




www.elsevier.com/locate/jconrel
Journal of Controlled Release 96 (2004) 309–323
GEN
ED
ELIV
ER
Y
Hydrophobized dextran-spermine conjugate as potential vector for
in vitro gene transfection
Tony Azzama, Hagit Eliyahua, Arik Makovitzkia,Michal Linialb, Abraham J. Domba,*,1
aDepartment of Medicinal Chemistry and Natural Products, Faculty of Medicine, School of Pharmacy, Hebrew University of Jerusalem,
Jerusalem 91120, IsraelbDepartment of Biological Chemistry, Hebrew University of Jerusalem, Jerusalem 91905, Israel
Received 11 August 2003; accepted 13 January 2004
Abstract
Dextran polysaccharide was grafted by reductive-amination with mixtures of spermine and other natural/synthetic oligoamines
of two to four amine groups. The transfection efficiencies of the polycations thus obtained were assessed in various cell lines, and
found to depend on the spermine contents. Higher spermine ratios of grafted oligoamines resulted in high gene expression,
whereas low to negligible expressions were obtained with lower spermine contents. The effect was explained by spermine residues
which exhibit altered buffering capacity in comparison to other substituted oligoamines. Hydrophobization of dextran-spermine
(D-SPM) was achieved by treating the polymer with N-hydroxysuccinimide derivatives of cholesterol and fatty acids in a mixture
of water/THF. The degree of hydrophobization was in the range of 1–30%mol/mol (hydrophobicmoieties/primary amine) and the
coupling yields were >95% as determined by 1H-NMR. The oleate-modified D-SPM remarkably enhanced the gene expression in
serum rich media, in marked contrast to unmodified D-SPM which resulted with a drastic decrease in the transfection yields.
Modified D-SPM derivatives of other fatty acids and cholesterol showed improved transfection yields in comparison to
unmodified D-SPM, but to a lower extent when compared to oleate modification. The improvement in cell transfection was
attributed to oleate residues which probably play a role in increasing stability and uptake of polycation–DNA complexes.
D 2004 Published by Elsevier B.V.
Keywords: Dextran; Spermine; Reductive-amination; Gene delivery; Hydrophobic
1. Introduction
DNA can be delivered into the cell nucleus using
physical means or using specific carriers that carry the
0168-3659/$ - see front matter D 2004 Published by Elsevier B.V.
doi:10.1016/j.jconrel.2004.01.022
* Corresponding author. Tel.: +972-2-675-7573; fax: +972-2-
675-8959.
E-mail address: [email protected] (A.J. Domb).1 A.J.D. is affiliated with the David R. Bloom Center for
Pharmacy and with the Alex Grass Center for Drug Design and
Synthesis at the Hebrew University of Jerusalem.
genes into the cells for gene expression. Of the
various methods developed for delivering genes, gene
carriers have been extensively investigated as trans-
fecting agents for therapeutic genes in Gene Therapy.
Gene carriers are divided into two main groups: viral
carriers where the DNA to be delivered is inserted into
a virus, and cationic molecular carriers that form
electrostatic interactions with DNA. Successful gene
therapy depends on the efficient delivery of genetic
materials into the cells nucleus and its effective

T. Azzam et al. / Journal of Controlled Release 96 (2004) 309–323310
GEN
ED
ELIV
ER
Y
expression within these cells [1]. Although at present,
the in vivo expression levels of synthetic molecular
gene vectors are lower than for viral vectors and gene
expression is transient, these vehicles are likely to
present several advantages including safety, low im-
munogenicity, capacity to deliver large genes, and
large-scale production at low cost. The two leading
classes of synthetic gene delivery systems that have
been mostly investigated involve the use of either
cationic lipids or cationic polymers [2].
Cationic polymers, commonly named polycations,
are a leading class of molecular gene-delivery sys-
tems, in part because of their molecular diversity that
can be modified to fine-tune their physicochemical
properties [3,4]. Polyelectrolyte complexes (PEC)
formed between DNA and polycations have been
shown to tightly pack the DNA in the PEC complex,
so that the entrapped DNA is shielded from contact
with DNase [5]. Polycations commonly used in gene
delivery and transfection include polyethylenimine
[6], poly(L-lysine) [7], cationic dendrimers [8], poly-
brene [9], gelatin [10], tetraminofullerene [11],
poly(L-histidine)-graft-poly(L-lysine) [12], and cation-
ic polysaccharides. Although PEC systems have some
advantages over viral vectors, e.g. low immunogenic-
ity and easy manufacture [13,14], several problems
such as toxicity, lack of biodegradability, low bio-
compatibility and, in particular, low transfection effi-
ciency need to be solved prior to practical use [15].
Polycations used in gene delivery are polyamines that
become cationic at physiological conditions. All pol-
ymers contain primary, secondary, tertiary, or quater-
nary amino groups capable of forming electrostatic
complexes with DNA under physiological conditions.
Most polycations are toxic to cells and non-biode-
gradable, while the polymers based on amino acids
such as poly(L-lysines) are immunogenic [16]. More
advanced polymeric gene delivery systems employ
macromolecules with high cationic charge density that
act as endosomal buffering systems, thus suppressing
the endosomal enzymes activity and protecting the
DNA from degradation [6]. Among the various poly-
cations used in gene delivery and transfection, cation-
ic polysaccharides are considered to be the most
attractive candidates. They are natural, non-toxic,
biodegradable, and biocompatible materials and can
be modified easily for improved physicochemical
properties [17,18].
In recent publications [19,20], we reported on a
new type of biodegradable polycation based on
grafted oligoamine residues on natural polysacchar-
ides, which are effective in delivering plasmids for a
high biological effect. The use of biodegradable
polysaccharide carriers is especially suitable for trans-
fection and biological applications because they are
water soluble, can be readily transported to cells in
vivo by known biological processes, and act as
effective vehicles for transporting agents complexed
with them [21]. More than 300 different polycations
were prepared starting from various polysaccharides
of different molecular weights and oligoamines hav-
ing two to four amine groups. Although most of these
conjugates formed stable complexes with DNA as
determined by ethidium bromide quenching assay
[22], only the dextran-spermine (D-SPM) polycations
of defined molecular weights were found to be active
in transfecting a wide range of cell lines in vitro. The
reason for the transfection of D-SPM conjugate was
attributed to spermine residues, which play a crucial
role in cell transfection.
Success of non-viral gene delivery depends on the
type of carrier materials to bind plasmid DNA and
facilitate the cell uptake of carrier–DNA complex
[23–25]. One of the major obstacles limiting the in
vitro and in vivo gene delivery is the interaction of
carrier–DNA with extracellular fluids. This phenom-
enon is well known and is attributed to proteins
(serum components) adsorption on the surface of the
carrier–DNA complex, which in part could induce the
aggregation/deactivation of the complex and finally to
the reduction in the transfection efficiencies [26–28].
In an attempt to alter the characteristics of the devel-
oped D-SPM, PEGylated derivatives of the polymer
were recently synthesized and tested for their trans-
fection efficiencies in serum rich media. Such modi-
fication resulted in a substantial increase in gene
expression compared to unmodified D-SPM. Also,
in contrast to unmodified D-SPM, PEGylated-D-
SPM complexed with pSV-hGal was shown to induce
gene expression in the liver of mice after i.v. admin-
istration [29]. The use of PEG for this purpose is
attractive since PEG polymers are hydrophilic, safe,
cheap and do not interact with plasma components
[30].
The complexes formed between D-SPM and plas-
mid DNAs is expected to exhibit a hydrophilic surface

T. Azzam et al. / Journal of Controlled Release 96 (2004) 309–323 311
GEN
ED
ELIV
ER
Y
due to the saccharides building units of the polymer
backbone. Such hydrophilic nature is thought to limit
the cell uptake of the complexes due to the hydro-
phobic nature of the cell membrane. The chemical
modification of D-SPM polycation is expected to
exploit its potential in cell transfection. The objectives
of the present study was to further study the impor-
tance of grafted spermine residues in the success of
transfection, and to test a new series of hydrophobized
D-SPM derivatives in terms of enhancing transfection
and complex stability in serum rich media.
2. Materials and methods
2.1. Materials
All solvents and reagents were of analytical
grade and were used as received. Tetrahydrofuran
(THF) was dried by distillation over sodium/benzo-
phenone. A sage-metering pump model-365 (Orion,
NJ, USA) was used for slow and reproducible
addition of reactants. Dextran with an average
molecular weight of 40 kDa was obtained from
Sigma Chemical Co. (St. Louis, MO, USA). Sper-
mine, spermidine, potassium periodate, sodium bo-
rohydride, cholesteryl chloroformate, octanoyl
chloride, lauroyl chloride, myristoyl chloride,
stearoyl chloride, oleoyl chloride (technical 85%),
and diisopropylethylamine (DIEA) were obtained
from Fluka Chemie (Buchs, Switzerland). N,N-
bis(3-aminopropyl)-1,3-propanediamine [3:3:3],
N,N-bis(3-aminopropyl)-1,2-ethanediamine [3:2:3],
butanediamine, N,N-dimethylpropylenediamine, and
N-hydroxysuccinimide (NHS) were obtained from
Aldrich (Milwaukee, WI, USA). 1H-NMR spectra
were recorded on a Varian 300MHz instrument
using CDCl3, D2O or DMSO-d6 as solvents. Values
were recorded as ppm relative to internal standard
(TMS). Molecular weights of starting polymers and
conjugates were determined on GPC Spectra Phys-
ics instrument (Darmstadt, Germany) containing a
pump, column (Shodex KB-803, Japan) and refrac-
tive index (RI) detector. Molecular weights were
determined according to Pullulan standards (PSS,
Mainz, Germany) with molecular weights between
1700 and 212,000. Eluents used were 0.05 M
NaNO3 for the uncharged polymers and 0.25 M
sodium phosphate buffer (pH 4) for the cationic
polymers [31]. Elemental microanalysis was per-
formed on a Perkin–Elmer 2400/II analyzer.
2.2. Oxidation of dextran
Oxidized dextran was prepared and characterized
as described elsewhere [19,20].
2.3. Dextran-oligoamine conjugates
Dextran grafted with various oligoamines was
synthesized by the reductive amination method as
described earlier [19,20]. In brief, a solution of
dialdehyde dextran (6.25 mmol of aldehyde groups)
in 100 ml of DDW was slowly added over 5 h (sage
metering pump) to a basic solution containing 1.25
molar equivalents of spermine (or other oligoamine
mixtures) dissolved in 50 ml of borate buffer (0.1 M,
pH 11). The mixture was gently stirred at room
temperature for 24 h, NaBH4 (1 g, 4 equimolar)
was added, and stirring was continued for 48 h under
the same conditions. The reduction was repeated with
an additional portion of NaBH4 (1 g) and with stirring
for 24 h under the same conditions. The resulting
light-yellow solution was poured into a dialysis
membrane (3500 cutoff, Membrane Filtration Prod-
ucts Inc., San Antonio, TX, USA) and dialyzed
against DDW (6� 5 l) at 4 jC for 2 days. The
dialysate was gravimetrically filtered to remove
insolubles and lyophilized to dryness.
Yield: 40% (w/w).
FT-IR (KBr): 1468 (UCH2U, aliphatic), 1653
(UNH2, primary amine), 2935 (CUC, aliphatic),
and 3297 (secondary amine and UOH groups) cm� 1.
2.4. Determination of primary amines by the TNBS
method
The primary amine content was determined accord-
ing to standard protocol [32] with a slight modifica-
tion. In brief, a total of 20 Al of freshly prepared
aqueous TNBS solution (15 mg ml� 1) was separately
added to marked tubes containing up to 0.2 Amol
spermine (or other soluble oligoamine) dissolved in
600 Al of DDW. The mixtures were separately diluted
with 200 Al of sodium bicarbonate buffer (0.8 M, pH
8.5), vortexed for 1 min, and incubated for 2 h at 37

T. Azzam et al. / Journal of Controlled Release 96 (2004) 309–323312
GEN
ED
ELIV
ER
Y
jC. Then, 600 Al of 1 N HCl aqueous solution was
added to each tube, vortexed for 1 min, and gently
sonicated for 2 min to remove bubbles. Absorbances
of samples were recorded at 410 nm. A sample
containing the same composition (without the oligo-
amine) was used as a reference in the absorbance
measurements. Weighed conjugates (50–500 Ag,depending on the degree of conjugation) were treated
as above, and the primary amine content was calcu-
lated according to the calibration curve.
2.5. Synthesis of cholesterol–NHS carbonate
derivative
To 0.5 g of cholesteryl chloroformate (1.12 mmol)
dissolved in 25 ml of anhydrous THF was added 0.4
ml of anhydrous DIEA (2.5 equimolar) under nitrogen
atmosphere. The mixture was cooled to 0 jC and 200
mg of solid NHS (1.5 equimolar to chloroformate)
was added, and stirring was continued for 2 h at 0 jCand overnight at room temperature. The solvent was
removed under reduced pressure and the crude was
dissolved in 50 ml of diethyl ether and washed with
saturated aqueous NaCl solution (2� 20 ml) and
water (2� 20 ml). The ethereal phase was dried over
anhydrous MgSO4, filtered and evaporated to dryness.
The crude compound was crystallized in dichloro-
methane/methanol (1:5), stored at � 20 jC for 24 h,
filtered and dried in vacuum over P2O5.
Yield: 0.5 g (f 85%); Rf 0.65 (1% methanol in
dichloromethane).1H-NMR (CDCl3): 0.672 (s, 3H), 0.847 (d, 3H),
0.897 (d, 3H), 0.94–2.1 (m, 32H), 2.492 (d, 2H),
2.830 (s, 4H, UCH2CH2U of NHS group), 4.597 (m,
1H), and 5.422 (m, 1H) ppm.
2.6. Synthesis of fatty acid–NHS ester (general
method)
1.33 g of NHS (11.56 mmol) and 1.35 ml of DIEA
(9.5 mmol) were dissolved in 50 ml of anhydrous
THF under inert atmosphere. The mixture was cooled
to 0 jC and 7.8 mmol of the corresponding fatty acid
chloride in 50 ml of anhydrous THF was slowly added
during 1 h. The mixture was stirred at 0 jC for 2 h and
overnight at room temperature. The resulting precip-
itate salt was discarded by filtration and the filtrate
was evaporated under reduced pressure, redissolved in
small amount of dichloromethane and purified over
silica-gel using dichloromethane as eluent. Fractions
containing the product were collected and solvent was
removed under reduced pressure to obtain colorless
oil. The fatty acid–NHS ester was crystallized from
hot ethanol, collected by filtration and vacuum-dried
over P2O5.
The follows summarize the yields, Rf (TLC) and1H-NMR of all NHS-derivatives:
2.6.1. NHS-oleate
Yield: 92%; Rf 0.35 (CHCl3).1H-NMR(CDCl3): 0.874 (t, 3H), 1.38 (m, 20H),
1.74 (m, 2H), 2.004 (m, 4H), 2.594 (t, 2H), 2.83 (s,
4H) and 5.34 (m, 2H) ppm.
2.6.2. NHS-octanoate
Yield: 85%; Rf 0.43 (CHCl3).1H-NMR (CDCl3): 0.874 (t, 3H), 1.210–1.480 (m,
8H), 1.738 (m, 2H), 2.595 (t, 2H) and 2.832 (s, 4H)
ppm.
2.6.3. NHS-laurate
Yield: 90%; Rf 0.34 (CHCl3).1H-NMR (CDCl3): 0.874 (t, 3H), 1.214–1.435 (m,
18H), 1.736 (m, 2H), 2.594 (t, 2H) and 2.827 (s, 4H)
ppm.
2.6.4. NHS-myristate
Yield: 83%; Rf 0.32 (CHCl3).1H-NMR (CDCl3): 0.877 (t, 3H), 1.210–1.4930
(m, 20H), 1.765 (m, 2H), 2.597 (t, 2H) and 2.835 (s,
4H) ppm.
2.6.5. NHS-stearate
Yield: 95%; Rf 0.42 (CHCl3).1H-NMR (CDCl3): 0.867 (t, 3H), 1.234–1.453 (m,
28H), 1.735 (m, 2H), 2.589 (t, 2H) and 2.828 (s, 4H)
ppm.
2.7. Modification of dextran-spermine with oleic acid
D-SPM conjugate (20 mg, f 26 Amol of primary
amine, TNBS method) was dissolved in 1.5 ml
mixture of DDW:THF (1:2). The solution was vigor-
ously stirred at room temperature using a microstirrer
and 1–30% mol/mol (to primary amine) of stock
NHS-oleate solution in anhydrous THF was added.

T. Azzam et al. / Journal of Controlled Release 96 (2004) 309–323 313
GEN
ED
ELIV
ER
Y
The mixture was stirred at room temperature for 24
h and THF was removed by a flux of nitrogen, diluted
with DDW and dialyzed against DDW to remove
NHS and unbound fatty acids. The remaining mixture
was freeze-dried and the lyophilate was stored under
nitrogen atmosphere.
Average yield: 18 mg (f 90% w/w).1H-NMR (D2O): 0.696 (m, terminal methyl
group of oleate), 1.095 (m, oleate methylene groups),
1.437 (m, Dextran-NHCH2CH2CH2NHCH2CH2CH2
CH2NHCH2CH2CH2CH2NH2), 1.617 (m, Dextran-
NHCH2CH2CH2NHCH2CH2CH2CH2NHCH2CH2
CH2CH2NH2), 2.15–3.26 (m, Dextran-NHCH2CH2
CH2NHCH 2CH2CH2CH2NHCH2CH2 CH2CH2NH2),
3.30–4.45 (m, polysaccharide hydrogens), 4.773 (m,
anomeric hydrogens of glycosidic linkages), and 5.140
(m, double bond of oleate residues) ppm.
D-SPM derivatives modified with 20% mol/mol (to
primary amine) of other fatty acids and cholesterol
were similarly prepared and characterized by 1H-NMR.
2.8. Transfection studies
2.8.1. Preparation of plasmid DNAs
Plasmid DNAs used in the transfection experi-
ments were pCMV-GFP encoding green fluorescence
protein, pSV-hGal encoding h-galactosidase, and
pLNC-luc encoding luciferase. Plasmid DNAs were
separately amplified in a transformant of Escherichia
coli bacteria and isolated from the bacteria by
Qiagen Maxi kit-25 (Qiagen K.K., Tokyo, Japan).
Briefly, the grown bacteria were harvested and lysed
in an aqueous solution of 1 wt.% sodium dodecyl
sulfate (SDS) and 0.1 wt.% RNase A solution in
NaOH (pH 8), and the lysate was neutralized by the
addition of 3 M potassium acetate (pH 5.5). After
separation of the insoluble portion by use of QIA
filter cartridge, the lysate was applied to the Qiagen-
tip (anion-exchange resin), followed by washing with
a buffer containing 1 M NaCl to remove traces of
RNAs and proteins. The plasmid DNA was eluted
with an elution buffer containing 1.25 M NaCl at pH
8.5, de-salted, and precipitated by 2-propanol. The
precipitated plasmid DNA was centrifuged at 14,000
rpm for 10 min at 4 jC and washed twice with 70%-
ethanol aqueous solution to substitute 2-propanol for
ethanol. After centrifugation (14,000 rpm, 2 min, 4
jC), the resulting plasmid DNA was air-dried and
dissolved in a small volume of 10 mM Tris–HCl
and 1 mM EDTA buffer solution. When measured to
assess the purity of plasmid DNA obtained, the ratio
of the absorbance at 260 nm to that at 280 nm
ranged from 1.8 to 2.0.
2.8.2. In vitro transfection
Human embryonal kidney (HEK293), mouse
fibroblasts (NIH3T3), and cervical cancer (HeLa)
cells were used in the transfection experiments. 0.5
Ag of purified plasmid (Qiagen kit) per well of
transfected cells was mixed with D-SPM (or other
cationic derivatives) at a various weight-mixing ratios
ranging from 1 to 20 w/w (polycation/DNA). The
polycation/DNA complex mixtures were diluted to a
final volume of 200 Al with serum-free medium
(SFM) and allowed to stand at room temperature
for 30 min. Twenty-four-well plates, seeded 24 h be-
fore transfection with 1.5� 105 cells per well, were
washed with SFM and the solution of the complexes
was added to the cell wells and cultured at 37 jC in
95% air/5% CO2. 4 h post transfection, cell medium
was replaced with Dulbecco’s modified Eagle’s Me-
dium (DMEM) containing 10% fetal calf serum
(FCS), and the cells were cultured for 48–72 h under
growth conditions. At this time growth medium was
removed, and cell lysates were formed and analyzed
for gene expression. h-Gal kit (Invitrogen Co., USA)
and luciferase assay kit (Promega) were used accord-
ing to manufacturer’s protocols for the evaluation of
h-galactosidase and luciferase activity, respectively.
Cells transfected with pCMV-GFP were analyzed for
gene expression using fluorescence microscope in-
strument (model Axiovert 35, Zeiss, Jena, Germany).
The yield of transfection (% transfection) was calcu-
lated by counting the fluorescent cells in a field of a
particular well, and dividing the number of fluores-
cent cells by the number of total cells in the same
field. In some cases, the degree of gene expression
was normalized to total protein content using the
standard BCA assay kit (Pierce, USA). For the
transfection experiments in serum-containing medi-
um, SFM was replaced with 10% FCS at the com-
plexation and uptake stages, followed by the same
procedure described above.
Cell transfection with Calcium phosphate reagent
(Sigma) was performed according to a well docu-
mented protocol [33,34]. Transfection applying

T. Azzam et al. / Journal of Controlled Release 96 (2004) 309–323314
GEN
ED
ELIV
ER
Y
DOTAP/Chol 1/1 (Avanti Polar lipids Inc., Alabama,
USA) and FuGen 6 (Roche Diagnostics, Indianapolis,
IN) based lipid formulations were performed accord-
ing to manufacturer’s protocols.
3. Results and discussion
3.1. Chemistry
3.1.1. Dextran-oligoamine based conjugates: synthe-
sis and characterization
Grafting of the representative oligoamine or
oligoamines mixtures to oxidized dextran was per-
formed by means of reductive amination method as
described earlier [19,20]. The oligoamines used for
conjugation were the naturally occurring spermine
and spermidine, and other oligoamines of two to
four amine groups. Also, dextran-oligoamine con-
jugates substituted with two different oligoamines at
various mole ratios were prepared and characterized
(Scheme 1).
Scheme 1. Reductive amination of typical dextran gra
The polycations were characterized by nitrogen
elemental analysis (%N), primary amine content
(TNBS) and average molecular weights (GPC) as
shown in Table 1. Group A represents seven dextran
derivatives conjugated to spermine (#1), spermidine
(#7), and spermine/spermidine mixed oligoamines
(#2–6) at varying mole ratios ranging from 9/1 to
1/9, respectively. The average nitrogen (%N) and
primary amine contents found for this group was
11.0F 0.30 and 1.25F 0.08 (Amol mg� 1), respec-
tively. The remarkable decrease in the average
molecular weights of conjugates in comparison to
starting dextran (40 kDa) is explained by the
random and extensive aminolysis of glycoside link-
ages during conjugation [35].
Group B of Table 1 shows a series of cationic
dextran derivatives conjugated to various commercial
oligoamines and their mixtures with spermine. The
derivative conjugated to N,N-dimethyl-1,3-propanedi-
amine (#7, Group B) was shown to exhibit a low and
negligible TNBS value as expected due to the absence
of primary amine groups. On the other hand, when
fted to mixed spermine/spermidine oligoamines.

Table 1
Chemical characterization of dextran derivatives grafted with various oligoamine mixturesa
# Substituted oligoamine(s)b % SPMc %Nd Amol mg� 1 (TNBS)e Mw (kD) Mn (kD) P
Group A
1 SPM 100 11.28 1.37F 0.05 10.07 4.50 2.23
2 SPM/SPD 90 10.89 1.29F 0.04 11.74 4.77 2.46
3 SPM/SPD 70 11.35 1.20F 0.12 9.43 4.10 2.29
4 SPM/SPD 50 10.59 1.10F 0.10 11.08 4.35 2.54
5 SPM/SPD 30 11.34 1.26F 0.05 8.87 3.92 2.26
6 SPM/SPD 10 10.68 1.36F 0.03 11.50 4.68 2.45
7 SPD 0 10.96 1.22F 0.02 10.80 4.47 2.42
Group B
1 [3:3:3] 0 9.58 1.07F 0.08 6.14 4.19 1.46
2 SPM/[3:3:3] 70 10.51 1.37F 0.02 7.42 3.78 1.96
3 [3:2:3] 0 9.67 1.28F 0.05 6.95 5.19 1.34
4 SPM/[3:2:3] 70 11.29 1.41F 0.03 10.48 4.34 2.41
5 [4] 0 9.05 2.43F 0.12 5.70 4.68 1.22
6 SPM/[4] 70 9.24 1.17F 0.04 8.89 4.02 2.21
7 N-[3]-N(CH3)2 0 9.68 0.05F 0.02 13.72 11.03 1.24
8 SPM/N-[3]-N(CH3)2 70 10.76 1.09F 0.04 11.05 4.60 2.40
a Reaction conditions: oxidized dextran (f 50% dialdehyde) and the appropriate oligoamine or oligoamine mixtures (1/1.25, aldehyde/(total
oligoamine)) were allowed to react under similar conditions as described in the Experimental section.b Abbreviations: SPM (spermine); SPD (spermidine); [3:3:3] (N,N-bis(3-aminopropyl)-1,3-propanediamine); [3:2:3] (N,N-bis(3-amino-
propyl)-1,2-ethanediamine); [4] (1,4-butanediamine); N-[3]-N(CH3)2 (N,N-dimethyl-1,3-propanediamine).c Percent of spermine content in dextran– (mixed oligoamine) conjugate.d Found nitrogen content (elemental analysis).e Amount of primary amine (Amol mg� 1) determined by the TNBS method (meanF S.D.). Mw, Mn, and polydispersity ( P=Mw/Mn) were
determined by GPC as described in the Experimental section for the polycationic polymers.
T. Azzam et al. / Journal of Controlled Release 96 (2004) 309–323 315
GEN
ED
ELIV
ER
Y
spermine was added in excess to N,N-dimethyl-1,3-
propanediamine oligoamine (#8, Group B), a relative-
ly high primary amine content was obtained (f 1.1
Amol mg� 1).
3.1.2. Hydrophobization of dextran-spermine with
fatty acids and cholesterol
Preliminary hydrophobization attempts of D-SPM
conjugate with fatty acids chlorides resulted in a low
degree of modification, probably due to rapid hydro-
lysis of the acyl chloride groups in aqueous media
(data not shown). Therefore, NHS-activated fatty
acids were prepared and applied for the modification
of D-SPM. Such derivatives were previously applied
and with high coupling yields in the modification of
monoclonal antibodies [36] and more recently to
poly(L-lysine)s [37,38]. Saturated fatty acid chlorides
of C8–C18 and oleyl chloride were treated with
NHS under anhydrous conditions and in the presence
of base to obtain the corresponding NHS-ester deriv-
atives in relatively high yields. These derivatives
were purified by silica-gel column chromatography
and crystallized from hot ethanol. The purity of the
compounds was >99% as determined by TLC and1H-NMR. Cholesterol activated with NHS-carbonate
group was similarly prepared from NHS and choles-
teryl chloroformate.
D-SPM conjugate was hydrophobized with in-
creasing amounts of fatty acid (1–30% mol/mol, fatty
acid/primary amine) by adding the corresponding
fatty-acid NHS ester to a concentrated solution of
D-SPM in a mixture of water/THF (Scheme 2).
Dialysis was proved efficient to purify the modified
polycations from liberated NHS and traces of un-
bound fatty acids as evaluated by TLC. More than
95% binding (relative to starting feed) was obtained
as determined by 1H-NMR. At low degrees of
fatty acid/cholesterol modification (1 to about 20%
mol/mol), water-soluble derivatives were obtained.
Higher degrees of modification (20% and 30% mol/
mol) resulted in slightly turbid solutions in aqueous
media.

Scheme 2. Hydrophobization of D-SPM conjugate with NHS-oleate.
T. Azzam et al. / Journal of Controlled Release 96 (2004) 309–323316
GEN
ED
ELIV
ER
Y
3.2. Transfection studies
3.2.1. Role of grafted spermine residues in cell
transfection
Various cationic polysaccharide derivatives hav-
ing multiple amine functionalities of various
grafted oligoamines were previously developed
and tested for their transfection efficiencies in a
wide range of cell lines [19]. Although most of
these conjugates formed stable complexes with
DNAs as determined by the ethidium bromide
quenching assay [22], only the D-SPM based
polycation was found to be active in cell transfec-
tion. The reason for the transfection efficiency of
certain polycations (i.e. D-SPM) is probably related
to the unique complexation properties and the
buffering capacity of grafted spermine residues
[39]. To emphasize the importance of grafted
spermine moieties in cell transfection, a series of
polycations were synthesized by grafting dextran
with mixtures of spermine and other oligoamines
as illustrated in Table 1.
In the first experiment, dextran polycations
grafted with a mixture of spermine/spermidine at
varying mole ratios (Group A, Table 1) were tested
for their transfection efficiencies. The mole ratios of
grafted spermine/spermidine were varied from 9/1 to
1/9, respectively. In addition, two control polycations
were grafted with either spermine or spermidine. The
transfection efficiencies of these polymers were eval-
uated in NIH3T3/pLNC-luc system as a function of
spermine percentages to total grafted oligoamine, and
the results were recorded at the optimal weight-
mixing ratio (polymer/DNA) as shown in Fig. 1a.
When spermine (without spermidine) was grafted to
dextran polysaccharide, high luciferase activity
(f 30,000 RLU/mg protein) was obtained similar
to a DOTAP/Chol 1/1 cationic lipid control (data not
shown). Incorporation of spermidine at 1/9 mole
ratio (i.e. 90% spermine) resulted in a 20% decrease
in luciferase expression. Higher spermidine substitu-
tions resulted in sharp loss of the luciferase expres-
sion (Fig. 1a). In a similar transfection experiment,
conjugates grafted with mixtures of spermine and
other oligoamines (Group B, Table 1) were tested for
their transfection efficiencies in a HEK293/pSV-hGalsystem (Fig. 1b). Dextran conjugated to N,N-bis(3-
aminopropyl)-1,3-propanediamine ([3:3:3], #1), N,N-
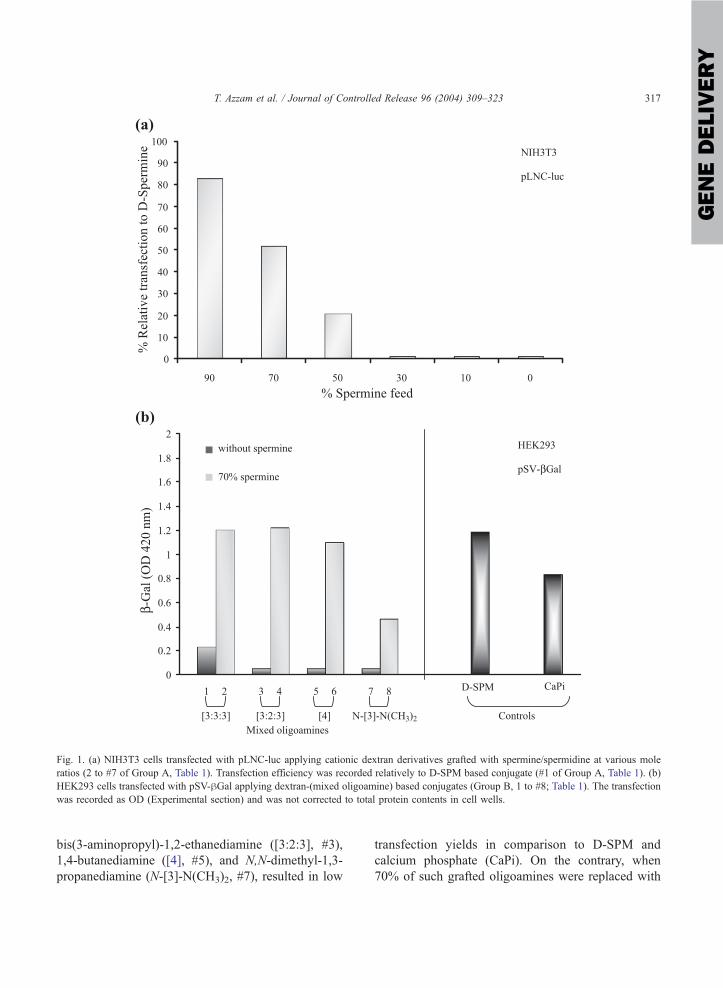
Fig. 1. (a) NIH3T3 cells transfected with pLNC-luc applying cationic dextran derivatives grafted with spermine/spermidine at various mole
ratios (2 to #7 of Group A, Table 1). Transfection efficiency was recorded relatively to D-SPM based conjugate (#1 of Group A, Table 1). (b)
HEK293 cells transfected with pSV-hGal applying dextran-(mixed oligoamine) based conjugates (Group B, 1 to #8; Table 1). The transfection
was recorded as OD (Experimental section) and was not corrected to total protein contents in cell wells.
T. Azzam et al. / Journal of Controlled Release 96 (2004) 309–323 317
GEN
ED
ELIV
ER
Y
bis(3-aminopropyl)-1,2-ethanediamine ([3:2:3], #3),
1,4-butanediamine ([4], #5), and N,N-dimethyl-1,3-
propanediamine (N-[3]-N(CH3)2, #7), resulted in low
transfection yields in comparison to D-SPM and
calcium phosphate (CaPi). On the contrary, when
70% of such grafted oligoamines were replaced with
T. Azzam et al. / Journal of Controlled Release 96 (2004) 309–323318
GEN
ED
ELIV
ER
Y
spermine (polymers 2, 4, 6 and #8; Fig. 1b), high
luciferase expression was obtained similar to D-SPM
and calcium phosphate. These results strongly indi-
cate the effect of the oligoamine structure on the
transfection yield, with spermine being the most
effective.
3.2.2. Hydrophobized dextran-spermine derivatives
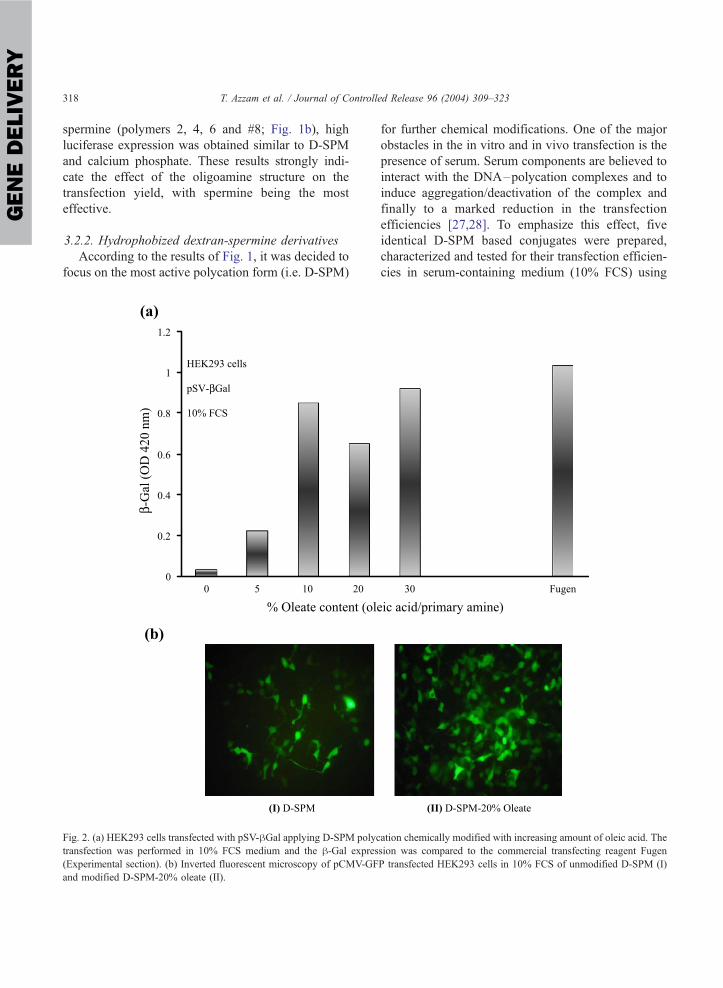
According to the results of Fig. 1, it was decided to
focus on the most active polycation form (i.e. D-SPM)
Fig. 2. (a) HEK293 cells transfected with pSV-hGal applying D-SPM polyc
transfection was performed in 10% FCS medium and the h-Gal expres(Experimental section). (b) Inverted fluorescent microscopy of pCMV-GF
and modified D-SPM-20% oleate (II).
for further chemical modifications. One of the major
obstacles in the in vitro and in vivo transfection is the
presence of serum. Serum components are believed to
interact with the DNA–polycation complexes and to
induce aggregation/deactivation of the complex and
finally to a marked reduction in the transfection
efficiencies [27,28]. To emphasize this effect, five
identical D-SPM based conjugates were prepared,
characterized and tested for their transfection efficien-
cies in serum-containing medium (10% FCS) using
ation chemically modified with increasing amount of oleic acid. The
sion was compared to the commercial transfecting reagent Fugen
P transfected HEK293 cells in 10% FCS of unmodified D-SPM (I)

T. Azzam et al. / Journal of Controlled Release 96 (2004) 309–323 319
GEN
ED
ELIV
ER
Y
the NIH3T3/pLNC-luc system. The presence of 10%
FCS remarkably reduced the luc expression by about
80–85% compared with the expression in SFM (data
not shown).
In order to improve the transfection in the pres-
ence of serum, fatty side groups were attached to the
polycation to protect the polymer–DNA complex
from serum proteins, and facilitate cellular uptake
of hydrophobic complexes through the cell mem-
brane [40,41]. The majority of cationic lipids com-
monly used in cell transfection are composed of di-
oleate hydrophobic tails connected via various
spacers to different cationic head groups [42–45].
In analogy to these constructs, the leading polycation
(i.e. D-SPM) was modified with increasing amounts
of oleic acid to evaluate the transfection efficiencies
of these modified derivatives as functions of oleate
and serum contents.
Fig. 2a summarizes the effect of oleate content of
D-SPM on the transfection efficiencies. The transfec-
tion was performed in 10% FCS medium applying the
HEK293/pSV-hGal system. Unmodified D-SPM (i.e.
0% oleate) resulted in low expression yields (OD
0.032) as expected. The highest hGal activities whereobtained with 10–30% oleate contents where nearly
equal expression to the FuGen 6 (Roche Diagnostics)
control was obtained. Fig. 2b shows a typical fluo-
rescent imaging of pCMV-GFP transfected HEK293
cells using D-SPM conjugate (I), and D-SPM-20%
oleate (II) in 10% FCS. It can be clearly concluded
Fig. 3. Effect of oleate content of D-SPM on the transfection efficiencies in
the activity obtained with unmodified D-SPM in SFM. The data was reco
from both images that oleate modification significant-
ly increased the transfection yields. Similar substantial
increase in the transfection efficiency of D-SPM-20%
oleate was obtained with the HeLa cell line (data not
shown).
The effect of oleate modification of D-SPM on
the transfection efficiencies in higher serum-contain-
ing medium (i.e. 20% FCS) was evaluated in
NIH3T3 cells applying pLNC-luc as the marker
gene. Fig. 3 shows the transfection efficiencies of a
series of three D-SPM conjugates modified with
increasing amounts of oleate (10–30% mol/mol).
The transfection yields were recorded relative to
luciferase readings obtained with unmodified D-
SPM in parallel transfection experiment conducted
with the same cell line and in SFM. The transfection
yields in this experiment were recorded as
meanF S.D. (n = 5). At 10% and 20% oleate mod-
ifications, nearly 25% and 45%, respectively, of the
gene expression was retained relative to the gene
expression obtained with unmodified D-SPM in
SFM. Modification of the polymer with 30% oleate
resulted in nearly 75% preservation of the gene
expression in comparison to D-SPM in SFM.
The substantial increase in the transfection effi-
ciencies of the hydrophobized D-SPM in serum-rich
media was attributed, as stated earlier, to the chemi-
cally bound oleate moieties which act as shielding
agents and keep the complexes intact from interaction
with serum components. To emphasize the importance
20% FCS medium. The luciferase activity was recorded relatively to
rded as meanF S.D. (n= 5).

Table 2
NIH3T3 transfected pLNC-luc applying D-SPM conjugate modified
in two forms with increasing amounts of oleic acid
% Oleate contenta Luciferase (RLU per mg protein)b
Chemical
modificationcPhysical
formulationd
5 1493 Low ( < 25)
10 1650 Low ( < 25)
20 2817 Low ( < 25)
30 10,240 Low ( < 25)
DOTAP/Chol (1/1)e 9980
The transfection applying all derivatives and formulations were
performed in 20% FCS medium unless sited otherwise.a Degree of modification in mol/mol (oleic acid/primary amine).b Relative Light Units (RLU) of luciferase normalized to 1 mg
protein (Experimental section).c Chemically bound oleic acid via amide linkage.d Physically bound oleic acid via ionic interactions.e DOTAP/Chol (1/1) lipid control was evaluated in 10% FCS
medium.
T. Azzam et al. / Journal of Controlled Release 96 (2004) 309–323320
GEN
ED
ELIV
ER
Y
of the chemically bound oleate moieties in increasing
the stability against serum components, D-SPM for-
mulations with increasing amounts of unbound oleic
acid were prepared and tested for their transfection
activities in serum-rich media. Such formulations
were prepared by simply mixing a water solution of
the D-SPM with an appropriate amount of oleic acid
in THF. The mixtures were allowed to stir at room
Fig. 4. HEK293 cells transfected with pCMV-GFP applying D-SPM conju
various saturated fatty acids (C8–C18), and cholesterol. Transfection w
qualitatively by fluorescent microscopy (Experimental section). D-SPM-2
temperature for at least 1 h, followed by evaporation
of THF by a flux of nitrogen gas and lyophilization to
dryness. The resulting formulations exhibit oleate
moieties connected to D-SPM by weak acid–base
salt interactions (UNH3+ �OOCU), which could be
easily replaced upon interaction with DNA.
Table 2 shows the transfection efficiencies in 20%
FCS medium (NIH3T3/pLNC-luc) obtained with D-
SPM polycations covalently-modified or ionically-
bound with increasing amounts of oleic acid (5–30%
mol/mol). Chemical modification with oleate resulted
with an increase in the transfection efficacy, whereas
when the ionically-bound oleate formulations were
applied, negligible luciferase readings were obtained
with all oleate contents. The results strongly empha-
size that chemically bound oleate moieties act as
shielding agents that increase the stability of poly-
mer–DNA complex against serum proteins. DOTAP/
Chol (1:1) in 10% FCS medium showed similar
luciferase readings compared to D-SPM-20% oleate
in 20% FCS medium system.
To further evaluate the importance of oleate
moieties to the altered transfection characteristics,
D-SPM was chemically modified with other types of
hydrophobic moieties and the transfection efficien-
cies of these derivatives were qualitatively evaluated
in HEK293/pCMV-GFP system (Fig. 4). The hydro-
gate chemically modified with 20% mol/mol (to primary amine) of
as performed in 10% FCS and the%transfection was determined
0% oleate and Fugen were used as positive controls.

T. Azzam et al. / Journal of Controlled Release 96 (2004) 309–323 321
GEN
ED
ELIV
ER
Y
phobic group content in all derivatives was chosen
to be 20% (mol/mol) based on the result described
above. The hydrophobic moieties used in the mod-
ification were saturated fatty acids of C8–C18, and
cholesterol. D-SPM-20% oleate and FuGen 6 were
used as positive controls. Although these derivatives
showed improved transfection characteristics in com-
parison to unmodified D-SPM, no substantial in-
crease in the transfection efficiencies was observed.
These results suggest that oleate moieties enhance
cellular uptake in marked extent compared to other
fatty acids and cholesterol. Current studies focus on
understanding the mechanism of action of D-SPM-
oleate by means of cellular uptake and trafficking.
4. Conclusions
A series of cationic polysaccharides were synthe-
sized by grafting dextran with spermine in combination
with other oligoamines of two to four amino groups.
The transfection efficiencies of these polycations were
assessed in various cell lines and marker genes, and
found to depend on the content of grafted spermine.
Polycations grafted with higher spermine content
resulted in high gene expression, whereas lower sper-
mine content resulted in low transfection yields. The
results emphasize the importance of spermine moieties
in cell transfection, probably due to the altered buffer-
ing capacity and complexation of spermine residues
compared with other grafted oligoamines.
The leading D-SPM polycation was further modi-
fied with increasing amounts of hydrophobic moieties
including C8–C18 saturated fatty acids, oleic acid and
cholesterol. Although hydrophobized polycations
showed altered transfection characteristics compared
to the unmodified D-SPM, only the oleate-modified
polycations showed significant increase in the trans-
fection efficiencies. The oleate-modified D-SPM fa-
cilitated transfection in serum containing medium (up
to 20% FCS), in marked contrast to unmodified D-
SPM which resulted in low to negligible transfection
yields in 10% and 20% FCS. The altered transfection
yields obtained with oleate-modified D-SPM were
explained by the unique hydrophobic nature of oleate
residues which enhances on the one hand cellular
uptake, and on the other hand reduces the interaction
with serum components.
Acknowledgements
Tony Azzam is grateful to the Ministry of Science,
Israel, for the financial assistance. This work was
supported in part by the AFIRST, French–Israeli
Cooperation on Gene Therapy, and by the US–Israel
Binational Fund (BSF).
References
[1] J. Gaucheron, C. Boulanger, C. Santaella, N. Sbirrazzuoli, O.
Boussif, P. Vierling, In vitro cationic lipid-mediated gene de-
livery with fluorinated glycerophosphoethanolamine helper
lipids, Bioconjugate Chem. 12 (6) (2001) 949–963.
[2] A.V. Kabanov, Taking polycation gene delivery systems from
in vitro to in vivo, Pharm. Sci. Technol. Today 2 (9) (1999)
365–372.
[3] A.J. Domb, M. Levy, Polymers in Gene Therapy: Frontiers in
Biological Polymer Application, Technomic, Lancaster, PA,
1999.
[4] D. Putnam, C.A. Gentry, D.W. Pack, R. Langer, Polymer-
based gene delivery with low cytotoxicity by a unique balance
of side-chain termini, Proc. Natl. Acad. Sci. USA 98 (3)
(2001) 1200–1205.
[5] T. Sato, T. Kawakami, N. Shirakawa, Y. Okahata, Preparation
and characterization of DNA–lipoglutamate complexes, Bull.
Chem. Soc. Jpn. 68 (9) (1995) 2709–2715.
[6] O. Boussif, F. Lezoualch, M.A. Zanta, M.D. Mergny, D.
Scherman, B. Demeneix, J.P. Behr, A versatile vector for gene
and oligonucleotide transfer into cells in culture and in-vivo-
polyethylenimine, Proc. Natl. Acad. Sci. USA 92 (16) (1995)
7297–7301.
[7] D. Oupicky, K.A. Howard, C. Konak, P.R. Dash, K. Ulbrich,
L.W. Seymour, Steric stabilization of poly-L-lysine/DNA com-
plexes by the covalent attachment of semitelechelic poly N-(2-
hydroxypropyl)methacrylamide, Bioconjugate Chem. 11 (4)
(2000) 492–501.
[8] A.U. Bielinska, A. Yen, H.L. Wu, K.M. Zahos, R. Sun,
N.D. Weiner, J.R. Baker, B.J. Roessler, Application of mem-
brane-based dendrimer/DNA complexes for solid phase
transfection in vitro and in vivo, Biomaterials 21 (9)
(2000) 877–887.
[9] R.J. Mumper, J.G. Duguid, K. Anwer, M.K. Barron, H. Nitta,
A.P. Rolland, Polyvinyl derivatives as novel interactive poly-
mers for controlled gene delivery to muscle, Pharm. Res. 13
(5) (1996) 701–709.
[10] K.W. Leong, H.Q. Mao, V.L. Truong-Le, K. Roy, S.M. Walsh,
J.T. August, DNA-polycation nanospheres as non-viral
gene delivery vehicles, J. Control. Release 53 (1–3) (1998)
183–193.
[11] H. Isobe, S. Sugiyama, K. Fukui, Y. Iwasawa, E. Nakamura,
Atomic force microscope studies on condensation of plasmid
DNA with functionalized fullerenes, Angew. Chem. Int. Ed.
40 (18) (2001) 3364–3367.

T. Azzam et al. / Journal of Controlled Release 96 (2004) 309–323322
GEN
ED
ELIV
ER
Y
[12] J.M. Benns, J.S. Choi, R.I. Mahato, J.S. Park, S.W. Kim, pH-
sensitive cationic polymer gene delivery vehicle: N-Ac-
poly(L-histidine)-graft-poly(L-lysine) comb shaped polymer,
Bioconjugate Chem. 11 (5) (2000) 637–645.
[13] D. Deshpande, P. Blezinger, R. Pillai, J. Duguid, B. Freimark,
A. Rolland, Target specific optimization of cationic lipid-
based systems for pulmonary gene therapy, Pharm. Res. 15
(9) (1998) 1340–1347.
[14] D. Oupicky, A.L. Parker, L.W. Seymour, Laterally stabilized
complexes of DNAwith linear reducible polycations: strategy
for triggered intracellular activation of DNA delivery vectors,
J. Am. Chem. Soc. 124 (1) (2002) 8–9.
[15] C.W. Pouton, L.W. Seymour, Key issues in non-viral gene
delivery, Adv. Drug Deliv. Rev. 46 (1–3) (2001) 187–203.
[16] S. Vanderkerken, T. Vanheede, V. Toncheva, E. Schacht, M.A.
Wolfert, L. Seymour, A. Urtti, Synthesis and evaluation of
poly(ethylene glycol)-polylysine block copolymers as carriers
for gene delivery, J. Bioact. Compat. Polym. 15 (2) (2000)
115–138.
[17] P.C. Berscht, B. Nies, A. Liebendorfer, J. Kreuter, In-vitro
evaluation of biocompatibility of different wound dressing
materials, J. Mater. Sci. Mater. Med. 6 (4) (1995) 201–205.
[18] B. Carreno Gomez, R. Duncan, Evaluation of the biological
properties of soluble chitosan and chitosan microspheres, Int.
J. Pharm. 148 (2) (1997) 231–240.
[19] T. Azzam, H. Eliyahu, L. Shapira, M. Linial, Y. Barenholz,
A.J. Domb, Polysaccharide-oligoamine based conjugates for
gene delivery, J. Med. Chem. 45 (9) (2002) 1817–1824.
[20] T. Azzam, A. Raskin, A. Makovitzki, H. Brem, P. Vierling,
M. Lineal, A.J. Domb, Cationic polysaccharides for gene
delivery, Macromolecules 35 (27) (2002) 9947–9953.
[21] C. Larsen, in: C. Larsen (Ed.), Dextran Prodrugs, Christesen
VA, Copenhagen, Denmark, 1990.
[22] T. Azzam, H. Eliyahu, A. Makovitzki, A.J. Domb, Dextran-
spermine conjugate: an efficient vector for gene delivery,
Macromol. Symp. 195 (2003) 247–261.
[23] R.I. Mahato, Y. Takakura, M. Hashida, Nonviral vectors for in
vivo gene delivery: physicochemical and pharmacokinetic
considerations, Crit. Rev. Ther. Drug Carrier Syst. 14 (2)
(1997) 133–172.
[24] E. Tomlinson, A.P. Rolland, Controllable gene therapy—phar-
maceutics of non-viral gene delivery systems, J. Control. Re-
lease 39 (2–3) (1996) 357–372.
[25] C.W. Pouton, L.W. Seymour, Key issues in non-viral gene
delivery, Adv. Drug Deliv. Rev. 34 (1) (1998) 3–19.
[26] M. Ogris, S. Brunner, S. Schuller, R. Kircheis, E. Wagner,
PEGylated DNA/transferrin–PEI complexes: reduced interac-
tion with blood components, extended circulation in blood and
potential for systemic gene delivery, Gene Ther. 6 (4) (1999)
595–605.
[27] D. Oupicky, M. Ogris, K.A. Howard, P.R. Dash, K. Ulbrich,
L.W. Seymour, Importance of lateral and steric stabilization of
polyelectrolyte gene delivery vectors for extended systemic
circulation, Mol. Ther. 5 (4) (2002) 463–472.
[28] M. Kursa, G.F. Walker, V. Roessler, M. Ogris, W. Roedl,
R. Kircheis, E. Wagner, Novel shielded transferrin-polyeth-
ylene glycol-polyethylenimine/DNA complexes for systemic
tumor-targeted gene transfer, Bioconjugate Chem. 14 (1)
(2003) 222–231.
[29] H. Hosseinkhani, T. Azzam, Y. Tabata, A.J. Domb, Dextran-
spermine polycation: an efficient non-viral vector for in
vitro and in vivo gene transfection, Gene Ther. 11 (2004)
194–203.
[30] R.B. Pepinsky, R.I. Shapiro, S.S. Wang, A. Chakraborty, A.
Gill, D.J. Lepage, D.Y. Wen, P. Rayhorn, G.S.B. Horan, F.R.
Garber, E.A. Garber, A. Galdes, T.M. Engber, Long-acting
forms of sonic hedgehog with improved pharmacokinetic
and pharmacodynamic properties are efficacious in a nerve
injury model, J. Pharm. Sci. 91 (2) (2002) 371–387.
[31] N. Kumar, T. Azzam, A.J. Domb, Molecular mass distribution
of polycations and dextrans by high-performance size exclu-
sion chromatography, Polym. Adv. Technol. 13 (10 –12)
(2002) 1071–1077.
[32] S.L. Snyder, P.Z. Sobocinski, An improved 2,4,6-trinitroben-
zenesulfonic acid method for the determination of amines,
Anal. Biochem. 64 (1) (1975) 284–288.
[33] M. Wigler, A. Pellicer, S. Silverstein, R. Axel, Biochemical
transfer of single-copy eukaryotic genes using total cellular
DNA as donor, Cell 14 (3) (1978) 725–731.
[34] M. Wigler, A. Pellicer, S. Silverstein, R. Axel, G. Urlaub, L.
Chasin, DNA-mediated transfer of the adenine phosphoribo-
syltransferase locus into mammalian-cells, Proc. Natl. Acad.
Sci. USA 76 (3) (1979) 1373–1376.
[35] O. Siiman, J. Wilkinson, A. Burshteyn, P. Roth, S. Ledis,
Fluorescent neoglycoproteins: antibody–aminodextran–phy-
cobiliprotein conjugates, Bioconjugate Chem. 10 (6) (1999)
1090–1106.
[36] A. Huang, L. Huang, S.J. Kennel, Monoclonal-antibody
covalently coupled with fatty-acid—a reagent for in vitro
liposome targeting, J. Biol. Chem. 255 (17) (1980)
8015–8018.
[37] W. Wang, L. Tetley, I.F. Uchegbu, A new class of amphiphilic
poly-L-lysine based polymers forms nanoparticles on probe
sonication in aqueous media, Langmuir 16 (20) (2000)
7859–7866.
[38] W. Wang, L. Tetley, I.F. Uchegbu, The level of hydrophobic
substitution and the molecular weight of amphiphilic poly-L-
lysine-based polymers strongly affects their assembly into
polymeric bilayer vesicles, J. Colloid Interf. Sci. 237 (2)
(2001) 200–207.
[39] H. Hosseinkhani, Y. Tabata, In vitro gene expression by cat-
ionized derivatives of an artificial protein with repeated RGD
sequences, pronectin (R), J. Control. Release 86 (1) (2003)
169–182.
[40] I. Koltover, T. Salditt, J.O. Radler, C.R. Safinya, An inverted
hexagonal phase of cationic liposome–DNA complexes relat-
ed to DNA release and delivery, Science 281 (5373) (1998)
78–81.
[41] I. Koltover, K. Wagner, C.R. Safinya, DNA condensation in
two dimensions, Proc. Natl. Acad. Sci. USA 97 (26) (2000)
14046–14051.
[42] P.L. Felgner, T.R. Gadek, M. Holm, R. Roman, H.W. Chan,
M. Wenz, J.P. Northrop, G.M. Ringold, M. Danielsen, Lip-
ofection—a highly efficient, lipid-mediated DNA—transfec-

T. Azzam et al. / Journal of Controlled Release 96 (2004) 309–323 323
EN
ED
ELIV
ER
Y
tion procedure, Proc. Natl. Acad. Sci. USA 84 (21) (1987)
7413–7417.
[43] I. Solodin, C.S. Brown, M.S. Bruno, C.Y. Chow, E.H. Jang,
R.J. Debs, T.D. Heath, A novel series of amphiphilic imida-
zolinium compounds for in-vitro and in-vivo gene delivery,
Biochemistry 34 (41) (1995) 13537–13544.
[44] N.S. Templeton, D.D. Lasic, P.M. Frederik, H.H. Strey, D.D.
Roberts, G.N. Pavlakis, Improved DNA: liposome complexes
for increased systemic delivery and gene expression, Nat. Bio-
technol. 15 (7) (1997) 647–652.
[45] J. Gaucheron, C. Santaella, P. Vierling, Highly fluorinated
lipospermines for gene transfer: synthesis and evaluation of
their in vitro transfection efficiency, Bioconjugate Chem. 12
(1) (2001) 114–128.
G




























