



The role of innate immunity in the pathogenesis of Serious Non-AIDS Events
Suzanne CroweAssociate Director, Burnet Institute
Consultant Infectious Diseases Physician, The Alfred Hospital
Melbourne

Today’s presentation
HIV, LPS trigger inflammation
Contribution of monocyte activationPro-inflammatory cytokines
Involvement of innate immune pathways inFibrosis
Atherosclerotic plaque formationPro-coagulant activity
Metabolic dysregulationImmune senescence
Serious non-AIDS Events
How HIV & inflammation are linked
How breaches in gut mucosa trigger inflammation
Links between monocytemetabolism &inflammation
How monocyte activationImpacts these pathways
Innate immune senescence& risk of events---

Today’s presentation
HIV, LPS trigger inflammation
Contribution of monocyte activationPro-inflammatory cytokines
Involvement of innate immune pathways inFibrosis
Atherosclerotic plaque formationPro-coagulant activity
Metabolic dysregulationImmune senescence
Serious non-AIDS Events

Chronic inflammation and immune activation
• Chronic immune activation & inflammation is central to HIV pathogenesis– In both treated and untreated individuals – Starts during seroconversion
• Pro-inflammatory pathways triggered by– HIV replication and viremia– Co-infection eg with CMV– Microbial translocation across the gut mucosa

Stacey et al J Virol 2009 83:3719-3733
Intense cytokine storm in acute HIV infection prior to peak viremia
Cytokine response in acute HIV Stacey 2009
Ch
ang
es
in p
lasm
a c
ytok
ine
leve
ls
Time since HIV infection (days)
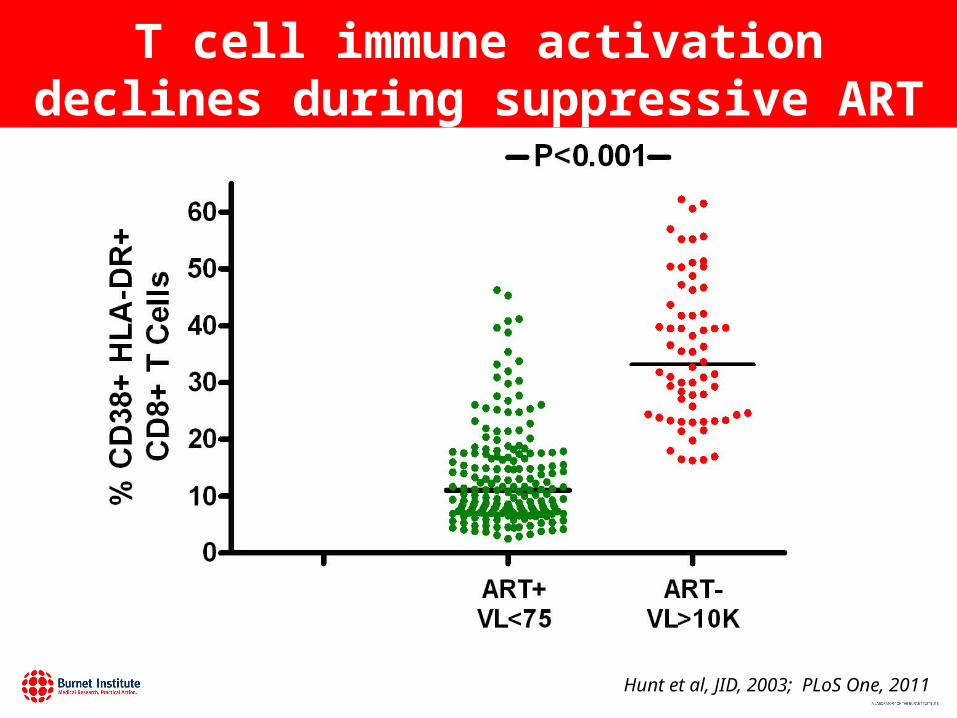
T cell immune activation declines during suppressive ART
Hunt et al, JID, 2003; PLoS One, 2011

But T cell activation remains elevated during suppressive ART
Hunt et al, JID, 2003; PLoS One, 2011

What about Innate immunity and pro-inflammatory pathways?

What is meant by pro-inflammatory pathways?
• Activation of cells eg monocytes & mϕ that produce pro-inflammatory cytokines
• Pro-inflammatory cytokines include IL-6, TNF-alpha
• Chronic inflammation leads to fibrosis and end-organ disease
R Medzhitov Cell 2010; 140:771

Differing pathogenetic roles of T cells & monocytes in HIV infection
• T cell activation – Main effect: Central role in T cell decline
• Monocyte activation– Main effect: Central role in chronic inflammation
– Important in pathogenesis of longer-term, non-AIDS morbidity
– CD16+ pro-inflammatory monocytes increased in HIV & only partly normalized by cART
1. Westhorpe C et al Immunol & Cell Biol 2014;92:1338; 2.Tenorio AR et al J Infect Dis 2014; 3. Lederman MM et al Adv Immunol 2013; 119”51; 4. Sandler &Sereti Curr Opin HIV AIDS 2014;9:72
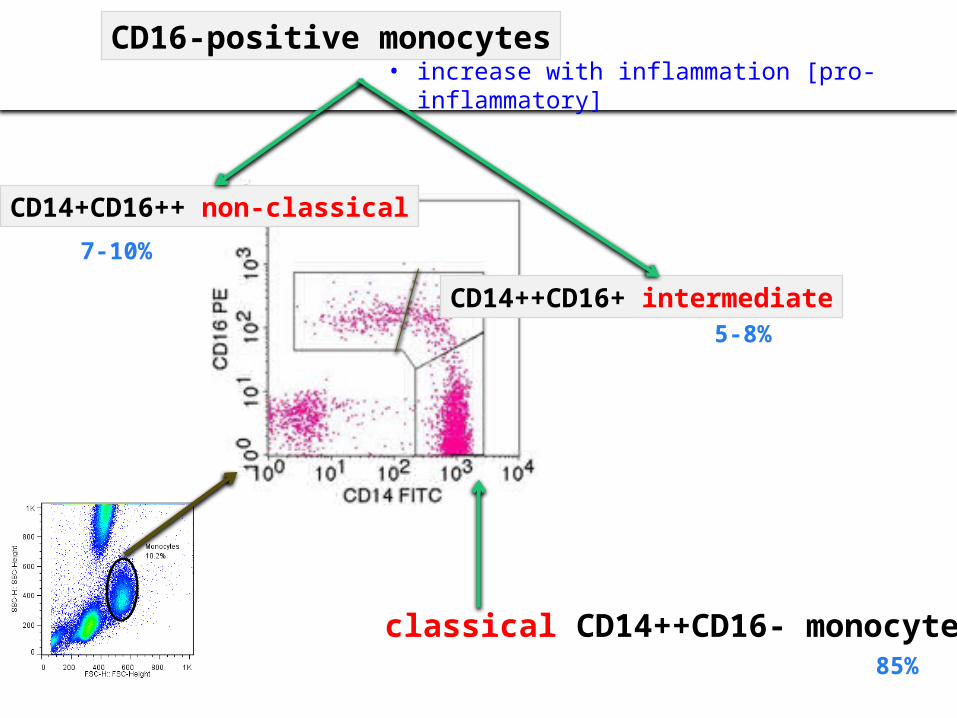
• increase with inflammation [pro-inflammatory] CD16-positive monocytes
classical CD14++CD16- monocytes
CD14+CD16++ non-classical
CD14++CD16+ intermediate
7-10%
5-8%
85%

Traditional Inflammatory Patrolling
1. Funderburg NT et al Blood 2012 120: 45992. Krishnan S et al J Infect Dis 2014; 209:931
CD16+ monocyte subsets are increased in uncontrolled viremia & may normalize with ART
CD14++CD16-
Classical
CD14++ CD16+
Intermediate
CD14+CD16++
Non classical
HIV infection increases the proportions of CD16+ monocytes in blood
cART restores the proportions of monocytes in blood1
o CD16+ monocytes remain elevated in elite controllers2
HIV- HIV+ VL<400HIV+ VL>400
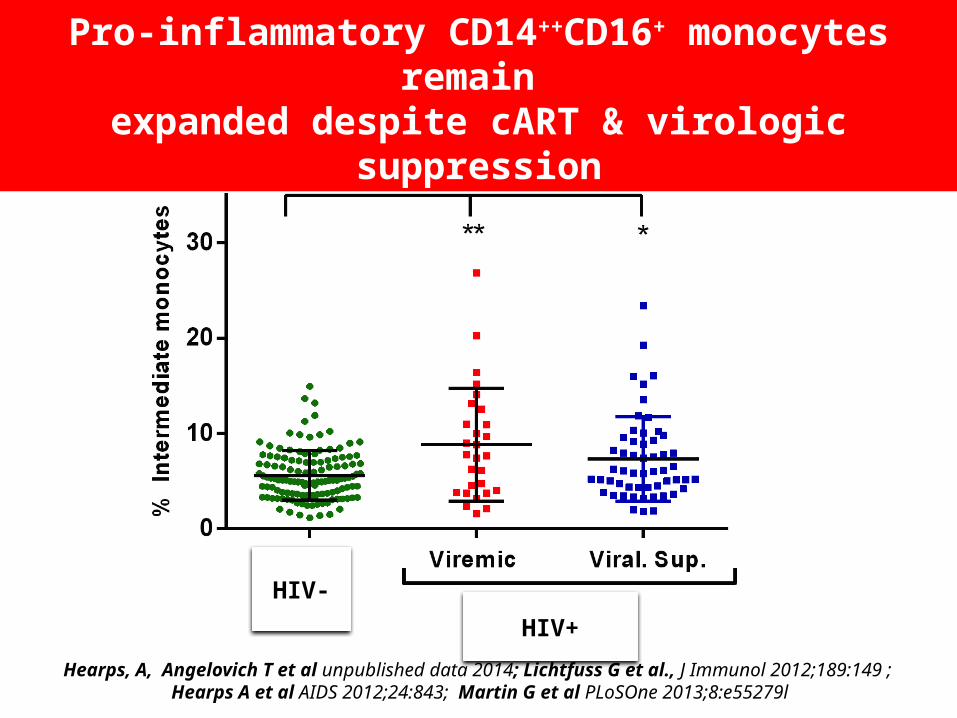
Hearps, A, Angelovich T et al unpublished data 2014; Lichtfuss G et al., J Immunol 2012;189:149 ; Hearps A et al AIDS 2012;24:843; Martin G et al PLoSOne 2013;8:e55279l
Pro-inflammatory CD14++CD16+ monocytes remain expanded despite cART & virologic suppression
HIV-
HIV+
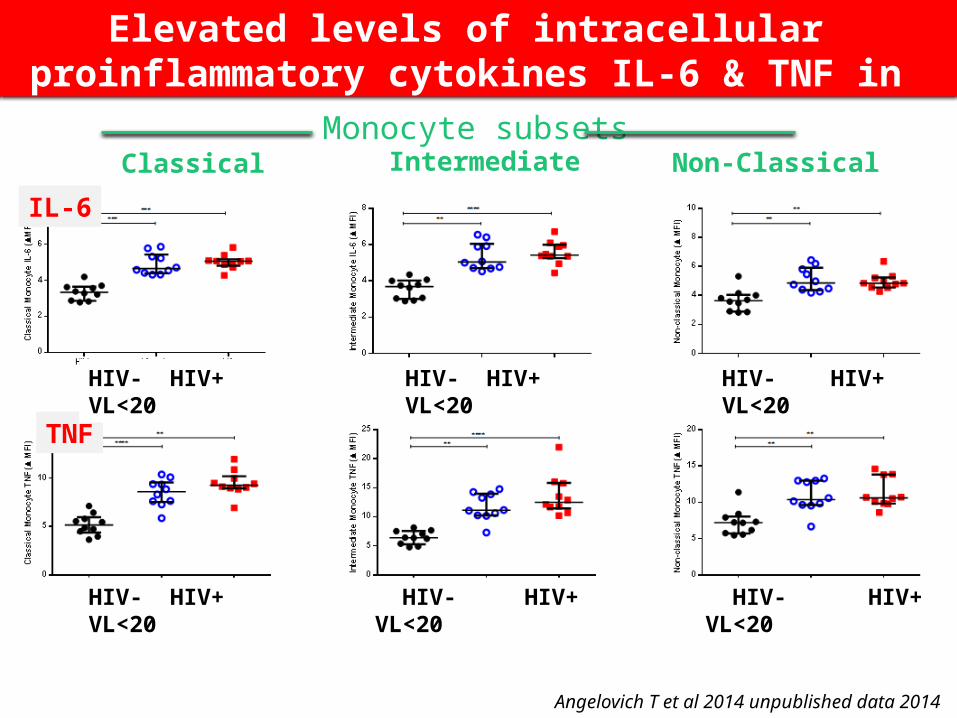
Intermediate Non-ClassicalClassical
Angelovich T et al 2014 unpublished data 2014
Elevated levels of intracellular proinflammatory cytokines IL-6 & TNF in monocytes from HIV+ pts
IL-6
TNF
HIV- HIV+ VL<20 HIV- HIV+ VL<20 HIV- HIV+ VL<20
HIV- HIV+ VL<20 HIV- HIV+ VL<20 HIV- HIV+ VL<20
Monocyte subsets

• Case-control study in virologically suppressed patients– Cases: non-AIDS events (AMI, CVA, non-AIDS
cancers, bacterial infection, death)
– Controls: well matched (age, sex, pre-ART CD4, ART regimen)
• Blood taken for inflammatory markers – at baseline, 1 yr post ART initiation, pre-event
• Conditional logistic regression analysis
Tenorio AR et al J Infect Dis 2014 May;
Markers of inflammation and coagulation, but not T-cell activation, predict non-AIDS events

Markers of inflammation & coagulation, but not T-cell activation, predict non-AIDS events
IL6
Unadjusted <.0012.58 (1.91-3.48)
Adjusted* <.0012.48 (1.83-3.35)
IP-10
Unadjusted 0.0021.49 (1.16-1.91)
Adjusted* 0.0071.42 (1.10-1.84)
sTNFr-I
Unadjusted <.0011.99 (1.49-2.66)
Adjusted* <.0011.94 (1.45-2.60)
sTNFr-II
Unadjusted <.0011.88 (1.44-2.46)
Adjusted* <.0011.81 (1.38-2.38)
Soluble CD14
Unadjusted <.0011.74 (1.29-2.35)
Adjusted* <.0011.67 (1.23-2.27)
D-Dimer
Unadjusted <.0012.41 (1.78-3.27)
Adjusted* <.0012.38 (1.75-3.25)
CD8+ %DR+38+
Unadjusted 0.5161.06 (0.88-1.28)
Adjusted* 0.8630.98 (0.80-1.20)
0.50 1.00 4.00
Pre-event Marker P ValueOdds Ratio per 1 IQR increase
*Adjusted by CD4 c
OR at baseline for:
Death CA MI/Stroke
20.9** 3.1** 2.2**19.9** 2.9** 2.1**
1.9 1.7* 1.7*
1.8 1.5 1.7*
3.3** 2.3** 2.1**
3.3* 2.2** 2.1**
2.6** 2.1** 1.9**
2.9** 1.9* 2*
2.7* 1.5 1.7
2.8* 1.4 1.7
8.4** 3.2** 2.6**
8.1** 3.1** 2.6**
1.4 1 1
0.8 0.9 0.9
**
**
**
**
**
**
**
**
**
**
**
**
Tenorio AR et al J Infect Dis 2014 May ahead of print
Death Ca AMI/stroke Pre-event marker Odds ratio 1 IQR increase p value OR at baseline for
IL-6
IP-10
sTNFR-I
sTNFR-II
sCD14
D-dimer
CD38+DR+CD8

• Increased innate immune inflammatory markers are strongly associated with non-AIDS morbidity
• T cell activation is less related to non-AIDS morbidity
Tenorio AR et al J Infect Dis 2014 May

So where does this inflammation come from?
(HIV,co-pathogens, and…)

Early loss of CD4+ cells in gut-associated lymphoid tissue results in systemic inflammation
Guadalupe M et al J Virol 2003 77:11708; Ciccone E et al J Virol 2011;85:5880 Brenchley et al Nat Med 2006 12:1365;. Allers K et al JID 2014 209:739-48
HIV-
HIV+
•Mϕ accumulate in gut mucosa * in untreated HIV
Further drive pro-inflammatory cytokine production
• Loss of CD4+ T cells in GALT o Not in elite controllers
Starts in acute HIV infection Permanently damages gut
mucosa Microbial translocation
o Systemic inflammation Incompletely restored by ART
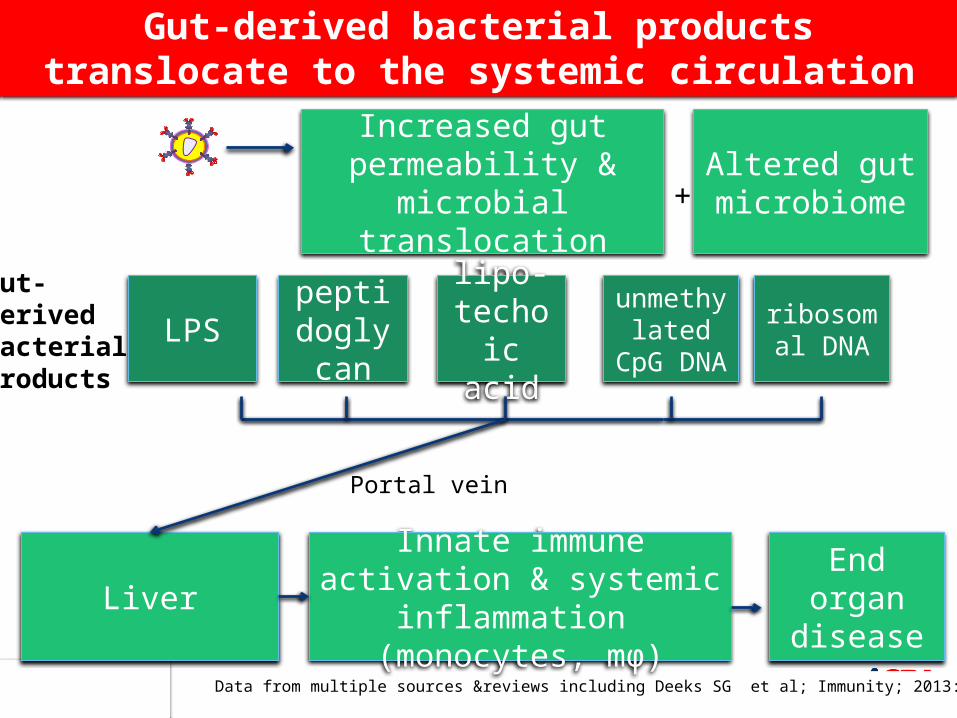
Data from multiple sources &reviews including Deeks SG et al; Immunity; 2013: 39 :633
Gut-derived bacterial products translocate to the systemic circulation
Increased gut permeability & microbial
translocation
Innate immune activation & systemic inflammation
(monocytes, mϕ)
LPS
Liver
End organ disease
ribosomal DNA
unmethylated CpG
DNA
peptidoglycan
lipo-techoic
acid
Altered gut microbiome+
Portal vein
Gut-derived bacterial products
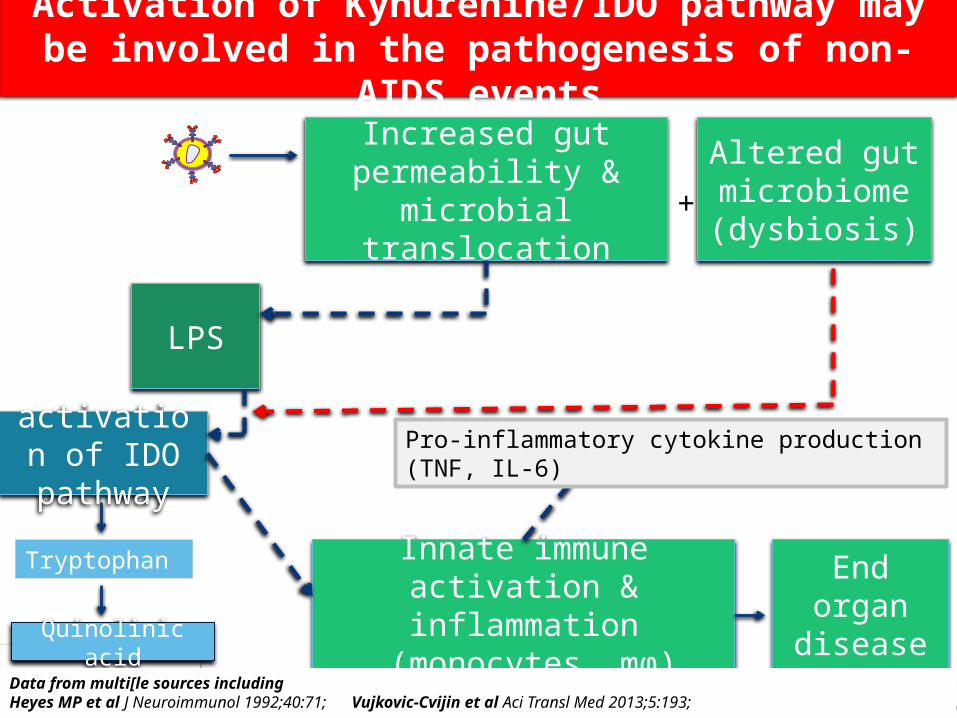
Activation of Kynurenine/IDO pathway may be involved in the pathogenesis of non-AIDS events
Increased gut permeability & microbial
translocation
Innate immune activation & inflammation
(monocytes, mϕ)
LPS
End organ disease
activation of IDO pathway
Altered gut microbiome(dysbiosis)
+
Quinolinic acid
Tryptophan
Pro-inflammatory cytokine production (TNF, IL-6)
Data from multi[le sources including Heyes MP et al J Neuroimmunol 1992;40:71; Vujkovic-Cvijin et al Aci Transl Med 2013;5:193;
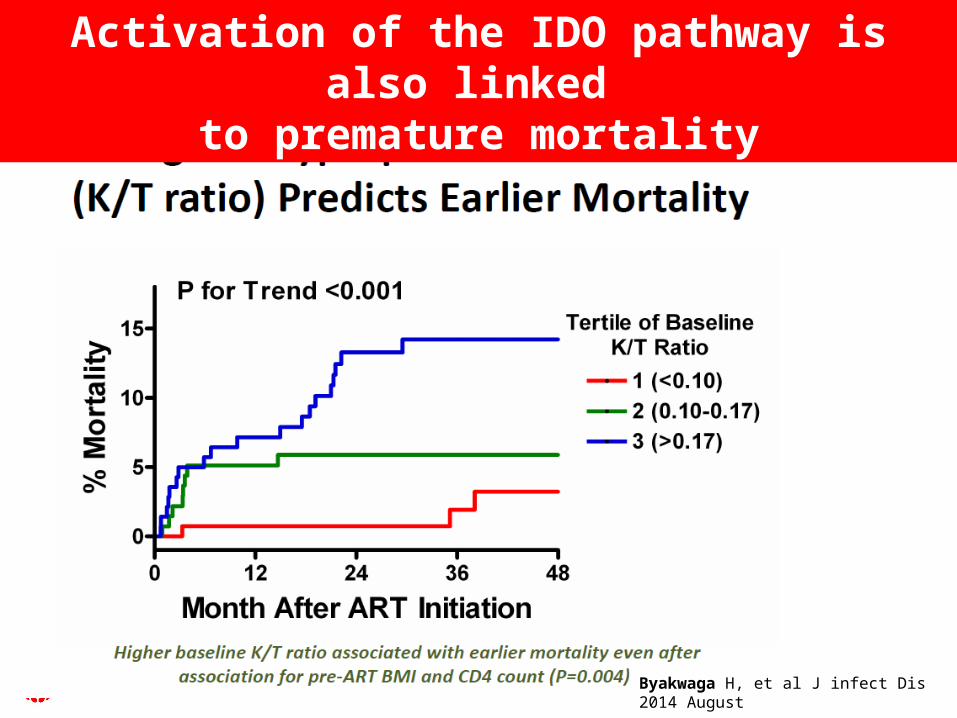
Byakwaga H, et al J infect Dis 2014 August
Activation of the IDO pathway is also linked to premature mortality

Tacke F& Zimmerman H J Hepatol 2014;60:1090; Balagopal A et al Gastroenterology 2008 135:226
Multi-pronged liver attack by HIV, co-pathogens and gut-derived bacterial products
Increased gut permeability & microbial
translocation
Activation of innate pro-inflammatory &
pro-fibrotic pathways
LPS
Kupffer mϕ& hepatic
stellate cell activation
End organ diseaseIncluding
liver fibrosis
ribosomal DNA
unmethylated CpG
DNA
peptidoglycan
lipo-techoic
acid
Portal veinLiver damage
clearanceLPS
Gut-derived bacterial products

Tacke F& Zimmerman H J Hepatol 2014;60:1090; Balagopal A et al Gastroenterology 2008 135:226
Multi-pronged liver attack via HIV, co-pathogens, gut-derived bacterial products, etc
Increased gut permeability & microbial
translocation
Activation of innate pro-inflammatory &
pro-fibrotic pathways
LPS
Kupffer mϕHepatic
stellate cellactivation
End organ diseaseeg
liver fibrosis
ribosomal DNA
unmethylated CpG
DNA
peptidoglycan
lipo-techoic
acid
Portal vein Impaired hepatic function
clearanceLPS, etc
Gut-derived bacterial products
CoinfectionEg HCV

Redd AD et al AIDS Res Hum Retroviruses 2013:29:1026
Monocyte activation is associated with liver fibrosis in HIV+ patients in Uganda
• Retrospective Case/Control study – in Rakai Uganda
• HIV+ & HIV- subjects with liver fibrosis (transient elastography >9.3kPa)
– Cases n = 133– Controls n =133
• matched for age, gender, HIV, ART
– Less than 5% infected with HBV

Patients with monocyte activation had higher odds of liver fibrosis
Soluble CD14 (ng/ml)
Overall population 1,770 (+/- 1163)
Cases (>9.3kPa) 2,029 (+/- 1320)
Controls (<9.3 kPa) 113 (+/-77.8)
HIV+ 1,840 (+/- 1369)
HIV- 1,682 (+/- 793)
• Higher sCD14 levels were associated with – 19% increased odds of having liver fibrosis (adj OR 1.19 p 0.002)
• In HIV+ higher sCD14 was associated with– 54% increased odds of having liver fibrosis (adj OR 1.54, p <0.001)
Redd AD et al AIDS Res Hum Retroviruses 2013:29:1026

How do monocytes contributeto the development of cardiovascular disease in HIV+?

Contributing factors to CVD in HIV+ patients
Traditional risk factors
cART toxicityCo-infection with eg CMV
Cardiovascular disease
monocyte & mϕactivation
chronic inflammation
other pro-inflammatory & pro-coagulant pathways

Role of monocytes in atheromatous plaque development
HIV activates monocytes & endothelial cells (in conjunction with proatherogenic lipids), Increase monocyte transmigration Increase uptake of oxLDL Promote differentiation into foam cells And contribute to atherosclerotic plaque formation Campbell J et al AIDS 2014 in press
Slide produced by G Martin
Adhesion m’cules

HIV promotes monocyte foam cell formation in vitro
• Sub-endothelial monocytes take up ox LDL via CD36 and develop into foam cells after migrating across TNF-activated endothelial cells1
– Endothelial activation is critical for this process1
– Foam cell formation by activated monocytes from HIV+2
1. Westhorpe C et al Exp Mol Pathol 2012;93:2202. Maisa A et al submitted 2014
monocyte macrophage Foam cell

How does monocyte metabolism augmentinflammationin HIV+ patients?

Glucose metabolism in monocytes
• Glucose is important substrate for ATP production
• Glut1 is major glucose transporter
• Activated monocytes dramatically increase Glut1 expression & glucose uptake
• Change from oxidative to glycolytic metabolism
Palmer CS & Crowe SM AIDS Res Human Retro 2014 30:335
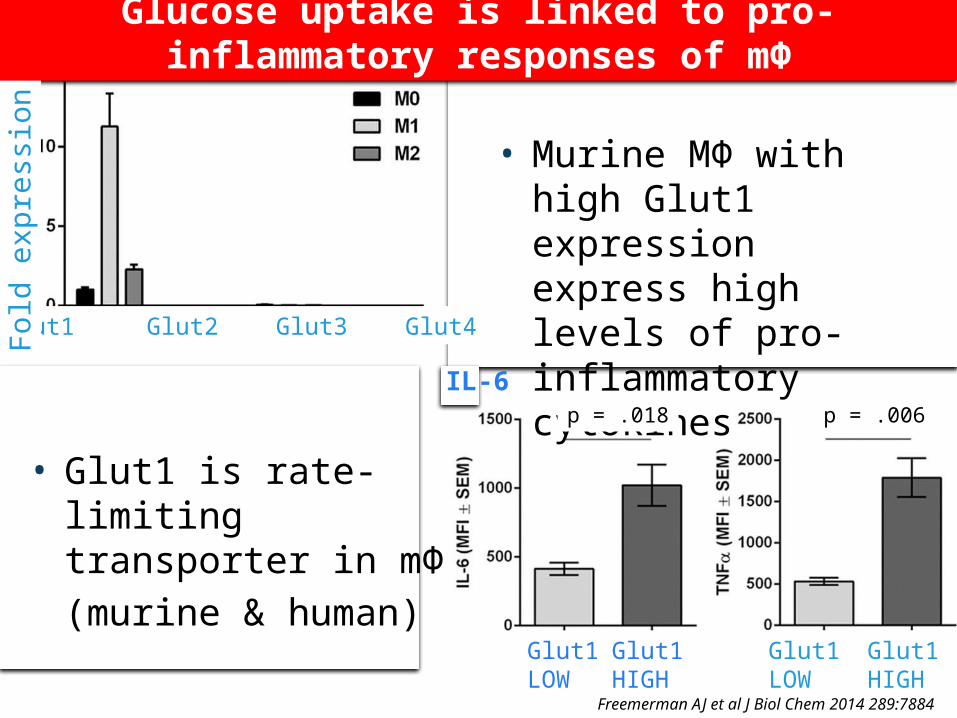
• Glut1 is rate-limiting transporter in mΦ
• (murine & human)
• Murine MΦ with high Glut1 expression express high levels of pro-inflammatory cytokines
Freemerman AJ et al J Biol Chem 2014 289:7884
IL-6 TNF
Glut1LOW
Glut1HIGH
Glut1HIGH
Glut1LOW
Glut1 Glut2 Glut3 Glut4
p = .018 p = .006
Fol
d ex
pres
sion
Glucose uptake is linked to pro-inflammatory
responses of mΦ

%G
lut1
+ c
ell
s o
nm
on
oc
yte
su
bp
op
ula
tio
ns
NC I C NC I C NC I C0
20
40
60
80
100
HIV- HIV+/naïve HIV+/cARTn= 15 14 8
p= 0.047p=0.021
p=0.018p=0.002
HIV increases glucose metabolic activity in monocytes, linked to inflammation
Palmer C , Anzinger J et al submitted, 2014See POSTER MOPE-006 & TODAY WEPE-009
• The increased Glut1 expression is on intermediate and non-classical (NC) monocytes in HIV+• Not restored by ART
%C
D14
+G
lut1
+
HIV-
HIV+/
naïve
HIV+/
cART
0
5
10
15
2030405060
p=0.005 p= 0.48
p=0.008
n= 16 17 11
% G
lut1
+ m
onoc
ytes
% G
lut1
+ m
onoc
ytes
NC I C NC I C NC I C HIV- HIV+ naïve HIV+cART+
HIV- HIV+ naïve HIV+cART+
N = 18 17 11
• Increase in proportion of Glut1+ monocytes in HIV+• Not restored by ART
• Correlates with monocyte activation & D-dimer expression

Does prematureInnate immune senescence contribute topremature risk of non-AIDS morbidities ?

Ageing HIV
CVD, cancers, kidney, bone & liver disease,
neurocognitive impairment
:
Shared immunologic features between HIV infection & ageing
Immune senescence
Chronic immune activation/inflamm
ation
Immunologic changes
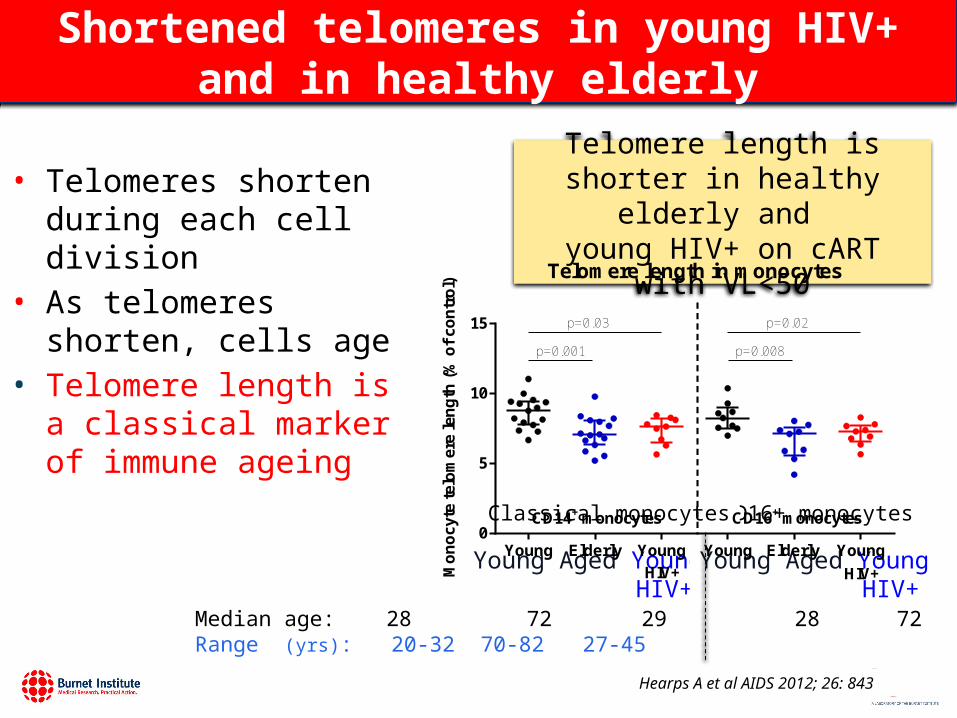
Shortened telomeres in young HIV+ and in healthy elderly
Hearps A et al AIDS 2012; 26: 843
• Telomeres shorten during each cell division
• As telomeres shorten, cells age
• Telomere length is a classical marker of immune ageing
Median age: 28 72 29 28 72 29 yrsRange (yrs): 20-32 70-82 27-45
Telomere length in monocytes
Young Elderly Young Young Elderly Young0
5
10
15
CD14+ monocytes CD16+ monocytes
HIV+ HIV+
p=0.001
p=0.03
p=0.008
p=0.02
Mo
no
cyte
tel
om
ere
len
gth
(%
of
con
tro
l)
Telomere length is shorter in healthy elderly and
young HIV+ on cART with VL<50
Young Aged Young HIV+
Young Aged Young HIV+
CD16+ monocytesClassical monocytes

HIV accelerates innate immune changes
• HIV induces age-related changes in monocytes– In HIV+ men1
– In HIV+ women2
– Changes appear approx 10-14 years earlier in HIV+ compared with HIV- women2.
• Does the clinical phenotype of ageing emerge earlier in HIV+ patients with innate senescence?– No clear evidence
Hearps A et al AIDS. 2012;24:843-53Martin G et al 2013;8(1):e55279;
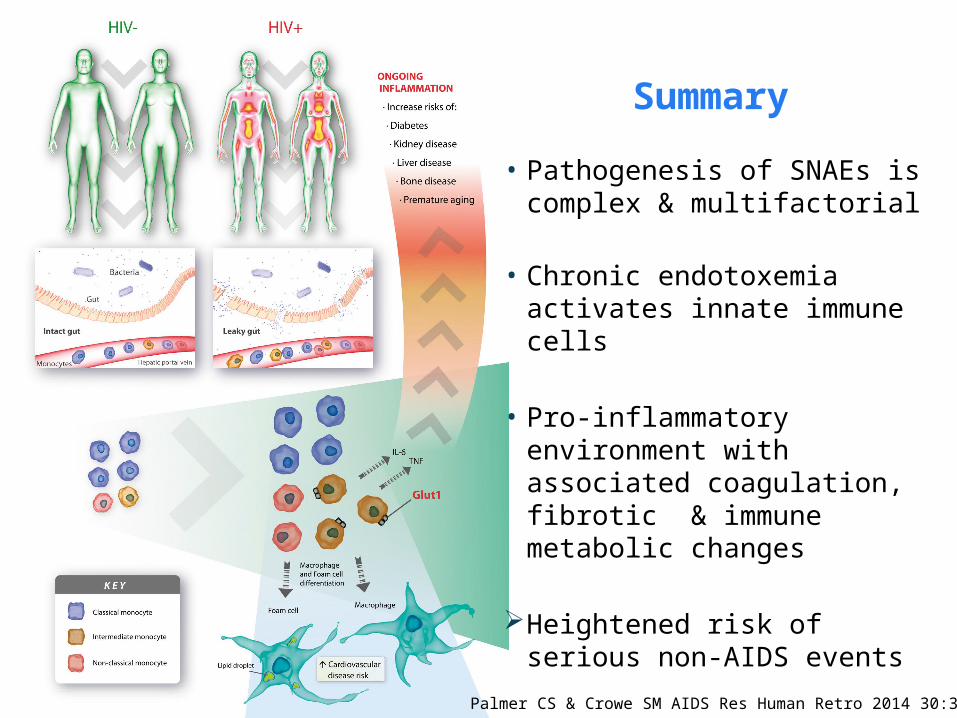
Palmer CS & Crowe SM AIDS Res Human Retro 2014 30:335
• Pathogenesis of SNAEs is complex & multifactorial
• Chronic endotoxemia activates innate immune cells
• Pro-inflammatory environment with associated coagulation, fibrotic & immune metabolic changes
Heightened risk of serious non-AIDS events
Summary

Conclusions: Strong role of innate immune pathways in pathogenesis of Serious Non-AIDS Events
• Most innate immune changes in HIV+ only partly reversed by ART– Residual immune dysregulation syndrome – Persistent immune activation, inflammation & coagulation
• Circulating biomarkers of inflammation are strongest predictors of non-AIDS events
• Less evidence for T cell involvement
• Innate immune senescence occurs earlier in HIV+– Not clear whether this translates into premature risk of
SNAEs

Anthony JaworowskiAnna HearpsGenevieve MartinClovis PalmerTom AngelovichAnna MaisaGregor LichtfussWan-Jung ChengJingling ZhouTim SpelmanM Gouillou
Infectious DiseasesSharon LewinJenny HoyJulian ElliottKate CherryJanine Trevillyon
Cardiovascular MedicineAnthony DartLiz DewarSofie Karapanagiotidis
Dmitri Sviridov
Alan Landay
The HaCH Study volunteers
Acknowledgements
William Muller
Funding
Univ West indies, Jamaica
Josh Anzinger
Clare Westhorpe
“The Ageing Team” Also with grateful thanks to colleagues who contributed slides for this presentation, some
not included because of time limitations
Mike McCune






























