Technische Universitt München Wissenschaftszentrum Weihenstephan für Ernhrung, Landnutzung und Umwelt Department für Tierwissenschaften Fachgebiet Tierernhrung Experimentelle Untersuchungen zur Ableitung des Tryptophanbedarfs laktierender Zuchtsauen anhand verschiedener Leistungs- und Stoffwechselparameter Franz Gabriel Pampuch Vollstndiger Abdruck der von der Fakultt Wissenschaftszentrum Weihenstephan für Ernhrung, Landnutzung und Umwelt der Technischen Universitt München zur Erlangung des akademischen Grades eines Doktors der Agrarwissenschaften (Dr. agr.) genehmigten Dissertation. Vorsitzender: Univ.-Prof. Dr. H.-R. Fries Prüfer der Dissertation: 1. Univ.-Prof. Dr. D. A. Roth-Maier 2. Univ.-Prof. Dr. J. Bauer Die Dissertation wurde am 09.09.2003 bei der Technischen Universitt München eingereicht und durch die Fakultt Wissenschaftszentrum Weihenstephan für Ernhrung, Landnutzung und Umwelt am 20.10.2003 angenommen.

Welcome message from author
This document is posted to help you gain knowledge. Please leave a comment to let me know what you think about it! Share it to your friends and learn new things together.
Transcript

Technische Universität München
Wissenschaftszentrum Weihenstephan für Ernährung, Landnutzung und Umwelt
Department für Tierwissenschaften
Fachgebiet Tierernährung
Experimentelle Untersuchungen zur Ableitung des
Tryptophanbedarfs laktierender Zuchtsauen
anhand verschiedener Leistungs- und Stoffwechselparameter
Franz Gabriel Pampuch
Vollständiger Abdruck der von der Fakultät Wissenschaftszentrum Weihenstephan
für Ernährung, Landnutzung und Umwelt der Technischen Universität München zur
Erlangung des akademischen Grades eines
Doktors der Agrarwissenschaften
(Dr. agr.)
genehmigten Dissertation.
Vorsitzender: Univ.-Prof. Dr. H.-R. Fries
Prüfer der Dissertation:
1. Univ.-Prof. Dr. D. A. Roth-Maier
2. Univ.-Prof. Dr. J. Bauer
Die Dissertation wurde am 09.09.2003 bei der Technischen Universität München eingereicht und
durch die Fakultät Wissenschaftszentrum Weihenstephan für Ernährung, Landnutzung und Umwelt
am 20.10.2003 angenommen.


Mein herzlicher Dank gilt Frau Prof. Dr. D.A. Roth-Maier für die Überlassung des Themas, die
wissenschaftliche Anleitung sowie die stets gewährte Unterstützung und die freundliche Betreuung der
Arbeit.
Bei Frau Dr. B.R. Paulicks möchte ich mich sehr herzlich für die freundliche und konstruktive
Betreuung der Arbeit sowie ihre stete Gesprächsbereitschaft und Unterstützung bedanken.
Weiterhin gilt mein ganz besonderer Dank Frau Viola Löbnitz und Herrn Steffen Löbnitz, die mich mit
überdurchschnittlichem Engagement stets freundlich und gewissenhaft bei der Durchführung des
Versuchs unterstützt haben.
Für die Aminosäurenanalysen sowie für die Überlassung der kristallinen Aminosäuren möchte ich
mich bei der Firma Lohmann Animal Health GmbH & Co. KG, Cuxhaven sehr bedanken.
Der Firma DLD Diagnostika GmbH, Hamburg danke ich sehr für die kostenlose Durchführung der
Serotoninanalysen.
Für die gewissenhafte und kostenlose Untersuchung der Sauenmilch möchte ich dem Milchprüfring
Bayern e.V., Wolnzach meinen Dank aussprechen.
Danken möchte ich auch allen Mitarbeiterinnen und Mitarbeitern der Fachgebiete Tierernährung sowie
des Versuchgutes Hirschau, die mich bei meiner Arbeit tatkräftig und stets freundlich unterstützt
haben.


1
Inhaltsverzeichnis 1. Einleitung und Problemstellung 132. Material und Methoden 162.1. Versuchsbetrieb 162.1.1. Tiermaterial 162.1.2. Aufstallung der Tiere 162.1.3. Fütterung der Tiere 172.1.4. Behandlung der Tiere 172.2. Minimierung systematischer Einflussgrößen 182.2.1. Lebendmasse, Alter und Genetik 182.2.2. Jahreszeit 192.2.3. Ferkelzahl je Wurf 192.3. Fütterung 202.3.1. Trächtigkeitsfütterung 202.3.2. Laktationsfütterung 212.3.3. Ferkelfütterung 262.4. Verdauungsversuche 272.4.1. Ermittlung der Verdaulichkeit der Rohnährstoffe 272.4.2. Ermittlung der ilealen Verdaulichkeit der Aminosäuren 282.5. Erfassung der Untersuchungsparameter 302.5.1. Lebendmasse der Sauen 302.5.2. Futteraufnahme der Sauen 302.5.3. Milchleistung der Sauen 302.5.4. Lebendmasse der Ferkel 332.5.5. Futterverbrauch der Ferkel 342.6. Probenahme und Analysenverfahren 342.6.1. Futter- und Kotproben 342.6.1.1. Nährstoffanalyse 342.6.1.2. Aminosäurenbestimmung 342.6.1.2.1. Tryptophanbestimmung 342.6.1.2.2. Bestimmung der restlichen Aminosäuren 362.6.1.3. Energiegehalt 372.6.2. Milchproben 372.6.2.1. Fett-, Eiweiß- und Laktosebestimmung 382.6.2.2. Harnstoffbestimmung 382.6.2.3. Trockensubstanz- und Aminosäurenbestimmung 392.6.2.3.1. Trockensubstanzbestimmung 392.6.2.3.2. Tryptophanbestimmung 392.6.2.3.3. Bestimmung der restlichen Aminosäuren 39

2
2.6.2.4. Energiegehalt 392.6.3. Blutproben 392.6.3.1. Aminosäuren- und Harnstoffbestimmung 402.6.3.2. Hämatokrit- und Thrombozytenbestimmung 402.6.3.3. Serotoninbestimmung 412.7. Statistische Auswertung und Ergebnisdarstellung 423. Ergebnisse 443.1. Verdaulichkeiten 443.1.1. Verdaulichkeit und Energiegehalt des Laktationsfutters 443.1.2. Ileale Verdaulichkeit des Rohproteins und ausgewählter
Aminosäuren 45
3.2. Futter-, Nährstoff- und Energieaufnahme der Sauen 453.2.1. Futteraufnahme 453.2.2. Proteinaufnahme 463.2.3. Tryptophanaufnahme 473.2.4. Energieaufnahme 483.3. Lebendmasse und -entwicklung der Sauen 493.4. Milchleistung der Sauen 503.5. Inhaltsstoffe und Energiegehalt der Sauenmilch 503.5.1. Trockensubstanz in der Sauenmilch 503.5.2. Fettgehalt und -ausscheidung 513.5.3. Eiweißgehalt und -ausscheidung 523.5.4. Laktosegehalt und -ausscheidung 533.5.5. Harnstoffgehalt und -ausscheidung 543.5.6. Aminosäurengehalt und -ausscheidung 553.5.7. Energiegehalt und -ausscheidung 583.6. Blutparameter 583.6.1. Aminosäurengehalt 583.6.1.1. Essentielle und semiessentielle Aminosäuren 583.6.1.2. Nicht-essentielle Aminosäuren 603.6.2. Harnstoffgehalt 613.6.3. Hämatokritwert 623.6.4. Thrombozytengehalt 623.6.5. Serotoningehalt im Serum 623.6.6. Serotoningehalt im Vollblut 633.6.7. Serotoningehalt je 109 Thrombozyten 643.7. Futterverbrauch der Ferkel 643.8. Lebendmasse und -entwicklung der Ferkel 654. Diskussion 674.1. Modelle zur Bedarfsableitung 69

3
4.1.1. Abschnittsweise definierte Modelle 694.1.1.1. Das Broken-Line Modell 704.1.1.2. Das Reading Modell 714.1.2. Das quadratische Modell 724.1.3. Asymptotische Modelle 734.2. Tryptophan 744.2.1. Struktur 744.2.2. Stoffwechsel 754.2.2.1. Allgemeiner Stoffwechsel 764.2.2.2. Tryptophan und Serotonin 804.2.2.3. Tryptophan und Niacin 834.2.3. Scheinbare ileale Verdaulichkeit in Futtermitteln 854.3. Auswirkung des Tryptophangehaltes im Futter auf die
verschiedenen untersuchten Versuchsparameter 87
4.3.1. Futteraufnahme der Sauen 874.3.2. Lebendmasse der Sauen 904.3.3. Milchabgabe der Sauen 954.3.3.1. Milchmenge 994.3.3.2. Trockenmasse 1014.3.3.3. Fettgehalt 1014.3.3.4. Eiweißgehalt 1024.3.3.5. Laktosegehalt 1044.3.3.6. Harnstoffgehalt 1054.3.3.7. Energiegehalt 1084.3.3.8. Aminosäurenzusammensetzung 1094.3.4. Blutparameter der Sauen 1114.3.4.1. Aminosäurengehalt im Plasma 1114.3.4.1.1. Tryptophan 1114.3.4.1.2. Essentielle Aminosäuren 1134.3.4.1.3. Nicht-essentielle Aminosäuren 1144.3.4.2. Harnstoffgehalt im Plasma 1154.3.4.3. Serotoningehalt 1184.3.5. Lebendmasse der Ferkel 1224.3.6. Futterverbrauch der Ferkel 124
5. Schlussbetrachtung 1266. Zusammenfassung 1337. Summary 1368. Literaturverzeichnis 1389. Anhang 173

4
Tabellenverzeichnis Tabelle 1: Zulage und Gesamtgehalt an Tryptophan im Laktationsfutter 14
Tabelle 2: Messzeitpunkte für die verschiedenen Untersuchungsparameter 15
Tabelle 3: Durchschnittliche Lebendmasse der Sauen am 110. Trächtigkeitstag
und durchschnittliche Wurfzahl
18
Tabelle 4: Jahreszeitliche Verteilung der Abferkelungen 19
Tabelle 5: Durchschnittlich geborene Ferkel je Wurf 19
Tabelle 6: Zusammensetzung des Trächtigkeitsfutters TF 60 20
Tabelle 7: Analysierte Inhaltsstoffe des Trächtigkeitsfutters TF 60 21
Tabelle 8: Zugabe und Gesamtgehalt an Tryptophan im Laktationsfutter 21
Tabelle 9: Sollgehalte im Laktationsfutter 23
Tabelle 10: Rohprotein- und Aminosäurengehalt ausgewählter Komponenten des
Laktationsfutters in g/kg FM
24
Tabelle 11: Zusammensetzung des Laktationsfutters in der Basisration 24
Tabelle 12: Tryptophan- und Glutaminsäurezulage (g/kg FM) in Abhängigkeit
der Futtergruppe
25
Tabelle 13: Gehalte an Rohnährstoffen, Stärke, Zucker und Aminosäuren im
Laktationsfutter
26
Tabelle 14: Verdaulichkeit der Rohnährstoffe und Gehalt an umsetzbarer
Energie
44
Tabelle 15: Scheinbare ileale Verdaulichkeit von Rohprotein und ausgewählten
Aminosäuren im Laktationsfutter I in %
45
Tabelle 16: Tägliche Futteraufnahme der Sauen in kg 46
Tabelle 17: Tägliche Proteinaufnahme der Sauen in g 47
Tabelle 18: Tägliche Tryptophanaufnahme je Sau in g 47
Tabelle 19: Tägliche Energieaufnahme der Sauen in MJ ME 48
Tabelle 20: Lebendmassen der Sauen am 110. Trächtigkeitstag sowie am 1. und
28. Laktationstag in kg
49
Tabelle 21: Masseverlust der Sauen während der Laktation 49
Tabelle 22: Tägliche Milchleistung der Sauen in g 5
Tabelle 23: Trockensubstanzgehalt in der Sauenmilch am 20./21. Laktationstag
in %
51
Tabelle 24: Fettgehalt der Sauenmilch in % 51
Tabelle 25: Fettausscheidung mit der Sauenmilch in g/Tag 51

5
Tabelle 26: Eiweißgehalt der Sauenmilch in % 52
Tabelle 27: Eiweißausscheidung mit der Sauenmilch in g/Tag 53
Tabelle 28: Laktosegehalt der Sauenmilch in % 53
Tabelle 29: Laktoseausscheidung mit der Milch in g/Tag 54
Tabelle 30: Harnstoffgehalt der Sauenmilch in mg/l 54
Tabelle 31: Harnstoffausscheidung mit der Sauenmilch in mg/Tag 54
Tabelle 32: Tryptophangehalt in der Sauenmilch in g/kg FS sowie g/16g N und
Tryptophanausscheidung in g/Tag
55
Tabelle 33: Gehalte an essentiellen Aminosäuren in der Milch der Gruppen I, III
und VI
56
Tabelle 34: Gehalte an nicht essentiellen Aminosäuren in der Milch der Gruppen
I, III und VI
56
Tabelle 35: Aminosäurenausscheidung mit der Milch in g/Tag 57
Tabelle 36: Energiegehalt der Sauenmilch in MJ/kg FS 58
Tabelle 37: Energieausscheidung mit der Sauenmilch in MJ/Tag 58
Tabelle 38: Gehalt an essentiellen und semiessentiellen Aminosäuren im Plasma
in mg/l
59
Tabelle 39: Gehalt an nicht-essentiellen Aminosäuren im Plasma in mg/l 60
Tabelle 40: Harnstoffgehalt im Plasma in mg/l 61
Tabelle 41: Hämatokritwert in % und Veränderung in % 61
Tabelle 42: Thrombozyten in 109/l und Veränderung in % 62
Tabelle 43: Serotonin im Serum in ng/ml und Veränderung in % 63
Tabelle 44: Serotoningehalt im Vollblut in ng/ml und Veränderung in % 63
Tabelle 45: Serotoningehalte in ng je 109 Thrombozyten und Veränderung in % 64
Tabelle 46: Saugferkelbeifutterverbrauch je Wurf in g 65
Tabelle 47: Lebendmassen der Ferkel in g 65
Tabelle 48: Lebendmassezunahmen der Ferkel in g/Tag 66
Tabelle 49: Grundsätzliche Parameter zur Ermittlung des Tryptophanbedarfs
laktierender Zuchtsauen
67
Tabelle 50: Aminosäuretransportsysteme (LÖFFLER und PETRIDES, 1997) 77
Tabelle 51: ATP-Äquivalente beim Abbau ausgewählter Verbindungen (nach:
LÖFFLER und PETRIDES, 1997; verändert und ergänzt)
78
Tabelle 52: Literaturangaben zur ilealen Verdaulichkeit von Tryptophan beim
Schwein
85

6
Tabelle 53: Gesamtgehalt und scheinbar ileal verdauliches Tryptophan im Futter
in % im Vergleich von Kalkulations- und Untersuchungsergebnissen
86
Tabelle 54: Einflussfaktoren auf den Futterverzehr von Sauen während der
Laktation (Auswahl; Literaturübersicht)
87
Tabelle 55: Einflussfaktoren auf die Milchleistung von Sauen (Auswahl;
Literaturübersicht)
95
Tabelle 56: Literaturübersicht der Zwischensäugeintervalle bei Sauen 96
Tabelle 57: Korrektur für Masseverluste der Ferkel während der Säugung in der
Literatur
98
Tabelle 58: Literaturangaben zum Trockensubstanzgehalt der Sauenmilch 101
Tabelle 59: Neuere Literaturangaben zum Fettgehalt in der Sauenmilch 102
Tabelle 60: Neuere Literaturangaben zum Proteingehalt in der Sauenmilch 103
Tabelle 61: Literaturangaben zum Laktosegehalt in der Sauenmilch 104
Tabelle 62: Literaturangaben zum Harnstoffgehalt in der Sauenmilch 106
Tabelle 63: Neuere Literaturangaben zum Energiegehalt in der Sauenmilch 108
Tabelle 64: Literaturangaben zum Aminosäureverhältnis in g AS/100 g Lysin bei
Sauenmilch
110
Tabelle 65: Tryptophangehalt, Standardabweichung absolut und Standard-
abweichung prozentual im Sauenplasma
113
Tabelle 66: Korrelationen zwischen Milchleistung der Sau und Ferkelzunahmen
(neuere Untersuchungen; Auswahl)
122
Tabelle 67: Untersuchungsparameter und resultierende Bedarfswerte in
Abhängigkeit des angewandten Regressionsmodells
126
Tabelle 68: Rationszusammensetzung und Gehalte an scheinbar ileal
verdaulichem Tryptophan in Praxisrationen (alle Angaben in % bzw.
Euro)
131

7
Abbildungsverzeichnis Abbildung 1: Skizze einer Abferkelbucht 17
Abbildung 2: Schematische Darstellung einer End-zu-End Ileorectal-
Anastomose (a = Jejunum, b = Ileum mit Ostium ileocaecale, c =
Caecum, d = Colon, e = Rectum)
28
Abbildung 3: Typisches HPLC-Chromatogramm mit den Peaks von Tryptophan
(Mitte) und internem Standard (rechts)
36
Abbildung 4: Beispielsgraph einer Broken-Line Regressionsgleichung 70
Abbildung 5: Beispielsgraph einer Reading Regressionsgleichung 71
Abbildung 6: Beispielsgraph einer quadratischen Regressionsgleichung 72
Abbildung 7: Beispielsgraph einer asymptotischen Regressionsgleichung 73
Abbildung 8: Strukturformeln von D- und L-Tryptophan 74
Abbildung 9: Stoffwechsel des Tryptophan (nach: VOET und VOET, 1994;
verändert und ergänzt)
76
Abbildung 10: Allgemeines Schema des Aminosäuren-Katabolismus in
Säugetieren (NELSON und COX, 2001)
79
Abbildung 11: Entstehung von Serotonin aus Tryptophan (VOET und VOET,
1994; verändert)
80
Abbildung 12: Schema der fütterungsbedingten Serotoninbeeinflussung im
Gehirn (nach FERNSTROM und WURTMAN, 1972; verändert
und ergänzt)
83
Abbildung 13: Biosynthese von Nicotinsäure und �amid aus Tryptopan
(HARPER et al., 1987)
84
Abbildung 14: Biosynthese von NAD aus Nicotinsäure (*beim NADP
phosphoryliert) (HARPER et al., 1987)
84
Abbildung 15: Mittlere tägliche Futteraufnahme der Sauen während der
Laktation
88
Abbildung 16: Ableitung des Tryptophanbedarfs anhand der Futteraufnahme der
Sauen in der 4. LW nach dem quadratischen Modell
89
Abbildung 17: Masseverlust der Sauen während der gesamten Laktation in g/Tag 91
Abbildung 18: Ableitung des Tryptophanbedarfs anhand der Lebendmasse-
verluste der Sauen während der gesamten Laktationsdauer nach
dem asymptotischen Modell
92

8
Abbildung 19: Ableitung des Tryptophanbedarfs anhand der Lebendmasse-
verluste der Sauen während der gesamten Laktationsdauer nach
dem Reading-Modell
93
Abbildung 20: Ableitung des Tryptophanbedarfs anhand der Lebendmasse-
verluste der Sauen während der gesamten Laktationsdauer nach
dem Broken-Line-Modell
93
Abbildung 21: Durchschnittliche tägliche Milchleistung der Sauen über die
gesamte Laktation
99
Abbildung 22: Ableitung des Tryptophanbedarfs anhand der Milchleistung der
Sauen während der gesamten Laktationsdauer nach dem
quadratischen Modell
100
Abbildung 23: Mittlerer Harnstoffgehalt der Sauenmilch 106
Abbildung 24: Ableitung des Tryptophanbedarfs anhand der Harnstoffgehalte in
der Sauenmilch am 20./21. LT nach dem quadratischen Modell
107
Abbildung 25: Tryptophangehalt im Sauenplasma am 22. Laktationstag 112
Abbildung 26: Harnstoffgehalt im Plasma der Sauen am 22. Laktationstag 116
Abbildung 27: Ableitung des Tryptophanbedarfs anhand des Harnstoffgehalts im
Sauenplasma am 22. Laktationstag nach dem quadratischen
Modell
118
Abbildung 28: Mittlere Konzentrationsabnahme beim Serotoningehalt im
Sauenplasma vom 85. TT auf den 28. LT
121
Abbildung 29: Mittlere Lebendmassezunahmen der Ferkel während der
Laktation
123
Abbildung 30: Saugferkelbeifutterverbrauch je Wurf während der 4.
Laktationswoche
124
Abbildung 31: Ableitung des Tryptophanbedarfs anhand der Futteraufnahme der
Sauen in der 4. LW (ohne Gruppe VI) nach dem asymptotischen
Modell
128
Abbildung 32: Ableitung des Tryptophanbedarfs anhand der Milchleistung der
Sauen während der gesamten Laktation (ohne Gruppe VI) nach
dem asymptotischen Modell
128

9
Abkürzungsverzeichnis Abw. Abweichung
Ala Alanin
Arg Arginin
AR-T atrophische Rhinitis
AS Aminosäure
Asn Asparagin
Asp Asparaginsäure
ATP Adenosintriphosphat
berechn. berechnet
BFS bakteriell fermentierbare Substanz
BHT Butylated Hydroxytoluene (Antioxidans)
C Kohlenstoff
Ca Kalzium
Cl Chlorid
CoA Coenzym A
CP crude protein
Cys Cystin
DLG Deutsche Landwirtschaftsgesellschaft
DXF verdauliche Rohfaser
DXL verdauliches Rohfett
DXP verdauliches Rohprotein
DXX verdauliche Stickstoff-freie Extraktionsstoffe
ed. editor(s)
essent. essentiell
e.V. eingetragener Verein
Fa. Firma
FFA Free Fatty Acids
FM Frischmasse
FS Frischsubstanz
g Gramm
g Erdbeschleunigung
Geh. Gehalt
ges. gesamt

10
GfE Gesellschaft für Ernährungsphysiologie
Gln Glutamin
Glu Glutaminsäure
Gly Glycin
H Wasserstoff
His Histidin
HPLC High Performance Liquid Cromatography
Hrsg. Herausgeber
Hyp Hydroxyprolin
I Iod
I.E. Internationale Einheiten
Ile Isoleucin
K Kalium
Kartoff.eiweiß Kartoffeleiweiß
kg Kilogramm
l Liter
LA Langzeitantibiotikum
Lakt. Laktation
Leu Leucin
LF Laktationsfutter
LM Lebendmasse
LM0,75 metabolische Lebendmasse
LNAA Large Neutral Amino Acids
LT Laktationstag
LW Laktationswoche
Lys Lysin
M Mol
ME umsetzbare Energie
Met Methionin
Mg Magnesium
mg Miligramm
min Minuten
min. mindestens
Mittelw. Mittelwert

11
MJ Megajoul
ml Milliliter
n Stichprobenumfang
N Stickstoff
Na Natrium
NAD Nicotinamid-Adenin-Dinukleotid
NADP Nicotinamid-Adenin-Dinukleotid-Phosphat
NfE Stickstoff-freie Extraktstoffe
ng Nanogramm
NRC National Research Council
O Sauerstoff
OM organische Masse
OR organischer Rest
P Phosphat
p Irrtumswahrscheinlichkeit
Phe Phenylalanin
plt platelets
PPi pyrophosphate
PPriboseP 5-phosphoribosyl-1-pyrophosphate (enzyme)
Pro Prolin
r Korrelationskoeffizient
r2 Bestimmtheitsmaß
RIA Radio Immuno Assay
RNA Ribonukleinsäure
RP Reversed Phase
S Standardfehler
sch. scheinbar
Ser Serin
Sojaex.schot Sojaextraktionsschrot
St.abw. Standardabweichung
TF Trächtigkeitsfutter
Thr Threonin
Trockenschn. Trockenschnitzel
Trp Tryptophan

12
Trp. Tryptophan
TS Trockensubstanz
TT Trächtigkeitstag
Tyr Tyrosin
TZ Thrombozyten
unters. untersucht
Val Valin
verd. verdaulich
W0,75 Metabolische Körpermasse
XF Rohfaser
XL Rohfett
XP Rohprotein
XS Stärke
XZ Zucker
µg Mikrogramm
µl Mikroliter
µm Mikrometer
°C Grad Celsius
% Prozent

13
1. Einleitung und Problemstellung
Die Forschung und die großtechnische Produktion von Aminosäuren haben in jüngerer Zeit bei der
Zusammenstellung von Futterrationen für Schweine auch in der Praxis den Übergang von der reinen
Rohproteinbetrachtung zu einer gezielten Aminosäurenbetrachtung ermöglicht. Hierbei kommt den
Aminosäuren Lysin, Methionin/Cystin, Threonin und Tryptophan in den heute üblicherweise
verfütterten Rationen auf Getreide-Soja-Basis eine besondere Bedeutung zu, da sie in der Regel als
erste leistungsbegrenzend sind. Der stetige züchterische Fortschritt sowie die gestiegenen
Anforderungen an die Wurfgröße und die daraus resultierende Forderung nach hohen Milchleistungen
auch bei der Zuchtsau machen eine Überprüfung der Versorgungsempfehlungen für essentielle
Aminosäuren von Zeit zu Zeit notwendig. So wurden die Empfehlungen zum Lysingehalt im Futter
laktierender Sauen innerhalb von 10 Jahren von 0,6 % (NRC, 1988) durch neuere Untersuchungen
(z.B.: SAUBER et al., 1994; KNABE et al., 1996) auf 1,0 % angehoben (NRC, 1998). Für die
Aminosäuren Methionin/Cystin (SCHNEIDER et al., 1992; 1992B; SCHWARZ et al., 1993) und
Threonin (WESTERMEIER et al., 1998; PAULICKS et al., 1998; 1998B) wurden ebenfalls in neuerer
Zeit genaue Studien zum Bedarf bei laktierenden Zuchtsauen durchgeführt. Zum Tryptophanbedarf der
säugenden Sau liegen dagegen bis jetzt keine umfassenden Untersuchungen unter Berücksichtigung
dieser neuen Ergebnisse vor. Die aktuellen Empfehlungen für die Tryptophankonzentration im
Laktationsfutter liegen je nach erwartetem Lebendmasseverlust der Zuchtsau während der Laktation
und den erwarteten täglichen Ferkelzunahmen zwischen 0,14 % und 0,19 % (NRC, 1998). Diese
Empfehlungen basieren auf Untersuchungen von LEWIS und SPEER (1974) sowie MENNENGA und
EASTER (1985). Als Parameter für die Bedarfsherleitung haben die Autoren Gewichtsveränderungen
von Sauen und Ferkeln (beide) sowie die Stickstoffbilanz und Plasmametaboliten (LEWIS und
SPEER, 1974) herangezogen. Wichtige Parameter wie Milchmenge und �zusammensetzung wurden
dabei nicht berücksichtigt. Bei der GfE (1987) sind keine Empfehlungen zum Tryptophanbedarf
säugender Sauen aufgeführt.
Mit der vorliegenden Arbeit sollte daher der Einfluss eines weit variierenden Tryptophangehalts von
0,12 % bis 0,42 % im Futter auf Leistungs- und Stoffwechselparameter untersucht werden. Besonderes
Augenmerk wurde dabei auf die Milchmenge und die Untersuchung der Milchinhaltsstoffe gelegt, da
diese in der Ferkelerzeugung leistungsbestimmend sind und Bedarfsableitungen genaugenommen nur
für den Parameter gelten, für den sie gewonnen wurden.

14
Grundsätzlich gibt es drei verschiedene Methoden zur Ableitung des Aminosäurenbedarfs:
- Variation der Anteile verschiedener nativer Proteinträger mit Ermittlung des Aminosäuren-
musters in der `besten´ Ration
- Grundration mit variierenden Zulagen an einer oder mehreren synthetischen Aminosäuren und
Bestimmung der `günstigsten´ Zulage
- ausschließliche Verwendung synthetischer Eiweißträger mit Variation bei einer oder mehreren
Aminosäuren und Aufzeigen der `effektivsten´ Zusammenstellung
Im vorliegenden Versuch wurde die zweite Methode gewählt. Es wurde ausschließlich die Aminosäure
Tryptophan variiert. Die Stufen des Tryptophangehalts im Futter wurden in sechs Schritten zwischen
0,12 % und 0,42 % gewählt, um einen weiten Versorgungsbereich vom vermutlichen Mangel bis zum
Überschuss abzudecken. Aus Tabelle 1 sind die verschiedenen Tryptophanzulagen und �gesamtgehalte
im Futter zu entnehmen.
Tabelle 1: Zulage und Gesamtgehalt an Tryptophan im Laktationsfutter
Gruppe I II III IV V VI Tryptophanzulage (%) 0,00 0,03 0,06 0,12 0,18 0,30 Tryptophan ges. (%) 0,12 0,15 0,18 0,24 0,30 0,42
Die weiteren Nährstoffgehalte wurden konstant gehalten, um ceteris paribus Bedingungen zu schaffen.
Pro Behandlung waren 12 Wiederholungen vorgesehen. Als Versuchsparameter wurden folgende
Daten erhoben:
• Lebendmasse der Sauen
• Futteraufnahme der Sauen
• Milchleistung der Sauen
• Fett-, Eiweiß-, Laktose-, Harnstoff- und Aminosäurekonzentration der Sauenmilch
• Harnstoff- und Aminosäurenkonzentration im Blutplasma der Sauen
• Serotoninkonzentration im Blutserum der Sauen
• Lebendmasse der Ferkel
• Beifutteraufnahme der Ferkel

15
Die Messzeitpunkte der untersuchten Parameter sind aus Tabelle 2 ersichtlich.
Tabelle 2: Messzeitpunkte für die verschiedenen Untersuchungsparameter
Parameter Trächtig-keitstag Laktationstag
60. 85. 110. 1. 13. 14. 20. 21. 22. 28. Masse der Sauen • • • • Futteraufnahme der Sauen täglich Milchleistung der Sauen • • • • Milchprobenentnahme • • • • Blutentnahme bei den Sauen • • • Masse der Ferkel • • • • • • Einwaage Ferkelbeifutter • Rückwaage Ferkelbeifutter •

16
2. Material und Methoden
2.1. Versuchsbetrieb Der Versuch wurde von Oktober 2001 bis September 2002 in der Versuchsstation für Futterbau und
Tierernährung, Versuchsgut Hirschau der TU-München, Freising-Weihenstephan durchgeführt. Auf
dem Betrieb wurde unter anderem eine Herde Zuchtsauen mit Ferkelaufzucht und angeschlossener
Jungsauenproduktion gehalten.
2.1.1. Tiermaterial
Zur Durchführung der Untersuchungen standen etwa 70 Herdbuchsauen der Deutschen Landrasse zur
Verfügung. Für den Versuch wurden nur Sauen herangezogen, die mindestens in der zweiten Laktation
waren. Bei deren künstlicher Besamung wurde ausschließlich Sperma von Ebern der Deutschen
Landrasse verwendet. Das Sperma wurde von der Schweineprüf- und Besamungsstation Schwaben
und Oberbayern e.V. in Bergheim bezogen und die Besamung durch Mitarbeiter des Versuchsgutes
durchgeführt. Insgesamt wurden 72 Laktationsperioden von 61 Sauen ausgewertet.
2.1.2. Aufstallung der Tiere
Während der Güstzeit wurden die Sauen in Einzellaufbuchten mit Teilspaltenboden und direktem
Eberkontakt gehalten. Die Haltung während der Trächtigkeit erfolgte in Kastenständen mit
planbefestigtem Boden und leichter Häckselstroheinstreu. Fünf Tage vor dem Geburtstermin wurden
die Sauen in eine Abferkelbucht umgestallt. Hier diente ebenfalls Häckselstroh als Einstreu auf dem
planbefestigten Boden. Aus der Skizze in Abbildung 1 sind die weiteren Details ersichtlich.
Das Ferkelnest war mit einer Heizliegeplatte sowie mit einem Infrarotstrahler ausgestattet, wodurch
die für neugeborene Ferkel notwendige Temperatur von 35°C erreicht wurde. Im Wartebereich wurde
eine Temperatur von ca. 18 � 22°C, im Abferkelstall von ca. 20 - 25°C gemessen. Vor jeder Belegung
wurden die Abferkelbuchten gereinigt und mit Orbivet (Fa. Schülke & Mayr, Norderstedt) desinfiziert.

17
Abbildung 1: Skizze einer Abferkelbucht
2.1.3. Fütterung der Tiere
Durch die permanente Einzeltierhaltung und abgetrennte Futtertröge konnte eine gezielte individuelle
Fütterung der Sauen von Hand durchgeführt werden. Die Futtervorlage bei den tragenden Sauen
erfolgte an Wochentagen etwa um 7 und um 15 Uhr. Die Sauen in den Abferkelbuchten wurden um 6
und um 14 Uhr gefüttert. Bei anstehenden Blutprobennahmen wurden die jeweiligen Sauen
zeitversetzt gefüttert (siehe: 2.6.3.). An Sonn- und Feiertagen erfolgte bei allen Sauen nur eine
einmalige Fütterung zwischen sechs und sieben Uhr. Den Ferkeln wurde am Morgen des 22.
Lebenstages 1500 g Prestarter in den Futterautomat eingewogen und bei Bedarf während der Tage bis
zum Absetzen nachgefüllt. In allen Stallbereichen stand den Sauen über Nippeltränken Wasser zur
freien Aufnahme zur Verfügung, ebenso den Ferkeln.
2.1.4. Behandlung der Tiere
Die Versuchssauen wurden während der Güstzeit mit 2 ml Erysorb Parvo (Fa. Intervet, Boxmeer,
Niederlande) gegen Rotlauf und Parvovirose geimpft. In der achten Trächtigkeitswoche wurden die
Sauen mit Flubenol (Fa. Janssen, Neuss) gegen Endoparasiten behandelt. Jeweils acht und sechs
Wochen vor der Geburt erfolgte eine gleichzeitige Immunisierung der Sauen mit 2 ml Gletvax plus
(Fa. Essex, München) gegen Escherichia-Coli und Ferkelenteritis sowie mit 2 ml Porcilis AR-T (Fa.
Intervet, Boxmeer, Niederlande) gegen die progressive Rhinitis atropicans. Vor dem Umstallen in die
Abferkelbucht wurden die Sauen gewaschen und mit Sebacil (Fa. Bayer, Monheim) gegen
Ektoparasiten behandelt. Bei Wehenschwäche wurden 3 ml Depotocin (Fa. Veyx, Schwarzenborn) zur

18
Geburtserleichterung gespritzt. Beim Auftreten von leichtem Fieber beziehungsweise Vaginalausfluss
nach der Geburt wurden 20 ml Frieso-Gent (Wirkstoff: Gentamicinsulfat; Fa. Essex, München)
injiziert. Hatten Sauen schwerwiegendere Krankheiten, wurden sie aus dem Versuch genommen.
Direkt nach der Geburt wurde den Ferkeln der Schwanz kupiert und sie wurden im Ohr zur
eindeutigen Identifikation tätowiert. Am dritten Lebenstag erhielten sie 2 ml Eisendextran (Fa.
AniMedica, Senden-Bösensell) sowie 0,5 ml Terramycin/LA (Wirkstoff: Oxytetracyclin-Dihydrat; Fa.
Pfizer, Karlsruhe) zur Stärkung der Vitalität; männliche Ferkel wurden kastriert. Die Impfung gegen
chronische Pneumonien verursacht durch Mycoplasma Hyopneumoniae erfolgte am 23. Tag nach der
Geburt mit 2 ml Stellamune Mycoplasma (Fa. Pfizer, Karlsruhe). Gleichzeitig erhielten die Ferkel ihre
Ohrmarke, und die Stammnummer wurde ins Ohr tätowiert. Gelenkentzündungen und
Hautabschürfungen wurden mit 0,5 ml Tardomyocel (Wirkstoff: Penicillin-Streptomycin-
Kombination; Fa. Bayer, Monheim) behandelt. Bei leichtem Durchfall der Ferkel wurden 0,5 ml
Baytril 5% (Wirkstoff: Enrofloxacin; Fa. Bayer, Monheim) verabreicht. Schwerwiegend erkrankte
Ferkel schieden aus dem Versuch aus.
Weder bei den Sauen noch bei den Ferkeln war ein Einfluss der Futtergruppe auf die Häufigkeit einer
Behandlung mit Medikamenten vorhanden.
2.2. Minimierung systematischer Einflussgrößen
2.2.1. Lebendmasse, Alter und Genetik
Damit der Effekt der verschiedenen Futtergruppen nicht durch systematische Einflüsse überlagert
wird, wurde versucht, diese Einflüsse gleichmäßig über alle Versuchsgruppen zu verteilen.
Um den Einfluss des Alters der Sauen auszuschalten, wurde auf eine annähernd gleichmäßige
Aufteilung der Wurfnummern über die Gruppen geachtet. Ebenso sollte bei den Lebendmassen der
Sauen eine gleichmäßige Verteilung vorliegen. Die einzelnen Werte sind in Tabelle 3 dargestellt.
Tabelle 3: Durchschnittliche Lebendmasse der Sauen am 110. Trächtigkeitstag und
durchschnittliche Wurfzahl
Gruppe I II III IV V VI Trp.-Geh. in % 0,12 0,15 0,18 0,24 0,30 0,42 Mittel Lebendmasse in kg 260 254 259 262 256 266 260
±27 ±26 ±24 ±31 ±33 ±30 ±29 Wurfzahl 4,4 3,4 3,4 3,8 3,9 4,4 3,9
±1,7 ±1,6 ±1,5 ±2,0 ±2,1 ±v2,6 ±1,9
19
Alle Sauen wurden aus eigener Aufzucht in den Bestand genommen, wodurch sich Auswirkungen
unterschiedlicher Herkünfte vermeiden ließen. Bei den Besamungen wurde auf eine gleichmäßige
Verteilung der Eber geachtet, um genetische Einflüsse auf die Leistung der Ferkel weitgehend
ausschließen zu können.
2.2.2. Jahreszeit
Um den Effekt jahreszeitlicher Schwankungen bei den Leistungen auszugleichen, wurden die
Abferkelungen jeder Gruppe über das ganze Jahr verteilt, wie aus Tabelle 4 ersichtlich wird.
Tabelle 4: Jahreszeitliche Verteilung der Abferkelungen
Gruppe I II III IV V VI Trp.-Geh. in % 0,12 0,15 0,18 0,24 0,30 0,42 Mittel Okt. - Dez. 3 2 1 3 3 2 14 Jan. - März 4 4 3 3 2 3 19 Apr. - Juni 3 3 4 3 5 4 22 Juli - Sep. 2 3 4 3 2 3 17 ges. 12 12 12 12 12 12 72
2.2.3. Ferkelzahl je Wurf
Die Milchleistung der Sau wird auch durch die Anzahl der Ferkel beeinflusst (ELSLEY, 1971; VAN
DER STEEN, 1985; KING et al., 1989; AULDIST et al., 1994). Aus diesem Grund wurde die
Wurfgröße je Sau auf zehn bis zwölf Ferkel standardisiert. Dazu mussten direkt nach der Geburt
überzählige Ferkel weggenommen beziehungsweise fehlende durch gleich alte ersetzt werden. Es
wurden je Wurf jedoch nur maximal drei Ferkel ausgetauscht. Bis zum zwölften Laktationstag konnten
schwer erkrankte oder verendete Ferkel durch gleichwertige ersetzt werden. Traten danach
Krankheiten oder Ausfälle der Ferkel auf, so wurde die Sau aus dem Versuch genommen. In Tabelle 5
sind die durchschnittlichen Wurfgrößen der verschiedenen Behandlungsgruppen eingetragen.
Tabelle 5: Durchschnittlich geborene Ferkel je Wurf
Gruppe I II III IV V VI Trp.-Geh. in % 0,12 0,15 0,18 0,24 0,30 0,42 Mittel Ferkelzahl je Wurf 10,25 10,25 10,33 10,17 10,42 10,50 10,32
±0,62 ±0,45 ±0,49 ±0,39 ±0,67 ±0,80 ±0,57

20
2.3. Fütterung
2.3.1. Trächtigkeitsfütterung
Bis zum 59. Trächtigkeitstag wurden die Sauen kombiniert gefüttert. Die Ration setzte sich aus 1,5 kg
eines getreidereichen Ergänzungsfutters (14 % XP, 4 % XF, 15,7 MJ ME/kg FS) und etwa 4 kg
Maissilage zusammen. Ab dem Beginn des Versuchszeitraumes am 60. Trächtigkeitstag bekamen alle
Versuchstiere ein Alleinfutter für tragende Sauen (TF 60).
Tabelle 6: Zusammensetzung des Trächtigkeitsfutters TF 60
Komponente
Anteil in %
Gerste
Trockenschnitzel
Haferschälkleie
Sojaextraktionsschrot
Pflanzenöl
vitaminiertes Mineralfutter1)
Aminosäurenvormischung2)
Vitaminvormischung3)
61,0
18,0
14,0
2,0
1,0
2,0
1,0
1,01)Zusammensetzung nach Herstellerangaben, siehe Anhangstabelle 1 2)Zusammensetzung siehe Anhangstabelle 4 3)Zusammensetzung siehe Anhangstabelle 5
Damit wurden Schwankungen bei der Nährstoffversorgung, wie sie bei der kombinierten Fütterung
verstärkt auftreten können, vermieden. In Tabelle 6 sind die Anteile der verschiedenen
Futterkomponenten des Trächtigkeitsfutters, in Tabelle 7 die analysierten Inhaltsstoffe eingetragen.
Vom 60. bis zum 85. TT bekamen die Sauen 2,4 kg und bis zum 100. TT 2,8 kg Futter je Tag. Bei
einem Energiegehalt von 11,5 MJ/kg FS war damit eine tägliche Energieversorgung von 28
beziehungsweise 32 MJ ME je Sau sichergestellt. Die Empfehlungen für die tägliche
Energieversorgung von trächtigen Sauen, ausgesprochen von der GESELLSCHAFT FÜR
ERNÄHRUNGSPHYSIOLOGIE (1987) sowie der DEUTSCHE(n) LANDWIRTSCHAFTS-
GESELLSCHAFT (1996), wurden somit eingehalten.

21
Tabelle 7: Analysierte Inhaltsstoffe des Trächtigkeitsfutters TF 60
Inhaltsstoffe
Anteil in %
Trockenmasse
Rohasche
Rohprotein
Rohfaser
Rohfett
Stärke
Zucker
Lysin
Tryptophan
Energie (in MJ ME/kg FM)1)
90,4
6,0
9,5
9,6
2,3
31,6
6,1
0,68
0,12
11,51)berechnet nach KIRCHGESSNER und ROTH (1983)
2.3.2. Laktationsfütterung
Ab dem 111. Trächtigkeitstag wurde den Sauen ein Laktationsfutter verabreicht, das sich zwischen den
Behandlungsgruppen nur im Tryptophangehalt unterschied. Ausgehend von der Basisration wurden für
die verschiedenen Gruppen ansteigende Mengen an L-Tryptophan zugesetzt. Insgesamt ergaben sich
sechs Futtergruppen. Aus Tabelle 8 sind die unterschiedlichen Zugaben und Gehalte an Tryptophan in
den Futtergruppen ersichtlich.
Tabelle 8: Zugabe und Gesamtgehalt an Tryptophan im Laktationsfutter
Gruppe I II III IV V VI Tryptophanzugabe (%) 0,00 0,03 0,06 0,12 0,18 0,30 Tryptophangehalt (%) 0,12 0,15 0,18 0,24 0,30 0,42 ileal verd. Trp (%) 0,09 0,12 0,15 0,21 0,27 0,39
Bei den gängigen Modellen zur Bedarfsableitung (siehe 4.1.) sind mindestens 2 Gruppen unter und
über dem aktuellen Bedarf erforderlich. Dies ist sowohl beim absoluten Gehalt im Futter als auch beim
Gehalt an ileal verdaulichem Tryptophan zu berücksichtigen. Die aktuelle Empfehlung liegt für
laktierende Sauen mit 10 Ferkeln und einem tolerierten Masseverlust während der Säugezeit von 10 kg
bei 0,17 % gesamt oder 0,14 % ileal verdaulichem Tryptophan im Futter (NRC, 1998). Das Futter der
Basisgruppe enthielt 0,12 % Tryptophan, da zum einen die Gefahr bestand, dass bei noch niedrigeren

22
Gehalten die Futteraufnahme durch die Sauen unter ein physiologisch vertretbares Niveau abfällt
(HENRY et al., 1992). MEISINGER und SPEER berichten 1979 sogar von einer totalen
Futteraufnahmeverweigerung bei Sauen, denen eine Ration mit starkem Tryptophanmangel vorgelegt
wurde. Zum anderen war es unter annähernd praktischen Bedingungen nicht möglich, einen
niedrigeren Tryptophanwert im Futter zu erreichen. Die Gruppe III kam der aktuellen Empfehlung sehr
nahe (Die Gehalte wurden um 0,01 % erhöht, um aufgrund der geringen Differenz zwischen 0,12 %
und 0,17 % eine etwas größere Abstufung zu erreichen, bei der Leistungsunterschiede besser
hervortreten.). Die Gruppe II lag bei beiden Gehalten dazwischen. Für die Gruppen IV und V wurde
die Abstufung zwischen den Gruppen von 0,03 % auf 0,06 % verdoppelt. Im überhöhten Bereich
wurde die Spreizung noch einmal auf 0,12 % vergrößert, um eventuelle Effekte einer sehr hohen
Dosierung noch erfassen zu können.
Alle anderen Nährstoffe wurden bei der Rationsberechnung bedarfsdeckend kalkuliert. Bei den drei
erstlimitierenden Aminosäuren und Valin wurde ein Sicherheitszuschlag von 10 % gegeben, da für
diese Aminosäuren neuere Untersuchungen vorlagen. Der Lysingehalt von 10 g/kg FM orientierte sich
an den Empfehlungen des NRC (1998) für laktierende Sauen. Der von SCHNEIDER (1992) ermittelte
Methionin- und Cystinbedarf von 6,5 g/kg FM wurde ebenso berücksichtigt, wie der von
WESTERMEIER (1997) empfohlene Threoninbedarf von 7 g/kg FM. Für Valin wurden die
Ergebnisse von OTT (2000) mit einem Bedarf von 6,5 g/kg FM mit einbezogen. Bei allen anderen
Aminosäuren wurde ein Sicherheitszuschlag von 20 % festgelegt, um limitierende Wirkungen anderer
Aminosäuren sicher ausschließen zu können. Die Gehalte an den restlichen Aminosäuren wurden
anhand der Relationen des `idealen Protein´ zu Lysin nach WANG und FULLER (1989) festgelegt.
Damit die Stickstoffzufuhr insgesamt nicht zu einem begrenzenden Faktor im Versuch wird, wurde ein
Verhältnis der essentiellen zu den nicht essentiellen Aminosäuren von 1 : 1,2 bis 1 : 1,3 angestrebt, das
nach WANG und FULLER (1989) beziehungsweise GOTTERBARM et al. (1997) im optimalen
Bereich liegt.
Bei der Festlegung des Energiegehaltes des Laktationsfutters wurde ein Wert von mindestens 13 MJ
ME/kg FM angestrebt. Damit sollte einer starken energetischen Unterversorgung vorgebeugt werden,
wenn die Futteraufnahme relativ gering ist. Nach TROTTIER und EASTER (1995) kann dies vor
allem in den ersten Tagen nach der Geburt der Fall sein.
Die Gehalte an Mengen- und Spurenelementen sowie an Vitaminen richteten sich nach den
Empfehlungen der GESELLSCHAFT FÜR ERNÄHRUNGSPHYSIOLOGIE (1987). In Tabelle 9 sind
die Sollgehalte im Laktationsfutter dargestellt.

23
Tabelle 9: Sollgehalte im Laktationsfutter
Inhaltsstoff
Sollgehalt
Einheit je kg FM
Rohprotein
Rohfett
Rohfaser
Stärke
Lysin
Methionin + Cystin
Threonin
Phenylalanin + Tyrosin
Leucin
Isoleucin
Histidin
Valin
Calcium
Phosphor
Natrium
Zink
Vitamin A
Vitamin D
Energie
min.
max.
max.
min.
min.
min.
min.
min.
min.
min
min.
160
80
70
330
9,9
7,2
7,7
13,0
11,9
6,5
4,1
7,2
8,0
6,0
2,5
50
5000
625
13,0
g
g
g
g
g
g
g
g
g
g
g
g
g
g
g
mg
I.E.
I.E.
MJ ME
Das Laktationsfutter sollte eine möglichst praxisrelevante Zusammensetzung haben und dennoch im
Tryptophangehalt nicht höher als 0,12 % liegen. Um dies zu erreichen, wurden Komponenten
verwendet, die von Haus aus wenig Tryptophan im Verhältnis zum Rohproteingehalt haben. Dafür
waren besonders Maiskleber, Mais, Trockenschnitzel und Erbsen geeignet, die auch unter
Praxisbedingungen eingesetzt werden. Zusätzlich wurden Kartoffeleiweiß und Sojaextraktionsschrot
eingemischt. Die oben genannten Komponenten wurden vor der Verwendung auf ihren Rohprotein-,
Lysin-, Methionin-, Cystin-, Threonin- und Tryptophangehalt untersucht. Die Tabelle 10 zeigt die
Resultate.
Ausgehend von diesen Ergebnissen und den Anforderungen an das Laktationsfutter, wurde mit Hilfe
des linearen Optimierungsprogrammes `Single Mix´ (Format Inc., 1990) die Rationsberechnung
durchgeführt.

24
Tabelle 10: Rohprotein- und Aminosäurengehalt ausgewählter Komponenten des
Laktationsfutters in g/kg FM
Komponente
Rohprotein
Lysin
Methionin
Cystin
Threonin
Tryptophan
Maiskleber
Mais
Trockenschn.
Erbsen
Kartoff.eiweiß
Sojaex.schrot
192
87
87
227
689
426
8,0
2,7
4,5
16,7
49,0
27,9
3,0
2,0
1,2
2,1
14,2
5,6
4,0
1,8
0,6
2,8
7,3
5,6
7,2
3,3
2,9
8,1
39,0
17,0
0,92
0,64
0,79
2,00
10,30
5,96
Tabelle 11: Zusammensetzung des Laktationsfutters in der Basisration
Komponente
Anteil in %
Mais
Erbsen
Maiskleber
Trockenschnitzel
Kartoffeleiweiß
Sojaextraktionsschrot
Pflanzenöl
Cellulosepulver
Futterzucker
vitaminiertes Mineralfutter1)
kohlensaurer Kalk
Monocalciumphosphat
Viehsalz
Glutaminsäure
Aminosäurenmischung2)
Cholinchlorid
48,00
13,50
16,93
3,93
2,60
3,00
1,00
2,23
1,80
1,40
1,00
0,50
0,50
2,29
1,24
0,081)Zusammensetzung nach Herstellerangaben, siehe Anhangstabelle 1 2)Zusammensetzung siehe Anhangstabelle 6

25
Aufgrund der Restriktionen war es nicht möglich ein passendes Futter nur aus praxisüblichen
Komponenten zu erstellen. Daher wurden zur Abdeckung des Bedarfes zusätzlich essentielle
Aminosäuren eingesetzt. Um einen für den Verdauungsapparat ausreichenden Rohfaseranteil zu
erreichen, wurde Cellulosepulver eingemischt. Futterzucker fand wegen der Verbesserung der
Schmackhaftigkeit und wegen der Erhöhung der Energiekonzentration im Futter Anwendung. Die
Zusammensetzung der Basisration ist in Tabelle 11 dargestellt.
Die steigenden Zulagen an Tryptophan bei den Gruppen II � VI gingen zu Lasten des Anteils an
Glutaminsäure. Es wurde L-Tryptophan verwendet. Den Zusammenhang zeigt Tabelle 12.
Tabelle 12: Tryptophan- und Glutaminsäurezulage (g/kg FM) in Abhängigkeit der
Futtergruppe
Gruppe
I
II
III
IV
V
VI
L-Tryptophan
0,0
0,3
0,6
1,2
2,8
3,0
Glutaminsäure 22,9 22,6 22,3 21,7 20,1 19,9
Summe
22,9
22,9
22,9
22,9
22,9
22,9
Das Laktationsfutter wurde in der Versuchsanlage der Fachgebiete Tierernährung der TU München-
Weihenstephan in einem Präzisionsmischer (Modell: FKM 1200, Fa. Lödige, Paderborn) gemischt.
Die Analysenergebnisse der jeweiligen Futter sind in Tabelle 13 aufgeführt.
Bei den Rohnährstoff-, Stärke-, Zucker- und Aminosäurengehalten traten mit Ausnahme des
Tryptophangehaltes keine Unterschiede auf. Die erwünschten Abstufungen bei der Tryptophan-
konzentration wurden durch die Analyseergebnisse bestätigt.
Am 111. Trächtigkeitstag erhielten die Sauen täglich 4 kg Laktationsfutter. Vom 112. Trächtigkeitstag
an wurde die Futtermenge bis zum berechneten Abferkeltag täglich um 1 kg reduziert und am
Abferkeltag wurden die Sauen nicht gefüttert. Danach bekamen die Sauen ausgehend von einer
Anfangsmenge von 2 kg täglich etwa 0,75 kg mehr Futter, bis zur ad libitum Aufnahme.
26
Tabelle 13: Gehalte an Rohnährstoffen, Stärke, Zucker und Aminosäuren im
Laktationsfutter
berechnet analysiert Gruppe I I II III IV V VI Trp.-Geh. in % 0,12 0,12 0,15 0,18 0,24 0,30 0,42 Inhaltsstoffe g/kg FM
Trockenmasse 915 903 905 907 906 907 905
Rohasche 60 58 60 59 59 62 59
Rohprotein 160 156 158 160 154 156 156
Rohfett 39 32 34 35 34 35 34
Rohfaser 62 53 54 54 55 55 57
Stärke1) 398 411 411 411 411 411 411
Zucker1) 42 44 44 44 44 44 44
Tryptophan 1,2 1,2 1,5 1,8 2,4 2,9 4,0
Lysin 10,1 10,6 10,4 10,5 10,8 10,7 10,5
Methionin 4,9 5,4 5,3 5,1 5,6 5,9 5,7
Cystin 2,3 2,6 2,4 2,6 2,5 2,5 2,6
Methionin + Cystin 7,2 8,0 7,7 7,7 8,1 8,4 8,3
Threonin 7,7 6,9 7,1 7,1 7,4 7,3 7,8
Isoleucin 6,5 5,2 5,5 6,2 6,0 6,0 5,9
Histidin 4,1 3,6 3,7 3,8 3,9 3,8 3,8
Leucin 14,4 12,4 12,3 12,8 13,1 13,0 13,1
Phenylalanin 10,1 11,4 11,0 12,1 11,9 11,8 12,2
Tyrosin 5,6 4,8 4,7 5,0 5,1 4,7 5,0
Phenylalanin + Tyrosin 15,7 16,2 15,7 17,1 17,0 16,5 17,2
Valin 7,3 7,6 7,2 8,0 7,6 7,9 8,0 1)aliquote Mischprobe der Gruppen I - VI
2.3.2. Ferkelfütterung
Das Beifutter, das die Ferkel ab dem 22. Lebenstag zur freien Aufnahme erhielten, war ein pelletiertes
Fertigfutter der Fa. Deuka, Deutsche Kraftfutterwerke GmbH & Co, Düsseldorf. Dieses Futter war
speziell für die Verfütterung an junge Saugferkel konzipiert. Die Zusammensetzung, die Inhalts- und
Zusatzstoffe des Futters sind aus der Anhangstabelle 2 ersichtlich.

27
2.4. Verdauungsversuche
2.4.1. Ermittlung der Verdaulichkeit der Rohnährstoffe
Um die scheinbare Verdaulichkeit der Rohnährstoffe im Laktationsfutter zu bestimmen, wurde am
Fachgebiet Tierernährung des Departments für Tierwissenschaften der TU München, Freising-
Weihenstephan, ein Verdauungsversuch durchgeführt.
Dazu standen 8 kastrierte männliche Schweine einer Zwei-Rassen-Kreuzung, Piétrain x Deutsche
Landrasse, zur Verfügung. Im Versuch wurde die Methode der quantitativen Sammeltechnik in
Anlehnung an SCHIEMANN (1981) angewandt. Dabei wurden die Tiere in Stoffwechselkäfigen
gehalten, die eine quantitative Erfassung der verzehrten Futtermenge sowie der ausgeschiedenen
Kotmenge ermöglichten. Während des gesamten Versuches herrschte im vollklimatisierten Stall eine
konstante Temperatur von 20°C bei einer relativen Luftfeuchtigkeit von 60 %. Jeweils 4 Versuchstiere
erhielten das Laktationsfutter der Gruppe I beziehungsweise IV.
Der Versuch gliederte sich in eine 9-tägige Vorperiode und eine 8-tägige Sammelperiode. Die
durchschnittliche Lebendmasse der Tiere betrug zu Beginn der Sammelperiode bei Gruppe I 62,9 kg
und bei Gruppe IV 63,3 kg. Zu Beginn der Sammelperiode wurde das Futter für den gesamten
restlichen Versuchszeitraum eingewogen, die Trockensubstanz ermittelt und Proben für die
Rohnährstoffanalyse entnommen. Für die Futterzuteilung wurde ein täglicher Energiebedarf von 1,0
MJ ME/kg LM0,75 unterstellt. Bei einer rechnerischen Energiedichte im Laktationsfutter von 13,2 MJ
ME (berechnet nach GESELLSCHAFT FÜR ERNÄHRUNGSPHYSIOLOGIE, 1987) ergaben sich
somit Tagesfuttermengen je Schwein von 1600 bis 1800 Gramm. Damit wurden die Empehlungen der
GESELLSCHAFT FÜR ERNÄHRUNGSPHYSIOLOGIE (1987) zum Bedarf an umsetzbarer Energie
eingehalten. Wegen schlechter Futteraufnahme bei zwei Tieren der Gruppe I musste bei diesen die
tägliche Futtervorlage auf 1500 Gramm begrenzt werden.
Die Fütterung erfolgte täglich um 7.00 und um 15.30 Uhr. Das mehlige Futter wurde dazu mit Wasser
feuchtkrümelig angemischt. Nach dem Fressen wurde den Tieren Wasser zur beliebigen Aufnahme
vorgesetzt. Der abgesetzte Kot jeden Tieres wurde gesammelt und in einem luftdicht verschlossenen
Plastikeimer bei 6°C im Kühlraum gelagert. Am Ende der Sammelperiode wurde der gesamte Kot
gewogen und eine Probe zur Stickstoffanalyse entnommen. Zur späteren Bestimmung von
Trockensubstanz und Rohnährstoffen wurde je ein Kilogramm Kot gefriergetrocknet.

28
c
d
e
ba Chymus
Gase
Die Verdauungsquotienten der Rohnährstoffe wurden nach der folgenden Formel berechnet.
VQ = ( I � F ) / I � 100
VQ = scheinbare Verdaulichkeit in %
I = mit dem Futter aufgenommene Nährstoffmenge
F = mit dem Kot ausgeschiedene Nährstoffmenge
2.4.2. Ermittlung der ilealen Verdaulichkeit der Aminosäuren
Zur Ermittlung der ilealen Verdaulichkeit der Aminosäuren wurde am Fachgebiet Tierernährung des
Departments für Tierwissenschaften der TU München, Freising-Weihenstephan, ein weiterer
Verdauungsversuch durchgeführt. Zu diesem Zweck standen drei kastrierte männliche Schweine einer
Zweirassenkreuzung, Piétrain x Deutsche Landrasse, zur Verfügung. Diesen wurde eine Ileocaecal-
Anastomose in End-zu-End-Technik unter Erhalt der Ileorectalklappe angelegt, so dass der
Nahrungsbrei nach der Passage des Dünndarmes unter Umgehung des Dickdarmes über den Mastdarm
austrat und für Untersuchungszwecke gesammelt werden konnte (ROTH-MAIER et al., 1998).
Abbildung 2: Schematische Darstellung einer End-zu-End Ileorectal-Anastomose (a = Jejunum,
b = Ileum mit Ostium ileocaecale, c = Caecum, d = Colon, e = Rectum)

29
Das operative Vorgehen orientierte sich wesentlich an den Methoden von HENNIG et al. (1990) und
LAPLACE et al. (1994) und wurde vom Team des Instituts für Experimentelle Chirurgie der
Technischen Universität München unter Leitung von Herrn Prof. Dr. W. Erhart durchgeführt. Die
Einzelhaltung der anastomierten Tiere erfolgte in speziellen Stoffwechselkäfigen, die eine quantitative
Erfassung des verzehrten Futters sowie des ausgeschiedenen Chymus ermöglichten. Auf der Seite des
flexiblen Darmkanülenaustrittes bestand die Käfigwand aus einer Plexiglasplatte, die die
Verletzungsgefahr der Tiere durch Hängenbleiben am Gitter verringerte. Die Käfige waren leicht zu
reinigen und zu desinfizieren, wodurch der erforderliche Hygienestandard während der Experimente
sichergestellt werden konnte. Die Raumtemperatur im vollklimatisierten Stall betrug konstant 21°C bei
einer relativen Luftfeuchtigkeit von 50 %. Die Beleuchtung war in der Zeit von 7.00 bis 17.00 Uhr
eingeschaltet. Die Fütterung erfolgte um 7.00 und um 15.30 Uhr mit jeweils nachfolgender
Chymussammlung. Der 7 Tage dauernden Vorperiode schloss sich eine 5-tägige Sammelperiode an.
Zu Beginn der Sammelperiode wurde das Futter für alle Tiere eingewogen und eine Probe für die
Analysen entnommen. Alle Tiere erhielten pro Mahlzeit 750 g des Laktationsfutters der Gruppe I mit
75 g Futterzucker, 150 ml Elektrolytlösung (mit 3,218 g Na, 0,195 g K, 0,036 g Mg, 0,010 g Ca je
1000 ml H2O) und 1500 ml Wasser zu einem Brei verrührt. Nach der Futteraufnahme wurde den
Tieren Wasser in ausreichender Menge vorgesetzt. Die Hälfte des gesammelten Chymus wurde sofort
zur Ermittlung der Trockensubstanz und zur Analyse gefriergetrocknet und daraus am Ende der
Sammelperiode eine aliquote Mischprobe erstellt. Für die Stickstoffuntersuchung wurde der Rest des
Chymus im Kühlraum bei 6°C gelagert und bei Versuchsende ebenfalls eine aliquote Mischprobe
genommen.
Für Aminosäuren, die dem Futter nicht in synthetischer Form beigemischt wurden, errechnete sich die
scheinbare ileale Verdaulichkeit nach untenstehender Formel.
VQi = ( I � C ) / I � 100
VQi = scheinbare ileale Verdaulichkeit in %
I = mit dem Futter aufgenommene Aminosäurenmenge
C = mit dem Chymus ausgeschiedene Aminosäurenmenge

30
Da bei diesem Versuch das Hauptinteresse auf die Ermittlung der Verdaulichkeit der Aminosäuren in
den natürlichen Futterkomponenten gerichtet war, wurde für Aminosäuren, die dem Futter in
synthetischer Form beigemischt wurden folgende Formel verwendet.
VQi = [( I � S ) - C ] / ( I � S ) � 100
VQi = scheinbare ileale Verdaulichkeit in %
I = mit dem Futter aufgenommene Aminosäurenmenge
S = mit dem Futter aufgenommene synthetische Aminosäurenmenge
C = mit dem Chymus ausgeschiedene Aminosäurenmenge
Dabei wurde davon ausgegangen, dass die scheinbare ileale Verdaulichkeit der synthetischen
Aminosäuren bei 100 % liegt (KIRCHGESSNER und ROTH, 1985; SCHUTTE et al., 1988).
2.5. Erfassung der Untersuchungsparameter
2.5.1. Lebendmasse der Sauen
Die Lebendmasse der Sauen wurde am 60. und 110. Trächtigkeitstag sowie am 1. und 28.
Laktationstag erfasst. Die Wiegung erfolgte etwa eine Stunde nach der morgendlichen Fütterung
mittels einer elektronischen Viehwaage (Fa. Baumann, Modell 2005, Thiersheim). Der Messbereich
der Waage lag zwischen 50 und 1000 kg und die Messgenauigkeit betrug ±1 kg.
2.5.2. Futteraufnahme der Sauen
Zur Ermittlung der täglichen Futteraufnahme der Sauen während der Säugezeit wurden morgens vor
der Fütterung die Futterreste der vorigen Tages aus dem Trog entfernt und eingefroren (-20°C)
wochenweise gesammelt. Die Futterreste wurden danach im Trockenschrank bei 70°C bis zur
Gewichtskonstanz getrocknet. Aus der Differenz von vorgelegtem Futter und Futterrückwaage wurde
die durchschnittliche Futteraufnahme je Sau errechnet.
2.5.3. Milchleistung der Sauen
Am 13. und 14. sowie am 20. und 21. Tag der Laktation wurde die Milchleistung der Sauen mit der
�weigh-suckle-weigh� Methode ermittelt. Diese wurde von RÄDER et al. (1990) modifiziert und
weiterentwickelt sowie durch KIRCHGESSNER et al. (1992) und PAULICKS et al. (1998) ergänzt.

31
Dabei wurde die Milchleistung über den Masseanstieg der Ferkel während der Säugungen ermittelt.
Dazu mussten die Ferkel während des Messzeitraumes von der Sau getrennt werden und nur zum
Säugen wurden sie der Sau wieder zugesetzt. Die Trennung der Ferkel von der Sau erfolgte mittels
eines Absperrgatters, das den Zugang der Ferkel vom Aktivitätsbereich zur Sau verhinderte. Um den
Trennungsstress für Ferkel und Sau gering zu halten, war das Gatter teildurchsichtig. Da der natürliche
Abstand zwischen zwei Säugungen nach BERGE und INDERBOE (1953) etwa 55 bis 65 Minuten
beträgt, wurden die Wiegungen stündlich durchgeführt. Um eine ausreichend genaue Schätzung der
Milchleistung zu erreichen sind nach Räder et al. (1990) sieben Stunden notwendig. Damit
Schwankungen der Milchabgabe in der Eingewöhnungsphase bei der ersten Messung am Tag die
Ergebnisse nicht beeinflussten, wurde die erste erfolgreiche Messung nicht in die Auswertung mit
einbezogen (in Anlehnung an: SPEER und COX 1984). Die Ferkel waren an Messtagen von 6 bis
15.00 Uhr von der Sau abgesperrt. Die erste Messung fand um sieben Uhr statt und wiederholte sich
stündlich. Ließ die Sau die Ferkel bei der ersten Messung nicht saufen, so verschob sich das ganze
Schema um eine Stunde. Spätestens bei der zweiten Messung erfolgte bei allen Sauen eine Säugung.
Um die von den Ferkeln aufgenommene Milchmenge bestimmen zu können, wurden die Ferkel
unmittelbar vor und nach der Säugung mit einer elektronischen Präzisionswaage (Modell EB
Economy-Serie, Fa. Sartorius, Göttingen) mit Mittelwertbildung von 20 Einzelwiegungen und einer
Anzeigegenauigkeit von ±1 g gewogen. Dazu wurden die Ferkel in eine flüssigkeitsundurchlässige
Kunststoffwanne mit Stroheinstreu gesetzt. Um einen stabilen Wert zu erhalten, muss nach
SCHNEIDER (1992) ein Mittelwert aus mindestens hundert Einzelwiegungen vorliegen. Aus diesem
Grund wurde jede Messung 6-mal wiederholt und somit ein Mittelwert aus 120 Einzelwiegungen
verwendet. Koteten oder harnten die Tiere während des Wiegens, wurde dies erfasst und bei der
Berechnung der Milchmenge berücksichtigt. Der stündliche Zyklus bei der �weigh-suckle-weigh�
Methode gliederte sich folgendermaßen:
1.
Herausnehmen der Ferkel aus dem Aktivitätsbereich,Wiegen vor der
Säugung und Zusetzen zur Sau
ca.
5 min
2.
Säugephase mit Erfassung der Kotungen und Harnungen
ca.
10 min
3.
Wegnehmen der Ferkel von der Sau, Wiegen nach der Säugung und
Zurücksetzen in den Aktivitätsbereich
ca.
5 min
4.
Verbleib der Ferkel im Aktivitätsbereich von der Sau getrennt
ca.
40 min

32
Die gewonnenen Ergebnisse wurden um Speichel-, Verdunstungs- und Stoffwechselverluste der Ferkel
während des Säugens korrigiert. Dabei wurde folgende Formel nach SCHNEIDER (1992) verwendet:
Speichel- , Verdunstungs- und Stoffwechselverluste:
s = 1,15 � LM0,75 + 0,418
s
LM0,75
=
=
geschätzter Masseverlust während des Säugens durch Speichel- ,
Verdunstungs- und Stoffwechselverluste in g
metabolische Lebendmasse des Ferkels in kg
Für Kot und Harn, der von den Ferkeln während der Säugephase abgesetzt wurde, wurde ebenfalls
eine Korrektur bei der Milchleistungsberechnung vorgenommen. Dazu wurde das Absetzen von Kot
und Harn anzahlsmäßig erfasst. Die entsprechenden Verluste wurden in Abhängigkeit der
metabolischen Lebendmasse mit folgenden Formeln geschätzt:
Kotverluste:
k = - 1,86 + 0,0132 � W0,75
k
W0,75
=
=
geschätzter Masseverlust während des Säugens durch einmal Koten in g
metabolische Lebendmasse des Ferkels in g
r2 = 20,5 % n = 43 S = 3,2
Harnverluste:
h = 1,23 + 0,0723 � W0,75
h
W0,75
=
=
geschätzter Masseverlust während des Säugens durch einmal Harnen in g
metabolische Lebendmasse des Ferkels in g
r2 = 19,4 % n = 46 S = 19,6

33
Die Einzeldaten zur Ableitung der Regressionen wurden bei der Erfassung der Ferkeleinzelgewichte
am 13./14. und am 20./21. Lebenstag gewonnen, wenn die Ferkel während des Wiegens Kot oder Harn
absetzten. Aus der Anhangstabelle 3 kann weiteres entnommen werden. Die Regressionsgleichungen
wurden trotz ihres geringen Bestimmtheitsmaßes verwendet, da sie den Gesamtfehler bei der
Milchleistungsschätzung geringer halten als ein pauschaler Mittelwert für Kot und Harn.
Aus den stündlichen Masseanstiegen ließ sich die Tagesmilchleistung der Sauen errechnen. Folgende
Formel beschreibt die mathematische Vorgehensweise:
7
MLd = 24/7 � Σ ( ∆mi + gwi + si + nki � ki + nhi � hi ) i=1
MLd = Milchleistung je Tag in g
∆mi = Differenz der Lebendmasse der Ferkel vor und nach Säugung
gwi = Kot- und Harnmassen während des Wiegens vor Säugung
si = Speichel- , Verdunstungs- und Stoffwechselverluste
nki = Anzahl der Kotverluste
ki = geschätzte Kotmasse
nhi = Anzahl der Harnverluste
hi = geschätzte Harnmasse
2.5.4. Lebendmasse der Ferkel
Jeweils am 1., 14., 21. und 28. Lebenstag wurden die Ferkel gewogen. Am ersten Lebenstag erfolgte
die Wiegung innerhalb der dritten und der elften Stunde nach der Geburt, da nach KIRCHGESSNER
und MERK (1984) in diesem Zeitraum noch keine Schwankungen bei der Lebendmasse auftreten. Da
die Geburtsüberwachung von etwa 5.50 bis 22.30 Uhr stattfand, konnte dies bei allen Würfen
eingehalten werden. Das Geburtsende wurde dabei mit dem Abgang der Nachgeburt definiert. Am 14.
und 21. Tag erfolgte die Wiegung um 6.15 Uhr, am 28. Tag in der Regel etwa um 12 Uhr. Zur
Ermittlung der Lebendmasse der Ferkel stand eine elektronische Präzisionswaage (Fa. Sartorius,
Modell EB Economy-Serie, Göttingen) mit Mittelwertbildung von 20 Einzelwiegungen und einer
Anzeigegenauigkeit von ±1 g zur Verfügung.

34
2.5.5. Futterverbrauch der Ferkel
Mittels eines in der Abferkelbucht eingebauten Ferkelfutterautomaten erhielten die Ferkel ab dem 22.
Lebenstag Futter zur freien Aufnahme. Beim ersten Mal wurde der Automat mit 1500 g Futter
beschickt, und bei Bedarf wurden dann jeweils weitere 1500 g nachgefüllt. Die Futterrückwaage
erfolgte zum Zeitpunkt des Absetzens am 28. Laktationstag. Aus der Differenz zur Einwaage ließ sich
der Futteraufverbrauch errechnen. Eine Korrektur um auftretende Futterverluste während des Fressens
der Ferkel wurde nicht durchgeführt.
2.6. Probenahme und Analyseverfahren
2.6.1. Futter- und Kotproben
Für die Untersuchung von Futter und Kot wurde jeweils eine aliquote Mischprobe aus den bei jeder
Charge entnommenen Einzelproben erstellt. Diese wurde beim Futter mit einer Trommelmühle
(Modell ZM 1, Fa. Retsch, Haan) und gefriergetrocknet beim Kot mit einer Schneidmühle (Modell E
380, Fa. Brabender, Duisburg) auf die erforderliche Teilchengröße zerkleinert.
2.6.1.1. Nährstoffanalyse
Die auf 1 mm gemahlenen Proben wurden nach der Weender Futtermittelanalyse untersucht. Die
Bestimmung des Gesamtstickstoffes wurde nach Kjeldahl (bei den Kotproben aus Frischmaterial)
durchgeführt. Nach einer Salzsäurehydrolysierung erfolgte die Bestimmung des Gesamtfettgehaltes bei
den Verdauungsversuchen durch Extraktion mit Petrolether in der Soxhlet-Apparatur. Die
Rohfettanalyse bei den restlichen Proben erfolgte direkt durch eine Extraktion mit Petrolether. Für die
Untersuchung auf Zucker- und Stärkegehalt wurden die Proben auf 0,5 mm gemahlen. Der Gehalt an
Zucker wurde nach dem Verfahren von Luff-Schoorl analysiert (alles nach: NAUMANN und
BASSLER, 1988). Mittels polarimetrischer Methode erfolgte die Bestimmung des Stärkegehaltes nach
der im AMTSBLATT DER EUROPÄISCHEN GEMEINSCHAFT (1972) erschienenen Vorschrift.
2.6.1.2. Aminosäurenbestimmung
Für die Aminosäurenanalyse wurden die Futterproben auf eine Partikelgröße von ≤ 0,5 mm gemahlen.
2.6.1.2.1. Tryptophanbestimmung
Die Untersuchung des Tryptophangehaltes erfolgte am Fachgebiet Tierernährung, Department für
Tierwissenschaften, der TU München, Freising-Weihenstephan. Dazu wurden 225 mg der Probe in
einer Polypropylenflasche mit 8,4 g Bariumhydroxid und 12 ml Wasser versetzt. Die aufgerührte

35
Lösung wurde 20 Stunden lang bei 110°C und 0,4 bar im Autoklaven hydrolysiert. Der heißen
Hydrolysatlösung wurden zunächst 30 ml Wasser und dann genau 2,00 ml der internen Standardlösung
(273 mg α-Methyl-DL-Tryptophan in 500 ml 0.1 M Salzsäure gelöst) zugegeben. Nachdem die
Lösung 15 Minuten unter Rühren im Eisbad gekühlt worden war, wurden nacheinander 5 ml 0,5 M
Phosphorsäure und 5 ml 6 M Salzsäure hinzugegeben. Danach wurde das Hydrolysat mit 1 mol/l HCl
auf einen pH-Wert von 3.00 eingestellt und der Ansatz mit Wasser auf 100 ml verdünnt. Ein Teil des
Hydrolysates wurde über ein Faltenfilter gegeben und 5 ml des Filtrates mit 20 ml 30 %igem Methanol
verdünnt. Die über eine 0,45 µm Membrane gefilterte Lösung wurde innerhalb von 24 Stunden für die
Analyse verwendet.
Für alle Proben wurde eine HPLC-Anlage der Fa. Merck-Hitachi, Darmstadt verwendet, die aus
folgenden Gerätebauteilen bestand:
- Probeaufgabesystem ( Modell L-7200, Autosampler)
- Hochdruckpumpe, isokratisch (Modell L- 7100)
- Vorsäule (RP 18; 5 µm; 17 x 4,6 mm)
- Trennsäule (RP 18; 5 µm; 250 x 4 mm)
- Fluoreszenzdetektor (Modell L-7480)
- PC-gestützter Integrator (Modell D-7000)
Die Steuerung der Geräte und die Auswertung der Ergebnisse erfolgte mit Hilfe der `Chromatography
Data Station Software; HPLC System Manager´ (Version 3.0, Fa. Hitachi, San Jose, USA). Als mobile
Phase diente eine Lösung von 0,01 mol/l KH2PO4 mit 8 % Methanol bei einer konstanten Flussrate
von 1,2 ml/min. Die Injektion der Proben wurde mit einem automatischen Probenaufgabesystem
durchgeführt. Die injizierte Probenmenge betrug immer 20 µl. In einem Wasserbad wurde die
Temperatur der Trennsäule gleichbleibend bei 37°C gehalten. Die Detektion des Tryptophan durch den
Fluoreszenzdetektor erfolgte bei einer Anregungswellenlänge von 280 nm und einer
Emissionswellenlänge von 355 nm. Die Retentionszeit für den Tryptophanpeak betrug zwischen 10
und 11 Minuten. Die Berechnung der einzelnen Tryptophanwerte erfolgte durch einen relativen
Flächenvergleich der Peaks von Tryptophan und internem Standard. Die Retentionszeit des internen
Standards betrug etwa 16 Minuten. Die Detektionszeit je Probe betrug 20 Minuten. Nach jeder
Analyse wurde die Trennsäule mit Methanol gewaschen. In Abbildung 3 ist ein typisches HPLC-
Chromatogramm dargestellt.

36
Abbildung 3: Typisches HPLC-Chromatogramm mit den Peaks von Tryptophan (Mitte) und
internem Standard (rechts)
2.6.1.2.2. Bestimmung der restlichen Aminosäuren
Für die Analyse der restlichen Aminosäuren wurden die Futterproben in Polyflaschen zur Firma
Lohmann Animal Health GmbH & Co. KG nach Cuxhaven geschickt, die die Untersuchung vornahm.
Zur quantitativen Bestimmung der Aminosäuren diente ein Ionenaustausch-Chromatograph (Modell:
LC 2000, Fa. Biotronic, Maintal). Dazu wurden 250 mg der Probe in einer Pyrex Glasflasch mit 25 ml
Hydrolysegemisch (492 ml HCl unter Zusatz von 1 g Phenol auf 1l mit H2O aufgefüllt) versetzt und 24
Stunden bei 110°C getrocknet. Das Hydrolysat wurde im Eisbad gekühlt und mit Pufferlösung
(Citratpufferlösung, pH 2,20, 0,2 mol/l: 19,61 g Natriumcitrat, 1g Phenol, 16,50 ml HCl, 5 ml
Thiodiglycol in 1l H2O lösen und auf pH 2,20 einstellen) quantitativ in ein 250 ml Becherglas
überführt. Unter weiterer Kühlung kamen 20 ml 7,5 mol/l Natronlauge unter Rühren hinzu, und der
pH-Wert wurde mit 2 mol/l Natronlauge auf exakt 2,20 eingestellt. Mit Pufferlösung wurde der Inhalt
in einen 200 ml Messkolben überspült und mit 2 ml internem Standard (0,656 g Norleucin in 250 ml
Messkolben einwiegen, in Pufferlösung auflösen und bis zur Marke auffüllen) versetzt und mit
Pufferlösung bis zur Marke aufgefüllt. Von der Lösung wurden 100 ml durch einen Membranfilter
filtriert. 50 µl des Hydrolysates wurden in den Aminosäurenanalyser appliziert. Da bei dieser
Probenvorbereitung ein Teil der schwefelhaltigen Aminosäuren zerstört wurde, war eine besondere
Vorbereitung für Methionin und Cystin notwendig. Dazu wurden 250 mg Probenmaterial in eine
100:ml Pyrex Glasflasche gegeben und 5 ml Oxydationsmischung (0,5 ml Wasserstoffperoxid mit 4-5
ml 88 %iger phenolhaltiger Ameisensäure und 25 mg Phenol vermischt) zugegeben. Die verschlossene

37
Flasche wurde für 24 Stunden im Eisbad bei 0°C gekühlt. Nach der Zugabe von 0,90 g
Natriumdisulfid wurde die Probe wie oben beschrieben hydrolysiert. Für Hydroxyprolin und Prolin
erfolgte die Extinktionsmessung bei 440 nm, für die anderen Aminosäuren bei 570 nm. Die Fläche der
durch die Messung gewonnenen Peaks des Chromatogramms wurden berechnet und daraus die
Aminosäuregehalte ermittelt.
2.6.1.3. Energiegehalt
Mit den Ergebnissen der Rohnährstoffanalyse konnte mit nachfolgender Gleichung nach
KIRCHGESSNER und ROTH (1983) der Gehalt an umsetzbarer Energie im Trächtigkeitsfutter
geschätzt werden. Als Einheit in der Gleichung werden dabei g/kg verwendet.
ME (MJ/kg TS) = (22,3 XP + 34,1 XL + 17,0 XS + 16,8 XZ + 7,4 OR � 10,9 XF) � 10-3
wobei OR = OM - XP - XL - XS - XZ - XF
Um für die Berechnung des Energiegehaltes im Laktationsfutter eine höhere Genauigkeit zu erzielen,
wurden in einem Verdauungsversuch (siehe 2.4.) die Verdaulichkeitsquotienten der Rohnährstoffe für
das Laktationsfutter ermittelt. Mit der folgenden Gleichung der GESELLSCHAFT FÜR
ERNÄHRUNGSPHYSIOLOGIE (1987) war es möglich, die umsetzbare Energie aus den verdaulichen
Nährstoffen zu berechnen. In der Gleichung werden als Einheit g/kg TS verwendet.
ME (MJ/kg TS) =
0,021 DXP + 0,0374 DXL + 0,0144 DXF + 0,0171 DXX - 0,0014 XZ � 0,0068 (BFS � 100)
Die Korrektur um Zucker (XZ) erfolgt dabei nur bei einem Gehalt von größer oder gleich 80 g/kg TS
(im vorliegenden Versuch war das nicht notwendig) und die Berücksichtigung der bakteriell
fermentierbaren Substanz (BFS) ist nur bei einem Gehalt von mehr als 100 g/kg TS notwendig.
2.6.2. Milchproben
Eine Stunde nach der letzten Milchmengenerfassung wurden jeweils am 13., 14., 20. und 21.
Laktationstag bei jeder Sau 100 ml Milch gemolken. Zu diesem Zweck wurden der Sau 1,5 ml
Oxytocin (Fa. Veyx) intramuskulär zur Erleichterung der Milchabgabe in den Ohrgrund injiziert. Bei
dieser Menge an Oxytocin kommt es nach RÄDER et al. (1990) und DEN HARTOG et al. (1987) zu

38
keiner signifikanten Beeinflussung der Milchinhaltsstoffe. Nach der Reinigung des Gesäuges mit
einem feuchten Lappen erfolgte die Milchentnahme. Es wurde darauf geachtet, dass von jeder aktiven
Zitze etwa die gleiche Menge ermolken wurde. Dies war wichtig, da der Unterschied bei den
Inhaltsstoffen der Milch zwischen dem Vor-, Haupt- und Nachgemelk nicht wesentlich (LENKEIT et
al, 1955), zwischen den einzelnen Drüsenabteilungen jedoch stark schwankt (BRABANT und
SCHULZ, 1968). Das Gesamtgemelk wurde durch Glaswolle (Fa. Merck, Darmstadt) filtriert und
abgefüllt. Für die wöchentlich stattfindende Fett-, Eiweiß-, Laktose- und Harnstoffbestimmung wurden
30 ml mit Natriumazid konserviert und bei + 6°C gelagert. Der Rest wurde bis zur Aminosäureanalyse
bei � 20°C tiefgefroren.
2.6.2.1. Fett-, Eiweiß- und Laktosebestimmung
Die Fett-, Eiweiß- und Laktoseanalyse bei den Milchproben wurde vom Milchprüfring Bayern e.V. in
Wolnzach durchgeführt. Dabei kam ein Infrarotspektralphotometer (Milco-Scan FT 6000, Fa. Foss,
Hamburg) zum Einsatz. Das Messverfahren basiert auf der Infrarotabsorption der zu bestimmenden
Milchinhaltsstoffe im Bereich ihres Absorptionsmaximums. Die Eichung des Gerätes erfolgte mit
Kuhmilch, was nach SCHNEIDER (1992) keinen gravierenden Einfluss auf die Ergebnisse hat.
2.6.2.2. Harnstoffbestimmung
Zur Bestimmung des Harnstoffgehaltes in der Sauenmilch wurden die Proben bei Raumtemperatur
über Nacht aufgetaut und eine aliquote Mischprobe der Milch des 20. und 21. Laktationstages
hergestellt. Die Enteiweißung der Proben fand in einer Kühlzentrifuge (Modell: RC2-B automatic
refrigerated, Fa. Sorvall, Superspeed New Town /Connecticut, USA) bei 7000 g und 8°C statt. Das
Fett setzte sich oben ab und der fettfreie Anteil wurde abdekantiert. Anschließend wurden 100 µl der
fettfreien Milch mit 1000 µl URAC (1,6 g/l Uranylacetat, Fa. Roche Diagnostics GmbH, Mannheim)
enteiweißt und mit einer Tischzentrifuge (Modell: Z 229, Fa. BHG B. Hermle GmbH, Gosheim)
getrennt. Der gewonnene Überstand wurde mit einem Testkit (Urea Nitrogen, Fa. Sigma Diagnostics,
St. Louis, USA) aufbereitet und mit einem Spektralphotometer (Modell: UVIKON 933 Double Beam
UV/VIS Spectrophotometer, Fa. Kontron Instruments, Milano, Italy) bei einer Wellenlänge von 570
nm auf den Harnstoffgehalt hin untersucht.

39
2.6.2.3. Trockensubstanz- und Aminosäurenbestimmung
2.6.2.3.1. Trockensubstanzbestimmung
Zur Bestimmung der Trockensubstanz wurde eine aliquote Mischprobe des 20. und 21.
Laktationstages mit etwa 85 ml Volumen erstellt. Diese wurde anschließend in der Gefriertrocknung
(Modell: Unitop 1000L, Fa. Virtis, New York) 72 Stunden getrocknet. Aus der Gewichtsdifferenz ließ
sich die Trockensubstanz errechnen. Danach wurden die getrockneten Proben gemörsert und in
Polyflaschen abgefüllt.
2.6.2.3.2. Tryptophanbestimmung
Die Bestimmung des Tryptophans erfolgte analog der Analyse im Futter (siehe 2.6.1.2.1.).
2.6.2.3.3. Bestimmung der restlichen Aminosäuren
Für die Analyse der restlichen Aminosäuren wurden die Milchproben in Polyflaschen zur Firma
Lohmann Animal Health GmbH & Co. KG nach Cuxhaven geschickt, welche die Untersuchung
vornahm (siehe 2.6.1.2.2.).
2.6.2.4. Energiegehalt
Der Energiegehalt der Sauenmilch wurde rechnerisch mittels der bei KIRCHGESSNER et al. (1992)
aufgeführten Funktion ermittelt:
Milchenergie (MJ/kg FS) = 0,346 � Fett (%) + 0,268 � Eiweiß (%) + 1,138
Bei Fett und Eiweiß wurde der photometrisch ermittelte Wert verwendet.
2.6.3. Blutproben
Die Blutentnahme bei den Sauen fand am 85. Trächtigkeitstag sowie am 22. und 28. Laktationstag aus
der Halsvene (vena jugularis externa) statt. Dazu wurden die Sauen im Kastenstand beziehungsweise
im Abferkelstand mittels einer Rüsselschlinge fixiert.
Am 85. Trächtigkeitstag und am 28. Laktationstag wurde den nüchternen Sauen morgens um ca. 7.30
Uhr Blut zur Gewinnung von Serum (10 ml Serum-Gel-Monovette; Fa. Sarstedt, Nümbrecht) und zur
sofortigen Untersuchung des Hämatokrit- und Thrombozytenwertes (10 ml lithiumheparinisierte
Monovette; Fa. Sarstedt, Nümbrecht) entnommen. Die Serum-Gel-Monovetten wurde sofort nach der

40
Blutentnahme bei 1500 g zentrifugiert (Modell: Universal, Fa. Hettich) und das abpipettierte Serum
bei -80°C bis zur Analyse eingefroren. Nach der Blutentnahme wurde den Sauen das morgendliche
Futter vorgelegt.
Die Blutprobenentnahme am 22. Laktationstag erfolgte 3 Stunden nach der Fütterung zur Gewinnung
von Plasma (10 ml lithiumheparinisierte Monovette; Fa. Sarstedt, Nümbrecht). Die
lithiumheparinisierte Monovette wurde sofort nach der Blutentnahme bei 1500 g zentrifugiert (Modell:
Universal, Fa. Hettich) und das abpipettierte Plasma bei -80°C bis zur Analyse eingefroren. War bei
mehreren Sauen Blut zu entnehmen, so erfolgte eine zeitversetzte Fütterung im Abstand von 10
Minuten. Diese Zeitspanne war in etwa für die Blutentnahme bei einer Sau notwendig.
2.6.3.1. Aminosäuren- und Harnstoffbestimmung
Die Aminosäuren- und Harnstoffbestimmung in den Plasmaproben erfolgte am Fachgebiet
Tierernährung und Leistungsphysiologie, Department für Tierwissenschaften, der TU München,
Freising-Weihenstephan. Nach dem Auftauen der Proben wurden 900 µl Plasma mit 100 µl des
internen Standards Norleucin versetzt. Davon wurden 800 µl mit 200 µl einer zehnprozentigen
Sulfosalicylsäure enteiweißt um störende Proteine zu entfernen. Anschließend wurde die Lösung bei
4°C für 30 Minuten gekühlt und der Niederschlag abzentrifugiert (Modell: Zentrifuge 3200, Fa.
Eppendorf). Der Überstand wurde 1:2 mit einem Probenverdünnungspuffer vermischt. Die Lösung
wurde in den Ionenaustauschchromatographen (Model LC 3000, Fa. Biotronic, Maintal) zur Analyse
der Aminosäuren- und Harnstoffkonzentration gegeben. Die Bestimmung der Konzentrationen erfolgte
aus dem Verhältnis der Peakflächen der analysierten Stoffe zur Peakfläche des Standards.
2.6.3.2. Hämatokrit- und Thrombozytenbestimmung
Aus dem am 85. Trächtigkeits- und am 22. Laktationstag genommenen lithiumheparinisierten Blut
wurde der Hämatokritwert und die Thrombozytenanzahl ermittelt. Dazu stand auf der Versuchsstation
für Futterbau und Tierernährung, Versuchsgut Hirschau der TU-München, Freising-Weihenstephan,
ein Analyser (Modell: Coulter-Counter T 840 for veterinary use, Fa. Coulter Elektronics, Krefeld) des
Fachgebietes Tierernährung, Department für Tierwissenschaften, der TU München, Freising-
Weihenstephan zur Verfügung. Das Blut in den Monovetten wurde sofort nach der Blutentnahme bei
den Sauen durch leichtes Schwenken homogenisiert und 1 ml in ein Eppendorf-Hütchen pipettiert. Aus
diesem entnahm der Autoanalyser die notwendige Menge Blut zur Untersuchung. Da je Sau zwei
Monovetten Blut entnommen und jede zweimal beprobt wurde, stellen die Ergebnisse einen Mittelwert
aus vier Einzelwerten dar. Nach einer Eichung anhand menschlichen Kontrollblutes wies das Gerät
nach Herstellerangaben auch für Schweineblut eine ausreichend hohe Genauigkeit auf.

41
2.6.3.3. Serotoninbestimmung
Die Serotoninmessung in den Serumproben des 85. Trächtigkeitstages und des 28. Laktationstages
wurde im Labor der Firma DLD Diagnostica GmbH im Hamburg durchgeführt. Es kam ein 125I-
Radioimmunoassay für die quantitative Bestimmung von Serotonin im Serum (Fa. DLD Diagnostica
GmbH, Hamburg) zum Einsatz.
Nach dem Auftauen der Proben wurden 25 µl Serum in Polypropylenröhrchen pipettiert. Dazu wurden
250 µl Acylierungspuffer und 25 µl Acylierungsreagenz eingemischt, 30 Minuten bei Raumtemperatur
inkubiert und im Anschluss 2 ml bidestilliertes Wasser zur Verdünnung hinzugegeben. Je 25 µl der so
vorbereiteten Proben wurden in RIA-Röhrchen (Polystyrol) gefüllt und dazu 50 µl 125I-Tracer
einpipettiert. Nach der Zugabe von 50 µl Antiserum wurde die Lösung gut gemischt und eine Minute
bei 500 g zentrifugiert. Im Anschluss wurden die Röhrchen 90 Minuten lang bei 2 � 8°C inkubiert.
Nach Zugabe von 500 µl kaltem (2 � 8°C) praezipitierendem Antiserum erfolgte eine abermalige 15-
minütige Inkubation bei 2 � 8°C. Unter Kühlung wurden die Röhrchen 15 Minuten bei 3000 g
zentrifugiert und der Überstand abgesaugt. Die Proben wurden eine Minute in einem Gamma-Counter
(Fa. Berthold, Bad Wildbach) gemessen.
Nach folgender Formel wurde aus dem untersuchten Serotoningehalt im Serum und dem
Hämatokritwert der Serotoningehalt im Vollblut ermittelt.
Sv = Ss : [1 : (1 � HCT)]
Sv
Ss
HCT
=
=
=
Serotonin in ng / ml Vollblut
Serotonin in ng / ml Serum
Hämatokritwert in Dezimalschreibweise
Aus den Ergebnissen der Hämatokrit- und Thrombozytenbestimmung sowie der oben beschriebenen
Analyse wurde der Gehalt auf ng Serotonin je 109 Thrombozyten umgerechnet. Folgende Formel
beschreibt die mathematische Vorgehensweise.

42
St = {Ss : [1 : (1 � HCT)]} : PLT
St
Ss
HCT
PLT
=
=
=
=
Serotonin in ng / 109 Thrombozyten
Serotonin in ng / ml Serum
Hämatokritwert in Dezimalschreibweise
Thrombozytenanzahl in 109
2.7. Statistische Auswertung und Ergebnisdarstellung Der Versuch wurde als unvollständige Blockanlage geplant und mittels der Varianzanalyse
ausgewertet. Unvollständig war die Blockanlage daher, da nicht jedes Versuchstier allen sechs
Behandlungen unterzogen wurde und somit ein Block von mehreren Sauen gebildet wurde.
Das varianzanalytische Muster wird durch nachfolgende Gleichung beschrieben:
yijk = µ + ai + bj + ck + eijk
yijk = Beobachtungswert des Blockes i der Behandlung j zum Zeitpunkt k
µ = Mittelwert aller Tiere
ai = Effekt des i-ten Blockes
bj = Effekt der j-ten Behandlung
ck = Effekt des k-ten Messzeitpunktes
eijk = Zufällige Abweichung des Blockes i bei der Behandlung j zum
Messzeitpunkt k
Die ermittelten Messwerte wurden zuerst auf die Nullhypothese `es existieren keine Unterschiede
zwischen den Behandlungen´ geprüft. Dabei galten die Faktoren Behandlung, Messzeitpunkt und
Block als fix und wurden jeweils gegen den Rest getestet. Interaktionen wurden im Modell nicht
berücksichtigt. War der F-Test statistisch gesichert (p<0,05), erfolgte ein multipler Mittelwertvergleich
für den Faktor Behandlung nach der Student-Newman-Keuls Methode.
In den Tabellen sind die Mittelwerte sowie die Standardabweichungen der Einzelwerte eingetragen.
Wurden signifikante Unterschiede (p<0,05) zwischen den Gruppenmittelwerten festgestellt, erfolgte

43
eine Kennzeichnung der entsprechenden Werte mit unterschiedlichen kleinen Hochbuchstaben. Bei
Bedarf wurde auch ein Mittelwertvergleich für die verschiedenen Messzeitpunkte durchgeführt. Traten
signifikante Unterschiede (p<0,05) auf, wurden diese mit verschiedenen großen Hochbuchstaben
gekennzeichnet.
Die Auswertung der Daten nach statistischen Gesichtspunkten wurde mit Unterstützung des
Programmpaketes Statistical Analysis System, Version 8.2 (SAS Institute Inc., 1999, Cary, North
Carolina, USA) durchgeführt.
Die Regressions- und Korrelationsanalysen wurden nach Standardmethoden (PRECHT und KRAFT,
1993) durchgeführt.

44
3. Ergebnisse
3.1. Verdaulichkeiten
3.1.1. Verdaulichkeit und Energiegehalt des Laktationsfutters
Die Verdauungsquotienten der Rohnährstoffe in den beiden Futtermischungen für Gruppe I und IV
und die daraus errechneten Gehalte an umsetzbarer Energie sind in Tabelle 14 angegeben.
Tabelle 14: Verdaulichkeit der Rohnährstoffe und Gehalt an umsetzbarer Energie
Futter Tryptophangehalt in %
I 0,12
IV 0,24
VQ in %
Trockensubstanz
organische Substanz
Rohprotein
Rohasche
Rohfaser
Gesamtfett
NfE
Energie in MJ / kg FS1)
ME
82,67 ±1,91
85,01 ±1,72
81,35 ±2,43
46,24 ±6,66
52,89 ±6,91
63,82 ±2,88
89,71 ±1,55
13,2
81,82 ±1,68
83,94 ±1,72
82,77 ±2,56
49,54 ±3,03
49,15 ±6,60
66,06 ±1,51
88,44 ±1,00
13,2 1) berechnet nach GESELLSCHAFT FÜR ERNÄHRUNGSPHYSIOLOGIE (1987)
Zwischen den Gruppen traten bei den Verdauungsquotienten und dem Energiegehalt keine
signifikanten Unterschiede auf. Die Verdaulichkeit der organischen Substanz lag mit durchschnittlich
84 % über den 80 Prozent, die für ein Alleinfuttermittel für Zuchtsauen erforderlich sind
(KIRCHGESSNER, 1997).
Der Energiegehalt im Laktationsfutter, errechnet anhand der von der GESELLSCHAFT FÜR
ERNÄHRUNGSPHYSIOLOGIE (1987) benannten Formel für Schweinemischfuttermittel aus den
verdaulichen Nährstoffen (siehe: 3.6.1.3.), betrug 13,2 MJ ME/kg FS. Der ermittelte Wert lag somit im
angestrebten Bereich.

45
3.1.2. Ileale Verdaulichkeit des Rohproteins und ausgewählter Aminosäuren
In Tabelle 15 sind die Ergebnisse der scheinbaren ilealen Verdaulichkeit verschiedener Aminosäuren
dargestellt, wie sie im vorliegenden Versuch ermittelt wurden.
Tabelle 15: Scheinbare ileale Verdaulichkeit von Rohprotein und ausgewählten Aminosäuren
im Laktationsfutter I in %
Rohprotein
75,8 ±1,88
Lysin 68,3 ±3,07
Methionin 85,5 ±0,65
Cystin 80,2 ±4,32
Threonin 77,0 ±4,70
Tryptophan 68,5 ±3,13
Die scheinbare ileale Verdaulichkeit lag beim Rohprotein bei 76 % und schwankte bei den fünf
erstlimitierenden Aminosäuren zwischen 68 % und 86 %
3.2. Futter- Nährstoff- und Energieaufnahme der Sauen
3.2.1. Futteraufnahme
In Tabelle 16 sind die Behandlungsmittelwerte der Futterverzehrserfassung eingetragen. Während der
ersten Laktationswoche waren bei der Futteraufnahme der Sauen noch keine statistisch gesicherten
Unterschiede zu erkennen. Jedoch fiel die Gruppe I auf, die mit 2,7 kg einen um etwa 0,8 kg
geringeren Verzehr hatte als die restlichen Gruppen mit durchschnittlich 3,5 kg. In der zweiten Woche
war ein signifikanter Unterschied zwischen Gruppe I und IV zu erkennen, wobei die absolute
Differenz bereits über 1,1 kg betrug. Die anderen Gruppen unterschieden sich nur unwesentlich und
lagen zwischen den beiden Extremen. Der anfängliche Trend der Gruppen I und IV mit den Sauen des
geringsten beziehungsweise höchsten Futterverzehrs in der ersten und zweiten Woche wurde in der
dritten und vierten Woche sowie beim Gesamtdurchschnitt signifikant. Während der dritten
Laktationswoche war die Futteraufnahme der Gruppe III etwas geringer als die der Gruppen II, V und
VI.
Nachdem die Futteraufnahme der Gruppen II und VI in der vierten Laktationswoche ebenfalls unter
der der Gruppen IV und V lag, hatte Gruppe IV über die gesamte Laktation mit durchschnittlich 5,5 kg
den höchsten Futterverzehr. Somit zeigte der Tryptophangehalt von 0,24 % im Futter einen
signifikanten, den Futterverzehr steigernden Effekt.

46
Tabelle 16: Tägliche Futteraufnahme der Sauen in kg
Gruppe I II III IV V VI Trp.-Geh. in % 0,12 0,15 0,18 0,24 0,30 0,42 Mittel
1. Laktationswoche 2,746 3,535 3,577 3,930 3,414 3,278 3,413 ±1,044 ±1,091 ±1,078 ±0,963 ±0,929 ±1,349 ±1,076
2. Laktationswoche 3,426b 4,052ab 4,277ab 5,562a 4,451ab 4,109ab 4,313 ±1,325 ±0,987 ±1,295 ±1,050 ±1,506 ±1,787 ±1,325
3. Laktationswoche 3,231c 4,917ab 4,789b 6,247a 4,892ab 5,128ab 4,867 ±1,127 ±1,376 ±1,576 ±0,920 ±1,117 ±1,370 ±1,248
4. Laktationswoche 3,575c 5,057b 4,912b 6,365a 5,815ab 4,832b 5,093 ±0,945 ±1,367 ±1,652 ±0,924 ±1,029 ±1,094 ±1,169
1.- 4. Lakt.woche 3,245c 4,386b 4,393b 5,532a 4,657b 4,342b 4,426 ±0,910 ±0,810 ±1,193 ±0,661 ±0,823 ±1,144 ±0,924
Die geringste Fresslust hatten die Tiere der Gruppe I. Der Tryptophangehalt von 0,12 % dieser
Futtergruppe zeigte zu jedem Zeitpunkt der Laktation (unter Einbeziehung des nicht statistisch
abgesicherten Trends in der ersten Laktationswoche) einen restriktiven Einfluss auf den täglichen
Futterverzehr der Sauen. Die Futteraufnahme der Gruppen II, III, V und VI lag mit durchschnitlich 4,4
kg zwischen den Werten der Gruppen I und IV. Die von den einzelnen Sauen verzehrten Futtermengen
sind in der Anhangstabelle 19 eingetragen.
3.2.2. Proteinaufnahme
Wie aus der Tabelle 17 ersichtlich ist, traten während der ersten Säugewoche zwischen den Gruppen
keine signifikanten Unterschiede bei der Proteinaufnahme über das Futter auf. Bei einer näheren
Betrachtung der Werte fällt jedoch auf, dass die Gruppe I einen gegenüber den anderen Gruppen
tendenziell niedrigeren Wert aufwies. Dieser Abstand wurde, auch statistisch gesehen, mit jeder
Laktationswoche größer. Während in der zweiten Woche nur Gruppe IV signifikant von I verschieden
war, waren es ab der dritten Laktationswoche alle anderen Gruppen. Auffallend ist, dass in der dritten
Woche die Gruppen II � VI statistisch betrachtet ein Plateau bilden, wobei jedoch die Proteinaufnahme
der Gruppe IV den höchsten Wert aufweist. Aufgrund der weitgehend gleichen Proteingehalte in den
verschiedenen Futtermischungen spiegelte die Proteinaufnahme im wesentlichen den Futterverzehr
wider.

47
Tabelle 17: Tägliche Proteinaufnahme der Sauen in g
Gruppe I II III IV V VI Trp.-Geh. in % 0,12 0,15 0,18 0,24 0,30 0,42 Mittel
1. Laktationswoche 428 559 572 605 533 511 535 ±162 ±172 ±172 ±148 ±145 ±210 ±168
2. Laktationswoche 534b 640ab 684ab 857a 694ab 641ab 675 ±207 ±156 ±207 ±162 ±245 ±279 ±209
3. Laktationswoche 504b 777a 766a 962a 763a 800a 762 ±176 ±217 ±252 ±142 ±174 ±214 ±196
4. Laktationswoche 558c 799ab 786ab 980a 907ab 754b 797 ±147 ±216 ±264 ±142 ±161 ±171 ±184
1.- 4. Lakt.woche 506c 693ab 703ab 852a 726ab 677b 693 ±142 ±128 ±191 ±102 ±128 ±178 ±145
In der vierten Woche und beim Gesamtdurchschnitt hatten die Tiere der Gruppe IV mit 980
beziehungsweise 852 g/Tag die höchste Proteinaufnahme. Die Aufnahme der Gruppe VI war im
Vergleich dazu abgefallen und unterschied sich signifikant von den Gruppen I und IV. Die anderen
Gruppen lagen bei der Proteinaufnahme zwischen den Gruppen IV und VI.
3.2.3. Tryptophanaufnahme
In der Tabelle 18 ist die mittlere tägliche Tryptophanaufnahme in Gramm je Sau eingetragen. Die
durchschnittliche Tryptophanaufnahme je Tag über die gesamte Laktationsdauer stieg von Gruppe I
mit 3,89 g bis zur Gruppe VI auf 17,24 g an.
Tabelle 18: Tägliche Tryptophanaufnahme je Sau in g
Gruppe I II III IV V VI Trp.-Geh. in % 0,12 0,15 0,18 0,24 0,30 0,42 Mittel 1. Laktationswoche 3,30d 5,27cd 6,47c 9,31b 9,80b 13,01a 7,86 ±1,25 ±1,63 ±1,95 ±2,28 ±2,67 ±5,36 ±2,52 2. Laktationswoche 4,11b 6,04b 7,74b 13,18a 12,77a 16,31a 10,03 ±1,59 ±1,47 ±2,34 ±2,49 ±4,32 ±7,09 ±3,22 3. Laktationswoche 3,88d 7,33c 8,67c 14,81b 14,04b 20,36a 11,51 ±1,35 ±2,05 ±2,85 ±2,18 ±3,21 ±5,44 ±2,85 4. Laktationswoche 4,29d 7,53c 8,89c 15,09b 16,69b 19,18a 11,95 ±1,13 ±2,04 ±2,99 ±2,19 ±2,95 ±4,34 ±2,61 1.- 4. Lakt.woche 3,89d 6,54c 7,95c 13,11b 13,37b 17,24a 10,35 ±1,09 ±1,21 ±2,16 ±1,57 ±2,36 ±4,54 ±2,15

48
3.2.4. Energieaufnahme
In Tabelle 19 sind die Mittelwerte der täglichen Energieaufnahme je Sau eingetragen. Während der
ersten Laktationswoche war bei der Energieaufnahme der Sauen noch kein statistisch gesicherter
Unterschied zu erkennen. Jedoch fiel Gruppe I auf, die mit 36 MJ eine um etwa 16 MJ geringere
Energieversorgung aus dem Futter hatte als die Gruppe IV mit 52 MJ. In der zweiten Woche war
dieser Unterschied bereits signifikant, wobei die absolute Differenz jetzt schon über 28 MJ betrug. Die
anderen Gruppen unterschieden sich nur unwesentlich voneinander und lagen zwischen den beiden
Extremen. Der Unterschied zwischen den Gruppen I und IV mit den Sauen, die die geringste
beziehungsweise die höchste Energieaufnahme hatten, war in der dritten und vierten Woche, sowie
beim Gesamtdurchschnitt signifikant. Während der dritten Laktationswoche war die
Energieversorgung der Gruppe III etwas geringer als die der Gruppen II, V und VI.
Tabelle 19: Tägliche Energieaufnahme der Sauen in MJ ME
Gruppe I II III IV V VI Trp.-Geh. in % 0,12 0,15 0,18 0,24 0,30 0,42 Mittel
1. Laktationswoche 36,2 46,7 47,2 51,9 45,1 43,3 45,1 ±13,8 ±14,4 ±14,2 ±12,7 ±12,3 ±17,8 ±14,2
2. Laktationswoche 45,2b 53,5ab 56,5ab 73,4a 58,8ab 54,2ab 56,9 ±17,5 ±13,0 ±17,1 ±13,9 ±19,9 ±23,6 ±17,5
3. Laktationswoche 42,6c 64,9ab 63,2b 82,5a 64,6ab 67,7ab 64,2 ±14,9 ±18,2 ±20,8 ±12,1 ±14,7 ±18,0 ±16,5
4. Laktationswoche 47,2c 66,8b 64,8b 84,0a 76,8ab 63,8b 67,2 ±12,5 ±18,0 ±21,8 ±12,2 ±13,6 ±14,4 ±15,4
1.- 4. Lakt.woche 42,8c 57,9b 58,0b 73,0a 61,5b 57,3b 58,4 ±12,0 ±10,7 ±15,7 ±8,7 ±10,9 ±15,1 ±12,1
Nachdem die Energieaufnahme der Gruppen II und VI in der vierten Laktationswoche ebenfalls hinter
der der Gruppen IV und V blieb, hatte die Gruppe IV im Durchschnitt über die gesamte Laktation mit
73 MJ die höchste Energieversorgung. So lag die Energieaufnahme der Gruppen II, III, V und VI mit
durchschnitlich 59 MJ zwischen den Werten der Gruppen I und IV. Aufgrund der gleichen
Energiegehalte in den verschiedenen Futtermischungen spiegelte die Energieaufnahme im
wesentlichen den Futterverzehr wieder.

49
3.3. Lebendmasse und -entwicklung der Sauen In Tabelle 20 sind die ermittelten durchschnittlichen Lebendmassen der Sauen zu den verschiedenen
Wiegeterminen aufgeführt. Die Einzeldaten sind aus der Anhangstabelle 20 ersichtlich. Sowohl
zwischen den Gruppen als auch zwischen den Messzeitpunkten bestanden bei den Lebendmassen zu
keinem Zeitpunkt signifikante Unterschiede.
Tabelle 20: Lebendmassen der Sauen am 110. Trächtigkeitstag (TT) sowie am 1. und 28.
Laktationstag (LT) in kg
Gruppe I II III IV V VI Trp.-Geh. in % 0,12 0,15 0,18 0,24 0,30 0,42 Mittel
Sauenmasse 110. TT 260 254 259 262 256 266 260 ±27 ±26 ±24 ±31 ±33 ±30 ±29
Sauenmasse 1. LT 239 241 239 249 238 246 242 ±25 ±27 ±27 ±31 ±35 ±32 ±30
Sauenmasse 28. LT 208 224 220 240 224 233 225 ±37 ±29 ±30 ±34 ±31 ±32 ±32
Die Lebendmasseverluste der Sauen unter dem Einfluss des Tryptophangehaltes im Futter sind in der
Tabelle 21 aufgeführt. Den größten Masseverlust während der Laktation hatte die Gruppe I mit über 30
kg, was 1100 g/Tag entsprach. Die Abnahme an Körpersubstanz war somit signifikant höher als die
der Gruppen IV bis VI, die im Mittel nur 12 kg abnahmen. Die Tiere mit der Basisration nahmen somit
über 3-mal mehr ab als die Tiere der Gruppe IV mit 339 g/Tag. Die geringste Lebendmasseabnahme
von nur 9,5 kg war bei Gruppe IV zu beobachten, was jedoch statistisch nicht abzusichern war. Die
Gruppen II und III lagen mit 16,9 beziehungsweise 19,2 kg zwischen den Extremen. Über alle
Gruppen beobachtet betrug der durchschnittliche Masseverlust 17, 3 kg, was 617 g/Tag entsprach.
Tabelle 21: Masseverlust der Sauen während der Laktation
Gruppe I II III IV V VI Trp.-Geh. in % 0,12 0,15 0,18 0,24 0,30 0,42 Mittel
Masseverlust ges. in kg 30,8a 16,9ab 19,2ab 9,5b 14,1b 13,1b 17,3 ±15,6 ±20,2 ±18,7 ±11,4 ±7,5 ±14,1 ±14,6
Masseverlust in g/Tag 1100a 604ab 686ab 339b 504b 468b 617 ±557 ±721 ±668 ±407 ±v268 ±504 ±521
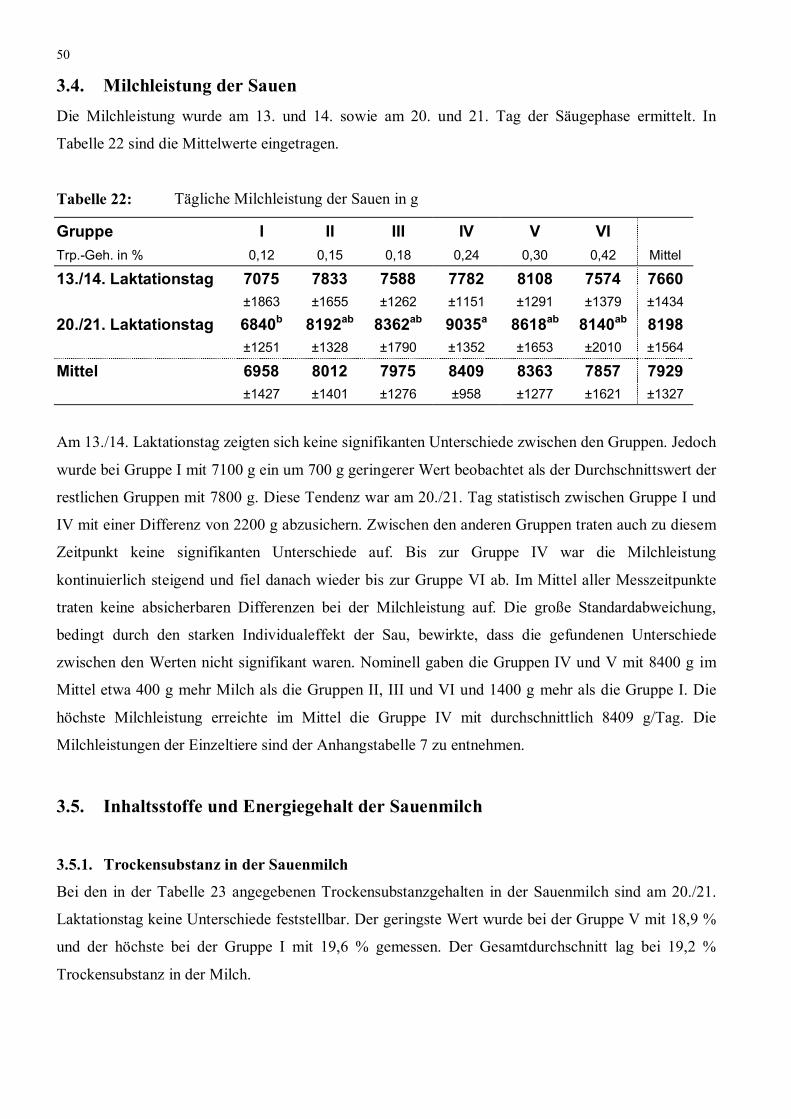
50
3.4. Milchleistung der Sauen Die Milchleistung wurde am 13. und 14. sowie am 20. und 21. Tag der Säugephase ermittelt. In
Tabelle 22 sind die Mittelwerte eingetragen.
Tabelle 22: Tägliche Milchleistung der Sauen in g
Gruppe I II III IV V VI Trp.-Geh. in % 0,12 0,15 0,18 0,24 0,30 0,42 Mittel
13./14. Laktationstag 7075 7833 7588 7782 8108 7574 7660 ±1863 ±1655 ±1262 ±1151 ±1291 ±1379 ±1434
20./21. Laktationstag 6840b 8192ab 8362ab 9035a 8618ab 8140ab 8198 ±1251 ±1328 ±1790 ±1352 ±1653 ±2010 ±1564
Mittel 6958 8012 7975 8409 8363 7857 7929 ±1427 ±1401 ±1276 ±958 ±1277 ±1621 ±1327
Am 13./14. Laktationstag zeigten sich keine signifikanten Unterschiede zwischen den Gruppen. Jedoch
wurde bei Gruppe I mit 7100 g ein um 700 g geringerer Wert beobachtet als der Durchschnittswert der
restlichen Gruppen mit 7800 g. Diese Tendenz war am 20./21. Tag statistisch zwischen Gruppe I und
IV mit einer Differenz von 2200 g abzusichern. Zwischen den anderen Gruppen traten auch zu diesem
Zeitpunkt keine signifikanten Unterschiede auf. Bis zur Gruppe IV war die Milchleistung
kontinuierlich steigend und fiel danach wieder bis zur Gruppe VI ab. Im Mittel aller Messzeitpunkte
traten keine absicherbaren Differenzen bei der Milchleistung auf. Die große Standardabweichung,
bedingt durch den starken Individualeffekt der Sau, bewirkte, dass die gefundenen Unterschiede
zwischen den Werten nicht signifikant waren. Nominell gaben die Gruppen IV und V mit 8400 g im
Mittel etwa 400 g mehr Milch als die Gruppen II, III und VI und 1400 g mehr als die Gruppe I. Die
höchste Milchleistung erreichte im Mittel die Gruppe IV mit durchschnittlich 8409 g/Tag. Die
Milchleistungen der Einzeltiere sind der Anhangstabelle 7 zu entnehmen.
3.5. Inhaltsstoffe und Energiegehalt der Sauenmilch
3.5.1. Trockensubstanz in der Sauenmilch
Bei den in der Tabelle 23 angegebenen Trockensubstanzgehalten in der Sauenmilch sind am 20./21.
Laktationstag keine Unterschiede feststellbar. Der geringste Wert wurde bei der Gruppe V mit 18,9 %
und der höchste bei der Gruppe I mit 19,6 % gemessen. Der Gesamtdurchschnitt lag bei 19,2 %
Trockensubstanz in der Milch.

51
Tabelle 23: Trockensubstanzgehalt in der Sauenmilch am 20./21. Laktationstag in %
Gruppe I II III IV V VI Trp.-Geh. in % 0,12 0,15 0,18 0,24 0,30 0,42 Mittel
Trockensubstanz 19,6 19,2 19,4 19,0 18,9 19,3 19,2 ±1,5 ±0,9 ±1,6 ±0,7 ±1,0 ±1,6 ±1,2
In der Anhangstabelle 18 können die einzelnen Werte nachgelesen werden.
3.5.2. Fettgehalt und �ausscheidung
In Tabelle 24 ist der prozentuale Fettgehalt der Sauenmilch dargestellt.
Tabelle 24: Fettgehalt der Sauenmilch in %
Gruppe I II III IV V VI Trp.-Geh. in % 0,12 0,15 0,18 0,24 0,30 0,42 Mittel
13./14. Laktationstag 8,30 8,34 7,97 8,14 7,84 8,72 8,22A
±1,48 ±1,01 ±1,46 ±0,89 ±0,80 ±1,35 ±1,17
20./21. Laktationstag 7,94 7,58 7,97 7,17 7,25 7,52 7,57B
±1,44 ±1,10 ±1,77 ±0,81 ±0,85 ±1,15 ±1,19 Mittel 8,12 7,96 7,97 7,66 7,54 8,12 7,90 ±1,37 ±0,76 ±1,48 ±0,76 ±0,70 ±1,13 ±1,03
Zu keinem Zeitpunkt konnte ein Zusammenhang zwischen der Tryptophanzulage im Futter und dem
Milchfettgehalt festgestellt werden. Jedoch ließ sich ein signifikanter Einfluss des Messzeitpunktes
innerhalb der Laktation auf den Fettgehalt nachweisen. So verringerte sich der mittlere Fettgehalt über
alle Gruppen vom 13./14. Laktationstag mit 8,22 % auf 7,57 % am 20./21. Laktationstag.
Aus Tabelle 25 kann die tägliche Fettausscheidung der Sauen über die Milch ersehen werden.
Tabelle 25: Fettausscheidung mit der Sauenmilch in g/Tag
Gruppe I II III IV V VI Trp.-Geh. in % 0,12 0,15 0,18 0,24 0,30 0,42 Mittel
13./14. Laktationstag 588 646 601 640 636 649 627 ±183 ±118 ±139 ±141 ±118 ±91 ±132
20./21. Laktationstag 542 625 651 647 623 606 616 ±128 ±159 ±159 ±114 ±140 ±154 ±142 Mittel 565 635 626 642 630 628 621 ±144 ±123 ±130 ±96 ±107 ±103 ±117

52
Hier ist zwar kein signifikanter Einfluss des Tryptophangehaltes im Futter auf die Fettausscheidung zu
erkennen, die Werte der Gruppe I lagen jedoch zu jedem Untersuchungstermin etwa 11 % unter dem
Niveau der restlichen Gruppen. Im Mittel der Messzeitpunkte hatte die Gruppe IV mit einer
durchschnittlichen Fettausscheidung von 642 g/Tag die höchste, Gruppe I mit 565 g/Tag die niedrigste
Fettausscheidung mit der Sauenmilch. Die Werte der einzelnen Sauen können der Anhangstabelle 11
entnommen werden.
3.5.3. Eiweißgehalt und �ausscheidung
Beim Eiweißgehalt der Sauenmilch war weder ein Einfluss der Futtergruppe noch einer des
Messzeitpunktes festzustellen, wie aus Tabelle 26 ersichtlich ist. Die Mittelwerte lagen am ersten
Messtermin bei 4,66 % und beim zweiten bei 4,72 %. Der Gesamtdurchschnitt beim Eiweißgehalt
betrug im Mittel 4,68 %.
Tabelle 26: Eiweißgehalt der Sauenmilch in %
Gruppe I II III IV V VI Trp.-Geh. in % 0,12 0,15 0,18 0,24 0,30 0,42 Mittel
13./14. Laktationstag 4,63 4,61 4,70 4,65 4,61 4,73 4,66 ±0,36 ±0,33 ±0,40 ±0,27 ±0,31 ±0,38 ±0,34
20./21. Laktationstag 4,68 4,72 4,80 4,72 4,62 4,77 4,72 ±0,40 ±0,27 ±0,24 ±0,30 ±0,36 ±0,52 ±0,35 Mittel 4,66 4,67 4,75 4,69 4,62 4,75 4,68 ±0,35 ±0,29 ±0,24 ±0,26 ±0,31 ±0,43 ±0,31
Bei der täglichen Proteinausscheidung (siehe Tabelle 27) am 13./14. Laktationstag traten keine
signifikanten Unterschiede zwischen den Gruppen auf. Den höchsten Wert hatte die Gruppe V mit 371
g/Tag. Auffallend war der Wert der Gruppe I, der sich zwar nicht signifikant von den restlichen
Gruppen unterschied, jedoch mit 325 g/Tag um 10 % niedriger lag als das Niveau der anderen Werte
mit 360 g/Tag. Am 20./21. Laktationstag war das Verhältnis ähnlich, jedoch hatte hier die Gruppe IV
mit 428 g/Tag den höchsten Wert und unterschied sich damit signifikant von der Gruppe I mit 319
g/Tag. Im Mittel der Messzeitpunkte hatte Gruppe IV ebenfalls die höchste tägliche
Eiweißausscheidung mit 394 g. Der Einfluss des Laktationstages war zwischen dem 13./14. und
20./21. Tag signifikant und führte zu einer Steigerung der täglichen Proteinabgabe um 31 g von 354 g
auf 385 g. Die Werte der einzelnen Sauen können aus Anhangstabelle 11 ersehen werden.

53
Tabelle 27: Eiweißausscheidung mit der Sauenmilch in g/Tag
Gruppe I II III IV V VI Trp.-Geh. in % 0,12 0,15 0,18 0,24 0,30 0,42 Mittel
13./14. Laktationstag 325 360 354 361 371 355 354B
±80 ±70 ±54 ±51 ±47 ±50 ±59
20./21. Laktationstag 319b 386ab 400ab 428a 396ab 381ab 385A
±57 ±65 ±81 ±77 ±70 ±75 ±71 Mittel 322b 373ab 377ab 394a 383ab 368ab 370 ±64 ±64 ±56 ±50 ±52 ±59 ±58
3.5.4. Laktosegehalt und �ausscheidung
Beim Laktosegehalt der Sauenmilch war, wie aus Tabelle 28 ersichtlich ist, weder zwischen den
Gruppen, noch zwischen den Messzeitpunkten ein signifikanter Unterschied zu finden. Der
Laktosegehalt lag am 13./14. Laktationstag im Durchschnitt aller Gruppen bei 5,69 % und am 20./21.
Tag bei 5,81 %.
Tabelle 28: Laktosegehalt der Sauenmilch in %
Gruppe I II III IV V VI Trp.-Geh. in % 0,12 0,15 0,18 0,24 0,30 0,42 Mittel
13./14. Laktationstag 5,74 5,74 5,58 5,71 5,68 5,70 5,69 ±0,20 ±0,18 ±0,26 ±0,09 ±0,28 ±0,20 ±0,20
20./21. Laktationstag 5,76 5,77 5,70 5,88 5,89 5,83 5,81 ±0,20 ±0,17 ±0,22 ±0,15 ±0,14 ±0,25 ±0,19 Mittel 5,75 5,75 5,64 5,79 5,78 5,77 5,76 ±0,18 ±0,15 ±0,21 ±0,09 ±0,16 ±0,21 ±0,16
Die tägliche Laktoseausscheidung (siehe Tabelle 29) war am 13./14. Laktationstag zwischen allen
Gruppen nicht signifikant unterschiedlich. Beim zweiten Beobachtungszeitpunkt war die Differenz
zwischen Gruppe IV mit dem höchsten Wert, und der Gruppe I mit dem geringsten Wert, signifikant.
Die restlichen Gruppen lagen mit durchschnittlich 484 g/Tag dazwischen. Im Mittel der
Messzeitpunkte hatte Gruppe IV den höchsten Wert mit einer durchschnittlichen täglichen
Laktoseausscheidung von 487 g.
Der Einfluss des Laktationstages war zwischen dem 13./14. und 20./21. Laktationstag signifikant und
führte zu einer Steigerung der täglichen Laktoseabgabe um 40 g von 437 g auf 477 g. Die Werte der
einzelnen Sauen können aus Anhangstabelle 11 ersehen werden.

54
Tabelle 29: Laktoseausscheidung mit der Milch in g/Tag
Gruppe I II III IV V VI Trp.-Geh. in % 0,12 0,15 0,18 0,24 0,30 0,42 Mittel
13./14. Laktationstag 406 451 425 444 462 433 437B
±108 ±105 ±81 ±65 ±87 ±84 ±88
20./21. Laktationstag 395b 473ab 479ab 531a 509a 476ab 477A
±79 ±80 ±111 ±79 ±105 ±124 ±96 Mittel 401 462 452 487 485 454 457 ±85 ±86 ±81 ±57 ±81 ±99 ±82
3.5.5. Harnstoffgehalt und �ausscheidung
Wie aus der Tabelle 30 entnommen werden kann, waren bei den Harnstoffgehalten in der Sauenmilch
keine signifikanten Unterschiede festzustellen. Anzumerken ist jedoch, dass die Werte bis zur Gruppe
V kontinuierlich abfallend waren, um danach wieder anzusteigen. Insgesamt lag der Durchschnitt des
Harnstoffgehaltes über alle Gruppen bei 190 mg/l. Die Harnstoffgehalte der jeweiligen Sauen können
in der Anhangstabelle 11 nachgelesen werden.
Tabelle 30: Harnstoffgehalt der Sauenmilch in mg/l
Gruppe I II III IV V VI Trp.-Geh. in % 0,12 0,15 0,18 0,24 0,30 0,42 Mittel
20./21. Laktationstag 211 201 185 184 173 188 190 ±28 ±84 ±56 ±25 ±46 ±42 ±47
Bei den täglichen Harnstoffausscheidungen (siehe Tabelle 31) unterschieden sich die Gruppen nicht
signifikant voneinander. Der durchschnittliche Wert lag bei einer Milchharnstoffausscheidung von
1553 mg je Tag.
Tabelle 31: Harnstoffausscheidung mit der Sauenmilch in mg/Tag
Gruppe I II III IV V VI Trp.-Geh. in % 0,12 0,15 0,18 0,24 0,30 0,42 Mittel
20./21. Laktationstag 1443 1647 1547 1662 1491 1530 1553 ±190 ±681 ±468 ±226 ±397 ±342 ±384
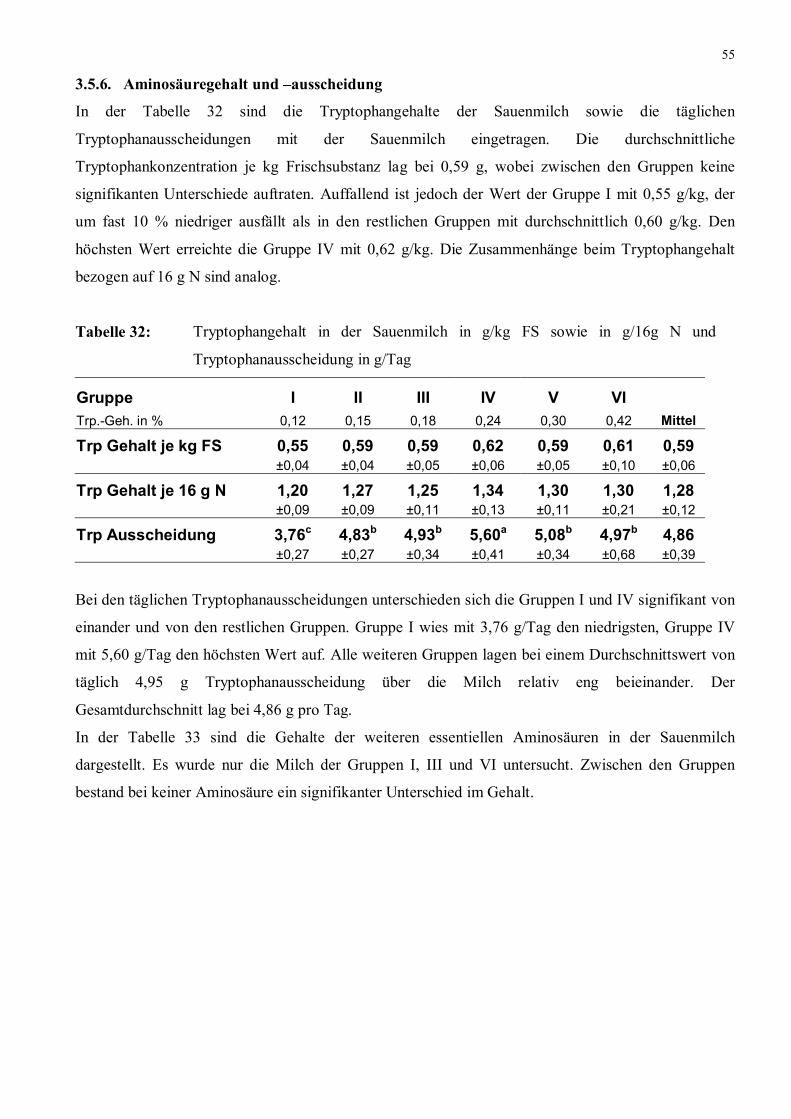
55
3.5.6. Aminosäuregehalt und �ausscheidung
In der Tabelle 32 sind die Tryptophangehalte der Sauenmilch sowie die täglichen
Tryptophanausscheidungen mit der Sauenmilch eingetragen. Die durchschnittliche
Tryptophankonzentration je kg Frischsubstanz lag bei 0,59 g, wobei zwischen den Gruppen keine
signifikanten Unterschiede auftraten. Auffallend ist jedoch der Wert der Gruppe I mit 0,55 g/kg, der
um fast 10 % niedriger ausfällt als in den restlichen Gruppen mit durchschnittlich 0,60 g/kg. Den
höchsten Wert erreichte die Gruppe IV mit 0,62 g/kg. Die Zusammenhänge beim Tryptophangehalt
bezogen auf 16 g N sind analog.
Tabelle 32: Tryptophangehalt in der Sauenmilch in g/kg FS sowie in g/16g N und
Tryptophanausscheidung in g/Tag
Gruppe I II III IV V VI Trp.-Geh. in % 0,12 0,15 0,18 0,24 0,30 0,42 Mittel Trp Gehalt je kg FS 0,55 0,59 0,59 0,62 0,59 0,61 0,59 ±0,04 ±0,04 ±0,05 ±0,06 ±0,05 ±0,10 ±0,06
Trp Gehalt je 16 g N 1,20 1,27 1,25 1,34 1,30 1,30 1,28 ±0,09 ±0,09 ±0,11 ±0,13 ±0,11 ±0,21 ±0,12
Trp Ausscheidung 3,76c 4,83b 4,93b 5,60a 5,08b 4,97b 4,86 ±0,27 ±0,27 ±0,34 ±0,41 ±0,34 ±0,68 ±0,39
Bei den täglichen Tryptophanausscheidungen unterschieden sich die Gruppen I und IV signifikant von
einander und von den restlichen Gruppen. Gruppe I wies mit 3,76 g/Tag den niedrigsten, Gruppe IV
mit 5,60 g/Tag den höchsten Wert auf. Alle weiteren Gruppen lagen bei einem Durchschnittswert von
täglich 4,95 g Tryptophanausscheidung über die Milch relativ eng beieinander. Der
Gesamtdurchschnitt lag bei 4,86 g pro Tag.
In der Tabelle 33 sind die Gehalte der weiteren essentiellen Aminosäuren in der Sauenmilch
dargestellt. Es wurde nur die Milch der Gruppen I, III und VI untersucht. Zwischen den Gruppen
bestand bei keiner Aminosäure ein signifikanter Unterschied im Gehalt.

56
Tabelle 33: Gehalte an essentiellen Aminosäuren in der Milch der Gruppen I, III und VI
Gruppe I III VI I III VI Trp. % 0,12 0,18 0,42 Mittel 0,12 0,18 0,42 Mittel in g/kg Milch FS in g/16g N
Lys 3,37 3,47 3,43 3,42 7,34 7,37 7,33 7,35 ±0,33 ±0,21 ±0,37 ±0,30 ±0,72 ±0,45 ±0,79 ±0,65
Met 0,88 0,88 0,88 0,88 1,92 1,87 1,88 1,89 ±0,09 ±0,06 ±0,09 ±0,08 ±0,20 ±0,13 ±0,19 ±0,17
Cys 0,53 0,56 0,59 0,56 1,15 1,19 1,26 1,20 ±0,04 ±0,05 ±v0,08 ±0,06 ±0,09 ±0,11 ±0,17 ±0,12
Thr 1,82 1,86 1,89 1,86 3,96 3,95 4,04 3,98 ±0,14 ±0,11 ±0,22 ±0,16 ±0,30 ±0,23 ±0,47 ±0,34
Leu 3,83 3,83 3,92 3,86 8,34 8,13 8,37 8,28 ±0,29 ±v0,24 ±0,44 ±0,32 ±0,63 ±0,51 ±0,94 ±0,69
Ile 1,87 1,92 1,92 1,90 4,07 4,08 4,10 4,08 ±0,20 ±0,11 ±0,20 ±0,17 ±0,44 ±0,23 ±0,43 ±0,37
Val 2,31 2,35 2,38 2,35 5,03 4,99 5,08 5,04 ±0,19 ±0,14 ±0,28 ±0,20 ±0,41 ±0,30 ±0,60 ±0,44
Phe 1,85 1,89 1,90 1,88 4,03 4,01 4,06 4,03 ±0,15 ±0,10 ±0,22 ±0,16 ±0,33 ±0,21 ±0,47 ±0,34
Tyr 1,72 1,82 1,79 1,78 3,75 3,86 3,82 3,81 ±0,19 ±0,11 ±0,23 ±0,18 ±0,41 ±0,23 ±0,49 ±0,38
His 1,22 1,25 1,26 1,24 2,66 2,65 2,69 2,67 ±0,11 ±0,08 ±0,13 ±0,11 ±0,24 ±0,17 ±0,28 ±0,23
Auch bei den nicht essentiellen Aminosäurengehalten in der Sauenmilch, die in Tabelle 34
zusammengefasst sind, bestanden keine signifikanten Unterschiede zwischen den Gruppen.
Tabelle 34: Gehalte an nicht essentiellen Aminosäuren in der Milch der Gruppen I, III und VI
Gruppe I III VI I III VI Trp. % 0,12 0,18 0,42 Mittel 0,12 0,18 0,42 Mittel in g/kg Milch FS in g/16g N
Ala 1,58 1,64 1,67 1,63 3,44 3,48 3,57 3,50 ±0,11 ±0,10 ±0,21 ±0,14 ±0,24 ±0,21 ±0,45 ±0,30
Arg 2,10 2,16 2,19 2,15 4,57 4,59 4,68 4,61 ±0,17 ±0,11 ±0,27 ±0,18 ±0,37 ±0,23 ±0,58 ±0,39
Asn 3,63 3,75 3,81 3,73 7,91 7,96 8,14 8,00 ±0,28 ±0,21 ±0,43 ±0,31 ±0,61 ±0,45 ±0,92 ±0,66
Gln 9,67 9,68 9,55 9,63 21,06 20,55 20,40 20,67 ±0,90 ±0,54 ±0,85 ±0,76 ±1,96 ±1,15 ±1,82 ±1,64
Gly 1,50 1,53 1,53 1,52 3,27 3,25 3,27 3,26 ±0,13 ±0,12 ±0,19 ±0,15 ±0,28 ±0,25 ±0,41 ±0,31
Ser 2,28 2,35 2,35 2,33 4,97 4,99 5,02 4,99 ±0,20 ±0,12 ±0,26 ±0,19 ±0,44 ±0,25 ±0,56 ±0,42

57
Die tierindividuellen Werte der Aminosäurenkonzentrationen in der Milch können den
Anhangstabellen 15 und 16 entnommen werden.
In der nachfolgenden Tabelle 35 sind die Werte für die täglichen Aminosäurenausscheidungen mit der
Sauenmilch aufgeführt. Bei allen untersuchten Aminosäuren war ein Einfluss des Tryptophangehaltes
im Futter auf die Menge der über die Milch ausgeschiedenen Aminosäuren-Mengen feststellbar. So
waren die Werte der Basisgruppe immer signifikant geringer als die der Gruppen III und VI, zwischen
denen sich keine Unterschiede zeigten. Mit Ausnahme von Cystin hatte die Gruppe III immer die
höchste Ausscheidung.
Tabelle 35 Aminosäurenausscheidung mit der Milch in g/Tag
Gruppe I III VI Tryptophangehalt in % 0,12 0,18 0,42 Mittel essentielle AS Lys 23,1b 29,0a 27,9 a 26,7 ±2,5 ±1,6 ±2,9 ±2,3 Met 6,0b 7,4a 7,2a 6,9 ±0,7 ±0,5 ±0,7 ±0,6 Cys 3,6b 4,7a 4,8a 4,4 ±0,3 ±0,4 ±0,6 ±0,4 Thr 12,5b 15,5a 15,4a 14,5 ±1,1 ±0,9 ±1,7 ±1,2 Leu 26,2b 32,0a 31,9a 30,0 ±2,2 ±v1,9 ±3,4 ±2,5 Ile 12,8b 16,1a 15,7a 14,9 ±1,5 ±0,9 ±1,6 ±1,3 Val 15,8b 19,6a 19,4a 18,3 ±1,5 ±1,1 ±2,2 ±1,6 Phe 12,6b 15,8a 15,5a 14,6 ±1,2 ±0,8 ±1,7 ±1,2 Tyr 11,8b 15,2a 14,5a 13,8 ±1,5 ±0,9 ±1,8 ±1,4 His 8,3b 10,4a 10,2a 9,6 ±0,8 ±0,6 ±1,0 ±0,8 nicht essent. AS Ala 10,8b 13,7a 13,6a 12,7 ±0,8 ±0,8 ±1,6 ±1,1 Arg 14,4b 18,1a 17,8a 16,8 ±1,3 ±0,9 ±2,1 ±1,4 Asn 24,8b 31,3a 31,0a 29,0 ±2,2 ±1,6 ±v3,3 ±2,4 Gln 66,2b 81,0a 77,7a 75,0 ±6,9 ±4,2 ±6,6 ±5,9 Gly 10,2b 12,8a 12,5a 11,9 ±1,0 ±0,9 ±1,5 ±1,1 Ser 15,6b 19,7a 19,2a 18,2 ±1,5 ±0,9 ±2,0 ±1,5
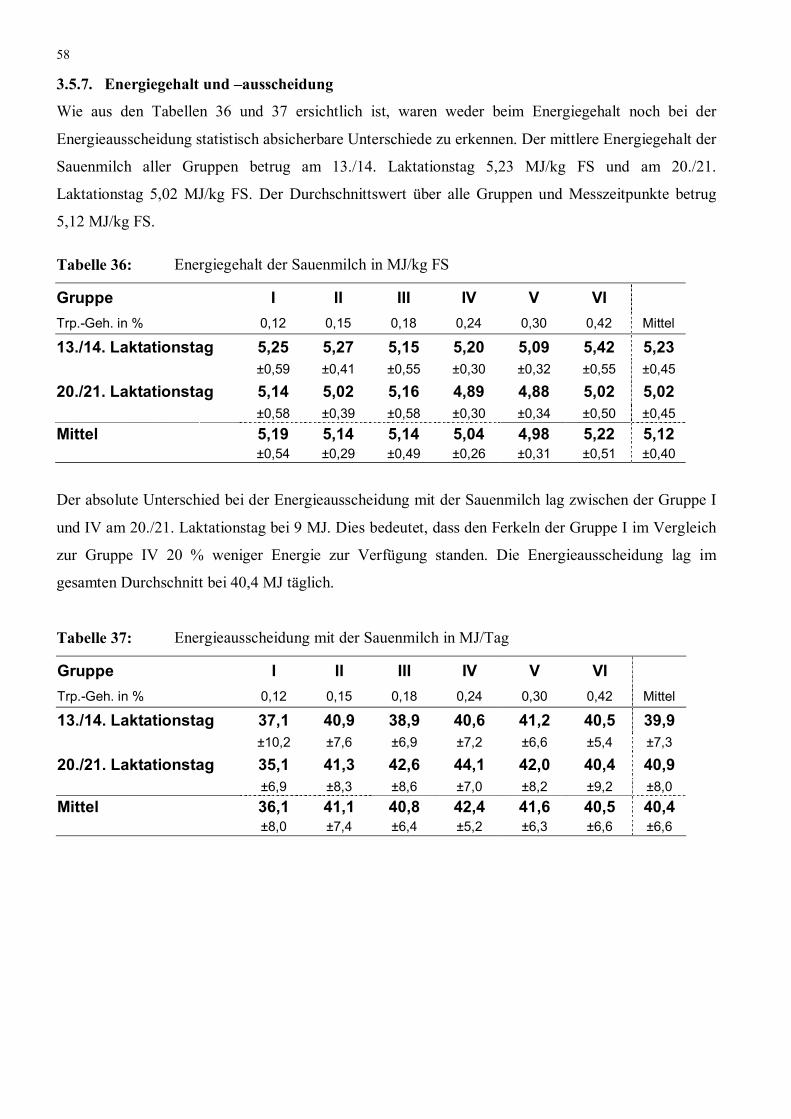
58
3.5.7. Energiegehalt und �ausscheidung
Wie aus den Tabellen 36 und 37 ersichtlich ist, waren weder beim Energiegehalt noch bei der
Energieausscheidung statistisch absicherbare Unterschiede zu erkennen. Der mittlere Energiegehalt der
Sauenmilch aller Gruppen betrug am 13./14. Laktationstag 5,23 MJ/kg FS und am 20./21.
Laktationstag 5,02 MJ/kg FS. Der Durchschnittswert über alle Gruppen und Messzeitpunkte betrug
5,12 MJ/kg FS. Tabelle 36: Energiegehalt der Sauenmilch in MJ/kg FS
Gruppe I II III IV V VI Trp.-Geh. in % 0,12 0,15 0,18 0,24 0,30 0,42 Mittel
13./14. Laktationstag 5,25 5,27 5,15 5,20 5,09 5,42 5,23 ±0,59 ±0,41 ±0,55 ±0,30 ±0,32 ±0,55 ±0,45
20./21. Laktationstag 5,14 5,02 5,16 4,89 4,88 5,02 5,02 ±0,58 ±0,39 ±0,58 ±0,30 ±0,34 ±0,50 ±0,45 Mittel 5,19 5,14 5,14 5,04 4,98 5,22 5,12
±0,54 ±0,29 ±0,49 ±0,26 ±0,31 ±0,51 ±0,40
Der absolute Unterschied bei der Energieausscheidung mit der Sauenmilch lag zwischen der Gruppe I
und IV am 20./21. Laktationstag bei 9 MJ. Dies bedeutet, dass den Ferkeln der Gruppe I im Vergleich
zur Gruppe IV 20 % weniger Energie zur Verfügung standen. Die Energieausscheidung lag im
gesamten Durchschnitt bei 40,4 MJ täglich.
Tabelle 37: Energieausscheidung mit der Sauenmilch in MJ/Tag
Gruppe I II III IV V VI Trp.-Geh. in % 0,12 0,15 0,18 0,24 0,30 0,42 Mittel
13./14. Laktationstag 37,1 40,9 38,9 40,6 41,2 40,5 39,9 ±10,2 ±7,6 ±6,9 ±7,2 ±6,6 ±5,4 ±7,3
20./21. Laktationstag 35,1 41,3 42,6 44,1 42,0 40,4 40,9 ±6,9 ±8,3 ±8,6 ±7,0 ±8,2 ±9,2 ±8,0 Mittel 36,1 41,1 40,8 42,4 41,6 40,5 40,4 ±8,0 ±7,4 ±6,4 ±5,2 ±6,3 ±6,6 ±6,6

59
3.6. Blutparameter
3.6.1 Aminosäurengehalt
3.6.1.1. Essentielle und semiessentielle Aminosäuren
In Tabelle 38 sind die durchschnittlichen Konzentrationen an essentiellen und semiessentiellen
Aminosäuren im Blutplasma der verschiedenen Gruppen dargestellt. Mit Ausnahme von Tryptophan,
Methionin und Valin waren keine signifikanten Unterschiede zwischen den verschiedenen
Futtergruppen feststellbar. Mit ansteigender Tryptophanzulage war ein gerichteter Einfluss auf die
Tryptophangehalte im Plasma zu erkennen. Die Gruppe VI hatte mit 23,91 mg/l fast die 9-fache
Konzentration der Gruppe I, die mit 2,74 mg/l den niedrigsten Wert aufwies. Die restlichen Werte
stiegen zwischen der Gruppe I und VI kontinuierlich an.
Tabelle 38: Gehalt an essentiellen und semiessentiellen Aminosäuren im Plasma in mg/l
Gruppe I II III IV V VI Trp.-Geh. in % 0,12 0,15 0,18 0,24 0,30 0,42 Mittel
Trp 2,74d 5,51cd 8,67c 15,54b 15,73b 23,91a 12,02 ±1,14 ±2,66 ±2,95 ±3,39 ±4,97 ±7,53 ±3,83
Lys 42,98 49,93 42,93 49,83 37,75 35,74 43,19 ±17,53 ±17,18 ±13,56 ±12,55 ±6,34 ±14,82 ±13,66
Met 13,37b 21,76a 23,43a 24,13a 22,10a 18,67a 20,58 ±3,58 ±6,76 ±4,28 ±6,03 ±4,08 ±5,81 ±5,09
Cys 3,14 1,61 1,91 1,86 1,34 1,77 1,94 ±2,49 ±1,12 ±1,49 ±0,53 ±0,47 ±1,48 ±1,26
Thr 40,58 41,18 34,98 35,67 29,87 31,79 35,68 ±7,61 ±12,38 ±7,14 ±6,66 ±5,67 ±7,66 ±7,85
Leu 22,39 24,76 26,37 29,10 21,17 25,06 24,81 ±5,95 ±7,71 ±5,78 ±6,43 ±4,33 ±5,56 ±5,96
Ile 14,90 15,64 14,64 15,91 10,58 13,78 14,24 ±4,18 ±5,19 ±3,02 ±3,51 ±2,45 ±3,47 ±3,64
Val 23,40ab 23,21ab 21,18ab 24,19a 14,92b 17,74ab 20,77 ±7,22 ±8,53 ±6,50 ±7,40 ±2,28 ±7,26 ±6,53
Phe 17,88 22,33 24,92 23,90 20,64 19,48 21,52 ±5,05 ±10,55 ±4,17 ±4,58 ±2,73 ±5,01 ±5,35
Tyr 17,84 19,59 19,03 22,29 16,70 17,82 18,88 ±4,96 ±4,90 ±3,27 ±5,48 ±2,49 ±4,10 ±4,20
His 17,46 18,94 18,09 17,79 17,18 16,94 17,73 ±2,23 ±2,65 ±1,94 ±3,37 ±1,99 ±2,58 ±2,46
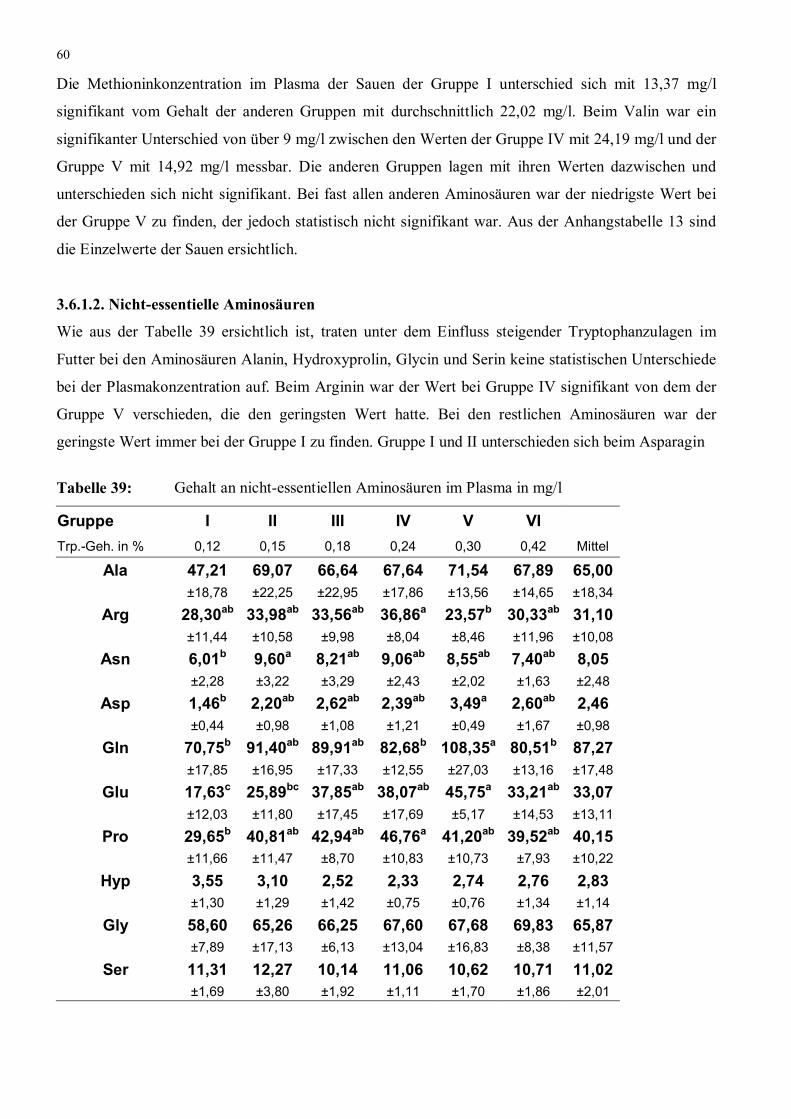
60
Die Methioninkonzentration im Plasma der Sauen der Gruppe I unterschied sich mit 13,37 mg/l
signifikant vom Gehalt der anderen Gruppen mit durchschnittlich 22,02 mg/l. Beim Valin war ein
signifikanter Unterschied von über 9 mg/l zwischen den Werten der Gruppe IV mit 24,19 mg/l und der
Gruppe V mit 14,92 mg/l messbar. Die anderen Gruppen lagen mit ihren Werten dazwischen und
unterschieden sich nicht signifikant. Bei fast allen anderen Aminosäuren war der niedrigste Wert bei
der Gruppe V zu finden, der jedoch statistisch nicht signifikant war. Aus der Anhangstabelle 13 sind
die Einzelwerte der Sauen ersichtlich.
3.6.1.2. Nicht-essentielle Aminosäuren
Wie aus der Tabelle 39 ersichtlich ist, traten unter dem Einfluss steigender Tryptophanzulagen im
Futter bei den Aminosäuren Alanin, Hydroxyprolin, Glycin und Serin keine statistischen Unterschiede
bei der Plasmakonzentration auf. Beim Arginin war der Wert bei Gruppe IV signifikant von dem der
Gruppe V verschieden, die den geringsten Wert hatte. Bei den restlichen Aminosäuren war der
geringste Wert immer bei der Gruppe I zu finden. Gruppe I und II unterschieden sich beim Asparagin
Tabelle 39: Gehalt an nicht-essentiellen Aminosäuren im Plasma in mg/l
Gruppe I II III IV V VI Trp.-Geh. in % 0,12 0,15 0,18 0,24 0,30 0,42 Mittel
Ala 47,21 69,07 66,64 67,64 71,54 67,89 65,00 ±18,78 ±22,25 ±22,95 ±17,86 ±13,56 ±14,65 ±18,34
Arg 28,30ab 33,98ab 33,56ab 36,86a 23,57b 30,33ab 31,10 ±11,44 ±10,58 ±9,98 ±8,04 ±8,46 ±11,96 ±10,08
Asn 6,01b 9,60a 8,21ab 9,06ab 8,55ab 7,40ab 8,05 ±2,28 ±3,22 ±3,29 ±2,43 ±2,02 ±1,63 ±2,48
Asp 1,46b 2,20ab 2,62ab 2,39ab 3,49a 2,60ab 2,46 ±0,44 ±0,98 ±1,08 ±1,21 ±0,49 ±1,67 ±0,98
Gln 70,75b 91,40ab 89,91ab 82,68b 108,35a 80,51b 87,27 ±17,85 ±16,95 ±17,33 ±12,55 ±27,03 ±13,16 ±17,48
Glu 17,63c 25,89bc 37,85ab 38,07ab 45,75a 33,21ab 33,07 ±12,03 ±11,80 ±17,45 ±17,69 ±5,17 ±14,53 ±13,11
Pro 29,65b 40,81ab 42,94ab 46,76a 41,20ab 39,52ab 40,15 ±11,66 ±11,47 ±8,70 ±10,83 ±10,73 ±7,93 ±10,22
Hyp 3,55 3,10 2,52 2,33 2,74 2,76 2,83 ±1,30 ±1,29 ±1,42 ±0,75 ±0,76 ±1,34 ±1,14
Gly 58,60 65,26 66,25 67,60 67,68 69,83 65,87 ±7,89 ±17,13 ±6,13 ±13,04 ±16,83 ±8,38 ±11,57
Ser 11,31 12,27 10,14 11,06 10,62 10,71 11,02 ±1,69 ±3,80 ±1,92 ±1,11 ±1,70 ±1,86 ±2,01

61
sowie I und V bei der Asparaginsäure bedeutsam. Den höchsten Gehalt an Glutamin im Plasma hatte
die Gruppe V. Sie unterschied sich signifikant von den Gruppen II, IV und VI. Die Tiere der
Behandlung V wiesen beim Glutaminsäuregehalt den höchsten Wert auf, der sich deutlich vom Wert
der Gruppen II und I absetzte. Die Gruppe II lag zwischen den Gruppen I und III mit IV. Beim Prolin
wies die Gruppe IV mit 47 mg/l den höchsten Wert auf, der sich von dem der Gruppe I mit nur 30 mg/l
signifikant unterschied. Die Werte der restlichen Gruppen lagen mit durchschnittlich 41 mg/l
dazwischen.Aus der Anhangstabelle 14 gehen die Einzelwerte der jeweiligen Tiere hervor.
3.6.2. Harnstoffgehalt
Wie aus Tabelle 40 hervorgeht, traten bei der Harnstoffkonzentration im Plasma zwischen den
Gruppen keine nachweisbaren Unterschiede auf. Den höchsten Wert erreichte Gruppe I mit einer
Konzentraion von 272 mg/l und den niedrigsten Gruppe V mit 213 mg/l. Die restlichen Werte
bewegten sich im Bereich um 240 mg/l.
Tabelle 40: Harnstoffgehalt im Plasma in mg/l
Gruppe I II III IV V VI Trp.-Geh. in % 0,12 0,15 0,18 0,24 0,30 0,42 Mittel
Harnstoff 272 243 231 229 213 235 240 ±52 ±55 ±53 ±48 ±23 ±47 ±53
Die Einzelergebnisse der Harnstoffuntersuchung im Blutplasma der Sauen sind aus Anhangstabelle 14
zu ersehen.
3.6.3. Hämatokritwert
Wie in der Tabelle 41 dargestellt ist, waren beim Hämatokritwert zwischen den Gruppen keine
signifikanten Unterschiede festzustellen. Jedoch hatte der Zeitpunkt der Probenahme einen Einfluss
auf die Höhe des Hämatokritwertes. So war eine deutliche Abnahme vom 85. Trächtigkeitstag mit Tabelle 41: Hämatokritwert in % und Veränderung in %
Gruppe I II III IV V VI Trp.-Geh. in % 0,12 0,15 0,18 0,24 0,30 0,42 Mittel 85. TT 37,9 36,5 37,2 33,6 37,0 37,3 36,6A
±4,5 ±5,1 ±4,0 ±4,2 ±4,8 ±3,1 ±4,3 28. LT 34,8 34,0 34,2 32,8 32,0 32,5 33,4B
±4,1 ±3,3 ±2,8 ±2,4 ±1,8 ±2,3 ±2,8 Veränderung in % -7,2 -5,0 -7,5 -0,9 -12,7 -12,3 -7,6 ±13,7 ±18,1 ±8,9 ±15,1 ±9,4 ±8,5 ±12,3

62
36,6 % zum 28. Laktationstag mit 33,4 % von über 3 % festzustellen. Am 85. Trächtigkeitstag und 28.
Laktationstag traten zwischen den Gruppen keine signifikanten Veränderungen auf. Die
durchschnittliche Veränderung zwischen den beiden Messzeitpunkten lag relativ bei �7,6 %. Die
Werte der einzelnen Sauen können in der Anhangstabelle 9 nachgelesen werden.
3.6.4. Thrombozytengehalt
Bei der Untersuchung des Sauenblutes hinsichtlich der Thrombozytenzahl (TZ; siehe Tabelle 42) war
am 85. Trächtigkeitstag ein signifikanter Unterschied zwischen der Gruppe II mit 109 TZ�109/l und der
Gruppe IV mit 160 � 109 TZ/l messbar. Die Werte der anderen Gruppen lagen dazwischen. Am 28.
Laktationstag traten keine Unterschiede zwischen den Gruppen auf. Signifikant war der Einfluss des
Messtages. Mit 196 � 109 TZ/l war die mittlere Konzentration im Blut am 28. Laktationstag mehr als
1,5-mal so groß wie am 85. Trächtigkeitstag mit 126 � 109 TZ/l. Eine signifikante Veränderung der
Thrombozytenzahl zwischen den Gruppen war nicht erkennbar. Insgesamt war ein Anstieg der
Thrombozytenzahl um über 70 % zu beobachten. Die Einzelwerte der verschiedenen Gruppen sind aus
der Anhangstabelle 9 ersichtlich.
Tabelle 42: Thrombozyten in 109/l und Veränderung in %
Gruppe I II III IV V VI Trp.-Geh. in % 0,12 0,15 0,18 0,24 0,30 0,42 Mittel 85. TT 128ab 109b 118ab 160a 111ab 131ab 126B
±33 ±35 ±35 ±27 ±43 ±53 ±38 28. LT 180 200 186 214 182 212 196A
±50 ±59 ±58 ±81 ±19 ±64 ±55 Veränderung in % 43,8 104,3 62,1 36,6 85,7 90,2 70,5 ±31,3 ±89,9 ±46,9 ±57,3 ±74,1 ±98,8 ±66,4
3.6.5. Serotonin im Serum
In der Tabelle 43 sind die Serotoningehalte im Blutserum der Sauen eingetragen. Am 85.
Trächtigkeitstag war zwischen den Gruppen kein signifikanter Unterschied feststellbar. Bei der
Messung am 28. Laktationstag waren die Gruppen I und II mit durchschnittlich 267 ng/ml und die
restlichen Gruppen mit im Mittel 511 ng/ml deutlich verschieden. Zwischen den Messtagen trat keine
signifikante Differenz auf.
Die Werte für den Serotoningehalt im Serum sind bei der Gruppe I am 28. Laktationstag um 40 %
geringer als am 85. Trächtigkeitstag. Mit zunehmendem Tryptophangehalt ist diese negative
Veränderung jedoch weniger stark ausgebildet und kehrt sich ab der Gruppe IV in eine positive um.
Die Gruppen I und II unterscheiden sich dabei signifikant von den restlichen Gruppen. Der
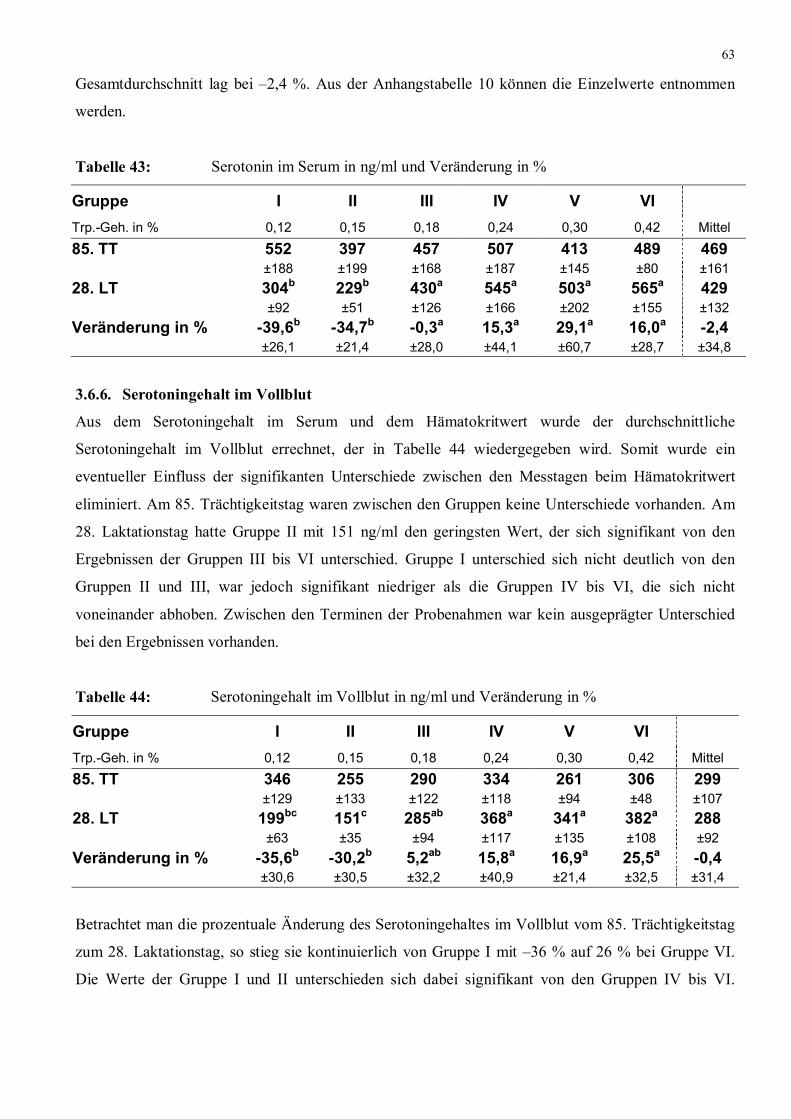
63
Gesamtdurchschnitt lag bei �2,4 %. Aus der Anhangstabelle 10 können die Einzelwerte entnommen
werden.
Tabelle 43: Serotonin im Serum in ng/ml und Veränderung in %
Gruppe I II III IV V VI Trp.-Geh. in % 0,12 0,15 0,18 0,24 0,30 0,42 Mittel 85. TT 552 397 457 507 413 489 469 ±188 ±199 ±168 ±187 ±145 ±80 ±161 28. LT 304b 229b 430a 545a 503a 565a 429 ±92 ±51 ±126 ±166 ±202 ±155 ±132 Veränderung in % -39,6b -34,7b -0,3a 15,3a 29,1a 16,0a -2,4 ±26,1 ±21,4 ±28,0 ±44,1 ±60,7 ±28,7 ±34,8
3.6.6. Serotoningehalt im Vollblut
Aus dem Serotoningehalt im Serum und dem Hämatokritwert wurde der durchschnittliche
Serotoningehalt im Vollblut errechnet, der in Tabelle 44 wiedergegeben wird. Somit wurde ein
eventueller Einfluss der signifikanten Unterschiede zwischen den Messtagen beim Hämatokritwert
eliminiert. Am 85. Trächtigkeitstag waren zwischen den Gruppen keine Unterschiede vorhanden. Am
28. Laktationstag hatte Gruppe II mit 151 ng/ml den geringsten Wert, der sich signifikant von den
Ergebnissen der Gruppen III bis VI unterschied. Gruppe I unterschied sich nicht deutlich von den
Gruppen II und III, war jedoch signifikant niedriger als die Gruppen IV bis VI, die sich nicht
voneinander abhoben. Zwischen den Terminen der Probenahmen war kein ausgeprägter Unterschied
bei den Ergebnissen vorhanden.
Tabelle 44: Serotoningehalt im Vollblut in ng/ml und Veränderung in %
Gruppe I II III IV V VI Trp.-Geh. in % 0,12 0,15 0,18 0,24 0,30 0,42 Mittel 85. TT 346 255 290 334 261 306 299 ±129 ±133 ±122 ±118 ±94 ±48 ±107 28. LT 199bc 151c 285ab 368a 341a 382a 288 ±63 ±35 ±94 ±117 ±135 ±108 ±92 Veränderung in % -35,6b -30,2b 5,2ab 15,8a 16,9a 25,5a -0,4 ±30,6 ±30,5 ±32,2 ±40,9 ±21,4 ±32,5 ±31,4
Betrachtet man die prozentuale Änderung des Serotoningehaltes im Vollblut vom 85. Trächtigkeitstag
zum 28. Laktationstag, so stieg sie kontinuierlich von Gruppe I mit �36 % auf 26 % bei Gruppe VI.
Die Werte der Gruppe I und II unterschieden sich dabei signifikant von den Gruppen IV bis VI.

64
Gruppe III lag dazwischen. Die entsprechenden Einzelwerte können der Anhangstabelle 10
entnommen werden.
3.6.7. Serotoningehalt je 109 Thrombozyten
Nach OCHSENKÜHN (1988) ist ein Großteil des im Blut befindlichen Serotonins an die
Thrombozyten gebunden. Somit kann eine Konzentrationsänderung beim freien Serotonin durch
thrombozytäres Serotonin ausgeglichen werden. Um diesen Einfluss auf den Gehalt an Serotonin
herauszurechnen, sind in Tabelle 45 die aus dem Hämatokrit-, dem Thrombozyten- und dem
Serotoninwert im Serum der Sauen errechneten Werte für den Serotoningehalt je 109 Trombozyten für
den 85. TT und den 28. LT eingetragen. Beim ersten Messzeitpunkt traten zwischen den Gruppen
keine signifikanten Unterschiede auf. Signifikante Differenzen waren am 28. Laktationstag zwischen
der Gruppe II und den Gruppen III bis VI zu erkennen. Der Wert der Gruppe I unterschied sich nicht
signifikant von den Werten der restlichen Gruppen. Der Gesamtdurchschnitt lag hier bei 1606
ng/109)TZ, der signifikant niedriger war, als der Wert am 85. Trächtigkeitstag mit 2631 ng/109 TZ.
Tabelle 45: Serotoningehalte in ng je 109 Thrombozyten und Veränderung in %
Gruppe I II III IV V VI Trp.-Geh. in % 0,12 0,15 0,18 0,24 0,30 0,42 Mittel Serotonin 85. TT 2930 2462 2789 2161 2531 2910 2631A
±554 ±1488 ±940 ±976 ±840 ±1814 ±1102 Serotonin 28. LT 1158ab 805b 1791a 2064a 1945a 1870a 1606B
±281 ±279 ±411 ±1666 ±403 ±429 ±578 Veränderung in % -54,7b -63,2b -32,6ab -6,1a -16,0a -21,0a -32,3 ±17,5 ±11,9 ±23,2 ±37,4 ±40,1 ±35,4 ±27,6
Bei den Gruppen I und II nahm die Serotoninkonzentration am 28. Laktationstag im Durchschnitt um
59 % des Ausgangswertes am 85. Trächtigkeitstag ab. Den geringsten Abfall beim Serotoningehalt
zeigt die Gruppe IV mit nur 6 %. Danach nahmen die Verringerungen der Konzentration wieder bis
auf 21 % bei Gruppe VI zu. Die Einzelergebnisse sind in der Anhangstabelle)8 aufgeführt.
3.7. Futterverbrauch der Ferkel In Tabelle 46 ist der Futterverbrauch der Ferkel in der vierten Laktationswoche dargestellt. Er betrug
im Mittel aller Würfe 800 g. Zwischen den Gruppen waren keine signifikanten Unterschiede
erkennbar, was durch die große Streuung zu erklären war, die teilweise größer als der Mittelwert selbst
war. Dies lässt auf einen starken Individualeffekt des einzelnen Wurfes schließen.

65
Tabelle 46: Saugferkelbeifutterverbrauch je Wurf in g
Gruppe I II III IV V VI Trp.-Geh. in % 0,12 0,15 0,18 0,24 0,30 0,42 Mittel
Beifutterverbrauch 703 708 941 947 624 565 798 ±727 ±837 ±636 ±822 ±565 ±474 ±252
In der Anhangstabelle 17 sind die einzelnen Mengen je Wurf dargestellt.
3.8. Lebendmasse und -entwicklung der Ferkel Die am 1., 14., 21. und 28. Lebenstag erfassten Massen der Ferkel sind als Durchschnittswerte in der
Tabelle 47 zusammengefasst.
Tabelle 47: Lebendmassen der Ferkel in g
Gruppe I II III IV V VI Trp.-Geh. in % 0,12 0,15 0,18 0,24 0,30 0,42 Mittel
1. Lebenstag 1473 1436 1577 1422 1502 1510 1487 ±164 ±196 ±190 ±153 ±206 ±171 ±180
14. Lebenstag 3563 3774 3810 3567 3948 3736 3733 ±501 ±576 ±701 ±247 ±358 ±445 ±471
21. Lebenstag 4732 5124 5236 5138 5217 5031 5080 ±621 ±738 ±818 ±380 ±470 ±608 ±606
28. Lebenstag 5854 6596 6704 6706 6663 6460 6497 ±628 ±868 ±1019 ±470 ±639 ±827 ±742
Die mittlere Geburtsmasse betrug 1490 g, wobei zwischen den einzelnen Gruppen keine Unterschiede
auftraten. Die Durchschnittsmasse der Ferkel betrug am 14. Tag 3730 g, am 21. Tag 5080 g und beim
Absetzen am 28. Tag 6497 g. Auch zu diesen Wiegeterminen traten keine signifikanten Unterschiede
zwischen den Gruppen auf, allerdings waren die Ferkel der Gruppe I mit 5854 g um über 850 g
beziehungsweise 13 % leichter als die Tiere der Gruppe IV mit 6706 g. Die Lebendmassen der
Einzeltiere sind in der Anhangstabelle 12 enthalten.
Bei den Lebendmassezunahmen, wie sie in Tabelle 48 ersichtlich sind, traten in den ersten zwei
Lebenswochen der Ferkel keine Unterschiede auf. Das Zunahmeniveau lag bei durchschnittlich 160
g/Tag. In der dritten Lebenswoche nahmen die Ferkel der Gruppe III mit 210 g/Tag signifikant besser
zu als die Ferkel der Gruppe I mit 166 g/Tag. Die Zunahmen der restlichen Gruppen lagen mit
durchschnittlich 198 g/Tag relativ eng beieinander. Zwischen den Gruppen II beziehungsweise III und
I bestand in der vierten Woche ein statistisch abgesicherter Unterschied. Die Tageszunahmen von

66
Gruppe II und III lagen im Mittel bei 220 g/Tag während es bei Gruppe I lediglich 170 g/Tag waren.
Die Gruppen II, V und VI lagen mit einem Durchschnittswert von 200 g/Tag dazwischen.
Tabelle 48: Lebendmassezunahmen der Ferkel in g/Tag
Gruppe I II III IV V VI Trp.-Geh. in % 0,12 0,15 0,18 0,24 0,30 0,42 Mittel
1. - 14. Lebenstag 147 167 159 156 171 159 160 ±28 ±28 ±38 ±15 ±18 ±29 ±26
15. - 21. Lebenstag 166b 196ab 204ab 210a 198ab 193ab 195 ±33 ±37 ±35 ±25 ±34 ±48 ±35
22. - 28. Lebenstag 168b 208ab 216a 224a 203ab 197ab 203 ±43 ±32 ±53 ±27 ±40 ±31 ±38
1. - 21. Lebenstag 153 176 174 175 179 170 171 ±26 ±27 ±31 ±17 ±18 ±31 ±25
1. - 28. Lebenstag 157 184 184 187 185 177 179 ±26 ±26 ±33 ±15 ±21 ±27 ±25
Bei den Zunahmen vom 1. bis 21. beziehungsweise 1. bis 28. Laktationstag traten keine statistischen
Unterschiede auf. Jedoch ist zu erkennen, dass sich die Werte der Gruppe I mit 153 g/Tag und 157
g/Tag (entspricht �13 % beziehungsweise -14 %) relativ stark vom Mittel der anderen Gruppen mit
175 g/Tag und 183 g/Tag unterschieden.

67
4. Diskussion Mit der vorliegenden Arbeit sollte der Einfluss einer im Tryptophangehalt variierenden Fütterung auf
Leistungs- und Stoffwechselveränderungen untersucht werden. Ausgehend von den Ergebnissen sollte
die Ableitung eines Wertes für den Tryptophanbedarf laktierender Zuchtsauen erstellt werden.
Als Bedarf wird die Minimalzufuhr an Tryptophan bezeichnet, die notwendig ist, um das Maximum
einer bestimmten Leistung inklusive der Erhaltung des Tieres zu erreichen. Zur Ermittlung des
Tryptophanbedarfs laktierender Zuchtsauen kommt grundsätzlich die Untersuchung der in Tabelle 49
dargestellten Parameter in Frage.
Tabelle 49: Grundsätzliche Parameter zur Ermittlung des Tryptophanbedarfs laktierender
Zuchtsauen
Parameter
für die Leistung
Parameter
des N-Stoffwechsels
Parameter
des Aminosäurenstoffwechsels
- Milchleistung
- Futteraufnahme der Sau
- Masseverlust der Sau in der
Laktation
- Ferkelzuwachs
- Anzahl aufgezogener Ferkel
- Absetzgewicht der Ferkel
- Beifutteraufnahme der
Ferkel
- Konzeptionsrate
- Leertage
- N-Bilanz
- Harnstoffauscheidung über
Urin und Milch
- Harnstoffgehalt in
Blutplasma und Milch
- Aminosäurengehalte in
Blutplasma und Milch
- Freie Aminosäuren im
Blutplasma
- Serotoningehalt im
Blutserum
- Niacingehalt im Blutplasma
Die Sauenmilch stellt für Ferkel in den ersten Lebenswochen die einzige Nahrungsquelle dar. Aus
diesem Grund sind Milchmenge und -inhaltsstoffe die entscheidenden Parameter für hohe Zunahmen
und Absetzgewichte sowie für geringe Verluste bei den Ferkeln. Um die Auswirkungen einer
variierenden Tryptophanversorgung zu ermitteln, wurden die Milchleistung und �inhaltsstoffe sowie
die Ferkelzunahmen erfasst. Da die verzehrte Menge an Saugferkelbeifutter indirekt Auskunft über die
Milchversorgung der Ferkel geben kann, wurde auch diese ermittelt. Die Ermittlung der
Lebendmasseverluste der Sauen wurde durchgeführt, da nach Untersuchungen von BLACK et al.
(1986), KING (1987) und JOHNSTON et al. (1989) hohe Masseverluste während der Laktation
negativ mit Fruchtbarkeitsparametern korreliert sind und somit eine indirekte Aussage möglich

68
erschien. Ebenso scheint dies nach HENRY et al. (1984), HUGHES et al. (1984) und KIRKWOOD et
al. (1988) anhand der Futteraufnahme während der Säugezeit möglich zu sein, da diese mit
Reproduktionskennzahlen korrelieren zu scheint. Nach MEISINGER und SPEER (1979) korrelieren
der Tryptophangehalt in der Ration mit der Futteraufnahme, weshalb die Futteraufnahme der Sauen
erfasst wurde. Die Aminosäuren- und Harnstoffkonzentrationen im Blut wurden ermittelt, da sie in
engem Zusammenhang mit der Aminosäurenversorgung über das Futter stehen (LEWIS und SPEER,
1973; 1974; 1975; ROUSSELOW und SPEER, 1980; KIRCHGESSNER et al., 1993; 1998; RICHERT
et al., 1996; OTT, 2000). Auch die Aminosäuren- und Harnstoffgehalte in der Milch wurden
gemessen, da diese durch eine veränderte Protein- beziehungsweise Aminoäurenversorgung der Sau
beeinflusst werden können (LEWIS und SPEER, 1973; ROUSSELOW et al., 1979; ROUSSELOW
und SPEER, 1980; WESTERMEIER, 1997; OTT, 2000).

69
4.1. Modelle zur Bedarfsableitung Bei der Bedarfsableitung aufgrund von Dosis-Wirkungs-Beziehungen wird die Wirkung eines
geeigneten Kriteriums in Abhängigkeit von der Dosis, die sinnvollerweise den Versorgungsbereich
suboptimal bis subtoxisch erfassen sollte, untersucht (KIRCHGESSNER, 1987). Da beim
vorliegenden Versuch eine definierte Grundration mit ansteigender Tryptophansupplementierung vom
Mangel bis zur Überversorgung eingesetzt wurde, lässt sich der Bedarf aus der Dosis-Wirkungs-
Beziehung ableiten. Neuere Untersuchungen haben bestätigt, dass mit einer zunehmenden
Aminosäurenversorgung kein unbegrenzt linearer Anstieg der Tierleistungen zu erwarten ist (ZHANG,
1991; SCHNEIDER, 1992; MÖHN, 1993; WESTERMEIER, 1997; OTT, 2000). Bei Erreichen einer
bestimmten Aminosäurenkonzentration im Futter kommt es zu einer Verflachung der Dosis-Wirkungs-
Kurve, der sogenannten Plateaubildung. Zur mathematischen Berechnung der Dosis-Wirkungs-Kurve
beziehungsweise zur Bedarfsableitung von Aminosäuren sind in der Literatur verschiedene Modelle
beschrieben (ROUSSELOW und SPEER, 1980; LELLIS und SPEER, 1987; SUSENBETH, 2002):
- Broken-Line Modell
- Reading Modell
- Quadratisches Modell
- Asymptotisches Modell
Die Berechnung wurde für alle Modelle mit dem Programmpaket Statistical Analysis System, Version
8.2 (SAS Institute Inc., 1999, Cary, North Carolina, USA) durchgeführt.
4.1.1. Abschnittsweise definierte Modelle
Wenn davon ausgegangen wird, dass die Tierleistungen mit zunehmendem Aminosäureneinsatz im
Futter nicht unbegrenzt ansteigen, kann die Bedarfsableitung bei einer Plateaubildung gut mit
abschnittsweise definierten Bedarfskurven vorgenommen werden. Dazu werden in der Literatur zwei
Modelle aufgeführt. Zum einen das Broken-Line Modell und zum anderen das Reading-Modell.

70
4.1.1.1. Das Broken-Line Modell
Von den zwei Abschnitten, in die sich die Funktion (Abbildung 4) gliedert, wird der erste mit den
ansteigenden Leistungen durch eine Gerade mit einer bestimmten Steigung beschrieben (y = ax + b).
Der zweite Abschnitt wird durch eine Parallele zur x-Achse dargestellt, die das Plateau widerspiegelt
(y = c). Der Abszissenwert des Schnittpunktes der zwei Geraden kennzeichnet den Bedarf. Dabei wird
der Schnittpunkt und die Funktion der ersten Geraden solange iterativ variiert, bis der Standardfehler
der im Versuch gewonnenen Ergebnisse zu den Geraden den geringsten Wert erreicht. Der Vorteil des
Broken-Line Modells liegt darin, dass es den am Anfang meist linear verlaufenden Leistungsanstieg
klar widerspiegelt und das Plateu eindeutig bestimmt. Durch den Schnittpunkt der beiden Geraden
wird auch der Bedarf exakt definiert. Nachteilig ist, dass der kurvilineare Teil der Dosis-Wirkungs-
Kurve im Bereich des Bedarfes nicht wiedergegeben wird (SUSENBETH, 2002). Nach ROBBINS et
al. (1979) kann mit diesem Modell noch eine Bedarfsableitung erfolgen, wenn aufgrund des
Datenmaterials oder anderer Überlegungen kein anderes Modell mehr sinnvoll erscheint. Dies ist vor
allem bei einer sehr starken Streuung des Datenmaterials der Fall. Grundsätzlich sollte jedoch
kurvilinearen Modellen der Vorzug gegeben werden, da MORRIS (1983) mathematisch nachwies,
dass die tatsächliche Dosis-Wirkungs-Funktion kurvilinear sein muss. Des weiteren kommt der Autor
zu dem Schluss, dass das Broken-Line Modell den Bedarf aufgrund der methodischen Vorgehensweise
stets unterschätzt.
Broken-Line Modell
Aminosäure im Futter
Leis
tung
Bedarf
Abbildung 4: Beispielsgraph einer Broken-Line Regressionsgleichung

71
4.1.1.2. Das Reading Modell
Dieses Modell (Abbildung 5) gliedert sich analog zum Broken-Line Modell ebenfalls in zwei
Abschnitte. Wie beim Broken-Line Modell erfolgt die Darstellung des Plateau-Bereiches durch eine
Gerade (y = d). Der Unterschied besteht darin, dass der Bedarfsgraph im Bereich des
Leistungsanstieges durch eine quadratische Funktion beschrieben wird (y = ax2 + bx + c). Dies
ermöglicht nach SUSENBETH (2002) eine bessere Anpassung an den linearen Anstieg der Kurve im
unteren Versorgungsbereich und den kurvilinearen Verlauf im Bereich des optimalen Bedarfes. Die
Ermittlung des Schnittpunktes erfolgt ebenfalls iterativ (siehe auch 4.1.1.1.). Der Schnittpunkt von
quadratischer Funktion und Gerade wird hier als Bedarf definiert. Dieses Modell entspricht in hohem
Maße den sich mit steigenden Zulagen ständig ändernden Stoffwechselprozessen im Organismus, da
der Leistungsanstieg bis zum Bedarfspunkt degressiv ansteigt. Es besteht jedoch die Gefahr, dass der
Bedarf mathematisch überschätzt wird (WESTERMEIER, 1997).
Reading Modell
Aminosäure im Futter
Leis
tung
Bedarf
Abbildung 5: Beispielsgraph einer Reading Regressionsgleichung

72
4.1.2. Das quadratische Modell
Beim quadratischen Modell (Abbildung 6) wird die Dosis-Wirkungs-Beziehung durch eine Parabel
(y)= ax2 + bx + c) beschrieben (ROTH und KIRCHGESSNER, 1987) und der Bedarf als Maximum
definiert. Bei diesem Modell fallen die Bedarfswerte nach dem Erreichen des Maximums jedoch
wieder ab. Dies lässt sich aus physiologischer Sicht nur bei sehr hohen Zulagen oder bei
Nebeneffekten der Aminosäuren auf andere Stoffwechselvorgänge erklären, da der Organismus nach
KIRCHGESSNER et al. (1987) in der Lage ist, ein Nährstoffüberangebot bis zu einer bestimmten
Grenze durch homöostatische Stoffwechselvorgänge unter Beibehaltung des Leistungsniveaus zu
regulieren. POPPE und WIESEMÜLLER (1968) zeigten, dass das quadratische Modell den Bedarf bei
Vorhandensein eines Plateaus systematisch überschätzt, da die Funktion den Hochpunkt in dessen
Mitte legt. Mit Abdeckung eines sehr weiten Bereiches bei der Zulagenmenge relativiert sich jedoch
der systematische Fehler.
Quadratisches Modell
Aminosäure im Futter
Leis
tung
Bedarf
Abbildung 6: Beispielsgraph einer quadratischen Regressionsgleichung

73
4.1.3. Asymptotische Modelle
Die Bedarfsableitung durch asymptotische Modelle (Abbildung 7) erfolgt anhand einer
Exponentialfunktion (im vorliegenden Versuch: y = a + b (1 � e-c (x-d))). Dabei streben die Werte im
Plateaubereich gegen einen Grenzwert, ohne ihn zu erreichen (MORRIS und BLACKBURN, 1982).
Da bei diesem Modell auch Zulagen noch eine Steigerung bei der Dosis-Wirkungskurve verursachen,
die in vivo keinen Effekt mehr zeigen, ist die Festlegung eines konkreten Bedarfspunktes auf dem
Graph schwierig (SUSENBETH, 2002). D�MELLO und LEWIS (1970) schlagen vor, den
Abszissenwert der Kurve als Bedarf zu nehmen, bei dem der Ordinatenwert 90 % des Grenzwertes
entspricht. ROBBINS et al. (1979) setzten den Bedarf bei 95 % des Grenzwertes an. Problematisch ist,
dass die Kurve im Bedarfsbereich aufgrund des abnehmenden Grenznutzens einer weiteren Zulage
sehr flach verläuft und eine geringe Änderung des angesetzten Prozentsatzes zu einer hohen
Schwankung beim Bedarfswert führt. Bei der vorliegenden Auswertung wurden die Ergebnisse immer
bei 95 % des Grenzwertes errechnet.
Asymptotisches Modell
Aminosäure im Futter
Leis
tung
Bedarf 95 %Bedarf 90 %
Abbildung 7: Beispielsgraph einer asymptotischen Regressionsgleichung
Wie bereits der Beschreibung der verschiedenen Modelle entnommen werden kann, ist eine
Bedarfsableitung auf mathematischem Weg problematisch. Sie kann in jedem Fall nur eine
Annäherung an den tatsächlichen physiologischen Bedarf eines Tieres darstellen. Um der Wirklichkeit
jedoch so nahe wie möglich zu kommen, wurde für alle relevanten Parameter eine Bedarfsableitung
nach allen vier Modellen vorgenommen. Es wurden jedoch nur Modelle bei der Auswertung
berücksichtigt, bei denen das Bestimmtheitsmaß (r) größer als 70 % war. Für den Fall, dass mehrere

74
Modelle mit in die Auswertung einbezogen wurden, kann man den geringsten ermittelten Bedarf als
Untergrenze und den höchsten als Obergrenze des Bereiches auffassen, bei dem die Leistungen ein
Maximum erreichen.
4.2. Tryptophan
4.2.1. Struktur
Tryptophan wurde erstmals von HOPKINS und COLE (1901) entdeckt. Dabei handelt es sich um eine
aromatische α-Aminosäure mit einem Indolring an einer Methylengruppe. Neben Kohlenstoff und
Wasserstoff enthält die Seitenkette auch noch ein Stickstoffatom. Die von der chemischen Struktur her
größte Aminosäure ist stark hydrophob und kommt in zwei Formen vor, dem D- und dem L-
Tryptophan, deren chemische Struktur Abbildung 8 zeigt. Dabei handelt es sich um die optischen
Isomere.
D-Tryptophan
L-Tryptophan
Abbildung 8:
Strukturformeln von D- und L-Tryptophan
In den Proteinen vorkommende Aminosäuren gehören der L-Form an. Bei der Produktion von
synthetischen Aminosäuren kommen jedoch beide Formen vor. Werden dem tierischen Organismus
diese in racemischer Form zugeführt, so muss die D- in die L-Form überführt werden. Dies ist über
eine Desaminierung zur α-Ketoform und anschließender Aminierung zur L-Aminosäure möglich. Die
Effizienz der Umwandlung ist jedoch von der Tierart und der jeweiligen Aminosäure abhängig. Beim
Schwein läuft die Umwandlung bei Methionin und Tryptophan so effektiv ab, dass aus
ernährungsphysiologischer Sicht auf eine Trennung der optischen Isomere verzichtet werden kann
(KIRCHGESSNER und ROTH, 1985; SCHUTTE et al., 1988). Trotzdem wurde in den verschiedenen
Futtermischungen des vorliegenden Versuches für Tryptophan nur die L-Form bei den synthetischen

75
Aminosäuren eingesetzt. Ausser DL-Methionin gehörten auch alle restlichen zugesetzten Aminosäuren
der L-Reihe an.
4.2.2. Stoffwechsel
Der höhere Organismus ist nicht in der Lage Tryptophan zu synthetisieren. Aus diesem Grund zählt
Tryptophan neben Lysin, Methionin und Threonin beim Schwein zu den wichtigsten essentiellen
Aminosäuren. Die Essentialität von Tryptophan beim Schwein wurde von BEESON et al. (1948) bei
der Verfütterung einer Ration nachgewiesen, in der Fischmehl als einzige Eiweißquelle vorkam, in
welchem das Tryptophan durch eine saure Hydrolyse denaturiert worden war.
Neben seiner Hauptaufgabe als essentielle Aminosäure für die Proteinsynthese im intermediären
Stoffwechsel stellt Tryptophan auch die Vorstufe für eine Reihe physiologisch wichtiger
Verbindungen dar. Tryptophan stimuliert die Biosynthese in der Leber und spielt in ihrem
Stoffwechsel eine wesentliche Rolle. Diese Stimulation spiegelt sich in einer vermehrten Aggregation
von Ribosomen zu Polyribosomen wider, wodurch eine Schlüsselfunktion für die Synthese größerer
Proteinbausteine gegeben ist (MUNRO, 1970; SIDRANSKY et al., 1971; MC GROWN et al., 1973).
Im Falle eines Tryptophanmangels wird das Gegenteil bewirkt und es kommt zur verminderten
hepatischen Proteinsynthese (YOKOGOSHI und YOSHIDA, 1980). Entsprechend zerfallen
Polysomen zu Ribosomen, die dann einer verstärkten Abbaurate unterliegen (JAHN, 2000). Ein
Tryptophanmangel führt in erster Linie, aufgrund einer gestörten Biosynthese, zu einer negativen N-
Bilanz (LELLIS und SPEER, 1985). Damit ist eine erhöhte Ausscheidung von Stickstoff im Harn, ein
verringerter Stickstoffansatz, eine Vergrößerung der Schilddrüse und ein verminderter
Hämoglobingehalt im Blut verbunden (BEESON et al. 1949, BECKER et al. 1955, BERTRAM und
BERENDE 1983, EDMUND und ANDERSON 1983). Untersuchungen von LIN et al. (1988) führen
diese Störungen auf eine verminderte Aktivität der Ribosome im Muskelgewebe zurück. Die
unterdrückende Wirkung eines Tryptophanmangels auf die Eiweißbiosynthese ist bei Ferkeln in
verschiedenen Geweben nachgewiesen. Auch eine Reduzierung der RNA-Aktivität ist bestätigt
(PONTER et al., 1994). Die Auswirkungen von Tryptophan auf einige Enzyme ist ebenfalls bekannt.
So steigert es in der Leber die Tätigkeit der Tryptophanoxygenase (Tryptophanabbau), der
Tyrosintransaminase (Tyrosinabbau), der Serin-Threonin-Dehydratase (Threoninabbau) und der
Phosphoenolpyruvatcarboxylase (Gluconeogenese) (LÖFFLER und PETRIDES, 1997; JAHN, 2000).
Daneben blockiert es die Neubildung der Leberglucokinase, eines Enzyms, das zur Glycogenbildung
beiträgt (JAHN, 2000). Durch einen Tryptophanmangel im Futter wird auch die Futteraufnahme
negativ beeinflusst (TROTTIER und EASTER, 1995; JAHN, 2000). Der oben erwähnte verringerte

76
Stickstoffansatz ist nach GRUBER und MENKE (1984) sowie LEATHWOOD (1987) teilweise auch
eine Folge der verringerten Futteraufnahme.
Tryptophan
Kynurenin 5-Hydroxytryptophan
Kynurensäure
3-Hydroxykynurenin Serotonin
Xanthurensäure
Alanin + 3-Hydroxyanthranilsäure Melatonin
2-Acroleyl-β-Aminofumarat
Chinolinsäure
α-Hydroxymuconsäure-Semialdehyd Nicotinsäure
γ-Oxalcrotonat Picolinsäure
Nicotinamid Glutaryl-CoA
Acetyl-CoA
Abbildung 9: Stoffwechsel des Tryptophan (nach: VOET und VOET, 1994; verändert und
ergänzt)
Wie aus der Abbildung 9 ersichtlich ist, gehen aus dem Tryptophanstoffwechsel vielfältige
Metaboliten hervor. Beim Abbau von Tryptophan handelt es sich im tierischen Gewebe um den
komplexesten aller Abbauwege für Aminosäuren. Dabei werden Teile des Tryptophanmoleküls unter
Abspaltung von Alanin zu Acetyl-CoA abgebaut. Dies stellt den Hauptabbauweg des Tryptophans dar.
Einige Zwischenprodukte des Tryptophan-Katabolismus stellen Vorstufen für andere Biosynthesen
dar. Dazu gehört die Bildung des Serotonin, Melatonin, Tryptamin und Niacin (LEHNINGER et al.,
2001).
4.2.2.1. Allgemeiner Stoffwechsel
Für den aktiven Transport von Aminosäuren existieren im Körper der höheren Lebewesen spezielle
Systeme, welche nach der Michaelis-Menten-Kinetik funktionieren (SCHARRER, 1986). Das

77
bedeutet, dass die Systeme konzentrationsspezifisch arbeiten und in ihrer Leistungsfähigkeit
eingeschränkt sind. Zusätzlich sind die verschiedenen Transportsysteme substratspezifisch und somit
auf wenige Aminosäuren beschränkt. Aus diesen Gründen kann es bei einem Ungleichgewicht der
Aminosäuren zur Konkurrenz um das entsprechende Transportsystem kommen. Tabelle 50 gibt eine
Übersicht zu den verschiedenen Transportsystemen und den damit bevorzugt transportierten
Aminosäuren.
Tabelle 50: Aminosäuretransportsysteme (LÖFFLER und PETRIDES, 1997)
Aminosäuretransportsystem
Bevorzugt transportierte Aminosäuren
A-
Glycin, Alanin, Serin, Prolin, Methionin
ASCP- Alanin, Serin, Cystein, Prolin
L- Verzweigtkettige Aminosäuren (Leu, Ile, Val),
aromatische Aminosäuren (Phe, Tyr, Trp), Methionin
Ly- Lysin, Arginin, Ornithin, Histidin
Dicarboxylat- Aspartat, Glutamat
β- Taurin, β-Alanin
N- Glutamin, Asparagin, Histidin
Die verzweigtkettigen Aminosäuren (Leucin, Isoleucin und Valin), Methionin und die aromatischen
Aminosäuren (Phenylalanin, Tyrosin und Tryptophan) werden auch als �large neutral amino acids�
(LNAA) bezeichnet. Diese teilen sich ein Transportsystem, das als L-Aminosäurentransportsystem
bezeichnet wird (LÖFFLER und PETRIDES, 1997). Tryptophan wird, im Protein gebunden, mit dem
Futter zugeführt. Bei der Eiweißhydrolyse im Verdauungstrakt werden die Proteine bis zu ihren
Grundbausteinen, den Aminosäuren aufgespalten und somit wird das Tryptophan frei. Durch aktiven
Transport mit dem L-System gelangt es in das Pfortaderblut. Bei diesem Vorgang besteht nach MÖHN
(1993) keine Konkurrenz zwischen den das L-System nutzenden Aminosäuren. Im Unterschied zu den
anderen Aminosäuren wird das Tryptophan im Blut zu einem großen Teil an Albumin gebunden
transportiert. Als freie Aminosäure liegen nur etwa 10 bis 30 % vor (WURTMAN, 1980; BENDER,
1985; LEATHWOOD, 1987). Dabei konkurrieren die freien Fettsäuren im Plasma (FFA) mit dem
freien Tryptophan um ihre gewöhnliche Bindung an die Plasmaproteine, speziell das Albumin
(CURZON, 1973; HYLDGAARD-JENSEN, 1976; STEINHART, 1979). Die Blutspiegel der meisten
Aminosäuren steigen nach dem Verzehr eiweißreicher Futtermittel stark an. Durch die Arbeit der
Leber wird jedoch eine Überflutung der peripheren Organe mit freien Eiweißbausteinen verhindert.
Dies ist darauf zurückzuführen, dass ein Großteil der resorbierten Aminosäuren in der Leber

78
aufgenommen wird. Eine Ausnahme stellen Valin, Leucin und Isoleucin dar, die die Leber passieren
und vorwiegend von der Muskulatur aufgenommen werden. In der Leber werden die Aminosäuren
entweder zur Biosynthese von Proteinen (Leber- und Plasmaproteine) herangezogen oder unter
Ammoniakabspaltung in Ketocarbonsäuren umgebaut. Die Ketocarbonsäuren können in Fettsäuren
oder Glucose umgewandelt oder zur Adenosin-Triphosphat-Bildung oxidiert werden. Tabelle 51 zeigt
den Energiegewinn in ATP-Äquivalenten bei der vollständigen Oxidation ausgewählter essentieller
Aminosäuren zu Kohlendioxid und Wasser. Zum Vergleich ist zusätzlich die Energieausbeute beim
Abbau von Glucose und Stearinsäure eingefügt. Die Energieausbeute beim Abbau von essentiellen
Aminosäuren ist demnach nicht unerheblich. Aminosäuren tragen durch ihren oxidativen Abbau
wesentlich zur Erzeugung von Stoffwechselenergie bei (NELSON und COX, 2001).
Tabelle 51: ATP-Äquivalente beim Abbau ausgewählter Verbindungen (nach: LÖFFLER
und PETRIDES, 1997; verändert und ergänzt)
Verbindung
Gewonnene ATP-Äquivalente
Stearinsäure
148
Tryptophan 44
Tyrosin 44
Phenylalanin 41
Glucose 38
Lysin 35
Threonin 26
Methionin 22
Im tierischen Organismus gibt es drei verschiedene Stoffwechselbedingungen, unter denen
Aminosäuren einem oxidativen Abbau unterliegen.
1. Aminosäuren werden während der normalen Auf- und Abbauvorgänge zellulärer
Proteine nicht für die Synthese neuer Eiweiße benötigt.
2. Mit der Nahrung werden mehr Aminosäuren angeflutet, als der Organismus für die
Proteinsynthese benötigt und die Leber kurzfristig speichern kann.
3. Aufgrund mangelhafter Nährstoffversorgung werden zelluläre Proteine zur
Energielieferung herangezogen.

79
Dabei haben Tryptophan und Tyrosin die höchste und Methionin die geringste Ausbeute beim Abbau
der essentiellen Aminosäuren. Glucose liegt im Vergleich dazu im oberen Drittel der essentiellen
Aminosäuren. Der ATP-Ertrag beim Abbau von Stearinsäure ist aufgrund der höheren Anzahl an
Kohlenstoffatomen erwartungsgemäß deutlich größer. Eine vollständige Oxidation von Aminosäuren
in der Leber würde zu einem nicht verbrauchbaren Überschuss an ATP führen. Daher werden die
Eiweißbausteine nur zu einem Teil oxidiert und zum anderen in Glucose und Acetacetat überführt, das
nach einer Oxidation indirekt anderen Organen als Energielieferant zur Verfügung steht (LÖFFLER
und PETRIDES 1997). Ammoniak, der nicht der Synthese stickstoffhaltiger Substanzen dient, wird in
der Leber zu Harnstoff umgewandelt und über die Nieren mit dem Harn ausgeschieden. In der
Abbildung 10 ist der Stoffwechselweg der Aminosäuren allgemein dargestellt.
Abbildung 10: Allgemeines Schema des Aminosäuren-Katabolismus in Säugetieren (NELSON
und COX, 2001)
Die Leber hat, auch bedingt durch ihre zentrale anatomische Lage, eine Pufferfunktion für
Aminosäuren. Dies ermöglicht eine kontinuierliche Abgabe von Eiweißbausteinen zwischen den
Futteraufnahmen, so dass die Versorgung der Gewebe über den ganzen Tag sichergestellt ist
(LÖFFLER und PETRIDES, 1997). Die aktive Aufnahme von Tryptophan in die Leber unterliegt
einer Konkurrenz mit den LNAA (SALTER et al., 1985). Dieser Sachverhalt wird jedoch durch die
Möglichkeit einer Diffusion, beziehungsweise einer erleichterten Diffusion von Tryptophan in die
Leber relativiert (BENDER 1985; POGSON et al., 1984). Tryptophan ist normalerweise die
Aminosäure mit der geringsten freien Konzentration in der Leber (LÖFFLER und PETRIDES, 1997).

80
Da ein großer Teil des aufgenommenen Tryptophans in der Leber metabolisiert wird (WURTMAN et
al., 1981), ist der Gehalt im peripheren Blut geringer als im Pfortaderblut (HYLGAARD-JENSEN et
al., 1976) und abhängig von der Aufnahme in der Leber (SMITH und POGSON, 1980). Die
Konzentrationen der Aminosäuren im peripheren Blut korrelieren nach YOKOGOSHI und
WURTMAN (1986) jedoch mit der Aminosäurenzusammensetzung des Futters. Das Gehirn ist vom
allgemeinen Blutkreislauf weitgehend durch einen selektiven Filter, die sogenannte Blut-Hirn-
Schranke, getrennt (VOET und VOET, 1994). Aus diesem Grund muss das Tryptophan auch hier aktiv
mit dem L-System transportiert werden. Die Transportkapazität in der Blut-Hirn-Schranke ist aber
geringer als in anderen Geweben (WURTMANN, 1980). Dies hat zur Folge, dass das Tryptophan mit
den LNAA im Wettstreit um diesen `Carrier´ steht (MEISINGER und SPEER, 1979; McGIVAN und
PASTOR-ANGLADA, 1994; TROTTIER und EASTER, 1995; SIDRANSKY, 2002). Dieser
Wettstreit ist von größerer Bedeutung als bei anderen Geweben (STEINHART, 1984), da das
Tryptophan im Gehirn den Serotoningehalt beeinflusst (WURTMANN, 1980).
4.2.2.2. Tryptophan und Serotonin
Beim Serotonin handelt es sich um ein biogenes Monoamin. Es geht in zwei Stufen aus Tryptophan
hervor. Abbildung 11 zeigt die Syntheseschritte von Tryptophan zu Serotonin.
Abbildung 11: Entstehung von Serotonin aus Tryptophan (VOET und VOET, 1994; verändert)

81
Diese Synthese findet in der Hypophyse, in der Milz, in der Lunge und im Darmtrakt statt. Schon seit
vielen Jahren haben Wissenschaftler vermutet, dass es im Gehirn und im restlichen Körper getrennte
Wege für die Synthese von Serotonin geben müsse, da Serotonin die Blut-Hirn-Schranke nicht
überwinden kann (HYLGAARD-JENSEN, 1976; VOET und VOET, 1994). Sowohl im Gehirn als
auch im restlichen Körper war jedoch immer die Tryptophanhydroxylase für die Serotoninbildung
zuständig (WURTMAN, 1980). Eine Klärung des Sachverhaltes ist erst in der jüngsten Vergangenheit
WALTHER et al. (2003) gelungen. Die Forscher konnten nachweisen, dass es zwei Gene (THP) gibt
(THP1: war schon bekannt; THP2: neu entdeckt), die eine aktive Tryptophanhydroxylase produzieren.
Der Unterschied besteht im Wirkungsort des Enzyms. Das THP2 veranlasst die Produktion von
Tryptophanhydroxylase ausschließlich im Gehirn, während das vom THP1 produzierte Enzym für die
Herstellung von Serotonin im restlichen Körper verantwortlich ist. Ebenfalls konnte nachgewiesen
werden, dass bei allen Wirbeltieren das THP2 vorkommt.
Beim Serotonin handelt es sich um ein Hormon, das vasokonstriktorisch wirkt (OCHSENKÜHN,
1988) und die glatte Muskulatur der Gefäße, des Respirations- und des Gastrointestinaltraktes anregt
(KARLSON et al., 1994). Ferner ist Serotonin im Gehirn als Neurotransmitter wirksam, wo es das
Fressverhalten (HENRY et al., 1992; TROTTIER und EASTER, 1995), den Schlaf-Wach-Rhythmus
sowie die Stimmungslage (SCHAHRZAD, 1994) beeinflusst. Dabei beeinflusst die Konzentration des
Tryptophans im Gehirn zu jeder Zeit die Synthese und Freisetzung von Serotonin als Neurotransmitter
(RICHARD und WURTMAN, 1980). EDMONDS et al. (1987) zeigten, dass bei Schweinen ein
Überschuss von Tryptophan im Futter zu Verzehrsdepressionen führt.
Die Tryptophankonzentration im Gehirn ist von mehreren Faktoren abhängig. Einmal indirekt vom
Gehalt an LNAA im Plasma, was darauf zurückzuführen ist, dass mit zunehmender Konzentration von
LNAA im Plasma diese mit Tryptophan um die Aufnahme über das L-System durch die Blut-Hirn-
Schranke konkurrieren (RICHARD und WURTMAN, 1980). Da diese Konkurrenz mit zunehmendem
Verhältnis von Tryptophan/LNAA zugunsten des Tryptophans abnimmt, steigt folglich die Passagerate
durch die Blut-Hirn-Schranke, und es erhöht sich somit der Tryptophangehalt im Gehirn (RICHARD
und WURTMAN, 1980; FERNSTROM, 1991). WURTMAN und FERNSTROM (1974),
WURTMAN (1980) sowie WURTMAN und WURTMAN (1996) zeigten, dass mit dem Verzehr einer
kohlenhydratreichen Diät die Gehalte an LNAA im Blut fallen, der Tryptophangehalt jedoch nicht.
Dies beruht auf der Tatsache, dass durch den infolge der Kohlenhydratverdauung ansteigenden
Blutzuckerspiegel die Freisetzung von Insulin angeregt wird, das die Aufnahme von langkettigen
Aminosäuren in vielen Geweben und Organen anregt. Betroffen davon sind zum Beispiel Valin,
Leucin und Isoleucin, die zur Gruppe der LNAA gehören. Da Tryptophan nicht betroffen ist,
vergrößert sich das Verhältnis von Tryptophan/LNAA (YOKOGOSHI und WURTMAN, 1986),

82
wodurch die Passagerate von Tryptophan durch die Blut-Hirn-Schranke begünstigt wird und seine
Konzentration im Gehirn ansteigt. Damit ist ein erhöhter Serotoningehalt im Gehirn erklärbar. Der
Tryptophangehalt im Gehirn wird aber auch direkt durch die Konzentration im Plasma, die aus dem
freien und dem gebundenen Tryptophan besteht, beeinflusst.
Vom freien Tryptophan wird angenommen, dass es die Fraktion im Blut repräsentiert, die bestimmend
für die Transportrate des Tryptophans durch die Blut-Hirn-Schranke ist (FERNSTROM, 1991). So
konnten mehrere Arbeitsgruppen eine deutliche Abhängigkeit der Tryptophangehalte im Gehirn von
der Konzentration an freiem Tryptophan im Plasma aufzeigen (KNOTT und CURZON, 1972;
TAGLIAMONTE et al., 1973; HILLER et al., 1974). Im Unterschied zu den anderen Aminosäuren
wird Tryptophan im Blut zu einem grossen Teil an Albumin gebunden transportiert (WURTMAN,
1980; BENDER, 1985; LEATHWOOD, 1987). Dabei konkurrieren die freien Fettsäuren (FFA) im
Plasma mit dem freien Tryptophan um ihre gewöhnliche Bindung an das Albumin (STEINHART,
1979), was zu den oben beschriebenen nicht unerheblichen Schwankungen beim Gehalt an freiem
Tryptophan im Blut führen kann. Albumin weist zum einen gegenüber Tryptophan eine sehr große
Bindungskapazität auf. Zum anderen ist die Affinität gegenüber dieser Aminosäure relativ gering.
Daher führt ein Anstieg der freien Fettsäuren im Plasma indirekt zu einem steigenden Gehalt an freiem
Tryptophan im Blut. Befindet sich der Organismus im Hungerzustand, erfolgt eine gesteigerte
Anflutung von freien Fettsäuren im Blut. Bei Ratten fanden KNOTT und CURZON (1972) nach 24-
stündiger Nüchterung eine Zunahme bei den freien Fettsäuren um 159 %, die mit einem gleichzeitigen
Anstieg beim freien Tryptophan um 150 % einherging. Die Zusammenhänge sind in der Abbildung 12
dargestellt.

83
Abbildung 12: Schema der fütterungsbedingten Serotoninbeeinflussung im Gehirn (nach
FERNSTROM und WURTMAN, 1972; verändert und ergänzt)
4.2.2.3. Tryptophan und Niacin
Der Begriff Niacin bezeichnet ein Vitamin der B-Gruppe und wird als Sammelbegriff für Nicotinamid
und Nicotinsäure sowie für die Coenzyme Nicotinamid-Adenin-Dinukleotid (NAD) und Nicotinamid-
Adenin-Dinukleotid-Phosphat (NADP) verwendet. Ihre Biosynthesewege sowie die chemische
Struktur sind aus den Abbildungen 13 und 14 ersichtlich. Bei Niacin handelt es sich um ein
Pyridinderivat, das beim Schwein aus Tryptophan synthetisiert werden kann (HARPER et al., 1987).
Die Nicotinamid-Nucleotide NAD und NADP dienen als Coenzyme bei einer Vielzahl reversibler
Redoxreaktionen. Die Wiederherstellung der normalen Nicotinamid-Nucleotid-Konzentration im Blut
ist nach der Proteinsynthese die zweitwichtigste Aufgabe des Tryptophanstoffwechsels
(SCHAHRZAD, 1994).
Fütterung
LNAA - TRP + Kohlenhydrate + FFA +
Plasma FFA + freies Plasma TRP + (geringe Wirkung)
Plasma LNAA -
Plasma TRP/LNAA +
Gehirn TRP +
Gehirn Serotonin +
(Insulin)
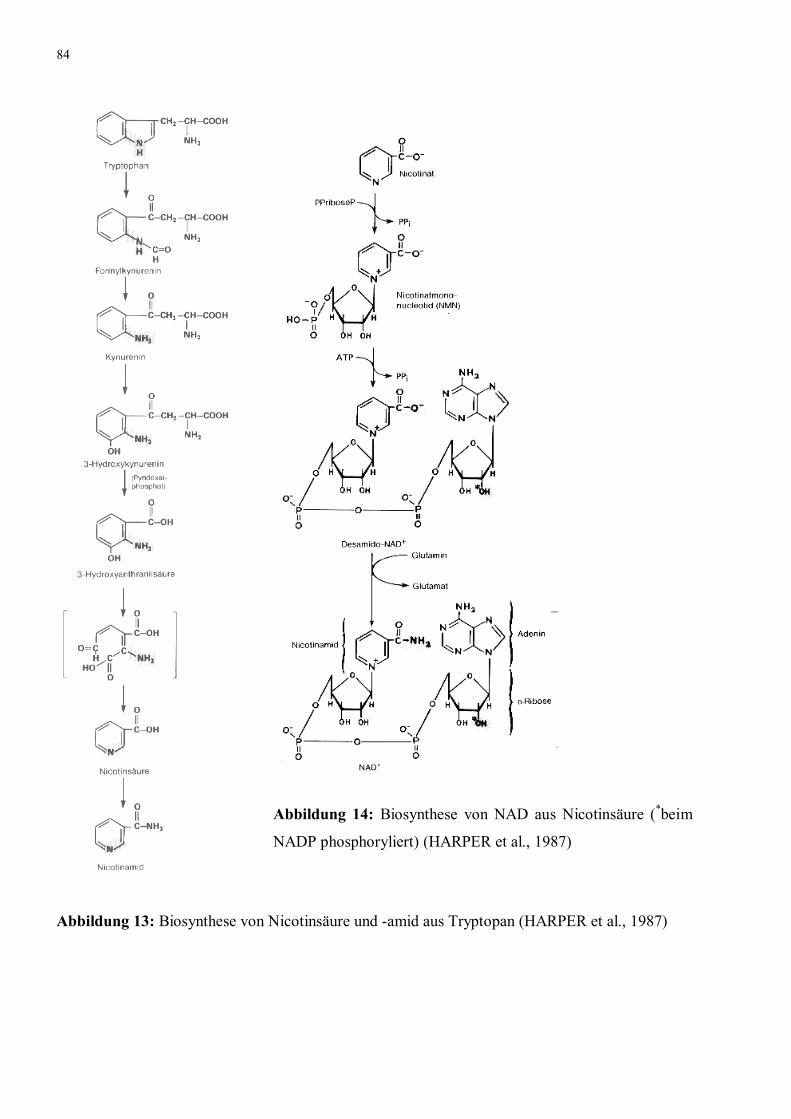
84
Abbildung 14: Biosynthese von NAD aus Nicotinsäure (*beim
NADP phosphoryliert) (HARPER et al., 1987)
Abbildung 13: Biosynthese von Nicotinsäure und -amid aus Tryptopan (HARPER et al., 1987)

85
4.2.3. Scheinbare ileale Verdaulichkeit in Futtermitteln
Für ein festgelegtes Leistungsniveau ist die Kenntnis der scheinbaren ilealen Verdaulichkeit des
Tryptophans aus der Nahrung für eine bedarfsgerechte Versorgung unumgänglich. Unter der
scheinbaren ilealen Verdaulichkeit versteht man diejenige Menge eines Stoffes, die aus dem
Dünndarm resorbiert werden kann, wobei endogene Ausscheidungen nicht berücksichtigt werden. Der
Einfluss mikrobieller Umsetzungen im Dickdarm wird dabei weitestgehend ausgeschlossen. Unter der
Annahme, dass verdaute und aufgenommene Aminosäuren dem Stoffwechsel zur Verfügung stehen,
führt dies zu einer exakteren Leistungsprognose im Vergleich zur Angabe des Gesamtgehaltes.
Besonders deutlich wird dies, wenn in Rationen Futtermittel eingesetzt werden, deren ileal
verdaulicher Gehalt einer essentiellen Aminosäure große Unterschiede zum absoluten Gehalt aufweist.
So entsprechen 10 g Lysin aus Erbsen 8,4 g scheinbar ileal verdaulichem Lysin, während es beim
Roggen nur 6,4 g sind (NRC, 1998). Dies entspricht einer Differenz von 20 % bezogen auf den
Gesamtgehalt an Lysin. An diesem Beispiel wird deutlich, warum neben dem Gesamtbedarf an
Aminosäuren auch die Angabe auf Basis der scheinbaren ilealen Verdaulichkeit erfolgen muss.
Tabelle 52: Literaturangaben zur ilealen Verdaulichkeit von Tryptophan beim Schwein
Futtermittel
Verdaulichkeit in %
Quelle
Erbsen
81
70
NRC (1998)
DEGUSSA (1999)
Kartoffeleiweiß 80 DEGUSSA (1999)
Mais 48
70
64
76
VAN LEEUWEN et al. (1987)
NRC (1988)
NRC (1998)
DEGUSSA (1999)
Maiskleberfutter 33
47
66
EUROLYSINE (1988)
NRC (1998)
DEGUSSA (1999)
Sojaex.schrot (44 % XP) 81
87
80
87
TANKSLEY et al. (1984)
VAN LEEUWEN et al. (1987)
NRC (1998)
DEGUSSA (1999)
Trockenschnitzel 50 DEGUSSA (1999)

86
In der Literatur sind Daten zur ilealen Verdaulichkeit von Tryptophan in den unterschiedlichsten
Futtermitteln zu finden. Während man bei synthetisch hergestellten Aminosäuren der L-Form von
einer Verfügbarkeit aus dem Futter von nahezu 100 % ausgehen kann (KIRCHGESSNER und ROTH,
1985; SCHUTTE et al., 1988), schwankt die Verfügbarkeit in nativen Rohstoffen beträchtlich. Tabelle
52 gibt die Literaturwerte für die Verdaulichkeit der im vorliegenden Versuch eingesetzten
Futtermittel an.
Dabei ist auffallend, dass bei verschiedenen Untersuchungen die Werte für einzelne Futtermittel zum
Teil sehr große Schwankungen aufweisen. Beispielhaft sei hier das Maiskleberfutter genannt, für
welches in verschiedenen Versuchen ileale Verdaulichkeitswerte mit relativen Abweichungen von 100
% ermittelt wurden. Zur Überprüfung des im vorliegenden Versuch rechnerisch ermittelten Wertes für
die scheinbare ileale Verdaulichkeit des Gesamttryptophans bei der Basisgruppe von 75 % wurde
daher ein Kontrollversuch durchgeführt, bei dem der heute als Methode der Wahl angesehene
Verdauungsversuch mit ileorektalanastomierten Tieren zur Ermittlung der scheinbaren ilealen
Verdaulichkeit durchgeführt wurde (MOUGHAN und SMITH, 1996; GfE, 2002; siehe auch 2.4.2.).
Die gefundenen Ergebnisse beim scheinbaren ileal verdaulichen Tryptophan bestätigten die
rechnerischen Werte (vergleiche Tabelle 53). Die berechneten Gesamttryptophangehalte stimmten
ebenfalls gut mit den Analysenergebnissen der Futtermischungen überein.
Tabelle 53: Gesamtgehalt und scheinbar ileal verdauliches Tryptophan im Futter in % im
Vergleich von Kalkulations- und Untersuchungsergebnissen
Gruppe I II III IV V VI Tryptophangehalt ges. berechn. 0,12 0,15 0,18 0,24 0,30 0,42 Tryptophangehalt ges. unters. 0,12 0,15 0,18 0,24 0,29 0,40 sch. ileal verd. Trp berechnet 0,09 0,12 0,15 0,21 0,27 0,39 sch. ileal verd. Trp untersucht 0,08 0,11 0,14 0,20 0,26 0,38
Die Werte in der letzten Zeile wurden ab der Gruppe II rechnerisch ermittelt, da zur Grundration nur
noch synthetisches L-Tryptophan zugelegt wurde. Dies hat nach KIRCHGESSNER und ROTH (1985)
sowie SCHUTTE et al. (1988) eine scheinbare ileale Verdaulichkeit von 100 %. Da sich die Gruppen
II bis VI nur im Gehalt an synthetischem Tryptophan unterschieden, war eine rechnerische
Weiterführung der Ergebnisse angesichts des hohen Aufwandes für weitere in vivo Untersuchungen
vertretbar.

87
4.3. Auswirkung des Tryptophangehaltes im Futter auf die verschiedenen
untersuchten Versuchsparameter
4.3.1. Futteraufnahme der Sauen
Eine unzureichende Futteraufnahme während der Laktation ist ein limitierender Faktor für gute
Leistungen in der Ferkelproduktion, da sie die Milchleistung und die Lebendmasseverluste der Sauen
sowie die Tageszunahmen der Ferkel negativ beeinflusst (O�GRADY et al, 1985; LYNCH, 1988;
TROTTIER und EASTER, 1995; VERSTEGEN et al., 1998). Auch negative Auswirkungen auf die
Überlebensrate im frühen Embryonalstadium (HUGHES et al., 1984) und die tatsächliche Wurfgröße
einer nachfolgenden Trächtigkeit sind in der Literatur beschrieben (KIRKWOOD et al., 1988). Je nach
Textquelle wird die Ursache in einer mangelnden Eiweißversorgung (O�GRADY und HANRAHAN,
1975; WALKER et al, 1979; BRENDEMUHL et al., 1987; 1989) oder Energieaufnahme (REESE et
al., 1982; Johnston et al., 1989) beziehungsweise in beidem (KING und WILLIAMS, 1984) gesehen.
Grundsätzlich beeinflussen viele verschiedene Faktoren die Futteraufnahme der Sau. In Tabelle 54 ist
eine Übersicht der in der Literatur beschriebenen Faktoren gegeben.
Tabelle 54: Einflussfaktoren auf den Futterverzehr von Sauen während der Laktation
(Auswahl; Literaturübersicht)
Faktor
in der Literatur benannt bei
Sauenrasse
POND et al. (1981)
Trächtigkeitsfütterung LODGE (1972); KIRCHGESSNER et al. (1991); WELDON
et al. (1993)
Gesundheitszustand der Sau NELSON (1979); O�GRADY et al. (1985)
Laktationsstadium JOHNSTON et al. (1989)
Energiegehalt im Futter BOYD et al. (1978); O�GRADY et al. (1984); HOUPT
(1984)
Eiweißgehalt im Futter MAHAN und GRIFO (1975); MAHAN und MANGAN
(1975); SHIELDS (1984)
Futterkonsistenz O�GRADY und LYNCH (1978)
Stalltemperatur LYNCH (1977); O�GRADY (1985)
Aufstallungsform ROWLINSON und BRYANT (1982)

88
Damit der Einfluss einer variierenden Aminosäurenzulage im Futter bestimmt werden konnte, wurden
die restlichen, die Futteraufnahme beeinflussenden Faktoren im vorliegenden Versuch konstant
gehalten (siehe auch: 2.2.). Besonders wurde darauf geachtet, dass die Sauen während der Trächtigkeit
nicht überversorgt wurden, da eine zu hohe Nährstoffaufnahme zu vermehrter Körperfettbildung führt,
die wiederum in der Säugephase aufgrund homöorhetischer Regulation verringerte Verzehrsmengen
verursacht (WILLIAMS, 1998). Vom 60. bis zum 85. TT bekamen die Sauen 2,4 kg und bis zum 100.
TT 2,8 kg Futter je Tag. Bei einem Energiegehalt von 11,5 MJ/kg FS war damit die tägliche
Energieversorgung auf 28 beziehungsweise 32 MJ ME je Sau beschränkt. Die Empfehlungen für die
tägliche Energieversorgung von trächtigen Sauen der GfE (1987) sowie der DLG (1996) wurden somit
nicht überschritten.
Die Rolle der Aminosäuren bei der Kontrolle der Futteraufnahme wird schon seit längerer Zeit
beobachtet (GRUBER, 1985; TEIK-NG und ANDERSON, 1992). Ein den Futterverzehr
beeinflussender Effekt trat vor allem dann auf, wenn im vorgelegten Futter die Aminosäuren-
Relationen unausgeglichen waren. In der Literatur wird darüber bei Valin (HUTCHISON et al., 1983;
JAHN, 2000), Phenylalanin (LELLIS und SPEER, 1987), beim Verhältnis von Tryptophan / BCAA
(TROTTIER und EASTER, 1995) und bei Tryptophan (MEISINGER und SPEER, 1979) berichtet.
tägliche Futteraufnahme der Sauen
1
2
3
4
5
6
7
0,07 0,12 0,17 0,22 0,27 0,32 0,37 0,42 0,47
Tryptophangehalt im Futter in %
Futte
rauf
nahm
e in
kg/
Tag
Abbildung 15: Mittlere tägliche Futteraufnahme der Sauen während der Laktation
Da die Relationen der einzelnen Aminosäuren mit Ausnahme des Tryptophans bei den verschiedenen
Futtergruppen des vorliegenden Versuches anhand der Relationen des `idealen Protein´ zu Lysin nach
WANG und FULLER (1989) und GOTTERBARM et al. (1997) festgelegt wurden und ceteris paribus
Bedingungen vorherrschten, müssen Verzehrsdepressionen aus der Aminosäuren-Imbalance zwischen

89
Tryptophan und den restlichen Aminosäuren resultieren. In Abbildung 15 sind die Gruppenmittelwerte
der Versuchssauen bezüglich der Futteraufnahme während der 28 Tage dauernden Laktation in
Abhängigkeit von den verschiedenen Tryptophankonzentrationen im Laktationsfutter dargestellt.
Trotz der relativ großen Standardabweichung, die durch den Individualeffekt der Sau verursacht
wurde, war ein Einfluss des Tryptophangehaltes im Futter auf die Verzehrsmenge zu erkennen. Die
höchste Futteraufnahme wies Gruppe IV mit über 5,5 kg/Tag auf und übertraf somit Gruppe I, die mit
3,2 kg/Tag den geringsten Futterverzehr hatte, um mehr als 70 %. Die restlichen Gruppen lagen mit
durchschnittlich 4,4 kg/Tag dazwischen; statistisch gesehen unterschieden sie sich signifikant von den
Gruppen I und IV. Der Gesamtdurchschnitt der Futteraufnahme lag im vorliegenden Versuch bei 4,4
kg/Tag. Bei anderen in der Literatur beschriebenen Versuchen war die durchschnittliche
Futteraufnahme ähnlich hoch (SCHNEIDER et al., 1992: 4,6 kg/Tag; WESTERMEIER, 1998: 4,3
kg/Tag; OTT, 2000: 4,4 kg/Tag).
Futteraufnahme der Sauen in der 4. LW
y = -82,4x2 + 47,6x - 0,68r = 0,86 n = 72
2
3
4
5
6
0,07 0,12 0,17 0,22 0,27 0,32 0,37 0,42 0,47
Tryptophangehalt im Futter in %
Futte
rauf
nahm
e in
kg/
Tag
Bedarf = 0,29
Abbildung 16: Ableitung des Tryptophanbedarfs anhand der Futteraufnahme in der 4. LW nach
dem quadratischen Modell
Führt man eine Bedarfsableitung nach dem quadratischen Modell durch, so liegt der optimale
Tryptophangehalt zur Erzielung eines größtmöglichen Verzehrs bei 0,29 % im Futter (siehe Abbildung
16). In der Literatur wird ein Anstieg der Futteraufnahme bei zunehmenden Tryptophangehalten im
Futter beschrieben. So fanden SOHAIL et al. (1978) eine zunehmende Futteraufnahme bei einer
Steigerung des Tryptophangehaltes in der Laktationsration von 0,12 % auf 0,17 %. Dies konnten

90
LIBAL et al. (1997) in einem ähnlichen Versuch bestätigen. Da auch in der vorliegenden Arbeit im
Bereich dieser Tryptophangehalte ein Anstieg bei der Futteraufnahme zu verzeichnen war, decken sich
die Ergebnisse gut. Die Veränderungen bei der Futteraufnahme können auch auf die Fähigkeit der
Schweine zur Auswahl eines Futters mit annähernd bedarfsgerechter Tryptophankonzentration
zurückgeführt werden. Die Grundlage für diese Annahme bilden Futterwahlversuche bei Ferkeln von
ETTLE und ROTH (2003) und ROTH und ETTLE (2003), die diese Fähigkeit bestätigen.
MENNENGA und EASTER (1984) sowie JONES und STAHLY (1999) fanden einen Einfluss des
Lysingehaltes, TROTTIER und EASTER (1995) des Gehaltes an verzweigtkettigen Aminosäuren und
LELLIS und SPEER (1985 und 1987) des Gehaltes an aromatischen Aminosäuren auf die
Futteraufnahme. Bei anderen Versuchen traten keine Einflüsse auf (Lysin: BOOMGAARD et al.,
1972; CHEN et al., 1978; MENNENGA und EASTER, 1985; ETIENNE et al., 1989;
Methionin/Cystin: SCHNEIDER et al., 1992; Threonin: WESTERMEIER et al., 1998; Valin:
RICHERT et al., 1996; OTT, 2000).
Beim Protein- und Energiegehalt wiesen die verschiedenen Futter zwischen den Gruppen keine
Unterschiede auf. Somit ist die Protein- und Energieaufnahme direkt von der Futteraufnahme
abhängig. Die Zusammenhänge sind daher analog denen bei der Futteraufnahme.
Da nach HENRY et al. (1984), HUGHES et al. (1984) und KIRKWOOD et al. (1988) ein enger
Zusammenhang zwischen der Futteraufnahme während der Säugezeit und den anschließenden
Zuchtleistungen besteht, ist mit steigender Futteraufnahme auch eine Verbesserung der
Reproduktionsleistungen zu erwarten. Somit ist eine maximale Futteraufnahme nicht nur für die direkt
messbaren Parameter wie Milchleistung und Ferkelzunahmen von Bedeutung, sondern auch für eine
nachhaltige Aufrechterhaltung hoher Zuchtleistungen.
4.3.2. Lebendmasse der Sauen
Bei der Untersuchung der Auswirkungen einer variierenden Tryptophanzulage im Futter auf die
Entwicklung der Lebendmassen der Sauen sind vor allem zwei Aspekte von Bedeutung. Zum einen
soll der Abbau von Lebendmasse bei Zuchtsauen während der Säugephase so gering wie möglich
ausfallen, da dies zur Erhaltung einer hohen und langfristigen Zuchtleistung entscheidend beiträgt
(DOURMAD et al., 1994; WILLIAMS, 1998). Besonders negativ ist dieser Einfluss auf die
Fruchtbarkeit der Sauen (BRENDEMUHL et al., 1987; KIRKWOOD et al., 1987; KING, 1987). Zum
anderen kann der Energie- und Nährstoffbedarf der Sau, vor allem zu Beginn der Laktation, nicht
durch die Futteraufnahme allein gedeckt werden. Dies ist auch bei uneingeschränktem Futterangebot
der Fall (NOBLET et al., 1990; DOURMAD, 1991). In der Regel herrscht also bei Sauen während der
Säugephase eine negative Energie- und Eiweißbilanz vor (JOHNSTON et al., 1993; KING et al.,

91
1993B; EVERTS und DEKKER, 1994). Zur Verhinderung einer hohen Sterberate der Ferkel wegen
Agalaktie bei Nahrungsmangel der Muttersau verfügt die Milchdrüse in der Laktationszeit zusammen
mit dem Gehirn und dem Zentralnervensystem über die höchste Priorität bei der Nährstoffversorgung
der Körpergewebe (KIRCHGESSNER, 1997). Daher werden als teilweiser Ausgleich bei einer
Mangelernährung Körperreserven mobilisiert, was zwangsläufig zu einer Abnahme der Lebendmasse
führt, da alle anderen anabolen Stoffwechselvorgänge bereits vorher eingestellt werden (KING und
DUNKIN, 1986; BRENDEMUHL et al., 1989; SCHNEIDER et al., 1992; KIRCHGESSNER, 1997).
Nach COFFEY et al. (1994) und DOURMAD et al. (1994) führt eine Überversorgung mit Energie und
Nährstoffen während der Tragezeit zu erhöhten Lebendmasseverlusten während der Säugephase. Im
vorliegenden Versuch wurde dies jedoch ausgeschlossen (siehe auch 3.3.1. und 5.3.1.) und somit
konnte ein Einfluss der Trächtigkeitsfütterung auf die Höhe der Lebendmasseverluste nicht auftreten.
In der Abbildung 17 sind die aufgetretenen täglichen Masseverluste der Sauen in Abhängigkeit der
Futtergruppe eingetragen.
täglicher Masseverlust der Sauen
-500
0
500
1000
1500
2000
0,07 0,12 0,17 0,22 0,27 0,32 0,37 0,42 0,47
Tryptophangehalt im Futter in %
Mas
seve
rlust
in g
/Tag
Abbildung 17: Masseverlust der Sauen während der gesamten Laktation in g/Tag
Bei der Basisgruppe mit 0,12 % Tryptophan im Futter traten signifikant höhere tägliche Masseverluste
auf als bei den Gruppen IV bis VI. Diese betrugen im Mittel über 660 g/Tag und zwischen den
Extremen sogar über 760 g/Tag. Absolut bedeutete dies einen Unterschied bei den Masseverlusten
über die gesamte Laktation von 21,3 kg im Durchschnitt. Die starke verzehrsvermindernde Wirkung
eines Tryptophangehalts von 0,12 % im Futter spiegelte sich in einer hohen Mobilisierung von
Körperreserven wider. Die Verluste bei den Gruppen II und III lagen mit durchschnittlich 640 g/Tag
statistisch zwischen der Mangelgruppe und den Gruppen IV bis VI. Auffallend waren die im Vergleich

92
zu den Gruppen IV und V hohen Standardabweichungen der restlichen Gruppen. Dies lässt zum einen
auf große Unterschiede bei der tierindividuellen Fähigkeit zur Nährstoffmobilisation aus
Körperreserven schließen, zum anderen scheint die Notwendigkeit einer verstärkten Mobilisation bei
den Gruppen IV und V nicht so stark gegeben zu sein. Die wiederum große Standardabweichung bei
Gruppe VI lässt sich zusätzlich durch die Belastung des Stoffwechsels aufgrund der überhöhten
Tryptophanzulage von 0,42 % in Verbindung mit der individuellen Fähigkeit der Tiere zur
Verstoffwechselung erklären. Unter Einbeziehung dieser Tatsache sowie dem Auftreten von
signifikanten Unterschieden und den Ergebnissen der Futteraufnahme scheint eine Bedarfsableitung
anhand dieses Parameters als durchaus sinnvoll. Zusätzlich spricht für eine Ermittlung des
Tryptophanbedarfs anhand der Lebendmasseverluste der Sauen die Korrelation mit verschiedenen
Fruchtbarkeitsparametern, die die nachfolgende Trächtigkeit betreffen. So verringert sich mit
abnehmendem Fett- (KING, 1987) und Eiweißverlust (KING und WILLIAMS, 1984) die Dauer
zwischen Absetzen und Östrus. BLACK et al. (1986) und KING (1987) fanden eine negative
Korrelation zwischen Masseverlust und Konzeptionsrate beziehungsweise Güstzeit.
täglicher Masseverlust der Sauen
0
500
1000
1500
0,07 0,12 0,17 0,22 0,27 0,32 0,37 0,42 0,47Tryptophangehalt im Futter in %
Mas
seve
rlust
der
Sau
en in
g/T
ag
Bedarf = 0,21
Abbildung 18: Ableitung des Tryptophanbedarfs anhand der Lebendmasseverluste der Sauen
während der gesamten Laktationsdauer nach dem asymptotischen Modell
y = [1,087 � 0,629 � (1 � e-33,8 � (x-0,12))] � 1000 Grenzwert = 458 r2 = 0,87 n = 72

93
täglicher Masseverlust der Sauen
0
500
1000
1500
0,07 0,12 0,17 0,22 0,27 0,32 0,37 0,42 0,47Tryptophangehalt im Futter in %
Mas
seve
rlust
der
Sau
en in
g/T
ag
Bedarf = 0,26
y = 37652x² - 19216x + 2803 Plateau = 351r² = 0,85 n = 72
Abbildung 19: Ableitung des Tryptophanbedarfs anhand der Lebendmasseverluste der Sauen
während der gesamten Laktationsdauer nach dem Reading-Modell
täglicher Masseverlust der Sauen
0
500
1000
1500
0,07 0,12 0,17 0,22 0,27 0,32 0,37 0,42 0,47Tryptophangehalt im Futter in %
Mas
seve
rlust
der
Sau
en in
g/T
ag
Bedarf = 0,20
y = -6900x + 1832 Plateau = 438r² = 0,80 n = 72
Abbildung 20: Ableitung des Tryptophanbedarfs anhand der Lebendmasseverluste der Sauen
während der gesamten Laktationsdauer nach dem Broken-Line-Modell

94
Zur Ermittlung der Fraktionen, aus denen sich der Körpermasseverlust der Sauen zusammensetzt, sind
in der Literatur verschiedene Versuche beschrieben. GUETTE und LENKEIT (1960) führten den
Masseverlust hauptsächlich auf einen Proteinabbau zurück, während WHITTEMORE und ELSLEY
(1976) feststellten, dass nur Fett abgebaut wurde. ALTAVINI und FERNANDES (1990) setzten Sauen
gezielt einem Energiemangel aus und stellten fest, dass nur Fettreserven eingeschmolzen wurden. Bei
einer Unterversorgung mit Futterprotein beobachteten DOURMAD et al. (1991) einen zunehmenden
Abbau von Körpereiweiß (Muskelgewebe). TOKACH et al. (1992) fanden, dass sich ein
Energiemangel stärker auf die Lebendmasse der Sauen auswirkte als ein Lysinmangel. Somit wird die
Höhe und die Zusammensetzung der abgebauten Körpermasse wesentlich durch die Menge und die
Gehalte an Energie und Protein des aufgenommenen Laktationsfutters bestimmt (REVELL et al.,
1998; DOURMAD et al., 1998; JONES und STAHLY, 1999). Bei einer restriktiven Fütterung oder
einer verminderten Futteraufnahme infolge antinutritiver Inhaltsstoffe ist folglich bei einer
ausgewogenen Ration davon auszugehen, dass sich der Körpermasseabbau bei laktierenden Sauen aus
Fett und Eiweiß zusammensetzt. Dies wurde in Versuchen von MULLEN und WILLIAMS (1990),
DOURMAD (1991), ROJKITTIKUHN et al. (1992) sowie EVERTS und DEKKER (1994) bestätigt.
Das Verhältnis von Fett- zu Proteinabbau bewegt sich je nach Kondition der Sau von 100 : 53 bis 100 :
35 (VAN DER PEET-SCHWERING et al., 1998). Dies ist dadurch zu erklären, dass einem
Organismus nur begrenzt Fettreserven zur Verfügung stehen, die bei einem sehr hohen Masseverlust
rasch aufgebraucht sind. Da der limitierende Faktor nun die Energie ist, geht der Körper dazu über, die
zusätzlich notwendige Energie aus der Verstoffwechselung von Proteinen zu gewinnen. Diese Form
der Energiegewinnung ist jedoch weniger effektiv (LÖFFLER und PETRIDES 1997; siehe auch
Tabelle 51), wodurch sich überhöhte Masseverluste bei der Basisgruppe zusätzlich erklären lassen.
Abgemildert wurde dieser Effekt jedoch durch die Tatsache, dass für diesen Vorgang relativ viel
resorbierte Aminosäuren aus dem Verdauungstrakt zur Verfügung standen, die aufgrund des
Tryptophanmangels im Futter nicht für anabole Stoffwechselvorgänge verwendet werden konnten.
Zusammenfassend kann gesagt werden, dass die Lebendmasseabnahmen in Art und Höhe bei
laktierenden Sauen in erster Linie von der Menge und Zusammensetzung des aufgenommenen Futters
abhängen (JONES und STAHLY, 1999). In der vorliegenden Arbeit wurde ein bedarfsgerechtes Futter
verwendet, das lediglich beim Tryptophangehalt variierte. Dieser hatte einen starken Einfluss auf die
Futteraufnahme, was bedeutete, dass die Futteraufnahme von Gruppe I bis Gruppe IV zunahm und
danach wieder abfiel. Da die Versorgung der Ferkel mit Milch jedoch vom maternalen Organismus
nach Möglichkeit aufrecht erhalten wird, werden Körperfett und -eiweiß abgebaut, wenn die
erforderliche Energie- und Nährstoffmenge über die Futteraufnahme nicht mehr sichergestellt ist. Im
vorliegenden Versuch ergab sich ein Korrelationskoeffizient von r = -0,92 zwischen den

95
Lebendmasseverlusten und der Futteraufnahme der Sauen während der gesamten Säugedauer.
Zwischen den Masseverlusten und der durchschnittlichen Milchleistung über alle Messtage betrug der
Koeffizient ebenfalls -0,92.
4.3.3. Milchabgabe der Sauen
Die Sauenmilch stellt für die Ferkel in den ersten Lebenswochen bis zur Beifütterung die einzige
Nahrungsquelle dar. Aus diesem Grund ist die Milchleistung der Sau in Verbindung mit der
Zusammensetzung der Inhaltsstoffe als wichtigste Voraussetzung für ein optimales Wachstum der
Ferkel zu sehen. Die Höhe der Milchproduktion wird bei der Sau durch eine Vielzahl von Faktoren
beeinflusst. Tabelle 55 gibt eine Übersicht der in der Literatur diskutierten Wirkungen auf die
Milchleistung.
Tabelle 55: Einflussfaktoren auf die Milchleistung von Sauen (Auswahl; Literaturübersicht)
Faktor
in der Literatur benannt bei
Laktationsstadium
KING et al. (1993B)
Ferkelanzahl KING et al. (1989); AULDIST und KING (1995);
TONER et al. (1996)
Körpermasse der Sau AULDIST und KING (1995)
Saugintensität der Ferkel AULDIST und KING (1995)
Stalltemperatur MULLAN et al. (1992)
Eiweiß- und Energieaufnahme mit
dem Futter
LENKEIT und GUETTE (1957); VERSTEGEN et al.
(1985); NOBLET und ETIENNE (1986); TOKACH et al.
(1992); KIRCHGESSNER et al. (1992); KING et al. (1993)
Um die tägliche Milchabgabe der Sauen zu erfassen, werden in der Literatur vier verschiedene
Methoden beschrieben:
1. Bestimmung der Milchleistung über die täglichen Zunahmen der Ferkel (LEWIS et al.,
1978; KING et al., 1989)
2. Ausmelken der Sau von Hand (HARTMANN et al., 1962) oder mit einer Melkmaschine
(SMITH et al., 1951; LODGE, 1957; HARTMAN und POND, 1960)

96
3. Isotopenverdünnungsmethode mit schwerem Wasser (YANG et al., 1980; PETTIGREW
et al., 1985)
4. Wiegen der Ferkel vor und nach dem Säugen: weigh-suckle-weigh Methode (LENKEIT
und GUETTE, 1955, 1955B, 1957; VAN SPAENDONCK und VAN SCHOUBROEK,
1964; RÄDER, 1990; SCHNEIDER, 1992; WESTERMEIER, 1997)
Aufgrund der großen Abweichungen bei der Bestimmung der Milchleistung über die täglichen
Zunahmen der Ferkel sowie den Schwierigkeiten beim Melken der Sau von Hand oder mit der
Maschine (HARTMANN et al., 1962) und bei der Verwendung der Isotopenverdünnungsmethode
wurde in vorliegendem Versuch die weigh-suckle-weigh Methode angewendet. Obwohl es sich dabei
um die mittlerweile am meisten angewendete Technik zur Abschätzung der Milchleistung der Sau
handelt (HARTMANN und HUGHES, 1996; ETIENNE et al., 1998), wurde noch keines der
Verfahren standardisiert. Die Ursache könnte darin liegen, dass noch verschiedene Punkte bei der
weigh-suckle-weigh Methode in der Literatur kritisch diskutiert werden. Die verwendete Methode
wurde von WOEHLBIER (1928) entwickelt, von RÄDER et al. (1990) modifiziert und
weiterentwickelt, sowie durch SCHNEIDER (1992) ergänzt. Dazu wurden die Ferkel während des
Messzeitraumes von der Sau abgetrennt und nur zum Säugen der Sau wieder zugesetzt (siehe: 2.5.3.).
Die Wiegungen wurden stündlich durchgeführt, da der natürliche Rhythmus zwischen den Säugungen
etwa 60 Minuten im Durchschnitt betrug. In der Literatur sind die in Tabelle 56 aufgeführten Werte zu
Zwischensäugeintervallen zu finden.
Tabelle 56: Literaturübersicht der Zwischensäugeintervalle bei Sauen
Quelle
Laktationstag
Minuten
BERGE und INDERBOE (1953)
1 bis 28
55 bis 65
BARBER et al. (1955) 6 und 51 51 und 63
NEWBERRY und WOOD-GUSH (1984) 1 bis 42 29 bis 78
AULDIST und KING (1994) 10 bis 24 48 bis 52
WECHSLER und BRODMANN (1996) 5 bis 14 52
SPINKA et al. (1997) 3 76
eigene Beobachtungen 12 bis 24 50 bis 70
Es ist zu erkennen, dass sich die beobachteten natürlichen Zwischensäugeintervalle in der Regel alle
im Bereich um 60 Minuten bewegen. Aus diesem Grund wurden die Messungen stündlich
durchgeführt. Während LODGE (1957) und BURGHART (1957) Messungen über 24 Stunden hinweg

97
durchführten, ist nach RÄDER et al. (1990) für eine ausreichend genaue Schätzung der
Tagesmilchleistung die Auswertung von 7 stündlich aufeinanderfolgenden Messungen genug. Dies
wurde auch im vorliegenden Versuch für ausreichend befunden. Insgesamt wurden jedoch mehr
Messungen vorgenommen, da SPEER und COX (1984) zeigten, dass die gemessenen Werte bei der
ersten Wiegung geringer ausfielen als der Durchschnitt der restlichen. Dies wird durch die
Eingewöhnungsphase verursacht, in welcher sich Sau und Ferkel erst an die Vorgehensweise
adaptieren müssen. Auch im vorliegenden Versuch kam es vor, dass sich Sauen bei der ersten
Datenerhebung oft gar nicht niederlegten oder verfrüht wieder aufsprangen und die Ferkel somit nicht
genug Zeit für eine sättigende Milchaufnahme hatten. Daher wurde die erste erfolgreiche Messung
nicht in die Auswertung miteinbezogen (siehe auch: 3.5.3.).
Ein weiteres Problem stellt die Aktivität der Ferkel dar, die während des Wiegevorganges große Fehler
durch die auftretenden Torsionskräfte bewirken kann, sowie die im Verhältnis zur Wurfmasse geringe
Milchaufnahme. Für die Wiegung der Würfe wurde daher eine elektronische Präzisionswaage (Modell
EB Economy-Serie, Fa. Sartorius, Göttingen) mit Mittelwertbildung von 20 Einzelwiegungen und
einer Anzeigegenauigkeit von ±1 g verwendet. Um den Wiegefehler auf ein vertretbares Maß zu
reduzieren reicht es nach den Untersuchungen von SCHNEIDER (1992), wenn man für die
Auswertung den Wiegevorgang 5-mal wiederholt (= Mittelwert aus 100 Einzelwiegungen). Zur
Erreichung eines noch geringeren Fehlers wurde im vorliegenden Versuch der Wiegevorgang 6-mal
wiederholt (= Mittelwert aus 120 Einzelwiegungen).
Um die Milchleistung der Sau so genau wie möglich bestimmen zu können, müssen darüber hinaus
alle Veränderungen der Wurfmasse, die nicht von der Milchaufnahme herrühren, erfasst und bei der
Berechnung entsprechend korrigiert werden. Dazu zählen Speichel-, Verdunstungs-, und
Stoffwechselverluste sowie abgesetzter Kot und Harn während der Säugephase. Werden diese
Korrekturen nicht in die Berechnung mit einbezogen, so verringert sich die ermittelte
Tagesmilchmenge (KLAVER et al., 1981; DEN HARTOG et al., 1984). SCHNEIDER (1992)
errechnete bei verschiedenen Autoren einen Anteil der Korrekturmassen von 7,5 % bis 29 % an der
Tagesmilchmenge. Daraus wird deutlich, dass eine Vernachlässigung dieser Größe zu einer nicht
unerheblichen Beeinflussung der ermittelten Milchmengen führt. In der vorliegenden Untersuchung
lag der Wert bei durchschnittlich 16 %. In Tabelle 57 sind die unterschiedlichen Ansätze zur Erfassung
der Masseverluste je Ferkel während des Säugens bei verschiedenen Autoren eingetragen.

98
Tabelle 57: Korrektur für Masseverluste der Ferkel während der Säugung in der Literatur
Quelle Harn Kot sonstige1)
KLAVER et al. (1981)
18,7 + 2,9 � LM0,75
nicht korrigiert
0,22 + 0,072 � t
DEN HARTOG et al. (1984) 19,4 + 5,3 � LM0,75 3,15 � 2,08 � t + 0,1 � t2 0,0443 + 0,019 � t
SPEER und COX (1984) 10 g 5 g nicht korrigiert
PETTIGREW et al. (1985) nicht korrigiert nicht korrigiert nach KLAVER et al. (1981)
NOBLET, ETIENNE (1986) durch Einzeltierrückwiegung bei Kot und Harnverlust 0,21 � LM0,75 � T
SCHNEIDER (1992) 15 g 15 g 0,418 + 1,15 � LM0,75
VERTESY (1996) 23 g 5 g nicht korrigiert
WESTERMEIER (1997) 18 g 7 g nach SCHNEIDER (1992)
OTT (2000) 21 g 5 g nach SCHNEIDER (1992)
vorliegende Untersuchung 1,23 + 0,0723 � W0,75 - 1,86 + 0,0132 � W0,75 nach SCHNEIDER (1992) 1) t LM0,75
W0,75
T
= = = = =
Speichel- , Verdunstungs- und Stoffwechselverluste Lebensalter des Ferkels in Tagen metabolische Lebendmasse des Ferkels in kg metabolische Lebendmasse des Ferkels in g Dauer der Säugephase in Minuten
Die Korrektur der Harn- und Kotverluste erfolgte bei der Mehrzahl der Autoren mit einem konstanten
Wert (SPEER und COX, 1984; SCHNEIDER, 1992; VERTESY, 1996; WESTERMEIER, 1997; OTT,
2000). KLAVER et al. (1981) führten beim Kot und PETTIGREW et al. (1985) bei Kot und Harn gar
keine Berichtigung durch. Die Einzeltierrückwaage bei NOBLET und ETIENNE (1986) stellt einen
Einzelfall mit hohem Arbeitsaufwand dar. Für eine Bereinigung der Verluste beim Koten und Harnen
in Abhängigkeit von der Lebendmasse beziehungsweise vom Lebensalters des Ferkels entschieden
sich KLAVER et al. (1981) und DEN HARTOG et al. (1984).
In der vorliegenden Arbeit wurde die Korrektur bei allen drei Paramatern in Abhängigkeit der
Ferkelmasse durchgeführt. Dabei handelt es sich bei Kot und Harn um selbst ermittelte Regressionen.
Die Regressionsgleichungen wurden trotz ihres geringen Bestimmtheitsmaßes (bei Kot: 0,21 und bei
Harn: 0,19) verwendet, da sie den Gesamtfehler bei der Milchleistungsschätzung geringer halten als
ein pauschaler Mittelwert für Kot und Harn. Die Einzeldaten zur Ableitung der Regression wurden bei
der Erfassung der Ferkeleinzelgewichte am 13./14. und am 20./21. Lebenstag gewonnen, wenn die
Ferkel während des Wiegens Kot oder Harn absetzten. Die Gleichung für die Speichel-, Verdunstungs-
und Stoffwechselverluste wurde von SCHNEIDER (1992) übernommen. Dieser sowie KLAVER et al.
(1981), DEN HARTOG et al. (1984) und NOBLET und ETIENNE (1986) führten eine
regressionsgestützte Korrektur durch, während bei anderen Autoren die Speichel-, Verdunstungs- und
Stoffwechselverluste unberücksichtigt blieben (SPEER und COX, 1984; VERTESY, 1996). Insgesamt
scheint eine Bereinigung der Milchleistung um Speichel-, Verdunstungs- und Stoffwechselverluste für

99
die Genauigkeit des Ergebnisses am wichtigsten zu sein, da sie bei dieser Arbeit im Durchschnitt einen
Anteil von mehr als 75 % an der Gesamtkorrektur ausmachten.
4.3.3.1. Milchmenge
Die gemessene tägliche Milchmenge betrug im Durchschnitt aller Gruppen und Messtage 7929 g.
Diese Leistung stimmt in etwa mit den bei KIRCHGESSNER et al. (1992), PAULICKS et al. (1998)
und OTT (2000) gefundenen Werten von 7841 g, 7420 g beziehungsweise 8692 g überein. Beim ersten
Messtermin am 13. und 14. Tag der Laktation war zwischen den Gruppen kein signifikanter
Unterschied in der Höhe der Milchleistung zu erkennen. Jedoch war die Milchmenge der Gruppe I mit
durchschnittlich 7100 g um 700g geringer als der Durchschnitt der restlichen Gruppen. Statistische
Unterschiede traten deshalb nicht auf, da die Standardabweichung innerhalb der Gruppen mit über
1100 g im Vergleich zu den absoluten Unterschieden relativ groß war. Beim zweiten Termin und bei
der Gesamtbetrachtung bewegten sich die Standardabweichungen in ähnlicher Höhe, jedoch waren die
Differenzen zwischen den Gruppen größer, so dass sich ein gesicherter Unterschied zwischen den
Gruppen I und IV von 2200 g feststellen ließ. Die Leistungen der übrigen Gruppen bewegten sich
dazwischen, jedoch bis zur Gruppe IV mit ansteigenden Werten und danach abfallend. Bei den
gemittelten Werten über die ganze Säugephase setzte sich dieser Trend fort, der jedoch nach
statistischen Gesichtspunkten nicht abzusichern war. Eine Ausnahme bildete hier der Wert der Gruppe
III, der jetzt niedriger war als der der Gruppe II, jedoch den der Gruppe I noch um 1000 g übertraf. In
Abbildung 21 sind die Mittelwerte über die gesamte Laktation mit den dazugehörigen
Standardabweichungen eingezeichnet.
tägliche Milchleistung der Sauen
5
6
7
8
9
10
0,07 0,12 0,17 0,22 0,27 0,32 0,37 0,42 0,47
Tryptophangehalt im Futter in %
Milc
hlei
stun
g in
kg/
Tag
Abbildung 21: Durchschnittliche tägliche Milchleistung der Sauen über die gesamte Laktation
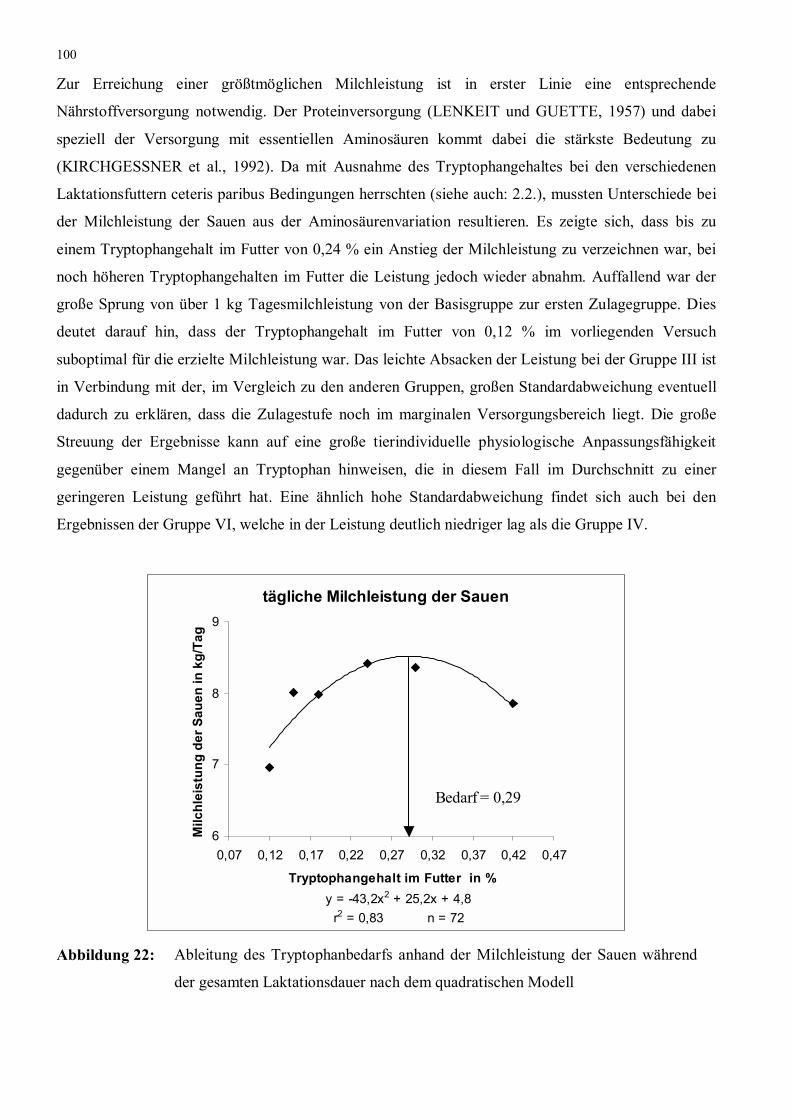
100
Zur Erreichung einer größtmöglichen Milchleistung ist in erster Linie eine entsprechende
Nährstoffversorgung notwendig. Der Proteinversorgung (LENKEIT und GUETTE, 1957) und dabei
speziell der Versorgung mit essentiellen Aminosäuren kommt dabei die stärkste Bedeutung zu
(KIRCHGESSNER et al., 1992). Da mit Ausnahme des Tryptophangehaltes bei den verschiedenen
Laktationsfuttern ceteris paribus Bedingungen herrschten (siehe auch: 2.2.), mussten Unterschiede bei
der Milchleistung der Sauen aus der Aminosäurenvariation resultieren. Es zeigte sich, dass bis zu
einem Tryptophangehalt im Futter von 0,24 % ein Anstieg der Milchleistung zu verzeichnen war, bei
noch höheren Tryptophangehalten im Futter die Leistung jedoch wieder abnahm. Auffallend war der
große Sprung von über 1 kg Tagesmilchleistung von der Basisgruppe zur ersten Zulagegruppe. Dies
deutet darauf hin, dass der Tryptophangehalt im Futter von 0,12 % im vorliegenden Versuch
suboptimal für die erzielte Milchleistung war. Das leichte Absacken der Leistung bei der Gruppe III ist
in Verbindung mit der, im Vergleich zu den anderen Gruppen, großen Standardabweichung eventuell
dadurch zu erklären, dass die Zulagestufe noch im marginalen Versorgungsbereich liegt. Die große
Streuung der Ergebnisse kann auf eine große tierindividuelle physiologische Anpassungsfähigkeit
gegenüber einem Mangel an Tryptophan hinweisen, die in diesem Fall im Durchschnitt zu einer
geringeren Leistung geführt hat. Eine ähnlich hohe Standardabweichung findet sich auch bei den
Ergebnissen der Gruppe VI, welche in der Leistung deutlich niedriger lag als die Gruppe IV.
tägliche Milchleistung der Sauen
y = -43,2x2 + 25,2x + 4,8r2 = 0,83 n = 72
6
7
8
9
0,07 0,12 0,17 0,22 0,27 0,32 0,37 0,42 0,47
Tryptophangehalt im Futter in %
Milc
hlei
stun
g de
r Sau
en in
kg/
Tag
Bedarf = 0,29
Abbildung 22: Ableitung des Tryptophanbedarfs anhand der Milchleistung der Sauen während
der gesamten Laktationsdauer nach dem quadratischen Modell

101
Ein optimaler Tryptophangehalt im Futter scheint demnach, in Hinblick auf die Milchleistung, im
Bereich der Gruppen IV und V mit 0,24 % beziehungsweise 0,30 % Tryptophan im Futter zu liegen.
Die Ableitung nach dem quadratischen Modell ergab einen Bedarf von 0,29 % (siehe Abb. 22).
4.3.3.2. Trockenmasse der Sauenmilch
Beim vorliegenden Versuch konnte kein Einfluss einer Tryptophanzulage im Laktationsfutter auf den
Trockensubstanzgehalt in der Sauenmilch festgestellt werden. Die durchschnittliche Festsubstanz in
der Milch lag bei 19,2 %, und reihte sich wie aus der Tabelle 58 ersichtlich wird sehr gut in die von
anderen Autoren in der Literatur aufgeführten Werte ein. Zwischen den Behandlungen traten keine
Unterschiede auf.
Tabelle 58: Literaturangaben zum Trockensubstanzgehalt der Sauenmilch
Quelle
Laktationstag
TS in%
Laktationstag
TS in%
KLOBASA et al. (1987)
14
18,2
21
18,7
VÈRTESY (1996) 13/14 18,6 20/21 18,2
CSAPÓ et al. (1996) 10 19,7 20 18,6
PLUSKE et al. (1998) 10-15 19,2 21-25 18,6
vorliegende Untersuchung 20/21 19,2
Nach Untersuchungen von KLOBASA et al. (1987) und CSAPÒ et al. (1996) fällt der
Trockensubstanzgehalt von etwa 25 % zum Geburtszeitpunkt in den ersten zwei Laktationswochen auf
etwa 18 � 19 % ab. Bis zum Ende der vierten Laktationswoche bleibt dieser Wert in etwa erhalten.
4.3.3.3. Fettgehalt
Tabelle 59 zeigt die von verschiedenen Autoren ermittelten Milchfettgehalte in Abhängigkeit des
Laktationstages. In der Literatur werden Werte von 5,4 % (SCHOENHERR et al., 1989) bis 8,5 %
(SCHNEIDER, 1992 und OTT, 2000) aufgeführt. KLOBASA et al. (1987) fanden, dass der
Milchfettgehalt in den ersten 72 Stunden nach der Geburt um etwa 1/3 ansteigt, dann bis zum 21.
Laktationstag nahezu konstant bleibt und danach mit zunehmender Säugedauer wieder abfällt.
Tendenziell fällt der Milchfettgehalt von der 2. auf die 3. Laktationswoche in allen aufgeführten
Arbeiten mit Ausnahme eines Ergebnisses bei HAYDON et al. (1990). Im vorliegenden Versuch war
ebenfalls ein signifikantes Abfallen des Milchfettgehaltes von der 2. auf die 3. Säugewoche um
durchschnittlich 0,6 Prozentpunkte von 8,2 % auf 7,6 % festzustellen. In der Literatur sind über die
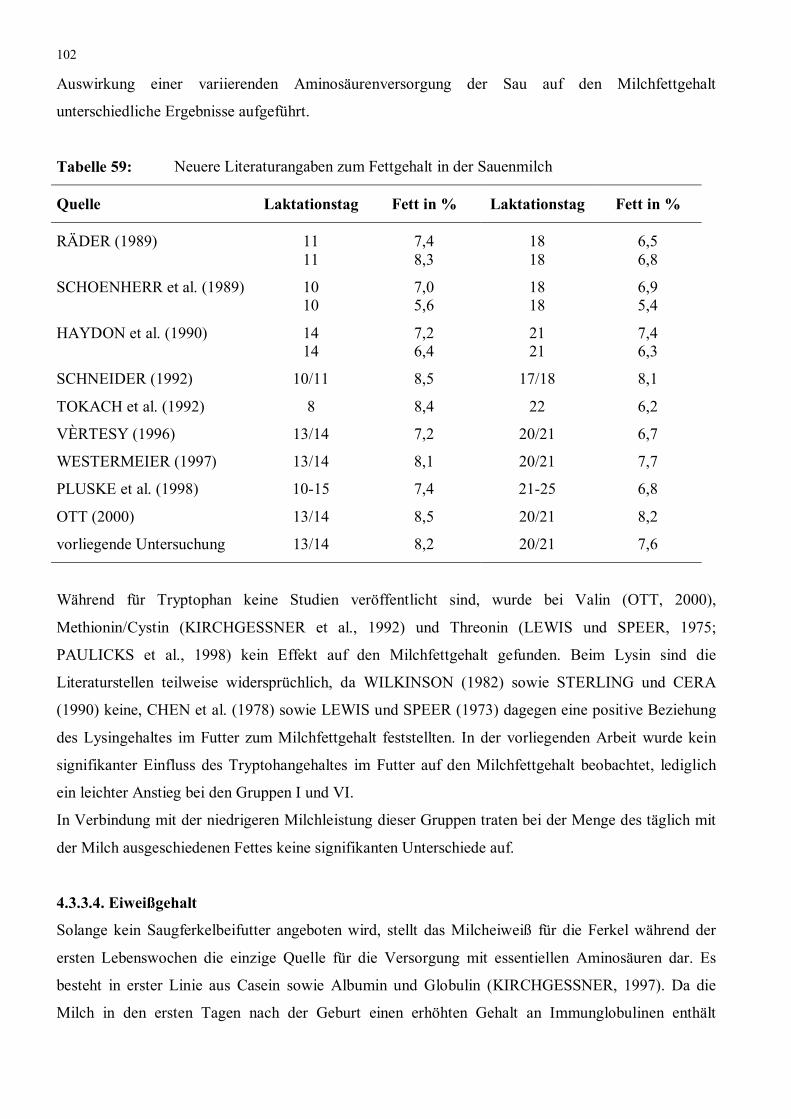
102
Auswirkung einer variierenden Aminosäurenversorgung der Sau auf den Milchfettgehalt
unterschiedliche Ergebnisse aufgeführt.
Tabelle 59: Neuere Literaturangaben zum Fettgehalt in der Sauenmilch
Quelle
Laktationstag
Fett in %
Laktationstag
Fett in %
RÄDER (1989)
11 11
7,4 8,3
18 18
6,5 6,8
SCHOENHERR et al. (1989)
10 10
7,0 5,6
18 18
6,9 5,4
HAYDON et al. (1990)
14 14
7,2 6,4
21 21
7,4 6,3
SCHNEIDER (1992)
10/11
8,5
17/18
8,1
TOKACH et al. (1992) 8 8,4 22 6,2
VÈRTESY (1996) 13/14 7,2 20/21 6,7
WESTERMEIER (1997) 13/14 8,1 20/21 7,7
PLUSKE et al. (1998) 10-15 7,4 21-25 6,8
OTT (2000) 13/14 8,5 20/21 8,2
vorliegende Untersuchung 13/14 8,2 20/21 7,6
Während für Tryptophan keine Studien veröffentlicht sind, wurde bei Valin (OTT, 2000),
Methionin/Cystin (KIRCHGESSNER et al., 1992) und Threonin (LEWIS und SPEER, 1975;
PAULICKS et al., 1998) kein Effekt auf den Milchfettgehalt gefunden. Beim Lysin sind die
Literaturstellen teilweise widersprüchlich, da WILKINSON (1982) sowie STERLING und CERA
(1990) keine, CHEN et al. (1978) sowie LEWIS und SPEER (1973) dagegen eine positive Beziehung
des Lysingehaltes im Futter zum Milchfettgehalt feststellten. In der vorliegenden Arbeit wurde kein
signifikanter Einfluss des Tryptohangehaltes im Futter auf den Milchfettgehalt beobachtet, lediglich
ein leichter Anstieg bei den Gruppen I und VI.
In Verbindung mit der niedrigeren Milchleistung dieser Gruppen traten bei der Menge des täglich mit
der Milch ausgeschiedenen Fettes keine signifikanten Unterschiede auf.
4.3.3.4. Eiweißgehalt
Solange kein Saugferkelbeifutter angeboten wird, stellt das Milcheiweiß für die Ferkel während der
ersten Lebenswochen die einzige Quelle für die Versorgung mit essentiellen Aminosäuren dar. Es
besteht in erster Linie aus Casein sowie Albumin und Globulin (KIRCHGESSNER, 1997). Da die
Milch in den ersten Tagen nach der Geburt einen erhöhten Gehalt an Immunglobulinen enthält

103
(KLOBASA et al., 1987), führt dies zu erhöhten Proteinwerten von bis zu 19 % im Kolostrum
(KIRCHGESSNER, 1997). Durch die Verringerung der Immunglobulinkonzentration nach den ersten
Säugetagen der Ferkel fällt die Eiweißkonzentration jedoch schnell (NOBLET und ETIENNE, 1986;
KLOBASA et al., 1987; CSAPO et al., 1996) auf einen Gehalt von etwa 4 � 5 % in der normalen
Milch. Etwa 97 % des in der Milch enthaltenen Stickstoffs entfällt auf in Proteinen gebundene
Aminosäuren. Der restliche nicht eiweißgebundene Stickstoff verteilt sich auf den Milchharnstoff
sowie die in freier Form vorliegenden Aminosäuren (KLOBASA et al., 1987). In der Tabelle 60 sind
neuere Literaturwerte für den Eiweißgehalt in normaler Sauenmilch aufgeführt.
Tabelle: 60: Neuere Literaturangaben zum Proteingehalt in der Sauenmilch
Quelle
Laktationstag
Protein in %
Laktationstag
Protein in %
RÄDER (1989)
11 11
4,5 4,5
18 18
4,7 4,3
SCHOENHERR et al. (1989)
10 10
4,8 4,7
18 18
4,5 4,6
HAYDON et al. (1990)
14 14
5,0 4,6
21 21
4,4 4,6
SCHNEIDER et al. (1992B)
10/11
4,4
17/18
4,2
TOKACH et al. (1992) 8 5,2 22 5,0
VÈRTESY (1996) 13/14 4,4 20/21 4,6
PAULICKS et al. (1998) 13/14 4,4 20/21 4,1
PLUSKE et al. (1998) 10-15 5,1 21-25 5,2
OTT (2000) 13/14 4,4 20/21 4,6
vorliegende Untersuchung 13/14 4,7 20/21 4,7
Wie aus Tabelle 60 ersichtlich ist, werden für den Eiweißgehalt in der Sauenmilch Werte von 4,1 %
(PAULICKS et al., 1998) bis 5,2 % (TOCKACH et al., 1992; PLUSKE et al., 1998) angegeben, die
von der 2. auf die 3. Laktationswoche in etwa gleich bleiben. Die in der vorliegenden Untersuchung
ermittelten Werte von durchschnittlich 4,7 % Milchprotein weichen somit nicht von den
Literaturwerten ab. Innerhalb der verschiedenen Futtergruppen waren keine signifikanten Unterschiede
beim Eiweißgehalt und auch kein gerichteter Behandlungseinfluss zu erkennen. Auch zwischen den
Probenahmeterminen trat keine Differenz auf. Somit zeigte die Tryptophanzulage im Laktationsfutter
keinerlei Einfluss auf den Milcheiweißgehalt.
Bezüglich der Wirkung steigender Aminosäurezulagen im Futter liegen in der Literatur
unterschiedliche Versuchsergebnisse vor. Bei Lysin beobachteten LEWIS und SPEER (1973) sowie
104
CHEN et al. (1978) bei zunehmender Versorgung einen Anstieg der produzierten Eiweißmenge,
während STERLING und CERA (1990) keinen Einfluss feststellen konnten. LEWIS und SPEER
(1975) sowie PAULICKS et al. (1998) konnten keine Auswirkung bei einer Threoninzulage im Futter
auf den Proteingehalt der Sauenmilch feststellen. SCHNEIDER et al. (1992B) ermittelten bei
Methionin und OTT (2000) sowie ROUSSELOW und SPEER (1980) bei Valin ansteigende
Milchproteingehalte bis zum Bedarfswert der Aminosäure im Futter. Jedoch erreichten RICHERT et
al. (1997) und MOSER et al. (2000) bei zunehmender Valinkonzentration im Futter keine
Veränderung beim Milcheiweiß. Bei Isoleucin fanden RICHERT et al. (1997) einen positiven Effekt
einer Zulage auf den Proteingehalt in der Milch. Da im vorliegenden Versuch keine nennenswerten Unterschiede beim Eiweißgehalt in der Milch
auftraten, spiegelten die täglichen Proteinausscheidungen die Verhältnisse der Milchleistung wider.
4.3.3.5. Laktosegehalt
Laktose stellt den überwiegenden Teil der in der Milch enthaltenen Kohlenhydrate (HARTMANN und
HOLMES, 1989). Andere Kohlenhydrate sind nur in sehr geringen Mengen vorhanden. Sie sind aus
Glucose und Galaktose, den Bestandteilen der Lactose, und Oligosacchariden aufgebaut (BOYD und
KENSINGER, 1998). Nach Untersuchungen von PERRIN (1955) und KLOBASA (1987) steigt der
Laktosegehalt in den ersten drei Tagen nach der Geburt um 80 % beziehungsweise 70 % an und bleibt
zwischen der zweiten und dritten Laktationswoche gleich.
Tabelle 61: Literaturangaben zum Laktosegehalt in der Sauenmilch
Quelle
Laktationstag
Laktose in %
Laktationstag
Laktose in %
KLAVER et al. (1981)
5
5,3
NOBLET u. ETIENNE (1986) 1-21 5,4
DEN HARTOG et al. (1989) 12 5,2 26 5,2
KLOBASA et al. (1987) 14 5,9 21 5,8
FOWLER und GILL (1989) 7 4,9
RÄDER et al. (1990) 5-22 5,6
TOKACH et al. (1992) 8 5,2 22 5,4
VÈRTESY (1996) 13/14 5,7 20/21 5,7
REVELL et al. (1998B) 4-6
4-6
5,2
5,5
25-27
25-27
5,3
5,4
PLUSKE et al. (1998) 10-15 5,5 21-25 5,6
vorliegende Untersuchung 13/14 5,7 20/21 5,8

105
Tabelle 61 führt Literaturstellen auf, in denen Laktosegehalte in der Sauenmilch benannt sind. Die
Werte zwischen der zweiten und dritten Woche weisen kaum Schwankungen auf und die in der
vorliegenden Arbeit gefundenen Werte stimmen gut mit denen der anderen Autoren überein. Laktose
ist der primär osmotisch wirkende Bestandteil der Milch (MACKENZIE und REVELL, 1998). Der
osmotische Druck der Milch ist gleich groß wie der anderer Körperflüssigkeiten (MACKENZIE und
REVELL, 1998). Daher muss ein eventueller Gradient zwischen Blut und Milch mit dem Übergang
von Wasser durch das semipermeable Milchdrüsenepithel ausgeglichen werden. Somit wird die
Milchmenge im wesentlichen durch die produzierte Menge an Laktose und damit durch die Menge an
aufgenommenem Wasser in der Milchdrüse bestimmt (PEAKER, 1977; 1978).
In der Literatur sind einige wenige Untersuchungen zur Auswirkung variierender Eiweißversorgungen
auf den Laktosegehalt bei Sauen zu finden. So fanden TOKACH et al. (1992) eine positive Korrelation
zwischen dem Lysingehalt im Futter und dem Milchzuckergehalt. In der vorliegenden Untersuchung
wurde jedoch kein Einfluss des Tryptophangehaltes im Laktationsfutter auf den Laktosegehalt in der
Milch festgestellt.
4.3.3.6. Harnstoffgehalt
Werden dem Organismus über das Futter mehr Eiweiß beziehungsweise Aminosäuren zugeführt als
für die Proteinsynthese benötigt werden, führt dies zu einem verstärkten Proteinabbau. Der dabei
anfallende Ammoniak wird durch die verstärkte Synthese von Harnstoff gebunden und somit in eine
für den Organismus unschädliche Form überführt und mit dem Harn oder der Milch ausgeschieden
(NELSON und COX, 2001). Anhand des Milchharnstoffgehaltes wird bei der Kuh schon seit längerem
auf die Proteinversorgung mit dem Futter geschlossen. Diese Zusammenhänge wurden von
PAULICKS (1985), KIRCHGESSNER und KREUZER (1985), KIRCHGESSNER et al. (1987) und
KIRCHGESSNER (1997) bereits eingehend beschrieben. Da es auch beim Schwein unter einer
variierenden Eiweißversorgung zu dosisabhängigen Schwankungen des Blutharnstoffgehaltes kommt
(PAULICKS et al., 1998B) und dieser auch von der Milchdrüse des Schweines absorbiert werden kann
(BOLDUAN und SCHNEIDER, 1986), besteht ein positiver Zusammenhang zwischen dem
Harnstoffgehalt im Blut und dem in der Milch. Aus den bei OTT (2000) aufgeführten Daten ließ sich
hierzu ein Korrelationskoeffizient von r = 0,79 (eigene Berechnung) ermitteln und BOLDUAN et al.
(1989) berichten von einer Korrelation mit r = 0,71. Der Zusammenhang im vorliegenden Versuch war
mit r = 0,97 sehr deutlich. Bei PAULICKS et al. (1998B) war mit r = 0,98 (eigene Berechnung)
ebenfalls eine enge Korrelation zwischen Blut- und Milchharnstoffkonzentration feststellbar. In der
Literatur sind nicht viele Ergebnisse zu Harnstoffgehalten in der Sauenmilch aufgeführt.
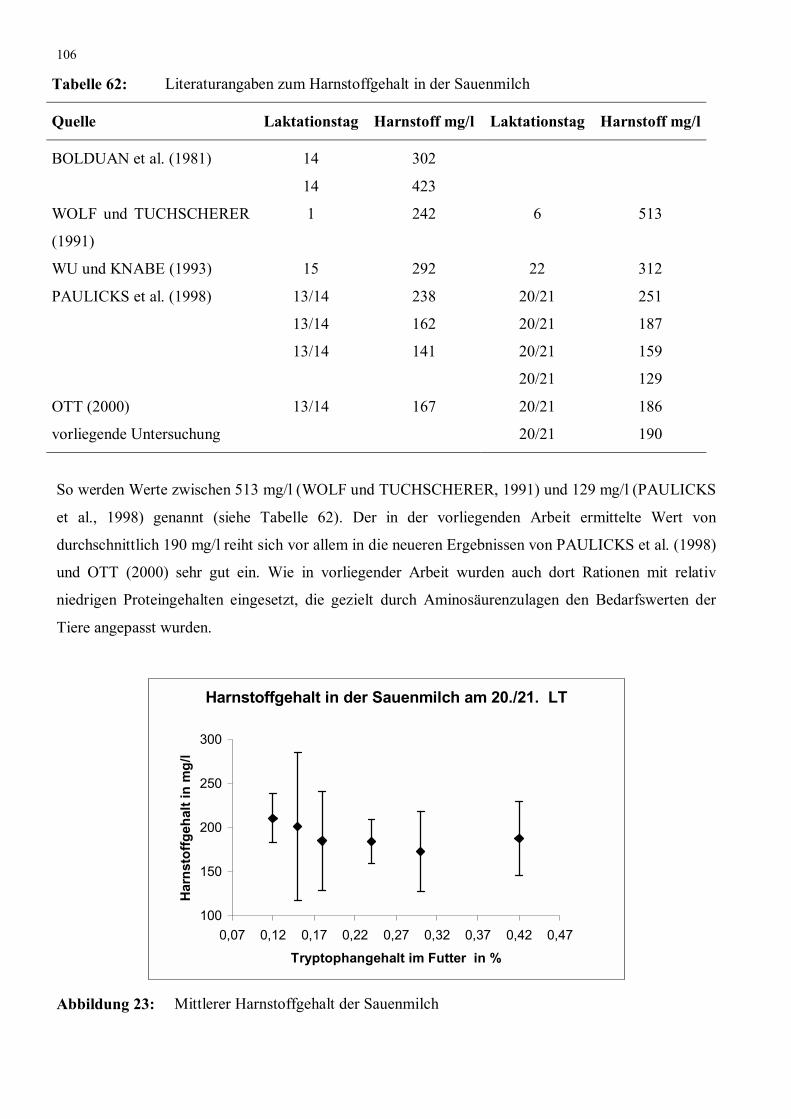
106
Tabelle 62: Literaturangaben zum Harnstoffgehalt in der Sauenmilch
Quelle
Laktationstag
Harnstoff mg/l
Laktationstag
Harnstoff mg/l
BOLDUAN et al. (1981)
14
14
302
423
WOLF und TUCHSCHERER
(1991)
1 242 6 513
WU und KNABE (1993) 15 292 22 312
PAULICKS et al. (1998) 13/14
13/14
13/14
238
162
141
20/21
20/21
20/21
20/21
251
187
159
129
OTT (2000) 13/14 167 20/21 186
vorliegende Untersuchung 20/21 190
So werden Werte zwischen 513 mg/l (WOLF und TUCHSCHERER, 1991) und 129 mg/l (PAULICKS
et al., 1998) genannt (siehe Tabelle 62). Der in der vorliegenden Arbeit ermittelte Wert von
durchschnittlich 190 mg/l reiht sich vor allem in die neueren Ergebnissen von PAULICKS et al. (1998)
und OTT (2000) sehr gut ein. Wie in vorliegender Arbeit wurden auch dort Rationen mit relativ
niedrigen Proteingehalten eingesetzt, die gezielt durch Aminosäurenzulagen den Bedarfswerten der
Tiere angepasst wurden.
Harnstoffgehalt in der Sauenmilch am 20./21. LT
100
150
200
250
300
0,07 0,12 0,17 0,22 0,27 0,32 0,37 0,42 0,47
Tryptophangehalt im Futter in %
Har
nsto
ffgeh
alt i
n m
g/l
Abbildung 23: Mittlerer Harnstoffgehalt der Sauenmilch

107
In der Abbildung 23 sind die mittleren Harnstoffgehalte in der Sauenmilch am 20./21. Laktationstag
dargestellt. Dabei ist ein Abfallen der Konzentration von 211 mg/l bei Gruppe I auf 173 mg/l bei der
Gruppe V zu erkennen. Danach steigt der Harnstoffgehalt bei Gruppe VI wieder an. Auffallend sind
die großen Unterschiede bei den Standardabweichungen, die jedoch auch schon in der Arbeit von OTT
(2000) beobachtet wurden.
Harnstoffgehalt in der Sauenmilch am 20./21. LT
y = 1026x2 - 627x + 271r2 = 0,93 n = 72
150
175
200
225
0,07 0,12 0,17 0,22 0,27 0,32 0,37 0,42 0,47
Tryptophangehalt im Futter in %
Har
nsto
ffgeh
alt i
n m
g/l
Bedarf = 0,31
Abbildung 24: Ableitung des Tryptophanbedarfs anhand der Harnstoffgehalte in der Sauenmilch
am 20./21. LT nach dem quadratischen Modell
Wie aus der Abbildung 24 hervorgeht, ergibt die Ableitung des Tryptophanbedarfs laktierender Sauen
aufgrund des Milchharnstoffgehaltes nach dem quadratischen Modell 0,31 % Tryptophan im Futter.
Bei diesem Tryptophangehalt im Futter ist davon auszugehen, dass die Relation von Tryptophan zu
den anderen essentiellen Aminosäuren optimal ist. Der im Falle eines Tryptophanmangels bei der
Proteinsynthese entstehende Überhang bei den restlichen Aminosäuren müsste letztendlich über
Harnstoff ins Plasma und von da in die Milch ausgeschieden werden. Mit zunehmendem
Tryptophangehalt verringert sich die Notwendigkeit andere Eiweißbausteine abzubauen und somit
sinkt der Harnstoffgehalt im Blut und damit in der Milch. Erreicht der Harnstoffgehalt im Plasma und
somit auch in der Milch sein Minimum, so ist die Relation von Tryptophan zu den restlichen
essentiellen Aminosäuren am besten. Da sich bei einer weiteren Zulage von Tryptophan dieses nach
Überschreiten des optimalen Verhältnisses zu den anderen essentiellen Aminosäuren im Überschuss
befindet, wird es verstärkt abgebaut, was wiederum zu einem Anstieg des Harnstoffgehaltes im

108
Plasma und letztendlich auch in der Milch führt. Der Anstieg des Harnstoffgehaltes ist dabei
erwartungsgemäß jedoch nicht so stark, da die Gesamtmenge an Eiweiß, die metabolisiert werden
muss vor Erreichen des Optimums größer ist als bei einer Überschreitung. Die Zusammenhänge sind
bei anderen essentiellen Aninosäuren ähnlich. Davon berichten auch PAULICKS et al. (1998) bei
Threonin und OTT (2000) bei Valin.
Bei den täglich über die Milch ausgeschiedenen absoluten Mengen an Harnstoff waren zwischen den
Gruppen keine signifikanten Unterschiede zu beobachten. Dies ist auf die Tatsache zurückzuführen,
dass die Gruppen mit geringer Milchleistung einen hohen Harnstoffgehalt aufwiesen und umgekehrt.
Bei der Berechnung der täglichen Ausscheidung waren somit die Werte ausgeglichen. Die mittlere
tägliche Harnstoffausscheidung mit der Milch betrug 1553 mg.
4.3.3.7. Energiegehalt
Aus den Werten der Fett- und Eiweißgehalte in der Sauenmilch wurde nach der Methode von
KIRCHGESSNER et al. (1992) der Energiegehalt rechnerisch ermittelt. Im Mittel aller Messungen
betrug dieser 5,12 MJ/kg FS. Nach COFFEY et al. (1982) fällt der Energiegehalt der Sauenmilch von
6,3 MJ/kg FS post partum auf 4,7 MJ/kg FS am 21. Laktationstag ab. Dies wird auch von NOBLET
und ETIENNE (1986) und DEN HARTOG et al. (1987) berichtet. In der vorliegenden Arbeit war der
Gehalt am 14. Laktationstag ebenfalls etwas höher als eine Woche später.
Tabelle 63: Neuere Literaturangaben zum Energiegehalt in der Sauenmilch
Quelle
Laktationstag
Energie MJ/kg
Laktationstag
Energie MJ/kg
RÄDER (1989)
11 11
5,2 5,3
18 18
5,1 5,1
SCHOENHERR et al.
(1989)1)
10 10
4,8 4,3
18 18
4,7 4,2
HAYDON et al. (1990) 1)
14 14
5,0 4,6
21 21
4,9 4,6
SCHNEIDER (1992)
10/11
5,3
17/18
4,9
TOKACH et al. (1992)1) 8 5,4 22 4,6
VÈRTESY (1996) 13/14 4,8 20/21 4,7
WESTERMAYER (1997) 13/14 5,1 20/21 4,9
OTT (2000) 13/14 5,2 20/21 5,1
vorliegende Untersuchung 13/14 5,2 20/21 5,0 1) = errechnet aus Fett- und Eiweißgehalt (nach KIRCHGESSNER et al., 1992); in der Arbeit nicht vermerkt

109
Betrachtet man die in der Tabelle 63 aufgeführten Werte neuerer Untersuchungen zum
Milchenergiegehalt, so sind sie mit den gefundenen Ergebnissen übereinstimmend. Zwischen den
Gruppen war wie im vorliegenden Versuch kein signifikanter Unterschied zu erkennen. Bei
Forschungen anderer Autoren mit den Aminosäuren Methionin (KIRCHGESSNER et al., 1992),
Threonin (PAULICKS et al., 1998) und Valin (OTT, 2000) konnte ebenfalls keine Auswirkung einer
Zulage im Futter auf die Energiekonzentration beobachtet werden. Im Gegensatz zu STERLING und
CERA (1990) fanden CHEN et al. (1978) mit zunehmendem Lysingehalt einen Anstieg der
Milchenergie.
Die mit der Milch täglich ausgeschiedenen Energiemengen spiegelten aufgrund der geringen
Gehaltsunterschiede die Verhältnisse bei den Tagesmilchmengen wider.
4.3.3.8. Aminosäurenzusammensetzung
Bei den in der Sauenmilch enthaltenen Aminosäuren kann grundsätzlich zwischen den im Protein
gebundenen und den freien Aminosäuren unterschieden werden (BENGTSSON, 1972; WU und
KNABE, 1994). Der Gehalt an freien Aminosäuren steigt dabei nach WU und KNABE (1994) vom 1.
bis zum 29. Laktationstag kontinuierlich von 0,3 auf 4,6 % an. Insgesamt werden also die meisten aus
dem Blut absorbierten Aminosäuren für die Synthese des Milcheiweißes verwendet. In der
vorliegenden Arbeit wurde nur der Gesamtgehalt der Aminosäuren betrachtet.
Das Aminosäurenmuster des Proteins in der Normalmilch wird durch die Fütterung nicht beeinflusst
(BEYER, 1986) und bleibt im Verlauf der Laktation konstant (WU und KNABE, 1994). Lediglich
zwischen der Kolostralmilch und der Normalmilch ist aufgrund der verschiedenen
Proteinzusammensetzung (siehe auch 4.3.3.3.) auch ein Unterschied im Aminosäurenmuster zu
beobachten (BEYER, 1986; WU und KNABE, 1994). Auch bei Versuchen mit unterschiedlicher
Aminosäurenversorgung wurden keine Auswirkungen auf die Zusammensetzung der Aminosäuren in
der Sauenmilch bei Methionin/Cystin (SCHNEIDER et al., 1992B), Threonin (WESTERMEIER,
1997) und Valin (OTT, 2000) beobachtet. Das Aminosäurenmuster in der Normalmilch scheint
demnach genetisch festgelegt zu sein. Bei der Untersuchung aller Gruppen hinsichtlich des
Tryptophangehaltes in der Milch ergaben sich keine signifikanten Differenzen. Aufgrund dieser
Tatsachen und des Umstandes, dass sich die Werte der Gruppen I, III und VI auch im vorliegenden
Versuch bei den restlichen Aminosäuren nicht signifikant unterschieden, wurde auf eine Untersuchung
der anderen Gruppen verzichtet und der Mittelwert für den vorliegenden Versuch in der Tabelle 64
eingetragen.
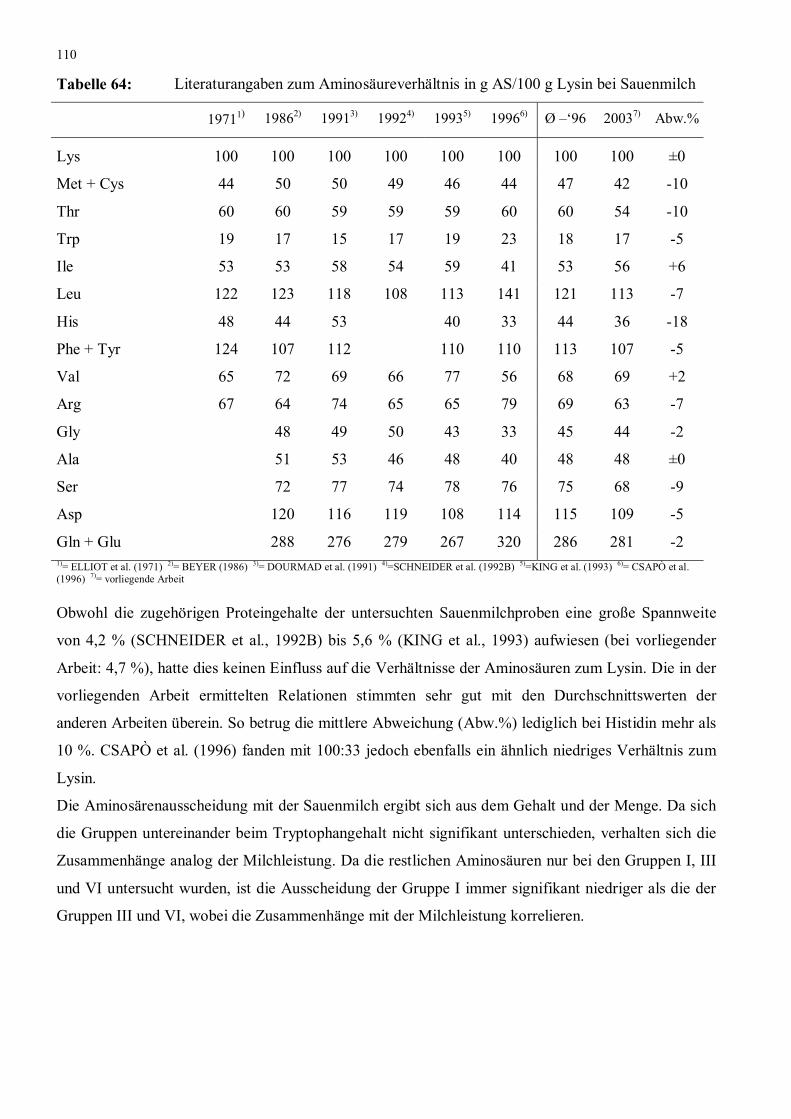
110
Tabelle 64: Literaturangaben zum Aminosäureverhältnis in g AS/100 g Lysin bei Sauenmilch
19711)
19862)
19913)
19924)
19935)
19966)
Ø ��96
20037)
Abw.%
Lys
100
100
100
100
100
100
100
100
±0
Met + Cys 44 50 50 49 46 44 47 42 -10
Thr 60 60 59 59 59 60 60 54 -10
Trp 19 17 15 17 19 23 18 17 -5
Ile 53 53 58 54 59 41 53 56 +6
Leu 122 123 118 108 113 141 121 113 -7
His 48 44 53 40 33 44 36 -18
Phe + Tyr 124 107 112 110 110 113 107 -5
Val 65 72 69 66 77 56 68 69 +2
Arg 67 64 74 65 65 79 69 63 -7
Gly 48 49 50 43 33 45 44 -2
Ala 51 53 46 48 40 48 48 ±0
Ser 72 77 74 78 76 75 68 -9
Asp 120 116 119 108 114 115 109 -5
Gln + Glu 288 276 279 267 320 286 281 -2 1)= ELLIOT et al. (1971) 2)= BEYER (1986) 3)= DOURMAD et al. (1991) 4)=SCHNEIDER et al. (1992B) 5)=KING et al. (1993) 6)= CSAPÒ et al. (1996) 7)= vorliegende Arbeit Obwohl die zugehörigen Proteingehalte der untersuchten Sauenmilchproben eine große Spannweite
von 4,2 % (SCHNEIDER et al., 1992B) bis 5,6 % (KING et al., 1993) aufwiesen (bei vorliegender
Arbeit: 4,7 %), hatte dies keinen Einfluss auf die Verhältnisse der Aminosäuren zum Lysin. Die in der
vorliegenden Arbeit ermittelten Relationen stimmten sehr gut mit den Durchschnittswerten der
anderen Arbeiten überein. So betrug die mittlere Abweichung (Abw.%) lediglich bei Histidin mehr als
10 %. CSAPÒ et al. (1996) fanden mit 100:33 jedoch ebenfalls ein ähnlich niedriges Verhältnis zum
Lysin.
Die Aminosärenausscheidung mit der Sauenmilch ergibt sich aus dem Gehalt und der Menge. Da sich
die Gruppen untereinander beim Tryptophangehalt nicht signifikant unterschieden, verhalten sich die
Zusammenhänge analog der Milchleistung. Da die restlichen Aminosäuren nur bei den Gruppen I, III
und VI untersucht wurden, ist die Ausscheidung der Gruppe I immer signifikant niedriger als die der
Gruppen III und VI, wobei die Zusammenhänge mit der Milchleistung korrelieren.

111
4.3.4. Blutparameter der Sauen
Im Vergleich zu Ergebnissen, die nur mit einem sehr großen Personal- und Technikaufwand zu
gewinnen sind (zum Beispiel die Milchleistungserfassung bei der Sau mit der weigh-suckle-weigh
Methode) stellt die Gewinnung von Blutproben kein großes Problem dar. Aus diesem Grund wurden in
sehr vielen Versuchen Kenngrößen, die aus dem Blut messbar sind, für die Auswertung herangezogen.
Bei Untersuchungen zum Bedarf von Aminosäuren wurden besonders die Konzentrationen der
einzelnen Aminosäuren und des Harnstoffs als Hilfsmittel verwendet (LEWIS und SPEER, 1973,
1974, 1975; CHEN et al., 1978; SOHAIL et al., 1978; ROUSSELOW et al., 1979; ROUSSELOW und
SPEER, 1980; WILKINSON et al., 1982; LELLIS und SPEER, 1985; KIRCHGESSNER et al., 1993;
RICHERT et al., 1996; KIRCHGESSNER et al., 1998).
4.3.4.1. Aminosäurengehalt im Plasma
Die Analyse der Aminosäurenkonzentrationen im Blutplasma der Sauen erfolgte am 22. Laktationstag.
Die Aminosäuren im Blut werden als sogenannter Pool bezeichnet, in welchen sie nach der Verdauung
eingespeist werden. Von dort aus werden sie für die einzelnen Stoffwechselvorgänge herangezogen.
4.3.4.1.1 Tryptophan
Mit steigender alimentärer Eiweißaufnahme steigt die Aminosäurenkonzentration im Blut. Nach
WURTMAN (1981) wird ein großer Teil des Tryptophans, das aus dem Portalblut absorbiert wird, in
der Leber metabolisiert. Somit vermag die Leber große Schwankungen bei der
Tryptophankonzentration des Pfortaderblutes teilweise abzupuffern (LÖFFLER und PETRIDES,
1997). HYLDGAARD-JENSEN (1976) schreibt, dass sich eine Erhöhung des Tryptophangehaltes im
Futter klar beim Tryptophangehalt sowohl im Pfortaderblut, als auch im peripheren Blutkreislauf
widerspiegelt. Jedoch ist die Auswirkung auf das Pfortaderblut schneller und stärker. Die Blutproben
wurden bei der vorliegenden Arbeit aus versuchstechnischen Gründen nur aus dem peripheren
Blutkreislauf (vena jugularis externa) gezogen. Aufgrund der oben beschriebenen Zusammenhänge
wurde das Plasma erst 3 Stunden nach der Fütterung genommen, um die zeitverzögerte
Konzentrationsschwankung (HYLDGAARD-JENSEN, 1976) dennoch erfassen zu können. Abbildung
25 gibt die Ergebnisse der Tryptophangehalte im Sauenplasma in Abhängigkeit der
Tryptophanaufnahme mit dem Futter in der 3. Laktationswoche wieder.
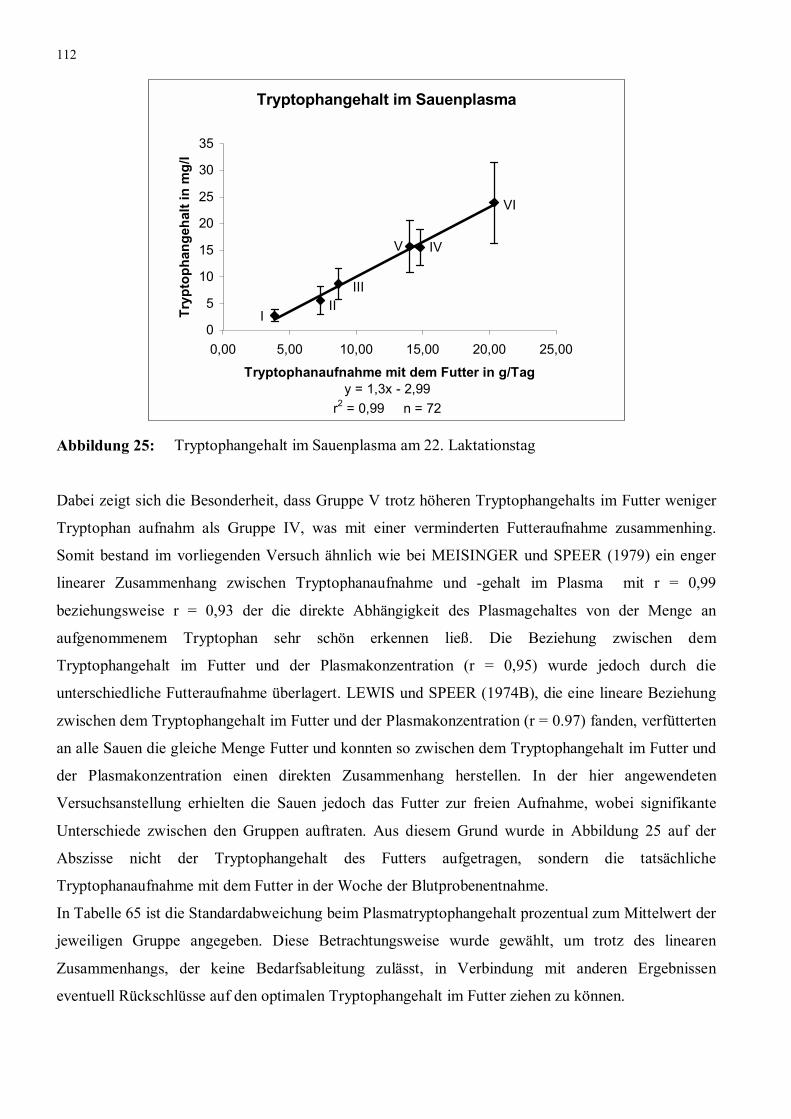
112
Tryptophangehalt im Sauenplasma
VI
V IV
IIIII
I
y = 1,3x - 2,99r2 = 0,99 n = 72
0
5
10
15
20
25
30
35
0,00 5,00 10,00 15,00 20,00 25,00
Tryptophanaufnahme mit dem Futter in g/Tag
Tryp
toph
ange
halt
in m
g/l
Abbildung 25: Tryptophangehalt im Sauenplasma am 22. Laktationstag
Dabei zeigt sich die Besonderheit, dass Gruppe V trotz höheren Tryptophangehalts im Futter weniger
Tryptophan aufnahm als Gruppe IV, was mit einer verminderten Futteraufnahme zusammenhing.
Somit bestand im vorliegenden Versuch ähnlich wie bei MEISINGER und SPEER (1979) ein enger
linearer Zusammenhang zwischen Tryptophanaufnahme und -gehalt im Plasma mit r = 0,99
beziehungsweise r = 0,93 der die direkte Abhängigkeit des Plasmagehaltes von der Menge an
aufgenommenem Tryptophan sehr schön erkennen ließ. Die Beziehung zwischen dem
Tryptophangehalt im Futter und der Plasmakonzentration (r = 0,95) wurde jedoch durch die
unterschiedliche Futteraufnahme überlagert. LEWIS und SPEER (1974B), die eine lineare Beziehung
zwischen dem Tryptophangehalt im Futter und der Plasmakonzentration (r = 0.97) fanden, verfütterten
an alle Sauen die gleiche Menge Futter und konnten so zwischen dem Tryptophangehalt im Futter und
der Plasmakonzentration einen direkten Zusammenhang herstellen. In der hier angewendeten
Versuchsanstellung erhielten die Sauen jedoch das Futter zur freien Aufnahme, wobei signifikante
Unterschiede zwischen den Gruppen auftraten. Aus diesem Grund wurde in Abbildung 25 auf der
Abszisse nicht der Tryptophangehalt des Futters aufgetragen, sondern die tatsächliche
Tryptophanaufnahme mit dem Futter in der Woche der Blutprobenentnahme.
In Tabelle 65 ist die Standardabweichung beim Plasmatryptophangehalt prozentual zum Mittelwert der
jeweiligen Gruppe angegeben. Diese Betrachtungsweise wurde gewählt, um trotz des linearen
Zusammenhangs, der keine Bedarfsableitung zulässt, in Verbindung mit anderen Ergebnissen
eventuell Rückschlüsse auf den optimalen Tryptophangehalt im Futter ziehen zu können.

113
Tabelle 65: Tryptophangehalt, Standardabweichung absolut und Standardabweichung
prozentual im Sauenplasma
Gruppe I II III IV V VI Trp.-Geh. in % 0,12 0,15 0,18 0,24 0,30 0,42
Trp.-Gehalt in mg/l
2,74d
5,51cd
8,67c
15,54b
15,73b
23,91a
Standardabweichung 1,14 2,66 2,95 3,39 4,97 7,53
St.abw. in % vom Mittelw. 42 48 34 21 32 32
Gruppe I hatte absolut die geringste Standardabweichung. Bei der prozentualen Standardabweichung
war der geringste Wert bei Gruppe IV zu verzeichnen. Folgende Punkte waren bei der prozentualen
Standardabweichung auffallend:
Bei Gruppe I würde man eventuell einen höheren Wert als bei Gruppe II erwarten, da dann die Werte
der Gruppen I bis IV kontinuierlich fallend wären. Eine Erklärung, warum dies nicht so ist, könnte
darin bestehen, dass mit dem Plasmatryptophangehalt von 2,7 mg/l eine so niedrige Konzentration
erreicht ist, dass ein weiteres Fallen des Plasmaspiegels unter Aufrechterhaltung einigermaßen
normaler physiologischer Stoffwechselvorgäge nicht mehr ohne weiteres möglich ist. Der geringste in
der vorliegenden Untersuchung gemessene Tryptophangehalt im Plasma lag bei 0,53 mg/l. Daher ist
die Streuung der einzelnen Werte durch einen eventuell vorhandenen Mindestwert begrenzt. Eine
andere Erklärung liegt sicher auch darin, dass der Tryptophangehalt bei einer Mangelversorgung gegen
einen Minimalwert strebt, wobei keine negativen Werte auftreten können. Die Standardabweichung
wird dadurch zwangsläufig kleiner.
Gruppe IV weist bei der relativen Standardabweichung den geringsten Wert auf. Damit ist die
Streuung der individuellen Stoffwechselantwort auf die variierenden Tryptophangehalte im Futter am
geringsten. Da das Verhältnis zwischen Tryptophanaufnahme über das Futter und Tryptophangehalt im
Plasma direkt proportional zu sein scheint (siehe Abb. 25), ist eine Bedarfsableitung nicht möglich.
Nachdem aber anhand der gefundenen Ergebnisse der optimale Tryptophangehalt im Futter im Bereich
der Gruppe IV zu liegen scheint, lässt die gefundene geringe Standardabweichung darauf schließen,
dass es sich hierbei um einen stoffwechselphysiologisch günstigen Tryptophangehalt für den
Organismus der Muttersau handelt.
4.3.4.1.2. Essentielle Aminosäuren
Mit Ausnahme von Tryptophan, Methionin und Valin waren keine signifikanten Einflüsse einer
Tryptophanzulage im Futter auf die Konzentration der essentiellen Aminosäuren erkennbar.

114
Die mit dem Futter aufgenommene Methioninmenge korrelierte eng mit dem Gehalt im Plasma
(r)=)0,83). Ähnliche Zusammenhänge sind bei Methionin in den Arbeiten von SCHNEIDER (1992)
mit r = 0,96, WESTERMEIER (1998) mit r = 0,66 und OTT (2000) mit r =0,91 (alles eigene
Berechnungen) zu finden, obwohl jeweils andere essentielle Aminosäuren im Futter variiert wurden.
Der Einfluss auf Valin war in der vorliegenden Arbeit nicht gerichtet. So hatte zwar Gruppe IV mit 24
mg/l die höchste Konzentration aufzuweisen, jedoch war die niedrigste bei der Gruppe V mit 15 mg/l
zu finden. Die anderen Gruppen lagen dazwischen. Auch LIBAL et al. (1997) fanden bei
Tryptophangehalten von 0,12 % und 0,17 % im Laktationsfutter Auswirkungen auf den Valingehalt
im Plasma (16 mg/l beziehungsweise 22 mg/l). Bemerkenswert ist die Tatsache, dass mit Ausnahme
von Tryptophan, Lysin, Methionin und Phenylalanin bei Gruppe V immer die niedrigste Konzentration
messbar war. Dies lässt vermuten, dass die Proteinsynthese bei einem Tryptophangehalt von etwa 0,3
% im Futter am besten abläuft, da hier die meisten Aminosäuren für anabole Stoffwechselprozesse,
wie zum Beispiel den Aufbau von Milcheiweiß, verwendet und somit aus dem Blut entfernt wurden.
Dies wird vor allem deutlich, wenn man die Gruppe V mit den Gruppen II und III vergleicht, die in der
dritten Laktationswoche eine ähnliche Futter- (4,9 kg/Tag und 4,8 kg/Tag beziehungsweise 4,9
kg/Tag) und somit Proteinaufnahme (Ausnahme: Tryptophan) hatten. Auch andere Autoren berichten
von einer Verminderung des Gehalts an essentiellen Aminosäuren im Blut bei Zulage einer sich im
Mangel befindlichen essentiellen Aminosäure (LEWIS und SPEER, 1973; 1974; 1975;
KIRCHGESSNER et al., 1993; 1998; OTT, 2000). Eine andere Erklärung könnte der erhöhte Abbau
von Körpersubstanz, in diesem Fall speziell der Muskelmasse, sein, durch die der Organismus versucht
einem Mangel bei einer essentiellen Aminosäure entgegenzuwirken (KING und DUNKIN, 1986;
BRENDEMUHL et al., 1989; siehe auch: 5.5.5.). Da sich im vorliegenden Versuch nur das
Tryptophan im Mangel befand, kam es zu einer Anreicherung der anderen freigesetzten essentiellen
Aminosäuren.
4.3.4.1.3. Nicht-essentielle Aminosäuren
In der vorliegenden Untersuchung war ein signifikanter Einfluss auf die meisten nicht essentiellen
Aminosäuren messbar. Lediglich bei Alanin, Hydroxyprolin, Glycin und Serin traten keine
signifikanten Unterschiede zwischen den Gruppen auf. Bei den anderen Aminosäuren stieg der Gehalt
bis zur Gruppe IV oder V an, um danach wieder abzufallen (bei Asparagin war das Maximum bei
Gruppe II messbar). Eine Erklärung für diese Beobachtung könnte darin liegen, dass sich mit
zunehmender Tryptophanzulage die Aminosäurenrelation im Futter bis zur Erreichung des Optimums
verbessert. Da dadurch auch die Proteinbiosyntheserate erhöht wird, besteht keine Notwendigkeit mehr
für den Organismus Aminosäuren zu verstoffwechseln. Ganz im Gegenteil scheint der Körper mit

115
zunehmender Annäherung an die ideale Aminosäurenrelation bestrebt zu sein, nicht essentielle
Proteinbausteine in ausreichender Menge zur Verfügung zu stellen. Vor allem bei den glucogenen
Aminosäuren Alanin, Asparaginsäure, Glutamin und Glutaminsäure war dieser Effekt zu beobachten.
Aus diesen Eiweißbausteinen setzt sich zum großen Teil das Muskelprotein zusammen (YOUNG und
MARCHINI, 1989; ADEOLA, 1995) und aus ihnen kann beim Abbau Glucose gewonnen werden
(NELSON und COX, 2001). Diese wird von der Milchdrüse aus dem Blut aufgenommen und zur
Synthese der Milchinhaltsstoffe, unter anderem auch von Laktose, verwendet (SPINCER et al., 1968;
LINZELL und MEPHAM, 1969). Da die gebildete Menge Lactose entscheidend für die Milchleistung
ist (MACKENZIE und REVELL, 1998), könnte sich ein erhöhter Gehalt an glucogenen Aminosäuren
positiv auf die Milchleistung ausgewirkt haben. In der vorliegenden Untersuchung korrelierte der
Gesamtgehalt an glucogenen Aminosäuren (Methionin, Cystin, Threonin, Valin, Histidin, Alanin,
Arginin, Asparagin, -säure, Glutamin, -säure, Prolin, Glycin, Serin) eng mit der Milchleistung mit r =
0,95 sehr eng korreliert. Der entsprechende Korrelationskoeffizient betrug bei WESTERMEIER
(1998) 0,96 und bei OTT (2000) 0,94 (alles eigene Berechnungen).
4.3.4.2. Harnstoffgehalt im Plasma
Herrscht im Stoffwechsel ein Eiweißüberschuss vor, werden Proteine über zahlreiche
Stoffwechselprozesse zu Ammonium-Ionen (bei den schwefelhaltigen Aminosäuren auch
Schwefelwasserstoff) sowie Kohlenstoffdioxid und Wasser abgebaut. Im Körperstoffwechsel kann
man bei den Aminosäuren in Versuchen zur Bedarfsermittlung grundsätzlich zwei Arten der
Überschusssituation finden:
1. Die Mehrzahl der Aminosäuren liegt in bedarfsgerechter Menge vor und die
zu untersuchende essentielle Aminosäure befinden sich im Mangel.
→ Es kann nur so lange eine Proteinsynthese erfolgen, bis die zu untersuchende
essentielle Aminosäure limitierend wird. Darüber hinaus stellen die restlichen
Aminosäuren einen Überschuss dar.
2. Die Mehrzahl der Aminosäuren liegt in bedarfsgerechter Menge vor und die
zu untersuchende essentielle Aminosäure ist im Überfluss vorhanden.
→ Es kann nur so lange eine Proteinsynthese erfolgen, bis die Mehrzahl der
bedarfsgerecht vorliegenden Aminosäuren aufgebraucht ist. Darüber hinaus
stellt die zu untersuchende essentielle Aminosäure einen Überschuss dar.

116
Somit hat die Aminosäurenzusammensetzung des Futters eine entscheidende Wirkung auf die
Harnstoffkonzentration im Blut. Weil Ammoniak für den tierischen Körper ein Zellgift ist (vor allem
für das Gehirn), stellt seine katabolische Erzeugung in biochemischer Hinsicht ein Problem dar. Der
Stoffwechsel muss das Ammoniak in eine unschädliche Form überführen. Die beim Abbau von
Aminosäuren entstehenden Aminogruppen werden im Muskel und anderen betroffenen Geweben
durch Transaminierung in Form von Glutamat gesammelt und in verschiedenen Verbindungen mit
dem Blut in die Leber transportiert (NELSON und COX, 2001). Dort erfolgt eine enzymatische
Umwandlung des Ammoniaks zu Harnstoff, der über den Urin oder die Milch ausgeschieden wird. Da
der Transport von der Leber zur Niere über das Blut erfolgt, kann die Blutharnstoffkonzentration als
Indikator für die Höhe der Abbaurate an überschüssigen Aminosäuren gewertet werden. Nach
EGGUM (1970) sowie MALMLÖF und ASKBRANT (1988) werden um so weniger Aminosäuren
desaminiert, je höher die biologische Wertigkeit des Futtereiweißes ist. Desto niedriger ist dann die
Harnstoffkonzentration im Blut. Der Blutharnstoffgehalt wird jedoch nie gleich Null sein, da auch
ohne Überschusssituation bei den Aminosäuren Ammoniak durch den normalen Proteinumsatz anfällt.
Deshalb haben verschiedene Autoren mit Erreichen einer minimalen Harnstoffkonzentration im Blut
den Gehalt der limitierenden Aminosäure als bedarfsdeckend erachtet (ROUSSELOW und SPEER,
1979; ROUSSELOW und SPEER, 1980; WILKINSON et al., 1982; LELLIS und SPEER, 1985;
KIRCHGESSNER et al., 1993; KIRCHGESSNER et al., 1998).
In der Abbildung 26 sind die Blutharnstoffgehalte in Abhängigkeit von der Tryptophankonzentration
im Futter dargestellt.
Harnstoffgehalt im Sauenplasma
100
200
300
400
0,07 0,12 0,17 0,22 0,27 0,32 0,37 0,42 0,47
Tryptophangehalt im Futter in %
Har
nsto
ffgeh
alt i
n m
g/l
Abbildung 26: Harnstoffgehalt im Plasma der Sauen am 22. Laktationstag

117
Der Harnstoffgehalt war von den Gruppen I bis V fallend und stieg bei Gruppe VI wieder an. Den
höchsten Wert hatte Gruppe I mit 272 mg/l, den niedrigsten Gruppe V mit 213 mg/l. Die abfallenden
Werte sind auf eine Verbesserung der Relation von Tryptophan zu den restlichen essentiellen
Aminosäuren zurückzuführen. Die Abnahme der Harnstoffkonzentration war dabei von der
Basisgruppe zu Gruppe II mit über 28 mg/l am deutlichsten ausgeprägt. Die Abstufungen bis zu
Gruppe V waren hingegegen nicht mehr so stark. Der Anstieg von Gruppe V auf Gruppe VI war
jedoch mit über 22 mg/l relativ groß. Daraus lässt sich ableiten, dass die Tryptophangehalte im Futter
der Gruppen I und VI mit 0,12 % und 0,42 % einen relativ hohen Proteinüberschuss im Stoffwechsel
hervorriefen, der sich in erhöhten Harnstoffkonzentrationen im Plasma widerspiegelte.
In der Literatur schwanken die Werte der Harnstoffkonzentration zwischen 24,9 mg/l (LEWIS und
SPEER, 1974) und 300,6 mg/l (KIRCHGESSNER et al., 1993). Da sich eine Vielzahl von Faktoren,
wie zum Beispiel die Proteinqualität oder die aufgenommene Eiweißmenge, auf die Harnstoffwerte im
Blut auswirken, ist ein Vergleich nicht ohne weiteres möglich. Auch der Zeitpunkt der Blutentnahme
ist entscheidend, da die Harnstoffkonzentrationen postprandialen Schwankungen unterliegen. Die
Empfehlung von MENKE (1980) sowie HERRMANN und SCHNEIDER (1986), die Probenahme im
Zeitraum von 3 bis 5 Stunden nach der Fütterung durchzuführen, wurde im vorliegendem Versuch
eingehalten.
In der Abbildung 27 ist die quadratische Regression des Harnstoffgehaltes im Plasma am 22.
Laktationstag auf den Tryptophangehalt im Futter dargestellt. Nach dieser Regression ergab sich für
den Parameter Harnstoffgehalt im Plasma ein Bedarf von 0,30 % Tryptophan im Futter, der exakt dem
Tryptophangehalt im Futter von Versuchsgruppe V entsprach. Dieser Wert liegt etwas höher als die
aufgrund anderer Parameter ermittelten Gehalte mit Ausnahme des Wertes, der anhand des
Milchharnstoffgehaltes ermittelt wurde (0,31 %) und der sehr gut mit dem obigen Wert
zusammenpasst. Die Tatsache, dass die Konzentrationen der meisten essentiellen Aminosäuren im
Plasma ebenfalls bei der Gruppe V am niedrigsten waren (siehe 4.3.4.1.2.), stützt zusätzlich das mit
dieser Regression gefundene Ergebnis sowie das der Milchharnstoffuntersuchung.
LEWIS und SPEER (1974B) fanden bei Tryptophangehalten im Futter von 0,046 % bis 0,161 %
tendenziell abnehmende Harnstoffgehalte im Plasma. Bei einer Untersuchung von LIBAL et al. (1997)
trat bei einer Zunahme des Futtertryptophangehaltes von 0,12 % auf 0,17 % ebenfalls noch eine
Abnahme des Plasmaharnstoffgehaltes auf. Diese Ergebnisse stehen somit im Einklang mit denen des
vorliegenden Versuchs.
118
Harnstoffgehalt im Sauenplasma
y = 1512x2 - 914x + 353r2 = 0,89 n=72
150
200
250
300
0,07 0,12 0,17 0,22 0,27 0,32 0,37 0,42 0,47
Tryptophangehalt im Futter in %
Har
nsto
ffgeh
alt i
n m
g/l
Bedarf = 0,30
Abbildung 27: Ableitung des Tryptophanbedarfs anhand des Harnstoffgehalts im Sauenplasma
am 22. Laktationstag nach dem quadratischen Modell
4.3.4.3. Serotoningehalt
Ein Zusammenhang zwischen Serotonin und Futteraufnahme wird seit langem diskutiert. Die Basis
dafür bilden Experimente, bei denen eine Verabreichung von Serotoninagonisten (zum Beispiel: 5-
Carboxamidotryptamin) und Serotoninwiederaufnahmehemmern (zum Beispiel: Imipramin) direkt in
das Gehirn eine Verringerung der Futteraufnahme zur Folge hatte (BLUNDELL, 1986; LEIBOWITZ
et al., 1988). Da Tryptophan eine Vorstufe des Serotonins ist, haben verschiedene Autoren den
Zusammenhang zwischen der Tryptophanaufnahme und dem Serotoningehalt im Gehirn beim Schwein
untersucht (u.a. HENRY et al., 1992; ADEOLA und BALL, 1992; UTTECHT et al., 1992; HENRY et
al., 1996) und eine positive Korrelation festgestellt. Jedoch wurden auch gegenteilige Ergebnisse
gewonnen (BLUNDELL, 1984). Innerhalb physiologischer Grenzen beeinflusst die
Tryptophankonzentration im Plasma den Tryptophangehalt im Gehirn und somit die Serotoninbildung
dort (ANDERSON und COVEN, 1991). Da die Aufnahme von Tryptophan ins Gehirn aber nicht nur
vom Gehalt im Plasma, sondern auch noch von der Blutkonzentration an LNAA und FFA abhängig ist
(siehe auch 5.2.2.2.), sind die Zusammenhänge sehr komplex. Dadurch sind möglicherweise auch die
oben genannten gegensätzlichen Ergebnisse zum Zusammenhang zwischen Tryptophanaufnahme und
Serotoningehalt im Gehirn zu erklären.
Eine Untersuchung des Serotoningehalts im Gehirn ist nur am toten Tier praktizierbar und war daher
im vorliegenden Versuch nicht möglich. Da nach POLLOCK und ROWLAND (1981) sowie

119
FLETCHER und BURTON (1984) jedoch auch eine Veränderung des Serotoninspiegels im peripheren
Blutkreislauf Auswirkungen auf die Futteraufnahme hat, wurde der Serotoningehalt im Sauenblut
ermittelt.
Die Gehalte an Serotonin im Blut sind einem sehr starken Individualeffekt unterworfen und weisen
daher sehr große Schwankungen auf (SARRIAS et al., 1991). Teilweise scheint der individuelle
Serotoninspiegel genetisch festgelegt zu sein (HYLGAARD-JENSEN 1976). Zur Ausschaltung dieses
Individualeffektes wurde ein einzeltierbezogener Vergleich der Werte vom 28. Laktationstag und 85.
Trächtigkeitstag vorgenommen. Dabei fiel die Konzentration an Gesamtserotonin im Vollblut von
durchschnittlich 299 ng/ml (85. Trächtigkeitstag) auf 288 ng/ml (28. Laktationstag) nur um 0,4 % ab.
Zwischen den Gruppen waren jedoch große Unterschiede zu erkennen. So betrug der Abfall bei den
Gruppen I und II 36 % beziehungsweise 30 %. Bei den restlichen Gruppen war dagegen fast keine
Veränderung (Gruppe III) oder sogar ein Anstieg feststellbar (im Mittel 19 %).
Für Serotoningehalte im Schweineblut sind in der Literatur Werte von 200 ng/ml bis 300 ng/ml
angegeben (ERSPAMER, 1954; TOPEL, 1973). Somit fügen sich die in der vorliegenden Arbeit
gefundenen Mittelwerte von 299 ng/ml und 288 ng/ml sehr gut ein.
Nach OCHSENKÜHN (1988) ist ein Großteil des im Blut befindlichen Serotonins an die
Thrombozyten gebunden (85 % beim Menschen, SEBEKOVA et al., 2001; 95 % bei der Ratte,
FARDAEE et al.,1998; 80 % bei 21 Wochen alten weiblichen Schweinen, HYLGAARD-JENSEN
1976). Veränderungen des Thrombozytenwertes beeinflussen damit auch die Serotoninwerte
(BOUILLON und O´BRIEN 1971; MURPHY et al. 1973; TSCHOPP und ZUCKER 1972), so dass als
Messwert die Serotoninkonzentration je 109 Thrombozyten verwendet wurde, die nach dem aktuellen
Wissensstand am besten für Aussagen zum Serotonin im peripheren Blutkreislauf geeignet ist.
GERDES et al. (1992) fanden bei Untersuchungen an Schweinen mit durch Halothan induzierter
maligner Hyperthermie, dass ein signifikanter Anstieg des Serotoningehaltes im thrombozytenhaltigen
Plasma mit einem gleichzeitigen Anstieg der Thrombozytenzahl zusammenhing und der
Serotoningehalt je 109 Thrombozyten nahezu unverändert blieb. Je nach Alter und Leistungsstadium
werden beim Schwein Thrombozytenzahlen von 255 � 109 TZ/l bis 540 � 109 TZ/l beim Ferkel
(TANEN et al., 2003; ROTHWELL et al., 2002) beziehungsweise 180 � 109 TZ/l bis 360 � 109 TZ/l
beim Mastschwein (ZWISCHENBERGER et al., 2001) gefunden. Die in der vorliegenden
Untersuchung gefundenen Werte von durchschnittlich 126 � 109 TZ/l am 85. Trächtigkeitstag und 196
� 109 TZ/l am 28. Laktationstag entsprechen in der Größenordnung diesen Angaben.
Demzufolge ergaben sich die Serotoninwerte mit 2600 ng/109 TZ (85. Trächtigkeitstag)
beziehungsweise 1600 ng/109 TZ (28. Laktationstag). Auch diese Werte entsprechen in der

120
Größenordnung den bei 19 kg bis 30 kg schweren Schweinen beschriebenen Zahlen von 3500 ng/109
TZ (GERDES et al., 1992).
Im vorliegenden Versuch war eine Korrelation zwischen Futteraufnahme und Serotoninveränderung je
109 Thrombozyten mit r = 0,72 zu erkennen. Mit steigender Serotoninkonzentration stieg auch die
Futteraufnahme zumindest bis Gruppe IV an. Danach fielen die Serotoninkonzentration ebenso wie die
Verzehrsmenge wieder bis Gruppe VI ab. Dieses Ergebnis widerspricht augenscheinlich der in der
Literatur vorherrschenden Meinung, dass mit steigendem Serotoningehalt im peripheren Blutkreislauf
die Futteraufnahme zurückgeht (POLLOCK und ROWLAND, 1981; FLETCHER und BURTON,
1984). Eine Erklärung dafür, warum dies im vorliegenden Fall nicht so war, könnte darin liegen, dass
der Effekt des Serotonins durch andere Mechanismen überlagert wurde. Naheliegend dabei ist, dass
durch die ausgeprägte Aminosäuren-Imbalanz im Futter besonders bei Gruppe I über andere
Regulationsvorgänge, vor allem im Gehirn (PENG et al., 1972; ROGERS und LEUNG, 1973;
HARPER und PETERS, 1989), die Futteraufnahme reduziert wurde. Hierbei spielt vermutlich der
Chemosensor des Körpers für Aminosäuren in der Prepyriform Cortex im Gehirn die entscheidende
Rolle (LEUNG und ROGERS, 1973; BEVERLY et al., 1990; 1990B). Die in der Literatur gefundenen
Ergebnisse zur Beeinflussung der Futteraufnahme aufgrund von erhöhten Serotoningehalten im
peripheren Blutkreislauf beruhten immer auf der Basis von Serotonininjektionen, wobei das Futter
bedarfsgerechte Inhaltsstoffe aufwies. Da dies im vorliegenden Versuch nicht der Fall war können
Überlagerungen der Effekte aufgetreten sein.
Insgesamt scheint die Futteraufnahme am meisten durch Aminosäuren-Imbalanzen im Gehirn
beeinflusst zu sein (PENG et al., 1972; ROGERS und LEUNG, 1973; HARPER und PETERS, 1989).
Vor allem wenn das Aminosäurenmuster im Futter extrem unausgeglichen ist, werden auch die
Grenzen des Regulationssystems, das eine ausgewogene Aminosäurenkonzentration im Gehirn
aufrechterhält, überschritten (ANDERSON et al., 1968; HARPER und TEWS, 1988).

121
Serotoninveränderung 85. TT auf 28. LT
-100
-80
-60
-40
-20
0
20
40
0,07 0,12 0,17 0,22 0,27 0,32 0,37 0,42 0,47
Tryptophangehalt im Futter in %
Kon
zent
ratio
nsän
deru
ng in
%
Abbildung 28: Mittlere Konzentrationsabnahme beim Serotoningehalt je 109 Thrombozyten vom
85. TT auf den 28. LT
Es hat sich gezeigt, dass, wie auch immer die Rolle des Serotonin bei der Kontrolle der
Futteraufnahme definiert wird, Variationsversuche mit Serotonin nicht automatisch zu einer
bestimmten Veränderung der Futteraufnahme führten. Die appetithemmende Wirksamkeit von
Serotonin im Körper wird durch viele Variablen reguliert. Besonders sind diese beim Futter zu suchen.
In der Literatur sind dabei als Einflüsse aufgeführt:
- Menge der verfügbaren Kohlenhydrate (MOSES und WURTMAN, 1984; LAWTON
und BLUNDELL, 1989)
- Struktur des Futters (McARTHUR und BLUNDELL, 1986)
- Verfügbarkeit zusätzlicher Nahrungsquellen bei Wahlversuchen (ORTHEN-GAMBILL
und KANAREK, 1982)
- Aufschlussgrad der Kohlenhydrate (LAWTON und BLUNDELL, 1989B)
- zyklische Bevorzugung besonderer Nahrungsmittel (LEIBOWITZ et al., 1989;
SIMANSKY, 1989)
Somit kann nicht einfach eine direkte Dosis-Wirkungs-Beziehung hergestellt werden, sondern die
Höhe einer Verzehrsdepression ist als Zusammenspiel vieler Faktoren zu verstehen.

122
4.5.3. Lebendmasse der Ferkel
Die Lebendmassezunahmen der Ferkel stellen ein entscheidendes Kriterium bei der Ferkelproduktion
dar. Sie werden neben der Geburtsmasse in erster Linie von der Milchleistung der Sau beeinflusst,
insbesondere solange diese die einzige Nahrungsquelle für das Ferkel darstellt und kein Beifutter
aufgenommen wird. So führte bei mehreren in der Literatur beschriebenen Versuchen eine
Unterversorgung der Muttersau mit essentiellen Aminsäuren zu einer verringerten Milchleistung mit
einhergehender Verminderung des Ferkelwachstums (LEWIS und SPEER 1973, 1974, 1975; LEWIS
et al. 1978; ROUSSELOW et al. 1979; ROUSSELOW und SPEER 1980; SCHWARZ et al. 1993;
WESTERMEIER et al. 1998; OTT 2000). Für eine maximale Milchmenge ist in erster Linie eine
optimale Fütterung der Sau entscheidend. Ein ebenfalls bedeutender Faktor auf die Zunahmen der
Ferkel ist die Geburtsmasse, die im vorliegenden Versuch im Durchschnitt der Gruppen jedoch keine
Unterschiede aufwies.
Damit stellt die Lebendmassezunahme der Ferkel auch ein geeignetes Nebenkriterium für die
Ermittlung des Tryptophanbedarfs der Zuchtsauen im vorliegenden Versuch dar. Die Milchleistung am
20./21. Laktationstag und die Ferkelzunahmen in der 3. Laktationswoche spiegeln mit einem
errechneten Korrelationskoeffizienten von 0,97 diesen engen Zusammenhang gut wider (Die
Ferkelbeifütterung erfolgte erst ab dem 22. Laktationstag).
Tabelle 66: Korrelationen zwischen Milchleistung der Sau und Ferkelzunahmen (neuere
Untersuchungen; Auswahl)
Quelle
Korrelationskoeffizient
SCHWARZ et al. (1993)
0,80
WESTERMEIER (1997) 0,92
OTT (2000) 0,90
vorliegende Untersuchung 0,97
Für den Vergleich des ermittelten Korrelationskoeffizienten mit Literaturwerten (vergleiche Tabelle
66) wurden 3 Untersuchungen herangezogen, denen fast identische Datenerhebungsmethoden
zugrunde lagen. Das Ergebnis der vorliegenden Untersuchung reiht sich gut in die Werte der anderen
Autoren ein.

123
tägliche Lebendmassezunahme der Ferkel
100
125
150
175
200
225
0,07 0,12 0,17 0,22 0,27 0,32 0,37 0,42 0,47
Tryptophangehalt im Futter in %
Zuna
hmen
in g
/Tag
Abbildung 29: Mittlere Lebendmassezunahmen der Ferkel während der Laktation
In der Abbildung 29 sind die täglichen Lebendmassezunahmen der Ferkel eingetragen. Obwohl
zwischen den einzelnen Gruppen beim Gesamtdurchschnitt keine statistisch absicherbaren
Unterschiede auftraten, lag Gruppe I mit Lebendmassezunahmen von nur 157 g/Tag um 14 %
niedriger als die restlichen Gruppen mit durchschnittlich 183 g/Tag. Die Ferkel aus Gruppe I
erreichten nach 3 Wochen mit durchschnittlich 4,73 kg nicht die nach KIRCHGESSNER (1997)
anzustrebende Lebendmasse von über 5 kg wie die Ferkel der anderen Gruppen. Ein Tryptophangehalt
im Sauenfutter von 0,12 % scheint demnach für eine erfolgreiche Ferkelaufzucht nicht ausreichend zu
sein. Da bei den Milchinhaltsstoffen keine Unterschiede zwischen den Gruppen erkennbar waren, sind
die Unterschiede bei den Tageszunahmen auf die unterschiedliche Milchleistung der jeweiligen
Gruppen zurückzuführen, die von der Tryptophanzulage abhängig waren.
Die Ferkel aus Gruppe IV hatten vom 1. bis zum 28. Laktationstag mit 187 g zwar die höchsten
Tageszunahmen, sie unterschieden sich jedoch nicht wesentlich von den anderen Behandlungen mit
Ausnahme von Gruppe I. Die Tiere der Gruppe I hatten zu jedem Zeitpunkt des Versuches eine
wesentlich geringere Zunahme als die Tiere der anderen Gruppen. Auch durch die Zufütterung
während der 4. Laktationswoche konnte Gruppe I bei den Tageszunahmen nicht aufholen.
Was Gruppe IV in erster Linie auszeichnete, war die Tatsache, dass sie zusätzlich zur höchsten
durchschnittlichen Tageszunahme die niedrigste Standardabweichung aller Gruppen aufwies. Da die
Standardabweichung in diesem Fall als ein gewisses Maß für das Auseinanderwachsen der Ferkel
gedeutet werden kann, ist davon auszugehen, dass bei einem Tryptophangehalt von 0,24 % im
Sauenfutter gleichmäßigere Absetzgruppen heranwachsen. Eine Bedarfsableitung anhand der

124
Ferkelzunahmen war aufgrund der nahezu gleichen Zunahmen der Ferkel der Gruppen II bis VI nicht
möglich
Für eine erfolgreiche Ferkelproduktion sind heute nicht nur viele abgesetzte Ferkel mit hohen
Zunahmen notwendig, sondern es ist auch von größter Wichtigkeit, dass sich diese Ferkel dann
gleichmäßig schnell entwickeln. Dies ist notwendig, da die programmierte Ferkelerzeugung mit exakt
aufeinander abgestimmten Produktionszyklen darauf angewiesen ist, dass alle Tiere eines
Wurfzeitpunktes möglichst gleichzeitig die angestrebte Lebendmasse erreichen. Vom Absetztermin
über die Ferkelaufzucht bis hin zum Schlachttermin ist dies von großem Vorteil, da sich konsequente
Rein-Raus-Systeme nur so effektiv betreiben lassen.
4.3.6. Futterverbrauch der Ferkel
Vom 22. bis zum 28. Laktationstag wurde den Ferkeln Beifutter zur freien Aufnahme angeboten. Die
Streuung der ermittelten Futteraufnahme zwischen den Würfen einer Gruppe war auffallend hoch und
teilweise größer als der Mittelwert selbst. In der Abbildung 30 sind die entsprechenden Werte
eingetragen.
Futterverbrauch der Ferkel in der 4. LW
0
500
1000
1500
2000
0,07 0,12 0,17 0,22 0,27 0,32 0,37 0,42 0,47
Tryptophangehalt im Futter in %
Futte
rver
brau
ch in
g je
Wur
f
Abbildung 30: Saugferkelbeifutterverbrauch je Wurf während der 4. Laktationswoche
Ein Zusammenhang mit Parametern wie der Milchleistung der Sau oder dem Tryptophangehalt im
Sauenfutter war nicht zu erkennen. Wider Erwarten fand auch kein Ausgleich einer mangelnden
Milchversorgung der Ferkel durch eine erhöhte Futteraufnahme in der 4. Laktationswoche bei Gruppe
I statt. Somit konnte auch kein kompensatorisches Wachstum beobachtet werden (siehe auch: 5.5.3.).
Der kurze Untersuchungszeitraum von nur einer Woche war dafür unter Umständen auch zu kurz. Die

125
Vermutung, dass die Futteraufnahme besser entwickelter Ferkel höher ist als die von Tieren mit
geringeren Tageszunahmen, ließ sich mit einem Korrelationskoeffizienten von r = 0,51 statistisch
ebenfalls nicht untermauern. Bei Untersuchungen zur Methionin/Cystin-Versorgung der Sau konnten
von SCHWARZ et al. (1992) ebenfalls keine signifikanten Einflüsse beobachtet werden. Auch
WESTERMEIER et al. (1998) konnten bei einer Arbeit zum Threoninbedarf laktierender Sauen
ebenso wie OTT (2000) bei der Valinbedarfsermittlung keine signifikanten Auswirkungen feststellen,
obwohl in diesen Versuchen die Ferkel erst nach 5 Wochen abgesetzt wurden und je Wurf 2 Wochen
Beifutteraufnahme möglich war. Lediglich CHEN et al. (1978) berichteten von einem erhöhten
Verbrauch an Saugferkelbeifutter infolge eines Lysinmangels im Laktationsfutter der Sauen.

126
5. Schlussbetrachtung Anhand der bisherigen Darstellung der Versuchsergebnisse ergibt sich kein einfaches und eindeutiges
Bild zur Festlegung des Tryptophanbedarfs laktierender Zuchtsauen. Als eindeutige Aussagen lassen
sich nur zwei Sachverhalte herausheben:
1. Die Sauen mit einem Tryptophangehalt von 0,12 % im Futter erzielten sowohl bei den
Leistungs- als auch bei den Stoffwechselparametern Ergebnisse, die in der Regel
signifikant unter denen der anderen Gruppen lagen. Somit ist ein Tryptophangehalt von 0,12 %
im Laktationsfutter (= Gruppe I) als zu gering zu bezeichnen.
2. Im Vergleich zu den Gruppen mit den besten Ergebnissen bei den Untersuchungsparametern
(in der Regel Gruppe IV oder V) war festzustellen, dass die Gruppe VI unter den Werten dieser
Gruppen lag. Daraus ist abzuleiten, dass ein Tryptophangehalt von 0,42 % im Futter oder
darüber zur Erzielung optimaler Ergebnisse bei der Ferkelproduktion zu hoch ist.
Auch die Bedarfsableitung nach verschiedenen gängigen mathematischen Modellen, die nach dem
höchsten Bestimmtheitsmaß ausgewählt wurden, ergab für die verschiedenen Parameter ein zunächst
etwas uneinheitliches Ergebnis, aus dem ein Tryptophanbedarf laktierender Sauen zwischen 0,20 %
und 0,31 % anzusetzen ist (siehe Tabelle 67).
Tabelle 67: Untersuchungsparameter und resultierende Bedarfswerte in Abhängigkeit des
angewandten Regressionsmodells
Parameter
Bedarfswert
Modell
Leistungsparameter
Futteraufnahme der Sauen 0,29 Quadratisch
Masseverlust der Sauen 0,21 Asymptotisch
0,26 Reading
0,20 Broken-Line
Milchleistung der Sauen 0,29 Quadratisch
Stoffwechselparameter
Harnstoffgehalt in der Sauenmilch 0,31 Quadratisch
Harnstoffgehalt im Sauenplasma 0,30 Quadratisch

127
Bei der Ableitung anhand mehrerer Parameter wird dann in der Regel der Bedarfswert herangezogen,
der am höchsten ist, da damit die erforderlichen Mindestgehalte der anderen Parameter ebenfalls
abgedeckt sind. Die hier mehrheitlich anhand quadratischer Modelle erfolgte mathematische
Bedarfsableitung ergab somit einen erforderlichen Gehalt von 0,31 % Tryptophan in der
Futterfrischmasse. Bei den Stoffwechselparametern war dieses Ergebnis mit den im Versuch
gewonnenen Daten übereinstimmend und es trat kein Unterschied zwischen den empirisch
gewonnenen Werten und dem Ergebnis der mathematischen Bedarfsableitung auf. So hatten die Tiere
der Gruppe V mit 0,30 % Tryptophan im Futter den niedrigsten Milch- und Blutharnstoffgehalt.
Werden nur die Leistungsparameter betrachtet, so zeigt sich ein anderes Bild. Hier wurde das �beste�
Ergebnis von den Tieren der Gruppe IV mit einem Tryptophangehalt im Futter von 0,24 % erzielt.
Somit ergab sich eine gewisse Diskrepanz zwischen den Ergebnissen der Bedarfsableitung und den im
Versuch gefundenen Ergebnissen, womit die durch Regression nach dem quadratischen Modell
ermittelten Bedarfswerte kritisch betrachtet werden sollten.
POPPE und WIESENMÜLLER (1968) zeigten nämlich, dass das quadratische Modell den Bedarf bei
Vorhandensein eines Plateaus systematisch überschätzt. Im vorliegenden Versuch war zwar ein
solches Plateau nicht ohne weiteres zu erkennen, aus anderen ähnlichen Arbeiten (schwefelhaltige
Aminosäuren: KIRCHGESSNER et al., 1992; SCHNEIDER et al., 1992; Threonin: PAULICKS et al.,
1998, 1998B; Valin: OTT, 2000) ist jedoch bekannt, dass es bei Variationsversuchen mit Aminosäuren
im Futter nach dem Erreichen des Bedarfswertes mit weiterer Aminosäurenzulage zu einer
Plateaubildung beim untersuchten Parameter kommt. Ein Plateau tritt dann nicht zu Tage, wenn zu
wenig Punkte der Dosis-Wirkungs-Kurve auf dem Plateau liegen. Dies kann an der gewählten
Abstufung bei den Tryptophangehalten im Futter und/oder an dem engen Versorgungsbereich liegen,
in dem ein solches Plateau auftritt. Das quadratische Modell beschreibt eine solche Plateauphase
unzutreffend, da es zu Beginn des Plateaus noch einen deutlichen Anstieg, in der Mitte ein Maximum
und danach einen Abfall aufweist (SUSENBETH, 2002). Die Aminosäurenaufnahme, bei der das
Maximum der quadratischen Funktion erreicht ist, liegt demnach bei Vorhandensein eines Plateaus
deutlich über dem Bedarf.
Daher ist beim vorliegenden Versuch zumindest bei den Parametern Futteraufnahme und
Milchleistung von einer Plateaubildung im Bereich der Gruppe V (0,30 % Typtophan im Futter)
auszugehen, die aufgrund der Wahl der Tryptophanabstufung und des engen Plateaubereiches durch
die erfassten Daten nicht deutlich wird.

128
tägliche Futteraufnahme der Sauen
2
3
4
5
6
7
0,07 0,12 0,17 0,22 0,27 0,32 0,37 0,42 0,47Tryptophangehalt im Futter in %
Futte
rauf
nahm
e de
r Sau
en in
kg/
Tag
Abbildung 31: Ableitung des Tryptophanbedarfs anhand der Futteraufnahme der Sauen in der 4.
LW (ohne Gruppe VI) nach dem asymptotischen Modell
tägliche Milchleistung der Sauen
6
7
8
9
0,07 0,12 0,17 0,22 0,27 0,32 0,37 0,42 0,47Tryptophangehalt im Futter in %
Milc
hlei
stun
g de
r Sau
en in
kg/
Tag
Abbildung 32: Ableitung des Tryptophanbedarfs anhand der Milchleistung der Sauen während
der gesamten Laktation (ohne Gruppe VI) nach dem asymptotischen Modell
Bedarf0,90 = 0,23 Bedarf0,95 = 0,26
y = 3,62 + 2,47 � (1 � e-20,86 � (x-0,12))] Grenzwert = 6,1 r2 = 0,87 n = 60
y = 6,98 + 1,37 � (1 � e-36,02 � (x-0,12))] Grenzwert = 8,3 r2 = 0,95 n = 60
Bedarf0,90 = 0,18 Bedarf0,95 = 0,20

129
Um dennoch den Bedarf näher zu bestimmen, wurde eine erneute Bedarfsableitung nach dem
asymptotischen Modell durchgeführt, wobei die Ergebnisse der Gruppe VI (0,42 % Tryptophan im
Futter) nicht mit einbezogen wurden, da davon auszugehen ist, dass es bei einem deutlichen
Tryptophanüberschuss zu Aminosäuren-Imbalanzen oder Aminosäuren-Antagonismen kommen kann,
wobei massive Stoffwechselbeeinträchtigungen auftreten können. Diese sind meist mit
Leistungsdepressionen verbunden, wobei sich die Effekte überlagern (KIRCHGESSNER, 1997).
Diese neue Bedarfsableitung nach dem asymptotischen Modell (siehe Abbildungen 31 und 32) ergab,
je nach dem ob 90 % oder 95 % des Maximums als Wert angesetzt wurden, einen erforderlichen
Tryptophangehalt im Laktationsfutter von 0,23 % bis 0,26 % beim Parameter Futteraufnahme und 0,18
% bis 0,20 % beim Parameter Milchleistung.
Jede Bedarfsableitung ist genaugenommen nur für die jeweiligen Bedingungen und den (Leistungs-)
Parameter gültig, bei dem sie durchgeführt wurde. Für die Erzielung einer höchstmöglichen
Milchmenge war nach vorliegendem Versuch ein Gehalt von 0,18 % bis 0,20 % Tryptophan im Futter
notwendig (Abbildung 32). Dieser Wert gilt aber nur für eine Laktation und für Sauen ab dem 2. Wurf.
Für eine maximale Futteraufnahme der Sauen während der Laktation ergibt sich dagegen ein Bedarf
von 0,23 % bis 0,26 % Tryptophan im Futter (Abbildung 31). Die niedrigeren Werte für die
Milchleistung sind so zu erklären, dass es durch die Mobilisierung von Körperreserven der Sau zu
einem �Einspareffekt� von Energie und Protein aus dem Futter kommt. Dies beruht auf der Tatsache,
dass im Rahmen der Homöorhese die Milchdrüse in der Laktationszeit zusammen mit dem
Zentralnervensystem über die höchste Priorität bei der Nährstoffversorgung verfügt und der
Organismus als teilweisen Ausgleich einer Mangelernährung Körperreserven mobilisiert. Bezogen auf
die Nährstoffaufnahme der Sau entspricht der Abbau von 1 kg Körpermasse einer Einsparung von
durchschnittlich 25 MJ ME und 200 g Protein aus dem Futter (KIRCHGESSNER, 1997) und somit
auch an Tryptophan. In größerem Umfang ist diese Möglichkeit des Ausgleichs über mehrere
Laktationen hinweg kaum möglich. Zudem stellt die Mobilisierung von Körperreserven eine
erhebliche Stoffwechselbelastung dar, die nicht über längere Zeit zumutbar ist, da laktierende
Zuchtsauen ohnehin metabolisch extrem gefordert sind. Der Nährstoffbedarf (Energie und Protein) bei
säugenden Sauen ist durch die Futteraufnahme meist ohnehin nicht gedeckt, insbesondere bei größeren
Würfen (>12 Ferkel). Im vorliegenden Versuch trat selbst bei Gruppe IV, die die höchste
Futteraufnahme hatte, im Vergleich zum Bedarf für Erhaltung und Milchleistung eine Unterversorgung
über das Futter von 8 % auf. Bei Gruppe I erreichte der Wert sogar 41 %. Daher ist die
Tryptophanversorgung auf eine möglichst hohe Futteraufnahme auszurichten. Ausgehend von den
vorliegenden Versuchsergebnissen ist somit von einem Tryptophanbedarf im Laktationsfutter von 0,23
% bis 0,26 % auszugehen. Bei diesem Gehalt im Futter ist anzunehmen, dass die Leistungen der Sauen

130
auch über mehrere Laktationen aufrecht erhalten werden. Der ermittelte Bedarf bezüglich der
Futteraufnahme fügt sich auch gut in das empirisch gewonnene Bild ein, bei dem die Sauen mit einem
Tryptophangehalt von 0,24 % im Futter die besten Leistungen aufwiesen.
Ausgehend von den Bedarfsermittlungen anhand der verschiedenen Parameter des vorliegenden
Versuches ist somit der Mindestbedarf laktierender Sauen auf 0,23 % Tryptophan im Futter
anzusetzen. Da sich bei Bedarfsempfehlungen auf der Basis des Gesamtgehalts einer Aminosäure
aufgrund der unterschiedlichen Verdaulichkeitswerte verschiedener Futtermitteln immer Probleme
ergeben (siehe auch 4.2.3.), müsste für Praxisempfehlungen ein Sicherheitszuschlag vorgenommen
werden und ein Mindestgehalt von 0,26 % Gesamttryptophan empfohlen werden. Zur Ausschaltung
dieser Problematik wurde als Basis für die Bedarfsangabe die scheinbare ileale Verdaulichkeit
gewählt.
Ausgedrückt in scheinbar ileal verdaulichem Tryptophan entsprechen 0,23 % Gesamttryptophan bei
einer scheinbaren ilealen Verdaulichkeit des nativen Futtertryptophans (0,12 % im Futter) von 69 %
(vorliegende Untersuchung; siehe: 3.1.2.) und einer scheinbaren ilealen Verdaulichkeit des zugesetzten
Tryptophans (0,11 % im Futter) von 100 % (KIRCHGESSNER und ROTH, 1985; SCHUTTE et al.,
1988) einem Gehalt von 0,19 % scheinbar ileal verdaulichem Tryptophan im Futter oder 0,21 % in der
Futtertrockenmasse. Im Vergleich dazu empfehlen DEGUSSA (2001) und NRC (1998) beim
Gesamttryptophan 0,20 % beziehungsweise 0,15 % bis 0,19 % und beim scheinbar ileal verdaulichen
Tryptophan 0,16 % beziehungsweise 0,11 % bis 0,14 %. Der Tryptophangehalt im Laktationsfutter
sollte demnach aufgrund der vorliegenden Ergebnisse angehoben werden. Dies gilt vor allem dann,
wenn stickstoffreduzierte Rationen zum Einsatz kommen, die aus betriebswirtschaftlicher Sicht oft
vorteilhaft und im Hinblick auf die Umwelt als positiv zu bewerten sind.
Für die praktische Fütterung stellt sich nun die Frage, ob dieser Bedarf mit herkömmlichen Rationen
auf den landwirtschaftlichen Betrieben gedeckt wird. Dazu wurden in der Praxis eingesetzte
Futterrationen hinsichtlich ihres Gehaltes an scheinbar ileal verdaulichem Tryptophan rechnerisch
überprüft. Die Grundlage dafür bildeten Tabellenwerte der �Standardisiert ileal verdauliche(n)
Aminosäuren für Schweine� (DEGUSSA, 1999). Die Rationen und deren Gehalte an scheinbar ileal
verdaulichem Tryptophan sind Tabelle 68 zu entnehmen.

131
Tabelle 68: Rationszusammensetzung und Gehalte an scheinbar ileal verdaulichem Tryptophan
in Praxisrationen (alle Angaben in % bzw. Euro)
Ration
I
II
III
IV
V
VI
s. i. v. Trp1)
Weizen
40
20
30
60
47
0,13
Gerste 37 27,5 40 42 8 25 0,09
Hafer 25 0,11
Mais 29 0,05
Grünmehl 5 0,13
Weizenkleie 7 6 5 5 0,17
Sojaex.schrot 10,5 18 19 18 12,5 18 0,50
Ackerbohnen 10 10 0,15
Mineralfutter 2,5 2,5 3 3 3 3
Pflanzenöl 3 2 1,5 2
scheinbar ileal
verdauliches Trp
0,153
0,181
0,156
0,176
0,170
0,183
Sollgehalt 0,190 0,190 0,190 0,190 0,190 0,190
zu ergänzen
0,037
0,009
0,034
0,014
0,020
0,007
Kosten je dt Futter2) 0,96 0,23 0,88 0,36 0,52 0,18
Kosten je Sau/Jahr3) 4,08 0,98 3,74 1,53 2,21 0,77 1)=Gehalt der Einzelfuttermittel an scheinbar ileal verdaulichem Tryptophan nach DEGUSSA (1999) 2)unterstellter Preis für synthetisches L-Tryptophan: 26,00 Euro/kg 3)unterstellter Laktationsfutterbedarf: 4,25 dt/Sau und Jahr
Der Gehalt an scheinbar ileal verdaulichem Tryptophan liegt bei diesen praktischen Rationen im
Bereich von 0,153 % bis 0,183 % in der Futterfrischmasse. Damit wird der erforderliche
Mindestgehalt von 0,190 % im Laktationsfutter nicht erreicht. Auch der Versuch einer Optimierung
hinsichtlich des Tryptophangehaltes (Ration VI) führte nicht zum gewünschten Ergebnis. Die Ursache
dafür liegt im geringen Gehalt an scheinbar ileal verdaulichem Tryptophan von Getreide, der zwischen
0,05 % (Mais) und 0,13 % (Weizen) liegt. Ein Ausgleich mit Futtermitteln, die höhere Werte
aufweisen (z.B. Kartoffeleiweiß, Leinschrot, Sojaextraktionsschrot), ist nur begrenzt möglich, da diese
auch meist durch einen höheren Rohproteingehalt gekennzeichnet sind und somit der
Gesamtproteingehalt der Rationen stark ansteigen würde. Dies widerspräche jedoch den Bemühungen
zur Stickstoffreduzierung in Futterrationen und den daraus resultierenden positiven Umweltaspekten.
Die einfachste Lösung wird der Einsatz eines mit großtechnisch hergestelltem Tryptophan versehenen

132
Mineralfutters sein, das eine eventuelle Lücke beim Tryptophangehalt schließt. Der Tryptophangehalt
eines solchen Mineralfutters sollte je nach Futterration und Höhe der Supplementierung zwischen
0,25)% und 1,25 % liegen.

133
6. Zusammenfassung Mit der vorliegenden Arbeit wurde der Einfluss einer über 6 Stufen variierten Tryptophanversorgung
(0,12 % bis 0,42 % Tryptophan im Futter) auf verschiedene Leistungs- und Stoffwechselparameter
laktierender Sauen und deren Ferkel untersucht. Der Versuch umfasste insgesamt 72
Laktationsperioden (12 Wiederholungen pro Behandlung) von je 28 Tagen Dauer von 61
verschiedenen Sauen der Deutschen Landrasse (mindestens eine abgeschlossene Laktation). Die
Wurfgröße wurde auf 10 � 12 (durchschnittlich 10,3) Ferkel standardisiert. Die Sauen erhielten ein als
Alleinfutter konzipiertes Versuchsträchtigkeitsfutter mit niedrigem aber bedarfsdeckendem
Tryptophangehalt (0,12 %) in Höhe von 2,4 kg je Tag ab dem 60. und 2,8 kg ab dem 85.
Trächtigkeitstag. Am 110. Trächtigkeitstag wurden die Tiere gleichmäßig nach Lebendmasse und
Wurfnummer in 6 Behandlungsgruppen eingeteilt und erhielten ein spezielles Laktationsfutter mit 15,6
% Rohprotein, 1,06 % Lysin und 13,2 MJ umsetzbarer Energie (ME) je kg Frischmasse. Die
Mischungen I bis VI unterschieden sich lediglich im Tryptophangehalt, wobei der Basismischung
(Gruppe I) L-Tryptophan bis zur jeweils angestrebten Konzentration zugelegt wurde. Die
Gesamttryptophangehalte (= scheinbar ileal verdauliches Tryptophan) betrugen in den Rationen von
Gruppe I 0,12 % (0,11 %), von Gruppe II 0,15 % (0,11 %), von Gruppe III 0,18 % (0,14 %), von
Gruppe IV 0,24 % (0,20 %), von Gruppe V 0,30 % (0,26 %) und von Gruppe VI 0,42 % (0,38 %). Die
Sauen wurden ad libitum gefüttert.
Die Tryptophankonzentration im Futter hatte einen erheblichen Einfluss auf den Futterverbrauch der
Sauen. Die Basisgruppe zeigte im Durchschnitt über die gesamte Laktation signifikant niedrigere
Futter-, Protein- und Energieaufnahmen als die Zulagegruppen. Gruppe IV wies immer den höchsten
Wert auf. Die durchschnittliche Aufnahme betrug im Mittel aller Gruppen beim Futter 4,4 kg/Tag (3,2
kg/Tag bei Gruppe I), beim Protein 690 g/Tag (510 g/Tag bei Gruppe I) und bei der Energie 58
MJ/Tag (43 MJ/Tag bei Gruppe I). Insgesamt stieg die Aufnahme von Gruppe I bis IV (0,24 %
Typtophan) an und fiel bis Gruppe VI wieder ab. Die tägliche Tryptophanaufnahme betrug bei Gruppe
I 3,9 g, bei Gruppe II 6,5 g , bei Gruppe III 8,0 g, bei Gruppe IV 13,1 g, bei Gruppe V 13,4 g und bei
Gruppe VI 17,2 g.
Die täglichen Lebendmasseverluste der Sauen von Gruppe I waren mit durchschnittlich 1100 g
signifikant höher als die der Gruppen IV bis VI mit einem Mittelwert von 440 g. Die Gruppen II und
III lagen mit 640 g im Mittel dazwischen. Die geringsten Lebendmasseverluste wies mit 340 g/Tag die
Gruppe IV auf.
Der Tryptophangehalt im Futter hatte auch einen entscheidenden Einfluss auf die Milchleistung der
Sauen. Mit 0,24 % Tryptophan im Futter war die tägliche Milchmenge am 20./21. Laktationstag (9035

134
g), die mittels weigh-suckle-weigh-Methode am 13./14. und 20./21. Laktationstag bestimmt wurde,
deutlich höher als bei 0,12 % Tryptophan im Futter (6840 g). Ein höherer Tryptophangehalt von 0,30
% oder 0,42 % brachte keine weitere signifikante Steigerung der Milchleistung. Der Fett- (7,9 %),
Eiweiß- (4,7 %), Laktose- (5,8 %), Energie- (5,1 MJ/kg FS) und Trockensubstanzgehalt (19,2 %) der
Milch wurde durch die Tryptophankonzentration im Futter nicht beeinflusst. Der Messzeitpunkt hatte
ebenfalls keine Auswirkung, lediglich beim Fettgehalt war ein signifikantes Absinken vom 13./14. auf
den 20./21. Laktationstag von 8,2 % auf 7,6 % zu beobachten. Der Harnstoffgehalt in der Sauenmilch
sank kontinuierlich von 211 mg/l bei Gruppe I (0,12 % Tryptophan) auf 173 mg/l bei Gruppe V (0,30
% Tryptophan) ab (Gruppe VI, 0,42 % Tryptophan: 188 mg/l). Die Aminosäurenzusammensetzung der
Milch wurde durch die verschiedenen Tryptophanzulagen nicht beeinflusst. Die Gehalte betrugen in
g/kg FS Milch: Lys 3,4, Met 0,9, Cys 0,6, Thr 1,9, Leu 3,9, Ile 1,9, Val 2,4, Phe 1,9, Tyr 1,8, His 1,3.
Zur Plasmagewinnung erfolgte bei allen Sauen eine Blutentnahme am 22. Laktationstag 3 Stunden
nach der Fütterung. Der Gehalt an freiem Tryptophan im Plasma korrelierte mit r = 0,99 sehr stark mit
der Tryptophanaufnahme über das Futter und stieg von 2,7 mg/l bei Gruppe I (0,12 % Tryptophan) auf
23,9 mg/l bei Gruppe VI (0,42 % Tryptophan) an. Die Gehalte an freien Aminosäuren betrugen am 22.
Laktationstag in mg/l: Lys 43,2, Met 20,6, Cys 1,9, Thr 35,7, Leu 24,8, Ile 14,2, Val 20,8, Phe 21,5,
Tyr 18,9, His 17,7. Der Harnstoffgehalt im Plasma fiel von Gruppe I mit 270 mg/l bis auf 210 mg/l bei
Gruppe V ab (Gruppe VI: 240 mg/l).
Zur Ermittlung des Hämatokrit-, Thrombozyten- und Serotoningehaltes erfolgte am 85.
Trächtigkeitstag und am 28. Laktationstag eine Blutentnahme morgens vor der Fütterung. Der
Hämatokritwert im Blut aller Gruppen fiel signifikant vom 85. Trächtigkeitstag mit 37 % auf 33 % am
28. Laktationstag. Gleichzeitig erhöhte sich der Thrombozytengehalt von 126 � 109 TZ/l auf 196 � 109
TZ/l. Der Serotoningehalt betrug am 85. Trächtigkeitstag im Mittel aller Gruppen 2630 ng/109 TZ. Am
28. Laktationstag war ein Einfluss der Tryptophanzulage im Futter erkennbar, der zu einer
signifikanten Abnahme des Serotoningehaltes bei den Gruppen I und II im Verhältnis zum 85.
Trächtigkeitstag von durchschnittlich 59 % (980 ng/109 TZ) führte. Die Zulagengruppen IV bis VI
wiesen dagegen nur eine mittlere Abnahme um 14 % (1960 ng/109 TZ) auf. Gruppe III lag mit 33 %
(1790 ng/109 TZ) dazwischen. Die niedrigste Serotoninabnahme wies die Gruppe IV mit nur 6 %
(2060 ng/109 TZ) auf.
Die Lebendmassen der Ferkel wurden am 1., 13./14., 20./21. und 28. Laktationstag erfasst. Die
Tageszunahmen lagen im Durchschnitt aller Gruppen über den gesamten Beobachtungszeitraum bei
180 g. Zwischen den Gruppen traten keine signifikanten Unterschiede auf, jedoch waren
Lebendmassen und Zunahmen der Ferkel aus Gruppe I zu jedem Zeitpunkt des Versuchs um etwa 13

135
% geringer als das Mittel der übrigen Gruppen. Der Futterverbrauch der Ferkel lag in der 4.
Laktationswoche bei 800 g je Wurf. Unterschiede zwischen den Gruppen traten nicht auf.
Für die Parameter Futteraufnahme, Milchleistung und Masseverlust der Sauen sowie
Harnstoffkonzentration in Milch und Sauenblut wurden Regressionen nach verschiedenen
mathematischen Modellen zur Bedarfsableitung errechnet. Ausgehend von den Ergebnissen dieser
Bedarfsermittlung wurde ein Mindestbedarf von 0,23 % Tryptophan im Laktationsfutter ermittelt. Auf
Basis der scheinbaren ilealen Verdaulichkeit entspricht dies einem Gehalt von 0,19 % (90,5 % TS).
In der Praxis werden diese Werte mit Rationen auf Getreide-Soja-Basis in der Regel nicht erreicht, so
dass eine entsprechende gezielte Supplementierung zu empfehlen ist. Eine Ergänzung von
Futtermischungen über ein Mineralfutter, das je nach Ration und Höhe der Supplementierung 0,25-
1,25 % großtechnisch hergestelltes Tryptophan enthalten sollte, erscheint zur Schließung einer
Bedarfslücke als die praktikabelste Lösung.

136
7. Summary The aim of the present study was to investigate the effects of varying supplemental levels of dietary
tryptophan (6 groups: 0.12 % - 0.42 %) on lactation performance and different physiological
parameters in lactating sows. In the experiment, 61 multiparous sows (German Landrace) were used
over a total of 72 lactation periods (12 replications per treatment) lasting 28 days each. The average
litter size was standardized with 10 � 12 piglets (average 10.3). The sows were fed daily 2.4 and 2.8 kg
of the experimental gestation diet from day 60 and day 85 of gravidity, respectively. On day 110 of
gestation, the sows were equally distributed to six treatments (I-VI) according to lactation number and
body weight. They were fed a basal diet with: CP 15.6 %; lys 1.06 %; 13.2 MJ ME/kg which was
supplemented using L-tryptophan to total tryptophan concentrations (= apparent ileal digestible basis)
of 0.12 % (0.08 %) in group I, 0.15 % (0.11 %) in group II, 0.18 % (0.14 %) in group III, 0.24 % (0.20
%) in group IV, 0.30 % (0.26 %) in group V and 0.42 % (0.38 %) in group VI. The sows were allowed
ad libitum access to the diets.
Dietary tryptophan considerably affected sows´ nutrient intakes. During the whole lactation period the
daily intake of feed, protein and energy of the sows was significantly depressed in treatment I with the
lowest dietary tryptophan content whereas the corresponding values of group IV were always the
highest. The average daily intake was 4.4 kg feed, 690 g protein and 58 MJ ME for all treatments and
3.2 kg feed, 510 g protein and 43 MJ ME for treatment I. Generally nutrient intake increased between
0.12 % and 0.24 % dietary tryptophan and decreased with 0.42 % dietary tryptophan. The sows had a
daily tryptophan intake of 3.9 g in group I, 6.5 g in group II, 8.0 g in group III, 13.1 g in group IV,
13.4 g in group V and 17.2 g in group VI.
The daily weight loss of sows in group I (1100 g) was significantly higher than in group IV, V and VI
(average 440 g) whereas groups II and III (average 640 g) were intermediate. Group IV has the lowest
weight loss of 340 g/d.
Milk yield as determined by the weigh-suckle-weigh-method on days 13/14 and 20/21 was also
affected by dietary tryptophan. With 0.24 % tryptophan in the diet, milk yield on day 20/21 (9035 g)
was significantly higher than with 0.12 % tryptophan (6840 g). Further dietary tryptophan
supplementations up to 0.30 % and 0.42 % tryptophan did not affect milk yield. Fat (7.9 %), protein
(4.7 %), lactose (5.8 %) and energy (5.1 MJ/kg) content as well as dry matter (19.2 %) in the milk
were not affected by dietary tryptophan. With the exception of fat content, the day of lactation had no
effect on the concentrations of milk nutrients. Fat content decreased significantly between days 13/14
and 20/21 from 8.2 % to 7.6 %. The concentration of urea in the milk on day 20/21 decreased
continuously from 211 mg/l in group I to less than 173 mg/l in group V and then, increased again to
188 mg/l in group VI. The concentrations of amino acids in the milk were not affected by the different

137
tryptophan supplementations. The contents (g/kg) in milk were: lys 3.4, met 0.9, cys 0.6, thr 1.9, trp
0.6, leu 3.9, ile 1.9, val 2.4, phe 1.9, tyr 1.8, his 1.2.
Blood samples were taken from all sows at day 22 of lactation 3 hours after feeding for plasma
preparation. The concentration of free plasma tryptophan was correlated (r = 0.99) with tryptophan
intake. It increased from 2.7 mg/l (group I) to 23.9 mg/l (group VI). The concentration of free amino
acids were in mg/l: lys 43.2, met 20.6, cys 1.9, thr 35.7, leu 24.8, ile 14.2, val 20.8, phe 21.5, tyr 18.9,
his 17.7. Plasma urea decreased from 270 mg/l in group I to 210 mg/l in group V and then increased
again to 240 mg/l in group VI.
On day 85 of gestation and day 28 of lactation before morning feeding of the sows blood samples were
taken to determine hematocrit, platelets and serotonin content. Hematocrit showed no differences
between the treatments but decreased significantly from 37 % on day 85 of gestation to 33 % on day
28 of lactation. Simultaneous the platelets increased from 126 � 109 plt/l to 196 � 109 plt/l. Serotonin
concentration on day 85 of gestation was 2630 ng/109 plt. On day 28 of lactation there was a effect of
dietary tryptophan leading to a significantly decrease of serotonin content in comparison to day 85 of
gestation with 59 % (980 ng/109 plt) in average of group I and II. In contrast the average decrease in
groups IV, V and VI was only 14 % (1960 ng/109 plt). With a value of 33 % (1790 ng/109 plt) group
III was intermediate. The lowest decrease was found in group IV with only 6 % (1060 ng/109 plt).
The mean body weight of the piglets was determined on day 1, 13/14, 20/21 and 28 of lactation. There
were no significant differences, although group I always had the lowest values. Daily weight gain of
the piglets averaged during the whole lactation period 180 g/d and did not differ significantly between
the groups. However, piglets of treatment I weighed 13 % less than the total mean. Average feed
consumption (800 g per litter in the fourth week of lactation) was also not significantly affected by the
dietary treatments.
Requirement for the parameters feed consumption, milk yield and weight loss as well as urea
concentration of milk and plasma was calculated using different regression models. Based on these
results a minimum requirement of 0.23 % tryptophan in lactation diet was determined. This is equal to
a content of 0.19 % on an apparent ileal digestible basis.
In practical feeding with corn-soybean meal diets these contents are usually not met. So a suitable
specific supplementation is recommended, which could best be done by enriching the mineral premix
to 0.25-1.25 % L-tryptophan depending on the basal diet and the percentage of supplementation.

138
8. Literaturverzeichnis
ADEOLA, O.; BALL, R.O.; 1992: Hypothalamic neurotransmitter concentrations and meat quality in
stressed pigs offered excess dietary tryptophan and tyrosine.
J. A. Sci. 70: 1888
ADEOLA, O.; 1995: Dietary lysine and threonine utilization by young pigs: Efficiency for carcass
growth.
Can. J. Anim. Sci. 75: 445-452
AMTSBLATT DER EUROPÄISCHEN GEMEINSCHAFT; 1972: Bestimmung von Stärke.
5. Jahrg., Nr.: L123: 7-9
ANDERSON, H.L.; BENEVENGA, N.J.; HARPER, A.E.; 1968: Associations among food and protein
intake, serine dehydratase and plasma amino acids.
Am. J. Physiol. 214: 1008-1013
ANDERSON, I.; COVEN, P.; 1991: Neuroendocrine response to l-tryptophan as an index of brain
serotonin function: Effect of weight loss. In: Kynurenine and serotonin pathways, Schwarcz et
al. (ed.)
Plenum Press, New York
ALTAVINI, A.C.C.; FERNANDES, L.C.O.; 1990: Reproductive and productive performance of
primiparous sows fed a diet restricted in energy.
J. Anim. Sci. 68 Suppl.: 367-368
ARBEITSGEMEINSCHAFT FÜR WIRKSTOFFE IN DER TIERERNÄHRUNG, Hrsg; 1998:
Aminosäuren in der Tierernährung.
Verlag Alfred Strothe, Holm
ARC, AGRICULTURAL RESEARCH COUNCIL, ed.; 1981: The nutrient requirements of pigs.
Commonwealth Agricultural Bureaux

139
AULDIST, D.E.; KING, R.H.; 1994: Piglets role in determining milk production in the sow. In:
Manipulation Pig Production V, Hennessy, D.P.; Cranwell, P.D., (ed.)
Australasian Pig Science Association, Werribee, Australia
BARBER, R.S.; BRAUDE, R.; MITCHELL, K.G.; 1955: Studies on milk production of large white
pigs.
J. Agric. Sci. 46: 97-118
BECKER, D.E.; NORTZOLD, R.A.; JENSEN, A.H.; TERRILL, S.W.; NORTON, H.W.; 1955:
The tryptophan requirements of the young pig.
J. Anim. Sci. 14: 664-673
BEESON, W.H.; MERTZ, E.T.; SHELTON, D.C.; 1948: The amino acid requirement of swine: I
Tryptophan.
Science 107: 599-600
BEESON, W.H.; MERTZ, E.T.; SHELTON, D.C.; 1949: Effect of tryptophan deficiency on the
pig.
J. Anim. Sci. 8: 532-540
BENDER, D.A.; 1985: Amino acid metabolism.
John Wiley & Sons, Chichester, U.K.
BENGTSSON, G.; 1972: Sow colostral free amino acids and their importance for the newborn piglets.
Acta Vet. Scand. 13: 7-13
BERGE, S.; INDERBOE, T.; 1953: Milk production by sows.
Meldiger fra Norges Landbrukshoegskole 33: 389-424
BERTRAM, H.L.; BERENDE, P.L.M.; 1983: Tryptopanbebarf des Mastschweines im Bereich 12 -
40 kg Lebendgewicht.
Kraftfutter 66: 46-50

140
BEVERLY, J.L.; GIETZEN, D.W.; ROGERS, Q.R.; 1990: Effect of dietary limiting amino acid in
prepyriform cortex on food intake.
Am. J. Physiol. 259: R709-R715
BEVERLY, J.L.; GIETZEN, D.W.; ROGERS, Q.R.; 1990B: Effect of dietary limiting amino acid in
prepyriform cortex on meal patterns.
Am. J. Physiol. 259: R716-R723
BEYER, M.; 1986: Untersuchungen zum Energie- und Stoffumsatz von graviden und laktierenden
Sauen sowie Saugferkeln � ein Beitrag zur Präzisierung des Energie- und Proteinbedarfes.
Dissertation am Forschungszentrum für Tierproduktion, Dummerstorf-Rostock
BLACK, J.L.; CAMPBELL, R.G.; WILLIAMS, I.H.; JAMES, K.J.; DAVIES, G.T.; 1986:
Stimulation of energy and amino acid utilisation in the pig.
Research and Development in Agriculture 3: 121-145
BLUNDELL, J.E.; 1984: Serotonin and appetite.
Neuropharmacology 23: 1537-1551
BLUNDELL, J.E.; 1986: Serotonin manipulations and the structure of feeding behavior.
Appetite 7 (Suppl.): 39-56
BLUNDELL, J.E.; LAWTON, C.L.; HALFORD, J.C.G.; 1995: Serotonin, eating bahavior, and fat
intake.
Obesty Research Vol. 3 Suppl. 4 Nov.: 471-476
BOCH, H.-D.; KOWALCYK, J.; 1976: Zum Problem des Harnstoffeinsatzes in der
Schweinefütterung.
Arch. Tierernährg. 26 (11): 777
BOLDUAN, G.; SCHNEIDER, R.; 1986: Einflüsse auf die Blutharnstoffkonzentration beim
Schwein.
Mh. Vet.-Med. 41: 524-527

141
BOLDUAN, G.; SCHNABEL, E.; MORGENTHUM, R.; 1989: Ernährungsregime für säugende
Sauen und Saugferkel.
F/E Bericht, Rostock-Dummerstorf
BOOMGAARD, J.; BAKER, D.H.; JENSEN, A.H.; HARMON, B.G.; 1972: Effect of dietary lysine
levels on 21-day lactation performance of first litter sows.
J. Anim. Sci. 34: 408-410
BOUILLON, D.J.; O´BREIN, R.A.; 1971: Abnormalities of 5-hydroxytryptamine uptake and binding
by blood platelets from children with Down´s syndrome.
J. Physiol. 212: 377-392
BOYD, R.D.; MOSER, B.D.; PEO, E.R.; CUNNINGHAM, P.J.; 1978: Effect of energy source
prior to parturation and during lactation on piglet survival and growth and on milk lipids.
J. Anim. Sci. 47: 883-892
BOYD, R.D.; KENSINGER, R.S.; 1998: Metabolic precursors for milk synthesis. In: The lactating
sow. Verstegen, M.W.A.; Moughan, P.J.; Schrama, J.W.; (ed.)
Wageningen Per, Wageningen
BRABANT, W.; SCHULZ, J.; 1968: Beiträge zur Physiologie und Pathologie des Schweines. II.
Untersuchung von Sauenmilch aus einzelnen Drüsenabteilungen.
Fortpfl. Haust. 4: 60-71
BRENDEMUHL, J.H.; LEWIS, A.J.; PEO, E.R.; 1987: Effect of protein and energy intake by
primiparous sows during lactation on sow and litter performance and sow serum thyroxine
and urea concentrations.
J. Anim. Sci. 64: 1060-1069
BRENDEMUHL, J.H.; LEWIS, A.J.; PEO, E.R.; 1989: Influence of energy and protein intake during
lactation on body composition of primiparous sows.
J. Anim. Sci. 67: 1478-1488

142
BURGHART, M.; 1957: Untersuchungen über die Milchleistung von Sauen unter besonderer
Berücksichtigung der Leistung der Einzeltiere.
Bay. Lw. Jahrb. 34: 485-504
CHEN, S.Y.; D�MELLO, J.P.F.; ELSLEY, F.W.H.; TAYLOR, A.G.; 1978: Effect of dietary lysine
levels on performance, nitrogen metabolism and plasma amino acid concentration of lactatin
sows.
Anim. Prod. 27: 331-344
COFFEY, M.T.; SEERLEY, R.W.; MABRY, J.W.; 1982: The effect of source of supplemental
dietary energy on sow milk yield, milk composition and litter performance.
J. Anim. Sci. 55: 1388-1394
COFFEY, M.T.; DIGS, B.G.; HANDLIN, D.L.; KNABE, D.A.; MAXWELL, C.V.; NOLAND,
P.R.; PRINCE, T.J.; CROMWELL, G.L.; 1994: Effects of dietary energy during gestation
and lactation on reproductive performance of sows: a cooperative study.
J. Anim. Sci. 72: 4-9
CSAPÒ, J.; MARTIN, T.G.; CSAPÒ-KISS, Z.S.; HÀZAS, Z.; 1996: Protein, fat, vitamin and
mineral concentrations in porcine colostrum and milk from parturition to 60 days.
Int. Dairy J. 6: 881-902
CURZON, G.; KANTAMANENI, B.D.; WINCH, J.; ROJAS-BUENO, A.; MURRAY-LYON,
I.M.; WILLIAMS, R.; 1973: Plasma and brain tryptophan changes in experimental acute
hepatic failure.
J. Neurochem. 21: 137-145
DAVIS, R.B.; WHITE, J.G.; 1968: Localization of 5-hydroxytryptamine in blood platelets. A
radioautographic and ultrastructural study.
Br. J. Haematol. 15: 93-99
DEGUSSA, Ed.; 1999: Standardisiert ileal verdauliche Aminosäuren für Schweine.
Degussa-Hüls AG, Frankfurt am Main, Selbstverlag

143
DEGUSSA, Ed.; 2001: Amino Acid Recommendation.
Degussa-Hüls AG, Frankfurt am Main, Selbstverlag
DEN HARTOG, L.A.; VERSTEGEN, M.W.A.; HERMANS, H.A.T.M.; NORDEWIER, G.J.; VAN
KEMPEN, G.J.M.; 1984: Some factors associated with determination of milk production in
sows by weighing piglets.
Z. Tierphysiol. Tierernährg. u. Futtermittelkd. 51: 148-157
DEN HARTOG, L.A.; BOER, H.; BOSCH, M.W.; KLASSEN, G.J.; VAN DER STEEN, H.A.M.;
1987: The effect of feeding level, stage of lactation and metho of milk sampling on the
composition of milk(fat) in sows.
J. Anim. Physiol. a. Anim. Nutr. 58: 253-261
(DLG) DEUTSCHE LANDWIRTSCHAFTSGESELLSCHAFT; 1996: Leistungs- und
qualitätsgerechte Schweinefütterung; Teil B: Sauen und Ferkel.
DLG-Information 2/1996
DE VRIES, J.X.; GÜNTHERT, W.; DING, R.; 1980: Determination of nicotinamide in human plasma
and urine by ion-pair reversed-phase high-performance liquid chromatography.
Journal of Chromatography, 221: 161-165
D�MELLO, J.P.F.; LEWIS, D.; 1970: Amino acids interactions in chick nutrition. 3.
Interdependence in amino acids requirements.
Br. Poult. Sci. 11: 367-385
DOURMAD, J.Y.; 1991: Effect of feeding level in the gilt during pregnancy on voluntary feed intake
during lactation and changes in body composition during gestation and lactation.
Livest. Prod. Sci. 27: 309-319
DOURMAD, J.Y.; ETIENNE, M; NOBLET, J.; 1991: A contribution to the study of amino acid
requirement for lactation in sows.
Journèes de Recherches Porcines en France, 23: 61-68

144
DOURMAD, J.Y.; 1993: Standing and feeding behavior of the lactating sow: effect of feeding level
during pregnancy.
Appl. Anim. Behav. Sci. 37: 311-319
DOURMAD, J.Y.; ETIENNE, M; PRUNIER, A.; NOBLET, J.; 1994: The effect of energy and protein
intake of sows on their longevity.
A. review. Livestock. Prod. Sci. 40: 87-97
DOURMAD, J.Y.; NOBLET, J.; ETIENNE, M.; 1998: Effect of protein and lysine supply on
performance, nitrogen balance and dody composition changes of sows during lactation.
J. Anim. Sci. 76: 542-550
EDMONDS, M.S.; GONYOU, H.W.; BAKER, D.H.; 1987: Effect of excess levels of methionine,
tryptophan, arginine, lysine or threonine on growth and dietary choice in the pig.
J. Anim. Sci. 65: 179-185
EDMUND, T.S.LI.; ANDERSON, G.H.; 1983: Amino acids in the regulation for food intake.
Nutr. Abstr. Rev. Clin. Nutr. 53 (3): 169-181
EGGUM, B.; 1970: Blood urea measurement as a technique assessing protein quality.
Br. J. Nutr. 24: 983-988
ELLIOT, R.F.; VAN DER NOOT, G.W.; GILLBREATH, R.L.; FISHER, H.; 1971: Effect of dietary
protein level on composition changes in sow colostrum and milk.
J. Anim. Sci. 32: 1128-1137
ELSLEY, F.W.H.; 1971: Nutrition and lactation in the sow. In: Lactation,Falconer, I.R., (ed.)
Butterworths, London
ETIENNE, M.; NOBLET, J.; DOURMAD, J.Y.; FORTUNE, H.; 1989: Etude du besoin en lysine
des truies en lactation.
J. Rech. Procine en France 7: 115-120

145
ETIENNE, M.; DOURMAD, J.Y.; NOBLET, J.; 1998: The influence of some sow and piglet
characteristics and of enviromental conditions on milk production. In: The lactating sow.
Verstegen, M.W.A.; Moughan, P.J.; Schrama, J.W.; (ed.)
Wageningen Per, Wageningen
ETTLE, T.; ROTH, F.X.; 2003: Evidence of a specific dietary selection for tryptophan by the piglet.
Proc. Soc. Nutr. Physiol. 12: 84
EVERTS, H.; DEKKER, R.A.; 1994: Effect of nitrogen supply on nitrogen and energy metabolism
in lactating sows.
Anim. Prod. 59: 445-454
FARDAEE, M.; PANJEHSHAHIN, M.R.; OWJI, A.A.; VASSEI, M.; 1998: Serotonin levels in
plasma and platelets of cyclosporin A treated rats.
Irn. J. Med. Sci. 23(3&4): 89-93
FERNSTROM, J.D.; WURTMAN, R.J.; 1972: Tryptophan and Serotonin
Science 178: 414-416
FERNSTROM, J.D.; 1991: Effects of the diet and other metabolic phenomena on brain tryptophan
uptake and serotomim synthesis. In: Kynurenine and serotonin pathways, Schwarcz et al.
(ed.)
Plenum Press, New York
FLETCHER, P.J.; BURTON, M.J.; 1984: Effects of manipulations of peripheral serotonin on feeding
and drinking in the rat.
Pharmacology Biochemistry & Behavior 20: 835-840
FOWLER, V.R.; GILL, B.P.; 1989: Voluntary food intake in the young pig. In: The Voluntary Food
Intake of Pigs, Frobes, J.M.; Varley, M.A.; Lawrence, T.J.; (ed.) Occasional Publication No.
13: 51-60.
British Society of Animal Production, Edinburgh, U.K.

146
FROBISH, L.T.; LILLIE, R.J.; 1976: Sow iron injection and pig hematology In: 4. International pig
veterinary society congress; International pig veterinary society (ed.)
Selbstverlag, Ames, Iowa, USA
GERDES, C.; RICHTER, A.; ANNIES, R.; LÖSCHER, W.; 1992: Increase of serotonin in plasma
during onset of halothane-induced malignant hyperthermia in pigs.
European Journal of Pharmacology 220: 91-94
(GfE) GESELLSCHAFT FÜR ERNÄHRUNGSPHYSIOLOGIE; 1987: Energie- und
Nährstoffbedarf landwirtschaftlicher Nutztiere, Nr. 4 Schweine.
DLG-Verlag, Frankfurt (Main)
(GfE) GESELLSCHAFT FÜR ERNÄHRUNGSPHYSIOLOGIE; 2002: Bestimmung der praecaecalen
Verdaulichkeit von Aminosäuren beim Schwein � Empfehlung zur standardisierten
Versuchsdurchführung.
Proc. Soc. Nutr. Physiol. 11: 233-243
GOTTERBARM, G.G.; ROTH, F.X.; KIRCHGESSNER, M.; 1998: Influence of the ratio of
indispensable: dispensable amino acids on whole-body protein turnover in growing pigs.
J. Anim. Physiol. a. Anim. Nutr. 79 (3/4): 174-183
GRUBER, F.; MENKE, K.H.; 1984: Einfluß von Aminosäuren-Zulage auf die Futteraufnahme und
die Proteinverwertung von Ferkeln.
Z. Tierphysiol. Tierernährg. u. Futtermittelkunde 51 (4): 214-228
GRUBER, F.; 1985: Der Einfluß von Aminosäuren auf die Futteraufnahme von omnivoren
Monogastriden.
Übers. Tierernähr. 13: 129-154
GUETTE, J.O.; LENKEIT, W.; 1960: Energieumsatz und Nährstoffbedarf säugender Sauen. 9.
Mitteilung der �Langfristigen Untersuchung zum äußeren und inneren Stoffwechsel des
graviden und laktierenden Schweines�.
Z. Tierphysiol. Tierernährg. Futtermittelk. 15: 165-191

147
HARPER, A.E.; TEWS, J.K.; 1988: Nutritional and metabolic control of brain amino acid
concentrations. In: Amino acid availability and brain funktion in health and disease. Huether,
G. (ed.) NATO ASI Series vol. H20.
Springer-Verlag, Berlin
HARPER, A.E.; PETERS, J.C.; 1989: Protein intake, brain amino acid and serotonin concentrations
and protein self-selection.
J. Nutr. 119: 677-689
HARPER, H.A.; MARTIN, D.W.; MAYES, P.A.; RODWELL, V.W.; unter Mitarbeit von:
GRODSKY, G.M.; NESTLE, M.; 1987: Medizinische Biochemie.
2. korrigierte Auflage, Springer Verlag, Berlin, Heidelberg, New York
HARTMAN, D.A.; POND, W.G.; 1960: Design and use of the milking machine for sows.
J. Anim. Sci. 19: 780-785
HARTMAN, D.A.; LUDWICK, T.M.; WILSON, R.F.; 1962: Certain aspects of lactating
performance in sows.
J. Anim. Sci. 21: 883-886
HARTMANN, P.E.; HOLMES, M.A.; 1989: Sow lactation. In: Manipulating pig production II.
Proceedings of the Australasian Pig Science Association, Barret, J.L.; Hennessy, D.P., (ed.)
Australasian Pig Science Association, Werribee, Australia
HARTMANN, P.E.; HUGHES, P.E.; 1996: Lactation. In: World animal science, C 10, Pig
production; Taverner, M.R., Dunkin, A.C. (eds.)
Elsevier, Amsterdam
HAYDON, K.D.; NEWTON, G.L.; DOVE, C.R.; HOBBS, S.E.;.1990: Effect of roasted or raw
peanut kernels on lactation performance an milk composition of swine.
J. Anim. Sci. 68: 2591-2597

148
HENNIG, U.; SOUFFRANT, W.B.; HERRMANN, U.; 1990: Use of ileorectostomy and
fistulation on pigs for studies into precaecal digestion of crude protein and amino acid
absorbtion.
Arch. Exper. Vet. Med., Leipzig 44, 35-44
HENRY, R.W.; PICKARD, D.W.; HUGHES, P.E.; 1984: The effects of lactation length and food
level on subsequent reproductive performance in the sow.
Anim. Prod. 38: 527
HENRY, Y.; SÈVE, B.; COLLÈAUX, Y.; GANIÈR, P.; SALIGAUT, C.; JÈGO, P; 1992:
Interactive effects of dietary levels of tryptophan and protein on voluntary feed intake and
growth performance in pigs, in relation to plasma free amino acids and hypothalamic
serotonin.
J. Anim. Sci. 70: 1873-1887
HENRY, Y.; SÈVE, B.; MOUNIER, A.; GANIER, P.; 1996: Growth performance and brain
neurotransmitters in pigs as afected by tryptophan, protein, and sex.
J. Anim. Sci. 74: 2700-2710
HERRMANN, U.; SCHNEIDER, R.; 1981: Die Blutharnstoffkonzentration, ein Parameter zur
Beurteilung des Proteinstofwechsels bei tragenden Sauen.
Arch. Tierernährg. 31: 471-479
HILLER, J.; HILLER, J.G.; REDFERN, P.H.; 1074: The effect of hydrocortisone on tryptophan
metabolism in the rat.
J Pharm. Pharmac. 26, Suppl.: 118P-119P
HOPKINS, F.G.; COLE, S.W.; 1901: On the proteid reaction of Adamkiewicz, with contribution to
the chemistry of glyoxylic.
Proc. Royal soc. London 68: 21-33
HOUPT, R.T.; 1984: Controls of feeding in pigs.
J. Anim. Sci. 59: 1345

149
HUGHES, P.E.; HENRY, R.W.; PICKARD, D.W.; 1984: The effects of lactation food level on
subsequent ovulation rate and early embryonic survival in the sow.
Anim. Prod. 38: 527
HUTCHISON, S.N.; ZARGHAMI, N.S.; CUSICK, P.K.; LONGENECKER, J.B.; HASKELL, B.E.;
1983: The effect of valine deficiency on neutral amino acid pattern in plasma and brain of the
rat.
J. Nutr. 113: 2164-2170
HYLGAARD-JENSEN, J.; CMILJANIC, R.; WEGGER, I.; ANDRIC, R.; 1976: Tryptophan
metabolism in pigs. Relationship to synthesis and function of serotonin
Acta Vet. Scand. 17: 117-130
JAHN, B.M.; 2000: Stickstoff-Bilanzstudien zum Erhaltungsbedarf an essentiellen Aminosäuren
bei ausgewachsenen Sauen.
Dissertation TU München-Weihenstephan, Institut für Ernährungswissenschaften
JOHNSTON, L.J.; FOGWELL, R.L.; WELDON, W.C.; AMES, N.K.; ULLREY, D.E.; MILLER,
E.R.; 1989: Relationship between body fat and postweaning interval to estrus in primiparous
sows.
J. Anim. Sci. 67: 943-956
JOHNSTON, L.J.; PETTIGREW, J.E.; RUST, J.W.; 1993: Response of maternal-line sows to dietary
protein concentration during lactation.
J. Anim. Sci. 71: 2151-2156
JONES, D.B.; STAHLY, T.S.; 1999: Impact of amino acid nutrition during lactation on body
nutrient mobilization and milk nutrient output in primiparous sows.
J. Anim. Sci. 77: 1513-1522
KARLSON, P.; DOENECKE, D.; KOOLMAN, J.; 1994: Kurzes Lehrbuch der Biochemie für
Mediziner und Naturwissenschaftler.
14. Auflage, Georg Thieme Verlag, Stuttgart, New York

150
KING, R.H.; WILLIAMS, I.H.; 1984: The effect of nutrition on the reproductive performance of
first litter sows. 2. Protein and energy intakes during lactation.
Anim. Prod. 38: 249-256
KING, R.H.; DUNKIN, A.C.; 1986: The effect of nutrition on the reproductive performance of first
litter sows. 3. The relative effects of energy and protein intakes during lactation on the
performance of sowa and their piglets.
Anim. Prod. 43: 319-325
KING, R.H.; 1987: Nutritional anoestrus in young sows.
Pig News and Information 8: 15-22
KING, R.H.; TONER, M.S.; DOVE, H.; 1989: Pattern of milk production in sows. In: Manipulation
Pig Production II und V, Barret, J.L.; Hennessy, D.P., (ed.)
Australasian Pig Science Association, Werribee, Australia
KING, R.H.; RAYNER, C.J.; KERR, M.; 1993: A note on the amino acid composition of sows milk.
Anim. Prod. 57: 500-502
KING, R.H.; TONER, M.S.; DOVE, H.; ATWOOD, C.S.; BROWN, W.G.; 1993B: The response
of first-litter sows to dietary protein level during lactation.
J. Anim. Sci 71: 2457-2463
KIRCHGESSNER, M.; ROTH, F.X.; 1983: Schätzgleichung zur Ermittlung des energetischen
Futterwertes von Mischfuttermitteln für Schweine.
Z. Tierphysiol. Tierernährg. u. Futtermittelkunde 50: 270-275
KIRCHGESSNER, M.; MERK, L.; 1984: Vergleichende Untersuchungen zum Einfluß konstanter
und gestaffelter Energieversorgung in der Gravidität auf einige Reproduktionskriterien bei
Sauen.
J. Anim. Physiol. a. Anim. Nutr. 52: 124-136

151
KIRCHGESSNER, M.; KREUZER, M.; 1985: Harnstoff und Allantoin in der Milch von Kühen
während und nach der Verfütterung zu hoher und zu niedriger Proteinmengen.
Z. Tierphysiol. Tierernährg. u. Futtermittelkunde 54: 141-151
KIRCHGESSNER, M.; ROTH, F.X.; 1985: Biologische Wirksamkeit von DL-Tryptophan bei
Mastschweinen.
Z. Tierphysiol. Tierernährg. u. Futtermittelkunde 54: 135-141
KIRCHGESSNER, M.; 1987: Experimentelle Ergebnisse aus der ernährungsphysiologischen und
metabolischen Spurenelementforschung.
Übers. Tierernährg. 15: 153-192
KIRCHGESSNER, M.; PAULICKS, B.; SCHWARZ, F. J.; 1987: Veränderungen im
Harnstoffgehalt der Kuhmilch bei unzureichender und überhöhter Proteinversorgung.
J. Anim. Physiol. a. Anim. Nutr. 59: 79-84
KIRCHGESSNER, M.; RÄDER, G.; ROTH-MAIER, D.A.; 1991: Zum Einfluß einer variierten
Energieversorgung in der Gravidität und der Laktation auf die Milchleistung von Sauen.
Züchtungskunde 63: 136-145
KIRCHGESSNER, M.; SCHNEIDER, R.; SCHWARZ, F. J.; PAULICKS, B.; 1992: Milchleistung
sowie Milchfett- und energiegehalte bei Sauen mit unterschiedlicher Methioninversorgung.
2. Mitteilung zum Bedarf laktierender Sauen an schwefelhaltigen Aminosäuren.
J. Anim. Physiol. a. Anim. Nutr. 68: 244-253
KIRCHGESSNER, M.; SCHNEIDER, R.; SCHWARZ, F. J.; PAULICKS, B.; 1993: Aminosäuren-
und Harnstoffgehalte im Blutplasma laktierender Sauen bei unterschiedlicher
Methioninversorgung. 5. Mitteilung zum Bedarf laktierender Sauen an schwefelhaltigen
Aminosäuren.
J. Anim. Physiol. a. Anim. Nutr. 69: 194-202
KIRCHGESSNER, M.; 1997: Tierernährung: Leitfaden für Studium, Beratung und Praxis.
10.; neubearb. Aufl., BLV-Verl.-Ges., München

152
KIRCHGESSNER, M.; PAULICKS, B.R.; WESTERMEIER, C.; 1998: Konzentration an freien
Aminosäuren und Harnstoff im Blutplasma laktierender Sauen in Abhängigkeit von der
Threoninversorgung. 3. Mitteilung zum Threoninbedarf laktierender Sauen.
J. Anim. Physiol. a. Anim. Nutr. 80: 31-39
KIRKWOOD, R.N.; BAIDOO, S.K.; AHERNE, F.X.; SATHER, A.P.; 1987: The influence of level
of weight and backfat change during lactation on the subsequent reproductive performance
of sows.
Agricult. Forestry Bulletin 66: 119-120
KIRKWOOD, R.N.; MITARU, B.N.; GOONERATNE, A.D.; BLAIR, R.; THACKER, P.A.; 1988:
The influence of dietary energy intake during successive lactations on sow prolificacy.
Can. J. Anim. Sci. 68: 283-290
KLAVER, J.; VAN KAMPEN, G.J.M.; DE LANGE, P.G.B.; VERSTEGEN, M.W.A.; BOER, H.;
1981: Milk composition and daily yield of different milk components as affected by sows
condition and lactating/feeding regimen.
J. Anim. Sci. 52: 1091-1097
KLOBASA, F.; WERHAHN, E.; BUTLER, J.E.; 1987: Composition of sow milk during lactation.
J. Anim. Sci. 64: 1458-1466
KNABE, D.A.; BRENDEMUHL, J.H.; CHIBA, L.I.; DOVE, C.R.; 1996: Supplemental lysine for
sows nursing large litters.
J. Anim. Sci. 74: 1635-1640
KNOTT, P.J.; CURZON, G.; 1972: Free tryptophan in plasma and brain tryptophan metabolism.
Nature 239: 452-453
LAPLACE, J.P.; SOUFFRANT, W.B.; HENNING, U.; CHAUBEAUTI, E. ; FEVRIER, C. ; 1994 :
Measurement of precaecal dietary protein and plant cell wall digestion in ileorectostomized
pigs. Comparison of four surgical procedures.
Livestock Prod. Sci. 40, 313-328

153
LAWTON, C.L.; BLUNDELL, J.E.; 1989: Pharmacological manipulation of 5-HT: effect on intake of
diets supplemented with either sweet or bland carbohydrates. In: Behavioural pharmacology of
5-HT. Bevan, P.; Cools, A.R.; Archer, T. (eds.)
Lawrence Erlbaum; Hilldale, NJ
LAWTON, C.L.; BLUNDELL, J.E.; 1989B: Serotonin and carbohydrate demand � effects modulated
by degree of hydration of test diets.
Appetite 12: 222
LEATHWOOD, P.D.; 1987: Tryptophan availability and serotonin synthesis.
Proc. Nutr. Soc. 46: 143-156
LEHNINGER, A.; NELSON, D.; COX, M.; 2001: Lehninger Biochemie.
3. Auflage; Springer Verlag; Berlin
LEIBOWITZ, S.F.; WEISS, G.F.; SHOR-POSNER, G.; 1988: Hypothalamic serotonin:
pharmacological, biochemical and behavioral analyses of its feeding-suppressing action.
Clin. Neuropharmacol. 11: 51-71
LEIBOWITZ, S.F.; WEISS, G.F.; WALSH, U.A.; VISWANATH, D.; 1989: Medial hypothalamic
serotonin: role in circadian patterns of feeding and macronutrient selection.
Brain Res. 503: 132-140
LELLIS, W.A.; SPEER, V.C.; 1985: Aromatic amino acids requirement of lactating swine.
J. Anim. Sci. 61 (Suppl 1): 103
LELLIS, W.A.; SPEER, V.C.; 1987: Phenylalanine requirement of lactating sow.
J. Anim. Sci. 63: 1006-1012
LENKEIT, W.; GUETTE, J.O.; 1955: Langfristige Untersuchungen zum äußeren und inneren
Stoffwechsel des graviden und laktierenden Schweines. 1. und 2. Mitteilung.
Z. Tierphysiol., Tierern. u. Futtermittelk. 10: 94-102, 228-237

154
LENKEIT, W.; GUETTE, J.O; 1957: Leistungsbestimende Faktoren während der Laktation des
Schweines. 6. Mitteilung der �langfristigen Untersuchungen zum äußeren und inneren
Stoffwechsel des graviden und laktierenden Schweines�.
Z. Tierphysiol., Tierern. u. Futtermittelk. 12: 58-63
LEUNG, P.M.B.; ROGERS, Q.R.; 1973: Effect of amygdaloid lesions on dietary intake of
disproportionate amounts of amino acids.
Physiol. & Behav. 11: 221-226
LEWIS, A.J.; SPEER V.C.; 1973: Lysine requirement of the lactating sow.
J. Anim. Sci. 37: 104-110
LEWIS, A.J.; SPEER V.C.; 1974: Tryptophan requirement of the lactating sow.
J. Anim. Sci. 38 (4): 778-784
LEWIS, A.J.; SPEER V.C.; 1974B: Plasma amino acid response curves in lactating sows.
J. Anim. Sci. 38 (4): 785-789
LEWIS, A.J.; SPEER V.C.; 1975: Threonin requirement of the lactating sow.
J. Anim. Sci. 40: 895-899
LEWIS, A.J.; SPEER, V.C.; HAUGHT, G.T.; 1978: Relationship between yield and composition of
sow�s milk and weight gains of nursing pigs.
J. Anim. Sci. 47: 634-638
LIBAL G.W.; UTTRECHT, D.J.; HAMILTON, C.R.; 1997: Tryptophan needs of lactating sows
fed diets supplemented with crystalline lysine.
J. Anim. Sci. 75: 417-422
LIN, F.D.; SMITH, T.K.; BAYLEY, H.S.; 1988: A role for tryptophan in regulation of protein
synthesis in porcine muscle.
J. Nutr. 118: 445-449

155
LINZELL, J.L.; MEPHAM, T.B.; 1969: Mammary metabolism in lactating sows: arteriovenous
differences of milk precursors and the mammary metabolism of [14C]glucose and
[14C]acetate.
Br. J. Nutr. 23: 319-333
LODGE, G.A.; 1957: Milking sows by machine.
J. Agric. Sci. 49: 127-128
LODGE, G.A.; 1972: Quantitative aspects of nutrition in pregnancy and lactation.
Pig Production, Butterworth, London: 399-426
LÖFFLER, G.; PETRIDES, P.E.; 1997: Biochemie und Pathobiochemie. 5.Auflage.
Springer Verlag, Berlin Heidelberg New York
LYNCH, P.B.; 1977: Effect of enviromental temperature on lactating sows and their litters.
Ir. J. Agric. Res. 16: 123-130
LYNCH, P.B.; 1988: Voluntary food intake of sows and gilts. In: The voluntary food intake of pigs;
Forbes, M.A. (ed.)
BSAP Occ. Publ. No. 13: 71-77
MACKENZIE, D.D.S.; REVELL, D.K.; 1998: Genetic influences on milk quantity. In: The
lactating sow, Verstegen, M.W.A.; Moughan, P.J.; Schrama, J.W.; (ed.)
Wageningen Per, Wageningen
MAHAN, D.C.; GRIFO, A.P.; 1975: Effects of dietary protein levels during lactation to first-litter
sows fed a fortified corn gestation diet.
J. Anim. Sci. 41: 1362-1367
MAHAN, D.C.; MANGAN, L.T.; 1975: Evaluation of various protein sequences on the nutritional
carry-over from gestation to lactation with first litter sows.
J. Nutr. 105: 1291-1298

156
MALMLÖF, K.; ASKBRANT, S.; 1988: A note on the potential of systematic plasma urea
measurements as a basic for determining optimal supplementation levels of lysine and
threonine in pigs diet.
Swed. J. Agrc. Res. 18: 191-193
MAAß, N.; 2002: Experimentelle Untersuchungen zum alimentären Einsatz von Echinacea purpurea-
Cobs bei Schwein und Geflügel � Auswirkungen auf Leistungs- und Immunparameter.
Dissertation TU München-Weihenstephan, Department für Tierwissenschaften, Fachgebiet
Tierernährung
McARTHUR, R.A.; BLUNDELL, J.E.; 1986: Dietary self-selection and intake of protein and energy
is altered by the texture of the diets.
Physiol. Behav. 38: 315-319
McGIVAN, J.D.; PASTOR-ANGLADA, M.; 1994: Regulatory and molecular aspects of
mammalian amino acid transport.
Biochem. J. 299 (Pt. 2): 321
McGOWN, E.; RICHARDSON, A.G.; HENDERSON, L.M.; SWAN, P.B.; 1973: Effect of amino
acids in ribosome aggregation and protein synthesis in perfused rat liver.
J. Nutr. 103: 109-116
MEISINGER D.J.; SPEER V.C.; 1979: Tryptophan requirement for reproduction in swine.
J. Anim. Sci. 48 (3): 559-569
MENKE, K.-H.: 1980: Proteinverwertung und Blutharnstoffkonzentration.
Arch. Tierernährg. 30: 51-54
MENNENGA, V.L.O.; EASTER, R.A.; 1984: Lysine supplementation of low protein corn-soybean
meal diets for lactating swine.
J. Anim. Sci. 59: 246-247

157
MENNENGA, V.L.O.; EASTER, R.A.; 1985: Lysine and tryptophan supplementation of low protein
corn-soybean meal diets for lactating swine.
J. Anim. Sci. 61 (Supl. 1): 317
MERK, L.; 1982: Niveau- und Verteilungseinfluß einer in der Gravidität variierten
Energieversorgung auf einige Reproduktionskriterien bei Sauen.
Dissertation TU München-Weihenstephan, Institut für Ernährungsphysiologie
MÖHN, S.; 1993: Wirkung des Tryptophans auf Wachstum, Stoffansatz und Plasmakonzentration
von Hormonen beim Schwein.
Dissertation Universität Hohenheim, Fakultät IV � Agrarwissenschaften II
MORRIS, T.R.; 1983: The interpretation of response data from animal feedings trials. In: Recent
advances in animal nutrition; Haresign, W. (Hrsg.)
Butterworths, London: 13-23
MORRIS, T.R.; BLACKBURN, H.A.; 1982: The shape of the response curve relating protein intake to
egg output for flocks of laying hens.
Br. Poult. Sci. 23: 405-424
MOSER, S.A.; TOCKACH, M.D.; GOODBAND, R.D.; NELSSEN, J.L.; LOUGHMILLER, J.A.;
2000: The effects of branched-chain amino acids on sow and litter performance.
J. Anim. Sci. 78: 658-667
MOSES, P.L.; WURTMAN, R.J.; 1984: The ability of certain anorexic drugs to supress food
consumption depends on the nutrient composition of the test diet.
Life Sci. 35: 1297-1300
MOUGHAN, P.J.; SMITH, W.C.; 1996: Lactation. In: World animal science, C 10, Pig production;
Taverner, M.R., Dunkin, A.C. (eds.)
Elsevier, Amsterdam

158
MULLAN, B.P.; WILLIAMS, I.H.; 1989: The effect of body reserves at farrowing on the
reproductive performance of first-litter sows.
Anim. Prod. 48: 449-457
MULLAN, B.P.; WILLIAMS, I.H.; 1990: The chemical composition of sows during their first
lactation.
Anim. Prod. 51: 375-387
MULLAN, B.P.; BROWN, W.; KERR, M.; 1992: The response of the lactating sow to ambient
temperature.
Proc. Nutr. Soc. Aust. 17: 215
MUNRO, H.N.; 1970: Free amino acid pools and their role in regulation. In: Mammalian protein
metabolism; Munro H.N., (ed.), vol 4.
Academic Press, London, New York
MURPHY, D.L.; MENDELL, J.R.; ENGEL, W.K.; 1973: Serotonin and platelet function in Duchenne
Muscular Dystrophy.
Arch. Neurol. (Chic.) 28: 239-243
NAUMANN, C.; BASSLER, R.; unter Mitarbeit von SEIBOLD, R.; BARTH, C.; 1976 mit 1. und
2. Ergänzungslieferung 1983 und 1988: Methodenbuch Band 3. Die chemische Untersuchung
von Futtermitteln.
Verlag J. Neudamm-Neumann
NELSON, D.L.; COX, M.M.; 2001: Lehninger Biochemie. 3., vollständig überarbeitete und
erweiterte Auflage.
Springer-Verlag, Berlin Heidelberg New York
NELSON, G.H.; 1979: Biological effects of mycotoxins (other than aflatoxin) on swine.
In: Interactions of mycotoxins in animal production. Seite: 77-86
N. A. S., Washington D.C.

159
NEWBERRY; R.C.; WOOD-GUSH, D.G.M.; 1984: The Suckling Behavior of Domestic Pigs in a
Semi-Natural Enviroment.
Behav. 95: 11-25
NOBLET, J.; ETIENNE, M.; 1986: Effect of energy level in lactating sows on yield and
composition of milk and nutritent balance in pigs.
J. Anim. Sci. 63: 1888-1896
NOBLET, J.; DOURMAD, J.Y.; ETIENNE, M.; 1990: Energy utilization in pregnant and lactating
sows: Modeling of energy requirements.
J. Anim. Sci. 68: 562-572
NRC; NATIONAL RESEARCH COUNCIL, (ed.); 1988: Nutrient requirements of swine,
National Academy Press, ninth revised edition, Washington D.C.
NRC; NATIONAL RESEARCH COUNCIL, (ed.); 1998: Nutrient requirements of swine,
National Academy Press, tenth revised edition, Washington D.C.
OCHSENKÜHN, A.; 1988: Interaktion zwischen Adrenalin und Serotonin an der Isolierten
Ohrrandvene des Schweines.
Dissertation an der Medizinischen Fakultät der LMU München
O�GRADY, J.F.; HANRAHAN, T.J.; 1975: Influence of protein level and amino acid
supplementation of diets fed in lactation on the performance of sows and their litters. 1. Sow
and litter performance.
Ir. J. Agric. Res. 14: 127-135
O�GRADY, J.F.; LYNCH, P.B.; 1978: Voluntary feed intake by lactating sows: Influence of system
of feeding and nutrient density of the diet.
Ir. J. Agric. Res. 17: 1-5
O�GRADY, J.F.; LYNCH, P.B.; KEARNEY, P.A.; 1985: Voluntary feed intake by lactating sows.
Livest. Prod. Sci. 12: 355

160
OLDMAN, A.D.; WALSH, A.E.S.; SALKOVSKIS, P.; LAVER, D.A.; COWEN, P.J.; 1994: Effect of
acute tryptophan depletion on mood and appetite in healthy female volunteers.
Journal of Psychopharmacology 8 (1): 8-13
ORTHEN-GAMBILL, N.; KANAREK, R.B.: Differential effects of amphetamine and fenfluramine
on dietary self-selection in rats.
Pharmacol. Biochem. & Behav. 16: 303-309
OTT, H.; 2000: Experimentelle Untersuchungen zum Valinbedarf laktierender Zuchtsauen.
Dissertation TU München-Weihenstephan, Institut für Ernährungsphysiologie
PAULICKS, B.R.; 1985: Experimentelle Untersuchungen an laktierenden Kühen zur Veränderung
von Milchmenge und Milchinhaltsstoffen bei Proteinfehlernährung und nachfolgend
bedarfsgerechter Versorgung.
Dissertation TU München-Weihenstephan, Institut für Ernährungsphysiologie
PAULICKS, B.R.; WESTERMEIER, C.; KIRCHGESSNER, M.; 1998: Milchmenge und
Milchinhaltsstoffe bei Sauen in Abhängigkeit von der Threoninversorgung. 2. Mitteilung
zum Threoninbedarf laktierender Sauen.
J. Anim. Physoil. a. Anim. Nutr. 79: 102-111
PAULICKS, B.R.; WESTERMEIER, C.; KIRCHGESSNER, M.; 1998B: Abgestufte
Threoninversorgung bei laktierenden Sauen und Ihre Auswirkungen auf Milchleistung sowie
Harnstoff- und Threoninkonzentration in Milch und Blutplasma.
Proc. Soc. Nutr. Physiol. 7: 115
PEAKER, M.; 1977: The aqueous phase of milk: ion & water transport.
Symp. Zool. Soc. Lond. 41: 113-134
PEAKER, M.; 1978: Ion & water transport in the mammary gland. In: Lactation. A comprehensive
treatise. Vol. IV.; Larson, B.L. (ed.)
Academic Press, New York, London

161
PENG, Y.; TEWS, J.K.; HARPER, A.E.; 1972: Amino acid imbalance, protein intake and changes in
rat brain and plasma amino acids.
Am. J. Physiol. 222: 314-321
PERRIN, D.R.; 1955: The chemical composition of the colostrum and milk of the sow.
J. Dairy Res. 22: 103-107
PETTIGREW, J.E.; SOWER, A.F.; CORNELIUS, S.G.; MOSER, R.L.; 1985: A comparison of
isotope dilution and weigh-suckle-weigh methods for estimating milk intake by pigs.
Can. J. Anim. Sci. 65: 989-992
PLUSKE, J.R.; WILLIAMS, I.H.; ZAK, L.J.; CLOWES, E.J.; CEGIELSKI, A.C.; AHERNE, F.X.;
1998: Feeding primiparous lactating sows to induce three divergent metabolic states. III.
Milk production and piglet growth.
J. Anim. Sci. 76: 1165-1171
POGSON, C.I.; MUNOZ-CLARES, R.A.; COOK, F.S.; SMITH, S.A.; 1984: Tryptophan
metabolism and its control in mammalian liver. In: Progress in Tryptophan and Serotonin
Research, Seiten: 625-632
de Gruyter & Co., Berlin, New York
POLLOCK, J.D.; ROWLAND, N.; 1981: Peripherally administered serotonin decreases food intake in
rats.
Pharmacology Biochemistry & Behavior 15: 179-183
POND, W.G.; YEN, J.T.; MAURER, R.R.; CHRISTENSON, R.K.; 1981: Effect of doubling daily
energy intake during the last two weeks of pregnancy on pig birth weight, survival and
weaning weight.
J. Anim. Sci. 52: 535-541
PONTER, A.A.; CORTAMIRA, N.O.; SEVE, B.; SALTER, D.N.; MORGAN, L.M.; 1994: The
effects of energy source and tryptophan on the rate of protein synthesis and on hormones of
the enteroinsular axis in the piglet.
Br. J. Nutr. 71: 661-674

162
POPPE, S.; WIESEMÜLLER, W.; 1968: Untersuchungen über den Aminosäurenbedarf
wachsender Schweine. 1. Mitteilung: Bestimmung des Lysinbedarfes mit Hilfe der N-
Bilanzmethode
Arch. Tierern. 18: 392-404
PRECH, M.; KRAFT, R.; 1993: Biostatistik 2.
5., völlig überarb. Aufl., Oldenbourg Verlag, München
PRELUSKY, D.B.; 1994: The effect of deoxynivalenol on serotonergic neurotransmitter levels in pig
blood.
J. Environ. Sci. Health B29(6): 1203-1218
RÄDER, G.; 1989: Untersuchungen zur Milchleistung der Sau bei variierter Energieversorgung in
der Gravidität, bzw. bei Argininzulage in der Laktation.
Dissertation TU München-Weihenstephan, Institut für Ernährungsphysiologie
RÄDER, G.; ROTH-MAIER, D.A.; KIRCHGESSNER, M..; 1990: Untersuchungen zur Erfassung der
Milchleistung und der Milchinhaltsstoffe bei Sauen.
Agribiol. Res. 43: 191-199
REESE, D.E.; MOSER, B.D.; PEO, E.R.; LEWIS, A.J.; 1982: Influence of energy intake during
lactation on subsequent gestation, lactation and postweaning performance of sows.
J. Anim. Sci. 55: 867
REVELL, D.K.; WILLIAMS, I.H.; MULLAN, B.P.; RANFORD, J.L.; SMITS, R.J.; 1998: Body
composition at farrowing and nutrition during lactation affect the performance of
primiparous sows: I. Voluntary feed intake, weight loss, and plasma metabolites.
J. Anim. Sci. 76: 1729-1737
REVELL, D.K.; WILLIAMS, I.H.; MULLAN, B.P.; RANFORD, J.L.; SMITS, R.J.; 1998B: Body
composition at farrowing and nutrition during lactation affect the performance of
primiparous sows: II. Milk composition, milk yield, and pig growth.
J. Anim. Sci. 76: 1738-1743

163
RICHARD, J.; WURTMAN, M.D.; 1980: Nutritional control of brain tryptophan and serotonin. In:
Biochemical an medical aspects of tryptophan metabolism, Hayaishi, O.; Ishimura, Y.; Kido,
R.; (eds.)
Elsevier/North-Holland Biomedical Press, Amsterdam
RICHERT, B.T.; TOKACH, M.D.; GOODBAND, R.D.; NELSSEN, J.L.; PETTIGREW, J.E.;
WALKER, R.D.; JOHNSTON, L.J.; 1996: Valine requirement of high-producing lactating
sow.
J. Anim. Sci. 74: 1307-1313
RICHERT, B.T.; GOODBAND, R.D.; TOKACH, M.D.; NELSSEN, J.L.; 1997: Increasing valine,
leucine, and total branched amino acids for the lactating sow.
J. Anim. Sci. 75: 2117-2128
ROBBINS, K.R.; NORTON, H.W.; BAKER, D.H.; 1979: Estimation of Nutrient Requirements
from Growth Data.
J. Nutr. 109: 1710-1714
ROGERS, Q.R.; LEUNG, P.M.B.; 1973: The influence of amino acids on the neuroregulation of food
intake.
Fed. Proc. 32: 1709-1719
ROTH, F.X.; KIRCHGESSNER, M.; 1987: Biological efficiency of dietary methionine or cystine
supplementation with growing pigs. � A contribution to the requirement for S-containing
amino acids.
J. Anim. Physiol. a. Anim. Nutr. 58: 267-280
ROTH, F.X.; ETTLE, T.; 2003: Specific dietary selection for tryptophan by the piglet.
noch unveröffentlichtes Manuskript (Fachgebiet Tierernährung und Leistungsphysiologie,
Technische Universität München, Hochfeldweg 6, D-85350 Freising)

164
ROTH-MAIER, D.A.; KIRCHGESSNER, M.; ERHART, W.; HENKE, J.; HENNING, U.; 1998:
Comparative studies for the determination of precaecal digestibility as a measure for the
availability of B-vitamins.
J. Anim. Physiol. A. Anim. Nutr. 79: 198-209
ROTHWELL, S.W.; FUDGE, J.M.; CHEN, W-K.; REID, T.J.; KRISHNAMURTI, C.; 2002: Addition
of a propyl gallate-based procoagulant to a fibrin bandage improves hemostatic performance in
a swine arterial bleeding model.
Thrombosis research 108 (5-6): 335-340
ROJKITTIKUHN, T.; EINARSSON, S.; UVNÄS-MOBERG, K.; EDQVIST, L.-E.; 1992: Body
weight loss during lactation in relation to energy and protein metabolism in standard-fed
primiparous sows.
J. Vet. Med. 40: 249-257
ROUSSELOW, D.L.; SPEER, V.C.; HAUGHT, D.G.; 1979: Leucin requirement of the lactating
sow.
J. Anim. Sci. 49: 498-506
ROUSSELOW, D.L.; SPEER, V.C.; 1980: Valine requirement of lactating sow.
J. Anim. Sci. 49: 498-506
ROWLINSON, P.; BRYANT, M.J.; 1982: Lactational oestrus in the sow. 2. Influence of group
housing, boar presence and feeding level upon the occurrence of oestrus in lactating sows.
Anim. Prod. 34: 283-290
SALTER, M.S.; BENDER, D.A.; POGSON, C.I.; 1985: Leucine and Tryptophan metabolism in
rats.
Biochem. J. 225: 277-281
SARRIAS, M.J.; MARTINEZ, E.; CELADA, P.; UDINA, C.; ALVAREZ, E.; ARTIGAS, F.; 1991:
Plasma free 5HT and platelet 5HT in depression: case control studies and the effect of
antidepressant therapy. In: Kynurenine and serotonin pathways, Schwarcz et al., (ed.)
Plenum Press, New York

165
SAUBER, T.E.; STAHLY, T.S.; EWAN, R.C.; WILLIAMS, N.H.; 1994: Interactive effects of sow
genotype and dietary amino acid intake on lactational performance of sows nursing large litters.
J. Anim. Sci. 72 (Suppl. 2): 66
SCHAHRZAD, H.; 1994: Untersuchungen zur Methodik der Tryptophananalytik in tierischen
Geweben sowie zum Tryptophanumsatz laktierender Sauen.
Dissertation an der Humboldt-Universität zu Berlin, Landwirtschaftlich-Gärtnerische
Fakultät
SCHARRER, E.; 1986: Epithelialer Transport von Aminosäuren und Peptiden.
Übers. Tierernährg. 14: 233-250
SCHIEMANN, R.; 1981: Methodische Richtlinien zur Durchführung von Verdauungsversuchen für
die Futterwertschätzung.
Arch. Tierernährung, Bd. 31, H1, 1-19
SCHNEIDER, R.; 1992: Experimentelle Untersuchungen zum Bedarf schwefelhaltiger Aminosäuren
laktierender Zuchtsauen.
Dissertation TU München-Weihenstephan, Institut für Ernährungsphysiologie
SCHNEIDER, R.; KIRCHGESSNER, M.; SCHWARZ, F.J.; PAULICKS, B.R.; 1992:
Futteraufnahme und Lebendmasseentwicklung von Sauen während der Laktation in
Abhängigkeit von der Methioninversorgung. 1. Mitteilung zum Bedarf laktierender Sauen
an schwefelhaltigen Aminosäuren.
J. Anim. Physiol. a. Anim. Nutr. 68: 235-243
SCHNEIDER, R.; KIRCHGESSNER, M.; PAULICKS, B.R.; SCHWARZ, F.J.; 1992B: Eiweiß-
und Aminosäurengehalte in der Sauenmilch bei unterschiedlicher Methioninversorgung. 3.
Mitteilung zum Bedarf laktierender Sauen an schwefelhaltigen Aminosäuren.
J. Anim Physiol. a. Amin. Nutr. 68: 254-262

166
SCHOENHERR, W.D.; STAHLY, T.S. CROMWELL, G.L.; 1989: The effects of dietary fat or
fiber addition on yield and composition of milk from sows housed in a warm or hot
environment.
J. Anim. Sci. 67: 482-495
SCHUTTE, J.B.; VAN WEERDEN, E.J.; KOCH, F.; 1988: Utilization of DL- and L-Tryptophan in
young pigs.
Anim. Prod. 46: 447-452
SCHWARZ, F.J.; SCHNEIDER, R.; PAULICKS, B.R.; KIRCHGESSNER, M.; 1993:
Lebendgewichtsentwicklung und Beifutteraufnahme von Saugferkeln in Abhängigkeit von
der Methioninversorgung der Sau. 4. Mitteilung zum Bedarf laktierender Sauen an
schwefelhaltigen Aminosäuren.
J. Anim. Physiol. a. Anim. Nutr. 69: 156-161
SEBEKOVA, K.; SPUSTOVA, V.; OPATRNY, K.(Jr.); DZURIK, R.; 2001: Serotonin and 5-
hydroxyindole-acetic acid.
Bratisl. Lek. Listy 102 (8): 351-356
SHIELDS, R.G.; MAHAN, D.C.; MAXSON, P.F.; 1985: Effect of dietary gestation and lactation
protein levels on reproductive performance and body composition of first-litter female swine.
J. Anim. Sci. 60: 179-189
SIDRANSKY, H.; VERNEY, E.; SHARMA, D.S.R.; 1971: Effect of tryptophan on polysomes and
protein synthesis in liver.
Am. J. Clin. Nutr. 24: 779-785
SIMANSKY, K.J.; 1989: Modulation of nocturnal free feeding by drugs acting on serotoninergic
neuronal processes in rats.
Ann. NY Acad. Sci. 575: 572-574
SMITH, D.M.; WHITTLESTONE, W.G.; ALLEN, J.; 1951: The design and use of a milking
machine for sows.
J. Dairy Res. 18: 31-34

167
SMITH, S.A.; POGSON, C.I.; 1980: The metabolism of L-Tryptophan by isolated rat liver cells.
Biochem. J. 186: 977-986
SOHAIL, M.A.; COLE, D.J.A.; LEWIS, D.; 1978: Amino acid requirements of the breeding sow:
The dietary lysine requirement of the lactating sow.
Br. J. Nutr. 40: 369-376
SPEER, V.C.; COX, D.F.; 1984: Estimating milk yields of sows.
J. Anim. Sci. 59: 1281-1285
SPINCER, J.; ROOK, J.A.F.; TOWERS, K.G.; 1968: The uptake of plasma constituents by the
mammary gland of sows.
Biochem. J. 111: 727-733
SPINKA, M.; ILLMANN, G.; ALGERS, B.; STETKOVA, Z.; 1997: The role of nursing frequency in
milk production in domestic pigs.
J. Anim. Sci. 75: 1223-1228
STEINHART, H.; 1979: Tryptophan-Bestimmung in biologischem Material, zugleich ein Beitrag
zur Toxizität von Tryptophan-Oxidationsprodukten im tierischen Organismus.
Habilitation TU München-Weihenstephan, Institut für Ernährungsphysiologie
STEINHART, H.; 1984: Tryptophan requirement of farm animals. in:
Progress in Tryptophan and Serotonin Research, Seiten: 537-544
de Gruyter & Co., Berlin, New York
STERLING, L.G.; CERA, K.R.; 1990: The effect of dietary lysine level during lactation on milk
composition and litter gain efficiency over successive reproductive cycles.
J. Anim. Sci. 68 (Suppl.): 365-366
SUSENBETH, A.; 2002: Ein Beitrag zur Frage der Proteinbewertung und Aminosäurenversorgung
beim Schwein. In: Tagungsband: 7. Tagung Schweine- und Geflügelernährung, 26.-28.
November 2002 Lutherstadt Wittenberg; Rodehutscord, M. (Hrsg.)
Universitätsdruckerei Martin-Luther-Universität Halle Wittenberg

168
TAGLIAMONTE, A.; BIGGIO, G.; VARGIU, L.; GESSA, G.L.; 1973: Free tryptophan in serum
controls brain tryptophan level an serotonin synthesis.
Life Sci. 12 Part II: 277-287
TANEN, D.A.; DANISH, D.C.; CLARK, R.F.; 2003: Crotalidae polyvalent immune fab antivenom
limits the decrease in perfusion pressure of the anterior leg compartment in a porcine crotaline
envenomation model.
Annals of Emergency Medicine 41 (3): 384-390
TANKSLEY, T.D.; KNABE, D.A.; 1984: Ileal digestibilities of amino acids in pig feeds and their
use in formulating diets. In: Recent Advances in Animal Nutrition.
Butterworth, London
TEIK-NG, L.; ANDERSON, G.H.; 1992: Route of administration of tryptophan and tyrosine affects
short-term food intake and plasma and brain amino acid concentrations in the rat.
J. Nutr. 122: 283
TOKACH, M.D.; PETTIGREW, J.E.; CROOKER, B.A.; DIAL, G.D.; SOWER A.F.; 1992:
Quantitative influence of lysine and energy intake on yield of milk components in the
primiparous sow.
J. Anim. Sci. 70: 1864-1872
TONER, M.S.; KING, R.H.; DUNSHEA, F.R.; DOVE, H.; ATWOOD, C.S.; 1996: The effect of
exogenous somatotropin on lactation performance of first litter sows.
J. Anim. Sci. 70: 167-172
TROTTIER N.L.; EASTER R.A.; 1995: Dietary and plasma branched-chain amino acids in relation
to tryptophan: effect on voluntary feed intake and laktation metabolism in the primiparous
sow.
J. Anim. Sci. 73: 1086-1092
TSCHOPP, T.B.; ZUCKER, M.B.; 1972: Hereditary defect in platelet function in rats.
Blood 40: 217-226

169
UTTECHT, D.J.; HAMILTON, C.R.; WEAVER, E.M.; LIBAL, G.W.; 1992: Relationship between
dietary tryptophan and other large neutral amino acids for growing and finishing pigs.
J. Anim. Sci. 70 (Suppl. 1): 65
VAN DER PEET-SCHWERING, C.M.C.; SWINKELS, J.W.G.M.; DEN HARTOG, L.A.; 1998:
Nutritional strategy and reproduction. In: The lactatin sow, Verstegen, M.W.A.; Moughan,
P.J.; Schrama, J.W.; (ed.)
Wageningen Per, Wageningen
VAN DER STEEN, H. A. M.; 1985: Maternal influence mediated by litter size during the suckling
period on reproduction traits in pigs.
Livest. Prod. Sci. 13: 147-158
VAN LEEUWEN, P.; SAUER, W.C.; HUISMAN, J.;VAN WEERDEN, E.J.; VAN KLEEF, D.;
DEN HARTOG, L.; 1987: Methodological aspects for the determination of amino acid
digestibilities in pigs fitted with ileorectal re-entrant cannulas.
J. Anim. Physiol. a. Anim. Nutr. 58: 122-133
VAN SPAENDONK, R.A.; VAN SCHOUBROEK, F.; 1964: Determination of the milk yield of
sows and correction for loss of weight due to metabolic process of piglets during suckling.
Anim. Prod. 6: 119-123
VERTESY, E.; 1996: Experimentelle Untersuchungen zu Selenstatus, Milchmenge, -
zusammensetzung und Aufzuchtleistung von Zuchtsauen bei unterschiedlicher Selenzufuhr
während der Laktation.
Dissertation TU München-Weihenstephan, Institut für Ernährungsphysiologie
VERSTEGEN, M.W.A.; MESU, J.; VAN KEMPEN, G.J.M.; GEERSE, C.; 1985: Energy balances
of lactating sows in relation to feeding level and stage of lactation.
J. Anim. Sci. 60: 731-740
VERSTEGEN, M.W.A.; MOUGHAN, P.J.; SCHRAMA, J.W.; (ed.); 1998: The lactating sow.
Wageningen Per, Wageningen

170
VOET, D.; VOET, J.G.; 1994: Biochemie.
1. korregierter Nachdruck der 1. Auflage; VCH Verlagsgesellschaf, Weinheim
WALKER, N.; WATT, D.; MACLEOD, A.S.; JOHNSON, C.L.; BOAZ, T.G.; CALDERA, A.F.C.;
1979: The effect of weaning at 10, 25 or 40 days on the reproductive performance of sows
from the first to the fifth parity.
J. agric. Sci., Camb. 92: 449-456
WALTHER, D.J.; PETER, J.-U.; BASHAMMAKH, S.; HÖRTNAGEL, H.; VOITS, M.; FINK, H.;
BADER, M.; 2003: Synthesis of serotonin by a second tryptophan hydroxylase isoform.
Science 299 (5603): 76
WANG, T.C.; FULLER, M.F.; 1989: The optimum dietary amino acid pattern for growing pigs. 1.
Experiments by amino acid deletion.
Br. J. Nutr. 62: 77-89
WECHSLER, B.; BRODMANN, N.; 1996: The synchronisation of nursing bouts in group housed
sows.
Appl. Anim. Behav. Sci. 47: 191-199
WELDON, W.C.; LEWIS, A.J.; LOUIS, G.F.; KOVAR, J.L.; GIESEMANN, M.A.; MILLER, P.S.;
1993: Postpartum hypophagia in primiparous sows: I. Effects of gestation feeding level on
feed intake, feeding behavior and plasma metabolite concentrations during lactation.
J. Anim. Sci. 72: 387-394
WESTERMEIER, C.; 1997: Experimentelle Untersuchungen zum Threoninbedarf laktierender
Zuchtsauen.
Dissertation TU München-Weihenstephan, Institut für Ernährungsphysiologie
WESTERMEIER, C.; PAULICKS, B.R.; KIRCHGESSNER, M.; 1998: Zur Threoninversorgung
laktierender Sauen und ihrem Einfluß auf die Lebendmasseentwicklung von Sauen und
Ferkeln.
Proc. Soc. Nutr. Physiol. 7: 120

171
WHITE, J.G.; 1968: The dense bodies of human platelets: Origin of serotonin particles from platelet
granules.
Am. J. Path. 53: 791-808
WHITTEMORE, C.T.; ELSLEY, F.W.H.; 1976: Practical pig nutrition.
Farming Press, Ipswich
WILKINSON, R.H.; COLE, D.J.A.; LEWIS. D.; 1982: Amino acid nutrition of the lactating sow:
the requirement for dietary lysine.
Anim. Prod. 35: 15-23
WILLIAMS, I.H.; 1998: Nutritional effects during lactation and during the interval from weaning to
oestrus. In: The lactating sow. Verstegen, M.W.A.; Moughan, P.J.; Schrama, J.W.; (ed.)
Wageningen Per, Wageningen
WOEHLBIER, W.; 1928: Stoffwechselversuche zum Eiweißansatz bei säugenden Ferkeln.
Biochem. Ztschr. 202: 29-69
WU, G.; KNABE, D.A.; 1994: Free and protein bound amino acids in sow´s colostrum and milk.
J. Nutr. 124: 415-424
WURTMAN, R.J.; 1980: Nutritional control of brain tryptophan and serotonin. In: Biochemical and
medical aspects of tryptophan metabolism. Seiten: 31-46 Hayaishi, O.; Ishimura, Y.; Kido, R.;
(eds.)
Elsevier, North Holland
WURTMAN, R.J.; HEFTI, F.; MELAMED, E.; 1981: Precursor control of neurotransmitter
synthesis.
Pharmacol. Rev. 32 (4): 315-335
WURTMAN, R.J.; WURTMAN, J.J.; 1996: Brain serotonin, carbohydrate-craving, obesty and
depression. In: Recent advances in tryptophan research, Graziella Filippini et al., (ed.)
Plenum Press, New York

172
YANG, T.S.; HOWARD, B.; MAC FARLANE, W.V.; 1980: A note on milk intake of piglets
measuren by tritium dilution.
Anim. Prod. 31: 201-203
YOKOGOSHI, H.; YOSHIDA, A.; 1980: Effects of supplementation and depletion of a single
essential amino acid on hepatic polyribosome profile in rats.
J. Nutr. 110: 375-382
YOKOGOSHI, M.T.; WURTMAN, R.J.; 1986: Meal composition and plasma amino acid ratios:
Effect of various proteins or carbohydrates, and of various protein concentrations.
Metabolism 35 (9): 837-842
YOUNG, V.R.; MARCHINI, J.S.; 1990: Mechanisms and nutritional significance of metabolic
responses to altered intakes of protein and amino acids, with reference tonutritional
adaptation in humans.
Am. J. Clin. Nutr. 51: 270-289
ZHANG, R.; 1991: Untersuchungen zum Tryptophanbedarf von Ferkeln bei unterschiedlicher
Lysinversorgung.
Dissertation Universität Hohenheim, Fakultät IV � Agrarwissenschaften II
ZWISCHENBERGER, J.B.; TAO, W.; DEYO, D.J.; VERTREES, R.A.; ALPARD, S.K.;
SHULMAN, G.; 2001: Safety and efficacy of a heparin removal device: a prospective
randomized preclinical outcomes study.
The Annals of Thoracic Surgery 71 (1): 270-277

173
9. Anhang Anhangstabelle 1: Zusammensetzung, Inhalts- und Zusatzstoffe des Mineralfutters
für tragende und laktierende Zuchtsauen
Mineralfuttermittel*) für Schweine
Inhaltsstoffe
Calcium Phosphor Natrium Magnesium
22,0 % 6,0 % 5,0 % 1,0 %
Zusatzstoffe
Vitamin A Vitamin D3 Vitamin E Vitamin K3 Vitamin B1 Vitamin B2 Vitamin B6 Vitamin B12 Nikotinsäure Pantothensäure Folsäure Cholinchlorid Biotin Eisen Zink Kupfer Mangan Jod Selen
600000 I.E. 60000 I.E.
2000 mg 35 mg 35 mg
125 mg 75 mg
1000 mcg 600 mg 300 mg 100 mg
15000 mg 8000 mg 5000 mg 5000 mg 1000 mg 2000 mg
60 mg 13 mg
Zusammensetzung
Calciumcarbonat Natriumchlorid Mono-Calcium-Phosphat Calcium-Magnesium-Phosphat Mono-Di-Calcium-Phosphat Natrium-Calcium-Magnesium-Phosphat Spurenelementvormischung Vitaminvormischung
*): �MIN 20 ZUCHT�; Raiffeisen Kraftfutterwerke Süd GmbH; Würzburg

174
Anhangstabelle 2: Zusammensetzung, Inhalts- und Zusatzstoffe des
Ergänzungsfuttermittels für Ferkel laut Herstellerangaben
Ergänzungsfutter*) für Ferkel
Inhaltsstoffe
Rohprotein Lysin Rohfett Rohfaser Rohasche Gesamtzucker Calcium Phosphor Natrium
20,00 %1,60 %8,50 %2,50 %6,00 %
10,00 %0,75 %0,60 %0,40 %
Zusatzstoffe je kg FM Vitamin A Vitmin D3 Vitamin E Kupfer Selen Antioxidans BHT Fumarsäure Zitronensäure Milchsäure Einheiten (FTU) Phytase Mischung aus Bacillus licheniformis (DSM5749) und Bacillus subtilis (DSM5750) im Verhältnis 1:1
225002000200 165
0
500
1,28E+
I.E.I.E.Mgmg
,4 mg 0,9 KBE
Zusammensetzung Haferflocken Mais (aufgeschlossen) Sojaproteinkonzentrat Weizen (aufgeschlossen) Magermilchpulver Gerste Saccarose Kartoffeleiweiß Molkenpulver (teilentzuckert) Volleipulver Maiskleber
Molke-Fett-Konzentrat Pflanzenöl Johannisbrot Vormischung Monodicalciumphosphat Calciumcarbonat Natriumchlorid Lysin-HCL DL-Methionin L-Threonin L-Tryptophan
*): �DEUKA PRIMO WEAN�; deuka Deutsche Kraftfutterwerke GmbH & Co; Düsseldorf

175
Anhangstabelle 3: Abgesetzte Kot- und Harnmassen in Abhängigkeit der Ferkelmasse in g
Ferkel Masse Kotmasse Ferkel Masse Harnmasse 1 1878 4 1 2457 10 2 2446 2 2 2682 45 3 2454 2 3 2904 25 4 2905 5 4 3017 25 5 2938 3 5 3046 35 6 3264 2 6 3441 30 7 3427 2 7 3455 25 8 3749 5 8 3687 45 9 3818 4 9 3758 40
10 3935 3 10 3832 25 11 4043 3 11 3939 40 12 4065 2 12 4026 35 13 4136 8 13 4102 40 14 4317 3 14 4102 45 15 4343 7 15 4164 50 16 4405 3 16 4283 65 17 4452 7 17 4405 30 18 4496 5 18 4504 20 19 4530 5 19 4621 33 20 4644 2 20 4748 35 21 4670 6 21 4891 60 22 4673 9 22 4935 60 23 4982 6 23 4985 95 24 5143 7 24 5017 35 25 5156 7 25 5144 10 26 5227 4 26 5268 40 27 5251 15 27 5318 45 28 5292 3 28 5381 40 29 5301 10 29 5399 35 30 5395 10 30 5448 55 31 5454 4 31 5463 30 32 5503 4 32 5501 65 33 5573 4 33 5878 35 34 5641 8 34 5891 40 35 5724 10 35 6203 60 36 5783 15 36 6231 80 37 5891 2 37 6345 50 38 6059 10 38 6345 50 39 6263 7 39 6413 55 40 6442 7 40 6449 40 41 6532 6 41 6973 40 42 6743 6 42 7007 120 43 6901 5 43 7133 45
44 7140 50 45 7142 45 46 7340 45

176
Anhangstabelle 4: Zusammensetzung der Amionosäurenvormischung des Trächtigkeits-
futters
Komponente
Anteil in %
L-Lysin HCl
DL-Methionin
L-Threonin
Wintergerste
29,71
2,99
2,83
64,47 Anhangstabelle 5: Zusammensetzung der Vitaminvormischung des Trächtigkeitsfutters
Komponente
Anteil in %
Vitamin B12
Cholinchlorid
Monocalciumphosphat
Natriumchlorid
Wintergerste
0,05
10,08
29,12
19,11
41,64 Anhangstabelle 6: Zugesetzte Aminosäuren je kg Laktationsfutter in der Grundration
Komponente
Zusatz in g
L-Lysin HCl
DL-Methionin
L-Threonin
L-Isoleucin
L-Phenylalanin
3,8
2,6
2,2
0,8
3,0

177
Anhangstabelle 7: Milchleistung der Sauen in g
Gruppe Sau 13. LT 14. LT 20. LT 21. LT
I Stilla 3452 3502 4264 3158
I Alfi 5881 5772 6283 6422
I Alpin 7091 7107 8574 8561
I Amper 6648 6053 7235 7032
I Arne 7934 8964 7123 6207 I Lupine 7893 8706 7462 5027
I Anneli II 8263 9078 6972 7558
I Isabell 10527 8793 8467 7006
I Silvi II 6113 6562 7111 8392
I Alfreda 8466 8650 7603 6961
I Raupe 4710 4046 5943 5594
I Asti 7444 8138 7837 7348
II Irka 5454 5131 6986 5904 II Siegrid 8182 10563 9132 11074
II Ronja 7113 8796 7231 7611
II Saba 9209 9590 10999 9233
II Alfa 5976 5624 7853 7369
II Saale 8281 8096 8338 8339
II Rubi II 8288 8476 9665 9047
II Alpe 7091 7107 8574 8561
II Angela 10443 9467 7558 9926 II Ida II 6087 6375 7578 6978
II Alida 9956 9963 8593 8279
II Samanta 6511 6214 6124 5639
III Alpha 7154 7705 8652 10026
III Luzi 6705 7412 9769 8922
III Anne 7944 9209 8851 9847
III Albi 6656 8614 9065 10170 III Ilse II 7055 7834 5139 3158
III Almi 6183 6220 7574 7069
III Anka 10522 9085 8264 7737
III Sasse 7236 6414 8997 9935
III Sina 8699 8205 8675 9412
III Reni 8052 8757 8583 8469
III Anna 4855 5060 5924 5789
III Rapunzel 7628 8902 10295 10344

178
Gruppe Sau 13. LT 14. LT 20. LT 21. LT
IV Antje 9289 9250 9250 9101
IV Anni 6696 9022 9022 7057
IV Randa 7985 7678 7678 10356
IV Sinta 6424 7476 7476 11136 IV Anja 7733 6239 6239 9287
IV Siegrid II 8480 8195 8195 8638
IV Isolde 6895 7083 7083 5823
IV Sabrina 9580 9894 9894 9903
IV Simone 8753 7315 7315 8717
IV Amper II 6068 6111 6111 9593
IV Leila 8323 9549 9549 9183 IV Alfa II 6225 6501 6501 8684
V Rubi 6996 7149 6942 7444
V Ilse 9926 9659 7424 8491
V Rassel 8419 10040 10911 12078
V Ida 6127 7466 5445 6390
V Andex 7154 8603 7091 6445
V Rosl 8053 9843 10605 10988
V Inka 6750 6663 10019 8999 V Rasso 8472 7214 8832 8450
V Luna 6605 6086 8960 7412
V Luzi II 9582 9517 9932 10514
V VS 8 6852 7801 8316 7827
V Struppi 9952 9649 9244 8070
VI Anneli 8011 8267 8406 8858
VI Silvi 6713 7324 8514 8558 VI Lunte 9246 8391 10438 10062
VI Iska 6341 5750 3767 4180
VI Astrit 5671 6413 7852 8341
VI Sterzi 6332 5461 7020 6447
VI Alfi II 7096 7708 8976 7113
VI Rosita 9227 10380 11109 11446
VI Rosa 8253 10448 9223 10015
VI Sandra 8846 7235 8739 9958 VI Renate 8735 7826 7703 7384
VI Albi II 5921 6156 5430 5802

179
Anhangstabelle 8: Serotoningehalt am 85. TT und am 28. LT in ng / 109 Thrombozyten
Gruppe Sau 85. TT 28. LT relativ in % I Silvi 6971 2142 -69,3 I Amper 2436 1122 -53,9 I Arne 2752 1215 -55,9 I Lupine 2393 1446 -39,6 I Anneli II 3911 885 -77,4 I Isabell 3043 627 -79,4 I Alfreda 1485 766 -48,4 I Asti 1847 1183 -36,0 I Raupe 1529 1035 -32,3 II Ronja 1294 541 -58,2 II Saba 4808 914 -81,0 II Alfa 2318 1216 -47,6 II Saale 1231 608 -50,7 II Rubi II 1319 400 -69,6 II Alpe 4367 946 -78,3 II Angela 2276 874 -61,6 II Ida 1504 700 -53,5 II Alida 3003 770 -74,3 II Saale 2499 1083 -56,7 III Alpha 927 651 -29,7 III Luzi 2544 1145 -55,0 III Anne 2306 887 -61,5 III Albi 1754 1082 -38,3 III Ilse II 4008 1867 -53,4 III Almi 2713 1594 -41,3 III Anka 1976 1536 -22,3 III Saba 4455 4804 7,8 III Sina 6734 3566 -47,0 III Reni 1568 1555 -0,8 III Anne 3483 1853 -46,8 III Rapunzel 984 952 -3,3
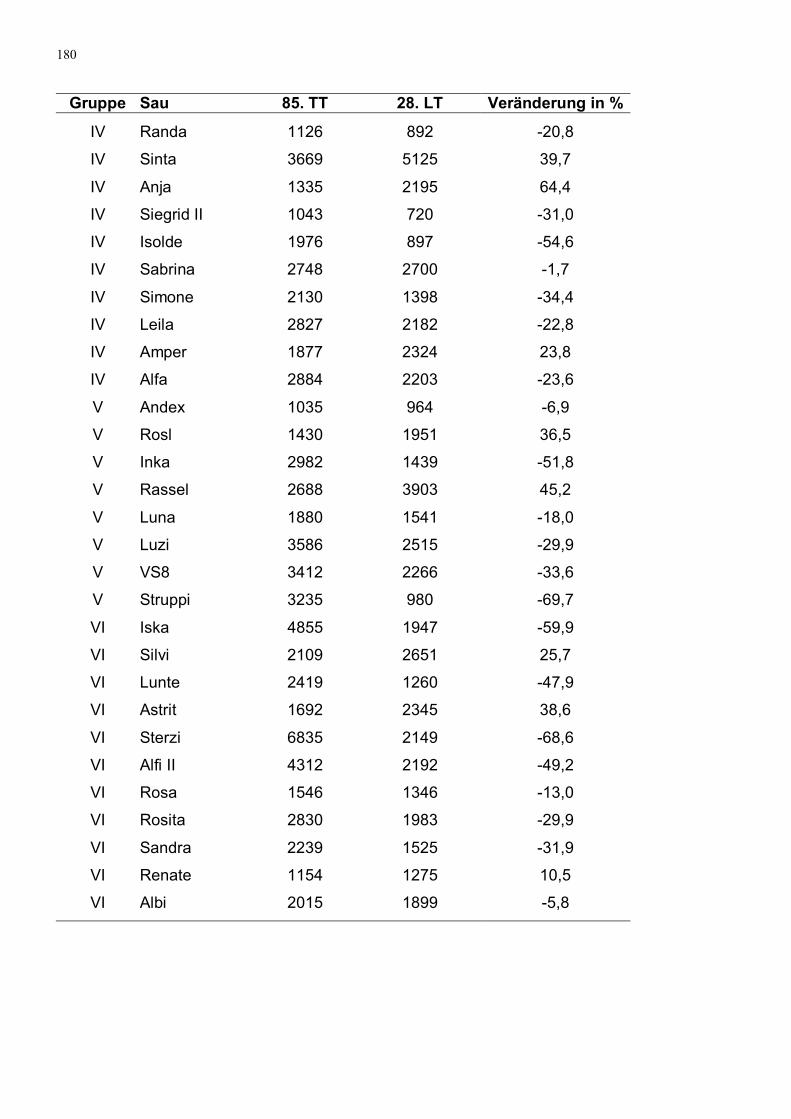
180
Gruppe Sau 85. TT 28. LT Veränderung in %
IV Randa 1126 892 -20,8
IV Sinta 3669 5125 39,7
IV Anja 1335 2195 64,4
IV Siegrid II 1043 720 -31,0
IV Isolde 1976 897 -54,6
IV Sabrina 2748 2700 -1,7
IV Simone 2130 1398 -34,4
IV Leila 2827 2182 -22,8
IV Amper 1877 2324 23,8
IV Alfa 2884 2203 -23,6
V Andex 1035 964 -6,9
V Rosl 1430 1951 36,5
V Inka 2982 1439 -51,8
V Rassel 2688 3903 45,2
V Luna 1880 1541 -18,0
V Luzi 3586 2515 -29,9
V VS8 3412 2266 -33,6
V Struppi 3235 980 -69,7
VI Iska 4855 1947 -59,9
VI Silvi 2109 2651 25,7
VI Lunte 2419 1260 -47,9
VI Astrit 1692 2345 38,6
VI Sterzi 6835 2149 -68,6
VI Alfi II 4312 2192 -49,2
VI Rosa 1546 1346 -13,0
VI Rosita 2830 1983 -29,9
VI Sandra 2239 1525 -31,9
VI Renate 1154 1275 10,5
VI Albi 2015 1899 -5,8

181
Anhangstabelle 9: Hämatokritwert in Dezimalschreibweise (links) und Thrombozytenzahl in
109/ml Vollblut (rechts) jeweils am 85. Trächtigkeitstag und 28. Laktationstag
sowie die prozentuale Veränderung
Gruppe Sau 85. TT 28. LT Veränd. in % 85. TT 28. LT Veränd. in %I Silvi 0,341 0,251 -26,4 0,083 0,121 45,8 I Amper 0,420 0,345 -17,9 0,175 0,230 31,4 I Arne 0,345 0,382 10,7 0,119 0,117 -1,7 I Lupine 0,346 0,379 9,5 0,161 0,146 -9,3 I Anneli II 0,307 0,328 6,8 0,112 0,196 75,0 I Isabell 0,395 0,385 -2,5 0,100 0,158 58,0 I Alfreda 0,414 0,358 -13,5 0,150 0,233 55,3 I Asti 0,436 0,360 -17,4 0,156 0,251 60,9 I Raupe 0,405 0,347 -14,3 0,093 0,166 78,5 II Ronja 0,438 0,269 -38,6 0,076 0,261 243,4 II Saba 0,347 0,323 -6,9 0,113 0,243 115,0 II Alfa 0,382 0,366 -4,2 0,157 0,145 -7,6 II Saale 0,405 0,354 -12,6 0,144 0,202 40,3 II Rubi II 0,400 0,347 -13,3 0,096 0,318 231,3 II Alpe 0,287 0,318 10,8 0,072 0,191 165,3 II Angela 0,355 0,369 3,9 0,136 0,174 27,9 II Ida 0,395 0,315 -20,3 0,138 0,186 34,8 II Alida 0,366 0,379 3,6 0,053 0,133 150,9 II Saale 0,279 0,355 27,2 0,101 0,143 41,6 III Alpha 0,351 0,355 1,1 0,147 0,204 38,8 III Luzi 0,431 0,362 -16,0 0,119 0,219 84,0 III Anne 0,389 0,363 -6,7 0,142 0,181 27,5 III Albi 0,332 0,348 4,8 0,152 0,294 93,4 III Ilse II 0,377 0,333 -11,7 0,060 0,159 165,0 III Almi 0,385 0,368 -4,4 0,085 0,184 116,5 III Anka 0,374 0,362 -3,2 0,121 0,140 15,7 III Saba 0,343 0,279 -18,7 0,082 0,101 23,2 III Sina 0,301 0,298 -1,0 0,090 0,101 12,2 III Reni 0,390 0,337 -13,6 0,163 0,211 29,4 III Anne 0,351 0,358 2,0 0,101 0,177 75,2 III Rapunzel 0,441 0,342 -22,4 0,159 0,262 64,8

182
Gruppe Sau 85. TT 28. LT Veränd. in % 85. TT 28. LT Veränd. in %
IV
Randa
0,402
0,330
-17,9
0,162
0,190
17,3
IV Sinta 0,323 0,316 -2,2 0,143 0,067 -53,1
IV Anja 0,268 0,335 25,0 0,176 0,176 0,0
IV Siegrid II 0,298 0,361 21,1 0,179 0,245 36,9
IV Isolde 0,357 0,355 -0,6 0,178 0,379 112,9
IV Sabrina 0,391 0,297 -24,0 0,166 0,163 -1,8
IV Simone 0,341 0,317 -7,0 0,189 0,274 45,0
IV Leila 0,297 0,293 -1,3 0,094 0,233 147,9
IV Amper 0,341 0,322 -5,6 0,165 0,215 30,3
IV Alfa 0,341 0,353 3,5 0,149 0,195 30,9
V Andex 0,433 0,325 -24,9 0,161 0,121 11,8
V Rosl 0,412 0,335 -18,7 0,081 0,230 125,9
V Inka 0,307 0,311 1,3 0,086 0,117 111,6
V Rassel 0,391 0,331 -15,3 0,145 0,146 3,4
V Luna 0,295 0,290 -1,7 0,177 0,196 19,2
V Luzi 0,385 0,304 -21,0 0,095 0,158 85,3
V VS8 0,372 0,346 -7,0 0,083 0,233 104,8
V Struppi 0,367 0,315 -14,2 0,063 0,251 223,8
VI Iska 0,342 0,346 1,2 0,074 0,166 73,0
VI Silvi 0,374 0,268 -28,3 0,149 0,261 27,5
VI Lunte 0,333 0,310 -6,9 0,115 0,243 90,4
VI Astrit 0,348 0,329 -5,5 0,148 0,145 -41,2
VI Sterzi 0,408 0,342 -16,2 0,044 0,202 302,3
VI Alfi II 0,355 0,330 -7,0 0,070 0,318 241,4
VI Rosa 0,423 0,336 -20,6 0,212 0,191 42,0
VI Rosita 0,352 0,335 -4,8 0,109 0,174 109,2
VI Sandra 0,358 0,308 -14,0 0,170 0,186 67,1
VI Renate 0,398 0,336 -15,6 0,180 0,133 33,3
VI Albi 0,410 0,339 -17,3 0,166 0,143 47,0

183
Anhangstabelle 10: Serotoningehalt im Serum in ng/ml (links) und Serotoningehalt im Vollblut
in ng/ml (rechts) jeweils am 85. Trächtigkeitstag und 28. Laktationstag
sowie die prozentuale Veränderung
Gruppe Sau 85. TT 28. LT Veränd. in % 85. TT 28. LT Veränd. in %I Silvi 878 346 -60,6 579 259 -55,2 I Amper 735 394 -46,4 426 258 -39,5 I Arne 500 230 -54,0 328 142 -56,6 I Lupine 589 340 -42,3 385 211 -45,2 I Anneli II 632 258 -59,2 438 173 -60,4 I Isabell 503 161 -68,0 304 99 -67,5 I Alfreda 380 278 -26,8 223 178 -19,9 I Asti 511 464 -9,2 288 297 3,0 I Raupe 239 263 10,0 142 172 20,8 II Ronja 175 193 10,3 98 141 43,4 II Saba 832 328 -60,6 543 222 -59,1 II Alfa 589 278 -52,8 364 176 -51,6 II Saale 298 190 -36,2 177 123 -30,8 II Rubi II 211 195 -7,6 127 127 0,6 II Alpe 441 265 -39,9 314 181 -42,5 II Angela 480 241 -49,8 310 152 -50,9 II Ida 343 190 -44,6 208 130 -37,3 II Alida 251 165 -34,3 159 102 -35,6 II Saale 350 240 -31,4 252 155 -38,7 III Alpha 210 206 -1,9 136 133 -2,5 III Luzi 532 393 -26,1 303 251 -17,2 III Anne 536 252 -53,0 327 161 -51,0 III Albi 399 488 22,3 267 318 19,4 III Ilse II 386 445 15,3 240 297 23,4 III Almi 375 464 23,7 231 293 27,2 III Anka 382 337 -11,8 239 215 -10,1 III Saba 556 673 21,0 365 485 32,8 III Sina 867 513 -40,8 606 360 -40,6 III Reni 419 495 18,1 256 328 28,4 III Anne 542 511 -5,7 352 328 -6,7 III Rapunzel 280 379 35,4 157 249 59,3
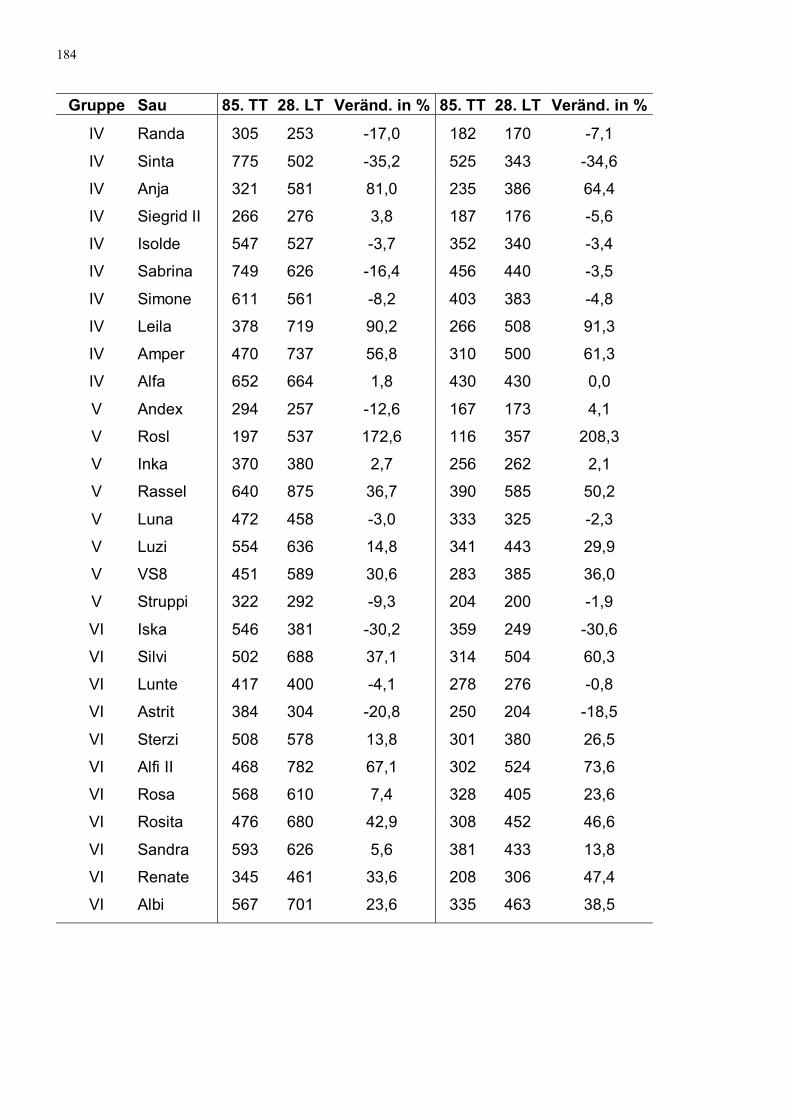
184
Gruppe Sau 85. TT 28. LT Veränd. in % 85. TT 28. LT Veränd. in %
IV
Randa
305
253
-17,0
182
170
-7,1
IV Sinta 775 502 -35,2 525 343 -34,6
IV Anja 321 581 81,0 235 386 64,4
IV Siegrid II 266 276 3,8 187 176 -5,6
IV Isolde 547 527 -3,7 352 340 -3,4
IV Sabrina 749 626 -16,4 456 440 -3,5
IV Simone 611 561 -8,2 403 383 -4,8
IV Leila 378 719 90,2 266 508 91,3
IV Amper 470 737 56,8 310 500 61,3
IV Alfa 652 664 1,8 430 430 0,0
V Andex 294 257 -12,6 167 173 4,1
V Rosl 197 537 172,6 116 357 208,3
V Inka 370 380 2,7 256 262 2,1
V Rassel 640 875 36,7 390 585 50,2
V Luna 472 458 -3,0 333 325 -2,3
V Luzi 554 636 14,8 341 443 29,9
V VS8 451 589 30,6 283 385 36,0
V Struppi 322 292 -9,3 204 200 -1,9
VI Iska 546 381 -30,2 359 249 -30,6
VI Silvi 502 688 37,1 314 504 60,3
VI Lunte 417 400 -4,1 278 276 -0,8
VI Astrit 384 304 -20,8 250 204 -18,5
VI Sterzi 508 578 13,8 301 380 26,5
VI Alfi II 468 782 67,1 302 524 73,6
VI Rosa 568 610 7,4 328 405 23,6
VI Rosita 476 680 42,9 308 452 46,6
VI Sandra 593 626 5,6 381 433 13,8
VI Renate 345 461 33,6 208 306 47,4
VI Albi 567 701 23,6 335 463 38,5

185
Anhangstabelle 11: Fett-, Eiweiß-, Laktose- und Harnstoffgehalte der Sauenmilch
13./14. LT
20./21. LT Gruppe Sau Fett Eiweiß Laktose Fett Eiweiß Laktose Harnstoff
I Stilla 6,29 4,56 5,91 6,52 4,58 5,66 219 I Alfi 9,17 4,76 5,66 8,38 4,48 5,68 213
I Alpin 7,60 4,30 5,82 7,62 4,18 5,89 279
I Amper 9,67 4,83 5,71 7,86 4,46 5,91 220
I Arne 7,12 4,25 5,81 6,81 4,25 5,91 181
I Lupine 9,60 4,51 5,55 10,13 5,25 5,31 182 I Anneli II 6,77 4,31 6,01 5,60 4,50 6,08 174
I Isabell 9,51 4,73 5,66 8,37 4,97 5,78 246
I Silvi II 6,49 4,36 6,07 6,86 4,49 5,92 185
I Alfreda 8,21 4,72 5,74 9,74 5,19 5,61 223
I Raupe 10,77 5,60 5,39 9,69 5,37 5,56 196
I Asti 8,39 4,63 5,59 7,62 4,48 5,79 218
II Irka 8,99 4,26 5,51 7,63 4,40 5,49 182
II Siegrid 8,04 4,49 5,75 7,73 4,68 5,87 142 II Ronja 9,34 4,75 5,73 6,81 4,99 5,87 149
II Saba 8,34 4,90 5,75 9,44 4,86 5,68 209
II Alfa 10,80 5,11 5,39 7,68 4,88 5,79 451
II Saale 8,44 4,35 5,82 5,91 4,60 6,05 154
II Rubi II 7,48 4,83 5,76 7,17 4,94 5,85 192
II Alpe 7,19 4,36 5,82 7,94 4,48 5,70 197
II Angela 7,46 4,17 5,99 6,32 4,30 5,89 149 II Ida II 8,80 5,09 5,52 8,11 5,18 5,56 235
II Alida 7,88 4,33 5,90 9,31 4,56 5,56 192
II Samanta 7,46 4,71 5,91 6,86 4,78 5,94 158
III Alpha 7,16 5,04 5,77 6,75 4,96 5,83 223
III Luzi 6,74 4,61 5,66 6,00 4,90 5,78 235
III Anne 7,64 4,37 5,79 7,49 4,43 5,88 197
III Albi 8,07 4,12 5,16 7,95 5,00 5,62 212
III Ilse II 8,63 4,27 5,56 10,61 4,87 5,48 160 III Almi 7,60 4,94 5,30 7,37 4,59 5,72 277
III Anka 7,41 4,80 5,82 6,07 4,85 5,93 166
III Sasse 6,84 4,95 5,82 7,59 4,73 5,85 139
III Sina 6,36 4,79 5,91 5,84 4,90 5,84 214
III Reni 7,64 4,13 5,57 9,96 4,80 5,38 200
III Anna 10,08 5,45 5,24 9,94 5,19 5,29 59
III Rapunzel 11,47 4,90 5,29 9,44 4,33 5,82 143

186
13./14. LT
20./21. LT Gruppe Sau Fett Eiweiß Laktose Fett Eiweiß Laktose Harnstoff
IV Antje 9,93 4,59 5,66 8,52 4,83 5,67 187
IV Anni 8,26 4,09 5,78 7,27 4,32 5,97 221
IV Randa 7,29 4,98 5,64 6,41 4,98 5,78 170 IV Sinta 6,94 4,56 5,61 6,47 5,08 5,86 168
IV Anja 7,55 4,67 5,64 7,63 5,14 5,80 209
IV Siegrid II 9,31 4,64 5,74 6,71 4,67 5,95 168
IV Isolde 7,74 4,75 5,65 6,72 4,87 5,89 187
IV Sabrina 8,63 4,47 5,63 6,66 4,36 6,00 199
IV Simone 7,76 4,35 5,83 7,32 4,18 6,04 204
IV Amper II 8,79 4,81 5,87 8,84 4,85 5,61 180 IV Leila 7,49 4,85 5,74 6,39 4,77 6,14 130
IV Alfa II 8,06 5,02 5,68 7,15 4,64 5,83 184
V Rubi 8,24 4,85 5,57 8,20 4,95 5,68 221
V Ilse 8,11 4,28 5,69 7,30 4,33 5,85 139
V Rassel 8,24 4,53 5,89 7,38 4,69 6,09 180
V Ida 9,11 5,25 5,49 7,69 5,42 5,79 132
V Andex 7,70 4,59 5,97 6,96 4,90 5,85 154
V Rosl 8,32 4,98 5,71 8,43 4,75 5,78 151 V Inka 6,36 4,60 4,94 7,34 4,02 5,96 178
V Rasso 8,91 4,47 5,82 7,63 4,48 5,97 187
V Luna 7,24 4,68 5,49 6,20 4,57 6,02 158
V Luzi II 7,35 4,33 5,88 5,20 4,42 6,10 149
V VS 8 7,39 4,66 5,85 7,19 4,50 5,79 132
V Struppi 7,10 4,13 5,83 7,45 4,43 5,80 293
VI Anneli 7,76 4,37 5,81 6,81 4,38 5,93 212 VI Silvi 8,41 4,74 5,71 7,74 4,78 5,82 144
VI Lunte 7,52 4,57 5,71 6,76 4,48 5,85 179
VI Iska 11,37 5,13 5,60 7,29 5,84 5,49 168
VI Astrit 11,10 5,45 5,18 10,19 5,40 5,35 235
VI Sterzi 8,41 4,85 5,87 7,74 4,72 6,00 174
VI Alfi II 8,54 4,90 5,59 7,51 4,58 5,99 163
VI Rosita 8,50 4,50 5,77 7,59 4,25 5,92 182
VI Rosa 7,29 4,69 5,62 7,15 4,72 5,70 164 VI Sandra 7,10 4,37 5,95 5,66 4,45 6,15 188
VI Renate 9,18 4,12 5,88 6,75 4,22 6,14 147
VI Albi II 9,24 5,06 5,78 8,96 5,48 5,57 293

187
Anhangstabelle 12: Lebendmasse der Ferkel in g
Gruppe Sau Ferkel 1.LT 13.LT 14. LT 20.LT 21. LT 28. LT
I Alfa 1 1298 2520 2685 3603 3795 4975
2 1399 3571 3842 4963 5223 7647
3 1524 3748 3965 4823 4988 6917
4 1907 3847 4061 4801 4969 6037
5 1657 3683 3761 4544 4703 6588 6 1911 4628 4915 6237 6485 8807
7 1519 2389 2508 3142 3200 4543
8 1720 3465 3340 3442 3430 3200
9 1620 2380 2375 2945 3137 4540
10 1680 2632 2820 3143 3235 4343
I Stilla 1 1859 3295 3492 4225 4473 6288
2 1665 2240 2379 2773 2952 3435
3 1795 3036 3309 4771 4692 5398 4 1716 3244 3385 4451 4709 6167
5 1399 2823 2912 3855 4001 5212
6 1545 2793 2878 3860 4008 4712
7 1237 2803 3070 4358 4340 4921
8 1863 3445 3665 4586 4568 5636
9 1495 3241 3366 4576 4567 5896
10 1830 2513 2500 3020 3033 3621
I Amper 1 1390 2810 2840 4180 4395 5883 2 875 1885 1905 2830 2908 3833
3 1155 2075 2163 2940 3106 4457
4 1740 3373 3547 4296 4503 5907
5 1280 2380 2505 3381 3536 4809
6 1400 2710 2855 3665 3783 5191
7 1403 4003 4197 5509 2991 4143
8 1240 1827 1935 2855 4683 6169 9 1360 3057 3215 4500 4130 5296
10 1695 3187 3320 4077 5798 6995
I Alpin 1 1597 4563 4807 6093 6285 7687
2 1592 4285 4415 5980 6187 7693
3 1566 4001 4150 5173 5330 6020
4 1271 2852 3007 3948 4120 5005
5 1605 4930 5190 6750 6947 8645
6 725 2926 2778 2400 2250 2700 7 1384 3350 3485 4835 5030 6805

188
Gruppe Sau Ferkel 1.LT 13.LT 14. LT 20.LT 21. LT 28. LT
8 1794 4477 4630 5490 5685 6993 9 1850 3010 3155 4300 4445 6037
10 1433 4495 4815 6120 6340 8077
I Arne 1 1295 3305 3416 4599 4832 6258
2 1820 4320 4435 5430 5798 7022
3 1270 3198 3330 4495 4682 5812
4 1225 3867 4051 5376 5586 6952 5 1290 3513 3677 4800 5020 6160
6 870 2255 2405 3615 3865 4785
7 925 2312 2445 3370 3370 4350
8 1475 2635 2680 3182 3275 4188
9 1500 4643 4876 6218 6483 7733
10 1460 2965 3068 4133 4347 5430
I Lupine 1 1668 3542 3734 5560 5661 6862
2 1421 3260 3515 5445 5583 6765 3 1690 3035 3240 4016 4065 4419
4 999 1590 1511 1937 2035 2462
5 1633 3663 3871 5506 5653 6940
6 1495 3880 4163 6170 6367 7885
7 1310 3494 3743 5723 5875 7380
8 1286 3090 3315 4891 5017 6230
9 1152 2353 2507 3876 4000 5118 10 1645 3005 3163 4317 4429 5873
I Isabell 1 1525 4402 4672 5891 6071 6545
2 1375 3998 4137 5292 5463 5868
3 1690 4550 4533 5573 5821 6227
4 1640 4999 5163 6296 6467 6876
5 1380 5001 5201 6385 6603 6661
6 1420 4921 5062 6208 6374 6622
7 1585 5227 5416 6473 6681 7136 8 1335 3264 3514 4509 4689 5073
9 1370 4958 5135 6532 6743 7223
10 1420 3024 3147 4069 4250 4444
I Silvi 1 1955 4497 4729 6037 6223 7695
2 1565 3258 3357 4431 4592 5750
3 1420 2938 3007 3133 3343 3821
4 1215 3046 3156 3993 4165 5170
5 1705 2822 2857 3427 3509 5487 6 1620 3013 3091 3792 3819 5427
189
Gruppe Sau Ferkel 1.LT 13.LT 14. LT 20.LT 21. LT 28. LT
7 1760 4319 4593 6017 6217 7783 8 1550 2408 2496 3103 3150 4170
9 1490 3510 3616 4858 5102 6163
10 1340 2041 2172 2976 3185 4726
11 1340 3576 3654 3745 3735 3941
12 1435 2687 2755 3188 3217 4673
I Alfreda 1 1501 4136 4422 5355 5572 6686 2 1558 3998 4247 5610 5817 6852
3 1597 4091 4317 5715 5928 6195
4 1905 4398 4601 5825 6027 6535
5 1585 4002 4036 5314 5536 6537
6 1190 3246 3401 4400 4445 4862
7 1513 3556 3631 4840 5071 6360
8 1925 4616 4673 5305 5454 6785
9 1809 4681 4780 6045 6150 6996 10 1660 4099 3963 4960 4916 5655
11 1748 4487 4330 5475 5648 6715
I Asti 1 1515 4499 4831 6578 6785 8463
2 1185 3304 3508 4743 4937 6390
3 1430 4298 4586 6152 6401 8163
4 1055 3692 3930 5308 5452 6916
5 920 2745 2997 4216 4410 5780 6 1625 4304 4541 5802 5962 7451
7 1125 3472 3745 5448 5656 7419
8 930 3092 3318 4618 4802 6303
9 840 2280 2478 3692 3856 5302
10 698 1380 1515 2321 2463 3661
I Raupe 1 1820 3899 4062 5374 5583 6656
2 2110 3440 3561 4891 5206 5830
3 1920 3858 3916 4886 5182 5920 4 2255 3310 3308 3606 3697 4155
5 1765 4416 4574 4462 4371 4325
6 1260 2051 2076 2580 2694 2535
7 2050 5093 5042 5243 5292 5685
8 955 2257 2278 2456 2555 2935
9 1990 3623 3687 4600 4871 5645
10 930 2361 2472 2930 3081 3735
I Anneli 1 1720 3738 3944 4797 4876 6797 2 1710 4018 4223 5169 5257 6542

190
Gruppe Sau Ferkel 1.LT 13.LT 14. LT 20.LT 21. LT 28. LT
3 1725 3229 3404 4100 4255 6017 4 1465 2907 3050 3792 3856 5180
5 1200 3068 3240 4204 4304 6037
6 1185 2644 2771 3456 3482 4703
7 1355 2912 3087 3890 3977 5571
8 1360 2899 3017 3758 3858 5316
9 945 2240 2397 3050 3121 3550 10 1120 2953 3113 3971 4120 5753
II Irka 1 1545 3865 4175 5643 5990 7843
2 1744 4225 4520 5650 5905 7525
3 1309 2750 2940 3963 4235 5415
4 1606 3875 4025 4950 5305 7333
5 853 1990 2010 2280 2325 3180
6 1515 3655 3915 5230 5365 6905
7 1540 3756 4040 5665 6095 7900 8 1230 3025 3245 4155 4310 5175
9 850 1562 1695 2393 2505 3080
10 1789 3520 3790 5237 5560 7285
II Ronja 1 1114 2770 2890 3913 3922 4675
2 1345 3882 4088 5653 5880 7665
3 971 3042 3207 4306 4350 4965
4 1317 4166 4340 5587 5643 7125 5 1345 4233 4437 6162 6333 8330
6 1202 3981 4225 5760 5991 7640
7 1077 3435 3595 4537 4693 6192
8 1665 2905 3047 4256 4370 4950
9 1580 3853 3980 5146 5350 6210
10 1370 4097 4311 5801 6073 8093
II Siegrid 1 1340 3152 3317 4165 4518 6415
2 1380 3710 3893 5575 5850 7956 3 1357 4232 4423 6293 6538 9031
4 1615 2667 2765 4004 4183 5533
5 1130 3313 3433 4842 4987 7070
6 1590 4443 4572 6053 6333 8395
7 1363 3532 3748 5615 5987 8605
8 1285 4173 4433 6322 6618 9227
9 1236 2637 2805 3825 4015 5630
10 1603 4361 4585 6641 6986 9451 II Saba 1 1751 4612 4780 6260 6245 7805

191
Gruppe Sau Ferkel 1.LT 13.LT 14. LT 20.LT 21. LT 28. LT
2 1538 4371 4515 6230 6425 7847 3 1487 3570 3863 5095 5330 6631
4 1480 4095 4345 5560 5685 7066
5 1673 4966 5190 6583 6745 7882
6 1724 3697 3752 5199 5610 7573
7 1476 3102 3173 4725 4799 6340
8 1350 4323 4586 5655 5805 7366 9 1235 3832 4066 5357 5365 6860
10 1165 3697 3713 4653 4991 6509
11 1195 3637 3851 5040 5265 6737
II Alfa 1 2040 3846 4056 5317 5318 6773
2 1400 3366 3647 5587 5823 7135
3 1365 3045 3287 5062 5277 6840
4 1915 3853 4105 5912 6182 8082
5 1785 4312 4553 6031 6256 7409 6 1975 2675 2683 3390 3553 4971
7 1495 2790 2940 4315 4525 5909
8 1630 3947 4234 5543 5425 5520
9 1440 3260 3486 4990 5240 6893
10 1360 3720 3365 3637 3637 4520
II Saale 1 1260 4647 4962 6123 6233 7823
2 1230 3839 3932 5000 5195 6775 3 1115 3538 3717 4753 4855 6208
4 830 3070 3331 4225 4357 5934
5 1363 4843 5151 6813 7010 8813
6 1170 3782 3957 5291 5411 6915
7 1245 2948 3148 4517 4720 6402
8 1065 3680 3863 4639 4820 5799
9 1035 3061 3364 4621 4761 6116
10 750 1500 1617 2475 2581 3711 11 730 1513 1611 2247 2295 2636
II Alpin 1 1764 3612 3800 5142 5296 7090
2 1957 4194 4354 5641 5872 7695
3 1606 3306 3403 4837 4948 6755
4 1564 2045 2193 3207 3301 4400
5 1150 3302 3335 4343 4496 5855
6 1508 4330 4519 6083 6306 8085
7 1560 2803 2923 4102 4251 5180 8 1580 3488 3583 4617 4845 6510
192
Gruppe Sau Ferkel 1.LT 13.LT 14. LT 20.LT 21. LT 28. LT
9 1487 3135 3247 4452 4622 6380 10 1505 2141 1985 2398 2538 3530
II Angela 1 1977 4383 4563 5915 6098 7309
2 1850 4940 5081 6962 7221 9137
3 1813 4345 4586 5406 5417 6372
4 1850 4798 5086 6311 6490 7852
5 2036 5642 5843 7516 7752 9481 6 1950 4956 5124 6923 7170 8483
7 1798 5502 5766 6760 6848 8293
8 1860 6229 6542 7923 7992 9409
9 1670 6182 6312 8271 8495 10520
10 1430 3437 3612 4672 4930 6221
II Ida 1 1690 4199 4395 5630 5793 7935
2 1765 4302 4470 5731 5952 7662
3 1365 2836 3045 4152 4376 6182 4 1085 3500 3746 4817 5062 6790
5 1280 3110 3241 4216 4405 5713
6 1240 2804 2984 4446 4657 6123
7 1320 4062 4242 5650 5902 7740
8 1685 4036 4285 5673 5856 7654
9 1385 4083 4319 5227 5406 7116
10 970 2981 3142 3679 3884 5625 II Alida 1 1690 4672 4955 5663 5920 8056
2 1515 4172 4452 5391 5573 7676
3 1610 3962 4176 5484 5709 7572
4 1300 3114 3308 4211 4322 5987
5 1525 3857 4125 4685 4843 6212
6 1585 4239 4498 5241 5355 7190
7 1330 3866 4125 5067 5341 7092
8 1125 2791 2966 3384 3602 5398 9 1790 1598 1600 1765 1690 1962
10 1620 3781 4040 5332 5572 7198
11 1420 2893 3018 3437 3543 4175
II Rubi 1 1725 4335 4379 5143 5331 6471
2 1880 4628 4793 5736 6009 7857
3 1705 3950 4195 5596 5878 8057
4 1245 3130 2970 3828 4053 4555
5 1280 3843 4081 5027 5164 5757 6 1565 3985 4128 4998 5268 6803

193
Gruppe Sau Ferkel 1.LT 13.LT 14. LT 20.LT 21. LT 28. LT
7 1515 3935 4165 5202 5416 7161 8 1175 2427 2612 3468 3687 4942
9 1025 3266 3502 4509 4622 5263
10 1475 4598 4756 4982 5123 7228
II Samanta 1 1400 2494 2696 3464 3555 5067
2 785 1656 1825 2962 3013 3565
3 1445 2908 3164 3779 4001 5593 4 1497 3182 3433 3769 3864 5476
5 1115 2513 2710 3250 3317 4632
6 1030 1689 1856 2626 2768 3781
7 945 2351 2590 3923 4136 5310
8 897 2392 2646 3212 3272 3480
9 1384 2260 2453 3082 3127 4405
10 1759 3601 4241 5904 6070 7352
III Ilse 1 1527 3179 3312 4732 4840 6276 2 987 1820 1953 2945 3027 3982
3 893 1743 1945 2876 2900 3099
4 1270 2914 3019 4093 4066 5082
5 1055 2682 2825 4201 4226 5343
6 1125 2835 3021 4631 4756 6196
7 970 2260 2457 3306 3461 5145
8 925 1740 1800 2927 2895 3215 9 1860 3652 3942 5199 5327 6746
10 1590 2692 2905 4434 4613 6211
III Alpha 1 1785 3722 3848 5061 5232 6007
2 1605 4183 4353 5674 5872 7265
3 2008 4923 5119 6543 6765 8367
4 1505 3777 3967 5209 5443 6485
5 1455 3460 3640 4912 5080 6025
6 1907 4443 4623 6225 6517 8105 7 1735 4558 4765 6362 6655 8543
8 1598 4441 4680 6271 6532 7810
9 1885 4605 4797 6352 6580 7651
10 1202 2565 2705 3853 4012 5014
11 1847 3813 3985 5468 5754 7043
III Almi 1 1740 2550 2566 3404 3534 4225
2 1925 3562 3951 5298 5615 7265
3 1655 2625 2875 4196 4422 6369 4 1475 2891 3159 4545 4783 5506

194
Gruppe Sau Ferkel 1.LT 13.LT 14. LT 20.LT 21. LT 28. LT
5 1580 2826 3017 4452 4656 5375 6 1545 2471 2430 2895 2962 4080
7 1680 2983 2966 4102 3965 3225
8 1547 3483 3765 5100 5051 6022
9 1670 2763 3045 4675 4972 6543
10 1435 1862 1936 3378 3602 4970
III Luzi 1 1453 3532 3840 5309 5765 8192 2 1352 3839 4056 5625 5940 7905
3 1255 3592 3834 4420 5709 5765
4 1203 3267 3277 4835 5093 6838
5 959 2334 2541 3520 4183 6347
6 1196 3077 3287 4840 5081 6782
7 1433 1670 1845 2465 2645 3247
8 1248 1943 2142 3309 3585 4979
9 1375 2901 3215 4847 5132 6687 10 1273 1920 1785 2405 2455 2905
III Anne 1 1880 4477 4765 5687 5891 6643
2 1683 5002 5067 6798 6973 8553
3 1805 5392 5632 7542 7779 9363
4 1765 4183 4243 6209 6375 7157
5 1585 4893 5082 5913 7133 8417
6 1782 3515 3659 4952 5093 5920 7 1705 4255 4351 6059 6345 7177
8 1737 5047 5202 7140 7340 8570
9 1650 4900 5048 6879 7142 8522
10 1530 3517 3616 4530 4621 5455
11 1530 4523 4700 6255 6413 7260
III Albi 1 2070 3883 3979 5437 5678 8036
2 1774 3518 3754 5622 5957 8370
3 1771 3383 3597 5144 5276 7235 4 1980 3377 3565 4977 5227 7327
5 1717 3668 3875 5261 5488 7033
6 1691 3378 3614 4921 5115 6581
7 1520 3169 3357 4063 4211 4966
8 1518 3353 3612 4748 4893 6525
9 1870 4791 5075 4888 7190 9668
10 2005 3868 4136 5613 5875 8309
III Anka 1 1934 4792 5057 6141 6327 8297 2 1824 5078 5381 7007 6680 7450

195
Gruppe Sau Ferkel 1.LT 13.LT 14. LT 20.LT 21. LT 28. LT
3 1969 5227 5487 6708 6911 9355 4 1723 5063 5368 7049 7277 9622
5 1934 4802 5045 6703 6934 9487
6 1824 5492 5802 7297 7505 9562
7 1630 5016 5346 6580 6402 8663
8 1430 4556 4741 6442 6512 8385
9 1408 4177 4373 5736 5951 7211 10 1165 3631 3896 4950 5173 6972
III Sasse 1 1300 3361 3562 4786 4983 6967
2 1785 3572 3916 5030 5340 8077
3 1875 4100 4091 4774 5042 7255
4 1490 3732 3983 5097 5419 7749
5 1515 3015 3255 4206 4403 6651
6 1520 3774 3983 4792 5038 7468
7 1295 1876 1891 2630 2440 2235 8 1465 3268 3319 4053 4310 6532
9 1840 2917 2942 3577 3798 5581
10 1875 2770 2820 3263 3440 4969
III Sina 1 1790 4307 4492 5641 5736 7948
2 1145 2370 2564 4262 4317 5358
3 1920 4578 4873 6932 7128 9352
4 2130 4832 5078 6302 6441 7742 5 1970 3659 3882 5392 5645 7656
6 1265 2734 2846 4433 4686 6583
7 1735 4561 4761 5861 5930 7642
8 1645 3471 3536 5264 5432 7192
9 825 2480 2620 3520 3669 4717
10 1880 4812 5043 6407 6588 8082
11 1625 3236 3410 4162 4225 4558
III Reni 1 1420 3564 3834 4935 5142 6163 2 1880 4238 4531 6062 6346 7463
3 1205 2341 2463 2946 3100 4066
4 1150 3233 3423 4736 4897 6071
5 1150 3264 3534 4364 4573 5940
6 1208 3625 3604 5112 5358 6455
7 784 1969 2045 2779 2780 3940
8 1480 3700 3796 5337 5726 6962
9 1510 4957 5251 6173 6351 7793 10 1275 3330 3425 4140 4095 4996

196
Gruppe Sau Ferkel 1.LT 13.LT 14. LT 20.LT 21. LT 28. LT
III Anna 1 1860 3761 3920 4962 5102 5972 2 1280 2978 3032 3244 3412 4370
3 1700 3662 3661 4538 4602 5271
4 1625 3611 3673 4351 4451 5054
5 1645 3030 3191 3631 3762 4882
6 1620 3572 3747 4863 5050 6490
7 1970 3952 4103 5223 5341 6772 8 1770 3322 3536 4502 4662 6035
9 1935 3384 3457 4167 4235 5173
10 1300 3814 3934 5077 5184 6456
11 1030 2375 2374 2746 2830 3381
III Rapunzel 1 1076 3883 4092 5682 5824 7753
2 1395 3801 3922 5204 5302 6875
3 2170 3926 4093 4928 5062 6573
4 2100 4682 4891 6729 6984 9372 5 1405 2941 3074 3821 3866 5498
6 1760 4036 4230 5300 5431 7285
7 1915 4105 4291 5092 5287 6751
8 2020 4721 5018 5842 5973 7038
9 1995 4887 5102 6783 6916 8917
10 2015 4355 4555 5528 5605 6972
IV Anni 1 1495 3332 3467 4509 4640 5885 2 1600 3440 3595 4483 4523 6085
3 1575 2262 2160 3360 3610 4293
4 1657 4425 4691 6235 6383 8063
5 1470 2966 3165 4250 4310 5887
6 1225 3758 3990 5325 5420 6727
7 1363 3080 3220 4028 4209 4630
8 1375 2355 2433 2805 3053 3947
9 1197 2633 2883 4730 4925 6805 10 1415 3547 3687 4630 4760 6492
IV Randa 1 850 2960 3145 4079 4182 5515
2 1033 3225 3455 4725 4944 6647
3 1190 2663 2821 3589 3800 4930
4 923 2405 2590 3505 3745 5185
5 1557 4182 4402 5275 5423 7253
6 1395 4101 4345 6456 6707 8942
7 1353 4125 4430 5132 5332 6407 8 1193 3860 4114 5308 5467 6707

197
Gruppe Sau Ferkel 1.LT 13.LT 14. LT 20.LT 21. LT 28. LT
9 1057 3725 3925 5325 5492 6885 10 1423 3836 3978 5120 5321 7230
IV Antje 1 1261 2970 3142 5300 5620 6760
2 1261 2895 3180 5033 5235 6532
3 1004 2470 2685 4145 4312 5521
4 1290 2995 3195 5347 5433 6145
5 1671 3600 3807 5598 5860 7175 6 1690 4145 4421 6927 6925 8423
7 1385 3273 3535 5802 5900 7087
8 1325 3047 3223 5198 5440 7006
9 1440 3374 3653 6093 6435 7460
10 1200 3140 3356 5478 5585 7130
11 1090 2515 2732 4444 4605 5908
IV Sinta 1 1613 3392 3640 4997 5205 7253
2 1495 3000 3187 4870 5135 7323 3 1760 3960 4170 6169 6437 8791
4 1850 3885 3855 4930 5253 6890
5 1587 4477 4812 6215 6467 8387
6 1270 2854 3049 3307 3410 4137
7 1645 3657 3775 5432 5773 7767
8 1450 2997 3245 4410 4709 6435
9 1620 3884 4100 5932 6267 8339 10 1280 2840 3035 4852 5203 7492
IV Anja 1 1755 4387 4537 5846 6185 7801
2 1710 3827 4055 5120 5359 7165
3 1585 3072 3285 3895 3915 4670
4 1895 4568 4799 6229 6497 8108
5 1795 4563 4810 6197 6367 7635
6 1825 4561 4761 6210 6517 8609
7 1920 3284 3477 4185 4335 6150 8 1840 4266 4500 5623 5831 7423
9 1500 3937 4102 5373 5672 7987
10 1410 2950 3030 3700 3871 5178
IV Siegrid 1 1360 3041 3226 4973 4986 6181
2 1430 3485 3703 5892 6140 8242
3 1535 3431 3640 5774 6093 7682
4 1245 3512 3782 6068 6244 8091
5 1300 2834 2891 4502 4698 6181 6 1560 3848 4093 6310 6496 8319
198
Gruppe Sau Ferkel 1.LT 13.LT 14. LT 20.LT 21. LT 28. LT
7 1437 3842 4056 6482 6697 8669 8 1265 3755 4009 6475 6696 8608
9 1045 2513 2711 4716 4930 6557
10 950 2280 2360 3585 3350 4292
IV Isolde 1 1640 3374 3504 4900 5054 6463
2 1401 3363 3582 5205 5335 6711
3 1421 3318 3509 5263 4935 6167 4 1533 3940 4155 6182 6274 7480
5 1428 3311 3463 5061 5236 6516
6 1402 3442 3635 5068 5110 6301
7 1475 3280 3412 4821 4937 6310
8 1233 2635 2811 4203 4249 5572
9 1259 3196 3280 4601 4782 6226
10 1530 3587 3765 5384 5500 7579
IV Sabrina 1 1505 4092 4377 6058 6263 8013 2 1495 3832 4164 5266 5351 6260
3 1455 4263 4510 6028 6263 8000
4 1425 2513 2768 4249 4467 6028
5 1310 3269 3522 4786 4946 6018
6 1105 3206 3576 5139 5350 7581
7 1155 2988 3154 4312 4504 6267
8 1080 3247 3532 4876 5083 6845 9 1340 4043 4320 5887 6112 8034
10 970 2579 2838 4291 4411 6150
IV Simone 1 1415 3043 3264 3983 4152 5103
2 1220 3397 3531 4590 4781 6533
3 1310 3418 3621 4721 4982 6750
4 1530 3986 4163 5178 5369 6785
5 1410 3157 3290 4387 4404 6042
6 1525 2302 2490 3333 3595 5087 7 1530 3300 3432 4393 4581 6365
8 1008 3687 3916 5022 4948 6081
9 870 2570 2723 3510 3679 4816
10 1335 2979 3202 4296 4492 6452
IV Amper 1 1555 3484 3607 4668 4692 6032
2 1655 4946 5213 6866 7056 8943
3 1612 3142 3388 4463 4643 6240
4 1440 4053 4153 5526 5696 7415 5 1620 4410 4558 5644 5869 7555

199
Gruppe Sau Ferkel 1.LT 13.LT 14. LT 20.LT 21. LT 28. LT
6 1095 2852 2917 3723 3792 4469 7 1700 3086 3340 4519 4657 5800
8 1465 4559 4764 5708 5925 7233
9 1480 3221 3396 4612 4846 6498
10 1375 2852 2862 4120 4329 6035
IV Alfa 1 1495 3263 3371 4742 4742 6263
2 1540 3359 3620 4636 4753 7217 3 1925 2726 2716 3761 3852 6208
4 1860 3798 4041 5187 5337 8549
5 1730 3157 3096 4900 4789 6251
6 1535 3462 3604 5011 4952 7456
7 1545 2701 2782 3531 3635 5841
8 1430 3401 3610 4793 4983 7623
9 1550 3250 3464 4023 5215 7992
10 1610 2987 3245 4302 4441 7211 IV Leila 1 1272 3192 3442 5416 5672 6972
2 1410 3635 3825 5762 5932 7167
3 1461 3331 3532 5302 5461 6357
4 1283 2923 3062 4431 4582 5484
5 1378 3017 3183 4779 4888 5824
6 1578 3186 3357 5208 5278 6248
7 1305 3698 3940 6223 6383 7396 8 1435 3142 3310 5060 5202 6652
9 1355 2947 3128 4732 4940 5837
10 1177 2973 3144 4704 4856 5973
11 1340 2961 3105 4506 4620 5686
V Ilse 1 1597 4727 4995 6137 6180 7905
2 1395 3958 4227 5467 5537 6583
3 1706 4565 4828 6613 6833 8177
4 1727 4625 4870 6677 6793 8633 5 1489 4355 4515 5550 5825 7535
6 1389 4340 4675 5328 5601 7003
7 1433 3940 4285 5405 5295 5927
8 1256 3595 3870 4683 4865 5910
9 1341 3935 4205 5583 5875 7390
10 1300 4085 4325 5512 5537 6983
11 850 2800 2875 3770 3900 4787
V Rassel 1 1655 4718 4975 6748 6942 8773 2 1750 4385 4627 6395 6733 8950

200
Gruppe Sau Ferkel 1.LT 13.LT 14. LT 20.LT 21. LT 28. LT
3 1315 3110 3390 5093 5391 7605 4 1130 3555 3793 5555 5802 7748
5 1605 4317 4535 6478 6752 9027
6 1390 4050 4271 5903 6174 8327
7 1270 3587 3872 5185 5255 6700
8 1463 3568 3800 4841 5040 6568
9 1440 4554 4927 6443 6643 8555 10 1605 4623 4850 6355 6521 8298
V Rubi 1 1725 4335 4379 5143 5331 6471
2 1880 4628 4793 5736 6009 7857
3 1705 3950 4195 5596 5878 8057
4 1245 3130 2970 3828 4053 4555
5 1280 3843 4081 5027 5164 5757
6 1565 3985 4128 4998 5268 6803
7 1515 3935 4165 5202 5416 7161 8 1175 2427 2612 3468 3687 4942
9 1025 3266 3502 4509 4622 5263
10 1475 4598 4756 4982 5123 7228
V Ida 1 1410 3682 3852 4497 4672 6600
2 1087 3395 3660 4245 4500 5475
3 1320 3262 3472 5083 5221 6365
4 1160 3295 3530 4658 4820 5786 5 1225 3535 3751 5022 5040 6394
6 1190 3072 3335 3475 3540 5311
7 907 2723 2955 4175 4165 5548
8 1295 4597 4950 6357 6563 7877
9 1300 2300 2340 3290 3580 4098
10 1425 2125 2145 2225 2230 2422
V Rosl 1 1640 1895 1790 1741 1775 2323
2 1390 3943 4201 5897 6156 8630 3 1270 3633 3827 4965 5243 7974
4 1335 2725 2902 4112 4410 6782
5 1285 3398 3557 5022 5331 7781
6 1465 3776 4026 5655 5883 8631
7 1210 2837 3023 4347 4646 7058
8 1400 3692 3922 5084 5021 5030
9 1240 3703 3775 5179 5386 8072
10 1365 3650 3898 5337 5579 8003 V Inka 1 1567 3241 3410 4488 4746 5795

201
Gruppe Sau Ferkel 1.LT 13.LT 14. LT 20.LT 21. LT 28. LT
2 1452 3693 3832 4982 5269 6147 3 1165 2778 2896 4039 4172 5234
4 1530 4349 4613 6310 6694 7952
5 1556 4472 4843 6660 6961 7523
6 1285 3462 3702 4116 4206 6011
7 1650 3713 3887 5386 5732 7165
8 1465 3900 4054 5132 5455 6630 9 1040 2319 2493 3826 4076 5173
10 1825 3332 3451 4096 3931 4992
V Rasso 1 1498 3788 4075 5443 5783 7736
2 1543 4170 4368 4947 5030 6459
3 1621 4990 5217 6604 6937 9280
4 1390 4462 4735 5691 5724 7004
5 1490 3842 4051 4922 5032 6732
6 1050 3465 3742 4582 4676 6192 7 1422 3576 3625 3535 3671 4748
8 1501 4491 4820 5395 5448 6377
9 1480 4064 4370 5000 5027 6089
10 1410 4513 4808 5936 6225 7974
11 1311 4012 4320 5178 5251 6613
V Luna 1 2114 4287 4453 6049 6173 8130
2 1420 2382 2486 3194 3320 3525 3 1463 4181 4472 5837 6002 8065
4 1260 2479 2555 3562 3700 4455
5 1555 3426 3615 4916 5307 6639
6 1780 3962 4101 5393 5617 7054
7 1835 3528 3715 4958 5115 6545
8 1370 3288 3417 4624 4713 6600
9 1705 3797 3974 5446 5625 7310
10 1227 3533 3721 5401 5616 7325 V Luzi 1 1775 4405 4610 5603 5697 7462
2 1439 4300 4509 5316 5481 7115
3 1112 3455 3611 4149 4306 5635
4 1657 4145 4280 5080 5214 7060
5 1578 3441 3705 4504 4746 6315
6 1741 4738 4730 5594 5763 7483
7 1295 3611 3542 4092 4276 5520
8 1210 3192 3325 3773 3792 5042 9 1433 3387 3436 3996 4023 5740

202
Gruppe Sau Ferkel 1.LT 13.LT 14. LT 20.LT 21. LT 28. LT
10 1433 2904 3034 3752 3924 4773 11 935 3046 3247 4134 4243 5178
12 1207 3209 3321 3928 4013 4964
V VS 8 1 1830 4058 4244 5414 5619 6295
2 1750 5031 5251 6725 6878 8531
3 1635 3997 4250 5962 6052 6395
4 1880 4368 4653 6334 6652 8115 5 1855 3954 4172 5317 5258 6173
6 1680 3732 3896 4863 4997 6664
7 1630 3833 4052 5254 5392 6836
8 1520 4020 4132 5169 5361 6887
9 1390 2838 2938 3722 3819 4674
10 1180 3433 3604 4523 4755 5992
V Andex 1 1525 3548 3741 4644 4834 6192
2 1670 4017 4237 5500 5767 7205 3 1820 4373 4605 5156 5351 5960
4 935 3049 3234 4268 4464 7183
5 1727 4638 4843 6231 6449 7771
6 1600 3455 3530 3749 3818 5245
7 1690 3930 4133 5318 5501 6803
8 1535 4009 4216 5374 5596 7237
9 1320 3927 4133 5412 5614 7155 10 1140 3646 4817 4935 5088 6485
V Struppi 1 1975 3699 3771 4808 4905 6331
2 2305 4882 4864 5421 5483 7041
3 2425 4427 4557 6399 6645 9050
4 2185 5023 5182 6861 6998 7872
5 2140 4248 4455 5994 6217 8645
6 2340 4435 4528 5275 5356 7420
7 2160 4026 4148 5392 5568 7905 8 1615 3864 4033 5278 5426 6723
9 1780 4265 4304 5273 5567 6545
10 1675 3010 3112 4076 4195 4490
VI Silvi 1 1715 3805 3960 4996 5227 7371
2 1732 4103 4402 5748 5949 8307
3 1503 2785 3077 4513 4785 6907
4 1687 3233 3438 4547 4795 6550
5 1783 3815 4105 5651 5978 8297 6 1525 2767 2910 3550 3626 4401

203
Gruppe Sau Ferkel 1.LT 13.LT 14. LT 20.LT 21. LT 28. LT
7 1535 2400 2500 3068 3095 3754 8 1486 2547 2505 2720 2810 3970
9 1425 2723 3055 4211 4445 6606
10 1525 4175 4460 6237 6487 8668
VI Anneli 1 1235 2705 2865 3694 3875 5047
2 1535 3140 3275 4533 4745 5993
3 1315 3279 3440 4662 4850 6004 4 1500 3807 4015 5456 5583 6943
5 1512 3705 3845 5795 5865 7307
6 1441 3153 3350 4612 4828 6230
7 1375 2847 2975 4173 4355 5717
8 1229 2153 2220 2982 3045 3822
9 1179 2863 2988 4414 4538 5790
10 870 2458 2600 3642 3745 4857
11 923 2345 2430 3551 3753 4763 12 623 1941 2073 3207 3273 4187
VI Lunte 1 1504 2453 2595 3221 3290 4591
2 1648 2677 2897 4097 4317 5325
3 1466 3207 3542 5337 5583 8817
4 1266 3252 3635 5334 5598 7353
5 2060 3275 3381 4702 4980 7335
6 1812 3423 3767 5455 5744 8060 7 1596 3512 3785 5533 5891 7932
8 1935 3842 4163 5413 5646 7730
9 1681 3851 4105 5612 5917 7541
10 1683 4202 4568 6258 6515 8464
VI Iska 1 1575 4463 4675 5198 5381 6963
2 1475 4755 5097 5644 5805 7035
3 1275 3165 3412 3895 3960 4248
4 1590 4103 4303 5030 5153 6108 5 910 1975 1917 2155 2218 2845
6 1290 3402 3572 4097 4217 5015
7 1090 2703 2940 3183 3303 3585
8 1650 4001 4160 4490 4555 4951
9 1545 4203 4535 5003 5086 6350
10 1025 2785 2991 3699 3812 4417
11 800 1125 1140 1383 1485 2075
VI Alfi 1 1910 4529 4724 6326 6641 8107 2 1793 3190 3431 5137 5240 6855

204
Gruppe Sau Ferkel 1.LT 13.LT 14. LT 20.LT 21. LT 28. LT
3 2021 4659 4884 6527 6883 8603 4 1771 3790 3986 5211 5522 6303
5 1693 2922 2972 3783 4015 4885
6 1621 4203 4403 5900 6162 7614
7 1823 3508 3527 4714 4811 5805
8 1410 3653 3932 5616 5875 7575
9 1154 2748 2975 4526 4757 6069 10 1567 2158 2305 3625 3904 5256
VI Sterzi 1 1765 2407 2446 3210 3347 4612
2 1725 1617 2575 3167 3303 4585
3 1960 4382 4881 6043 6304 8479
4 1850 4241 4462 5949 6140 8317
5 1740 1878 1921 2318 2430 3306
6 1630 4102 4313 5473 5682 7360
7 1305 2143 2076 2343 2454 3405 8 1585 4516 4788 6090 6305 8356
9 1255 3130 3203 4378 4551 6213
10 1865 3840 3967 4500 4650 6710
VI Astrit 1 1705 4853 5150 6901 7083 8913
2 1925 4501 4800 6403 6691 8822
3 1645 3790 3992 5252 5430 6967
4 1465 4133 4367 5885 6121 7877 5 1415 3985 4245 5679 5835 7283
6 1645 4185 4456 5682 5917 7068
7 1600 3730 3920 5254 5425 6925
8 1607 3975 4213 5399 5570 7053
9 1560 3499 3662 4733 4830 6191
10 943 1530 1637 1958 2030 2830
VI Rosa 1 1710 4009 4035 4378 4496 5506
2 1515 3710 3847 4768 5036 6649 3 1525 3935 4163 5461 5755 7638
4 1453 4173 4435 5855 5920 8045
5 1542 4122 4304 5304 5555 7805
6 1434 3737 3941 4684 4927 6292
7 1490 3686 3838 5250 5702 7953
8 1220 3354 3571 4710 4967 6430
9 1528 4776 5047 6482 6875 7652
10 1650 4638 4861 6458 6760 8687 11 1434 3581 3731 4768 5049 6548

205
Gruppe Sau Ferkel 1.LT 13.LT 14. LT 20.LT 21. LT 28. LT
VI Rosita 1 1713 4399 4626 5817 6102 8134 2 1389 3837 4088 5291 5573 7181
3 1436 4461 4628 5750 6026 7106
4 1102 3575 3758 4912 5107 6441
5 1050 3233 3496 4316 4418 5615
6 1790 4494 4750 5754 5970 7576
7 1758 4697 4920 5860 5924 7239 8 1404 4222 4382 5501 5870 6903
9 1728 4875 5183 6463 6745 7890
10 1328 4096 4405 5416 5372 6840
11 1070 3736 3976 4984 5151 6592
12 895 3073 3221 4260 4336 5400
VI Sandra 1 1750 4887 5087 6678 6892 8380
2 1725 4909 5291 6832 7134 8935
3 1455 4587 4800 5747 5990 7530 4 1540 4591 4770 5441 5613 6975
5 1450 4335 4542 5936 6127 7255
6 1370 3631 3626 5059 5133 6635
7 1020 3402 3560 4816 4956 6174
8 1370 4308 4496 5945 6081 7420
9 1713 5637 5899 7198 7374 9170
10 1095 3602 3745 4783 4917 6165 VI Renate 1 1800 4422 4611 6004 6199 7946
2 910 2215 2355 3312 3406 4716
3 1150 2884 3060 4306 4412 5763
4 1025 2617 2761 3832 4008 5297
5 1135 3172 3398 4911 4984 6825
6 1370 3552 3704 4863 5032 6661
7 1535 3684 3834 5130 5281 6785
8 1865 3860 4047 5669 5860 7774 9 1550 3862 4106 5412 5679 7640
10 1105 3103 3282 4370 4576 6075
VI Albi 1 1830 4551 4692 5203 5367 6576
2 2050 3564 3697 4281 4286 5099
3 1890 3154 3296 3948 3942 4165
4 1870 2633 2716 3237 3418 4392
5 1165 2187 2399 2741 2779 3243
6 1710 3596 3622 5253 5562 7328 7 1790 3967 4071 5273 5434 6977

206
Gruppe Sau Ferkel 1.LT 13.LT 14. LT 20.LT 21. LT 28. LT
8 1850 3453 3633 4150 4311 5132 9 1385 3201 2970 2864 2881 2765
10 1580 3715 3911 4500 4612 3985

207
Anhangstabelle 13: Konzentrationen der essentiellen Aminosäuren im Sauenplasma in mg/l
Gr Sau Lys Met Cys Thr Trp Leu Ile Val Phe Tyr His
I Stilla 47,75 15,59 6,18 36,65 3,19 23,68 16,48 26,50 18,35 16,99 17,87
Amper 55,62 17,51 2,98 31,78 1,73 30,33 16,99 29,77 20,76 17,99 16,06
Arne 22,81 12,30 1,31 43,06 3,26 16,49 10,34 17,65 14,52 16,10 15,26
Lupine 25,04 8,91 0,97 32,17 3,14 16,78 9,14 17,13 12,52 16,39 16,87
Anneli 60,86 18,16 8,11 41,81 4,13 27,75 15,45 25,37 21,44 26,40 19,86
Isabell 30,46 14,57 0,75 38,18 2,91 16,74 13,19 16,16 18,27 14,81 19,01
Silvi 40,87 16,49 1,88 37,94 0,53 19,60 14,59 21,05 19,86 20,40 18,59
Alfreda 25,47 10,85 1,89 29,60 4,14 15,64 10,74 15,48 11,25 10,04 21,27
Asti 75,57 11,14 2,21 55,56 1,64 28,11 20,33 27,02 28,02 25,04 14,55
Raupe 45,35 8,21 5,11 32,69 2,74 28,79 21,71 37,85 13,76 14,24 15,26
II Irka 49,41 19,45 1,48 32,15 4,86 19,88 12,32 20,78 19,24 14,04 18,30
Ronja 30,45 20,74 0,52 47,60 4,28 22,06 13,94 18,57 21,10 19,09 17,58
Saba 73,98 30,14 0,27 50,31 11,47 32,46 23,79 31,34 32,08 19,79 20,36
Alfa 61,72 27,85 0,98 45,92 6,85 33,40 19,44 32,45 30,05 23,89 16,88
Saale 23,36 8,69 2,48 17,72 4,51 20,28 11,87 22,47 12,61 15,78 17,22
Rubi 35,34 17,25 4,58 29,28 1,44 17,82 9,67 15,08 15,94 14,94 15,52
Alpin 63,51 28,89 1,22 59,93 6,06 35,15 22,56 32,86 35,21 26,47 22,25
Angela 66,95 28,18 1,15 55,90 4,72 25,65 17,12 22,19 29,89 22,21 22,58
Ida 38,82 16,94 2,43 36,80 5,51 13,58 9,28 9,92 17,35 15,35 16,78
Alida 40,23 16,46 0,83 36,15 3,73 18,19 11,92 14,68 0,49 15,98 17,80
Saale 65,49 24,76 1,78 41,16 7,20 33,95 20,11 35,00 31,64 27,90 23,04
III Luzi 70,05 28,11 1,06 42,71 13,38 32,71 19,86 31,71 29,16 24,32 20,66
Anne 43,05 20,29 0,40 33,65 10,21 26,46 15,58 20,39 23,39 17,89 19,55
Albi 53,18 22,82 1,91 33,89 10,60 37,63 17,89 34,15 25,87 25,59 16,83
Ilse 37,66 18,41 2,21 29,11 5,95 27,21 12,82 23,48 21,93 22,40 15,64
Almi 33,64 21,39 1,43 23,64 8,12 25,63 15,38 23,10 21,70 15,68 19,81
Anka 46,69 30,92 1,59 50,68 5,31 25,47 15,07 15,75 30,31 14,84 18,86
Saba 45,74 27,16 0,50 38,52 5,84 25,38 13,67 17,09 31,35 19,36 20,55
Sina 40,71 23,78 1,78 34,05 4,18 31,25 16,67 18,64 23,58 22,28 16,61
Reni 29,24 15,47 1,96 26,59 9,19 20,15 9,69 12,31 16,14 18,53 19,59
Anne 16,92 23,43 6,27 34,98 8,67 16,23 9,63 14,74 24,92 20,16 16,27
Rapunzel 42,10 26,04 1,97 37,23 13,07 20,84 13,50 20,19 26,04 18,98 17,22
Alpha 56,22 23,36 1,79 34,68 9,56 27,50 15,89 22,65 24,62 21,85 15,45

208
Gr Sau Lys Met Cys Thr Trp Leu Ile Val Phe Tyr His
IV Anni 54,01 22,89 1,86 33,75 15,84 26,35 14,58 27,46 28,80 30,42 18,31
Randa 54,80 30,74 1,32 45,82 21,09 38,62 22,32 37,01 26,26 24,56 21,49
Sina 53,47 25,52 1,77 35,20 15,02 34,32 17,95 25,56 25,39 24,26 15,63
Anja 69,30 22,65 2,46 36,48 19,32 35,09 19,44 31,43 27,07 25,56 20,17
Sigrid 36,42 18,00 2,36 29,45 9,42 27,02 15,21 22,78 19,25 14,30 11,77
Isolde 58,77 29,77 0,78 42,91 16,53 29,07 15,30 22,31 29,32 25,67 20,32
Simone 50,86 33,36 2,02 44,31 16,39 34,76 18,12 28,69 23,90 25,13 22,59
Leila 24,31 14,31 2,51 27,20 11,52 18,77 11,24 13,43 15,56 13,47 15,12
Amper 42,47 19,06 1,67 29,11 14,01 25,10 13,01 18,45 18,83 17,50 16,68
Alfa 53,92 25,03 1,86 32,43 16,25 21,92 11,97 14,79 24,63 22,06 15,85
V Rassel 47,66 25,88 0,56 36,45 21,36 22,32 11,65 14,87 22,82 17,74 15,08
Ida 38,58 14,92 1,89 23,43 9,52 21,53 11,78 15,69 15,23 14,64 17,26
Andechs 33,05 21,48 1,34 32,09 11,50 25,68 11,69 17,83 17,96 18,06 13,95
Rosl 42,20 20,03 1,37 25,12 22,18 23,14 14,07 17,57 20,15 13,83 16,24
Inka 30,19 26,95 0,70 34,50 17,15 14,56 8,42 8,45 18,83 17,39 19,68
Rassel 28,45 25,35 0,93 31,74 19,46 15,96 7,94 11,65 23,91 15,87 17,47
Luna 34,28 20,70 1,67 23,68 15,17 24,05 10,88 14,10 22,51 13,13 17,18
Luzi 39,80 25,36 1,67 30,41 19,63 18,44 8,91 12,03 23,28 16,99 20,84
VS8 37,75 16,61 1,45 23,14 8,46 18,01 6,82 8,79 20,21 17,58 17,03
Struppi 45,55 23,72 1,82 38,15 12,91 27,96 13,61 15,62 21,51 21,75 17,05
VI Silvi 39,79 15,51 0,75 31,61 22,10 23,61 13,34 19,80 17,76 19,35 19,46
Luna 52,70 25,19 0,47 29,74 30,85 20,76 14,45 19,79 24,40 13,98 17,77
Iska 24,77 9,68 1,14 18,55 16,19 18,41 6,25 11,06 13,03 17,51 13,08
Astrit 37,83 19,99 2,74 30,35 29,36 16,87 13,80 16,73 16,95 16,75 18,49
Sterzi 42,29 19,43 1,77 31,20 26,98 20,87 10,11 12,92 20,14 13,94 18,91
Alfi 52,77 25,76 1,99 32,51 36,76 25,33 14,45 19,49 28,34 18,66 15,97
Rosa 11,78 13,55 1,08 17,25 14,54 25,88 12,54 16,94 13,36 11,57 17,09
Rosita 13,57 13,29 1,04 18,67 17,03 21,04 10,18 18,07 16,31 9,89 12,44
Renate 34,07 26,56 1,20 39,00 17,35 18,83 9,13 7,75 20,12 20,42 20,10
Albi 47,84 17,72 5,54 36,08 27,95 36,22 18,59 34,87 24,40 22,94 16,12
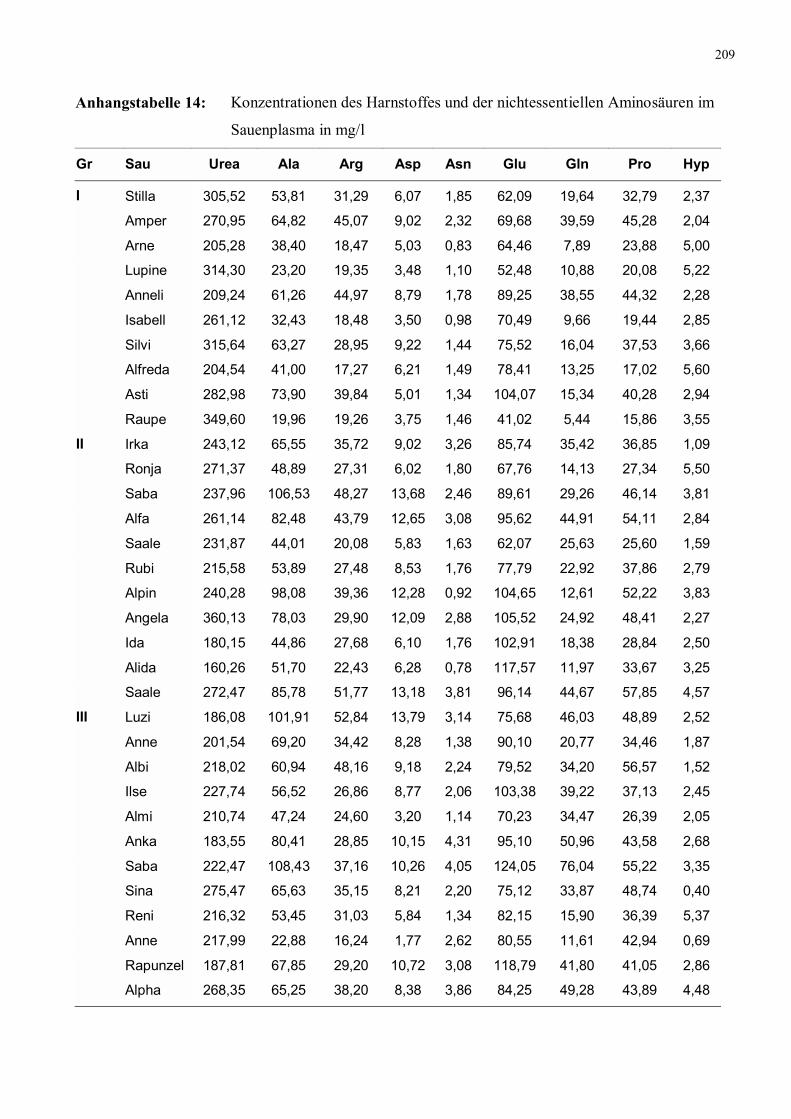
209
Anhangstabelle 14: Konzentrationen des Harnstoffes und der nichtessentiellen Aminosäuren im
Sauenplasma in mg/l
Gr Sau Urea Ala Arg Asp Asn Glu Gln Pro Hyp
I Stilla 305,52 53,81 31,29 6,07 1,85 62,09 19,64 32,79 2,37
Amper 270,95 64,82 45,07 9,02 2,32 69,68 39,59 45,28 2,04
Arne 205,28 38,40 18,47 5,03 0,83 64,46 7,89 23,88 5,00
Lupine 314,30 23,20 19,35 3,48 1,10 52,48 10,88 20,08 5,22
Anneli 209,24 61,26 44,97 8,79 1,78 89,25 38,55 44,32 2,28
Isabell 261,12 32,43 18,48 3,50 0,98 70,49 9,66 19,44 2,85
Silvi 315,64 63,27 28,95 9,22 1,44 75,52 16,04 37,53 3,66
Alfreda 204,54 41,00 17,27 6,21 1,49 78,41 13,25 17,02 5,60
Asti 282,98 73,90 39,84 5,01 1,34 104,07 15,34 40,28 2,94
Raupe 349,60 19,96 19,26 3,75 1,46 41,02 5,44 15,86 3,55
II Irka 243,12 65,55 35,72 9,02 3,26 85,74 35,42 36,85 1,09
Ronja 271,37 48,89 27,31 6,02 1,80 67,76 14,13 27,34 5,50
Saba 237,96 106,53 48,27 13,68 2,46 89,61 29,26 46,14 3,81
Alfa 261,14 82,48 43,79 12,65 3,08 95,62 44,91 54,11 2,84
Saale 231,87 44,01 20,08 5,83 1,63 62,07 25,63 25,60 1,59
Rubi 215,58 53,89 27,48 8,53 1,76 77,79 22,92 37,86 2,79
Alpin 240,28 98,08 39,36 12,28 0,92 104,65 12,61 52,22 3,83
Angela 360,13 78,03 29,90 12,09 2,88 105,52 24,92 48,41 2,27
Ida 180,15 44,86 27,68 6,10 1,76 102,91 18,38 28,84 2,50
Alida 160,26 51,70 22,43 6,28 0,78 117,57 11,97 33,67 3,25
Saale 272,47 85,78 51,77 13,18 3,81 96,14 44,67 57,85 4,57
III Luzi 186,08 101,91 52,84 13,79 3,14 75,68 46,03 48,89 2,52
Anne 201,54 69,20 34,42 8,28 1,38 90,10 20,77 34,46 1,87
Albi 218,02 60,94 48,16 9,18 2,24 79,52 34,20 56,57 1,52
Ilse 227,74 56,52 26,86 8,77 2,06 103,38 39,22 37,13 2,45
Almi 210,74 47,24 24,60 3,20 1,14 70,23 34,47 26,39 2,05
Anka 183,55 80,41 28,85 10,15 4,31 95,10 50,96 43,58 2,68
Saba 222,47 108,43 37,16 10,26 4,05 124,05 76,04 55,22 3,35
Sina 275,47 65,63 35,15 8,21 2,20 75,12 33,87 48,74 0,40
Reni 216,32 53,45 31,03 5,84 1,34 82,15 15,90 36,39 5,37
Anne 217,99 22,88 16,24 1,77 2,62 80,55 11,61 42,94 0,69
Rapunzel 187,81 67,85 29,20 10,72 3,08 118,79 41,80 41,05 2,86
Alpha 268,35 65,25 38,20 8,38 3,86 84,25 49,28 43,89 4,48

210
Gr Sau Urea Ala Arg Asp Asn Glu Gln Pro Hyp
IV Anni 252,07 40,99 42,34 7,96 0,94 75,31 18,19 43,94 2,33
Randa 244,69 75,13 37,01 10,53 2,98 97,07 44,78 61,14 2,27
Sina 207,96 89,21 42,50 11,42 2,27 78,90 35,04 58,08 3,31
Anja 287,25 70,22 51,95 9,24 2,65 69,71 45,03 46,86 1,97
Sigrid 250,78 54,62 27,64 5,64 1,45 63,46 22,63 31,17 2,75
Isolde 271,27 63,61 40,18 12,82 0,97 85,83 20,16 46,03 3,01
Simone 369,63 94,54 35,79 11,40 4,47 98,60 67,27 59,80 3,13
Leila 158,16 50,05 23,80 6,46 2,26 86,95 28,18 30,51 0,97
Amper 241,75 54,71 35,25 6,64 1,77 73,12 33,29 41,22 2,05
Alfa 147,70 83,34 32,16 8,45 4,10 97,87 66,11 48,83 1,50
V Rassel 204,99 92,90 34,13 12,40 4,11 91,50 39,90 48,25 2,74
Ida 181,22 65,89 27,82 6,01 2,52 90,24 36,33 36,75 4,09
Andechs 185,05 74,31 17,10 8,47 3,11 102,95 44,23 65,77 1,82
Rosl 240,62 64,24 36,33 9,09 4,01 100,81 53,47 28,61 2,91
Inka 213,46 86,35 18,94 10,84 3,23 147,76 52,09 36,84 2,28
Rassel 203,98 75,52 20,25 9,22 3,87 120,85 46,80 36,79 3,25
Luna 222,90 64,01 23,90 7,71 3,49 74,57 43,35 38,87 1,39
Luzi 169,84 65,88 21,12 6,03 3,88 120,02 45,75 33,87 2,95
VS8 219,78 45,24 7,79 7,02 3,20 154,30 48,00 36,12 3,20
Struppi 247,77 81,04 28,28 8,71 3,50 80,47 47,62 50,15 2,75
VI Silvi 250,19 78,29 26,80 8,84 2,14 70,30 20,46 41,68 2,98
Luna 222,56 96,33 32,10 10,38 2,60 84,82 37,50 39,41 4,39
Iska 198,90 71,17 16,71 5,98 2,47 94,41 26,98 34,94 3,69
Astrit 197,18 77,71 49,17 8,32 1,39 95,96 21,69 34,09 3,83
Sterzi 184,28 65,62 28,77 8,09 5,55 87,19 58,43 38,68 4,04
Alfi 266,47 70,68 41,12 6,15 2,30 86,62 37,01 50,56 1,01
Rosa 225,81 44,22 25,56 5,79 1,31 68,35 25,72 26,82 2,98
Rosita 304,68 52,87 19,23 5,14 0,68 52,90 19,92 32,35 1,52
Renate 172,23 63,58 16,71 7,38 5,58 80,51 58,06 51,49 2,70
Albi 312,93 58,46 47,09 7,91 1,93 84,02 26,37 45,16 0,50

211
Anhangstabelle 15: Gehalte an essentiellen Aminosäuren in der Sauenmilch in g/kg FS
Gr. Sau Lys Met Cys Thr Trp Leu Ile Val Phe Tyr His
I Stilla 0,33 0,084 0,051 0,18 0,049 0,36 0,18 0,22 0,18 0,17 0,12
Alfi 0,38 0,099 0,052 0,20 0,053 0,42 0,21 0,26 0,20 0,20 0,14
Alpin 0,29 0,079 0,049 0,16 0,044 0,35 0,16 0,21 0,16 0,15 0,11
Amper 0,33 0,087 0,058 0,18 0,054 0,37 0,19 0,22 0,18 0,17 0,12
Arne 0,33 0,084 0,057 0,18 0,053 0,39 0,18 0,23 0,18 0,16 0,12
Lupine 0,30 0,078 0,047 0,17 0,047 0,34 0,16 0,21 0,16 0,15 0,11
Anneli II 0,37 0,094 0,054 0,19 0,053 0,40 0,20 0,24 0,20 0,18 0,13
Isabell 0,38 0,101 0,055 0,20 0,054 0,42 0,21 0,25 0,20 0,20 0,14
Silvi II 0,33 0,089 0,055 0,18 0,049 0,39 0,18 0,23 0,19 0,17 0,12
Alfreda 0,31 0,082 0,054 0,17 0,050 0,37 0,17 0,22 0,18 0,15 0,11
Raupe 0,39 0,102 0,056 0,20 0,059 0,43 0,22 0,26 0,21 0,20 0,14
Asti 0,31 0,082 0,048 0,17 0,045 0,36 0,17 0,22 0,18 0,17 0,12
III Alpha 0,35 0,092 0,058 0,19 0,058 0,39 0,19 0,24 0,19 0,19 0,13
Luzi 0,34 0,083 0,060 0,18 0,056 0,37 0,18 0,23 0,18 0,17 0,12
Anne 0,38 0,095 0,059 0,19 0,059 0,40 0,20 0,25 0,20 0,19 0,14
Albi 0,37 0,092 0,056 0,20 0,057 0,40 0,20 0,24 0,20 0,19 0,13
Ilse II 0,31 0,078 0,049 0,17 0,048 0,34 0,17 0,21 0,17 0,16 0,11
Almi 0,35 0,088 0,061 0,20 0,059 0,39 0,19 0,24 0,20 0,19 0,13
Anka 0,36 0,091 0,062 0,20 0,057 0,40 0,20 0,24 0,20 0,18 0,13
Sasse 0,33 0,084 0,053 0,18 0,055 0,37 0,19 0,23 0,19 0,18 0,12
Sina 0,36 0,091 0,056 0,19 0,055 0,40 0,20 0,25 0,19 0,18 0,13
Reni 0,35 0,090 0,053 0,18 0,051 0,38 0,20 0,23 0,19 0,19 0,12
Anna 0,36 0,096 0,058 0,19 0,052 0,41 0,20 0,25 0,20 0,19 0,13
Rapunzel 0,31 0,078 0,047 0,17 0,046 0,33 0,17 0,21 0,17 0,17 0,11
VI Anneli 0,33 0,085 0,056 0,18 0,054 0,37 0,18 0,22 0,18 0,17 0,12
Silvi 0,31 0,082 0,056 0,18 0,049 0,37 0,17 0,22 0,17 0,15 0,11
Lunte 0,38 0,100 0,066 0,21 0,061 0,44 0,21 0,27 0,21 0,20 0,14
Iska 0,42 0,107 0,077 0,24 0,075 0,48 0,24 0,30 0,24 0,23 0,15
Astrit 0,32 0,084 0,055 0,18 0,051 0,36 0,18 0,22 0,18 0,17 0,12
Sterzi 0,35 0,089 0,058 0,19 0,054 0,39 0,20 0,24 0,19 0,17 0,13
Alfi II 0,32 0,079 0,047 0,17 0,047 0,34 0,18 0,21 0,17 0,16 0,11
Rosa 0,33 0,085 0,061 0,18 0,055 0,39 0,18 0,23 0,19 0,18 0,12
Rosita 0,34 0,086 0,057 0,19 0,056 0,39 0,19 0,23 0,19 0,18 0,12
Sandra 0,32 0,083 0,057 0,17 0,051 0,38 0,18 0,23 0,17 0,16 0,12
Renate 0,31 0,077 0,049 0,17 0,049 0,34 0,17 0,21 0,17 0,16 0,11
Albi II 0,40 0,099 0,067 0,22 0,064 0,45 0,22 0,28 0,22 0,22 0,14

212
Anhangstabelle 16: Gehalte an nicht essentiellen Aminosäuren in der Sauenmilch in g/kg FS
Gr. Sau Ala Arg Asp Glu Gly Ser
I Stilla 0,15 0,20 0,35 0,93 0,14 0,22
Alfi 0,16 0,23 0,39 1,10 0,16 0,25
Alpin 0,14 0,19 0,33 0,86 0,14 0,20
Amper 0,16 0,20 0,36 0,89 0,13 0,22
Arne 0,16 0,20 0,36 0,94 0,14 0,23
Lupine 0,14 0,18 0,33 0,88 0,13 0,21
Anneli II 0,16 0,22 0,38 1,04 0,15 0,24
Isabell 0,17 0,23 0,40 1,05 0,16 0,25
Silvi II 0,16 0,21 0,36 0,97 0,15 0,23
Alfreda 0,16 0,21 0,35 0,90 0,16 0,22
Raupe 0,18 0,24 0,41 1,12 0,17 0,26
Asti 0,15 0,20 0,33 0,91 0,15 0,21
III Alpha 0,17 0,22 0,39 0,93 0,14 0,23
Luzi 0,16 0,21 0,37 0,90 0,14 0,22
Anne 0,17 0,23 0,40 1,00 0,15 0,25
Albi 0,17 0,21 0,38 0,98 0,14 0,25
Ilse II 0,15 0,20 0,34 0,91 0,15 0,22
Almi 0,17 0,23 0,38 0,95 0,15 0,24
Anka 0,17 0,23 0,39 0,98 0,16 0,24
Sasse 0,16 0,22 0,37 0,96 0,15 0,23
Sina 0,16 0,22 0,38 1,02 0,16 0,24
Reni 0,16 0,21 0,36 0,97 0,15 0,23
Anna 0,16 0,23 0,39 1,09 0,18 0,25
Rapunzel 0,15 0,19 0,33 0,91 0,14 0,21
VI Anneli 0,16 0,20 0,36 0,91 0,14 0,22
Silvi 0,15 0,20 0,35 0,88 0,14 0,22
Lunte 0,18 0,25 0,41 1,09 0,18 0,26
Iska 0,21 0,28 0,48 1,11 0,19 0,29
Astrit 0,15 0,21 0,36 0,90 0,15 0,22
Sterzi 0,16 0,22 0,39 0,97 0,15 0,24
Alfi II 0,14 0,20 0,34 0,89 0,14 0,21
Rosa 0,17 0,22 0,37 0,93 0,16 0,23
Rosita 0,17 0,22 0,39 0,95 0,16 0,24
Sandra 0,16 0,21 0,36 0,91 0,14 0,22
Renate 0,15 0,18 0,33 0,86 0,13 0,21
Albi II 0,20 0,25 0,43 1,06 0,17 0,27

213
Anhangstabelle 17: Saugferkelbeifutterverbrauch in g je Wurf während der 4. Laktationswoche
Gruppe Sau Verbrauch Gruppe Sau Verbrauch I Stilla 322 IV Antje 184 Alfi 1020 Anni 50 Alpin 260 Randa 255 Amper 730 Sinta 380 Arne 1280 Anja 465 Lupine 65 Isolde 1295 Anneli II 705 Siegrid II 295 Isabell 185 Sabrina 1300 Silvi II 2535 Simone 1500 Alfreda 110 Amper II 1292 Raupe 50 Leila 1500 Asti 1175 Alfa II 2846 II Irka 365 V Rubi 269 Siegrid 540 Ilse 260 Ronja 1075 Rassel 370 Saba 111 Ida 100 Alfa 80 Andex 1500 Saale 435 Rosl 115 Rubi II 130 Inka 215 Alpe 246 Luna 70 Angela 2985 Rasso 670 Ida II 1455 Luzi II 1284 Alida 180 VS 8 1396 Samanta 895 Struppi 1240
III Alpha 865 VI Anneli 998 Luzi 115 Silvi 55 Anne 1165 Lunte 205 Albi 1480 Iska 70 Ilse II 170 Astrit 476 Almi 110 Sterzi 1180 Anka 0 Alfi II 410 Sasse 1214 Rosa 560 Sina 645 Rosita 410 Reni 905 Sandra 855 Anna 1425 Renate 1500 Rapunzel 2260 Albi II 60
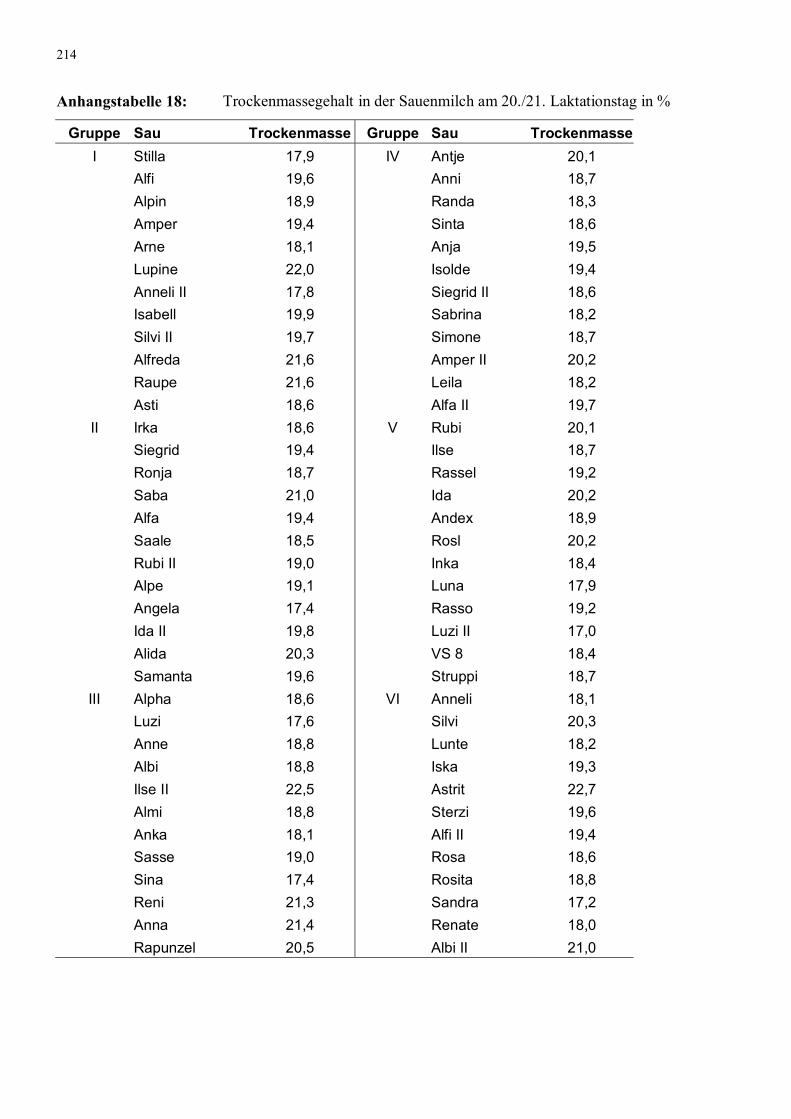
214
Anhangstabelle 18: Trockenmassegehalt in der Sauenmilch am 20./21. Laktationstag in %
Gruppe Sau Trockenmasse Gruppe Sau Trockenmasse I Stilla 17,9 IV Antje 20,1 Alfi 19,6 Anni 18,7 Alpin 18,9 Randa 18,3 Amper 19,4 Sinta 18,6 Arne 18,1 Anja 19,5 Lupine 22,0 Isolde 19,4 Anneli II 17,8 Siegrid II 18,6 Isabell 19,9 Sabrina 18,2 Silvi II 19,7 Simone 18,7 Alfreda 21,6 Amper II 20,2 Raupe 21,6 Leila 18,2 Asti 18,6 Alfa II 19,7 II Irka 18,6 V Rubi 20,1 Siegrid 19,4 Ilse 18,7 Ronja 18,7 Rassel 19,2 Saba 21,0 Ida 20,2 Alfa 19,4 Andex 18,9 Saale 18,5 Rosl 20,2 Rubi II 19,0 Inka 18,4 Alpe 19,1 Luna 17,9 Angela 17,4 Rasso 19,2 Ida II 19,8 Luzi II 17,0 Alida 20,3 VS 8 18,4 Samanta 19,6 Struppi 18,7
III Alpha 18,6 VI Anneli 18,1 Luzi 17,6 Silvi 20,3 Anne 18,8 Lunte 18,2 Albi 18,8 Iska 19,3 Ilse II 22,5 Astrit 22,7 Almi 18,8 Sterzi 19,6 Anka 18,1 Alfi II 19,4 Sasse 19,0 Rosa 18,6 Sina 17,4 Rosita 18,8 Reni 21,3 Sandra 17,2 Anna 21,4 Renate 18,0 Rapunzel 20,5 Albi II 21,0
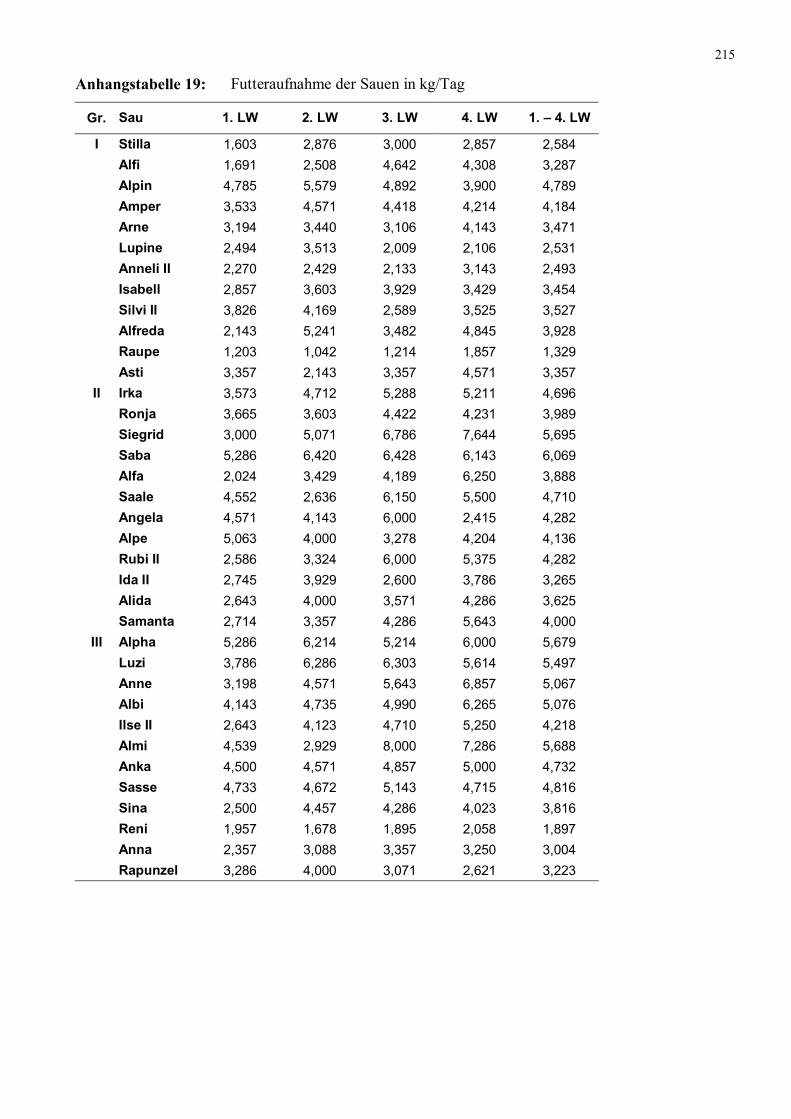
215
Anhangstabelle 19: Futteraufnahme der Sauen in kg/Tag
Gr. Sau 1. LW 2. LW 3. LW 4. LW 1. � 4. LW
I Stilla 1,603 2,876 3,000 2,857 2,584 Alfi 1,691 2,508 4,642 4,308 3,287 Alpin 4,785 5,579 4,892 3,900 4,789 Amper 3,533 4,571 4,418 4,214 4,184 Arne 3,194 3,440 3,106 4,143 3,471 Lupine 2,494 3,513 2,009 2,106 2,531 Anneli II 2,270 2,429 2,133 3,143 2,493 Isabell 2,857 3,603 3,929 3,429 3,454 Silvi II 3,826 4,169 2,589 3,525 3,527 Alfreda 2,143 5,241 3,482 4,845 3,928 Raupe 1,203 1,042 1,214 1,857 1,329 Asti 3,357 2,143 3,357 4,571 3,357
II Irka 3,573 4,712 5,288 5,211 4,696 Ronja 3,665 3,603 4,422 4,231 3,989 Siegrid 3,000 5,071 6,786 7,644 5,695 Saba 5,286 6,420 6,428 6,143 6,069 Alfa 2,024 3,429 4,189 6,250 3,888 Saale 4,552 2,636 6,150 5,500 4,710 Angela 4,571 4,143 6,000 2,415 4,282 Alpe 5,063 4,000 3,278 4,204 4,136 Rubi II 2,586 3,324 6,000 5,375 4,282 Ida II 2,745 3,929 2,600 3,786 3,265 Alida 2,643 4,000 3,571 4,286 3,625 Samanta 2,714 3,357 4,286 5,643 4,000
III Alpha 5,286 6,214 5,214 6,000 5,679 Luzi 3,786 6,286 6,303 5,614 5,497 Anne 3,198 4,571 5,643 6,857 5,067 Albi 4,143 4,735 4,990 6,265 5,076 Ilse II 2,643 4,123 4,710 5,250 4,218 Almi 4,539 2,929 8,000 7,286 5,688 Anka 4,500 4,571 4,857 5,000 4,732 Sasse 4,733 4,672 5,143 4,715 4,816 Sina 2,500 4,457 4,286 4,023 3,816 Reni 1,957 1,678 1,895 2,058 1,897 Anna 2,357 3,088 3,357 3,250 3,004 Rapunzel 3,286 4,000 3,071 2,621 3,223
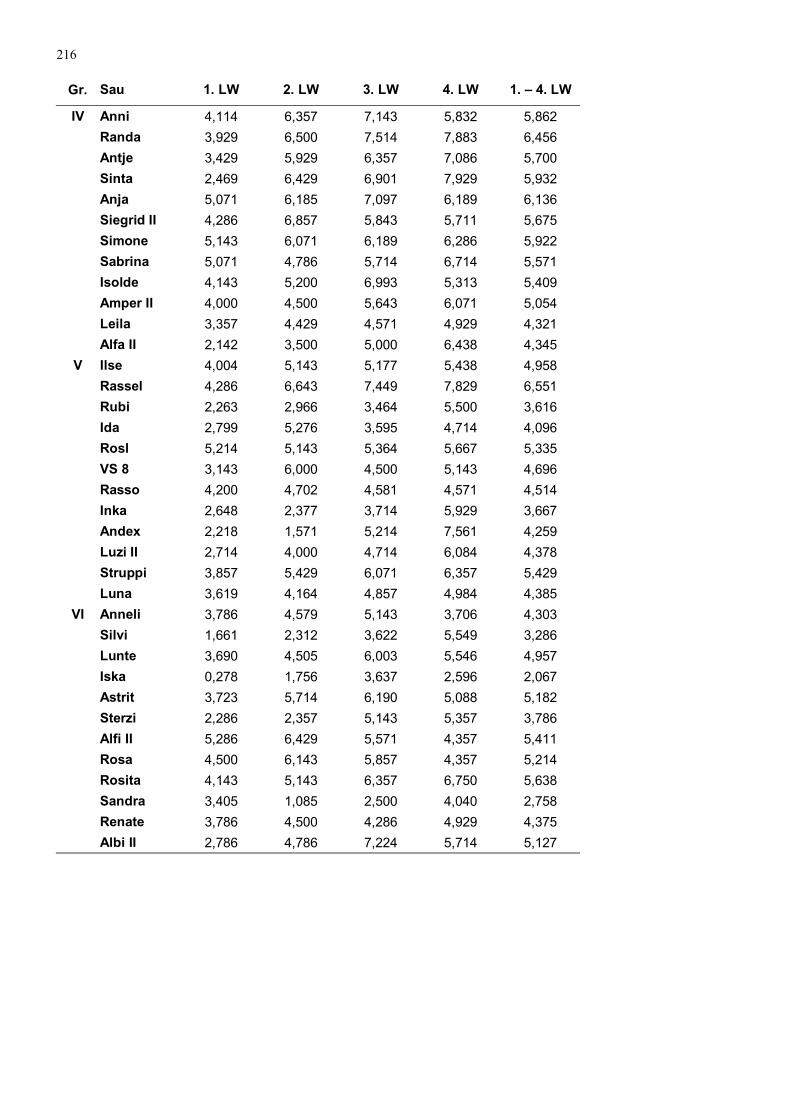
216
Gr. Sau 1. LW 2. LW 3. LW 4. LW 1. � 4. LW
IV Anni 4,114 6,357 7,143 5,832 5,862 Randa 3,929 6,500 7,514 7,883 6,456 Antje 3,429 5,929 6,357 7,086 5,700 Sinta 2,469 6,429 6,901 7,929 5,932 Anja 5,071 6,185 7,097 6,189 6,136 Siegrid II 4,286 6,857 5,843 5,711 5,675 Simone 5,143 6,071 6,189 6,286 5,922 Sabrina 5,071 4,786 5,714 6,714 5,571 Isolde 4,143 5,200 6,993 5,313 5,409 Amper II 4,000 4,500 5,643 6,071 5,054 Leila 3,357 4,429 4,571 4,929 4,321 Alfa II 2,142 3,500 5,000 6,438 4,345
V Ilse 4,004 5,143 5,177 5,438 4,958 Rassel 4,286 6,643 7,449 7,829 6,551 Rubi 2,263 2,966 3,464 5,500 3,616 Ida 2,799 5,276 3,595 4,714 4,096 Rosl 5,214 5,143 5,364 5,667 5,335 VS 8 3,143 6,000 4,500 5,143 4,696 Rasso 4,200 4,702 4,581 4,571 4,514 Inka 2,648 2,377 3,714 5,929 3,667 Andex 2,218 1,571 5,214 7,561 4,259 Luzi II 2,714 4,000 4,714 6,084 4,378 Struppi 3,857 5,429 6,071 6,357 5,429 Luna 3,619 4,164 4,857 4,984 4,385
VI Anneli 3,786 4,579 5,143 3,706 4,303 Silvi 1,661 2,312 3,622 5,549 3,286 Lunte 3,690 4,505 6,003 5,546 4,957 Iska 0,278 1,756 3,637 2,596 2,067 Astrit 3,723 5,714 6,190 5,088 5,182 Sterzi 2,286 2,357 5,143 5,357 3,786 Alfi II 5,286 6,429 5,571 4,357 5,411 Rosa 4,500 6,143 5,857 4,357 5,214 Rosita 4,143 5,143 6,357 6,750 5,638 Sandra 3,405 1,085 2,500 4,040 2,758 Renate 3,786 4,500 4,286 4,929 4,375 Albi II 2,786 4,786 7,224 5,714 5,127

217
Anhangstabelle 20: Lebendmassen der Sauen am 60. und 110. TT sowie am 1. und
28. LT in kg
Gr. Sau 60. TT 110. TT 1. LT 28. LT
I Stilla 272 295 269 257 Alfi 259 279 238 212 Alpin 208 235 217 191 Amper 272 274 270 255 Arne 198 238 228 183 Lupine 202 220 202 162 Anneli II 250 263 245 234 Isabell 276 293 264 242 Silvi II 272 294 276 241 Alfreda 228 247 223 180 Raupe 212 240 212 148 Asti 222 238 222 192
II Irka 208 234 219 198 Ronja 273 294 277 254 Siegrid 203 228 223 251 Saba 244 254 245 240 Alfa 253 294 279 265 Saale 242 251 254 239 Angela 191 213 190 180 Alpe 205 235 223 202 Rubi II 217 238 226 184 Ida II 262 277 270 223 Alida 238 260 241 206 Samanta 256 274 244 246
III Alpha 220 238 225 215 Luzi 244 255 241 249 Anne 248 258 242 219 Albi 205 220 194 186 Ilse II 235 260 234 232 Almi 219 245 224 220 Anka 265 291 275 251 Sasse 258 272 263 249 Sina 291 310 288 265 Reni 220 243 209 180 Anna 255 266 245 184 Rapunzel 221 253 269 257
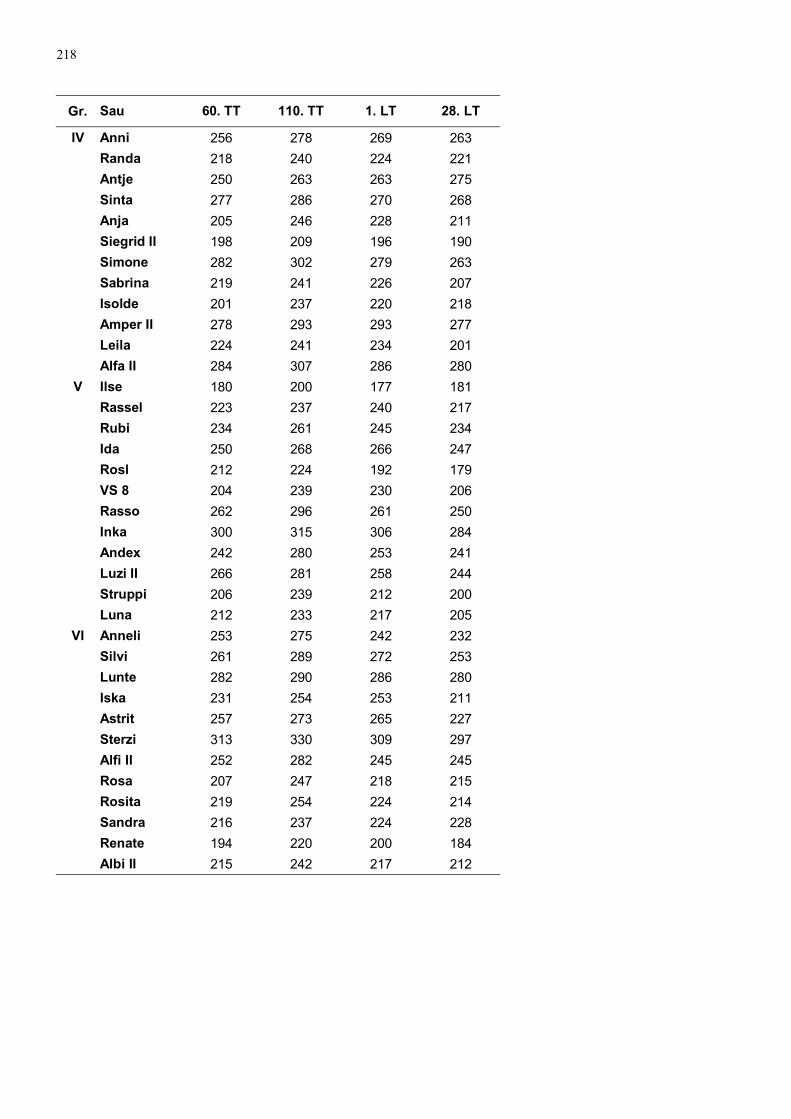
218
Gr. Sau 60. TT 110. TT 1. LT 28. LT
IV Anni 256 278 269 263 Randa 218 240 224 221 Antje 250 263 263 275 Sinta 277 286 270 268 Anja 205 246 228 211 Siegrid II 198 209 196 190 Simone 282 302 279 263 Sabrina 219 241 226 207 Isolde 201 237 220 218 Amper II 278 293 293 277 Leila 224 241 234 201 Alfa II 284 307 286 280
V Ilse 180 200 177 181 Rassel 223 237 240 217 Rubi 234 261 245 234 Ida 250 268 266 247 Rosl 212 224 192 179 VS 8 204 239 230 206 Rasso 262 296 261 250 Inka 300 315 306 284 Andex 242 280 253 241 Luzi II 266 281 258 244 Struppi 206 239 212 200 Luna 212 233 217 205
VI Anneli 253 275 242 232 Silvi 261 289 272 253 Lunte 282 290 286 280 Iska 231 254 253 211 Astrit 257 273 265 227 Sterzi 313 330 309 297 Alfi II 252 282 245 245 Rosa 207 247 218 215 Rosita 219 254 224 214 Sandra 216 237 224 228 Renate 194 220 200 184 Albi II 215 242 217 212

Lebenslauf
Persönliche Daten
Name, Vornamen: Pampuch, Franz Gabriel
Geburtsdatum: 29.06.1976
Geburtsort: Neresheim
Familienstand: ledig
Schulausbildung
1982 - 1986 Grundschule, Weisingen
1986 - 1996 Albertus-Gymnasium, Lauingen
Wehrdienst
1996 - 1997 Grundwehrdienst und
Ausbildung zum Militärkraftfahrer
Studium und beruflicher Werdegang
1997 - 2001 Studium der Landwirtschaft an der
Fachhochschule Weihenstephan;
Studienschwerpunkt Tierische Erzeugung
seit 2001 Wissenschaftlicher Mitarbeiter am Department für
Tierwissenschaften; Fachgebiet Tierernährung
Related Documents











