Introduction to Transcriptional
Machinery

"DNA makes RNA, RNA makes protein, and proteins make us." Francis Crick

Central Dogma of Molecular Biology

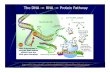
RNA Polymerase of E. Coli• Transcribes all mRNA, rRNAs and tRNAs• 7,000 molecules per cell• 5,000 molecules are synthesizing RNA at any
given time• M.W. of the holoenzyme is ~465 Kd
RNA Polymerase of E. Coli
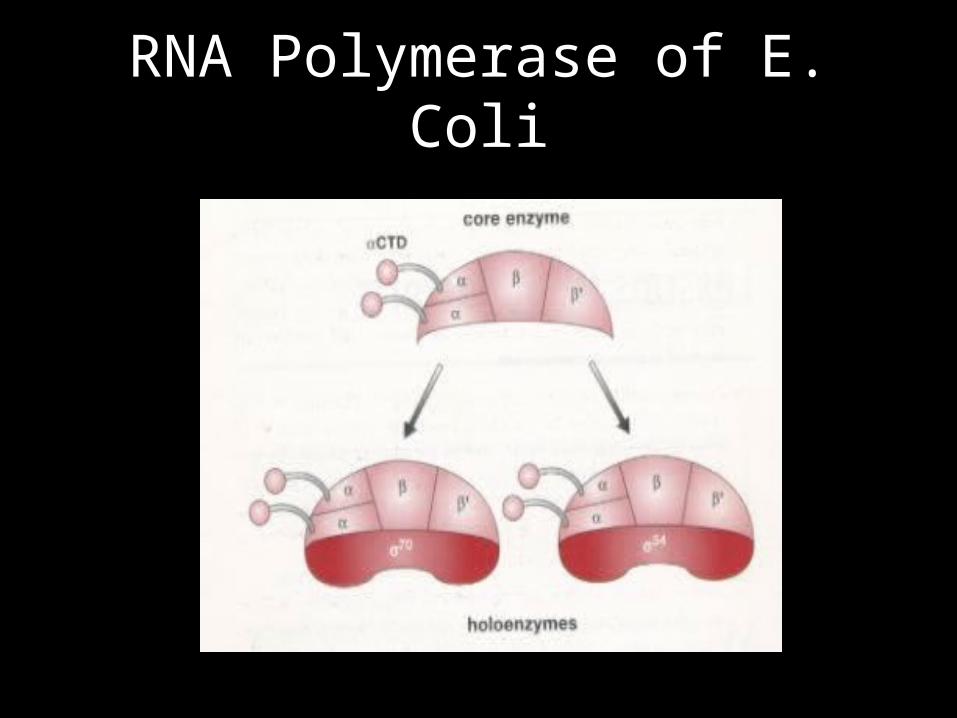
Factor Controls Specificity

Holoenzyme & Core Enzyme
• Holoenzyme binds promoters with half lives of hours - 1,000 time higher than core enzyme.
• Holoenzyme has a drastically reduced ability to recognize “loose binding sites” - half life of <1sec – 104 time lower than core enzyme.

Transcription Initiation

Promoter Elements in E. Coli
• -35: recognition domain• -10: unwinding domain• Seperating distances• UP element• Start Point: purine in 90% of the genes
16-19

T80A95T45A60A50T96
First Level of Regulation
• ~100 fold variation in the binding rate of RNA Pol to different promoters in vitro.
• Binding rates correlate with the frequencies of transcription in vivo.

E. Coli has several Factors

Factors Recognize Promoters by Consensus Sequences
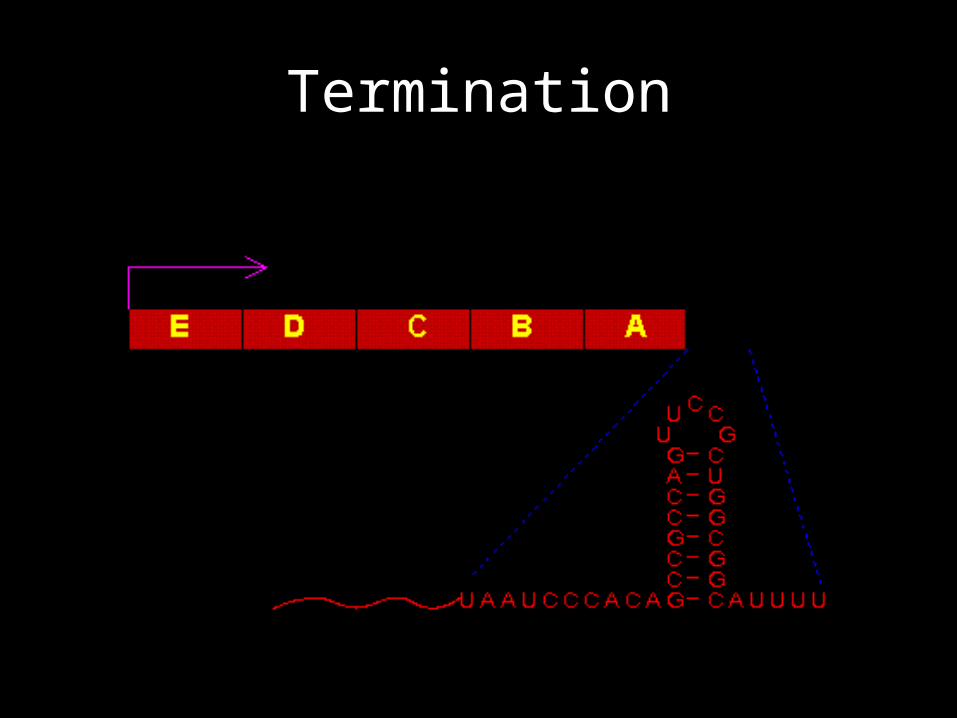
Termination

What was known in the 1960’s
• Jacob and Monod 1961 – genetic control mechanisms in prokaryotes
• Anticipation for Eukarotes…• Eukaryotes – genomic complexity –
reiterated DNA sequences• Lack of genetic approach

February 1969, Strait of Juan de Fuca

“Eureka!”
Taken from: The eukaryotic tarnscriptional machinery, Robert G Roeder

3 RNA Polymerases
• Pol I localized within nucleoli – the sites of rRNA gene transcription
• Pol II and Pol III restricted to the nucleoplasm

• Roberto Weinmann - 1974• Differential sensitivities to the
mushroom toxin - amanitin• Pol I – rRNA synthesis• Pol II – adenovirus pre-mRNA• Pol III – cellular 5S and tRNA
3 RNA Polymerases

RNA Polymerases of Eukaryotes
• Pol I - transcribes pre-ribosomal RNA (18S, 5.8S, 28S)• Pol II - mRNAs• Pol III - tRNAs, 5S RNAs and some specialized small RNAs.

RNA Polymerase II
• 2002 – RNA Pol II structure• 2003 – transcription complex
structure (RNA Pol II + TFIIS), ’, I, II, - conserved in yeast
and bacteria – evolutionary conserved mechanism of transcription

Significant homology between eukaryotic and bacterial RNA polymerases in their structure

Transcription Mechanism
• RNA Pol II can catalyze RNA synthesis but cannot initiate.• Assembly• Initiation• Elongation• Termination

Transcription
Mechanism

TBP
• Only GTF that creates sequence specific contact with DNA
• Unusual Binding in minor groove• Causes DNA bending

• 80% conserved between yeast and man• Large outer surface binds proteins• Deformation of DNA structure, but no strand separation
TBP

The transcriptional machinery
• Initiation begins with the formation of the first phosphodiester bond and phosphorylation of Ser5 on the CTD by TFIIH.
• mRNA passes through a positively charged exit channel, and once the RNA is approximately 18n long it becomes accessible to the RNA processing machinery.
• Consistent with the coupling of transcript capping to early transcription events

Pre-mRNA Processing
• Addition of 5’ cap
• Splicing – removal of intron sequences
• Generation of 3’ poly-A tail.
• 3’ cleavage
• RNA serveillance by the exosome
• Packaging of the mRNA for export
Occurs (most efficiently) co-transcriptionally

Transcription Regulating Elements
• GTFs - required at any Pol II promoter• Enhancers – sequences, increase transcription• Transactivators - bind enhancers • Co-activators - act indirectly, not by binding to
DNA, communication between transactivators and RNA PolII + GTS
• Mediator - 20 proteins, Interacts with CTD

Major Differences between Pro & Eu
• Prokaryotes RNA Pol has access to promoters and initiates transcription even in the absence of activators and repressors.
• Eukaryotes - promoters are generally inactive in vivo
• Transcription in eukaryotes is seperated in both space and time from translation
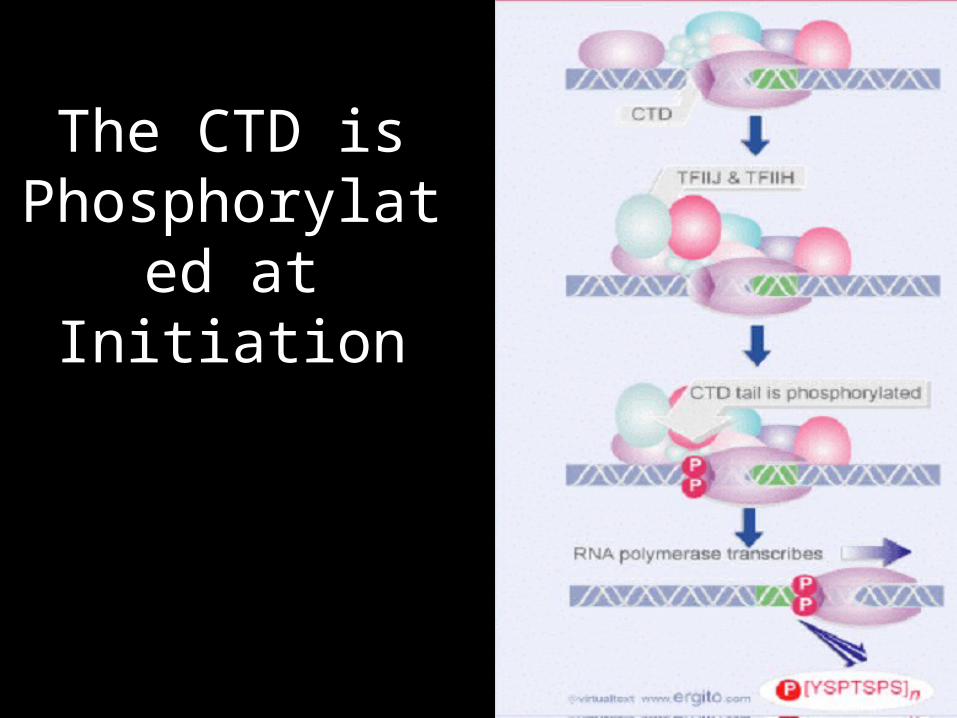
The CTD is Phosphorylated at Initiation

CTD
• Highly conserved tandemly repeated heptapeptide motif (YSPTSPS)
• Platform for ordered assembly of the different families of pre-mRNA processing machinery
• Undergoes phosphorylation and dephosphorylation during the transcription cycle

CTD
• P-TEFb contains CDK9 and cyclin T• It couples RNA processing to
transcription by phosphorylating Ser2 of CTD
• RNA Pol II is recycled through dephosphorylation of Ser2 by the phosphatase activity of Fcp1

CTD Phosphorylation During Transcription

Splicing (& Alternative Splicing)

Expansive role of Transcription
• RNA surveillance – Exosome associates with Spt6 EF
• Coupling of transcription to mRNA export
• 19S particle of the Proteosome recruited to active promoters – important for efficient RNA Pol II elongation

Translation and Post-Translation
• Bacteria – translation occurs as the nascent transcript emerges from the RNA polymerase
• It is assumed that in eukaryotes transcription and translation are spatially separated events
• Protein synthesis – solely a cytoplasmic event? (1977 – Gozes et al, 2001 lborra et al)

Traditional View of Gene Expression

Contemporary View of Gene Expression

The Sister Chromatids
of a Mitotic Pair
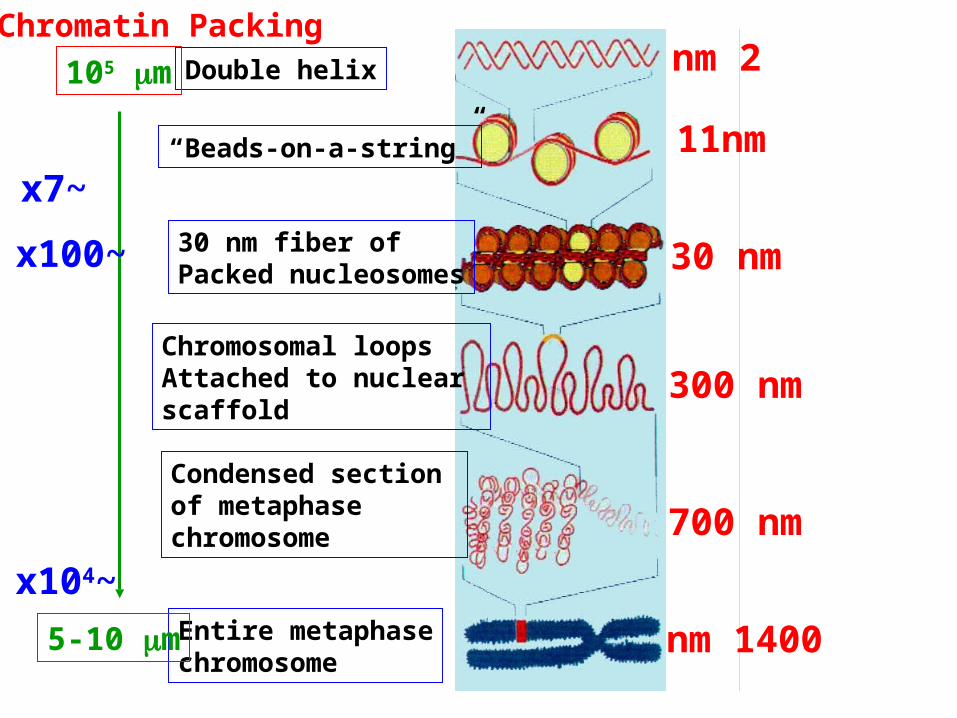
2 nm
11nm
30 nm
300 nm
700 nm
1400 nm
30 nm fiber of Packed nucleosomes
Chromosomal loopsAttached to nuclear scaffold
Condensed section of metaphasechromosome
Entire metaphasechromosome
“Beads-on-a-string”
Chromatin PackingDouble helix105 m
5-10 m
~x104
~x7
~x100

Chromatin Structure
• DNA accessibility – a major challenge in a chromatin environment
• Nucleosomes –
building
blocks of
chromatin
AT Pairs Are Preferred
GC Pairs Are Preferred
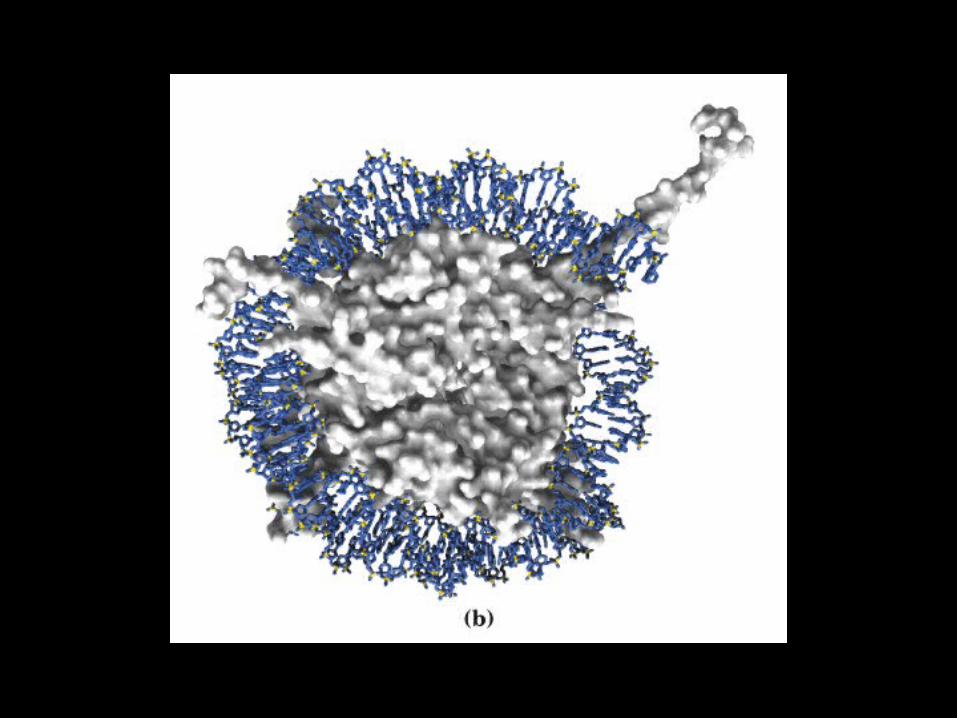
Histone Core DNA

•146 bp are wrapped around the histone core 1.75 times
• ~0-80 bp in the linker sequences between nucleosomes
• Human genome (~6x109 bp) contains ~3x107 nucleosomes • The histone core (octamer) consists of two copies of:
• Histones H2A, H2B, H3 and H4• Histone H1 binds in the spacing linker sequence
Structure of the Nucleosome

H2B
H2A
H4
H3
H2B
H3
H2A
DNA
HistoneCore
The Nucleosome


Histones•Highly conserved throughout eukaryotic evolution
• Mutations in histones encoding genes are often lethal
• Highly abundant (~60 million copies/cell)
• Additional non-histone proteins play a role in the chromatin structure and function

Types and Properties of Histones
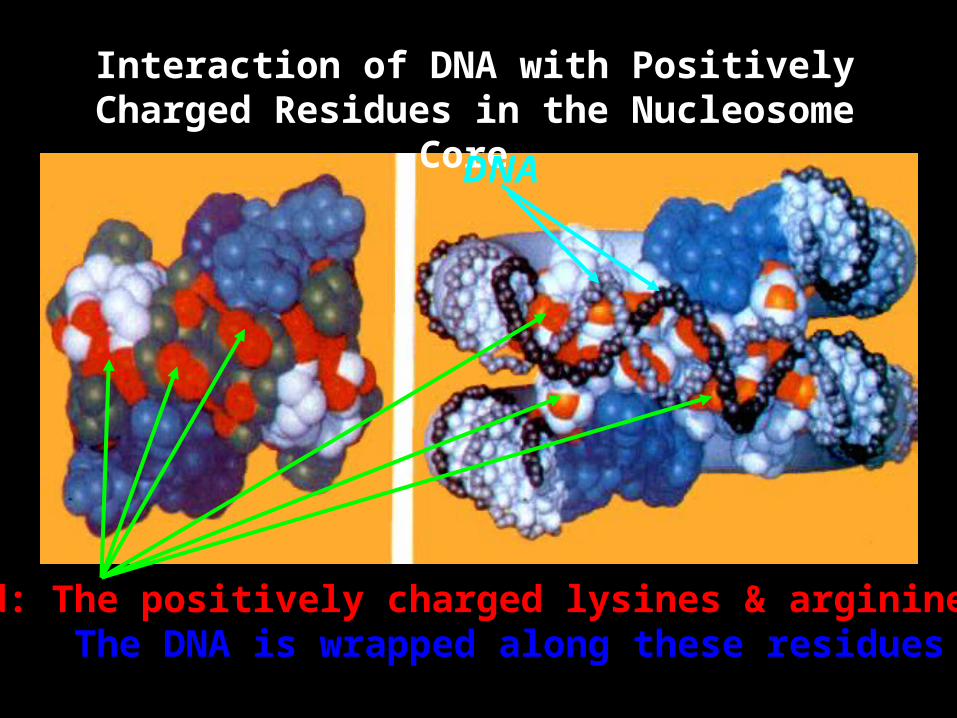
Interaction of DNA with Positively Charged Residues in the Nucleosome
Core
Red: The positively charged lysines & arginines The DNA is wrapped along these residues
DNA
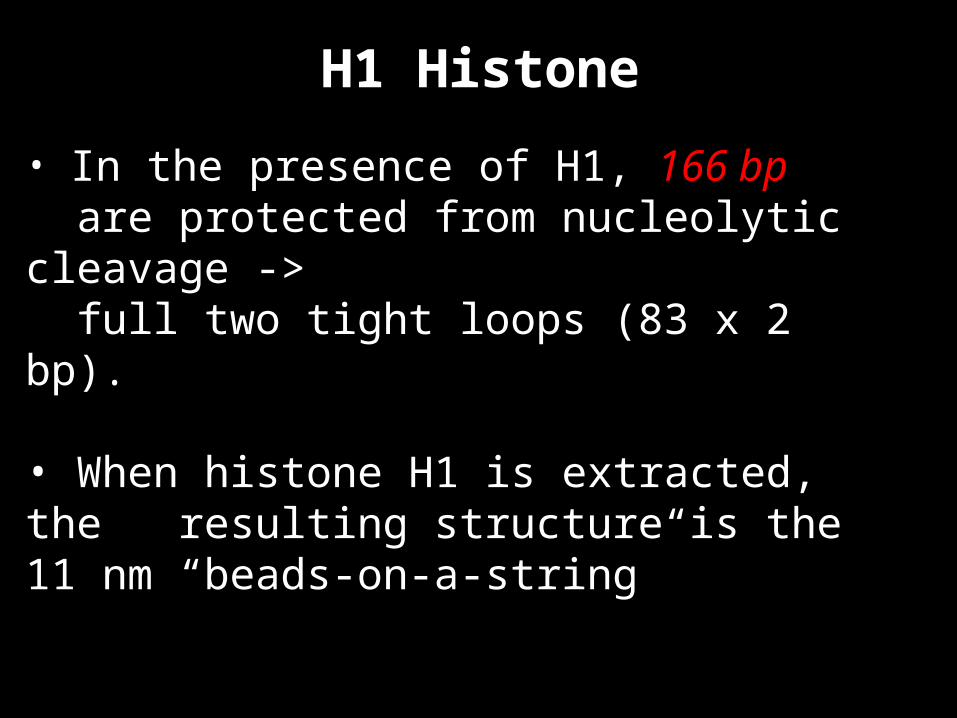
H1 Histone
• In the presence of H1, 166 bp are protected from nucleolytic cleavage -> full two tight loops (83 x 2 bp).
• When histone H1 is extracted, the resulting structure is the 11 nm “beads-on-a-string”

H1 Histone
DNA
CoreHistone
View Along the Axis of One Turn of the 30nm Fiber

DNA
Side View of the 30nm Fiber
Histone core
Nucleosome Histone H1
30 nm fiber
11 nm fiber
Histone H1

• heterochromatin and euchromatin
• How do TFs access the DNA in the first place?
• Example: GR, NF1 and MMTV gene (Di Groce et al., 1999)
“Chicken and Egg” Scenario

Histone Code Hypothesis
• language of covalent post-translational histone modifications
• acetylation• phosphorylation • methylation • ubiquitylation • ADP-ribosylation and • glycosylation

• Sequence elements• Post-translational modifications• Nucleosome remodeling complexes• Transcriptional Elongation
Regulation of Nucleosome Stability
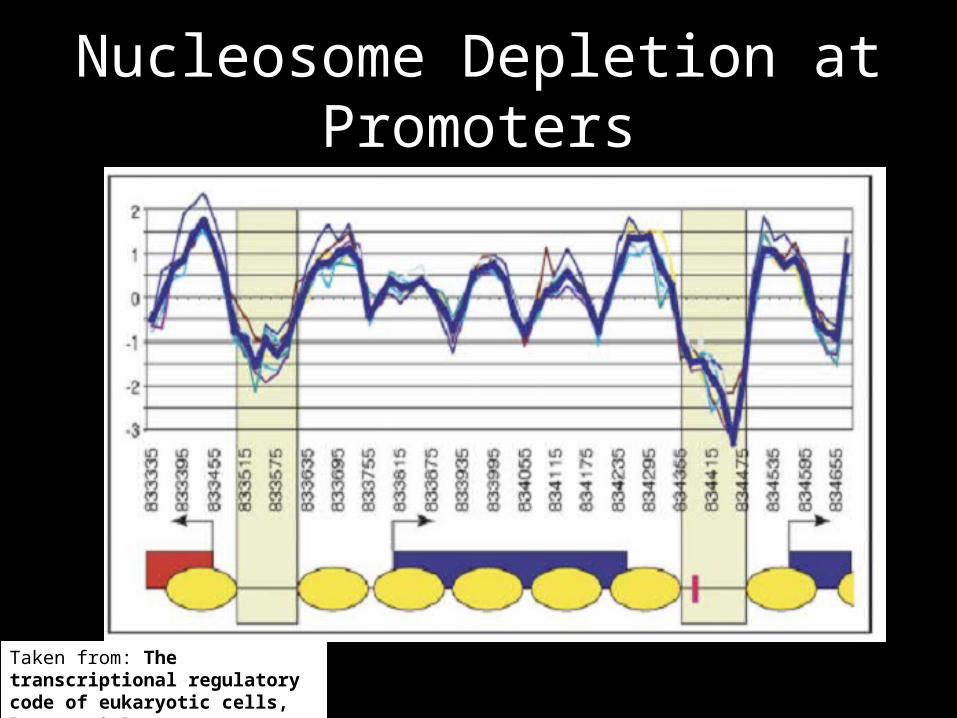
Nucleosome Depletion at Promoters
Taken from: The transcriptional regulatory code of eukaryotic cells, Barrera & Ren

Dynamic Histone Methylation
• Histone methylation is irreversible!• Methylation is dynamic - alterations
in H3-K4 and H3-K9 methylation – (Martinowich et al. 2003)
• Required: a mechanism for removal of long- term histone modifications!

Histone Variants
• H2AZ prevents spread of heterochromatin and gene silencing in transcriptionally active regions
• H3.3 enriched in histone modifications that correspond to transcriptional activation

Histone Exchane
• SWI/SNF and the RSC exchange H2A-H2B dimers
• FACT - EF that removes one H2A-H2B dimer from the nucleosome
• SWR1 (ATPase) selectively exchanges H2A histone variants
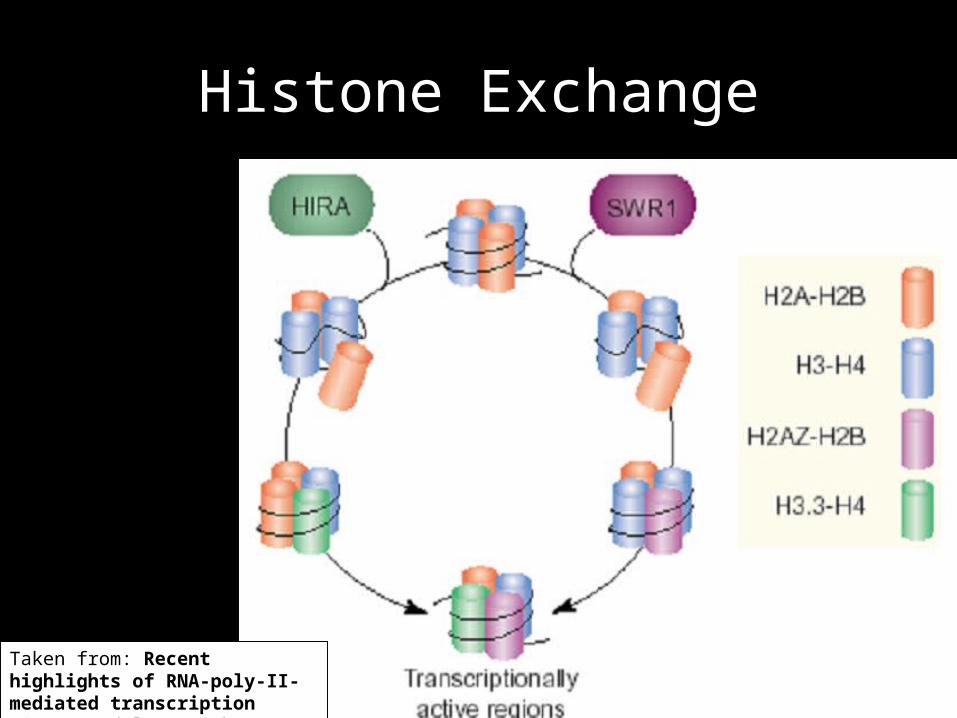
Histone Exchange
Taken from: Recent highlights of RNA-poly-II-mediated transcription Sims, Mandal & Reinberg
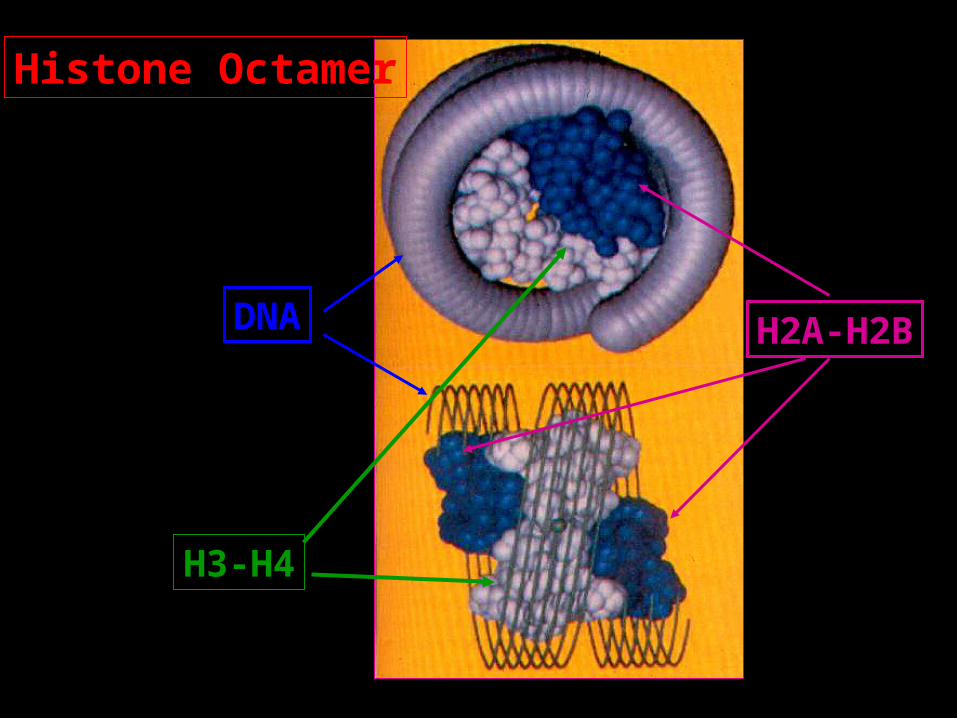
Histone Octamer
DNA H2A-H2B
H3-H4

How this Helps Transcription?
Taken from: Recent highlights of RNA-poly-II-mediated transcription Sims, Mandal & Reinberg

Take Home Message
• Complexity of the transcription is the rule, not the exception.
• Transcription is coupled to mRNA processing, RNA surveillance and export, among other cellular processes.
• Chromatin structure – transcription regulatory code.