The Epigenetic Factor Kmt2a/Mll1 Regulates NeuralProgenitor Proliferation and Neuronal and GlialDifferentiation
Yin-Cheng Huang,1,2 Hung-Yu Shih,3 Sheng-Jia Lin,3 Ching-Chi Chiu,4 Tsu-Lin Ma,3
Tu-Hsueh Yeh,2,4,5 Yi-Chuan Cheng3
1 Department of Neurosurgery, Chang Gung Memorial Hospital at Linkou Medical Center,Taoyuan, Taiwan
2 College of Medicine, Chang Gung University, Taoyuan, Taiwan
3 Graduate Institute of Biomedical Sciences, College of Medicine, Chang Gung University,Taoyuan, Taiwan
4 Neuroscience Research Center, Chang Gung Memorial Hospital at Linkou Medical Center, Taoyuan,Taiwan
5 Section of Movement Disorders, Department of Neurology, Chang Gung Memorial Hospital at LinkouMedical Center, Taoyuan, Taiwan
Received 5 June 2014; revised 14 September 2014; accepted 29 September 2014
ABSTRACT: Multiple epigenetic factors play a
critical role in cell proliferation and differentiation. How-
ever, their function in embryogenesis, especially in neural
development, is currently unclear. The Trithorax group
(TrxG) homolog KMT2A (MLL1) is an important epige-
netic regulator during development and has an especially
well-defined role in hematopoiesis. Translocation and
aberrant expression of KMT2A is often observed in many
tumors, indicating its proto-oncogenic character. Here,
we show that Kmt2a was essential for neural develop-
ment in zebrafish embryos. Disrupting the expression of
Kmt2a using morpholino antisense oligonucleotides and
a dominant-negative variant resulted in neurogenic phe-
notypes, including downregulated proliferation of neural
progenitors, premature differentiation of neurons, and
impaired gliogenesis. This study therefore revealed a
novel function of Kmt2a in cell proliferation and differ-
entiation, providing further insight into the function of
TrxG proteins in neural development and brain
tumors. VC 2014 Wiley Periodicals, Inc. Develop Neurobiol 00: 000–
000, 2014
Keywords: Kmt2a; neural progenitors; proliferation;
differentiation; zebrafish
INTRODUCTION
Epigenetic regulatory mechanisms play a critical role
in brain development and in the development of vari-
ous diseases and tumors. The Trithorax group (TrxG)
and Polycomb group (PcG) proteins are the major
chromatin modulators that activate or silence target
gene expression, and defects in these proteins cause
Additional Supporting Information may be found in the onlineversion of this article.
Correspondence to: Y.-C. Cheng ([email protected]) orT.-H. Yeh ([email protected]).
Yin-Cheng Huang and Hung-Yu Shih contributed equally tothis work.
Contract grant sponsor: Chang Gung Memorial Hospital; con-tract grant numbers: CMRPD3B0041, CMRPD3B0042.
Contract grant sponsor: National Science Council of Taiwan;contract grant number: 102-2311-B-182-002-MY3.� 2014 Wiley Periodicals, Inc.Published online 00 Month 2014 in Wiley Online Library(wileyonlinelibrary.com).DOI 10.1002/dneu.22235
1

homeotic transformations. The Lysine (K)-specific
methyltransferase 2A [KMT2A, also known as mixed
lineage leukemia 1 (MLL1)] has been identified as
the mammalian ortholog of Drosophila trithorax(trx) and belongs to the Trithorax group (TrxG) of
proteins (Djabali et al., 1992). The encoded protein
contains 9–10 zinc-finger motifs and a highly con-
served SET [Su(var), Enhancer of zeste, trx] domain.
KMT2A mediates chromatin modifications through
its histone H3 lysine 4 methyltransferase activity and
is known to directly regulate homeotic genes. Multi-
ple chromosomal translocations involving this gene
are the cause of certain acute lymphoid leukemia and
acute myeloid leukemia (Mohan et al., 2010).
The in vivo function of KMT2A has been analyzed
in invertebrates and vertebrates. Drosophila mutant of
trx showed homeotic transformation (Mozer and
Dawid, 1989). The Kmt2a homozygous knockout was
embryonically lethal, and most of the mice died before
embryonic day 12.5. By contrast, heterozygous knock-
out mice survived and exhibited growth retardation,
hematopoietic defects, and skeletal malformation (Yu
et al., 1995). The phenotypical differences between
the homozygous and the heterozygous littermates sug-
gested a dosage-sensitive regulation by the KMT2A
protein. Mice harboring a truncation of Kmt2a exhibit
various phenotypes, such as failure of preimplantation
(Ayton et al., 2001), fetal liver hematopoiesis (Yagi
et al., 1998), and acute leukemia (Dobson et al., 1999),
depending on the particular allele knocked out and the
tissues in, which Kmt2a was deleted. Furthermore,
knockdown of Kmt2a expression in zebrafish embryos
resulted in hematopoietic defects, which is a conserved
phenotype observed in mammals (Wan et al., 2011).
These studies demonstrated the important role of
KMT2A in hematopoiesis at both physiological and
pathological levels.
The enriched and ubiquitous expression of
KMT2A suggests its role in regulating cellular proc-
esses in tissues in addition to the hematopoietic sys-
tem. Previously, a conditional KMT2A knockout
study showed impaired neuronal differentiation in
postnatal mouse brain, demonstrating an essential
role of KMT2A in neurogenesis (Lim et al., 2009).
More recently, two studies revealed that KMT2A is
expressed in hypoxic conditions (Heddleston et al.,
2012) and is required for the growth of glioblastoma
stem cells (Gallo et al., 2013), suggesting that
KMT2A is associated with glial-derived tumors and
may have a potential role in gliogenesis. However,
the role of KMT2A in brain malignancies still
requires further characterization. The prenatal func-
tion of KMT2A in the developing nervous system
remains especially unclear, and its study may provide
valuable information toward elucidating the role of
KMT2A in brain tumorigenesis.
In this study, we interfered with the expression of
Kmt2a in zebrafish embryos to study the endogenous
role during neural development. Because the study of
mice suggested that KMT2A acts in a dosage-sensitive
manner and exhibit lethal actions in homozygous knock-
out mice (Yu et al., 1995), we knocked down without
completely abolishing the endogenous Kmt2a expression
using a kmt2a antisense morpholino. In addition, we
used a dominant-negative kmt2a variant to confirm the
results of the knockdown experiment further. The results
revealed that the embryos with Kmt2a deficiency exhib-
ited decreased neural progenitor cell proliferation, prema-
ture differentiation of neurons, and defected gliogenesis.
Our data show a novel function for Kmt2a in the regula-
tion of cellular processes, providing further insights into
the diverse roles of Kmt2a in neural development and a
possible mechanism in brain tumor formation.
MATERIALS AND METHODS
Ethics Statement
All experiments were performed in strict accordance with
standard guidelines for zebrafish work and approved by the
Institutional Animal Care and Use Committee of Chang
Gung University (IACUC approval number: CGU12–039).
Fish Maintenance and Mutants
T€u (wild type) and Tg(gfap:egfp) zebrafish embryos were
purchased from the Zebrafish International Resource Center
(Oregon) and were raised, maintained, and paired under
standard conditions. The embryos were staged according to
the number of somites, hours postfertilization, and days
postfertilization (Kimmel et al., 1995).
Generation of Constructs
kmt2aN84 was polymerase chain reaction (PCR)-amplified
with high fidelity Pfu and with primers (F: 50-GGATCCGCCGCCACCATGGCGCACAGCTGTCGGT
GGC-30and R: 50-GGATCCCTATTATTCCTCCTC
CCCGCTACTGGAGCC-30) according to the GenBank
sequence (accession number: XM_005157583.1). The Kmt2amorpholino binding sequence was inserted upstream of an
enhanced green fluorescent protein (eGFP) reporter in the
pCS21 vector to create 50kmt2a-EGFP construct to evaluate
the specificity and efficiency of morpholino.
RNA and Morpholino Injection
Capped RNA encoding the full coding sequence of
kmt2aN84 was prepared as described previously (Chung
et al., 2011). Antisense morpholino oligonucleotides were
2 Huang et al.
Developmental Neurobiology

purchased from Gene Tools, LLC (Oregon). A morpholino
against kmt2a (TGCTGAGATCGCTCGTTCGGGGCTA)
that corresponds to 272 to 248 to the translation start site
was used. Basic Local Alignment Search Tool (Blast) anal-
ysis revealed homology of less than 20 bp identity for
kmt2a morpholino to other genomic sequences, none of
which corresponded to 50 UTR or exon–intron splicing site
of predicted or characterized genes, suggesting that the
morpholino would be specific for kmt2a. As a control
experiment, a morpholino designed with a random nucleo-
tide sequence not found in the zebrafish genome (50-CCTCTTACCTCAGTTACAATTTATA-30; Gene Tools)
or a morpholino with five bases mismatch to kmt2a mor-
pholino (50-TAcCCCCcAAgGAGCcATCTCAcCA-30; mis-
matched bases are indicated by small letters) was injected
in equal amounts to the kmt2a morpholino. All injections
were performed at the one- to two-cell stage and cRNAs or
morpholinos were introduced into blastomeres.
Histological Analysis
Digoxigenin-UTP labeled riboprobes were synthesized
according to the manufacturer’s instructions (Roche) and insitu hybridizations were performed as described previously
(Cheng et al., 2013). The color reaction was carried out using
the NBT/BCIP substrate (Roche). For immunohistochemistry,
the embryos were blocked in 5% goat serum and incubated
with mouse anti-HuC/HuD monoclonal 16A11 antibody (1/
500 dilution, Invitrogen), rabbit phospho-histone H3 antibody
(1:500, Millipore), or rabbit monoclonal antiactive caspase-3
(1:200, Abcam). Fluorochrome-conjugated antibodies Alexa
Fluor 488 goat anti-mouse (Invitrogen) was used to detect the
primary antibodies. Embryos were mounted with Vectashield
mounting medium (Vector Laboratories).
Quantitative Analysis
For quantitative real-time PCR (qPCR), embryos were
homogenized in TRIzol reagent (Invitrogen), and total
RNA was extracted using a standard method. cDNA was
synthesized from total RNA with random hexamer priming
using RevertAid First Strand cDNA Synthesis Kit (Fermen-
tas). qPCR was performed on an ABI StepOneTM Real-
Time PCR System (Applied Biosystems) with SYBR green
fluorescent label (Fermentas). Primers for sox2 (F: 50-CG
GAAAATGGCACAGGAGAA-30; R: 50-GTAATCCGGGT
GTTCCTTCATG-30), neurogenin1 (F: 50-CGCACACGGA
TGATGAAGACTCGCG-30; R: 50-CGGTTCTTCTTCAC-
GACGTGCACAGTGG-30), slc1a3 (F: 50-GTAACGGGGA
GACGCGTCTGCAGCG-30; R: 50-GATTATTCCCACGA
TGACGGCGGCG-30), gata1 (F: 50-ACACAGTCCAGTT
CGCCAAGT-30; R: 50-TGGAGAGGTGTTTTTGGGAA
A-30), and gapdh (F: 50-ACCCGTGCTGCTTTCTTGAC-30;R: 50-GACCAGTTTGCCGCCTTCT-30) were used. Gene
expression levels were normalized to gapdh and assessed
using the comparative CT (40 cycles) according to the manu-
facturer’s instructions (Applied Biosystems).
For Western blot analysis, embryos were homogenized
in sodium dodecyl sulfate (SDS) lysis buffer. Sixty micro-
grams was loaded on a 12% SDS polyacrylamide gel, trans-
ferred to a polyvinylidene difluoride (PVDF) membrane
and detected with anti-GFP antibody (1:1000, Invitrogen)
or tubulin (1:5000, Sigma). After washes, membranes were
incubated with goat anti-Mouse horseradish peroxidase
(HRP)-conjugated secondary Ab (Chemicon) and devel-
oped with ECL (Millipore). Band intensities were quanti-
fied using Multi Gaugre analysis software.
Statistical analysis was performed using Student’s t-test
in Microsoft ExcelVR 2007. The significance level was set at
p< 0.05. All reactions were performed in triplicate for each
sample.
RESULTS
kmt2a Morpholinos and the Dominant-Negative kmt2a Variant Effectively Inter-fere Kmt2a Expression
The sequence and expression of kmt2a have been
reported previously (Robinson et al., 2011; Wan
et al., 2011). We performed expression analysis with
more focus on the developing nervous system and
found that kmt2a was ubiquitously expressed in the
entire embryo from four-cell stage but later restricted
in the brain from 48-hours postfertilization (hpf), as
previously described [Fig. 1(A)].
To study the role of Kmt2a in the developing nerv-
ous system, we used a morpholino knockdown
approach to interfere with the endogenous expression
of Kmt2a. Two morpholinos were used in the previ-
ous study to knockdown Kmt2a expression, but these
morpholinos induced off-target cell apoptosis (Wan
et al., 2011). Therefore, we synthesized another anti-
sense morpholino that targeted the alternative transla-
tion start site of kmt2a mRNA to block protein
production. This morpholino showed no off-target
effects, as described later. BLAST analysis detected
<20 bp identity between the kmt2a morpholino and
other genomic sequences, none of which corre-
sponded to the 50-UTR or exon–intron splicing site of
the predicted or characterized genes, suggesting that
the morpholino may act specifically on kmt2a. The
specificity of the kmt2a morpholino was further con-
firmed by coinjection of a reporter construct that
fused with the kmt2a morpholino binding sequences
upstream of an eGFP reporter. Specific knockdown
of the translation of this construct (evident by eGFP
loss) was observed on coinjection with the kmt2amorpholinos but not with the 5-mismatch control
morpholino [Supporting Information Fig. 1(A)]. A
previous study showed that Kmt2a is essential for
hematopoiesis in zebrafish embryos (Wan et al.,
2011). Downregulation of the hematopoietic marker
Kmt2a Regulates Neurodevelopment 3
Developmental Neurobiology
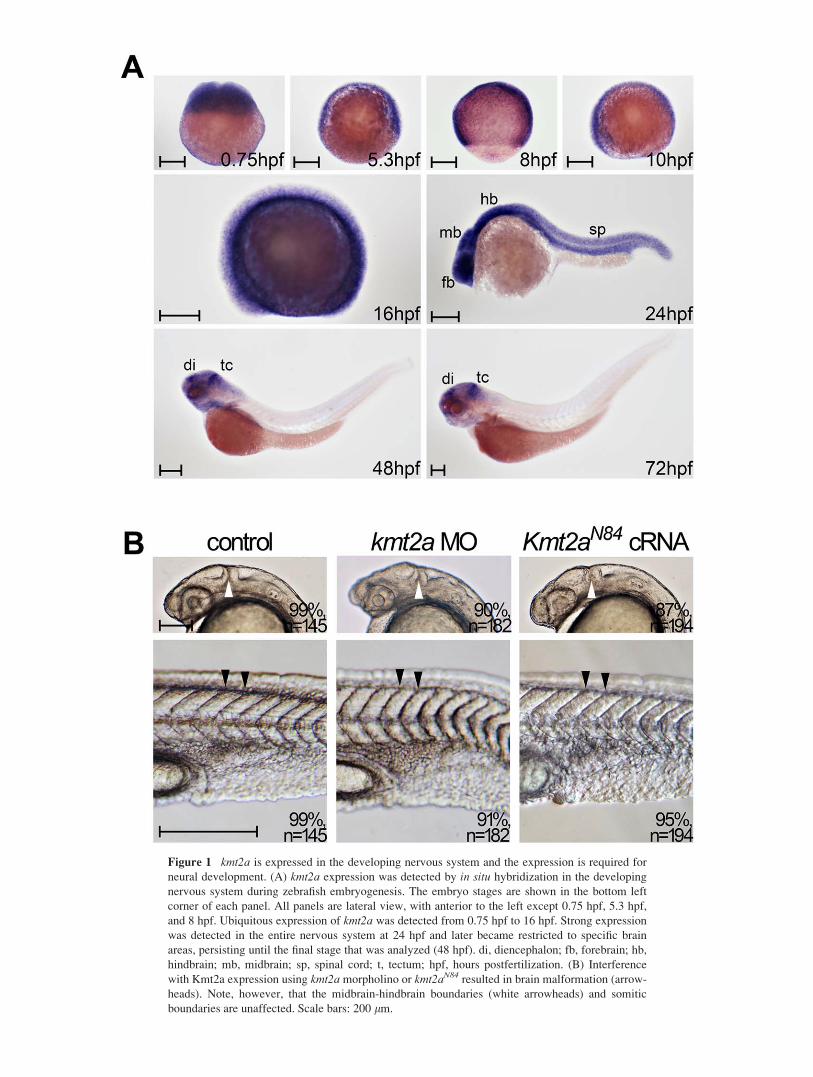
Figure 1 kmt2a is expressed in the developing nervous system and the expression is required for
neural development. (A) kmt2a expression was detected by in situ hybridization in the developing
nervous system during zebrafish embryogenesis. The embryo stages are shown in the bottom left
corner of each panel. All panels are lateral view, with anterior to the left except 0.75 hpf, 5.3 hpf,
and 8 hpf. Ubiquitous expression of kmt2a was detected from 0.75 hpf to 16 hpf. Strong expression
was detected in the entire nervous system at 24 hpf and later became restricted to specific brain
areas, persisting until the final stage that was analyzed (48 hpf). di, diencephalon; fb, forebrain; hb,
hindbrain; mb, midbrain; sp, spinal cord; t, tectum; hpf, hours postfertilization. (B) Interference
with Kmt2a expression using kmt2a morpholino or kmt2aN84 resulted in brain malformation (arrow-
heads). Note, however, that the midbrain-hindbrain boundaries (white arrowheads) and somitic
boundaries are unaffected. Scale bars: 200 lm.
gata1 as previously reported [Supporting Information
Fig. 1(B,C)] indicated the effectiveness of the kmt2amorpholino.
The specificity of the morpholino could not be con-
firmed by rescue experiments using kmt2a cRNA due
to the extra-long coding sequence (12,703 bp) that
made it difficult to amplify the cDNA, which also
could not be transcribed in vitro. Therefore, we con-
structed a deletion variant that contained only the 84-
amino acid N terminus of kmt2a (kmt2aN84). This N-
terminal minipeptide contains a highly conserved
Menin-binding motif and has been demonstrated to act
as a dominant negative form that disrupts normal func-
tioning of kmt2a during hematopoiesis (Wan et al.,
2011). Injection of kmt2aN84 downregulated gata1expression in a manner identical to that observed in
kmt2a knockdown embryos [Supporting Information
Fig. 1(B,C) and Fig. 1(B)]. Therefore, kmt2aN84 injec-
tion confirmed the specificity of the kmt2a morpholino
and was used in each experiment as described later.
Kmt2a Depletion is Sufficient to ReduceNeural Progenitor Proliferation
Embryos injected with kmt2a morpholino or
kmt2aN84 were first analyzed at 24 hpf for morpho-
logical defects. The injection of kmt2a morpholino or
kmt2aN84 produced an identical phenotype exhibiting
brain malformation, particularly thickening and
abnormally folded structures of the neural tube, indi-
cating that Kmt2a is required for neural development
[Fig. 1(B)]. A previous study showed aberrant seg-
mental boundaries of spinal ganglia and somites in a
Kmt2a deficient mouse (Yu et al., 1998). In contrast,
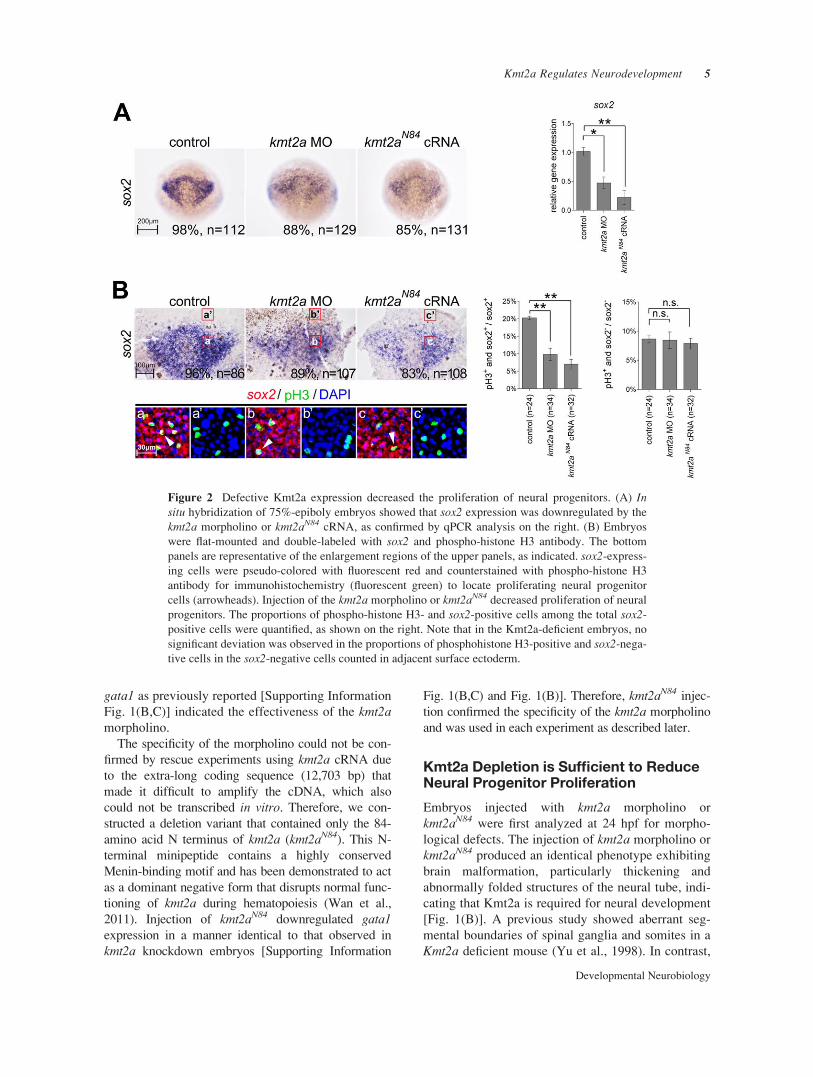
Figure 2 Defective Kmt2a expression decreased the proliferation of neural progenitors. (A) Insitu hybridization of 75%-epiboly embryos showed that sox2 expression was downregulated by the
kmt2a morpholino or kmt2aN84 cRNA, as confirmed by qPCR analysis on the right. (B) Embryos
were flat-mounted and double-labeled with sox2 and phospho-histone H3 antibody. The bottom
panels are representative of the enlargement regions of the upper panels, as indicated. sox2-express-
ing cells were pseudo-colored with fluorescent red and counterstained with phospho-histone H3
antibody for immunohistochemistry (fluorescent green) to locate proliferating neural progenitor
cells (arrowheads). Injection of the kmt2a morpholino or kmt2aN84 decreased proliferation of neural
progenitors. The proportions of phospho-histone H3- and sox2-positive cells among the total sox2-
positive cells were quantified, as shown on the right. Note that in the Kmt2a-deficient embryos, no
significant deviation was observed in the proportions of phosphohistone H3-positive and sox2-nega-
tive cells in the sox2-negative cells counted in adjacent surface ectoderm.
Kmt2a Regulates Neurodevelopment 5
Developmental Neurobiology

the brain and somitic boundaries appeared unaffected
by the injection of kmt2a morpholino or kmt2aN84
[Fig. 1(B)].
During development, the heterogeneous neurons
and glial cells are derived from neural stem cells and
progenitor cells originating from the neuroectoderm
in spatial- and temporal-related fashions. Therefore,
we first analyzed the effects of Kmt2a knockdown
using the neural progenitor marker sox2 during neural
induction. The result of whole-mount in situ hybrid-
ization showed significantly decreased sox2 expres-
sion [Fig. 2(A)]. Quantitative real-time PCR (qPCR)
analysis confirmed a 2-fold reduction in sox2 [Fig.
2(A)]. kmt2aN84 injection also downregulated the
expression of sox2 by 4-fold [Fig. 2(A)]. We ques-
tioned whether the reduced number of neural progen-
itors in Kmt2a deficient embryos resulted from
induction of apoptosis or inhibition of proliferation
and further analyzed cell proliferation using a phos-
phohistone H3 antibody and counterstaining with
sox2 to localize the proliferating neural progenitors.
The result revealed a decreased number of proliferat-
ing neural progenitors in embryos injected with the
kmt2a morpholino or kmt2aN84, an observation that
was further confirmed by a count of the proliferating
cells in the sox2-positive and sox2-negative popula-
tions [Fig. 2(B)]. This result suggests that Kmt2a is
required for the proliferation of neural progenitors.
To investigate whether the loss of sox2-positive
neural progenitors was due to cell death, apoptotic
neural precursors were analyzed by looking for the
presence of proteolytic activation of the effector
caspase-3 by immunohistochemistry on Kmt2a defi-
cient embryos. Following in situ hybridization with
sox2 RNA probe, embryos were subjected to immu-
nohistochemical staining to determine whether apo-
ptosis was localized to neural progenitors (Fig. 3).
Injection of kmt2a morpholino or kmt2aN84 did not
cause a significant alteration in the number of apopto-
tic neural progenitors, suggesting that altered Kmt2a
expression had no effect on cell survival (Fig. 3).
This result also confirmed that the effect of the kmt2amorpholino was due to specific Kmt2a knockdown
but not off-target cell apoptosis.
Impaired Kmt2a Expression CausesPrecocious Neuronal Differentiation
After the formation of neural progenitors, the next
step in neurogenesis is the specification of neuronal
precursors within the neurogenic region, a process
regulated by proneural genes. We examined the role
of Kmt2 in neuronal precursors using the proneural
marker neurogenin1. The results showed a 1.9 to 2.9-
fold decrease in neurogenin1 expression in embryos
injected with kmt2a morpholino or kmt2aN84 ana-
lyzed at the bug stage [Fig. 4(A,B)]. Although this
effect could be explained by the decreased neural
progenitors as described earlier and, therefore,
Figure 3 kmt2a morpholino or kmt2aN84 cRNA injection had no effect on neural progenitor apo-
ptosis. Apoptotic neural progenitor cells were labeled for sox2 (fluorescent red) and activated
caspase-3 antibody (fluorescent green). As shown in E, apoptotic neural progenitor cells were quan-
tified by counting the proportions of activated caspase-3- and sox2-positive (or -negative) cells
among the total sox2-positive (or -negative) cells. *p< 0.05; **p< 0.01; n.s., not significant. Scale
bar: 100 lm, applies to all panels.
6 Huang et al.
Developmental Neurobiology

consequently caused the loss of neuronal precursors,
immunohistochemistry analysis using the postmitotic
neuronal marker HuC/D antibody revealed significant
upregulation of HuC/D in Kmt2a deficient embryos at
24 hpf [Fig. 4(C)]. Ectopic HuC/D-positive cells were
also observed in several regions where normal neuro-
nal differentiation would not occur [arrows in Fig.
4(C)]. This effect was confirmed by Western blot anal-
ysis showing a 3- to 4-fold increase in HuC/D expres-
sion in kmt2a deficient embryos [Fig. 4(D)].
Concurrently with the increased HuC/D expression,
the expression of neurogenin1 was significantly down-
regulated at 24 hpf [Fig. 4(E,F)]. These results sug-
gested that injection of the kmt2a morpholino or
kmt2aN84 could cause excessive neurogenesis. We
thus performed a time course analysis to investigate
whether impaired Kmt2a expression could elicit pre-
mature differentiation of neurons. The result showed
that the number of HuC/D-positive neurons was unal-
tered in Kmt2a deficient embryos in comparison to
control embryos at 14 hpf. However, HuC/D-positive
neurons started to be significantly upregulated in
Kmt2a deficient embryos from 16 hpf and onwards
(Fig. 5). This result indicated that neurons were pre-
maturely differentiated from neurogenein1-positive
precursors into HuC/D-positive differentiating neurons
and that this effect is separable from the decreased
proliferation of neural progenitors. The well-
organized, ladder-like arrays of neurons in the devel-
oping hindbrain were unaffected by the kmt2a mor-
pholino or kmt2aN84 cRNA injection (Figs. 4(C) and
5), suggesting that Kmt2a deficiency does not affect
Figure 4 Disrupted Kmt2a expression causes aberrant formation of neuronal precursors and
mature neurons. (A) At the bud stage, the expression level of neurogenin1 was significantly
decreased in embryos injected with the kmt2a morpholino or kmt2aN84 cRNA injected embryos in
comparison to the controls. (B) qPCR analysis confirmed the results obtained by in situ hybridiza-
tion in A. (C and E) The upper panels are enlargements of the hindbrain region, anterior to the top;
and the bottom panels present lateral views of the enlargements of the 3–9-somite levels of the spi-
nal cord. (C) HuC/D-expressing post-mitotic neurons increased massively in kmt2a morpholino or
kmt2aN84 cRNA injected embryos, as shown by immunohistochemical analysis with anti-HuC/D
antibody at 24 hpf. Note the ectopic HuC/D expression in the ventricular zone (arrowheads). (D)
Levels of HuC/D expression were confirmed by Western blot analysis and were quantified. (E) At
24 hpf, neurogenin1 expression was reduced significantly by the kmt2a morpholino and kmt2aN84.
(F) qPCR analysis further confirmed the decreased expression of neurogenein1 at 24 hpf in E.
*p< 0.05; **p< 0.01.
Kmt2a Regulates Neurodevelopment 7
Developmental Neurobiology

neural migration and patterning. In addition, we exam-
ined whether Kmt2a deficiency affected the proliferat-
ing neurons using phosphohistone H3 antibody and
counterstaining with neurogenin1. The results demon-
strated that neuronal proliferation remained intact in
embryos injected with kmt2a morpholino or kmt2aN84
(Supporting Information Fig. 2), indicating that Kmt2a
was not required for neuronal proliferation.
Kmt2a is Required for Gliogenesis
In addition to producing neurons, neural progenitors
generate glial derivatives such as astrocytes and oli-
godendrocytes. We accordingly analyzed the role of
Kmt2a in gliogenesis. Injection of the kmt2a morpho-
lino or kmt2aN84 cRNA downregulated the expression
of the early glial marker slc1a3a (Glast in mammals)
Figure 5 Kmt2a-deficient embryos exhibit upregulated HuC/D expression. Significant upregula-
tion of HuC/D signals, as analyzed by immunohistochemistry using an anti-HuC/D antibody, can
be detected at 16 hpf and 18 hpf in embryos injected with either the kmt2a morpholino or kmt2aN84
cRNA. Quantitative data are presented as the mean 6 standard deviation normalized to the number
of controls. *p< 0.05; **p< 0.01; n.s., not significant. Scale bar: 100 lm, applies to all panels.
8 Huang et al.
Developmental Neurobiology
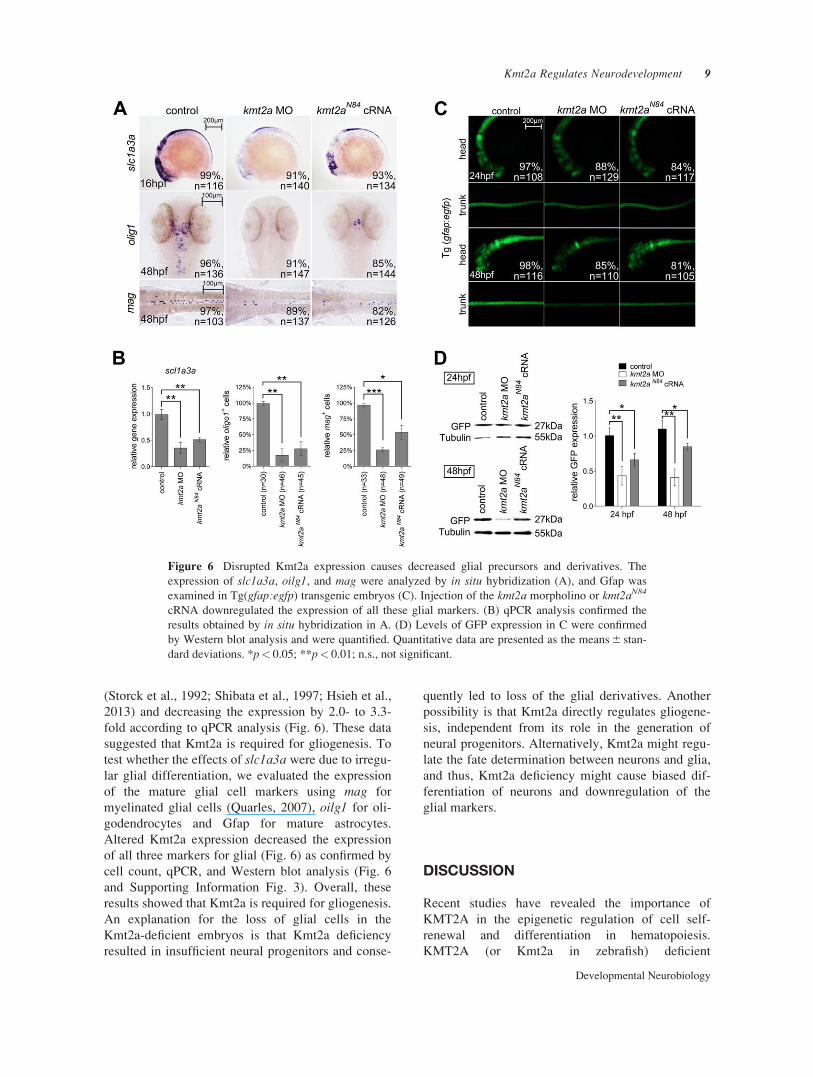
(Storck et al., 1992; Shibata et al., 1997; Hsieh et al.,
2013) and decreasing the expression by 2.0- to 3.3-
fold according to qPCR analysis (Fig. 6). These data
suggested that Kmt2a is required for gliogenesis. To
test whether the effects of slc1a3a were due to irregu-
lar glial differentiation, we evaluated the expression
of the mature glial cell markers using mag for
myelinated glial cells (Quarles, 2007), oilg1 for oli-
godendrocytes and Gfap for mature astrocytes.
Altered Kmt2a expression decreased the expression
of all three markers for glial (Fig. 6) as confirmed by
cell count, qPCR, and Western blot analysis (Fig. 6
and Supporting Information Fig. 3). Overall, these
results showed that Kmt2a is required for gliogenesis.
An explanation for the loss of glial cells in the
Kmt2a-deficient embryos is that Kmt2a deficiency
resulted in insufficient neural progenitors and conse-
quently led to loss of the glial derivatives. Another
possibility is that Kmt2a directly regulates gliogene-
sis, independent from its role in the generation of
neural progenitors. Alternatively, Kmt2a might regu-
late the fate determination between neurons and glia,
and thus, Kmt2a deficiency might cause biased dif-
ferentiation of neurons and downregulation of the
glial markers.
DISCUSSION
Recent studies have revealed the importance of
KMT2A in the epigenetic regulation of cell self-
renewal and differentiation in hematopoiesis.
KMT2A (or Kmt2a in zebrafish) deficient
Figure 6 Disrupted Kmt2a expression causes decreased glial precursors and derivatives. The
expression of slc1a3a, oilg1, and mag were analyzed by in situ hybridization (A), and Gfap was
examined in Tg(gfap:egfp) transgenic embryos (C). Injection of the kmt2a morpholino or kmt2aN84
cRNA downregulated the expression of all these glial markers. (B) qPCR analysis confirmed the
results obtained by in situ hybridization in A. (D) Levels of GFP expression in C were confirmed
by Western blot analysis and were quantified. Quantitative data are presented as the means 6 stan-
dard deviations. *p< 0.05; **p< 0.01; n.s., not significant.
Kmt2a Regulates Neurodevelopment 9
Developmental Neurobiology

experiments have been performed in vivo using
knockout mice (Yu et al., 1995) and morpholino
knockdown zebrafish (Wan et al., 2011), which
revealed the essential role of KMT2A (Kmt2a) in
embryonic hematopoiesis; however, no previous
study has described the role of KMT2A (Kmt2a) defi-
ciency in causing any abnormal development in the
nervous systems. Conditional knockout of Kmt2a in
the neural stem cells of the subventricular zone of the
postnatal nervous system demonstrated that although
the neural stem cells survive, proliferate, and effi-
ciently differentiate into glial lineages, neuronal dif-
ferentiation is severely impaired (Lim et al., 2009).
By contrast, other studies have suggested a potential
role of KMT2A in gliogenesis by stating that
KMT2A is required for the growth of the glioblas-
toma cells (Heddleston et al., 2012; Gallo et al.,
2013). Therefore, the role of KMT2A in the nervous
system, particularly in cell progression including pro-
liferation, survival, and differentiation, remains
unclear. This study evaluated the physiological role
of the Kmt2a protein in the development of the nerv-
ous system by disrupting the Kmt2a expression using
the kmt2a morpholino and the dominant-negative
kmt2a variant kmt2aN84. Disrupted endogenous
expression of Kmt2a in the zebrafish embryos
decreased the proliferation of neural progenitor cells,
which resemble neural stem cells in many ways but
undergo a limited number of replication cycles invivo, indicating that Kmt2a is essential for the prolif-
eration of neural progenitors or stem cells and might
thus regulate the proliferation of neural progenitor
and brain tumor stem cells through a similar mecha-
nism. Moreover, the Kmt2a-deficient embryos exhib-
ited premature differentiation of neurons and
defective gliogenesis, which indicated that Kmt2a is
essential for the inhibition of neuronal differentiation
as well as appropriate glial differentiation; this fur-
ther strengthens the explanations regarding the onco-
genic role of Kmt2a in brain tumor formation.
The molecular regulation of KMT2A (Kmt2a) in
neural progenitor proliferation, neurogenesis, or glio-
genesis is not clear. The best established target of
KMT2A are the Homeobox genes (HOX genes),
which are fundamental for segmental identity during
early development (Krivtsov and Armstrong, 2007).
We found that hoxa9a, one of the well-characterized
genes downstream of Kmt2a, was downregulated in
Kmt2a-deficient embryos (unpublished data), sug-
gesting that Kmt2a also positively regulates hoxa9a.
However, hoxa9a is not expressed in neural progeni-
tor populations, suggesting that Kmt2a-mediated neu-
ral progenitor proliferation is regulated by other
molecules. Some potential candidates are the compo-
nents in the Notch signaling pathway. Research in
Drosophila and mammals suggests that the Notch
pathway is regulated by TrxG proteins such as the
histone demethylase UTX and the chromatin remod-
eler BRM (Schuettengruber et al., 2011). In particu-
lar, the Drosophila homologue of Kmt2a, trithorax,
was shown to collaborate with Notch in gene activa-
tion (Bejarano and Milan, 2009). Notch-mediated
signaling plays a fundamental role in a variety of
neural developmental processes and in the pathoge-
nesis of several human cancers (Roy et al., 2007). In
the developing nervous system, Notch signaling posi-
tively regulates the maintenance of neural progenitors
and later governs the decision between neuronal and
glial lineages. Deficient Notch signaling resulted in
decreased proliferation of neural progenitors, prema-
ture neuronal differentiation, and defected gliogene-
sis (Chung et al., 2011; Cheng et al., 2013), which
highly resembles the phenotypes observed in Kmt2a-
deficient embryos. Therefore, it is worthwhile to fur-
ther investigate the regulatory mechanism between
KMT2A (Kmt2a) and the Notch signaling pathway in
neural development and tumorigenesis.
The authors thank David Wilkinson for neurogenin1and Paul Scotting for the sox2 constructs used in making
the riboprobes. The authors are also grateful to the Taiwan
Zebrafish Core facility at ZeTH and the Zebrafish Core in
Academia Sinica for providing fish.
REFERENCES
Ayton P, Sneddon SF, Palmer DB, Rosewell IR, Owen MJ,
Young B, Presley R, et al. 2001. Truncation of the Mll gene
in exon 5 by gene targeting leads to early preimplantation
lethality of homozygous embryos. Genesis 30:201–212.
Bejarano F, Milan M. 2009. Genetic and epigenetic mecha-
nisms regulating hedgehog expression in the Drosophila
wing. Dev Biol 327:508–515.
Cheng YC, Scotting PJ, Hsu LS, Lin SJ, Shih HY, Hsieh
FY, Wu HL, et al. 2013. Zebrafish rgs4 is essential for
motility and axonogenesis mediated by Akt signaling.
Cell Mol Life Sci 70:935–950.
Chung PC, Lin WS, Scotting PJ, Hsieh FY, Wu HL, Cheng
YC. 2011. Zebrafish Her8a is activated by Su(H)-depend-
ent notch signaling and is essential for the inhibition of
neurogenesis. PLoS One 6:e19394.
Djabali M, Selleri L, Parry P, Bower M, Young BD, Evans
GA. 1992. A trithorax-like gene is interrupted by chromo-
some 11q23 translocations in acute leukaemias. Nat
Genet 2:113–118.
Dobson CL, Warren AJ, Pannell R, Forster A, Lavenir I,
Corral J, Smith AJ, et al. 1999. The mll-AF9 gene fusion
in mice controls myeloproliferation and specifies acute
myeloid leukaemogenesis. EMBO J 18:3564–3574.
10 Huang et al.
Developmental Neurobiology

Gallo M, Ho J, Coutinho FJ, Vanner R, Lee L, Head R,
Ling EK, et al. 2013. A tumorigenic MLL-homeobox net-
work in human glioblastoma stem cells. Cancer Res 73:
417–427.
Heddleston JM, Wu Q, Rivera M, Minhas S, Lathia JD,
Sloan AE, Iliopoulos O, et al. 2012. Hypoxia-induced
mixed-lineage leukemia 1 regulates glioma stem cell
tumorigenic potential. Cell Death Differ 19:428–439.
Hsieh FY, Ma TL, Shih HY, Lin SJ, Huang CW, Wang
HY, Cheng YC. 2013. Dner inhibits neural progenitor
proliferation and induces neuronal and glial differentia-
tion in zebrafish. Dev Biol 375:1–12.
Kimmel CB, Ballard WW, Kimmel SR, Ullmann B,
Schilling TF. 1995. Stages of embryonic development of
the zebrafish. Dev Dyn 203:253–310.
Krivtsov AV, Armstrong SA. 2007. MLL translocations,
histone modifications and leukaemia stem-cell develop-
ment. Nat Rev Cancer 7:823–833.
Lim DA, Huang YC, Swigut T, Mirick AL, Garcia-
Verdugo JM, Wysocka J, Ernst P, et al. 2009. Chromatin
remodelling factor Mll1 is essential for neurogenesis
from postnatal neural stem cells. Nature 458:529–533.
Mohan M, Lin C, Guest E, Shilatifard A. 2010. Licensed to
elongate: A molecular mechanism for MLL-based leu-
kaemogenesis. Nat Rev Cancer 10:721–728.
Mozer BA, Dawid IB. 1989. Cloning and molecular charac-
terization of the trithorax locus of Drosophila mela-
nogaster. Proc Natl Acad Sci USA 86:3738–3742.
Quarles RH. 2007. Myelin-associated glycoprotein (MAG):
Past, present and beyond. J Neurochem 100:1431–1448.
Robinson BW, Germano G, Song Y, Abrams J, Scott M,
Guariento I, Tiso N, et al. 2011. mll ortholog containing
functional domains of human MLL is expressed through-
out the zebrafish lifespan and in haematopoietic tissues.
Br J Haematol 152:307–321.
Roy M, Pear WS, Aster JC. 2007. The multifaceted role of
notch in cancer. Curr Opin Genet Dev 17:52–59.
Schuettengruber B, Martinez AM, Iovino N, Cavalli G.
2011. Trithorax group proteins: Switching genes on and
keeping them active. Nat Rev Mol Cell Biol 12:799–814.
Shibata T, Yamada K, Watanabe M, Ikenaka K, Wada K,
Tanaka K, Inoue Y. 1997. Glutamate transporter GLAST
is expressed in the radial glia-astrocyte lineage of devel-
oping mouse spinal cord. J Neurosci 17:9212–9219.
Storck T, Schulte S, Hofmann K, Stoffel W. 1992. Struc-
ture, expression, and functional analysis of a Na(1)-
dependent glutamate/aspartate transporter from rat brain.
Proc Natl Acad Sci USA 89:10955–10959.
Wan X, Hu B, Liu JX, Feng X, Xiao W. 2011. Zebrafish
mll gene is essential for hematopoiesis. J Biol Chem 286:
33345–33357.
Yagi H, Deguchi K, Aono A, Tani Y, Kishimoto T, Komori
T. 1998. Growth disturbance in fetal liver hematopoiesis
of Mll-mutant mice. Blood 92:108–117.
Yu BD, Hanson RD, Hess JL, Horning SE, Korsmeyer SJ.
1998. MLL, a mammalian trithorax-group gene, func-
tions as a transcriptional maintenance factor in morpho-
genesis. Proc Natl Acad Sci USA 95:10632–10636.
Yu BD, Hess JL, Horning SE, Brown GA, Korsmeyer SJ.
1995. Altered Hox expression and segmental identity in
Mll-mutant mice. Nature 378:505–508.
Kmt2a Regulates Neurodevelopment 11
Developmental Neurobiology