



PATHOGENS OCCURRING IN THE WINTER PEA – MAIZE – WINTER
WHEAT ROTATION, THEIR HOST SPECIFICITY AND THE
POTENTIAL OF COMPOST IN SUPPRESSING FOOT AND ROOT
DISEASE OF PEAS
DISSERTATION
ZUR ERLANGUNG DES AKADEMISCHEN GRADES EINES DOKTORS DER AGRARWISSENSCHAFTEN (DR. AGR.)
EINGEREICHT AM FACHBEREICH ÖKOLOGISCHE AGRARWISSENSCHAFTEN DER UNIVERSITÄT KASSEL
VON
JELENA BAĆANOVIĆ
January 2015

Supervisor:
1 Prof. Dr. Maria R. Finckh
2 Prof. Dr. Gunter Backes
Defense date: 15.04.2015

To a loving memory of my sister
Milena Baćanović
(10.01.1985 – 07.04. 2009)
You will always be in my heart and my thoughts

TABLE OF CONTENTS
IV
Table of Contents
Table of Contents ............................................................................................................... IV
List of Abbreviations......................................................................................................... VII
List of Figures ................................................................................................................. VIII
List of Tables .................................................................................................................. XIII
1. GENERAL INTRODUCTION ............................................................................................ 1
1.1 Effects of modern agriculture and climate change on soil health and quality ........... 1
1.2 Possibilities to enhance soil health and quality and system resilience ..................... 3
1.3 Legumes as important components in sustainable agricultural systems .................. 4
1.4 Objectives and thesis outline ................................................................................... 6
1.5 References .............................................................................................................. 7
2. INCIDENCE OF FUSARIUM SPP. IN THE CROP ROTATION WINTER PEA – MAIZE – WINTER WHEAT AND THE POTENTIAL OF YARD WASTE COMPOST TO SUPPRESS FOOT AND ROOT ROT OF PEAS UNDER FIELD CONDITIONS .......................................10
Abstract ............................................................................................................................10
2.1 Introduction ............................................................................................................11
2.2 Material and Methods .............................................................................................13
2.2.1 Experimental site and setup ............................................................................13
Inoculum preparation and inoculation ............................................................................17
2.2.2 Assessments ........................................................................................................19
Soil N-dynamics ............................................................................................................19
Plant development and yields ........................................................................................19
Disease assessments, pathogen isolation and identification .........................................19
2.3 Data processing and analysis ................................................................................22
2.4 Results ..................................................................................................................23
2.4.1 Weather data ........................................................................................................23
2.4.2 Crop performance .................................................................................................25
2.4.3 Foot and root rot of peas and wheat .....................................................................28
2.4.4 Occurrence of Fusarium spp. in the tree crops in the rotation ...............................30
2.5 Discussion ..................................................................................................................34
2.6 References .................................................................................................................37
3. AGGRESSIVENESS OF FOUR PATHOGENS CAUSING FOOT AND ROOT ROT OF PEA (PISUM SATIVUM L.) ON A SPRING AND A WINTER PEA CULTIVAR UNDER VARIABLE TEMPERATURE CONDITIONS ........................................................................40
Abstract ............................................................................................................................40

TABLE OF CONTENTS
V
3.1 Introduction .................................................................................................................41
3.2 Materials and Methods ................................................................................................45
3.2.1 Pathogenicity of Fusarium avenaceum, Fusarium solani, Phoma medicaginis and Didymella pinodes, in sterile sand and in non-sterilized field soil ...................................45
3.2.2 Effect of the temperature on the aggressiveness of F. avenaceum and P. medicaginis ...................................................................................................................47
3.2.3 Pathogenicity of F. avenaceum .........................................................................47
3.2.4 Measurement of plant biomass and assessment of disease .................................48
3.2.5 Data processing and analysis ...............................................................................48
3.3 Results ........................................................................................................................49
3.3.1 Comparison of aggressiveness of F. avenaceum, F. solani, P. medicaginis and D. pinodes on two pea varieties in sand and field soil ........................................................49
3.3.2 Effect of temperature on disease severity caused by F. avenaceum and P. medicaginis ...................................................................................................................56
3.3.3 Pathogenicity of Fusarium avenaceum .................................................................57
3.4 Discussion ..................................................................................................................60
3.5 References .................................................................................................................63
4. EFFECTS OF COMPOST APPLICATION ON FOOT AND ROOT ROT OF SPRING AND WINTER PEA VARIETIES CAUSED BY FUSARIUM AVENACEUM, FUSARIUM
SOLANI, PHOMA MEDICAGINIS AND DIDYMELLA PINODES .........................................66
Abstract ............................................................................................................................66
4.1 Introduction ............................................................................................................67
4.2 Material and Methods .............................................................................................69
4.2.1 Potential of yard waste compost to suppress foot and root rot of peas caused by F. avenaceum, F. solani, P. medicaginis and D. pinodes ...................................................69
4.2.2 Effect of compost application rate on disease suppression ...................................70
4.2.3 Effect of reduced temperature on compost induced disease suppression .............70
4.2.4 Measurement of plant biomass and assessment of disease .................................71
4.2.5 Data processing and analysis ...............................................................................71
4.3 Results ...................................................................................................................72
4.3.1 Potential of yard waste compost to suppress foot and root rot of peas caused by F. avenaceum, F. solani, P. medicaginis and D. pinodes ...................................................72
4.3.2 Effect of compost application rate on disease suppression ...................................75
4.3.3 Temperature effect on compost induced disease suppression .............................77
4.4 Discussion ..............................................................................................................79
4.5 References .................................................................................................................82
5. GENERAL DISCUSSION ..............................................................................................84
5.1 References .................................................................................................................89

TABLE OF CONTENTS
VI
APPENDIX ...........................................................................................................................90
Chapter 2 ..........................................................................................................................91
Chapter 3 ........................................................................................................................ 100
Chapter 4 ........................................................................................................................ 110
Summary ........................................................................................................................... 116
Acknowledgments ............................................................................................................ 120
Erklärung .......................................................................................................................... 121

LIST OF ABBREVIATIONS
VII
List of Abbreviations SOM Soil organic matter
YWC Yard waste compost
IPCC Intergovernmental Panel on Climate Change
EEG Renewable Energy Sources Act
FHB Fusarium head blight
TGW Thousand grain weight
OA Organic amendments
Nmin Mineral nitrogen
ANOVA Analysis of Variance
DI Disease index
HSD Honest significant differences
ÖNORM Österreichisches Normungsinstitut
VDLUFA Verband deutscher landwirtschaftlicher Untersuchungs- und
Forschungsanstalten
SD Standard deviation
Pm Phoma medicaginis
Dp Didymella pinodes
Fs Fusarium solani
Fo Fusarium oxysporum
Fa Fusarium avenaceum
BOFRU Projekt: „Steigerung der Wertschöpfung ökologisch angebauter
Marktfrüchte durch Optimierung des Managements der
Bodenfruchtbarkeit“
Df Degrees of freedom
PDA Potato dextrose agar
SNA Synthetic Nutrient Agar
PPA Pentachloronitrobenzene Peptone Agar
DM Dry matter
KLIFF KLimaFolgenForschung in Niedersachsen

LIST OF FIGURES
VIII
List of Figures
Figure 2.1. Section of the crop rotation that was studied in the field experiment. .................14
Figure 2.2. Main plot divided in four experimental plots presenting four different treatments
O=Control, I=Inoculated with P. medicaginis, C=Compost and I+C= Inoculated + Compost. 15
Figure 2.3. Inoculum of Phoma medicaginis grown on oat seeds.........................................17
Figure 2.4. Monthly mean temperatures (°C) and precipitation (mm) in the period between
2009 and 2013, compared with thirty year average (1970-2000). .........................................24
Figure 2.5. Percentage of the emerged pea plants that survived the winters 2010, 2011 and
2012. Treatments O = Control; I = Inoculated with P. medicaginis; C= Compost; I+C=
Inoculated with P. medicaginis + Compost. The error bars represent the ± 1 SD.................25
Figure 2.6. Dry matter yield of peas (A) and maize (B), and grain yield of wheat (14%
moisture) (C). For peas O= control, I= inoculated with P. medicaginis, C= compost, I+C=
inoculated with P. medicaginis + compost. For maize and wheat C= with compost, O=
without compost. Different small letters within one crop indicate significant differences in yield
among years at P < 0.05 (Tukey’s HSD test). The error bars represent the ± 1 SD. .............26
Figure 2.7. Effect of compost application on pea fresh matter yield in May 2010 and 2011..
.............................................................................................................................................27
Figure 2.8. Mineral nitrogen (Nmin) content in soil at 0-60 cm depth. The error bars
represent the ± 1 SD. n.a. stand for not available. ................................................................28
Figure 2.9. Foot Disease Index (DI) on peas in March and May 2010-2012. For the year
2012 disease was assessed only in May. Different small letters within one year indicate
significant differences in DI at P < 0.05 (Tukey’s HSD test). Different capital letters indicate
significant differences in DI in May among years at P < 0.05 (Tukey’s HSD test). The error
bars represent the ± 1 SD. ...................................................................................................29
Figure 2.10. Incidence of P. medicaginis (Pm), D. pinodes (Dp), F. solani (Fs), F. oxysporum
(Fo) and F. avenaceum (Fa) isolated from pea in May of 2010, 2011, 2012. Different small
letters within one pathogen indicate significant differences in incidence at P < 0.05 (Tukey’s
HSD test). .............................................................................................................................29
Figure 2.11. Foot rot disease of wheat caused by Fusarium spp., Pseudocercosporella
herpotrichoides and Rhizoctonia cerealis in treatments without (O) and with compost (C).
The error bars represent the ± 1 SD. ....................................................................................30
Figure 2.12. Incidence of Fusarium spp. isolated from three crops in the rotation. WP =
winter peas, M = maize and WW = winter wheat; time of samplings, month/year, is given in
the brackets. .........................................................................................................................32
Figure 3.1. Disease index (DI) of Santana and EFB33 plants grown in sand and soil and
inoculated with five isolates of F. avenaceum (A), F. solani (B), D. pinodes (C) and P.

LIST OF FIGURES
IX
medicaginis (D). Different small letters within variety and substrate are indicating significant
differences in DI at P < 0.05 (Tukey’s HSD test). The horizontal line in the boxplot shows the
median, the bottom and tops of the box the 25th and 75th percentiles and the vertical lines
the minimum and maximum values; outliers as single points. Mean values of DI are marked
with triangles. .......................................................................................................................50
Figure 3.2. Fresh weights of Santana and EFB33 plants inoculated with five isolates of F.
avenaceum and grown in sterile sand or non-sterilized field soil. Different letters within
variety and substrate are indicating significant differences in DI at P < 0.05 (Tukey’s HSD
test). The error bars represent the ± 1 SD. ...........................................................................51
Figure 3.3. Fresh weights of Santana and EFB33 plants inoculated with five isolates of P.
medicaginis and grown in sterile sand or non-sterilized field soil. Different letters within
variety and substrate are indicating significant differences in DI at P < 0.05 (Tukey’s HSD
test). The error bars represent the ± 1 SD. ...........................................................................51
Figure 3.4. Typical symptoms of F. solani infection on the stem base of Santana (A) and
EFB33 (B) plants. .................................................................................................................52
Figure 3.5. Wilting symptoms of Santana caused by F. avenaceum with the formation of
bright orange sporodochia (marked with the arrow). .............................................................53
Figure 3.6. Disease index (DI) of Santana and EFB33 plants grown in sand and soil and
inoculated with F. avenaceum (Fa), F. solani (Fs), P. medicaginis (Pm) and D. pinodes (Dp).
O is the non-inoculated control. The horizontal line in the boxplot shows the median, the
bottom and tops of the box the 25th and 75th percentiles and the vertical lines the minimum
and maximum values; outliers as single points. Mean values of DI are marked with triangles.
Different letters within variety and substrate are indicating significant differences in DI at P <
0.05 (Tukey’s HSD test). ......................................................................................................54
Figure 3.7. Fresh weights of Santana and EFB33 inoculated with F. avenaceum (Fa), F.
solani (Fs), P. medicaginis (Pm) and D. pinodes (Dp). Different letters within variety and
substrate are indicating significant differences in DI at P < 0.05 (Tukey’s HSD test). The error
bars represent the ± 1 SD. ...................................................................................................54
Figure 3.8. Disease index of Santana and EFB33 inoculated with F. avenaceum (Fa) and P.
medicaginis (Pm) and grown under different temperature regimes. O is the non-inoculated
control. The horizontal line in the boxplot shows the median, the bottom and tops of the box
the 25th and 75th percentiles and the vertical lines the minimum and maximum values;
outliers as single points. Mean values of DI are marked with triangles. Different letters within
variety and temperature are indicating significant differences in DI at P < 0.05 (Tukey’s HSD
test). .....................................................................................................................................56
Figure 3.9. Mean values of fresh weights of Santana and EFB33 plants grown at three
temperature regimes and inoculated with F. avenaceum (Fa) and P. medicaginis (Pm). O is

LIST OF FIGURES
X
the non-inoculated control. Different capital letters within one variety indicate significant
differences in fresh weights among temperature regimes, whereas small letters indicate
significant differences among treatments within variety and temperature regime at P < 0.05
(Tukey’s HSD test). The error bars represent the ± 1 SD. ....................................................57
Figure 4.1. Effect of compost application (no-without, N-not sterilized and S-sterilized
compost) on fresh weight per plant of Santana in sand and soil inoculated with F. avenaceum
(Fa), F. solani (Fs), P. medicaginis (Pm) and D. pinodes (Dp). Different letters within one
substrate and one pathogen indicate significant differences in fresh weights among compost
treatments at P < 0.05 (Tukey’s HSD test). The error bars represent the ± 1 SD. .................74
Figure 4.2. Disease index of Santana and EFB33 inoculated with F. avenaceum (Fa), F.
solani (Fs), P. medicaginis (Pm) and D. pinodes (Dp) and amended with compost (no-
without, low- 3.5% v/v and high- 20% v/v). O is the non-inoculated control. The horizontal line
in the boxplot shows the median, the bottom and tops of the box the 25th and 75th
percentiles and the vertical lines the minimum and maximum values; outliers as single points.
Mean values of DI are marked with triangles. Different letters within variety and pathogen are
indicating significant differences in DI at P < 0.05 (Tukey’s HSD test). .................................75
Figure 4.3. Effect of compost application rate (no-without, low- 3.5% v/v and high- 20% v/v)
on the fresh weight of (A) Santana and (B) EFB33 plants inoculated with 105 spores g-1 of
substrate of F. avenaceum (Fa), F. solani (Fs), P. medicaginis (Pm) and D. pinodes (Dp).
Different letters within pathogen are indicating significant differences in fresh weights at P <
0.05 (Tukey’s HSD test). The error bars represent the ± 1 SD. .............................................76
Figure 4.4. Effect of compost application (no-without, compost- 20% v/v) on the fresh
weights of Santana (A) and EFB33 (B) plants inoculated with F. avenaceum (Fa) and P.
medicaginis (Pm) and grown under different temperature regimes (low - 16/12°C and high -
19/16°C). Asterisk indicates significant differences at P < 0.05 among treatments with and
without compost within one temperature regime and pathogen. The error bars represent the
± 1 SD. .................................................................................................................................78
Figure A 2.1. Effect of treatments on pea emergence one month after sowing presented as
percentage of sowing densities. The error bars represent the ± 1 SD. ..................................92
Figure A. 2.2. Emergence (Autumn) and winter survival (Spring) of peas as a percentage of
sown plants in 2011/12. The error bars represent the ± 1 SD. ..............................................92
Figure A. 2.3. Fresh weight per plant of maize in 2010, 2011 and 2012. Different letters
indicate significant differences at P < 0.05 (Tukey’s HSD test). The error bars represent the ±
1 SD. ....................................................................................................................................94
Figure A. 2.4. Thousand grain weights (g) (A) and number of the heads per m- 2 (B) of wheat
in treatments with (+ compost) and without compost (- compost) in three experimental years

LIST OF FIGURES
XI
(2011 - 2013). Different letters indicate significant differences at P < 0.05 (Tukey’s HSD test).
The error bars represent the ± 1 SD. ....................................................................................94
Figure A. 2.5. Incidence (%) of pathogens isolated from pea in different treatments in March
2010 and 2011. O = Control; I = Inoculated with P. medicaginis; C= Compost; I+C=
Inoculated with P. medicaginis + Compost. ..........................................................................96
Figure A. 2.6. Incidence (%) of pathogens isolated from pea in different treatments in May of
three experimental years. O = Control; I = Inoculated with P. medicaginis; C= Compost; I+C=
Inoculated with P. medicaginis + Compost. ..........................................................................96
Figure A. 2.7. Pathogen incidence on pea in March and May of 2010 and 2011. Pm = P.
medicaginis, Dp = D. pinodes, Fs = F. solani and Fo = F. oxysporum. .................................98
Figure A. 2.8. External (A) and Internal (B) disease scores of pea in May of all three
experimental years. Score 1 stands for healthy plant; score 9 for dead plant. O = Control; I =
Inoculated with P. medicaginis; C= Compost; I+C= Inoculated with P. medicaginis +
Compost. ..............................................................................................................................97
Figure A. 2.9. External (A) and Internal (B) disease scores of pea in March and May 2010.
Score 1 stands for healthy plant; score 9 for dead. O = Control; I = Inoculated with P.
medicaginis; C= Compost; I+C= Inoculated with P. medicaginis + Compost. .......................98
Figure A. 2.10. External (A) and Internal (B) disease scores of pea in March and May 2011.
Score 1 stands for healthy plant; score 9 for dead plant. O = Control; I = Inoculated with P.
medicaginis; C= Compost; I+C= Inoculated with P. medicaginis + Compost. .......................99
Figure A. 2.11. Incidence of different Fusarium spp. isolated from maize (A) and wheat (B) in
treatments with (+ compost) and without compost (- compost) from 2009-2013. For wheat
n=30 in 2011 and n=20 in 2012 and 2013. For maize n=60 for all three years. ................... 100
Figure A. 3.1. External lesion scores for all of the isolates of all tested pathogens on
Santana .............................................................................................................................. 103
Figure A. 3.2. External lesion scores for all of the isolates of all tested pathogens on EFB33
........................................................................................................................................... 104
Figure A. 3.3. Fresh weight of Santana and EFB33 plants inoculated with five isolates of F.
solani and grown in sterile sand or non-sterilized field soil. The error bars represent the ± 1
SD. ..................................................................................................................................... 105
Figure A. 3.4. Fresh weight of Santana and EFB33 plants inoculated with five isolates of D.
pinodes and grown in sterile sand or non-sterilized field soil. The error bars represent the ± 1
SD. ..................................................................................................................................... 105
Figure A. 3.5. Correlation between fresh weight and disease severity of Santana grown in
sterile sand (A) and non-sterilized field soil (B) and inoculated with different pathogens. Only
significant regressions are indicated with regression lines. ................................................. 108

LIST OF FIGURES
XII
Figure A. 3.6. Correlation between fresh weight and disease severity of EFB33 grown in
sterile sand (A) and non-sterilized field soil (B) and inoculated with different pathogens. Only
significant regressions are indicated with regression lines. ................................................. 109
Figure A. 4.1. Disease index of EFB33 in sand and soil inoculated with F. avenaceum (Fa),
F. solani (Fs), P. medicaginis (Pm) and D. pinodes (Dp) and amended with compost (no-
without, N-not sterilized and S-sterilized compost). The horizontal line in the boxplot shows
the median, the bottom and tops of the box the 25th and 75th percentiles and the vertical
lines the minimum and maximum values; outliers as single points. Mean values of DI are
marked with triangles.......................................................................................................... 111
Figure A. 4.2. Effect of compost application (no-without, N-not sterilized and S-sterilized
compost) on fresh weight per plant of EFB33 in sterile sand and non-sterilized field soil
inoculated with F. avenaceum (Fa), F. solani (Fs), P. medicaginis (Pm) and D. pinodes (Dp).
Different letters within one substrate and one pathogen indicate significant differences in
fresh weights among compost treatments at P < 0.05 (Tukey’s HSD test). The error bars
represent the ± 1 SD. ......................................................................................................... 111
Figure A. 4.3. Effect of compost application (no-without, N-not sterilized and S-sterilized
compost) on dry weight per plant of Santana in sterile sand and non-sterilized field soil
inoculated with F. avenaceum (Fa), F. solani (Fs), P. medicaginis (Pm) and D. pinodes (Dp).
Different letters within one substrate and one pathogen indicate significant differences in
fresh weights among compost treatments at P < 0.05 (Tukey’s HSD test). The error bars
represent the ± 1 SD. ......................................................................................................... 112
Figure A. 4.4. Effect of compost application (no-without, N-not sterilized and S-sterilized
compost) on dry weight per plant of EFB33 in sterile sand and non-sterilized field soil
inoculated with F. avenaceum (Fa), F. solani (Fs), P. medicaginis (Pm) and D. pinodes (Dp).
Different letters within one substrate and one pathogen indicate significant differences in
fresh weights among compost treatments at P < 0.05 (Tukey’s HSD test). The error bars
represent the ± 1 SD. ......................................................................................................... 112
Figure A. 4.5. Effect of compost application rate (no-without, low- 3.5% v/v and high- 20%
v/v) on the dry weight per plant of Santana and EFB33 plants inoculated with F. avenaceum
(Fa), F. solani (Fs), P. medicaginis (Pm) and D. pinodes (Dp). Different letters within one
variety and one pathogen indicate significant differences in fresh weights among compost
treatments at P < 0.05 (Tukey’s HSD test). The error bars represent the ± 1 SD. ............... 114
Figure A. 4.6. Effect of compost application (no-without, compost- 20% v/v) on the dry
weights of Santana (A) and EFB33 (B) plants inoculated with F. avenaceum (Fa) and P.
medicaginis (Pm) and grown under different temperature regimes (low – 12/16°C, high –
16/19°C). Asterisk indicates significant differences in fresh weights between two compost
treatments within one pathogen. ......................................................................................... 115

LIST OF TABLES
XIII
List of Tables
Table 2.1. Chemical characteristic of composts used in the field from 2009-2013. ...............16
Table 2.2. Treatments applied in the field experiment. .........................................................17
Table 2.3. Timing of field operations. ...................................................................................18
Table 2.4. Scoring scheme for assessment of root and foot rot of peas (Pflughöft, 2008). ...21
Table 2.5. Number and average temperatures of the frost and icing days in February. ........23
Table 2.6. Incidence of different Fusarium spp. in the maize and wheat in 2010 - 2013.
Different letters within one species indicate significant differences in incidence among three
experimental years at P < 0.05 (Tukey’s HSD test). .............................................................33
Table 3.1. Pathogens and isolates used in experiments. ......................................................46
Table 3.2. Temperature regimes used in the experiment .....................................................47
Table 3.3. Plant species, varieties and sowing density used in the experiment. ...................47
Table 3.4. Disease Index (DI), coefficient of determination (R2) and significance for the
correlation between Disease Index and fresh weights of EFB33 and Santana plants grown in
sterile sand and not sterilized field soil. ................................................................................55
Table 3.5. Emergence, percentage of emerged plants that wilted, fresh weight per pot and
plant height of nine plant species inoculated with two F. avenaceum isolates, (Fa1 – isolate
from wheat, Fa4 – isolate from pea). ....................................................................................59
Table 4.1. Chemical properties of substrates and composts used in the experiment. ...........70
Table 4.2. Disease Index (DI) of Santana in sterile sand and non-sterilized field soil in non-
inoculated control and treatments inoculated with F. avenaceum, F. solani, P. medicaginis
and D. pinodes and amended with compost (no-without, N-not sterilized and S-sterilized
compost). Different letters within one pathogen and one substrate indicate significant
differences in DI among compost treatments at P < 0.05 (Tukey’s HSD test). ......................73
Table 4.3. Disease index of Santana and EFB33 inoculated with F. avenaceum and P.
medicaginis, amended with compost (no-without, compost - 20% v/v) and grown under two
temperature regimes (low - 16/12°C and high - 19/16°C). Different letters within one variety,
temperature and pathogen are indicating significant differences in DI among compost
treatments at P < 0.05 (Tukey’s HSD test). Percent change presents difference in DI in low
temperature treatment compared to high. .............................................................................77
Table A. 2.1. Mineral nitrogen (Nmin) content in the soil at 0-60 cm depth. O=Control,
I=Inoculated, C=Compost, I+C=Inoculated+Compost. ..........................................................93
Table A. 2.2. Fresh and dry weights of pea and weeds in 2010 and 2011. O = Control, I =
Inoculated, C = Compost, I+C = Inoculated + Compost ........................................................93
Table A. 2.3. F-values for dry matter yields of peas and maize, and grain yield of wheat in
field experiment from 2009-2013. .........................................................................................94

LIST OF TABLES
XIV
Table A. 2.4. F-values for the disease index (DI) for root and foot rot of peas in 2010 and
2011. ....................................................................................................................................95
Table A. 3.1. F-values for DI for isolate comparison of four pathogens in three-way ANOVA.
........................................................................................................................................... 101
Table A. 3.2. F-values for DI of Santana and EFB33 inoculated with five isolates of four
pathogens in sand and soil. ................................................................................................ 101
Table A. 3.3. F-values for fresh weights of Santana and EFB33 grown in sand and soil and
inoculated with five isolates of four pathogens. ................................................................... 101
Table A. 3.4. F-values for dry weights of Santana and EFB33 grown in sand and soil and
inoculated with five isolates of four pathogens. ................................................................... 102
Table A. 3.5. F-values for the DI of pea inoculated with four different pathogens in three-way
ANOVA............................................................................................................................... 106
Table A. 3.6. F-values for DI for pathogen comparison for Santana and EFB33 in sterile sand
and non-sterilized field soil. ................................................................................................ 106
Table A. 3.7. ANOVA table and F values for fresh and dry weights per plant of Santana and
EFB33 inoculated with four different pathogens and grown in sterile sand and non-sterilized
field soil. ............................................................................................................................. 106
Table A. 3.8. F-values for DI for Santana and EFB33 inoculated with P. medicaginis and F.
avenaceum and grown under three different temperature regimes. .................................... 107
Table A. 3.9. F-values for fresh and dry weights for Santana and EFB33 inoculated with P.
medicaginis and F. avenaceum and grown under three different temperature regimes. ..... 107
Table A. 4.1. F-values for fresh and dry weights and DI for Santana and EFB33 inoculated
with F. avenaceum, F. solani, P. medicaginis and D. pinodes and amended with 20% v/v of
non-sterilized and γ sterilized YWC and grown in sterile sand and non-sterilized field soil. 110
Table A. 4.2. F-values for DI, fresh and dry weights of Santana and EFB33 inoculated with
four pathogens and amended with two rates of compost (low – 3.5% v/v and high – 20% v/v)
and grown in sterile sand in three-way ANOVA. ................................................................. 113
Table A. 4.3. F-values for DI, fresh and dry weights of Santana and EFB33 inoculated with
four pathogens and amended with two rates of compost (low – 3.5% v/v and high – 20% v/v)
and grown in sterile sand. ................................................................................................... 113
Table A. 4.4. F-values for DI, fresh and dry weights of Santana and EFB33 inoculated with F.
avenaceum and P. medicaginis, amended with 20% v/v YWC and grown under two
temperature regimes (low - 16/12°C and high – 19/16°C) in sterile sand in three-way ANOVA
………………………………………………………………………………………………………114

GENERAL INTRODUCTION
1
1. GENERAL INTRODUCTION
1.1 Effects of modern agriculture and climate change on soil health and quality
Agricultural systems are considered to be sustainable if they can sustain themselves over a
long period of time, meaning they are economically viable, environmentally safe and socially
fair (Lichtfouse et al., 2009). At the center of each sustainable system is the soil, a resource
that is finite, unequally distributed, nonrenewable and prone to degradation (Lal, 2009).
Over the past half century, agriculture has become highly dependent on synthetically
produced agrochemicals resulting in changes in agricultural practices with respect to soil
management that have led to a general decline in soil quality (Bailey and Lazarovits, 2003).
Thus, in many modern industrialized agricultural systems traditional practices of crop rotation
and the use of animal manures or plant residues have been abandoned in favor of short
rotations, monocultures, intensive tillage, synthetic fertilizers and pesticides. In the long term,
this approach adversely affects soil fertility, quality and health (Sturz and Christie, 2003) as
the balance between beneficial and detrimental members of the resident microbial population
in the soil shifts in favor of the latter (Bailey and Lazarovits, 2003; Hoitink and Boehm, 1999;
Katan, 1997), leading to a decline in system productivity and an increase in plant diseases
(Doran and Zeiss, 2000). Nearly 40 % of the world’s agricultural land is showing human-
induced degradation as a result of soil erosion, pollution, over-grazing, salinization,
desertification, etc. (Doran, 2002). Because of these adverse effects of industrialized
agriculture on the environment, soil, water, and air quality there is a need to change the
existing short term approach in modern agriculture into long-term oriented, holistic and
sustainable systems geared towards improved soil quality and health.
Soil quality has been defined as “the capacity of a specific kind of soil to function, within
natural or managed ecosystem boundaries, to sustain plant and animal productivity, maintain
or enhance water and air quality, and support human health and habitation” (Schjønning et
al., 2004). Similarly, soil health has been defined as “the continued capacity of soil to function
as a vital living system, within ecosystem and land-use boundaries, to sustain biological
productivity, maintain the quality of air and water environments, and promote plant, animal,
and human health” (Doran et al., 1996). These definitions emphasize the multiple functions
soil resources have to fulfil in the modern world – i) production of food to meet the growing
needs of the world’s population, ii) energy production through growth of energy plants, iii)
sequestration of carbon, iv) improvement of water quality and use efficiency, v) waste
disposal medium, vi) habitat for diverse organisms, etc. (Lal, 2008). Soil organic matter
(SOM) is a key for maintaining soil quality and essential for the sustainability of agricultural
systems (Dick and Gregorich, 2004; Lal, 2009). Many soil properties such as microbial

GENERAL INTRODUCTION
2
activity, soil structure, cation exchange capacity, water to air ratio, etc., are directly or
indirectly affected by SOM content (Dick and Gregorich, 2004).
In addition to the need for high and stable production to feed a growing world population,
current agriculture also has to cope with global climatic changes and extreme weather events
of considerable magnitude (Crutzen, 2002; Walther et al., 2002; Parmesan and Yohe, 2003;
Root et al., 2003) that require mitigation and adaptation strategies. This needs to be based
on a solid understanding of soil processes and how these are affected by climate and
agricultural practices. The impacts of a changing climate on the system productivity are a
combination of direct effects on the growing environment, changes in the geography and
prevalence of pests and diseases, as well as changes in fertility and biological function of
soils (Jarvis et al., 2010). Thus, any environmental change that alters soil nutrient availability
changes the trophic structure in the soil and can affect crop productivity (Pritchard, 2011).
Moreover, these changes lead to changes in geographic distribution of host plants and their
pathogens and to changes in host-pathogen interactions (Coakley et al., 1999).
Climate change and climate variability are important drivers in the epidemiology of plant
diseases (Juroszek and Tiedemann, 2011). Working group I reported in the Third
Assessment Report of The Intergovernmental Panel on Climate Change (IPCC) that average
diurnal temperature had increased by 0.6°C in the 20th century. Sea levels increased
between 0.1 and 0.2 m. Overall annual changes in precipitation have been 1 % per decade.
However, frequency and intensity of heavy rainfall events as well as occurrence of extreme
weather conditions and cloud cover increased (IPCC, 2001). Climate change scenarios for
Germany exhibit a definite warming trend. By the year 2080 the annual average temperature
is predicted to increase by 1.6 – 3.8°C, compared to 1990. Although only small changes in
average annual precipitation are predicted, significant changes in the annual distribution of
precipitation are expected. Winter precipitation will increase between about 7 and 30%, while
summer precipitation is expected to decrease by about 5 to 33% by 2080 (Schröter et al.,
2005).
Factors that are most likely to influence disease development, spread and severity are
increased atmospheric CO2, extreme weather conditions (heavy rains, sudden drops of
temperature, drought, cyclones) and warmer winter temperatures (Cannon, 1998;
Chakraborty et al., 2000; Pimentel et al., 2001). Changed environmental conditions cause
alteration in morphology and physiology of plants such as stoma morphology, increased net
photosynthesis, carbohydrate accumulation in plant tissues, increased fiber content and
cuticular waxes, etc. that can modify host resistance (Chakraborty et al., 2000; Juroszek and
Tiedemann, 2011).

GENERAL INTRODUCTION
3
Despite the considerable body of literature available on the potential effects of the global
environmental changes on the above ground components of the ecosystem, by 2004, less
than 3 % of these articles were considering the belowground organisms or processes
(Pritchard, 2011; Wardle et al., 2004). Soil inhabiting organisms play an important role in the
dynamics of SOM decomposition and nutrient cycling. Thus, there is a need to understand
how changes in climate will affect the soil community. Predicted climatic changes will affect
soil microorganisms directly and indirectly through effects on plant growth and physiology.
Soil borne pathogens are already playing an important role, limiting agricultural production
worldwide (Pritchard, 2011). Increased winter temperatures will most likely reduce the
latency period and allow a higher number of generations of pathogens per season that can
result in more rapid buildup of pathogen populations in early spring (Garrett et al., 2006;
Pariaud et al., 2009). Effects of climate change on soil microbial processes are also likely to
affect pathogens. In moist and warm soils microbial diversity and biomass are higher and
that can lead to faster microbial turnover of organic material.
(Wardle et al., 2004) state that composition and functioning of soil organisms are closely
related to aboveground plant composition. Introduction of novel plant species into existing
ecosystems may alter the timing and type of inputs by rhizodeposition, introduce new
antimicrobial compounds, alter soil structure, and change the existing nutrient cycles (Wolfe
and Klironomos, 2005). Moreover, it can lead to introduction of new pests and pathogens
(Chakraborty et al., 2000). Newly introduced crops can act as alternative hosts to already
present pathogens and increase disease pressure for the succeeding crops. For example,
incidence of Fusarium graminearum a pathogen of Fusarium Head Blight (FHB) of wheat in
Western Europe has increased as a result of inclusion of maize in crop rotations (West et al.,
2012).
1.2 Possibilities to enhance soil health and quality and system resilience
Soil resilience and resistance are two fundamental components of soil quality (Seybold et al.,
1999). Soil resistance is “the capacity of a soil to continue to function without change
throughout a disturbance” (Herrick and Wander, 1997). Resilience is defined as “the capacity
of a system to absorb disturbance and re-organize while undergoing change so as to still
retain essentially the same function, structure, identity and feedbacks” (Walker et al., 2004).
A resilient soil can replace and rebuild functions that may have been compromised by
management and environmental changes. Both properties depend on the soil type, diversity
at all levels, land use, level of the disturbance, and climate (Seybold et al., 1999). Resilient
and resistant soils are bases on which resilient and sustainable agricultural systems should
be built on.

GENERAL INTRODUCTION
4
In recent decades, awareness of the dependence of sustainable agriculture on the
management of soil quality resulted in changes in soil tillage practices, introduction of cover
crops, living or dead mulches, application of organic amendments such as compost, green
manures or manures in the production system (Abawi and Widmer, 2000). These practices
that maintain or increase SOM content are improving soil functions and contributing to
sustainability of the system (Lal, 2008) as a soil’s biological, chemical, and physical
properties are functions of SOM (Abawi and Widmer, 2000). Soil is also the environment that
hosts soil borne pathogens; diseases caused by these are the result of a disturbed balance
between beneficial and harmful parts of the soil microbial community (Alabouvette et al.,
2004). Incorporation of different organic amendments not only increases SOM, provides
plant nutrients, and improves physical properties of soil, but it can also introduce beneficial
organisms which contribute to soil health and disease suppression (Boutler et al., 2000; Dick
and Gregorich, 2004; Hoitink et al., 1997; Hoitink and Fahy, 1986).
Composting of different biodegradable materials is an efficient and environmentally safe way
of dealing with organic wastes (Boutler et al., 2000) and, at the same time, the final product
is stable compost rich in humus, nutrients, and beneficial microorganisms. Compost induced
disease suppression can be attributed mainly to the biological properties of composts
(Hadar, 2011; Hoitink and Boehm, 1999). Characteristics of pathogen inhibiting microbial
populations in composts depend on the type of material used and conditions under which
composting was conducted (Boutler et al., 2000; Recycled Organics Unit, 2006).
Depending if the disease suppression is the result of the compost’s overall microbial biomass
or a specific group of microorganisms, two categories of disease suppression can be
distinguished – general and specific. General disease suppression is the result of total
microbial activity in a medium after compost addition which increases nutrient depletion
resulting in microstasis of pathogens (Aviles et al., 2011). In specific suppression, one or
more organisms are responsible for disease suppression. A range of different mechanisms
responsible for this type of suppression, such as competition for nutrients, infection sites,
direct antagonism, antibiotic production, parasitism, etc., have been described. Which of
these prevails depends on the type of pathogen and antagonistic organisms present in the
system (Hoitink et al., 1997; Hoitink and Fahy, 1986; Recycled Organics Unit, 2006).
1.3 Legumes as important components in sustainable agricultural systems
Enhanced diversity contributes to system resilience. Promoting the biodiversity in
agroecosystems, below ground as well as aboveground, provides ecological services that
are necessary for long-term sustainability of the system. However, this is only possible if the
diversification strategy takes into account the site specific conditions and is adjusted to local
conditions. Plants that not only have a role as a crop but also provide an ecological service to

GENERAL INTRODUCTION
5
the system are of great importance for the sustainability of agricultural systems. Because of
their ability for biological nitrogen fixation, leguminous plants can generally contribute to soil
fertility thus delivering an important ecological service. Due to the restrictions on the use of
mineral nitrogen fertilizers organic farming is highly dependent on legumes to build soil
fertility (Corre-Hellou and Crozat, 2005) and requires crop rotations with high frequencies of
leguminous crops. The amount of nitrogen fixed by different leguminous species is estimated
to be in the range between 20 and 350 kg N ha-1 (Karpenstein-Machan and Stuelpnagel,
2000; Peoples et al., 1995). In addition to nitrogen, legumes can also mobilize and
remineralize other nutrients (Eriksen and Thorup-Kristensen, 2002), thereby preventing their
leaching and making them available for following crops (Mueller and Thorup-Kristensen,
2001). Furthermore, used as a cover crop they can prevent soil erosion (Langdale et al.,
1991), increase water infiltration (Meisinger et al., 1991), improve the soil structure (Roldán
et al., 2003), and contribute to weed suppression (Hartwig and Ammon, 2002; Teasdale et
al., 2007).
Peas (Pisum sativum L.) together with faba beans (Vicia faba) are the most common grain
legumes grown in Germany (Pflughöft, 2008). Besides their ecological services provided in
crop rotations, peas and faba beans are important sources of proteins for humans and
animals. Despite their importance, production of peas is declining in Germany and many
other European countries (Sass, 2009; Urbatzka et al., 2011) and the same trend is present
in organic agriculture (Boehm, 2009). Low and instable yields together with low prices are
discouraging organic farmers from growing peas (Charles et al., 2007). The yield instability is
due to a range of abiotic and biotic factors (Fuchs et al., 2014). Abiotic factors limiting pea
productivity are unfavorable soil structure (Allmaras et al., 2003, 1988; Tu, 1994), lack of
nutrients (Fuchs et al., 2014), and toxic compounds in the soil (Narawal, 2000). Moreover,
the commonly used spring pea varieties are highly susceptible to soil-borne diseases
(Urbatzka et al., 2011a), compete poorly with weeds (Corre-Hellou et al., 2011), and they are
prone to lodging (Graß, 2003). Thus, legumes, peas in particular, are in principle able to
provide multiple services to the German agroecosystem, however, there are severe
constraints to pea production in Germany and in general. There is an urgent need to find
options to minimize the numerous factors limiting pea production and to optimize the
beneficial aspects of their inclusion into the crop rotation.
Growing peas in mixtures with cereals (Corre-Hellou et al., 2011) can be useful to a certain
extent, mainly for the purposes of better weed control and also improved N use efficiency
(Bedoussac and Justes, 2010; Saeed, 2013). Secondly, substitution of commonly grown
spring peas with winter pea varieties can be a good alternative as winter peas efficiently
compete with weeds and have a higher yield potential and stability (Stoddard et al., 2006;
Urbatzka et al., 2011a). As a catch crop they may prevent leaching of nitrogen during the

GENERAL INTRODUCTION
6
winter (Graß, 2003). Urbatzka et al., (2011b) showed that winter peas fix higher amounts of
N2 than spring varieties. However, little is known about their susceptibility to common soil
borne pathogens that currently present a major constraint to spring pea production
(Pflughöft, 2008).
1.4 Objectives and thesis outline
The main focus of the work presented in this thesis is soil health management with a specific
focus on soilborne diseases of peas. The central part of the research is an organic rotation
encompassing winter peas as green manure crop followed by maize and wheat. In the field
and in the laboratory the potential of improving system health with compost application was
explored. The results of the field experiments are presented in Chapter 2.
The key for the success of the field rotation described above is the ability of peas to fix
enough nitrogen. This depends crucially on pea health and until now little is known about the
importance and specificity of pathogens affecting winter peas in the German climate. Also,
there are open questions about the role of peas as alternative host for mycotoxin producing
Fusarium spp. pathogens of maize and wheat. Therefore, the most important pathogens of
the root and foot rot complex on pea identified in the field experiment were tested for their
aggressiveness on spring and winter peas under controlled conditions (Chapter 3). The
aggressiveness of Phoma medicaginis var pinodella, Didymella pinodes, Fusarium solani f.
sp. pisi and Fusarium avenaceum was assessed in sterile sand and in non-sterilized field
soil. Also, the pathogenicity of F. avenacuem on nine different plant species (including two
pea varieties) commonly grown in Germany was determined. P. medicaginis and F.
avenaceum were chosen because of their high virulence and wide host range for further
tests of temperature effects on the level of aggressiveness.
Chapter 4 deals with the potential of Yard Waste Compost (YWC) to suppress root and foot
rot of peas caused by P. medicaginis var pinodella, D. pinodes, F. solani f. sp. pisi and F.
avenaceum. In addition, effects of compost amounts and temperature were studied.
In the final Chapter 5, the results of Chapters 2 to 4 are summarized and discussed.
The research presented in this dissertation was conducted as the part of the “Climate change and production of healthy crops - processes and adaptation strategies by the
year 2030” in the frame of the research project “KLIFF - Climate Impact Research in
Lower Saxony".

GENERAL INTRODUCTION
7
1.5 References
Abawi, G.S., Widmer, T.L., 2000. Impact of soil health management practices on soilborne pathogens, nematodes and root diseases of vegetable crops. Appl. Soil Ecol. 15, 37–47.
Alabouvette, C., Backhouse, D., Steinberg, C., Donovan, N.J., Edel-Hermann, V., Burgess, L.W., 2004. Microbial diversity in soil - effects on crop health, in: Schjønning, P., Elmholt, S., Christensen, B.T. (Eds.), Managing Soil Quality: Challenges in Modern Agriculture. CABI, Wallingford, pp. 121–138.
Allmaras, R.R., Fritz, V.A., Pfleger, F.L., Copeland, S.M., 2003. Impaired internal drainage and Aphanomyces euteiches root rot of pea caused by soil compaction in a fine-textured soil. Soil Tillage Res. 70, 41–52.
Allmaras, R.R., Kraft, J.M., Miller, D.E., 1988. Effects of soil compaction and incorporated crop residue on root health. Annu. Rev. Phytopathol. 26, 219–243.
Aviles, M., Borrero, M., Trillas, M.I., 2011. Review on compost as inducer of disease suppression in plants grown in soilless culture. Dyn. Soil Dyn. Plant 5, 1–11.
Bailey, K.L., Lazarovits, G., 2003. Suppressing soil-borne diseases with residue management and organic amendments. Soil Tillage Res. 72, 169–180.
Bedoussac, L., Justes, E., 2010. The efficiency of a durum wheat-winter pea intercrop to improve yield and wheat grain protein concentration depends on N availability during early growth. Plant Soil 330, 19–35.
Boehm, H., 2009. Körnerleguminosen - Stand des Wissens sowie zukünftiger Forschungsbedarf aus Sicht des Ökologishen Landbaus. J. Für Kult. 61, 324–331.
Boutler, J.I., Boland, G.J., Trevors, J.T., 2000. Compost: A study of the development process and end-product potential for suppression of turfgrass disease. World J. Microbiol. Biotechnol. 16, 115–134.
Cannon, R.J.C., 1998. The implications of predicted climate change for insect pests in the UK, with emphasis on non-indigenous species. Glob. Change Biol. 4, 785–796.
Chakraborty, S., Tiedemann, A.., Teng, P.., 2000. Climate change: potential impact on plant diseases. Environ. Pollut. 108, 317–326.
Charles, R., Gaume, A., Richthofen, J.-S. von, 2007. Auswertung des Körnerleguminosenanbaus durch die Produzenten. AGRARForschung 14, 300–305.
Coakley, S.M., Scherm, H., Chakraborty, S., 1999. Climate change and plant disease management. Annu. Rev. Phytopathol. 37, 399–426.
Corre-Hellou, G., Crozat, Y., 2005. N2 fixation and N supply in organic pea (Pisum sativum L.) cropping systems as affected by weeds and peaweevil (Sitona lineatus L.). Eur. J. Agron. 22, 449–458.
Crutzen, P.J., 2002. Geology of mankind. Nature 415, 23. Dick, W.A., Gregorich, E.G., 2004. Developing and maintaining soil organic matter levels., in:
Schjønning, P., Elmholt, S., Christensen, B.T. (Eds.), Managing Soil Quality: Challenges in Modern Agriculture. CABI, Wallingford, pp. 103–120.
Doran, J.W., 2002. Soil health and global sustainability: translating science into practice. Agric. Ecosyst. Environ., Soil Health as an Indicator of Sustainable Management 88, 119–127.
Doran, J.W., Sarrantonio, M., Liebig, M.A., 1996. Soil Health and Sustainability, in: Donald L. Sparks (Ed.), Advances in Agronomy. Academic Press, pp. 1–54.
Doran, J.W., Zeiss, M.R., 2000. Soil health and sustainability: managing the biotic component of soil quality. Appl. Soil Ecol., Special issue: Managing the Biotic component of Soil Quality 15, 3–11.
Eriksen, J., Thorup-Kristensen, K., 2002. The effect of catch crops on sulphate leaching and availability of S in the succeeding crop, in: Book of Abstracts. p. 98.
Fuchs, J.G., Thuerig, B., Brandhuber, R., Bruns, C., Finckh, M.R., Fließbach, A., Mäder, P., Schmidt, H., Vogt-Kaute, W., Wilbois, K.-P., Lucius, T., 2014. Evaluation of the causes of legume yield depression syndrome using an improved diagnostic tool. Appl. Soil Ecol. 79, 26–36.

GENERAL INTRODUCTION
8
Garrett, K.A., Dendy, S.P., Frank, E.E., Rouse, M.N., Travers, S.E., 2006. Climate Change Effects on Plant Disease: Genomes to Ecosystems. Annu. Rev. Phytopathol. 44, 489–509.
Hadar, Y., 2011. Suppressive compost: when plant pathology met microbial ecology. Phytoparasitica 39, 311–314.
Herrick, J.E., Wander, M.M., 1997. Relationships between soil organic carbon and soil quality in cropped and rangeland soils: the importance of distribution, composition, and soil biological activity. Soil Process. Carbon Cycle 405–425.
Hoitink, H.A.J., Boehm, M.J., 1999. Biocontrol within the context of soil microbial communities: a substrate-dependent phenomenon. Annu. Rev. Phytopathol. 37, 427–446.
Hoitink, H.A.J., Fahy, P.C., 1986. Basis for the control of soilborne plant pathogens with compost. Annu. Rev. Phytopathol. 24, 93–114.
Hoitink, H.A.J., Stone, A.G., Han, D.Y., 1997. Suppression of plant disease by composts. HortScience 32, 184–189.
IPCC, 2001. Climate change 2001: The scientific basis, Third report. Intergovernmental Panel on Climate Change.
Jarvis, A., Ramirez, J., Anderson, B., Leibing, C., Aggarwal, P., 2010. Scenarios of climate change within the context of agriculture, in: Reynolds, M.P. (Ed.), Climate Change and Crop Production. CABI.
Juroszek, P., Tiedemann, A. von, 2011. Potential strategies and future requirements for plant disease management under a changing climate. Plant Pathol. 60, 100–112.
Karpenstein-Machan, M., Stuelpnagel, R., 2000. Biomass yield and nitrogen fixation of legumes monocropped and intercropped with rye and rotation effects on a subsequent maize crop. Plant Soil 218, 215–232.
Katan, J., 1997. Soil disinfestation: environmental problems and solutions, in: Rosen, D., Tel-Or, E., Hadar, Y., Chen, Y. (Eds.), Modern Agriculture and the Environment, Developments in Plant and Soil Sciences. Springer Netherlands, pp. 41–45.
Lal, R., 2009. Soils and food sufficiency. A review. Agron. Sustain. Dev. 29, 113–133. Lal, R., 2008. Soils and sustainable agriculture. A review. Agron. Sustain. Dev. 28, 57–64 Langdale, G.W., Blevins, R.., Karlen, D.L., McCool, D.K., Nearing, M.A., Skidmore, E.L.,
Thomas, A.W., Tyler, D.D., Williams, J.R., 1991. Cover crop effects on soil erosion by wind and water, in: Cover Crops for Clean Water. Jackson, Tennessee, pp. 15–40.
Lichtfouse, E., Navarrete, M., Debaeke, P., Véronique, S., Alberola, C. (Eds.), 2009. Sustainable Agriculture. Springer Netherlands, Dordrecht.
Meisinger, J.J., Hargrove, W.L., Mikkelsen, R.L., Williams, J.R., Benson, V.W., 1991. Effects of cover crops on groundwater quality. Cover Crops Clean Water Soil Water Conserv. Soc. Ankeny Iowa 266, 793–799.
Mueller, T., Thorup-Kristensen, K., 2001. N-Fixation of Selected Green Manure Plants in an Organic Crop Rotation. Biol. Agric. Hortic. 18, 345–363.
Narawal, S.S., 2000. Allelopathic interactions in multiple cropping systems, in: Allelopathy in Ecological Agriculture and Forestry. pp. 141–157.
Pariaud, B., Ravigné, V., Halkett, F., Goyeau, H., Carlier, J., Lannou, C., 2009. Aggressiveness and its role in the adaptation of plant pathogens. Plant Pathol. 58, 409–424.
Parmesan, C., Yohe, G., 2003. A globally coherent fingerprint of climate change impacts across natural systems. Nature 421, 37–42.
Peoples, M.B., Herridge, D.F., Ladha, J.K., 1995. Biological nitrogen fixation: An efficient source of nitrogen for sustainable agricultural production? Plant Soil 174, 3–28.
Pflughöft, O., 2008. Pilzkrankheiten in Körnerfuttererbsen (Pisum sativum L.) – Diagnose, Epidemiologie, Ertragsrelevanz und Bekämpfung. Der Fakultät für Agrarwissenschaften der Georg-August-Universität Göttingen.
Pimentel, D., McNair, S., Janecka, J., Wightman, J., Simmonds, C., O’Connell, C., Wong, E., Russel, L., Zern, J., Aquino, T., Tsomondo, T., 2001. Economic and environmental threats of alien plant, animal, and microbe invasions. Agric. Ecosyst. Environ. 84, 1–20.

GENERAL INTRODUCTION
9
Pritchard, S.G., 2011. Soil organisms and global climate change. Plant Pathol. 60, 82–99. Recycled Organics Unit, 2006. Compost use for pest and disease suppression in NSW. The
University of New South Wales, Australia. Roldán, A., Caravaca, F., Hernández, M.T., Garcia, C., Sánchez-Brito, C., Velásquez, M.,
Tiscareño, M., 2003. No-tillage, crop residue additions, and legume cover cropping effects on soil quality characteristics under maize in Patzcuaro watershed (Mexico). Soil Tillage Res. 72, 65–73.
Root, T.L., Price, J.T., Hall, K.R., Schneider, S.H., Rosenzweig, C., Pounds, J.A., 2003. Fingerprints of global warming on wild animals and plants. Nature 421, 57–60.
Saeed, M.F., 2013. Seed health in organic peas and faba beans and management approaches to improve pea production in organic rotations. University of Kassel, Witzenhausen.
Sass, O., 2009. Marktsituation und züchterische Aktivitäten bein Ackerbohnen und Körnererbsen in der EU. J. Für Kult. 61, 306–308.
Schjønning, P., Elmholt, S., Christensen, B.T., 2004. Soil Quality Management - Concepts and Terms, in: Schjønning, P., Elmholt, S., Christensen, B.T. (Eds.), Managing Soil Quality: Challenges in Modern Agriculture. CABI Publishing, Wallingford, UK, pp. 1–16.
Schröter, D., Zebisch, M., Grothman, T., 2005. Climate change in Germany - Vulnerability and Adaptation to Climate-Sensitive Sectors, Klimastatusbericht.
Seybold, C.A., Herrick, J.E., Brejda, J.J., 1999. Soil resilience: a fundamental component of soil quality. Soil Sci. 164, 224–234.
Sturz, A.V., Christie, B.R., 2003. Rationale for an holistic approach to soil quality and crop health. Soil Tillage Res., Soil Agroecosystems: Impacts of Management on Soil Health and Crop Diseases 72, 105–106.
Tu, J.C., 1994. Effects of soil compaction, temperature, and moisture on the development of the Fusarium root rot complex of pea in southwestern Ontario. Phytoprotection 75, 125–131.
Urbatzka, P., Graß, R., Haase, T., Schüler, C., Trautz, D., Heß, J., 2011. Grain yield and quality characteristics of different genotypes of winter pea in comparison to spring pea for organic farming in pure and mixed stands. Org. Agric. 1, 187–202.
Walker, B., Holling, C.S., Carpenter, S.R., Kinzig, A., 2004. Resilience, adaptability and transformability in social–ecological systems. Ecol. Soc. 9, 5.
Walther, G.R., Post, E., Convey, P., Menzel, A., Parmesan, C., Beebee, T.J.C., Fromentin, J.-M., Hoegh-Guldberg, O., Bairlein, F., 2002. Ecological responses to recent climate change. Nature 416, 389–395.
Wardle, D.A., Bardgett, R.D., Klironomos, J.N., Setälä, H., van der Putten, W.H., Wall, D.H., 2004. Ecological linkages between aboveground and belowground biota. Science 304, 1629–1633.
West, J.S., Townsend, J.A., Stevens, M., Fitt, B.D.L., 2012. Comparative biology of different plant pathogens to estimate effects of climate change on crop diseases in Europe. Eur. J. Plant Pathol. 133, 315–331. doi:10.1007/s10658-011-9932-x
Wolfe, B.E., Klironomos, J.N., 2005. Breaking New Ground: soil communities and exotic plant invasion. BioScience 55, 477–487.

FUSARIUM IN THE CROP ROTATION AND YWC DISEASE SUPPRESSION IN THE FIELD
10
2. INCIDENCE OF FUSARIUM SPP. IN THE CROP ROTATION WINTER PEA – MAIZE – WINTER WHEAT AND THE POTENTIAL OF YARD WASTE COMPOST TO SUPPRESS FOOT AND ROOT ROT OF PEAS UNDER FIELD CONDITIONS
“If it is green, there is some Fusarium that can grow on it, in it, or with it”
Leslie and Summerel, 2006
Abstract
Field experiments were carried out from 2009 until 2013 to assess crop performance and pathogen occurrence in the rotation winter pea-maize-winter wheat and if the application of composts can improve system performance. The winter peas were left untreated or inoculated with Phoma medicaginis, in the presence or absence of yard waste compost at rate of 5 t dry matter ha-1. A second application of compost was made to the winter wheat. Fusarium ssp. were isolated and identified from the roots of all three crops. In addition, the Ascochyta complex pathogens on peas were identified. Overall crop performance of peas, maize and wheat and N dynamics were strongly affected by seasonal climatic conditions but not by experimental treatments. Dry summers impeded N-uptake by maize resulting in relatively high soil N contents in fall and consequent risk of leaching. Application of composts overall stabilized crop performance but it did not lead to yield increases nor did it affect pathogen composition and occurrence. Pathogen occurrence was highly variable across the experimental field and among years. Foot rot of peas was moderate in 2010 and 2011 and severe in 2012 when the crop failed. Five different pathogens were isolated and identified from symptomatic plants with Phoma medicaginis dominating. It appears that higher winter temperatures combined with lower rainfall favored P. medicaginis on peas over other pathogens in 2011 and 2012 in comparison to 2010. In 2012, surviving pea plants were severely affected by Fusarium avenaceum. More than 15 different Fusarium species were isolated from maize and wheat. Overall frequencies of Fusarium spp. on maize and winter wheat were highest in 2011. Dominating species on maize in all three years were F.
graminearum and F. culmorum. In addition, F. proliferatum, Microdochium nivale, F.
crookwellense, F. sambucinum, F. equiseti were frequently isolated species. The most frequently isolated species from winter wheat were F. oxysporum, F. culmorum, F.
avenaceum and M. nivale. The Fusarium species dominating on pea plants were F.
oxysporum, F. avenaceum and F. solani, and they were isolated most frequent in 2012. The Fusarium species occurring in the three subsequent crops did not correlate well. Suggesting factors other than the crops themselves influencing species dominance in a given season and crop.
Keywords: Fusarium spp., Ascochyta, Phoma medicaginis, winter peas, maize, wheat, yard waste compost

FUSARIUM IN THE CROP ROTATION AND YWC DISEASE SUPPRESSION IN THE FIELD
11
2.1 Introduction
As a result of the Renewable Energy Sources Act (EEG) of the German Government in
2009, biogas production gained high economic competitiveness. By 2012, this led to an
increase in silage maize production of nearly 60% compared with the year 2000 (Meyer and
Priefer, 2012). Maize production in organic agriculture has also increased in the last decade
although to a lesser extent. Organically produced maize is partly used for biogas production
but mainly as a protein and energy rich animal feed. High frequency of maize in the crop
rotation can lead to various environmental problems such as nitrogen leaching, soil erosion
and need for intensive and expensive weed control (Graß, 2003) and to an increase of
Fusarium head blight in wheat (West et al., 2012).
The “Double cropping” system (Zweikulturnutzungssystem) is a production system consisting
of a winter pea catch and cover crop before maize that was developed in order to optimize
benefits of growing a winter legume in the crop rotation and at the same time minimize the
potential negative aspects of growing maize (Graß, 2003). Winter peas are sown in
September or October, grown overwinter until the end of May and then either mulched or
incorporated into the soil as green manure or harvested for use as animal feed or biogas.
Winter soil cover provided by peas prevents soil erosion, reduces early weed infestation in
the spring and preserves winter moisture. After pea harvest, maize can be sown directly or
after shallow tillage. Maize utilizes the available N left after peas preventing it from leaching.
Due to the reduced tillage intensity production costs are also reduced. Maize is harvested at
the end of September or beginning of October and used either for silage or in biogas
production. Under temperate conditions, winter wheat is often the only crop that can be sown
after maize. With respect to soil and plant health the pea-maize-wheat rotation is a system
that might increase the risk of infection with Fusarium fungi and thus mycotoxin
contamination (Dill-Macky and Jones, 2000).
Fusarium head blight of wheat (FHB) and Fusarium ear rot of maize are economically
important diseases causing yield losses and losses in quality due to contamination of the
grains with mycotoxins (Vigier et al., 1997). Stalk and crown rot of maize and wheat are also
caused by Fusarium spp. and although of less economic importance, they are wide spread
wherever these crops are grown (Ares et al., 2004; Gatch and Munkvold, 2002; Osunlaja,
1990). In the case of severe infection and symptoms, foot rot of wheat can lead to lodging or
emergence of sterile ears leading to yield reduction (T. Pettitt et al., 1996). Infection of roots
and stem bases with Fusarium spp., also enhances later survival of the fungi in the soil and
plant residues and increases soil borne inoculum (Cotten and Munkvold, 1998; Leplat et al.,
2013; Suárez-Estrella et al., 2004).

FUSARIUM IN THE CROP ROTATION AND YWC DISEASE SUPPRESSION IN THE FIELD
12
As well ear as foot diseases are caused by a complex of different Fusarium spp. Fusarium
graminearum, F. culmorum, F. verticillioides, F.crookwellense, F. subglutinans, F.
avenaceum and F. equiseti are the most commonly isolated species from stalk rot of maize
in Europe (Scauflaire et al., 2011). Fusarium crown rot of wheat is commonly caused by F.
pseudograminearum, F. graminearum, F. culmorum, F. avenaceum and Microdochium nivale
(Backhouse, 2014; Khudhair et al., 2014; Liu et al., 2010; Melloy et al., 2014; Moya-Elizondo
et al., 2011; Poole et al., 2013). F. avenaceum and F. culmorum together with F. solani f. sp
pisi, F. oxysporum f.sp pisi, are part of the pathogen complex causing foot and root rot of
peas, limiting pea production worldwide (Ali et al., 1993; Bretag and Ramsey, 2001; Kraft et
al., 1998; Oyarzun, 1993).
Prevalence and dominance of different species in the above mentioned pathogen complexes
are affected by environmental conditions, tillage and cultural practices (Backhouse, 2014;
Gaurilckiene and Cesnuleviciene, 2013; Khudhair et al., 2014; Munkvold, 2003; Munkvold et
al., 1997; Persson et al., 1997). Most Fusarium spp. are soil and seed borne fungi, with little
host specification, usually with saprophytic and parasitic phases in their life cycle. Many are
able to produce survival structures like chlamydospores that enable long survival in the soil,
making control and management of root and stem rot diseases of pea, maize and wheat
difficult. In addition, lack of an effective management option for the control of the numerous
wide host range Fusarium spp. affecting pea, maize and wheat undermine the effectiveness
of the rotation. As the diseases are caused by a dynamic complex rather than one pathogen,
finding resistant cultivars is a challenge. Removal or incorporation of maize residues may
decrease the risk of Fusarium ear infection on subsequent wheat (Kong, 2014). However, the
retention of crop residues on or near the soil surface positively affects soil quality by
increasing the soil microbial biomass, increasing soil organic matter (SOM) content,
improving nutrient cycling and soil structure (Kong, 2014; Liu et al., 2010). Moreover, residue
incorporation requires several tillage operations increasing production costs.
An alternative way to suppress soil borne diseases is the enhancement of microbial activity
e.g. through the use of organic amendments (OA) (Bailey and Lazarovits, 2003; Bonanomi et
al., 2007; Bulluck III and Ristaino, 2002; Osunlaja, 1990). In a review of 250 articles
published between 1940 and 2006, in 45% of the cases OA suppressed disease, in 35% of
the cases there were no significant effects, while in 20% disease incidence was enhanced
(Bonanomi et al., 2007). Among the OA tested, composts were the most effective and
increase in disease incidence and/or severity was rarely observed.
In pot trials successful disease suppression usually follows application of 30 to 50% v/v of
compost to the potting mixes (Noble and Coventry, 2005; Termorshuizen et al., 2006). In
contrast to greenhouse trials, the amounts of compost that can be applied in the field are

FUSARIUM IN THE CROP ROTATION AND YWC DISEASE SUPPRESSION IN THE FIELD
13
considerably lower, e.g. in organic farming in Germany field application of off-farm compost
is limited to 5 t DM ha-1 and year. (Litterick et al., 2004) reviewed the literature about the role
of different uncomposted and composted organic materials in the reduction of pests and
diseases of different horticultural and agricultural crops. They suggested that besides
compost composition, application rates in the field are usually inadequate to achieve the
desired level of suppression. However, even at 5 t DM ha-1 of high quality yard waste
compost in the field can successfully reduce severity of Rhizoctonia solani on potatoes
(Bohne et al., 2013; Schulte-Geldermann et al., 2009). Field experiments with row
applications of compost and the highly susceptible pea variety Santana were variable in
outcome though (Bruns, 2013).
The field experiments described above were all based on one time application of compost to
the fields. In organic farming systems OA are typically applied on a regular basis, changing
overall microbial dynamics. We therefore set up field experiments with the rotation winter
pea-maize-wheat including a yearly application of compost. The two-year rotation was run
three times in the period between 2009 and 2013, with the main focus on identifying
Fusarium species present on the root and/or stem base of all three crops. In addition, the
potential of the yard waste compost (YWC) application to suppress foot and root rot of peas
and improve the system health and performance was investigated. The following questions
were addressed:
1) What are the effects of compost applications on pea and wheat foot and root
diseases in the rotation?
2) How does the spectrum of Fusarium spp. on the crops change in the course of the
rotation and are there effects of compost applications?

FUSARIUM IN THE CROP ROTATION AND YWC DISEASE SUPPRESSION IN THE FIELD
14
2.2 Material and Methods
2.2.1 Experimental site and setup
The field experiments were conducted on the experimental farm of the University of Kassel in
Neu Eichenberg, at 51°22’ N.L. and 9°54’ E.L, at an elevation of 247 m asl. The soil type is a
deep Haplic Luvisol from loess with 78 soil points (Wildhagen, 1998). Average annual
temperature is 7.9°C and average annual precipitation is 619 mm. The experimental farm is
managed organically since 1984.
The experiments took place within a crop rotation of six years consisting of two years grass-
clover followed by winter wheat a cover crop of winter pea – maize – winter wheat followed
by a spring cereal. Since 2005, each year all crops of the rotation are grown in large plots of
17.5 x 50 m resulting in a total of 24 plots arranged in four blocks of six plots. The sequence
starting with a winter pea cover crop until the end of the second wheat crop was studied
three times starting in 2009, 2010 and 2011 (Fig. 2.1). The winter pea variety EFB33, the
maize hybrid Fabregas and wheat variety Achat were used in all three years. In the second
experimental year due to lack of seeds, Fabregas was sown in the three middle rows of the
plots and hybrids Ronaldino and Atletico were used for the rest of plots. All the sampling and
analysis were done only on Fabregas.
Figure 2.1. Section of the crop rotation that was studied in the field experiment.
Within the large plots four treatments were arranged as plots of 7.5 x 20 m (Fig. 2.2). A total
of 5 t of dry matter ha-1 yard waste compost (YWC) was applied by hand two times in each
rotation. The first application was on the day before sowing peas and the second right before
wheat was sown (Fig 2.1). Nutrient contents and other properties of the YWC are listed in
Table 2.1.
C+ YWC 1 C+ YWC 2
C+ YWC 2 C+ YWC 3
C+ YWC 3 C+ YWC 4

FUSARIUM IN THE CROP ROTATION AND YWC DISEASE SUPPRESSION IN THE FIELD
15
Figure 2.2. Main plot divided in four experimental plots presenting four different treatments O=Control, I=Inoculated with P. medicaginis, C=Compost and I+C= Inoculated + Compost.
In addition to the compost application, half of the plots were inoculated with laboratory
prepared inoculum of Phoma medicaginis (Fig. 2.3). Peas were cut in May before maize was
sown. In 2010, plant material was removed; in 2011 all of the material was incorporated into
the top 5 cm of the soil during seed bed preparation for maize. In 2012, the peas failed due
to the severe frost. Maize was harvested in the second week of October, and the same plots
that had already received compost before peas received compost before wheat was sown
(Tab. 2.2). All field operations are listed in the Table 2.3.
I
I+C O
C
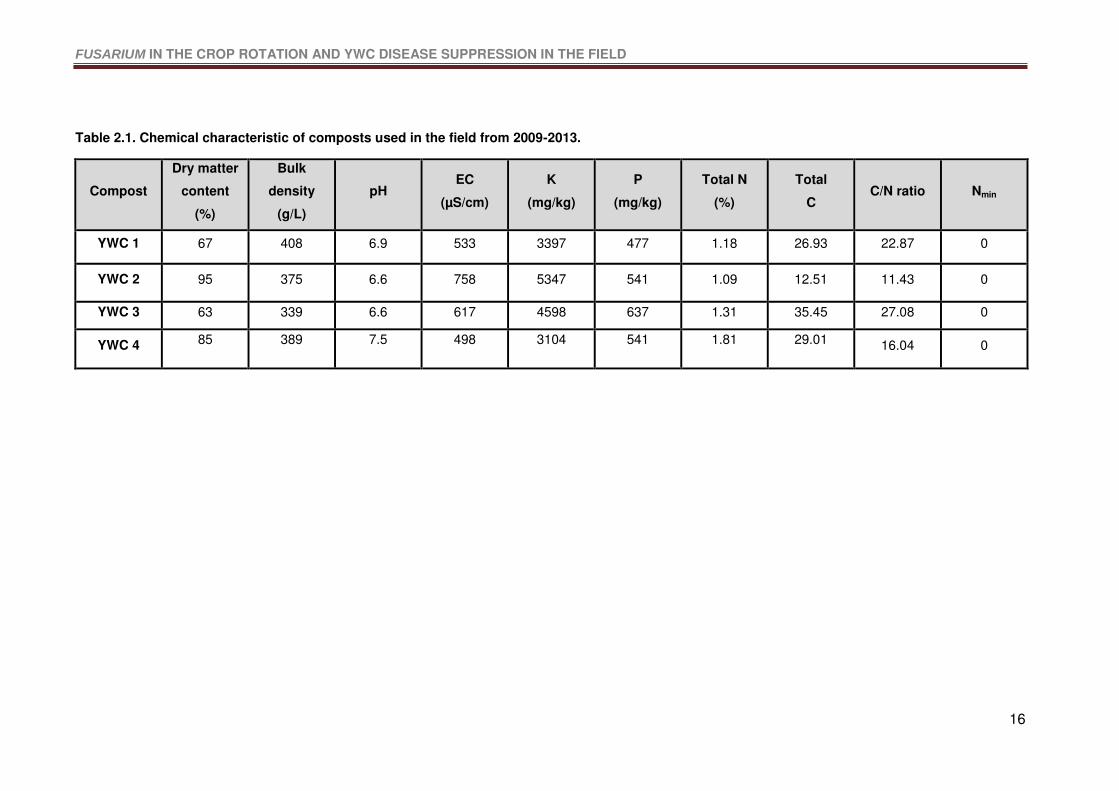
FUSARIUM IN THE CROP ROTATION AND YWC DISEASE SUPPRESSION IN THE FIELD
16
Table 2.1. Chemical characteristic of composts used in the field from 2009-2013.
Compost
Dry matter
content
(%)
Bulk
density
(g/L)
pH EC
(µS/cm)
K
(mg/kg)
P
(mg/kg)
Total N
(%)
Total
C C/N ratio Nmin
YWC 1 67 408 6.9 533 3397 477 1.18 26.93 22.87 0
YWC 2 95 375 6.6 758 5347 541 1.09 12.51 11.43 0
YWC 3 63 339 6.6 617 4598 637 1.31 35.45 27.08 0
YWC 4 85 389 7.5 498 3104 541 1.81 29.01 16.04 0

FUSARIUM IN THE CROP ROTATION AND YWC DISEASE SUPPRESSION IN THE FIELD
17
Table 2.2. Treatments applied in the field experiment.
Compost
application Inoculation Treatment name
Pea
- Compost - P. medicaginis Control (O)
+ P. medicaginis Inoculated (I)
+ Compost - P. medicaginis Compost (C)
+ P. medicaginis Inoculated + Compost (I+C)
Maize - Compost -* Control (O)
+ Compost - Compost (C)
Wheat - Compost - Control (O)
+ Compost - Compost (C)
*Compost was not directly applied before maize, but indicates plots that have received compost in the pea crop (see Fig. 2.1)
Inoculum preparation and inoculation
Inoculum of P. medicaginis, isolate Pm1, (Tab. 3.1, Chapter 3.2.1) was prepared on broken
oat grains. Approximately 40 kg of grains were soaked in water for 24 hours, autoclaved at
121°C for 15 minutes, and after another 24 hours autoclaved a second time. Agar plugs, 1
cm in diameter, of fungal culture from two fully covered Petri dishes were added and
incubated for two weeks at room temperature. During this time the material was regularly
stirred by hand to ensure homogenous fungal growth and prevent anaerobic conditions (Fig.
2.3). Inoculation in the field was done together with compost application by hand
broadcasting approximately 4 kg of inoculum per plot.
Figure 2.3. Inoculum of Phoma medicaginis grown on oat seeds.

FUSARIUM IN THE CROP ROTATION AND YWC DISEASE SUPPRESSION IN THE FIELD
18
Table 2.3. Timing of field operations.
Crop Rotation I
(2009/11)
Rotation II
(2010/12)
Rotation III
(2011/13)
Pea
Plough (15 cm)
Rotary disc
26.09.2009
29.09.2009
01.10.2010
04.10.2010
05.10.2011
05.10.2011
Compost application/ inoculation 28.09.2009 08.10.2010 17.10.2011
Sowing 29.09.2009 10.10. 2010 18.10.2011
Seed/m2 80 120 120
Emergence assessment 18.11.2009 26.11.2010 28.11.2011
Overwintering assessment 03.03.2010 05.04.2011 30.04.2012
Sampling for disease assessment
and pathogen ID
29.03.2010
18.05.2011
04.03.2011
18.05.2011
- 1
05.05.2012
Soil sampling for Nmin 19.03.2010
26.05.2010
05.04.2011
16.05.2011
26.04.2012
- 2
Harvest date 25.05.2010 18.05. 2011 No harvest 1
Maize
Plough (15 cm)
05.06.2010
18.05., 20.05.,
25.05.2011 09.05.2012
Sowing date 06.06.2010 25.05.2011 10.05.2012.
Sowing density in seed number/m2 11 11 16
Fertilization (hair pellets 14%N)
06.06.2010
56 kg N ha-1
in seed row
25.05.2011
56 kg N ha-1
in seed row
30.05.2012
42 kg N ha-1
broadcast
Weed control (hoe) 10.07.2010 29.06.2011 25.06.2012
Harvest date 15.10.2010 14.10.2011 03.10.2012.
Soil samples for Nmin - 3 15.10.2011 04.10.2012.
Wheat
Plough (15 cm)
Rotary disc
27.10.2010
28.10.2010
21.10.2011
23.10.2011
08.10.2012
11.10.2012
Compost application 28.10.2010 24.10.2011 11.10.2012
Sowing date 29.10.2010 25.10.2011 12.10.2012
Seed /m2 350 350 350
Sampling for pathogen ID 23.03.2011 26.03.2012 11.03.2013
Assessment foot disease 07.07.2011 16.07.2012 18.07.2013
Harvest date 29.07.2011 02.08.2012 15.08.2013
Soil samples for Nmin 02.08.2011 17.08.2012 22.08.2013
1 peas failed in 2011/12 due to severe frost
2 no early spring Nmin sampling was possible due to frozen soil
3 not measured

FUSARIUM IN THE CROP ROTATION AND YWC DISEASE SUPPRESSION IN THE FIELD
19
2.2.2 Assessments
Soil N-dynamics
Mineral nitrogen contents (Nmin) in soil depths of 0-30 cm and 30-60 cm were usually
measured in March in the peas, before and after maize and after wheat harvest (Tab. 2.3).
Ten soil cores were taken throughout each plot following a “W” sampling shape; bulk
samples were made and directly cooled down in the field. In the laboratory samples were
frozen at -18°C until analysis. Extraction and determination of Nmin was carried out according
to ÖNORM L-1091 (Österreichisches Normungsinstitut, 1999) and VDLUFA A6.1.4.1
(VDLUFA Methodenbuch, 1991). Extraction was done with 0.0125 M CaCl2 solution, and
measurement of NOx-N and NH4-N was done photometrically by absorption at 540 nm and
660 nm, respectively.
Plant development and yields
The number of pea and wheat plants was counted 2 to 4 weeks after sowing, depending on
the year. In addition, pea overwintering rate was recorded in early spring. The number of pea
plants per 0.1 m2 was counted in five randomly selected subplots per plot and average plant
density per m2 was calculated. For wheat, before harvest the number of head bearing tillers
was counted in five randomly chosen 1.1 m long rows and the mean density calculated.
Fresh mass of peas and weeds was measured right before mulching from three times 1 m2
per plot. After separation of weeds and peas they were dried at 105°C for 48 to 72 h for
determination of dry weights. Wheat was combine harvested and grain yield calculated
based on 14% moisture content and thousand grains weight determined. Maize was
harvested at BBCH 73-75 (Meier, 1997). Three 1 m row sections were cut by hand per plot,
number of plants counted and weighted. All harvested plants from one plot were afterwards
chopped into pieces of approximately 1-3 cm length and mixed well. Dry weights were
determined from subsamples of 300-500 g.
Disease assessments, pathogen isolation and identification
Pea plants were sampled two times for the assessment of root and foot rot severity and
pathogen identification. The first sampling was at BBCH stage 15/17, i.e. with five to seven
fully developed leaves (middle to end March) and the second at the beginning of flowering,
BBCH 59-61 (middle to end of May) (Tab. 2.3). Twenty plants together with roots were
selected following a “W” sampling route throughout each plot and taken to the laboratory for
the assessment and isolation of pathogens. Root and foot rot on peas was assessed using
the key of (Pflughöft, 2008) (Tab. 2.4). For pathogen isolations plants were surface
disinfected by placing them in 3% NaClO for 10 s. Afterwards they were thoroughly rinsed
under running distilled water and placed on filter paper under a laminar flow hood for 1 h to
dry. From the stem base and root of each plant three 1 cm pieces were cut, placed on
Coon’s agar media (Maltose 4 g, KNO3 2 g, MgSO4 1.20 g, KH2PO4 2.68 g, Agar 20 g , H2O

FUSARIUM IN THE CROP ROTATION AND YWC DISEASE SUPPRESSION IN THE FIELD
20
1L) and incubated for a week under UV light at 23 ± 3°C. After isolation, pathogenic fungi
belonging to Fusarium spp. and the Ascochyta complex, i.e. Ascochyta pisi, Phoma
medicaginis, Didymella pinodes, were identified to the species level. For that purpose the
one-week old mycelium from the original Coon’s plate was subcultured and incubated again
under the same conditions. Subculturing was done on half strength Potato Dextrose Agar
(PDA), for colony formation, and on Synthetic Nutrient Agar (SNA) medium (KH2PO4 1 g,
KNO3 1 g, MgSO4•7H2O 0.5 g, KCl 0.5 g, Glucose 0.2 g, Sucrose 0.2 g, Agar 20 g, H2O 1L)
for the sporulation of Fusarium spp., whereas for Ascochyta complex pathogens Coon’s agar
was used again. After 10 to 14 days pathogens were identified based on the colony
characteristic and microscopically based on the characteristics of spores (Leslie and
Summerell, 2006; Nasir et al., 1992).
At maize harvest, five to ten plants per plot were randomly selected and the lowest 20 cm of
the stem was cut, placed in paper bags and left to dry at room temperature. The sampling for
isolation and identification of Fusarium spp. on the stem basis and roots of wheat was done
in early spring in the seedling stage, BBCH 19-21. Ten to 15 plants per plot were taken to the
laboratory. At the late milk (BBCH 77) to early dough stage (BBCH 83) foot diseases on the
stem basis were assessed visually after removing all leaf materials. Symptoms caused by
the foot rot pathogens Fusarium spp., Pseudocercosporella herpotrichoides and Rhizoctonia
cerealis were assessed separately on the stem bases and severity scores from 0 to 3 were
given where 0 = no visible symptoms; 1 = less than 50% of the stem circumference affected;
2 = 50-100 % of stem circumference covered by a lesion; and 3 = stem is rotten or broken.
For Fusarium spp. isolation from wheat and maize, plants were washed with running tap
water for 1 h and placed under the laminar flow hood for 1 h to dry. Fusarium spp. selective
Pentachloronitrobenzene Peptone Agar (PPA) (Peptone 15 g, KH2PO4 1 g, MgSO4•7H2O 0.5
g, Pentachloronitrobenzene 750 mg, Agar 20 g, H2O 1 L) (Leslie and Summerell, 2006) was
used for the isolation. About 1 cm long plant pieces (three per plant) were placed on agar
media and incubated for 3 to 5 days under UV light at 23 ± 3°C, then subcultured on half
strength PDA and on SNA and again incubated for 10 to 14 days before identification as
described above.

FUSARIUM IN THE CROP ROTATION AND YWC DISEASE SUPPRESSION IN THE FIELD
21
Table 2.4. Scoring scheme for assessment of root and foot rot of peas (Pflughöft, 2008).
Score State of plant external tissues at root and stem
State of plant internal tissues at root and stem by cutting through the lesion
1 No visible symptoms No visible symptoms
2 Streaks at the transition zone or at epicotyl
or hypocotyl (not the discoloration of the seed and seed remaining attached)
Epidermis/rhizodermis is brownish to black
3 Brownish lesion covers <50% perimeter Brownish discoloration on cortical tissues
4 Brownish-black lesion covers >50%
perimeter Cortical tissues partially black, but center and
endodermis are still brownish or healthy 5 Black lesion covers 100% of stem Cortex tissue is completely black
6 Intensive color and black spread of lesion Cortex tissue begins to rot (bursting of
epicotyl or rhizodermis on the root)
7 Lesions spread up to the first lower leaf or
<3 cm on the tap root Cortex tissue is completely rotten
8 Lesions spread above the first lower leaf
and/or >3 cm on the tap root Shedding of the cortex tissue of endodermis
9 Dead plant Dead plant

FUSARIUM IN THE CROP ROTATION AND YWC DISEASE SUPPRESSION IN THE FIELD
22
2.3 Data processing and analysis As each pea plant was assessed with an external and internal score, a disease index was
calculated as the mean of external and internal lesion score and further analyses were done
on these values.
For foot rot of wheat a disease index (DI) based on a weighted mean was calculated as
follows:
�� = # ���� �� ��� + # ���� �� ��� ∗ + # ���� �� ℎ� ��� ∗n∗
where n= total number of assessed plants per sample
This results in DI values ranging between 0 and 100.
Data were analyzed with the statistical program R version 2.15.2. Analysis of Variance
(ANOVA) was conducted to analyze the effect of experimental factors on the plant growth
parameters, severity of disease and pathogen frequencies. Prior to ANOVA, data were
checked for normality with the Shapiro-Wilks-W-Test. Data for the pathogen incidence (%)
and DI were arcsine square root transformed to fulfil ANOVA requirements. Mean
separations were made by Tukey honest significant differences (HSD) test at P < 0.05.
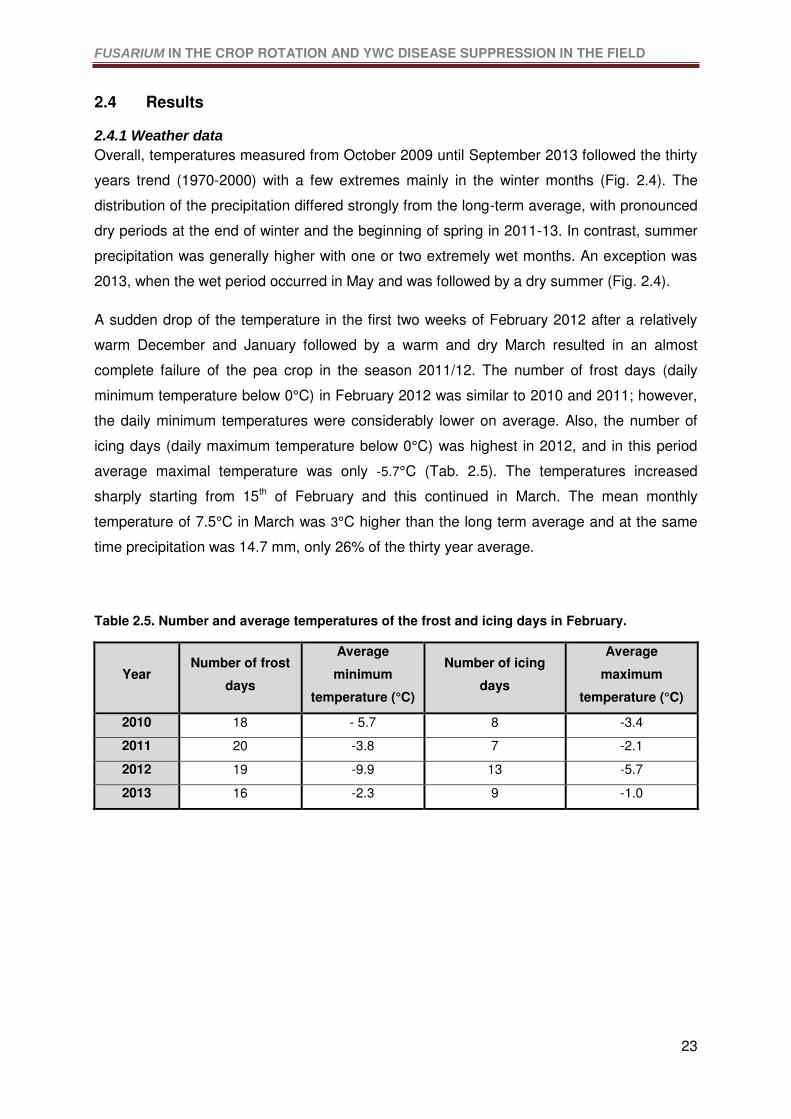
FUSARIUM IN THE CROP ROTATION AND YWC DISEASE SUPPRESSION IN THE FIELD
23
2.4 Results
2.4.1 Weather data
Overall, temperatures measured from October 2009 until September 2013 followed the thirty
years trend (1970-2000) with a few extremes mainly in the winter months (Fig. 2.4). The
distribution of the precipitation differed strongly from the long-term average, with pronounced
dry periods at the end of winter and the beginning of spring in 2011-13. In contrast, summer
precipitation was generally higher with one or two extremely wet months. An exception was
2013, when the wet period occurred in May and was followed by a dry summer (Fig. 2.4).
A sudden drop of the temperature in the first two weeks of February 2012 after a relatively
warm December and January followed by a warm and dry March resulted in an almost
complete failure of the pea crop in the season 2011/12. The number of frost days (daily
minimum temperature below 0°C) in February 2012 was similar to 2010 and 2011; however,
the daily minimum temperatures were considerably lower on average. Also, the number of
icing days (daily maximum temperature below 0°C) was highest in 2012, and in this period
average maximal temperature was only -5.7°C (Tab. 2.5). The temperatures increased
sharply starting from 15th of February and this continued in March. The mean monthly
temperature of 7.5°C in March was 3°C higher than the long term average and at the same
time precipitation was 14.7 mm, only 26% of the thirty year average.
Table 2.5. Number and average temperatures of the frost and icing days in February.
Year Number of frost
days
Average
minimum
temperature (°C)
Number of icing
days
Average
maximum
temperature (°C)
2010 18 - 5.7 8 -3.4
2011 20 -3.8 7 -2.1
2012 19 -9.9 13 -5.7
2013 16 -2.3 9 -1.0
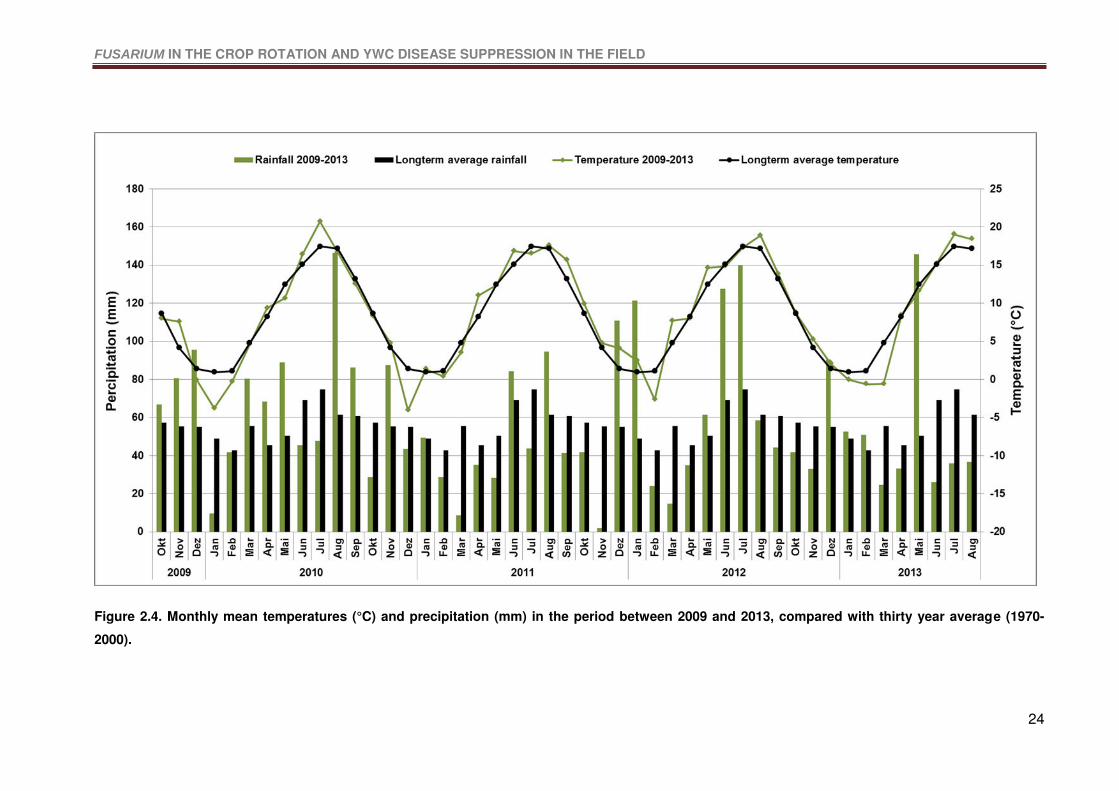
FUSARIUM IN THE CROP ROTATION AND YWC DISEASE SUPPRESSION IN THE FIELD
24
Figure 2.4. Monthly mean temperatures (°C) and precipitation (mm) in the period between 2009 and 2013, compared with thirty year average (1970-
2000).
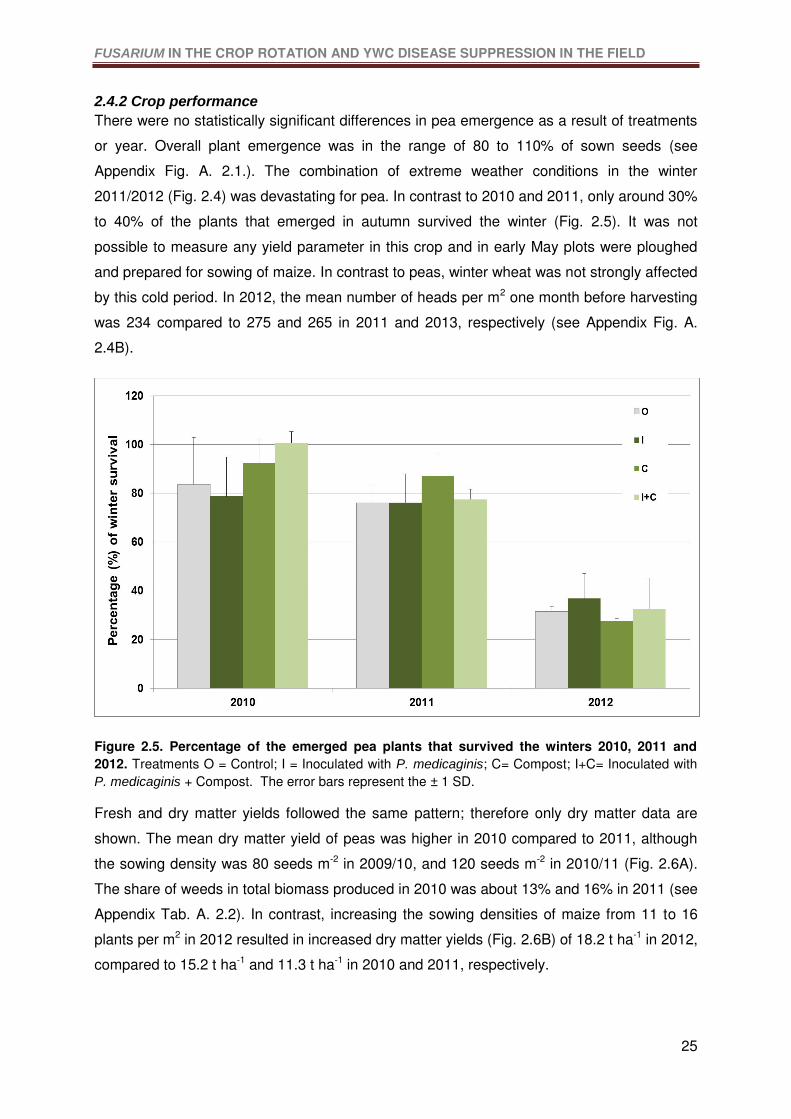
FUSARIUM IN THE CROP ROTATION AND YWC DISEASE SUPPRESSION IN THE FIELD
25
2.4.2 Crop performance
There were no statistically significant differences in pea emergence as a result of treatments
or year. Overall plant emergence was in the range of 80 to 110% of sown seeds (see
Appendix Fig. A. 2.1.). The combination of extreme weather conditions in the winter
2011/2012 (Fig. 2.4) was devastating for pea. In contrast to 2010 and 2011, only around 30%
to 40% of the plants that emerged in autumn survived the winter (Fig. 2.5). It was not
possible to measure any yield parameter in this crop and in early May plots were ploughed
and prepared for sowing of maize. In contrast to peas, winter wheat was not strongly affected
by this cold period. In 2012, the mean number of heads per m2 one month before harvesting
was 234 compared to 275 and 265 in 2011 and 2013, respectively (see Appendix Fig. A.
2.4B).
Figure 2.5. Percentage of the emerged pea plants that survived the winters 2010, 2011 and 2012. Treatments O = Control; I = Inoculated with P. medicaginis; C= Compost; I+C= Inoculated with P. medicaginis + Compost. The error bars represent the ± 1 SD.
Fresh and dry matter yields followed the same pattern; therefore only dry matter data are
shown. The mean dry matter yield of peas was higher in 2010 compared to 2011, although
the sowing density was 80 seeds m-2 in 2009/10, and 120 seeds m-2 in 2010/11 (Fig. 2.6A).
The share of weeds in total biomass produced in 2010 was about 13% and 16% in 2011 (see
Appendix Tab. A. 2.2). In contrast, increasing the sowing densities of maize from 11 to 16
plants per m2 in 2012 resulted in increased dry matter yields (Fig. 2.6B) of 18.2 t ha-1 in 2012,
compared to 15.2 t ha-1 and 11.3 t ha-1 in 2010 and 2011, respectively.

FUSARIUM IN THE CROP ROTATION AND YWC DISEASE SUPPRESSION IN THE FIELD
26
Figure 2.6. Dry matter yield of peas (A) and maize (B), and grain yield of wheat (14% moisture) (C). For peas O= control, I= inoculated with P. medicaginis, C= compost, I+C= inoculated with P.
medicaginis + compost. For maize and wheat C= with compost, O= without compost. Different small letters within one crop indicate significant differences in yield among years at P < 0.05 (Tukey’s HSD test). The error bars represent the ± 1 SD.
a b
A)
B)
C)
b c a
b b a

FUSARIUM IN THE CROP ROTATION AND YWC DISEASE SUPPRESSION IN THE FIELD
27
The weight per plant in 2012 was significantly lower than in 2010 when plant density had
been lower. In contrast, during the dry season 2011 plant weights were also lower than in
2010 despite the lower density (see Appendix Fig. A. 2.3). The years had statistically
significant effects on the grain yield of wheat (Fig. 6C) and thousand grain weight (TGW)
(see Appendix Fig. A. 2.4A). The highest yield of wheat was achieved in 2013 with 5.8 t ha-1
(TGW = 48 g) followed by 4.6 t ha-1 (TGW = 54 g) in 2011 and 4.3 t ha-1 (TGW = 50 g) in
2012.
Compost application did not have any statistically significant effect on yields of any of the
three crops in the rotation. Also, neither inoculation nor compost did have any measurable
effects on disease on peas. However, in 2009/10 performance of the peas varied among
main plots from 750 g m-2 fresh matter to 1800 g m-2. In the main plots with the lowest mean
yield the control yielded 930 g m-2 while the inoculated peas yielded only 212 g m-2. Plots that
had received compost did markedly better (Fig 2.7.) The differences were less marked for
the high yielding main plots. In 2011 overall yields were lower and less variable among plots.
Figure 2.7. Effect of compost application on pea fresh matter yield in May 2010 and 2011. Treatments: Control ; Inoculated ; Compost ; Inoculated + Compost .
Overall, the lowest contents of soil mineral N were measured in the first rotation sequence at
all sampling times (Fig. 2.8). In 2011 and 2012, relatively high amounts of mineral N were left
in the soil after maize was harvested, average 55 and 40 kg N ha-1, respectively, and about
30 kg N ha-1 after wheat in both rotations.
2010
2011
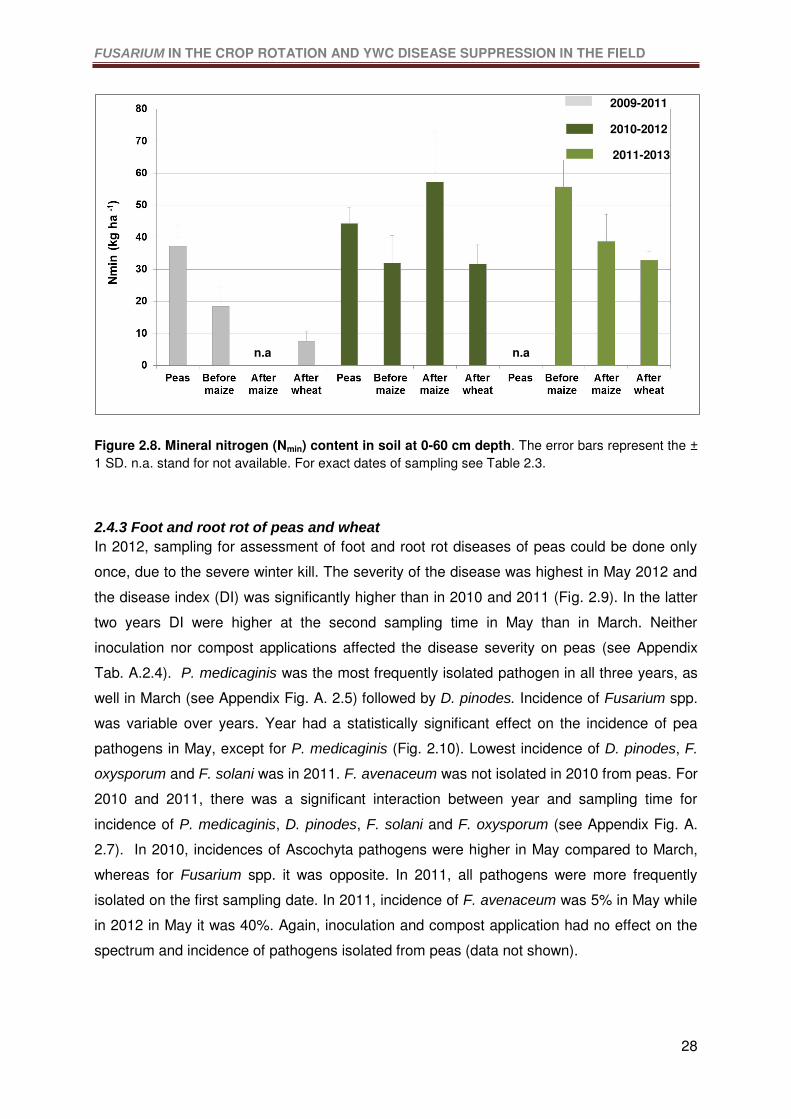
FUSARIUM IN THE CROP ROTATION AND YWC DISEASE SUPPRESSION IN THE FIELD
28
Figure 2.8. Mineral nitrogen (Nmin) content in soil at 0-60 cm depth. The error bars represent the ± 1 SD. n.a. stand for not available. For exact dates of sampling see Table 2.3.
2.4.3 Foot and root rot of peas and wheat
In 2012, sampling for assessment of foot and root rot diseases of peas could be done only
once, due to the severe winter kill. The severity of the disease was highest in May 2012 and
the disease index (DI) was significantly higher than in 2010 and 2011 (Fig. 2.9). In the latter
two years DI were higher at the second sampling time in May than in March. Neither
inoculation nor compost applications affected the disease severity on peas (see Appendix
Tab. A.2.4). P. medicaginis was the most frequently isolated pathogen in all three years, as
well in March (see Appendix Fig. A. 2.5) followed by D. pinodes. Incidence of Fusarium spp.
was variable over years. Year had a statistically significant effect on the incidence of pea
pathogens in May, except for P. medicaginis (Fig. 2.10). Lowest incidence of D. pinodes, F.
oxysporum and F. solani was in 2011. F. avenaceum was not isolated in 2010 from peas. For
2010 and 2011, there was a significant interaction between year and sampling time for
incidence of P. medicaginis, D. pinodes, F. solani and F. oxysporum (see Appendix Fig. A.
2.7). In 2010, incidences of Ascochyta pathogens were higher in May compared to March,
whereas for Fusarium spp. it was opposite. In 2011, all pathogens were more frequently
isolated on the first sampling date. In 2011, incidence of F. avenaceum was 5% in May while
in 2012 in May it was 40%. Again, inoculation and compost application had no effect on the
spectrum and incidence of pathogens isolated from peas (data not shown).
n.a n.a
2011-2013
2009-2011
2010-2012
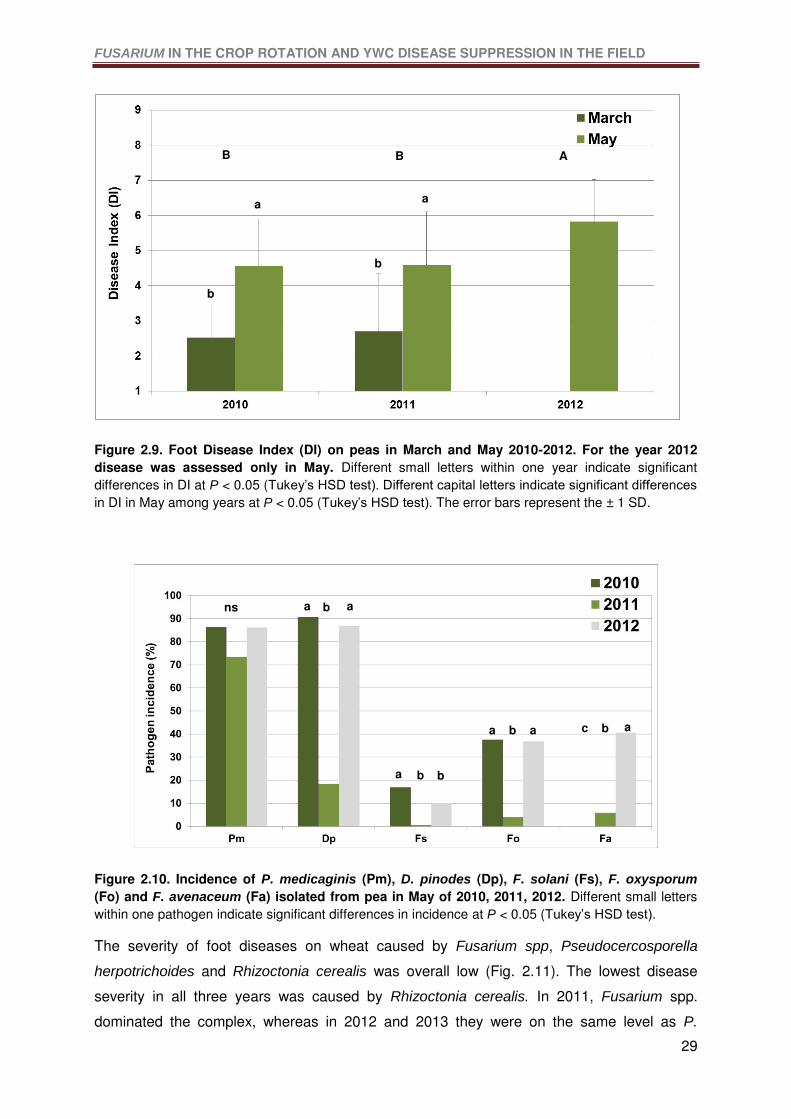
FUSARIUM IN THE CROP ROTATION AND YWC DISEASE SUPPRESSION IN THE FIELD
29
Figure 2.9. Foot Disease Index (DI) on peas in March and May 2010-2012. For the year 2012 disease was assessed only in May. Different small letters within one year indicate significant differences in DI at P < 0.05 (Tukey’s HSD test). Different capital letters indicate significant differences in DI in May among years at P < 0.05 (Tukey’s HSD test). The error bars represent the ± 1 SD.
Figure 2.10. Incidence of P. medicaginis (Pm), D. pinodes (Dp), F. solani (Fs), F. oxysporum (Fo) and F. avenaceum (Fa) isolated from pea in May of 2010, 2011, 2012. Different small letters within one pathogen indicate significant differences in incidence at P < 0.05 (Tukey’s HSD test).
The severity of foot diseases on wheat caused by Fusarium spp, Pseudocercosporella
herpotrichoides and Rhizoctonia cerealis was overall low (Fig. 2.11). The lowest disease
severity in all three years was caused by Rhizoctonia cerealis. In 2011, Fusarium spp.
dominated the complex, whereas in 2012 and 2013 they were on the same level as P.
ns
a b b
a a b c a b
a a b
b
a a
b
B B A
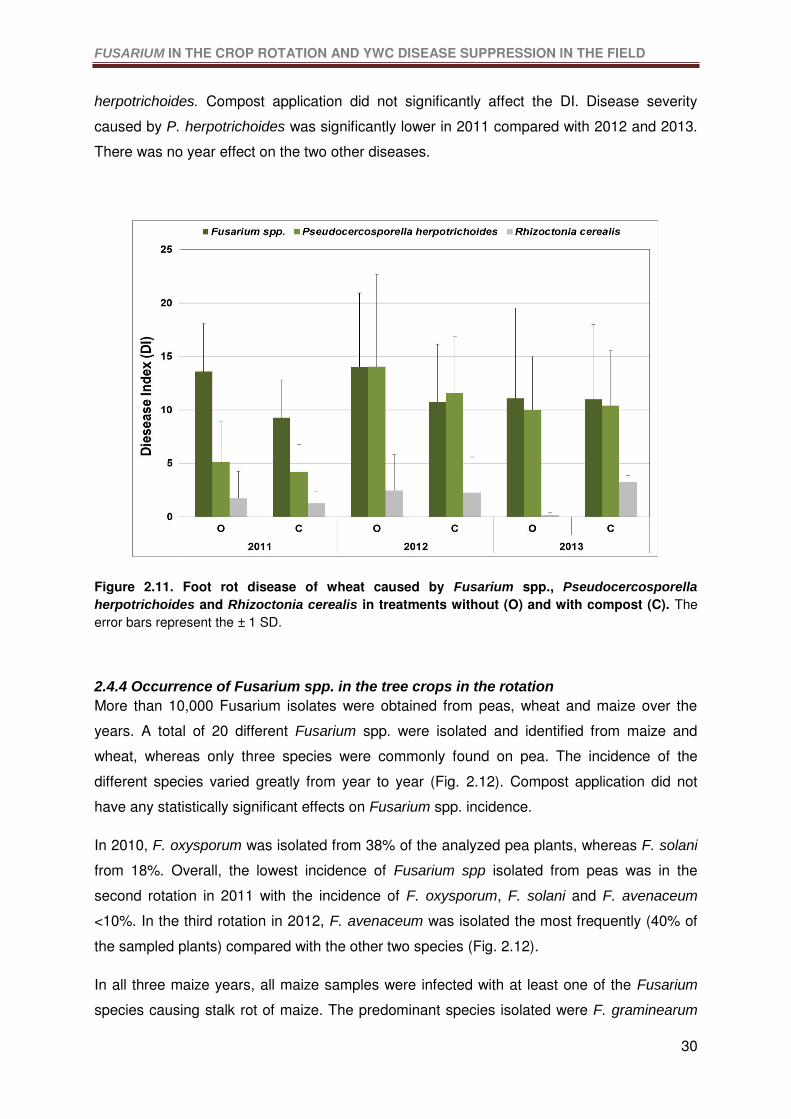
FUSARIUM IN THE CROP ROTATION AND YWC DISEASE SUPPRESSION IN THE FIELD
30
herpotrichoides. Compost application did not significantly affect the DI. Disease severity
caused by P. herpotrichoides was significantly lower in 2011 compared with 2012 and 2013.
There was no year effect on the two other diseases.
Figure 2.11. Foot rot disease of wheat caused by Fusarium spp., Pseudocercosporella
herpotrichoides and Rhizoctonia cerealis in treatments without (O) and with compost (C). The error bars represent the ± 1 SD.
2.4.4 Occurrence of Fusarium spp. in the tree crops in the rotation
More than 10,000 Fusarium isolates were obtained from peas, wheat and maize over the
years. A total of 20 different Fusarium spp. were isolated and identified from maize and
wheat, whereas only three species were commonly found on pea. The incidence of the
different species varied greatly from year to year (Fig. 2.12). Compost application did not
have any statistically significant effects on Fusarium spp. incidence.
In 2010, F. oxysporum was isolated from 38% of the analyzed pea plants, whereas F. solani
from 18%. Overall, the lowest incidence of Fusarium spp isolated from peas was in the
second rotation in 2011 with the incidence of F. oxysporum, F. solani and F. avenaceum
<10%. In the third rotation in 2012, F. avenaceum was isolated the most frequently (40% of
the sampled plants) compared with the other two species (Fig. 2.12).
In all three maize years, all maize samples were infected with at least one of the Fusarium
species causing stalk rot of maize. The predominant species isolated were F. graminearum

FUSARIUM IN THE CROP ROTATION AND YWC DISEASE SUPPRESSION IN THE FIELD
31
and F. culmorum (Fig. 2.12). Dominance among them varied over years. In the first rotation
in 2010, F. graminearum was isolated from nearly 50% of the samples and F. culmorum from
30%. In contrast, F. culmorum was isolated more frequently than F. graminearum, and in
2012 incidence of both pathogens was similar at about 40%. In 2012, with the exception of
those two species, overall incidence of Fusarium species isolated from maize was lowest
(Fig. 2.12). Microdochium nivale, F. proliferatum, F. oxysporum, F. sambucinum, F.
verticilioides and F. equiseti were commonly found on maize but at lower frequencies.
F. culmorum, F. oxysporum, F. avenaceum and F. equiseti were the most frequently isolated
species from wheat seedlings (Fig. 2.12). However, dominance and incidence of different
Fusarium spp. varied greatly among years. In the first rotation in 2011, incidence of all
isolated species was below 20% with dominance of F. proliferatum and F. oxysporum. In the
two subsequent years, incidence of F. aveanceum, F. culmorum, F. equiseti and M. nivale
increased reaching the maximum incidence in the last rotation sequence in 2013. There was
no obvious relationship between Fusarium communities found on maize and wheat neither
within year nor within rotation (Tab. 2.6) For example, incidence of F. graminearum on maize
was high and did not vary statistically significantly among years. On the following wheat it
was much lower and differences among years were statistically significant. F. crookwellense
occurred always in maize but only in 2013 on wheat. F. avenaceum was more prominent on
maize (15%) than on wheat (2.5%) in 2010/11, while in 2012/13 it was low on maize (5%)
and high on wheat (33%) (Tab. 2.6).

FUSARIUM IN THE CROP ROTATION AND YWC DISEASE SUPPRESSION IN THE FIELD
32
Figure 2.12. Incidence of Fusarium spp. isolated from three crops in the rotation. WP = winter peas, M = maize and WW = winter wheat; time of samplings, month/year, is given in the brackets.
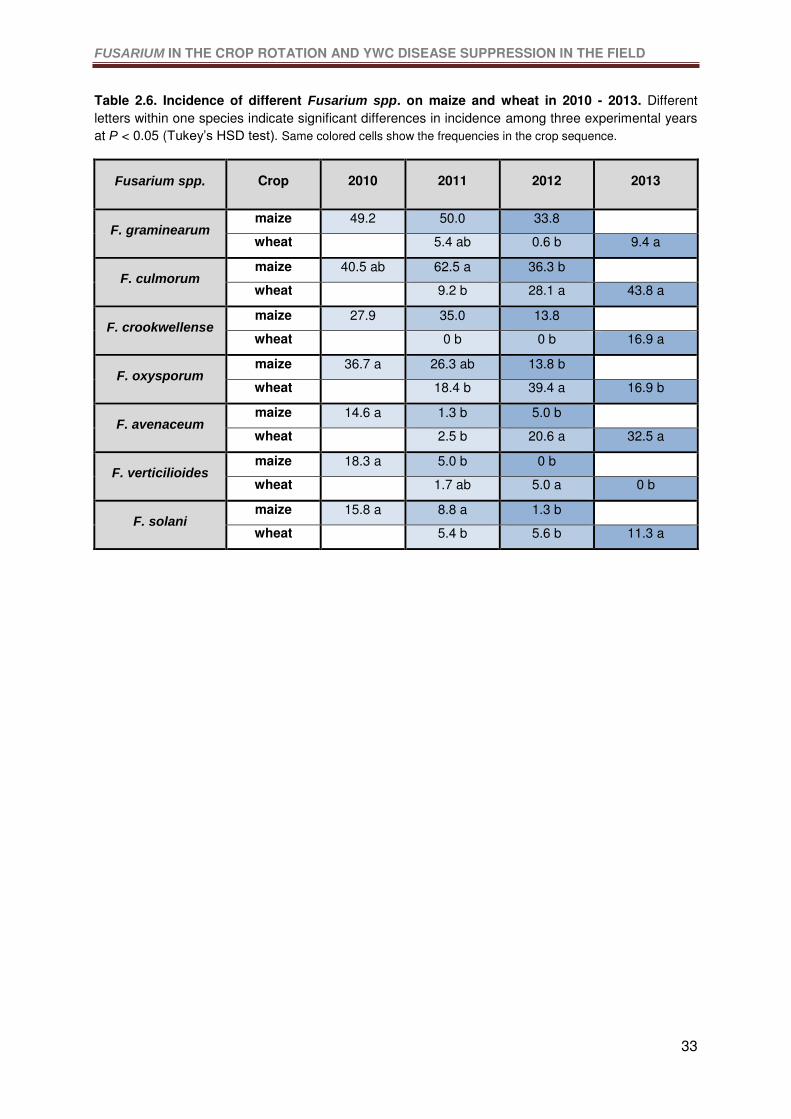
FUSARIUM IN THE CROP ROTATION AND YWC DISEASE SUPPRESSION IN THE FIELD
33
Table 2.6. Incidence of different Fusarium spp. on maize and wheat in 2010 - 2013. Different letters within one species indicate significant differences in incidence among three experimental years at P < 0.05 (Tukey’s HSD test). Same colored cells show the frequencies in the crop sequence.
Fusarium spp. Crop 2010 2011 2012 2013
F. graminearum maize 49.2 50.0 33.8
wheat
5.4 ab 0.6 b 9.4 a
F. culmorum maize 40.5 ab 62.5 a 36.3 b
wheat
9.2 b 28.1 a 43.8 a
F. crookwellense maize 27.9 35.0 13.8
wheat
0 b 0 b 16.9 a
F. oxysporum maize 36.7 a 26.3 ab 13.8 b
wheat
18.4 b 39.4 a 16.9 b
F. avenaceum maize 14.6 a 1.3 b 5.0 b
wheat
2.5 b 20.6 a 32.5 a
F. verticilioides maize 18.3 a 5.0 b 0 b
wheat
1.7 ab 5.0 a 0 b
F. solani maize 15.8 a 8.8 a 1.3 b
wheat
5.4 b 5.6 b 11.3 a

FUSARIUM IN THE CROP ROTATION AND YWC DISEASE SUPPRESSION IN THE FIELD
34
2.5 Discussion Overall crop performance of peas, maize and wheat was strongly affected by seasonal
climatic conditions but not by experimental treatments. As a result of drought in the summer
2011 yield of the maize was relatively low. Plants were not able to utilize available nitrogen
and high amounts were left in the soil after maize harvest. In the rotation with wheat following
maize, high nitrogen contents in the soil in the autumn pose an increased risk of leaching
losses as the demands of wheat for N in the early growth stages are relatively low. Excessive
N in late fall in wheat production may lead to N leaching to ground water, as wheat does not
take up much N before winter (Brown and Petrie, 2006).
Application of yard waste compost overall did not reduce the severity of foot rot of peas or
wheat in our study. Bruns et al. (2013) showed that concentrated application of compost in
the seed row together with pea seeds can increase efficiency of compost in suppressing root
rot on the susceptible spring variety Santana. Application of compost had a positive effect on
the pea biomass especially in the low performing plots in the trial in 2010. The winter and
early spring 2009/10 were unusually wet compared to the long-term average and also
considerably wetter than in the following years. The low productivity of peas in one of main
plots (Fig 2.7.) was most likely due to water logging that was apparent in this part of the field
but not in the others. It seems that under these conditions the inoculated peas did especially
poorly and that compost application helped to improve the situation.
Foot rot of peas was moderate in 2010 and 2011 and severe in 2012 when the crop failed.
Five different pathogens were isolated and identified from symptomatic plants with Phoma
medicaginis dominating. In most of the cases foot rot of peas is caused by mixed infections
with Ascochyta complex pathogens and several Fusarium spp. (Ali et al., 1993; Gaurilckiene
and Cesnuleviciene, 2013; Kraft et al., 1998; Oyarzun, 1993; Persson et al., 1997) and
dominance and importance of single pathogens in the complex vary greatly depending on
location, climate, and agricultural practice (Gaurilckiene and Cesnuleviciene, 2013; Jensen et
al., 2004; Persson et al., 1997). Severe foot rot in 2012 occurred on the few remaining plants
after the frost in early February. Especially the incidence of F. avenacuem increased
significantly from <10% in the previous years to 40% (Fig. 2.12). (Bateman et al., 1997)
found that poor survival of lupines in the UK was due to inadequate cold tolerance followed
by fungal infestation. F. avenaceum is a wide host range pathogen that can survive
saprotrophically in the soil. In a healthy and active soil, it is usually not posing a problem, as
it is suppressed by fast growing soil saprophytes (Fletcher et al., 1991). However, plants
under environmental stress are more susceptible to this opportunistic pathogen (Leslie et al.,
1990). F. avenaceum is together with F. tricinctum, F. poae, F. culmorum and F.
graminearum part of the complex causing Fusarium Head Blight (FHB) of small grain cereals

FUSARIUM IN THE CROP ROTATION AND YWC DISEASE SUPPRESSION IN THE FIELD
35
in Europe. Moreover, it has been shown that in recent years in Northern Europe F.
avenaceum is the dominating species of the complex (Uhlig et al., 2007). In our study, F.
avenaceum was isolated from all three crops in the crop rotation. Beside causing wilting and
rot symptoms in infected plants, F. avenaceum produces mycotoxins moniliformin, enniatin,
beauvericin and zearalenone (Bottalico, 1998; Langseth, 1998; Logrieco et al., 2002) that are
harmful for animals and humans. Mycotoxin production of fungi that cause diseases on roots
and stem bases of plants is usually not of a great concern, because they stay in the field as
plant residues after harvest. However, in the case of F. avenaceum the lack of host
specialization and saprophytic survival on plant residues increases the risk of building up
inoculum that can lead to subsequent infection and mycotoxin contamination of cereals.
More than fifteen Fusarium species were isolated from maize and wheat in our study and
most of them are mycotoxin producers. The spectrum of species found did not differ among
years; however, the incidence of different species of the complex did. In all three seasons F.
graminearum and F. culmorum dominated the pathogen complex on maize. They were found
on 28 to 62% of the sampled plants, depending on the year. These results are in agreement
with findings of (Scauflaire et al., 2011) who reported these species dominating in the
pathogen complex causing stalk rot of maize in Northern Europe. F. graminearum and F.
culmorum were also found on wheat seedlings but at much lower frequencies and the
dominance was neither connected to the Fusarium spp. in the preceding maize crop nor to
the communities found on maize in the same year (Fig. 2.12, Tab. 2.6). F. culmorum was
always isolated at higher frequencies than F. graminearum from wheat seedlings. In all three
rotations, the frequency of F. graminearum strongly decreased from maize to wheat. This
species was more often associated with FHB in Australia and North America, whereas stem-
base disease of wheat is mainly related with F. culmorum. F. culmorum has been dominating
the Fusarium complex on wheat in the cooler and humid conditions of Northern Europe and
America (Backhouse et al., 2001; Poole et al., 2013). These results are confirmed by our
data.
Maize residues are considered to be an important source of inoculum for the infection of
wheat ears with FHB pathogens (Osborne and Stein, 2007). The low incidence (< 10%) of F.
graminearum on wheat seedlings in our study should not be interpreted as an indicator of low
risk for FHB later in the season or as an indicator of low inoculum potential of maize
residues. It is known that growth and sporulation of F. graminearum as well as its teleomorph
Gibberella zeae is favored by warm and humid conditions (Melloy et al., 2014; Pettitt et al.,
2003; T. R. Pettitt et al., 1996). The optimum temperature for mycelial growth and sporulation
is 25°C, and growth of the fungus stops below 5°C (Leplat et al., 2013). Therefore, the low
level of infestation of wheat seedlings with F. graminearum in early spring is likely the result

FUSARIUM IN THE CROP ROTATION AND YWC DISEASE SUPPRESSION IN THE FIELD
36
of reduced activity of the fungi due to low temperatures rather than low inoculum potential.
The role of maize residues and the level of risk it poses for later infection of wheat ears are
highly dependent on a combination of agronomic practices and climatic conditions at the time
of anthesis (Kong, 2014; Munkvold, 2003).
The other species found on maize, F. sambucinum, F. crookwellense, F. equisety, F.
proliferatum and F. verticilioides have been reported as part of the pathogen complex
causing maize stalk rot worldwide (Cotten and Munkvold, 1998; Leslie et al., 1990; Leslie
and Summerell, 2006; Osborne and Stein, 2007; Scauflaire et al., 2011). The spectrum of
Fusarium species on wheat seedlings was also diverse and variable over years. Besides the
above mentioned F. avenaceum and F. culmorum, F. oxysporum, F. equiseti and M. nivale
were part of the pathogen complex on wheat seedlings. In our study, F. equiseti was found at
relatively high frequencies in the second and third rotation sequence on both maize and
wheat. It is a cosmopolitan soil-borne species that was for a long time considered a
saprophyte or weak pathogen of different plants (Gerlach and Nirenberg, 1982). However, F.
equiseti has been confirmed as a pathogen causing stalk rot of maize (Leslie and Summerell,
2006), root rot and FHB of wheat (Bottalico, 1998; Logrieco et al., 2002), blights and pre
emergence damping-off of cucurbits (Palmero et al., 2011) and tomato (Palmero et al.,
2011). Although it is usually associated with warmer areas, in the last decade it has
commonly been part of the Fusarium complex in Northern Europe and America (Leslie and
Summerell, 2006).
It is common that many different Fusarium species occur simultaneously in plants. Overall
diversity as well as relative prevalence of species in specific regions of the world is affected
by cultural practices, plant cultivars and weather patterns (Gatch and Munkvold, 2002;
Munkvold, 2003; Osunlaja, 1990; Scauflaire et al., 2011). The changing climate with more
frequent extreme weather events brings new complexity into an already complicated system.
General increase or decrease in temperatures and precipitation will affect certain Fusarium
species adversely and favor others. However, this should not be observed outside of the
existing agricultural systems that will also have to change as a result of climate change.
Changes in tillage practices, sowing times, crop and cultivar choice among others, make the
prediction of disease risk and pathogens associated with climate change difficult. Current
expansion of maize production in Germany is likely to increase the risk of damage caused by
Fusarium spp. and their mycotoxins in crop rotation as many of the species have a wide host
range (Meissle et al., 2009). Our results emphasize the importance of further research on the
effect of environmental and cultural practices on the Fusarium complex. This may include the
optimization of compost applications or other organic amendments in order to maximize
suppressiveness to soil-borne diseases.

FUSARIUM IN THE CROP ROTATION AND YWC DISEASE SUPPRESSION IN THE FIELD
37
2.6 References Ali, S.M., Sharma, B., Ambrose, M.J., 1993. Current status and future strategy in breeding
pea to improve resistance to biotic and abiotic stresses. Euphytica 73, 115–126. Ares, A.J.L., Ferro, A.R.C., Ramírez, C.L., González, M.J., 2004. Short communication.
Fusarium graminearum Schwabe, a maize root and stalk rot pathogen isolated from lodged plants in northwest Spain. Span. J. Agric. Res. 2, 249–252.
Backhouse, D., 2014. Modelling the behaviour of crown rot in wheat caused by Fusarium pseudograminearum. Australas. Plant Pathol. 43, 15–23.
Backhouse, D., Burgess, L.W., Summerell, B.A., 2001. Biogeography of Fusarium, in: Fusarium. Paul E. Nelson Memorial Symposium. APS Press, Bethesda, pp. 122–137.
Bailey, K.L., Lazarovits, G., 2003. Suppressing soil-borne diseases with residue management and organic amendments. Soil Tillage Res. 72, 169–180.
Bateman, G.L., Ferguson, A.W., Shield, I., 1997. Factors affecting winter survival of the florally determinate white lupin cv. Lucyane. Ann. Appl. Biol. 130, 349–359.
Bohne, B., Hensel, O., Bruns, C., 2013. Reihenapplikation von Komposten zur Kontrolle bodenbürtiger Krankheiten – technische Lösungen für Kartoffeln und Körnerleguminosen.
Bonanomi, G., Antignani, V., Pane, C., Scala, F., 2007. Suppression of soilborne fungal diseases with organic amendments. J. Plant Pathol. 89, 311–324.
Bottalico, A., 1998. Fusarium diseases of cereals: species complex and related mycotoxin profiles, in Europe. J. Plant Pathol. 85–103.
Bretag, T.W., Ramsey, M., 2001. Foliar diseases caused by fungi. Ascochyta spp., in: Compendium of Pea Diseases and Pests. Second Edition. APS Press, The American Phytopathological Society, St. Paul Minnesota, pp. 24–28.
Brown, B.D., Petrie, S., 2006. Irrigated hard winter wheat response to fall, spring, and late season applied nitrogen. Field Crops Res. 96, 260–268.
Bulluck III, L.R., Ristaino, J.B., 2002. Effect of synthetic and organic soil fertility amendments on southern blight, soil microbial communities, and yield of processing tomatoes. Phytopathology 92, 181–189.
Cotten, T.K., Munkvold, G.P., 1998. Survival of Fusarium moniliforme, F. proliferatum, and F. subglutinans in maize stalk residue. Phytopathology 88, 550–555.
Dill-Macky, R., Jones, R.K., 2000. The effect of previous crop residues and tillage on Fusarium head blight of wheat. Plant Dis. 84, 71–76.
Fletcher, J.D., Broadhurst, P.G., Bansal, R.K., 1991. Fusarium avenaceum: A pathogen of lentil in New Zealand. N. Z. J. Crop Hortic. Sci. 19, 207–210.
Gatch, E.W., Munkvold, G.P., 2002. Fungal species composition in maize stalks in relation to European corn borer injury and transgenic insect protection. Plant Dis. 86, 1156–1162.
Gaurilckiene, I., Cesnuleviciene, R., 2013. The susceptibility of pea (Pisum sativum L.) to Ascochyta blight under Lithuanian conditions. Zemdirb.-Agric. 100, 283–288.
Gerlach, W., Nirenberg, H., 1982. The genus Fusarium -- a pictorial atlas. 406 pp. Graß, R., 2003. Direkt- und Spätsaat von Silomais - Ein neues Anbausystem zur
Reduzierung von Umweltgefährdungen und Anbauproblemen bei Optimierung der Erträge (Dissertation). University of Kassel, Witzenhausen.
Jensen, B., Bødker, L., Larsen, J., Knudsen, J.C., Jørnsgaard, B., 2004. Specificity of soil-borne pathogens on grain legumes. Conference paper available at: http://orgprints.org/3522/
Khudhair, M., Melloy, P., Lorenz, D.J., Obanor, F., Aitken, E., Datta, S., Luck, J., Fitzgerald, G., Chakraborty, S., 2014. Fusarium crown rot under continuous cropping of susceptible and partially resistant wheat in microcosms at elevated CO2. Plant Pathol. 1–10.
Kong, L., 2014. Maize residues, soil quality, and wheat growth in China. A review. Agron. Sustain. Dev. 34, 405–416.
Kraft, J.M., Dunne, B., Goulden, D., Armstrong, S., 1998. A search for resistance in peas to Mycosphaerella pinodes. Plant Dis. 82, 251–253.

FUSARIUM IN THE CROP ROTATION AND YWC DISEASE SUPPRESSION IN THE FIELD
38
Langseth, W., 1998. Mycotoxin production and cytotoxicity of Fusarium strains isolated from Norwegian cereals. Mycopathologia 144, 103–113.
Leplat, J., Friberg, H., Abid, M., Steinberg, C., 2013. Survival of Fusarium graminearum, the causal agent of Fusarium head blight. A review. Agron. Sustain. Dev. 33, 97–111.
Leslie, J.F., Pearson, C.A.S., Nelson, P.E., Toussoun, T.A., 1990. Fusarium spp. from Corn, Sorghum, and Soybean Fields in the Central and Eastern United States. Phytopathology 80, 343–350.
Leslie, J.F., Summerell, B.A., 2006. The Fusarium laboratory manual. Blackwell Pub., Ames, Iowa.
Litterick, A.M., Harrier, L., Wallace, P., Watson, C.A., Wood, M., 2004. The role of uncomposted materials, composts, manures and compost extracts in reducing pest and disease incidence and severity in sustainable temperate agricultural and horticultural crop production – a review. Crit. Rev. Plant Sci. 23, 453–479.
Liu, Y.X., Yang, X.M., Ma, J., Wei, Y.M., Zheng, Y.L., Ma, H.X., Yao, J.B., Yan, G.J., Wang, Y.G., Manners, J.M., 2010. Plant height affects Fusarium crown rot severity in wheat. Phytopathology 100, 1276–1281.
Logrieco, A., Rizzo, A., Ferracane, R., Ritieni, A., 2002. Occurrence of beauvericin and enniatins in wheat affected by Fusarium avenaceum head blight. Appl. Environ. Microbiol. 68, 82–85.
Meissle, M., Mouron, P., Musa, T., Bigler, F., Pons, X., Vasileiadis, V.P., Otto, S., Antichi, D., Kiss, J., Pálinkás, Z., Dorner, Z., Van Der Weide, R., Groten, J., Czembor, E., Adamczyk, J., Thibord, J.-B., Melander, B., Nielsen, G.C., Poulsen, R.T., Zimmermann, O., Verschwele, A., Oldenburg, E., 2009. Pests, pesticide use and alternative options in European maize production: current status and future prospects: Pest management in European maize production. J. Appl. Entomol. 134, 357–375.
Melloy, P., Aitken, E., Luck, J., Chakraborty, S., Obanor, F., 2014. The influence of increasing temperature and CO2 on Fusarium crown rot susceptibility of wheat genotypes at key growth stages. Eur. J. Plant Pathol. 140, 19–37.
Meyer, R., Priefer, C., 2012. Ökologischer Landbau und Bioenergieerzeugung–Zielkonflikte und Lösungsansätze. Arbeitsbericht.
Moya-Elizondo, E.A., Rew, L.J., Jacobsen, B.J., Hogg, A.C., Dyer, A.T., 2011. Distribution and prevalence of Fusarium crown rot and common root rot pathogens of wheat in Montana. Plant Dis. 95, 1099–1108.
Munkvold, G.., 2003. Epidemiology of Fusarium diseases and their mycotoxins in maize ears. Eur. J. Plant Pathol. 109, 705–713.
Munkvold, G.P., Hellmich, R.L., Showers, W.B., 1997. Reduced Fusarium ear rot and symptomless infection in kernels of maize genetically engineered for European corn borer resistance. Phytopathology 87, 1071–1077.
Nasir, M., Hoppe, H.-H., Ebrahim-Nesbat, F., 1992. The development of different pathotype groups of Mycosphaerella pinodes in susceptible and partially resistant pea leaves. Plant Pathol. 41, 187–194.
Noble, R., Coventry, E., 2005. Suppression of soil-borne plant diseases with composts: A review. Biocontrol Sci. Technol. 15, 3–20.
Osborne, L.E., Stein, J.M., 2007. Epidemiology of Fusarium head blight on small-grain cereals. Int. J. Food Microbiol., Mycotoxins from the Field to the Table 119, 103–108.
Österreichisches Normungsinstitut, 1999. ÖNORM L 1091: Chemische Bodenuntersuchungen - Bestimmung von mineralischem Stickstoff - Nmin-Methode.
Osunlaja, S.O., 1990. Effect of organic soil amendments on the incidence of stalk rot of maize. Plant Soil 127, 237–241.
Oyarzun, P.J., 1993. Bioassay to assess root rot in pea and effect of root rot on yield. Neth. J. Plant Pathol. 99, 61–75.
Palmero, D., Cara, M. de, Iglesias, C., Gálvez, L., Tello, J.C., 2011. Comparative study of the pathogenicity of seabed isolates of Fusarium equiseti and the effect of the composition of the mineral salt medium and temperature on mycelial growth. Braz. J. Microbiol. 42, 948–953.

FUSARIUM IN THE CROP ROTATION AND YWC DISEASE SUPPRESSION IN THE FIELD
39
Persson, L., Bodker, L., Larsson-Wikström, M., 1997. Prevalence and pathogenicity of foot and root rot pathogens of pea in south Scandinavia. Plant Dis. 81, 171–174.
Pettitt, T., Parry, D., Polley, R., 1996. Effect of temperature on the incidence of nodal foot rot symptoms in winter wheat crops in England and Wales caused by Fusarium culmorum and Microdochium nivale. Agric. For. Meteorol. 79, 233–242.
Pettitt, T.R., Parry, D.W., Polley, R.W., 1996. Effect of temperature on the incidence of nodal foot rot symptoms in winter wheat crops in England and Wales caused by Fusarium culmorum and Microdochium nivale. Agric. For. Meteorol., Climate Change-From impact to interaction 79, 233–242.
Pettitt, T., Xu, X., Parry, D., 2003. Association of Fusarium species in the wheat stem rot complex, in: Xu, X., Bailey, J.A., Cooke, B.M. (Eds.), Epidemiology of Mycotoxin Producing Fungi. Springer Netherlands, pp. 769–774.
Pflughöft, O., 2008. Pilzkrankheiten in Körnerfuttererbsen (Pisum sativum L.) – Diagnose, Epidemiologie, Ertragsrelevanz und Bekämpfung. Der Fakultät für Agrarwissenschaften der Georg-August-Universität Göttingen.
Poole, G.J., Smiley, R.W., Walker, C., Huggins, D., Rupp, R., Abatzoglou, J., Garland-Campbell, K., Paulitz, T.C., 2013. Effect of climate on the distribution of Fusarium spp. causing crown rot of wheat in the Pacific northwest of the United States. Phytopathology 103, 1130–1140.
Scauflaire, J., Mahieu, O., Louvieaux, J., Foucart, G., Renard, F., Munaut, F., 2011. Biodiversity of Fusarium species in ears and stalks of maize plants in Belgium. Eur. J. Plant Pathol. 131, 59–66.
Schulte-Geldermann, E., Bruns, C., Hess, J., Finckh, M.R., 2009. Einfluss von ligninhaltigen Komposten und Pflanzgutgesundheit auf den Befall mit Rhizoctonia solani bei Kartoffeln.
Suárez-Estrella, F., Vargas-Garcıa, M.C., López, M.J., Moreno, J., 2004. Survival of Fusarium oxysporum f.sp. melonis on plant waste. Crop Prot. 23, 127–133.
Termorshuizen, A.J., van Rijn, E., van der Gaag, D.J., Alabouvette, C., Chen, Y., Lagerlöf, J., Malandrakis, A.A., Paplomatas, E.J., Rämert, B., Ryckeboer, J., Steinberg, C., Zmora-Nahum, S., 2006. Suppressiveness of 18 composts against 7 pathosystems: Variability in pathogen response. Soil Biol. Biochem. 38, 2461–2477.
Uhlig, S., Jestoi, M., Parikka, P., 2007. Fusarium avenaceum — The North European situation. Int. J. Food Microbiol. 119, 17–24.
VDLUFA Methodenbuch, 1991. Band I Die Untersuchung von Böden [WWW Document]. URL http://www.vdlufa.de/Methodenbuch/index.php?option=com_content&view=article&id=25&Itemid=7 (accessed 7.10.14).
Vigier, B., Reid, L.M., Seifert, K.A., Stewart, D.W., Hamilton, R.I., 1997. Distribution and prediction of Fusarium species associated with maize ear rot in Ontario. Can. J. Plant Pathol. 19, 60–65.
West, J.S., Townsend, J.A., Stevens, M., Fitt, B.D.L., 2012. Comparative biology of different plant pathogens to estimate effects of climate change on crop diseases in Europe. Eur. J. Plant Pathol. 133, 315–331.

FOOT AND ROOT ROT OF PEAS
40
3. AGGRESSIVENESS OF FOUR PATHOGENS CAUSING FOOT AND ROOT ROT OF PEA (PISUM SATIVUM L.) ON A SPRING AND A WINTER PEA CULTIVAR UNDER VARIABLE TEMPERATURE CONDITIONS
Abstract
A series of greenhouse trials was carried out to evaluate the susceptibility of the spring pea variety Santana and the winter variety EFB33 to Fusarium avenaceum, F. solani f. sp pisi, D.
pinodes and P. medicaginis under controlled conditions in sterile sand and field soil. Moreover, the effect of temperature on the aggressiveness of F. avenaceum and P.
medicaginis and pathogenicity of F. avenaceum on the agricultural crops that are commonly grown in rotation with peas in Germany was studied. Three weeks after sowing and inoculation disease symptoms were assessed and plant growth parameters measured. Regardless of pathogen and isolate used, as well as pea variety, plants grown in non-sterilized field soil were healthier than the plants grown in sterile sand. Santana was more susceptible than EFB33. All of the tested pathogens resulted in disease development. F.
avenaceum was the most aggressive pathogen causing severe wilting symptoms on Santana in sand (disease incidence (DI) 7.4). A strong negative correlation was observed between external root tissue damage and fresh weight for both varieties in both substrates. P.
medicaginis was the second most aggressive pathogen on Santana in sand (DI 5.7) and reduced fresh weight by 20% compared to control. Although the DI caused by D. pinodes
was somewhat lower than that of P. medicaginis, it caused significantly higher reductions of fresh weight in both substrates on Santana and in soil on EFB33. F. solani was the least aggressive of the four pathogens tested. Although there were no statistically significant effects of temperature on disease severity, there was a tendency that with decreasing temperature disease severity also decreased. In all treatments both varieties produced the highest biomass at the highest temperature. Out of eight plant species tested maize and wheat were least affected by inoculation with F. avenaceum. Stunted growth and reduction in biomass was observed for black oats, peas (Santana and EFB33), subterranean clover and Indian mustard, while white clover and white mustard exhibited severe wilting. This study implies that the tested pathogens differ in their aggressiveness and that the wide host range pathogen F. avenaceum can be highly aggressive causing severe disease and reduction of biomass on leguminous and no-leguminous hosts, especially Brassicaceae. Winter pea EFB33 is more tolerant to the pathogens tested and could be considered as alternative to susceptible summer varieties commonly grown in Germany and potentially contribute in overcoming recent decline in pea production in organic farming in Germany. Keywords: F. avenaceum, P. medicaginis, D. pinodes, F. solani, peas, temperature, foot rot

FOOT AND ROOT ROT OF PEAS
41
3.1 Introduction
Root and foot rot limits pea production worldwide (Ali et al., 1993; Bretag and Ramsey, 2001)
and is likely one of the reasons for the decline of pea production in Germany (Pflughöft et al.,
2012). It limits water and nutrient uptake, nitrogen fixation and causes stagnation of plant
growth (Oyarzun, 1993), preventing peas from reaching their full genetic potential (Kraft et
al., 1988). Yield losses of 10 - 30% are commonly reported for legume crops affected with
root rot, but yield loss potential under favorable conditions is even higher (Foroud et al.,
2014). A complex of more than 20 different species of soil-borne fungal pathogens can
cause foot and root rot of peas (Blume and Harman, 1979; Bodker et al., 1993; Kraft et al.,
1988; Olofsson, 1967; Persson et al., 1997). Many of the pathogens involved also attack a
range of legumes including chickpea, lentil, faba bean and lupine.
The most important species causing foot and root rot on peas are Didymella pinodes (syn
Mycospharella pinodes), Phoma medicaginis, Fusarium spp. (F. solani f. sp pisi, F.
oxysporum f.sp pisi, F. avenaceum, F. culmorum), Pythium spp., Thielaviopsis basicola,
Aphanomyces euteiches, and Rhizoctonia solani (Feng et al., 2010; Kraft et al., 1988; Nasir
et al., 1992; Oyarzun, 1993; Persson et al., 1997; Pflughöft et al., 2012).
Didymella pinodes (Berk. & A. Bloxam) Vestergr. (anamorph Ascochyta pinodes L. K. Jones)
together with Phoma medicaginis var. pinodela (Jones) and Ascochyta pisi Lib. are grouped
together in the “Ascochyta comlex” due to similar symptoms caused on pea (Wallen, 1974).
However, Ascochyta pisi rarely causes root rot. It is seed borne, does not produce
chlamydospores and causes mainly leaf and pod spot. The other two pathogens are seed
and soil borne and cause lesions on leaves, stems, and pods as well as foot rot (Ali et al.,
1993; Tivoli and Banniza, 2007).
Wallen (1974) stresses the importance of D. pinodes as a root rot pathogen on peas, as
infection of young seedlings can result in damping-off and increase lodging of older plants.
The main source of inoculum is infected stubble, in which the fungus can persist for several
cropping seasons (Gossen et al., 2010; Zhang et al., 2003). In seed, D. pinodes is able to
survive up to four years (Corbière et al., 1994) and in severe cases it may infect the embryo.
However, seed to seedling transmission is overall low (Bretag et al., 1995; Gossen et al.,
2010). Moussart et al. (1998) showed that low temperatures increase the frequency of this
type of transmission in peas. Even though seed infection is apparently of minor importance
for the epidemic development, it is often responsible for the introduction of the pathogen to
new areas (Gossen et al., 2010). In the case of blight, i.e. foliar and stem infection, air-borne
ascospores are considered to be the most important source of inoculum (Gossen et al.,
2010; Roger and Tivoli, 1996). Cool and wet conditions favor blight epidemics caused by D.
pinodes because they enhance ascospore dispersal and promote infection. Severe

FOOT AND ROOT ROT OF PEAS
42
epidemics can lead to yield losses of up to 50% (Bretag et al., 1995; Setti et al., 2009;
Wallen, 1974), reduce seed quality and limit the area in which field peas can be grown (Ali et
al., 1978; Le May et al., 2009; Roger et al., 1999).
Phoma medicaginis var. pinodella (teleomorph Didymella sp.) is also seedborne (Tivoli and
Banniza, 2007) but the main sources of inoculum in the field are pycnidiospores and
chlamydospores on crop debris in soil. There they can survive up to 3 years (Carr and Smith
1988). The main means of their distribution is rain splash. Ascospores produced by
Didymella sp. can be spread by wind over longer distances, and can lead to development of
severe epidemics (Bowen et al., 1997). P. medicagnis can cause severe foot rot symptoms
that can reduce plant growth and subsequent yield loss can reach 25% (Wallen, 1974). Ali et
al. (1994) report that P. medicaginis var pinodella isolates from soil are weakly virulent until
passage through the host plant and that is the reason why it is not usually epiphytotic in
nature. It has a wide host range within the Fabaceae family, causing the blights and root rot
on different Medicago, Vicia, Pisum, Melilotus, Glycine, Lupinus and Trifolium spp (Farr and
Rosmann, 2015).
F. solani f. sp pisi together with F. oxysporum f.sp pisi have been described as the most
frequent and most aggressive root rot causing fungi on peas worldwide in the last three
decades (Kraft et al., 1988; Oyarzun, 1993; Persson et al., 1997; Pflughöft et al., 2012). They
can reduce pea yields by 10% to 50% (Kraft et al., 1988; Rush and Kraft, 1986). Other
Fusarium spp. such as F. avenaceum and F. culmorum, are also frequently isolated from
roots (Persson et al., 1997).
F. oxysporum Schlecht. f. sp. pisi (Lindford) Snyder & Hansen occurs worldwide but the
severity of the damage depends on the race of the pathogen (Pflughöft, 2008). F. oxysporum
is a vascular wilt pathogen that usually infects plants through the side roots (Kraft et al.,
1988; Kraft and Pfleger, 2001). No sexual state has been found so far (Skovgaard et al.,
2002). This pathogen survives in the soil in the form of chlamydospores (Leslie and
Summerell, 2006).
F. solani (Mart.) Sacc. f. sp. pisi (Jones) Snyder & Hansen (teleomorph: Nectria
haematococca) is a very variable species that can be both saprophytic and parasitic (Ondrej
et al., 2008). It infects plants early with seed germination but symptoms usually appear later.
Biotic and abiotic stress conditions affect the incubation time of pathogen (Rush and Kraft,
1986). Initial symptoms on the roots consist of reddish-brown streaks starting from the
cotyledonary attachment area, which later spread and form black lesions on the roots and
stem base (Kraft et al., 1988; Kraft and Pfleger, 2001; Ondrej et al., 2008). Severely infected
plants are chlorotic and have stunted growth (Tu, 1987). In the plants infected with F. solani,

FOOT AND ROOT ROT OF PEAS
43
vascular tissue often develops intensive red color and this can be used as an indicator of its
presence. The pathogen survives in soil in the form of chlamydospores.
F. avenaceum (Fries) Sacc (sexual stage Gibberella avenacea) is predominantly a soilborne
fungus, that may be also seedborne on some legumes (Leslie and Summerell, 2006). It is
common in temperate regions throughout the world. F. avenaceum has a wide host range,
does not typically segregate into host-specific pathotypes (Schneider, 1958; Hwang et al.,
1994), and causes disease on a range of agricultural crops. F. avenaceum is morphologically
variable (Schneider, 1958) and genetically diverse (Miedaner et al., 2008). It has been
reported as a pathogen causing economically important diseases of canola (Berkenkamp
and Vaartnou, 1972; Hwang et al., 2000), lentil (Fletcher et al., 1991; Hwang et al., 2000),
subterranean clover (Greenhalgh and Siobhan,1984), red clover (Yli-Mattila et al. 2010), pea
(Feng et al., 2010; Fletcher et al., 1991b; Hwang et al., 2000; Persson et al., 1997), soybean
(Zhang et al., 2013), alfalfa (Cormack, 1951; Hwang et al., 2000), wheat, barley, oat (Cook,
1980; Hwang et al., 2000; Smiley and Patterson, 1996), etc.
F. avenaceum rarely produces chlamydospores and survives saprophyticaly in soils or on
plant residues (Forbes and Dickinson, 1977; Leslie and Summerell, 2006). It produces
secondary metabolites such as beauvericin, fusarin C, moniliformin, and enniatins A, B, and
B1 (Leslie and Summerell, 2006), which are toxic to animals and humans. Pre-emergence
damping off, post-emergence root rots, yellowing, stunting and wilting are symptoms of
infection with F. avenaceum (Bateman et al., 2007).
Usually, several pathogens are isolated from symptomatic pea roots. The role of single
pathogens in disease development and severity is not clear. Prevalence, dominance and
importance of the single pathogens vary greatly depending on location, climate, and
agricultural practice (Gaurilckiene and Cesnuleviciene, 2013; Jensen et al., 2004; Persson et
al., 1997) and some shifts in importance have occurred over time. Thus, overall disease
severity and yield loss due to foot and root rot are the combined effects of the pathogens and
it is practically impossible to relate symptoms on roots and reduction of yield under field
conditions to single pathogens (Tu, 1987). For that reason a series of greenhouse trials was
conducted in order to:
1) Examine the pathogenic potential of the four foot and root rot pathogens, F.
avenaceum, F. solani, P. medicaginis and D. pinodes on a spring and winter pea
variety in two substrates;
2) Examine the effect of temperature on disease severity caused by F. avenaceum and
P. medicaginis;

FOOT AND ROOT ROT OF PEAS
44
3) Investigate the pathogenicity of F. avenaceum on the agricultural crops that are
commonly grown in rotation with peas in Germany.

FOOT AND ROOT ROT OF PEAS
45
3.2 Materials and Methods All experiments were replicated five times in a completely randomized design.
3.2.1 Pathogenicity of Fusarium avenaceum, Fusarium solani, Phoma medicaginis and
Didymella pinodes, in sterile sand and in non-sterilized field soil
The winter pea variety EFB33 and spring variety pea Santana (KWS LOCHOW, Gmbh) were
used in the experiments. EFB33 is botanically Pisum sativum L. ssp. sativum convar.
speciosum (Dierb.) Alef. (formerly Pisum arvense L.). It is a regularly leafed, small seeded
variety with thousand seed weight of about 130 g and violet flowers. Santana is a spring pea
widely grown in Germany. Botanically it belongs to Pisum sativum L. ssp. sativum convar.
sativum. This cultivar is semi-leafless, white flowering with white seed coat and a thousand
seed weight of 250 to 300 g.
Prior to use pea seeds were surface disinfected with 70% ethanol for five minutes, rinsed
with running distilled water and air dried under a laminar flow cabinet. All defective and
damaged seeds were discarded. Plants were grown in 6 cm diameter and 8 cm deep round
plastic pots filled with 150 mL (~200 g) of sterile yellow sand or non-sterilized field soil as
growing media. Sand was autoclaved for 20 minutes at 121°C and stored in a closed
container until the beginning of the trial. It had a water content of 7%, water holding capacity
of 27.5 g per 100g dry sand and maximum water absorption of 17.5 mL per 100 g. The field
soil for the trial was collected from the experimental field of the University of Kassel at Neu-
Eichenberg. Peas were grown in this field a year before the soil was used for the greenhouse
experiments. The soil type is a deep Haplic Luvisol from loess with 78 soil points (Wildhagen,
1998). Four seeds were sown in each pot at 2 cm depth. Plants were grown for 21 days with
a day/night temperature regime of 19/16°C. Pots with sand as a substrate were fertilized with
complex N:P:K fertilizer Wuxal Super (8:8:6 + microelements). A total of 100 mg of N L-1
substrate was divided in two equal portions and added 10 and 15 days after sowing. No
fertilizer was used in the field soil.
Five different isolates of each pathogen were used in the experiment (Tab. 3.1). Isolates of F.
solani were collected from pea plants on different organic farms in Germany, in the period
between 2010 and 2012. Two isolates of D. pinodes and two of P. medicaginis were kindly
provided by Dr. B. Tivoli from the National Institute of Agronomic Research (INRA) France.
One isolate of D. pinodes and one of P. medicaginis were kindly provided by the Plant
Pathology Department of the Georg-August-University in Göttingen. F. avenaceum was
represented in the experiment by two isolates from winter, one isolate from subterranean
clover seeds and two from pea plants collected between 2011 and 2012. D. pinodes and P.
medicaginis isolates were multiplied in Petri dishes containing COON’s agar (Maltose 4 g,
KNO3 2 g, MgSO4 1.20 g, KH2PO4 2.68 g, Agar 20 g , H2O 1L) while F. solani and F.

FOOT AND ROOT ROT OF PEAS
46
avenaceum cultures were grown on half strength PDA. Petri dishes were incubated at 20˚C
and subjected to 12 hour cycles of UV light and dark for 15 to 20 days.
Inoculation was done with spore suspensions at the time of sowing. Spores were prepared
by flooding the surface of cultures with distilled water and gently scraping them. The spore
concentrations were determined with a Fuchs Rosenthal haemocytometer and adjusted to
105 spores g-1 of growing substrate. After inoculation, pots were watered and placed in the
greenhouse. Irrigation was done regularly according to need.
Table 3.1. Pathogens and isolates used in experiments.
Pathogen Isolate name Plant species Origin
Didymella pinodes
Dp4 Dp 91.31.12 Pea INRA, France
Dp5 Dp Gö Pea Göttingen
Dp6 Dp KLIFF Pea Neu Eichenberg
Dp3 Dp 96Ca2 Pea INRA, France
Dp7 Dp B2 Pea Neu Eichenberg
Fusarium avenaceum
Fa1 Fa W/VIII.11 Wheat Neu Eichenberg
Fa6 Fa W/III.13.2 Wheat Neu Eichenberg
Fa4 Fa NEB.12 Pea Neu Eichenberg
Fa7 Fa DFH Pea Domene Frankenhausen
Fa8 Fa SC Subclover seeds Italy
Phoma medicaginis
Pm1 Pm GÖ Pea Göttingen
Pm2 Pm KLIFF 12 Pea Neu Eichenberg
Pm3 Pm 94.56.3 Pea INRA, France
Pm4 B1.10 Pea Neu Eichenberg
Pm5 Pm 94.14.11 Pea INRA, France
Fusarium solani
Fs2 Fs GER 10III10 Pea East Germany
Fs3 Fs LIN V.12 Pea BOFRU
Fs4 Fs MÖL V.12 Pea BOFRU
Fs1 Fs KLIFF IV.12 Pea Neu Eichenberg
Fs5 Fs T19 3III.10 Pea North of Germany
FOOT AND ROOT ROT OF PEAS
47
3.2.2 Effect of temperature on the aggressiveness of F. avenaceum and P. medicaginis
Effects of reduced growing temperatures (Tab. 3.2) on the aggressiveness of P. medicaginis
and F. avenaceum were studied in the period between March and May 2012. P. medicaginis
isolate Pm1 and F. avenaceum isolate Fa1 (Tab. 3.1) were chosen for this test. The same
pea varieties and sterile sand were used as in the previous trial. Preparation of seeds,
sowing, inoculation, fertilization, watering and duration of experiment was as described
above (see 3.2.1.). Square pots of 12 x 12 x 11 cm were filled with 1 L (~1.3 kg) of yellow
sterile sand and eight pea seeds were sown per pot.
Table 3.2. Temperature regimes used in the experiment
Treatment Day temperature (°C) Night temperature (°C)
Low 13 10
Medium 16 12
High 19 16
3.2.3 Pathogenicity of F. avenaceum
The pathogenicity of two F. avenaceum isolates, Fa1 from wheat and Fa4 from pea (Tab.
3.1) was tested on eight plant species in addition to peas (Tab. 3.3). Plants were inoculated
at the time of sowing with 104 spores g-1 substrate and grown at a day/night temperature
regime of 19/16°C. Square pots of 12 x 12 x 11 cm with 1 L (~1.3 kg) of yellow sterile sand
were used. Preparation of seeds, sowing, inoculation, fertilization, watering and duration of
experiment was as described above (see 3.2.1).
Table 3.3. Plant species, varieties and sowing density used in the experiment.
Plant species Variety No. plants per pot Maize (Zea mays) Fabregas 5
Winter wheat (Triticum aestivum) Achat 5
Oat (Avena sativa) Dominik 5
Black oat (Avena strigosa) Pratex 5
Pea (Pisum sativum L. convar. sativum) Santana 8
Pea (Pisum sativum L. convar. speciosum) EFB33 8
Subterranean clover (Trifolium subterraneum) Dalkeith 12
White clover (Trifolium repens) Liflex 12
Indian mustard (Brassica juncea) Terrafit 12
White mustard (Sinapis alba) Samba 12

FOOT AND ROOT ROT OF PEAS
48
3.2.4 Measurement of plant biomass and assessment of disease
Plants were harvested 21 days after sowing in all experiments. They were carefully pulled
out of the substrate. Roots were cut off, washed with running tap water, and stored at 4°C
until use. Above ground plant parts of each pot were weighted and dried at 105°C until
constant weight. Root disease severity was assessed according to the key of Pflughöft
(2008) (Tab. 2.4, Chapter 2). Assessments were always performed within 48 hours following
harvest. Symptoms on the external plant tissue were evaluated followed by the assessment
of damage of the internal tissue. In the test for pathogenicity of F. avenaceum on nine plant
species, in addition to above ground biomass, roots were also weighted. As the symptoms
on roots were uniform within different plant species, no detailed assessment of the symptoms
was performed.
3.2.5 Data processing and analysis
A disease index (DI) for each plant was calculated as the average of the external and internal
lesion score. As all plants in sand controls emerged, missing plants in the treatments were
considered dead and score 9 was given for both external and internal tissue. The mean DI of
the eight plants in each pot was calculated to present one replication, and further analyses
were done on these values. Analyses of variance (ANOVA) were applied in order to analyze
the effect of the pathogens, substrate and temperature on the DI, plant weights and survival
rates. Prior to ANOVA, data were checked for normality with Shapiro-Wilks-W-Test. Mean
separations were made by Tukey honest significant differences (HSD) test at the P < 0.05.
All statistical analyses were done in the statistical program R version 2.15.2. Regressions of
fresh weights and DI were analyzed with Excel 2007.

FOOT AND ROOT ROT OF PEAS
49
3.3 Results
3.3.1 Comparison of aggressiveness of F. avenaceum, F. solani, P. medicaginis and D.
pinodes on two pea varieties in sand and field soil
3.3.1.1 Isolate comparison
Regardless of pathogen and isolate used, as well as pea variety, plants grown in non-
sterilized field soil were healthier than the plants grown in sterile sand. Santana was more
susceptible than EFB33. DI varied rarely among isolates within pathogens depending on the
test system (Fig. 3.1). Statistically significant isolate effects occurred only for F. avenaceum
isolates on Santana in sand (Fig. 3.1A) and for D. pinodes isolates on Santana grown in soil,
and on EFB33 in both substrates (Fig. 3.1C).
All of the tested F. avenaceum isolates significantly reduced the weight of Santana plants
grown in sand (Fig. 3.2). In contrast, EFB33 was not affected. There was even a tendency
that weights of inoculated treatments were higher than control. However, this difference was
not statistically significant (Fig. 3.2).
Three out of the five P. medicaginis caused significant reductions of fresh and dry weights
(data not shown) of Santana plants in sand (Fig. 3.3). There were no significant differences
in fresh weights among different D. pinodes or F. solani isolates neither on Santana nor on
EFB33 (see Appendix Fig. A. 3.3 and A. 3.4).

FOOT AND ROOT ROT OF PEAS
50
Figure 3.1. Disease index (DI) of Santana and EFB33 plants grown in sand and soil and inoculated with five isolates of F. avenaceum (A), F. solani (B), D. pinodes (C) and P. medicaginis (D). Different small letters within variety and substrate are indicating significant differences in DI at P < 0.05 (Tukey’s HSD test). The horizontal line in the boxplot shows the median, the bottom and tops of the box the 25th and 75th percentiles and the vertical lines the minimum and maximum values; outliers as single points. Mean values of DI are marked with triangles.
ab
a ab b b
ab b a b ab ab b ab a ab
b b ab a ab
A B
C D
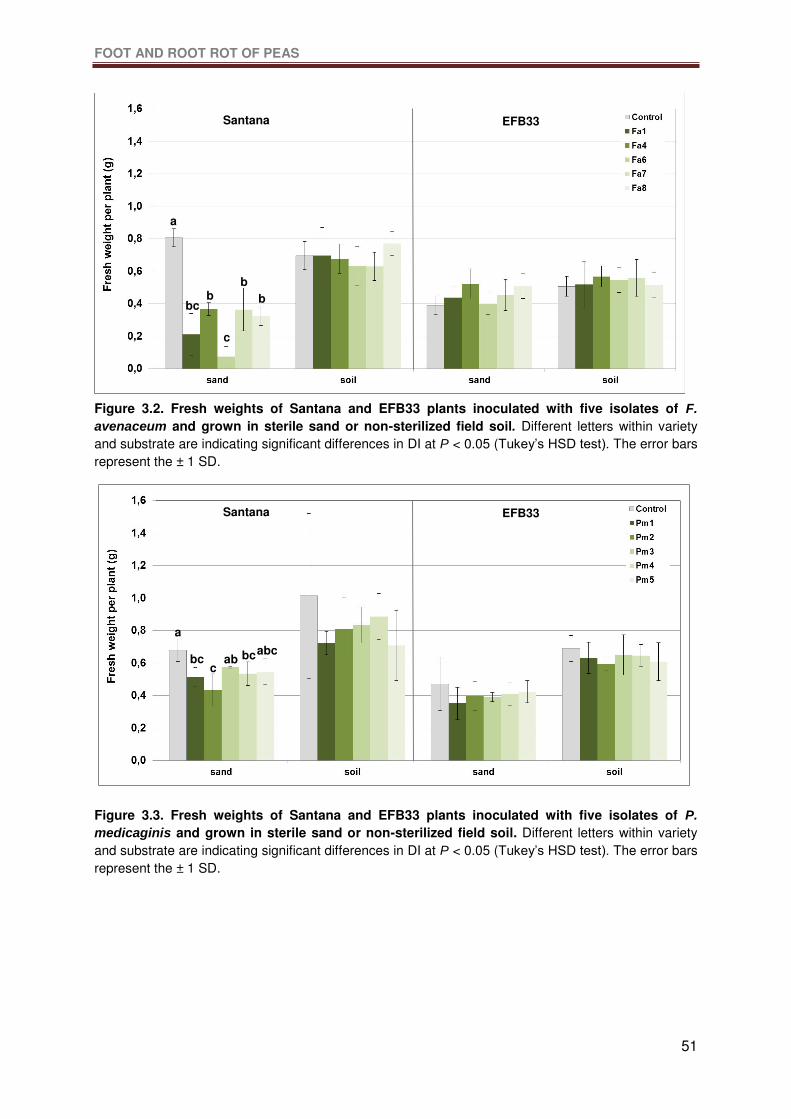
FOOT AND ROOT ROT OF PEAS
51
Figure 3.2. Fresh weights of Santana and EFB33 plants inoculated with five isolates of F.
avenaceum and grown in sterile sand or non-sterilized field soil. Different letters within variety and substrate are indicating significant differences in DI at P < 0.05 (Tukey’s HSD test). The error bars represent the ± 1 SD.
Figure 3.3. Fresh weights of Santana and EFB33 plants inoculated with five isolates of P.
medicaginis and grown in sterile sand or non-sterilized field soil. Different letters within variety and substrate are indicating significant differences in DI at P < 0.05 (Tukey’s HSD test). The error bars represent the ± 1 SD.
Santana EFB33
Santana EFB33
a
bc c
ab bc abc
a
bc b
b
c
b

FOOT AND ROOT ROT OF PEAS
52
3.3.1.2 Pathogen comparison
Unlike the other pathogens that caused compact black lesions that spread completely around
the plant stem accompanied by rot of epidermis/rhizodermis, symptoms of F. solani infection
appeared as a mass of single black streaks very close to each other but it was still possible
to see green healthy tissue in between (Fig. 3.4). During the disease assessment red color of
vascular tissue was observed in plants inoculated with both Fusarium spp.
Figure 3.4. Typical symptoms of F. solani infection on the stem base of Santana (A) and EFB33 (B) plants.
Interactions between pathogen and variety, as well as pathogen and substrate were highly
significant, P < 0.001 (see Appendix Tab. A. 3.5). F. avenaceum caused severe disease
symptoms on Santana plants grown in sand (Fig. 3.5) with mean DI of 7.38 (Fig. 3.6) and an
average reduction of fresh weight of 65.3% compared to the control (Fig. 3.7). Besides the
intensive black lesions on the epicotyl that spread up to the soil level, plants exhibited strong
wilting symptoms. First wilting symptoms usually appeared after 14 days and soon after the
plants died. Intensive orange sporodochia were formed on the dead plant tissue (Fig. 3.5).
A B

FOOT AND ROOT ROT OF PEAS
53
Figure 3.5. Wilting symptoms of Santana caused by F. avenaceum with the formation of bright orange sporodochia (marked with the arrow).
The second most aggressive pathogen on Santana in sand was P. medicaginis with a mean
DI of 5.7 (Fig. 3.6) and reduction of plant fresh weight by 20%. Although D. pinodes caused
less severe symptoms with DI 4.8, reduction of fresh weight caused by infection with this
pathogen was on average 34% (Fig. 3.7). F. solani was least aggressive and caused
moderate disease symptoms with DI 4.0 and an average fresh weight reduction of 8% (Fig.
3.7). Dry weights followed the same pattern (data not shown).

FOOT AND ROOT ROT OF PEAS
54
Figure 3.6. Disease index (DI) of Santana and EFB33 plants grown in sand and soil and inoculated with F. avenaceum (Fa), F. solani (Fs), P. medicaginis (Pm) and D. pinodes (Dp). O is the non-inoculated control. The horizontal line in the boxplot shows the median, the bottom and tops of the box the 25th and 75th percentiles and the vertical lines the minimum and maximum values; outliers as single points. Mean values of DI are marked with triangles. Different letters within variety and substrate are indicating significant differences in DI at P < 0.05 (Tukey’s HSD test).
Figure 3.7. Fresh weights of Santana and EFB33 inoculated with F. avenaceum (Fa), F. solani (Fs), P. medicaginis (Pm) and D. pinodes (Dp). Different letters within variety and substrate are indicating significant differences in DI at P < 0.05 (Tukey’s HSD test). The error bars represent the ± 1 SD.
c
ab b
a
c
a
ab b b c
b ab ab ab a
e d a
Santana EFB33
a
d
ab
c b
a ab
bc
a
c
ab
a
b ab a a a
b
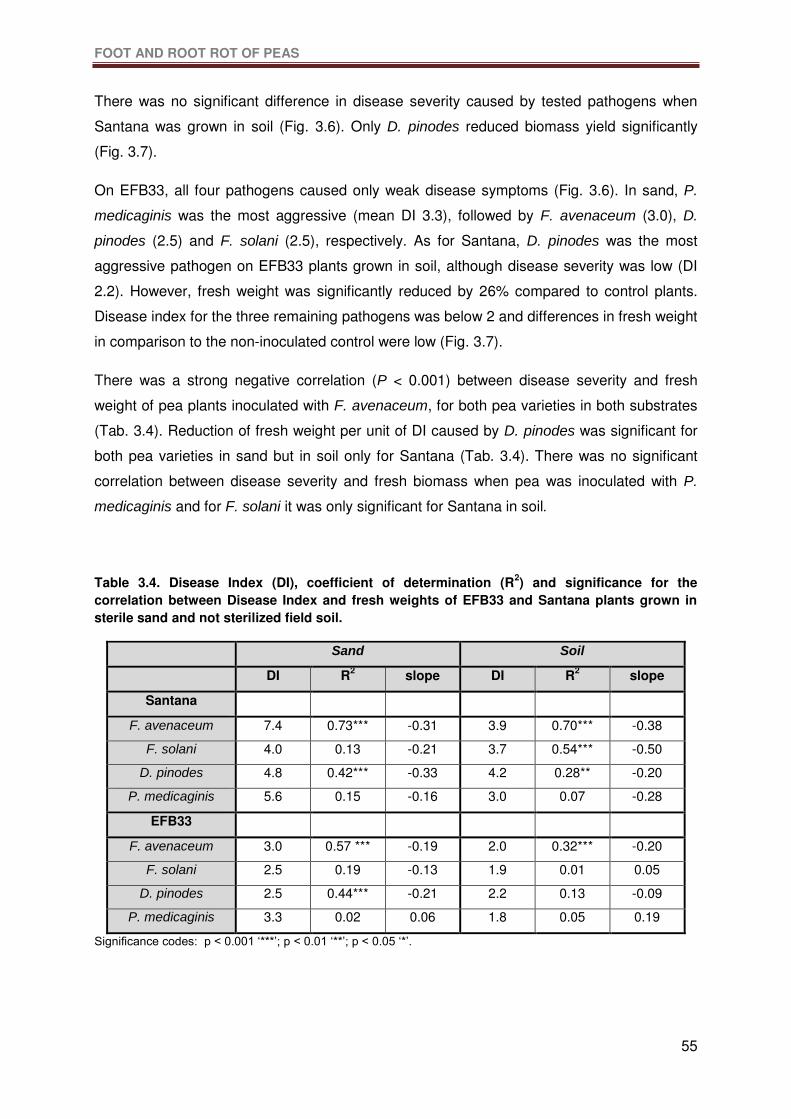
FOOT AND ROOT ROT OF PEAS
55
There was no significant difference in disease severity caused by tested pathogens when
Santana was grown in soil (Fig. 3.6). Only D. pinodes reduced biomass yield significantly
(Fig. 3.7).
On EFB33, all four pathogens caused only weak disease symptoms (Fig. 3.6). In sand, P.
medicaginis was the most aggressive (mean DI 3.3), followed by F. avenaceum (3.0), D.
pinodes (2.5) and F. solani (2.5), respectively. As for Santana, D. pinodes was the most
aggressive pathogen on EFB33 plants grown in soil, although disease severity was low (DI
2.2). However, fresh weight was significantly reduced by 26% compared to control plants.
Disease index for the three remaining pathogens was below 2 and differences in fresh weight
in comparison to the non-inoculated control were low (Fig. 3.7).
There was a strong negative correlation (P < 0.001) between disease severity and fresh
weight of pea plants inoculated with F. avenaceum, for both pea varieties in both substrates
(Tab. 3.4). Reduction of fresh weight per unit of DI caused by D. pinodes was significant for
both pea varieties in sand but in soil only for Santana (Tab. 3.4). There was no significant
correlation between disease severity and fresh biomass when pea was inoculated with P.
medicaginis and for F. solani it was only significant for Santana in soil.
Table 3.4. Disease Index (DI), coefficient of determination (R2) and significance for the correlation between Disease Index and fresh weights of EFB33 and Santana plants grown in sterile sand and not sterilized field soil.
Sand Soil
DI R2 slope DI R2 slope
Santana
F. avenaceum 7.4 0.73*** -0.31 3.9 0.70*** -0.38
F. solani 4.0 0.13 -0.21 3.7 0.54*** -0.50
D. pinodes 4.8 0.42*** -0.33 4.2 0.28** -0.20
P. medicaginis 5.6 0.15 -0.16 3.0 0.07 -0.28
EFB33
F. avenaceum 3.0 0.57 *** -0.19 2.0 0.32*** -0.20
F. solani 2.5 0.19 -0.13 1.9 0.01 0.05
D. pinodes 2.5 0.44*** -0.21 2.2 0.13 -0.09
P. medicaginis 3.3 0.02 0.06 1.8 0.05 0.19
Significance codes: p < 0.001 ‘***’; p < 0.01 ‘**’; p < 0.05 ‘*’.

FOOT AND ROOT ROT OF PEAS
56
3.3.2 Effect of temperature on disease severity caused by F. avenaceum and P.
medicaginis
Both pathogens caused moderate to severe disease with DI between 4.5 and 7.4 on Santana
(Fig. 3.8). DIs at 19°/16°C day/night regime were the same as in the first experiment (see
Fig. 3.6). They were generally higher than at the lower temperatures. Again, in contrast to
Santana, disease severity on EFB33 was overall low. Although there were no statistically
significant differences among different temperature regimes, there was a tendency that with
decresing temperature disease severity decreased. The highest DI on EFB33 was 3.0 in the
treatment inoculated with P. medicaginis and grown at the highest tested temperature (Fig.
3.8).
Figure 3.8. Disease index of Santana and EFB33 inoculated with F. avenaceum (Fa) and P.
medicaginis (Pm) and grown under different temperature regimes. O is the non-inoculated control. The horizontal line in the boxplot shows the median, the bottom and tops of the box the 25th and 75th percentiles and the vertical lines the minimum and maximum values; outliers as single points. Mean values of DI are marked with triangles. Different letters within variety and temperature are indicating significant differences in DI at P < 0.05 (Tukey’s HSD test).
b a a b a a c a b
b a a c b a b a a
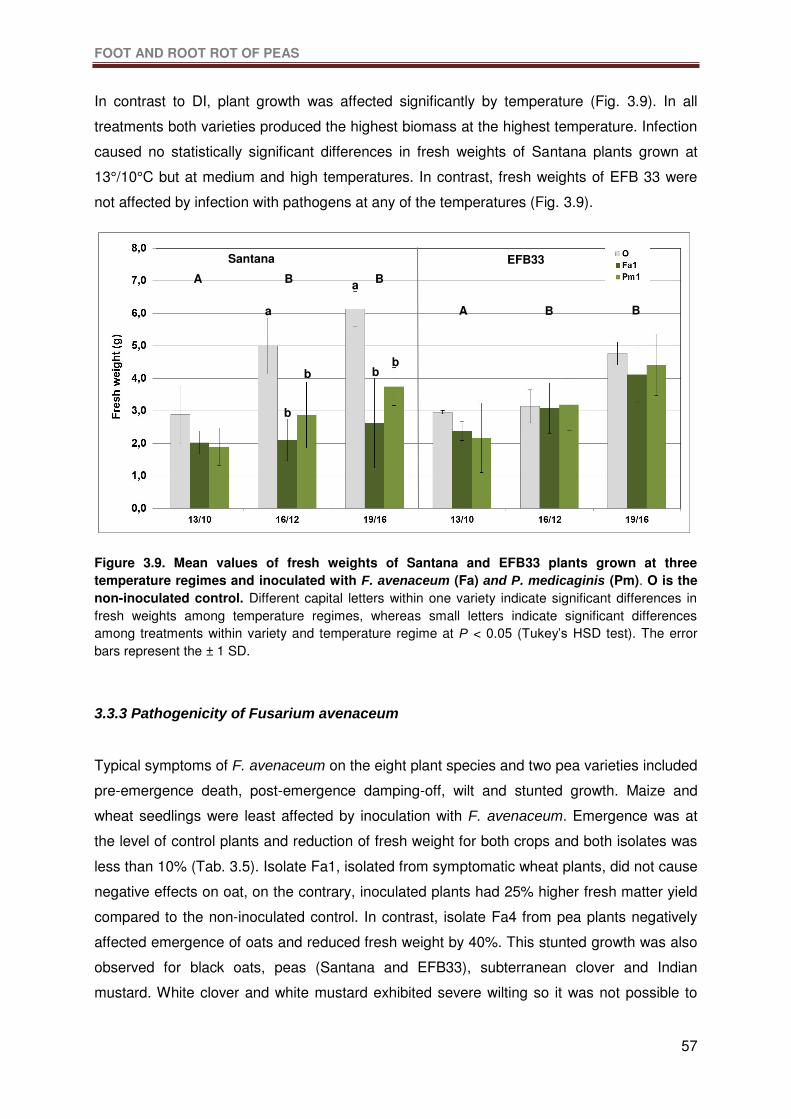
FOOT AND ROOT ROT OF PEAS
57
In contrast to DI, plant growth was affected significantly by temperature (Fig. 3.9). In all
treatments both varieties produced the highest biomass at the highest temperature. Infection
caused no statistically significant differences in fresh weights of Santana plants grown at
13°/10°C but at medium and high temperatures. In contrast, fresh weights of EFB 33 were
not affected by infection with pathogens at any of the temperatures (Fig. 3.9).
Figure 3.9. Mean values of fresh weights of Santana and EFB33 plants grown at three temperature regimes and inoculated with F. avenaceum (Fa) and P. medicaginis (Pm). O is the non-inoculated control. Different capital letters within one variety indicate significant differences in fresh weights among temperature regimes, whereas small letters indicate significant differences among treatments within variety and temperature regime at P < 0.05 (Tukey’s HSD test). The error bars represent the ± 1 SD.
3.3.3 Pathogenicity of Fusarium avenaceum
Typical symptoms of F. avenaceum on the eight plant species and two pea varieties included
pre-emergence death, post-emergence damping-off, wilt and stunted growth. Maize and
wheat seedlings were least affected by inoculation with F. avenaceum. Emergence was at
the level of control plants and reduction of fresh weight for both crops and both isolates was
less than 10% (Tab. 3.5). Isolate Fa1, isolated from symptomatic wheat plants, did not cause
negative effects on oat, on the contrary, inoculated plants had 25% higher fresh matter yield
compared to the non-inoculated control. In contrast, isolate Fa4 from pea plants negatively
affected emergence of oats and reduced fresh weight by 40%. This stunted growth was also
observed for black oats, peas (Santana and EFB33), subterranean clover and Indian
mustard. White clover and white mustard exhibited severe wilting so it was not possible to
Santana EFB33
A B B
A B B
a
b b
a
b
b

FOOT AND ROOT ROT OF PEAS
58
record yield data for these two species. Percentage of wilted plants was close to 80% for
white mustard and 55% for white clover inoculated with Fa4 and 70% for Fa1 (Tab. 3.5).
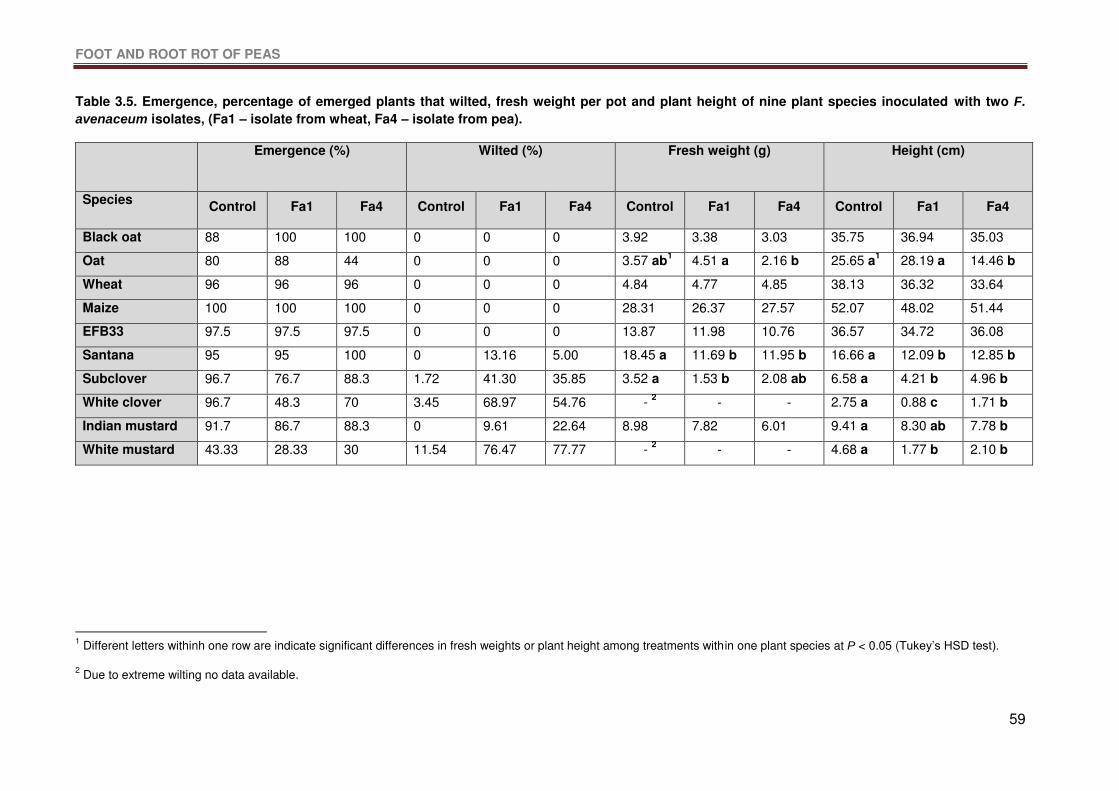
FOOT AND ROOT ROT OF PEAS
59
Table 3.5. Emergence, percentage of emerged plants that wilted, fresh weight per pot and plant height of nine plant species inoculated with two F.
avenaceum isolates, (Fa1 – isolate from wheat, Fa4 – isolate from pea).
Emergence (%) Wilted (%) Fresh weight (g) Height (cm)
Species Control Fa1 Fa4 Control Fa1 Fa4 Control Fa1 Fa4 Control Fa1 Fa4
Black oat 88 100 100 0 0 0 3.92 3.38 3.03 35.75 36.94 35.03
Oat 80 88 44 0 0 0 3.57 ab1 4.51 a 2.16 b 25.65 a1 28.19 a 14.46 b
Wheat 96 96 96 0 0 0 4.84 4.77 4.85 38.13 36.32 33.64
Maize 100 100 100 0 0 0 28.31 26.37 27.57 52.07 48.02 51.44
EFB33 97.5 97.5 97.5 0 0 0 13.87 11.98 10.76 36.57 34.72 36.08
Santana 95 95 100 0 13.16 5.00 18.45 a 11.69 b 11.95 b 16.66 a 12.09 b 12.85 b
Subclover 96.7 76.7 88.3 1.72 41.30 35.85 3.52 a 1.53 b 2.08 ab 6.58 a 4.21 b 4.96 b
White clover 96.7 48.3 70 3.45 68.97 54.76 - 2 - - 2.75 a 0.88 c 1.71 b
Indian mustard 91.7 86.7 88.3 0 9.61 22.64 8.98 7.82 6.01 9.41 a 8.30 ab 7.78 b
White mustard 43.33 28.33 30 11.54 76.47 77.77 - 2 - - 4.68 a 1.77 b 2.10 b
1 Different letters withinh one row are indicate significant differences in fresh weights or plant height among treatments within one plant species at P < 0.05 (Tukey’s HSD test).
2 Due to extreme wilting no data available.

FOOT AND ROOT ROT OF PEAS
60
3.4 Discussion
In sterile sand F. avenaceum was the most aggressive pathogen, followed by P. medicaginis,
D. pinodes, and F. solani. In contrast, in non-sterile field soil, overall aggressiveness of all
pathogens was greatly reduced and F. avenaceum and D. pinodes were similar in their
aggressiveness in that system. Overall, the spring pea variety Santana was considerably
more susceptible than the winter pea variety EFB33 in both systems. Lower temperatures
reduced plant growth and slightly increased disease on winter variety EFB33, with the
highest DI at low temperatures (13/10°C). In contrast, the highest disease severity on
Santana was observed at the highest temperature, 19/16°C. The two isolates of F.
avenaceum originating from wheat and pea caused severe symptoms on roots of all plant
species that were tested, especially on peas, white clover and subterranean clover as well as
mustard and to a lesser degree on Indian mustard. The pea isolate also affected growth of
oats, whereas the isolate from wheat had no negative effect on oats.
Zhang et al. (2013) showed that in steam-sterilized peat-soil potting mix, among nine fungal
species (F. oxysporum, F. graminearum, F. solani, F. avenaceum, F. tricinctum, F.
sporotrichioides, F. equiseti, F. poae and Rhizoctonia solani), F. avenaceum together with F.
graminearum and R. solani, was the most pathogenic on soybean seedlings. Persson et al
(1997) observed that inoculation with F. avenaceum resulted in high disease severity and low
plant weight of peas in sterilized 1:1 peat/sand mix. The observed decrease in
aggressiveness in non-sterilized soil can be explained by the low competitive ability of this
pathogen, that is unable to compete with fast growing soil saprophytes and can be
suppressed by the soil microbiota (Fletcher et al., 1991; Lin and Cook, 1979).
With respect to the rot symptoms, Phoma medicaginis was the second most aggressive
pathogen on Santana and EFB33 and caused moderate disease. P. medicaginis is important
pea pathogen in the Netherlands, Scandinavia, and Germany (Oyarzun and Loon, 1989;
Persson et al., 1997; Pflughöft et al., 2012). It was the most frequently isolated root pathogen
of peas in Sweden in the years 1993-1995 (Persson et al., 1997) and second most frequent
(after Fusarium spp.) in Germany in 2005-2007 (Pflughöft et al., 2012) and most frequent in
Germany from 2008-2012. In a field study that we conducted from 2009-2012 (see Chapter
2) P. medicaginis was the dominant pathogen on EFB33, isolated from 82% of the sampled
plants. It is part of the Ascochyta blight complex, and it can reduce pea yields by 25%
(Wallen, 1974). In our study, infection with P. medicaginis did not reduce fresh weights of
three week old plants, except for Santana in sterile sand. Similar results were reported by
Persson et al. (1997) who found that although the pathogen caused moderate symptoms on
the epicotyl of pea in a pot trial, reduction of plant fresh weight was small. Severity of disease
caused by P. medicaginis in the sterile system did not change under different temperature

FOOT AND ROOT ROT OF PEAS
61
conditions. Fresh weights of EFB33 plants did not differ from the control plants; in Santana
they were reduced by 35% on average regardless of the temperature applied.
In general, effects of pathogens in our experiments were more visible on the fresh than on
the dry weights. This is likely due to the fact that disease caused by tested pathogens firstly
impeded water uptake and before the intensive accumulation of dry matter in plants
occurred. As the plants were harvested after three weeks of growing effects on dry weights
were therefore expected to be less pronounced.
Fusarium solani and Didymella pinodes caused moderate disease symptoms on Santana
and low symptoms on EFB33. Nevertheless, inoculation with D. pinodes resulted in high
reductions of plant fresh weight of both pea varieties. Plants inoculated with this pathogen
had, after F. avenaceum, the lowest biomass yield. There are few data available about the
role of D. pinodes as a part of foot and root rot in the yield reduction of peas. In contrast to
our findings, (Persson et al., 1997) reported only small yield reductions after inoculation of
pea with D. pinodes in greenhouse trials. High genetic variability in the D. pinodes
populations and existence of different pathotypes can be a reason for this disagreement. As
part of the Ascochyta blight complex, D. pinodes is the most damaging (Boros and Wawer,
2009; Onfroy et al., 1999; Wallen, 1974) causing yield reductions of up to 70% (Le May et al.,
2009). Results obtained in our study strongly support Campbell and Neher (1994)
observation that in the case of root disease in order to observe the full effect of a pathogen,
assessment of severity of disease symptoms on plant roots must be correlated with
measurement of above-ground plant growth parameters (leaf area, weights, heights, leaf
number, respiration intensity, etc).
Fusarium solani has been recognized as one of the most prevalent and important parts of the
foot rot complex on peas (Kraft et al., 1988; Rush and Kraft, 1986). In our study, F. solani
caused the least damage compared to the three other pathogens. Besides a low DI, it did not
affect plant growth. Moreover, fresh weight of both varieties inoculated with F. solani in soil
was higher than in the control plants. However, this difference was not statistically significant.
Lack of external stress and the short time for colonization (3 weeks) of the root system by
this pathogen can be reasons for the low effects of F. solani. Rush and Kraft (1986) observed
that despite of severe rot symptoms at the cotyledon attachment area even 35 days after
emergence of peas, deterioration of roots and reduction of plant growth was still low. The
same rot symptoms caused severe loss of roots and stunting of plant growth 49 days after
emergence (beginning of flowering). Hence, they concluded that one of the most important
predictors of potential effects of F. solani f.sp. pisi is the timing of deterioration of roots and
not the severity of symptoms. Any factor that stresses the host root system will shorten the
time between infections and severe root rot (Thung and Rao, 1999). As F. solani f. sp pisi is

FOOT AND ROOT ROT OF PEAS
62
a slow colonizer of the roots (Huisman, 1982), it is very likely that three weeks for the
experiments were not long enough for the pathogen to colonize roots and cause more severe
damage. Besides that, in the bioassays done, the only stress factor for pea plants was the
pathogen itself, and this is rarely the situation in field conditions. It is more usual that in the
field plants are exposed to a combination of different environmental stress factors (drought,
frost, soil compaction, etc.).
F. avenaceum is known to cause severe disease on a range of legume species (Feng et al.,
2010; Fletcher et al., 1991; Lamprecht et al., 1986; Persson et al., 1997; Zhang et al., 2013)
as well as on canola (Berkenkamp and Vaartnou, 1972; Hwang et al., 2000), oat, barly and
wheat (Cook, 1980; Hwang et al., 2000; Smiley and Patterson, 1996). The high fresh weight
reduction observed in our study is in agreement with findings of Persson et al. (1997) who
reported biomass reductions of 66% in peas in comparison to the uninoculated control. Our
results suggest a great potential of F. avenaceum to damage some brassicaceous species at
least in a sterile system. Indeed, epidemics of F. avenaceum on Brassica napus have been
reported from Canada (Feng et al., 2010).
The optimal temperature for growth of F. avenaceum is in the range of 20° and 25°C
(Campbell and Lipps, 1998; Pettitt et al., 1996). Brennan et al. (2003) showed that at the
temperature range of 20-25°C F. avenaceum caused the greatest retardation in coleoptile
growth of wheat (> 89.3%). At the same time, in in vitro tests, growing rates of F. avenaceum
culture increased with increasing temperature until 20°C and then decreased. Hwang et al.
(2000) obtained similar results in a controlled-environment study on lentil seedlings. Root rot
symptoms were the most severe when lentil was grown in warmer temperatures from 20°C to
27.5°C. At lower or higher temperatures disease severity on seedlings declined.
Furthermore, Satyaprasad et al. (2000) reported the increase in mean disease scores
caused on wheat and lupines by F. avenaceum with rising temperature.
Our results suggest that due to increases of host growth at increased temperatures the
overall damage may still be less at higher temperatures. An important consideration is that F.
avenaceum is capable of continuous growth in wet and cool soils at a high carbon dioxide
partial pressure which gives it an advantage over other soil microorganisms and enables it to
escape potential antagonism (Forbes and Dickinson, 1977). Thus, warmer and wetter winter
conditions may change the importance of F. avenaceum in the future. We observed a severe
outbreak of F. avenaceum on EFB33 when a frost-free winter period was followed by severe
frost and subsequent higher than normal temperatures (see Chapter 2).

FOOT AND ROOT ROT OF PEAS
63
3.5 References Ali, S.M., Nitschke, L.F., Dube, A.J., Krause, M.R., Cameron, B., 1978. Selection of pea lines
for resistance to pathotypes of Ascochyta pinodes, A. pisi and Phoma medicaginis var. pinodella. Crop Pasture Sci. 29, 841–849.
Ali, S.M., Sharma, B., Ambrose, M.J., 1993. Current status and future strategy in breeding pea to improve resistance to biotic and abiotic stresses. Euphytica 73, 115–126.
Bateman, G.L., Gutteridge, R.J., Gherbawy, Y., Thomsett, M.A., Nicholson, P., 2007. Infection of stem bases and grains of winter wheat by Fusarium culmorum and F. graminearum and effects of tillage method and maize-stalk residues. Plant Pathol. 56, 604–615.
Berkenkamp, B., Vaartnou, H., 1972. Fungi associated with rape root rot in Alberta. Can. J. Plant Sci. 52, 973–976.
Blume, M.C., Harman, G.E., 1979. Thielaviopsis basicola: A component of the pea root rot complex in New York State. Phytopathology 69, 785–788.
Bodker, L., Leroul, N., Smedegaard-Petersen, V., 1993. Influence of pea cropping history on disease severity and yield depression. Plant Dis. 77, 896–900.
Boros, L., Wawer, A., 2009. Garden pea varietal susceptibility to Mycosphaerella pinodes and its effect on yield components of single plants. Veg. Crops Res. Bull. 70.
Bowen, J.K., Lewis, B.G., Matthews, P., 1997. Discovery of the teleomorph of Phoma medicaginis var pinodella in culture. Mycol. Res. 101, 80–84.
Bretag, T., Price, T., Keane, P., 1995. Importance of seed-borne inoculum in the etiology of the Ascochyta blight complex of field peas (Pisum sativum L.) grown in Victoria. Aust. J. Exp. Agric. 35, 525–530.
Bretag, T.W., Ramsey, M., 2001. Foliar diseases caused by fungi. Ascochyta spp., in: Compendium of Pea Diseases and Pests. Second Edition. APS Press, The American Phytopathological Society, St. Paul Minnesota, pp. 24–28.
Campbell, C.L., Neher, D.A., 1994. Estimating disease severity and incidence, in: Epidemiology and Management of Root Diseases. Springer Berlin Heidelberg, pp. 117–147.
Campbell, K.A.G., Lipps, P.E., 1998. Allocation of resources: sources of variation in Fusarium head blight screening nurseries. Phytopathology 88, 1078–1086.
Cook, R.J., 1980. Fusarium foot rot of wheat and its control in the pacific northwest. Plant Dis. 64, 1061–1066.
Corbière, R., Gelie, B., Molinero, V., Spire, D., Agarwal, V.K., 1994. Investigations on seedborne nature of Mycosphaerella pinodes in pea seeds. Seed Res. 22, 26–30.
Cormack, M.W., 1951. Variation in the cultural characteristics and pathogenicity of Fusarium avenaceum and F. arthrosporioides. Can. J. Bot. 29, 32–45.
Feng, J., Hwang, R., Chang, K.F., Hwang, S.F., Strelkov, S.E., Gossen, B.D., Conner, R.L., Turnbull, G.D., 2010. Genetic variation in Fusarium avenaceum causing root rot on field pea: Genetic variation in Fusarium avenaceum. Plant Pathol. 59, 845–852.
Fletcher, J.D., Broadhurst, P.G., Bansal, R.K., 1991. Fusarium avenaceum : A pathogen of lentil in New Zealand. N. Z. J. Crop Hortic. Sci. 19, 207–210.
Forbes, R.S., Dickinson, C.H., 1977. Behaviour of Fusarium avenaceum in soil growth analysis plates. Trans. Br. Mycol. Soc. 69, 197–205.
Foroud, N.A., Chatterton, S., Reid, L.M., Turkington, T.K., Tittlemier, S.A., Gräfenhan, T., 2014. Fusarium diseases of Canadian grain crops: Impact and disease management strategies, in: Goyal, A., Manoharachary, C. (Eds.), Future Challenges in Crop Protection Against Fungal Pathogens, Fungal Biology. Springer New York, pp. 267–316.
Gaurilckiene, I., Cesnuleviciene, R., 2013. The susceptibility of pea (Pisum sativum L.) to Ascochyta blight under Lithuanian conditions. Zemdirb.-Agric. 100, 283–288.
Gossen, B.D., McDonald, M.R., Conner, R.L., Hwang, S.F., Chang, K.F., 2010. Significance of seed infection on epidemics of Mycosphaerella blight in field pea. Can. J. Plant Pathol. 32, 458–467.

FOOT AND ROOT ROT OF PEAS
64
Huisman, O.C., 1982. Interrelations of root growth dynamics to epidemiology of root-invading fungi. Annu. Rev. Phytopathol. 20, 303–327.
Hwang, S.F., Gossen, B.D., Turnbull, G.D., Chang, K.F., Howard, R.J., Thomas, A.G., 2000. Effect of temperature, seeding date, fungicide seed treatment and inoculation with Fusarium avenaceum on seedling survival, root rot severity and yield of lentil. Can. J. Plant Sci. 80, 899–907.
Jensen, B., Bødker, L., Larsen, J., Knudsen, J.C., Jørnsgaard, B., 2004. Specificity of soil-borne pathogens on grain legumes.
Kraft, J.M., Haware, M.P., Hussein, M.M., 1988. Root rot and wilt diseases of food legumes, in: Summerfield, R.J. (Ed.), World Crops: Cool Season Food Legumes, Current Plant Science and Biotechnology in Agriculture. Springer Netherlands, pp. 565–575.
Kraft, J.M., Pfleger, F.L., 2001. Compendium of pea diseases and pests. APS Press. Lamprecht, S.C., Marasas, W.F.O., Thiel, P.G., Schneider, D.J., Knox-Davies, P.S., 1986.
Incidence and Toxigenicity of seedborne Fusarium species from annual Medicago species in south Africa. Phytopathology 76, 1040–1042.
Le May, C., Jumel, S., Schoney, A., Tivoli, B., 2009. Ascochyta blight development on a new winter pea genotype highly reactive to photoperiod under field conditions. Field Crops Res. 111, 32–38.
Leslie, J.F., Summerell, B.A., 2006. The Fusarium laboratory manual. Blackwell Pub., Ames, Iowa.
Lin, Y.S., Cook, R.J., 1979. Supression of Fusarium roseum “avenaceum” by soil microorganisms. Phytopathology 69, 384–388.
Moussart, A., Tivoli, B., Lemarchand, E., Deneufbourg, F., Roi, S., Sicard, G., 1998. Role of seed infection by the Ascochyta blight pathogen of dried pea (Mycosphaerella pinodes) in seedling emergence, early disease development and transmission of the disease to aerial plant parts. Eur. J. Plant Pathol. 104, 93–102.
Nasir, M., Hoppe, H.-H., Ebrahim-Nesbat, F., 1992. The development of different pathotype groups of Mycosphaerella pinocles in susceptible and partially resistant pea leaves. Plant Pathol. 41, 187–194.
Olofsson, J., 1967. Root rot of canning and freezing peas in Sweden. Acta Agric. Scand. 17, 101–107.
Ondrej, M., Dostalova, R., Trojan, R., 2008. Evaluation of virulence of Fusarium solani isolates on pea. Plant Prot. Sci. 44, 9–18.
Onfroy, C., Tivoli, B., Corbiere, R., Bouznad, Z., 1999. Cultural, molecular and pathogenic variability of Mycosphaerella pinodes and Phoma medicaginis var. pinodella isolates from dried pea (Pisum sativum) in France. Plant Pathol. 48, 218–229.
Oyarzun, P.J., 1993. Bioassay to assess root rot in pea and effect of root rot on yield. Neth. J. Plant Pathol. 99, 61–75.
Oyarzun, P., Loon, J. van, 1989. Aphanomyces euteiches as a component of the complex of foot and root pathogens of peas in Dutch soils. Neth. J. Plant Pathol. 95, 259–264.
Persson, L., Bodker, L., Larsson-Wikström, M., 1997. Prevalence and pathogenicity of foot and root rot pathogens of pea in south Scandinavia. Plant Dis. 81, 171–174.
Pettitt, T., Parry, D., Polley, R., 1996. Effect of temperature on the incidence of nodal foot rot symptoms in winter wheat crops in England and Wales caused by Fusarium culmorum and Microdochium nivale. Agric. For. Meteorol. 79, 233–242.
Pflughöft, O., 2008. Pilzkrankheiten in Körnerfuttererbsen (Pisum sativum L.) – Diagnose, Epidemiologie, Ertragsrelevanz und Bekämpfung. Der Fakultät für Agrarwissenschaften der Georg-August-Universität Göttingen.
Pflughöft, O., Merker, C., von Tiedemann, A., Schäfer, B.C., 2012. Zur Verbreitung und Bedeutung von Pilzkrankheiten in Körnerfuttererbsen (Pisum sativum L.) in Deutschland. Gesunde Pflanz. 64, 39–48.
Roger, C., Tivoli, B., 1996. Effect of culture medium, light and temperature on sexual and asexual reproduction of four strains of Mycosphaerella pinodes. Mycol. Res. 100, 304–306.

FOOT AND ROOT ROT OF PEAS
65
Roger, C., Tivoli, B., Huber, L., 1999. Effects of temperature and moisture on disease and fruit body development of Mycosphaerella pinodes on pea (Pisum sativum). Plant Pathol. 48, 1–9.
Rush, C.M., Kraft, J.M., 1986. Effects of inoculum density and placement on Fusarium root rot of peas. Phytopathology 76, 1325–1329.
Satyaprasad, K., Bateman, G.L., Ward, E., 2000. Comparison of Isolates F. avenaceum from white lupin and other crops by pathogenicity tests, DNA analyses and Vegetative compatibility tests. J. Phytopathol. 148, 211–219.
Setti, B., Bencheikh, M., Henni, J., Neema, C., 2009. Comparative aggressiveness of Mycosphaerella pinodes on peas from different regions in western Algeria. Phytopathol. Mediterr. 48.
Skovgaard, K., Bødker, L., Rosendahl, S., 2002. Population structure and pathogenicity of members of the Fusarium oxysporum complex isolated from soil and root necrosis of pea (Pisum sativum L.). FEMS Microbiol. Ecol. 42, 367–374.
Smiley, R.W., Patterson, L.-M., 1996. Pathogenic fung associated with Fusarium foot rot of winter wheat in the semiarid pacific northwest. Plant Dis. 80, 944–949.
Thung, M., Rao, I.M., 1999. Integrated management of abiotic stresses, in: Singh, S.P. (Ed.), Common Bean Improvement in the Twenty-First Century, Developments in Plant Breeding. Springer Netherlands, pp. 331–370.
Tivoli, B., Banniza, S., 2007. Comparison of the epidemiology of ascochyta blights on grain legumes. Eur. J. Plant Pathol. 119, 59–76.
Tu, J.C., 1987. Integrated control of the pea root rot disease complex in Ontario. Plant Dis. 9–13.
Wallen, V.R., 1974. Influence of three Ascochyta diseases of peas on plant development and yield. Can. Plant Dis. Surv. 54, 86–90.
Zhang, J.X., Fernando, W.G.D., Xue, A.G., 2003. Virulence and genetic variability among isolates of Mycosphaerella pinodes. Plant Dis. 87, 1376–1383.
Zhang, J.X., Xue, A.G., Cober, E.R., Morrison, M.J., Zhang, H.J., Zhang, S.Z., Gregorich, E., 2013. Prevalence, pathogenicity and cultivar resistance of Fusarium and Rhizoctonia species causing soybean root rot. Can. J. Plant Sci. 93, 221–236.

DISEASE SUPPRESSION WITH COMPOST APPLICATION
66
4. EFFECTS OF COMPOST APPLICATION ON FOOT AND ROOT ROT OF SPRING AND WINTER PEA VARIETIES CAUSED BY FUSARIUM AVENACEUM, FUSARIUM SOLANI, PHOMA
MEDICAGINIS AND DIDYMELLA PINODES
Abstract
In order to evaluate the potential of yard waste compost (YWC) to suppress foot rot disease of pea caused by Didymella pinodes, Phoma medicaginis, Fusarium solani f. sp pisi and F.
avenaceum, pot experiments were carried out under controlled conditions in sterile sand and non-sterilized field soil. In addition, effects of application rate of YWC and of reduced temperatures on the disease suppression were assessed. The spring pea Santana and winter pea EFB33 were inoculated with spore suspensions of the pathogens at sowing or left uninoculated, and grown in sterile sand and field soil amended with and without YWC (20% v/v). Part of the experiments included γ irradiated compost as additional controls. After three weeks disease severity was assessed and yield parameters measured. Application of compost to sterile sand significantly reduced the disease index (DI) of Santana caused by F.
avenaceum, P. medicaginis and D. pinodes. Disease caused by F. solani was low (mean DI 3.0) and no effect of compost application was observed. The suppressiveness of compost was reduced through γ irradiation for D. pinodes and P. medicaginis, while disease symptoms of F. avenaceum and F. solani were even higher than in inoculated control without compost. When plants were grown in non-sterilized soil symptoms due to F. avenaceum, P.
medicaginis and D. pinodes were lower than in the sterile sand while for F. solani they were on the same level. Compost application to soil did not have an effect on disease severity. However, when sterile compost was applied disease severity due to all pathogens was significantly increased compared to normal compost in soil. With the exception of the treatment inoculated with F. avenaceum, application of non- and sterilized compost had positive effects on the plant weight increasing it between 25 % and 60 % depending on pathogen and growing substrate. When lower rates of compost were applied (3.5% v/v) to sand, disease symptoms were significantly reduced. Temperature did not have significant effects on level of disease suppression by compost. The YWC used in the trials could suppress disease caused by the tested pathogens and the intensity of suppression depended on variety, pathogens and growing media. Results indicate that it is likely that suppression of disease caused by Fusarium spp. is biological in origin and for D. pinodes
and P. medicaginis chemical and physical properties of compost are also contributing to the suppression of foot and root rot disease.
Keywords: compost, F. avenaceum, P. medicaginis, D. pinodes, F. solani, foot rot, peas

DISEASE SUPPRESSION WITH COMPOST APPLICATION
67
4.1 Introduction
It is widely recognized that application of high quality compost can effectively suppress
diseases caused by a range of soil borne pathogens on a range of different crops (Aviles et
al., 2011; Bonanomi et al., 2007; Hadar, 2011; Hoitink, 2006; Hoitink et al., 1997; Hoitink and
Fahy, 1986; Noble, 2011; Noble and Coventry, 2005; Termorshuizen et al., 2006; Yogev et
al., 2006). Success of compost in disease suppression depends on the type of substrate,
maturity, and the composting process used (Hoitink and Fahy, 1986). Inconsistency in the
level of disease suppression between pot and field experiments is often reported (Hoitink et
al., 1997; Litterick et al., 2004; Noble and Roberts, 2003).
Suppressive effects of composts are mainly due to beneficial microorganisms (Hoitink and
Boehm, 1999) which has been supported by loss of suppressivness after heat treatment, e.g.
pasteurization or sterilization. Hoitink and Boehm (1999) described four mechanisms
involved in compost induced microbial disease suppression: competition for nutrients
between compost inhabiting microbes and pathogens, antibiosis due to production of
different metabolites by beneficial microorganisms, parasitism and predation of pathogens by
beneficial nematodes, mites or fungi, and induced resistance. Chemical and physical
properties as affected by compost age and level of organic matter decomposition, C:N ratio,
cellulose and lignin content, nitrogen content and form, pH, electrical conductivity (EC),
organic chemicals released by compost, etc., are also contributing to disease suppression
(Bonanomi et al., 2007; Hoitink, 2006; Hoitink et al., 1997; Hoitink and Fahy, 1986; Recycled
Organics Unit, 2006). Besides direct suppressive effects, compost application also improves
soil quality by adding organic matter and improving various physical soil properties.
Depending if the suppression is caused by the action of a specific organism or the total
microbial community two categories of suppression can be distinguished - specific and
general (Hoitink, 2006; Hoitink et al., 1997). These two types do not exclude each other.
General suppression is the result of the activity of overall microbial biomass (Recycled
Organics Unit, 2006), and it cannot be transferred. In contrast, in specific suppression one or
several groups of organisms are responsible for suppression and it can be transferred to
other substrates/soils by adding a small amount of suppressive substrate/soil (Aviles et al.,
2011).
Peas are strongly affected by root and foot rots that limit their production worldwide (Ali et al.,
1993; Bretag and Ramsey, 2001). A complex of more than 20 different species of soil-borne
fungal pathogens can cause foot and root rot of peas (Blume and Harman, 1979; Bodker et
al., 1993; Kraft et al., 1988; Olofsson, 1967; Persson et al., 1997). The most important
species causing foot and root rot on peas are Didymella pinodes (syn Mycospharella

DISEASE SUPPRESSION WITH COMPOST APPLICATION
68
pinodes), Phoma medicaginis, Fusarium spp. (F. solani f. sp pisi, F. oxysporum f.sp pisi, F.
avenaceum, F. culmorum), Pythium spp., Thielaviopsis basicola, Aphanomyces euteiches,
and Rhizoctonia solani (Feng et al., 2010; Kraft et al., 1988; Nasir et al., 1992; Oyarzun,
1993; Persson et al., 1997; Pflughöft et al., 2012). Prevalence, dominance and importance of
the single pathogens vary greatly depending on location, climate, and agricultural practice
(Gaurilckiene and Cesnuleviciene, 2013; Jensen et al., 2004; Persson et al., 1997).
Especially impaired soil structure through compaction and inadequate drainage play a crucial
role for pea health (Solaiman et al., 2007).
Regular applications of organic amendments, especially high quality composts, could help
improve soil structure and supressiveness against pathogen. However, suppressiveness of
compost against one pathogen does not necessarily mean that it will suppress another
pathogen. This variability in the disease suppressiveness has been reported also for
composts made of similar substrates and under similar conditions (Noble and Coventry,
2005). Given the high variation in the response Termorshuizen et al. (2006) emphasize that
prediction of potential suppression of disease should be made based on specific
pathosystems only. Also, in pot trials successful suppression is usually achieved with
application rates of 30 to 50% v/v of compost to the potting mixes (Noble and Coventry,
2005; Termorshuizen et al., 2006). However, in organic farming in Germany application rate
of externally brought compost is limited by law to only 5 t dry matter ha -1 per year. Even at
this low rate application of high quality composts have been successful in reducing R. solani
in potatoes in the field (Bohne et al., 2013; Schulte-Geldermann et al., 2009). Also, raw
application together with pea seeds was successful in suppressing P. medicaginis. The
research presented here was conducted to determine if a locally produced Yard Waste
Compost (YWC) can suppress root and foot rot disease of peas caused by the four
pathogens Fusarium avenaceum, Fusarium solani f. sp. pisi, Phoma medicaginis var.
pinodella, and Didymella pinodes. This was assessed in two growing substrates, sterile sand
and non-sterilized field soil on a spring and winter pea variety. In additional experiments the
effects of application rate of YWC and of reduced temperatures on the disease suppression
by compost application were assessed.

DISEASE SUPPRESSION WITH COMPOST APPLICATION
69
4.2 Material and Methods All experiments were replicated five times in a completely randomized design.
4.2.1 Potential of yard waste compost to suppress foot and root rot of peas caused by
F. avenaceum, F. solani, P. medicaginis and D. pinodes
The winter pea variety EFB33 and spring variety Santana (KWS LOCHOW, Gmbh) were
used for the experiments. EFB33 is a regularly leafed, small seeded variety with thousand
seed weight of about 130 g and violet flowers. Santana is a spring pea widely grown in
Germany. It is semi-leafless, white flowering with white seed coat and a thousand seed
weight of 250 to 300g. Prior to use, pea seeds were surface disinfected with 70% ethanol for
five minutes, rinsed with running distilled water and air dried under a laminar flow cabinet. All
defective and damaged seeds were discarded.
Sterilized yellow sand and non-sterilized field soil, without or amended with non-sterilized
and sterilized compost were used as growing media (Tab 4.1). Sand was autoclaved for 20
minutes at 121°C and stored in a closed container until the beginning of the trial. The field
soil for the trial was collected from the experimental field of the University of Kassel at the
Neu-Eichenberg. The soil type is a deep Haplic Luvisol from loess with 78 soil points
(Wildhagen, 1998). Locally produced mature YWC made of garden trimmings was used in
the trial. In order to determine whether the effects of YWC were biological in origin, half of the
compost used was gamma irradiated with 31.5 kGy one week prior to the experiment and
stored at 4°C until the beginning of the trial.
At the day of sowing, compost was thoroughly mixed with sand or soil at the rate of 20% v/v.
Approximately 1.3 kg of substrate was filled in square pots of 1 L (12 x 12 x 11 cm) and eight
pea seeds per pot were sown at 2 cm depth. Following sowing, pots were inoculated with
spore suspensions of F. avenaceum, F. solani, P. medicaginis and D. pinodes at a rate of
104 spores g-1 of substrate. Five isolates for each pathogen (Tab. 3.1, Chapter 3.2.1) were
used in a mixture. Preparation of inoculum was as described in section 3.2.
Pots were watered every second day and treatments with sole sand as a substrate were
fertilized with the complex N:P:K fertilizer Wuxal Super (8:8:6 + trace elements) in the
amount corresponding to a total of 100 mg of N L-1 substrate. Plants were grown for 21 days
with a day/night temperature regime of 19/16°C.
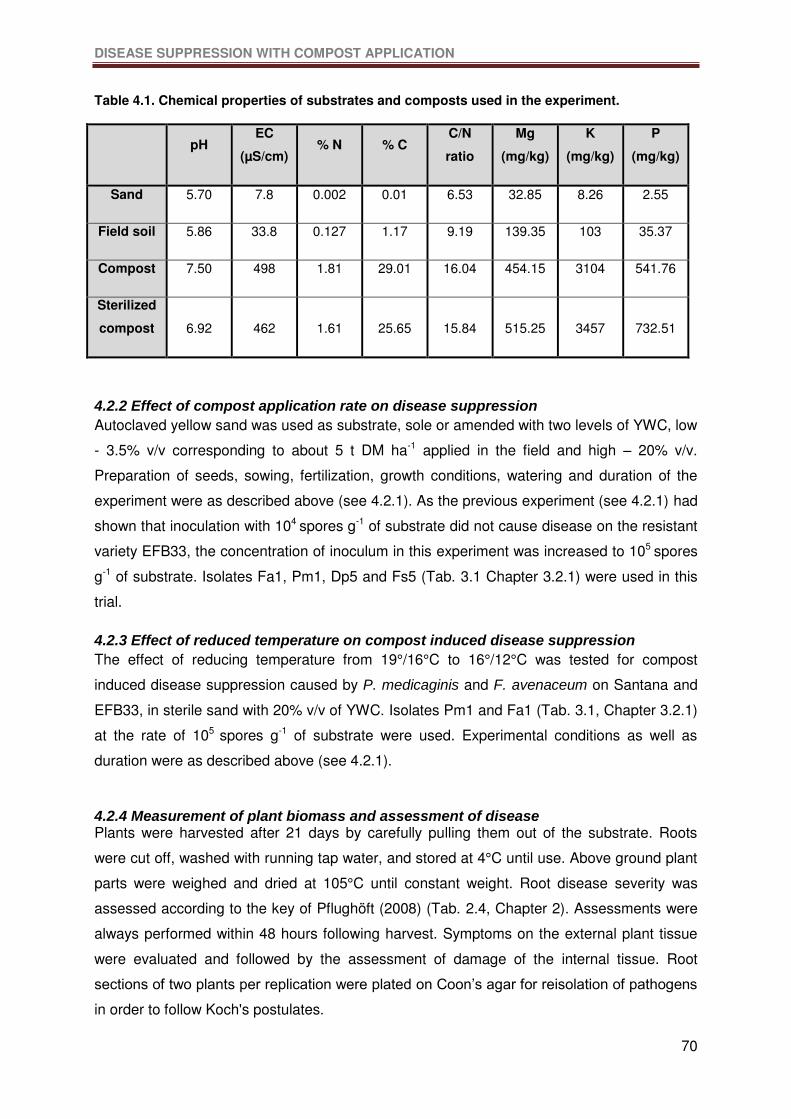
DISEASE SUPPRESSION WITH COMPOST APPLICATION
70
Table 4.1. Chemical properties of substrates and composts used in the experiment.
pH
EC
(µS/cm) % N % C
C/N
ratio
Mg
(mg/kg)
K
(mg/kg)
P
(mg/kg)
Sand 5.70 7.8 0.002 0.01 6.53 32.85 8.26 2.55
Field soil 5.86 33.8 0.127 1.17 9.19 139.35 103 35.37
Compost 7.50 498 1.81 29.01 16.04 454.15 3104 541.76
Sterilized
compost 6.92 462 1.61 25.65 15.84 515.25 3457 732.51
4.2.2 Effect of compost application rate on disease suppression
Autoclaved yellow sand was used as substrate, sole or amended with two levels of YWC, low
- 3.5% v/v corresponding to about 5 t DM ha-1 applied in the field and high – 20% v/v.
Preparation of seeds, sowing, fertilization, growth conditions, watering and duration of the
experiment were as described above (see 4.2.1). As the previous experiment (see 4.2.1) had
shown that inoculation with 104 spores g-1 of substrate did not cause disease on the resistant
variety EFB33, the concentration of inoculum in this experiment was increased to 105 spores
g-1 of substrate. Isolates Fa1, Pm1, Dp5 and Fs5 (Tab. 3.1 Chapter 3.2.1) were used in this
trial.
4.2.3 Effect of reduced temperature on compost induced disease suppression
The effect of reducing temperature from 19°/16°C to 16°/12°C was tested for compost
induced disease suppression caused by P. medicaginis and F. avenaceum on Santana and
EFB33, in sterile sand with 20% v/v of YWC. Isolates Pm1 and Fa1 (Tab. 3.1, Chapter 3.2.1)
at the rate of 105 spores g-1 of substrate were used. Experimental conditions as well as
duration were as described above (see 4.2.1).
4.2.4 Measurement of plant biomass and assessment of disease Plants were harvested after 21 days by carefully pulling them out of the substrate. Roots
were cut off, washed with running tap water, and stored at 4°C until use. Above ground plant
parts were weighed and dried at 105°C until constant weight. Root disease severity was
assessed according to the key of Pflughöft (2008) (Tab. 2.4, Chapter 2). Assessments were
always performed within 48 hours following harvest. Symptoms on the external plant tissue
were evaluated and followed by the assessment of damage of the internal tissue. Root
sections of two plants per replication were plated on Coon’s agar for reisolation of pathogens
in order to follow Koch's postulates.

DISEASE SUPPRESSION WITH COMPOST APPLICATION
71
4.2.5 Data processing and analysis
Disease index (DI) for each plant was calculated as the average of the external and internal
lesion score. As all plants in sand controls emerged, missing plants in the treatments were
considered dead and score 9 was given for both external and internal tissue. The mean DI of
all plants in each pot was calculated to present one replication, and further analyses were
done on these values. Biomass production was expressed as fresh and/or dry weight per
plant. Weights of all plants in one pot were measured together and then divided with the
number of plants per pot and represents one replication.
Analyses of variance (ANOVA) were applied in order to analyze the effect of the pathogens,
substrate and temperature on the DI, plant weights and survival rates. Prior to ANOVA, data
were checked for normality using Shapiro-Wilks-W-Test. Mean separations were made by
Tukey honest significant differences (HSD) test at the P < 0.05. All statistical analyses were
conducted with statistical program R version 2.15.2.

DISEASE SUPPRESSION WITH COMPOST APPLICATION
72
4.3 Results
4.3.1 Potential of yard waste compost to suppress foot and root rot of peas caused by
F. avenaceum, F. solani, P. medicaginis and D. pinodes
Plants grown in non-inoculated sand control with and without compost were healthy and had
mean DI below 2.5. In the treatment with γ irradiated compost, DI of the control plants was
significantly higher, however still below 3. In contrast, plants grown in the non-inoculated soil
controls had moderate to severe disease symptoms in treatments with and without compost
(Tab. 4.2).
Application of compost to sterile sand significantly reduced the DI of Santana plants
inoculated with F. avenaceum, P. medicaginis and D. pinodes (Tab. 4.2). In the treatment
inoculated with F. solani, disease severity was low (mean DI 3.0) and no effect of compost
application (mean DI 2.5) was observed. The disease suppressiveness of compost was
significantly reduced or lost through γ irradiation for D. pinodes and P. medicaginis,
respectively while disease symptoms of F. avenaceum and F. solani were significantly higher
than in the non-amended sand.
When non-sterilized soil was used as growing medium, the non-inoculated plants showed
moderate symptoms with a mean DI of 4.8. This was not affected by the compost application.
Symptoms due to F. avenaceum, P. medicaginis and D. pinodes were lower than in the
sterile sand while for F. solani the DI was 3.9 in soil compared to 3.0 in sand (Tab. 4.2).
When sterile compost was used, disease severity due to all pathogens was significantly
increased compared to normal compost in soil. Compared to sand, symptoms caused by F.
avenaceum were less severe in soil with sterile compost while for F. solani, P. medicaginis
and D. pinodes severities were higher in soil than in sand.

DISEASE SUPPRESSION WITH COMPOST APPLICATION
73
Table 4.2. Disease Index (DI) of Santana in sterile sand and non-sterilized field soil in non-inoculated control and treatments inoculated with F. avenaceum, F. solani, P. medicaginis and D. pinodes and amended with compost (no-without, N-not sterilized and S-sterilized compost). Different letters within one pathogen and one substrate indicate significant differences in DI among compost treatments at P < 0.05 (Tukey’s HSD test).
Pathogen Compost Sand Soil Control no 1.5 b 4.8
N 2.0 b 4.5 S 3.0 a 6.0
F. avenaceum no 5.1 b 3.8 b N 2.9 c 3.7 b S 8.3 a 5.9 a
F. solani no 3.0 b 3.9 ab N 2.5 b 3.0 b S 5.3 a 5.4 a
P. medicaginis no 4.7 a 3.6 b N 3.2 b 3.5 b S 4.4 a 6.8 a
D. pinodes no 4.6 a 4.2 ab N 2.9 c 3.4 b S 3.6 b 5.5 a
With the exception of the treatment inoculated with F. avenaceum, application of compost,
regardless if it was sterilized or not, had positive effects on the plant weights of Santana in
sand (Fig. 4.1). The fresh weight increases ranged between 35% for F. solani and 55% for P.
medicaginis and D. pinodes. Application of non-sterilized compost in the treatment with F.
avenaceum significantly increased fresh weight (50%) while with γ irradiated compost
biomass of plants was reduced by 60%. In soil, significant increases of the fresh weight
following non-sterile compost application were only observed in the non-inoculated controls
and when inoculted F. solani (Fig. 4.1). Sterilized compost had no significant effect on plant
weights. Disease severity of EFB33 was overall low (DI between 2 and 3) and was not
affected by compost application, regardless if sand or soil were used as growing substrate
(see Appendix Fig. A. 4.1). Both types of compost positively affected plant weights (see
Appendix Fig. A. 4.2). Increases in fresh weights were in the range between 25% and 50%,
depending on the pathogen and growing substrate. Dry weights of both varieties followed
similar pattern as the fresh weights (see Appendix Fig. A. 4.3 and A. 4.4).
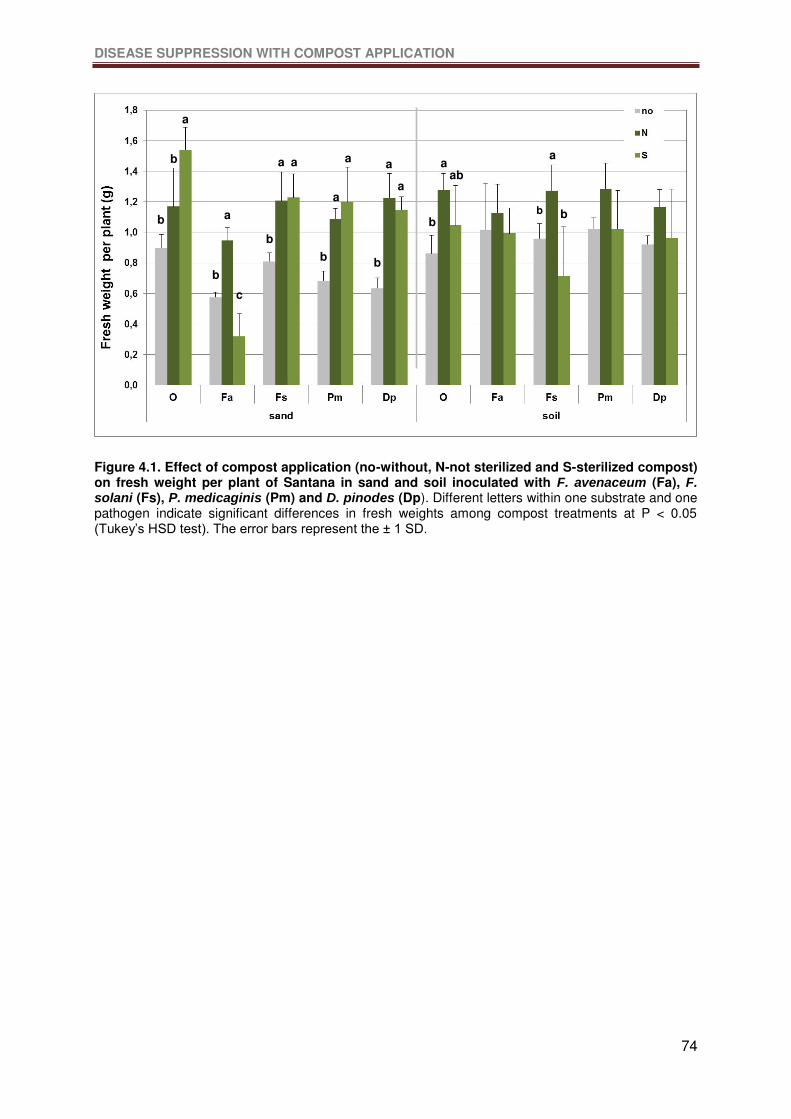
DISEASE SUPPRESSION WITH COMPOST APPLICATION
74
Figure 4.1. Effect of compost application (no-without, N-not sterilized and S-sterilized compost) on fresh weight per plant of Santana in sand and soil inoculated with F. avenaceum (Fa), F. solani (Fs), P. medicaginis (Pm) and D. pinodes (Dp). Different letters within one substrate and one pathogen indicate significant differences in fresh weights among compost treatments at P < 0.05 (Tukey’s HSD test). The error bars represent the ± 1 SD.
b
b
a
b
a a
b
a
a
b
a
a
c
b
a b
a ab
b
a
b

DISEASE SUPPRESSION WITH COMPOST APPLICATION
75
4.3.2 Effect of compost application rate on disease suppression
Increasing inoculum concentration to 105 spores g-1 of substrate resulted in slightly higher DI
for both varieties. All DIs of Santana were lower in the compost treatments than in the
controls (Fig. 4.2). The reductions were significant for P. medicaginis and the high amount of
compost, and for D. pinodes at both compost rates. Application of 20% v/v compost in the
system with F. avenaceum and EFB33 significantly reduced disease severity, while both
compost rates reduced P. medicaginis.
Figure 4.2. Disease index of Santana and EFB33 inoculated with F. avenaceum (Fa), F. solani (Fs), P. medicaginis (Pm) and D. pinodes (Dp) and amended with compost (no-without, low- 3.5% v/v and high- 20% v/v). O is the non-inoculated control. The horizontal line in the boxplot shows the median, the bottom and tops of the box the 25th and 75th percentiles and the vertical lines the minimum and maximum values; outliers as single points. Mean values of DI are marked with triangles. Different letters within variety and pathogen are indicating significant differences in DI at P < 0.05 (Tukey’s HSD test).
Compost application positively affected biomass of Santana plants, except when inoculated
with F. solani, where compost had no effect (Fig. 4.3). For treatments inoculated with F.
avenaceum and P. medicaginis increase in biomass was significant when compost was
applied at the high rate, and for D. pinodes at both compost treatments. In contrast to
Santana, weights of EFB33 were not affected by compost application (Fig. 4.3).
a ab b a b b
ab a b a b b

DISEASE SUPPRESSION WITH COMPOST APPLICATION
76
Figure 4.3. Effect of compost application rate (no-without, low- 3.5% v/v and high- 20% v/v) on the fresh weight of (A) Santana and (B) EFB33 plants inoculated with 105 spores g-1 of substrate
of F. avenaceum (Fa), F. solani (Fs), P. medicaginis (Pm) and D. pinodes (Dp). Different letters within pathogen are indicating significant differences in fresh weights at P < 0.05 (Tukey’s HSD test). The error bars represent the ± 1 SD.
b
b
a ab
a
b ab
a
b
a
b
a
A)
B)
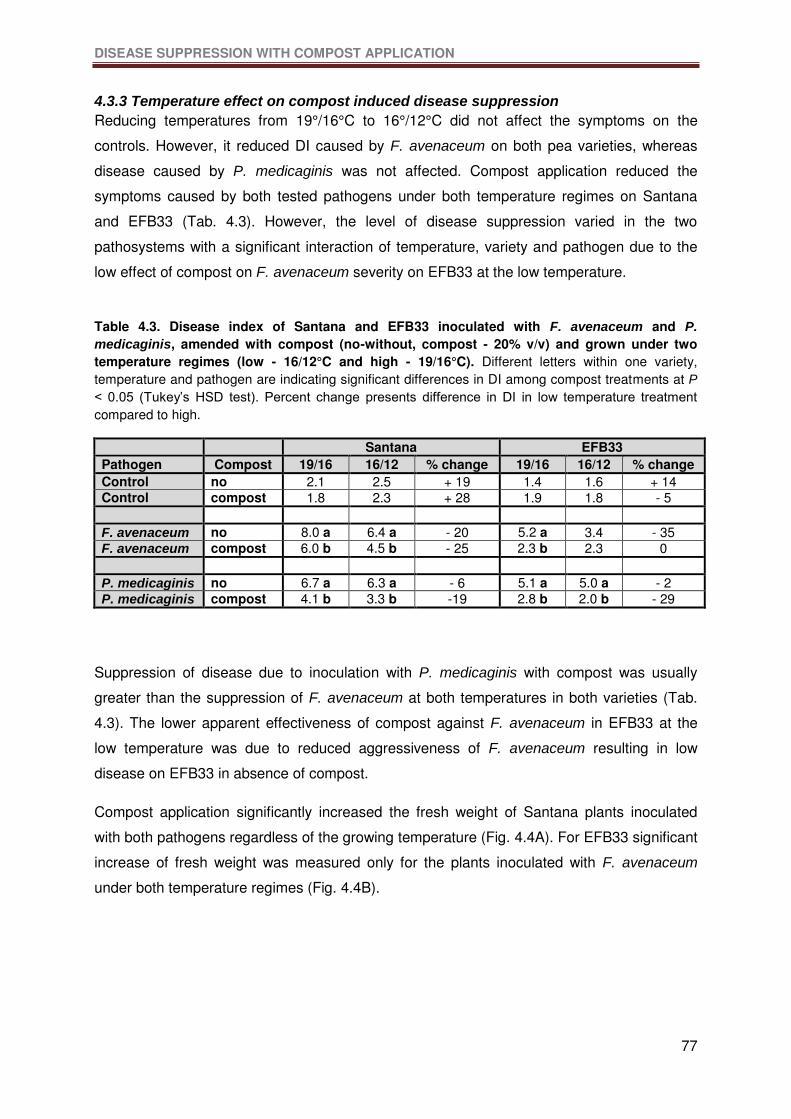
DISEASE SUPPRESSION WITH COMPOST APPLICATION
77
4.3.3 Temperature effect on compost induced disease suppression
Reducing temperatures from 19°/16°C to 16°/12°C did not affect the symptoms on the
controls. However, it reduced DI caused by F. avenaceum on both pea varieties, whereas
disease caused by P. medicaginis was not affected. Compost application reduced the
symptoms caused by both tested pathogens under both temperature regimes on Santana
and EFB33 (Tab. 4.3). However, the level of disease suppression varied in the two
pathosystems with a significant interaction of temperature, variety and pathogen due to the
low effect of compost on F. avenaceum severity on EFB33 at the low temperature.
Table 4.3. Disease index of Santana and EFB33 inoculated with F. avenaceum and P.
medicaginis, amended with compost (no-without, compost - 20% v/v) and grown under two temperature regimes (low - 16/12°C and high - 19/16°C). Different letters within one variety, temperature and pathogen are indicating significant differences in DI among compost treatments at P < 0.05 (Tukey’s HSD test). Percent change presents difference in DI in low temperature treatment compared to high.
Santana EFB33 Pathogen Compost 19/16 16/12 % change 19/16 16/12 % change Control no 2.1 2.5 + 19 1.4 1.6 + 14 Control compost 1.8 2.3 + 28 1.9 1.8 - 5 F. avenaceum no 8.0 a 6.4 a - 20 5.2 a 3.4 - 35 F. avenaceum compost 6.0 b 4.5 b - 25 2.3 b 2.3 0 P. medicaginis no 6.7 a 6.3 a - 6 5.1 a 5.0 a - 2 P. medicaginis compost 4.1 b 3.3 b -19 2.8 b 2.0 b - 29
Suppression of disease due to inoculation with P. medicaginis with compost was usually
greater than the suppression of F. avenaceum at both temperatures in both varieties (Tab.
4.3). The lower apparent effectiveness of compost against F. avenaceum in EFB33 at the
low temperature was due to reduced aggressiveness of F. avenaceum resulting in low
disease on EFB33 in absence of compost.
Compost application significantly increased the fresh weight of Santana plants inoculated
with both pathogens regardless of the growing temperature (Fig. 4.4A). For EFB33 significant
increase of fresh weight was measured only for the plants inoculated with F. avenaceum
under both temperature regimes (Fig. 4.4B).

DISEASE SUPPRESSION WITH COMPOST APPLICATION
78
Figure 4.4. Effect of compost application (no-without, compost- 20% v/v) on the fresh weights of Santana (A) and EFB33 (B) plants inoculated with F. avenaceum (Fa) and P. medicaginis (Pm) and grown under different temperature regimes (low - 16/12°C and high - 19/16°C). Asterisk indicates significant differences at P < 0.05 among treatments with and without compost within one temperature regime and pathogen. The error bars represent the ± 1 SD.
A
B
*
*
*
*
*
*
*

DISEASE SUPPRESSION WITH COMPOST APPLICATION
79
4.4 Discussion The Yard Waste Compost (YWC) we used could suppress root and foot rot of peas caused
by Fusarium avenaceum, Fusarium solani f. sp. pisi, Phoma medicaginis var. pinodella, and
Didymella pinodes. However, the intensity of suppression depended on pea varieties,
pathogens and growing media used. Disease severity on EFB33 was low and no effect of
compost application was observed. An increase of the inoculum concentration in the trials on
compost amount and temperature only slightly increased the severity of disease. A
temperature effect was observed only in the system with EFB33 and F. avenaceum where
the disease suppression through compost application was lower at the reduced temperature
because the overall aggressiveness of F. avenaceum was also low.
After γ sterilization of the compost, the suppressive effect was lost in the pathosystems with
Fusarium spp. suggesting that suppression was biological in origin. Moreover, severity of
symptoms caused by F. avenaceum and F. solani increased after incorporation of sterilized
compost. In contrast, part of the suppressive effects against P. medicaginis and D. pinodes
apparently remained even after γ sterilization.
In the case of root disease in order to observe the full effect of a pathogen, assessment of
severity of disease symptoms on plant roots and measurements of above-ground plant
growth parameters have to be considered (Campbell and Neher, 1994). Our results show
that when sterilized sand was used as growing substrate compost application successfully
reduced disease caused by F. avenaceum, P. medicaginis, and D. pinodes on Santana.
Furthermore, fresh weight was significantly increased with compost application in all
treatments including controls. While the overall effect of all three pathogens on the
susceptible pea variety was minimized through compost application, clearly, the mineral
fertilization could not compensate for additional beneficial effects of compost.
The increased disease caused by Fusarium spp. in sand amended with γ-irradiated compost
in our trial could be due to active colonization of the nutrient rich sterile compost by the
pathogens enhancing their activity and population buildup leading to more severe symptoms
in comparison to the inoculated control. The fact that in the systems with D. pinodes
sterilized compost decreased DI and the trend was similar with P. medicaginis suggests that
different mechanisms of suppression may be involved and that the observed reduction in
symptoms caused by Fusarium spp. is biological in origin, whereas chemical and physical
properties of compost are playing an additional role in the suppression of disease caused by
D. pinodes and P. medicaginis on peas. Physical and chemical characteristics are known to
affect the suppressive effects of composts. Incorporation of composts in growing substrates
increases air capacity and in addition to positively affecting plant growth, it also reduces
severity of root rots (Aviles et al., 2011).

DISEASE SUPPRESSION WITH COMPOST APPLICATION
80
When Santana was grown in soil, compost application did not result in significant reduction of
DI, and in the treatments with γ-irradiated compost disease symptoms were even more
severe than without compost application for all four pathogens. Serra-Wittling et al. (1996)
reported a higher decrease in the population of F. oxysporum f. sp lini in soil treated with
control compost compared with heat treated compost, however, intensity of disease
suppression was at the same level for both composts. In contrast to our results, Filippi and
Pera (1989) observed suppression of Fusarium wilt in carnations caused by Fusarium
oxysporum f. sp. dianthi when sterilized compost was incorporated in the soil. The disease
reduction was attributed to the activity of the fraction of native soil microorganisms that was
more competitive for nutrients than the pathogen population and acted as antagonists, and
not to the activity of the microflora present in compost. In our trial, relatively high severity of
disease symptoms in the non-inoculated control suggest that soil borne inoculum was
present in the field soil used in the trial. Fusarium solani was mainly isolated from
symptomatic plants grown in the non-inoculated soil. An explanation for increased disease in
soil amended with sterilized compost can be that the level of native microflora in the field soil
used in our trial was not high enough to compete with the existing inoculum and the
introduced pathogens for the available food source provided by sterilized compost.
Pathogens were most likely more efficient in the colonization of compost, which resulted in
faster population growth and consequently an increase in disease severity.
An additional factor that could have contributed to the loss of suppressiveness through γ-
irradiation are changes in the chemical composition and pH of the compost. The combination
of higher amounts of available Mg, K, and P in the sterilized compost, a lower pH, and
microbial-free organic matter could have increased the advantage of pathogens even more
and may thus have resulted in increased disease severity. Borrero et al. (2009, 2004)
showed a strong correlation between pH and disease suppression of Fusarium wilt in tomato
(F. oxysporum f. sp. lycopersici) and carnations (F. oxysporum f. sp dianthi). Namely,
substrates with low pH were conducive to wilt, whereas adding compost increased the pH of
the substrate and disease was successfully suppressed.
The disease suppressive effect of compost generally increased with rate of application, and
rates of at least 20% (v/v) are usually recommended to consistently obtain a disease
suppressive effect, especially when inert material like peat is used as growing substrate
(Noble and Coventry, 2005). Although we observed a tendency of lower DI in systems with
20% v/v of compost, there were no significant differences in the DI between treatments
amended with the two levels of compost. Lumsden et al. (1983) reported significant reduction
in the disease severity of Fusarium wilt in melons (Fusarium oxysporum f. sp. melonis),
crown rot (Phytophtora capsici) in peppers, blights (Pythium myriotylum) on bean and lettuce

DISEASE SUPPRESSION WITH COMPOST APPLICATION
81
drop caused by Sclerotinia minor after amending soil with 10% v/v composted sewage
sludge in a pot trial. In contrast, in the pea - Fusarium solani f. sp pisi pathosystem foot rot
increased by 76% compared to the control without compost application. Schüler et al. (1993)
showed that foot rot caused by D. pinodes could be suppressed by amending sand and also
field soil with 30% of compost, whereas an amount of 10% did not affect the disease.
However, an important difference to our study is that they introduced the pathogen in the
system by soaking pea seeds in the inoculum suspension. This led to seed borne inoculum
and infection may have started earlier compared to our study. Watering the substrate with
the spore suspension after sowing in our system gave more time for the physical, chemical
and microbial changes resulting from compost incorporation, potentially hindering infection or
at least slowing down the pathogens. This gave the compost a chance to suppress the
pathogens before infection occurred.
Our results show that the YW compost we used can suppress foot disease on the
susceptible variety Santana caused by F. avenaceum, P. medicaginis, and D. pinodes, by
reducing the intensity of disease symptoms and promoting plant growth. Although high
amounts of compost (30-50% v/v) are recommended for successful disease suppression,
application of 3.5% compost in sterile sand was enough to achieve suppression of disease
symptoms for all four pathogens. Moreover, results we obtained indicate that different
mechanisms of suppression may be involved in the reduction of symptoms caused by
Fusarium spp. and by D. pinodes and P. medicaginis on peas. It is likely that suppression of
disease caused by Fusarium spp. is biological in origin and for D. pinodes and P.
medicaginis in addition chemical and physical properties of compost were contributing to the
suppression of foot and root rot disease.

DISEASE SUPPRESSION WITH COMPOST APPLICATION
82
4.5 References Ali, S.M., Sharma, B., Ambrose, M.J., 1993. Current status and future strategy in breeding
pea to improve resistance to biotic and abiotic stresses. Euphytica 73, 115–126. Aviles, M., Borrero, M., Trillas, M.I., 2011. Review on compost as inducer of disease
suppression in plants grown in soilless culture. Dyn. Soil Dyn. Plant 5, 1–11. Blume, M.C., Harman, G.E., 1979. Thielaviopsis basicola : A component of the pea root rot
complex in New York State. Phytopathology 69, 785–788. Bodker, L., Leroul, N., Smedegaard-Petersen, V., 1993. Influence of pea cropping history on
disease severity and yield depression. Plant Dis. 77, 896–900. Bohne, B., Hensel, O., Bruns, C., 2013. Reihenapplikation von Komposten zur Kontrolle
bodenbürtiger Krankheiten – technische Lösungen für Kartoffeln und Körnerleguminosen.
Bonanomi, G., Antignani, V., Pane, C., Scala, F., 2007. Suppression of soilborne fungal diseases with organic amendments. J. Plant Pathol. 89, 311–324.
Borrero, C., Trillas, I., Aviles, M., 2009. Carnation Fusarium wilt suppression in four composts. Eur. J. Plant Pathol. 123, 425–433.
Borrero, C., Trillas, M.I., Ordovás, J., Tello, J.C., Avilés, M., 2004. Predictive factors for the suppression of Fusarium wilt of tomato in plant growth media. Phytopathology 94, 1094–1101.
Bretag, T.W., Ramsey, M., 2001. Foliar diseases caused by fungi. Ascochyta spp., in: Compendium of Pea Diseases and Pests. Second Edition. APS Press, The American Phytopathological Society, St. Paul Minnesota, pp. 24–28.
Campbell, C.L., Neher, D.A., 1994. Estimating disease severity and incidence, in: Epidemiology and Management of Root Diseases. Springer Berlin Heidelberg, pp. 117–147.
Chen, W., Hoitink, H.A.J., Schmitthenner, A.F., Touvinen, O.H., 1988. The role of microbial activity in suppression of damping-off caused by Pythium ultimum. Phytopathology 78, 314–322.
Feng, J., Hwang, R., Chang, K.F., Hwang, S.F., Strelkov, S.E., Gossen, B.D., Conner, R.L., Turnbull, G.D., 2010. Genetic variation in Fusarium avenaceum causing root rot on field pea: Genetic variation in Fusarium avenaceum. Plant Pathol. 59, 845–852.
Filippi, C., Pera, A., 1989. The role of telluric microflora in the control of Fusarium wilt in carnations grown in soils with bark compost. Biol. Wastes 27, 271–279.
Gaurilckiene, I., Cesnuleviciene, R., 2013. The susceptibility of pea (Pisum sativum L.) to Ascochyta blight under Lithuanian conditions. Zemdirb.-Agric. 100, 283–288.
Hadar, Y., 2011. Suppressive compost: when plant pathology met microbial ecology. Phytoparasitica 39, 311–314.
Hoitink, H.A., 2006. Compost use for disease suppression, in: Presentation at “Washington Organic Recycling Council 2006 Meeting”. Seattle, WA.
Hoitink, H.A.J., Boehm, M.J., 1999. Biocontrol within the context of soil microbial communities: a substrate-dependent phenomenon. Annu. Rev. Phytopathol. 37, 427–446.
Hoitink, H.A.J., Fahy, P.C., 1986. Basis for the control of soilborne plant pathogens with compost. Annu. Rev. Phytopathol. 24, 93–114.
Hoitink, H.A.J., Stone, A.G., Han, D.Y., 1997. Suppression of plant disease by composts. HortScience 32, 184–189.
Jensen, B., Bødker, L., Larsen, J., Knudsen, J.C., Jørnsgaard, B., 2004. Specificity of soil-borne pathogens on grain legumes.
Kraft, J.M., Haware, M.P., Hussein, M.M., 1988. Root rot and wilt diseases of food legumes, in: Summerfield, R.J. (Ed.), World Crops: Cool Season Food Legumes, Current Plant Science and Biotechnology in Agriculture. Springer Netherlands, pp. 565–575.
Kuter, G.A., Nelson, E.B., Hoitink, H.A.J., Madden, L.V., 1983. Fungal populations in container media amended with composted hardwood bark suppressive and conducive to Rhizoctonia damping-off. Phytopathology 73, 1450–1456.

DISEASE SUPPRESSION WITH COMPOST APPLICATION
83
Litterick, A.M., Harrier, L., Wallace, P., Watson, C.A., Wood, M., 2004. The role of uncomposted materials, composts, manures and compost extracts in reducing pest and disease incidence and severity in sustainable temperate agricultural and horticultural crop production – a review. Crit. Rev. Plant Sci. 23, 453–479.
Lumsden, R.D., Lewis, B.G., Millner, P.D., 1983. Effect of composted sewage sludge on several soilborne pathogens and diseases. Phytopathology 73, 1543–1548.
Nasir, M., Hoppe, H.-H., Ebrahim-Nesbat, F., 1992. The development of different pathotype groups of Mycosphaerella pinocles in susceptible and partially resistant pea leaves. Plant Pathol. 41, 187–194.
Noble, R., 2011. Risks and benefits of soil amendment with composts in relation to plant pathogens. Australas. Plant Pathol. 40, 157–167.
Noble, R., Coventry, E., 2005. Suppression of soil-borne plant diseases with composts: A review. Biocontrol Sci. Technol. 15, 3–20.
Noble, R., Roberts, S.J., 2003. A review of the literature on eradication of plant pathogens and nematodes during composting, disease suppression and detection of plant pathogens in compost. Published by: The Waste and Resources Action Programme, Banbury, Oxon, UK. 42pp. Oxon-UK Wastes Resour. Action Programme WRAP.
Olofsson, J., 1967. Root rot of canning and freezing peas in Sweden. Acta Agric. Scand. 17, 101–107.
Oyarzun, P.J., 1993. Bioassay to assess root rot in pea and effect of root rot on yield. Neth. J. Plant Pathol. 99, 61–75.
Persson, L., Bodker, L., Larsson-Wikström, M., 1997. Prevalence and pathogenicity of foot and root rot pathogens of pea in south Scandinavia. Plant Dis. 81, 171–174.
Pflughöft, O., Merker, C., von Tiedemann, A., Schäfer, B.C., 2012. Zur Verbreitung und Bedeutung von Pilzkrankheiten in Körnerfuttererbsen (Pisum sativum L.) in Deutschland. Gesunde Pflanz. 64, 39–48.
Recycled Organics Unit, 2006. Compost use for pest and disease suppression in NSW. The University of New South Wales, Australia.
Schüler, C., Pikny, J., Nasir, M., Vogtmann, H., 1993. Effects of composted organic kitchen and garden waste on Mycosphaerella pinodes (Berk. et blox) Vestergr., causal organism of foot rot on peas (Pisum sativum L.). Biol. Agric. Hortic. 9, 353–360.
Schulte-Geldermann, E., Bruns, C., Hess, J., Finckh, M.R., 2009. Einfluss von ligninhaltigen Komposten und Pflanzgutgesundheit auf den Befall mit Rhizoctonia solani bei Kartoffeln.
Serra-Wittling, C., Houot, S., Alabouvette, C., 1996. Increased soil suppressiveness to Fusarium wilt of flax after addition of municipal solid waste compost. Soil Biol. Biochem. 28, 1207–1214.
Solaiman, Z., Colmer, T.D., Loss, S.P., Thomson, B.D., Siddique, K.H.M., 2007. Growth responses of cool-season grain legumes to transient waterlogging. Aust. J. Agric. Res. 58, 406–412.
Termorshuizen, A.J., van Rijn, E., van der Gaag, D.J., Alabouvette, C., Chen, Y., Lagerlöf, J., Malandrakis, A.A., Paplomatas, E.J., Rämert, B., Ryckeboer, J., Steinberg, C., Zmora-Nahum, S., 2006. Suppressiveness of 18 composts against 7 pathosystems: Variability in pathogen response. Soil Biol. Biochem. 38, 2461–2477.
Yogev, A., Raviv, M., Hadar, Y., Cohen, R., Katan, J., 2006. Plant waste-based composts suppressive to diseases caused by pathogenic Fusarium oxysporum. Eur. J. Plant Pathol. 116, 267–278.

GENERAL DISCUSSION
84
5. GENERAL DISCUSSION
Seasonal climatic conditions strongly affected overall crop performance of peas, maize and
wheat in the field trials (Chapter 2). The dry summer of 2011 resulted in low maize yields
compared with the two other experimental years. The sudden drop of the temperature in the
first two weeks of February 2012 followed by a warm and dry March resulted in the failure of
the pea crop in 2011/12. The extreme weather events led to enhanced mineralization and
resulted in the relatively high amounts of about 55 kg ha-1 mineral nitrogen in the soil in
spring 2012 available for maize despite the failure of the peas. N inputs in organic systems
are low and catch crops are often used, N leaching is minimized in comparison to
conventional agricultural systems (Hansen et al., 2000). However, in the field trial between
2009 and 2013 N min residues in fall before wheat ranged between 40 and 55 kg ha-1. As
winter wheat does not require more than 17 – 30 kg N ha-1 before spring (Alley et al., 2009),
this means that there was a potential for N-leaching risks leading to N loses in the winter
wheat.
Field application of 5 t dry matter ha-1 of compost did not have any statistically significant
effects on yields of any of the three crops in the rotation (Chapter 2). An exception occurred
in main plot 24 (see Figure 2.7) which is located at the lower end of the experimental field.
Due to the very wet winter temporary water logging occurred in this plot. Field peas are
adversely affected by waterlogging and respond with a decrease of root growth and a decline
in nitrogen fixation (Solaiman et al., 2007). In this plot overall fresh matter production of peas
was low with about 250 g m-2 in the treatment inoculated with Phoma medicaginis and 900 g
m-2 in the non-inoculated control. Compost application resulted in strong increases in yield to
750 g m-2 in inoculated and 1100 g m-2 in the non-inoculated treatments (see Figure 2.7).
This suggests that application of high quality composts was able to minimize the negative
effects of the soil conditions on the pea performance and stabilize the yield.
A complex of different soil borne pathogens was isolated from peas and among them Phoma
medicaginis was dominating. Its incidence was not affected by inoculation, compost
application or seasonal climatic conditions (see Figure 2.10). P. medicaginis has been
reported as an important and commonly found pathogen causing foot rot of peas in Germany
(Pflughöft et al., 2012). In addition, Didymella pinodes, Fusarium oxysporum, Fusarium
solani and Fusarium avenaceum were commonly found on peas. The most aggressive pea
pathogen was F. avenaceum, followed by P. medicaginis, D. pinodes and F. solani. F.
avenaceum was also the pathogen that caused the highest reduction in fresh weights, 65%
compared to the non-inoculated control in pot experiments. These findings are in agreement

GENERAL DISCUSSION
85
with Persson et al. (1997) who reported reduction in pea biomass by 66% in greenhouse
trials. The violet flowering winter pea variety EFB33 used in the field trials usually had only
moderate disease symptoms both in the field and under controlled conditions. An exception
was the field in 2011/12 when the few plants that had survived the frost in February all were
severelly affected by root rot. In contrast, the white flowering spring variety Santana was
highly susceptible to all tested foot rot pathogens under controlled conditions (Chapter 3).
F. avenaceum is a wide host range, opportunistic pathogen that causes damage to plants
weakened by stress (Leslie et al., 1990). In healthy and active soils it is usually suppressed
by the native soil microbiota (Fletcher et al., 1991; Lin and Cook, 1979). F. avenaceum does
not produce chlamidospores and survives in the soil on crop residues (Leslie and Summerell,
2006). Agricultural practices such as reduced tillage, increased glyphosate application,
rotation of susceptible hosts in combination with changing climate and general reductions of
SOM have likely contributed to increased occurrence of F. avenaceum and damaging levels
in Northern Europe and Canada in recent years (Fernandez, 2007; Fernandez et al., 2008;
Foroud et al., 2014; Pflughöft et al., 2012). Moreover, due its genetic and ecological diversity
it can occupy several ecological niches allowing selection for different fitness traits for both
saprophytic and pathogenic phases of its life cycle (Foroud et al., 2014; Holtz et al., 2011).
Forbes and Dickinson (1977) reported that this pathogen is capable of continuous growth in
wet and cool soils which gives it an advantage over other soil microorganisms. It can also
thrive at relatively high partial pressures of carbon dioxide and reduced oxygen leveIs in the
wet soils. Incidence of F. avenaceum in the field in the season 2011/12 was higher than in
the other two years. This was most likely due to the late frost. After the prolonged frost
surface soil layers melted before subsoil resulting in water logging in the upper soil creating
unfavorable conditions for root growth relative to F. aveanceum.
D. pinodes also strongly affected growth of peas. Although it caused only moderate disease
symptoms with mean DI of 4.8, reduction of fresh weight was on average 34% compared to
mean DI of 5.7 due to P. medicaginis causing fresh weight reduction of 20%. Clearly, plant
growth parameters and disease severity need to be considered together in order to correctly
judge the damage potential of pathogens. Fusarium solani has been recognized as one of
the most prevalent and important pathogen of the foot rot complex on peas (Kraft et al.,
1988b; Rush and Kraft, 1986). This pathogen also had a high damage potential in the field
used in these studies as demonstrated by high DI in non-inoculated controls in Chapter 3
and 4. The reisolation from the control plants grown in field soil were dominated by F. solani.
However, under the testing conditions in Chapter 3 and 4, F. solani was the least damaging
pathogen causing low DI and no adverse effects on plant growth. F. solani f. sp pisi is a slow
colonizer of the roots (Huisman, 1982). After penetrating the epidermis, F. solani grows

GENERAL DISCUSSION
86
through the cortex until the Casparian strip and then degradation of the vascular system
starts (Foroud et al., 2014). Any factor that stresses the host root system will shorten the time
between infection and symptom development (Thung and Rao, 1999). The short time of
growing (21 days) and lack of external stress are likely reasons for the lack of symptoms on
peas and adverse effects due to F. solani in the greenhouse tests.
The most important Fusarium spp. isolated from maize and wheat were F. graminearum and
F. culmorum, however they were isolated at much lower frequencies from wheat seedlings. It
is common that many different Fusarium species occur simultaneously in plants and overall
diversity is affected by cultural practices, plant cultivars and weather patterns (Gatch and
Munkvold, 2002; Munkvold, 2003; Osunlaja, 1990; Scauflaire et al., 2011). Besides the
above mentioned F. avenaceum, F. gramineraum and F. culmorum, F. oxysporum, F.
equiseti, F. sambucinum and F. crookwellense were commonly isolated from maize and
wheat. Although these two crops shared a range of Fusarium species, there was no
correlation between isolation frequencies of different Fusarium spp. among them. Therefore,
no conclusion about the risk of increased Fusarium infestation by growing maize before
wheat can be made. However, the high diversity of species isolated from both crops
combined with the fact that Fusarium spp. are soil - and residue - borne pathogens able to
survive for a long time, clearly indicate the potential of the maize – wheat rotation to
contribute to an increase in the diversity and size of the soil borne Fusarium community. On
the long run, considering the changing climate with more frequent extreme weather events
predicted, this can lead to a comparative advantage of the Fusarium community over the
plants in agricultural systems leading to increased disease risk and mycotoxin contamination.
When Santana and EFB33 were grown in non-sterilized field soil, overall disease caused by
the pathogens tested was lower compared to sterilized sand. However, low to moderate
disease symptoms occurred on the non-inoculated soil controls suggesting that natural
inoculum of F. solani was present in the soil used as confirmed by reisolations. Soils used in
the greenhouse trials were collected from the control plots of the field experiments (no
compost application and no inoculation with P. medicaginis) during the early spring while
winter peas were grown. Peas had not been grown in these plots in the last five years prior to
the experiments; however, clover in the mixture with grass was grown for two years in a row,
a year before the field experiments. This could have contributed to the buildup of natural
inoculum in the soil as most of the root rot pathogens of pea are also pathogenic to clovers
(Farr and Rossman, 2015). When EFB33 was grown in the non-sterilized field soil in the
greenhouse trial, disease severity was considerably lower than on Santana just like in all
inoculation trials. This confirms the broad resistance of EFB33 towards wide range of
pathogens from the root and foot rot complex. Violet flowering pea varieties have been

GENERAL DISCUSSION
87
reported to be more resistant to soil-borne diseases, compared to the white flowering
varieties (Vogt-Kaute et al., 2013). However, Vogt-Kaute et al. (2013) could not find a
statistically significant correlation between flower color and susceptibility to foot and root rot
of twenty four pea lines and varieties. From the results it can be concluded that EFB33 could
be a good alternative to the commonly grown spring variety Santana with respect to
resistance foot and root rot. EFB33 should be included in pea breeding programs as it can
bring valuable insights in the pathogen-host interaction and resistance mechanisms.
Inconsistency in the level of compost induced disease suppression between pot and field
experiments has often been reported (Hoitink et al., 1997; Litterick et al., 2004; Noble and
Roberts, 2003) and was also observed in this work (Chapter 2 and 4). Application of yard
waste compost (YWC) did not reduce the severity of foot rot of winter peas or wheat in the
field (Chapter 2); however, different levels of suppression of foot rot of peas were observed
in the greenhouse experiments (Chapter 4). The YWC used suppressed foot disease
caused by F. avenaceum, P. medicaginis, and D. pinodes on the susceptible variety Santana
by reducing the intensity of disease symptoms and promoting plant growth. Although high
amount of compost (30-50% v/v) are recommended for successful disease suppression,
application of 3.5% compost in sterile sand was high enough to achieve suppression of
disease symptoms of all four pathogens. When non-sterilized field soil was used as growing
substrate compost application did not result in significant reduction. The complexity of
disease suppressiveness can be reflected by the various and complex interactions between
pathogens and antagonistic microflora, and between soil abiotic factors and microflora
(Pérez-Piqueres et al., 2006). Complexity of the system is the main reason why it is difficult
to clearly show the beneficial effect of short term compost application on the health and
disease reduction in field grown crops although bioassays under the controlled conditions
have been successful in reducing the disease of the numerous crops caused by the range of
soil-borne pathogens (Bailey and Lazarovits, 2003; Hoitink, 2006; Litterick et al., 2004; Noble
and Coventry, 2005; Termorshuizen et al., 2006). Although we did not observe reduction of
the severity of foot rot of winter peas or wheat in the field trial, a long term field application of
high quality composts might have multiple beneficial effects on overall soil quality that could
help in buildup of soil suppressiveness towards soil-borne diseases. Addition of composts
increases the level of soil organic matter (SOM) which is considered helpful for developing
soil suppressiveness for soil borne diseases as it supports greater microbial biomass and
activity (Stone et al., 2003). In addition, improvement in physical soil characteristics like
water holding capacity, aggregate stability and soil porosity allows plants to develop deep
and well developed root systems that can minimize detrimental effects of the infections with
soil-borne pathogens.

GENERAL DISCUSSION
88
Different mechanisms of suppression were possibly involved in reduction of symptoms
caused by Fusarium spp. and by D. pinodes and P. medicaginis on peas. After γ sterilization
of compost severity of symptoms caused by F. avenaceum and F. solani increased, whereas
in the systems with D. pinodes and P. medicaginis even sterilized compost reduced disease
to some extent. It is likely that suppression of disease caused by Fusarium spp. is biological
in origin and for D. pinodes and P. medicaginis in addition chemical and physical properties
of compost are contributing to the suppression of disease. The disease increase after
application of sterilized compost to field soil was likely due to a low level of native beneficial
microflora in the field soil that was not able to compete with the natural inoculum and the
introduced pathogens for the available food source provided by the sterilized compost.
Pathogens were most likely more efficient in colonizing the sterile compost which resulted in
faster population growth and consequently increase in disease severity.
Due to the complex interactions between site specific factors affecting agricultural production
it is difficult to predict the direct effects of changing climate on the agricultural systems. For
that reason the goal should be to aim at building sustainable agricultural systems that are
resilient to environmental stress. This should be achieved by maintaining active and healthy
soils, increasing above and belowground biodiversity through the long term application of
organic amendments such as compost, green manures or manures in the production system
and introduction of cover crops, living or dead mulches (Abawi and Widmer, 2000).

GENERAL DISCUSSION
89
5.1 References
Abawi, G.S., Widmer, T.L., 2000. Impact of soil health management practices on soilborne pathogens, nematodes and root diseases of vegetable crops. Appl. Soil Ecol. 15, 37–47.
Alley, M.M., Scharf, P., Brann, D.E., Hammons, J.L., 2009. Nitrogen management for winter wheat: principles and recommendations. Va. Coop. Ext. 424, 1–6.
Bailey, K.L., Lazarovits, G., 2003. Suppressing soil-borne diseases with residue management and organic amendments. Soil Tillage Res. 72, 169–180.
Fernandez, M.R., 2007. Fusarium populations in roots of oilseed and pulse crops grown in eastern Saskatchewan. Can. J. Plant Sci. 87, 945–952.
Fernandez, M.R., Huber, D., Basnyat, P., Zentner, R.P., 2008. Impact of agronomic practices on populations of Fusarium and other fungi in cereal and noncereal crop residues on the Canadian Prairies. Soil Tillage Res. 100, 60–71.
Fletcher, J.D., Broadhurst, P.G., Bansal, R.K., 1991. Fusarium avenaceum : A pathogen of lentil in New Zealand. N. Z. J. Crop Hortic. Sci. 19, 207–210.
Forbes, R.S., Dickinson, C.H., 1977. Behaviour of Fusarium avenaceum in soil growth analysis plates. Trans. Br. Mycol. Soc. 69, 197–205.
Foroud, N.A., Chatterton, S., Reid, L.M., Turkington, T.K., Tittlemier, S.A., Gräfenhan, T., 2014. Fusarium diseases of Canadian grain crops: Impact and disease management strategies, in: Goyal, A., Manoharachary, C. (Eds.), Future Challenges in Crop Protection Against Fungal Pathogens, Fungal Biology. Springer New York, pp. 267–316.
Gatch, E.W., Munkvold, G.P., 2002. Fungal species composition in maize stalks in relation to European corn borer injury and transgenic insect protection. Plant Dis. 86, 1156–1162.
Hansen, B., Kristensen, E.S., Grant, R., Høgh-Jensen, H., Simmelsgaard, S.E., Olesen, J.E., 2000. Nitrogen leaching from conventional versus organic farming systems — a systems modelling approach. Eur. J. Agron. 13, 65–82.
Hoitink, H.A., 2006. Compost use for disease suppression, in: Presentation at “Washington Organic Recycling Council 2006 Meeting”. Seattle, WA.
Hoitink, H.A.J., Stone, A.G., Han, D.Y., 1997. Suppression of plant disease by composts. HortScience 32, 184–189.
Holtz, M.D., Chang, K.F., Hwang, S.F., Gossen, B.D., Strelkov, S.E., 2011. Characterization of Fusarium avenaceum from lupin in central Alberta: genetic diversity, mating type and aggressiveness. Can. J. Plant Pathol. 33, 61–76.
Huisman, O.C., 1982. Interrelations of root growth dynamics to epidemiology of root-invading fungi. Annu. Rev. Phytopathol. 20, 303–327.
Kraft, J.M., Haware, M.P., Hussein, M.M., 1988. Root rot and wilt diseases of food legumes, in: Summerfield, R.J. (Ed.), World Crops: Cool Season Food Legumes, Current Plant Science and Biotechnology in Agriculture. Springer Netherlands, pp. 565–575.
Leslie, J.F., Pearson, C.A.S., Nelson, P.E., Toussoun, T.A., 1990. Fusarium spp. from Corn, Sorghum, and Soybean Fields in the Central and Eastern United States. Phytopathology 80, 343–350.
Leslie, J.F., Summerell, B.A., 2006. The Fusarium laboratory manual. Blackwell Pub., Ames, Iowa.
Lin, Y.S., Cook, R.J., 1979. Supression of Fusarium roseum “avenaceum” by soil microorganisms. Phytopathology 69, 384–388.
Litterick, A.M., Harrier, L., Wallace, P., Watson, C.A., Wood, M., 2004. The role of uncomposted materials, composts, manures and compost extracts in reducing pest and disease incidence and severity in sustainable temperate agricultural and horticultural crop production – a review. Crit. Rev. Plant Sci. 23, 453–479.
Munkvold, G.., 2003. Epidemiology of Fusarium diseases and their mycotoxins in maize ears. Eur. J. Plant Pathol. 109, 705–713.
Noble, R., Coventry, E., 2005. Suppression of soil-borne plant diseases with composts: A review. Biocontrol Sci. Technol. 15, 3–20.

GENERAL DISCUSSION
90
Noble, R., Roberts, S.J., 2003. A review of the literature on eradication of plant pathogens and nematodes during composting, disease suppression and detection of plant pathogens in compost. Published by: The Waste and Resources Action Programme, Banbury, Oxon, UK. 42pp. Oxon-UK Wastes Resour. Action Programme WRAP.
Osunlaja, S.O., 1990. Effect of organic soil amendments on the incidence of stalk rot of maize. Plant Soil 127, 237–241.
Pérez-Piqueres, A., Edel-Hermann, V., Alabouvette, C., Steinberg, C., 2006. Response of soil microbial communities to compost amendments. Soil Biol. Biochem. 38, 460–470.
Persson, L., Bodker, L., Larsson-Wikström, M., 1997. Prevalence and pathogenicity of foot and root rot pathogens of pea in south Scandinavia. Plant Dis. 81, 171–174.
Pflughöft, O., Merker, C., von Tiedemann, A., Schäfer, B.C., 2012. Zur Verbreitung und Bedeutung von Pilzkrankheiten in Körnerfuttererbsen (Pisum sativumL.) in Deutschland. Gesunde Pflanz. 64, 39–48.
Rush, C.M., Kraft, J.M., 1986. Effects of inoculum density and placement on Fusarium root rot of peas. Phytopathology 76, 1325–1329.
Scauflaire, J., Mahieu, O., Louvieaux, J., Foucart, G., Renard, F., Munaut, F., 2011. Biodiversity of Fusarium species in ears and stalks of maize plants in Belgium. Eur. J. Plant Pathol. 131, 59–66.
Solaiman, Z., Colmer, T.D., Loss, S.P., Thomson, B.D., Siddique, K.H.M., 2007. Growth responses of cool-season grain legumes to transient waterlogging. Aust. J. Agric. Res. 58, 406–412.
Stone, A.G., Vallad, G.E., Cooperband, L.R., Rotenberg, D., Darby, H.M., James, R.V., Stevenson, W.R., Goodman, R.M., 2003. Effect of organic amendments on soilborne and foliar diseases in field-grown snap bean and cucumber. Plant Dis. 87, 1037–1042.
Termorshuizen, A.J., van Rijn, E., van der Gaag, D.J., Alabouvette, C., Chen, Y., Lagerlöf, J., Malandrakis, A.A., Paplomatas, E.J., Rämert, B., Ryckeboer, J., Steinberg, C., Zmora-Nahum, S., 2006. Suppressiveness of 18 composts against 7 pathosystems: Variability in pathogen response. Soil Biol. Biochem. 38, 2461–2477.
Thung, M., Rao, I.M., 1999. Integrated management of abiotic stresses, in: Singh, S.P. (Ed.), Common Bean Improvement in the Twenty-First Century, Developments in Plant Breeding. Springer Netherlands, pp. 331–370.
Vogt-Kaute, W., Jorek, B., Tilcher, R., 2013. Gibt es bei Körnererbsen Sortenunterschiede in der Anfälligkeit gegenüber bodenbürtigen Krankheiten?

APPENDIX
91
APPENDIX

APPENDIX
92
Chapter 2
INCIDENCE OF FUSARIUM SPP. IN THE CROP ROTATION WINTER PEA – MAIZE – WINTER WHEAT AND THE POTENTIAL OF YARD WASTE COMPOST TO SUPPRESS FOOT AND ROOT ROT OF PEAS UNDER FIELD CONDITIONS
Figure A. 2.1. Effect of treatments on pea emergence one month after sowing presented as percentage of sowing densities. The error bars represent the ± 1 SD.
Figure A. 2.2. Emergence (Autumn) and winter survival (Spring) of peas as a percentage of sown plants in 2011/12. The error bars represent the ± 1 SD.
APPENDIX
93
Table A. 2.1. Mineral nitrogen (Nmin) content in the soil at 0-60 cm depth. O=Control, I=Inoculated, C=Compost, I+C=Inoculated+Compost.
Peas Before Maize After Maize After Wheat
Rotation I 19.03.2010 26.05.2010 - 02.08.2011
O 35.07 16.90 6.21
I 37.25 25.01 7.53
C 37.06 16.37 7.37
I+C 39.38 15.24 8.92
Rotation II 05.04.2011 16.05.2011 15.10.2011 17.08.2012
O 41.50 32.82 59.79 28.75
I 52.11 34.32 57.89 35.02
C 44.95 28.23 55.40 29.64
I+C 44.96 32.49 55.65 33.22
Rotation III - 26.04.2012 04.10.2012 22.08.2013
O 56.84 43.67 33.13
I 58.57 32.57 31.64
C 49.96 37.06 34.56
I+C 57.38 41.18 31.88
Table A. 2.2. Fresh and dry weights of pea and weeds in 2010 and 2011. O = Control, I = Inoculated, C = Compost, I+C = Inoculated + Compost
Year Fresh weight
pea
Fresh weight weeds Dry weight pea Dry weight
weeds
g m-1 SD g m-1 SD % of total FW g m-1
SD g m-1 SD
2010 O 1410 459 191 75 11 169 45 27 8
I 979 618 203 61 17 124 69 27 7
C 1395 454 170 70 12 173 49 25 10
I+C 1296 437 183 81 12 160 48 26 14
2011 O 914 222 175 86 17 127 29 27 14
I 962 284 169 63 15 115 31 36 17
C 982 276 202 57 15 130 34 32 11
I+C 1025 238 175 90 16 123 25 40 20
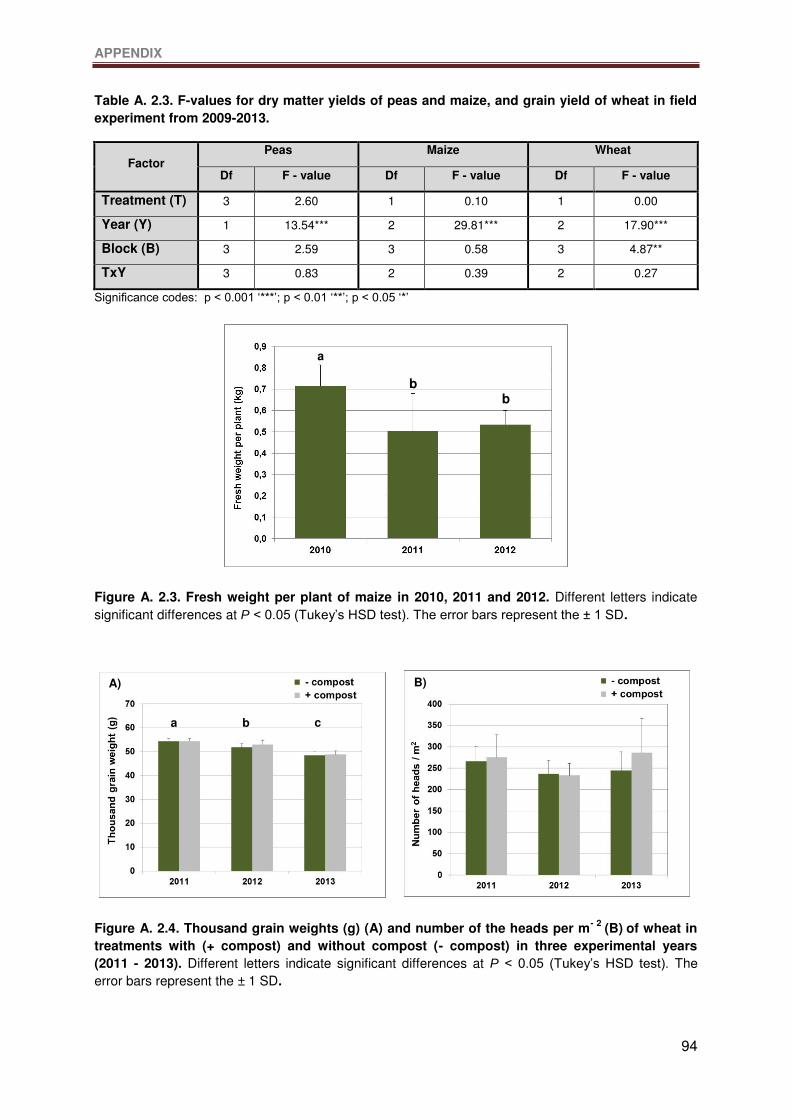
APPENDIX
94
Table A. 2.3. F-values for dry matter yields of peas and maize, and grain yield of wheat in field experiment from 2009-2013.
Factor Peas Maize Wheat
Df F - value Df F - value Df F - value
Treatment (T) 3 2.60 1 0.10 1 0.00
Year (Y) 1 13.54*** 2 29.81*** 2 17.90***
Block (B) 3 2.59 3 0.58 3 4.87**
TxY 3 0.83 2 0.39 2 0.27
Significance codes: p < 0.001 ‘***’; p < 0.01 ‘**’; p < 0.05 ‘*’
Figure A. 2.3. Fresh weight per plant of maize in 2010, 2011 and 2012. Different letters indicate significant differences at P < 0.05 (Tukey’s HSD test). The error bars represent the ± 1 SD.
Figure A. 2.4. Thousand grain weights (g) (A) and number of the heads per m- 2 (B) of wheat in treatments with (+ compost) and without compost (- compost) in three experimental years (2011 - 2013). Different letters indicate significant differences at P < 0.05 (Tukey’s HSD test). The error bars represent the ± 1 SD.
a b c
a
b b
A) B)
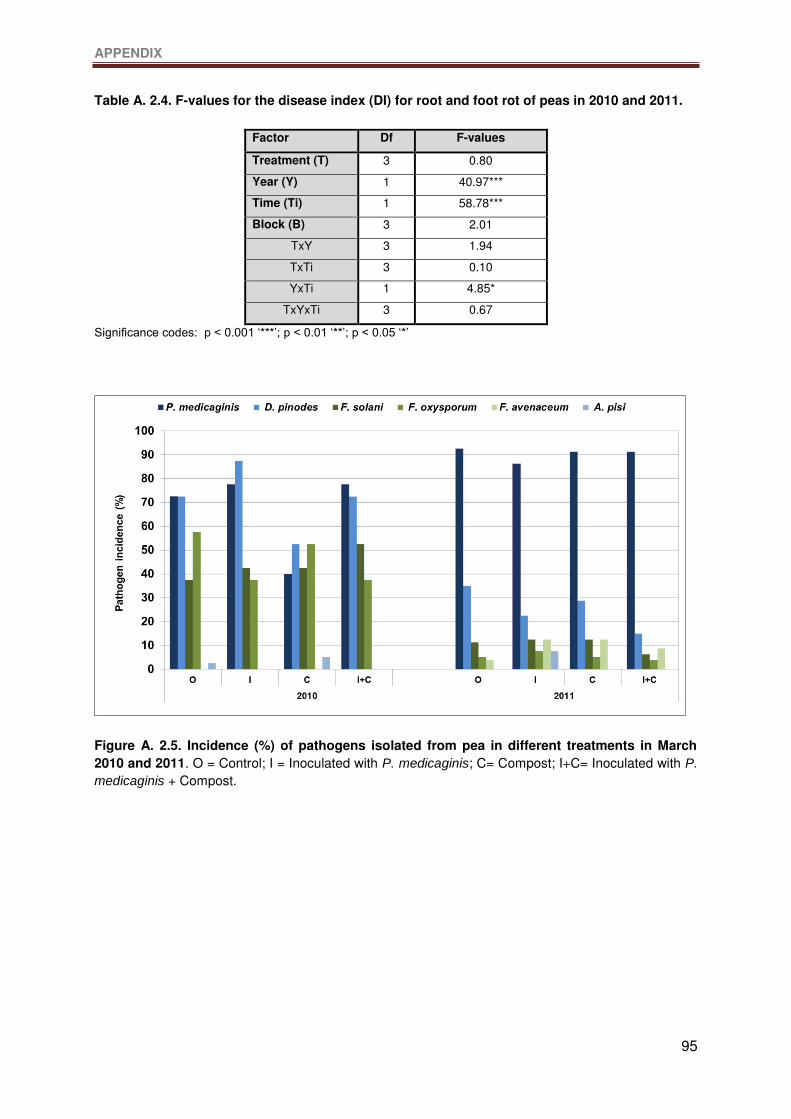
APPENDIX
95
Table A. 2.4. F-values for the disease index (DI) for root and foot rot of peas in 2010 and 2011.
Factor Df F-values
Treatment (T) 3 0.80
Year (Y) 1 40.97***
Time (Ti) 1 58.78***
Block (B) 3 2.01
TxY 3 1.94
TxTi 3 0.10
YxTi 1 4.85*
TxYxTi 3 0.67
Significance codes: p < 0.001 ‘***’; p < 0.01 ‘**’; p < 0.05 ‘*’
Figure A. 2.5. Incidence (%) of pathogens isolated from pea in different treatments in March 2010 and 2011. O = Control; I = Inoculated with P. medicaginis; C= Compost; I+C= Inoculated with P.
medicaginis + Compost.

APPENDIX
96
Figure A. 2.6. Incidence (%) of pathogens isolated from pea in different treatments in May of three experimental years. O = Control; I = Inoculated with P. medicaginis; C= Compost; I+C= Inoculated with P. medicaginis + Compost.
Figure A. 2.7. Pathogen incidence on pea in March and May of 2010 and 2011. Pm = P.
medicaginis, Dp = D. pinodes, Fs = F. solani and Fo = F. oxysporum.

APPENDIX
97
Figure A. 2.8. External (A) and Internal (B) disease scores of pea in May of all three experimental years. Score 1 stands for healthy plant; score 9 for dead plant. O = Control; I = Inoculated with P. medicaginis; C= Compost; I+C= Inoculated with P. medicaginis + Compost.
A)
B)

APPENDIX
98
Figure A. 2.9. External (A) and Internal (B) disease scores of pea in March and May 2010. Score 1 stands for healthy plant; score 9 for dead. O = Control; I = Inoculated with P. medicaginis; C= Compost; I+C= Inoculated with P. medicaginis + Compost.
A)
B)
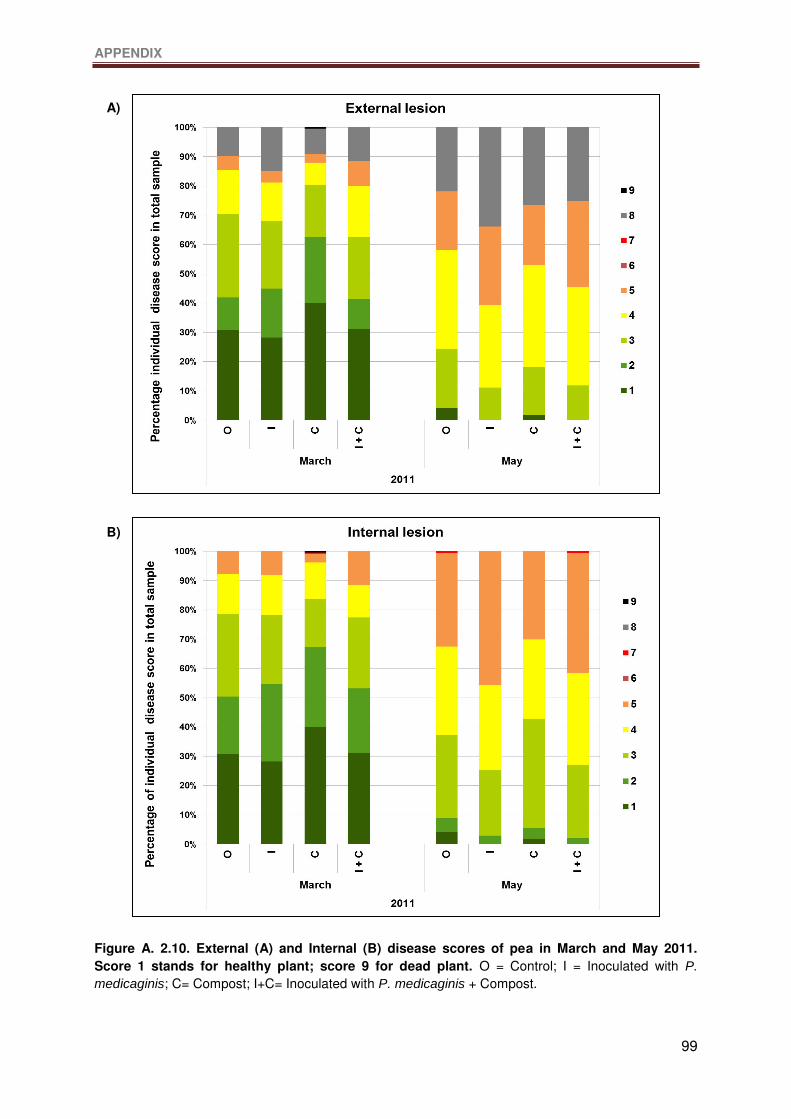
APPENDIX
99
Figure A. 2.10. External (A) and Internal (B) disease scores of pea in March and May 2011. Score 1 stands for healthy plant; score 9 for dead plant. O = Control; I = Inoculated with P.
medicaginis; C= Compost; I+C= Inoculated with P. medicaginis + Compost.
B)
A)
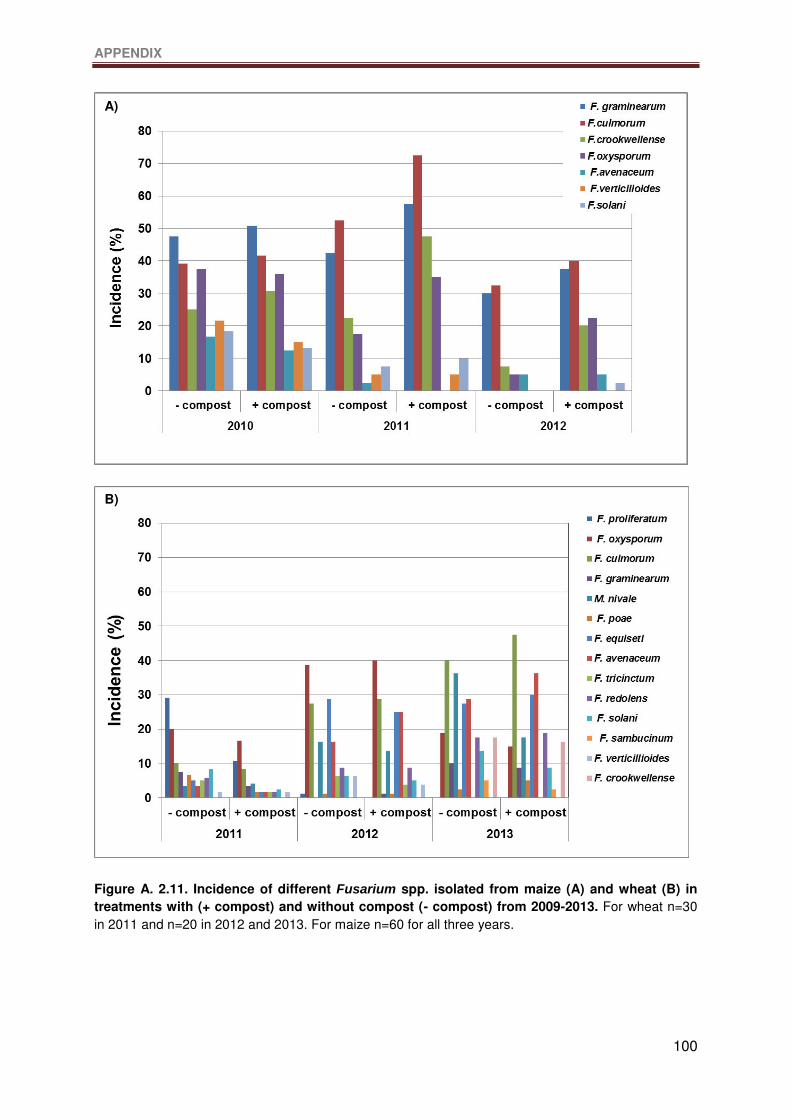
APPENDIX
100
Figure A. 2.11. Incidence of different Fusarium spp. isolated from maize (A) and wheat (B) in treatments with (+ compost) and without compost (- compost) from 2009-2013. For wheat n=30 in 2011 and n=20 in 2012 and 2013. For maize n=60 for all three years.
B)
A)

APPENDIX
101
Chapter 3
AGGRESSIVENESS OF FOUR PATHOGENS CAUSING FOOT AND ROOT ROT
OF PEA (PISUM SATIVUM L.) ON A SPRING AND A WINTER PEA CULTIVAR
Table A. 3.1. F-values for DI for isolate comparison of four pathogens in three-way ANOVA.
Factor Df F. avenaceum F. solani P. medicaginis D. pinodes
Variety (V) 1 193.3*** 127.7*** 169.5*** 140.4***
Substrate (S) 1 100.7*** 2.5 235.7*** 5.8**
Isolate (I) 4 4.1*** 0.4 3.2* 5.6***
VxS 1 30.0*** 0.8 15.6*** 0.4
VxI 4 1.5 0.8 0.6 3.3**
SxI 4 1.8 0.4 1.4 1.1
VxSxI 4 0.8 0.4 0.3 0.8
Table A. 3.2. F-values for DI of Santana and EFB33 inoculated with five isolates of four pathogens in sand and soil.
Factor Df F. avenaceum F. solani P. medicaginis D. pinodes
Sand
Variety (V) 1 193.0*** 57.95*** 124.5*** 87.96***
Isolate (I) 4 4.68** 0.7 3.35* 2.48
VxI 4 1.50 0.9 0.5 1.33
Soil
Variety (V) 1 34.57*** 69.73*** 48.77*** 56.79***
Isolate (I) 4 1.36 0.1 0.9 4.02**
VxI 4 1.07 0.4 0.4 2.70*
Table A. 3.3. F-values for fresh weights of Santana and EFB33 grown in sand and soil and inoculated with five isolates of four pathogens.
Significance codes: p < 0.001 ‘***’; p < 0.01 ‘**’; p < 0.05 ‘*’.
Factor Df F. avenaceum F. solani P. medicaginis D. pinodes
Sand
Variety (V) 1 19.51*** 86.98*** 41.65*** 16.06***
Isolate (I) 4 21.62*** 1.39 4.04*** 1.25
V x I 4 25.17*** 1.00 1.27 1.09
Soil
Variety (V) 1 31.31*** 53.38*** 15.42*** 24.49***
Isolate (I) 4 0.38 0.97 1.39 0.80
V x I 4 1.22 0.73 0.54 0.90

APPENDIX
102
Table A. 3.4. F-values for dry weights of Santana and EFB33 grown in sand and soil and inoculated with five isolates of four pathogens.
Significance codes: p < 0.001 ‘***’; p < 0.01 ‘**’; p < 0.05 ‘*’.
Factor Df F. avenaceum F. solani P. medicaginis D. pinodes
Sand
Variety (V) 1 25.68*** 10.06*** 1.25 0.08
Isolate (I) 4 6.83*** 1.61 9.25*** 5.29***
V x I 4 10.45*** 1.83 2.05* 1.91
Soil
Variety (V) 1 2.32 9.29*** 1.86 0.004
Isolate (I) 4 1.67 1.37 2.55** 3.45***
V x I 4 0.71 2.64** 0.77 0.67

APPENDIX
103
Figure A. 3.1. External lesion scores for all of the isolates of all tested pathogens on Santana.
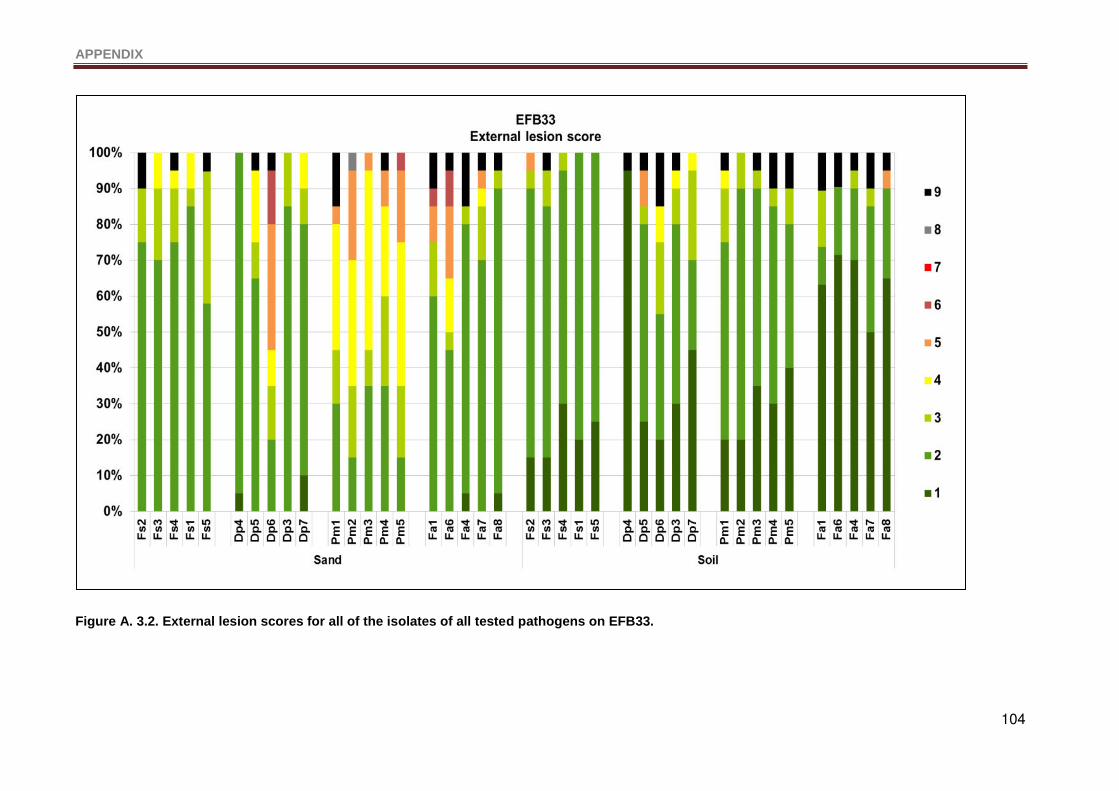
APPENDIX
104
Figure A. 3.2. External lesion scores for all of the isolates of all tested pathogens on EFB33.

APPENDIX
105
Figure A. 3.3. Fresh weight of Santana and EFB33 plants inoculated with five isolates of F.
solani and grown in sterile sand or non-sterilized field soil. The error bars represent the ± 1 SD.
Figure A. 3.4. Fresh weight of Santana and EFB33 plants inoculated with five isolates of D.
pinodes and grown in sterile sand or non-sterilized field soil. The error bars represent the ± 1 SD.
Santana EFB33
Santana EFB33

APPENDIX
106
Table A. 3.5. F-values for the DI of pea inoculated with four different pathogens in three-way
ANOVA.
Factor Df F-values
Pathogen (P) 3 9.94***
Variety (V) 1 435.77***
Substrate (S) 1 125.25***
PxV 3 6.38***
PxS 3 10.68***
VxS 1 0.03
PxVxS 3 1.32
Significance codes: p < 0.001 ‘***’; p < 0.01 ‘**’; p < 0.05 ‘*’
Table A. 3.6. F-values for DI for pathogen comparison for Santana and EFB33 in sterile sand
and non-sterilized field soil.
Factor Df Santana EFB33
Substrate (S) 1 111.00*** 34.18 ***
Pathogen (P) 3 24.06*** 5.16 **
SxP 3 22.95*** 2.32
Significance codes: p < 0.001 ‘***’; p < 0.01 ‘**’; p < 0.05 ‘*’
Table A. 3.7. ANOVA table and F values for fresh and dry weights per plant of Santana and
EFB33 inoculated with four different pathogens and grown in sterile sand and non-sterilized
field soil.
Df Fresh weight Dry weight
Santana
Pathogen (P) 4 35.09*** 24.39***
Substrate (S) 1 185.47*** 122.68***
PxS 4 12.66*** 8.41***
EFB33
Pathogen (P) 4 13.67*** 11.51***
Substrate (S) 1 166.09*** 129.74***
PxS 4 11.40*** 7.83***
Significance codes: p < 0.001 ‘***’; p < 0.01 ‘**’; p < 0.05 ‘

APPENDIX
107
Table A. 3.8. F-values for DI for Santana and EFB33 inoculated with P. medicaginis and F.
avenaceum and grown under three different temperature regimes.
Df DI
Santana
Pathogen (P) 2 0.012
Temperature (T) 2 6.32**
PxT 2 1.17
EFB33
Pathogen (P) 2 11.69**
Temperature (T) 2 0.27
PxT 2 0.78
Significance codes: p < 0.001 ‘***’; p < 0.01 ‘**’; p < 0.05 ‘*’.
Table A. 3.9. F-values for fresh and dry weights for Santana and EFB33 inoculated with P.
medicaginis and F. avenaceum and grown under three different temperature regimes.
Df Fresh weight Dry weight
Santana
Pathogen (P) 2 29.89*** 36.56***
Temperature (T) 2 72.35*** 26.05***
PxT 2 1.30 1.69
EFB33
Pathogen (P) 2 0.79ns 0.24
Temperature (T) 2 29.06*** 6.74***
PxT 2 1.93 1.54
Significance codes: p < 0.001 ‘***’; p < 0.01 ‘**’; p < 0.05 ‘*’.
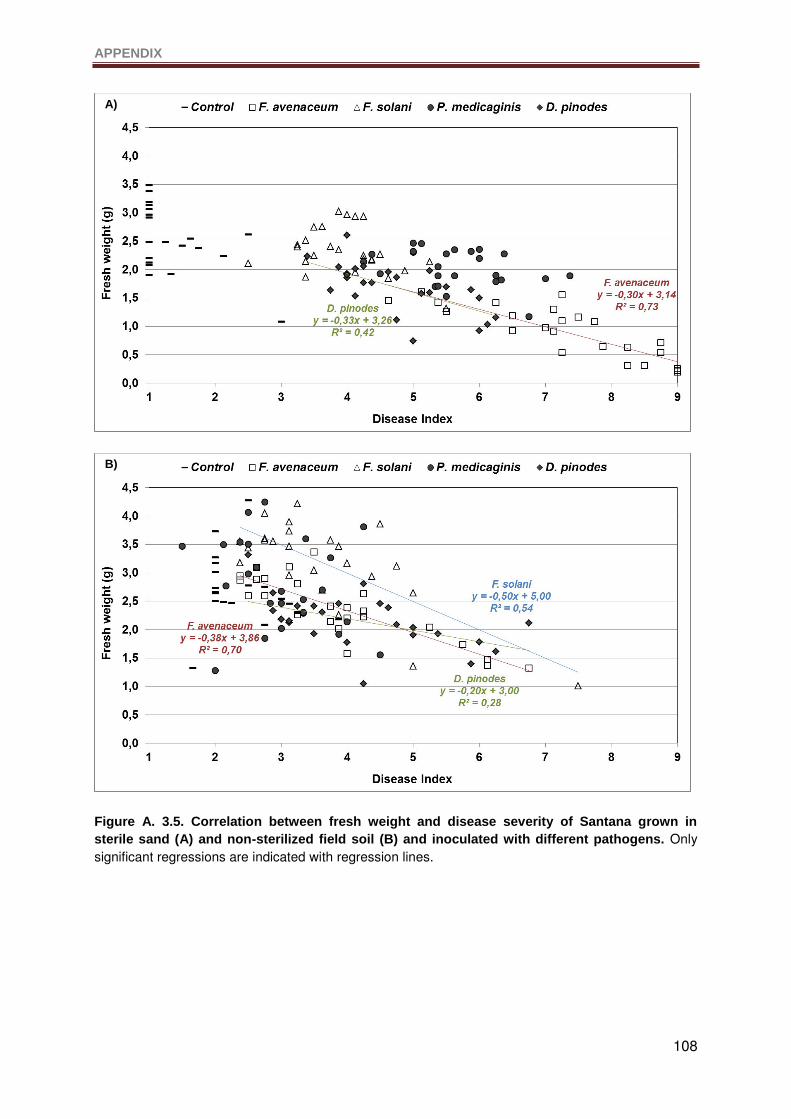
APPENDIX
108
Figure A. 3.5. Correlation between fresh weight and disease severity of Santana grown in
sterile sand (A) and non-sterilized field soil (B) and inoculated with different pathogens. Only significant regressions are indicated with regression lines.
A)
B)

APPENDIX
109
Figure A. 3.6. Correlation between fresh weight and disease severity of EFB33 grown in sterile
sand (A) and non-sterilized field soil (B) and inoculated with different pathogens. Only significant regressions are indicated with regression lines.
B)
A)

APPENDIX
110
Chapter 4
EFFECTS OF COMPOST APPLICATION ON FOOT AND ROOT ROT OF SPRING AND WINTER PEA VARIETIES CAUSED BY FUSARIUM AVENACEUM, FUSARIUM SOLANI, PHOMA MEDICAGINIS AND DIDYMELLA PINODES
Table A. 4.1. F-values for fresh and dry weights and DI for Santana and EFB33 inoculated with F. avenaceum, F. solani, P. medicaginis and D. pinodes and amended with 20% v/v of non-sterilized and γ sterilized YWC and grown in sterile sand and non-sterilized field soil.
Factor Df Fresh weight Dry weight DI
Pathogen (P) 4 7.89*** 5.98*** 19.21***
Compost (C) 2 148.30*** 71.55*** 12.71***
Substrate (S) 1 17.69*** 0.43 28.24***
Variety (V) 1 90.77*** 6.97*** 175.64***
PxC 4 3.88*** 3.67*** 1.27
PxS 4 12.09*** 4.50*** 21.19***
CxS 2 16.55*** 4.32** 0.00
PxV 4 15.47*** 7.30*** 2.56*
CxV 2 11.61*** 7.78*** 18.16***
SxV 1 1.04 4.34** 0.54
PxCxS 4 3.23*** 2.25** 2.03*
PxCxV 4 3.74*** 3.72*** 1.21
PxSxV 4 11.84*** 8.28*** 2.56*
CxSxV 2 5.55*** 1.55 0.05
PxCxSxV 4 3.19*** 1.16 1.37
Significance codes: p < 0.001 ‘***’; p < 0.01 ‘**’; p < 0.05 ‘*.’
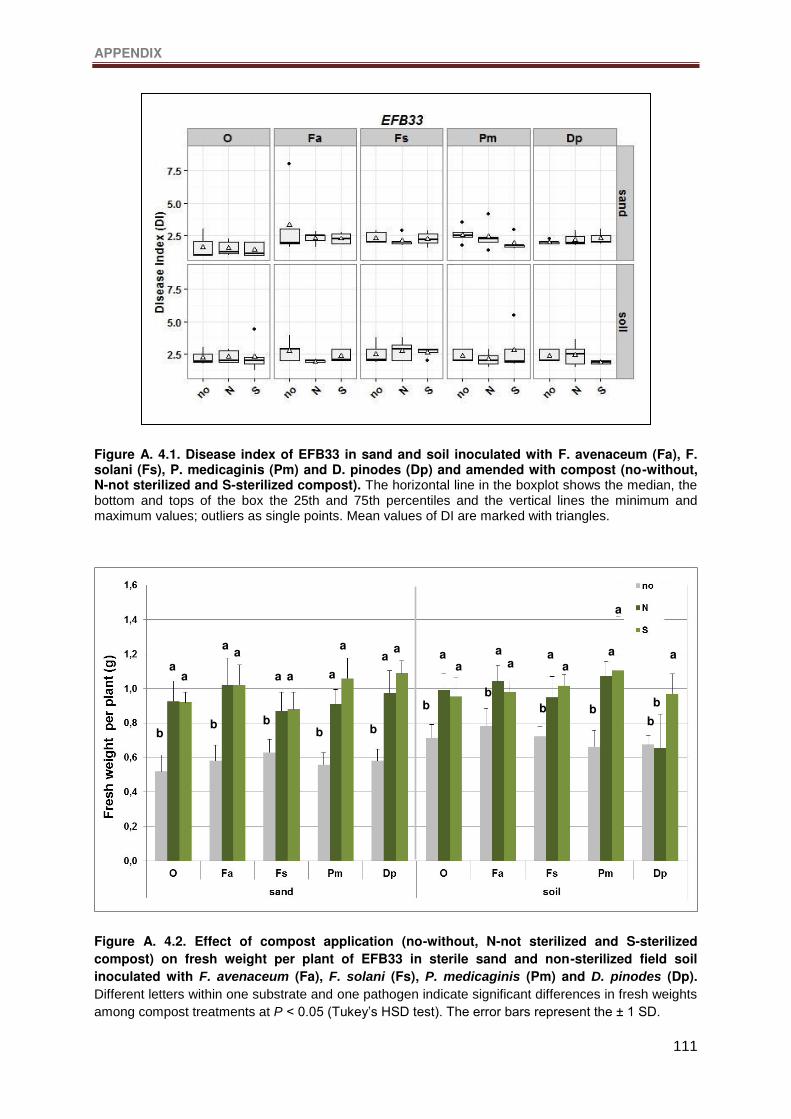
APPENDIX
111
Figure A. 4.1. Disease index of EFB33 in sand and soil inoculated with F. avenaceum (Fa), F. solani (Fs), P. medicaginis (Pm) and D. pinodes (Dp) and amended with compost (no-without, N-not sterilized and S-sterilized compost). The horizontal line in the boxplot shows the median, the bottom and tops of the box the 25th and 75th percentiles and the vertical lines the minimum and maximum values; outliers as single points. Mean values of DI are marked with triangles.
Figure A. 4.2. Effect of compost application (no-without, N-not sterilized and S-sterilized compost) on fresh weight per plant of EFB33 in sterile sand and non-sterilized field soil inoculated with F. avenaceum (Fa), F. solani (Fs), P. medicaginis (Pm) and D. pinodes (Dp). Different letters within one substrate and one pathogen indicate significant differences in fresh weights
among compost treatments at P < 0.05 (Tukey’s HSD test). The error bars represent the ± 1 SD.
b
a a
b
a
a
b
b
a
b
a a
a
b
a
b
a a
b
a
a
b
a a
b
a a a
b
a

APPENDIX
112
Figure A. 4.3. Effect of compost application (no-without, N-not sterilized and S-sterilized compost) on dry weight per plant of Santana in sterile sand and non-sterilized field soil inoculated with F. avenaceum (Fa), F. solani (Fs), P. medicaginis (Pm) and D. pinodes (Dp). Different letters within one substrate and one pathogen indicate significant differences in fresh weights
among compost treatments at P < 0.05 (Tukey’s HSD test). The error bars represent the ± 1 SD.
Figure A. 4.4. Effect of compost application (no-without, N-not sterilized and S-sterilized compost) on dry weight per plant of EFB33 in sterile sand and non-sterilized field soil inoculated with F. avenaceum (Fa), F. solani (Fs), P. medicaginis (Pm) and D. pinodes (Dp). Different letters within one substrate and one pathogen indicate significant differences in fresh weights
among compost treatments at P < 0.05 (Tukey’s HSD test). The error bars represent the ± 1 SD.
a
c
b a
c
a
b b
a
b
a
a a
b b b
ab
a
b
a
a
b
a ab
b
a
a
b
a
a
b
a a
b
a a
b
ab
a
b ab
a
b
a
a
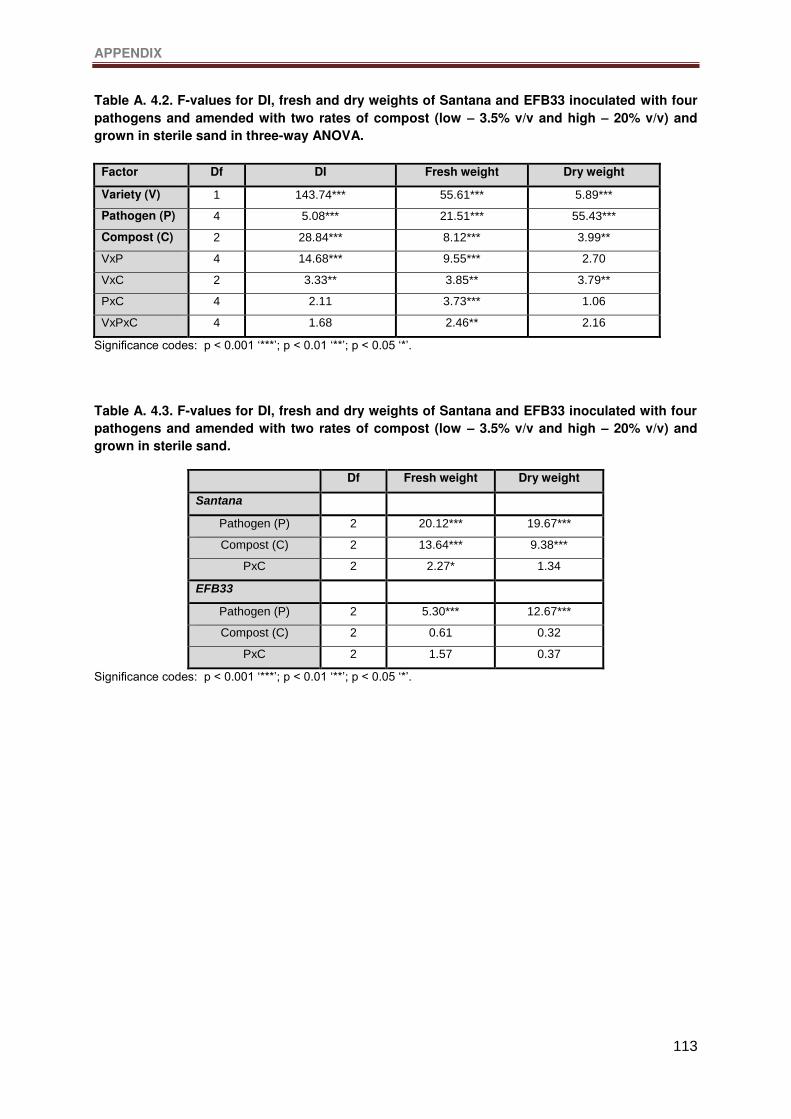
APPENDIX
113
Table A. 4.2. F-values for DI, fresh and dry weights of Santana and EFB33 inoculated with four pathogens and amended with two rates of compost (low – 3.5% v/v and high – 20% v/v) and grown in sterile sand in three-way ANOVA.
Factor Df DI Fresh weight Dry weight
Variety (V) 1 143.74*** 55.61*** 5.89***
Pathogen (P) 4 5.08*** 21.51*** 55.43***
Compost (C) 2 28.84*** 8.12*** 3.99**
VxP 4 14.68*** 9.55*** 2.70
VxC 2 3.33** 3.85** 3.79**
PxC 4 2.11 3.73*** 1.06
VxPxC 4 1.68 2.46** 2.16
Significance codes: p < 0.001 ‘***’; p < 0.01 ‘**’; p < 0.05 ‘*’.
Table A. 4.3. F-values for DI, fresh and dry weights of Santana and EFB33 inoculated with four pathogens and amended with two rates of compost (low – 3.5% v/v and high – 20% v/v) and grown in sterile sand.
Df Fresh weight Dry weight
Santana
Pathogen (P) 2 20.12*** 19.67***
Compost (C) 2 13.64*** 9.38***
PxC 2 2.27* 1.34
EFB33
Pathogen (P) 2 5.30*** 12.67***
Compost (C) 2 0.61 0.32
PxC 2 1.57 0.37
Significance codes: p < 0.001 ‘***’; p < 0.01 ‘**’; p < 0.05 ‘*’.

APPENDIX
114
Figure A. 4.5. Effect of compost application rate (no-without, low- 3.5% v/v and high- 20% v/v) on the dry weight per plant of Santana and EFB33 plants inoculated with F. avenaceum (Fa), F.
solani (Fs), P. medicaginis (Pm) and D. pinodes (Dp). Different letters within one variety and one
pathogen indicate significant differences in fresh weights among compost treatments at P < 0.05
(Tukey’s HSD test). The error bars represent the ± 1 SD.
Table A. 4.4. F-values for DI, fresh and dry weights of Santana and EFB33 inoculated with F.
avenaceum and P. medicaginis, amended with 20% v/v YWC and grown under two temperature regimes (low - 16/12°C and high – 19/16°C) in sterile sand in three-way ANOVA.
Factor Df DI Fresh weight Dry weight
Variety (V) 1 139.77*** 22.08*** 2.57
Temperature (T) 1 12.74*** 40.72*** 10.90***
Pathogen (P) 2 188.08*** 53.79*** 21.69***
Compost (C) 1 141.65** 109.60*** 19.80***
VxT 1 0.09 19.20*** 2.62
VxP 2 31.58*** 57.12*** 17.88***
TxP 2 11.93*** 4.23** 0.94
VxC 1 1.79 60.72*** 14.62***
TxC 1 0.37 0.93 0.22
PxC 2 36.59*** 14.23*** 6.77***
VxTxP 2 1.75 5.78*** 0.33
VxTxC 1 0.21 0.37 0.03
VxPxC 2 0.95 3.38** 0.72
TxPxC 2 3.05* 6.88*** 1.98
VxTxPxC 2 1.89 2.61* 0.10
Significance codes: p < 0.001 ‘***’; p < 0.01 ‘**’; p < 0.05 ‘*’.
a
ab
b b
b
a
b
a
a

APPENDIX
115
Figure A. 4.6. Effect of compost application (no-without, compost- 20% v/v) on the dry weights of Santana (A) and EFB33 (B) plants inoculated with F. avenaceum (Fa) and P. medicaginis (Pm) and grown under different temperature regimes (low – 12/16°C, high – 16/19°C). Asterisk
indicates significant differences in fresh weights between two compost treatments within one
pathogen.
B
A
*
* *
* *

SUMMARY
116
Summary
An alternative way to suppress soil borne diseases is the enhancement of microbial activity
e.g. through the use of organic amendments. Composts are the most effective organic
amendment for this purpose. In pot trials successful disease suppression usually follows
application of 30 to 50% v/v of compost to the potting mixes. In contrast to greenhouse trials,
the amounts of compost that can be applied in the field are considerably lower. In organic
farming field application of off-farm compost is limited to 5 t dry matter (DM) ha-1 and year.
This is equivalent to an application of 3.5% v/v in the top 10 cm of the soil. The work
presented was conducted as part of the “Climate change and production of healthy crops -
processes and adaptation strategies by the year 2030” in the frame of the research project
“KLIFF - Climate Impact Research in Lower Saxony". The overall aim was to evaluate soil
health management with a specific focus on soil borne diseases of peas. For that purpose
field, greenhouse, and laboratory experiments were conducted (i) to explore the disease
dynamics in an organic rotation encompassing winter peas as green manure crop followed
by maize and wheat and the potential of improving system health with a repeated compost
application. (ii) To study in detail the pathogens occurring in the field and (iii) to study the
interactions of these pathogens with a suppressive yard waste compost.
The rotation was followed three times encompassing each time two years starting in fall 2009
to 2011 and ending with the wheat harvest 2011 to 2013, respectively. The winter pea variety
EFB33, Maize Fabregas and winter wheat Achat were used. There were four treatments for
the peas. Two were left uninoculated and two were inoculated by distributing oats colonized
with a local isolate of Phoma medicaginis at sowing with the seed (4 kg 150 m-2). In addition,
two treatments received 5 t dry matter (DM) ha-1 of yard waste compost at sowing resulting in
a complete two-factorial. All field experiments were set up as randomized complete blocks
with four replications. Peas were mulched and incorporated in May followed by Maize. Before
sowing the winter wheat the same plots that had received compost before received the same
amount of compost again.
Overall crop performance of peas, maize and wheat was strongly affected by seasonal
climatic conditions but not by experimental treatments. Application of yard waste compost
overall did not reduce the severity of foot rot of peas or wheat. However, it had a positive
effect on the pea biomass in plots affected by water logging in 2010. Phoma medicaginis was
dominating the root rot pathogen complex on peas and its incidence was not affected by
compost application or year. Besides P. medicaginis, D. pinodes, F. oxysporum, F. solani
and F. avenaceum were isolated from peas. Severe foot rot occurred in 2012 after the frost

SUMMARY
117
in early February with a 40% incidence of F. avenacuem compared to <10 % in the previous
years.
Pathogens occurring on all three crops were sampled and identified microscopically in all
experimental years. Phoma medicaginis var pinodella, Didymella pinodes, Fusarium solani f.
sp. pisi and Fusarium avenaceum were identified as the most important pathogens of the
root and foot rot complex on peas in the field experiment. More than fifteen Fusarium species
were isolated from maize and wheat root and/or stem bases. The incidence of different
species of the complex differed among years while the spectrum of species did not. In all
three seasons F. graminearum and F. culmorum dominated the pathogen complex on maize
and wheat. F. sambucinum, F. crookwellense, F. equisety, F. proliferatum and F.
verticilioides were commonly found on maize, whereas F. avenaceum, F. oxysporum, F.
equiseti and M. nivale were common on wheat seedlings. There was no obvious relationship
between incidence and frequencies of the Fusarium spp. in peas, maize and wheat either
over the course of a given rotation or within year suggesting that the factors influencing
occurrence of the species may not have been connected to the soil, seed, or plant species
but rather to climatic and potentially other unknown environmental conditions.
P. medicaginis var pinodella, D. pinodes, F. solani f. sp. pisi and F. avenaceum were
assessed for their aggressiveness on the spring pea variety Santana and winter pea EFB33
under controlled conditions. Furthermore, in addition to peas, the pathogenicity of two
isolates of F. avenaceum originating from wheat and pea was determined on eight different
plant species commonly grown in Germany: Maize (Zea mays), Winter wheat (Triticum
aestivum), Oat (Avena sativa), Black oat (Avena strigosa), Subterranean clover (Trifolium
subterraneum), White clover (Trifolium repens), Indian mustard (Brassica juncea) and White
mustard (Sinapis alba). In addition, the potential of Yard Waste Compost (YWC) to suppress
root and foot rot of peas caused by P. medicaginis, D. pinodes, F. solani and F. avenaceum
was studied in greenhouse bioassays at different application rates.
Bioassays under controlled conditions using spore solutions as inoculum showed that the
spring pea variety Santana was considerably more susceptible to all four pathogens tested in
comparison to the winter pea variety EFB33 in both sterile sand and non-sterilized field soil.
In sterile sand F. avenaceum was the most aggressive pathogen, followed by P. medicaginis,
D. pinodes, and F. solani. Aggressiveness of all pathogens was greatly reduced in non-
sterile field soil. Reduction of growing temperatures from 19/16°C day/night to 16/12°C and
13/10°C reduced plant growth and slightly increased disease on EFB33 whereas the highest
disease severity on Santana was observed at the highest temperature, 19/16°C. The two
isolates of F. avenaceum caused severe symptoms on roots of all nine plant species tested.
Especially susceptible were White clover (Trifolium repens), Subterranean clover (Trifolium

SUMMARY
118
subterraneum), Indian mustard (Brassica juncea) and White mustard (Sinapis alba) in
addition to peas.
Application of 20% v/v of YWC reduced disease on peas due to F. avenaceum, F. solani, P.
medicaginis and D. pinodes. The intensity of suppression of root and foot rot of peas
depended on pea variety, pathogen and growing media used. As disease severity on EFB33
was overall low due to its high resistance no statistically significant effects of compost
application were observed in sterile sand or non-sterile field soil. In contrast, on Santana
when sterilized sand was used as growing substrate compost application successfully
reduced disease caused by F. avenaceum, P. medicaginis, and D. pinodes. Furthermore,
fresh weight was significantly increased with compost application in all treatments. In non-
sterilized field soil as well Santana as EFB33 were moderately diseased in the uninoculated
controls due to the presence of native inoculum of F. solani. At the same time, overall
disease in the inoculated plants was considerably lower than in the sterile sand suggesting
that a certain degree of natural suppression was occurring in the field soil. In order to
determine if YWC applied at the rate allowed in the field was able to suppress disease in
greenhouse trials under controlled conditions, tests were performed with compost application
rate of 3.5% v/v. Suppression was also achieved with this lower application rate, although
there was a tendency that plants were healthier when substrate was amended with 20% v/v
compost.
To investigate if the cause for the suppressive effects was of biotic origin experiments were
conducted comparing the effects of non-sterile with γ sterilized compost. The suppressive
effect was lost in the pathosystems with Fusarium spp. and sterile compost while in the
systems with D. pinodes and P. medicaginis sterilized compost still decreased disease. This
suggests that different mechanisms of suppression may be involved and that the
suppression of disease caused by Fusarium spp. is biological in origin, whereas chemical
and physical properties of compost are playing an additional role in the suppression of
disease caused by D. pinodes and P. medicaginis. In contrast to sterilized sand when non-
sterilized field soil was used as substrate, compost application did not result in significant
reduction of disease, and in treatments with γ-irradiated compost disease symptoms were
even more severe than without compost application. This suggests that the sterilized
compost may have served as a food base for the native inoculum present in the soil.
In conclusion, the winter pea variety EFB33 can be considered as a good alternative to the
commonly grown spring variety Santana due to its broad resistance towards predominant
root rot pathogens of peas in German conditions. Most field experiments are conducted with
one-time applications of compost. Compost based agriculture will apply compost on a regular
basis, however. Although inconsistency in the compost induced disease suppression

SUMMARY
119
between field and greenhouse tests was observed, YWC has the potential to improve pea
health. Longer term regular compost applications in the field should be tested for their
capacity to build resilient soils that are the basis of sustainable agricultural systems.

ACKNOWLEDGMENTS
120
Acknowledgments
First and foremost I want to thank my supervisor Professor Dr. Maria R. Finckh for all of her
support, guidance, motivation and contribution of time and ideas, especially during the final
stages of this PhD. I am thankful for the excellent example she has provided as a scientist
and professor. My gratitude goes also to Professor Dr. Gunter Backes for his kind and
valuable advices.
I gratefully acknowledge the project “KLIFF - Climate Impact Research in Lower Saxony" and
the University of Kassel for founding my research. Also, Dr. Tivoli from the National Institute
of Agronomic Research (INRA) France and Prof. Dr. von Tiedemann from the Plant
Pathology Department of the Georg-August-University in Göttingen for kindly providing
isolates of P. medicaginis and D. pinodes used in this work.
I cannot find adequate words to thank enough all my colleagues from the Department of
Ecological Plant Protection for all of their help and assistance during the last five years, but
even more I am grateful for enriching my life with new friends.
I would like to thank Adnan for his friendship and patience through my ups and downs,
especially in the last several months. I am grateful for his company and support and most of
all for making me laugh.
To my dearest friends Marija Pavleska Wood, Marija Kalentić and Miroslava Spernjak I am
eternally grateful for being always there for me, accompanying me through good and bad
times. You are truly the best friends one can wish for.
Lastly, I would like to thank my parents Radmila and Nemanja for their unconditional love
and encouragement. They raised me with a love of science and encouraged me to explore
the world with eyes wide open always having in mind that it is not only the destination that
matters rather the road you take.
Without all of you this would be just a work, but you all made it an experience that I would
cherish forever.
With all of my gratitude,
Jelena

ERKLÄRUNG
121
Erklärung
Hiermit versichere ich, dass ich die vorliegende Dissertation selbstständig, ohne unerlaubte
Hilfe Dritter angefertigt und andere als die in der Dissertation angegebenen Hilfsmittel nicht
benutzt habe. Alle Stellen, die wörtlich oder sinn-gemäß aus veröffentlichten oder
unveröffentlichten Schriften entnommen sind, habe ich als solche kenntlich gemacht. Dritte
waren an der inhaltlich-materiellen Erstellung der Dissertation nicht beteiligt; insbesondere
habe ich hierfür nicht die Hilfe eines Promotionsberaters in Anspruch genommen. Kein Teil
dieser Arbeit ist in einem anderen Promotions- oder Habilitationsverfahren verwendet
worden.
Witzenhausen, 22. Januar 2015
________________________________
Jelena Baćanović








![[Model names] PEA-RP200GAQ PEA-RP250GAQ PEA-RP400GAQ PEA …mitsubishitech.co.uk/Data/Mr-Slim_Indoor/PEA[H]-RP/... · PEA-RP200GAQ Fan Performance Curve 50Hz PEA-RP250GAQ Fan Performance](https://static.cupdf.com/doc/110x72/600812e007963a6f320df208/model-names-pea-rp200gaq-pea-rp250gaq-pea-rp400gaq-pea-h-rp-pea-rp200gaq.jpg)














![[Model names] PEA-RP200GAQ PEA-RP250GAQ PEA-RP400GAQ PEA …H]-RP/2016-2015/PEA... · PEA-RP200GAQ Fan Performance Curve 50Hz PEA-RP250GAQ Fan Performance Curve 50Hz PEA-RP400GAQ](https://static.cupdf.com/doc/110x72/5f80bcbd06056748830d6182/model-names-pea-rp200gaq-pea-rp250gaq-pea-rp400gaq-pea-h-rp2016-2015pea.jpg)


![PEA-RP250GA PEA-RP400GA PEA-RP500GA - …H]-RP/2010-2009/... · PEA-RP250GA PEA-RP400GA PEA-RP500GA ... Cautions for units utilising refrigerant R410A ... It is also possible to attach](https://static.cupdf.com/doc/110x72/5ad5679d7f8b9a075a8cd92b/pea-rp250ga-pea-rp400ga-pea-rp500ga-h-rp2010-2009pea-rp250ga-pea-rp400ga.jpg)



