a VHA-specific in-hibitor has been demonstrated to reduce Na uptake in ze-brafish (2) tilapia and carp (9) Previous studies have pro-posed VHA to be a part of the NHERh metabolon (41 50 51)however the placement of an apically oriented VHA is coun-terintuitive to the function of an NHE as it would exacerbatethe noted thermodynamic constraints Placement of a VHA inconcert with an apical ASIC is more plausible since it wouldnot only facilitate Na uptake from very low ionic strengthwater but could also provide a mechanism for gating ofASIC-mediated Na conductance It remains to be determinedif ASIC and VHA proteins colocalize to the same type ofMRCs and this warrants further investigation
Perspectives In summary we propose that ASCI4 is in-volved in mediating Na uptake from FW in rainbow troutacclimatized to low ionic strength low pH media This is thefirst demonstration of a nonneuronal function for any ASICfamily member and simultaneously provides an attractive so-lution to the debate surrounding the thermodynamic limitationsof the other proposed Na transport mechanisms and resolvesthe conundrum of the role of an apical VHA in Na transportThe possibility that other fish models may use similar mecha-nisms under different environmental conditions where modelssuch as the NHERh metabolon cannot be expected to functionremains to be explored
ACKNOWLEDGMENTS
We thank the staff in the Biosciences Aquatic Facilities the MolecularBiology Service Unit and the Cross Cancer Institute Cell Imaging Facility atthe University of Alberta for outstanding help throughout the study We alsothank Alex Clifford for advice and help with phylogenetic analysis and JamesEde for help with confocal microscopy
GRANTS
This study was supported by a Natural Sciences and Engineering ResearchCouncil of Canada Discovery Grant to G G Goss and an Alberta IngenuityStudent Scholarship to A K Dymowska
DISCLOSURES
No conflicts of interest financial or otherwise are declared by the authors
AUTHOR CONTRIBUTIONS
AKD AGS and GGG are responsible for conception and design ofthe research AKD AGS SDB and DC performed the experiments
AKD AGS SDB DC and GGG analyzed the data AKD AGSSDB DC and GGG interpreted the results of the experiments AKDAGS SDB and GGG prepared the figures AKD AGS SDB DCand GGG drafted the manuscript AKD AGS SDB DC and GGGedited and revised the manuscript AKD AGS SDB and GGGapproved the final version of the manuscript
REFERENCES
1 Avella M Bornancin M A new analysis of ammonia and sodiumtransport through the gills of the freshwater rainbow trout (Salmo gaird-neri) J Exp Biol 142 155ndash175 1989
2 Boisen AM Amstrup J Novak I Grosell M Sodium and chloridetransport in soft water and hard water acclimatized zebrafish (Daniorerio) Biochim Biophys Acta 1618 207ndash218 2003
3 Brix KV Grosell M Comparative characterization of Na transport inCyprinodon variegatus variegatus and Cyprinodon variegatus hubbsi amodel species complex for studying teleost invasion of freshwater J ExpBiol 215 1199ndash1209 2012
4 Chen X Polleichtner G Kadurin I Grunder S Zebrafish acid-sensingion channel (ASIC) 4 Characterization of homo- and heteromeric chan-nels and identification of regions important for activation by H J BiolChem 282 30406ndash30413 2007
5 Chen X Qiu L Li M Durrnagel S Orser BA Xiong ZG MacDonaldJF Diarylamidines high potency inhibitors of acid-sensing ion channelsNeuropharmacology 58 1045ndash1053 2010
6 Dymowska AK Hwang PP Goss GG Structure and function of iono-cytes in the freshwater fish gill Respir Physiol Neurobiol 184 282ndash2922012
7 Edwards SL Tse CM Toop T Immunolocalization of NHE3-likeimmunoreactivity in the gills of the rainbow trout (Oncorhynchus mykiss)and the blue-throated wrasse (Pseudolabrus tetrious) J Anat 195 465ndash469 1999
8 Evans DH Piermarini PM Choe KP The multifunctional gill domi-nant site of gas exchange osmoregulation acid-base regulation andexcretion of nitrogenous waste Physiol Rev 85 97ndash177 2005
9 Fenwick JC Bonga SE Flik G In vivo bafilomycin-sensitive Na
uptake in young freshwater fish J Exp Biol 202 3659ndash3666 199910 Galvez F Reid SD Hawkings G Goss GG Isolation and characteriza-
tion of mitochondria-rich cell types from the gill of freshwater rainbowtrout Am J Physiol Regul Integr Comp Physiol 282 R658ndashR668 2002
11 Goss G Orlowski J Grinstein S Coimmunoprecipitation of a 24-kDaprotein with NHE1 the ubiquitous isoform of the NaH exchanger AmJ Physiol Cell Physiol 270 C1493ndashC1502 1996
12 Goss GG Wood CJ Na and Cl uptake kinetics diffusive effluxes andacidic equivalent fluxes across the gills of rainbow trout 1 Responses toenvironmental hyperoxia J Exp Biol 152 521ndash547 1990
13 Grunder S Geissler HS Bassler EL Ruppersberg JP A new memberof acid-sensing ion channels from pituitary gland Neuroreport 11 1607ndash1611 2000
14 Harvey BJ Energization of sodium absorption by the H-ATPase pumpin mitochondria-rich cells of frog skin J Exp Biol 172 289ndash309 1992
15 Hirata T Kaneko T Ono T Nakazato T Furukawa N Hasegawa SWakabayashi S Shigekawa M Chang MH Romero MF Hirose SMechanism of acid adaptation of a fish living in low pH 35 lake Am JPhysiol Regul Integr Comp Physiol 284 R1199ndashR1212 2003
16 Holzer P Acid-sensitive ion channels and receptors Hand Exp Pharma-col 194 283ndash332 2009
17 Hung CY Tsui KN Wilson JM Nawata CM Wood CM Wright PARhesus glycoprotein gene expression in the mangrove killifish Kryptole-bias marmoratus exposed to elevated environmental ammonia levels andair J Exp Biol 210 2419ndash2429 2007
18 Hwang PP Lee TH Lin LY Ion regulations in fish gills recentprogresses in the cellular and molecular mechanisms Am J Physiol RegulIntegr Comp Physiol 301 R28ndashR47 2011
19 Ivanis G Esbaugh AJ Perry SF Branchial expression and localizationof SLC9A2 and SLC9A3 sodiumhydrogen exchangers and their possiblerole in acid-base regulation in freshwater rainbow trout (Oncorhynchusmykiss) J Exp Biol 211 2467ndash2477 2008
20 Jasti J Furukawa H Gonzales EB Gouaux E Structure of acid-sensingion channel 1 at 19A resolution and low pH Nature 449 316ndash323 2007
21 Jonz MG Nurse CA Epithelial mitochondria-rich cells and associatedinnervation in adult and developing zebrafish J Comp Neurol 497817ndash832 2006
C264 INVOLVEMENT OF ASICs IN FISH GILL Na TRANSPORT
AJP-Cell Physiol bull doi101152ajpcell003982013 bull wwwajpcellorg
22 Jonz MG Nurse CA New developments on gill innervation insightsfrom a model vertebrate J Exp Biol 211 2371ndash2378 2008
23 Kellenberger S Epithelial sodium and acid-sensing ion channels InSensing With Ion Channels edited by Martinac B New York Springer2008 chapt 11 p 225ndash246
24 Kellenberger S Schild L Epithelial sodium channeldegenerin family ofion channels a variety of functions for a shared structure Physiol Rev 82735ndash767 2002
25 Kleyman TR Cragoe EJ Amiloride and its analogs as tools in the studyof ion-transport J Membr Biol 105 1ndash21 1988
26 Krogh A The active absorption of ions in some freshwater animals ZVergl Physiol 25 335ndash350 1938
27 Levanti MB Guerrera MC Calavia MG Ciriaco E Montalbano GCobo J Germana A Vega JA Acid-sensing ion channel 2 (ASIC2) inthe intestine of adult zebrafish Neurosci Lett 494 24ndash28 2011
28 Lin H Randall DJ Evidence for the presence of an electrogenic protonpump on the trout gill epithelium J Exp Biol 161 119ndash134 1991
29 Maetz J Garcia Romeu F The mechanism of sodium and chlorideuptake by the gills of a fresh-water fish Carassius auratus II Evidencefor NH4
Na and HCO3Cl exchanges J Gen Physiol 47 1209ndash1227
196430 Nakada T Hoshijima K Esaki M Nagayoshi S Kawakami K Hirose
S Localization of ammonia transporter Rhcg1 in mitochondrion rich cellsof yolk sac gill and kidney of zebrafish and its ionic strength-dependentexpression Am J Physiol Regul Integr Comp Physiol 293 R1743ndashR17532007
31 Nawata CM Hung CC Tsui TK Wilson JM Wright PA Wood CMAmmonia excretion in rainbow trout (Oncorhynchus mykiss) evidence forRh glycoprotein and H-ATPase involvement Physiol Genomics 31463ndash474 2007
32 Parks SK Tresguerres M Galvez F Goss GG Intracellular pH regu-lation in isolated trout gill mitochondrion-rich (MR) cell subtypes evi-dence for NaH activity Comp Biochem Physiol A 155 139ndash1452010
33 Parks SK Tresguerres M Goss GG Interactions between Na channelsand Na-HCO3
cotransporters in the freshwater fish gill MRC a modelfor transepithelial Na uptake Am J Physiol Cell Physiol 292 C935ndashC944 2007
34 Parks SK Tresguerres M Goss GG Theoretical considerations under-lying Na uptake mechanisms in freshwater fishes Comp BiochemPhysiol C 148 411ndash418 2008
35 Paukert M Sidi S Russell C Siba M Wilson SW A family ofacid-sensing ion channels from zebrafish widespread expression in thecentral nervous system suggests a conserved role in neuronal communi-cation J Biol Chem 279 18783ndash18791 2004
36 Perry SF The chloride cell structure and function in the gill of freshwaterfish Annu Rev Physiol 59 325ndash347 1997
37 Perry SF Randall DJ Effects of amiloride and SITS on branchial ionfluxes in rainbow trout Salmo gairdneri J Exp Zool 215 225ndash228 1981
38 Preest MR Gonzales RJ Wilson RW Pharmacological examination ofNa and Cl transport in two species of freshwater fish Physiol BiochemZool 78 259ndash272 2005
39 Reid SD Hawkings GS Galvez F Goss GG Localization and charac-terization of phenamil-sensitive Na influx in isolated rainbow trout gillepithelial cells J Exp Biol 206 551ndash559 2003
40 Sakai H Lingueglia E Champigny G Mattei MG Lazdunski MCloning and functional expression of a novel degenerin-like Na channelgene in mammals J Physiol 519 323ndash333 1999
41 Shih TH Horng JL Liu ST Hwang PP Lin LY Rhcg1 and NHE3b areinvolved in ammonium-dependent sodium uptake by zebrafish larvaeacclimated to low-sodium water Am J Physiol Regul Integr Comp Physiol302 R84ndashR93 2012
42 Springauf A Grunder S An acid-sensing ion channel from shark(Squalus acanthias) mediates transient and sustained responses to protonsJ Physiol 588 809ndash820 2010
43 Spry DJ Wood CM Ion flux rates acid-base status and blood gases inrainbow trout Salmo gairdneri exposed to toxic zinc in natural soft waterCan J Fish Aquat Sci 42 1332ndash1337 1985
44 Sullivan GV Fryer JN Perry SF Immunolocalization of proton pumps(H-ATPase) in pavement cells of rainbow trout gill J Exp Biol 1982619ndash2629 1996
45 Tresguerres M Parks SK Katoh F Goss GG Microtubule-dependentrelocation of branchial V-H-ATPase to the basolateral membrane in thePacific spiny dogfish (Squalus acanthias) a role in base secretion J ExpBiol 209 599ndash609 2006
46 Vina E Parisi V Cabo R Laura R Lopez-Velasco S Lopez-Muniz AGarcia-Suarez O Germana A Vega JA Acid-sensing ion channels(ASICs) in the taste buds of adult zebrafish Neurosci Lett 536 35ndash402013
47 Waldmann R Lazdunski M H-gated cation channels neuronal acidsensors in the NaCDEG family of ion channels Curr Opin Neurobiol 8418ndash424 1998
48 Wilson JM Laurent P Tufts BL Benos DJ Donowitz M Vogl AWRandall DJ NaCl uptake by the branchial epithelium in freshwater teleostfish an immunological approach to ion-transport protein localization JExp Biol 203 2279ndash2296 2000
49 Wood CM Randall DJ Sodium balance in rainbow-trout (Salmo-gaird-neri) during extended exercise J Comp Physiol 82 235ndash256 1973
50 Wright PA Wood CM A new paradigm for ammonia excretion inaquatic animals role of rhesus (Rh) glycoproteins J Exp Biol 2122303ndash2312 2009
51 Yan JJ Chou MY Kaneko T Hwang PP Gene expression of NaH
exchanger in zebrafish H-ATPase-rich cells during acclimation to low-Na and acidic environments Am J Physiol Cell Physiol 293 C1814ndashC1823 2007
C265INVOLVEMENT OF ASICs IN FISH GILL Na TRANSPORT
AJP-Cell Physiol bull doi101152ajpcell003982013 bull wwwajpcellorg
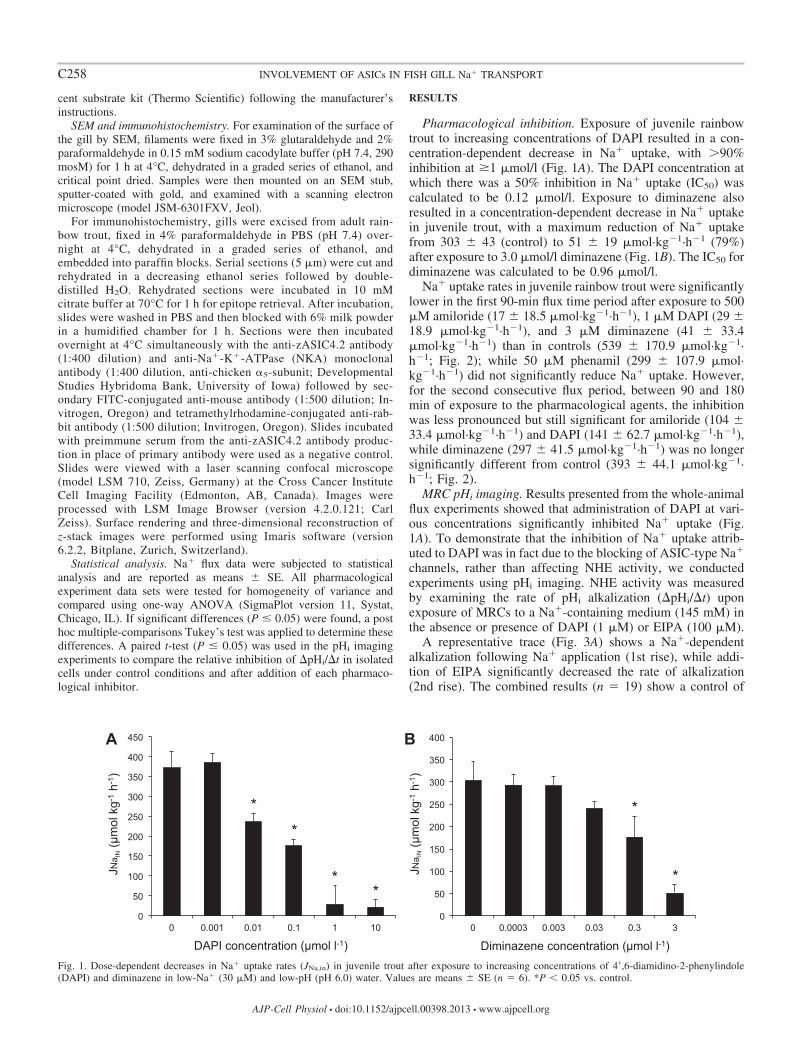
cent substrate kit (Thermo Scientific) following the manufacturerrsquosinstructions
SEM and immunohistochemistry For examination of the surface ofthe gill by SEM filaments were fixed in 3 glutaraldehyde and 2paraformaldehyde in 015 mM sodium cacodylate buffer (pH 74 290mosM) for 1 h at 4degC dehydrated in a graded series of ethanol andcritical point dried Samples were then mounted on an SEM stubsputter-coated with gold and examined with a scanning electronmicroscope (model JSM-6301FXV Jeol)
For immunohistochemistry gills were excised from adult rain-bow trout fixed in 4 paraformaldehyde in PBS (pH 74) over-night at 4degC dehydrated in a graded series of ethanol andembedded into paraffin blocks Serial sections (5 m) were cut andrehydrated in a decreasing ethanol series followed by double-distilled H2O Rehydrated sections were incubated in 10 mMcitrate buffer at 70degC for 1 h for epitope retrieval After incubationslides were washed in PBS and then blocked with 6 milk powderin a humidified chamber for 1 h Sections were then incubatedovernight at 4degC simultaneously with the anti-zASIC42 antibody(1400 dilution) and anti-Na-K-ATPase (NKA) monoclonalantibody (1400 dilution anti-chicken 5-subunit DevelopmentalStudies Hybridoma Bank University of Iowa) followed by sec-ondary FITC-conjugated anti-mouse antibody (1500 dilution In-vitrogen Oregon) and tetramethylrhodamine-conjugated anti-rab-bit antibody (1500 dilution Invitrogen Oregon) Slides incubatedwith preimmune serum from the anti-zASIC42 antibody produc-tion in place of primary antibody were used as a negative controlSlides were viewed with a laser scanning confocal microscope(model LSM 710 Zeiss Germany) at the Cross Cancer InstituteCell Imaging Facility (Edmonton AB Canada) Images wereprocessed with LSM Image Browser (version 420121 CarlZeiss) Surface rendering and three-dimensional reconstruction ofz-stack images were performed using Imaris software (version622 Bitplane Zurich Switzerland)
Statistical analysis Na flux data were subjected to statisticalanalysis and are reported as means SE All pharmacologicalexperiment data sets were tested for homogeneity of variance andcompared using one-way ANOVA (SigmaPlot version 11 SystatChicago IL) If significant differences (P 005) were found a posthoc multiple-comparisons Tukeyrsquos test was applied to determine thesedifferences A paired t-test (P 005) was used in the pHi imagingexperiments to compare the relative inhibition of pHit in isolatedcells under control conditions and after addition of each pharmaco-logical inhibitor
RESULTS
Pharmacological inhibition Exposure of juvenile rainbowtrout to increasing concentrations of DAPI resulted in a con-centration-dependent decrease in Na uptake with 90inhibition at 1 moll (Fig 1A) The DAPI concentration atwhich there was a 50 inhibition in Na uptake (IC50) wascalculated to be 012 moll Exposure to diminazene alsoresulted in a concentration-dependent decrease in Na uptakein juvenile trout with a maximum reduction of Na uptakefrom 303 43 (control) to 51 19 molmiddotkg1middoth1 (79)after exposure to 30 moll diminazene (Fig 1B) The IC50 fordiminazene was calculated to be 096 moll
Na uptake rates in juvenile rainbow trout were significantlylower in the first 90-min flux time period after exposure to 500M amiloride (17 185 molmiddotkg1middoth1) 1 M DAPI (29 189 molmiddotkg1middoth1) and 3 M diminazene (41 334molmiddotkg1middoth1) than in controls (539 1709 molmiddotkg1middoth1 Fig 2) while 50 M phenamil (299 1079 molmiddotkg1middoth1) did not significantly reduce Na uptake Howeverfor the second consecutive flux period between 90 and 180min of exposure to the pharmacological agents the inhibitionwas less pronounced but still significant for amiloride (104 334 molmiddotkg1middoth1) and DAPI (141 627 molmiddotkg1middoth1)while diminazene (297 415 molmiddotkg1middoth1) was no longersignificantly different from control (393 441 molmiddotkg1middoth1 Fig 2)
MRC pHi imaging Results presented from the whole-animalflux experiments showed that administration of DAPI at vari-ous concentrations significantly inhibited Na uptake (Fig1A) To demonstrate that the inhibition of Na uptake attrib-uted to DAPI was in fact due to the blocking of ASIC-type Na
channels rather than affecting NHE activity we conductedexperiments using pHi imaging NHE activity was measuredby examining the rate of pHi alkalization (pHit) uponexposure of MRCs to a Na-containing medium (145 mM) inthe absence or presence of DAPI (1 M) or EIPA (100 M)
A representative trace (Fig 3A) shows a Na-dependentalkalization following Na application (1st rise) while addi-tion of EIPA significantly decreased the rate of alkalization(2nd rise) The combined results (n 19) show a control of
0
50
100
150
200
250
300
350
400
450
0 0001 001 01 1 10
JNa I
N(μ
mol
kg-1
h-1 )
DAPI concentration (μmol l-1)
0
50
100
150
200
250
300
350
400
0 00003 0003 003 03 3
JNa I
N(μ
mol
k g-1
h-1 )
Diminazene concentration (μmol l-1)
A B
Fig 1 Dose-dependent decreases in Na uptake rates (JNain) in juvenile trout after exposure to increasing concentrations of 4=6-diamidino-2-phenylindole(DAPI) and diminazene in low-Na (30 M) and low-pH (pH 60) water Values are means SE (n 6) P 005 vs control
C258 INVOLVEMENT OF ASICs IN FISH GILL Na TRANSPORT
AJP-Cell Physiol bull doi101152ajpcell003982013 bull wwwajpcellorg
0642 0040 pHimin while the alkalization rate wasreduced by 62 in the presence of EIPA (0 240 0036pHimin P 0001 Fig 3B) For DAPI Na-dependentalkalization following Na application was similar in the
presence or absence of DAPI (Fig 3D) No differences inpHit were noted between control (0542 0042 pHimin) and DAPI-treated (0578 0047 pHimin) cells(P 0149 n 22 Fig 3D)
0
100
200
300
400
500
600
700
800
Cont Amil DAPI Dim Phen Cont Amil DAPI Dim Phen
JNa I
N(μ
mol
kg-1
h-1 )
Treatment
T0-90min T90-180min
Fig 2 Effects of pharmacological agents [control (Cont DMSO) amiloride (Amil 500 moll) DAPI (1 moll) diminazene (Dim 3 moll) and phenamil(Phen 50 moll)] during consecutive 90-min measurements of Na uptake in juvenile trout in low-Na (30 M) and low-pH (pH 60) water Values are means SE(n 6) P 005 vs control
65
7
75
8
85
9
0 100 200 300 400 500 600
pHi
Time (sec)
Na FreeNa Free Na
EIPA
Na
0
01
02
03
04
05
06
07
08
Na+ Na+EIPA
ΔpH
iΔM
in
Cell Alkalinization Treatments
65
7
75
8
85
9
0 100 200 300 400 500 600
pHi
Time (sec)
Na Free Na
DAPI
Na Na Free
0
01
02
03
04
05
06
07
08
Na+ Na+DAPI
ΔpH
iΔM
in
Cell Alkalinization Treatments
A
DC
B
Fig 3 Effect of ethylisopropylamiloride (EIPA) and DAPI on Na-induced alkalinization of peptide nucleic acid-positive (PNA) cells following Na-freeexposure at resting intracellular pH (pHi) A representative trace demonstrating inhibition of cell alkalinization by EIPA (100 M) B quantitative analysis ofcell alkalinization in the absence and presence of EIPA (62 inhibition) Values are means SE (n 19 cells) P 0001 (by paired t-test) C representativetrace demonstrating lack of effect on cell alkalinization in the presence of DAPI (1 M) D quantitative analysis of cell alkalinization in the absence and presenceof DAPI Values are means SE (n 22) P 005 (by paired t-test)
C259INVOLVEMENT OF ASICs IN FISH GILL Na TRANSPORT
AJP-Cell Physiol bull doi101152ajpcell003982013 bull wwwajpcellorg
Phylogenetic analysis and tissue distribution of ASIC1 andASIC4 Two distinct DNA fragments whose derived aminoacid sequences exhibit similarity to the ASIC family werecloned from the gill of rainbow trout To determine theiridentity we conducted a phylogenetic analysis by comparisonwith known ASIC homologs from a variety of species Thegenerated phylogenetic tree (Fig 4) showed that the twocloned rainbow trout ASIC genes (tASICs) were members ofthe ASIC1 and ASIC4 subunits respectively (Fig 4A) ThetASIC1 (partial clone) grouped most closely with ASIC1 fromsalmon (Salmo salar) while tASIC4 grouped with other iden-
tified ASIC4 family members from other fish and mammalspecies
Furthermore we conducted a tissue distribution analysis oftASIC1 and tASIC4 by RT-PCR We observed mRNA expres-sion of tASIC1 in all tested tissues gill MRCs head and trunkkidney brain and blood tASIC4 had a slightly differentdistribution it was present in the perfused gill MRCs brainand trunk kidney but it was absent in head kidney and bloodtissues
Immunoprecipitation and immunostaining with anti-zASIC42To determine if ASIC4 was present in the fish gill and to validate our
Fig 4 Phylogenetic analysis of amino acid sequences of cloned rainbow trout acid-sensing ion channels ASIC1 and ASIC4 and ASICs from various fish speciesPhylogenetic tree was constructed using the maximum-likelihood method and numbers indicate bootstrap values for 700 resamplings GenBank or Ensemblaccession numbers are as follows human ASIC2 NP_0010852 Western clawed frog ASIC2 XP_0029330862 Japanese pufferfish ASIC2 XP_0039648711zebrafish ASIC2 NP_9999531 three-spined stickleback ASIC2 ENSGACG00000009558 Atlantic cod ASIC2 ENSGMOT00000019145 spotted greenpufferfish ASIC2 ENSTNIG00000014943 Southern platyfish ASIC2 ENSXMAG00000008283 oyster toadfish ASIC2 AAP456331 coelacanth ASIC2ENSLACG00000007191 rat ASIC2 NP_0370242 Southern platyfish ASIC1c ENSXMAG00000018841 African clawed frog ASIC11 AEC486651 Africanclawed frog ASIC12 AEC486671 brown rat ASIC1 NP_0770681 human ASIC1 NP_0644232 three-spined stickleback ASIC1a ENSGACG00000003075Japanese pufferfish ASIC1 XP_0039635201 Southern platyfish ASIC1a XP_0058018011 Atlantic cod ASIC1a ENSGMOG00000009659 zebrafishASIC12 NP_9999551 rainbow trout ASIC1 KF964646 Atlantic salmon ASIC1 NP_0011334561 oyster toadfish ASIC1 Q7T1N41 elephant shark ASIC1AEC486681 three-spined stickleback AIC1c ENSGACG00000017974 Atlantic cod ASIC1c ENSGMOG00000003105 zebrafish ASIC13 NP_9999541river lamprey ASIC1 AAY289831 sea lamprey ASIC1b ENSPMAG00000003813 dogfish shark ASIC1 AAY289851 zebrafish ASIC11 NP_9999561human ASIC4 NP_8782672 rat ASIC4 NP_0715702 African clawed frog ASIC4 AEC486661 Japanese pufferfish ASIC4 XP_0039621271 spotted greenpufferfish ASIC4b ENSTNIG00000010994 Nile tilapia ASIC4 XP_0034454461 Southern platyfish ASIC4a ENSXMAG00000003624 three-spinedstickleback ASIC4a ENSGACG00000007052 Atlantic cod ASIC4a ENSGMOG00000012740 rainbow trout ASIC4 KF964645 zebrafish ASIC41NP_9999521 zebrafish ASIC42 NP_9999511
C260 INVOLVEMENT OF ASICs IN FISH GILL Na TRANSPORT
AJP-Cell Physiol bull doi101152ajpcell003982013 bull wwwajpcellorg
antibody for later immunocytochemistry we performed an im-munoprecipitation using an antibody raised against zASIC42on the basis of sequence similarly of 60 between zASIC42(aa 146 ndash160 PKSRKGHRPSELQYPP) and the clonedtASIC4 (aa 142ndash156 PKNREGHKPTDLDYPA) Using West-ern blotting we analyzed the precipitates for the presence ofASIC4 (see Fig 6A) and found a single band of 65 kDawhich approximates the predicted size for tASIC4 (604 kDa)The strong band at 55 kDa corresponds to the IgG heavychain
To determine the location of ASIC4 protein within therainbow trout gill tissue fixed sections were double-labeledwith anti-zASIC42 antibody (see Fig 6B) and anti-NKA andthen counterstained with the nuclear stain DAPI Immunore-activity was observed in cells located in the lamellae andinterlamellar region (see Fig 6B arrowhead) Moreover themajority of cells positive for zASIC42 also colocalized withNKA protein (Fig 6 C and D arrowhead) a known marker ofMRCs suggesting that in gill epithelium ASIC4 protein ismainly expressed in MRCs SEM analysis of the gill filamentdemonstrated that MRCs are clearly distinguished on the sur-face of the epithelium and have a broad apical convex openingto the environment (see Fig 7A arrowheads) The generatedthree-dimensional reconstructions of the staining immunoreac-tivity pattern for ASIC4 and NKA proteins revealed the pres-ence of ASIC4 and NKA in the same cells but in distinctregions of the cells NKA was present on the basolateralmembrane (arrows green staining) while ASIC4 was localizedto the broad apical surface of the cell (arrowheads red stain-ing) Figure 7B shows a generated image with a top view whileFig 7C is the same cell viewed from the side
DISCUSSION
The existence of an apical Na channel electrochemicallycoupled to a VHA mediating Na uptake from dilute FW hasbeen postulated on theoretical thermodynamic grounds for anumber of decades (1 36 34) However an inability todetermine the molecular identity of the Na channel has beena major drawback in the general acceptance of the modelUsing a combination of pharmacological blockade isotopicfluxes immunocytochemistry and molecular biology we pro-pose that an ASIC family member is potentially the long-sought-after apical Na channel responsible for Na uptake byrainbow trout acclimatized to water with low Na and low pHOur study provides the first evidence for a nonneuronal role forASICs in any vertebrate whereby in gills from FW fishesASICs are associated with the MRCs and tASIC4 localizes tothe apical membrane of NKA positive cells
ASICs are a broad family of Na-specific ion channels thathave been extensively studied in mammalian models (forreview see Ref 23) with only a few studies performed in anyfish species (35 42 46) ASICs have primarily been associatedwith neuronal tissues with studies focusing on a neuronalfunction for ASICs (23) In this study ASIC1 and ASIC4 werecloned in rainbow trout using PCR and they were expressed ina variety of tissues both neuronal and nonneuronal in origin(Fig 5) Importantly tASIC1 gene was present in all fiveinvestigated tissues including whole blood It appears that thetASIC4 gene has a more defined tissue distribution thantASIC1 with expression in the ionoregulatory tissues (trunk
kidney and perfused gill) and the brain Unlike tASIC1 therewas no apparent expression of tASIC4 in the head kidney orthe blood Therefore tASIC4 expression in MRCs and per-fused gill could not be attributed to blood contamination
Both tASIC1 and tASIC4 genes were expressed in the gillepithelium and more specifically were present in MRCs ofrainbow trout gills It is believed that similar to ENaCsfunctional ASICs are trimers (20) although their configurationhas not been resolved It has been demonstrated that with afew exceptions ASICs function as homomers (forming chan-nels of identical subunits) or heteromers (forming channelswith other ASIC subunits) (4 23) ASICs have been charac-terized by using oocyte expression in zebrafish (4 35) anddogfish shark (ASIC1b only) (42) It has been found thatcoexpression of zASIC41 with zASIC13 in Xenopus oocytesincreased the current amplitude and abundance (15-fold) ofthe channel at the cell surface indicating that these two ASICsubunits form a functional channel (4) Moreover thisASIC4113 heteromeric channel was more efficiently traf-ficked to the plasma membrane and had increased affinity forH Additionally zASIC41 when heterologously expressedin Xenopus oocytes has been shown to be also gated open bydecreases in extracellular Ca2 The signal responsible forgating of zASIC42 was unable to be determined in that study(4) It is therefore possible that tASIC1 and tASIC4 form aheteromeric channel in MRCs but this cannot be determined atthis time and would require confirmation by immunohisto-chemistry with an anti-ASIC1 antibody that is immunoreactivefor rainbow trout It is also possible that other ASIC subunitsandor regulatory elements may be involved in the function ofASICs in the FW trout gill A full characterization of tASIC4by oocyte expression and electrophysiology is required tounderstand the mechanisms of gating in low-Na low-Ca2
watersIn our study ASIC4 protein was colocalized with NKA and
was present on the apical surface of cells suggesting that it isfound primarily in MRCs (Fig 6 D and E) Previous studies onASICs in fishes have suggested that they are primarily associ-ated with neuronal tissue and sensory cells (27 35 42 46) Inadult and developing zebrafish morphological studies usingconfocal immunofluorescence techniques demonstrated thatgill MRCs come into direct contact with nerve fibers in thebasolateral region (21 22) However in our study ASIC4protein was clearly localized to the apical region of the MRCsas demonstrated by three-dimensional reconstruction of con-
Head k
idney
ASIC4
ASIC1
GAPDH
Gill MRCs
Brain
Trunk k
idney
Blood
No tem
plate
Fig 5 Tissue distribution analysis of ASIC4 and ASIC1 genes in adultrainbow trout as determined by PCR GAPDH was used as a reference geneMRCs mitochondrion-rich cells
C261INVOLVEMENT OF ASICs IN FISH GILL Na TRANSPORT
AJP-Cell Physiol bull doi101152ajpcell003982013 bull wwwajpcellorg
focal images ASIC4 immunoreactivity did not overlap withthe standard basolateral immunoreactive pattern associatedwith NKA (Fig 7 B and C) Therefore we conclude thatneuronally derived tissues cannot account for the expression ofASIC4 protein in MRCs We also corroborate the ASIC4staining pattern with the MRC external morphology whichshows that in FW rainbow trout raised in Edmonton tap water(500 M Na) the MRCs have wide apical openings (Fig7A) This apical localization of ASIC4 protein in the cells of atransporting epithelium is the first evidence for a nonneuronalfunction for any ASIC member and suggests that ASIC4 as aNa channel may be responsible for apical uptake of Na inFW fish gill
Pharmacological profiling has long been used to differentiatethe mechanisms of ion transport across epithelial tissues For Na
transport classical analysis has usually involved amiloride and itssubstituted derivatives (eg EIPA mercaptoisobutyric acid andphenamil) as a means to differentiate between the involvementof NHE isoforms and ENaC in Na transport (25) We inves-tigated two newly described ASIC-specific inhibitors dimina-zene and DAPI (5) for their potency in inhibiting unidirec-tional Na uptake in fish Diminazene and DAPI exhibited adose-dependent inhibitory relationship and both were veryeffective at low concentrations (Fig 1) Na uptake in rainbow
trout acclimatized to low-ionic-strength acidic water wasclearly inhibited by these diarylamidines at concentrationssimilar to those reported in a previous study of cultured rathippocampal neurons (5) However in our study DAPI provedto be more effective than diminazene in contrast to observa-tions in rat neurons The inhibition of 22Na uptake by DAPIand diminazene was also compared with other common Na
transport inhibitors amiloride and phenamil Amiloride wasobserved to be an effective inhibitor of Na uptake in rainbowtrout but only at relatively high concentrations (500 M)which is consistent with previous studies (2 37) Recentlyamiloride was also demonstrated to completely block ASIC-mediated currents (5 35 42) however it is also known toblock many other Na transporters including NHE and ENaC(25) Amiloride therefore cannot be used as a distinguishingpharmacological agent
While DAPI diminazene and amiloride almost completelyblocked Na transport in the present study phenamil had nosignificant effect on Na flux in rainbow trout This suggeststhat phenamil is a poor inhibitor of Na uptake in troutexposed to low-ionic-strength water (30 moll Na) Paststudies of the effects of phenamil on Na uptake in a variety ofFW fishes have reported inconsistent results For example inneon tetras acclimatized to low Na (50 moll) phenamil had
Fig 6 Immunoreactivity of anti-zebrafish ASIC42 (zASIC42) antibody in the gill of rainbow trout A immunoprecipitation Western blot from whole gillhomogenates with zASIC42 showing a distinct band of 65 kDa Lane 1 (L1) contains 300 mg of gill tissue and lane 2 (L2) contains 200 mg of gill tissueBndashE confocal images of gill sections labeled for ASIC4 and Na-K-ATPase (NKA) proteins B and C double-labeling with DAPI and anti-zASIC42 (B) andDAPI and anti-NKA (C) D merged image of B and C E higher-magnification image of a MRC showing that ASIC4 and NKA are present in distinct regionswithin the MRC F control micrograph with DAPI and no primary antibody Arrowheads (BndashD) indicate positive staining
C262 INVOLVEMENT OF ASICs IN FISH GILL Na TRANSPORT
AJP-Cell Physiol bull doi101152ajpcell003982013 bull wwwajpcellorg
no effect on Na transport (38) whereas it caused 70reduction in Na influx in Cyprinidon variegatus hubbsi (100moll Na) and 60 reduction in goldfish (300 moll Na)(2 38) These conflicting results from various phenamil studiessuggest that sensitivity to phenamil might be species- andorenvironment-specific Collectively the pharmacological anal-ysis suggests that the Na uptake in FW rainbow trout is mostlikely not mediated by ENaC given the lack of inhibition byphenamil but may be attributed to ASIC since Na uptakewas almost completely blocked by DAPI and diminazene bothof which inhibit ASICs but not ENaC
In our Na flux study with various pharmacological agentswe made an interesting observation that inhibition of 22Na
uptake was less pronounced for all tested inhibitors during thesecond consecutive measurement (90ndash180 min) (Fig 2) Toour knowledge this phenomenon where potency of the testeddrug diminishes over time has not been reported in anyprevious study on fish This observation may be explained bya decreased stabilityefficacy of the tested pharmacologicalagents in aqueous media metabolism of the drug by theanimal or activation of compensatory Na uptake mechanismsnot sensitive to amiloride DAPI or diminazene We believethat the decrease of the potency of amiloride DAPI anddiminazene over time demonstrates the importance of properexperimental design and choice of period of study when ionfluxes are examined using pharmacological inhibitors
While the two novel ASIC inhibitors have been tested fortheir potency in inhibiting ASICs and ENaC in neurons (5)they have not been tested for their potential effects on otherNa transporters implicated in Na uptake in fishes mostnotably the NHEs To verify that the Na uptake inhibitioncaused by DAPI and diminazene in rainbow trout was due toinhibition of ASICs rather than NHEs we tested the effect ofDAPI on MRCs isolated from the rainbow trout gill epithe-lium Our laboratory previously showed that rainbow troutpossess two populations of MRCs peanut lectin agglutinin(PNA) positive (PNA) and PNA negative (PNA) MRCs(10) It has been demonstrated that NHE3 and NHE2 areexpressed in PNA cells isolated from branchial epithelium of
FW rainbow trout (19) Therefore we investigated the effect ofDAPI on PNA cells and compared it with the effect of EIPAa potent and highly selective inhibitor of NHE (25) Earlierstudies in our laboratory using cell imaging determined thatPNA cells undergo NHE-dependent alkalinization (32 33)Here we demonstrate that while EIPA was effective at inhib-iting NHE function in PNA cells (Fig 3A) there was noeffect of high concentrations of DAPI on Na-dependent pHi
elevation demonstrating that NHE was not inhibited by DAPI(Fig 3B) Therefore the measured inhibition of Na uptake byDAPI that we see in whole animals (Fig 1) cannot be attrib-uted to inhibition of NHE
On the basis of the mRNA expression of tASIC1 andtASIC4 genes in the gill epithelium and isolated MRCs local-ization of ASIC4 protein to the apical region of the MRCs and
Fig 7 Three-dimensional views of freshwater(FW) rainbow trout MRCs A scanning elec-tron micrograph of FW rainbow trout gillepithelium showing broad apical openings ofMRCs (arrowheads) and mucous cells () Band C 3-dimensional reconstruction of z-stack images showing apical localization ofASIC4 protein (red arrowhead) and basolat-eral localization of NKA (green arrow)B top view C side view
PNA- MRC
Na+
ASIC
H+
VHA
NKA
Na+
K+
NBC
Na+ HCO3-
PNA+ MRC
NH3
Na+
H+
NHE Rhcg2
NKA
Na+
K+
Fig 8 Model for Na uptake in PNA and PNA MRCs in gill epithelium ofrainbow trout showing placement of proposed ASIC-mediated Na uptakemechanisms Rhcg2 rhesus glycoprotein NHE NaH exchanger VHAvacuolar H-ATPase NBC Na-HCO3
cotransporter
C263INVOLVEMENT OF ASICs IN FISH GILL Na TRANSPORT
AJP-Cell Physiol bull doi101152ajpcell003982013 bull wwwajpcellorg
pharmacological blockade of Na influx by ASIC inhibitors inadult rainbow trout we propose a revised model for transepi-thelial Na uptake in FW rainbow trout gills (Fig 8) wherebyone apical mode of Na entry is via an ASIC4-mediatedmechanism
A number of recent studies on Na uptake in FW fishes havedemonstrated the presence of the NHERh metabolon in thegills of various species (17 30 31) As mentioned previouslywe specifically targeted the rainbow trout as a model for thisstudy as it is native to oligotrophic Na-poor mountainstreams with low buffering capacity This type of environmentpresents key thermodynamic challenges to the NHERhmetabolon as Na restrictions cannot be alleviated by thepresence of an Rh protein (6) Moreover searches of rainbowtrout genomes and transcriptomes have failed to identify anyENaC subunits ( 13 or ) despite the fact that they are knownto actively take up Na from water with 50 M Na (49)Furthermore other research studies have suggested and dem-onstrated an apical localization of the VHA in the gills ofrainbow trout (44 48) and bafilomycin a VHA-specific in-hibitor has been demonstrated to reduce Na uptake in ze-brafish (2) tilapia and carp (9) Previous studies have pro-posed VHA to be a part of the NHERh metabolon (41 50 51)however the placement of an apically oriented VHA is coun-terintuitive to the function of an NHE as it would exacerbatethe noted thermodynamic constraints Placement of a VHA inconcert with an apical ASIC is more plausible since it wouldnot only facilitate Na uptake from very low ionic strengthwater but could also provide a mechanism for gating ofASIC-mediated Na conductance It remains to be determinedif ASIC and VHA proteins colocalize to the same type ofMRCs and this warrants further investigation
Perspectives In summary we propose that ASCI4 is in-volved in mediating Na uptake from FW in rainbow troutacclimatized to low ionic strength low pH media This is thefirst demonstration of a nonneuronal function for any ASICfamily member and simultaneously provides an attractive so-lution to the debate surrounding the thermodynamic limitationsof the other proposed Na transport mechanisms and resolvesthe conundrum of the role of an apical VHA in Na transportThe possibility that other fish models may use similar mecha-nisms under different environmental conditions where modelssuch as the NHERh metabolon cannot be expected to functionremains to be explored
ACKNOWLEDGMENTS
We thank the staff in the Biosciences Aquatic Facilities the MolecularBiology Service Unit and the Cross Cancer Institute Cell Imaging Facility atthe University of Alberta for outstanding help throughout the study We alsothank Alex Clifford for advice and help with phylogenetic analysis and JamesEde for help with confocal microscopy
GRANTS
This study was supported by a Natural Sciences and Engineering ResearchCouncil of Canada Discovery Grant to G G Goss and an Alberta IngenuityStudent Scholarship to A K Dymowska
DISCLOSURES
No conflicts of interest financial or otherwise are declared by the authors
AUTHOR CONTRIBUTIONS
AKD AGS and GGG are responsible for conception and design ofthe research AKD AGS SDB and DC performed the experiments
AKD AGS SDB DC and GGG analyzed the data AKD AGSSDB DC and GGG interpreted the results of the experiments AKDAGS SDB and GGG prepared the figures AKD AGS SDB DCand GGG drafted the manuscript AKD AGS SDB DC and GGGedited and revised the manuscript AKD AGS SDB and GGGapproved the final version of the manuscript
REFERENCES
1 Avella M Bornancin M A new analysis of ammonia and sodiumtransport through the gills of the freshwater rainbow trout (Salmo gaird-neri) J Exp Biol 142 155ndash175 1989
2 Boisen AM Amstrup J Novak I Grosell M Sodium and chloridetransport in soft water and hard water acclimatized zebrafish (Daniorerio) Biochim Biophys Acta 1618 207ndash218 2003
3 Brix KV Grosell M Comparative characterization of Na transport inCyprinodon variegatus variegatus and Cyprinodon variegatus hubbsi amodel species complex for studying teleost invasion of freshwater J ExpBiol 215 1199ndash1209 2012
4 Chen X Polleichtner G Kadurin I Grunder S Zebrafish acid-sensingion channel (ASIC) 4 Characterization of homo- and heteromeric chan-nels and identification of regions important for activation by H J BiolChem 282 30406ndash30413 2007
5 Chen X Qiu L Li M Durrnagel S Orser BA Xiong ZG MacDonaldJF Diarylamidines high potency inhibitors of acid-sensing ion channelsNeuropharmacology 58 1045ndash1053 2010
6 Dymowska AK Hwang PP Goss GG Structure and function of iono-cytes in the freshwater fish gill Respir Physiol Neurobiol 184 282ndash2922012
7 Edwards SL Tse CM Toop T Immunolocalization of NHE3-likeimmunoreactivity in the gills of the rainbow trout (Oncorhynchus mykiss)and the blue-throated wrasse (Pseudolabrus tetrious) J Anat 195 465ndash469 1999
8 Evans DH Piermarini PM Choe KP The multifunctional gill domi-nant site of gas exchange osmoregulation acid-base regulation andexcretion of nitrogenous waste Physiol Rev 85 97ndash177 2005
9 Fenwick JC Bonga SE Flik G In vivo bafilomycin-sensitive Na
uptake in young freshwater fish J Exp Biol 202 3659ndash3666 199910 Galvez F Reid SD Hawkings G Goss GG Isolation and characteriza-
tion of mitochondria-rich cell types from the gill of freshwater rainbowtrout Am J Physiol Regul Integr Comp Physiol 282 R658ndashR668 2002
11 Goss G Orlowski J Grinstein S Coimmunoprecipitation of a 24-kDaprotein with NHE1 the ubiquitous isoform of the NaH exchanger AmJ Physiol Cell Physiol 270 C1493ndashC1502 1996
12 Goss GG Wood CJ Na and Cl uptake kinetics diffusive effluxes andacidic equivalent fluxes across the gills of rainbow trout 1 Responses toenvironmental hyperoxia J Exp Biol 152 521ndash547 1990
13 Grunder S Geissler HS Bassler EL Ruppersberg JP A new memberof acid-sensing ion channels from pituitary gland Neuroreport 11 1607ndash1611 2000
14 Harvey BJ Energization of sodium absorption by the H-ATPase pumpin mitochondria-rich cells of frog skin J Exp Biol 172 289ndash309 1992
15 Hirata T Kaneko T Ono T Nakazato T Furukawa N Hasegawa SWakabayashi S Shigekawa M Chang MH Romero MF Hirose SMechanism of acid adaptation of a fish living in low pH 35 lake Am JPhysiol Regul Integr Comp Physiol 284 R1199ndashR1212 2003
16 Holzer P Acid-sensitive ion channels and receptors Hand Exp Pharma-col 194 283ndash332 2009
17 Hung CY Tsui KN Wilson JM Nawata CM Wood CM Wright PARhesus glycoprotein gene expression in the mangrove killifish Kryptole-bias marmoratus exposed to elevated environmental ammonia levels andair J Exp Biol 210 2419ndash2429 2007
18 Hwang PP Lee TH Lin LY Ion regulations in fish gills recentprogresses in the cellular and molecular mechanisms Am J Physiol RegulIntegr Comp Physiol 301 R28ndashR47 2011
19 Ivanis G Esbaugh AJ Perry SF Branchial expression and localizationof SLC9A2 and SLC9A3 sodiumhydrogen exchangers and their possiblerole in acid-base regulation in freshwater rainbow trout (Oncorhynchusmykiss) J Exp Biol 211 2467ndash2477 2008
20 Jasti J Furukawa H Gonzales EB Gouaux E Structure of acid-sensingion channel 1 at 19A resolution and low pH Nature 449 316ndash323 2007
21 Jonz MG Nurse CA Epithelial mitochondria-rich cells and associatedinnervation in adult and developing zebrafish J Comp Neurol 497817ndash832 2006
C264 INVOLVEMENT OF ASICs IN FISH GILL Na TRANSPORT
AJP-Cell Physiol bull doi101152ajpcell003982013 bull wwwajpcellorg
22 Jonz MG Nurse CA New developments on gill innervation insightsfrom a model vertebrate J Exp Biol 211 2371ndash2378 2008
23 Kellenberger S Epithelial sodium and acid-sensing ion channels InSensing With Ion Channels edited by Martinac B New York Springer2008 chapt 11 p 225ndash246
24 Kellenberger S Schild L Epithelial sodium channeldegenerin family ofion channels a variety of functions for a shared structure Physiol Rev 82735ndash767 2002
25 Kleyman TR Cragoe EJ Amiloride and its analogs as tools in the studyof ion-transport J Membr Biol 105 1ndash21 1988
26 Krogh A The active absorption of ions in some freshwater animals ZVergl Physiol 25 335ndash350 1938
27 Levanti MB Guerrera MC Calavia MG Ciriaco E Montalbano GCobo J Germana A Vega JA Acid-sensing ion channel 2 (ASIC2) inthe intestine of adult zebrafish Neurosci Lett 494 24ndash28 2011
28 Lin H Randall DJ Evidence for the presence of an electrogenic protonpump on the trout gill epithelium J Exp Biol 161 119ndash134 1991
29 Maetz J Garcia Romeu F The mechanism of sodium and chlorideuptake by the gills of a fresh-water fish Carassius auratus II Evidencefor NH4
Na and HCO3Cl exchanges J Gen Physiol 47 1209ndash1227
196430 Nakada T Hoshijima K Esaki M Nagayoshi S Kawakami K Hirose
S Localization of ammonia transporter Rhcg1 in mitochondrion rich cellsof yolk sac gill and kidney of zebrafish and its ionic strength-dependentexpression Am J Physiol Regul Integr Comp Physiol 293 R1743ndashR17532007
31 Nawata CM Hung CC Tsui TK Wilson JM Wright PA Wood CMAmmonia excretion in rainbow trout (Oncorhynchus mykiss) evidence forRh glycoprotein and H-ATPase involvement Physiol Genomics 31463ndash474 2007
32 Parks SK Tresguerres M Galvez F Goss GG Intracellular pH regu-lation in isolated trout gill mitochondrion-rich (MR) cell subtypes evi-dence for NaH activity Comp Biochem Physiol A 155 139ndash1452010
33 Parks SK Tresguerres M Goss GG Interactions between Na channelsand Na-HCO3
cotransporters in the freshwater fish gill MRC a modelfor transepithelial Na uptake Am J Physiol Cell Physiol 292 C935ndashC944 2007
34 Parks SK Tresguerres M Goss GG Theoretical considerations under-lying Na uptake mechanisms in freshwater fishes Comp BiochemPhysiol C 148 411ndash418 2008
35 Paukert M Sidi S Russell C Siba M Wilson SW A family ofacid-sensing ion channels from zebrafish widespread expression in thecentral nervous system suggests a conserved role in neuronal communi-cation J Biol Chem 279 18783ndash18791 2004
36 Perry SF The chloride cell structure and function in the gill of freshwaterfish Annu Rev Physiol 59 325ndash347 1997
37 Perry SF Randall DJ Effects of amiloride and SITS on branchial ionfluxes in rainbow trout Salmo gairdneri J Exp Zool 215 225ndash228 1981
38 Preest MR Gonzales RJ Wilson RW Pharmacological examination ofNa and Cl transport in two species of freshwater fish Physiol BiochemZool 78 259ndash272 2005
39 Reid SD Hawkings GS Galvez F Goss GG Localization and charac-terization of phenamil-sensitive Na influx in isolated rainbow trout gillepithelial cells J Exp Biol 206 551ndash559 2003
40 Sakai H Lingueglia E Champigny G Mattei MG Lazdunski MCloning and functional expression of a novel degenerin-like Na channelgene in mammals J Physiol 519 323ndash333 1999
41 Shih TH Horng JL Liu ST Hwang PP Lin LY Rhcg1 and NHE3b areinvolved in ammonium-dependent sodium uptake by zebrafish larvaeacclimated to low-sodium water Am J Physiol Regul Integr Comp Physiol302 R84ndashR93 2012
42 Springauf A Grunder S An acid-sensing ion channel from shark(Squalus acanthias) mediates transient and sustained responses to protonsJ Physiol 588 809ndash820 2010
43 Spry DJ Wood CM Ion flux rates acid-base status and blood gases inrainbow trout Salmo gairdneri exposed to toxic zinc in natural soft waterCan J Fish Aquat Sci 42 1332ndash1337 1985
44 Sullivan GV Fryer JN Perry SF Immunolocalization of proton pumps(H-ATPase) in pavement cells of rainbow trout gill J Exp Biol 1982619ndash2629 1996
45 Tresguerres M Parks SK Katoh F Goss GG Microtubule-dependentrelocation of branchial V-H-ATPase to the basolateral membrane in thePacific spiny dogfish (Squalus acanthias) a role in base secretion J ExpBiol 209 599ndash609 2006
46 Vina E Parisi V Cabo R Laura R Lopez-Velasco S Lopez-Muniz AGarcia-Suarez O Germana A Vega JA Acid-sensing ion channels(ASICs) in the taste buds of adult zebrafish Neurosci Lett 536 35ndash402013
47 Waldmann R Lazdunski M H-gated cation channels neuronal acidsensors in the NaCDEG family of ion channels Curr Opin Neurobiol 8418ndash424 1998
48 Wilson JM Laurent P Tufts BL Benos DJ Donowitz M Vogl AWRandall DJ NaCl uptake by the branchial epithelium in freshwater teleostfish an immunological approach to ion-transport protein localization JExp Biol 203 2279ndash2296 2000
49 Wood CM Randall DJ Sodium balance in rainbow-trout (Salmo-gaird-neri) during extended exercise J Comp Physiol 82 235ndash256 1973
50 Wright PA Wood CM A new paradigm for ammonia excretion inaquatic animals role of rhesus (Rh) glycoproteins J Exp Biol 2122303ndash2312 2009
51 Yan JJ Chou MY Kaneko T Hwang PP Gene expression of NaH
exchanger in zebrafish H-ATPase-rich cells during acclimation to low-Na and acidic environments Am J Physiol Cell Physiol 293 C1814ndashC1823 2007
C265INVOLVEMENT OF ASICs IN FISH GILL Na TRANSPORT
AJP-Cell Physiol bull doi101152ajpcell003982013 bull wwwajpcellorg
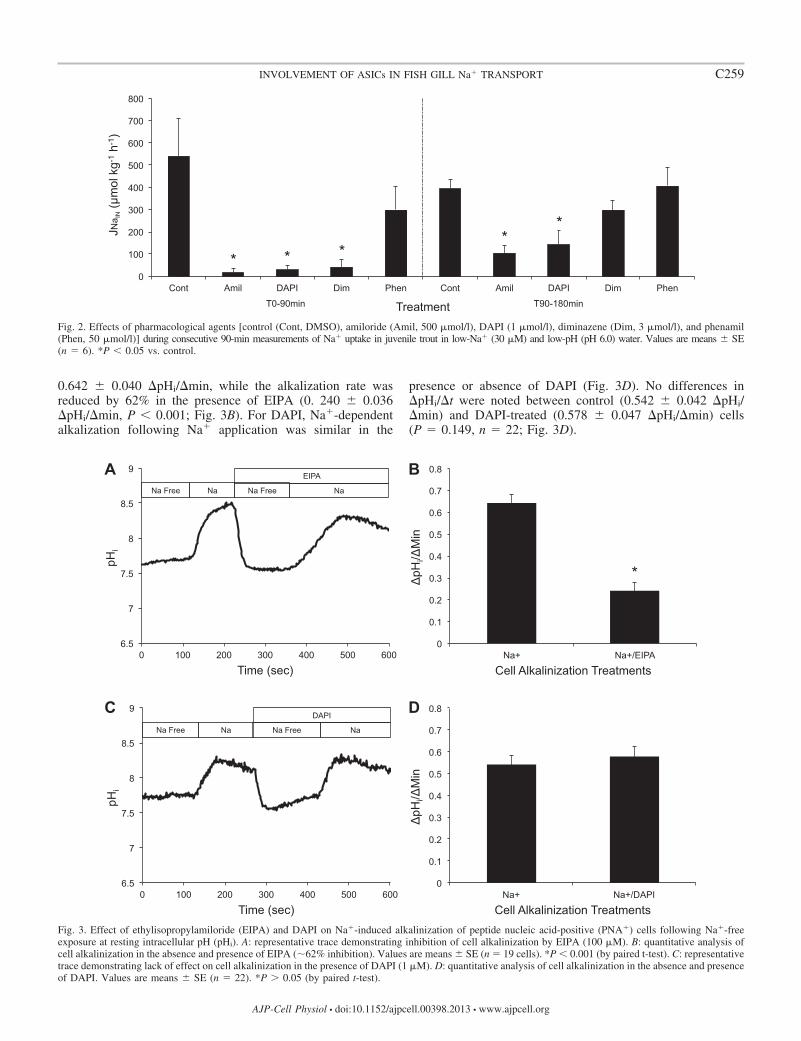
0642 0040 pHimin while the alkalization rate wasreduced by 62 in the presence of EIPA (0 240 0036pHimin P 0001 Fig 3B) For DAPI Na-dependentalkalization following Na application was similar in the
presence or absence of DAPI (Fig 3D) No differences inpHit were noted between control (0542 0042 pHimin) and DAPI-treated (0578 0047 pHimin) cells(P 0149 n 22 Fig 3D)
0
100
200
300
400
500
600
700
800
Cont Amil DAPI Dim Phen Cont Amil DAPI Dim Phen
JNa I
N(μ
mol
kg-1
h-1 )
Treatment
T0-90min T90-180min
Fig 2 Effects of pharmacological agents [control (Cont DMSO) amiloride (Amil 500 moll) DAPI (1 moll) diminazene (Dim 3 moll) and phenamil(Phen 50 moll)] during consecutive 90-min measurements of Na uptake in juvenile trout in low-Na (30 M) and low-pH (pH 60) water Values are means SE(n 6) P 005 vs control
65
7
75
8
85
9
0 100 200 300 400 500 600
pHi
Time (sec)
Na FreeNa Free Na
EIPA
Na
0
01
02
03
04
05
06
07
08
Na+ Na+EIPA
ΔpH
iΔM
in
Cell Alkalinization Treatments
65
7
75
8
85
9
0 100 200 300 400 500 600
pHi
Time (sec)
Na Free Na
DAPI
Na Na Free
0
01
02
03
04
05
06
07
08
Na+ Na+DAPI
ΔpH
iΔM
in
Cell Alkalinization Treatments
A
DC
B
Fig 3 Effect of ethylisopropylamiloride (EIPA) and DAPI on Na-induced alkalinization of peptide nucleic acid-positive (PNA) cells following Na-freeexposure at resting intracellular pH (pHi) A representative trace demonstrating inhibition of cell alkalinization by EIPA (100 M) B quantitative analysis ofcell alkalinization in the absence and presence of EIPA (62 inhibition) Values are means SE (n 19 cells) P 0001 (by paired t-test) C representativetrace demonstrating lack of effect on cell alkalinization in the presence of DAPI (1 M) D quantitative analysis of cell alkalinization in the absence and presenceof DAPI Values are means SE (n 22) P 005 (by paired t-test)
C259INVOLVEMENT OF ASICs IN FISH GILL Na TRANSPORT
AJP-Cell Physiol bull doi101152ajpcell003982013 bull wwwajpcellorg
Phylogenetic analysis and tissue distribution of ASIC1 andASIC4 Two distinct DNA fragments whose derived aminoacid sequences exhibit similarity to the ASIC family werecloned from the gill of rainbow trout To determine theiridentity we conducted a phylogenetic analysis by comparisonwith known ASIC homologs from a variety of species Thegenerated phylogenetic tree (Fig 4) showed that the twocloned rainbow trout ASIC genes (tASICs) were members ofthe ASIC1 and ASIC4 subunits respectively (Fig 4A) ThetASIC1 (partial clone) grouped most closely with ASIC1 fromsalmon (Salmo salar) while tASIC4 grouped with other iden-
tified ASIC4 family members from other fish and mammalspecies
Furthermore we conducted a tissue distribution analysis oftASIC1 and tASIC4 by RT-PCR We observed mRNA expres-sion of tASIC1 in all tested tissues gill MRCs head and trunkkidney brain and blood tASIC4 had a slightly differentdistribution it was present in the perfused gill MRCs brainand trunk kidney but it was absent in head kidney and bloodtissues
Immunoprecipitation and immunostaining with anti-zASIC42To determine if ASIC4 was present in the fish gill and to validate our
Fig 4 Phylogenetic analysis of amino acid sequences of cloned rainbow trout acid-sensing ion channels ASIC1 and ASIC4 and ASICs from various fish speciesPhylogenetic tree was constructed using the maximum-likelihood method and numbers indicate bootstrap values for 700 resamplings GenBank or Ensemblaccession numbers are as follows human ASIC2 NP_0010852 Western clawed frog ASIC2 XP_0029330862 Japanese pufferfish ASIC2 XP_0039648711zebrafish ASIC2 NP_9999531 three-spined stickleback ASIC2 ENSGACG00000009558 Atlantic cod ASIC2 ENSGMOT00000019145 spotted greenpufferfish ASIC2 ENSTNIG00000014943 Southern platyfish ASIC2 ENSXMAG00000008283 oyster toadfish ASIC2 AAP456331 coelacanth ASIC2ENSLACG00000007191 rat ASIC2 NP_0370242 Southern platyfish ASIC1c ENSXMAG00000018841 African clawed frog ASIC11 AEC486651 Africanclawed frog ASIC12 AEC486671 brown rat ASIC1 NP_0770681 human ASIC1 NP_0644232 three-spined stickleback ASIC1a ENSGACG00000003075Japanese pufferfish ASIC1 XP_0039635201 Southern platyfish ASIC1a XP_0058018011 Atlantic cod ASIC1a ENSGMOG00000009659 zebrafishASIC12 NP_9999551 rainbow trout ASIC1 KF964646 Atlantic salmon ASIC1 NP_0011334561 oyster toadfish ASIC1 Q7T1N41 elephant shark ASIC1AEC486681 three-spined stickleback AIC1c ENSGACG00000017974 Atlantic cod ASIC1c ENSGMOG00000003105 zebrafish ASIC13 NP_9999541river lamprey ASIC1 AAY289831 sea lamprey ASIC1b ENSPMAG00000003813 dogfish shark ASIC1 AAY289851 zebrafish ASIC11 NP_9999561human ASIC4 NP_8782672 rat ASIC4 NP_0715702 African clawed frog ASIC4 AEC486661 Japanese pufferfish ASIC4 XP_0039621271 spotted greenpufferfish ASIC4b ENSTNIG00000010994 Nile tilapia ASIC4 XP_0034454461 Southern platyfish ASIC4a ENSXMAG00000003624 three-spinedstickleback ASIC4a ENSGACG00000007052 Atlantic cod ASIC4a ENSGMOG00000012740 rainbow trout ASIC4 KF964645 zebrafish ASIC41NP_9999521 zebrafish ASIC42 NP_9999511
C260 INVOLVEMENT OF ASICs IN FISH GILL Na TRANSPORT
AJP-Cell Physiol bull doi101152ajpcell003982013 bull wwwajpcellorg
antibody for later immunocytochemistry we performed an im-munoprecipitation using an antibody raised against zASIC42on the basis of sequence similarly of 60 between zASIC42(aa 146 ndash160 PKSRKGHRPSELQYPP) and the clonedtASIC4 (aa 142ndash156 PKNREGHKPTDLDYPA) Using West-ern blotting we analyzed the precipitates for the presence ofASIC4 (see Fig 6A) and found a single band of 65 kDawhich approximates the predicted size for tASIC4 (604 kDa)The strong band at 55 kDa corresponds to the IgG heavychain
To determine the location of ASIC4 protein within therainbow trout gill tissue fixed sections were double-labeledwith anti-zASIC42 antibody (see Fig 6B) and anti-NKA andthen counterstained with the nuclear stain DAPI Immunore-activity was observed in cells located in the lamellae andinterlamellar region (see Fig 6B arrowhead) Moreover themajority of cells positive for zASIC42 also colocalized withNKA protein (Fig 6 C and D arrowhead) a known marker ofMRCs suggesting that in gill epithelium ASIC4 protein ismainly expressed in MRCs SEM analysis of the gill filamentdemonstrated that MRCs are clearly distinguished on the sur-face of the epithelium and have a broad apical convex openingto the environment (see Fig 7A arrowheads) The generatedthree-dimensional reconstructions of the staining immunoreac-tivity pattern for ASIC4 and NKA proteins revealed the pres-ence of ASIC4 and NKA in the same cells but in distinctregions of the cells NKA was present on the basolateralmembrane (arrows green staining) while ASIC4 was localizedto the broad apical surface of the cell (arrowheads red stain-ing) Figure 7B shows a generated image with a top view whileFig 7C is the same cell viewed from the side
DISCUSSION
The existence of an apical Na channel electrochemicallycoupled to a VHA mediating Na uptake from dilute FW hasbeen postulated on theoretical thermodynamic grounds for anumber of decades (1 36 34) However an inability todetermine the molecular identity of the Na channel has beena major drawback in the general acceptance of the modelUsing a combination of pharmacological blockade isotopicfluxes immunocytochemistry and molecular biology we pro-pose that an ASIC family member is potentially the long-sought-after apical Na channel responsible for Na uptake byrainbow trout acclimatized to water with low Na and low pHOur study provides the first evidence for a nonneuronal role forASICs in any vertebrate whereby in gills from FW fishesASICs are associated with the MRCs and tASIC4 localizes tothe apical membrane of NKA positive cells
ASICs are a broad family of Na-specific ion channels thathave been extensively studied in mammalian models (forreview see Ref 23) with only a few studies performed in anyfish species (35 42 46) ASICs have primarily been associatedwith neuronal tissues with studies focusing on a neuronalfunction for ASICs (23) In this study ASIC1 and ASIC4 werecloned in rainbow trout using PCR and they were expressed ina variety of tissues both neuronal and nonneuronal in origin(Fig 5) Importantly tASIC1 gene was present in all fiveinvestigated tissues including whole blood It appears that thetASIC4 gene has a more defined tissue distribution thantASIC1 with expression in the ionoregulatory tissues (trunk
kidney and perfused gill) and the brain Unlike tASIC1 therewas no apparent expression of tASIC4 in the head kidney orthe blood Therefore tASIC4 expression in MRCs and per-fused gill could not be attributed to blood contamination
Both tASIC1 and tASIC4 genes were expressed in the gillepithelium and more specifically were present in MRCs ofrainbow trout gills It is believed that similar to ENaCsfunctional ASICs are trimers (20) although their configurationhas not been resolved It has been demonstrated that with afew exceptions ASICs function as homomers (forming chan-nels of identical subunits) or heteromers (forming channelswith other ASIC subunits) (4 23) ASICs have been charac-terized by using oocyte expression in zebrafish (4 35) anddogfish shark (ASIC1b only) (42) It has been found thatcoexpression of zASIC41 with zASIC13 in Xenopus oocytesincreased the current amplitude and abundance (15-fold) ofthe channel at the cell surface indicating that these two ASICsubunits form a functional channel (4) Moreover thisASIC4113 heteromeric channel was more efficiently traf-ficked to the plasma membrane and had increased affinity forH Additionally zASIC41 when heterologously expressedin Xenopus oocytes has been shown to be also gated open bydecreases in extracellular Ca2 The signal responsible forgating of zASIC42 was unable to be determined in that study(4) It is therefore possible that tASIC1 and tASIC4 form aheteromeric channel in MRCs but this cannot be determined atthis time and would require confirmation by immunohisto-chemistry with an anti-ASIC1 antibody that is immunoreactivefor rainbow trout It is also possible that other ASIC subunitsandor regulatory elements may be involved in the function ofASICs in the FW trout gill A full characterization of tASIC4by oocyte expression and electrophysiology is required tounderstand the mechanisms of gating in low-Na low-Ca2
watersIn our study ASIC4 protein was colocalized with NKA and
was present on the apical surface of cells suggesting that it isfound primarily in MRCs (Fig 6 D and E) Previous studies onASICs in fishes have suggested that they are primarily associ-ated with neuronal tissue and sensory cells (27 35 42 46) Inadult and developing zebrafish morphological studies usingconfocal immunofluorescence techniques demonstrated thatgill MRCs come into direct contact with nerve fibers in thebasolateral region (21 22) However in our study ASIC4protein was clearly localized to the apical region of the MRCsas demonstrated by three-dimensional reconstruction of con-
Head k
idney
ASIC4
ASIC1
GAPDH
Gill MRCs
Brain
Trunk k
idney
Blood
No tem
plate
Fig 5 Tissue distribution analysis of ASIC4 and ASIC1 genes in adultrainbow trout as determined by PCR GAPDH was used as a reference geneMRCs mitochondrion-rich cells
C261INVOLVEMENT OF ASICs IN FISH GILL Na TRANSPORT
AJP-Cell Physiol bull doi101152ajpcell003982013 bull wwwajpcellorg
focal images ASIC4 immunoreactivity did not overlap withthe standard basolateral immunoreactive pattern associatedwith NKA (Fig 7 B and C) Therefore we conclude thatneuronally derived tissues cannot account for the expression ofASIC4 protein in MRCs We also corroborate the ASIC4staining pattern with the MRC external morphology whichshows that in FW rainbow trout raised in Edmonton tap water(500 M Na) the MRCs have wide apical openings (Fig7A) This apical localization of ASIC4 protein in the cells of atransporting epithelium is the first evidence for a nonneuronalfunction for any ASIC member and suggests that ASIC4 as aNa channel may be responsible for apical uptake of Na inFW fish gill
Pharmacological profiling has long been used to differentiatethe mechanisms of ion transport across epithelial tissues For Na
transport classical analysis has usually involved amiloride and itssubstituted derivatives (eg EIPA mercaptoisobutyric acid andphenamil) as a means to differentiate between the involvementof NHE isoforms and ENaC in Na transport (25) We inves-tigated two newly described ASIC-specific inhibitors dimina-zene and DAPI (5) for their potency in inhibiting unidirec-tional Na uptake in fish Diminazene and DAPI exhibited adose-dependent inhibitory relationship and both were veryeffective at low concentrations (Fig 1) Na uptake in rainbow
trout acclimatized to low-ionic-strength acidic water wasclearly inhibited by these diarylamidines at concentrationssimilar to those reported in a previous study of cultured rathippocampal neurons (5) However in our study DAPI provedto be more effective than diminazene in contrast to observa-tions in rat neurons The inhibition of 22Na uptake by DAPIand diminazene was also compared with other common Na
transport inhibitors amiloride and phenamil Amiloride wasobserved to be an effective inhibitor of Na uptake in rainbowtrout but only at relatively high concentrations (500 M)which is consistent with previous studies (2 37) Recentlyamiloride was also demonstrated to completely block ASIC-mediated currents (5 35 42) however it is also known toblock many other Na transporters including NHE and ENaC(25) Amiloride therefore cannot be used as a distinguishingpharmacological agent
While DAPI diminazene and amiloride almost completelyblocked Na transport in the present study phenamil had nosignificant effect on Na flux in rainbow trout This suggeststhat phenamil is a poor inhibitor of Na uptake in troutexposed to low-ionic-strength water (30 moll Na) Paststudies of the effects of phenamil on Na uptake in a variety ofFW fishes have reported inconsistent results For example inneon tetras acclimatized to low Na (50 moll) phenamil had
Fig 6 Immunoreactivity of anti-zebrafish ASIC42 (zASIC42) antibody in the gill of rainbow trout A immunoprecipitation Western blot from whole gillhomogenates with zASIC42 showing a distinct band of 65 kDa Lane 1 (L1) contains 300 mg of gill tissue and lane 2 (L2) contains 200 mg of gill tissueBndashE confocal images of gill sections labeled for ASIC4 and Na-K-ATPase (NKA) proteins B and C double-labeling with DAPI and anti-zASIC42 (B) andDAPI and anti-NKA (C) D merged image of B and C E higher-magnification image of a MRC showing that ASIC4 and NKA are present in distinct regionswithin the MRC F control micrograph with DAPI and no primary antibody Arrowheads (BndashD) indicate positive staining
C262 INVOLVEMENT OF ASICs IN FISH GILL Na TRANSPORT
AJP-Cell Physiol bull doi101152ajpcell003982013 bull wwwajpcellorg
no effect on Na transport (38) whereas it caused 70reduction in Na influx in Cyprinidon variegatus hubbsi (100moll Na) and 60 reduction in goldfish (300 moll Na)(2 38) These conflicting results from various phenamil studiessuggest that sensitivity to phenamil might be species- andorenvironment-specific Collectively the pharmacological anal-ysis suggests that the Na uptake in FW rainbow trout is mostlikely not mediated by ENaC given the lack of inhibition byphenamil but may be attributed to ASIC since Na uptakewas almost completely blocked by DAPI and diminazene bothof which inhibit ASICs but not ENaC
In our Na flux study with various pharmacological agentswe made an interesting observation that inhibition of 22Na
uptake was less pronounced for all tested inhibitors during thesecond consecutive measurement (90ndash180 min) (Fig 2) Toour knowledge this phenomenon where potency of the testeddrug diminishes over time has not been reported in anyprevious study on fish This observation may be explained bya decreased stabilityefficacy of the tested pharmacologicalagents in aqueous media metabolism of the drug by theanimal or activation of compensatory Na uptake mechanismsnot sensitive to amiloride DAPI or diminazene We believethat the decrease of the potency of amiloride DAPI anddiminazene over time demonstrates the importance of properexperimental design and choice of period of study when ionfluxes are examined using pharmacological inhibitors
While the two novel ASIC inhibitors have been tested fortheir potency in inhibiting ASICs and ENaC in neurons (5)they have not been tested for their potential effects on otherNa transporters implicated in Na uptake in fishes mostnotably the NHEs To verify that the Na uptake inhibitioncaused by DAPI and diminazene in rainbow trout was due toinhibition of ASICs rather than NHEs we tested the effect ofDAPI on MRCs isolated from the rainbow trout gill epithe-lium Our laboratory previously showed that rainbow troutpossess two populations of MRCs peanut lectin agglutinin(PNA) positive (PNA) and PNA negative (PNA) MRCs(10) It has been demonstrated that NHE3 and NHE2 areexpressed in PNA cells isolated from branchial epithelium of
FW rainbow trout (19) Therefore we investigated the effect ofDAPI on PNA cells and compared it with the effect of EIPAa potent and highly selective inhibitor of NHE (25) Earlierstudies in our laboratory using cell imaging determined thatPNA cells undergo NHE-dependent alkalinization (32 33)Here we demonstrate that while EIPA was effective at inhib-iting NHE function in PNA cells (Fig 3A) there was noeffect of high concentrations of DAPI on Na-dependent pHi
elevation demonstrating that NHE was not inhibited by DAPI(Fig 3B) Therefore the measured inhibition of Na uptake byDAPI that we see in whole animals (Fig 1) cannot be attrib-uted to inhibition of NHE
On the basis of the mRNA expression of tASIC1 andtASIC4 genes in the gill epithelium and isolated MRCs local-ization of ASIC4 protein to the apical region of the MRCs and
Fig 7 Three-dimensional views of freshwater(FW) rainbow trout MRCs A scanning elec-tron micrograph of FW rainbow trout gillepithelium showing broad apical openings ofMRCs (arrowheads) and mucous cells () Band C 3-dimensional reconstruction of z-stack images showing apical localization ofASIC4 protein (red arrowhead) and basolat-eral localization of NKA (green arrow)B top view C side view
PNA- MRC
Na+
ASIC
H+
VHA
NKA
Na+
K+
NBC
Na+ HCO3-
PNA+ MRC
NH3
Na+
H+
NHE Rhcg2
NKA
Na+
K+
Fig 8 Model for Na uptake in PNA and PNA MRCs in gill epithelium ofrainbow trout showing placement of proposed ASIC-mediated Na uptakemechanisms Rhcg2 rhesus glycoprotein NHE NaH exchanger VHAvacuolar H-ATPase NBC Na-HCO3
cotransporter
C263INVOLVEMENT OF ASICs IN FISH GILL Na TRANSPORT
AJP-Cell Physiol bull doi101152ajpcell003982013 bull wwwajpcellorg
pharmacological blockade of Na influx by ASIC inhibitors inadult rainbow trout we propose a revised model for transepi-thelial Na uptake in FW rainbow trout gills (Fig 8) wherebyone apical mode of Na entry is via an ASIC4-mediatedmechanism
A number of recent studies on Na uptake in FW fishes havedemonstrated the presence of the NHERh metabolon in thegills of various species (17 30 31) As mentioned previouslywe specifically targeted the rainbow trout as a model for thisstudy as it is native to oligotrophic Na-poor mountainstreams with low buffering capacity This type of environmentpresents key thermodynamic challenges to the NHERhmetabolon as Na restrictions cannot be alleviated by thepresence of an Rh protein (6) Moreover searches of rainbowtrout genomes and transcriptomes have failed to identify anyENaC subunits ( 13 or ) despite the fact that they are knownto actively take up Na from water with 50 M Na (49)Furthermore other research studies have suggested and dem-onstrated an apical localization of the VHA in the gills ofrainbow trout (44 48) and bafilomycin a VHA-specific in-hibitor has been demonstrated to reduce Na uptake in ze-brafish (2) tilapia and carp (9) Previous studies have pro-posed VHA to be a part of the NHERh metabolon (41 50 51)however the placement of an apically oriented VHA is coun-terintuitive to the function of an NHE as it would exacerbatethe noted thermodynamic constraints Placement of a VHA inconcert with an apical ASIC is more plausible since it wouldnot only facilitate Na uptake from very low ionic strengthwater but could also provide a mechanism for gating ofASIC-mediated Na conductance It remains to be determinedif ASIC and VHA proteins colocalize to the same type ofMRCs and this warrants further investigation
Perspectives In summary we propose that ASCI4 is in-volved in mediating Na uptake from FW in rainbow troutacclimatized to low ionic strength low pH media This is thefirst demonstration of a nonneuronal function for any ASICfamily member and simultaneously provides an attractive so-lution to the debate surrounding the thermodynamic limitationsof the other proposed Na transport mechanisms and resolvesthe conundrum of the role of an apical VHA in Na transportThe possibility that other fish models may use similar mecha-nisms under different environmental conditions where modelssuch as the NHERh metabolon cannot be expected to functionremains to be explored
ACKNOWLEDGMENTS
We thank the staff in the Biosciences Aquatic Facilities the MolecularBiology Service Unit and the Cross Cancer Institute Cell Imaging Facility atthe University of Alberta for outstanding help throughout the study We alsothank Alex Clifford for advice and help with phylogenetic analysis and JamesEde for help with confocal microscopy
GRANTS
This study was supported by a Natural Sciences and Engineering ResearchCouncil of Canada Discovery Grant to G G Goss and an Alberta IngenuityStudent Scholarship to A K Dymowska
DISCLOSURES
No conflicts of interest financial or otherwise are declared by the authors
AUTHOR CONTRIBUTIONS
AKD AGS and GGG are responsible for conception and design ofthe research AKD AGS SDB and DC performed the experiments
AKD AGS SDB DC and GGG analyzed the data AKD AGSSDB DC and GGG interpreted the results of the experiments AKDAGS SDB and GGG prepared the figures AKD AGS SDB DCand GGG drafted the manuscript AKD AGS SDB DC and GGGedited and revised the manuscript AKD AGS SDB and GGGapproved the final version of the manuscript
REFERENCES
1 Avella M Bornancin M A new analysis of ammonia and sodiumtransport through the gills of the freshwater rainbow trout (Salmo gaird-neri) J Exp Biol 142 155ndash175 1989
2 Boisen AM Amstrup J Novak I Grosell M Sodium and chloridetransport in soft water and hard water acclimatized zebrafish (Daniorerio) Biochim Biophys Acta 1618 207ndash218 2003
3 Brix KV Grosell M Comparative characterization of Na transport inCyprinodon variegatus variegatus and Cyprinodon variegatus hubbsi amodel species complex for studying teleost invasion of freshwater J ExpBiol 215 1199ndash1209 2012
4 Chen X Polleichtner G Kadurin I Grunder S Zebrafish acid-sensingion channel (ASIC) 4 Characterization of homo- and heteromeric chan-nels and identification of regions important for activation by H J BiolChem 282 30406ndash30413 2007
5 Chen X Qiu L Li M Durrnagel S Orser BA Xiong ZG MacDonaldJF Diarylamidines high potency inhibitors of acid-sensing ion channelsNeuropharmacology 58 1045ndash1053 2010
6 Dymowska AK Hwang PP Goss GG Structure and function of iono-cytes in the freshwater fish gill Respir Physiol Neurobiol 184 282ndash2922012
7 Edwards SL Tse CM Toop T Immunolocalization of NHE3-likeimmunoreactivity in the gills of the rainbow trout (Oncorhynchus mykiss)and the blue-throated wrasse (Pseudolabrus tetrious) J Anat 195 465ndash469 1999
8 Evans DH Piermarini PM Choe KP The multifunctional gill domi-nant site of gas exchange osmoregulation acid-base regulation andexcretion of nitrogenous waste Physiol Rev 85 97ndash177 2005
9 Fenwick JC Bonga SE Flik G In vivo bafilomycin-sensitive Na
uptake in young freshwater fish J Exp Biol 202 3659ndash3666 199910 Galvez F Reid SD Hawkings G Goss GG Isolation and characteriza-
tion of mitochondria-rich cell types from the gill of freshwater rainbowtrout Am J Physiol Regul Integr Comp Physiol 282 R658ndashR668 2002
11 Goss G Orlowski J Grinstein S Coimmunoprecipitation of a 24-kDaprotein with NHE1 the ubiquitous isoform of the NaH exchanger AmJ Physiol Cell Physiol 270 C1493ndashC1502 1996
12 Goss GG Wood CJ Na and Cl uptake kinetics diffusive effluxes andacidic equivalent fluxes across the gills of rainbow trout 1 Responses toenvironmental hyperoxia J Exp Biol 152 521ndash547 1990
13 Grunder S Geissler HS Bassler EL Ruppersberg JP A new memberof acid-sensing ion channels from pituitary gland Neuroreport 11 1607ndash1611 2000
14 Harvey BJ Energization of sodium absorption by the H-ATPase pumpin mitochondria-rich cells of frog skin J Exp Biol 172 289ndash309 1992
15 Hirata T Kaneko T Ono T Nakazato T Furukawa N Hasegawa SWakabayashi S Shigekawa M Chang MH Romero MF Hirose SMechanism of acid adaptation of a fish living in low pH 35 lake Am JPhysiol Regul Integr Comp Physiol 284 R1199ndashR1212 2003
16 Holzer P Acid-sensitive ion channels and receptors Hand Exp Pharma-col 194 283ndash332 2009
17 Hung CY Tsui KN Wilson JM Nawata CM Wood CM Wright PARhesus glycoprotein gene expression in the mangrove killifish Kryptole-bias marmoratus exposed to elevated environmental ammonia levels andair J Exp Biol 210 2419ndash2429 2007
18 Hwang PP Lee TH Lin LY Ion regulations in fish gills recentprogresses in the cellular and molecular mechanisms Am J Physiol RegulIntegr Comp Physiol 301 R28ndashR47 2011
19 Ivanis G Esbaugh AJ Perry SF Branchial expression and localizationof SLC9A2 and SLC9A3 sodiumhydrogen exchangers and their possiblerole in acid-base regulation in freshwater rainbow trout (Oncorhynchusmykiss) J Exp Biol 211 2467ndash2477 2008
20 Jasti J Furukawa H Gonzales EB Gouaux E Structure of acid-sensingion channel 1 at 19A resolution and low pH Nature 449 316ndash323 2007
21 Jonz MG Nurse CA Epithelial mitochondria-rich cells and associatedinnervation in adult and developing zebrafish J Comp Neurol 497817ndash832 2006
C264 INVOLVEMENT OF ASICs IN FISH GILL Na TRANSPORT
AJP-Cell Physiol bull doi101152ajpcell003982013 bull wwwajpcellorg
22 Jonz MG Nurse CA New developments on gill innervation insightsfrom a model vertebrate J Exp Biol 211 2371ndash2378 2008
23 Kellenberger S Epithelial sodium and acid-sensing ion channels InSensing With Ion Channels edited by Martinac B New York Springer2008 chapt 11 p 225ndash246
24 Kellenberger S Schild L Epithelial sodium channeldegenerin family ofion channels a variety of functions for a shared structure Physiol Rev 82735ndash767 2002
25 Kleyman TR Cragoe EJ Amiloride and its analogs as tools in the studyof ion-transport J Membr Biol 105 1ndash21 1988
26 Krogh A The active absorption of ions in some freshwater animals ZVergl Physiol 25 335ndash350 1938
27 Levanti MB Guerrera MC Calavia MG Ciriaco E Montalbano GCobo J Germana A Vega JA Acid-sensing ion channel 2 (ASIC2) inthe intestine of adult zebrafish Neurosci Lett 494 24ndash28 2011
28 Lin H Randall DJ Evidence for the presence of an electrogenic protonpump on the trout gill epithelium J Exp Biol 161 119ndash134 1991
29 Maetz J Garcia Romeu F The mechanism of sodium and chlorideuptake by the gills of a fresh-water fish Carassius auratus II Evidencefor NH4
Na and HCO3Cl exchanges J Gen Physiol 47 1209ndash1227
196430 Nakada T Hoshijima K Esaki M Nagayoshi S Kawakami K Hirose
S Localization of ammonia transporter Rhcg1 in mitochondrion rich cellsof yolk sac gill and kidney of zebrafish and its ionic strength-dependentexpression Am J Physiol Regul Integr Comp Physiol 293 R1743ndashR17532007
31 Nawata CM Hung CC Tsui TK Wilson JM Wright PA Wood CMAmmonia excretion in rainbow trout (Oncorhynchus mykiss) evidence forRh glycoprotein and H-ATPase involvement Physiol Genomics 31463ndash474 2007
32 Parks SK Tresguerres M Galvez F Goss GG Intracellular pH regu-lation in isolated trout gill mitochondrion-rich (MR) cell subtypes evi-dence for NaH activity Comp Biochem Physiol A 155 139ndash1452010
33 Parks SK Tresguerres M Goss GG Interactions between Na channelsand Na-HCO3
cotransporters in the freshwater fish gill MRC a modelfor transepithelial Na uptake Am J Physiol Cell Physiol 292 C935ndashC944 2007
34 Parks SK Tresguerres M Goss GG Theoretical considerations under-lying Na uptake mechanisms in freshwater fishes Comp BiochemPhysiol C 148 411ndash418 2008
35 Paukert M Sidi S Russell C Siba M Wilson SW A family ofacid-sensing ion channels from zebrafish widespread expression in thecentral nervous system suggests a conserved role in neuronal communi-cation J Biol Chem 279 18783ndash18791 2004
36 Perry SF The chloride cell structure and function in the gill of freshwaterfish Annu Rev Physiol 59 325ndash347 1997
37 Perry SF Randall DJ Effects of amiloride and SITS on branchial ionfluxes in rainbow trout Salmo gairdneri J Exp Zool 215 225ndash228 1981
38 Preest MR Gonzales RJ Wilson RW Pharmacological examination ofNa and Cl transport in two species of freshwater fish Physiol BiochemZool 78 259ndash272 2005
39 Reid SD Hawkings GS Galvez F Goss GG Localization and charac-terization of phenamil-sensitive Na influx in isolated rainbow trout gillepithelial cells J Exp Biol 206 551ndash559 2003
40 Sakai H Lingueglia E Champigny G Mattei MG Lazdunski MCloning and functional expression of a novel degenerin-like Na channelgene in mammals J Physiol 519 323ndash333 1999
41 Shih TH Horng JL Liu ST Hwang PP Lin LY Rhcg1 and NHE3b areinvolved in ammonium-dependent sodium uptake by zebrafish larvaeacclimated to low-sodium water Am J Physiol Regul Integr Comp Physiol302 R84ndashR93 2012
42 Springauf A Grunder S An acid-sensing ion channel from shark(Squalus acanthias) mediates transient and sustained responses to protonsJ Physiol 588 809ndash820 2010
43 Spry DJ Wood CM Ion flux rates acid-base status and blood gases inrainbow trout Salmo gairdneri exposed to toxic zinc in natural soft waterCan J Fish Aquat Sci 42 1332ndash1337 1985
44 Sullivan GV Fryer JN Perry SF Immunolocalization of proton pumps(H-ATPase) in pavement cells of rainbow trout gill J Exp Biol 1982619ndash2629 1996
45 Tresguerres M Parks SK Katoh F Goss GG Microtubule-dependentrelocation of branchial V-H-ATPase to the basolateral membrane in thePacific spiny dogfish (Squalus acanthias) a role in base secretion J ExpBiol 209 599ndash609 2006
46 Vina E Parisi V Cabo R Laura R Lopez-Velasco S Lopez-Muniz AGarcia-Suarez O Germana A Vega JA Acid-sensing ion channels(ASICs) in the taste buds of adult zebrafish Neurosci Lett 536 35ndash402013
47 Waldmann R Lazdunski M H-gated cation channels neuronal acidsensors in the NaCDEG family of ion channels Curr Opin Neurobiol 8418ndash424 1998
48 Wilson JM Laurent P Tufts BL Benos DJ Donowitz M Vogl AWRandall DJ NaCl uptake by the branchial epithelium in freshwater teleostfish an immunological approach to ion-transport protein localization JExp Biol 203 2279ndash2296 2000
49 Wood CM Randall DJ Sodium balance in rainbow-trout (Salmo-gaird-neri) during extended exercise J Comp Physiol 82 235ndash256 1973
50 Wright PA Wood CM A new paradigm for ammonia excretion inaquatic animals role of rhesus (Rh) glycoproteins J Exp Biol 2122303ndash2312 2009
51 Yan JJ Chou MY Kaneko T Hwang PP Gene expression of NaH
exchanger in zebrafish H-ATPase-rich cells during acclimation to low-Na and acidic environments Am J Physiol Cell Physiol 293 C1814ndashC1823 2007
C265INVOLVEMENT OF ASICs IN FISH GILL Na TRANSPORT
AJP-Cell Physiol bull doi101152ajpcell003982013 bull wwwajpcellorg

Phylogenetic analysis and tissue distribution of ASIC1 andASIC4 Two distinct DNA fragments whose derived aminoacid sequences exhibit similarity to the ASIC family werecloned from the gill of rainbow trout To determine theiridentity we conducted a phylogenetic analysis by comparisonwith known ASIC homologs from a variety of species Thegenerated phylogenetic tree (Fig 4) showed that the twocloned rainbow trout ASIC genes (tASICs) were members ofthe ASIC1 and ASIC4 subunits respectively (Fig 4A) ThetASIC1 (partial clone) grouped most closely with ASIC1 fromsalmon (Salmo salar) while tASIC4 grouped with other iden-
tified ASIC4 family members from other fish and mammalspecies
Furthermore we conducted a tissue distribution analysis oftASIC1 and tASIC4 by RT-PCR We observed mRNA expres-sion of tASIC1 in all tested tissues gill MRCs head and trunkkidney brain and blood tASIC4 had a slightly differentdistribution it was present in the perfused gill MRCs brainand trunk kidney but it was absent in head kidney and bloodtissues
Immunoprecipitation and immunostaining with anti-zASIC42To determine if ASIC4 was present in the fish gill and to validate our
Fig 4 Phylogenetic analysis of amino acid sequences of cloned rainbow trout acid-sensing ion channels ASIC1 and ASIC4 and ASICs from various fish speciesPhylogenetic tree was constructed using the maximum-likelihood method and numbers indicate bootstrap values for 700 resamplings GenBank or Ensemblaccession numbers are as follows human ASIC2 NP_0010852 Western clawed frog ASIC2 XP_0029330862 Japanese pufferfish ASIC2 XP_0039648711zebrafish ASIC2 NP_9999531 three-spined stickleback ASIC2 ENSGACG00000009558 Atlantic cod ASIC2 ENSGMOT00000019145 spotted greenpufferfish ASIC2 ENSTNIG00000014943 Southern platyfish ASIC2 ENSXMAG00000008283 oyster toadfish ASIC2 AAP456331 coelacanth ASIC2ENSLACG00000007191 rat ASIC2 NP_0370242 Southern platyfish ASIC1c ENSXMAG00000018841 African clawed frog ASIC11 AEC486651 Africanclawed frog ASIC12 AEC486671 brown rat ASIC1 NP_0770681 human ASIC1 NP_0644232 three-spined stickleback ASIC1a ENSGACG00000003075Japanese pufferfish ASIC1 XP_0039635201 Southern platyfish ASIC1a XP_0058018011 Atlantic cod ASIC1a ENSGMOG00000009659 zebrafishASIC12 NP_9999551 rainbow trout ASIC1 KF964646 Atlantic salmon ASIC1 NP_0011334561 oyster toadfish ASIC1 Q7T1N41 elephant shark ASIC1AEC486681 three-spined stickleback AIC1c ENSGACG00000017974 Atlantic cod ASIC1c ENSGMOG00000003105 zebrafish ASIC13 NP_9999541river lamprey ASIC1 AAY289831 sea lamprey ASIC1b ENSPMAG00000003813 dogfish shark ASIC1 AAY289851 zebrafish ASIC11 NP_9999561human ASIC4 NP_8782672 rat ASIC4 NP_0715702 African clawed frog ASIC4 AEC486661 Japanese pufferfish ASIC4 XP_0039621271 spotted greenpufferfish ASIC4b ENSTNIG00000010994 Nile tilapia ASIC4 XP_0034454461 Southern platyfish ASIC4a ENSXMAG00000003624 three-spinedstickleback ASIC4a ENSGACG00000007052 Atlantic cod ASIC4a ENSGMOG00000012740 rainbow trout ASIC4 KF964645 zebrafish ASIC41NP_9999521 zebrafish ASIC42 NP_9999511
C260 INVOLVEMENT OF ASICs IN FISH GILL Na TRANSPORT
AJP-Cell Physiol bull doi101152ajpcell003982013 bull wwwajpcellorg
antibody for later immunocytochemistry we performed an im-munoprecipitation using an antibody raised against zASIC42on the basis of sequence similarly of 60 between zASIC42(aa 146 ndash160 PKSRKGHRPSELQYPP) and the clonedtASIC4 (aa 142ndash156 PKNREGHKPTDLDYPA) Using West-ern blotting we analyzed the precipitates for the presence ofASIC4 (see Fig 6A) and found a single band of 65 kDawhich approximates the predicted size for tASIC4 (604 kDa)The strong band at 55 kDa corresponds to the IgG heavychain
To determine the location of ASIC4 protein within therainbow trout gill tissue fixed sections were double-labeledwith anti-zASIC42 antibody (see Fig 6B) and anti-NKA andthen counterstained with the nuclear stain DAPI Immunore-activity was observed in cells located in the lamellae andinterlamellar region (see Fig 6B arrowhead) Moreover themajority of cells positive for zASIC42 also colocalized withNKA protein (Fig 6 C and D arrowhead) a known marker ofMRCs suggesting that in gill epithelium ASIC4 protein ismainly expressed in MRCs SEM analysis of the gill filamentdemonstrated that MRCs are clearly distinguished on the sur-face of the epithelium and have a broad apical convex openingto the environment (see Fig 7A arrowheads) The generatedthree-dimensional reconstructions of the staining immunoreac-tivity pattern for ASIC4 and NKA proteins revealed the pres-ence of ASIC4 and NKA in the same cells but in distinctregions of the cells NKA was present on the basolateralmembrane (arrows green staining) while ASIC4 was localizedto the broad apical surface of the cell (arrowheads red stain-ing) Figure 7B shows a generated image with a top view whileFig 7C is the same cell viewed from the side
DISCUSSION
The existence of an apical Na channel electrochemicallycoupled to a VHA mediating Na uptake from dilute FW hasbeen postulated on theoretical thermodynamic grounds for anumber of decades (1 36 34) However an inability todetermine the molecular identity of the Na channel has beena major drawback in the general acceptance of the modelUsing a combination of pharmacological blockade isotopicfluxes immunocytochemistry and molecular biology we pro-pose that an ASIC family member is potentially the long-sought-after apical Na channel responsible for Na uptake byrainbow trout acclimatized to water with low Na and low pHOur study provides the first evidence for a nonneuronal role forASICs in any vertebrate whereby in gills from FW fishesASICs are associated with the MRCs and tASIC4 localizes tothe apical membrane of NKA positive cells
ASICs are a broad family of Na-specific ion channels thathave been extensively studied in mammalian models (forreview see Ref 23) with only a few studies performed in anyfish species (35 42 46) ASICs have primarily been associatedwith neuronal tissues with studies focusing on a neuronalfunction for ASICs (23) In this study ASIC1 and ASIC4 werecloned in rainbow trout using PCR and they were expressed ina variety of tissues both neuronal and nonneuronal in origin(Fig 5) Importantly tASIC1 gene was present in all fiveinvestigated tissues including whole blood It appears that thetASIC4 gene has a more defined tissue distribution thantASIC1 with expression in the ionoregulatory tissues (trunk
kidney and perfused gill) and the brain Unlike tASIC1 therewas no apparent expression of tASIC4 in the head kidney orthe blood Therefore tASIC4 expression in MRCs and per-fused gill could not be attributed to blood contamination
Both tASIC1 and tASIC4 genes were expressed in the gillepithelium and more specifically were present in MRCs ofrainbow trout gills It is believed that similar to ENaCsfunctional ASICs are trimers (20) although their configurationhas not been resolved It has been demonstrated that with afew exceptions ASICs function as homomers (forming chan-nels of identical subunits) or heteromers (forming channelswith other ASIC subunits) (4 23) ASICs have been charac-terized by using oocyte expression in zebrafish (4 35) anddogfish shark (ASIC1b only) (42) It has been found thatcoexpression of zASIC41 with zASIC13 in Xenopus oocytesincreased the current amplitude and abundance (15-fold) ofthe channel at the cell surface indicating that these two ASICsubunits form a functional channel (4) Moreover thisASIC4113 heteromeric channel was more efficiently traf-ficked to the plasma membrane and had increased affinity forH Additionally zASIC41 when heterologously expressedin Xenopus oocytes has been shown to be also gated open bydecreases in extracellular Ca2 The signal responsible forgating of zASIC42 was unable to be determined in that study(4) It is therefore possible that tASIC1 and tASIC4 form aheteromeric channel in MRCs but this cannot be determined atthis time and would require confirmation by immunohisto-chemistry with an anti-ASIC1 antibody that is immunoreactivefor rainbow trout It is also possible that other ASIC subunitsandor regulatory elements may be involved in the function ofASICs in the FW trout gill A full characterization of tASIC4by oocyte expression and electrophysiology is required tounderstand the mechanisms of gating in low-Na low-Ca2
watersIn our study ASIC4 protein was colocalized with NKA and
was present on the apical surface of cells suggesting that it isfound primarily in MRCs (Fig 6 D and E) Previous studies onASICs in fishes have suggested that they are primarily associ-ated with neuronal tissue and sensory cells (27 35 42 46) Inadult and developing zebrafish morphological studies usingconfocal immunofluorescence techniques demonstrated thatgill MRCs come into direct contact with nerve fibers in thebasolateral region (21 22) However in our study ASIC4protein was clearly localized to the apical region of the MRCsas demonstrated by three-dimensional reconstruction of con-
Head k
idney
ASIC4
ASIC1
GAPDH
Gill MRCs
Brain
Trunk k
idney
Blood
No tem
plate
Fig 5 Tissue distribution analysis of ASIC4 and ASIC1 genes in adultrainbow trout as determined by PCR GAPDH was used as a reference geneMRCs mitochondrion-rich cells
C261INVOLVEMENT OF ASICs IN FISH GILL Na TRANSPORT
AJP-Cell Physiol bull doi101152ajpcell003982013 bull wwwajpcellorg
focal images ASIC4 immunoreactivity did not overlap withthe standard basolateral immunoreactive pattern associatedwith NKA (Fig 7 B and C) Therefore we conclude thatneuronally derived tissues cannot account for the expression ofASIC4 protein in MRCs We also corroborate the ASIC4staining pattern with the MRC external morphology whichshows that in FW rainbow trout raised in Edmonton tap water(500 M Na) the MRCs have wide apical openings (Fig7A) This apical localization of ASIC4 protein in the cells of atransporting epithelium is the first evidence for a nonneuronalfunction for any ASIC member and suggests that ASIC4 as aNa channel may be responsible for apical uptake of Na inFW fish gill
Pharmacological profiling has long been used to differentiatethe mechanisms of ion transport across epithelial tissues For Na
transport classical analysis has usually involved amiloride and itssubstituted derivatives (eg EIPA mercaptoisobutyric acid andphenamil) as a means to differentiate between the involvementof NHE isoforms and ENaC in Na transport (25) We inves-tigated two newly described ASIC-specific inhibitors dimina-zene and DAPI (5) for their potency in inhibiting unidirec-tional Na uptake in fish Diminazene and DAPI exhibited adose-dependent inhibitory relationship and both were veryeffective at low concentrations (Fig 1) Na uptake in rainbow
trout acclimatized to low-ionic-strength acidic water wasclearly inhibited by these diarylamidines at concentrationssimilar to those reported in a previous study of cultured rathippocampal neurons (5) However in our study DAPI provedto be more effective than diminazene in contrast to observa-tions in rat neurons The inhibition of 22Na uptake by DAPIand diminazene was also compared with other common Na
transport inhibitors amiloride and phenamil Amiloride wasobserved to be an effective inhibitor of Na uptake in rainbowtrout but only at relatively high concentrations (500 M)which is consistent with previous studies (2 37) Recentlyamiloride was also demonstrated to completely block ASIC-mediated currents (5 35 42) however it is also known toblock many other Na transporters including NHE and ENaC(25) Amiloride therefore cannot be used as a distinguishingpharmacological agent
While DAPI diminazene and amiloride almost completelyblocked Na transport in the present study phenamil had nosignificant effect on Na flux in rainbow trout This suggeststhat phenamil is a poor inhibitor of Na uptake in troutexposed to low-ionic-strength water (30 moll Na) Paststudies of the effects of phenamil on Na uptake in a variety ofFW fishes have reported inconsistent results For example inneon tetras acclimatized to low Na (50 moll) phenamil had
Fig 6 Immunoreactivity of anti-zebrafish ASIC42 (zASIC42) antibody in the gill of rainbow trout A immunoprecipitation Western blot from whole gillhomogenates with zASIC42 showing a distinct band of 65 kDa Lane 1 (L1) contains 300 mg of gill tissue and lane 2 (L2) contains 200 mg of gill tissueBndashE confocal images of gill sections labeled for ASIC4 and Na-K-ATPase (NKA) proteins B and C double-labeling with DAPI and anti-zASIC42 (B) andDAPI and anti-NKA (C) D merged image of B and C E higher-magnification image of a MRC showing that ASIC4 and NKA are present in distinct regionswithin the MRC F control micrograph with DAPI and no primary antibody Arrowheads (BndashD) indicate positive staining
C262 INVOLVEMENT OF ASICs IN FISH GILL Na TRANSPORT
AJP-Cell Physiol bull doi101152ajpcell003982013 bull wwwajpcellorg
no effect on Na transport (38) whereas it caused 70reduction in Na influx in Cyprinidon variegatus hubbsi (100moll Na) and 60 reduction in goldfish (300 moll Na)(2 38) These conflicting results from various phenamil studiessuggest that sensitivity to phenamil might be species- andorenvironment-specific Collectively the pharmacological anal-ysis suggests that the Na uptake in FW rainbow trout is mostlikely not mediated by ENaC given the lack of inhibition byphenamil but may be attributed to ASIC since Na uptakewas almost completely blocked by DAPI and diminazene bothof which inhibit ASICs but not ENaC
In our Na flux study with various pharmacological agentswe made an interesting observation that inhibition of 22Na
uptake was less pronounced for all tested inhibitors during thesecond consecutive measurement (90ndash180 min) (Fig 2) Toour knowledge this phenomenon where potency of the testeddrug diminishes over time has not been reported in anyprevious study on fish This observation may be explained bya decreased stabilityefficacy of the tested pharmacologicalagents in aqueous media metabolism of the drug by theanimal or activation of compensatory Na uptake mechanismsnot sensitive to amiloride DAPI or diminazene We believethat the decrease of the potency of amiloride DAPI anddiminazene over time demonstrates the importance of properexperimental design and choice of period of study when ionfluxes are examined using pharmacological inhibitors
While the two novel ASIC inhibitors have been tested fortheir potency in inhibiting ASICs and ENaC in neurons (5)they have not been tested for their potential effects on otherNa transporters implicated in Na uptake in fishes mostnotably the NHEs To verify that the Na uptake inhibitioncaused by DAPI and diminazene in rainbow trout was due toinhibition of ASICs rather than NHEs we tested the effect ofDAPI on MRCs isolated from the rainbow trout gill epithe-lium Our laboratory previously showed that rainbow troutpossess two populations of MRCs peanut lectin agglutinin(PNA) positive (PNA) and PNA negative (PNA) MRCs(10) It has been demonstrated that NHE3 and NHE2 areexpressed in PNA cells isolated from branchial epithelium of
FW rainbow trout (19) Therefore we investigated the effect ofDAPI on PNA cells and compared it with the effect of EIPAa potent and highly selective inhibitor of NHE (25) Earlierstudies in our laboratory using cell imaging determined thatPNA cells undergo NHE-dependent alkalinization (32 33)Here we demonstrate that while EIPA was effective at inhib-iting NHE function in PNA cells (Fig 3A) there was noeffect of high concentrations of DAPI on Na-dependent pHi
elevation demonstrating that NHE was not inhibited by DAPI(Fig 3B) Therefore the measured inhibition of Na uptake byDAPI that we see in whole animals (Fig 1) cannot be attrib-uted to inhibition of NHE
On the basis of the mRNA expression of tASIC1 andtASIC4 genes in the gill epithelium and isolated MRCs local-ization of ASIC4 protein to the apical region of the MRCs and
Fig 7 Three-dimensional views of freshwater(FW) rainbow trout MRCs A scanning elec-tron micrograph of FW rainbow trout gillepithelium showing broad apical openings ofMRCs (arrowheads) and mucous cells () Band C 3-dimensional reconstruction of z-stack images showing apical localization ofASIC4 protein (red arrowhead) and basolat-eral localization of NKA (green arrow)B top view C side view
PNA- MRC
Na+
ASIC
H+
VHA
NKA
Na+
K+
NBC
Na+ HCO3-
PNA+ MRC
NH3
Na+
H+
NHE Rhcg2
NKA
Na+
K+
Fig 8 Model for Na uptake in PNA and PNA MRCs in gill epithelium ofrainbow trout showing placement of proposed ASIC-mediated Na uptakemechanisms Rhcg2 rhesus glycoprotein NHE NaH exchanger VHAvacuolar H-ATPase NBC Na-HCO3
cotransporter
C263INVOLVEMENT OF ASICs IN FISH GILL Na TRANSPORT
AJP-Cell Physiol bull doi101152ajpcell003982013 bull wwwajpcellorg
pharmacological blockade of Na influx by ASIC inhibitors inadult rainbow trout we propose a revised model for transepi-thelial Na uptake in FW rainbow trout gills (Fig 8) wherebyone apical mode of Na entry is via an ASIC4-mediatedmechanism
A number of recent studies on Na uptake in FW fishes havedemonstrated the presence of the NHERh metabolon in thegills of various species (17 30 31) As mentioned previouslywe specifically targeted the rainbow trout as a model for thisstudy as it is native to oligotrophic Na-poor mountainstreams with low buffering capacity This type of environmentpresents key thermodynamic challenges to the NHERhmetabolon as Na restrictions cannot be alleviated by thepresence of an Rh protein (6) Moreover searches of rainbowtrout genomes and transcriptomes have failed to identify anyENaC subunits ( 13 or ) despite the fact that they are knownto actively take up Na from water with 50 M Na (49)Furthermore other research studies have suggested and dem-onstrated an apical localization of the VHA in the gills ofrainbow trout (44 48) and bafilomycin a VHA-specific in-hibitor has been demonstrated to reduce Na uptake in ze-brafish (2) tilapia and carp (9) Previous studies have pro-posed VHA to be a part of the NHERh metabolon (41 50 51)however the placement of an apically oriented VHA is coun-terintuitive to the function of an NHE as it would exacerbatethe noted thermodynamic constraints Placement of a VHA inconcert with an apical ASIC is more plausible since it wouldnot only facilitate Na uptake from very low ionic strengthwater but could also provide a mechanism for gating ofASIC-mediated Na conductance It remains to be determinedif ASIC and VHA proteins colocalize to the same type ofMRCs and this warrants further investigation
Perspectives In summary we propose that ASCI4 is in-volved in mediating Na uptake from FW in rainbow troutacclimatized to low ionic strength low pH media This is thefirst demonstration of a nonneuronal function for any ASICfamily member and simultaneously provides an attractive so-lution to the debate surrounding the thermodynamic limitationsof the other proposed Na transport mechanisms and resolvesthe conundrum of the role of an apical VHA in Na transportThe possibility that other fish models may use similar mecha-nisms under different environmental conditions where modelssuch as the NHERh metabolon cannot be expected to functionremains to be explored
ACKNOWLEDGMENTS
We thank the staff in the Biosciences Aquatic Facilities the MolecularBiology Service Unit and the Cross Cancer Institute Cell Imaging Facility atthe University of Alberta for outstanding help throughout the study We alsothank Alex Clifford for advice and help with phylogenetic analysis and JamesEde for help with confocal microscopy
GRANTS
This study was supported by a Natural Sciences and Engineering ResearchCouncil of Canada Discovery Grant to G G Goss and an Alberta IngenuityStudent Scholarship to A K Dymowska
DISCLOSURES
No conflicts of interest financial or otherwise are declared by the authors
AUTHOR CONTRIBUTIONS
AKD AGS and GGG are responsible for conception and design ofthe research AKD AGS SDB and DC performed the experiments
AKD AGS SDB DC and GGG analyzed the data AKD AGSSDB DC and GGG interpreted the results of the experiments AKDAGS SDB and GGG prepared the figures AKD AGS SDB DCand GGG drafted the manuscript AKD AGS SDB DC and GGGedited and revised the manuscript AKD AGS SDB and GGGapproved the final version of the manuscript
REFERENCES
1 Avella M Bornancin M A new analysis of ammonia and sodiumtransport through the gills of the freshwater rainbow trout (Salmo gaird-neri) J Exp Biol 142 155ndash175 1989
2 Boisen AM Amstrup J Novak I Grosell M Sodium and chloridetransport in soft water and hard water acclimatized zebrafish (Daniorerio) Biochim Biophys Acta 1618 207ndash218 2003
3 Brix KV Grosell M Comparative characterization of Na transport inCyprinodon variegatus variegatus and Cyprinodon variegatus hubbsi amodel species complex for studying teleost invasion of freshwater J ExpBiol 215 1199ndash1209 2012
4 Chen X Polleichtner G Kadurin I Grunder S Zebrafish acid-sensingion channel (ASIC) 4 Characterization of homo- and heteromeric chan-nels and identification of regions important for activation by H J BiolChem 282 30406ndash30413 2007
5 Chen X Qiu L Li M Durrnagel S Orser BA Xiong ZG MacDonaldJF Diarylamidines high potency inhibitors of acid-sensing ion channelsNeuropharmacology 58 1045ndash1053 2010
6 Dymowska AK Hwang PP Goss GG Structure and function of iono-cytes in the freshwater fish gill Respir Physiol Neurobiol 184 282ndash2922012
7 Edwards SL Tse CM Toop T Immunolocalization of NHE3-likeimmunoreactivity in the gills of the rainbow trout (Oncorhynchus mykiss)and the blue-throated wrasse (Pseudolabrus tetrious) J Anat 195 465ndash469 1999
8 Evans DH Piermarini PM Choe KP The multifunctional gill domi-nant site of gas exchange osmoregulation acid-base regulation andexcretion of nitrogenous waste Physiol Rev 85 97ndash177 2005
9 Fenwick JC Bonga SE Flik G In vivo bafilomycin-sensitive Na
uptake in young freshwater fish J Exp Biol 202 3659ndash3666 199910 Galvez F Reid SD Hawkings G Goss GG Isolation and characteriza-
tion of mitochondria-rich cell types from the gill of freshwater rainbowtrout Am J Physiol Regul Integr Comp Physiol 282 R658ndashR668 2002
11 Goss G Orlowski J Grinstein S Coimmunoprecipitation of a 24-kDaprotein with NHE1 the ubiquitous isoform of the NaH exchanger AmJ Physiol Cell Physiol 270 C1493ndashC1502 1996
12 Goss GG Wood CJ Na and Cl uptake kinetics diffusive effluxes andacidic equivalent fluxes across the gills of rainbow trout 1 Responses toenvironmental hyperoxia J Exp Biol 152 521ndash547 1990
13 Grunder S Geissler HS Bassler EL Ruppersberg JP A new memberof acid-sensing ion channels from pituitary gland Neuroreport 11 1607ndash1611 2000
14 Harvey BJ Energization of sodium absorption by the H-ATPase pumpin mitochondria-rich cells of frog skin J Exp Biol 172 289ndash309 1992
15 Hirata T Kaneko T Ono T Nakazato T Furukawa N Hasegawa SWakabayashi S Shigekawa M Chang MH Romero MF Hirose SMechanism of acid adaptation of a fish living in low pH 35 lake Am JPhysiol Regul Integr Comp Physiol 284 R1199ndashR1212 2003
16 Holzer P Acid-sensitive ion channels and receptors Hand Exp Pharma-col 194 283ndash332 2009
17 Hung CY Tsui KN Wilson JM Nawata CM Wood CM Wright PARhesus glycoprotein gene expression in the mangrove killifish Kryptole-bias marmoratus exposed to elevated environmental ammonia levels andair J Exp Biol 210 2419ndash2429 2007
18 Hwang PP Lee TH Lin LY Ion regulations in fish gills recentprogresses in the cellular and molecular mechanisms Am J Physiol RegulIntegr Comp Physiol 301 R28ndashR47 2011
19 Ivanis G Esbaugh AJ Perry SF Branchial expression and localizationof SLC9A2 and SLC9A3 sodiumhydrogen exchangers and their possiblerole in acid-base regulation in freshwater rainbow trout (Oncorhynchusmykiss) J Exp Biol 211 2467ndash2477 2008
20 Jasti J Furukawa H Gonzales EB Gouaux E Structure of acid-sensingion channel 1 at 19A resolution and low pH Nature 449 316ndash323 2007
21 Jonz MG Nurse CA Epithelial mitochondria-rich cells and associatedinnervation in adult and developing zebrafish J Comp Neurol 497817ndash832 2006
C264 INVOLVEMENT OF ASICs IN FISH GILL Na TRANSPORT
AJP-Cell Physiol bull doi101152ajpcell003982013 bull wwwajpcellorg
22 Jonz MG Nurse CA New developments on gill innervation insightsfrom a model vertebrate J Exp Biol 211 2371ndash2378 2008
23 Kellenberger S Epithelial sodium and acid-sensing ion channels InSensing With Ion Channels edited by Martinac B New York Springer2008 chapt 11 p 225ndash246
24 Kellenberger S Schild L Epithelial sodium channeldegenerin family ofion channels a variety of functions for a shared structure Physiol Rev 82735ndash767 2002
25 Kleyman TR Cragoe EJ Amiloride and its analogs as tools in the studyof ion-transport J Membr Biol 105 1ndash21 1988
26 Krogh A The active absorption of ions in some freshwater animals ZVergl Physiol 25 335ndash350 1938
27 Levanti MB Guerrera MC Calavia MG Ciriaco E Montalbano GCobo J Germana A Vega JA Acid-sensing ion channel 2 (ASIC2) inthe intestine of adult zebrafish Neurosci Lett 494 24ndash28 2011
28 Lin H Randall DJ Evidence for the presence of an electrogenic protonpump on the trout gill epithelium J Exp Biol 161 119ndash134 1991
29 Maetz J Garcia Romeu F The mechanism of sodium and chlorideuptake by the gills of a fresh-water fish Carassius auratus II Evidencefor NH4
Na and HCO3Cl exchanges J Gen Physiol 47 1209ndash1227
196430 Nakada T Hoshijima K Esaki M Nagayoshi S Kawakami K Hirose
S Localization of ammonia transporter Rhcg1 in mitochondrion rich cellsof yolk sac gill and kidney of zebrafish and its ionic strength-dependentexpression Am J Physiol Regul Integr Comp Physiol 293 R1743ndashR17532007
31 Nawata CM Hung CC Tsui TK Wilson JM Wright PA Wood CMAmmonia excretion in rainbow trout (Oncorhynchus mykiss) evidence forRh glycoprotein and H-ATPase involvement Physiol Genomics 31463ndash474 2007
32 Parks SK Tresguerres M Galvez F Goss GG Intracellular pH regu-lation in isolated trout gill mitochondrion-rich (MR) cell subtypes evi-dence for NaH activity Comp Biochem Physiol A 155 139ndash1452010
33 Parks SK Tresguerres M Goss GG Interactions between Na channelsand Na-HCO3
cotransporters in the freshwater fish gill MRC a modelfor transepithelial Na uptake Am J Physiol Cell Physiol 292 C935ndashC944 2007
34 Parks SK Tresguerres M Goss GG Theoretical considerations under-lying Na uptake mechanisms in freshwater fishes Comp BiochemPhysiol C 148 411ndash418 2008
35 Paukert M Sidi S Russell C Siba M Wilson SW A family ofacid-sensing ion channels from zebrafish widespread expression in thecentral nervous system suggests a conserved role in neuronal communi-cation J Biol Chem 279 18783ndash18791 2004
36 Perry SF The chloride cell structure and function in the gill of freshwaterfish Annu Rev Physiol 59 325ndash347 1997
37 Perry SF Randall DJ Effects of amiloride and SITS on branchial ionfluxes in rainbow trout Salmo gairdneri J Exp Zool 215 225ndash228 1981
38 Preest MR Gonzales RJ Wilson RW Pharmacological examination ofNa and Cl transport in two species of freshwater fish Physiol BiochemZool 78 259ndash272 2005
39 Reid SD Hawkings GS Galvez F Goss GG Localization and charac-terization of phenamil-sensitive Na influx in isolated rainbow trout gillepithelial cells J Exp Biol 206 551ndash559 2003
40 Sakai H Lingueglia E Champigny G Mattei MG Lazdunski MCloning and functional expression of a novel degenerin-like Na channelgene in mammals J Physiol 519 323ndash333 1999
41 Shih TH Horng JL Liu ST Hwang PP Lin LY Rhcg1 and NHE3b areinvolved in ammonium-dependent sodium uptake by zebrafish larvaeacclimated to low-sodium water Am J Physiol Regul Integr Comp Physiol302 R84ndashR93 2012
42 Springauf A Grunder S An acid-sensing ion channel from shark(Squalus acanthias) mediates transient and sustained responses to protonsJ Physiol 588 809ndash820 2010
43 Spry DJ Wood CM Ion flux rates acid-base status and blood gases inrainbow trout Salmo gairdneri exposed to toxic zinc in natural soft waterCan J Fish Aquat Sci 42 1332ndash1337 1985
44 Sullivan GV Fryer JN Perry SF Immunolocalization of proton pumps(H-ATPase) in pavement cells of rainbow trout gill J Exp Biol 1982619ndash2629 1996
45 Tresguerres M Parks SK Katoh F Goss GG Microtubule-dependentrelocation of branchial V-H-ATPase to the basolateral membrane in thePacific spiny dogfish (Squalus acanthias) a role in base secretion J ExpBiol 209 599ndash609 2006
46 Vina E Parisi V Cabo R Laura R Lopez-Velasco S Lopez-Muniz AGarcia-Suarez O Germana A Vega JA Acid-sensing ion channels(ASICs) in the taste buds of adult zebrafish Neurosci Lett 536 35ndash402013
47 Waldmann R Lazdunski M H-gated cation channels neuronal acidsensors in the NaCDEG family of ion channels Curr Opin Neurobiol 8418ndash424 1998
48 Wilson JM Laurent P Tufts BL Benos DJ Donowitz M Vogl AWRandall DJ NaCl uptake by the branchial epithelium in freshwater teleostfish an immunological approach to ion-transport protein localization JExp Biol 203 2279ndash2296 2000
49 Wood CM Randall DJ Sodium balance in rainbow-trout (Salmo-gaird-neri) during extended exercise J Comp Physiol 82 235ndash256 1973
50 Wright PA Wood CM A new paradigm for ammonia excretion inaquatic animals role of rhesus (Rh) glycoproteins J Exp Biol 2122303ndash2312 2009
51 Yan JJ Chou MY Kaneko T Hwang PP Gene expression of NaH
exchanger in zebrafish H-ATPase-rich cells during acclimation to low-Na and acidic environments Am J Physiol Cell Physiol 293 C1814ndashC1823 2007
C265INVOLVEMENT OF ASICs IN FISH GILL Na TRANSPORT
AJP-Cell Physiol bull doi101152ajpcell003982013 bull wwwajpcellorg

antibody for later immunocytochemistry we performed an im-munoprecipitation using an antibody raised against zASIC42on the basis of sequence similarly of 60 between zASIC42(aa 146 ndash160 PKSRKGHRPSELQYPP) and the clonedtASIC4 (aa 142ndash156 PKNREGHKPTDLDYPA) Using West-ern blotting we analyzed the precipitates for the presence ofASIC4 (see Fig 6A) and found a single band of 65 kDawhich approximates the predicted size for tASIC4 (604 kDa)The strong band at 55 kDa corresponds to the IgG heavychain
To determine the location of ASIC4 protein within therainbow trout gill tissue fixed sections were double-labeledwith anti-zASIC42 antibody (see Fig 6B) and anti-NKA andthen counterstained with the nuclear stain DAPI Immunore-activity was observed in cells located in the lamellae andinterlamellar region (see Fig 6B arrowhead) Moreover themajority of cells positive for zASIC42 also colocalized withNKA protein (Fig 6 C and D arrowhead) a known marker ofMRCs suggesting that in gill epithelium ASIC4 protein ismainly expressed in MRCs SEM analysis of the gill filamentdemonstrated that MRCs are clearly distinguished on the sur-face of the epithelium and have a broad apical convex openingto the environment (see Fig 7A arrowheads) The generatedthree-dimensional reconstructions of the staining immunoreac-tivity pattern for ASIC4 and NKA proteins revealed the pres-ence of ASIC4 and NKA in the same cells but in distinctregions of the cells NKA was present on the basolateralmembrane (arrows green staining) while ASIC4 was localizedto the broad apical surface of the cell (arrowheads red stain-ing) Figure 7B shows a generated image with a top view whileFig 7C is the same cell viewed from the side
DISCUSSION
The existence of an apical Na channel electrochemicallycoupled to a VHA mediating Na uptake from dilute FW hasbeen postulated on theoretical thermodynamic grounds for anumber of decades (1 36 34) However an inability todetermine the molecular identity of the Na channel has beena major drawback in the general acceptance of the modelUsing a combination of pharmacological blockade isotopicfluxes immunocytochemistry and molecular biology we pro-pose that an ASIC family member is potentially the long-sought-after apical Na channel responsible for Na uptake byrainbow trout acclimatized to water with low Na and low pHOur study provides the first evidence for a nonneuronal role forASICs in any vertebrate whereby in gills from FW fishesASICs are associated with the MRCs and tASIC4 localizes tothe apical membrane of NKA positive cells
ASICs are a broad family of Na-specific ion channels thathave been extensively studied in mammalian models (forreview see Ref 23) with only a few studies performed in anyfish species (35 42 46) ASICs have primarily been associatedwith neuronal tissues with studies focusing on a neuronalfunction for ASICs (23) In this study ASIC1 and ASIC4 werecloned in rainbow trout using PCR and they were expressed ina variety of tissues both neuronal and nonneuronal in origin(Fig 5) Importantly tASIC1 gene was present in all fiveinvestigated tissues including whole blood It appears that thetASIC4 gene has a more defined tissue distribution thantASIC1 with expression in the ionoregulatory tissues (trunk
kidney and perfused gill) and the brain Unlike tASIC1 therewas no apparent expression of tASIC4 in the head kidney orthe blood Therefore tASIC4 expression in MRCs and per-fused gill could not be attributed to blood contamination
Both tASIC1 and tASIC4 genes were expressed in the gillepithelium and more specifically were present in MRCs ofrainbow trout gills It is believed that similar to ENaCsfunctional ASICs are trimers (20) although their configurationhas not been resolved It has been demonstrated that with afew exceptions ASICs function as homomers (forming chan-nels of identical subunits) or heteromers (forming channelswith other ASIC subunits) (4 23) ASICs have been charac-terized by using oocyte expression in zebrafish (4 35) anddogfish shark (ASIC1b only) (42) It has been found thatcoexpression of zASIC41 with zASIC13 in Xenopus oocytesincreased the current amplitude and abundance (15-fold) ofthe channel at the cell surface indicating that these two ASICsubunits form a functional channel (4) Moreover thisASIC4113 heteromeric channel was more efficiently traf-ficked to the plasma membrane and had increased affinity forH Additionally zASIC41 when heterologously expressedin Xenopus oocytes has been shown to be also gated open bydecreases in extracellular Ca2 The signal responsible forgating of zASIC42 was unable to be determined in that study(4) It is therefore possible that tASIC1 and tASIC4 form aheteromeric channel in MRCs but this cannot be determined atthis time and would require confirmation by immunohisto-chemistry with an anti-ASIC1 antibody that is immunoreactivefor rainbow trout It is also possible that other ASIC subunitsandor regulatory elements may be involved in the function ofASICs in the FW trout gill A full characterization of tASIC4by oocyte expression and electrophysiology is required tounderstand the mechanisms of gating in low-Na low-Ca2
watersIn our study ASIC4 protein was colocalized with NKA and
was present on the apical surface of cells suggesting that it isfound primarily in MRCs (Fig 6 D and E) Previous studies onASICs in fishes have suggested that they are primarily associ-ated with neuronal tissue and sensory cells (27 35 42 46) Inadult and developing zebrafish morphological studies usingconfocal immunofluorescence techniques demonstrated thatgill MRCs come into direct contact with nerve fibers in thebasolateral region (21 22) However in our study ASIC4protein was clearly localized to the apical region of the MRCsas demonstrated by three-dimensional reconstruction of con-
Head k
idney
ASIC4
ASIC1
GAPDH
Gill MRCs
Brain
Trunk k
idney
Blood
No tem
plate
Fig 5 Tissue distribution analysis of ASIC4 and ASIC1 genes in adultrainbow trout as determined by PCR GAPDH was used as a reference geneMRCs mitochondrion-rich cells
C261INVOLVEMENT OF ASICs IN FISH GILL Na TRANSPORT
AJP-Cell Physiol bull doi101152ajpcell003982013 bull wwwajpcellorg
focal images ASIC4 immunoreactivity did not overlap withthe standard basolateral immunoreactive pattern associatedwith NKA (Fig 7 B and C) Therefore we conclude thatneuronally derived tissues cannot account for the expression ofASIC4 protein in MRCs We also corroborate the ASIC4staining pattern with the MRC external morphology whichshows that in FW rainbow trout raised in Edmonton tap water(500 M Na) the MRCs have wide apical openings (Fig7A) This apical localization of ASIC4 protein in the cells of atransporting epithelium is the first evidence for a nonneuronalfunction for any ASIC member and suggests that ASIC4 as aNa channel may be responsible for apical uptake of Na inFW fish gill
Pharmacological profiling has long been used to differentiatethe mechanisms of ion transport across epithelial tissues For Na
transport classical analysis has usually involved amiloride and itssubstituted derivatives (eg EIPA mercaptoisobutyric acid andphenamil) as a means to differentiate between the involvementof NHE isoforms and ENaC in Na transport (25) We inves-tigated two newly described ASIC-specific inhibitors dimina-zene and DAPI (5) for their potency in inhibiting unidirec-tional Na uptake in fish Diminazene and DAPI exhibited adose-dependent inhibitory relationship and both were veryeffective at low concentrations (Fig 1) Na uptake in rainbow
trout acclimatized to low-ionic-strength acidic water wasclearly inhibited by these diarylamidines at concentrationssimilar to those reported in a previous study of cultured rathippocampal neurons (5) However in our study DAPI provedto be more effective than diminazene in contrast to observa-tions in rat neurons The inhibition of 22Na uptake by DAPIand diminazene was also compared with other common Na
transport inhibitors amiloride and phenamil Amiloride wasobserved to be an effective inhibitor of Na uptake in rainbowtrout but only at relatively high concentrations (500 M)which is consistent with previous studies (2 37) Recentlyamiloride was also demonstrated to completely block ASIC-mediated currents (5 35 42) however it is also known toblock many other Na transporters including NHE and ENaC(25) Amiloride therefore cannot be used as a distinguishingpharmacological agent
While DAPI diminazene and amiloride almost completelyblocked Na transport in the present study phenamil had nosignificant effect on Na flux in rainbow trout This suggeststhat phenamil is a poor inhibitor of Na uptake in troutexposed to low-ionic-strength water (30 moll Na) Paststudies of the effects of phenamil on Na uptake in a variety ofFW fishes have reported inconsistent results For example inneon tetras acclimatized to low Na (50 moll) phenamil had
Fig 6 Immunoreactivity of anti-zebrafish ASIC42 (zASIC42) antibody in the gill of rainbow trout A immunoprecipitation Western blot from whole gillhomogenates with zASIC42 showing a distinct band of 65 kDa Lane 1 (L1) contains 300 mg of gill tissue and lane 2 (L2) contains 200 mg of gill tissueBndashE confocal images of gill sections labeled for ASIC4 and Na-K-ATPase (NKA) proteins B and C double-labeling with DAPI and anti-zASIC42 (B) andDAPI and anti-NKA (C) D merged image of B and C E higher-magnification image of a MRC showing that ASIC4 and NKA are present in distinct regionswithin the MRC F control micrograph with DAPI and no primary antibody Arrowheads (BndashD) indicate positive staining
C262 INVOLVEMENT OF ASICs IN FISH GILL Na TRANSPORT
AJP-Cell Physiol bull doi101152ajpcell003982013 bull wwwajpcellorg
no effect on Na transport (38) whereas it caused 70reduction in Na influx in Cyprinidon variegatus hubbsi (100moll Na) and 60 reduction in goldfish (300 moll Na)(2 38) These conflicting results from various phenamil studiessuggest that sensitivity to phenamil might be species- andorenvironment-specific Collectively the pharmacological anal-ysis suggests that the Na uptake in FW rainbow trout is mostlikely not mediated by ENaC given the lack of inhibition byphenamil but may be attributed to ASIC since Na uptakewas almost completely blocked by DAPI and diminazene bothof which inhibit ASICs but not ENaC
In our Na flux study with various pharmacological agentswe made an interesting observation that inhibition of 22Na
uptake was less pronounced for all tested inhibitors during thesecond consecutive measurement (90ndash180 min) (Fig 2) Toour knowledge this phenomenon where potency of the testeddrug diminishes over time has not been reported in anyprevious study on fish This observation may be explained bya decreased stabilityefficacy of the tested pharmacologicalagents in aqueous media metabolism of the drug by theanimal or activation of compensatory Na uptake mechanismsnot sensitive to amiloride DAPI or diminazene We believethat the decrease of the potency of amiloride DAPI anddiminazene over time demonstrates the importance of properexperimental design and choice of period of study when ionfluxes are examined using pharmacological inhibitors
While the two novel ASIC inhibitors have been tested fortheir potency in inhibiting ASICs and ENaC in neurons (5)they have not been tested for their potential effects on otherNa transporters implicated in Na uptake in fishes mostnotably the NHEs To verify that the Na uptake inhibitioncaused by DAPI and diminazene in rainbow trout was due toinhibition of ASICs rather than NHEs we tested the effect ofDAPI on MRCs isolated from the rainbow trout gill epithe-lium Our laboratory previously showed that rainbow troutpossess two populations of MRCs peanut lectin agglutinin(PNA) positive (PNA) and PNA negative (PNA) MRCs(10) It has been demonstrated that NHE3 and NHE2 areexpressed in PNA cells isolated from branchial epithelium of
FW rainbow trout (19) Therefore we investigated the effect ofDAPI on PNA cells and compared it with the effect of EIPAa potent and highly selective inhibitor of NHE (25) Earlierstudies in our laboratory using cell imaging determined thatPNA cells undergo NHE-dependent alkalinization (32 33)Here we demonstrate that while EIPA was effective at inhib-iting NHE function in PNA cells (Fig 3A) there was noeffect of high concentrations of DAPI on Na-dependent pHi
elevation demonstrating that NHE was not inhibited by DAPI(Fig 3B) Therefore the measured inhibition of Na uptake byDAPI that we see in whole animals (Fig 1) cannot be attrib-uted to inhibition of NHE
On the basis of the mRNA expression of tASIC1 andtASIC4 genes in the gill epithelium and isolated MRCs local-ization of ASIC4 protein to the apical region of the MRCs and
Fig 7 Three-dimensional views of freshwater(FW) rainbow trout MRCs A scanning elec-tron micrograph of FW rainbow trout gillepithelium showing broad apical openings ofMRCs (arrowheads) and mucous cells () Band C 3-dimensional reconstruction of z-stack images showing apical localization ofASIC4 protein (red arrowhead) and basolat-eral localization of NKA (green arrow)B top view C side view
PNA- MRC
Na+
ASIC
H+
VHA
NKA
Na+
K+
NBC
Na+ HCO3-
PNA+ MRC
NH3
Na+
H+
NHE Rhcg2
NKA
Na+
K+
Fig 8 Model for Na uptake in PNA and PNA MRCs in gill epithelium ofrainbow trout showing placement of proposed ASIC-mediated Na uptakemechanisms Rhcg2 rhesus glycoprotein NHE NaH exchanger VHAvacuolar H-ATPase NBC Na-HCO3
cotransporter
C263INVOLVEMENT OF ASICs IN FISH GILL Na TRANSPORT
AJP-Cell Physiol bull doi101152ajpcell003982013 bull wwwajpcellorg
pharmacological blockade of Na influx by ASIC inhibitors inadult rainbow trout we propose a revised model for transepi-thelial Na uptake in FW rainbow trout gills (Fig 8) wherebyone apical mode of Na entry is via an ASIC4-mediatedmechanism
A number of recent studies on Na uptake in FW fishes havedemonstrated the presence of the NHERh metabolon in thegills of various species (17 30 31) As mentioned previouslywe specifically targeted the rainbow trout as a model for thisstudy as it is native to oligotrophic Na-poor mountainstreams with low buffering capacity This type of environmentpresents key thermodynamic challenges to the NHERhmetabolon as Na restrictions cannot be alleviated by thepresence of an Rh protein (6) Moreover searches of rainbowtrout genomes and transcriptomes have failed to identify anyENaC subunits ( 13 or ) despite the fact that they are knownto actively take up Na from water with 50 M Na (49)Furthermore other research studies have suggested and dem-onstrated an apical localization of the VHA in the gills ofrainbow trout (44 48) and bafilomycin a VHA-specific in-hibitor has been demonstrated to reduce Na uptake in ze-brafish (2) tilapia and carp (9) Previous studies have pro-posed VHA to be a part of the NHERh metabolon (41 50 51)however the placement of an apically oriented VHA is coun-terintuitive to the function of an NHE as it would exacerbatethe noted thermodynamic constraints Placement of a VHA inconcert with an apical ASIC is more plausible since it wouldnot only facilitate Na uptake from very low ionic strengthwater but could also provide a mechanism for gating ofASIC-mediated Na conductance It remains to be determinedif ASIC and VHA proteins colocalize to the same type ofMRCs and this warrants further investigation
Perspectives In summary we propose that ASCI4 is in-volved in mediating Na uptake from FW in rainbow troutacclimatized to low ionic strength low pH media This is thefirst demonstration of a nonneuronal function for any ASICfamily member and simultaneously provides an attractive so-lution to the debate surrounding the thermodynamic limitationsof the other proposed Na transport mechanisms and resolvesthe conundrum of the role of an apical VHA in Na transportThe possibility that other fish models may use similar mecha-nisms under different environmental conditions where modelssuch as the NHERh metabolon cannot be expected to functionremains to be explored
ACKNOWLEDGMENTS
We thank the staff in the Biosciences Aquatic Facilities the MolecularBiology Service Unit and the Cross Cancer Institute Cell Imaging Facility atthe University of Alberta for outstanding help throughout the study We alsothank Alex Clifford for advice and help with phylogenetic analysis and JamesEde for help with confocal microscopy
GRANTS
This study was supported by a Natural Sciences and Engineering ResearchCouncil of Canada Discovery Grant to G G Goss and an Alberta IngenuityStudent Scholarship to A K Dymowska
DISCLOSURES
No conflicts of interest financial or otherwise are declared by the authors
AUTHOR CONTRIBUTIONS
AKD AGS and GGG are responsible for conception and design ofthe research AKD AGS SDB and DC performed the experiments
AKD AGS SDB DC and GGG analyzed the data AKD AGSSDB DC and GGG interpreted the results of the experiments AKDAGS SDB and GGG prepared the figures AKD AGS SDB DCand GGG drafted the manuscript AKD AGS SDB DC and GGGedited and revised the manuscript AKD AGS SDB and GGGapproved the final version of the manuscript
REFERENCES
1 Avella M Bornancin M A new analysis of ammonia and sodiumtransport through the gills of the freshwater rainbow trout (Salmo gaird-neri) J Exp Biol 142 155ndash175 1989
2 Boisen AM Amstrup J Novak I Grosell M Sodium and chloridetransport in soft water and hard water acclimatized zebrafish (Daniorerio) Biochim Biophys Acta 1618 207ndash218 2003
3 Brix KV Grosell M Comparative characterization of Na transport inCyprinodon variegatus variegatus and Cyprinodon variegatus hubbsi amodel species complex for studying teleost invasion of freshwater J ExpBiol 215 1199ndash1209 2012
4 Chen X Polleichtner G Kadurin I Grunder S Zebrafish acid-sensingion channel (ASIC) 4 Characterization of homo- and heteromeric chan-nels and identification of regions important for activation by H J BiolChem 282 30406ndash30413 2007
5 Chen X Qiu L Li M Durrnagel S Orser BA Xiong ZG MacDonaldJF Diarylamidines high potency inhibitors of acid-sensing ion channelsNeuropharmacology 58 1045ndash1053 2010
6 Dymowska AK Hwang PP Goss GG Structure and function of iono-cytes in the freshwater fish gill Respir Physiol Neurobiol 184 282ndash2922012
7 Edwards SL Tse CM Toop T Immunolocalization of NHE3-likeimmunoreactivity in the gills of the rainbow trout (Oncorhynchus mykiss)and the blue-throated wrasse (Pseudolabrus tetrious) J Anat 195 465ndash469 1999
8 Evans DH Piermarini PM Choe KP The multifunctional gill domi-nant site of gas exchange osmoregulation acid-base regulation andexcretion of nitrogenous waste Physiol Rev 85 97ndash177 2005
9 Fenwick JC Bonga SE Flik G In vivo bafilomycin-sensitive Na
uptake in young freshwater fish J Exp Biol 202 3659ndash3666 199910 Galvez F Reid SD Hawkings G Goss GG Isolation and characteriza-
tion of mitochondria-rich cell types from the gill of freshwater rainbowtrout Am J Physiol Regul Integr Comp Physiol 282 R658ndashR668 2002
11 Goss G Orlowski J Grinstein S Coimmunoprecipitation of a 24-kDaprotein with NHE1 the ubiquitous isoform of the NaH exchanger AmJ Physiol Cell Physiol 270 C1493ndashC1502 1996
12 Goss GG Wood CJ Na and Cl uptake kinetics diffusive effluxes andacidic equivalent fluxes across the gills of rainbow trout 1 Responses toenvironmental hyperoxia J Exp Biol 152 521ndash547 1990
13 Grunder S Geissler HS Bassler EL Ruppersberg JP A new memberof acid-sensing ion channels from pituitary gland Neuroreport 11 1607ndash1611 2000
14 Harvey BJ Energization of sodium absorption by the H-ATPase pumpin mitochondria-rich cells of frog skin J Exp Biol 172 289ndash309 1992
15 Hirata T Kaneko T Ono T Nakazato T Furukawa N Hasegawa SWakabayashi S Shigekawa M Chang MH Romero MF Hirose SMechanism of acid adaptation of a fish living in low pH 35 lake Am JPhysiol Regul Integr Comp Physiol 284 R1199ndashR1212 2003
16 Holzer P Acid-sensitive ion channels and receptors Hand Exp Pharma-col 194 283ndash332 2009
17 Hung CY Tsui KN Wilson JM Nawata CM Wood CM Wright PARhesus glycoprotein gene expression in the mangrove killifish Kryptole-bias marmoratus exposed to elevated environmental ammonia levels andair J Exp Biol 210 2419ndash2429 2007
18 Hwang PP Lee TH Lin LY Ion regulations in fish gills recentprogresses in the cellular and molecular mechanisms Am J Physiol RegulIntegr Comp Physiol 301 R28ndashR47 2011
19 Ivanis G Esbaugh AJ Perry SF Branchial expression and localizationof SLC9A2 and SLC9A3 sodiumhydrogen exchangers and their possiblerole in acid-base regulation in freshwater rainbow trout (Oncorhynchusmykiss) J Exp Biol 211 2467ndash2477 2008
20 Jasti J Furukawa H Gonzales EB Gouaux E Structure of acid-sensingion channel 1 at 19A resolution and low pH Nature 449 316ndash323 2007
21 Jonz MG Nurse CA Epithelial mitochondria-rich cells and associatedinnervation in adult and developing zebrafish J Comp Neurol 497817ndash832 2006
C264 INVOLVEMENT OF ASICs IN FISH GILL Na TRANSPORT
AJP-Cell Physiol bull doi101152ajpcell003982013 bull wwwajpcellorg
22 Jonz MG Nurse CA New developments on gill innervation insightsfrom a model vertebrate J Exp Biol 211 2371ndash2378 2008
23 Kellenberger S Epithelial sodium and acid-sensing ion channels InSensing With Ion Channels edited by Martinac B New York Springer2008 chapt 11 p 225ndash246
24 Kellenberger S Schild L Epithelial sodium channeldegenerin family ofion channels a variety of functions for a shared structure Physiol Rev 82735ndash767 2002
25 Kleyman TR Cragoe EJ Amiloride and its analogs as tools in the studyof ion-transport J Membr Biol 105 1ndash21 1988
26 Krogh A The active absorption of ions in some freshwater animals ZVergl Physiol 25 335ndash350 1938
27 Levanti MB Guerrera MC Calavia MG Ciriaco E Montalbano GCobo J Germana A Vega JA Acid-sensing ion channel 2 (ASIC2) inthe intestine of adult zebrafish Neurosci Lett 494 24ndash28 2011
28 Lin H Randall DJ Evidence for the presence of an electrogenic protonpump on the trout gill epithelium J Exp Biol 161 119ndash134 1991
29 Maetz J Garcia Romeu F The mechanism of sodium and chlorideuptake by the gills of a fresh-water fish Carassius auratus II Evidencefor NH4
Na and HCO3Cl exchanges J Gen Physiol 47 1209ndash1227
196430 Nakada T Hoshijima K Esaki M Nagayoshi S Kawakami K Hirose
S Localization of ammonia transporter Rhcg1 in mitochondrion rich cellsof yolk sac gill and kidney of zebrafish and its ionic strength-dependentexpression Am J Physiol Regul Integr Comp Physiol 293 R1743ndashR17532007
31 Nawata CM Hung CC Tsui TK Wilson JM Wright PA Wood CMAmmonia excretion in rainbow trout (Oncorhynchus mykiss) evidence forRh glycoprotein and H-ATPase involvement Physiol Genomics 31463ndash474 2007
32 Parks SK Tresguerres M Galvez F Goss GG Intracellular pH regu-lation in isolated trout gill mitochondrion-rich (MR) cell subtypes evi-dence for NaH activity Comp Biochem Physiol A 155 139ndash1452010
33 Parks SK Tresguerres M Goss GG Interactions between Na channelsand Na-HCO3
cotransporters in the freshwater fish gill MRC a modelfor transepithelial Na uptake Am J Physiol Cell Physiol 292 C935ndashC944 2007
34 Parks SK Tresguerres M Goss GG Theoretical considerations under-lying Na uptake mechanisms in freshwater fishes Comp BiochemPhysiol C 148 411ndash418 2008
35 Paukert M Sidi S Russell C Siba M Wilson SW A family ofacid-sensing ion channels from zebrafish widespread expression in thecentral nervous system suggests a conserved role in neuronal communi-cation J Biol Chem 279 18783ndash18791 2004
36 Perry SF The chloride cell structure and function in the gill of freshwaterfish Annu Rev Physiol 59 325ndash347 1997
37 Perry SF Randall DJ Effects of amiloride and SITS on branchial ionfluxes in rainbow trout Salmo gairdneri J Exp Zool 215 225ndash228 1981
38 Preest MR Gonzales RJ Wilson RW Pharmacological examination ofNa and Cl transport in two species of freshwater fish Physiol BiochemZool 78 259ndash272 2005
39 Reid SD Hawkings GS Galvez F Goss GG Localization and charac-terization of phenamil-sensitive Na influx in isolated rainbow trout gillepithelial cells J Exp Biol 206 551ndash559 2003
40 Sakai H Lingueglia E Champigny G Mattei MG Lazdunski MCloning and functional expression of a novel degenerin-like Na channelgene in mammals J Physiol 519 323ndash333 1999
41 Shih TH Horng JL Liu ST Hwang PP Lin LY Rhcg1 and NHE3b areinvolved in ammonium-dependent sodium uptake by zebrafish larvaeacclimated to low-sodium water Am J Physiol Regul Integr Comp Physiol302 R84ndashR93 2012
42 Springauf A Grunder S An acid-sensing ion channel from shark(Squalus acanthias) mediates transient and sustained responses to protonsJ Physiol 588 809ndash820 2010
43 Spry DJ Wood CM Ion flux rates acid-base status and blood gases inrainbow trout Salmo gairdneri exposed to toxic zinc in natural soft waterCan J Fish Aquat Sci 42 1332ndash1337 1985
44 Sullivan GV Fryer JN Perry SF Immunolocalization of proton pumps(H-ATPase) in pavement cells of rainbow trout gill J Exp Biol 1982619ndash2629 1996
45 Tresguerres M Parks SK Katoh F Goss GG Microtubule-dependentrelocation of branchial V-H-ATPase to the basolateral membrane in thePacific spiny dogfish (Squalus acanthias) a role in base secretion J ExpBiol 209 599ndash609 2006
46 Vina E Parisi V Cabo R Laura R Lopez-Velasco S Lopez-Muniz AGarcia-Suarez O Germana A Vega JA Acid-sensing ion channels(ASICs) in the taste buds of adult zebrafish Neurosci Lett 536 35ndash402013
47 Waldmann R Lazdunski M H-gated cation channels neuronal acidsensors in the NaCDEG family of ion channels Curr Opin Neurobiol 8418ndash424 1998
48 Wilson JM Laurent P Tufts BL Benos DJ Donowitz M Vogl AWRandall DJ NaCl uptake by the branchial epithelium in freshwater teleostfish an immunological approach to ion-transport protein localization JExp Biol 203 2279ndash2296 2000
49 Wood CM Randall DJ Sodium balance in rainbow-trout (Salmo-gaird-neri) during extended exercise J Comp Physiol 82 235ndash256 1973
50 Wright PA Wood CM A new paradigm for ammonia excretion inaquatic animals role of rhesus (Rh) glycoproteins J Exp Biol 2122303ndash2312 2009
51 Yan JJ Chou MY Kaneko T Hwang PP Gene expression of NaH
exchanger in zebrafish H-ATPase-rich cells during acclimation to low-Na and acidic environments Am J Physiol Cell Physiol 293 C1814ndashC1823 2007
C265INVOLVEMENT OF ASICs IN FISH GILL Na TRANSPORT
AJP-Cell Physiol bull doi101152ajpcell003982013 bull wwwajpcellorg
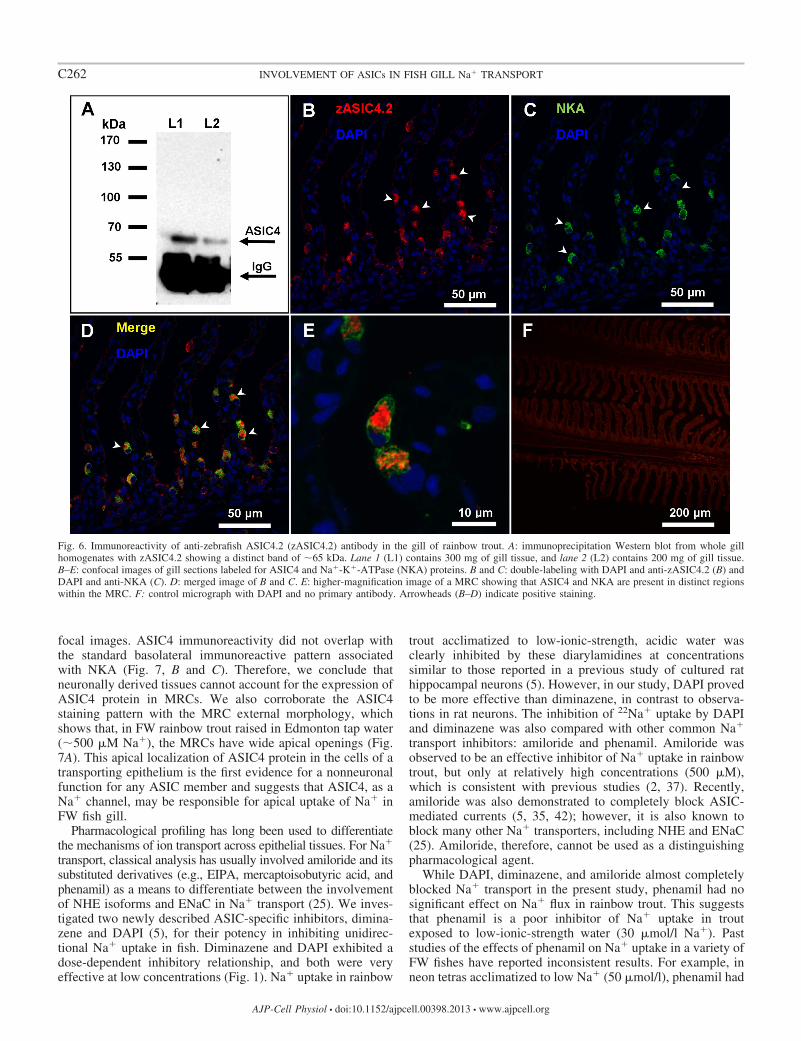
focal images ASIC4 immunoreactivity did not overlap withthe standard basolateral immunoreactive pattern associatedwith NKA (Fig 7 B and C) Therefore we conclude thatneuronally derived tissues cannot account for the expression ofASIC4 protein in MRCs We also corroborate the ASIC4staining pattern with the MRC external morphology whichshows that in FW rainbow trout raised in Edmonton tap water(500 M Na) the MRCs have wide apical openings (Fig7A) This apical localization of ASIC4 protein in the cells of atransporting epithelium is the first evidence for a nonneuronalfunction for any ASIC member and suggests that ASIC4 as aNa channel may be responsible for apical uptake of Na inFW fish gill
Pharmacological profiling has long been used to differentiatethe mechanisms of ion transport across epithelial tissues For Na
transport classical analysis has usually involved amiloride and itssubstituted derivatives (eg EIPA mercaptoisobutyric acid andphenamil) as a means to differentiate between the involvementof NHE isoforms and ENaC in Na transport (25) We inves-tigated two newly described ASIC-specific inhibitors dimina-zene and DAPI (5) for their potency in inhibiting unidirec-tional Na uptake in fish Diminazene and DAPI exhibited adose-dependent inhibitory relationship and both were veryeffective at low concentrations (Fig 1) Na uptake in rainbow
trout acclimatized to low-ionic-strength acidic water wasclearly inhibited by these diarylamidines at concentrationssimilar to those reported in a previous study of cultured rathippocampal neurons (5) However in our study DAPI provedto be more effective than diminazene in contrast to observa-tions in rat neurons The inhibition of 22Na uptake by DAPIand diminazene was also compared with other common Na
transport inhibitors amiloride and phenamil Amiloride wasobserved to be an effective inhibitor of Na uptake in rainbowtrout but only at relatively high concentrations (500 M)which is consistent with previous studies (2 37) Recentlyamiloride was also demonstrated to completely block ASIC-mediated currents (5 35 42) however it is also known toblock many other Na transporters including NHE and ENaC(25) Amiloride therefore cannot be used as a distinguishingpharmacological agent
While DAPI diminazene and amiloride almost completelyblocked Na transport in the present study phenamil had nosignificant effect on Na flux in rainbow trout This suggeststhat phenamil is a poor inhibitor of Na uptake in troutexposed to low-ionic-strength water (30 moll Na) Paststudies of the effects of phenamil on Na uptake in a variety ofFW fishes have reported inconsistent results For example inneon tetras acclimatized to low Na (50 moll) phenamil had
Fig 6 Immunoreactivity of anti-zebrafish ASIC42 (zASIC42) antibody in the gill of rainbow trout A immunoprecipitation Western blot from whole gillhomogenates with zASIC42 showing a distinct band of 65 kDa Lane 1 (L1) contains 300 mg of gill tissue and lane 2 (L2) contains 200 mg of gill tissueBndashE confocal images of gill sections labeled for ASIC4 and Na-K-ATPase (NKA) proteins B and C double-labeling with DAPI and anti-zASIC42 (B) andDAPI and anti-NKA (C) D merged image of B and C E higher-magnification image of a MRC showing that ASIC4 and NKA are present in distinct regionswithin the MRC F control micrograph with DAPI and no primary antibody Arrowheads (BndashD) indicate positive staining
C262 INVOLVEMENT OF ASICs IN FISH GILL Na TRANSPORT
AJP-Cell Physiol bull doi101152ajpcell003982013 bull wwwajpcellorg
no effect on Na transport (38) whereas it caused 70reduction in Na influx in Cyprinidon variegatus hubbsi (100moll Na) and 60 reduction in goldfish (300 moll Na)(2 38) These conflicting results from various phenamil studiessuggest that sensitivity to phenamil might be species- andorenvironment-specific Collectively the pharmacological anal-ysis suggests that the Na uptake in FW rainbow trout is mostlikely not mediated by ENaC given the lack of inhibition byphenamil but may be attributed to ASIC since Na uptakewas almost completely blocked by DAPI and diminazene bothof which inhibit ASICs but not ENaC
In our Na flux study with various pharmacological agentswe made an interesting observation that inhibition of 22Na
uptake was less pronounced for all tested inhibitors during thesecond consecutive measurement (90ndash180 min) (Fig 2) Toour knowledge this phenomenon where potency of the testeddrug diminishes over time has not been reported in anyprevious study on fish This observation may be explained bya decreased stabilityefficacy of the tested pharmacologicalagents in aqueous media metabolism of the drug by theanimal or activation of compensatory Na uptake mechanismsnot sensitive to amiloride DAPI or diminazene We believethat the decrease of the potency of amiloride DAPI anddiminazene over time demonstrates the importance of properexperimental design and choice of period of study when ionfluxes are examined using pharmacological inhibitors
While the two novel ASIC inhibitors have been tested fortheir potency in inhibiting ASICs and ENaC in neurons (5)they have not been tested for their potential effects on otherNa transporters implicated in Na uptake in fishes mostnotably the NHEs To verify that the Na uptake inhibitioncaused by DAPI and diminazene in rainbow trout was due toinhibition of ASICs rather than NHEs we tested the effect ofDAPI on MRCs isolated from the rainbow trout gill epithe-lium Our laboratory previously showed that rainbow troutpossess two populations of MRCs peanut lectin agglutinin(PNA) positive (PNA) and PNA negative (PNA) MRCs(10) It has been demonstrated that NHE3 and NHE2 areexpressed in PNA cells isolated from branchial epithelium of
FW rainbow trout (19) Therefore we investigated the effect ofDAPI on PNA cells and compared it with the effect of EIPAa potent and highly selective inhibitor of NHE (25) Earlierstudies in our laboratory using cell imaging determined thatPNA cells undergo NHE-dependent alkalinization (32 33)Here we demonstrate that while EIPA was effective at inhib-iting NHE function in PNA cells (Fig 3A) there was noeffect of high concentrations of DAPI on Na-dependent pHi
elevation demonstrating that NHE was not inhibited by DAPI(Fig 3B) Therefore the measured inhibition of Na uptake byDAPI that we see in whole animals (Fig 1) cannot be attrib-uted to inhibition of NHE
On the basis of the mRNA expression of tASIC1 andtASIC4 genes in the gill epithelium and isolated MRCs local-ization of ASIC4 protein to the apical region of the MRCs and
Fig 7 Three-dimensional views of freshwater(FW) rainbow trout MRCs A scanning elec-tron micrograph of FW rainbow trout gillepithelium showing broad apical openings ofMRCs (arrowheads) and mucous cells () Band C 3-dimensional reconstruction of z-stack images showing apical localization ofASIC4 protein (red arrowhead) and basolat-eral localization of NKA (green arrow)B top view C side view
PNA- MRC
Na+
ASIC
H+
VHA
NKA
Na+
K+
NBC
Na+ HCO3-
PNA+ MRC
NH3
Na+
H+
NHE Rhcg2
NKA
Na+
K+
Fig 8 Model for Na uptake in PNA and PNA MRCs in gill epithelium ofrainbow trout showing placement of proposed ASIC-mediated Na uptakemechanisms Rhcg2 rhesus glycoprotein NHE NaH exchanger VHAvacuolar H-ATPase NBC Na-HCO3
cotransporter
C263INVOLVEMENT OF ASICs IN FISH GILL Na TRANSPORT
AJP-Cell Physiol bull doi101152ajpcell003982013 bull wwwajpcellorg
pharmacological blockade of Na influx by ASIC inhibitors inadult rainbow trout we propose a revised model for transepi-thelial Na uptake in FW rainbow trout gills (Fig 8) wherebyone apical mode of Na entry is via an ASIC4-mediatedmechanism
A number of recent studies on Na uptake in FW fishes havedemonstrated the presence of the NHERh metabolon in thegills of various species (17 30 31) As mentioned previouslywe specifically targeted the rainbow trout as a model for thisstudy as it is native to oligotrophic Na-poor mountainstreams with low buffering capacity This type of environmentpresents key thermodynamic challenges to the NHERhmetabolon as Na restrictions cannot be alleviated by thepresence of an Rh protein (6) Moreover searches of rainbowtrout genomes and transcriptomes have failed to identify anyENaC subunits ( 13 or ) despite the fact that they are knownto actively take up Na from water with 50 M Na (49)Furthermore other research studies have suggested and dem-onstrated an apical localization of the VHA in the gills ofrainbow trout (44 48) and bafilomycin a VHA-specific in-hibitor has been demonstrated to reduce Na uptake in ze-brafish (2) tilapia and carp (9) Previous studies have pro-posed VHA to be a part of the NHERh metabolon (41 50 51)however the placement of an apically oriented VHA is coun-terintuitive to the function of an NHE as it would exacerbatethe noted thermodynamic constraints Placement of a VHA inconcert with an apical ASIC is more plausible since it wouldnot only facilitate Na uptake from very low ionic strengthwater but could also provide a mechanism for gating ofASIC-mediated Na conductance It remains to be determinedif ASIC and VHA proteins colocalize to the same type ofMRCs and this warrants further investigation
Perspectives In summary we propose that ASCI4 is in-volved in mediating Na uptake from FW in rainbow troutacclimatized to low ionic strength low pH media This is thefirst demonstration of a nonneuronal function for any ASICfamily member and simultaneously provides an attractive so-lution to the debate surrounding the thermodynamic limitationsof the other proposed Na transport mechanisms and resolvesthe conundrum of the role of an apical VHA in Na transportThe possibility that other fish models may use similar mecha-nisms under different environmental conditions where modelssuch as the NHERh metabolon cannot be expected to functionremains to be explored
ACKNOWLEDGMENTS
We thank the staff in the Biosciences Aquatic Facilities the MolecularBiology Service Unit and the Cross Cancer Institute Cell Imaging Facility atthe University of Alberta for outstanding help throughout the study We alsothank Alex Clifford for advice and help with phylogenetic analysis and JamesEde for help with confocal microscopy
GRANTS
This study was supported by a Natural Sciences and Engineering ResearchCouncil of Canada Discovery Grant to G G Goss and an Alberta IngenuityStudent Scholarship to A K Dymowska
DISCLOSURES
No conflicts of interest financial or otherwise are declared by the authors
AUTHOR CONTRIBUTIONS
AKD AGS and GGG are responsible for conception and design ofthe research AKD AGS SDB and DC performed the experiments
AKD AGS SDB DC and GGG analyzed the data AKD AGSSDB DC and GGG interpreted the results of the experiments AKDAGS SDB and GGG prepared the figures AKD AGS SDB DCand GGG drafted the manuscript AKD AGS SDB DC and GGGedited and revised the manuscript AKD AGS SDB and GGGapproved the final version of the manuscript
REFERENCES
1 Avella M Bornancin M A new analysis of ammonia and sodiumtransport through the gills of the freshwater rainbow trout (Salmo gaird-neri) J Exp Biol 142 155ndash175 1989
2 Boisen AM Amstrup J Novak I Grosell M Sodium and chloridetransport in soft water and hard water acclimatized zebrafish (Daniorerio) Biochim Biophys Acta 1618 207ndash218 2003
3 Brix KV Grosell M Comparative characterization of Na transport inCyprinodon variegatus variegatus and Cyprinodon variegatus hubbsi amodel species complex for studying teleost invasion of freshwater J ExpBiol 215 1199ndash1209 2012
4 Chen X Polleichtner G Kadurin I Grunder S Zebrafish acid-sensingion channel (ASIC) 4 Characterization of homo- and heteromeric chan-nels and identification of regions important for activation by H J BiolChem 282 30406ndash30413 2007
5 Chen X Qiu L Li M Durrnagel S Orser BA Xiong ZG MacDonaldJF Diarylamidines high potency inhibitors of acid-sensing ion channelsNeuropharmacology 58 1045ndash1053 2010
6 Dymowska AK Hwang PP Goss GG Structure and function of iono-cytes in the freshwater fish gill Respir Physiol Neurobiol 184 282ndash2922012
7 Edwards SL Tse CM Toop T Immunolocalization of NHE3-likeimmunoreactivity in the gills of the rainbow trout (Oncorhynchus mykiss)and the blue-throated wrasse (Pseudolabrus tetrious) J Anat 195 465ndash469 1999
8 Evans DH Piermarini PM Choe KP The multifunctional gill domi-nant site of gas exchange osmoregulation acid-base regulation andexcretion of nitrogenous waste Physiol Rev 85 97ndash177 2005
9 Fenwick JC Bonga SE Flik G In vivo bafilomycin-sensitive Na
uptake in young freshwater fish J Exp Biol 202 3659ndash3666 199910 Galvez F Reid SD Hawkings G Goss GG Isolation and characteriza-
tion of mitochondria-rich cell types from the gill of freshwater rainbowtrout Am J Physiol Regul Integr Comp Physiol 282 R658ndashR668 2002
11 Goss G Orlowski J Grinstein S Coimmunoprecipitation of a 24-kDaprotein with NHE1 the ubiquitous isoform of the NaH exchanger AmJ Physiol Cell Physiol 270 C1493ndashC1502 1996
12 Goss GG Wood CJ Na and Cl uptake kinetics diffusive effluxes andacidic equivalent fluxes across the gills of rainbow trout 1 Responses toenvironmental hyperoxia J Exp Biol 152 521ndash547 1990
13 Grunder S Geissler HS Bassler EL Ruppersberg JP A new memberof acid-sensing ion channels from pituitary gland Neuroreport 11 1607ndash1611 2000
14 Harvey BJ Energization of sodium absorption by the H-ATPase pumpin mitochondria-rich cells of frog skin J Exp Biol 172 289ndash309 1992
15 Hirata T Kaneko T Ono T Nakazato T Furukawa N Hasegawa SWakabayashi S Shigekawa M Chang MH Romero MF Hirose SMechanism of acid adaptation of a fish living in low pH 35 lake Am JPhysiol Regul Integr Comp Physiol 284 R1199ndashR1212 2003
16 Holzer P Acid-sensitive ion channels and receptors Hand Exp Pharma-col 194 283ndash332 2009
17 Hung CY Tsui KN Wilson JM Nawata CM Wood CM Wright PARhesus glycoprotein gene expression in the mangrove killifish Kryptole-bias marmoratus exposed to elevated environmental ammonia levels andair J Exp Biol 210 2419ndash2429 2007
18 Hwang PP Lee TH Lin LY Ion regulations in fish gills recentprogresses in the cellular and molecular mechanisms Am J Physiol RegulIntegr Comp Physiol 301 R28ndashR47 2011
19 Ivanis G Esbaugh AJ Perry SF Branchial expression and localizationof SLC9A2 and SLC9A3 sodiumhydrogen exchangers and their possiblerole in acid-base regulation in freshwater rainbow trout (Oncorhynchusmykiss) J Exp Biol 211 2467ndash2477 2008
20 Jasti J Furukawa H Gonzales EB Gouaux E Structure of acid-sensingion channel 1 at 19A resolution and low pH Nature 449 316ndash323 2007
21 Jonz MG Nurse CA Epithelial mitochondria-rich cells and associatedinnervation in adult and developing zebrafish J Comp Neurol 497817ndash832 2006
C264 INVOLVEMENT OF ASICs IN FISH GILL Na TRANSPORT
AJP-Cell Physiol bull doi101152ajpcell003982013 bull wwwajpcellorg
22 Jonz MG Nurse CA New developments on gill innervation insightsfrom a model vertebrate J Exp Biol 211 2371ndash2378 2008
23 Kellenberger S Epithelial sodium and acid-sensing ion channels InSensing With Ion Channels edited by Martinac B New York Springer2008 chapt 11 p 225ndash246
24 Kellenberger S Schild L Epithelial sodium channeldegenerin family ofion channels a variety of functions for a shared structure Physiol Rev 82735ndash767 2002
25 Kleyman TR Cragoe EJ Amiloride and its analogs as tools in the studyof ion-transport J Membr Biol 105 1ndash21 1988
26 Krogh A The active absorption of ions in some freshwater animals ZVergl Physiol 25 335ndash350 1938
27 Levanti MB Guerrera MC Calavia MG Ciriaco E Montalbano GCobo J Germana A Vega JA Acid-sensing ion channel 2 (ASIC2) inthe intestine of adult zebrafish Neurosci Lett 494 24ndash28 2011
28 Lin H Randall DJ Evidence for the presence of an electrogenic protonpump on the trout gill epithelium J Exp Biol 161 119ndash134 1991
29 Maetz J Garcia Romeu F The mechanism of sodium and chlorideuptake by the gills of a fresh-water fish Carassius auratus II Evidencefor NH4
Na and HCO3Cl exchanges J Gen Physiol 47 1209ndash1227
196430 Nakada T Hoshijima K Esaki M Nagayoshi S Kawakami K Hirose
S Localization of ammonia transporter Rhcg1 in mitochondrion rich cellsof yolk sac gill and kidney of zebrafish and its ionic strength-dependentexpression Am J Physiol Regul Integr Comp Physiol 293 R1743ndashR17532007
31 Nawata CM Hung CC Tsui TK Wilson JM Wright PA Wood CMAmmonia excretion in rainbow trout (Oncorhynchus mykiss) evidence forRh glycoprotein and H-ATPase involvement Physiol Genomics 31463ndash474 2007
32 Parks SK Tresguerres M Galvez F Goss GG Intracellular pH regu-lation in isolated trout gill mitochondrion-rich (MR) cell subtypes evi-dence for NaH activity Comp Biochem Physiol A 155 139ndash1452010
33 Parks SK Tresguerres M Goss GG Interactions between Na channelsand Na-HCO3
cotransporters in the freshwater fish gill MRC a modelfor transepithelial Na uptake Am J Physiol Cell Physiol 292 C935ndashC944 2007
34 Parks SK Tresguerres M Goss GG Theoretical considerations under-lying Na uptake mechanisms in freshwater fishes Comp BiochemPhysiol C 148 411ndash418 2008
35 Paukert M Sidi S Russell C Siba M Wilson SW A family ofacid-sensing ion channels from zebrafish widespread expression in thecentral nervous system suggests a conserved role in neuronal communi-cation J Biol Chem 279 18783ndash18791 2004
36 Perry SF The chloride cell structure and function in the gill of freshwaterfish Annu Rev Physiol 59 325ndash347 1997
37 Perry SF Randall DJ Effects of amiloride and SITS on branchial ionfluxes in rainbow trout Salmo gairdneri J Exp Zool 215 225ndash228 1981
38 Preest MR Gonzales RJ Wilson RW Pharmacological examination ofNa and Cl transport in two species of freshwater fish Physiol BiochemZool 78 259ndash272 2005
39 Reid SD Hawkings GS Galvez F Goss GG Localization and charac-terization of phenamil-sensitive Na influx in isolated rainbow trout gillepithelial cells J Exp Biol 206 551ndash559 2003
40 Sakai H Lingueglia E Champigny G Mattei MG Lazdunski MCloning and functional expression of a novel degenerin-like Na channelgene in mammals J Physiol 519 323ndash333 1999
41 Shih TH Horng JL Liu ST Hwang PP Lin LY Rhcg1 and NHE3b areinvolved in ammonium-dependent sodium uptake by zebrafish larvaeacclimated to low-sodium water Am J Physiol Regul Integr Comp Physiol302 R84ndashR93 2012
42 Springauf A Grunder S An acid-sensing ion channel from shark(Squalus acanthias) mediates transient and sustained responses to protonsJ Physiol 588 809ndash820 2010
43 Spry DJ Wood CM Ion flux rates acid-base status and blood gases inrainbow trout Salmo gairdneri exposed to toxic zinc in natural soft waterCan J Fish Aquat Sci 42 1332ndash1337 1985
44 Sullivan GV Fryer JN Perry SF Immunolocalization of proton pumps(H-ATPase) in pavement cells of rainbow trout gill J Exp Biol 1982619ndash2629 1996
45 Tresguerres M Parks SK Katoh F Goss GG Microtubule-dependentrelocation of branchial V-H-ATPase to the basolateral membrane in thePacific spiny dogfish (Squalus acanthias) a role in base secretion J ExpBiol 209 599ndash609 2006
46 Vina E Parisi V Cabo R Laura R Lopez-Velasco S Lopez-Muniz AGarcia-Suarez O Germana A Vega JA Acid-sensing ion channels(ASICs) in the taste buds of adult zebrafish Neurosci Lett 536 35ndash402013
47 Waldmann R Lazdunski M H-gated cation channels neuronal acidsensors in the NaCDEG family of ion channels Curr Opin Neurobiol 8418ndash424 1998
48 Wilson JM Laurent P Tufts BL Benos DJ Donowitz M Vogl AWRandall DJ NaCl uptake by the branchial epithelium in freshwater teleostfish an immunological approach to ion-transport protein localization JExp Biol 203 2279ndash2296 2000
49 Wood CM Randall DJ Sodium balance in rainbow-trout (Salmo-gaird-neri) during extended exercise J Comp Physiol 82 235ndash256 1973
50 Wright PA Wood CM A new paradigm for ammonia excretion inaquatic animals role of rhesus (Rh) glycoproteins J Exp Biol 2122303ndash2312 2009
51 Yan JJ Chou MY Kaneko T Hwang PP Gene expression of NaH
exchanger in zebrafish H-ATPase-rich cells during acclimation to low-Na and acidic environments Am J Physiol Cell Physiol 293 C1814ndashC1823 2007
C265INVOLVEMENT OF ASICs IN FISH GILL Na TRANSPORT
AJP-Cell Physiol bull doi101152ajpcell003982013 bull wwwajpcellorg
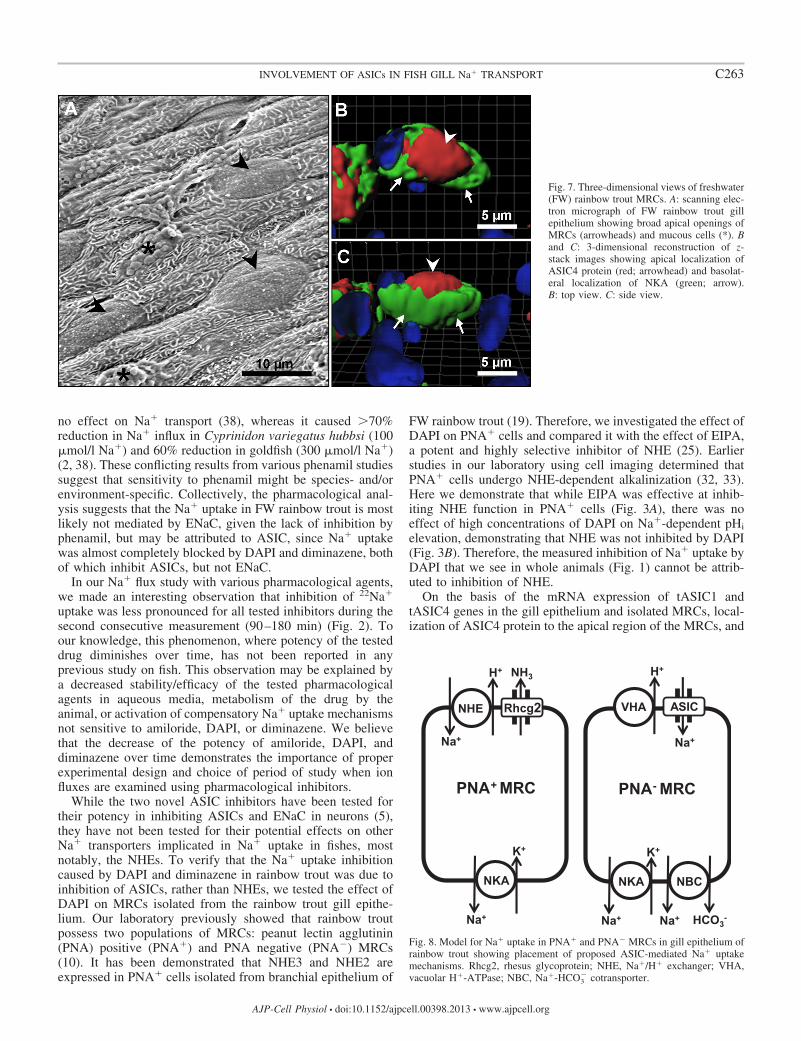
no effect on Na transport (38) whereas it caused 70reduction in Na influx in Cyprinidon variegatus hubbsi (100moll Na) and 60 reduction in goldfish (300 moll Na)(2 38) These conflicting results from various phenamil studiessuggest that sensitivity to phenamil might be species- andorenvironment-specific Collectively the pharmacological anal-ysis suggests that the Na uptake in FW rainbow trout is mostlikely not mediated by ENaC given the lack of inhibition byphenamil but may be attributed to ASIC since Na uptakewas almost completely blocked by DAPI and diminazene bothof which inhibit ASICs but not ENaC
In our Na flux study with various pharmacological agentswe made an interesting observation that inhibition of 22Na
uptake was less pronounced for all tested inhibitors during thesecond consecutive measurement (90ndash180 min) (Fig 2) Toour knowledge this phenomenon where potency of the testeddrug diminishes over time has not been reported in anyprevious study on fish This observation may be explained bya decreased stabilityefficacy of the tested pharmacologicalagents in aqueous media metabolism of the drug by theanimal or activation of compensatory Na uptake mechanismsnot sensitive to amiloride DAPI or diminazene We believethat the decrease of the potency of amiloride DAPI anddiminazene over time demonstrates the importance of properexperimental design and choice of period of study when ionfluxes are examined using pharmacological inhibitors
While the two novel ASIC inhibitors have been tested fortheir potency in inhibiting ASICs and ENaC in neurons (5)they have not been tested for their potential effects on otherNa transporters implicated in Na uptake in fishes mostnotably the NHEs To verify that the Na uptake inhibitioncaused by DAPI and diminazene in rainbow trout was due toinhibition of ASICs rather than NHEs we tested the effect ofDAPI on MRCs isolated from the rainbow trout gill epithe-lium Our laboratory previously showed that rainbow troutpossess two populations of MRCs peanut lectin agglutinin(PNA) positive (PNA) and PNA negative (PNA) MRCs(10) It has been demonstrated that NHE3 and NHE2 areexpressed in PNA cells isolated from branchial epithelium of
FW rainbow trout (19) Therefore we investigated the effect ofDAPI on PNA cells and compared it with the effect of EIPAa potent and highly selective inhibitor of NHE (25) Earlierstudies in our laboratory using cell imaging determined thatPNA cells undergo NHE-dependent alkalinization (32 33)Here we demonstrate that while EIPA was effective at inhib-iting NHE function in PNA cells (Fig 3A) there was noeffect of high concentrations of DAPI on Na-dependent pHi
elevation demonstrating that NHE was not inhibited by DAPI(Fig 3B) Therefore the measured inhibition of Na uptake byDAPI that we see in whole animals (Fig 1) cannot be attrib-uted to inhibition of NHE
On the basis of the mRNA expression of tASIC1 andtASIC4 genes in the gill epithelium and isolated MRCs local-ization of ASIC4 protein to the apical region of the MRCs and
Fig 7 Three-dimensional views of freshwater(FW) rainbow trout MRCs A scanning elec-tron micrograph of FW rainbow trout gillepithelium showing broad apical openings ofMRCs (arrowheads) and mucous cells () Band C 3-dimensional reconstruction of z-stack images showing apical localization ofASIC4 protein (red arrowhead) and basolat-eral localization of NKA (green arrow)B top view C side view
PNA- MRC
Na+
ASIC
H+
VHA
NKA
Na+
K+
NBC
Na+ HCO3-
PNA+ MRC
NH3
Na+
H+
NHE Rhcg2
NKA
Na+
K+
Fig 8 Model for Na uptake in PNA and PNA MRCs in gill epithelium ofrainbow trout showing placement of proposed ASIC-mediated Na uptakemechanisms Rhcg2 rhesus glycoprotein NHE NaH exchanger VHAvacuolar H-ATPase NBC Na-HCO3
cotransporter
C263INVOLVEMENT OF ASICs IN FISH GILL Na TRANSPORT
AJP-Cell Physiol bull doi101152ajpcell003982013 bull wwwajpcellorg
pharmacological blockade of Na influx by ASIC inhibitors inadult rainbow trout we propose a revised model for transepi-thelial Na uptake in FW rainbow trout gills (Fig 8) wherebyone apical mode of Na entry is via an ASIC4-mediatedmechanism
A number of recent studies on Na uptake in FW fishes havedemonstrated the presence of the NHERh metabolon in thegills of various species (17 30 31) As mentioned previouslywe specifically targeted the rainbow trout as a model for thisstudy as it is native to oligotrophic Na-poor mountainstreams with low buffering capacity This type of environmentpresents key thermodynamic challenges to the NHERhmetabolon as Na restrictions cannot be alleviated by thepresence of an Rh protein (6) Moreover searches of rainbowtrout genomes and transcriptomes have failed to identify anyENaC subunits ( 13 or ) despite the fact that they are knownto actively take up Na from water with 50 M Na (49)Furthermore other research studies have suggested and dem-onstrated an apical localization of the VHA in the gills ofrainbow trout (44 48) and bafilomycin a VHA-specific in-hibitor has been demonstrated to reduce Na uptake in ze-brafish (2) tilapia and carp (9) Previous studies have pro-posed VHA to be a part of the NHERh metabolon (41 50 51)however the placement of an apically oriented VHA is coun-terintuitive to the function of an NHE as it would exacerbatethe noted thermodynamic constraints Placement of a VHA inconcert with an apical ASIC is more plausible since it wouldnot only facilitate Na uptake from very low ionic strengthwater but could also provide a mechanism for gating ofASIC-mediated Na conductance It remains to be determinedif ASIC and VHA proteins colocalize to the same type ofMRCs and this warrants further investigation
Perspectives In summary we propose that ASCI4 is in-volved in mediating Na uptake from FW in rainbow troutacclimatized to low ionic strength low pH media This is thefirst demonstration of a nonneuronal function for any ASICfamily member and simultaneously provides an attractive so-lution to the debate surrounding the thermodynamic limitationsof the other proposed Na transport mechanisms and resolvesthe conundrum of the role of an apical VHA in Na transportThe possibility that other fish models may use similar mecha-nisms under different environmental conditions where modelssuch as the NHERh metabolon cannot be expected to functionremains to be explored
ACKNOWLEDGMENTS
We thank the staff in the Biosciences Aquatic Facilities the MolecularBiology Service Unit and the Cross Cancer Institute Cell Imaging Facility atthe University of Alberta for outstanding help throughout the study We alsothank Alex Clifford for advice and help with phylogenetic analysis and JamesEde for help with confocal microscopy
GRANTS
This study was supported by a Natural Sciences and Engineering ResearchCouncil of Canada Discovery Grant to G G Goss and an Alberta IngenuityStudent Scholarship to A K Dymowska
DISCLOSURES
No conflicts of interest financial or otherwise are declared by the authors
AUTHOR CONTRIBUTIONS
AKD AGS and GGG are responsible for conception and design ofthe research AKD AGS SDB and DC performed the experiments
AKD AGS SDB DC and GGG analyzed the data AKD AGSSDB DC and GGG interpreted the results of the experiments AKDAGS SDB and GGG prepared the figures AKD AGS SDB DCand GGG drafted the manuscript AKD AGS SDB DC and GGGedited and revised the manuscript AKD AGS SDB and GGGapproved the final version of the manuscript
REFERENCES
1 Avella M Bornancin M A new analysis of ammonia and sodiumtransport through the gills of the freshwater rainbow trout (Salmo gaird-neri) J Exp Biol 142 155ndash175 1989
2 Boisen AM Amstrup J Novak I Grosell M Sodium and chloridetransport in soft water and hard water acclimatized zebrafish (Daniorerio) Biochim Biophys Acta 1618 207ndash218 2003
3 Brix KV Grosell M Comparative characterization of Na transport inCyprinodon variegatus variegatus and Cyprinodon variegatus hubbsi amodel species complex for studying teleost invasion of freshwater J ExpBiol 215 1199ndash1209 2012
4 Chen X Polleichtner G Kadurin I Grunder S Zebrafish acid-sensingion channel (ASIC) 4 Characterization of homo- and heteromeric chan-nels and identification of regions important for activation by H J BiolChem 282 30406ndash30413 2007
5 Chen X Qiu L Li M Durrnagel S Orser BA Xiong ZG MacDonaldJF Diarylamidines high potency inhibitors of acid-sensing ion channelsNeuropharmacology 58 1045ndash1053 2010
6 Dymowska AK Hwang PP Goss GG Structure and function of iono-cytes in the freshwater fish gill Respir Physiol Neurobiol 184 282ndash2922012
7 Edwards SL Tse CM Toop T Immunolocalization of NHE3-likeimmunoreactivity in the gills of the rainbow trout (Oncorhynchus mykiss)and the blue-throated wrasse (Pseudolabrus tetrious) J Anat 195 465ndash469 1999
8 Evans DH Piermarini PM Choe KP The multifunctional gill domi-nant site of gas exchange osmoregulation acid-base regulation andexcretion of nitrogenous waste Physiol Rev 85 97ndash177 2005
9 Fenwick JC Bonga SE Flik G In vivo bafilomycin-sensitive Na
uptake in young freshwater fish J Exp Biol 202 3659ndash3666 199910 Galvez F Reid SD Hawkings G Goss GG Isolation and characteriza-
tion of mitochondria-rich cell types from the gill of freshwater rainbowtrout Am J Physiol Regul Integr Comp Physiol 282 R658ndashR668 2002
11 Goss G Orlowski J Grinstein S Coimmunoprecipitation of a 24-kDaprotein with NHE1 the ubiquitous isoform of the NaH exchanger AmJ Physiol Cell Physiol 270 C1493ndashC1502 1996
12 Goss GG Wood CJ Na and Cl uptake kinetics diffusive effluxes andacidic equivalent fluxes across the gills of rainbow trout 1 Responses toenvironmental hyperoxia J Exp Biol 152 521ndash547 1990
13 Grunder S Geissler HS Bassler EL Ruppersberg JP A new memberof acid-sensing ion channels from pituitary gland Neuroreport 11 1607ndash1611 2000
14 Harvey BJ Energization of sodium absorption by the H-ATPase pumpin mitochondria-rich cells of frog skin J Exp Biol 172 289ndash309 1992
15 Hirata T Kaneko T Ono T Nakazato T Furukawa N Hasegawa SWakabayashi S Shigekawa M Chang MH Romero MF Hirose SMechanism of acid adaptation of a fish living in low pH 35 lake Am JPhysiol Regul Integr Comp Physiol 284 R1199ndashR1212 2003
16 Holzer P Acid-sensitive ion channels and receptors Hand Exp Pharma-col 194 283ndash332 2009
17 Hung CY Tsui KN Wilson JM Nawata CM Wood CM Wright PARhesus glycoprotein gene expression in the mangrove killifish Kryptole-bias marmoratus exposed to elevated environmental ammonia levels andair J Exp Biol 210 2419ndash2429 2007
18 Hwang PP Lee TH Lin LY Ion regulations in fish gills recentprogresses in the cellular and molecular mechanisms Am J Physiol RegulIntegr Comp Physiol 301 R28ndashR47 2011
19 Ivanis G Esbaugh AJ Perry SF Branchial expression and localizationof SLC9A2 and SLC9A3 sodiumhydrogen exchangers and their possiblerole in acid-base regulation in freshwater rainbow trout (Oncorhynchusmykiss) J Exp Biol 211 2467ndash2477 2008
20 Jasti J Furukawa H Gonzales EB Gouaux E Structure of acid-sensingion channel 1 at 19A resolution and low pH Nature 449 316ndash323 2007
21 Jonz MG Nurse CA Epithelial mitochondria-rich cells and associatedinnervation in adult and developing zebrafish J Comp Neurol 497817ndash832 2006
C264 INVOLVEMENT OF ASICs IN FISH GILL Na TRANSPORT
AJP-Cell Physiol bull doi101152ajpcell003982013 bull wwwajpcellorg
22 Jonz MG Nurse CA New developments on gill innervation insightsfrom a model vertebrate J Exp Biol 211 2371ndash2378 2008
23 Kellenberger S Epithelial sodium and acid-sensing ion channels InSensing With Ion Channels edited by Martinac B New York Springer2008 chapt 11 p 225ndash246
24 Kellenberger S Schild L Epithelial sodium channeldegenerin family ofion channels a variety of functions for a shared structure Physiol Rev 82735ndash767 2002
25 Kleyman TR Cragoe EJ Amiloride and its analogs as tools in the studyof ion-transport J Membr Biol 105 1ndash21 1988
26 Krogh A The active absorption of ions in some freshwater animals ZVergl Physiol 25 335ndash350 1938
27 Levanti MB Guerrera MC Calavia MG Ciriaco E Montalbano GCobo J Germana A Vega JA Acid-sensing ion channel 2 (ASIC2) inthe intestine of adult zebrafish Neurosci Lett 494 24ndash28 2011
28 Lin H Randall DJ Evidence for the presence of an electrogenic protonpump on the trout gill epithelium J Exp Biol 161 119ndash134 1991
29 Maetz J Garcia Romeu F The mechanism of sodium and chlorideuptake by the gills of a fresh-water fish Carassius auratus II Evidencefor NH4
Na and HCO3Cl exchanges J Gen Physiol 47 1209ndash1227
196430 Nakada T Hoshijima K Esaki M Nagayoshi S Kawakami K Hirose
S Localization of ammonia transporter Rhcg1 in mitochondrion rich cellsof yolk sac gill and kidney of zebrafish and its ionic strength-dependentexpression Am J Physiol Regul Integr Comp Physiol 293 R1743ndashR17532007
31 Nawata CM Hung CC Tsui TK Wilson JM Wright PA Wood CMAmmonia excretion in rainbow trout (Oncorhynchus mykiss) evidence forRh glycoprotein and H-ATPase involvement Physiol Genomics 31463ndash474 2007
32 Parks SK Tresguerres M Galvez F Goss GG Intracellular pH regu-lation in isolated trout gill mitochondrion-rich (MR) cell subtypes evi-dence for NaH activity Comp Biochem Physiol A 155 139ndash1452010
33 Parks SK Tresguerres M Goss GG Interactions between Na channelsand Na-HCO3
cotransporters in the freshwater fish gill MRC a modelfor transepithelial Na uptake Am J Physiol Cell Physiol 292 C935ndashC944 2007
34 Parks SK Tresguerres M Goss GG Theoretical considerations under-lying Na uptake mechanisms in freshwater fishes Comp BiochemPhysiol C 148 411ndash418 2008
35 Paukert M Sidi S Russell C Siba M Wilson SW A family ofacid-sensing ion channels from zebrafish widespread expression in thecentral nervous system suggests a conserved role in neuronal communi-cation J Biol Chem 279 18783ndash18791 2004
36 Perry SF The chloride cell structure and function in the gill of freshwaterfish Annu Rev Physiol 59 325ndash347 1997
37 Perry SF Randall DJ Effects of amiloride and SITS on branchial ionfluxes in rainbow trout Salmo gairdneri J Exp Zool 215 225ndash228 1981
38 Preest MR Gonzales RJ Wilson RW Pharmacological examination ofNa and Cl transport in two species of freshwater fish Physiol BiochemZool 78 259ndash272 2005
39 Reid SD Hawkings GS Galvez F Goss GG Localization and charac-terization of phenamil-sensitive Na influx in isolated rainbow trout gillepithelial cells J Exp Biol 206 551ndash559 2003
40 Sakai H Lingueglia E Champigny G Mattei MG Lazdunski MCloning and functional expression of a novel degenerin-like Na channelgene in mammals J Physiol 519 323ndash333 1999
41 Shih TH Horng JL Liu ST Hwang PP Lin LY Rhcg1 and NHE3b areinvolved in ammonium-dependent sodium uptake by zebrafish larvaeacclimated to low-sodium water Am J Physiol Regul Integr Comp Physiol302 R84ndashR93 2012
42 Springauf A Grunder S An acid-sensing ion channel from shark(Squalus acanthias) mediates transient and sustained responses to protonsJ Physiol 588 809ndash820 2010
43 Spry DJ Wood CM Ion flux rates acid-base status and blood gases inrainbow trout Salmo gairdneri exposed to toxic zinc in natural soft waterCan J Fish Aquat Sci 42 1332ndash1337 1985
44 Sullivan GV Fryer JN Perry SF Immunolocalization of proton pumps(H-ATPase) in pavement cells of rainbow trout gill J Exp Biol 1982619ndash2629 1996
45 Tresguerres M Parks SK Katoh F Goss GG Microtubule-dependentrelocation of branchial V-H-ATPase to the basolateral membrane in thePacific spiny dogfish (Squalus acanthias) a role in base secretion J ExpBiol 209 599ndash609 2006
46 Vina E Parisi V Cabo R Laura R Lopez-Velasco S Lopez-Muniz AGarcia-Suarez O Germana A Vega JA Acid-sensing ion channels(ASICs) in the taste buds of adult zebrafish Neurosci Lett 536 35ndash402013
47 Waldmann R Lazdunski M H-gated cation channels neuronal acidsensors in the NaCDEG family of ion channels Curr Opin Neurobiol 8418ndash424 1998
48 Wilson JM Laurent P Tufts BL Benos DJ Donowitz M Vogl AWRandall DJ NaCl uptake by the branchial epithelium in freshwater teleostfish an immunological approach to ion-transport protein localization JExp Biol 203 2279ndash2296 2000
49 Wood CM Randall DJ Sodium balance in rainbow-trout (Salmo-gaird-neri) during extended exercise J Comp Physiol 82 235ndash256 1973
50 Wright PA Wood CM A new paradigm for ammonia excretion inaquatic animals role of rhesus (Rh) glycoproteins J Exp Biol 2122303ndash2312 2009
51 Yan JJ Chou MY Kaneko T Hwang PP Gene expression of NaH
exchanger in zebrafish H-ATPase-rich cells during acclimation to low-Na and acidic environments Am J Physiol Cell Physiol 293 C1814ndashC1823 2007
C265INVOLVEMENT OF ASICs IN FISH GILL Na TRANSPORT
AJP-Cell Physiol bull doi101152ajpcell003982013 bull wwwajpcellorg

pharmacological blockade of Na influx by ASIC inhibitors inadult rainbow trout we propose a revised model for transepi-thelial Na uptake in FW rainbow trout gills (Fig 8) wherebyone apical mode of Na entry is via an ASIC4-mediatedmechanism
A number of recent studies on Na uptake in FW fishes havedemonstrated the presence of the NHERh metabolon in thegills of various species (17 30 31) As mentioned previouslywe specifically targeted the rainbow trout as a model for thisstudy as it is native to oligotrophic Na-poor mountainstreams with low buffering capacity This type of environmentpresents key thermodynamic challenges to the NHERhmetabolon as Na restrictions cannot be alleviated by thepresence of an Rh protein (6) Moreover searches of rainbowtrout genomes and transcriptomes have failed to identify anyENaC subunits ( 13 or ) despite the fact that they are knownto actively take up Na from water with 50 M Na (49)Furthermore other research studies have suggested and dem-onstrated an apical localization of the VHA in the gills ofrainbow trout (44 48) and bafilomycin a VHA-specific in-hibitor has been demonstrated to reduce Na uptake in ze-brafish (2) tilapia and carp (9) Previous studies have pro-posed VHA to be a part of the NHERh metabolon (41 50 51)however the placement of an apically oriented VHA is coun-terintuitive to the function of an NHE as it would exacerbatethe noted thermodynamic constraints Placement of a VHA inconcert with an apical ASIC is more plausible since it wouldnot only facilitate Na uptake from very low ionic strengthwater but could also provide a mechanism for gating ofASIC-mediated Na conductance It remains to be determinedif ASIC and VHA proteins colocalize to the same type ofMRCs and this warrants further investigation
Perspectives In summary we propose that ASCI4 is in-volved in mediating Na uptake from FW in rainbow troutacclimatized to low ionic strength low pH media This is thefirst demonstration of a nonneuronal function for any ASICfamily member and simultaneously provides an attractive so-lution to the debate surrounding the thermodynamic limitationsof the other proposed Na transport mechanisms and resolvesthe conundrum of the role of an apical VHA in Na transportThe possibility that other fish models may use similar mecha-nisms under different environmental conditions where modelssuch as the NHERh metabolon cannot be expected to functionremains to be explored
ACKNOWLEDGMENTS
We thank the staff in the Biosciences Aquatic Facilities the MolecularBiology Service Unit and the Cross Cancer Institute Cell Imaging Facility atthe University of Alberta for outstanding help throughout the study We alsothank Alex Clifford for advice and help with phylogenetic analysis and JamesEde for help with confocal microscopy
GRANTS
This study was supported by a Natural Sciences and Engineering ResearchCouncil of Canada Discovery Grant to G G Goss and an Alberta IngenuityStudent Scholarship to A K Dymowska
DISCLOSURES
No conflicts of interest financial or otherwise are declared by the authors
AUTHOR CONTRIBUTIONS
AKD AGS and GGG are responsible for conception and design ofthe research AKD AGS SDB and DC performed the experiments
AKD AGS SDB DC and GGG analyzed the data AKD AGSSDB DC and GGG interpreted the results of the experiments AKDAGS SDB and GGG prepared the figures AKD AGS SDB DCand GGG drafted the manuscript AKD AGS SDB DC and GGGedited and revised the manuscript AKD AGS SDB and GGGapproved the final version of the manuscript
REFERENCES
1 Avella M Bornancin M A new analysis of ammonia and sodiumtransport through the gills of the freshwater rainbow trout (Salmo gaird-neri) J Exp Biol 142 155ndash175 1989
2 Boisen AM Amstrup J Novak I Grosell M Sodium and chloridetransport in soft water and hard water acclimatized zebrafish (Daniorerio) Biochim Biophys Acta 1618 207ndash218 2003
3 Brix KV Grosell M Comparative characterization of Na transport inCyprinodon variegatus variegatus and Cyprinodon variegatus hubbsi amodel species complex for studying teleost invasion of freshwater J ExpBiol 215 1199ndash1209 2012
4 Chen X Polleichtner G Kadurin I Grunder S Zebrafish acid-sensingion channel (ASIC) 4 Characterization of homo- and heteromeric chan-nels and identification of regions important for activation by H J BiolChem 282 30406ndash30413 2007
5 Chen X Qiu L Li M Durrnagel S Orser BA Xiong ZG MacDonaldJF Diarylamidines high potency inhibitors of acid-sensing ion channelsNeuropharmacology 58 1045ndash1053 2010
6 Dymowska AK Hwang PP Goss GG Structure and function of iono-cytes in the freshwater fish gill Respir Physiol Neurobiol 184 282ndash2922012
7 Edwards SL Tse CM Toop T Immunolocalization of NHE3-likeimmunoreactivity in the gills of the rainbow trout (Oncorhynchus mykiss)and the blue-throated wrasse (Pseudolabrus tetrious) J Anat 195 465ndash469 1999
8 Evans DH Piermarini PM Choe KP The multifunctional gill domi-nant site of gas exchange osmoregulation acid-base regulation andexcretion of nitrogenous waste Physiol Rev 85 97ndash177 2005
9 Fenwick JC Bonga SE Flik G In vivo bafilomycin-sensitive Na
uptake in young freshwater fish J Exp Biol 202 3659ndash3666 199910 Galvez F Reid SD Hawkings G Goss GG Isolation and characteriza-
tion of mitochondria-rich cell types from the gill of freshwater rainbowtrout Am J Physiol Regul Integr Comp Physiol 282 R658ndashR668 2002
11 Goss G Orlowski J Grinstein S Coimmunoprecipitation of a 24-kDaprotein with NHE1 the ubiquitous isoform of the NaH exchanger AmJ Physiol Cell Physiol 270 C1493ndashC1502 1996
12 Goss GG Wood CJ Na and Cl uptake kinetics diffusive effluxes andacidic equivalent fluxes across the gills of rainbow trout 1 Responses toenvironmental hyperoxia J Exp Biol 152 521ndash547 1990
13 Grunder S Geissler HS Bassler EL Ruppersberg JP A new memberof acid-sensing ion channels from pituitary gland Neuroreport 11 1607ndash1611 2000
14 Harvey BJ Energization of sodium absorption by the H-ATPase pumpin mitochondria-rich cells of frog skin J Exp Biol 172 289ndash309 1992
15 Hirata T Kaneko T Ono T Nakazato T Furukawa N Hasegawa SWakabayashi S Shigekawa M Chang MH Romero MF Hirose SMechanism of acid adaptation of a fish living in low pH 35 lake Am JPhysiol Regul Integr Comp Physiol 284 R1199ndashR1212 2003
16 Holzer P Acid-sensitive ion channels and receptors Hand Exp Pharma-col 194 283ndash332 2009
17 Hung CY Tsui KN Wilson JM Nawata CM Wood CM Wright PARhesus glycoprotein gene expression in the mangrove killifish Kryptole-bias marmoratus exposed to elevated environmental ammonia levels andair J Exp Biol 210 2419ndash2429 2007
18 Hwang PP Lee TH Lin LY Ion regulations in fish gills recentprogresses in the cellular and molecular mechanisms Am J Physiol RegulIntegr Comp Physiol 301 R28ndashR47 2011
19 Ivanis G Esbaugh AJ Perry SF Branchial expression and localizationof SLC9A2 and SLC9A3 sodiumhydrogen exchangers and their possiblerole in acid-base regulation in freshwater rainbow trout (Oncorhynchusmykiss) J Exp Biol 211 2467ndash2477 2008
20 Jasti J Furukawa H Gonzales EB Gouaux E Structure of acid-sensingion channel 1 at 19A resolution and low pH Nature 449 316ndash323 2007
21 Jonz MG Nurse CA Epithelial mitochondria-rich cells and associatedinnervation in adult and developing zebrafish J Comp Neurol 497817ndash832 2006
C264 INVOLVEMENT OF ASICs IN FISH GILL Na TRANSPORT
AJP-Cell Physiol bull doi101152ajpcell003982013 bull wwwajpcellorg
22 Jonz MG Nurse CA New developments on gill innervation insightsfrom a model vertebrate J Exp Biol 211 2371ndash2378 2008
23 Kellenberger S Epithelial sodium and acid-sensing ion channels InSensing With Ion Channels edited by Martinac B New York Springer2008 chapt 11 p 225ndash246
24 Kellenberger S Schild L Epithelial sodium channeldegenerin family ofion channels a variety of functions for a shared structure Physiol Rev 82735ndash767 2002
25 Kleyman TR Cragoe EJ Amiloride and its analogs as tools in the studyof ion-transport J Membr Biol 105 1ndash21 1988
26 Krogh A The active absorption of ions in some freshwater animals ZVergl Physiol 25 335ndash350 1938
27 Levanti MB Guerrera MC Calavia MG Ciriaco E Montalbano GCobo J Germana A Vega JA Acid-sensing ion channel 2 (ASIC2) inthe intestine of adult zebrafish Neurosci Lett 494 24ndash28 2011
28 Lin H Randall DJ Evidence for the presence of an electrogenic protonpump on the trout gill epithelium J Exp Biol 161 119ndash134 1991
29 Maetz J Garcia Romeu F The mechanism of sodium and chlorideuptake by the gills of a fresh-water fish Carassius auratus II Evidencefor NH4
Na and HCO3Cl exchanges J Gen Physiol 47 1209ndash1227
196430 Nakada T Hoshijima K Esaki M Nagayoshi S Kawakami K Hirose
S Localization of ammonia transporter Rhcg1 in mitochondrion rich cellsof yolk sac gill and kidney of zebrafish and its ionic strength-dependentexpression Am J Physiol Regul Integr Comp Physiol 293 R1743ndashR17532007
31 Nawata CM Hung CC Tsui TK Wilson JM Wright PA Wood CMAmmonia excretion in rainbow trout (Oncorhynchus mykiss) evidence forRh glycoprotein and H-ATPase involvement Physiol Genomics 31463ndash474 2007
32 Parks SK Tresguerres M Galvez F Goss GG Intracellular pH regu-lation in isolated trout gill mitochondrion-rich (MR) cell subtypes evi-dence for NaH activity Comp Biochem Physiol A 155 139ndash1452010
33 Parks SK Tresguerres M Goss GG Interactions between Na channelsand Na-HCO3
cotransporters in the freshwater fish gill MRC a modelfor transepithelial Na uptake Am J Physiol Cell Physiol 292 C935ndashC944 2007
34 Parks SK Tresguerres M Goss GG Theoretical considerations under-lying Na uptake mechanisms in freshwater fishes Comp BiochemPhysiol C 148 411ndash418 2008
35 Paukert M Sidi S Russell C Siba M Wilson SW A family ofacid-sensing ion channels from zebrafish widespread expression in thecentral nervous system suggests a conserved role in neuronal communi-cation J Biol Chem 279 18783ndash18791 2004
36 Perry SF The chloride cell structure and function in the gill of freshwaterfish Annu Rev Physiol 59 325ndash347 1997
37 Perry SF Randall DJ Effects of amiloride and SITS on branchial ionfluxes in rainbow trout Salmo gairdneri J Exp Zool 215 225ndash228 1981
38 Preest MR Gonzales RJ Wilson RW Pharmacological examination ofNa and Cl transport in two species of freshwater fish Physiol BiochemZool 78 259ndash272 2005
39 Reid SD Hawkings GS Galvez F Goss GG Localization and charac-terization of phenamil-sensitive Na influx in isolated rainbow trout gillepithelial cells J Exp Biol 206 551ndash559 2003
40 Sakai H Lingueglia E Champigny G Mattei MG Lazdunski MCloning and functional expression of a novel degenerin-like Na channelgene in mammals J Physiol 519 323ndash333 1999
41 Shih TH Horng JL Liu ST Hwang PP Lin LY Rhcg1 and NHE3b areinvolved in ammonium-dependent sodium uptake by zebrafish larvaeacclimated to low-sodium water Am J Physiol Regul Integr Comp Physiol302 R84ndashR93 2012
42 Springauf A Grunder S An acid-sensing ion channel from shark(Squalus acanthias) mediates transient and sustained responses to protonsJ Physiol 588 809ndash820 2010
43 Spry DJ Wood CM Ion flux rates acid-base status and blood gases inrainbow trout Salmo gairdneri exposed to toxic zinc in natural soft waterCan J Fish Aquat Sci 42 1332ndash1337 1985
44 Sullivan GV Fryer JN Perry SF Immunolocalization of proton pumps(H-ATPase) in pavement cells of rainbow trout gill J Exp Biol 1982619ndash2629 1996
45 Tresguerres M Parks SK Katoh F Goss GG Microtubule-dependentrelocation of branchial V-H-ATPase to the basolateral membrane in thePacific spiny dogfish (Squalus acanthias) a role in base secretion J ExpBiol 209 599ndash609 2006
46 Vina E Parisi V Cabo R Laura R Lopez-Velasco S Lopez-Muniz AGarcia-Suarez O Germana A Vega JA Acid-sensing ion channels(ASICs) in the taste buds of adult zebrafish Neurosci Lett 536 35ndash402013
47 Waldmann R Lazdunski M H-gated cation channels neuronal acidsensors in the NaCDEG family of ion channels Curr Opin Neurobiol 8418ndash424 1998
48 Wilson JM Laurent P Tufts BL Benos DJ Donowitz M Vogl AWRandall DJ NaCl uptake by the branchial epithelium in freshwater teleostfish an immunological approach to ion-transport protein localization JExp Biol 203 2279ndash2296 2000
49 Wood CM Randall DJ Sodium balance in rainbow-trout (Salmo-gaird-neri) during extended exercise J Comp Physiol 82 235ndash256 1973
50 Wright PA Wood CM A new paradigm for ammonia excretion inaquatic animals role of rhesus (Rh) glycoproteins J Exp Biol 2122303ndash2312 2009
51 Yan JJ Chou MY Kaneko T Hwang PP Gene expression of NaH
exchanger in zebrafish H-ATPase-rich cells during acclimation to low-Na and acidic environments Am J Physiol Cell Physiol 293 C1814ndashC1823 2007
C265INVOLVEMENT OF ASICs IN FISH GILL Na TRANSPORT
AJP-Cell Physiol bull doi101152ajpcell003982013 bull wwwajpcellorg

22 Jonz MG Nurse CA New developments on gill innervation insightsfrom a model vertebrate J Exp Biol 211 2371ndash2378 2008
23 Kellenberger S Epithelial sodium and acid-sensing ion channels InSensing With Ion Channels edited by Martinac B New York Springer2008 chapt 11 p 225ndash246
24 Kellenberger S Schild L Epithelial sodium channeldegenerin family ofion channels a variety of functions for a shared structure Physiol Rev 82735ndash767 2002
25 Kleyman TR Cragoe EJ Amiloride and its analogs as tools in the studyof ion-transport J Membr Biol 105 1ndash21 1988
26 Krogh A The active absorption of ions in some freshwater animals ZVergl Physiol 25 335ndash350 1938
27 Levanti MB Guerrera MC Calavia MG Ciriaco E Montalbano GCobo J Germana A Vega JA Acid-sensing ion channel 2 (ASIC2) inthe intestine of adult zebrafish Neurosci Lett 494 24ndash28 2011
28 Lin H Randall DJ Evidence for the presence of an electrogenic protonpump on the trout gill epithelium J Exp Biol 161 119ndash134 1991
29 Maetz J Garcia Romeu F The mechanism of sodium and chlorideuptake by the gills of a fresh-water fish Carassius auratus II Evidencefor NH4
Na and HCO3Cl exchanges J Gen Physiol 47 1209ndash1227
196430 Nakada T Hoshijima K Esaki M Nagayoshi S Kawakami K Hirose
S Localization of ammonia transporter Rhcg1 in mitochondrion rich cellsof yolk sac gill and kidney of zebrafish and its ionic strength-dependentexpression Am J Physiol Regul Integr Comp Physiol 293 R1743ndashR17532007
31 Nawata CM Hung CC Tsui TK Wilson JM Wright PA Wood CMAmmonia excretion in rainbow trout (Oncorhynchus mykiss) evidence forRh glycoprotein and H-ATPase involvement Physiol Genomics 31463ndash474 2007
32 Parks SK Tresguerres M Galvez F Goss GG Intracellular pH regu-lation in isolated trout gill mitochondrion-rich (MR) cell subtypes evi-dence for NaH activity Comp Biochem Physiol A 155 139ndash1452010
33 Parks SK Tresguerres M Goss GG Interactions between Na channelsand Na-HCO3
cotransporters in the freshwater fish gill MRC a modelfor transepithelial Na uptake Am J Physiol Cell Physiol 292 C935ndashC944 2007
34 Parks SK Tresguerres M Goss GG Theoretical considerations under-lying Na uptake mechanisms in freshwater fishes Comp BiochemPhysiol C 148 411ndash418 2008
35 Paukert M Sidi S Russell C Siba M Wilson SW A family ofacid-sensing ion channels from zebrafish widespread expression in thecentral nervous system suggests a conserved role in neuronal communi-cation J Biol Chem 279 18783ndash18791 2004
36 Perry SF The chloride cell structure and function in the gill of freshwaterfish Annu Rev Physiol 59 325ndash347 1997
37 Perry SF Randall DJ Effects of amiloride and SITS on branchial ionfluxes in rainbow trout Salmo gairdneri J Exp Zool 215 225ndash228 1981
38 Preest MR Gonzales RJ Wilson RW Pharmacological examination ofNa and Cl transport in two species of freshwater fish Physiol BiochemZool 78 259ndash272 2005
39 Reid SD Hawkings GS Galvez F Goss GG Localization and charac-terization of phenamil-sensitive Na influx in isolated rainbow trout gillepithelial cells J Exp Biol 206 551ndash559 2003
40 Sakai H Lingueglia E Champigny G Mattei MG Lazdunski MCloning and functional expression of a novel degenerin-like Na channelgene in mammals J Physiol 519 323ndash333 1999
41 Shih TH Horng JL Liu ST Hwang PP Lin LY Rhcg1 and NHE3b areinvolved in ammonium-dependent sodium uptake by zebrafish larvaeacclimated to low-sodium water Am J Physiol Regul Integr Comp Physiol302 R84ndashR93 2012
42 Springauf A Grunder S An acid-sensing ion channel from shark(Squalus acanthias) mediates transient and sustained responses to protonsJ Physiol 588 809ndash820 2010
43 Spry DJ Wood CM Ion flux rates acid-base status and blood gases inrainbow trout Salmo gairdneri exposed to toxic zinc in natural soft waterCan J Fish Aquat Sci 42 1332ndash1337 1985
44 Sullivan GV Fryer JN Perry SF Immunolocalization of proton pumps(H-ATPase) in pavement cells of rainbow trout gill J Exp Biol 1982619ndash2629 1996
45 Tresguerres M Parks SK Katoh F Goss GG Microtubule-dependentrelocation of branchial V-H-ATPase to the basolateral membrane in thePacific spiny dogfish (Squalus acanthias) a role in base secretion J ExpBiol 209 599ndash609 2006
46 Vina E Parisi V Cabo R Laura R Lopez-Velasco S Lopez-Muniz AGarcia-Suarez O Germana A Vega JA Acid-sensing ion channels(ASICs) in the taste buds of adult zebrafish Neurosci Lett 536 35ndash402013
47 Waldmann R Lazdunski M H-gated cation channels neuronal acidsensors in the NaCDEG family of ion channels Curr Opin Neurobiol 8418ndash424 1998
48 Wilson JM Laurent P Tufts BL Benos DJ Donowitz M Vogl AWRandall DJ NaCl uptake by the branchial epithelium in freshwater teleostfish an immunological approach to ion-transport protein localization JExp Biol 203 2279ndash2296 2000
49 Wood CM Randall DJ Sodium balance in rainbow-trout (Salmo-gaird-neri) during extended exercise J Comp Physiol 82 235ndash256 1973
50 Wright PA Wood CM A new paradigm for ammonia excretion inaquatic animals role of rhesus (Rh) glycoproteins J Exp Biol 2122303ndash2312 2009
51 Yan JJ Chou MY Kaneko T Hwang PP Gene expression of NaH
exchanger in zebrafish H-ATPase-rich cells during acclimation to low-Na and acidic environments Am J Physiol Cell Physiol 293 C1814ndashC1823 2007
C265INVOLVEMENT OF ASICs IN FISH GILL Na TRANSPORT
AJP-Cell Physiol bull doi101152ajpcell003982013 bull wwwajpcellorg
Related Documents











