PLEASE SCROLL DOWN FOR ARTICLE
This article was downloaded by: [Karolinska Institute]On: 12 December 2008Access details: Access Details: [subscription number 779857372]Publisher Informa HealthcareInforma Ltd Registered in England and Wales Registered Number: 1072954 Registered office: Mortimer House,37-41 Mortimer Street, London W1T 3JH, UK
StressPublication details, including instructions for authors and subscription information:http://www.informaworld.com/smpp/title~content=t713652982
Stress in cynomolgus monkeys (Macaca fascicularis) subjected to long-distance transport and simulated transport housing conditionsA. L. Fernström ab; W. Sutian ac; F. Royo a; K. Westlund d; T. Nilsson a; H. -E. Carlsson a; Y. Paramastri c; J.Pamungkas c; D. Sajuthi c; S. J. Schapiro e; J. Hau ab
a Department of Neuroscience, Comparative Medicine, Uppsala University, Uppsala, Sweden b Department ofExperimental Medicine, University of Copenhagen and State Hospital, The Panum Institute, 2200 NCopenhagen, Denmark c Primate Research Centre, Bogor Agricultural University, Bogor, 16151, Indonesia d
The Swedish Institute for Infectious Disease Control, Stockholm, Sweden e The University of Texas M. D.Anderson Cancer Center, Bastrop, TX, USA
First Published:2008
To cite this Article Fernström, A. L., Sutian, W., Royo, F., Westlund, K., Nilsson, T., Carlsson, H. -E., Paramastri, Y., Pamungkas, J.,Sajuthi, D., Schapiro, S. J. and Hau, J.(2008)'Stress in cynomolgus monkeys (Macaca fascicularis) subjected to long-distancetransport and simulated transport housing conditions',Stress,11:6,467 — 476
To link to this Article: DOI: 10.1080/10253890801903359
URL: http://dx.doi.org/10.1080/10253890801903359
Full terms and conditions of use: http://www.informaworld.com/terms-and-conditions-of-access.pdf
This article may be used for research, teaching and private study purposes. Any substantial orsystematic reproduction, re-distribution, re-selling, loan or sub-licensing, systematic supply ordistribution in any form to anyone is expressly forbidden.
The publisher does not give any warranty express or implied or make any representation that the contentswill be complete or accurate or up to date. The accuracy of any instructions, formulae and drug dosesshould be independently verified with primary sources. The publisher shall not be liable for any loss,actions, claims, proceedings, demand or costs or damages whatsoever or howsoever caused arising directlyor indirectly in connection with or arising out of the use of this material.

ORIGINAL RESEARCH REPORT
Stress in cynomolgus monkeys (Macaca fascicularis) subjectedto long-distance transport and simulated transport housing conditions
A. L. FERNSTROM1,5, W. SUTIAN1,3, F. ROYO1,†, K. WESTLUND2, T. NILSSON1,
H.-E. CARLSSON1, Y. PARAMASTRI3, J. PAMUNGKAS3, D. SAJUTHI3, S. J. SCHAPIRO4, &
J. HAU1,5
1Department of Neuroscience, Comparative Medicine, Uppsala University, BMC Box 572, 75123 Uppsala, Sweden,2The Swedish Institute for Infectious Disease Control, Stockholm, Sweden, 3Primate Research Centre, Bogor Agricultural
University, Jalan Lodaya II/5, Bogor, 16151, Indonesia, 4The University of Texas M. D. Anderson Cancer Center, Bastrop,
TX USA, and 5Department of Experimental Medicine, University of Copenhagen and State Hospital, The Panum Institute,
Blegdamsvej 3B, 2200 N Copenhagen, Denmark
(Received 24 July 2007; revised form 27 December 2007; accepted 8 January 2008)
AbstractThe stress associated with transportation of non-human primates used in scientific research is an important but almostunexplored part of laboratory animal husbandry. The procedures and routines concerning transport are not only importantfor the animals’ physical health but also for their mental health as well. The transport stress in cynomolgus monkeys (Macacafascicularis) was studied in two experiments. In Experiment 1, 25 adult female cynomolgus monkeys were divided into fivegroups of five animals each that received different diets during the transport phase of the experiment. All animals weretransported in conventional single animal transport cages with no visual or tactile contact with conspecifics. The animals weretransported by lorry for 24 h at ambient temperatures ranging between 208C and 358C. Urine produced before, during andafter transport was collected and analysed for cortisol by enzyme-linked immunosorbent assay (ELISA). All monkeysexhibited a significant increase in cortisol excretion per time unit during the transport and on the first day following transport.Although anecdotal reports concerning diet during transport, including the provision of fruits and/or a tranquiliser, wasthought likely to influence stress responses, these were not corrobated by the present study. In Experiment 2, behavioural datawere collected from 18 cynomolgus macaques before and after transfer from group cages to either single or pair housing, andalso before and after a simulated transport, in which the animals were housed in transport cages. The single housed monkeyswere confined to single transport cages and the pair housed monkeys were kept in their pairs in double size cages. Both pairhoused and singly housed monkeys showed clear behavioural signs of stress soon after their transfer out of their group cages.However, stress-associated behaviours were more prevalent in singly housed animals than in pair housed animals, and thesebehaviours persisted for a longer time after the simulated transport housing event than in the pair housed monkeys. Our dataconfirm that the transport of cynomolgus monkeys is stressful and suggest that it would be beneficial for the cynomolgusmonkeys to be housed and transported in compatible pairs from the time they leave their group cages at the source countrybreeding facility until they arrive at their final laboratory destination in the country of use.
Keywords: Behaviour, macaque, pair-housing, primate, urinary cortisol, welfare
Introduction
There is a strong scientific case for maintaining work on
non-human primates (NHPs) as models in carefully
selected research problems (Weatherall 2006), but
there is a critical shortage of captive-bred monkeys, in
particular Old World species, for use in biomedical
research (Cohen 2000; National Research Council
2003; Hau and Schapiro 2006). The present
inadequate supply of captive-bred NHPs in the USA
ISSN 1025-3890 print/ISSN 1607-8888 online q 2008 Informa USA, Inc.
DOI: 10.1080/10253890801903359
†Present address: Unit of Cytogenomics, CICBIOGUNE, Bizkaia Science Park, Derio, Spain
Correspondence: J. Hau, Department of Experimental Medicine, University of Copenhagen and State Hospital, 3 Blegdamsvej,2200 N Copenhagen, Denmark. Tel. þ45 35 32 73 63. Fax. þ45 35 32 73 99. Mobile: þ45 28 75 73 63. E-mail: [email protected]
Stress, November 2008; 11(6): 467–476
Downloaded By: [Karolinska Institute] At: 11:49 12 December 2008

and Europe necessitates the import of NHPs from
source countries in Asia and Africa. The use of primates
in scientific research is not expected to decrease
considerably in the coming years and the growing
demand in the USA and Europe will become less and
less attainable by production from domestic sources
(Carlsson et al. 2004; Weatherall 2006). As mentioned
in Weatherall (2006) report and recommended by Hau
and Schapiro (2006) establishment of accredited and
expert staffed NHP research laboratories in source
countries would benefit animal welfare and global
research. This is, however, a slow process and long
distance transportation of primates will thus continue,
and is associated with a number of animal welfare issues
(Honess et al. 2004; Guidelines for the Humane
Transportation of Research Animals 2006).
The typical transport event for a research NHP
approximates the following pattern. Shortly before
shipment, animals are removed from their social
groups (usually breeding groups) and placed in more
typical laboratory housing (single caging or perhaps
pair caging) for a period of pre-shipment monitoring
and conditioning. Following this period of pre-
shipping conditioning, the primates are normally
transported singly in small cages. Transport is
logistically complex, and the animals may travel
and/or wait for many hours under varying climatic
conditions. When the NHPs finally arrive at the
destination facility in a new country, they are again
quarantined – often in single cages. Long distance
transportation across multiple climate and time zones
is likely to induce substantial stress in the transported
animals (Wolfensohn 1997; Hau and Schapiro 2004;
Honess et al. 2004; Prescott and Jennings 2004).
While the effects of transport on farm animals have
been reasonably well investigated (Guide for Livestock
Exporters 1997), there have been few systematic
assessments of the impact of transportation on NHPs
(Wolfensohn 1997), although empirically derived
guidelines for the acquisition, care and breeding of
NHPs have been published by International Primato-
logical Society (IPS 2007). Monkeys caught from the
wild and confined to caging exhibit pronounced stress
(Moinde et al. 2004; Suleman et al. 2004), and
haematological parameters do not settle at new stable
values until after 6 months (Kagira et al. 2007).
However, little empirical data are available that identify
specific stress effects of transport in the literature,
although two recent publications have quantified
acclimation effects in NHPs brought into the
laboratory. Kim et al. (2005) have shown that
cynomolgus monkeys exhibited an increase in the
neutrophil-to-lymphocyte (N/L) ratio upon arrival
at their new laboratory. This response normalised a
week after arrival, suggesting a stress response to the
transportation. Similarly, Capitanio et al. (2006)
recently reviewed the effects of housing changes on a
variety of physiological parameters, and suggested that
captive NHPs may take up to 90 days to adapt to new
housing conditions, depending on the level of change
they experienced.
Provision of natural food items allows the animals to
focus on processing these and is generally rec-
ommended as an enhancement of environmental
complexity (Reinhardt 2002). Boinski et al. (1999)
demonstrated that access to a variety of foraging
enrichment positively affects behavioural and physio-
logical responses to stress and enhances psychological
well-being in singly housed brown capuchins. Conse-
quently, it may be possible to lower the stress
perception of transport in single housed monkeys by
providing them with different fruits and vegetables
during their journey.
It is well recognised that membership in a
compatible group of conspecifics provides a sense of
security for NHPs (European Council 2002). Social
living also provides opportunities for a wide range of
positive species-specific social activities (Jolly 1985;
Cheney et al. 1987). For instance, it has been
demonstrated that social grooming has a relaxing
effect on the animal receiving grooming, lowering its
heart rate (Boccia et al. 1989). Similarly, cage mates
seem to serve as a buffer against stress in NHPs
subjected to experimental procedures (Mason 1960;
Coe and Franklin 1982; Coelho et al. 1991). It is thus
likely that pair housing during shipment of primates
may reduce some of the stress-related responses
commonly observed during and after transportation.
Cynomolgus monkeys constitute one of the most
important NHP species in biomedical research – 54%
of all NHPs used in the UK in the mid-1990s (Owen
et al. 1997) – and the use of this species is increasing
(Carlsson et al. 2004). Cynomolgus monkeys seem,
however, to exhibit more pronounced stress responses
to confinement in transport cages than do rhesus
monkeys (Macaca mulatta) or bonnet macaques
(Macaca radiata) (Clarke et al. 1988). The present
experiments were designed to quantify physiological
stress responses to transport and to assess the effects of
one manipulation (simulated transport housing in
single vs. pair enclosures) on behavioural measures
potentially indicative of stress. In both experiments,
the duration of the transport episode was chosen to
represent a typical transport scenario from South East
Asia to Europe or the USA.
The aims were addressed through two experiments
to (i) examine the urinary cortisol excretion as a
measure of stress in cynomolgus monkeys subjected
to standard transport conditions in standard single
cages and (ii) to analyse the behavioural response
A. L. Fernstrom et al.468
Downloaded By: [Karolinska Institute] At: 11:49 12 December 2008

of cynomolgus monkeys to transfer to single and pair
caging from group caging, and whether animals who
were subjected to a quarantine period and simulated
transport housing conditions in pairs behaved differ-
ently from singly housed animals subjected to the same
procedures.
Materials and methods
Both of the experiments described in this paper were
conducted at the quarantine facility of the Primate
Research Center (PRC) of Bogor Agricultural Univer-
sity, Java, Indonesia.
Experiment 1
Twenty-five captive bred (captive bred parents) adult
(6–7 years old) female cynomolgus monkeys (Macaca
fascicularis) obtained from New Inquatex (Rumpin,
Indonesia) were randomly divided into five groups of
five individuals each (R0–R4). Except during trans-
port, all animals were kept indoors in single cages
under quarantine conditions under ambient con-
ditions with approximately 12 h of daylight and 12 h of
darkness for 90 days to allow them to habituate to
single housing. The cages (w: 0.39 m, l: 0.39 m and h:
0.75 m), were made of stainless steel, and were located
on an upper or a lower tier of a two-tiered caging
system. All subjects had visual, auditory, and olfactory
contact with the other monkeys in the same room. The
cages were cleaned by hosing the tray under the cage
twice daily at 8 a.m. and 1 p.m. Fresh water bottles
were supplied at both cleaning times.
During transport, the temperature ranged from
258C to 358C and humidity ranged from 75% to 85%.
Although UK and USA recommendations advocate a
temperature range between 158C and 258C, the
transport in source countries is often performed in
lorries with no means of climate control. All groups of
animals were fed a commercial diet (Monkey chow,
Charogen, Phokphan, Thailand) supplemented with
fruit (banana and guava) and water was available ad
libitum. For the purposes of this study, the maintenance
diet is defined as the combination of the commercial
monkey chow, banana and guava. The space under the
wire mesh bottoms of the cages was modified to
separate faeces and urine from the individual animals.
During that time, they were fed the maintenance diet,
and water was available ad libitum. Two days prior to
transport, each group of monkeys was adapted to the
diet they were to be fed during transport:
Group R0 received only monkey chow.
Group R1 received the maintenance diet: monkey
chow, banana and guava.
Group R2 received the maintenance diet (monkey
chow, banana and guava) plus multi-vitamins.
Group R3 received sugar cane, banana, guava, apple
and orange.
Group R4 received the maintenance diet plus multi-
vitamins and a tranquilizer (Acepromazine Maleat
Injectione, 0.4 mg/kg body weight, intra-muscular
injection (Boehringer Ingelheim Vetmedica, Inc.,
St Joseph, MO, USA).
The monkeys were subjected to transport by lorry for
24 h. They were transported in standard transport
cages (IATA, 2007; w: 0.24 m, l: 0.4 m and h: 0.5 m)
made of plywood (Figure 1). After transportation, the
monkeys were returned to their home indoor single
cages under quarantine conditions, fed the mainten-
ance diet and provided with water ad libitum. Urine
was collected at 8 h intervals, from 16 h before
transport started, during the 24-h transport, and for
32 h after transport ended. The samples were stored
frozen at 2208C until analysed.
Quantification of cortisol. Urine samples were thawed,
centrifuged (1000g for 2 min) and the supernatants
were analysed without extraction. Cortisol was
measured with a pan-specific cortisol ELISA kit
(DRG Diagnostics, Marburg, Germany). The inter-
and intra-assay coefficients of variation were 9.3% and
1.9%, respectively.
Experiment 2
Subjects included 18 female cynomolgus monkeys from
the semi-natural breeding facility on Tinjil Island,
Indonesia (Kyes 1998). Prior to the project, the subjects
were housed at the PRC for 2 months in 32 m2
Figure 1. A typical transport box consisting of two single cage
units. The centre wall of the unit was removed to accommodate pair
housed monkeys.
Transport stress in cynomolgus monkeys 469
Downloaded By: [Karolinska Institute] At: 11:49 12 December 2008

group cages together with other (10–20) cynomolgus
monkeys. The subjects’ ages ranged from 2.5 to 5.5
years and body weights ranged from 1.6 to 2.1 kg.
The experimentbeganwhenthemonkeyswere removed
from their group cages and placed in either single or pair
cages in the indoor quarantine room. Room tempera-
ture and relative humidity varied between 258C and
318C and between 75% and 85%, respectively.
The quarantine room had no windows and the lights
were on daily between 8 a.m. and 5 p.m. The project
consisted of three phases. Ethological observations were
performed only during phases 1 and 3.
Phase 1: The first 21 days after the monkeys were
transferred from group cages into the PRC’s indoor
quarantine facility.
Phase 2: The transport housing simulation (48 h).
Phase 3: The first 21 days after the monkeys were
returned to their pair or single cages after the
transport housing simulation.
Phase 1 (days 1–21). On day 1, subjects were
transferred from a group cage to a room in the
quarantine facility. Ten monkeys were selected at
random to live in single cages and 10 monkeys were
selected to live in pairs (due to a computer breakdown,
the data from two of the pair-housed monkeys were
lost). Both single and pair cages were of the same size
(w: 0.39 m, l: 0.39 m and h: 0.75 m), were made of
stainless steel, and were located on an upper or a lower
tier of a two-tiered caging system. All subjects had
visual, auditory and olfactory contact with the other
monkeys in the same room. The cages were cleaned by
hosing the tray under the cage twice daily at 8 a.m. and
1 p.m. The monkeys were fed after every cleaning
episode. Monkey chow (Charoen Phokphan, Bankok,
Thailand) was provided in the morning and fresh
fruits or vegetables were given in the afternoon. Fresh
water bottles were supplied at both cleaning times.
Phase 2 (days 22 and 23). A partial transport
simulation was performed by transferring the
subjects to transport cages within the facility,
avoiding potentially confounding effects associated
with real transportation (e.g. noise, vibrations,
temperature changes, odours and air pressure
changes). The transport boxes used for single caged
monkeys were standard transport boxes (Figure 1).
Single transport boxes were modified into pair
transport boxes by simply removing the central wall
between adjoining compartments to accommodate
two monkeys. The single housed monkeys were placed
in single transport boxes and the pair housed animals
were placed in pair transport boxes. The boxes were
put in a separate section of the facility for 48 h without
any contact with other animals. The single and pair
housed monkeys were separated into different, but
identical rooms during this phase of the study.
The monkeys were given water, fruits and vegetables
in the transport boxes at 9 a.m. at the start of the
simulation. Fresh water was provided daily at the usual
times (8 a.m. and 1 p.m.). Some fruit and vegetable
material was still present in the animals’ compartments
at the end of the transport simulation. Since the
monkeys were located in the smaller closed transport
boxes during this period and since we were attempting
to simulate transport conditions, no ethological
observations were conducted during Phase 2.
Phase 3 (days 24–44). After the 48-h transport
housing simulation, the monkeys were returned to
their original positions and social conditions in their
original home cages in the quarantine room and
observed for 21 days.
Behavioural observations
The ethogramme used (Table I) was derived from an
ethogramme designed to detect and monitor stress-
related behaviours in NHPs (Schapiro and Bloom-
smith 1994). An instantaneous time-sampling method
(Altmann 1974) was used to record appetitive,
affiliative and other active and inactive behaviours.
In an attempt to economically, yet representatively,
the sample activity from all of the subjects, behaviours
were recorded at 10 s intervals for 8 min daily
for each monkey. Additionally, since this technique
Table I. The ethogramme used in the study (modified from
Schapiro and Bloomsmith 1994).
Behavioural
categories Definition
Appetitive behaviours Eat, forage and drink
Affiliative behaviours Social groom, given and received
Autogroom Grooming own body
Locomotion Moving the whole body from one spot
to another
Rest/inactivity Sitting still with eyes not focusing
on object or other animal
Watch /exploratory
behaviours
Investigating object or other monkey
Abnormal behaviours Head toss, self plucking, self aggression,
faeces eating, and pacing
Aggressive behaviours Facial threat, approach and physical
aggression
Other behaviours Behaviours other than those described
above
A. L. Fernstrom et al.470
Downloaded By: [Karolinska Institute] At: 11:49 12 December 2008

over-represents states and under-represents events,
important events such as aggressive and abnormal
behaviours, were recorded using 1/0 sampling
(Altmann 1974). All observations were made by one
observer (ALF), and conducted between 8 a.m. and 3
p.m. and subjects for observation were randomised on
the first day (using Randomizer available at www.ran-
domizer.org), after which the order of observation
remained unchanged throughout the study. However,
observations for all subjects were evenly distributed
across time intervals during the daily observation
period. The monkeys were observed for a total of
101 h in the two phases (1 and 3).
Statistics
In Experiment 1, statistical analyses were performed
using SPSS v 12.0.1. Effect of transportation and diet
were tested by a mixed-model ANOVA with diet as a
between-subjects factor and time point (9 over the
3 days) as a within-subjects factor. The raw data were
not normally distributed and were subjected to a
square root transformation, normalising the data, for
analysis.
In Experiment 2, the data presented in results part
A (means of behavioural responses to caging and
transport simulation) were analysed with Fisher’s
exact test. The data presented in results part B
(adaptation to single and pair caging and effects of
transport housing simulation) were not normally
distributed and were analysed using Kruskal–Wallis
tests. P-values , 0.05 were considered significant for
all analyses.
Ethical approval
The protocols of the present studies were approved by
the Institutional Animal Care and Use Committee of
Bogor Agricultural University, Java, Indonesia.
Results
Experiment 1
There was a main effect of time (before vs. during vs.
after) on urinary cortisol concentration that increased
in all monkeys during and after transportation
(P ¼ 0.002) (Figure 2). Using polynomial contrast
analyses to compare groups R0, R1, R2, and R3 with
group R4, the last group had a significantly lower
concentration of cortisol in urine than the monkeys of
groups 1, 2, and 3 (P , 0.001). However, when
corrected for variation in urine volume voided, no
significant differences were found.
Experiment 2
Behavioural responses to caging and transport simulation
Appetitive behaviours. The singly housed monkeys
consistently spent more time engaged in appetitive
behaviours than did the pair housed animals during
both Phases 1 and 3 (Fishers test, two-tailed: P-values
,0.001). Little time was allocated to appetitive
behaviours on the first day of Phase 1. The monkeys
appeared to spend an increasing proportion of time in
appetitive activities throughout the entire observation
period.
Allogrooming. The pair housed monkeys spent on
average 1% of their time allogrooming during the
observation period. There was no adaptation effect
over-time during Phases 1 and 3 was no effect of the
transport housing simulation on allogrooming activity.
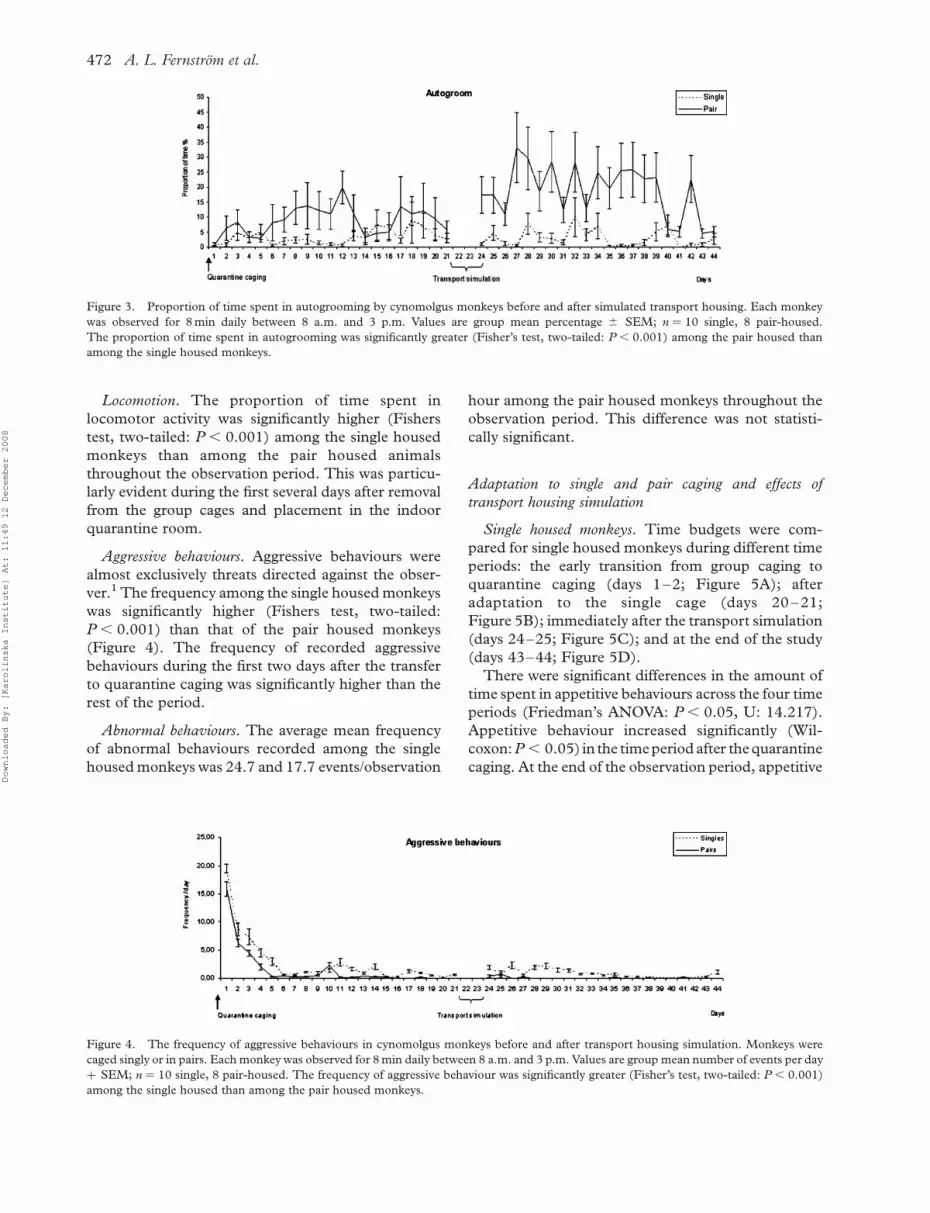
Autogrooming. The proportion of time spent in
autogrooming was significantly greater (Fishers test,
two-tailed: P , 0.001) among the pair housed
monkeys compared with that of the single housed
animals (Figure 3).
Watch/exploratory behaviour. The pair housed mon-
keys spent on average 58%, and the single housed
monkeys 68%, of their time engaged in exploratory/-
watch activity during the observation period. This
difference was not significant.
Figure 2. Urinary concentrations of cortisol in cynomolgus
monkeys fed different diets before, during and after transport by
lorry for 24 h. The transport period is represented as a solid line on
the x-axis. The diets were started 48 h before transport and
maintained until the end of the 24-h transport: group R0 received
only monkey chow; group R1 received the maintenance diet:
monkey chow, banana and guava; group R2 received the
maintenance diet (monkey chow, banana and guava) plus multi-
vitamins; group R3 received sugar cane, banana, guava, apple and
orange and group R4 received the maintenance diet plus multi-
vitamins and a tranquiliser. n ¼ 5 per group. Values are group means
^ SEM. Urinary cortisol concentration increased in all groups
during and after transportation (P ¼ 0.002).
Transport stress in cynomolgus monkeys 471
Downloaded By: [Karolinska Institute] At: 11:49 12 December 2008
Locomotion. The proportion of time spent in
locomotor activity was significantly higher (Fishers
test, two-tailed: P , 0.001) among the single housed
monkeys than among the pair housed animals
throughout the observation period. This was particu-
larly evident during the first several days after removal
from the group cages and placement in the indoor
quarantine room.
Aggressive behaviours. Aggressive behaviours were
almost exclusively threats directed against the obser-
ver.1 The frequency among the single housed monkeys
was significantly higher (Fishers test, two-tailed:
P , 0.001) than that of the pair housed monkeys
(Figure 4). The frequency of recorded aggressive
behaviours during the first two days after the transfer
to quarantine caging was significantly higher than the
rest of the period.
Abnormal behaviours. The average mean frequency
of abnormal behaviours recorded among the single
housed monkeys was 24.7 and 17.7 events/observation
hour among the pair housed monkeys throughout the
observation period. This difference was not statisti-
cally significant.
Adaptation to single and pair caging and effects of
transport housing simulation
Single housed monkeys. Time budgets were com-
pared for single housed monkeys during different time
periods: the early transition from group caging to
quarantine caging (days 1–2; Figure 5A); after
adaptation to the single cage (days 20–21;
Figure 5B); immediately after the transport simulation
(days 24–25; Figure 5C); and at the end of the study
(days 43–44; Figure 5D).
There were significant differences in the amount of
time spent in appetitive behaviours across the four time
periods (Friedman’s ANOVA: P , 0.05, U: 14.217).
Appetitive behaviour increased significantly (Wil-
coxon:P , 0.05) in the timeperiod after thequarantine
caging. At the end of the observation period, appetitive
Figure 3. Proportion of time spent in autogrooming by cynomolgus monkeys before and after simulated transport housing. Each monkey
was observed for 8 min daily between 8 a.m. and 3 p.m. Values are group mean percentage ^ SEM; n ¼ 10 single, 8 pair-housed.
The proportion of time spent in autogrooming was significantly greater (Fisher’s test, two-tailed: P , 0.001) among the pair housed than
among the single housed monkeys.
Figure 4. The frequency of aggressive behaviours in cynomolgus monkeys before and after transport housing simulation. Monkeys were
caged singly or in pairs. Each monkey was observed for 8 min daily between 8 a.m. and 3 p.m. Values are group mean number of events per day
þ SEM; n ¼ 10 single, 8 pair-housed. The frequency of aggressive behaviour was significantly greater (Fisher’s test, two-tailed: P , 0.001)
among the single housed than among the pair housed monkeys.
A. L. Fernstrom et al.472
Downloaded By: [Karolinska Institute] At: 11:49 12 December 2008

behaviours had increased significantly compared with
the time just after arrival (Wilcoxon: P , 0.05) in
quarantine and the time just after the transport housing
simulation (Wilcoxon: P , 0.05). There were signifi-
cant differences in the amount of time spent in
locomotor behaviours across the four time periods
(Friedman’s ANOVA: P , 0.05, U: 16.408). Loco-
motor behaviours decreased significantly after the
transfer to the quarantine room (Wilcoxon P , 0.05)
and increased again after the transport simulation
(Wilcoxon P , 0.05). Rest/inactive behaviours
occurred infrequently during the study.
Pair housed monkeys. Time budgets were compared
for pair housed monkeys during the different time
periods as above (Figure 6). There were significant
differences in the amount of time spent in appetitive
behaviours across the four time periods (Friedman’s
ANOVA: P , 0.001, U: 17.830). At the end of the
observation period, appetitive behaviours had
decreased significantly compared with the first two
days after arrival in the indoor pair cages (Wilcoxon
P , 0.05).
There were significant differences in the amount of
time spent in locomotor behaviours across the four
time periods (Friedman’s ANOVA: P , 0.05, U:
9.160). At the end of the observation period, the
proportion of time spent in locomotor behaviours had
decreased significantly (Wilcoxon P , 0.05).
Discussion
Experiment 1 confirmed that standard single cage
transport is associatedwithan increasedurinary cortisol
excretion in the cynomolgus monkey, indicating HPA
axis activation. These data are taken to be indicative of a
stress response in these monkeys subjected to lorry
transport for 24 h. In the present study, the addition of
fruit treats, multi-vitamins, and/or tranquilisers had no
significant effect on urinary cortisol concentration
during transport, indicating that the provision of
various fruits and vegetables probably does not affect
the monkeys’ perception of, or response to the transport
stressor.
The second part of this study (Experiment 2) was
designed to mimic the sequence of cage changes
commonly experienced by monkeys produced in
South East Asia when they are designated for use in
research, and are subsequently transported from the
breeder to a laboratory in Europe or the US. In order
to minimise confounding variables and disturbances,
urine or faeces were not collected for cortisol
measurement in this behavioural study.
An absence of appetitive activities (anorexia) is a
sign of severe distress among captive NHPs (Morton
and Griffiths 1985). The trend toward spending an
increasing proportion of time in appetitive activities
observed for all monkeys in the present study may
indicate that they were adapting successfully to the new
Figure 5. Time-budgets for behaviours before and after transport housing simulation for the single housed macaques. (A) The early
transition from group caging to quarantine caging (days 1–2). (B) After adaptation to the single cage (days 20–21). (C) Immediately after the
transport simulation (days 24–25). (D) At the end of the study (days 43–44); n ¼ 10 monkeys.
Transport stress in cynomolgus monkeys 473
Downloaded By: [Karolinska Institute] At: 11:49 12 December 2008

conditions (habituation). The levels of allogrooming
activities were low in the present study. Judge and
De Waal (1997) described how levels of all behaviours
increased – aggressive as well as affiliative – between
female rhesus monkeys when subjected to crowding,
and there was no indication in the allogrooming results
from the present study to suggest stress from pair
housing caused by space constraints. Autogrooming is
essential for the wellbeing of all non-human primates
and an animal displaying an absence of autogrooming
is a general cause of worry for animal care staff.
However, “over-grooming” may also be indicative of
distress. The low autogrooming activity of the singly
housed animals compared with the pair housed
monkeys contrasts with our previous studies (personal
observation) and may suggest that even the pair
housed animals in the present study experienced
substantial stress.
In the wild, cynomolgus monkeys have a home
range size of about 1.25 km2 and daily path length
varies greatly between 150 and 1900 m (Wheatley
1980; Rowe 1996), and locomotor activity in semi-
wild conditions and large group cages is much greater
than in small cages (personal observation). In the
present study, opportunities for expressing natural
locomotion were inhibited during the entire study due
to the small cages. However, the singly housed
monkeys spent a higher proportion of their time
engaged in locomotor behaviours than did the pair
housed monkeys.
The presence of aggressive behaviours among
captive NHPs may be a sign of stress (Heinz 1999).
The present results on aggression suggest that the pair
housed animals were less stressed than the single
housed monkeys throughout the observation period.
It is possible that the results indicate that the initial
transfer from group caging to quarantine caging was a
more stressful event than was the transfer to and
period in the transport cages. However, information
about the baseline behaviour before transfer to the
quarantine caging is not available, and a more likely
reason for the frequent aggressive bouts during the
first several days of the study was due to the presence
of the observer, to which the monkeys were not yet
accustomed. The transport housing simulation did
result in a modest increase in aggressive behaviour,
although the monkeys at that point in time had
habituated somewhat to the new cages. Among the
single housed monkeys the aggressive behaviours
remained elevated for a longer period of time
compared with those of the pair housed animals in
which aggressive behaviours decreased rapidly and
reached a baseline faster than in the single housed
animals. This seems to support the interpretation that
a companion animal functions as a buffer against
stress (Mason 1960; Coe and Franklin 1982; Coelho
et al. 1991).
There was no significant difference in the abnormal
behaviours between single housed and pair housed
monkeys, and they were present throughout the
Figure 6. Time-budgets for behaviours before and after transport housing simulation for the pair housed macaques. (A) The early transition
from group caging to quarantine caging (days 1–2). (B) After adaptation to the single cage (days 20–21). (C) Immediately after the transport
simulation (days 2425). (D) at the end of the study (days 43–44); n ¼ 8 monkeys.
A. L. Fernstrom et al.474
Downloaded By: [Karolinska Institute] At: 11:49 12 December 2008

observation period. However, the presence of abnor-
mal behaviour such as stereotypies is interpreted to
reflect a reduced level of well-being and an inadequate
environment (Maple 1979; Bayne et al. 1992; Broom
and Johnson 1993; Olfert et al. 1993; IPS 2007).
In conclusion, Experiment 1 confirmed that trans-
port procedures result in activation of the HPA axis as
measured by urinary cortisol, and that the provision of
food treats and/or a tranquiliser associated with the
transport event had no significant influence on the
monkeys’ stress responses. Experiment 2 demonstrated
that both pair housed and singly housed monkeys
showed behavioural signs that may indicate elevated
stress levels just after transfer from group cages to
single- and pair-housing in indoor rooms. However,
stress-associated behaviours were in general more
prominent in singly housed than in pair housed
monkeys, and these behaviours remained consistently
more prominent throughout the observation period.
Although transport simulation, which in this case was
limited to 48 h in quiet, unmoved transport boxes,
seems to be a stressful event, it may not be as stressful as
the initial transfer from a group cage to a smaller single
or pair cage as judged by the behaviour of the animals in
the present study. At the time of the transport
simulation, the monkeys had had three weeks to adjust
to their new caging. This suggests that it may be
valuable for formerly group housed monkeys destined
for long distance transportation to spend some time in
cages, simply for the purpose of habituation, prior to
shipment. However, it has been described that
maintaining NHPs in extended social groups until
immediately prior to transportation may lower the
overall stress (magnitude and duration) associated with
transferringagroupofmonkeys fromonesocialgroupin
the source country to the same social grouping in the
receiving laboratory (Wolfensohn and Honess 2005).
Based on most of the behavioural data in the present
study, transport simulation seemed more stressful for
monkeys kept singly in transport cages compared with
monkeys kept housed in 3-week-old pairs. Also, after
transportation, the stress-related behaviours, with the
exception of the autogrooming results, were more
prominent in the single housed than in the pair housed
monkeys. However, the transport simulation of the
present study only involved the subjects being placed in
transport cages in quiet surroundings and not subjected
to noise, movements of the cage, changes in tempera-
ture and humidity, which they would experience during
real transport. The present approach was chosen in
order to avoid an over-abundance of confounding
variables in a single study. A real transportation episode
is thus likely to be more stressful.
Primates showphysiological and behavioural distress
reactions when exposed to life-threatening situations
over which they have no control (Reinhardt 2002).
Therefore, being removed from a familiar environment
and placed in a strange one may cause distress for the
animals (Hau and Schapiro 2004). Being housed with
another or other member(s) of the same species is
recognised as the single most important enrichment
practice for NHPs (Hau and Schapiro 2004).
The present study supports the contention that single
housed monkeys exhibit more stress-related behaviour
than pair housed monkeys. Our data suggest that it
would be beneficial for cynomolgus monkeys to be
housed and transported in compatible pairs from the
time they leave the group cages at the breeder until they
arrive at their final destination in the new laboratory.
Acknowledgements
This study was generously financed by the Swedish
National Board for Laboratory Animals (CFN).
We are grateful for the help provided by Diah Ariyanti,
and the staff at the quarantine facility of Primate
Research Center of Bogor Agricultural University.
Ingegerd and Viking Olof Bjork are acknowledged for
funding parts of Anna-Linnea Fernstrom’s travel
costs.
Note
1. Only on very rare occasions were threats observed to be aimedat other animals.
References
Altmann J. 1974. Observational study of behaviour: Sampling
methods. Behaviour 91:449–459.
Bayne KAL, Hurst JK, Dexter SL. 1992. Evaluation of the
preference to and behavioral effects of an enriched environment
on male rhesus monkeys. Lab Anim Sci 42:38–45.
Boccia ML, Reite M, Laudenslager M. 1989. On the physiology of
grooming in a pigtail macaque. Physiol Beh 45:667–670.
Boinski S, Swing SP, Gross TS, Davis JK. 1999. Environmental
enrichment of brown capuchins (Cebus apella): Behavioral and
plasma and fecal cortisol measures of effectiveness. Am J
Primatol 48:49–68.
Broom DM, Johnson KG. 1993. Stress and animal welfare.
London: Chapman and Hall.
Capitanio JP, Kyes RC, Fairbanks LA. 2006. Considerations in the
selection and conditioning of Old World monkeys for laboratory
research: Animals from domestic sources. Ins Lab Anim Resour J
47:294–306.
Carlsson HE, Schapiro SJ, Farah IO, Hau J. 2004. The use of
primates in research: A global overview. Am J Primatol 63:
225–237.
Cheney DL, Seyfarth RM, Smuts BB, Wrangham RW. 1987.
The study of primate societies. In: Smuts BB, Cheney DL,
Seyfarth RM, Wrangham RW, Struhsaker TT, editors. Primate
societies. Chicago, IL: University of Chicago Press. p 1–8.
Clarke S, Mason WA, Moberg GP. 1988. Interspecific contrasts in
responses of macaques to transport cage training. Lab Anim Sci
38:305–309.
Transport stress in cynomolgus monkeys 475
Downloaded By: [Karolinska Institute] At: 11:49 12 December 2008

Coe CL, Franklin D. 1982. Hormonal responses accompanying fear
and agitation in the squirrel monkey. Physiol Beh 29:
1051–1057.
Coelho AM, Carey KD, Shade RE. 1991. Assessing the effects of
social environment on blood pressure and heart rates of baboons.
Am J Primatol 23:257–267.
Cohen J. 2000. Vaccine studies stymied by shortage of animals.
Science :959–960.
European Council. 2002. http://conventions.coe.int/Treaty/EN/
Treaties/Html/123-A.htm European Convention ETS 123
Appendix A
Guide for Livestock Exporters. 1997. USDA AMS Transportation &
Marketing Livestock Transportation Program. Washington, DC:
http://www.ams.usda.gov/tmd/livestock/feedback2.htm USDA-
AMS.
Guidelines for the Humane Transportation of Research Animals.
2006. Guidelines for the Humane Transportation of Research
Animals. Washington, DC: Institute for Laboratory Animal
Research. National Research Council.
Hau J, Schapiro SJ. 2004. The welfare of non-human primates. In:
Kaliste E, editor. The welfare of laboratory animals, series:
Animal welfare. 2. Cambridge: Springer.
Hau J, Schapiro SJ. 2006. Non-human primates in biomedical
research. Scand J Lab Anim Sci 33:9–12.
Heinz A. 1999. Serotonergic dysfunction after social isolation –
implications for the development of aggressiveness and alcohol
dependence. Nervenarzt 70:780–789.
Honess PE, Johnson PJ, Wolfensohn S. 2004. A study of behavioural
responses of non-human primates to air transport and
re-housing. Lab Anim 38:119–132.
IATA. 2007. http://www.iata.org/ps/publications/lar.htm
IPS. 2007. http://www.internationalprimatologicalsociety.org/docs/
IPS_International_Guidelines_for_the_Acquisition_Care_and_
Breeding_of_Nonhuman_Primates_Second_Edition_2007.pdf
Jolly A. 1985. Evolution of primate behavior. 2, New York:
MacMillan. p 526.
Judge PG, De Waal FBM. 1997. Rhesus monkeys behaviour under
diverse population densities: Coping with long term crowding.
Anim Beh 54:643–662.
Kagira JM, Ngotho M, Thuita JK, Maina NW, Hau J. 2007.
Hematological changes in vervet monkeys (Chlorocebus aethiops)
during eight months’ adaptation to captivity. Am J Primatol 69:
1053–1063.
Kim C-Y, Han JS, Suzuki T, Han S-S. 2005. Indirect indicator of
transport stress in haematological values in newly acquired
cynomolgus monkeys. J Med Primatol 34:188–192.
Kyes R. 1998. Management of a natural habitat breeding colony of
long tailed macaques. Trop Biodiv 5:127–137.
Maple TL. 1979. Great apes in captivity: The good, the bad, and the
ugly. In: Erwin J, Maple T, Mitchell G, editors. Captivity and
behavior: Primates in breeding colonies, laboratories and zoos.
New York: Van Nostrand Reinhold. p 231–232.
Mason WA. 1960. Socially mediated reduction in emotional
responses of young rhesus monkeys. J Abnorm Soc Psychol 60:
100–104.
Moinde NN, Suleman MA, Higashi H, Hau J. 2004. Habituation,
capture and relocation of Sykes monkeys (Cercopithecus mitis
albotorquatus) on the coast of Kenya. Anim Welf 13:343–353.
Morton DB, Griffiths PHM. 1985. Guidelines on the recognition of
pain, distress, and discomfort in experimental animals and an
hypothesis for assessment. Vet Rec 116(431):436.
National Research Council. 2003. International perspectives: The
future of nonhuman primate resources. Washington, DC:
National Research Council.
Olfert ED, Cross BM, McWilliam AA. 1993. Guide to the care and
use of experimental animals. 1. 2, Ottawa: Canadian Council on
Animal Care. p 211.
Owen S, Thomas C, West P, Wolfensohn S, Wood M. 1997. Report
on primate supply for biomedical scientific work in the UK. Lab
Anim 31:289–297.
Prescott MJ, Jennings M. 2004. Ethical and welfare implications of
the acquisition and transport of non-human primates for use in
research and testing. Altern Lab Anim 32(1):323–327.
Reinhardt V. 2002. Comfortable quarters for primates in research
institutions. Comfortable quarters for laboratory animals. 9,
Washington, DC: Animal Welfare Institute. p 65–77.
Rowe N. 1996. The pictorial guide to the living primates. East
Hampton, NY: Pogonias Press. p 263.
Schapiro SJ, Bloomsmith MA. 1994. Behavioral effects of
enrichment on pair-housed juvenile rhesus monkeys. Am J
Primatol 32:159–170.
Suleman MA, Wango BVM, Sapolsky RM, Odongo H, Hau J. 2004.
Physiologic manifestations of stress from capture and restraint of
free-ranging male African green monkeys (Cercopithecus
aethiops). J Zoo Wildl Med 35:20–24.
Weatherall D. 2006. The report of Sir David Weatherall’s working
group: ’The use of non-human primates in research’,
The Academy of Medical Sciences. Medical Research Council,
Royal Society and Wellcome Trust.
Wheatley BP. 1980. Feeding and ranging of east Bornean Macaca
fascicularis. In: Lindburg DG, editor. The macaques: studies in
ecology, behavior, and evolution. New York: Van Nostrand
Reinhold. p 215–246.
Wolfensohn SE. 1997. Brief review of scientific studies of the welfare
implications of transporting primates. Lab Anim 31:303–305.
Wolfensohn SE, Honess PE. 2005. Handbook of primate husbandry
and welfare. Oxford: Blackwell Publishing Ltd.
A. L. Fernstrom et al.476
Downloaded By: [Karolinska Institute] At: 11:49 12 December 2008

















![[XLS] · Web view1336 gs Caenorhabditis briggsae Echinococcus granulosus Galaxea fascicularis Ovis aries Rhodopirellula baltica Suberites domuncula Schistosoma mansoni Steinernema](https://static.cupdf.com/doc/110x72/5b075a377f8b9a58148e2bed/xls-view1336-gs-caenorhabditis-briggsae-echinococcus-granulosus-galaxea-fascicularis.jpg)