



Morphology, rbcL phylogeny and distribution of distromatic Ulva(Ulvophyceae, Chlorophyta) in Ireland and southern Britain
CIARAN J. LOUGHNANE1,2,3*, LYNNE M. MCIVOR
1,2,3, FABIO RINDI2,{, DAGMAR B. STENGEL
1,2,3AND MICHAEL D. GUIRY
2,3
1Department of Botany, National University of Ireland, Galway, Ireland2Martin Ryan Institute, National University of Ireland, Galway, Ireland
3Environmental Change Institute, National University of Ireland, Galway, Ireland4Department of Biological Sciences, The University of Alabama, Box 870345, Tuscaloosa, AL 35487, USA
C.J. LOUGHNANE, L.M. MCIVOR, F. RINDI, D.B. STENGEL AND M.D. GUIRY. 2008. Morphology, rbcL phylogeny anddistribution of distromatic Ulva (Ulvophyceae, Chlorophyta) in Ireland and southern Britain. Phycologia 47: 416–429.DOI: 10.2216/07-61.1
Species of Ulva occur worldwide in all aquatic habitats from freshwater through brackish to fully saline environments.The scarcity of stable morphological characters and the morphological plasticity linked to environmental conditionstypical of species referred to this genus make identification and circumscription problematic. In Ireland and Britain thetaxonomy of Ulva species has been uncertain, and only three species have been reported: Ulva lactuca, U. rigida and U.
olivascens (later reassigned to Umbraulva olivascens). We reassessed the taxonomic status of this genus by combiningmorphological investigations with analyses of rbcL sequence data for 24 selected specimens from the Irish and southernBritish shores. This combination of methods allowed the identification of seven lineages, which were considered torepresent the following species: Ulva rigida, U. scandinavica, U. lactuca, U. gigantea, U. rotundata, U. californica andUmbraulva olivascens. Of these, U. scandinavica, U. gigantea, U. rotundata and U. californica are newly reported; thedistribution of U. olivascens was shown to be much wider than previously thought. The use of morphological featuresproved to be largely inconclusive and of limited value for circumscription of species. In the rbcL phylogeny, U.
olivascens formed a sister clade to all other species. The results support the conspecificity of U. rigida, U. scandinavica
and U. armoricana and the conspecificity of U. pseudocurvata and U. compressa already suggested in previous studies.
KEY WORDS: Atlantic, Chlorophyta, Ireland, Britain, Molecular systematics, Morphology, Phylogeny, rbcL gene, Ulva,Ulvophyceae
INTRODUCTION
The genus Ulva Linnaeus (Ulvophyceae, Ulvales) is
cosmopolitan in its distribution, with species occurring in
all aquatic habitats from freshwater through brackish to
fully saline environments (Guiry & Guiry 2007). Ulva
species have either a monostromatic tubular (previously
regarded as Enteromorpha Link; Hayden et al. 2003) or
distromatic, foliose thallus that can vary in length, when
mature, from a few centimetres to over 1 m. As presently
circumscribed, the genus includes more than 100 species
(Guiry & Guiry 2007). The diversity and species-level
delimitation of this genus in Ireland and Britain has been
surrounded by considerable uncertainty. In the most
comprehensive monograph currently available of the green
seaweeds of this area, Burrows (1991) reported three species
of distromatic Ulva for Ireland and Britain: U. lactuca
Linnaeus, U. rigida C. Agardh and U. olivascens P.J.L.
Dangeard [later reassigned as Umbraulva olivascens (P.J.L.
Dangeard) Bae & I.K. Lee, 2001]. Hardy & Guiry (2003)
reported that both U. lactuca and U. rigida were recorded
from most shores in Ireland and Britain; however, they
noted that ‘members of this genus [Ulva] are extremely
difficult to separate morphologically’ and consequently that
records of U. lactuca, in particular, ‘probably represent
several different entities’. Umbraulva olivascens was record-
ed from only three locations in Ireland (Hardy & Guiry
2003): Carna and Killary Harbour, Co. Galway, and New
Quay, Co. Clare. These authors also questioned the lack of
records and explained that this may be due to its confusion
with U. lactuca. Bliding (1968) recorded eight species (two
new and one recombination) for Europe: U. lactuca, U.
rigida, U. olivascens, U. scandinavica Bliding, U. rotundata
Bliding, U. gigantea (Kutzing) Bliding, U. curvata (Kutzing)
De Toni and U. neapolitana Bliding. Koeman & van den
Hoek (1981) reported five species from the Netherlands, one
of which was new: U. pseudocurvata Koeman & Hoek, and
Dion et al. (1998) described a new species, U. armoricana
Dion, de Reviers & Coat, from Brittany, France. Thus, at
present, there are nine species of distromatic Ulva reported
in coastal waters of northwest Europe.
As Ulva has a very simple morphology, delineation of
species by morphology is notoriously difficult (Bliding
1963; Tanner 1979; Koemann & van den Hoek 1981;
Hoeksema & van den Hoek 1983; Blomster et al. 1999; Tan
et al. 1999; Hayden & Waaland 2002; Hayden et al. 2003;
Hayden & Waaland 2004). The morphological characters
traditionally used for identification show considerable
plasticity in response to changing environmental conditions
(Titylanov et al. 1975; Tanner 1979, 1986; Malta et al.
1999). Morphology can vary with the age of the thallus, the
time of the year, salinity and whether the thallus is attached
* Corresponding author ([email protected]).{Present address: Department of Biological Sciences, The
University of Alabama, Box 870345, Tuscaloosa, AL 35487, USA.
Phycologia (2008) Volume 47 (4), 416–429 Published 3 July 2008
416

or floating (e.g. Malta et al. 1999). Thalli of Ulva fenestrata
Postels & Ruprecht varied in size, shape and thickness
depending on wave exposure, vertical position and the time
of year (Titlyanov et al. 1975; Tanner 1979, 1986). The
blade length, width and habit of Ulva californica Wille
varied with latitude, season, wave exposure and tempera-
ture (Tanner 1986). In U. taeniata (Setchell) Setchell & N.L.
Gardner the blade thickness increased, and the length and
number of marginal teeth decreased with increasing water
temperature. This temperature-related morphological var-
iation explains distinct seasonal variation in the morphol-
ogy of both species (Tanner 1979). Pyrenoid number was
first considered by Bliding (1968) to be an important
taxonomic character. In addition, Koeman & van den
Hoek (1981), Hoeksema & van den Hoek (1983) and Dion
et al. (1998) have also regarded it to be a reliable character
in the identification of European Ulva. However, Tanner
(1986) deemed the use of pyrenoid numbers as too variable,
and Malta et al. (1999) found the number of pyrenoids in
U. scandinavica from the Netherlands was greater than one
in 50% of cells in early May and was just one pyrenoid in
nearly all cells by the end of the growing season. These
findings are in stark contrast to Koeman & van den Hoek
(1981) and Hoeksema & van den Hoek (1983), who
maintained that U. scandinavica consistently possesses two
or more pyrenoids.
In recent decades, with the increasing use of molecular
data to resolve the taxonomic status of various algal
groups (Medlin et al. 2007), the application of molecular
systematics to resolve and identify Ulva species has
developed (Blomster et al. 1998, 1999; Coat et al. 1998;
Malta et al. 1999; Tan et al. 1999; Bae & Lee 2001;
Hayden & Waaland 2002; Hayden et al. 2003; Shimada et
al. 2003; Hayden & Waaland 2004). In response to the
difficulty encountered in accurately identifying species of
Ulva (particularly distromatic species) from Ireland and
Britain, this study aimed to clarify the taxonomic status of
the members of this group by using morphological
investigations and rbcL sequence data. In addition to
providing new information about the species composition
on Irish and English shores, the feasibility of using
morphology-based techniques in the identification of these
ubiquitous algae is discussed.
MATERIAL AND METHODS
Over 210 specimens of Ulva were collected from sites in
Ireland and the south coast of Britain from 2000 to 2003
(Fig. 1). All specimens were pressed and dried on herbar-
ium sheets for morphological analysis, and a portion of the
thallus was preserved in silica gel for molecular analysis.
For the present study, representative morphotypes were
selected for morphological and molecular analyses (Ta-
ble 1). Voucher specimens were deposited in the phycolog-
ical herbarium of the National University of Ireland,
Galway (GALW).
For the morphological analysis, sections were cut from
the apical, middle and basal regions of the thallus and
Fig. 1. Map of Ireland and southern Britain showing the sites (N) sampled in the present study.
Loughnane et al.: Distromatic Ulva in the northeastern Atlantic 417

rehydrated in sterilized seawater for 30 min prior to
examination (after Tanner 1986). Where it was difficult to
establish pyrenoid number, specimens were stained with
iodine in potassium iodide (I2KI) to highlight the cell walls
and pyrenoids. The morphological characters used for
species identification were chosen following Bliding (1968),
Koeman & van den Hoek (1981), Hoeksema & van den
Hoek (1983), Tanner (1986) and Dion et al. (1998) and were
as follows: thallus form, texture and colour; shape and form
of the basal region; shape, size and arrangement of the cells;
shape and position of the chloroplast; number of pyrenoids;
thickness of the thallus; and the presence or absence of
teeth-like projections on the blade edge (Table 2). Micro-
scopic observations were carried out using a Nikon
Optiphot II Microscope, and photographs were taken
using a Nikon DXM1200 digital camera.
DNA was extracted from fresh or silica-dried material of
selected morphotypes (Table 1) either by using the Qiagen
DNeasy Plant Mini Kit (Qiagen GmbH, Hilden, Germany)
in accordance with the manufacturer’s instructions or by
the CTAB method, modified after Doyle & Doyle (1987).
Prior to extraction, silica-dried material was rehydrated in
sterile seawater. In both fresh and rehydrated silica-dried
material c. 1 cm2 of the thallus was excised using a sterile
blade and ground in liquid nitrogen in a 1.5-ml micro-
centrifuge tube using a sterile pellet-pestle. Using the CTAB
method, 600 ml of CTAB extraction buffer (0.1 M Tris,
0.02 M EDTA, 1.4 M NaCl, 2% CTAB) were added along
with 1% PVP and 1.5 ml b-mercaptoethanol. The tubes
were incubated at 60uC for a minimum of 1 h with constant
gentle mixing. This was followed by two to three
purification steps with 24 : 1 chloroform/isoamyl alcohol
and precipitated with 220uC isopropanol. DNA was
dissolved in 100 ml 0.1 M TE. The quantity and quality of
the extracted DNA was assessed on a 1% agarose gel prior
to PCR amplification.
The plastid-encoded rbcL gene was PCR amplified using
the primers RH1 (59-ATGTCACCACAAACAGAAAC-
TAAAGC-39) and 1385r (59-AATTCAAATTTAATTT-
CTTTCC-39) (Manhart 1994). All reactions were run in a
PTC-200 DNA Engine thermal cycler (MJ Research Inc.,
Waltham, MA, USA). Total genomic DNA (10–20 ng) was
added to 25-ml PCR reactions each containing final
concentrations of 2.5 mM MgCl2, 200 mM dNTPs,
Table 1. Specimens and sampling locations, including GenBank and NUI, Galway Herbarium (GALW), accession numbers used for rbcLanalysis. n/a 5 not applicable.
Sample Species Location and collection date GenBank GALW
Ua21 U. fasciata Faial, Azores (38u219N, 28u259W) (18 Aug. 2002) EU484418 015524U16 U. lactuca Dun Laoghaire, Co. Dublin, Ireland (53u179N, 06u079W) (08 Jun. 2001) EU484399 015506U19 U. lactuca Dun Laoghaire, Co. Dublin, Ireland (53u179N, 06u079W) (08 Jun. 2001) EU484400 015508U29 U. rotundata Casla, Co. Galway, Ireland (53u17.39N, 09u33.59W) (18 Sep. 2001) EU484401 015512U31 U. gigantea Oranmore, Co. Galway, Ireland (53u16.09N, 08u56.09W) (21 Sep. 2001) EU484402 015521U33 U. gigantea Oranmore, Co. Galway, Ireland (53u16.09N, 08u56.09W) (21 Sep. 2001) EU484403 015519U34 U. rotundata Cashel Bay, Co. Galway, Ireland (53u25.09N, 09u48.99W) (20 Sep. 2001) EU484404 015513U38 Umbraulva olivascens Carna, Co. Galway, Ireland (53u199N, 09u509W) (20 Sep. 2001) EU484405 015529U41 U. rotundata Ballyvaughan, Co. Clare, Ireland (53u07.29N, 09u09.29W) (21 Sep. 2001) EU484406 015522U46 Umbraulva olivascens New Quay, Co. Clare, Ireland (53u09.39N, 09u04.59W) (1 Apr. 2002) EU484407 015530U601 U. rigida Killary Harbour, Co. Galway, Ireland (53u36.09N, 09u48.89W) (18 May 2002) EU484408 015523U72 U. lactuca Crosshaven, Co. Cork, Ireland (51u489N, 08u169W) (25 Jun. 2002) EU484409 015509U76 Umbraulva olivascens East Ferry, Co. Cork, Ireland (51u529N, 08u129W) (25 Jun. 2002) EU484410 015528U95 Umbraulva olivascens Portmagee, Co. Kerry, Ireland (51u539N, 10u219W) (27 Jun. 2002) EU484411 015531U111 U. scandinavica Langstone Harbour, Hampshire, UK (49u509N, 1u009W) (20 Jul. 2001) EU484412 015518U125 U. lactuca Lulworth Cove, Dorset, UK (50u229N, 2u099W) (21 Jul. 2001) EU484413 015511U135 U. gigantea Port Isaac Bay, Cornwall, UK (50u35.69N, 4u49.99W) (24 Jul. 2001) EU484414 015520U157 U. californica Dingle Harbour, Co. Kerry, Ireland (52u08.39N, 10u16.69W) (27 Jun. 2002) EU484415 015517U168 U. scandinavica Kilbrittan, Co. Cork, Ireland (51u39.09N, 08u40.79W) (08 Aug. 2002) EU484416 015516U311 U. rigida Kilmore Quay, Co. Wexford, Ireland (52u099N, 06u359W) (24 Sep. 2002) EU484417 015525Aran1 U. rigida Inis Mor, Aran Islands, Ireland (53u079N, 09u409W) (25 Aug. 2002) EU484395 015515CashA U. rotundata Cashel Bay, Co. Galway, Ireland (53u25.09N, 09u48.89W) (5 Apr. 2003) EU484396 015514CashB U. compressa Cashel Bay, Co. Galway, Ireland (53u25.09N, 09u48.89W) (5 Apr. 2003) EU484397 015526E102 U. lactuca Finavarra, Co. Clare, Ireland (53u09.59N, 09u07.19W) (7 Apr. 2003) EU484398 015510GenBank U. armoricana Yokohama, Japan (Shimada et al. 2003) AB097632 n/aGenBank U. californica California, USA (Hayden & Waaland 2004) AY422565 n/aGenBank U. compressa Portaferry, N. Ireland (Blomster et al. 1998) AY255859 n/aGenBank U. curvata Delaware, USA (Sherwood et al. 2000) AF189071 n/aGenBank U. fasciata Hawaii, USA (Hayden & Waaland 2004) AY422565 n/aGenBank U. fenestrata Washington, USA (Hayden & Waaland 2002) AF499668 n/aGenBank U. intestinalis Karlskrona, Sweden (Shimada et al. 2003) AB097617 n/aGenBank U. intestinalis London, UK (Blomster et al. 1998) AY255860 n/aGenBank U. lactuca Alaska, USA (Hayden & Waaland 2004) AY422546 n/aGenBank U. pseudocurvata California, USA (Hayden & Waaland 2004) AY422553 n/aGenBank U. pseudocurvata Ythan Estuary, Scotland (Tan et al. 1999) AF255866 n/aGenBank U. rigida Pelluco Beach, Chile (Hayden & Waaland 2004) AY422564 n/aGenBank U. scandinavica Wadden Sea, Netherlands (Shimada et al. 2003) AB097629 n/aGenBank Umbraulva olivascens Portaferry, N. Ireland (Hayden et al. 2003) AY255876 n/a
1 Collected by Dr. Stefan Kraan, Irish Seaweed Centre, Martin Ryan Institute, NUI, Galway.
418 Phycologia, Vol. 47 (4), 2008
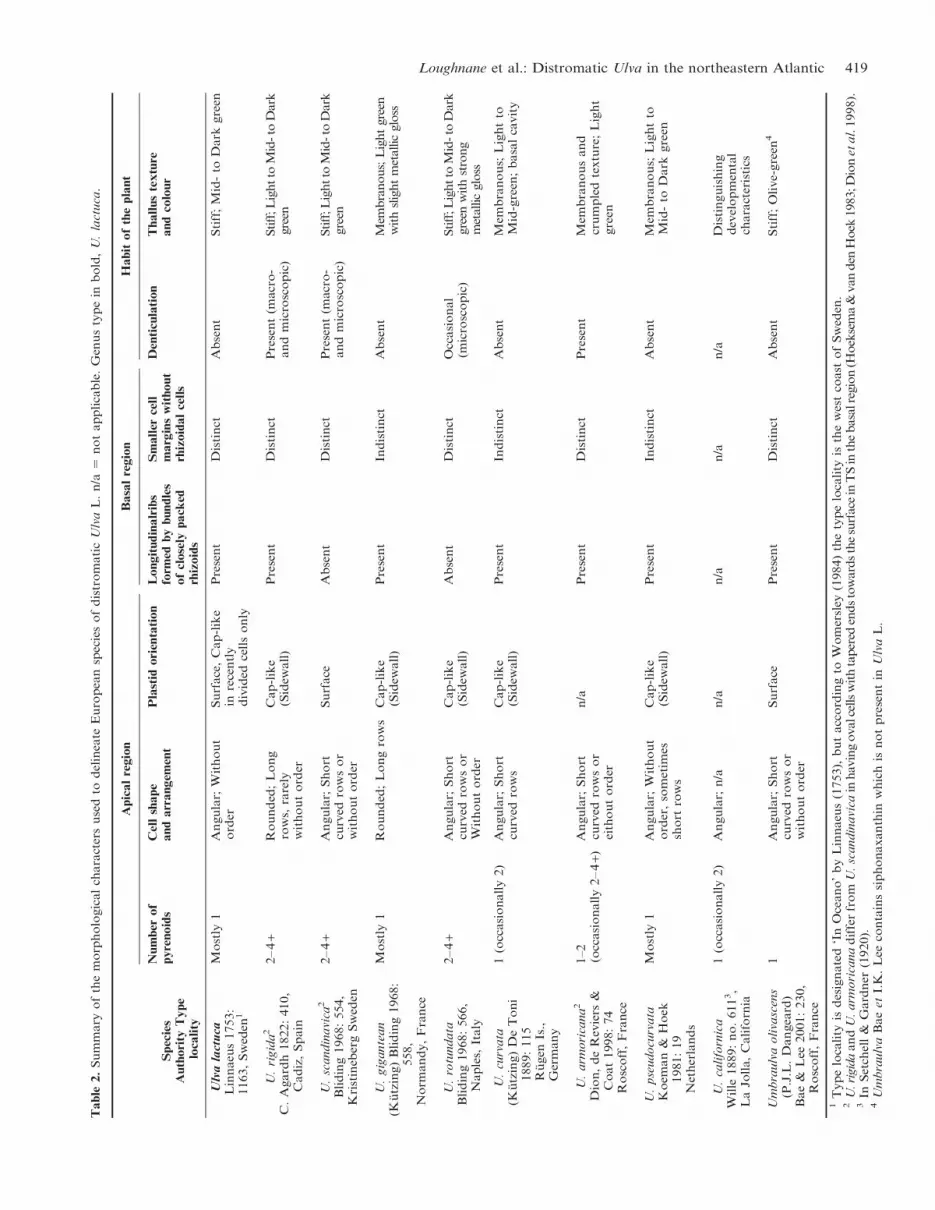
Table
2.
Su
mm
ary
of
the
mo
rph
olo
gic
al
chara
cter
su
sed
tod
elin
eate
Eu
rop
ean
spec
ies
of
dis
tro
mati
cU
lva
L.
n/a
5n
ot
ap
pli
cab
le.
Gen
us
typ
ein
bo
ld,
U.
lact
uca
.
Apic
al
regio
nB
asa
lre
gio
nH
abit
of
the
pla
nt
Spec
ies
Auth
ori
tyT
ype
loca
lity
Num
ber
of
pyre
noid
sC
ell
shape
and
arr
angem
ent
Pla
stid
ori
enta
tion
Longit
udin
alr
ibs
form
edby
bundle
sof
close
lypack
edrh
izoid
s
Sm
all
erce
llm
arg
ins
wit
hout
rhiz
oid
al
cell
s
Den
ticu
lati
on
Thall
us
tex
ture
and
colo
ur
Ulv
ala
ctuca
Lin
naeu
s1753:
1163,
Sw
eden
1
Mo
stly
1A
ngu
lar;
Wit
ho
ut
ord
erS
urf
ace
,C
ap
-lik
ein
rece
ntl
yd
ivid
edce
lls
on
ly
Pre
sen
tD
isti
nct
Ab
sen
tS
tiff
;M
id-
toD
ark
gre
en
U.
rigid
a2
C.
Agard
h1822:
410,
Cad
iz,
Sp
ain
2–
4+
Ro
un
ded
;L
on
gro
ws,
rare
lyw
ith
ou
to
rder
Cap
-lik
e(S
idew
all
)P
rese
nt
Dis
tin
ctP
rese
nt
(macr
o-
an
dm
icro
sco
pic
)S
tiff
;L
igh
tto
Mid
-to
Dark
gre
en
U.
scandin
avi
ca2
Bli
din
g1968:
554,
Kri
stin
eber
gS
wed
en
2–
4+
An
gu
lar;
Sh
ort
curv
edro
ws
or
wit
ho
ut
ord
er
Su
rface
Ab
sen
tD
isti
nct
Pre
sen
t(m
acr
o-
an
dm
icro
sco
pic
)S
tiff
;Lig
ht
toM
id-
toD
ark
gre
en
U.
gig
ante
an
(Ku
tzin
g)
Bli
din
g1968:
558,
No
rman
dy,
Fra
nce
Mo
stly
1R
ou
nd
ed;
Lo
ng
row
sC
ap
-lik
e(S
idew
all
)P
rese
nt
Ind
isti
nct
Ab
sen
tM
emb
ran
ou
s;L
igh
tgre
enw
ith
slig
ht
met
allic
glo
ss
U.
rotu
ndata
Bli
din
g1968:
566,
Nap
les,
Italy
2–
4+
An
gu
lar;
Sh
ort
curv
edro
ws
or
Wit
ho
ut
ord
er
Cap
-lik
e(S
idew
all
)A
bse
nt
Dis
tin
ctO
ccasi
on
al
(mic
rosc
op
ic)
Sti
ff;L
igh
tto
Mid
-to
Dark
gre
enw
ith
stro
ng
met
allic
glo
ss
U.
curv
ata
(Ku
tzin
g)
De
To
ni
1889:
115
Ru
gen
Is.,
Ger
man
y
1(o
ccasi
on
all
y2)
An
gu
lar;
Sh
ort
curv
edro
ws
Cap
-lik
e(S
idew
all
)P
rese
nt
Ind
isti
nct
Ab
sen
tM
emb
ran
ou
s;L
igh
tto
Mid
-gre
en;
basa
lca
vit
y
U.
arm
ori
cana
2
Dio
n,
de
Rev
iers
&C
oat
1998:
74
Ro
sco
ff,
Fra
nce
1–2
(occ
asi
on
all
y2–
4+)
An
gu
lar;
Sh
ort
curv
edro
ws
or
eith
ou
to
rder
n/a
Pre
sen
tD
isti
nct
Pre
sen
tM
emb
ran
ou
san
dcr
um
ple
dte
xtu
re;
Lig
ht
gre
en
U.
pse
udocurv
ata
Ko
eman
&H
oek
1981:
19
Net
her
lan
ds
Mo
stly
1A
ngu
lar;
Wit
ho
ut
ord
er,
som
etim
essh
ort
row
s
Cap
-lik
e(S
idew
all
)P
rese
nt
Ind
isti
nct
Ab
sen
tM
emb
ran
ou
s;L
igh
tto
Mid
-to
Dark
gre
en
U.
cali
forn
ica
Wil
le1889:
no
.611
3,
La
Joll
a,
Cali
forn
ia
1(o
ccasi
on
all
y2)
An
gu
lar;
n/a
n/a
n/a
n/a
n/a
Dis
tin
gu
ish
ing
dev
elo
pm
enta
lch
ara
cter
isti
cs
Um
bra
ulv
aoli
vasc
ens
(P.J
.L.
Dan
gea
rd)
Bae
&L
ee2001:
230,
Ro
sco
ff,
Fra
nce
1A
ngu
lar;
Sh
ort
curv
edro
ws
or
wit
ho
ut
ord
er
Su
rface
Pre
sen
tD
isti
nct
Ab
sen
tS
tiff
;O
live-
gre
en4
1T
yp
elo
cali
tyis
des
ign
ate
d‘I
nO
cean
o’
by
Lin
naeu
s(1
753),
bu
tacc
ord
ing
toW
om
ersl
ey(1
984)
the
typ
elo
cali
tyis
the
wes
tco
ast
of
Sw
eden
.2
U.ri
gid
aan
dU
.arm
ori
cana
dif
fer
fro
mU
.sc
andin
avic
ain
havin
go
valc
ells
wit
hta
per
eden
ds
tow
ard
sth
esu
rface
inT
Sin
the
basa
lreg
ion
(Ho
ekse
ma
&van
den
Ho
ek1983;D
ion
et
al.
1998).
3In
Set
chel
l&
Gard
ner
(1920).
4U
mbra
ulv
aB
ae
et
I.K
.L
eeco
nta
ins
sip
ho
naxan
thin
wh
ich
isn
ot
pre
sen
tin
Ulv
aL
.
Loughnane et al.: Distromatic Ulva in the northeastern Atlantic 419

1.0 mM of each primer, 0.5 units of Taq polymerase
(Bioline, London, UK) and 103 PCR reaction buffer. The
PCR cycle consisted of an initial denaturing phase of 3 min
at 94uC, followed by 35 cycles of 94uC for 1 min, 45uC for
2 min and 65uC for 2 min (after Hayden et al. 2003).
The success of the PCR reaction was assessed on a 1%
agarose gel, stained with ethidium bromide (EtBr) and
visualized under UV illumination. The PCR products were
purified using the Roche High PureTM PCR Product
Purification Kit (Roche Diagnostics Ltd, Lewes, East
Sussex, UK) and sequenced commercially by MWG
Biotech (UK) Ltd (Milton Keynes, UK).
The DNA sequence alignments were constructed by eye
using MacClade v. 4.05 (Maddison & Maddison 2002). The
in-group taxa (n 5 38) are listed in Table 1; other rbcL
sequences of Ulva were retrieved from GenBank and added
to the samples sequenced in the course of this study. These
sequences were selected as additional in-groups in order to
provide a better resolution at the species level and a more
comprehensive representation of the variation of the rbcL
gene in the genus. Sequence data were analysed with PAUP
v. 4.0b10 (Swofford 2003) for maximum parsimony (MP),
maximum likelihood (ML) and neighbour joining (NJ),
using a maximum likelihood distance algorithm as input,
and posterior probabilities were estimated by Bayesian
analysis using MrBayes v. 3.1 (Ronquist & Huelsenbeck
2003). All phylogenetic trees were rooted using Monostroma
nitidum Wittrock, Blidingia minima (Nageli ex Kutzing)
Kylin, Derbesia marina (Lyngbye) Kjellman and Bryopsis
plumosa (Hudson) C. Agardh. As outgroups Blidingia was
resolved as sister taxon to Ulva and Umbraulva in previous
studies (Hayden & Waaland 2002); the other taxa were
selected in order to have an adequate representation of
ulvophycean taxa more and less closely related to Ulva in
the analyses. Modeltest v. 3.7 (Posada & Crandall 1998)
was used to determine the most appropriate evolutionary
model for the ML analysis. It specified a general time
reversible plus gamma distributed model (GTR+G) using
the Akaike information criterion. The rate matrix was
specified as
[A-C] 5 1.2393, [A-G] 5 3.0300, [A-T] 5 3.5636, [C-G] 5
1.7686, [C-T] 5 9.8624 and [G-T] 5 1.0000, with base
frequencies at A 5 0.2852, C 5 0.1520, G 5 0.2267 and T
5 0.3355 and a gamma distribution shape parameter (G) 5
0.2842. The robustness of the data set was determined by
bootstrapping (Felsenstein 1995), with 1000 resamplings for
both NJ and MP. Bayesian analysis of phylogenies is
similar to maximum likelihood in that there is a predefined
evolutionary model and the program (MrBayes v. 3.1;
Ronquist & Huelsenbeck 2003) searches for the best trees
that are consistent with both the model and the data (Hall
2001). Posterior probabilities estimated by Bayesian infer-
ence were calculated under the parameters specified by
Modeltest. Four independent runs each using four simul-
taneous Markov chains were run for 1,000,000 generations,
sampling every 100 generations, with a specified burn-in of
25,000 generations to allow for stabilisation of the
likelihood value. The results were compared from the four
independent runs to ensure that the likelihood had reached
convergence. A 50% majority rule consensus tree was then
created from the trees generated.
RESULTS
Morphological study
The results of the morphological investigations of species of
distromatic Ulva from Ireland and southern Britain are
summarised in Table 3. Based on the morphological
characters used by Bliding (1968), Koeman & van den
Hoek (1981), Hoeksema & van den Hoek (1983), Tanner
(1986) and Dion et al. (1998) (Table 2), seven putative
species from Ireland and southern Britain were identified
from the specimens examined: these corresponded to Ulva
rigida, U. scandinavica, U. lactuca, U. gigantea, U.
rotundata, U. californica and Umbraulva olivascens.
The first species group circumscribed on morphology was
the U. rigida group and included the samples Aran1
(Fig. 2), U60 and U311. Morphological characters for this
group were very stable and corresponded to published
information used to delineate this species. All specimens
examined had more than two pyrenoids per cell, and cells
were rounded in shape and formed long rows that were
particularly evident in the middle and apical regions
(Fig. 9). In the middle region, the lengths of the cells varied
between 15 and 25 mm and the cell widths between 9 and
31 mm. The plastid was orientated to the outer cell wall in
surface view. In the basal region, longitudinal rows or ribs
formed by closely packed groups of rhizoidal cells were
evident, as were the distinct margins of normal vegetative
cells that lacked any rhizoidal cells. Cells in transverse
section from the basal region were spindle-shaped with
tapered ends (Fig. 10). The formation of denticulations at
the edge of the thallus was observed in all specimens;
however, they occurred only in the basal region in U60.
Thalli were generally stiff in texture when fresh, sometimes
perforated, and varied in colour from light to dark green.
The second species group identified included samples
U111 and U168 (Fig. 3), both of which were sampled from
‘‘green tides’’ in Ireland and southern Britain, and were
identified as U. scandinavica. Specimens examined had
either one to two or . 2 pyrenoids per cell. Cells were
polygonal in shape with rounded corners and were
arranged in no particular order (Fig. 11). The lengths of
the cells in the middle region varied between 14 and 25 mm,
and the cell widths were 15–24 mm. The green-tide species
included in this study were unattached and thus did not
allow for the determination of the morphological charac-
teristics of the basal region. However, observations on
other specimens (results not shown) indicated that U.
scandinavica lacks longitudinal ribs formed by closely
packed groups of cells. Additionally, no distinct margin
of vegetative cells lacking rhizoidal cells was found. The
cells from the lower middle/basal region were oval shaped
with rounded ends in TS (Fig. 12). While denticulations
were found on attached specimens (results not shown),
green-tide plants completely lacked denticulations. Ulva
scandinavica plants were membranous in texture (attached
plants were stiffer), and the thallus colour ranged from light
to mid-green.
The third species group was identified as the U. lactuca
group, U16, U19, U72, U125 (Fig. 4) and E102. This group
exhibited the greatest variation in morphological characters
420 Phycologia, Vol. 47 (4), 2008
of all species groups. Ulva lactuca usually has one to two
pyrenoids, but specimens with . 2 pyrenoids were not
uncommon (Fig. 13). The cells were round in shape or
polygonal with rounded corners and were generally
arranged in long rows or in short curved rows. The lengths
of the cells in the middle region varied between 12 and
22 mm, and the cell widths were between 15 and 24 mm. The
orientation of the chloroplast was mainly to the sidewall,
but it was sometimes observed covering the whole outer cell
wall. In TS the cells were oval with rounded apices
(Fig. 14). The basal region was characterised by having
longitudinal ribs of closely packed rhizoids, but these were
indistinct in some specimens. A distinct margin of
vegetative cells without rhizoidal cells was common;
although, this region was indistinct in some specimens.
While the majority of specimens lacked denticulations,
some had marginal microscopic protuberances. These were
distinguished from the larger denticulations on the basis
that they only contained two to four cells and were visible
only with the aid of a microscope. Thallus texture varied
from stiff to membranous, and coloration differed from
light to mid- to dark green.
Specimens provisionally identified as U. gigantea (U31,
U33 and U135; Fig. 8) were characterised by having only
one pyrenoid per cell. Ulva gigantea differs from the
morphologically similar U. pseudocurvata in possessing cells
that are polygonal with rounded corners and arranged in
long or short curved rows (Koeman & van den Hoek 1981;
Fig. 15). In the middle region of the thallus the cell lengths
varied from 8 to 19 mm, and the cell widths were between 8
and 21 mm. The chloroplast was always appressed to the
outer cell wall. Longitudinal ribs formed by closely packed
groups of rhizoids were evident, if not always clearly
distinct, and a margin of only vegetative cells was indistinct.
Cells from the lower middle/basal region appeared oval in
TS (Fig. 16). Microscopic denticulations were found in only
one specimen (U31). The thalli were membranous in texture
and ranged in colour from mid- to dark green.
Ulva rotundata specimens (CashA, U29, U34 and U41;
Fig. 5) could be readily identified in the field by the
presence of a strong metallic gloss on submerged thalli.
Cells possessed one, two or . 2 pyrenoids, were mostly
polygonal in shape with rounded corners and occurred
either in long distinct rows or without any order (Fig. 17).
Cells in the middle region varied from 16 to 31 mm in length
to 16 to 32 mm in width. The chloroplast was appressed to
the sidewall, giving a cap-like appearance. Longitudinal
ribs formed by groups of closely packed rhizoids and
margins characterised by a lack of rhizoidal cells were
indistinct or absent in most specimens. The majority of
specimens examined lacked denticulations, but microscopic
protuberances were found in the basal region of specimen
U29. The cells from the basal region appeared oval with flat
ends in cross section (Fig. 18). The thalli were membranous
in texture and ranged from light to mid-green in colour.
Mature thalli of Umbraulva olivascens were easy to
distinguish from Ulva spp. because of their strong olive-
green colour, which is particularly evident in the field.
Numerous specimens were identified, of which four were
used in the present study (U38, U46, U76 and U95; Fig. 6).
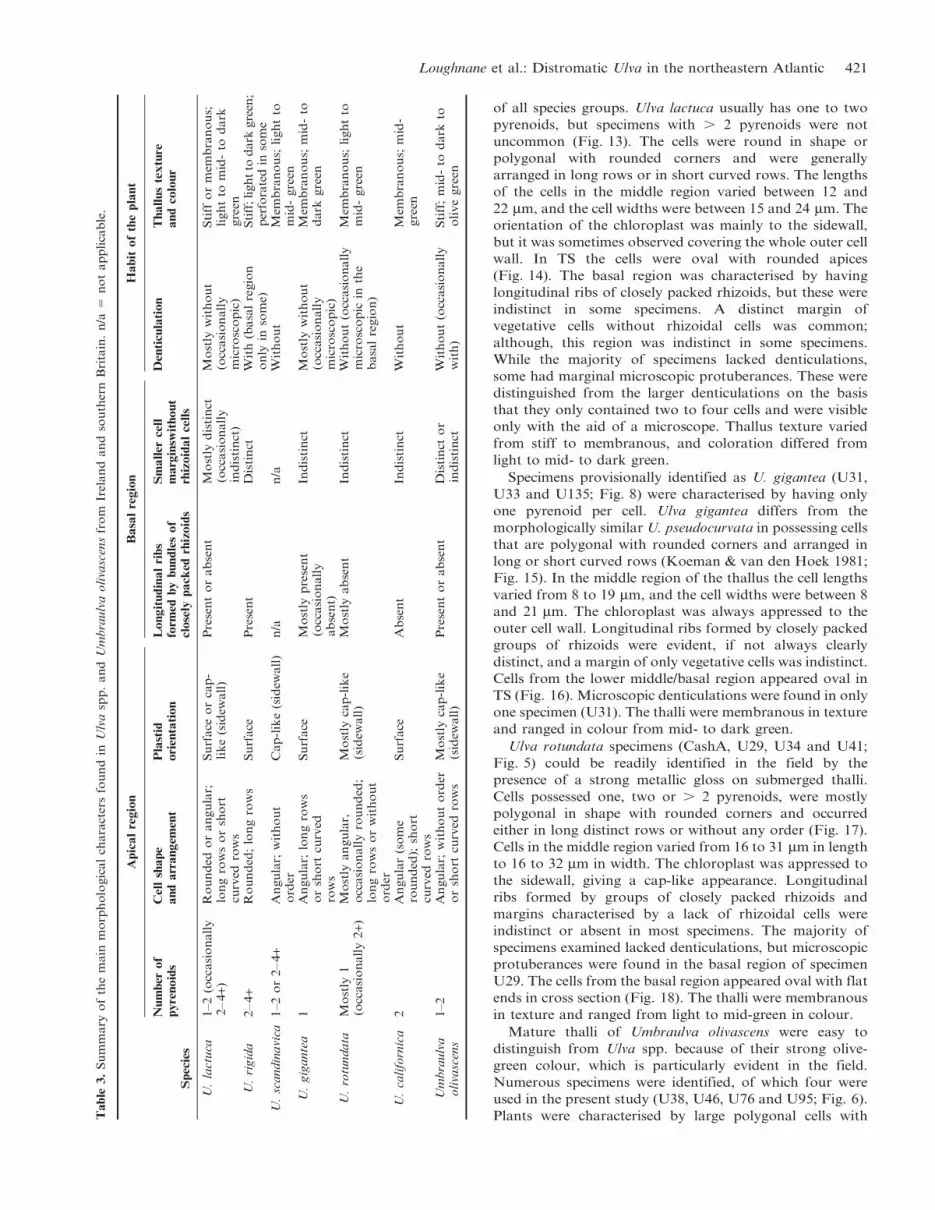
Plants were characterised by large polygonal cells withTable
3.
Su
mm
ary
of
the
main
mo
rph
olo
gic
al
chara
cter
sfo
un
din
Ulv
asp
p.
an
dU
mbra
ulv
aoli
vasc
ens
fro
mIr
elan
dan
dso
uth
ern
Bri
tain
.n
/a5
no
tap
pli
cab
le.
Apic
al
regio
nB
asa
lre
gio
nH
abit
of
the
pla
nt
Spec
ies
Num
ber
of
pyre
noid
sC
ell
shape
and
arr
angem
ent
Pla
stid
ori
enta
tion
Longit
udin
al
ribs
form
edby
bundle
sof
close
lypack
edrh
izoid
s
Sm
all
erce
llm
arg
insw
ithout
rhiz
oid
al
cell
s
Den
ticu
lati
on
Thall
us
tex
ture
and
colo
ur
U.
lactu
ca1–2
(occ
asi
on
all
y2–
4+)
Ro
un
ded
or
an
gu
lar;
lon
gro
ws
or
sho
rtcu
rved
row
s
Su
rface
or
cap
-li
ke
(sid
ewall
)P
rese
nt
or
ab
sen
tM
ost
lyd
isti
nct
(occ
asi
on
all
yin
dis
tin
ct)
Mo
stly
wit
ho
ut
(occ
asi
on
all
ym
icro
sco
pic
)
Sti
ffo
rm
emb
ran
ou
s;li
gh
tto
mid
-to
dark
gre
enU
.ri
gid
a2–
4+
Ro
un
ded
;lo
ng
row
sS
urf
ace
Pre
sen
tD
isti
nct
Wit
h(b
asa
lre
gio
no
nly
inso
me)
Sti
ff;li
gh
tto
dark
gre
en;
per
fora
ted
inso
me
U.
scandin
avi
ca1–2
or
2–
4+
An
gu
lar;
wit
ho
ut
ord
erC
ap
-lik
e(s
idew
all
)n
/an
/aW
ith
ou
tM
emb
ran
ou
s;li
gh
tto
mid
-gre
enU
.gig
ante
a1
An
gu
lar;
lon
gro
ws
or
sho
rtcu
rved
row
s
Su
rface
Mo
stly
pre
sen
t(o
ccasi
on
all
yab
sen
t)
Ind
isti
nct
Mo
stly
wit
ho
ut
(occ
asi
on
all
ym
icro
sco
pic
)
Mem
bra
no
us;
mid
-to
dark
gre
en
U.
rotu
ndata
Mo
stly
1(o
ccasi
on
all
y2+)
Mo
stly
an
gu
lar,
occ
asi
on
all
yro
un
ded
;lo
ng
row
so
rw
ith
ou
to
rder
Mo
stly
cap
-lik
e(s
idew
all
)M
ost
lyab
sen
tIn
dis
tin
ctW
ith
ou
t(o
ccasi
on
all
ym
icro
sco
pic
inth
eb
asa
lre
gio
n)
Mem
bra
no
us;
ligh
tto
mid
-gre
en
U.
cali
forn
ica
2A
ngu
lar
(so
me
rou
nd
ed);
sho
rtcu
rved
row
s
Su
rface
Ab
sen
tIn
dis
tin
ctW
ith
ou
tM
emb
ran
ou
s;m
id-
gre
en
Um
bra
ulv
aoli
vasc
ens
1–2
An
gu
lar;
wit
ho
ut
ord
ero
rsh
ort
curv
edro
ws
Mo
stly
cap
-lik
e(s
idew
all
)P
rese
nt
or
ab
sen
tD
isti
nct
or
ind
isti
nct
Wit
ho
ut
(occ
asi
on
all
yw
ith
)S
tiff
;m
id-
tod
ark
too
live
gre
en
Loughnane et al.: Distromatic Ulva in the northeastern Atlantic 421

rounded corners that had a cap-like chloroplast with one or
two pyrenoids (Fig. 19). The cells were arranged either in
short, curved rows or without any order. In the middle
region of the thallus, cell lengths varied between 13 and
26 mm, and the cell widths were between 8 and 21 mm. In
TS, the large cells were oval with rounded ends (Fig. 20).
The characteristics of the basal region were variable – with
or without longitudinal ribs formed by rows of closely
packed groups of rhizoids – and the margin devoid of
rhizoidal cells was either distinct or indistinct. The majority
of thalli lacked marginal denticulations. The thalli were stiff
in texture and predominantly olive-green or occasionally
dark green.
Plants from Dingle Harbour, Co. Kerry (see U157;
Fig. 7), could be distinguished from other putative species
groups, except U. lactuca, on the basis of the morphological
characteristics of the cells and thalli (Figs 21, 22). Cells
were polygonal with rounded corners (some cells were
round) and were arranged in short, curved rows. Cells
observed in the middle region of the thallus were 10–13 mm
in length and 10–18 mm in width. The chloroplast was
positioned to the outer cell wall and had no more than two
pyrenoids per cell. The basal region lacked any longitudinal
ribs formed by rows of closely packed rhizoids, and there
was no distinct margin of vegetative cells devoid of
rhizoidal cells. Thalli lacked marginal denticulations, were
membranous in texture and mid-green in colour.
Molecular analyses
Between 1272 and 1346 base pairs (bp) of rbcL sequences
were obtained from the material sequenced. Published data
obtained from GenBank (Table 1) were combined to give a
1346-bp sequence alignment for analyses. No insertions or
Figs 2–8. Habit of Ulva spp. Bars 5 1 cm.Fig. 2. U. rigida, Aran1, Inis Mor, Aran Islands, Co. Galway, GALW015515.Fig. 3. U. scandinavica, U168, Kilbrittan, Co. Cork, GALW015516.Fig. 4. U. lactuca, U125, Lulworth Cove, Dorset, England, GALW015511.Fig. 5. U. rotundata, CashA, Cashel Bay, Co. Galway, GALW015514.Fig. 6. Umbraulva olivascens, U76, East Ferry, Cobh Island, Co. Cork, GALW015528.Fig. 7. U. californica, U157, Dingle Harbour, Co. Kerry, GALW015517.Fig. 8. U. gigantea, U135, Port Isaac, Cornwall, England, GALW015520.
422 Phycologia, Vol. 47 (4), 2008

deletions were found in the in-group taxa; however, a 3-bp
insertion was found at position 229–232 in the out-group
taxa (Monostroma nitidum, Blidingia minima, Derbesia
marina and Bryopsis plumosa). Only unambiguously aligned
regions were used for the phylogenetic analyses. The
alignment contained 282 parsimony-informative sites and
911 invariable positions.
All analyses produced trees of generally congruent
topology. Maximum parsimony analysis produced a single
most parsimonious tree (length 5 724, consistency index 5
0.749, retention index 5 0.846, homoplasy index 5 0.251).
A single maximum likelihood tree was found (2lnL 5
5365.57198); this tree, with bootstrap values for NJ, MP
and the posterior probabilities estimated by Bayesian
inference overlaid on the nodes, is shown in Fig. 25.
In all analyses, the members of the genus Ulva resolved into
a monophyletic clade with strong bootstrap and Bayesian
support (BP 5 97–97%, BI 5 0.99). A well-supported
monophyletic clade (BP 5 100–100%, BI 5 1.00), contain-
ing specimens identified as Umbraulva olivascens, was sister
to all other in-group taxa. The out-group taxa formed two
well-supported separate clades (M. nitidium with B. minima,
and D. marina with B. plumosa). Sequence divergence
between the in-group Ulva taxa (except for Umbraulva)
and the out-group taxa was between 12.22% and 19.67%.
Within the in-group taxa (except for Umbraulva), sequence
divergence ranged from 0% to 0.56% for conspecific taxa
and from 0.67% to 4.45% between different lineages.
Within the large Ulva clade, eight different lineages were
resolved in the phylogenetic analyses. Their relative
Figs 9–24. Microscopic details of Ulva spp. Bars 5 30 mm.Fig. 9. Surface view through middle region of the thallus of U. rigida, Aran1, GALW015515.Fig. 10. Transverse section (TS) through middle region of the thallus of U. rigida, Aran1, GALW015515.Fig. 11. Surface view through middle region of the thallus of U. scandinavica, U168, GALW015516.Fig. 12. TS through middle region of the thallus of U. scandinavica, U168, GALW015516.Fig. 13. Surface view through middle region of the thallus of U. lactuca, U125, GALW015511.Fig. 14. TS through middle region of the thallus of U. lactuca, U125, GALW015511.Fig. 15. Surface view through middle region of the thallus of U. gigantea, U135, GALW015520.Fig. 16. TS through middle region of the thallus of U. gigantea, U135, GALW015520.Fig. 17. Surface view through middle region of the thallus of U. rotundata, CashA, GALW015514.Fig. 18. TS through middle region of the thallus of U. rotundata, CashA, GALW015514.Fig. 19. Surface view through middle region of the thallus of Umbraulva olivascens, U46, GALW015528.Fig. 20. TS through middle region of the thallus of Umbraulva olivascens, U46, GALW015528.Fig. 21. Surface view through middle region of the thallus of U. californica, U157, GALW015517.Fig. 22. TS through middle region of the thallus of U. californica, U157, GALW015517.Fig. 23. Surface view through middle region of the monostromatic thallus of U. compressa, CashB, GALW015526.Fig. 24. TS through middle region of the monostromatic thallus of U. compressa, CashB, GALW015526.
Loughnane et al.: Distromatic Ulva in the northeastern Atlantic 423

positions were not clearly resolved, as several nodes in the
internal parts of the tree had very low support. Individually,
however, the eight lineages formed well-supported mono-
phyletic groups with high bootstrap values and posterior
probabilities, and their composition was identical in
different inference methods. In general, they showed good
correspondence with species defined on morphological
features; for this reason, herein we refer to them with the
names of the morphological species to which they
correspond.
A subclade containing three species groups, U. gigantea,
U. fasciata and a group containing U. rigida, U. scandina-
vica and U. armoricana, was resolved with low bootstrap
support (BP 5 55–64%, BI 5 0.98). Both the U. gigantea
and the U. fasciata clade were strongly supported (BP 5
99–100%, BI 5 1.00), and the clade containing U. rigida/U.
scandinavica/U. armoricana was resolved with moderate
support (BP 5 70–75%, BI , 0.5). Sequence divergence
within the U. gigantea group was 0.39–0.56%, U. fasciata
was 0% and U. rigida/U. scandinavica/U. armoricana clade
ranged from 0% to 0.22%.
A sister clade containing specimens identified as U.
californica and U. curvata with strong bootstrap support
(BP 5 100–100%, BI 5 1.00) was separated basally from
the U. rigida and U. fasciata clades with weak bootstrap
support (BP 5 , 50–52%, BI 5 0.98). Specimen U157 from
Dingle Harbour, Co. Kerry, was conspecific with U.
californica from the type locality in California, USA (0%
divergence), and both sequences are identical to a specimen
identified as U. curvata from Delaware, USA.
A second subclade contained the U. pseudocurvata and
U. compressa species group with strong bootstrap support
(BP 5 100%, BI 5 1.00). GenBank specimens identified as
U. intestinalis formed a strongly supported species group
(BP 5 100–100%, BI 5 1.00). Low bootstrap support was
found for a clade containing the U. pseudocurvata/U.
compressa group and U. intestinalis (BP 5 , 50–56%, BI
5 0.98). Sequence divergence within the U. pseudocurvata/
U. compressa group ranged from 0% to 0.15%.
A third subclade containing the U. lactuca species group
was resolved with strong bootstrap support (BP 5 100–
100%, BI 5 1.00), and sequence divergence ranged from 0–
0.37%. Ulva fenestrata from Washington, USA, was also
found in this subclade with strong support, and sequence
divergence from other taxa in this group ranged from 0% to
0.29%.
A fourth subclade with strong bootstrap support (BP 5
100–100%, BI 5 1.00) contained the U. rotundata species
group with sequence divergence ranging from 0% to 0.6%.
DISCUSSION
The combined approach of traditional morphological and
cytological techniques of identification and molecular
phylogenetic analyses has revealed at least six distinct
lineages of distromatic Ulva in Ireland and southern
Britain. On morphological grounds, we conclude that four
represent previously unreported species: U. scandinavica
(Ireland only), U. gigantea (Ireland and Britain), U.
rotundata (Ireland only) and U. californica (Ireland only).
It can be expected that many of these entities reported only
for Ireland occur in Britain also.
The arrangement of species within Ulva as inferred from
rbcL sequence data generally concurs with that of previous
investigations (Hayden & Waaland 2002, 2004; Hayden et
al. 2003; Shimada et al. 2003). Sequence divergence between
conspecifics (0–0.56%) and in-group taxa ranged between
0% and 4.45%. This is consistent with other studies on Ulva
using rbcL sequence data where the range of sequence
divergence amongst conspecific taxa was between 0% and
0.4% (Hayden et al. 2003; Hayden & Waaland 2004) and up
to 4.9% between in-group taxa (Hayden & Waaland 2004).
Morphological characters considered typical of U. rigida
by other authors (e.g., Bliding 1963; Koemann & van den
Hoek 1981; Hoeksema & van den Hoek 1983; Dion et al.
1998) were found to be consistently useful in identifying this
species in Ireland and Britain. Bliding (1963) reported U.
rigida as the most common Ulva species on Mediterranean
Fig. 25. Maximum likelihood tree with bootstrap proportions formaximum parsimony (top) and neighbour joining (middle) andposterior probabilities from Bayesian analysis (bottom) are given atthe nodes. Bootstrap support less than 50% and posteriorprobabilities less than 0.95 are not given, and branch lengths aredrawn proportional to sequence change.
424 Phycologia, Vol. 47 (4), 2008

and Atlantic coasts. Material referred by Bliding (1963) to
this species was characterized by rectangular or polygonal
cells, with slightly rounded corners, arranged in longitudi-
nal and occasionally transversal rows in most of the thallus.
In adult plants, the dimensions of cells in surface view
ranged from 15 to 22 3 11 to 17 mm. Elongated cells with
tapered ends towards the surface, as viewed in TS, were
deemed characteristic of the species by Bliding (1963).
Indeed, specimens were readily identified by the presence of
the spindle-shaped cells with tapered ends in the basal and
lower middle region from France (Hoeksema & van den
Hoek 1983; Dion et al. 1998) and the Netherlands
(Koemann & van den Hoek 1981). Plants examined in this
study were identified as U. rigida sensu Bliding primarily on
the basis of this anatomical character. However, this
character is not consistent with the type specimen as
described by C. Agardh (1822) and redescribed by J.
Agardh (1883) (see Papenfuss 1960; Phillips 1988). Cells
with tapered or narrowed ends in TS are distinctive of U.
laetevirens Areschoug 1854, a common species found in
Australia, New Zealand, southern California, the Black Sea
and the Mediterranean (Phillips 1988; Guiry & Guiry 2007).
In a study on Australian Ulva, Phillips (1988) examined
both the lectotype of U. rigida C. Agardh and the holotype
of U. rigida var. fimbriata J. Agardh, which was subse-
quently reduced to synonymy with U. laetevirens. Applying
the anatomical character of spindle-shaped or tapered cells
in the basal region, Phillips noted that specimens recorded
and described by Bliding (1968) were misidentified, and she
concluded that U. rigida sensu Bliding should in fact be
referred to U. laetevirens. Since Bliding’s work, many
investigations conducted in Europe have recorded the
presence of U. rigida sensu Bliding, including the present
study. Further work, perhaps incorporating sequences
obtained from the lectotype specimen of U. rigida and
from Bliding’s herbarium material, is needed for a definitive
clarification of this taxonomic problem.
Two specimens identified as U. scandinavica sampled
from green tides were used in this investigation: U111 from
Langstone Harbour, UK, and U168 from Kilbrittan, Co.
Cork. Ulva scandinavica from Cork represents the first
report for Ireland, although it has been recorded previously
for Britain (Tan et al. 1999). Morphological analyses and
subsequent grouping by phylogenetic methods revealed
inconsistencies in characters used to identify this species.
The pyrenoid number varied from one per cell in U111 to .
2 in U168. Malta et al. (1999) found that the number of
pyrenoids varied not only spatially but also temporally.
Plants identified as U. scandinavica in the Netherlands were
found to have . 1 pyrenoid in 50% of cells in May, while at
the end of the growing season all the cells had one pyrenoid.
Specimens from the Netherlands, France and the Adriatic
had . 2 pyrenoids in more than 50% of cells (Koeman &
van den Hoek 1981; Hoeksema & van den Hoek 1983;
Battelli & Tan 1998). Ulva scandinavica is separated from
U. rigida sensu Bliding and U. armoricana by the absence of
spindle-shaped cells in TS from the basal region and from
other Ulva species by the absence of longitudinal ribs
formed by rows of closely packed bundles of rhizoidal cells
(Coat et al. 1998; Dion et al. 1998). Cell shape in the apical
and middle regions of the thallus in surface view
distinguishes U. armoricana (polygonal cells) from U. rigida
(rounded cells) (Coat et al. 1998).
Sequence divergence between samples identified as both
U. rigida sensu Bliding, U. scandinavica and GenBank
sequences for U. armoricana was within the range of
conspecificity, namely, 0–0.22% (e.g. U. lactuca ranges from
0% to 0.37%). This is in agreement with other studies using
rbcL data, which found that rbcL sequence divergence
between U. rigida and U. scandinavica samples was about
0.2% (Hayden & Waaland 2004). These authors remarked
that the morphological and anatomical variation observed
in these three species was not diagnostic, and they
speculated that further study may show U. armoricana
and U. scandinavica to be conspecific with U. rigida sensu
Bliding. However, the taxonomic arrangement of species in
this group will remain unresolved without full consideration
of the status of U. rigida C. Agardh and U. laetevirens
Areschoug, which, as stated previously, should include
comprehensive morphological and molecular examination
of both type specimens.
Ulva californica was originally described from La Jolla,
California, by Wille (1889). Tanner (1979) concluded that
the ubiquitous and morphologically variable entity com-
monly called U. californica is a distinct entity at the species
rank, occupying a wide range of habitats with a large
variation in thallus shape and size. Based on developmental
and morphological parallels, he placed into synonymy with
U. californica two other taxa: U. angusta Setchell & N.L.
Gardner and U. scagelii Chihara (Tanner 1986). This
conclusion was subsequently corroborated by phylogenetic
analyses based on ITS and rbcL sequences (Hayden &
Waaland 2004). Ulva californica was considered to be
restricted to the Pacific coast of North America (Scagel et
al. 1989), but Hayden & Waaland (2004), using a
combination of ITS and rbcL data, confirmed that U.
californica was present in Scotland (U. californica EU as
Ulva sp. in Tan et al. 1999); it has also been reported from
North Senegal on the west coast of Africa (John et al.
2003); although, this latter record is somewhat uncertain. A
GenBank sequence from Delaware, USA (AF189071;
Sherwood et al. 2000), identified as U. curvata was also
found to be conspecific with U. californica from California
and Co. Kerry (BP 5 100%, BI 5 1.00) with 0% sequence
divergence. Ulva curvata, a North Atlantic species found in
the Mediterranean and from Morocco to the Baltic as well
as eastern North America (Guiry & Guiry 2007), is
characterised by a hollow thallus immediately above the
holdfast (Bliding 1968; Koemann & van den Hoek 1981).
This important and distinctive morphological feature had
not been fully assessed (Sherwood et al. 2000), and it seems
likely, based on the phylogenetic evidence presented here,
that U. curvata from Delaware as deposited in GenBank
(AF189071) represents a misidentification. A specimen
from Dingle Harbour, Co. Kerry (Fig. 7), was also
conspecific with U. californica from the type locality with
strong bootstrap support (BP 5 100%, BI 5 1.00) and 0%
sequence divergence. While this is not the first record of U.
californica from Europe (Tan et al. 1999; Hayden &
Waaland 2004), it is a new record for Ireland. It may have
been overlooked or misidentified in the past because of its
broad range of habit, form and morphological and
Loughnane et al.: Distromatic Ulva in the northeastern Atlantic 425

cytological characteristics (Tanner 1979, 1986). Ulva
californica from Co. Kerry was linear to oblanceolate,
30 cm in length, membranous with strongly ruffled
margins. Cells in surface view were angular, occurring in
short curved rows, and the chloroplast with one to two
pyrenoids covered the entire surface wall. The basal region
and stipe lacked a central cavity that is characteristic of U.
curvata (Koemann & van den Hoek 1981; Hoeksema & van
den Hoek 1983). This description closely matches the
description of U. angusta, a putative synonym of U.
californica (Abbott & Hollenberg 1976). On the Pacific
coast of North America, U. lactuca often is misidentified as
the more common U. californica as the definitive characters
that separate these entities are developmental (Hayden &
Waaland 2004). It would not be unreasonable to assume
that the converse situation could occur in Europe, that is,
that specimens of U. californica could be misidentified as U.
lactuca (see Hardy & Guiry 2003), and as such this species
could be more common.
Ulva gigantea is notoriously difficult to separate from U.
pseudocurvata on morphological grounds alone (B. de
Reviers, Museum National d’Histoire Naturelle, Paris,
France, pers. comm.). While both species share many
morphological characteristics, they do differ in the arrange-
ment of the cells, in surface view, in the middle and basal
regions. Bliding (1968) described polygonal cells with
rounded corners arranged in long rows or in short curved
rows in U. gigantea. Ulva pseudocurvata was described as a
new species by Koeman & van den Hoek (1981) from the
Netherlands on the basis of the different arrangement of the
cells in surface view (distinctly polygonal without rounded
corners, forming indistinct groups or without order). Ulva
gigantea is currently known only from Greece and the
Atlantic coast of France (Guiry & Guiry 2007), and
specimens in this study represent the first records for
Ireland and Britain. No rbcL gene sequences have been
published for U. gigantea, a situation that does not allow
for comparisons with sequences obtained here.
Ulva compressa (formerly E. compressa), with a tubular
monostromatic structure, is a common intertidal alga in
Ireland and Britain. It is morphologically similar to U.
intestinalis; however, previous studies using crossing
experiments (Larsen 1981), and phylogenetic analysis of
ITS nrDNA sequences (Blomster et al. 1998) and rbcL
sequences (Hayden et al. 2003) have shown them to be
distinct species. The present study mirrors these conclu-
sions, viz., that U. compressa and U. intestinalis are distinct
species, with sequence divergence of 1.73–1.94%, well above
the limits of conspecific taxa. Ulva compressa from Galway
(Figs 23, 24) resolved into a monophyletic clade with U.
pseudocurvata from Scotland and Washington State, USA,
with strong bootstrap support (BP 5 100%, BI 5 1.00).
Sequence divergence, of between 0% and 0.15%, supports
the hypothesis that these two entities are conspecific. Some
morphological synapomorphies define this clade: both taxa
have polygonal cells that form indistinct groups in the
middle and apical region, and the chloroplast is cap-like in
appearance (appressed to the side wall of the cell in surface
view), containing one pyrenoid per cell (Koeman & van den
Hoek 1981, 1982). However, U. compressa has a mono-
stromatic, tubular, variously branched habit, and U.
pseudocurvata is distromatic and often expanded in habit.
Tan et al. (1999) suggested that a developmental switch may
exist in some members of Ulva and that this genetic
encoding allows the plant to switch morphologies between
monostromatic tubular and distromatic blades. They also
noted that it is most probable that a combination of
environmental prompts (salinity gradient, presence or
absence of bacterial types) may activate the nonreversible
switch between morphological forms from one generation
to the next. Using ITS nrDNA data, these authors also
found that sequence divergence between the partially
distromatic Ulva linza (as E. linza) and the highly branched
monostromatic tubular Ulva procera (as E. procera) were
also within the range of conspecifics. Blomster et al. (2002)
examined green tides on the Baltic coast of Finland and
found that monostromatic tubular U. intestinalis (as E.
intestinalis) formed vast proliferations of monostromatic
sheets that had previously been misidentified as Mono-
stroma. Morphological plasticity is not unknown within the
Ulvales: formation of branches in unbranched U. intesti-
nalis (Reed & Russell 1978), formation of second-genera-
tion tubular plants in axenic culture from first-generation
distromatic plants (Gayral & Lepailleur 1968; Bonneau
1977; Marshall et al. 2006) and variable bacterial flora can
induce the formation of tubular progeny from distromatic
Ulva (Provasoli & Pintner 1980). This plasticity in form
may lead to the description of various species when, in fact,
they are not phylogenetically distinct entities.
The generitype (lectotype) species of Ulva Linnaeus is U.
lactuca, which is presumed to be a common cosmopolitan
species, known from the Arctic Ocean, North and South
Atlantic, Indian Ocean, and the Pacific Rim (Guiry &
Guiry 2007). Bliding (1968) described U. lactuca as having
polygonal or rounded cells that form slightly curved rows
and have one pyrenoid. Koeman & van den Hoek (1981)
agreed with this diagnosis yet did not mention round cells in
surface view but did catalogue a diagnostic feature of newly
divided cells in the apical region having cap-like chloro-
plasts that lie opposite each other. This feature is absent in
older cells in the middle region where the chloroplast covers
the surface of most of the cells. Pyrenoid number, the shape
of the cells in surface view, plastid orientation, denticula-
tion, presence or absence of longitudinal ribs formed by
rows of closely packed rhizoids and distinct or indistinct
margins of vegetative cells devoid of rhizoids were all
variable characteristics in Irish and British populations of
U. lactuca.
Plasticity of morphological characters in the Ulvales is
not uncommon and is often attributed to variable
environmental conditions (Titylanov et al. 1975; Tanner
1979, 1986; Malta et al. 1999). Pyrenoid number was
considered by some workers to be a useful taxonomic
character (Bliding 1968; Koeman & van den Hoek 1981;
Hoeksema & van den Hoek 1983; Dion et al. 1998) but
unreliable and too variable by others (Tanner 1979, 1986).
The arrangement of the cells in surface view was considered
by Bliding (1968) to be characteristic for a species, but
others (e.g. Hoeksema & van den Hoek 1983) deemed it to
be too variable to be used alone. Chloroplast position,
again considered to be a useful character by many authors,
often in conjunction with other characters (Koeman & van
426 Phycologia, Vol. 47 (4), 2008

den Hoek 1981; Hoeksema & van den Hoek 1983; Malta et
al. 1999), has also been demonstrated to alter position due
to photon irradiance levels (Titlyanov et al. 1975), a
circadian rhythm (Britz & Briggs, 1976) or in recently
divided cells (Løvlie 1964). It is thus not surprising to find
considerable variation in characters that were traditionally
used to delineate U. lactuca. In this study, anatomical
features of the basal region were also found to be highly
variable. Indeed, resolution of this taxon was not conclusive
without the use of molecular phylogenetic-based methods.
A specimen originally identified as U. fenestrata
(AF49966; Hayden et al. 2003) but redesignated U. lactuca
by Hayden & Waaland (2004) was conspecific with Irish,
English and Alaskan U. lactuca (0–0.29% divergence).
There is some confusion over U. fenestrata, as it is
morphologically similar to U. lactuca (Hayden et al. 2003;
Hayden & Waaland 2004). Tanner (1979) concluded that it
was a separate entity from U. lactuca based on crossing
experiments. He concluded that a delay in fusion and a
reduced number of fused gametes was a negative response.
Evidence presented here corroborates other studies (Hay-
den & Waaland 2002, 2004; Hayden et al. 2003) that
consider U. fenestrata as conspecific with U. lactuca.
Bliding (1968) originally described U. rotundata from
specimens he had earlier collected (1958) from Naples,
Italy. In his survey of European Ulva, he subsequently
discovered that U. rotundata was common in the Canary
Islands and in Norway. It is now considered a ubiquitous
North Atlantic and Mediterranean species found from
West Africa to Scandinavia and the eastern United States
(Guiry & Guiry 2007). The main diagnostic feature of this
taxon is the presence of a strong metallic gloss in the field
and the presence of relatively large cells in the middle region
(Hoeksema & van den Hoek 1983; B. de Reviers, personal
communication). Ulva rotundata from Ireland corresponds
well with French specimens (Hoeksema & van den Hoek
1983), with large polygonal or angular cells arranged in
long rows or without order with the plastids mostly
oriented to the sidewall. In contrast to French specimens,
Irish specimens generally had only a single pyrenoid per
cell, and one specimen had microscopic denticules in the
basal region. Ulva rotundata from Galway and Clare are the
first records of this species from Ireland.
Umbraulva olivascens (synonym Ulva olivascens) is the
only member of the genus occurring in Europe (Guiry &
Guiry 2007) and was originally known from only three sites
in Ireland (Hardy & Guiry 2003). Umbraulva forms a sister
clade to Ulva with strong bootstrap support (Fig. 25) and is
not monophyletic with Ulva (Bae & Lee 2001). Siphonax-
anthin, an accessory pigment, is found in Umbraulva and
not in Ulva, giving members of this genus its distinctive
olive-green colour (Bae & Lee 2001). It was assumed that
the paucity of records for this taxon in Ireland was due to
its resemblance to U. lactuca (Hardy & Guiry 2003),
particularly in immature thalli (C.J. Loughnane, personal
observations). The present study has confirmed that its
distribution is much wider than previously recorded, with
specimens identified from counties Wexford, Cork, Kerry,
Clare, Galway and Mayo. When mature, olive-green thalli
of U. olivascens are very distinctive and easily identifiable in
the field. Our rbcL analyses confirm with strong bootstrap
support (BP 5 100%; BI 5 1.00) that Umbraulva does not
form a monophyletic group with Ulva and is a separate
genus (Bae & Lee 2001). Recent evidence suggests that the
previously unreported Pacific endemic U. pertusa Kjellman
was recently introduced to the northwestern coast of
Iberian Peninsula (Baamonde Lopez et al. 2007). While
comprehensive and appropriate analyses of available
molecular data are required to clarify this record, it does
seem likely that the number of species within the genus Ulva
recorded in European waters will continue to grow.
To date, this study is the largest and most comprehensive
investigation of the distromatic members of the genus Ulva
in Ireland and southern Britain. The results indicate that
the use of traditional morphological techniques in the
identification of species in this genus is often inconclusive.
Previous unrecorded taxa in Ireland were possibly over-
looked due to the variability and unreliability of accepted
morphological characters, and without the use of sequence
data, some species groups would remain misidentified. Yet,
without morphological assessment (i.e. spindle-shaped cells
in TS), the delimitation of U. rigida sensu Bliding from U.
scandinavica would have proved difficult. Thus, the
concomitant use of a morphological- and molecular-based
approach to species delimitation and identification in this
genus is recommended. Additions to the Irish marine flora
include U. scandinavica, U. gigantea, U. rotundata and U.
californica and expanded distributional records of Um-
braulva olivascens.
ACKNOWLEDGEMENTS
We would like to thank Robert Wilkes, Ana Felga, Bruno
de Reviers, Christine Maggs and Stefan Kraan for
collecting samples, providing sequences and miscellaneous
data and general discussions. The study was funded by the
Higher Education Authority’s PRTLI Cycle 2 funding for
the Environmental Change Institute, National University
of Ireland, Galway, Ireland.
REFERENCES
ABBOTT I.A. & HOLLENBERG G.J. 1976. Marine algae of California.827 pp. Stanford University Press, Stanford, California, USA.
AGARDH C.A. 1822. Species algarum rite cognitae, cum synonymis,differentiis specificis et descriptionibus succinctis. Berling, Lund.Vol. 1, part 2, fasc. 1, pp. 169–398.
AGARDH J.G. 1883. Till algernes systematik. Nya bidrag. (Tredjeafdelningen.) Lunds Universitets Ars-Skrift, Afdelningen forMathematik och Naturvetenskap 19(2): 1–177, 4 plates.
ARESCHOUG J.E. 1854. Phyceae novae et minus cognitae in maribusextraeuropaeis collectae. Nova Acta Regiae Societatis Scien-tiarum Upsaliensis, Ser. 3, 1: 329–372.
BAAMONDE LOPEZ S., BASPINO FERNANDEZ I., BARREIRO LOZANO
R. & CREMADES UGARTE J. 2007. Is the cryptic alien seaweedUlva pertusa (Ulvales, Chlorophyta) widely distributed alongEuropean Atlantic coasts? Botanica Marina 50: 267–274.
BAE E.H. & LEE I.K. 2001. Umbraulva, a new genus based on Ulvajaponica (Holmes) Papenfuss (Ulvaceae, Chlorophyta). Algae 16:217–231.
BATTELLI C. & TAN I.H. 1998. Ulva scandinavica Bliding,(Chlorophyta): a new species for the Adriatic Sea. Annales:Annals for Istran and Mediterranean Studies 13: 121–124.
Loughnane et al.: Distromatic Ulva in the northeastern Atlantic 427

BLIDING C. 1963. A critical survey of European taxa in Ulvales.Part I. Capsosiphon, Percursaria, Blidingia, Enteromorpha. OperaBotanica 8(3): 1–160.
BLIDING C. 1968. A critical survey of European taxa in Ulvales. II.Ulva, Ulvaria, Monostroma, Kornmannia. Botaniska Notisker121: 535–629.
BLOMSTER J., MAGGS C.A. & STANHOPE M.J. 1998. Molecular andmorphological analysis of Enteromorpha intestinalis and E.compressa (Chlorophyta) in the British Isles. Journal ofPhycology 34: 319–340.
BLOMSTER J., MAGGS C.A. & STANHOPE M.J. 1999. Extensiveintraspecific morphological variation in Enteromorpha muscoides(Chlorophyta) revealed in molecular analysis. Journal ofPhycology 35: 575–586.
BLOMSTER J., BACK S., FEWER D.P., KIIRIKKI M., LEHVO A.,MAGGS C.A. & STANHOPE M.J. 2002. Novel morphology inEnteromorpha (Ulvophyceae) forming green tides. AmericanJournal of Botany 89: 1756–1763.
BONNEAU E.R. 1977. Polymorphic behaviour of Ulva lactuca(Chlorophyta) in axenic culture. I. Occurrence of Enteromorpha-like plants in haploid clones. Journal of Phycology 13: 133–140.
BRITZ S.J. & BRIGGS W.R. 1976. Circadian rhythms of chloroplastorientation and photosynthetic capacity in Ulva. Plant Physiol-ogy 58: 22–27.
BURROWS E.M. 1991. Seaweeds of the British Isles. Volume 2.Chlorophyta. Natural History Museum Publications, London.238 pp.
COAT G., DION P., NOAILLES M.-C., DE REVIERS, B., FONTAINE J.-M., BERGER-PERROT Y. & LOISEAUX-DE GOER, S. 1998. Ulvaarmoricana (Ulvales, Chlorophyta) from the coasts of Brittany(France). II. Nuclear rDNA ITS sequence analysis. EuropeanJournal of Phycology 33: 81–86.
DE TONI G.B. 1889. Sylloge algarum omnium hucusque cognitarum.Vol. I. Chlorophyceae. Sumptibus auctoris, Patavii [Padua], 1315pp.
DION P., DE REVIERS, B. & COAT G. 1998. Ulva armoricana sp. nov.(Ulvales, Chlorophyta) from the coasts of Brittany (France). I.Morphological identification. European Journal of Phycology 33:73–80.
DOYLE J.J. & DOYLE J.L. 1987. A rapid DNA isolation procedurefor small quantities of fresh leaf tissue. Phytochemical Bulletin 19:11–15.
FELSENSTEIN J. 1995. Confidence limits on phylogenies: anapproach using the bootstrap. Evolution 39: 783–791.
GAYRAL P. & LEPAILLEUR H. 1968. Une rare Chlorophyceeunicellulaire marine: Heterogonium salinum P.A. Dangeard.Revue Algologique 9: 131–134.
GUIRY M.D. & GUIRY G.M. 2007. AlgaeBase version 4.2. World-wide electronic publication, National University of Ireland,Galway. Available at: http://www.algaebase.org (Accessed 2 May2007).
HALL B.G. 2001. Phylogenetic trees made easy: a how-to manual formolecular biologists. Sinauer Associates, Sunderland, MA, USA.179 pp.
HARDY F.G. & GUIRY M.D. 2003. A Check-list and Atlas of theSeaweeds of Britain and Ireland. British Phycological Society,London. 435 pp.
HAYDEN H.S. & WAALAND J.R. 2002. Phylogenetic systematics ofthe Ulvaceae (Ulvales, Ulvophyceae) using chloroplast andnuclear DNA sequences. Journal of Phycology 38: 1200–1212.
HAYDEN H.S. & WAALAND J.R. 2004. A molecular systematicstudy of Ulva (Ulvaceae, Ulvales) from the northeast Pacific.Phycologia 43: 364–382.
HAYDEN H.S., BLOMSTER J., MAGGS C.A., SILVA P.C., STANHOPE
M.J. & WAALAND J.R. 2003. Linnaeus was right all along: Ulvaand Enteromorpha are not distinct genera. European Journal ofPhycology 38: 277–294.
HOEKSEMA B.W. & VAN DEN HOEK, C. 1983. The taxonomy ofUlva (Chlorophyceae) from the coastal region of Roscoff(Brittany, France). Botanica Marina 26: 65–86.
JOHN D.M., LAWSON G.W. & AMEKA G.K. 2003. The marinemacroalgae of the Tropical West Africa Subregion. Beihefte zurNova Hedwigia 125: 1–217.
KOEMAN R.P.T. & VAN DEN HOEK, C. 1981. The taxonomy of Ulva(Chlorophyceae) in the Netherlands. British Phycological Journal16: 9–53.
KOEMAN R.P.T. & VAN DEN HOEK, C. 1982. The taxonomy ofEnteromorpha Link, 1820, (Chlorophyceae) in the Netherlands. I.The section Enteromorpha. Archiv fur Hydrobiologie, Supplement-band 63(Algological Studies 32): 279–330.
LARSEN J. 1981. Crossing experiments with Enteromorpha intesti-nalis and E. compressa from different European localities. NordicJournal of Botany 1: 128–136.
LINNAEUS C. 1753. Species plantarum. Stockholm. 1200 pp.LØVLIE A. 1964. Genetic control of division rate and morphogen-
esis in Ulva mutabilis Føyn. Comptes Rendus des Travaux duLaboratoire Carlsberg 34: 77–168.
MADDISON D.R. & MADDISON W.P. 2002. MacClade 4: analysis ofphylogeny and character evolution, Sinauer Associates, Sunder-land, MA, USA.
MALTA E.-J., DRAISMA S.G.A. & KAMERMANS P. 1999. Free-floating Ulva in the southwest Netherlands: species or morpho-types? A morphological, molecular and ecological comparison.European Journal of Phycology 34: 443–454.
MANHART J. 1994. Phylogenetic analysis of green plant rbcLsequences. Molecular Phylogeny and Evolution 3: 114–127.
MARSHALL K., JOINT I., CALLOW M. & CALLOW J. 2006. Effect ofmarine bacterial isolates on the growth and morphology ofaxenic plantlets of the green alga Ulva linza. Microbial Ecology52: 302–310.
MEDLIN L.K., MEFTIES K., JOHN U. & OLSEN J.L. 2007. Algalmolecular systematics: a review of the past and prospects for thefuture. In: Unravelling the algae: the past, present and future ofalgal systematics (Ed. by J. Brodie & J. Lewis), SystematicsAssociation Special Volume Series 75. CRC Press, Boca Raton,FL, USA. pp. 341–353.
PAPENFUSS G.F. 1960. On the genera of the Ulvales and the statusof the order. Botanical Journal of the Linnaean Society 56:303–318.
PHILLIPS J.A. 1988. Field, anatomical and developmental studieson southern Australian species of Ulva (Ulvaceae, Chlorophyta).Australian Systematic Botany 1: 411–456.
POSADA D. & CRANDALL K.A. 1998. Modeltest: testing the modelof DNA substitution. Bioinformatics 14: 817–818.
PROVASOLI L. & PINTNER I.J. 1980. Bacteria induced polymor-phism in an axenic laboratory strain of Ulva lactuca (Chlor-ophyceae). Journal of Phycology 16: 196–201.
REED R.H. & RUSSELL G. 1978. Salinity fluctuations and theirinfluence on ‘‘bottle brush’’ morphogenesis in Enteromorphaintestinalis (L.) Link. British Phycological Journal 13: 149–153.
RONQUIST F. & HUELSENBECK J.P. 2003. MRBAYES 3: Bayesianphylogenetic inference under mixed models. Bioinformatics 19:1572–1574.
SCAGEL R.F., GABRIELSON P.W., GARBARY D.J., GOLDEN L.,HAWKES M.W., LINDSTROM S.C., OLIVEIRA J.C. & WIDDOWSON
T.B. 1989. A synopsis of the benthic marine algae of BritishColumbia, southeast Alaska, Washington and Oregon. Phycolo-gical Contributions [University of British Columbia] 3: vi + 532 pp.
SETCHELL W.A. & GARDNER N.L. 1920. Phycological contribu-tions. Part I. University of California Publications in Botany 7:279–324.
SHERWOOD A.R., GARBARY D.J. & SHEATH R.G. 2000. Assessingthe phylogenetic position of the Prasiolales (Chlorophyta) usingrbcL and 18S rRNA gene sequence data. Phycologia 39: 139–146.
SHIMADA S., HIRAOKA M., NABATA S., LIMA M. & MASUDA M.2003. Molecular phylogenetic analyses of the Japanese Ulva andEnteromorpha (Ulvales, Ulvophyceae), with special reference tothe free-floating Ulva. Phycological Research 51: 99–108.
SWOFFORD D.L. 2003. PAUP*: phylogenetic analysis usingparsimony (*and other methods), Sinauer Associates, Sunder-land, MA, USA.
TAN I.H., BLOMSTER J., HANSEN G., LESKINEN E., MAGGS C.A.,MANN A., SLUIMAN H.J. & STANHOPE M.J. 1999. Molecularphylogenetic evidence for a reversible morphogenetic switchcontrolling the gross morphology of two common genera ofgreen seaweeds, Ulva and Enteromorpha. Molecular Biology andEvolution 16: 1011–1018.
428 Phycologia, Vol. 47 (4), 2008

TANNER C.E. 1979. The taxonomy and morphological variation ofdistromatic Ulvaceous algae (Chlorophyta) from the northeastPacific. PhD thesis. University of British Columbia, Vancouver.249 pp.
TANNER C.E. 1986. Investigations of the taxonomy and morpho-logical variation of Ulva (Chlorophyta): Ulva californica Wille.Phycologia 25: 510–520.
TITLYANOV E.A., GEEBOVA N.T. & KOTLYAROVA L.S. 1975.Seasonal changes in structure of the thalli of Ulva fenestrata P.et R. Ekologiya 6: 320–324.
WILLE N. 1889. In: Phycotheca Boreali-Americana (F.S. Collins, I.Holden & W.A. Setchell). Malden, MA, USA. Fasicle XIII, no.611.
WOMERSLEY H.B.S. 1984. The marine benthic flora of southernAustralia. Part I. Government Printer, South Australia, Ade-laide, Australia. 329 pp.
Received 1 August 2007; accepted 26 March 2008
Associate editor: Paul Gabrielson
Loughnane et al.: Distromatic Ulva in the northeastern Atlantic 429






























