



M I N I R E V I E W
Identifying infection-associatedgenesofCandidaalbicans in thepostgenomiceraDuncan Wilson1, Sascha Thewes2, Katherina Zakikhany3, Chantal Fradin4, Antje Albrecht1,Ricardo Almeida1, Sascha Brunke1, Katharina Grosse1, Ronny Martin1, Francois Mayer1, Ines Leonhardt1,Lydia Schild1, Katja Seider1, Melanie Skibbe1, Silvia Slesiona1, Betty Waechtler1, Ilse Jacobsen1 &Bernhard Hube1,5
1Department of Microbial Pathogenicity Mechanisms, Leibniz Institute for Natural Product Research and Infection Biology, Hans-Knoell-Institute, Jena,
Germany; 2Institute for Biology, Free University, Berlin, Germany; 3Department of Microbiology, Tumor and Cell Biology, Karolinska Institute, Stockholm,
Sweden; 4Laboratory of Fundamental and Applied Mycology, University of Lille, Lille, France; and 5Friedrich Schiller University, Jena, Germany
Correspondence: Bernhard Hube,
Department of Microbial Pathogenicity
Mechanisms, Leibniz Institute for Natural
Product Research and Infection Biology, Hans
Knoell Institute (HKI), Beutenbergstrasse 11a,
07745 Jena, Germany. Tel.: 149 3641 532
1401; fax: 149 3641 532 0810;
e-mail: [email protected]
Received 3 March 2009; revised 9 April 2009;
accepted 10 April 2009.
Final version published online 18 May 2009.
DOI:10.1111/j.1567-1364.2009.00524.x
Editor: Teun Boekhout
Keywords
Host-pathogen interactions; invasion; gene
expression; infection models.
Abstract
The human pathogenic yeast Candida albicans can cause an unusually broad range
of infections reflecting a remarkable potential to adapt to various microniches
within the human host. The exceptional adaptability of C. albicans is mediated by
rapid alterations in gene expression in response to various environmental stimuli
and this transcriptional flexibility can be monitored with tools such as micro-
arrays. Using such technology it is possible to (1) capture a genome-wide portrait
of the transcriptome that mirrors the environmental conditions, (2) identify
known genes, signalling pathways and transcription factors involved in pathogen-
esis, (3) identify new patterns of gene expression and (4) identify previously
uncharacterized genes that may be associated with infection. In this review, we
describe the molecular dissection of three distinct stages of infections, covering
both superficial and invasive disease, using in vitro, ex vivo and in vivo infection
models and microarrays.
Introduction
The polymorphic yeast Candida albicans is the most im-
portant fungal pathogen of humans. However, as well as
being a successful pathogen, C. albicans exists as part of the
normal human microbiota and is not found in environ-
mental reservoirs such as soil. In the context of the rest
of the fungal kingdom, this lifestyle is remarkable: of the
estimated 1.5 million (over 100 000 of which have been
confirmed) fungal species, only a very small percentage
(150–200 species) is capable of causing infections in humans.
Some of these species are specialized for infections of the skin
(dermatophytes or Malassezia species), while others are also
able to cause more serious, systemic infections. Most of the
major pathogens in the latter group (such as Aspergillus
fumigatus, Cryptococcus neoformans and Histoplasma capsula-
tum) are environmental fungi, capable of exogenously infecting
susceptible individuals. Candida albicans on the other hand is,
under normal circumstances, a benign colonizer of human
mucosal surfaces and, therefore, highly specialized for life on or
within the human host. However, certain alterations to the host
environment can result in the transition from a commensal to a
pathogenic phase, permitting infection of virtually every organ
of the human body and resulting in severe infections. Even a
mildly compromised immune system or a minor imbalance of
the microbiota can be sufficient for C. albicans to cause
infections of the skin or of mucosal surfaces. These superficial
infections are extremely common – for example, c. 75% of all
women experience vulvovaginal candidosis during their life-
time and a significant proportion suffer from recurrent infec-
tions. In addition, oral and oesophageal candidosis are
particularly common in HIV-positive individuals and, without
intervention with highly active antiretroviral therapy, occurred
in up to 90% of HIV patients (Calderone, 2002). The severity of
candidosis, however, increases dramatically in patient popula-
tions with predisposing factors such as severely impaired
FEMS Yeast Res 9 (2009) 688–700c� 2009 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
YEA
ST R
ESEA
RC
H

immunity (e.g. neutropenia), cancer (e.g. leukaemia), disrup-
tion of natural barriers (e.g. by burn injury or disruption of gut
mucosal barriers by abdominal surgery), the presence of
indwelling catheters, dialysis or solid organ transplantation
(Ruhnke & Maschmeyer, 2002; Perlroth et al., 2007). These
mostly hospital-acquired (nosocomial) bloodstream and inva-
sive infections are life-threatening diseases and candidaemia is
now the third most common form of nosocomial bloodstream
infection responsible for 9% of such infections (Wisplinghoff
et al., 2004; Perlroth et al., 2007). Not only are invasive Candida
infections common in intensive care units of hospitals, they are
also difficult to treat (Perlroth et al., 2007). Even with first-line
antifungal therapy, disseminated candidosis has an attributable
mortality of up to 50% in some patient populations, depending
on the underlying illness (Perlroth et al., 2007). The mortality of
severe sepsis caused by Candida is 4 50%, and is therefore
higher than the mortality from sepsis due to any bacterium,
including Pseudomonas aeruginosa. Invasive candidosis is usual-
ly due to fungal entry into the blood resulting in candidaemia
or haematogenous dissemination to internal organs. Candida
albicans can also cause candiduria, perotinitis, endocarditis,
pericarditis, endophthlamitis, meningitis and pneumonia (Cal-
derone, 2002). It is this extraordinary ability of C. albicans to
successfully infect virtually every anatomical site of the human
host that makes it such an important organism to study – both
from a medical and biological perspective.
The exceptional adaptability of C. albicans is mediated by
rapid alterations in gene expression in response to environ-
mental stimuli, such as changes in nutrient availability, pH,
osmolarity, temperature or attack by cells of the immune
system (Fradin et al., 2003; Prigneau et al., 2003; Rubin-
Bejerano et al., 2003; Bensen et al., 2004; Hube, 2004; Brown
et al., 2007; Kumamoto, 2008) and this transcriptional
flexibility can be monitored with tools such as microarrays.
Indeed, this decade has been witness to numerous genome-
wide studies on C. albicans gene expression. These, mainly
in vitro studies have described the transcriptional response
of C. albicans to different environmental conditions such as
the presence of inducers of hyphal development, various
stresses or treatment with antifungal agents (Nantel et al.,
2002; Enjalbert et al., 2006). They have also been essential in
the identification of targets of regulatory circuits (e.g. the
regulon repressed by Tup1 – Garcia-Sanchez et al., 2005).
More recently, we and other groups have used microarray
technology in the context of C. albicans infection biology to (1)
capture a genome-wide portrait of the transcriptome that
mirrors the environmental conditions (thus enabling us to
examine the pathogenic processes in detail), (2) identify
known genes, signalling pathways and transcription factors
involved in pathogenesis, (3) identify new patterns of gene
expression and (4) identify previously uncharacterized genes
(unknown function genes) that may be associated with infec-
tion. As described above, the range of tissue types that
C. albicans can infect is extensive; however, generally infections
can be grouped into two major types: mucosal or systemic
candidosis (also referred to as superficial or invasive infections,
respectively). In our laboratory, we use a cyclical approach for
dissection of the mechanisms of host–pathogen interactions
during distinct types of C. albicans infections and identifying
genes involved in the infection process. First, infection models
are established and investigated using a combination of micro-
scopic and biochemical techniques to determine the temporal
phases of infection. Based on this, appropriate time points are
selected and global transcriptional profiling performed. From
the expression data, genes of interest are selected, deleted and
the resultant knockout mutants tested for attenuation in the
infection model in question. If attenuated, further in-depth
analysis of gene functions will be performed. In this review, we
describe the molecular dissection of three distinct types or
stages of infections covering both superficial and invasive
disease: oral candidosis (mucosal), bloodstream and liver
infection (systemic/invasive).
Mucosal infections: oral candidosis
Recently, we have focused on characterizing oral candidosis
and the molecular mechanisms underpinning this type of
infection using a combination of different in vitro infection
models [e.g. the reconstituted human oral epithelium (RHE)
and monolayers of oral epithelial cells] and by comparing
these data with results generated from clinical samples (mainly
from HIV-positive patients suffering from oral candidosis).
Based on our observations made in the experimental infection
models, we identified three different substages during the
pathogenesis of oral infections: an early/colonization phase,
characterized by adhesion of the fungus to the upper layers of
the host tissue and fungal proliferation; an invasion phase,
associated with hyphal formation and penetration of the
upper cell layers of the oral tissue; and a late phase, associated
with substantial tissue destruction (Fig. 1a and b). By combin-
ing histological analysis, scanning electron microscopy, cellu-
lar cytotoxicity measurements and global gene-expression
analysis, we were able to study these different stages of oral
candidosis at both the cellular and molecular level. Of course,
naturally, the time scale of oral candidosis is different. Here,
initial colonization can precede the onset of infection by many
years and it is only upon compromise of the host that oral
candidosis manifests. However, despite the artificiality of the
RHE model, it appears to at least partially mimic the clinical
setting as the majority of genes constitutively expressed in the
RHE were also upregulated by C. albicans infecting the oral
cavity of HIV-positive patients (Zakikhany et al., 2007).
Early-phase oral candidosis
The early phase (0–3 h) in the experimental setting repre-
sents the establishment of infection: inoculated C. albicans
FEMS Yeast Res 9 (2009) 688–700 c� 2009 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
689Identifying infection-associated genes of Candida albicans
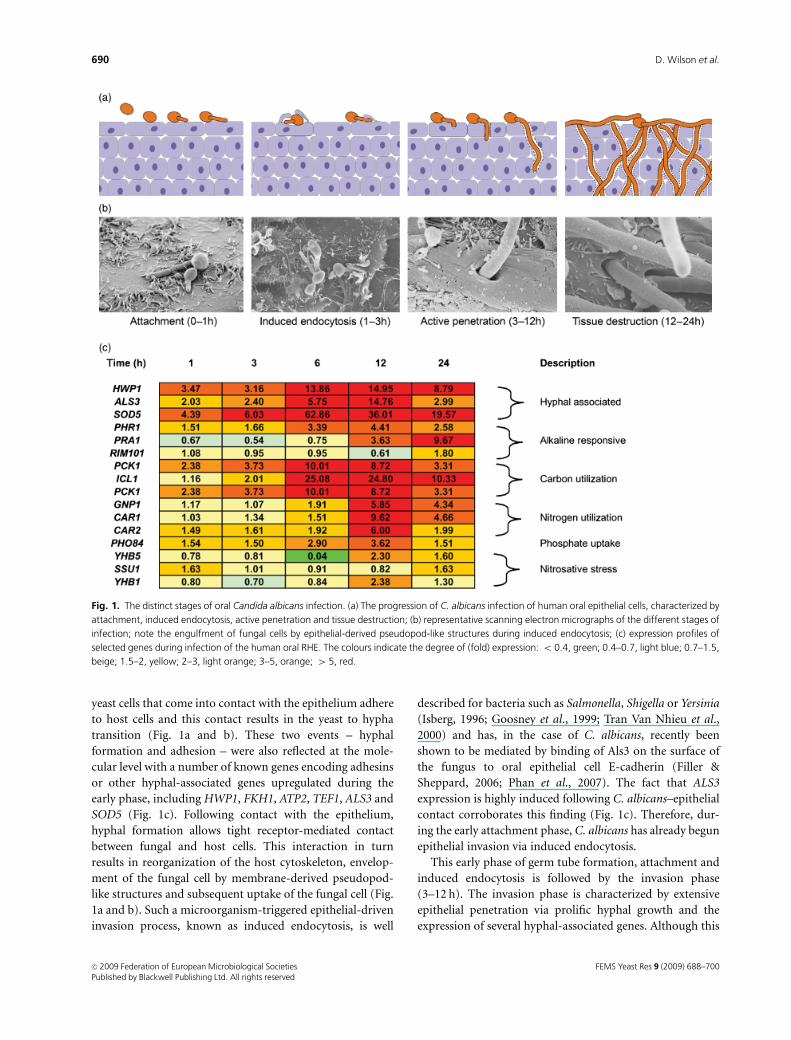
yeast cells that come into contact with the epithelium adhere
to host cells and this contact results in the yeast to hypha
transition (Fig. 1a and b). These two events – hyphal
formation and adhesion – were also reflected at the mole-
cular level with a number of known genes encoding adhesins
or other hyphal-associated genes upregulated during the
early phase, including HWP1, FKH1, ATP2, TEF1, ALS3 and
SOD5 (Fig. 1c). Following contact with the epithelium,
hyphal formation allows tight receptor-mediated contact
between fungal and host cells. This interaction in turn
results in reorganization of the host cytoskeleton, envelop-
ment of the fungal cell by membrane-derived pseudopod-
like structures and subsequent uptake of the fungal cell (Fig.
1a and b). Such a microorganism-triggered epithelial-driven
invasion process, known as induced endocytosis, is well
described for bacteria such as Salmonella, Shigella or Yersinia
(Isberg, 1996; Goosney et al., 1999; Tran Van Nhieu et al.,
2000) and has, in the case of C. albicans, recently been
shown to be mediated by binding of Als3 on the surface of
the fungus to oral epithelial cell E-cadherin (Filler &
Sheppard, 2006; Phan et al., 2007). The fact that ALS3
expression is highly induced following C. albicans–epithelial
contact corroborates this finding (Fig. 1c). Therefore, dur-
ing the early attachment phase, C. albicans has already begun
epithelial invasion via induced endocytosis.
This early phase of germ tube formation, attachment and
induced endocytosis is followed by the invasion phase
(3–12 h). The invasion phase is characterized by extensive
epithelial penetration via prolific hyphal growth and the
expression of several hyphal-associated genes. Although this
Fig. 1. The distinct stages of oral Candida albicans infection. (a) The progression of C. albicans infection of human oral epithelial cells, characterized by
attachment, induced endocytosis, active penetration and tissue destruction; (b) representative scanning electron micrographs of the different stages of
infection; note the engulfment of fungal cells by epithelial-derived pseudopod-like structures during induced endocytosis; (c) expression profiles of
selected genes during infection of the human oral RHE. The colours indicate the degree of (fold) expression: o 0.4, green; 0.4–0.7, light blue; 0.7–1.5,
beige; 1.5–2, yellow; 2–3, light orange; 3–5, orange; 4 5, red.
FEMS Yeast Res 9 (2009) 688–700c� 2009 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
690 D. Wilson et al.

phase is associated with considerable invasion of the epithe-
lium, substantial tissue damage is not observed until the late
phase (12–24 h), suggesting that initial invasion alone is
not sufficient to cause damage and not the only factor
contributing to tissue destruction. Invasion during the mid
and late phases is mediated predominantly by active pene-
tration (Fig. 1a and b). Active penetration is an invasion
mechanism distinct from induced endocytosis as it does not
rely on the host’s cellular machinery, but exclusively on
fungal attributes possibly including physical pressure ap-
plied by the advancing hyphal tip and the secretion of
extracellular hydrolases, which have been presumed to assist
in the invasion process via the degradation of host cell
components (Schaller et al., 2005).
Late-phase oral candidosis
Finally, the late phase, characterized by substantial tissue
destruction, was – at the transcriptional level – associated
with numerous adaptive responses (Fig. 1c). By 24 h,
expression of the two alkaline-responsive genes, PHR1 and
PRA1, was induced, as was that of the alkaline-responsive
transcription factor, RIM101, suggesting that the fungal cells
were in an environment of neutral-alkaline pH. Although it
is possible that the induction of these genes was influenced
by the pH of the RHE maintenance medium, it is probable
that these genes are truly induced during oral epithelial
tissue destruction, as their transcript levels were 4 1.5-fold
upregulated in patient samples. Candida albicans appears to
face a glucose-poor environment in the oral tissue as
indicated by the upregulation of the glucose and maltose
transporter genes HGT12 and MAL31 and key components
of the gluconeogenesis pathway and glyoxylate cycle (PCK1,
MLS1 and ICL1). A number of genes involved in amino acid
sensing and transport (GNP1, CAR1 and CAR2) were also
upregulated suggesting that C. albicans cells sense a nitro-
gen-poor environment on or within the oral tissue. In
addition, strong induction of the high-affinity phosphate
transporter PHO84 suggested that ready access to phosphate
sources was also limited in this tissue. Apart from the
limitation of certain nutrients, the only other clear stress
condition encountered by the fungus appeared to be nitro-
sative stress as indicated by the upregulation of the marker
genes YHB5, SSU1 and YHB1 involved in detoxification of
nitric oxide (Hromatka et al., 2005). It is known that
epithelial cells produce nitrogen monoxide radicals as part
of their innate immune response against microorganisms
and it would therefore appear that the experimental RHE
model is capable of mounting at least this innate defence
mechanism against C. albicans infection. Surprisingly, many
well-known genes involved in iron acquisition – a major
virulence determinant of virtually all pathogenic microor-
ganisms (Andrews et al., 2003; Howard, 2004; Sutak et al.,
2008) – were not upregulated in oral RHE tissue. It is
possible that the experimental set-up of the RHE allowed
exposure of C. albicans to unnaturally high levels of iron
from the surrounding medium. Alternatively, C. albicans
may be utilizing a novel iron source during invasion of this
tissue. Our group has recently demonstrated that C. albicans
can utilize iron from host ferritin during oral infections and
have shown that this event is mediated by the multifunctional
cell surface protein Als3 (Almeida et al., 2008). Given the high
expression of ALS3 during our models of oral infection, this
novel iron-acquisition strategy is a likely possibility.
Molecular analysis of an oral candidosis-associated gene
Among the genes that were upregulated during oral infec-
tion were a substantial number with no known function or
with no homologue in the brewer’s yeast Saccharomyces
cerevisiae. We reasoned that these unknown function infec-
tion-associated genes constitute good candidates for novel
virulence factors in C. albicans. One such gene, orf19.7561
(renamed EED1) has no obvious homologues in any other
sequenced organism and was upregulated in patient samples
and during both the early and the late phases of RHE
infection. Given the expression profile of this gene we
predicted that it might play a role (1) during the onset of
and (2) in the maintenance/persistence of infections of the
oral cavity. For functional characterization of the role of
EED1 during oral infections, both copies of the gene were
deleted. The resultant eed1D mutant had severe hyphal-
formation defects, growing as yeast or short chains of
pseudohyphae under standard laboratory hyphal-induction
conditions. Growth in the presence of very strong stimuli
(e.g. RPMI with 10% serum) resulted in elongated pseudo-
hyphal germ tube formation of eed1D; however, following
extended incubation time (5 h), the eed1D cells were unable
to maintain filamentous growth and switched back to the
yeast morphology. These results suggested that EED1 is
required for both the initiation and maintenance of fila-
mentous growth. As hyphal development is a prerequisite
for invasion of oral epithelial cells, it was predicted that the
eed1D mutant would be highly attenuated in our oral
infection models. Surprisingly, despite the observed in vitro
filamentation defects, upon infection of TR146 oral cells the
eed1D mutant cells switched to filamentous growth, reinfor-
cing the view that fungal contact with epithelial cells itself is
a potent inducer of filamentation and bypasses the require-
ment for EED1. Moreover, these filaments were able to
invade the epithelial cells via induced endocytosis. Despite
initial filamentation and invasion, by 24 h, eed1D had
reverted to the yeast morphology and existed within intrae-
pithelial inclusion bodies while wild-type cells disseminated
throughout the epithelial tissue via extensive hyphal
FEMS Yeast Res 9 (2009) 688–700 c� 2009 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
691Identifying infection-associated genes of Candida albicans

formation. The gene was therefore named EED1 for epithe-
lial escape and dissemination. Although other genes (EFG1
and CPH1) have been shown to be required for escape from
professional phagocytic cells, such as macrophages, EED1 is
the first example of a fungal gene required for escape from
and dissemination within oral epithelial tissue.
Systemic infections: survival in the bloodand liver invasion
Systemic infections are characterized by three major events:
dissemination via the bloodstream followed by escape from
the bloodstream and the infection of deep-seated organs. In
order to study these events, we have established bloodstream
and liver infection models. A number of mechanisms as to
how C. albicans enters the bloodstream have been proposed;
these include so-called ‘natural’ routes, such as via the
penetration of epithelial cells at mucosal surfaces, or iatro-
genic (artificial) routes, such as implantation of medical
devices, surgery, trauma or depletion of the natural micro-
bial flora by antibiotic treatment (Mavor et al., 2005). Once
inside the bloodstream, C. albicans is able to disseminate
and can potentially infect almost every organ of the host;
however, for this to occur, the fungus must first survive
within the bloodstream and then escape via traversal of the
endothelium.
Survival in blood
The bloodstream is a harsh environment for any pathogenic
microorganism due to the presence of numerous immu-
noactive cells and molecules; however, in order to cause
systemic infections, pathogenic microorganisms must pos-
sess mechanisms to resist attack by the immune system. In
order to investigate the fungal response to the hostile
environment of the blood, we incubated C. albicans with
human blood and measured the transcriptional response of
the fungus over a time course experiment (10, 20, 30 and
60 min) using microarrays compromising 2002 genes (Fra-
din et al., 2003). In this study, we showed that C. albicans
rapidly adapts to the blood environment. Such rapid adap-
tation relies on the expression of a distinct subset of genes
and translation into the corresponding proteins necessary to
meet the requirements of the new environment. This was
reflected by the strong upregulation of genes related to
protein synthesis at 10 min. This early adaptation event was
followed by the upregulation of genes involved in the
glyoxylate cycle, fermentation, glycolysis and response to
oxidative stress. In addition, several known hyphal-asso-
ciated genes were upregulated upon exposure to blood, in
agreement with microscopy observations that 42% of cells
had undergone the yeast to hypha transition during the time
course of the experiment (Fig. 2a). The fact that genes of the
glyoxylate cycle were upregulated at the same time as genes
involved in glycolysis is surprising: in general, the glycolytic
pathway and the glyoxylate cycle are utilized in the absence
or presence of carbohydrates, respectively, and not at the
same time by the same cell. One explanation for this is that
the fungal cells existed in at least two distinct subpopula-
tions, one with and one without ready access to sugars. This
split-population hypothesis may also be supported by the
morphogenic heterogeneity of fungal cells exposed to blood
and may be explained by the fact that blood consists of a
heterogeneous mixture of different cell types, which may act
differentially on distinct fungal cells.
In order to dissect which blood factors are involved in
combating C. albicans, and which fungal factors may resist
this assault, we further analysed the cellular and transcrip-
tional response of C. albicans to whole blood and to various
blood fractions enriched in particular host cell types (Fradin
et al., 2005). One major virulence trait – the yeast to hypha
transition – is also presumed to aid in C. albicans escape
from the bloodstream by assisting in traversal of the
endothelial lining of blood vessels (Filler & Sheppard,
2006) and has been shown to mediate escape following
phagocytosis by macrophages (Lorenz & Fink, 2001). The
morphology of C. albicans exposed to the various blood
fractions for 30 min was therefore determined. In blood
fractions lacking neutrophils (erythrocyte, monocyte/
lymphocyte and plasma fractions) most cells (80–85%)
formed germ tubes. However, exposure of C. albicans to the
polymorphonuclear (PMN) neutrophil fraction almost
completely blocked hyphal development with 96.5% of cells
remaining in the yeast morphology (Fig. 2a). This dominant
effect of PMNs on the morphology of C. albicans was also
reflected at the transcriptional level: cluster analysis showed
that cells incubated in whole blood or in the PMN fraction
shared a similar profile that was distinct from cells incubated
in the presence of plasma, erythrocytes, monocytes or in
whole blood depleted of PMNs.
In the absence of neutrophils, numerous genes associated
with protein synthesis, glycolysis and hyphal formation
were expressed, suggesting that cells were metabolically
active and in concordance with observation that the
majority of cells had undergone the yeast to hypha transi-
tion. In stark contrast, in the presence of neutrophils, fungal
cells underwent growth arrest, and hyphal morphogenesis
was almost completely blocked. Nutrient starvation cer-
tainly appeared to contribute to the observed growth arrest.
The environmental nitrogen level of cells exposed to neu-
trophils was low as indicated by the upregulation of the
ammonium permeases MEP2 and MEP3. Furthermore, the
amino acid starvation-responsive transcriptional regulator
GCN4, as well as several genes associated with arginine,
leucine, lysine and methionine biosynthetic pathways were
induced as described previously by Rubin-Bejerano et al.
(2003). Vacuolar proteases (Prb1, Prb2 and Apr1) and
FEMS Yeast Res 9 (2009) 688–700c� 2009 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
692 D. Wilson et al.

carboxypeptidases (Prc1 and Prc2) – known to be involved
in the utilization of endogenous nitrogen sources – were also
upregulated in response to neutrophils. In addition to this
clear nitrogen-starvation response, C. albicans also appeared
to face a carbohydrate-poor environment, as genes associated
with glycolysis were downregulated while components of the
glyoxylate cycle were strongly induced (Fig. 2c).
One of the proposed antimicrobial mechanisms of neutro-
phils is the production of reactive oxygen species, which
contribute to the killing of pathogenic microorganisms by
attacking multiple cellular components such as DNA, proteins
and lipids. It appears that neutrophils exert substantial
oxidative stress on C. albicans as a number of genes involved
in detoxification of reactive oxygen species – such as super-
oxide dismutases, catalase, glutathione peroxidase, glutathione
reductase, glutathione S-transferase and thioredoxin – were
strongly induced in response to incubation with neutrophils
(Fig. 2c). To further investigate the role of oxidative stress on
C. albicans during interactions with neutrophils we fused a
green fluorescent protein (GFP) reporter to the oxidative
stress-responsive SOD5 promoter (Martchenko et al., 2004).
The SOD5 reporter was found to be induced in yeast cells that
were either attached to or phagocytosed by neutrophils. This
finding suggests that neutrophils elicit an oxidative stress
response, which does not rely on phagocytosis of the fungal
cell. Finally, it was shown that sod5D mutant cells had reduced
survival following incubation with neutrophils, reinforcing the
view that an appropriate oxidative stress response is critical for
Fig. 2. Interaction of Candida albicans with blood. (a) The effect of blood or blood fractions on C. albicans morphology, note the heterogenous
morphology of fungal cells in whole blood, repression of hyphal formation in the PMN (neutrophil) fraction and hyphal development in the MNC
(monocyte/lymphocyte) and red blood cells (RBC) (erythrocyte) fractions; (b) representative micrographs of human blood cells; (c) expression profiles of
selected genes following 30-min incubation with indicated blood component. The colours indicate the degree of (fold) expression: o 0.4, green;
0.4–0.7, light blue; 0.7–1.5, beige; 1.5–2, yellow; 2–3, light orange; 3–5, orange.
FEMS Yeast Res 9 (2009) 688–700 c� 2009 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
693Identifying infection-associated genes of Candida albicans
fungal survival in the hostile environment of the blood and
subsequent dissemination throughout the body.
Liver invasion
Having survived the bloodstream for a certain time period,
C. albicans cells must escape from the circulation via
traversal across the endothelial lining of blood vessels in
order to avoid the killing activities of blood cells. Following
escape from the blood stream, the fungus has the potential
to infect virtually every internal organ. As a model of
infection of a deep-seated organ we analysed C. albicans
invasion of the liver. In order to identify genes associated
with tissue invasion of organs, we designed two comparative
experiments (Thewes et al., 2007a, b): (1) comparison of
C. albicans gene expression following in vivo intraperitoneal
infection of mouse liver and ex vivo infection of perfused pig
liver to identify genes commonly associated with infection
of this organ; (2) analysis of genes expressed in the liver
model by the invasive strain (SC5314) but not by the
noninvasive strain (ATCC10231) to more stringently define
those genes actually involved in the invasion process, as
opposed to those that are simply more highly expressed in
the host than in broth culture. Preliminary histological
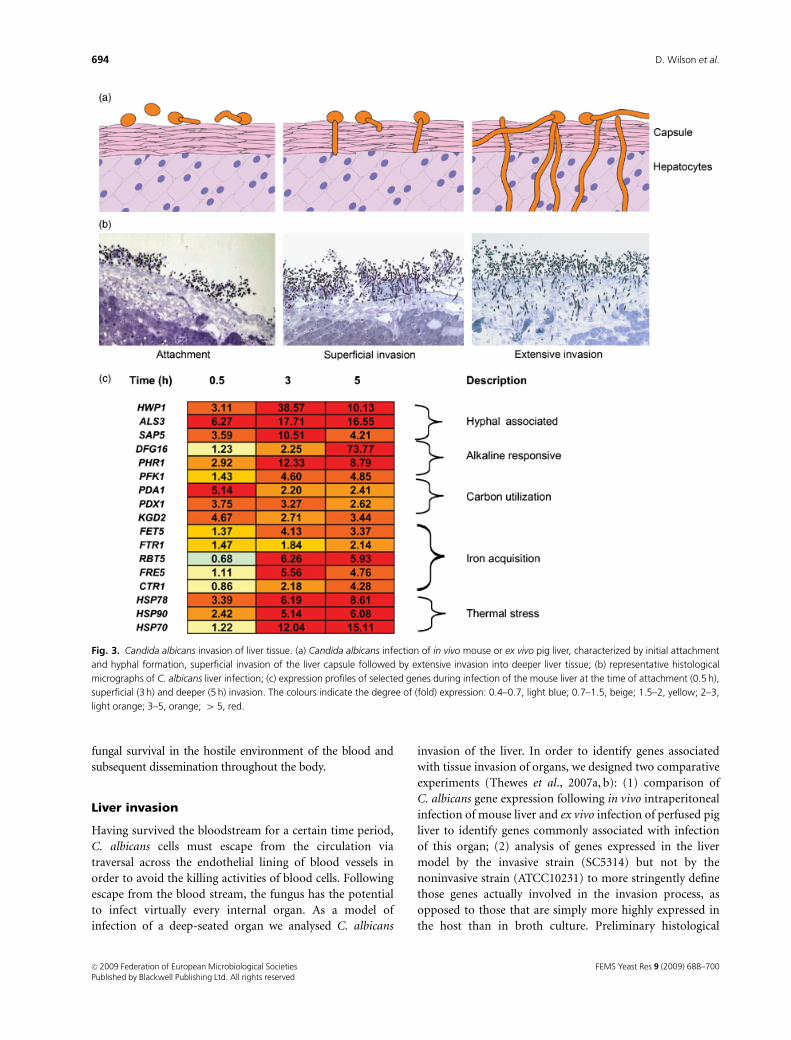
Fig. 3. Candida albicans invasion of liver tissue. (a) Candida albicans infection of in vivo mouse or ex vivo pig liver, characterized by initial attachment
and hyphal formation, superficial invasion of the liver capsule followed by extensive invasion into deeper liver tissue; (b) representative histological
micrographs of C. albicans liver infection; (c) expression profiles of selected genes during infection of the mouse liver at the time of attachment (0.5 h),
superficial (3 h) and deeper (5 h) invasion. The colours indicate the degree of (fold) expression: 0.4–0.7, light blue; 0.7–1.5, beige; 1.5–2, yellow; 2–3,
light orange; 3–5, orange; 4 5, red.
FEMS Yeast Res 9 (2009) 688–700c� 2009 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
694 D. Wilson et al.

analysis revealed that in both infection models, the infection
process of the invasive strain was characterized by initial
attachment and hyphal formation followed by penetration
of the liver capsule and invasion into the tissue by hyphal
cells (Thewes et al., 2007b; Fig. 3a and b).
Based on the expression profile of so-called ‘marker
genes’, a number of inferences could be drawn about
the nutritional state of the environment encountered by
C. albicans during liver invasion (Thewes et al., 2007a;
Fig. 3c). For example, as opposed to the situation in the oral
cavity, there appeared to be sufficient access to sugar for
utilization as a carbon source as indicated by upregulation
of PFK1 (encoding a key enzyme of glycolysis), PDA1 and
PDX1 (involved in acetyl-CoA biosynthesis) and KGD1 and
KGD2 (encoding key enzymes of the tricarboxylic acid
cycle). Similarly, there appeared to be sufficient levels of
nitrogen in the liver as genes associated with amino acid
starvation were not upregulated. In spite of this apparent
abundance of carbon and nitrogen sources, C. albicans
appeared to face severe iron limitation upon liver invasion
as indicated by the upregulation of FET5, FTR1, ZRT1,
CFL1, RBT5, FRE5 and CTR1 – all genes associated with an
iron-poor environment or with iron acquisition. There did
not appear to be any specific response to oxidative, nitrosa-
tive or osmotic stress in the liver, although genes associated
with general/thermal stress such as HSP78, HSP90, HSP104,
HSP12 and HSP70 were upregulated. Whether this is due to
actual thermal stress or cross-protection against an as-yet
unknown stress in the liver remains to be determined.
Upregulation of well-known hyphae-associated genes such
as SAP5, ALS3 and HWP1 was in accordance with the
histological observation that cells were growing predomi-
nantly in the hyphal morphology. Finally, upregulation of
the alkaline-responsive PHR1 indicated that the majority of
cells encountered a neutral-alkaline pH.
Molecular analysis of a gene involved in invasivecandidosis
The next step of this study involved dissection of the
transcriptional data to identify genes intimately associated
with the process of liver tissue invasion. To focus our analysis
in this direction, we looked for genes that were upregulated
by the invasive SC5314 during both mouse (3 and 5 h
postinfection) and pig (12 h postinfection) liver invasion but
not by the noninvasive strain, ATCC10231. This detailed
analysis yielded five genes, one of which (DFG16) was selected
for further analysis. DFG16 encodes a member of the PalH/
RIM21 super-family, which constitutes putative pH sensors
and has been shown to function in the Rim101 pathway
(Barwell et al., 2005). To characterize the function of this
gene, both alleles were deleted and the resultant dfg16Dmutant tested. In accordance with the protein’s predicted role
as a pH sensor, dfg16D behaved normally at acidic pH but was
unable to grow under iron- or phosphate-limited conditions
at alkaline pH. Moreover, dfg16D had reduced osmotic stress
tolerance at alkaline, but not acidic, pH and failed to form
filaments at pH 8. Finally, the virulence of dfg16D was
attenuated in a mouse model of haematogenously dissemi-
nated candidosis (Thewes et al., 2007a) and dfg16D cells had
reduced potential to invade liver tissue following intraperito-
neal infection (our unpublished data).
The fact that transcript levels of DFG16, encoding a
putative pH sensor, are increased during liver invasion
underscores the importance of environmental pH sensing
and adaptation during the progression of systemic candido-
sis. Unlike environmental human fungal pathogens, which
receive clear host-associated signals to initiate infection (e.g.
a shift to 37 1C), C. albicans must be able to dramatically
reprogramme its behaviour based on more subtle environ-
mental cues. Probably the most extensively studied trait of
C. albicans, the yeast to hypha transition, is under the
control of an extensive network of signalling pathways,
which integrate the receipt of a wide range of environmental
signals (temperature, pH, oxygen, CO2, nutrients and physical
contact to name only a few) to control cellular morphology.
Why must C. albicans sense such a vast array of environmental
signals to determine its morphology? Firstly, unlike many
environmental pathogenic fungi, which form either non-
pathogenic/saprophytic or host-associated/pathogenic mor-
phologies, the pathogenicity of C. albicans relies on the
reversible yeast to hypha transition (mutants unable to form
yeast also have reduced virulence). Secondly, because
C. albicans is continuously in contact with the host (even in
the nonpathogenic stage as commensal) and, therefore, in an
environment of physiological temperature, the formation of a
single morphological state in response to temperature alone
would be inappropriate. Thirdly, although the gross morphol-
ogy of C. albicans is dependent on certain, sometimes quite
different, combinations of environmental signals, any given
subset of signals may specifically result in the expression of a
different subset of genes not strictly coexpressed with a given
morphology; these discrete transcriptomes reflecting the given
requirements encountered at particular anatomical niches: for
example, RBT5, encoding the cell surface-localized haemoglo-
bin-binding protein (Weissman & Kornitzer, 2004), is highly
expressed by hyphal cells invading liver tissue but not by
hyphal cells invading oral tissue.
Given the highly dynamic response of C. albicans to its
environment, combined with the diverse niches it occupies, it
is not surprising that the genome of this fungus contains a
large number of genes encoding described and putative
sensors. The correct sensing of – and response to – environ-
mental signals is crucial and relies on the expression of
relevant sensor-encoding genes in a given microenvironment.
The reduced virulence of dfg16D demonstrates that
FEMS Yeast Res 9 (2009) 688–700 c� 2009 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
695Identifying infection-associated genes of Candida albicans

perturbations in the sensing of a single environmental factor
(pH) can block the pathogenicity of C. albicans and illustrates
the delicacy of the cross-talk between the host and the
pathogen.
Discussion and outlook: the future ofC. albicans infection models andidentification of infection-associatedgenes
In summary, we have presented three examples of how,
using carefully designed infection models combined with
global gene expression analysis, we can confirm presumed
features of pathogenicity, identify genes that are expressed
during infection and discover novel aspects of host–patho-
gen interactions during representative C. albicans infections.
The transcriptome of C. albicans as a tool toexplore the physiological environment duringinfection
One possible application of genome-wide profiling of a
pathogenic microorganism during infection is the concept
of the transcriptome as a biosensor, which may enable us
to monitor the physiological conditions encountered by
the microorganism. For example, we verified that when
C. albicans was exposed to blood, neutrophils elicit a strong
oxidative stress response. This was not the case in the liver or
oral cavity, where general or nitrosative stress responses,
respectively, dominated, thus demonstrating that C. albicans
faces diverse challenges depending on the anatomical niche
in which it finds itself. On the other hand, some features of
the host environment appeared to be common and there
was significant transcriptional overlap between different
infection types. For example, key components of the glyox-
ylate cycle were upregulated during both oral and blood-
stream (but not liver) infections. Similarly numerous
hyphal-associated genes were upregulated in both oral and
liver infections, but not during interaction with neutrophils.
Interestingly, very few genes were strongly induced under all
infection conditions, suggesting that, in general, there is no
general response to infection-associated growth and that the
transcriptome of C. albicans within the human host is
overall niche specific. This may be explained by the com-
mensal lifestyle of C. albicans. Environmental fungi receive
clear signals upon the onset of infection. For example,
synthesis of one of the major virulence factors of C. neofor-
mans, the polysaccharide capsule, is induced upon infection
via the fungus sensing a shift to an environment of iron
limitation and physiological temperature (Jung & Kronstad,
2008). Candida albicans on the other hand, outside of the
laboratory, is in constant contact with the human host and
while colonizing mucosal surfaces is constitutively exposed
to an environment of physiological temperature and limited
iron. Only disturbance of the normal bacterial microbiota
and/or a weakened immune system triggers activity asso-
ciated with infection and the colonization of new niches
within the human body. Given the niche specificity of
C. albicans, the principle of using the transcriptome as a
biosensor may be applicable for studying the conditions
faced by the fungus at diverse body sites. For example, based
on detailed in vitro studies, the expression of certain marker
genes such as PHR1, PRA1, PHR2, SOD5, CAT1, YHB1,
ICL1 and MEP2 may indicate alkaline or acidic pH, oxida-
tive stress, nitrosative stress, low glucose or nitrogen condi-
tions, respectively (De Bernardis et al., 1998; Sentandreu
et al., 1998; Lorenz & Fink, 2001; Martchenko et al., 2004;
Biswas & Morschhauser, 2005; Corvey et al., 2005; Hromat-
ka et al., 2005). However, further in vitro gene expression
experiments are needed to identify not only single marker
genes, but also sets of genes (regulons) and signatures
associated with a certain physiological situation. For exam-
ple, the identification of particular transcriptional signa-
tures associated with growth on defined nutrient sources
could be used to overlay the transcriptome during infection
and may actually pinpoint what nutrients are being utilized
in vivo. Bignell and colleagues (McDonagh et al., 2008) have
recently used such an approach to study the initiation of
infection during pulmonary aspergillosis. By comparing
transcriptional signatures obtained from specific in vitro
conditions to the expression pattern observed upon infec-
tion of the murine lung, they demonstrate that A. fumigatus
adaptation to the host is associated with iron limitation,
alkaline stress and nutrient deprivation.
Unknown function genes
One major obstacle in interpreting such global studies is the
overall lack of annotation of a large set of genes in the
C. albicans genome. Although d’Enfert et al. (2005) assigned
‘tentative functional assignments’ for 92% of the C. albicans
genome, the true number of genes with known function is
probably much lower. Braun et al. (2005) reported gene
ontology (GO) terms (excluding unknown function) for only
56% of the genome. Moreover, 19% of genes do not share
significant sequence homology with other organisms (Braun
et al., 2005). Because C. albicans is highly specialized to exist
in association with warm-blooded mammals, it is reasonable
to hypothesize that C. albicans possesses unique factors
involved in interactions with its host environment. Although
these host-interaction factors likely evolved to maintain a
commensal life-style, when the host environment undergoes
certain changes (antibiotic treatment, immune compromise,
etc.), they may become virulence factors involved in the onset
and progression of invasive infections. Therefore, one of our
central premises is that unknown function genes
FEMS Yeast Res 9 (2009) 688–700c� 2009 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
696 D. Wilson et al.

transcriptionally upregulated during infection constitute pro-
mising host-interaction/virulence factors, required for the
pathogenesis of C. albicans. For example, we have shown that
EED1, a previously undescribed gene with no obvious
sequence homology in any other sequenced organism is
induced during in vivo and in vitro oral infections and was
essential for dissemination within oral tissue (Zakikhany
et al., 2007). This example therefore illustrates that there exist
genes of unknown function, induced during infection, which
are essential for the pathogenesis of C. albicans. We are
therefore continuing to explore the role of unknown function
genes in the pathobiology of C. albicans.
Experimental design
A major issue to consider during the interpretation of
current transcriptional profiling studies is the choice of
control. In our group we cohybridize experimental RNA
against a ‘common control’ of RNA from cells grown in YPD
at 37 1C to mid-logarithmic phase. The rationale behind this
is that virtually all genes are expressed under this condition,
and many infection-associated genes should be more highly
expressed during infection than in batch culture. However,
care should be taken when using this approach as ‘upregula-
tion’ is dependent on the relative expression in the control
and thus genes expressed at the same or even lower level
in vivo compared with YPD may still be important for the
infection process.
Recent work by Walker et al. (2009) analysing gene
expression of C. albicans infecting the rabbit kidney further
highlighted certain issues associated with the choice of
control. The authors report a strong downregulation in the
expression of well characterized and reportedly ‘hyphae-
associated’ genes such as ALS3 and ECE1, despite histologi-
cal observations suggesting hyphal growth in the kidney
lesions. However, in this study the control chosen was RNA
from cells grown in RPMI 1640 at 37 1C and, as the authors
discuss, the downregulation of these genes does not neces-
sarily mean that they are not being transcribed in vivo, but
simply that the expression is significantly higher in the
control cells.
Two strategies can be used to circumvent these issues. The
first is to design a temporal experiment where the transcrip-
tome is captured at different time points during the infec-
tion process including the preculture. The dynamics of
expression can then be analysed over time, effectively
independent of the control signal. The second approach is
to perform parallel experiments where only one variable
is changed between the two, for example, the comparison of
C. albicans infecting reconstituted human epithelium either
with or without the presence of neutrophils (Schaller et al.,
2004). It should be noted that, for some experimental
designs, one colour labelling, may be more appropriate.
Readers are directed to two excellent reviews describing the
design principles of microarray experiments (Yang & Speed,
2002; Bryant et al., 2004).
Although microarrays represent the current ‘gold stan-
dard’ for unbiased global analysis of gene expression in
C. albicans, other technologies may be more appropriate for
certain applications. For example, a number of reporter
systems have been described that can monitor expression of
single genes during models of infection. For example, in vivo
expression can be measured very sensitively by placing the
site-specific recombinase FLP under control of the promoter
of interest. Using this method, transient expression of
the gene of interest can be detected during animal models
of infection at the single cell level (Staib et al., 1999). A
second single cell profiling approach involves fusion of the
promoter of interest to a GFP reporter. This method is
particularly effective for monitoring gene expression in a
mixed population of cells as demonstrated by Barelle et al.
(2006).
Finally, although the C. albicans genome now stands at its
21st assembly (van het Hoog et al., 2007), it is likely that
numerous small transcripts, potentially involved in patho-
genesis, have been overlooked and, therefore, not included
on current microarrays. This and the above-discussed
problems may be overcome by serial analysis of gene
expression of C. albicans during infection as has recently
been described for C. neoformans infecting the murine lung
(Hu et al., 2008). Using this approach, it will be possible to
determine which genes are actually expressed at high levels
during infection and to uncover truly novel transcripts
involved in the infection process.
The future of C. albicans infection models
The past half decade has witnessed the establishment of
numerous postgenomic technologies and refined infection
models for the study of C. albicans, and this review has
described the use of such technologies to characterize the
behaviour of this fungus in different niches of the human
host. However, this is only part of the story, as all forms of
candidosis (whether superficial or invasive infections) are
dependent on the host status and the host response must
always be considered as an element of pathogenesis
(Richardson & Rautemaa, 2009). Therefore, we need to also
investigate the specific host responses for the different types
of infection and the different stages of disease development
to understand the underlying mechanisms that contribute
to C. albicans infections.
Based on infection models such as those presented in this
review, it should be possible to extend current studies on
C. albicans host–pathogen interactions to more comprehen-
sively cover infection-associated parameters: both fungal
pathogenicity mechanisms and host immune factors. For
FEMS Yeast Res 9 (2009) 688–700 c� 2009 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
697Identifying infection-associated genes of Candida albicans

example, we have shown that in blood, it is neutrophils that
have the greatest impact on C. albicans morphology, viabi-
lity and gene expression (Fradin et al., 2003, 2005) lending
direct experimental evidence to clinical observations that
neutrophils are the primary defence mechanism against
systemic candidosis (Chauhan et al., 2006). As an example
of extension of such a study, Fradin et al. (2007) went on to
characterize the transcriptional response of neutrophils
exposed to C. albicans, identifying an enrichment in genes
involved in proinflammatory cell–cell signalling. And while
the innate immune system is vital for the first line of
protection against systemic candidosis, T-cell-mediated cel-
lular immunity is generally considered particularly impor-
tant at oral mucosa (reviewed in Saunus et al., 2008).
Recently, Schaller and colleagues (Schaller et al., 2004;
Weindl et al., 2007) have utilized the RHE model to show that
neutrophils induced a protective T-helper type 1 immune
response in human oral epithelial cells and that this protection
is directly mediated by TLR4 receptors.
The future of C. albicans infection biology will surely
extend such approaches as those reviewed here, simulta-
neously integrating analysis of pathogen and host factors to
assemble both a detailed and global picture of host–patho-
gen interactions. The inherent complexity of these interac-
tions means that systems biology must also play a role in
shaping our understanding of this delicate cross-talk.
Attaining such an inclusive portrait of candidosis will un-
questionably drive forward the discovery of novel diagnostic
tools and the development of effective antifungal therapies.
Acknowledgements
Our own research was supported by the Robert Koch-Institute
(RKI), the Hans-Knoell-Institute (HKI), the Deutsche
Forschungsgemeinschaft (DFG), the European Union (EU),
and the Federal Ministry of Education and Research (BMBF).
We would also like to thank the reviewers of this manuscript
for their helpful suggestions and Brice Enjalbert for interesting
discussions on data interpretation.
References
Almeida RS, Brunke S, Albrecht A, Thewes S, Laue M, Edwards
JE, Filler SG & Hube B (2008) The hyphal-associated adhesin
and invasin Als3 of Candida albicans mediates iron acquisition
from host ferritin. PLoS Pathog 4: e1000217.
Andrews SC, Robinson AK & Rodriguez-Quinones F (2003)
Bacterial iron homeostasis. FEMS Microbiol Rev 27: 215–237.
Barelle CJ, Priest CL, Maccallum DM, Gow NA, Odds FC &
Brown AJ (2006) Niche-specific regulation of central
metabolic pathways in a fungal pathogen. Cell Microbiol 8:
961–971.
Barwell KJ, Boysen JH, Xu W & Mitchell AP (2005) Relationship
of DFG16 to the Rim101p pH response pathway in
Saccharomyces cerevisiae and Candida albicans. Eukaryot Cell
4: 890–899.
Bensen ES, Martin SJ, Li M, Berman J & Davis DA (2004)
Transcriptional profiling in Candida albicans reveals new
adaptive responses to extracellular pH and functions for
Rim101p. Mol Microbiol 54: 1335–1351.
Biswas K & Morschhauser J (2005) The Mep2p ammonium
permease controls nitrogen starvation-induced filamentous
growth in Candida albicans. Mol Microbiol 56: 649–669.
Braun BR, van Het Hoog M, d’Enfert C et al. (2005) A human-
curated annotation of the Candida albicans genome. PLoS
Genet 1: 36–57.
Brown AJ, Odds FC & Gow NA (2007) Infection-related gene
expression in Candida albicans. Curr Opin Microbiol 10:
307–313.
Bryant PA, Venter D, Robins-Browne R & Curtis N (2004) Chips
with everything: DNA microarrays in infectious diseases.
Lancet Infect Dis 4: 100–111.
Calderone RA (2002) Candida and Candiosis. ASM Press,
Washington, DC.
Chauhan N, Latge JP & Calderone R (2006) Signalling and
oxidant adaptation in Candida albicans and Aspergillus
fumigatus. Nat Rev Microbiol 4: 435–444.
Corvey C, Koetter P, Beckhaus T, Hack J, Hofmann S, Hampel M,
Stein T, Karas M & Entian KD (2005) Carbon Source-
dependent assembly of the Snf1p kinase complex in Candida
albicans. J Biol Chem 280: 25323–25330.
De Bernardis F, Muhlschlegel FA, Cassone A & Fonzi WA (1998)
The pH of the host niche controls gene expression in and
virulence of Candida albicans. Infect Immun 66: 3317–3325.
d’Enfert C, Goyard S, Rodriguez-Arnaveilhe S et al. (2005)
CandidaDB: a genome database for Candida albicans
pathogenomics. Nucleic Acids Res 33: D353–D357.
Enjalbert B, Smith DA, Cornell MJ, Alam I, Nicholls S, Brown AJ
& Quinn J (2006) Role of the Hog1 stress-activated protein
kinase in the global transcriptional response to stress in the
fungal pathogen Candida albicans. Mol Biol Cell 17:
1018–1032. Epub 7 December 2005.
Filler SG & Sheppard DC (2006) Fungal invasion of normally
non-phagocytic host cells. PLoS Pathog 2: e129.
Fradin C, Kretschmar M, Nichterlein T, Gaillardin C, d’Enfert C
& Hube B (2003) Stage-specific gene expression of Candida
albicans in human blood. Mol Microbiol 47: 1523–1543.
Fradin C, De Groot P, MacCallum D, Schaller M, Klis F, Odds FC
& Hube B (2005) Granulocytes govern the transcriptional
response, morphology and proliferation of Candida albicans in
human blood. Mol Microbiol 56: 397–415.
Fradin C, Mavor AL, Weindl G, Schaller M, Hanke K, Kaufmann
SH, Mollenkopf H & Hube B (2007) The early transcriptional
response of human granulocytes to infection with Candida
albicans is not essential for killing but reflects cellular
communications. Infect Immun 75: 1493–1501. Epub 4
December 2006.
FEMS Yeast Res 9 (2009) 688–700c� 2009 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
698 D. Wilson et al.

Garcia-Sanchez S, Mavor AL, Russell CL, Argimon S, Dennison P,
Enjalbert B & Brown AJ (2005) Global roles of Ssn6 in Tup1-
and Nrg1-dependent gene regulation in the fungal pathogen,
Candida albicans. Mol Biol Cell 16: 2913–2925.
Goosney DL, Knoechel DG & Finlay BB (1999) Enteropathogenic
E. coli, Salmonella, and Shigella: masters of host cell
cytoskeletal exploitation. Emerg infect dis 5: 216–223.
Howard DH (2004) Iron gathering by zoopathogenic fungi.
FEMS Immunol Med Mic 40: 95–100.
Hromatka BS, Noble SM & Johnson AD (2005) Transcriptional
response of Candida albicans to nitric oxide and the role of the
YHB1 gene in nitrosative stress and virulence. Mol Biol Cell 16:
4814–4826.
Hu G, Cheng PY, Sham A, Perfect JR & Kronstad JW (2008)
Metabolic adaptation in Cryptococcus neoformans during early
murine pulmonary infection. Mol Microbiol 69: 1456–1475.
Hube B (2004) From commensal to pathogen: stage- and tissue-
specific gene expression of Candida albicans. Curr Opin
Microbiol 7: 336–341.
Isberg RR (1996) Uptake of enteropathogenic Yersinia by
mammalian cells. Curr Top Microbiol 209: 1–24.
Jung WH & Kronstad JW (2008) Iron and fungal pathogenesis: a
case study with Cryptococcus neoformans. Cell Microbiol 10:
277–284.
Kumamoto CA (2008) Niche-specific gene expression during
C. albicans infection. Curr Opin Microbiol 11: 325–330.
Lorenz MC & Fink GR (2001) The glyoxylate cycle is required for
fungal virulence. Nature 412: 83–86.
Martchenko M, Alarco AM, Harcus D & Whiteway M (2004)
Superoxide dismutases in Candida albicans: transcriptional
regulation and functional characterization of the hyphal-
induced SOD5 gene. Mol Biol Cell 15: 456–467.
Mavor AL, Thewes S & Hube B (2005) Systemic fungal infections
caused by Candida species: epidemiology, infection process
and virulence attributes. Curr Drug Targets 6: 863–874.
McDonagh A, Fedorova ND, Crabtree J et al. (2008) Sub-
telomere directed gene expression during initiation of invasive
aspergillosis. PLoS Pathog 4: e1000154.
Nantel A, Dignard D, Bachewich C et al. (2002) Transcription
profiling of Candida albicans cells undergoing the yeast-to-
hyphal transition. Mol Biol Cell 13: 3452–3465.
Perlroth J, Choi B & Spellberg B (2007) Nosocomial fungal
infections: epidemiology, diagnosis, and treatment. Med Mycol
45: 321–346.
Phan QT, Myers CL, Fu Y, Sheppard DC, Yeaman MR, Welch
WH, Ibrahim AS, Edwards JE & Filler SG (2007) Als3 is a
Candida albicans invasin that binds to cadherins and induces
endocytosis by host cells. PLoS Biol 5: e64.
Prigneau O, Porta A, Poudrier JA, Colonna-Romano S, Noel T &
Maresca B (2003) Genes involved in beta-oxidation, energy
metabolism and glyoxylate cycle are induced by Candida
albicans during macrophage infection. Yeast 20: 723–730.
Richardson M & Rautemaa R (2009) How the host fights against
Candida infections. Front Biosci 14: 4363–4375.
Rubin-Bejerano I, Fraser I, Grisafi P & Fink GR (2003)
Phagocytosis by neutrophils induces an amino acid
deprivation response in Saccharomyces cerevisiae and Candida
albicans. P Natl Acad Sci USA 100: 11007–11012.
Ruhnke M & Maschmeyer G (2002) Management of mycoses in
patients with hematologic disease and cancer – review of the
literature. Eur J Med Res 7: 227–235.
Saunus J M, Kazoullis A & Farah CS (2008) Cellular and
molecular mechanisms of resistance to oral Candida albicans
infections. Front Biosci 13: 5345–5358.
Schaller M, Boeld U, Oberbauer S, Hamm G, Hube B & Korting
HC (2004) Polymorphonuclear leukocytes (PMNs) induce
protective Th1-type cytokine epithelial responses in an in vitro
model of oral candidosis. Microbiology 150: 2807–2813.
Schaller M, Borelli C, Korting HC & Hube B (2005) Hydrolytic
enzymes as virulence factors of Candida albicans. Mycoses 48:
365–377.
Sentandreu M, Elorza MV, Sentandreu R & Fonzi WA (1998)
Cloning and characterization of PRA1, a gene encoding a novel
pH-regulated antigen of Candida albicans. J Bacteriol 180:
282–289.
Staib P, Kretschmar M, Nichterlein T, Kohler G, Michel S, Hof H,
Hacker J & Morschhauser J (1999) Host-induced, stage-
specific virulence gene activation in Candida albicans during
infection. Mol Microbiol 32: 533–546.
Sutak R, Lesuisse E, Tachezy J & Richardson DR (2008) Crusade
for iron: iron uptake in unicellular eukaryotes and its
significance for virulence. Trends Microbiol 16: 261–268.
Thewes S, Kretschmar M, Park H, Schaller M, Filler SG & Hube B
(2007a) In vivo and ex vivo comparative transcriptional
profiling of invasive and non-invasive Candida albicans
isolates identifies genes associated with tissue invasion. Mol
Microbiol 63: 1606–1628.
Thewes S, Reed HK, Grosse-Siestrup C, Groneberg DA, Meissler
M, Schaller M & Hube B (2007b) Haemoperfused liver as an ex
vivo model for organ invasion of Candida albicans. J Med
Microbiol 56: 266–270.
Tran Van Nhieu G, Bourdet-Sicard R, Dumenil G, Blocker A &
Sansonetti PJ (2000) Bacterial signals and cell responses
during Shigella entry into epithelial cells. Cell Microbiol 2:
187–193.
van het Hoog M, Rast TJ, Martchenko M et al. (2007) Assembly
of the Candida albicans genome into sixteen supercontigs
aligned on the eight chromosomes. Genome Biol 8: R52.
Walker LA, MacCallum DM, Bertram G, Gow NA, Odds FC &
Brown AJ (2009) Genome-wide analysis of Candida albicans
gene expression patterns during infection of the mammalian
kidney. Fungal Genet Biol 46: 210–219.
Weindl G, Naglik JR, Kaesler S, Biedermann T, Hube B, Korting
HC & Schaller M (2007) Human epithelial cells establish direct
antifungal defense through TLR4-mediated signaling. J Clin
Invest 117: 3664–3672.
Weissman Z & Kornitzer D (2004) A family of Candida cell
surface haem-binding proteins involved in haemin and
FEMS Yeast Res 9 (2009) 688–700 c� 2009 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
699Identifying infection-associated genes of Candida albicans

haemoglobin-iron utilization. Mol Microbiol 53:
1209–1220.
Wisplinghoff H, Bischoff T, Tallent SM, Seifert H, Wenzel RP &
Edmond MB (2004) Nosocomial bloodstream infections in US
hospitals: analysis of 24,179 cases from a prospective
nationwide surveillance study. Clin Infect Dis 39: 309–317.
Yang YH & Speed T (2002) Design issues for cDNA microarray
experiments. Nat Rev Genet 3: 579–588.
Zakikhany K, Naglik JR, Schmidt-Westhausen A, Holland G,
Schaller M & Hube B (2007) In vivo transcript profiling of
Candida albicans identifies a gene essential for interepithelial
dissemination. Cell Microbiol 9: 2938–2954.
FEMS Yeast Res 9 (2009) 688–700c� 2009 Federation of European Microbiological SocietiesPublished by Blackwell Publishing Ltd. All rights reserved
700 D. Wilson et al.






























