



This article was downloaded by:[CSIC]On: 28 November 2007Access Details: [subscription number 770351778]Publisher: Informa HealthcareInforma Ltd Registered in England and Wales Registered Number: 1072954Registered office: Mortimer House, 37-41 Mortimer Street, London W1T 3JH, UK
Biocatalysis and BiotransformationPublication details, including instructions for authors and subscription information:http://www.informaworld.com/smpp/title~content=t713454445
Microscopy studies reveal delignification and sterolremoval from eucalypt kraft pulps by laccase-HBTM. Speranza ab; D. Ibarra b; J. Romero a; A. T. Martínez b; M. J. Martínez b; S.Camarero ba Centro de Investigación y Tecnología, Pontevedra, Spainb Centro de Investigaciones Biológicas, Madrid, Spain
First Published on: 27 June 2007To cite this Article: Speranza, M., Ibarra, D., Romero, J., Martínez, A. T.,Martínez, M. J. and Camarero, S. (2007) 'Microscopy studies reveal delignificationand sterol removal from eucalypt kraft pulps by laccase-HBT', Biocatalysis andBiotransformation, 25:2, 251 - 259To link to this article: DOI: 10.1080/10242420701444348
URL: http://dx.doi.org/10.1080/10242420701444348
PLEASE SCROLL DOWN FOR ARTICLE
Full terms and conditions of use: http://www.informaworld.com/terms-and-conditions-of-access.pdf
This article maybe used for research, teaching and private study purposes. Any substantial or systematic reproduction,re-distribution, re-selling, loan or sub-licensing, systematic supply or distribution in any form to anyone is expresslyforbidden.
The publisher does not give any warranty express or implied or make any representation that the contents will becomplete or accurate or up to date. The accuracy of any instructions, formulae and drug doses should beindependently verified with primary sources. The publisher shall not be liable for any loss, actions, claims, proceedings,demand or costs or damages whatsoever or howsoever caused arising directly or indirectly in connection with orarising out of the use of this material.

Dow
nloa
ded
By:
[CS
IC] A
t: 15
:05
28 N
ovem
ber 2
007
ORIGINAL ARTICLE
Microscopy studies reveal delignification and sterol removal fromeucalypt kraft pulps by laccase�HBT
M. SPERANZA1,2, D. IBARRA2, J. ROMERO1, A. T. MARTINEZ2, M. J. MARTINEZ2, &
S. CAMARERO2
1Centro de Investigacion y Tecnologıa, ENCE, Carretera de Campano s/n, E-36157 Pontevedra, Spain and 2Centro de
Investigaciones Biologicas, CSIC, Ramiro de Maetzu 9, E-28040 Madrid, Spain
AbstractFungal laccases in the presence of mediators are powerful biocatalysts to degrade lignin. Pycnoporus cinnabarinus laccase and1-hydroxybenzotriazole (HBT) have been successfully used to delignify eucalypt kraft pulp once integrated in a totallychlorine-free bleaching sequence. Real time delignification of kraft pulp by laccase�HBT was verified in situ by monitoringthe loss of lignin autofluorescence during the enzymatic treatment using confocal laser scanning microscopy. The highestdelignification of pulp fibers occurred over a very short time-span (5 min). Moreover, we demonstrate the removal of sterols,responsible for pitch deposits in hardwood kraft pulps, as an additional effect of laccase�HBT. Spherical structures betweenpulp fibers localized by low temperature scanning electron microscopy were removed by laccase�HBT. The use of filipin, aspecific stain, revealed the sterol nature of many of these structures. At the end of the enzyme-aided bleaching sequence, thefluorescent sterols�filipin signals were almost completely absent.
Keywords: Laccase�HBT, lignin, sterols, pitch, eucalypt pulp, filipin staining, microscopy
Introduction
The pulp and paper industry’s concern to reduce
consumption of chemicals and improve pulp quality
has led to the introduction of enzyme-aided tech-
nologies in different parts of the manufacturing
process (Bajpai 2004). During kraft pulping, most
lignin from wood is removed, but residual lignin
remains in the cellulose fibers. This residual lignin,
responsible for the kraft pulp color, must be
removed during the bleaching process. Traditionally,
chlorine and chlorine dioxide have been used as
bleaching agents, but their use in industrial bleach-
ing sequences of most pulp types has been elimi-
nated or considerably reduced due to the toxicity of
the chlorinated compounds released. Modern,
totally chlorine-free (TCF) bleaching is environmen-
tally friendly, but less efficient in attaining high and
permanent pulp brightness due to the lower deligni-
fication power of oxygen and hydrogen peroxide
compared to chlorine reagents.
However, the problem of so-called pitch deposits
in pulp increased with the use of TCF bleaching
because wood extractives survive this type of pro-
cess. The accumulation of wood extractives in pitch
deposits reduces pulp quality and produces pro-
blems at the mill (Back & Allen 2000). Lipases are
being used successfully for pitch biocontrol in soft-
wood pulps, but are not effective in hardwood pulps,
such as eucalypt pulps, since in this case, sterols
(mainly sitosterol and stigmastanol) are the principal
responsible for pitch deposits (del Rıo et al. 1999). A
challenge of recent years has been to find biological
agents for delignification and control of pitch
deposits in paper pulps.
Laccases are multicopper oxidases secreted by
white-rot fungi (among other organisms) that oxi-
dize a wide range of aromatic compounds using
molecular oxygen as final electron acceptor (Thur-
ston 1994; Claus 2004). However, the oxidation
capability of laccases is limited by their relatively low
redox potential (0.4�0.8 V). This limitation is over-
come in the presence of certain compounds acting as
redox mediators that expand laccase activity towards
high redox potential substrates. Once oxidized by the
Correspondence: Susana Camarero, Centro de Investigaciones Biologicas, CSIC, Ramiro de Maeztu 9, 28040 Madrid, Spain. E-mail:
Biocatalysis and Biotransformation, March�August 2007; 25(2�4): 251�259
ISSN 1024-2422 print/ISSN 1029-2446 online # 2007 Informa UK Ltd
DOI: 10.1080/10242420701444348

Dow
nloa
ded
By:
[CS
IC] A
t: 15
:05
28 N
ovem
ber 2
007
enzyme, the mediator radicals undertake the
oxidation of the target aromatic substrate by differ-
ent mechanisms (Baiocco et al. 2003). Laccase�mediator systems constitute powerful biocatalysts
for lignin degradation, and have been thoroughly
assayed for delignification of paper pulps (Bourbon-
nais et al. 1997; Call & Mucke 1997). In this sense,
Pycnoporus cinnabarinus laccase, in the presence of
1-hydroxybenzotriazole (HBT) as mediator, has
been used successfully as a delignifying and bleach-
ing agent to produce high quality paper pulp from
non-wood (Camarero et al. 2004) and eucalypt
pulps (Ibarra et al. 2006a). The integration of this
system in a TCF industrial-like bleaching sequence
in combination with chemicals produced significant
improvement of pulp properties, including higher
brightness and lower kappa number values than
those attained by the standard chemical bleaching
sequence (Ibarra et al. 2006b).
Structural studies of cellulose pulps using different
microscopic techniques provide useful information
about the physical and chemical properties of this
material submitted to different treatments (Daniel
et al. 2002). Low temperature scanning electron
microscopy (LTSEM) allows structural studies avoid-
ing the artifacts caused by the pretreatment of samples
as in conventional SEM. In addition, it is possible to
detect pitch deposition in eucalypt pulps by fluores-
cence microscopy (FM) using filipin staining
(Speranza et al. 2002). Filipin is a polyene antibiotic
used as a probe to detect cholesterol in animals and
ergosterol in fungal membranes (Milhaud et al. 1988;
Severs 1997). This antibiotic reacts specifically with
those 3b-hydroxysterols, including an 8�10 carbon
side-chain and a flat tetracyclic nucleus (Elias et al.
1979; Clejan & Bittman 1985), and, more efficiently,
when a double bond is present at C5 (Milhaud et al.
1988). The complexes formed with filipin have
absorption and emissions bands at 357 and 480 nm,
respectively. Therefore, they can be localized after
ultraviolet excitation using FM. Since these com-
plexes are formed in a 1:1 stoichiometric reaction
(Milhaud et al. 1988), the presence and distribution
of free sterols can be quantitatively analyzed.
In this study, we proposed to disclose a dual effect
of the laccase�HBT system, the removal of sterols
together with the removal of lignin from eucalypt
kraft pulps, by means of different microscopic
techniques.
Materials and methods
Pulp samples
Eucalyptus globulus kraft pulp was produced at the
ENCE mill in Pontevedra (Spain). Brown (un-
bleached) pulp with 15.8 kappa number and oxygen
delignified pulp with 9.7 kappa number were used for
laccase�HBT treatments.
Laccase
Laccase was produced by Beldem (Andenne,
Belgium) from P. cinnabarinus strain ss3 (Herpoel
et al. 2000). Laccase activity was determined by
measuring the oxidation of 5 mM 2,2?-azino-bis (3-
ethylbenzothiazoline-6-sulphonic acid) (ABTS) buf-
fered with 100 mM sodium acetate (pH 5) at 248C.
One unit of activity was defined as the amount of
enzyme that transforms 1 mmol of ABTS to its cation
radical (o436�29,300 M�1 cm�1) per minute.
Pulp bleaching
Pulp treatment with P. cinnabarinus laccase in the
presence of HBT (Sigma-Aldrich), once optimized
for eucalypt pulp (Ibarra et al. 2006a), was inte-
grated in an industrial-type TCF bleaching sequence
(Figure 1) (Ibarra et al. 2006b). Pulp bleaching was
carried out in 4-L reactors with 200 g of eucalypt
pulp (dry weight) at 10% consistency. The TCF
industrial-type bleaching sequence O-O-Q-PoP in-
cluded: two alkaline oxygen stages (O-O), a chela-
tion stage (Q) with diethylenetriaminepenta-acetic
acid (DTPA) and an alkaline peroxide stage (PoP)
including a first step under pressurized O2 and an
atmospheric step. Laccase (20 U g�1 of pulp) and
HBT (1.5% of pulp dry weight) were applied after
the double oxygen stage (O-O-L-Q-PoP) at pH 4 for
2 h at 508C, with stirring for 1 min (60 rev min�1)
each 30 min. The same bleaching sequence with
stage under the same conditions as the L stage but
without enzyme was used as control. Oxygen de-
lignified pulp treated with laccase�HBT (O-O-L)
and full bleached pulp at the end of the enzyme-
aided bleaching sequence (O-O-L-Q-PoP) were
used for microscopy studies, and compared with
their corresponding controls (O-O and O-O-Q-PoP
pulps). Laccase�HBT treatment was also applied on
the unbleached (brown) kraft pulp (kraft-L). Pulp
samples were filtered and washed with distilled water
before storing at �208C.
Low temperature scanning electron microscopy
Ultrastructural characterization of kraft-L, O-O-L
and O-O-L-Q-PoP pulps, with their respective
controls, was carried out using LTSEM. Hydrated
fibers were mounted with O.C.T. (Optimal Cutting
Temperature BDH) on a specimen holder at room
temperature and plunged into subcooled liquid
nitrogen (�2048C) under vacuum using an Ox-
ford CT 1500 Cryotransfer system. The specimens
252 M. Speranza et al.

Dow
nloa
ded
By:
[CS
IC] A
t: 15
:05
28 N
ovem
ber 2
007
were then immediately transferred to the cryo-
preparation chamber, etched (�908C for 2 min),
and gold sputter-coated under pressure (4�10�1
thorr). The observations were carried out on a
LT-SEM Zeiss DSM960, at �135/�1508C,
operated at 15 kV and varying working distances
(8�20 mm).
Sterol staining
Filipin, a specific stain for sterols, was used to
probe and localize the action of laccase�HBT using
FM, and to determine the spatial distribution of the
main lipophilic extractives during the bleaching
process (Speranza et al. 2002). Filipin solution
was prepared by dissolving 2.5 mg of filipin (Sigma)
in 1 mL of dimethylformamide (Merck), and mix-
ing with 50 mL of PBS (Sarig et al. 1994). Filipin
solution was applied to the hydrated pulp samples
with their respective controls, mounted in glass
slides, and incubated in the dark for 20 min at
room temperature. The preparations were rinsed
with PBS to remove the stain, and mounted in
glycerol containing 1% Mowiol 40�88 (Aldrich) as
antifade agent. They were immediately examined
with transmitted light, using phase contrast, and
epifluorescence with an Axioplan Zeiss microscope.
Filipin was excited with UV light using a 360/40D
filter, the fluorescence emitted was analyzed
through a 460-nm barrier filter, and the image
captured digitally with a CCD device (Leica DFC
350 FX). Similar exposure times were used in all
cases to facilitate sterol distribution analysis.
Extractive-free controls were prepared by extracting
pulp samples with acetone in a Soxhlet apparatus
for 8 h, followed by water and PBS washing.
Positive controls for filipin staining were prepared
using sitosterol and stigmastanol solutions (1 mg
mL�1) in acetone.
Confocal microscopy
Real time delignification of brown kraft pulp by
laccase�HBT was monitored in situ for 45 min using
a CLSM Leica TCS microscope with a spectro-
photometer detection system (SP2) (Leica Micro-
systems, Mannheim, Germany). Eucalypt fibers
were deposited in a glass bottom culture dish
(P35G-1.5-14C, MatTek Corporation, USA), and
treated with 20 U laccase and 1.5% HBT per gram
of pulp, in tartrate buffer pH 4, at room tempera-
ture. Pulp buffered with sodium tartrate (pH 4) in
the presence of 1.5% HBT was used as control. The
culture dish was placed immediately in the CLSM
inverted microscope, and lignin autofluorescence
emission between 530 and 560 nm was collected
using an FITC filter, and using blue Ar (488,
20 mw) ion laser as excitation line. Laser excitation
power and emission detection conditions were se-
lected to prevent photo-induced quenching, and
were not altered throughout the experiments (Bars-
berg & Nielsen 2003). Digitally transmitted light
signal was collected simultaneously with the confocal
images. Time course observations of the same fibers
from 0 time (3�5 min after enzyme�mediator was
applied to pulp) and each 5 min during 45 min of
incubation were carried out. Before real time experi-
ments, pulp samples were analyzed using Lambda
scan function to determine the emission spectrum of
autofluorescence for emission wavelengths between
385 and 700 using UV Ar (351 and 364 nm) and
blue Ar (488) as light sources. Lambda image stack
was obtained by scanning the same x �y optical
section with a bandwidth of 20 nm.
O O
Brownpulp
StockL
Q Po PFilter Filter Filter Filter Filter
Laccase/HBT
O2 O2DTPAH2SO4
H2O2NaOH
O2
Figure 1. Scheme of the enzyme-aided TCF sequence used for bleaching eucalypt kraft pulp, resulting from the integration of a laccase�mediator stage (L) in an industrial-type sequence (O-O-Q-PoP), adapted from Ibarra et al. (2006a). The L stage was integrated after the
double oxygen delignification (O-O) and followed by chelation (Q) and hydrogen peroxide (PoP) stages.
Microscopy reveals delignification and sterol removal 253
Dow
nloa
ded
By:
[CS
IC] A
t: 15
:05
28 N
ovem
ber 2
007
Fluorescence analyses
Due to the displacement of the focal plane in the z
axis during the experiments, a single fluorescence
measurement included 15�20 2D-optical sections
successively covering the z direction. The fluores-
cence intensity was measured with Leica Confocal
Software (version 2871-7A), using the region of
interest (ROI) function, the fluorescence intensity of
5�10 ROIs were measured, and the mean fluores-
cence intensity (MFI) was calculated for each cell.
Results and discussion
Integration of an enzymatic treatment with
P. cinnabarinus laccase in the presence of HBT as
mediator in a industrial-type TCF bleaching se-
quence produces a significant decrease in the final
lignin content of eucalypt kraft pulp together with
higher and more permanent pulp brightness (Ibarra
et al. 2006b). The industrial TCF sequence
(O-O-Q-PoP) was based on the use of oxygen and
hydrogen peroxide as delignifying and bleaching
agents. The point where the enzymatic stage was
integrated in the bleaching sequence is indicated in
Figure 1. Pulp samples after different bleaching
stages were analyzed by different microscopic tech-
niques (see Materials and methods for pulp nomen-
clature). Brown (unbleached) kraft pulp treated with
laccase�mediator system was also analyzed for
comparison.
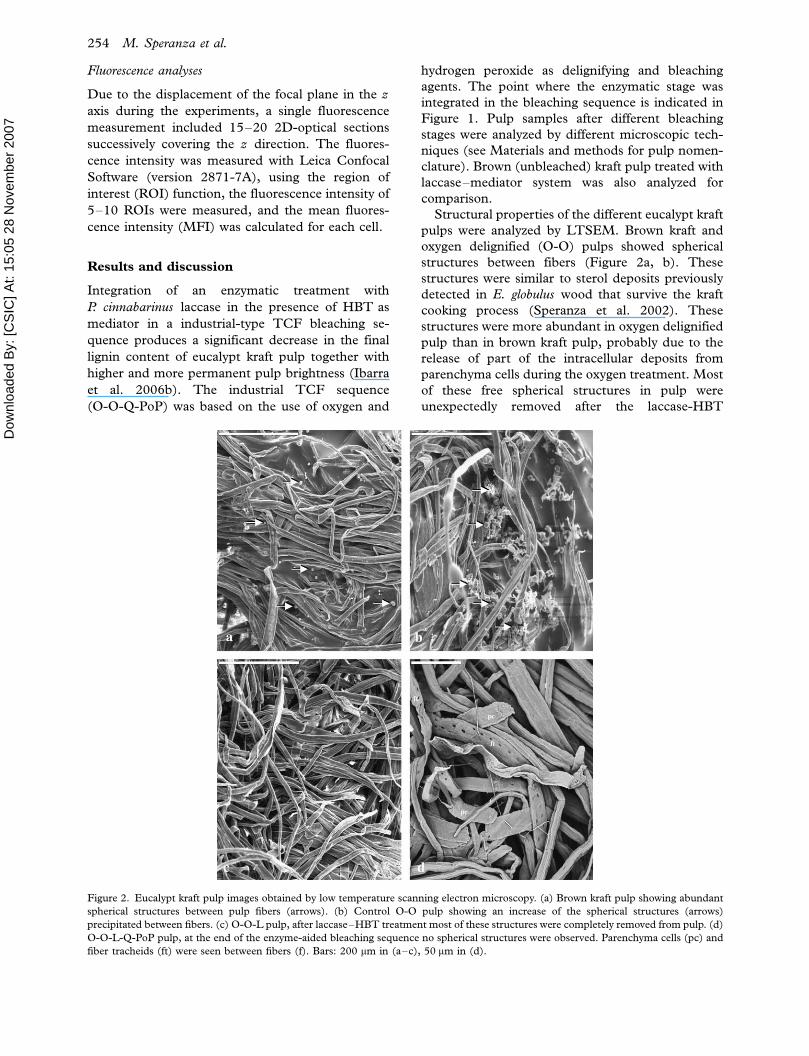
Structural properties of the different eucalypt kraft
pulps were analyzed by LTSEM. Brown kraft and
oxygen delignified (O-O) pulps showed spherical
structures between fibers (Figure 2a, b). These
structures were similar to sterol deposits previously
detected in E. globulus wood that survive the kraft
cooking process (Speranza et al. 2002). These
structures were more abundant in oxygen delignified
pulp than in brown kraft pulp, probably due to the
release of part of the intracellular deposits from
parenchyma cells during the oxygen treatment. Most
of these free spherical structures in pulp were
unexpectedly removed after the laccase-HBT
Figure 2. Eucalypt kraft pulp images obtained by low temperature scanning electron microscopy. (a) Brown kraft pulp showing abundant
spherical structures between pulp fibers (arrows). (b) Control O-O pulp showing an increase of the spherical structures (arrows)
precipitated between fibers. (c) O-O-L pulp, after laccase�HBT treatment most of these structures were completely removed from pulp. (d)
O-O-L-Q-PoP pulp, at the end of the enzyme-aided bleaching sequence no spherical structures were observed. Parenchyma cells (pc) and
fiber tracheids (ft) were seen between fibers (f). Bars: 200 mm in (a�c), 50 mm in (d).
254 M. Speranza et al.

Dow
nloa
ded
By:
[CS
IC] A
t: 15
:05
28 N
ovem
ber 2
007
treatment (Figure 2c), and at the end of the enzyme-
aided bleaching sequence (O-O-L-Q-PoP) were
completely absent (Figure 2d).
To verify the removal of sterols during the enzy-
matic treatment, FM studies were carried out using
a methodology previously developed for sterol loca-
lization in pulp (Speranza et al. 2002). The use of
filipin staining allowed the localization of sitosterol
and stigmastanol, two of the main wood extractives
responsible for pitch deposits in eucalypt pulp.
During chemical bleaching, the strong-fluorescent
signals from the sterols�filipin complexes remained
(Figure 3a, c, e), confirming that these compounds
can survive TCF bleaching (del Rıo et al. 1999).
However, in the eucalypt pulps treated with laccase-
HBT, reduction of fluorescent sterols�filipin signals
was evident, revealing sterol removal by the enzy-
matic system (Figure 3b, d, f).
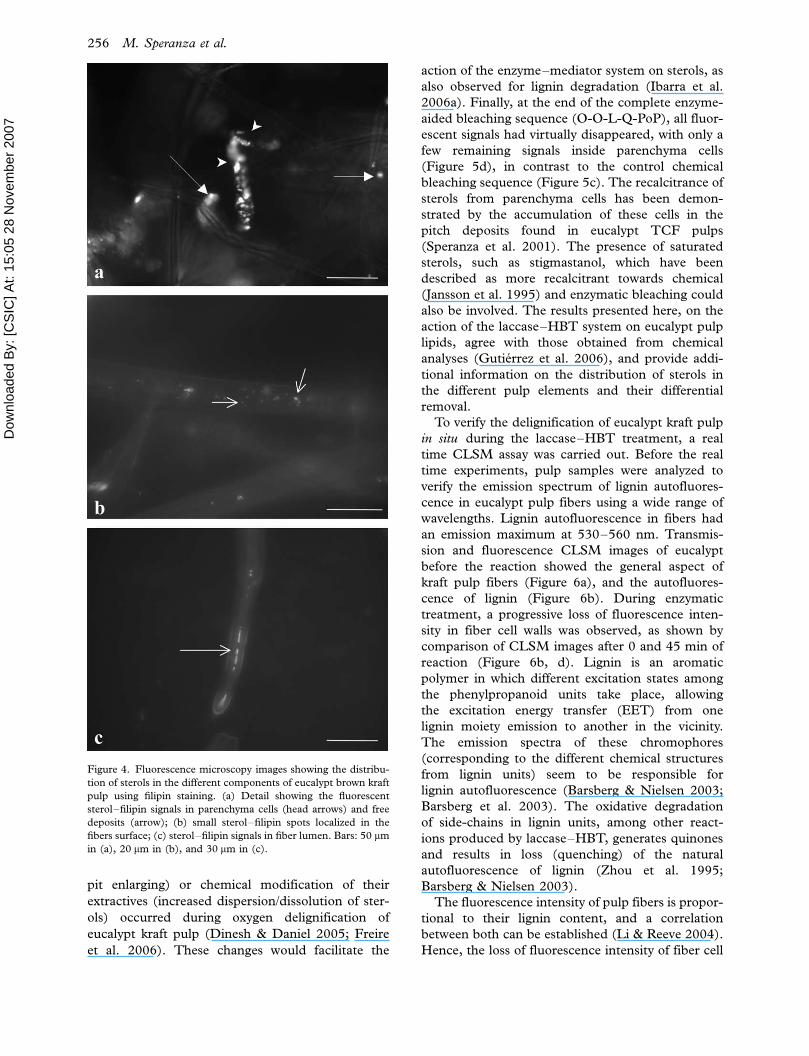
In brown kraft pulp, the strongest fluorescent
signals of the sterol�filipin complexes were found in
the parenchyma cells and free deposits (Figure 4a),
but fluorescence was also localized in the fiber lumen
(Figure 4c) and fiber cell wall surface (Figure 4b).
During laccase-HBT treatment of kraft pulp, sig-
nificant reduction of sterol-filipin fluorescence in
fibers was observed, with the signals inside parench-
yma cells remaining (data not shown).
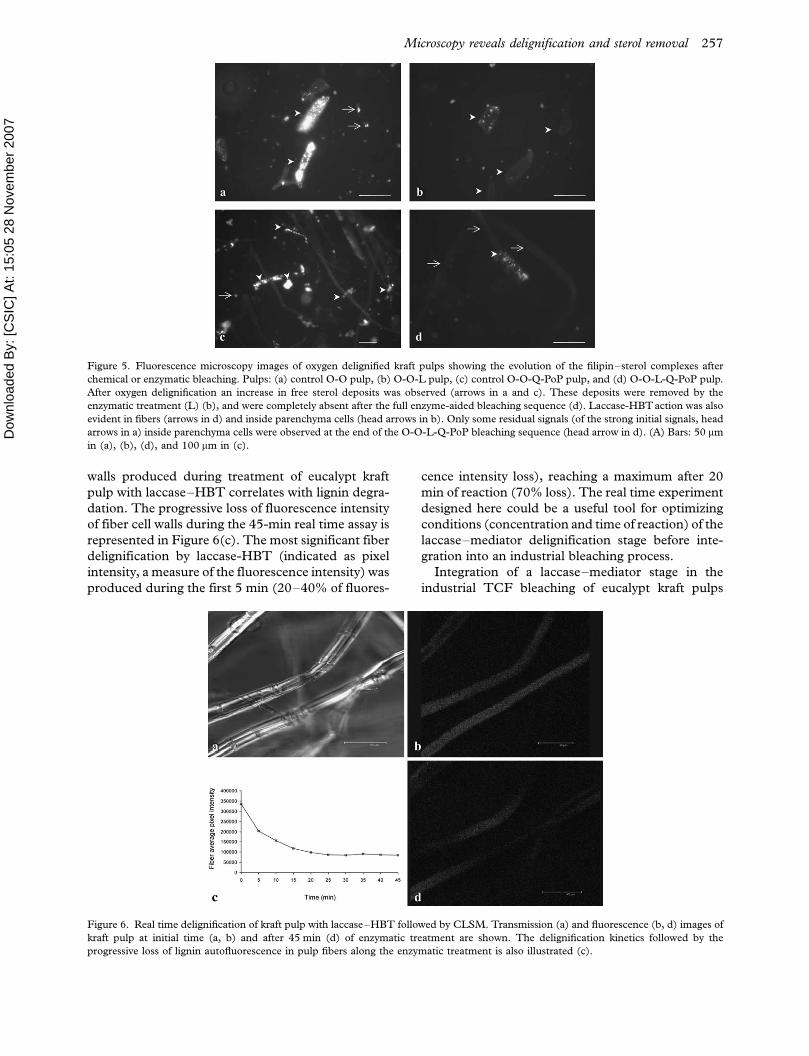
In the control oxygen delignified pulp, the stron-
gest sterol�filipin signals were also found inside
parenchyma cells. However, a high level of fluores-
cence was also detected in free deposits, correlating
with the spherical structures released during
O-O treatment previously observed by LTSEM
(Figure 5a). This fluorescence pattern was main-
tained until the end of the chemical bleaching
sequence (Figure 5c). During laccase-HBT treat-
ment of O-O pulp, significant reduction in the
general fluorescence corresponding to sterol�filipin
signals in free deposits was observed (Figure 5b).
Additionally, the less accessible internal sterol de-
posits from unbroken parenchyma cells, that
retained their fluorescence intensity after laccase-
HBT treatment of brown kraft pulp, were removed
to a greater extent from oxygen delignified pulp by
the enzymatic treatment (Figure 5b). It seems as if
structural changes of the parenchyma cells (such as
Figure 3. Images from fluorescence microscopy showing a general aspect of sterol�filipin complexes in different eucalypt kraft pulps and
the loss of fluorescent signals after laccase�HBT treatment (L). Brown-L (b), O-O-L (d) and O-O-L-Q-PoP (f) pulps were compared with
their corresponding controls: brown kraft pulp (a), O-O pulp (c) and O-O-Q-PoP pulp (e). Inserts correspond to transmission images of the
same fields. Bars: 200 mm.
Microscopy reveals delignification and sterol removal 255
Dow
nloa
ded
By:
[CS
IC] A
t: 15
:05
28 N
ovem
ber 2
007
pit enlarging) or chemical modification of their
extractives (increased dispersion/dissolution of ster-
ols) occurred during oxygen delignification of
eucalypt kraft pulp (Dinesh & Daniel 2005; Freire
et al. 2006). These changes would facilitate the
action of the enzyme�mediator system on sterols, as
also observed for lignin degradation (Ibarra et al.
2006a). Finally, at the end of the complete enzyme-
aided bleaching sequence (O-O-L-Q-PoP), all fluor-
escent signals had virtually disappeared, with only a
few remaining signals inside parenchyma cells
(Figure 5d), in contrast to the control chemical
bleaching sequence (Figure 5c). The recalcitrance of
sterols from parenchyma cells has been demon-
strated by the accumulation of these cells in the
pitch deposits found in eucalypt TCF pulps
(Speranza et al. 2001). The presence of saturated
sterols, such as stigmastanol, which have been
described as more recalcitrant towards chemical
(Jansson et al. 1995) and enzymatic bleaching could
also be involved. The results presented here, on the
action of the laccase�HBT system on eucalypt pulp
lipids, agree with those obtained from chemical
analyses (Gutierrez et al. 2006), and provide addi-
tional information on the distribution of sterols in
the different pulp elements and their differential
removal.
To verify the delignification of eucalypt kraft pulp
in situ during the laccase�HBT treatment, a real
time CLSM assay was carried out. Before the real
time experiments, pulp samples were analyzed to
verify the emission spectrum of lignin autofluores-
cence in eucalypt pulp fibers using a wide range of
wavelengths. Lignin autofluorescence in fibers had
an emission maximum at 530�560 nm. Transmis-
sion and fluorescence CLSM images of eucalypt
before the reaction showed the general aspect of
kraft pulp fibers (Figure 6a), and the autofluores-
cence of lignin (Figure 6b). During enzymatic
treatment, a progressive loss of fluorescence inten-
sity in fiber cell walls was observed, as shown by
comparison of CLSM images after 0 and 45 min of
reaction (Figure 6b, d). Lignin is an aromatic
polymer in which different excitation states among
the phenylpropanoid units take place, allowing
the excitation energy transfer (EET) from one
lignin moiety emission to another in the vicinity.
The emission spectra of these chromophores
(corresponding to the different chemical structures
from lignin units) seem to be responsible for
lignin autofluorescence (Barsberg & Nielsen 2003;
Barsberg et al. 2003). The oxidative degradation
of side-chains in lignin units, among other react-
ions produced by laccase�HBT, generates quinones
and results in loss (quenching) of the natural
autofluorescence of lignin (Zhou et al. 1995;
Barsberg & Nielsen 2003).
The fluorescence intensity of pulp fibers is propor-
tional to their lignin content, and a correlation
between both can be established (Li & Reeve 2004).
Hence, the loss of fluorescence intensity of fiber cell
Figure 4. Fluorescence microscopy images showing the distribu-
tion of sterols in the different components of eucalypt brown kraft
pulp using filipin staining. (a) Detail showing the fluorescent
sterol�filipin signals in parenchyma cells (head arrows) and free
deposits (arrow); (b) small sterol�filipin spots localized in the
fibers surface; (c) sterol�filipin signals in fiber lumen. Bars: 50 mm
in (a), 20 mm in (b), and 30 mm in (c).
256 M. Speranza et al.
Dow
nloa
ded
By:
[CS
IC] A
t: 15
:05
28 N
ovem
ber 2
007
walls produced during treatment of eucalypt kraft
pulp with laccase�HBT correlates with lignin degra-
dation. The progressive loss of fluorescence intensity
of fiber cell walls during the 45-min real time assay is
represented in Figure 6(c). The most significant fiber
delignification by laccase-HBT (indicated as pixel
intensity, a measure of the fluorescence intensity) was
produced during the first 5 min (20�40% of fluores-
cence intensity loss), reaching a maximum after 20
min of reaction (70% loss). The real time experiment
designed here could be a useful tool for optimizing
conditions (concentration and time of reaction) of the
laccase�mediator delignification stage before inte-
gration into an industrial bleaching process.
Integration of a laccase�mediator stage in the
industrial TCF bleaching of eucalypt kraft pulps
Figure 5. Fluorescence microscopy images of oxygen delignified kraft pulps showing the evolution of the filipin�sterol complexes after
chemical or enzymatic bleaching. Pulps: (a) control O-O pulp, (b) O-O-L pulp, (c) control O-O-Q-PoP pulp, and (d) O-O-L-Q-PoP pulp.
After oxygen delignification an increase in free sterol deposits was observed (arrows in a and c). These deposits were removed by the
enzymatic treatment (L) (b), and were completely absent after the full enzyme-aided bleaching sequence (d). Laccase-HBTaction was also
evident in fibers (arrows in d) and inside parenchyma cells (head arrows in b). Only some residual signals (of the strong initial signals, head
arrows in a) inside parenchyma cells were observed at the end of the O-O-L-Q-PoP bleaching sequence (head arrow in d). (A) Bars: 50 mm
in (a), (b), (d), and 100 mm in (c).
Figure 6. Real time delignification of kraft pulp with laccase�HBT followed by CLSM. Transmission (a) and fluorescence (b, d) images of
kraft pulp at initial time (a, b) and after 45 min (d) of enzymatic treatment are shown. The delignification kinetics followed by the
progressive loss of lignin autofluorescence in pulp fibers along the enzymatic treatment is also illustrated (c).
Microscopy reveals delignification and sterol removal 257

Dow
nloa
ded
By:
[CS
IC] A
t: 15
:05
28 N
ovem
ber 2
007
could be advantageous from two points of view: (i)
to decrease lignin content of pulp, providing higher
degrees of brightness and lower brightness reversion
than obtained by standard sequences, and (ii) for
pitch control, by removing the sterols accumulated
in pulp during TCF bleaching sequences. Both
enzymatic effects should enhance pulp quality with
respect to standard chemical TCF bleaching, as
revealed by chemical analyses (Gutierrez et al.
2006). Moreover, the use of natural mediators
obtained from cheap sources, such as lignin
(Camarero et al. 2005), instead of synthetic ones,
might significantly increase the feasibility of laccase�mediator systems in biotechnological applications
of environmental interest (Camarero et al. 2007;
Gutierrez et al. 2007).
Conclusions
. Free HBT radicals generated by oxidation with
laccase are able to concomitantly remove both
lignin and lipids in eucalypt kraft pulps, as
revealed by microscopic techniques.
. Filipin staining localization of sterols in the
different pulp components after laccase�HBT
treatment, demonstrated the ability of the
enzymatic system to degrade the recalcitrant
spherical structures from parenchyma cells, that
are mainly responsible for the pitch problem in
eucalypt pulps.
. CLSM analyses allowed for the first time in situ
demonstration of lignin degradation in eucalypt
fibers by the laccase�HBT system, and deter-
mination of the process kinetics.
Acknowledgements
This study has been funded by the Spanish
CTQ2005-08925-C02-02/PPQ, BIO2005-03569
and BIO2003-00621 projects, the EU project
NMPQ-CT-2006-026456 and ENCE-CSIC con-
tract. The authors thank M. Asther (INRA
Marseille) for the fungal strain, Beldem (Andenne,
Belgium) for laccase samples, F. Pinto (CCMA,
CSIC) and M.T. Seisdedos and S. Hernandez (CIB,
CSIC) for technical assistance in LTSEM and
CLSM, respectively. M.S and S.C. are acknowl-
edged for TQ and R&C contracts of the Spanish
MEC, and D. Ibarra thank the Spanish CSIC for
I3P fellowship.
References
Back EL, Allen LH. 2000. Pitch Control, Wood Resin and
Deresination. Atlanta, GA: TAPPI Press.
Baiocco P, Barreca AM, Fabbrini M, Galli C, Gentili P. 2003.
Promoting laccase activity towards non-phenolic substrates: A
mechanistic investigation with some laccase�mediator systems.
Org Biomol Chem 1(1):191�197.
Bajpai P. 2004. Biological bleaching of chemical pulps. Crit Rev
Biotechnol 24(1):1�58.
Barsberg S, Nielsen KA. 2003. Oxidative quenching of spruce
thermomechanical pulp fiber autofluorescence monitored in
real time by confocal laser scanning microscopy: Implica-
tions for lignin autofluorescence. Biomacromolecules
4(1):64�69.
Barsberg S, Elder T, Felby C. 2003. Lignin�quinone interactions:
Implications for optical properties of lignin. Chem Mater
15(3):649�655.
Bourbonnais R, Paice MG, Freiermuth B, Bodie E, Borneman S.
1997. Reactivities of various mediators and laccases with kraft
pulp and lignin model compounds. Appl Environ Microbiol
63(12):4627�4632.
Call HP, Mucke I. 1997. History, overview and applications of
mediated lignolytic systems, especially laccase�mediator sys-
tems (Lignozym(R)-process). J Biotechnol 53(2�3):163�202.
Camarero S, Garcıa O, Vidal T, Colom J, del Rıo JC, Gutierrez A,
Gras JM, Monje R, Martınez MJ, Martınez AT. 2004. Efficient
bleaching of non-wood high-quality paper pulp using laccase�mediator system. Enzyme Microb Technol 35(2/3):113�120.
Camarero S, Ibarra D, Martınez MJ, Martınez AT. 2005. Lignin-
derived compounds as efficient laccase mediators for decolor-
ization of different types of recalcitrant dyes. Appl Environ
Microbiol 71(4):1775�1784.
Camarero S, Ibarra D, Martınez AT, Romero J, Gutierrez A, del
Rio JC. 2007. Paper pulp delignification using laccase and
natural mediators. Enzyme Microb Technol 40:1264�1271.
Claus H. 2004. Laccases: structure, reactions, distribution.
Micron 35(1�2):93�96.
Clejan S, Bittman R. 1985. Rates of amphotericin B and filipin
association with sterols. A study of changes in sterol structure
and phospholipid composition of vesicles. J Biol Chem
260(5):2884�2889.
Daniel G, Duchesne I, Tokoh C, Bardage S. 2002. The surface
and intracellular nanostructure of wood fibres: Electron
microscope methods and applications. In: Schmitt U, Ander
P, Barnett J, Emons AMC, Jeronimidis G, Saranpaa P, Tschegg
S, editors. Wood fibre cell walls: Methods to study their
formation, structure and properties. COST European Coop-
eration in the Field of Scientific and Technical Research ed.
Cost Action E20. p 87�104.
del Rıo JC, Gutierrez A, Gonzalez-Vila FJ. 1999. Analysis of
impurities occurring in a totally chlorine-free bleached kraft
pulp. J Chromatogr 830:227�232.
Dinesh F, Daniel G. 2005. The state and spatial distribution of
extractives during birch kraft pulping, as evaluated by staining
techniques. Nordic Pulp Paper J 20(4):383�391.
Elias PM, Friend DS, Goerke J. 1979. Membrane sterol hetero-
geneity: Freeze�fracture detection with saponins and filipin.
J Histochem Cytochem 27:1247�1260.
Freire CSR, Silvestre AJD, Neto CP, Evtuguin DV. 2006. Effect of
oxygen, ozone and hydrogen peroxide bleaching stages on the
contents and composition of extractives of Eucalyptus globulus
kraft pulps. Bioresour Technol 97:420�428.
Gutierrez A, del Rıo JC, Ibarra D, Rencoret J, Romero J, Speranza
M, Camarero S, Martınez MJ, Martınez AT. 2006. Enzymatic
removal of free and conjugated sterols forming pitch deposits in
environmentally sound bleaching of eucalypt paper pulp.
Environ Sci Technol 40(10):3416�3422.
Gutierrez A, Rencoret J, Ibarra D, Molina S, Camarero S,
Romero J, del Rıo JC, Martınez AT. 2007. Removal of
lipophilic extractives from paper pulp by laccase and lignin-
derived phenols as natural mediators. Environ Sci Technol
41:4124�4129.
258 M. Speranza et al.

Dow
nloa
ded
By:
[CS
IC] A
t: 15
:05
28 N
ovem
ber 2
007
Herpoel I, Moukha S, Lesage-Meessen L, Sigoillot JC, Asther M.
2000. Selection of Pycnoporus cinnabarinus strains for laccase
production. FEMS Microbiol Lett 183(2):301�306.
Ibarra D, Romero J, Martınez MJ, Martınez AT, Camarero S.
2006a. Exploring the enzymatic parameters for optimal de-
lignification of eucalypt kraft pulp by laccase�mediator.
Enzyme Microb Technol 39:1319�1327.
Ibarra D, Camarero S, Romero J, Martınez MJ, Martınez AT.
2006b. Integrating laccase�mediator treatment into an indus-
trial-type sequence for totally chlorine free bleaching eucalypt
kraft pulp. J Chem Technol Biotechnol 81:1159�1165.
Jansson MB, Wormald P, Dahlman O. 1995. Reactions of wood
extractives during ECF and TCF bleaching of kraft pulp. Pulp
Paper Can 96(4):T134�T137.
Li K, Reeve DW. 2004. Fluorescent labeling of lignin in the wood
pulp fiber wall. J Wood Chem Technol 24(2):169�181.
Milhaud J, Bolard J, Benveniste P, Hatrmann MA. 1988.
Interaction of the polyene antibiotic filipin with model and
natural membranes containing plant sterols. Biochim Biophys
Acta 943:315�325.
Sarig S, Weiss TA, Katz I, Kahana F, Azoury R, Okon E, Kruth
HS. 1994. Detection of cholesterol associated with calcium
mineral using confocal fluorescence microscopy. Lab Invest
71(5):782�787.
Severs NJ. 1997. Cholesterol cytochemistry in cell biology and
disease. In: Bittman R, editor. Cholesterol: Its functions and
metabolism in biology and medicine. New York, NY: Plenum
Press. p 477�505.
Speranza M, Martınez MJ, Martınez AT. 2001. Wood and pulp
localization of sterols involved in pitch deposition using
fluorescent filipin staining. Proceedings of the 11th ISWPC,
Nice, 11�14 June.
Speranza M, Martınez MJ, Gutierrez A, del Rıo JC, Martınez AT.
2002. Wood and pulp localization of sterols involved in pitch
deposition using filipin fluorescent staining. J Pulp Paper Sci
28(9):292�297.
Thurston CF. 1994. The structure and function of fungal
laccases. Microbiology 140:19�26.
Zhou JH, Olmstead JA, Gray DG. 1995. Fluorescent detection of
o-quinones formed in lignin-containing pulps during irradia-
tion. J Wood Chem Technol 15(1):43�64.
Microscopy reveals delignification and sterol removal 259






























